Factors determining the dynamics of toxic blooms of Alexandrium minutum during a 10-year study along the shallow southwestern Mediterranean coasts Q3 Moufida Abdenadher a , Asma Hamza a , Wafa Fekih a , Imen Hannachi b , Amel Zouari Bellaaj a , Mohamed Nejmeddine Bradai a , Lotfi Aleya c, * a Institut National des Sciences et Technologie de la Mer, Centre de Sfax, BP 1035, Sfax 3018, Tunisie b Université de Sfax, Faculté des Sciences de Sfax, Département des Sciences de la Vie, Unité de recherche 00/UR/0907 Ecobiologie, Planctonologie and Microbiologie des Ecosystèmes Marins, Route Soukra Km 3.5, BP 1171, CP 3000 Sfax, Tunisie c Université de Franche-Comté, Laboratoire de Chrono-Environnement, UMR CNRS 6249, 1 Place Leclerc, F-25030 Besançon cedex, France article info Article history: Received 24 July 2011 Accepted 28 April 2012 Available online xxx Keywords: Alexandrium minutum distribution beach samples southwestern Mediterranean HAB abstract Many blooms of the toxic dinoflagellate Alexandrium minutum have been recorded since 1990 in the Gulf of Gabes (southwestern Mediterranean Sea). To understand the determining factors of bloom formation, we studied the distribution of A. minutum in relation to environmental factors in samples taken at shallow sandy (<1 m) beach stations and a lagoon between 1997 and 2006. This was accompanied by laboratory experiments to identify A. minutum. The species forms harmful algal blooms (HABs) in stations subjected to anthropogenic eutrophication and in confined lagoons, living under conditions of varying salinity which gives it a unique opportunity to broaden its physiological tolerance and increase its colonisation potential. Increases in phosphorus appear to be more important than nitrogen or temperature in the control of A. minutum. The stations sampled all along the coast present specific hydrographic properties (shallowness, turbulence) suggesting that factors other than temperature and nutrients influence Alexandrium distribution, the exception being Boughrara Lagoon where the species developed in accordance with published data. Our findings and their interpretations indicate that the mechanism of the sudden A. minutum blooms along the nearshore of the Gulf of Gabes was complex and differed from that of true coastal ecosystems. Ó 2012 Elsevier Ltd. All rights reserved. 1. Introduction The southeast coast of Tunisia has rich aquatic resources accounting for approximately 65% of the country’s fish production (DGPA, 2005e2009). Various pollutants from liquid and solid wastes discharged untreated from domestic and industrial activi- ties (crude phosphate treatment, chemical and mineral processing plants, tanneries, textile mills) have severely degraded this coastal ecosystem in the last decade, particularly in the Gulf of Gabes (Mezghani-Chaari et al., 2011; Rekik et al., 2012). As a result, a decrease in fish production has continually been observed since 1990 (Hamza-Chaffai et al., 1997, 2003), in parallel with the large- scale decline of seagrass beds (Ben Brahim et al., 2010) while many blooms of the toxic dinoflagellate Alexandrium minutum have been recorded (Hamza, 2003; Drira et al., 2008). Through an increasing awareness of the consequences for both the developing aquaculture industry and tourism (which is Tunisia’s largest foreign currency earner), and also because of the human impact on the coastal waters, Tunisia undertook, from 1997 to 2006, the Tunisian national monitoring network programme of phytoplankton and phytotoxins (REPHY) in order to monitor microalgae and biotoxins and thus ensure public safety. Such programmes have been implemented in many countries (Anderson et al., 2005; McGillicuddy et al., 2005). The armoured dinoflagellate genus Alexandrium Halim, known to produce paralytic shellfish poisoning (PSP) saxitoxins and a number of related derivatives, comprises more than 30 species (Balech, 1995; Moestrup et al., 2002) distributed worldwide (Anderson et al., 1994, 2012). In the Medi- terranean Alexandrium forms harmful algal blooms (HABs) in many coastal areas (Vila et al., 2001; Guillou et al., 2002; Bravo et al., 2010), most of which have been found in harbours or confined water areas subject to increased eutrophication (Smayda, 1990). In northern Tunisia, the presence of intensely green-brown-coloured water along the coast of the Gulf of Tunis has been reported since 1982, associated with the bloom of A. minutum Halim (Daly Yahia- Kefi et al., 2001). To date, monitoring efforts in the Gulf of Gabes * Corresponding author. E-mail address: lotfi[email protected] (L. Aleya). Contents lists available at SciVerse ScienceDirect Estuarine, Coastal and Shelf Science journal homepage: www.elsevier.com/locate/ecss 0272-7714/$ e see front matter Ó 2012 Elsevier Ltd. All rights reserved. doi:10.1016/j.ecss.2012.04.029 Estuarine, Coastal and Shelf Science xxx (2012) 1e10 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 YECSS3863_proof ■ 19 May 2012 ■ 1/10 Please cite this article in press as: Abdenadher, M., et al., Factors determining the dynamics of toxic blooms of Alexandrium minutum during a 10- year studyalong the shallowsouthwestern Mediterranean coasts, Estuarine, Coastal and Shelf Science (2012), doi:10.1016/j.ecss.2012.04.029

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Q3
at SciVerse ScienceDirect
Estuarine, Coastal and Shelf Science xxx (2012) 1e10
12345678910111213141516171819202122232425262728293031323334353637383940414243444546474849505152535455
YECSS3863_proof ■ 19 May 2012 ■ 1/10
Contents lists available
Estuarine, Coastal and Shelf Science
journal homepage: www.elsevier .com/locate/ecss
56575859606162636465666768
Factors determining the dynamics of toxic blooms of Alexandrium minutum duringa 10-year study along the shallow southwestern Mediterranean coasts
Moufida Abdenadher a, Asma Hamza a, Wafa Fekih a, Imen Hannachi b, Amel Zouari Bellaaj a,Mohamed Nejmeddine Bradai a, Lotfi Aleya c,*
a Institut National des Sciences et Technologie de la Mer, Centre de Sfax, BP 1035, Sfax 3018, TunisiebUniversité de Sfax, Faculté des Sciences de Sfax, Département des Sciences de la Vie, Unité de recherche 00/UR/0907 Ecobiologie, Planctonologie and Microbiologie des EcosystèmesMarins, Route Soukra Km 3.5, BP 1171, CP 3000 Sfax, TunisiecUniversité de Franche-Comté, Laboratoire de Chrono-Environnement, UMR CNRS 6249, 1 Place Leclerc, F-25030 Besançon cedex, France
697071727374757677787980
a r t i c l e i n f o
Article history:Received 24 July 2011Accepted 28 April 2012Available online xxx
Keywords:Alexandrium minutumdistributionbeach samplessouthwestern MediterraneanHAB
* Corresponding author.E-mail address: [email protected] (L. Aley
0272-7714/$ e see front matter � 2012 Elsevier Ltd.doi:10.1016/j.ecss.2012.04.029
81828384
Please cite this article in press as: Abdenadheyear study along the shallow southwestern
a b s t r a c t
Many blooms of the toxic dinoflagellate Alexandrium minutum have been recorded since 1990 in the Gulfof Gabes (southwestern Mediterranean Sea). To understand the determining factors of bloom formation,we studied the distribution of A. minutum in relation to environmental factors in samples taken atshallow sandy (<1 m) beach stations and a lagoon between 1997 and 2006. This was accompanied bylaboratory experiments to identify A. minutum. The species forms harmful algal blooms (HABs) instations subjected to anthropogenic eutrophication and in confined lagoons, living under conditions ofvarying salinity which gives it a unique opportunity to broaden its physiological tolerance and increaseits colonisation potential. Increases in phosphorus appear to be more important than nitrogen ortemperature in the control of A. minutum. The stations sampled all along the coast present specifichydrographic properties (shallowness, turbulence) suggesting that factors other than temperature andnutrients influence Alexandrium distribution, the exception being Boughrara Lagoon where the speciesdeveloped in accordance with published data. Our findings and their interpretations indicate that themechanism of the sudden A. minutum blooms along the nearshore of the Gulf of Gabes was complex anddiffered from that of true coastal ecosystems.
� 2012 Elsevier Ltd. All rights reserved.
8586 87 888990919293949596979899100101102103104
1. Introduction
The southeast coast of Tunisia has rich aquatic resourcesaccounting for approximately 65% of the country’s fish production(DGPA, 2005e2009). Various pollutants from liquid and solidwastes discharged untreated from domestic and industrial activi-ties (crude phosphate treatment, chemical and mineral processingplants, tanneries, textile mills) have severely degraded this coastalecosystem in the last decade, particularly in the Gulf of Gabes(Mezghani-Chaari et al., 2011; Rekik et al., 2012). As a result,a decrease in fish production has continually been observed since1990 (Hamza-Chaffai et al., 1997, 2003), in parallel with the large-scale decline of seagrass beds (Ben Brahim et al., 2010) whilemany blooms of the toxic dinoflagellate Alexandrium minutum havebeen recorded (Hamza, 2003; Drira et al., 2008). Through anincreasing awareness of the consequences for both the developing
a).
All rights reserved.
