Abbau von Polyethylenterephthalat mit PET-Hydrolasen aus Thermobifida fusca KW3 Von der Fakultät für Maschinenbau der Technischen Universität Chemnitz Genehmigte Dissertation zu Erlangung des akademischen Grades Doktor-Ingenieur Dr.-Ing. vorgelegt von Dipl.-Ing. Susan Billig geboren am 12.01.1978 in Marienberg eingereicht am 06.09.2011 Gutachter: Prof. Dr. Wolfgang Zimmermann Prof. Dr.-Ing. habil. Bernd Platzer Dr. Vincent A. Nierstrasz Chemnitz, den 06.09.2011

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Abbau von Polyethylenterephthalat mit PET-Hydrolasen aus
Thermobifida fusca KW3
Von der Fakultät für Maschinenbau der Technischen Universität Chemnitz
Genehmigte
Dissertation
zu Erlangung des akademischen Grades
Doktor-Ingenieur
Dr.-Ing.
vorgelegt
von Dipl.-Ing. Susan Billig
geboren am 12.01.1978 in Marienberg
eingereicht am 06.09.2011
Gutachter:
Prof. Dr. Wolfgang Zimmermann
Prof. Dr.-Ing. habil. Bernd Platzer
Dr. Vincent A. Nierstrasz
Chemnitz, den 06.09.2011

II
Bibliographische Beschreibung
Autor Billig, Susan
Thema Abbau von Polyethylenterephthalat mit PET-Hydrolasen aus Thermobifida fusca KW3
Dissertation an der Fakultät für Maschinenbau der Technischen Universität Chemnitz, Institut für
Technische Thermodynamik, Chemnitz, 06.09.2011
Seitenzahl: 200, Anzahl der Abbildungen: 91, Anzahl der Tabellen: 37, Anzahl der Literaturzitate: 203
Referat
Der Actinomycet T. fusca KW3, isoliert aus Kompost, bildete während der Kultivierung im
Mineralsalz-Spurenelement-Vitamin-Minimalmedium nach Zusatz von PET-Fasern eine 52 kDa
Carboxylesterase (TfCa), welche effizient zyklische PET Trimere (CTR) hydrolysiert. Die TfCa besitzt
einen pI von 4,8, eine Substratspezifität gegenüber kurzkettigen p-Nitrophenyl-Estern und wird durch
Phenylmethylsulfonylfluorid (PMSF) und Tosyl-L-Phenylalanin-Chloromethylketon (TPCK) in der
Aktivität gehemmt. Die Carboxylesterase hydrolysiert kein Cutin oder Poly-ε-caprolacton (PCL). CTR
hingegen wurden durch die TfCa mit einem Km von 0,5 mM und einer Vmax von 9,3 µmol/min/mg bei
optimalen Bedingungen (60°C, pH 6) hydrolysiert. Das aktive Zentrum der Carboxylesterase besteht
aus den Aminosäuren Ser185, Glu319 und His415, wobei das Serin in das katalytische Motiv G-E-S-A-
G eingebettet ist.
Während der Reaktion setzte die TfCa auch Hydrolyseprodukte aus PET-Fasern und -Filmen frei. Der
Nachweis der Hydrolyse erfolgte durch Umkehrphasen-Hochleistungsflüssigkeitschromatographie
der Abbauprodukte und bei den PET-Filmen zusätzlich mittels Rasterelektronenmikroskopie. Dabei
zeigte die Carboxylesterase verglichen mit anderen PET-Hydrolasen eine geringere Effizienz, was
durch die Lage des aktiven Zentrums in einer Bindungstasche und der daraus folgenden schlechten
Zugänglichkeit für polymere Substrate begründet werden kann. Bei der Hydrolyse der viel kleineren
CTR war die TfCa deutlich effektiver, was auf eine höhere Spezifität gegenüber kurzkettigen PET
Substraten hinweist.
Schlagworte
Polyethylenterephthalat, Thermobifida fusca, Carboxylesterase, PET-Hydrolase, Hydrolyse,
Actinomycet, enzymatische PET-Modifikation, enzymatischer PET-Abbau

III
Ich weiß, dass ich nichts weiß.
Sokrates (469-399 v. Chr.)
Was wir wissen, ist ein Tropfen; was wir nicht wissen, ein Ozean.
Wenn ich fähig war, weiter zu sehen als andere,
dann deshalb, weil ich auf den Schultern von Riesen stand.
Isaac Newton (1643-1727)
Eigentlich weiß man nur, wenn man wenig weiß;
Mit dem Wissen wächst der Zweifel.
Johann Wolfgang von Goethe (1749-1832)
O glücklich, wer noch hoffen kann,
aus diesem Meere des Irrtums aufzutauchen!
Was man nicht weiß, das eben braucht man,
und was man weiß, kann man nicht brauchen.
Johann Wolfgang von Goethe (1749-1832)
Der Zweifel ist der Beginn der Wissenschaft. Wer nichts anzweifelt, prüft nichts.
Wer nichts prüft, entdeckt nichts. Wer nichts entdeckt, ist blind und bleibt blind.
Teilhard de Chardin (1881-1955)

IV
Danksagung
Hiermit möchte ich allen Personen für Ihre Unterstützung, sowohl körperlich als auch mental,
danken, die damit zur Fertigstellung der vorliegenden Dissertation beigetragen haben.
Als allererstes gilt mein Dank Herrn Prof. Dr. W. Zimmermann, AG Mikrobiologie und
Bioverfahrenstechnik der Universität Leipzig, für die Unterstützung bei der Themenerstellung und bei
der Durchführung der Studien sowie seinem beständigen Interesse an der Entwicklung der Arbeit.
Die Erstellung der Dissertation fand im Rahmen eines Promotionsstipendiums der Deutschen
Bundesstiftung Umwelt (AZ 20004/730) statt, der ich hiermit für diese Finanzierung danken möchte.
Herrn Prof. Dr.-Ing. habil. B. Platzer, Professur Technische Thermodynamik an der Technischen
Universität Chemnitz, danke ich für die Übernahme der Zweitbegutachtung.
Frau Dr. Claudia Birkemeyer, AG Massenspekrometrie, danke ich sehr für die Unterstützung bei den
MS-Messungen, für die vielen interessanten Diskussionen und Gespräche sowie für Ihre
selbstverständliche Hilfsbereitschaft bei der Lösung verschiedenster Fragen oder Probleme.
Weiterhin danke ich Frau R. Oehme, AG Massenspekrometrie, Frau Dr. I. Neuendorf und Herrn Dr. J.
Stichel, AG Biochemie und Bioorganische Chemie, für die Unterstützung bei den durchgeführten
GC/MS, Sequenzierungs- und MALDI Tof/Tof Analysen. Ebenfalls bedanke ich mich bei Frau PD Dr. M.
Grunow, AG Biochemie und Molekularbiologie, für die Unterstützung bei der biochemischen
Datenauswertung.
Für die Durchführung eines wissenschaftlichen Auslandsaufenthalts in den Niederlanden möchte ich
mich bei der COST 868 Action für die finanzielle Unterstützung bedanken. Des Weiteren gilt mein
Dank Herrn Dr. V.A. Nierstrasz, Herrn Dr. Pramod B. Agrawal und Herrn Prof. Dr. M.M.C.G.
Warmoeskerken für die Hilfe bei der Realisierung meiner Studien. Ein besonderer Dank geht
ebenfalls an die gesamte Arbeitsgruppe „Engineering of Fibrous Smart Materials“ der Universität
Twente, die dazu beigetragen haben, dass ich mich auch weit weg wie zu Hause gefühlt habe.
Ein herzlicher Dank geht an meine Kolleginnen und Kollegen der AG Mikrobiologie und
Bioverfahrenstechnik: Fr. Blesz, Christina, Karen, Mohd, Nicole, Ren, René, Sandra, Simone, Susanne
und Thorsten, für das beflügelnde Arbeitsklima und die ständige Unterstützung. Besonders in
Erinnerung bleiben werden die gemeinsamen Feiern, Ausflüge und geselligen Beisammen sein.
Besonders herzlich bedanken möchte ich mich bei Nicole, meiner Mitdoktorandin, Tischnachbarin
und meinem „Fels im stürmischen Laboralltag“. Sie hatte immer ein offenes Ohr für alle Probleme
und Fragen und stets einen hilfreichen Tipp. Ich danke Ihr für die tolle Zusammenarbeit, die vielen
Gespräche mit oder ohne Kaffee, die gemeinsamen Tagungsbesuche und die hoffentlich noch lange
weiter bestehende Freundschaft.
Abschließend danke ich meiner Familie und meinen Freunden für ihre Unterstützung und Geduld mit
mir und meiner, für sie meist unverständlichen, Arbeit.

V
Inhaltsverzeichnis
1 Einführung 1
1.1 Actinomyceten 1
1.1.1 Klassifizierung 1
1.1.2 Thermobifida (Thermomonospora) fusca 2
1.2 Biopolyester Cutin und Suberin 4
1.2.1 Bestandteile des Cutins 4
1.2.2 Bestandteile des Suberins 5
1.2.3 Struktur der Biopolymere 7
1.3 Hydrolasen 12
1.3.1 Struktur und katalytischer Mechanismus 12
1.3.2 Carboxylesterasen 16
1.3.3 Lipasen 19
1.3.4 Cutinasen 22
1.4 Polyethylenterephthalat 23
1.4.1 Konventionelle PET-Faserbehandlung 24
1.4.2 Charakterisierung von PET 26
1.4.3 Biofunktionalisierung von PET 30
1.4.4 PET-Hydrolasen 38
1.5 Zielsetzung 49
2 Material und Methoden 50
2.1 Materialien 50
2.1.1 Verwendete Mikroorganismen 50
2.1.2 Verwendete Enzyme 50
2.1.3 Größenstandards 51
2.1.3.1 Low Molecular Weight Marker (GE Healthcare) 51
2.1.3.2 Roti®-Mark Standard (Fa. Carl Roth GmbH) 51
2.1.3.3 SpectraTM
Multicolor Broad range Protein Ladder (Fermentas) 51
2.1.3.4 PageRulerTM
plus prestained protein ladder (Fermentas) 51
2.1.3.5 Kalibrierungskit für pI Bestimmung (pH 3-10, GE Healthcare) 52
2.1.3.6 Kalibrierungskit für die Größenausschlusschromatographie (LMW, GE Healthcare) 52
2.1.4 Chemikalien 53
2.1.5 Geräte und Materialien 55

VI
2.1.6 Software 58
2.1.7 Nährmedien 58
2.1.8 Suberin- und Cutin-Präparationen 60
2.1.9 PET-Substrate für die Abbauuntersuchungen 60
2.1.10 Puffer und Lösungen 60
2.2 Mikrobiologische Methoden 65
2.2.1 Stammhaltung und Kultivierung 65
2.2.2 Mikroskopische Untersuchungen 65
2.2.3 Trockengewichtsbestimmung 65
2.3 Proteinchemische Methoden 66
2.3.1 Proteinaufreinigung der Wildtyp TfCa 66