105106107108
r, M., et al., Factors determiniMediterranean coasts, Estuar
aquaculture industry and tourism (which is Tunisia’s largest foreigncurrency earner), and also because of the human impact on thecoastal waters, Tunisia undertook, from 1997 to 2006, the Tunisiannational monitoring network programme of phytoplankton andphytotoxins (REPHY) in order to monitor microalgae and biotoxinsand thus ensure public safety. Such programmes have beenimplemented in many countries (Anderson et al., 2005;McGillicuddy et al., 2005). The armoured dinoflagellate genusAlexandrium Halim, known to produce paralytic shellfish poisoning(PSP) saxitoxins and a number of related derivatives, comprisesmore than 30 species (Balech, 1995; Moestrup et al., 2002)distributed worldwide (Anderson et al., 1994, 2012). In the Medi-terranean Alexandrium forms harmful algal blooms (HABs) in manycoastal areas (Vila et al., 2001; Guillou et al., 2002; Bravo et al.,2010), most of which have been found in harbours or confinedwater areas subject to increased eutrophication (Smayda, 1990). Innorthern Tunisia, the presence of intensely green-brown-colouredwater along the coast of the Gulf of Tunis has been reported since1982, associated with the bloom of A. minutum Halim (Daly Yahia-Kefi et al., 2001). To date, monitoring efforts in the Gulf of Gabes
109110
ng the dynamics of toxic blooms of Alexandriumminutum during a 10-ine, Coastal and Shelf Science (2012), doi:10.1016/j.ecss.2012.04.029

M. Abdenadher et al. / Estuarine, Coastal and Shelf Science xxx (2012) 1e102
111112113114115116117118119120121122123124125126127128129130131132133134135136137138139140141142143144145146147148149150151152153154155156157158159160161162163164165166167168169170171172173174175
176177178179180181182183184185186187188189190191192193194195196197198199200201202203204205206207
YECSS3863_proof ■ 19 May 2012 ■ 2/10
have focused on the distribution of dinoflagellates in the open sea(Drira et al., 2008, 2010; Bel Hassen et al., 2009), but there havebeen no reports over the past 10 years on the dynamics of dino-flagellates occurring along the very shallow (<1 m in depth)nearshore transect extending from the Libyan border in the southto the zones north of the city of Sfax in the north and passingthrough the Gulf of Gabes (Fig. 1). This is of note since this transectpresents an opportunity to compare the dynamics of A. minutumwithin an ecosystem composed of ecologically distinct regions. Forexample, the Boughrara Lagoon, subjected to the arid conditions ofthe south near the Sahara desert, may be ideal not only as seedingstock for Alexandrium cysts which retain the capability to germinatefor several years (Figueroa et al., 2007; Bravo et al., 2010), but alsofor growth as the lagoon receives iron rich dust particles known totrigger red tide blooms (Wells et al., 1991; Estrada et al., 2010). Inaddition, this lagoon is a quasi-closed lagoon with calm waters,connected to the Mediterranean by only two channels thus leadingto lagoon water exchange properties (Chapman, 2012; De Pascaliset al., 2012) important for the spread of Alexandrium along thewhole coastline of the Gulf of Gabes. The water salinity in the southof the Gulf, near the inland flat areas called sabkhas, is greater(46 salinity) than in the north (36e37 salinity) (Kchaou et al., 2009;Khemakhem et al., 2010), thus changes in the distribution ofA. minutum are expected (Hwang and Lu, 2000; Grzebyk et al.,2003).
This paper presents the results of 10 years of routine monitoringat 20 very shallow (<1m) beach locations along the beach coastlineof the Gulf of Gabes for the presence of harmful algae in order toassess the potential for bloom formation. We first attempted todetermine the spatio-temporal distribution of Alexandrium minu-tum in relation to environmental factors. Secondly, as no study hadyet been conducted in this area to accurately identify A. minutum,
Fig. 1. Geographical map focussing on the monitoring network phytoplanktonic and clam samS3 (4: Mahres), S4 (5: Ras Younga), S5 (6: Jaboussa), S6 (7: Skhira), G1 (8: El Hicha, 9: Tarf El M15: Hassi Jerbi), M3 (16: Borj Djelijel, 17: Houmet Essouk), M4 (18: COTUSAL channel), M5
Please cite this article in press as: Abdenadher, M., et al., Factors determiniyear study along the shallow southwestern Mediterranean coasts, Estuar
we isolated several strains of this taxon from HAB blooms andattempted morphological identification through minute details ofapical tabulation (Taylor et al., 1995) in bloom conditions. It is ofnote that this is the first study of long-term variability in thedistribution of A. minutum along the nearshore of the southeasternMediterranean coasts.
2. Materials and methods
2.1. Study site
The study was conducted in the Gulf of Gabes in an areaextending 262 km along the coastal locations of three cities,Medenine, Djerba Island and Sfax, all located in the south of Tunisiain the southern Ionian Sea (between 35�N and 33�N) (Fig. 1). Theclimate is dry (average annual precipitation: 210 mm year�1) andsunny with strong easterly winds resulting in severe aeolianerosion and particles transported into the sea. The Gulf of Gabes isopen to the offshore area, influenced by the regional water circu-lation (Ben Ismail et al., 2012) and has a wide continental shelfextending from “Ras Kapoudia” at the 35�N parallel level to theTunisian-Libyan border and sheltering the Kerkena Islands in thenortheast and Djerba Island in the southeast (Fig. 1). The tide issemidiurnal being among the highest in the Mediterranean witha maximum range of about 2 m (Drira et al., 2008).
2.2. Sampling
Sampling stations were selected through the Tunisian nationalmonitoring network of phytoplankton and phycotoxins (REPHY),with consideration given to already-existing mollusc collectionsites and followed up by field reconnaissance. The mostly sandy
pling stations in the Gulf of Gabes, Tunisia. S1 (1: Ellouza, 2: El Aouabed), S2 (3: Tabia),a), G2 (10: Port Gabes), G3 (11: Zarrat), M1 (12: Ajim), M2 (13: Boughrara, 14: Karkoub;(19: Bibane Islets), M6 (20: Macif Jetty).
208209210211212213214215216217218219220221222223224225226227228229230231232233234235236237238239240
ng the dynamics of toxic blooms of Alexandriumminutum during a 10-ine, Coastal and Shelf Science (2012), doi:10.1016/j.ecss.2012.04.029

Table 1Size and growth rates of Alexandrium minutum strainsisolated from Jaboussa station.
Strains Length/width (mm)
Am.1 25/20Am.2 25/20Am.3 25/20Am.4 24/20Am.5 25/19Am.6 25/17Am.7 25/20Am.8 25/20Am.9 25/20Am.10 23/17Am.11 25/20Am.12 25/21Am.13 25/21Am.14 25/21Am.15 26/21Am.16 23/17Am.17 25/20Am.18 25/20Am.19 25/20Am.20 24/20Am.21 24/20Am.22 24/20Am.23 23/17Am.24 23/17Am.25 23/17Am.26 25/17Am.27 25/17
M. Abdenadher et al. / Estuarine, Coastal and Shelf Science xxx (2012) 1e10 3
241242243244245246247248249250251252253254255256257258259260261262263264265266267268269270271272273274275276277278279280281282283284285286287288289290291292293294295296297298299300301302303304305
306307308309310311312313314315316317318319320321322323324325326327328329330331332333334335336337338339340341342343344345346347348349350351352353354355356357358359360361362363364365366367368369370
YECSS3863_proof ■ 19 May 2012 ■ 3/10
coastal locations (20 stations) are labelled here, from north tosouth, as S for Sfax, G for Gabes and M for Medenine. Sfax (S) has 6locations: S1 with two stations (1: Ellouza, 2: El Aouabed), S2 withone station (3: Tabia), S3 with one station (4: Mahres), S4 with onestation (5: Ras Younga), S5 with one station (6: Jaboussa), and S6with one station (7: Skhira). For Gabes (G), we took samples at 3locations, G1with 2 stations (8: El Hicha, 9: Tarf El Ma), G2with onestation (10: Gabes Harbour), G3 with one station (11: Zarrat); thesestations were likely to be important sites as they are located insemi-enclosed waters, adjacent to Gabes Harbour which is subjectto intense marine traffic inducing pollution. For Medenine (M), wetook samples at 6 locations, M1 with one station (12: Ajim station,in the NW of Djerba Island), M2 with 3 stations (13: Boughrara, 14:Karboub, 15: Hassi Jerbi), M3 with 2 stations (16: Borj Djelijel, 17:Houmet Essouk in the southeast of Djerba Island), M4 (18:COTUSAL channel), M5 (19: Bibane Islets) and M6 (20: Macif Jetty).Surrounded by a saline area called the Meider Sabkha (Latitude33�270000 N, Longitude 10�450000 E) that forms a narrow beltalongside the stations, the M sites were the saltiest (Kchaou et al.,2009). Among the three stations of M2, Boughrara station waschosen as representing true lagoon conditions. It is a quasi-closedparalic environment, communicating with the Mediterranean viatwo channels, the El Kantara (width: 5 km, average depth: 1 m) tothe northeast and the Ajim channel to the northwest (width:2.2 km, average depth: 15 m). At these stations net evaporation ishigh and soil washing is negligible due to limited freshwatersupplies coupledwith scanty rainfall and poor soil drainage. StationM3 is located in the tourist centre of Djerba Island which is trulymarine, subject to no influence of river discharge.
Between 1997 and 2006, phytoplankton samples from the 20very shallow (<1 m) beach stations were taken weekly fromOctober to May, and monthly the rest of the year (Fig. 1). Watersamples were collected with a Van Dorn bottle at a depth of10e20 cm. Two replicates were taken on individual sampling dates.Associated temperature and salinity parameters were measuredwith a mercury thermometer and microprocessor conductivitymeter WTW Model LF197. Samples for nutrients (nitrite: NO2
�,nitrate: NO3
�, ammonium: NH4þ and orthophosphate: PO4
3�) werestored at �20 �C before analysis with a BRAN and LUEBBE type 3analyser. Phytoplankton cells were preserved with Lugol’s iodinesolution. A 50 ml aliquot was allowed to settle in a countingchamber (Uthermöhl, 1958) for 1 day. Cell counts were madeunder an inverted microscope (Olympus CK40).
2.3. Laboratory experiments
During toxic dinoflagellate outbreaks, samples were taken eachday. Clonal cultures of Alexandrium minutum (n ¼ 27, labelled fromAm.1 to Am.27) (Table 1) were obtained from samples collected on19 January 2006 at Jaboussa station. Samples were withdrawnthrough pipetting and placed on microscope slides for taxonomicidentification which for the Alexandrium species was based on thedescription of Balech (1995). Alexandrium minutum strains weredifferentiated by the presence or absence of a ventral pore on thefirst apical plate (10) and the shapes of the sixth precingular (600)and posterior sulcal plates (sp) (Kofoid notation). Armoured Alex-andrium were stained with calcofluor white (Fritz and Triemer,1985) and examined at 1000� magnification with an invertedmicroscope equipped with epifluoresence optics (Olympus BX41).
2.4. Statistics
Principal components analysis (PCA) was performed on stand-ardised data of biotic and abiotic variables for the years 2005 and2006, using Ade4 (Dray and Dufour, 2007). Direct correlations
Please cite this article in press as: Abdenadher, M., et al., Factors determiniyear study along the shallow southwestern Mediterranean coasts, Estuar
between Alexandriumminutum concentrations and abiotic variableswere estimated using the Spearman correlation coefficient. Allstatistical analyses were performed using R2.9.2. (R DevelopmentCore Team, 2008).