2.3.2 Proteinaufreinigung der rekombinanten TfCa, TfCut1 und TfCut2 67
2.4 Analytische Methoden 67
2.4.1 Esterase-Aktivitätsbestimmung 67
2.4.1.1 Bestimmung mittels Spektrophotometer 68
2.4.1.2 Bestimmung mittels Plattenleser 68
2.4.2 Cutinase-Aktivitätsbestimmung 68
2.4.3 PCL-Abbauuntersuchung 69
2.4.3.1 Bestimmung mittels Hofbildung 69
2.4.3.2 Bestimmung mittels Plattenleser 69
2.4.4 Quantitative Protein-Bestimmung nach Bradford (1976) 69
2.4.5 SDS Polyacrylamidgelelektrophorese (SDS-PAGE) 70
2.4.5.1 Esteraseaktivitäts-Färbung 70
2.4.5.2 Coomassie-Färbung 71
2.4.5.3 Silberfärbung der Proteine 71
2.4.6 Bestimmung des pI 71
2.4.7 Bestimmung der Molaren Masse 72
2.4.8 Bestimmung der Temperatur und pH-Wert Stabilität 72
2.4.9 Bestimmung der Stabilität gegenüber Inhibitoren 72
2.4.10 Bestimmung der kinetischen Konstanten für die Hydrolyse von p-NP Ester 72
2.4.11 Bestimmung der kinetische Konstanten für die Hydrolyse von CTR 73
2.4.12 Bestimmung optimaler Temperatur und pH-Wert für die Hydrolyse von CTR 73
2.4.13 N-terminale Sequenzierung 73
2.4.14 MALDI-TOF Sequenzierung 74
2.4.15 Abbaustudien 74

VII
2.4.16 Analytik der PET Abbauprodukte 75
2.4.17 Konzentrationabhängige CTR-Abbaustudien 75
2.5 Homologie-Modelling der TfCa und weitere PET-Hydrolasen 76
3 Ergebnisse und Diskussion 77
3.1 Screening nach PET-Hydrolasen aus T. fusca 77
3.1.1 Wachstum von T. fusca KW3 im Czapek-Medium 77
3.1.2 Wachstum von T. fusca KW3 im MSV-Medium 78
3.1.2.1 Esterasebildung mit verschiedenen synthetischen und natürlichen Polyestern 78
3.1.2.2 Esterasebildung mit einer Suberinpräparation 81
3.1.2.3 Esterasebildung mit PET-Fasern 83
3.1.3 Esterasebildung bei T. fusca KW3 DSM 6013, T. fusca DSM 43792 und DSM 43793 mit
PET-Fasern und Diethylterephthalat 85
3.2 Charakterisierung der PET-Hydrolasen aus T. fusca KW3 95
3.2.1 Aufreinigung 95
3.2.1.1 Aufreinigung der TfCa 95
3.2.1.2 Aufreinigung der rekombinanten PET-Hydrolasen 105
3.2.2 pI der TfCa 113
3.2.3 Molare Masse der TfCa 113
3.2.4 Temperatur- und pH-Stabilität der TfCa 114
3.2.5 Wirkung von Inhibitoren auf die TfCa 117
3.2.6 Kinetik der Hydrolyse von verschiedenen Esterasesubstraten durch die TfCa 118
3.2.7 Optimale Temperatur und optimaler pH-Wert der CTR-Hydrolyse durch die TfCa 119
3.2.8 Cutinaseaktivität der TfCa 120
3.2.9 PCL-Abbau durch die TfCa 121
3.2.10 N-terminale Sequenz der TfCa 122
3.2.11 MALDI-TOF Sequenzierung der TfCa 123
3.2.12 Homologie-Modeling der TfCa und Vergleich der Struktur mit anderen Hydrolasen 126
3.2.13 Vergleich der TfCa mit bekannten PET-Hydrolasen 128
3.3 Hydrolyse von PET Substraten durch prokaryotische und eukaryotische Hydrolasen 138
3.3.1 Partielle Hydrolyse von APET-Filmen durch die PET-Hydrolasen 140
3.3.2 Partielle Hydrolyse von PET-Fasern durch die PET-Hydrolasen 148
3.3.3 Hydrolyse von PET-Trimeren durch die PET-Hydrolasen 151
4 Zusammenfassung 165

VIII
5 Literaturverzeichnis 166
6 Publikationen 181
7 Poster und Vorträge 182
8 Lebenslauf 183

IX
Abbildungsverzeichnis Seite
Abbildung 1: Phylogenetischer Stammbaum des Lebens auf Grundlage der 16S rDNA Untersuchungen. Die Gruppe der Actinobakterien ist im linken oberen Bereich angeordnet (www.lisci.kitasato-u.ac.jp/me/images/phylogenetic.jpg). 1
Abbildung 2: T. fusca KW3 mit 1000facher Vergrößerung (links, Färbung mit Safranin) und Makroskopisches Aussehen nach Wachstum auf Czapek-Medium (rechts). 3
Abbildung 3: Schematische Darstellung der Einlagerung von Cutin in einem Blatt. Die Stärke, Struktur und Zusammen-setzung der einzelnen Schichten der Cuticula können stark variieren abhängig von Spezies, Organen und Entwicklungs-phase. Die epicuticulären Wachse (EW) bedecken die Cuticula (C) komplett, die aus Cutin und darin eingelagerten intracuticulären Wachsen besteht. Die cuticulären Schichten (CL) bestehen höchstwahrscheinlich aus Cutin und Polysacchariden der Zellwand, könnten aber vielleicht auch intracuticuläre Wachse enthalten. (PW: primäre Zellwand, Cy: Cytoplasma, V: Vakuole) (Pollard et al. 2008). 5