3. Results
3.1. Physico-chemical environment
The range of temperature variation recorded in this studyillustrates a pattern classically observed in the arid to semi-aridzone of the northern hemisphere with warming starting in springand reaching a maximum from the end of July to early August,followed by cooling and a seasonal minimum in January (Table 2).Mean water temperature showed similar variations at thedifferent stations during the entire survey period. The lowesttemperature (19.4 �C) (mean � SD ¼ 21.34 � 1.09) was recorded in1997 in Gabes Harbour (G2, 10) and the maximum (23.6 �C)(mean � SD ¼ 22.16 � 0.77) in 2006 at Karboub (station 18 at M2).Thermal stratification was not established due to hydrodynamicsand the shallowness of the sampled stations (<1 m). The highestsalinity (44.4) was found in 1999 in Karboub (station 18 at M2) andthe lowest (36.7) in 2000 at Jaboussa (station 6 at S5). Overall, highsalinity was clearly detected in Medenine (from Hassi Jerbi: station15 at M2, to Macif: station 20 at M6), especially at the M2, M3, M4and M5 stations which receive salt input from sabkhas.
3.2. Identification of A. minutum
This was based on both phase contrast and UV epifluoresencemicroscopy. Cells were single, rounds and small-sized, 24.51 � 0.36 mm long (min ¼ 23 mm; max ¼ 26 mm) and19.22 � 0.79 mmwide (min ¼ 17 mm; max ¼ 21 mm). The four maindistinctive characteristics described by Halim (1960) and Balech(1995) are shown in Fig. 2. A characteristic ventral pore (V.p.) islocated on the slender and rhomboidal 10 plate (Fig. 2a, b, e). The
ng the dynamics of toxic blooms of Alexandriumminutum during a 10-ine, Coastal and Shelf Science (2012), doi:10.1016/j.ecss.2012.04.029
1
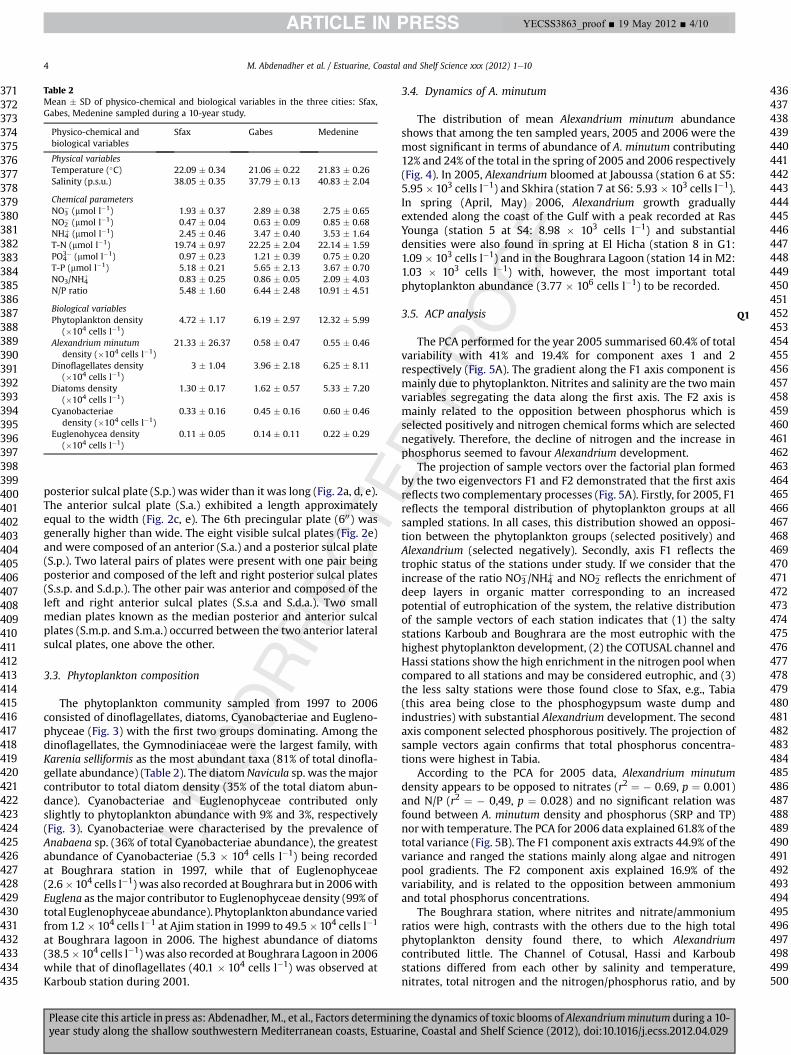
Table 2Mean � SD of physico-chemical and biological variables in the three cities: Sfax,Gabes, Medenine sampled during a 10-year study.
Physico-chemical andbiological variables
Sfax Gabes Medenine
Physical variablesTemperature (�C) 22.09 � 0.34 21.06 � 0.22 21.83 � 0.26Salinity (p.s.u.) 38.05 � 0.35 37.79 � 0.13 40.83 � 2.04
Chemical parametersNO3
� (mmol l�1) 1.93 � 0.37 2.89 � 0.38 2.75 � 0.65NO2
� (mmol l�1) 0.47 � 0.04 0.63 � 0.09 0.85 � 0.68NH4
þ (mmol l�1) 2.45 � 0.46 3.47 � 0.40 3.53 � 1.64T-N (mmol l�1) 19.74 � 0.97 22.25 � 2.04 22.14 � 1.59PO4
3� (mmol l�1) 0.97 � 0.23 1.21 � 0.39 0.75 � 0.20T-P (mmol l�1) 5.18 � 0.21 5.65 � 2.13 3.67 � 0.70NO3/NH4
þ 0.83 � 0.25 0.86 � 0.05 2.09 � 4.03N/P ratio 5.48 � 1.60 6.44 � 2.48 10.91 � 4.51
Biological variablesPhytoplankton density
(�104 cells l�1)4.72 � 1.17 6.19 � 2.97 12.32 � 5.99
Alexandrium minutumdensity (�104 cells l�1)
21.33 � 26.37 0.58 � 0.47 0.55 � 0.46
Dinoflagellates density(�104 cells l�1)
3 � 1.04 3.96 � 2.18 6.25 � 8.11
Diatoms density(�104 cells l�1)
1.30 � 0.17 1.62 � 0.57 5.33 � 7.20
Cyanobacteriaedensity (�104 cells l�1)
0.33 � 0.16 0.45 � 0.16 0.60 � 0.46
Euglenohycea density(�104 cells l�1)
0.11 � 0.05 0.14 � 0.11 0.22 � 0.29
M. Abdenadher et al. / Estuarine, Coastal and Shelf Science xxx (2012) 1e104
371372373374375376377378379380381382383384385386387388389390391392393394395396397398399400401402403404405406407408409410411412413414415416417418419420421422423424425426427428429430431432433434435
436437438439440441442443444445446447448449450451452453454455456457458459460461462463464465466467468469470471472473474475476477478479480481482483484485486487488489490491492493494495496497498499500
YECSS3863_proof ■ 19 May 2012 ■ 4/10
posterior sulcal plate (S.p.) was wider than it was long (Fig. 2a, d, e).The anterior sulcal plate (S.a.) exhibited a length approximatelyequal to the width (Fig. 2c, e). The 6th precingular plate (600) wasgenerally higher than wide. The eight visible sulcal plates (Fig. 2e)and were composed of an anterior (S.a.) and a posterior sulcal plate(S.p.). Two lateral pairs of plates were present with one pair beingposterior and composed of the left and right posterior sulcal plates(S.s.p. and S.d.p.). The other pair was anterior and composed of theleft and right anterior sulcal plates (S.s.a and S.d.a.). Two smallmedian plates known as the median posterior and anterior sulcalplates (S.m.p. and S.m.a.) occurred between the two anterior lateralsulcal plates, one above the other.
3.3. Phytoplankton composition
The phytoplankton community sampled from 1997 to 2006consisted of dinoflagellates, diatoms, Cyanobacteriae and Eugleno-phyceae (Fig. 3) with the first two groups dominating. Among thedinoflagellates, the Gymnodiniaceae were the largest family, withKarenia selliformis as the most abundant taxa (81% of total dinofla-gellate abundance) (Table 2). The diatom Navicula sp. was themajorcontributor to total diatom density (35% of the total diatom abun-dance). Cyanobacteriae and Euglenophyceae contributed onlyslightly to phytoplankton abundance with 9% and 3%, respectively(Fig. 3). Cyanobacteriae were characterised by the prevalence ofAnabaena sp. (36% of total Cyanobacteriae abundance), the greatestabundance of Cyanobacteriae (5.3 � 104 cells l�1) being recordedat Boughrara station in 1997, while that of Euglenophyceae(2.6�104 cells l�1) was also recorded at Boughrara but in 2006withEuglena as themajor contributor to Euglenophyceae density (99% oftotal Euglenophyceae abundance). Phytoplanktonabundance variedfrom 1.2� 104 cells l�1 at Ajim station in 1999 to 49.5�104 cells l�1
at Boughrara lagoon in 2006. The highest abundance of diatoms(38.5�104 cells l�1) was also recorded at Boughrara Lagoon in 2006while that of dinoflagellates (40.1 � 104 cells l�1) was observed atKarboub station during 2001.