Abbildung 4: Schematische Darstellung der Einlagerung von Suberin in der Endodermis einer Wurzel. Abgebildet sind zwei angrenzende Zellen mit Mittellamelle (ML) und Pektinschicht zwischen ihren primären Zellwänden (PW). Die Suberinlamellen befinden (SL) sich auf der Innenseite der primären Zellwand. (SW: sekundäre Zellwand, PM: Plasmamembran, Cy: Cytoplasma, V: Vakuole) (Pollard et al. 2008). 6
Abbildung 5: Modell der möglichen Bindungsstrukturen im Biopolyester Cutin (Kolattukudy 2002). 7 Abbildung 6: Modell der hypotetischen Struktur der Suberinschicht der Kartoffel. P: phenolisches Monomer, C:
Kohlenhydrat, S: Suberin (aromatisch oder aliphatisch) (Bernards 2002). 9 Abbildung 7: Darstellung der möglichen Verbindungen der Monomere der Biopolyester Cutin und Suberin. a) Hauptsächlich
treten dabei primäre Esterbindungen auf (roter Pfeil), wodurch lange Ketten gebildet werden. Die sekundären Esterbindungen (blauer Pfeil) stellen zusätzliche Verzweigungspunkte dar. b) Anordnung der Fettsäure- und ω-Hydroxyfettsäure-Monomere unter Bildung eines Dendrimers. c) Anordnung der α,ω-Dicarbonsäure- (DCS) und Glycerin-Monomere unter Bildung einer Dendrimer Struktur, angeordnet um freie OH Gruppen um eine Bindung an einige Glycerin Monomere zu ermöglichen. d) Anordnung der DCS- und Glycerin-Monomere unter Bildung einer quervernetzten Domäne. (Pollard et al. 2008). 11
Abbildung 8: α/β Hydrolase-Faltungsmotiv der Hydrolasen ist eine „doubly wound α/β superfold“ bestehend aus 8 zentral parallel angeordneten β-Faltblättern (rot) und 6 umgebenden stabilisierenden α-Helices (blau). Im Speziellen kann die Anzahl der Elemente variieren, das allgemeine Gerüst ist aber hochkonserviert. Das aktive Zentrum bilden 3 Aminosäuren (grün), das Nucleophil befindet sich im hoch konservierten Bereich zwischen der αC-Helix und dem β5-Faltblatt, dem „nucleophilic elbow“ (Ollis et al. 1992). 13
Abbildung 9: Der katalytische Reaktionsmechanismus der Hydrolasen am Beispiel der Spaltung eines Esters in die dazugehörige Säure und Alkohol. Die Reaktion erfolgt ohne Verwendung eines Cofaktors und ist somit sehr einfach und effizient. Die Bildung des Acylenzyms ist ein schneller Reaktionsschritt, die Freisetzung der Säure und des Enzyms ist der geschwindigkeitsbestimmende Reaktionsschritt. (Holmquist 2000). 14
Abbildung 10: Übersicht über die katalysierten hydrolytischen Spaltungen der Enzymgruppen der Carboxylesterase, Triacylglycerol Hydrolasen und der Cutin Hydrolasen im wässrigen Milieu. 15
Abbildung 11: Reaktionskinetik der Carboxylesterasen (links) nach Michaelis-Menten und der sigmoide Kurvenverlauf der Reaktionskinetik der Lipasen (rechts) (Pütz 2006). 16
Abbildung 12: Struktur der offenen und geschlossenen Lipase von Burkholderia cepacia BcL. Die offene Kristallstruktur ist blau dargestellt und das geschlossene Homologie-Modell grün, die gelbe Kugel stellt ein Ca
2+-Ion dar. Die katalytische
Triade besteht aus Serin, Histidin und Asparaginsäure (Trodler et al. 2009). 20 Abbildung 13: Polykondensationsreaktion von Terephthalsäure und Ethylenglycol zu Polyethylenterephthalat. 23 Abbildung 14: Struktur eines zyklischen PET-Trimers. 24 Abbildung 15: Schematische Darstellung des Schmelzspinnverfahrens von PET-Fibrillen (links) und Darstellung einer
beheizten Schmelzspinndüse (Jahreiß 2005). 26 Abbildung 16: Anordnung der PET-Ketten vor (links) und nach (rechts) dem Verstrecken unter Ausbildung von geordneten
kristallinen Bereichen (Jahreiß 2005). 27 Abbildung 17: Darstellung der Wasserstoffbrückenbindungen zwischen den durch Verstrecken entstandenen geordneten
PET-Ketten (grün). 27 Abbildung 18: Darstellung möglicher Faserquerschnitte und Oberflächen (links) und die PET-Fasern vor und nach dem
Texturieren (rechts) (Jahreiß 2005). 28 Abbildung 19: DSC-Kurve einer PET-Folie (http://web.utk.edu/~mse/Textiles/Polymer%20Crystallinity.htm). 29 Abbildung 20: Darstellung der chemischen Struktur der Polyethylenterephthalat Polymerkette, der zyklischen Trimere und
der enzymatisch freigesetzten Hydrolyseprodukte (Zimmermann und Billig 2011). 31 Abbildung 21: Darstellung der chemischen Struktur der Polytrimethylenterephthalat Polymerkette, der zyklischen Dimere