Please cite this article in press as: Abdenadher, M., et al., Factors determiniyear study along the shallow southwestern Mediterranean coasts, Estuar
3.4. Dynamics of A. minutum
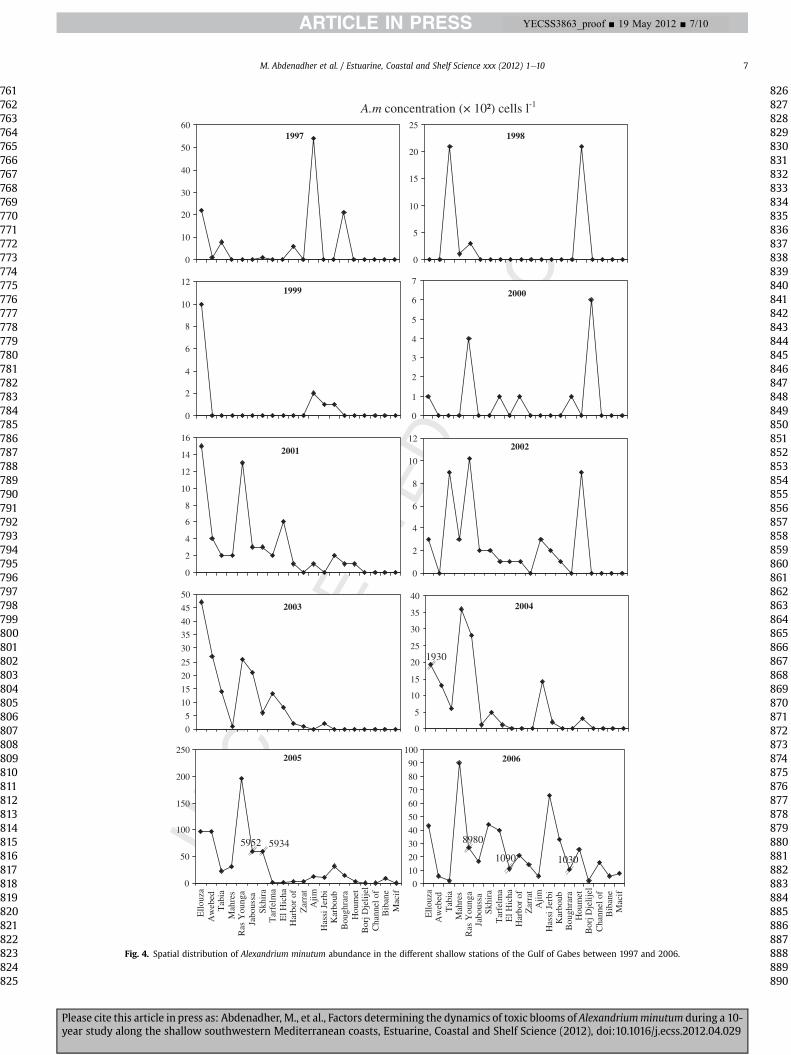
The distribution of mean Alexandrium minutum abundanceshows that among the ten sampled years, 2005 and 2006 were themost significant in terms of abundance of A. minutum contributing12% and 24% of the total in the spring of 2005 and 2006 respectively(Fig. 4). In 2005, Alexandrium bloomed at Jaboussa (station 6 at S5:5.95�103 cells l�1) and Skhira (station 7 at S6: 5.93� 103 cells l�1).In spring (April, May) 2006, Alexandrium growth graduallyextended along the coast of the Gulf with a peak recorded at RasYounga (station 5 at S4: 8.98 � 103 cells l�1) and substantialdensities were also found in spring at El Hicha (station 8 in G1:1.09 � 103 cells l�1) and in the Boughrara Lagoon (station 14 in M2:1.03 � 103 cells l�1) with, however, the most important totalphytoplankton abundance (3.77 � 106 cells l�1) to be recorded.
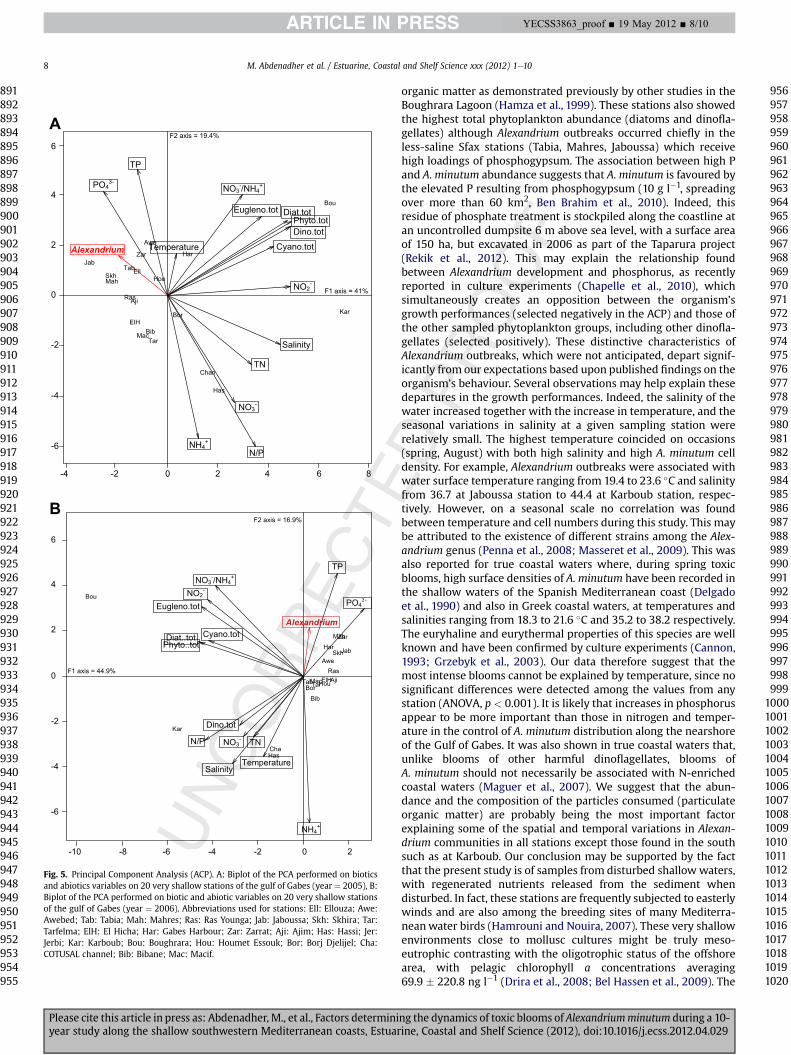
3.5. ACP analysis Q
The PCA performed for the year 2005 summarised 60.4% of totalvariability with 41% and 19.4% for component axes 1 and 2respectively (Fig. 5A). The gradient along the F1 axis component ismainly due to phytoplankton. Nitrites and salinity are the twomainvariables segregating the data along the first axis. The F2 axis ismainly related to the opposition between phosphorus which isselected positively and nitrogen chemical forms which are selectednegatively. Therefore, the decline of nitrogen and the increase inphosphorus seemed to favour Alexandrium development.
The projection of sample vectors over the factorial plan formedby the two eigenvectors F1 and F2 demonstrated that the first axisreflects two complementary processes (Fig. 5A). Firstly, for 2005, F1reflects the temporal distribution of phytoplankton groups at allsampled stations. In all cases, this distribution showed an opposi-tion between the phytoplankton groups (selected positively) andAlexandrium (selected negatively). Secondly, axis F1 reflects thetrophic status of the stations under study. If we consider that theincrease of the ratio NO3
�/NH4þ and NO2
� reflects the enrichment ofdeep layers in organic matter corresponding to an increasedpotential of eutrophication of the system, the relative distributionof the sample vectors of each station indicates that (1) the saltystations Karboub and Boughrara are the most eutrophic with thehighest phytoplankton development, (2) the COTUSAL channel andHassi stations show the high enrichment in the nitrogen pool whencompared to all stations and may be considered eutrophic, and (3)the less salty stations were those found close to Sfax, e.g., Tabia(this area being close to the phosphogypsum waste dump andindustries) with substantial Alexandrium development. The secondaxis component selected phosphorous positively. The projection ofsample vectors again confirms that total phosphorus concentra-tions were highest in Tabia.
According to the PCA for 2005 data, Alexandrium minutumdensity appears to be opposed to nitrates (r2 ¼ � 0.69, p ¼ 0.001)and N/P (r2 ¼ � 0,49, p ¼ 0.028) and no significant relation wasfound between A. minutum density and phosphorus (SRP and TP)nor with temperature. The PCA for 2006 data explained 61.8% of thetotal variance (Fig. 5B). The F1 component axis extracts 44.9% of thevariance and ranged the stations mainly along algae and nitrogenpool gradients. The F2 component axis explained 16.9% of thevariability, and is related to the opposition between ammoniumand total phosphorus concentrations.
The Boughrara station, where nitrites and nitrate/ammoniumratios were high, contrasts with the others due to the high totalphytoplankton density found there, to which Alexandriumcontributed little. The Channel of Cotusal, Hassi and Karboubstations differed from each other by salinity and temperature,nitrates, total nitrogen and the nitrogen/phosphorus ratio, and by
ng the dynamics of toxic blooms of Alexandriumminutum during a 10-ine, Coastal and Shelf Science (2012), doi:10.1016/j.ecss.2012.04.029
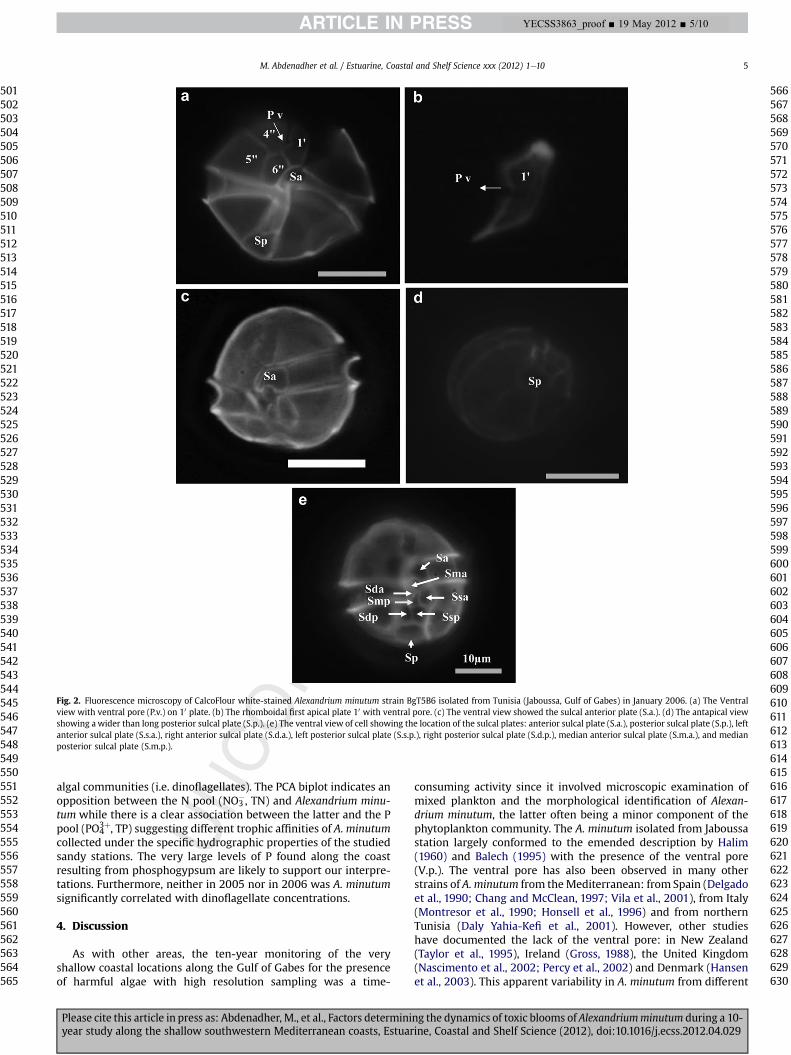
Fig. 2. Fluorescence microscopy of CalcoFlour white-stained Alexandrium minutum strain BgT5B6 isolated from Tunisia (Jaboussa, Gulf of Gabes) in January 2006. (a) The Ventralview with ventral pore (P.v.) on 10 plate. (b) The rhomboidal first apical plate 10 with ventral pore. (c) The ventral view showed the sulcal anterior plate (S.a.). (d) The antapical viewshowing a wider than long posterior sulcal plate (S.p.). (e) The ventral view of cell showing the location of the sulcal plates: anterior sulcal plate (S.a.), posterior sulcal plate (S.p.), leftanterior sulcal plate (S.s.a.), right anterior sulcal plate (S.d.a.), left posterior sulcal plate (S.s.p.), right posterior sulcal plate (S.d.p.), median anterior sulcal plate (S.m.a.), and medianposterior sulcal plate (S.m.p.).