und der enzymatisch freigesetzten Hydrolyseprodukte (Zimmermann und Billig 2011). 32 Abbildung 22: Darstellung der chemischen Struktur der angewandten Polyethylenterephthalat Modellsubstrate
(Zimmermann und Billig 2011). 32 Abbildung 23: Kalibrierungskit für die pI Bestimmung, Breiter pI Bereich (pH 3-10, rechte Spur, GE Healthcare). 52 Abbildung 24: Elutionsprofil des Kalibrierungskit für die Größenausschlusschromatographie eluiert von einer XK 26/60
Säule, Superdex 200 prep grade, 320 ml SV, 50 mM Tris HCl pH7,2 150 mM NaCl, 2,5 ml/min, 280 nm und dazugehörige Kalibrierkurve (kleine Abbildung). 53

X
Abbildung 25: Wachstumskurve von T. fusca KW3 im Czapek-Medium bei 50°C und 150 rpm. 77 Abbildung 26: SDS-PAGE und Aktivitätsfärbung mit den Esterasen in den zellfreien Überständen des MSV-Mediums nach 10
tägiger Inkubation bei 50°C mit PET-Fasern (Spur 1), recyceltes PET-Granulat (Spur 2), einer PET-Oligomerpräparation (Spur 3), einer zyklischen PET-Oligomerpräparation (Spur 4), BHET (Spur 5), PBT-Fasern (Spur 6), einer Suberinpräparation (Spur 7) und einer Cutinpräparation (Spur 8). 79
Abbildung 27: Esteraseproduktion durch T. fusca KW3 mit recyceltem PET-Granulat, PET-Oligomerpräparation, zyklische PET-Oligomerpräparation, BHET, PBT und PET-Fasern über einen Zeitraum von 10 Tagen im MSV-Medium. 80
Abbildung 28: Bestimmung der Esteraseaktivitäten (◊) und der spezifischen Esteraseaktivitäten () während des Wachstums im MSV-Medium unter Zugabe von Suberin. 82
Abbildung 29: SDS-PAGE der zellfreien Überstände nach Inkubation mit Suberin im MSV-Medium, links Coomassie-gefärbtes Gel; rechts Gel mit Aktivitätsfärbung mit Fast Red. Spur 1, 7: Roti
®-Mark SDS Proteinstandard; Spur 2:
Nullwert; Spur 3, 8: zellfreier Überstand nach 2 Tagen Kultivierung; Spur 4: zellfreier Überstand nach 4 Tagen Kultivierung; Spur 5: zellfreier Überstand nach 7 Tagen Kultivierung; Spur 6: zellfreier Überstand nach 10 Tagen Kultivierung. 82
Abbildung 30: Biotrockenmasseproduktion () und Esteraseaktivität (◊) von T. fusca KW3 im MSV-Medium. 83 Abbildung 31: Graphische Darstellung der Esteraseaktivität (◊) und der spezifischen Esteraseaktivität () gebildet während
der 10-tägigen Kultivierung im MSV-Medium mit PET-Fasern. 84 Abbildung 32: SDS-PAGE der zellfreien Überstände nach Inkubation mit PET-Fasern im MSV-Medium, links Coomassie-
gefärbtes Gel; rechts Gel mit Aktivitätsfärbung mit Fast Red. Spur 1 und 7: Roti®-Mark SDS Proteinstandard; Spur 2:
Nullwert; Spur 3: zellfreier Überstand nach 2 Tagen Kultivierung; Spur 4: zellfreier Überstand nach 4 Tagen Kultivierung; Spur 5: zellfreier Überstand nach 8 Tagen Kultivierung; Spur 6, 8: zellfreier Überstand nach 10 Tagen Kultivierung. 84
Abbildung 33: SDS-PAGE der Rohenzymlösungen nach Kultivierung mit unbehandelten PET-Fasern bzw. DET im MSV-Medium, Gel gefärbt mit Coomassie- und mit Aktivitätsfärbung. Spur 1, 8: Low Molecular Weight Längenstandard; Spur 2: T. fusca DSM 43792 mit PET-Fasern kultiviert; Spur 3: T. fusca DSM 43792 mit DET kultiviert; Spur 4: T. fusca DSM 43793 mit PET-Fasern kultiviert; Spur 5: T. fusca DSM 43793 mit DET kultiviert; Spur 6: T. fusca KW3 DSM 6013 mit PET-Fasern kultiviert; Spur 7: T. fusca KW3 DSM 6013 mit DET kultiviert. 86
Abbildung 34: Esteraseproduktion durch T. fusca KW3 mit DET (◊) und PET-Fasern () über einen Zeitraum von 13 Tagen im MSV-Medium. 87
Abbildung 35: Elutionsprofil der Enzymlösung während des zweiten Aufreinigungsschritts (Phenylsepharose, Säulenvolumen 25 ml, Puffer A: 20 mM Phosphatpuffer pH 8, Puffer B: 30% Isopropanol in 20 mM Phosphatpuffer pH 8). Dargestellt sind das Proteinprofil (grau), die Esteraseaktivität (schwarz) und der Gradient der Elution (gestrichelt). 96
Abbildung 36: Elutionsprofil der Enzymlösung während des dritten Aufreinigungsschritts (DEAE-Sepharose, Säulenvolumen 54 ml, Puffer A: 20 mM Phosphatpuffer pH 8, Puffer B: 1 M NaCl in 20 mM Phosphatpuffer pH 8). Dargestellt sind das Proteinprofil (grau), die Esteraseaktivität (schwarz) und der Gradient der Elution (gestrichelt). 96