M. Abdenadher et al. / Estuarine, Coastal and Shelf Science xxx (2012) 1e10 5
501502503504505506507508509510511512513514515516517518519520521522523524525526527528529530531532533534535536537538539540541542543544545546547548549550551552553554555556557558559560561562563564565
566567568569570571572573574575576577578579580581582583584585586587588589590591592593594595596597598599600601602603604605606607608609610611612613614615616617618619620621622623624625626627628629630
YECSS3863_proof ■ 19 May 2012 ■ 5/10
algal communities (i.e. dinoflagellates). The PCA biplot indicates anopposition between the N pool (NO3
�, TN) and Alexandrium minu-tum while there is a clear association between the latter and the Ppool (PO4
3þ, TP) suggesting different trophic affinities of A. minutumcollected under the specific hydrographic properties of the studiedsandy stations. The very large levels of P found along the coastresulting from phosphogypsum are likely to support our interpre-tations. Furthermore, neither in 2005 nor in 2006 was A. minutumsignificantly correlated with dinoflagellate concentrations.
4. Discussion
As with other areas, the ten-year monitoring of the veryshallow coastal locations along the Gulf of Gabes for the presenceof harmful algae with high resolution sampling was a time-
Please cite this article in press as: Abdenadher, M., et al., Factors determiniyear study along the shallow southwestern Mediterranean coasts, Estuar
consuming activity since it involved microscopic examination ofmixed plankton and the morphological identification of Alexan-drium minutum, the latter often being a minor component of thephytoplankton community. The A. minutum isolated from Jaboussastation largely conformed to the emended description by Halim(1960) and Balech (1995) with the presence of the ventral pore(V.p.). The ventral pore has also been observed in many otherstrains of A. minutum from theMediterranean: from Spain (Delgadoet al., 1990; Chang and McClean, 1997; Vila et al., 2001), from Italy(Montresor et al., 1990; Honsell et al., 1996) and from northernTunisia (Daly Yahia-Kefi et al., 2001). However, other studieshave documented the lack of the ventral pore: in New Zealand(Taylor et al., 1995), Ireland (Gross, 1988), the United Kingdom(Nascimento et al., 2002; Percy et al., 2002) and Denmark (Hansenet al., 2003). This apparent variability in A. minutum from different
ng the dynamics of toxic blooms of Alexandriumminutum during a 10-ine, Coastal and Shelf Science (2012), doi:10.1016/j.ecss.2012.04.029
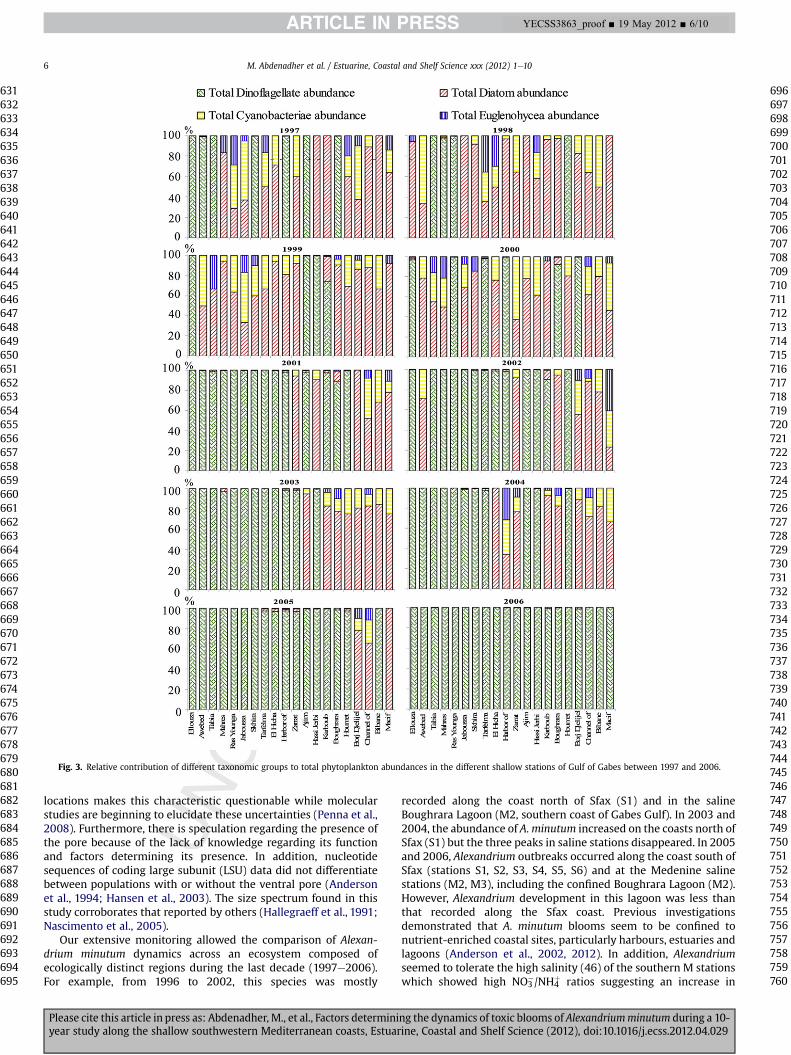
Fig. 3. Relative contribution of different taxonomic groups to total phytoplankton abundances in the different shallow stations of Gulf of Gabes between 1997 and 2006.
M. Abdenadher et al. / Estuarine, Coastal and Shelf Science xxx (2012) 1e106
631632633634635636637638639640641642643644645646647648649650651652653654655656657658659660661662663664665666667668669670671672673674675676677678679680681682683684685686687688689690691692693694695
696697698699700701702703704705706707708709710711712713714715716717718719720721722723724725726727728729730731732733734735736737738739740741742743744745746747748749750751752753754755756757758759760
YECSS3863_proof ■ 19 May 2012 ■ 6/10
locations makes this characteristic questionable while molecularstudies are beginning to elucidate these uncertainties (Penna et al.,2008). Furthermore, there is speculation regarding the presence ofthe pore because of the lack of knowledge regarding its functionand factors determining its presence. In addition, nucleotidesequences of coding large subunit (LSU) data did not differentiatebetween populations with or without the ventral pore (Andersonet al., 1994; Hansen et al., 2003). The size spectrum found in thisstudy corroborates that reported by others (Hallegraeff et al., 1991;Nascimento et al., 2005).
Our extensive monitoring allowed the comparison of Alexan-drium minutum dynamics across an ecosystem composed ofecologically distinct regions during the last decade (1997e2006).For example, from 1996 to 2002, this species was mostly
Please cite this article in press as: Abdenadher, M., et al., Factors determiniyear study along the shallow southwestern Mediterranean coasts, Estuar
recorded along the coast north of Sfax (S1) and in the salineBoughrara Lagoon (M2, southern coast of Gabes Gulf). In 2003 and2004, the abundance of A. minutum increased on the coasts north ofSfax (S1) but the three peaks in saline stations disappeared. In 2005and 2006, Alexandrium outbreaks occurred along the coast south ofSfax (stations S1, S2, S3, S4, S5, S6) and at the Medenine salinestations (M2, M3), including the confined Boughrara Lagoon (M2).However, Alexandrium development in this lagoon was less thanthat recorded along the Sfax coast. Previous investigationsdemonstrated that A. minutum blooms seem to be confined tonutrient-enriched coastal sites, particularly harbours, estuaries andlagoons (Anderson et al., 2002, 2012). In addition, Alexandriumseemed to tolerate the high salinity (46) of the southern M stationswhich showed high NO3
�/NH4þ ratios suggesting an increase in
ng the dynamics of toxic blooms of Alexandriumminutum during a 10-ine, Coastal and Shelf Science (2012), doi:10.1016/j.ecss.2012.04.029
A.m concentration (× 10²) cells l-1
1997
0
10
20
30
40
50
601998
0
5
10
15
20
25
1999
0
2
4
6
8
10
122000
0
1
2
3
4
5
6
7
2001
0
2
4
6
8
10
12
14
162002
0
2
4
6
8
10
12
2003
0
5
10
15
20
25
30
35
40
45
50
2005
0
50
100
150
200
250
Ello
uza
Aw
ebed
Tab
iaM
ahre
sR
as Y
oung
aJa
bous
saSk
hira
Tar
felm
aE
l Hic
haH
arbo
r of
Zar
rat
Ajim
Has
si J
erbi
Kar
boub
Bou
ghra
raH
oum
etB
orj D
jelij
elC
hann
el o
fB
iban
eM
acif
2006
0
10
20
30
40
50
60
70
80
90
100
Ello
uza
Aw
ebed
Tab
iaM
ahre
sR
as Y
oung
aJa
bous
saSk
hira
Tar
felm
aE
l Hic
haH
arbo
r of
Zar
rat
Ajim
Has
si J
erbi
Kar
boub
Bou
ghra
raH
oum
etB
orj D
jelij
elC
hann
el o
fB
iban
eM
acif
2004
0
5
10
15
20
25
30
35
40
8980
1090 1030
5952 5934
1930
Fig. 4. Spatial distribution of Alexandrium minutum abundance in the different shallow stations of the Gulf of Gabes between 1997 and 2006.