Abbildung 37: Elutionsprofil der Enzymlösung während des vierten Aufreinigungsschritts (Octyl-Sepharose, Säulenvolumen 1 ml, Puffer A: 20 mM Phosphatpuffer pH 8, Puffer B: 30% Isopropanol in 20 mM Phosphatpuffer pH 8). Dargestellt sind das Proteinprofil (grau), die Esteraseaktivität (schwarz) und der Gradient der Elution (gestrichelt). 97
Abbildung 38: Elutionsprofil der Enzymlösung während des fünften Aufreinigungsschritts (Sephadex 200, Säulenvolumen 110 ml, Puffer: 0,15 M NaCl in 20 mM Tris/HCl pH 8). Dargestellt sind das Proteinprofil (grau) und die Esteraseaktivität (schwarz). 97
Abbildung 39: SDS-PAGE der Aufreinigung der TfCa aus T. fusca KW3. Spur 1: Enzymlösung nach der Dialyse; Spur 2: Enzymlösung nach der HIC 1; Spur 3: Enzymlösung nach der AIEX; Spur 4: Enzymlösung nach der HIC 2; Spur 5: Enzymlösung nach der SEC; Spur 6: Low Molecular Weight Längenstandard (Billig et al. 2010). 98
Abbildung 40: Elutionsprofil der TfCa während des ersten Aufreinigungsschritts (HisTrap, Säulenvolumen 10 ml, Puffer A: 50 mM NaH2PO4, 300 mM NaCl; pH 8,0, Puffer B: 50 mM NaH2PO4, 300 mM NaCl, 250mM Imidazol, pH 8). Dargestellt sind das Proteinprofil (grau), die p-NPB Aktivität (schwarz) und der Gradient der Elution (gestrichelt). 106
Abbildung 41: Elutionsprofil der TfCa während des zweiten Aufreinigungsschritts (Sephadex 200 prep grade, Säulenvolumen 320 ml, Puffer: 0,15 M NaCl, 20 mM Phosphatpuffer pH 8). Dargestellt sind das Proteinprofil (grau), die p-NPB Aktivität (schwarz). 106
Abbildung 42: Elutionsprofil der TfCut2 während des ersten Aufreinigungsschritts (HisTrap, Säulenvolumen 10 ml, Puffer A: 50 mM NaH2PO4, 300 mM NaCl; pH 8,0, Puffer B: 50 mM NaH2PO4, 300 mM NaCl, 250 mM Imidazol, pH 8). Dargestellt sind das Proteinprofil (grau), die p-NPB Aktivität (schwarz) und der Gradient der Elution (gestrichelt). 107
Abbildung 43: Elutionsprofil der TfCut2 während des zweiten Aufreinigungsschritts (Sephadex 200 prep grade, Säulenvolumen 320 ml, Puffer: 0,15 M NaCl, 20 mM Phosphatpuffer pH 8). Dargestellt sind das Proteinprofil (grau), die p-NPB Aktivität (schwarz). 107
Abbildung 44: SDS-PAGE der Aufreinigung der rekombinant produzierten TfCa in E. coli (A) und der rekombinant produzierten TfCut2 in E. coli (B). Spur M: Low Molecular Weight Längenstandard; Spur 1, 4: Rohenzymlösung nach Zellaufschluss; Spur 2, 5: Enzymlösung nach Ni-Affinitätschromatographie; Spur 3, 6: Enzymlösung nach SEC. 108
Abbildung 45: PhastGel der Bestimmung des isoelektrischen Punktes der TfCa. (linkes Gel PhastGel IEF 3-9, rechtes Gel PhastGel IEF 4-6,5) Spur 1: 26,7 ng TfCa; Spur 2: 66,7 ng TfCa; Spur 3: 133 ng TfCa; Spur 4: 26,7 ng TfCa; Spur 5: 53,4 ng TfCa; Spur 6: Kalibrierungskit für die pI Bestimmung, Breiter pI Bereich (pH 3-10, GE Healthcare). 113
Abbildung 46: Laufstrecken der Proteine des Roti®-Mark SDS Proteinstandards (Fa. Carl Roth GmbH) aufgetragen gegen den
Logarithmus ihres Molekulargewichtes. 114

XI
Abbildung 47: Esteraseaktivitäten der TfCa über einen Zeitraum von 7 Tagen bei Inkubation in unterschiedlichen pH-Werten von 4 bis 11 bestimmt mit p-NPB als Substrat. 115
Abbildung 48: Esteraseaktivitäten der TfCa nach 24 Stunden Inkubation in verschiedenen pH-Werten von 3 bis 11 bestimmt mit p-NPB als Substrat. 115
Abbildung 49: Esteraseaktivitäten der TfCa über einen Zeitraum von 7 Tagen bei Inkubation bei 40°C, 50°C und 55°C bestimmt mit p-NPB als Substrat (Billig et al. 2010). 116
Abbildung 50: Esteraseaktivitäten der TfCa nach 24 Stunden Inkubation bei 30°C, 40°C, 50°C und 55°C bestimmt mit p-NPB als Substrat. 116
Abbildung 51: Lineweaver-Burk Darstellung für die enzymkatalysierte Hydrolyse der CTR durch die TfCa. Mit Hilfe der doppeltreziproken graphischen Darstellung konnten die Geschwindigkeitskonstanten Vmax und Km bestimmt werden. 119
Abbildung 52: pH und Temperaturoptimum der TfCa beim Abbau von zyklischen PET-Trimeren, bestimmt per RP HPLC (Billig et al. 2010). 119
Abbildung 53: Gaschromatographische Analyse des Blindwerts (ohne Enzym) der Cutinhydrolyse. 120 Abbildung 54: Gaschromatographische Analyse des Ansatzes mit 1 U TfCa der Cutinhydrolyse. 120 Abbildung 55: Gaschromatographische Analyse des Ansatzes mit 2 U TfCa der Cutinhydrolyse. 121 Abbildung 56: Gaschromatographische Analyse des Ansatzes mit 4 U TfCa der Cutinhydrolyse. 121 Abbildung 57: PCL-Agarplatten nach 24 stündiger Inkubation mit TfCa (linkes Photo), mit dem Kulturüberstand des Suberin-