M. Abdenadher et al. / Estuarine, Coastal and Shelf Science xxx (2012) 1e10 7
761762763764765766767768769770771772773774775776777778779780781782783784785786787788789790791792793794795796797798799800801802803804805806807808809810811812813814815816817818819820821822823824825
826827828829830831832833834835836837838839840841842843844845846847848849850851852853854855856857858859860861862863864865866867868869870871872873874875876877878879880881882883884885886887888889890
YECSS3863_proof ■ 19 May 2012 ■ 7/10
Please cite this article in press as: Abdenadher, M., et al., Factors determining the dynamics of toxic blooms of Alexandriumminutum during a 10-year study along the shallow southwestern Mediterranean coasts, Estuarine, Coastal and Shelf Science (2012), doi:10.1016/j.ecss.2012.04.029
Temperature Alexandrium
Ell
Awe
Mah
Tar
ElH
Has
Bor
Chan
Bib Mac
Tab
Ras
Har
Aji
Bou
Hou Skh
Jab Zar
Kar
Salinity
NH4+
NO2-
NO3-
PO43-
NO3-/NH4
+
N/P
TN
TP
Phyto.tot Dino.tot
Diat.tot
Cyano.tot
Eugleno.tot
2 -4 -2 0 4 6 8
6
4
2
0
-2
-4
-6
F2 axis = 19.4%
F1 axis = 41%
Diat..tot
Tab
Mah
Tar ElH Aji
Awe Ras
Zar
Jab Skh Har
Has
Hou Bor
Kar
Cha
Bib
Mac
Bou
Temperature Salinity
NH4+
NO2-
NO3-
NO3-/NH4
+
N/P TN
TP
Alexandrium
Phyto..tot
Dino.tot
Cyano.tot
Eugleno.tot
-10 -8 -6 -4 -2 0 2
6
4
2
0
-2
-4
-6
PO43-
F2 axis = 16.9%
F1 axis = 44.9%
A
B
Fig. 5. Principal Component Analysis (ACP). A: Biplot of the PCA performed on bioticsand abiotics variables on 20 very shallow stations of the gulf of Gabes (year ¼ 2005), B:Biplot of the PCA performed on biotic and abiotic variables on 20 very shallow stationsof the gulf of Gabes (year ¼ 2006). Abbreviations used for stations: Ell: Ellouza; Awe:Awebed; Tab: Tabia; Mah: Mahres; Ras: Ras Younga; Jab: Jaboussa; Skh: Skhira; Tar:Tarfelma; ElH: El Hicha; Har: Gabes Harbour; Zar: Zarrat; Aji: Ajim; Has: Hassi; Jer:Jerbi; Kar: Karboub; Bou: Boughrara; Hou: Houmet Essouk; Bor: Borj Djelijel; Cha:COTUSAL channel; Bib: Bibane; Mac: Macif.
M. Abdenadher et al. / Estuarine, Coastal and Shelf Science xxx (2012) 1e108
891892893894895896897898899900901902903904905906907908909910911912913914915916917918919920921922923924925926927928929930931932933934935936937938939940941942943944945946947948949950951952953954955
956957958959960961962963964965966967968969970971972973974975976977978979980981982983984985986987988989990991992993994995996997998999
100010011002100310041005100610071008100910101011101210131014101510161017101810191020
YECSS3863_proof ■ 19 May 2012 ■ 8/10
Please cite this article in press as: Abdenadher, M., et al., Factors determiniyear study along the shallow southwestern Mediterranean coasts, Estuar
organic matter as demonstrated previously by other studies in theBoughrara Lagoon (Hamza et al., 1999). These stations also showedthe highest total phytoplankton abundance (diatoms and dinofla-gellates) although Alexandrium outbreaks occurred chiefly in theless-saline Sfax stations (Tabia, Mahres, Jaboussa) which receivehigh loadings of phosphogypsum. The association between high Pand A. minutum abundance suggests that A. minutum is favoured bythe elevated P resulting from phosphogypsum (10 g l�1, spreadingover more than 60 km2, Ben Brahim et al., 2010). Indeed, thisresidue of phosphate treatment is stockpiled along the coastline atan uncontrolled dumpsite 6 m above sea level, with a surface areaof 150 ha, but excavated in 2006 as part of the Taparura project(Rekik et al., 2012). This may explain the relationship foundbetween Alexandrium development and phosphorus, as recentlyreported in culture experiments (Chapelle et al., 2010), whichsimultaneously creates an opposition between the organism’sgrowth performances (selected negatively in the ACP) and those ofthe other sampled phytoplankton groups, including other dinofla-gellates (selected positively). These distinctive characteristics ofAlexandrium outbreaks, which were not anticipated, depart signif-icantly from our expectations based upon published findings on theorganism’s behaviour. Several observations may help explain thesedepartures in the growth performances. Indeed, the salinity of thewater increased together with the increase in temperature, and theseasonal variations in salinity at a given sampling station wererelatively small. The highest temperature coincided on occasions(spring, August) with both high salinity and high A. minutum celldensity. For example, Alexandrium outbreaks were associated withwater surface temperature ranging from 19.4 to 23.6 �C and salinityfrom 36.7 at Jaboussa station to 44.4 at Karboub station, respec-tively. However, on a seasonal scale no correlation was foundbetween temperature and cell numbers during this study. This maybe attributed to the existence of different strains among the Alex-andrium genus (Penna et al., 2008; Masseret et al., 2009). This wasalso reported for true coastal waters where, during spring toxicblooms, high surface densities of A. minutum have been recorded inthe shallow waters of the Spanish Mediterranean coast (Delgadoet al., 1990) and also in Greek coastal waters, at temperatures andsalinities ranging from 18.3 to 21.6 �C and 35.2 to 38.2 respectively.The euryhaline and eurythermal properties of this species are wellknown and have been confirmed by culture experiments (Cannon,1993; Grzebyk et al., 2003). Our data therefore suggest that themost intense blooms cannot be explained by temperature, since nosignificant differences were detected among the values from anystation (ANOVA, p < 0.001). It is likely that increases in phosphorusappear to be more important than those in nitrogen and temper-ature in the control of A. minutum distribution along the nearshoreof the Gulf of Gabes. It was also shown in true coastal waters that,unlike blooms of other harmful dinoflagellates, blooms ofA. minutum should not necessarily be associated with N-enrichedcoastal waters (Maguer et al., 2007). We suggest that the abun-dance and the composition of the particles consumed (particulateorganic matter) are probably being the most important factorexplaining some of the spatial and temporal variations in Alexan-drium communities in all stations except those found in the southsuch as at Karboub. Our conclusion may be supported by the factthat the present study is of samples from disturbed shallow waters,with regenerated nutrients released from the sediment whendisturbed. In fact, these stations are frequently subjected to easterlywinds and are also among the breeding sites of many Mediterra-neanwater birds (Hamrouni and Nouira, 2007). These very shallowenvironments close to mollusc cultures might be truly meso-eutrophic contrasting with the oligotrophic status of the offshorearea, with pelagic chlorophyll a concentrations averaging69.9 � 220.8 ng l�1 (Drira et al., 2008; Bel Hassen et al., 2009). The
ng the dynamics of toxic blooms of Alexandriumminutum during a 10-ine, Coastal and Shelf Science (2012), doi:10.1016/j.ecss.2012.04.029

Q2
M. Abdenadher et al. / Estuarine, Coastal and Shelf Science xxx (2012) 1e10 9
10211022102310241025102610271028102910301031103210331034103510361037103810391040104110421043104410451046104710481049105010511052105310541055105610571058105910601061106210631064106510661067106810691070107110721073107410751076107710781079108010811082108310841085
10861087108810891090109110921093109410951096109710981099110011011102110311041105110611071108110911101111111211131114111511161117111811191120112111221123112411251126112711281129113011311132113311341135113611371138113911401141114211431144114511461147114811491150
YECSS3863_proof ■ 19 May 2012 ■ 9/10
above observations suggest that factors other than temperatureand nutrients influenced Alexandrium distribution at these stations,thus contrasting with studies cited from elsewhere in the Medi-terranean (Vila et al., 2005; Llaveria et al., 2010). In addition, at oursampled stations we found that all kinds of particles from bacteria,nanoflagellates, microalgae, and microzooplankton to eggs ofcopepods were usable by A. minutum (Tillmann, 2004; Kchaouet al., 2009; Drira et al., 2010). The dominance of A. minutum mayalso be supported by the finding showing its allelopathy throughinhibitory effects on other phytoplankton taxa (Lelong et al., 2011),its toxin production which is stimulated by water movements andits mixotrophy (Zubkov and Tarran, 2008). If we consider only thesemi-enclosed, saline and arid Boughrara Lagoon, we recorda prevalence of A. minutum outbreaks (1.03 � 103 cells l�1) corre-lating with nitrogen forms as also reported from nutrient-enrichedcoastal sites, particularly harbours, estuaries and lagoons (Giacobbeet al., 1996; Collos et al., 2007; Pereira et al., 2012).
We infer that the Boughrara Lagoon may act as an accumulatorof neritic phytoplankton. The sporadic connections of lagoons withcoastal waters (Pereira et al., 2012) can introduce phytoplankton,possibly including HAB species, into the lagoons where they maygrow and become highly abundant. This is supported by the alreadyreported annual blooms of harmful dinoflagellate Karenia sell-iformis (Guelorget, 1991; Hamza et al., 1999) in this lagoon afterintroduction via coastal waters (Drira et al., 2008). This coastallagoon receives negligible tidal input, but is occasionally floodedduring sea storms after which it usually recovers its confinedcharacter without marine exchange (Hamza et al., 1999).
5. Conclusions
Our study indicated that the variability of environmental factorsinfluence both timing and location of Alexandrium outbreaks alongthe nearshore (<1 m) of the Gulf of Gabes. Alexandrium minutumforms HABs in stations impacted by anthropogenic eutrophication(Sfax Harbour) and confined lagoons reflecting a high physiologicalflexibility. Our findings and their interpretations also serve toillustrate that the mechanism of the sudden A. minutum bloomsalong the nearshore of the Gulf of Gabes is complex. In addition,living in variable salinity regimes gives A. minutum a uniqueopportunity to broaden its physiological tolerance and increase itscolonisation potential together with planozygote and cyst forma-tion (Figueroa et al., 2011). Our study revealed that A. minutum, incomparison to other microalgae living under true, stronger coastalenvironmental conditions, is significantly distinct in its behaviourand dynamic sensitivity to environmental variables providing newinsights into its capabilities to take optimal advantage of nicheopportunities.
To improve our early warning capabilities, we are currentlystudying the cyst distribution in surface sediments known to beabundant near mollusc-harvesting stations (Wang et al., 2012) withregard to environmental variables and the appearance ofA. minutum blooms. It is also of relevance to relate the appearanceof A. minutum to PSP toxin contents in molluscs as well as the studyof toxin production in the cultures.
Uncited reference
Bravo et al., 2008.
Acknowledgements
This work is the result of a collaboration project involving theINSTM (Institut National des Sciences et Technologie de la Mer,centre de Sfax) and the University of Franche-Comté, Chrono-
Please cite this article in press as: Abdenadher, M., et al., Factors determiniyear study along the shallow southwestern Mediterranean coasts, Estuar
Environnement, CNRS 6249, France, as part of the REPHY project(Tunisian National Monitoring Network of Phytoplankton andPhytotoxins in the Golf of Gabes). We thank the assistance of theanonymous reviewers who contributed to improving our paper.
References
Anderson, D.M., Kulis, D.M., Doucette, G.J., Gallagher, J.C., Balech, E., 1994. Bioge-ography of toxic dinoflagellates in the genus Alexandrium from the northeasternUnited States and Canada. Marine Biology 120, 467e478.
Anderson, D.M., Glibert, P.M., Burkholder, J.M., 2002. Harmful algal blooms andeutrophication: nutrient sources, composition and consequences. Estuaries 25,704e726.
Anderson, D.M., Kulis, D.M., Keafer, B.A., Gribble, K.E., Marin, R., Scholin, C.A., 2005.Identification and enumeration of Alexandrium spp. from the Gulf of Maineusing molecular probes. In: The Ecology and Oceanography of Toxic Alexan-drium Fundyense Blooms in the Gulf of Maine. Deep Sea Research Part II:Topical Studies in Oceanography, vol. 52, pp. 2467e2490.
Anderson, D.M., Alpermann, T.J., Cembella, A.D., Collos, Y., Masseret, E.,Montresor, M., 2012. The globally distributed genus Alexandrium: multifacetedroles in marine ecosystems and impacts on human health. Harmful Algae 14,10e35.
Balech, E., 1995. The Genus Alexandrium Halim (Dinoflagellata). Sherkin IslandMarine Station, Cork, Ireland, pp. 151.
Bel Hassen, M., Drira, Z., Hamza, A., Ayadi, H., Akrout, F., Messaoudi, S., Issaoui, H.,Aleya, L., Bouaïn, A., 2009. Plankton-pigment signatures and their relationshipto springesummer stratification in the South-eastern Mediterranean. Estuarine,Coastal and Shelf Science 83, 296e306.
Ben Brahim, M., Hamza, A., Hannachi, I., Rebai, A., Jarboui, O., Bouain, A., Aleya, L.,2010. Variability in the structure of epiphytic assemblages of Posidonia oceanicain relation to human interferences in the Gulf of Gabes, Tunisia. Marine Envi-ronmental Research 70, 411e421.
Ben Ismail, S., Sammari, C., Gasparini, G.P., Béranger, K., Brahim, M., Aleya, L., 2012.Water masses exchanged through the Channel of Sicily: evidence for thepresence of new water masses on the Tunisian side of the Channel. Deep SeaResearch I 63, 65e81.
Bravo, I., Vila, M., Maso, M., Figueroa, R.I., Ramilo, I., 2008. Alexandrium catenella andAlexandrium minutum blooms in the Mediterranean Sea: toward the identifi-cation of ecological niches. Harmful Algae 7, 515e522.
Bravo, I., Figueroa, R.I., Garcés, E., Fraga, S., Massanet, A., 2010. The intricacies ofdinoflagellate pellicle cysts: the example of Alexandrium minutum cysts froma bloom-recurrent area (Bay of Baiona, NW Spain). Deep Sea Research II 57,166e174.
Cannon, J.A., 1993. Growth in culture of the toxic dinoflagellate Alexandrium min-utum from the Port River, South Australia. In: Smayda, T.J., Shimizu, Y. (Eds.),Toxic Phytoplankton Blooms in the Sea. Elsevier, Amsterdam, pp. 741e745.
Chang, F.H., McClean, M., 1997. Growth responses of Alexandrium minutum (Dino-phyceae) as a function of three different nitrogen sources and irradiance. NewZealand Journal of Marine and Freshwater Research 31, 1e7.
Chapelle, A., Labry, C., Sourisseau, M., Lebreton, C., Youenoua, A., Crassous, M.P.,2010. Alexandrium minutum growth controlled by phosphorus: an appliedmodel. Journal of Marine Systems 83, 181e191.
Chapman, P.M., 2012. Management of coastal lagoons under climate change. Estu-arine, Coastal and Shelf Science. doi:10.1016/j.ecss.2012.01.010.
Collos, Y., Vaquer, A., Laabir, M., Abadie, E., Laugier, T., Pastoureaud, A., Souchu, P.,2007. Contribution of several nitrogen sources to growth of Alexandrium cat-enella during blooms in Thau lagoon, southern France. Harmful Algae 6,781e789.
Daly Yahia-Kefi, O., Nézan, E., Daly Yahia, M.N., 2001. Sur la présence du genreAlexandrium Halim (Dinoflagellés) dans la baie de Tunis (Tunisie). OceanologicaActa 24, 17e25.
Delgado, M., Estrada, M., Camp, J., Fernandez, J.V., Santmarti, M., Lleti, C., 1990.Development of a toxic Alexandrium minutum Halim (Dinophyceae) bloom inthe harbour of Sant Carles de la Rapita (Ebro Delta, northwestern Mediterra-nean). Scientia Marina 54, 1e7.
De Pascalis, F., Ruzafa, A.P., Gilabert, J., Marcos, C., Georg Umgiesser, G., 2012.Climate change response of the Mar Menor coastal lagoon (Spain) usinga hydrodynamic finite element model. Estuarine, Coastal and Shelf Science.doi:10.1016/j.ecss.2011.12.002.
DGPA, 2005e2009. annuaire statistique. Direction Générale de la pêche et del’aquaculture, Ministère de l’agriculture, Tunisie.
Dray, S., Dufour, A.B., 2007. The ade4 package: implementing the duality diagramfor ecologists. Journal of Statistical Software 22, 1e20.
Drira, Z., Hamza, A., Bel Hassen, M., Ayadi, H., Bouaïn, A., Aleya, L., 2008. Dynamicsof dinoflagellates and environmental factors during the summer in the Gulf ofGabes (Tunisia, Eastern Mediterranean Sea). Scientia Marina 72, 59e71.
Drira, Z., Bel Hassen, M., Hamza, A., Rebai, A., Bouaïn, A., Ayadi, H., Aleya, L., 2010.Coupling of phytoplankton community structure to nutrients, ciliates andcopepods in the Gulf of Gabes (South Ionian Sea, Tunisia). Journal of the MarineBiological Association, UK 190, 1203e1215.
Estrada, M., Sole, J., Angles, S., Garces, E., 2010. The role of resting cysts in Alexan-drium minutum population dynamics. Deep Sea Research II 57, 308e321.
ng the dynamics of toxic blooms of Alexandriumminutum during a 10-ine, Coastal and Shelf Science (2012), doi:10.1016/j.ecss.2012.04.029

M. Abdenadher et al. / Estuarine, Coastal and Shelf Science xxx (2012) 1e1010
1151115211531154115511561157115811591160116111621163116411651166116711681169117011711172117311741175117611771178117911801181118211831184118511861187118811891190119111921193119411951196119711981199120012011202120312041205120612071208
1209121012111212121312141215121612171218121912201221122212231224122512261227122812291230123112321233123412351236123712381239124012411242124312441245124612471248124912501251125212531254125512561257125812591260126112621263
YECSS3863_proof ■ 19 May 2012 ■ 10/10
Figueroa, R.I., Garces, E., Bravo, I., 2007. Comparative study of the life cycles ofAlexandrium tamutum and Alexandrium minutum (Gonyaulacales, Dinophyceae)in culture. Journal of Phycology 43, 1039e1053.
Figueroa, R.I., Vázquez, J.A., Massanet, A., Murado, M.A., Bravo, I., 2011. Interactiveeffects of salinity and temperature on planozygote and cyst formation ofAlexandrium minutum (Dinophyceae) in culture. Journal of Phycology 47, 13e24.
Fritz, L., Triemer, R.E., 1985. A rapid technique utilizing calcofluor white M2R for thevisualization of dinoflagellate thecal plates. Journal of Phycology 21, 662e664.
Giacobbe, M.G., Oliva, F.D., Maimone, G., 1996. Environmental factors and seasonaloccurrence of the dinoflagellate Alexandrium minutum, a PSP potentialproducer, in a Mediterranean lagoon. Estuarine, Coastal and Shelf Science 42,539e549.
Gross, J., 1988. A red tide outbreak in Cork Harbour, July 1987. Red Tide Newsletter 1,4. Sherkin Island Marine Station.
Grzebyk, D., Bechemin, C., Ward, C.J., Verite, C., Codd, G.A., Maestrini, S.Y., 2003.Effects of salinity and two coastal waters on the growth and toxin content of thedinoflagellate Alexandrium minutum. Journal of Plankton Research 25,1185e1199.
Guelorget, O., 1991. Etude de la crise survenue en mer Boughrara au cours de l’été1991. Rapport C.G.P. p. 7.
Guillou, L., Nézan, E., Cueff, V., Erard-Le Denn, E., Cambon-Bonavita, M.A.,Barbier, G., 2002. Genetic diversity and molecular detection of three toxicdinoflagellate genera (Alexandrium, Dinophysis, and Karenia) from Frenchcoasts. Protist 153, 223e238.
Halim, Y., 1960. Alexandrium minutum, n. gen. n. sp. dinoflagellé induisant des eauxrouges. Vie et Milieu 11, 102e105.