MSV-Mediums (mittleres Photo) und mit Kulturüberstand des Cutin-MSV-Mediums (rechtes Photo). 122 Abbildung 58: Lokalisation des Tfu_2427 Gens im Genom von T. fusca YX (Quelle: http://www.ncbi.nlm.nih.gov/). 122 Abbildung 59: Proteinsequenz von Tfu_2427 aus T. fusca YX, die Aminosäuren des aktiven Zentrums und das GXSXG Motiv
sind gelb markiert. Die ersten 10 Aminosäuren am N-Terminus, anhand derer die Identifikation erfolgte, sind unterstrichen (Billig et al. 2010). 123
Abbildung 60: Proteinsequenz der TfCa aus T. fusca KW3, die Aminosäuren des aktiven Zentrums und das GXSXG Motiv sind gelb markiert, die identifizierten Unterschiede zu Tfu_2427 aus T. fusca YX sind rot markiert (Billig et al. 2010). 124
Abbildung 61: Alignment der Carboxylesterase TfCa aus T. fusca KW3 (tfu), der Carboxylesterase Arth_3983 aus Arthrobacter FB24 (art), der Carboxylesterase SCO6127 aus Streptomyces coelicolor (sco), der putativen Carboxylesterase FRAAL0556 aus Frankia alni (fal) und der putativen p-Nitrobenzylesterase SACE_2933 aus Saccharopolyspora erythraea (sen) (Billig et al. 2010). 125
Abbildung 62: Tertiärstruktur der Carboxylesterase TfCa aus T. fusca KW3. Rot sind die α-Helices gekennzeichnet und gelb die β-Faltblattstrukturen. Die grünen Bereiche sind Sequenzen ohne eine spezielle Sekundärstruktur. Die drei blau gekennzeichneten Aminosäuren sind die katalytische Triade (Billig et al. 2010). 126
Abbildung 63: Ausschnitt aus der Tertiärstruktur der Carboxylesterase TfCa aus T. fusca KW3. Rot sind die α-Helices gekennzeichnet und gelb die β-Faltblattstrukturen. Die grünen Bereiche sind Sequenzen ohne eine spezielle Sekundärstruktur. Die drei blau gekennzeichneten Aminosäuren sind die katalytische Triade. 126
Abbildung 64: Homologie-Modelle der Carboxylesterase TfCa aus T. fusca KW3 (a) verglichen mit verschiedenen PET-Cutinasen. Die Strukturen der Cutinasen aus H. insolens (b), F. solani sp. pisi (c), der Cutinasen Tfu_0882 (d) und Tfu_0883 (e) aus T. fusca KW3 und der Cutinase aus P. mendocina (f) sind dargestellt (Billig et al. 2010). 127
Abbildung 65: SDS-PAGE der für die Abbauuntersuchungen eingesetzten Hydrolasen, linkes Gel mit Aktivitätsfärbung, rechtes Gel Aktivitäts- und Coomassie-Färbung. Spur 1: Carboxylesterase TfCa aus T. fusca KW3; Spur 2: rekombinante Cutinase 1 von T. fusca WSH03-11 (Chen et al. 2010, Chen J et al. 2008); Spur 3: rekombinante Cutinase 2 von T. fusca WSH03-11 (Chen et al. 2010, Chen J et al. 2008); Spur 4: rekombinante Cutinase von F. solani pisi (Chen et al. 2010); Spur 5: Cutinase aus Fusarium solani sp. pisi (Vertommen et al. 2005); Spur 6: Cutinase von Novozymes. Zum Vergleich die Kulturüberstände verschiedener MSV-Medien, Spur 7: Cutin-induzierte Hydrolase aus T. fusca KW3; Spur 8: Suberin-induzierte Hydrolase aus T. fusca KW3; St
1: Spectra™ Multicolor Broad Range Protein Ladder (Fermentas); St
2:
PageRulerTM
Plus Prestained Protein Ladder (Fermentas). 138 Abbildung 66: PCL Abbautest der für die Abbauuntersuchungen eingesetzten Hydrolasen (obere Reihe: Carboxylesterase
TfCa aus T. fusca KW3 (links), rekombinante Cutinase 1 von T. fusca WSH03-11 (Mitte, Chen et al. 2010, Chen J et al. 2008), rekombinante Cutinase 2 von T. fusca WSH03-11 (rechts, Chen et al. 2010, Chen J et al. 2008); mittlere Reihe: rekombinante Cutinase von F. solani pisi (links, Chen et al 2010), Cutinase aus F. solani sp. pisi (Mitte, Vertommen et al. 2005), Cutinase von Novozymes (rechts); untere Reihe : rekombinant produzierten Hydrolasen aus T. fusca KW3: Carboxylesterase TfCa (links), Cutinase TfCut1 (Mitte), Cutinase TfCut2 (rechts). 139
Abbildung 67: Detektierte Hydrolyseprodukte nach Inkubation des APET-Films mit TfCa aus T. fusca KW3. 140 Abbildung 68: Detektierte Hydrolyseprodukte nach Inkubation des APET-Films mit der Cutinase 2 aus T. fusca WSH03-11