Hallegraeff, G.M., Bolch, S.J.C., Blackburn, S.I., Oshima, Y., 1991. Species of the toxi-genic dinoflagellate genus Alexandrium in southeastern Australian waters.Botanica Marina 10, 575e587.
Hamrouni, K., Nouira, S., 2007. Diversité et organisation spatiale de l’avifaune deThyna, Sfax (Tunisie). Ostrich 78, 517e522.
Hamza, A., 2003. Le statut du phytoplancton dans le Golfe de Gabes. Thèse dedoctorat en Sciences Biologiques, 298 pp.
Hamza, A., Bouain, A., El Abed, A., 1999. Le Gymnodinium sp. phytoplancton toxiquedu golfe de Gabès. Actes des 3èmes journées tunisiennes des Sciences de la mer.Bulletin de l’Institut National des Sciences et Technologie de la Mer, 49e51.
Hamza-Chaffai, A., Amiard-Triquet, C., El Abed, A., 1997. Metallothionein-likeprotein, is it an efficient biomarker of metal contamination? A case study basedon fish from the Tunisian coast. Archives of Environmental Contamination andToxicology 33, 53e62.
Hamza-Chaffai, A., Pellerin, J., Amiard, J.C., 2003. Health assessment of marinebivalve (Ruditapes decussatus) from the gulf of Gabès (Tunisia). EnvironmentInternational 28, 609e617.
Hansen, G., Daugbjerg, N., Franco, J.M., 2003. Morphology, toxin composition andLSU rDNA phylogeny of Alexandrium minutum (Dinophyceae) from Denmark,with some morphological observations on other European strains. HarmfulAlgae 2, 317e335.
Honsell, G., Poletti, R., Pompei, M., Sidari, L., Milandri, A., Casadei, C., Viviani, R.,1996. Alexandrium minutum Halim and PSP contamination in the NorthernAdriatic Sea (Mediterranean Sea). In: Yasumoto, T., Oshima, T., Fukuyo, Y. (Eds.),Harmful and Toxic Algae. IOC, UNESCO, pp. 77e80.
Hwang, D.F., Lu, Y.H., 2000. Influence of environmental and nutritional factors ongrowth, toxicity, and toxin profile of dinoflagellate Alexandrium minutum.Toxicon 38, 1491e1503.
Kchaou, N., Elloumi, J., Drira, Z., Hamza, A., Ayadi, H., Bouain, A., Aleya, L., 2009.Distribution of ciliates in relation to environmental factors along the coastlineof the Gulf of Gabes, Tunisia. Estuarine, Coastal and Shelf Science 83, 414e424.
Khemakhem, H., Elloumi, J., Moussa, M., Aleya, L., Ayadi, H., 2010. The concept ofecological succession applied to phytoplankton over four consecutive years infive ponds featuring a salinity gradient. Estuarine, Coastal and Shelf Science 88,33e44.
Lelong, A., Haberkorn, H., Le Goïc, N., Hégaret, H., Soudant, P., 2011. A new insightinto allelopathic effects of Alexandrium minutum on photosynthesis and respi-ration of the diatom Chaetoceros neogracile revealed by photosynthetic-performance analysis and flow cytometry. Microbial Ecology 62, 919e930.
Llaveria, G., Garcés, E., Ross, O.N., Figueroa, R.I., Sampedro, N., Berdalet, E., 2010.Small-scale turbulence can reduce parasite infectivity to dinoflagellates. MarineEcology Progress Series 412, 45e56.
Maguer, J.F., L’Helguen, S., Madec, C., Claire Labry, C., Le Corre, P., 2007. Nitrogenuptake and assimilation kinetics in Alexandrium minutum (Dinophyceae): effectof N-limited growth rate on nitrate and ammonium interactions. Journal ofPhycology 43, 295e303.
Please cite this article in press as: Abdenadher, M., et al., Factors determiniyear study along the shallow southwestern Mediterranean coasts, Estuar
Masseret, E., Grzebyk, D., Nagai, S., Genovesi, B., Bernard Lasserre, B., MohamedLaabir, M., Yves Collos, Y., André Vaquer, A., Patrick Berrebi, P., 2009. Unex-pected genetic diversity among and within populations of the toxic dinofla-gellate Alexandrium catenella as revealed by nuclear microsatellite markers.Applied and Environmental Microbiology 75, 2037e2045.
McGillicuddy Jr., D.J., Anderson, D.M., Lynch, D.R., Townsend, D.W., 2005. Mecha-nisms regulating large-scale seasonal fluctuations in Alexandrium fundyensepopulations in the Gulf of Maine: results from a physicalebiological model.Deep Sea Research II 52, 2698e2714.
Mezghani-Chaari, S., Hamza, A., Hamza-Chaffai, A., 2011. Mercury contamination inhuman hair and some marine species from Sfax coasts of Tunisia: levels andrisk assessment. Environmental Monitoring and Assessment 180, 477e487.
Moestrup, Ø, Codd, G.A., Elbrächter, M., Faust, M.A., Fraga, S., Fukuyo, Y.,Cronberg, G., Halim, Y., Taylor, F.J.R., Zingone, A., 2002. IOC Taxonomic ReferenceList of Toxic Plankton Algae. http://www.bi.ku.dk/ioc/.
Montresor, M., Marino, D., Zingone, A., Dafnis, G., 1990. Three Alexandrium speciesfrom coastal Tyrrhenian waters. In: Graneli, E., Sundstrom, B., Edler, L.,Anderson, D. (Eds.), Toxic Marine Phytoplankton. Elsevier, New York, pp. 82e87.
Nascimento, S.M., Purdie, D.A., Larsen, J., Morris, S., 2002. A new isolate of Alex-andrium minutum from a coastal lagoon in southern England. In: Steidinger, K.A.(Ed.), Book of Abstracts. Xth International Conference on Harmful Algae, p. 212.
Nascimento, S.M., Purdie, D.A., Lilly, E.L., Larsen, J., Morris, S., 2005. Toxin profile,pigment composition, and large subunit rDNA phylogenetic analysis of anAlexandrium minutum (Dinophyceae) strain isolated from the Fleet Lagoon,United Kingdom. Journal of Phycology 41, 343e353.
Penna, A., Fraga, S., Maso, M., Giacobbe, M.G., Bravo, I., Garces, E., Vila, M.,Bertozzini, E., Andreoni, F., Luglie, A., Vernesi, C., 2008. Phylogenetic relation-ships among the Mediterranean Alexandrium (Dinophyceae) species based onsequences of 5.8S gene and internal transcript spacers of the rRNA operon.European Journal of Phycology 43, 163e178.
Percy, L., Lewis, J., Morris, S., Stone, D.M., Higman, W., 2002. The relationshipsbetween Alexandrium species in water and PSP toxins in shellfish from the Falestuary, United Kingdom. In: Steidinger, K.A. (Ed.), Tenth International Confer-ence on Harmful Algae, Book of Abstracts, St. Petersburg, FL, p. 229.
Pereira, P., Botelho, M.J., Cabrita, M.T., Vale, C., Maria Teresa Moita, M.T., CéliaGonçalves, C., 2012. Winteresummer nutrient composition linkage to algae-produced toxins in shellfish at a eutrophic coastal lagoon (Óbidos lagoon,Portugal). Estuarine, Coastal and Shelf Science. doi:10.1016/j.ecss.2011.07.016.
R Development Core Team, 2008. R: A Language and Environment for StatisticalComputing. R Foundation for Statistical Computing, Vienna, Austria, ISBN3-900051-07-0. http://www.R-project.org.
Rekik, A., Drira, Z., Guermazi, W., Elloumi, J., Maalej, S., Aleya, L., Ayadi, H., 2012.Impacts of an uncontrolled phosphogypsum dumpsite on summer distributionof phytoplankton, copepods and ciliates in relation to abiotic variables along thenear-shore of the southwestern Mediterranean coast. Marine Pollution Bulletin64, 336e346.
Smayda, T.J., 1990. Novel and nuisance phytoplankton blooms in the sea: evidencefor a global epidemic. In: Granéli, E., Sundström, P.B., Edler, L., Anderson, D.M.(Eds.), Toxic Marine Phytoplankton. Elsevier, New York, pp. 29e41.
Taylor, F.J.R., Fukuyo, Y., Larsen, J., 1995. Taxonomy of harmful dinoflagellates. In:Hallegraeff, G.M., Anderson, D.M., Cembella, A.D. (Eds.), Manual on HarmfulMarine Microalgae. IOC Manual & Guides No. 33. UNESCO, pp. 283e317.
Tillmann, U., 2004. Interactions between planktonic microalgae and protozoangrazers. Journal of Eukaryotic Microbiology 51, 156e168.
Uthermöhl, H., 1958. Zur Vervollkommung der quantitativen PhytoplanktonMethodik. Mitteilungen Internationale Vereinigung für Theoretische undAngewandte Limnologie 9, 1e38.
Vila, M., Camp, J., Garcès, E., Maso, M., Delgado, M., 2001. High resolution spatio-temporal detection of potentially harmful dinoflagellates in confined watersof the NW Mediterranean. Journal of Plankton Research 23, 497e514.
Vila, M., Giacobbe, M.G., Maso, M., Gangemi, E., Penna, A., Sanpedro, N., Azzaro, F.,Camp, J., Galluzzi, L., 2005. A comparative study on recurrent blooms of Alex-andrium minutum in two Mediterranean coastal areas. Harmful Algae 4,673e695.
Wang, Z.H., Mu, D.H., Li, Y.F., Cao, Y., Zhang, Y.J., 2012. Recent eutrophication andhuman disturbance in Daya Bay, the South China Sea: dinoflagellate cyst andgeochemical evidence. Estuarine, Coastal and Shelf Science. doi:10.1016/j.ecss.2011.01.015.
Wells, M.L., Mayer, L.M., Guillard, R.R.L., 1991. Evaluation of iron as a triggeringfactor for red tide blooms. Marine Ecology Progress Series 69, 93e102.
Zubkov, M.V., Tarran, G.A., 2008. High bacterivory by the smallest phytoplankton inthe North Atlantic Ocean. Nature 455, 224e226.
126412651266
ng the dynamics of toxic blooms of Alexandriumminutum during a 10-ine, Coastal and Shelf Science (2012), doi:10.1016/j.ecss.2012.04.029
Related Documents