von Chen et al. 2010 und Chen J et al. 2008. 140 Abbildung 69: Detektierte Hydrolyseprodukte nach Inkubation des APET-Films mit der Cutinase aus F. solani pisi von
Vertommen et al. 2005. 141 Abbildung 70: Detektierte Hydrolyseprodukte nach Inkubation des APET-Films mit der Cutinase aus F. solani pisi von Chen
et al. 2010. 141 Abbildung 71: Detektierte Hydrolyseprodukte nach Inkubation des APET-Films mit der Cutinasepräparation von
Novozymes. 142 Abbildung 72: Rasterelektronenmikroskopische Analyse der enzymatisch behandelten APET-Film Oberflächen:
Unbehandelte Kontrolle (A), Inkubation mit TfCa bei 50°C 74 h (B) und Inkubation mit Cutinase FsC bei 30°C 74 h (C). 144
Abbildung 73: Chromatogramm der HPLC Analyse des enzymatischen APET Abbaus durch die TfCa. 146

XII
Abbildung 74: MS-Spektren der Hydrolyseprodukte: TPA (m/z: 121,0; 164,9; 352,9), MHET (m/z: 120,9; 208,8; 440,9), BHET (m/z: 120,9; 252,9), EBT (m/z: 357,0) und EMT (m/z: 401,0). 147
Abbildung 75: Detektierte Hydrolyseprodukte nach Inkubation der PET-Fasern mit TfCa aus T. fusca KW3. 148 Abbildung 76: Detektierte Hydrolyseprodukte nach Inkubation der PET-Fasern mit der Cutinase 2 aus T. fusca WSH03-11
von Chen et al. 2010, Chen J et al. 2008. 148 Abbildung 77: Detektierte Hydrolyseprodukte nach Inkubation der PET-Fasern mit der Cutinase aus F. solani pisi von
Vertommen et al. 2005. 149 Abbildung 78: Detektierte Hydrolyseprodukte nach Inkubation der PET-Fasern mit der Cutinase aus F. solani pisi von Chen
et al. 2010. 149 Abbildung 79: Detektierte Hydrolyseprodukte nach Inkubation der PET-Fasern mit der Cutinasepräparation von Novozymes.
150 Abbildung 80: Detektierte Hydrolyseprodukte nach Inkubation der zyklischen PET-Trimeren mit TfCa aus T. fusca KW3. 152 Abbildung 81: Detektierte Hydrolyseprodukte nach Inkubation der zyklischen PET-Trimeren mit der Cutinase 1 aus T. fusca
WSH03-11 von Chen et al. 2010, Chen J et al. 2008. 152 Abbildung 82: Detektierte Hydrolyseprodukte nach Inkubation der zyklischen PET-Trimeren mit der Cutinase 2 aus T. fusca
WSH03-11 von Chen et al. 2010, Chen J et al. 2008. 153 Abbildung 83: Detektierte Hydrolyseprodukte nach Inkubation der zyklischen PET-Trimeren mit rekombinanter TfCut1. 153 Abbildung 84: Detektierte Hydrolyseprodukte nach Inkubation der zyklischen PET-Trimeren mit rekombinanter TfCut2. 154 Abbildung 85: Detektierte Hydrolyseprodukte nach Inkubation der zyklischen PET-Trimeren mit der Cutinase aus
F. solani pisi von Vertommen et al. 2005. 154 Abbildung 86: Detektierte Hydrolyseprodukte nach Inkubation der zyklischen PET-Trimeren mit der Cutinase aus
F. solani pisi von Chen et al. 2010. 155 Abbildung 87: Detektierte Hydrolyseprodukte nach Inkubation der zyklischen PET-Trimeren mit der Cutinase-Präparation
von Novozymes. 155 Abbildung 88: Effekt der rTfCa Konzentration auf die Hydrolyse von PET-Trimeren (Reaktionszeit 4 Stunden). 159 Abbildung 89: Effekt der rTfCut2 Konzentration auf die Hydrolyse von PET-Trimeren (Reaktionszeit 4 Stunden). 160 Abbildung 90: Effekt der Reaktionszeit auf die Hydrolyse von PET-Trimeren mit rTfCa. 161 Abbildung 91: Effekt der Reaktionszeit auf die Hydrolyse von PET-Trimeren mit rTfCut2. 161
Tabellenverzeichnis
Tabelle 1: Vertreter der Actinomyceten-Gruppe (Madigan et al. 2009). 2 Tabelle 2: Hauptmonomere des Cutins: C16: Hexadecansäure, 16-Hydroxyhexadecansäure, Dihydroxyhexadecansäure; 5 C18: Octadeca-9-ensäure, 18-Hydroxyoctadeca-9-ensäure, 18-Hydroxy-cis-9,10-epoxyoctadecansäure, 9,10,18-
Trihydroxyoctadecansäure. (*Δ12 ungesättigte Analoga können auftreten; y = 8,7,6,oder 5; x+y = 13) (Kolattukudy 2002). 5
Tabelle 3: Struktur der Monomere Suberins: Aliphatische Monomere: 1-Alkanole; Alkansäuren; ω-Hydroxyalkansäuren; α,ω-Dialkansäuren; 9(10),ω-Dihydroxyalkansäure; 9,10-Dihydroxyalkansäure; 9,10,18-Trihydroxyalkansäure; 9,10-Dihydroxy-α,ω-Dialkansäure; 9,10-Epoxy-ω-Hydroxyalkansäure; 9,10-Epoxy-α,ω-Dialkansäure; Ferulasäure-Ester (mit 1-Alkanole); Glycerin. Aromatische Monomere: a) R1=R2=H, p-Cumarsäure; R1=OH, R2=H, Kaffeesäure; R1=OCH3, R2=H, Ferulasäure; R1=R2=OCH3, Sinapinsäure. b) R1=R2=H, p-Cumarylalkohol; R1=OCH3, R2=H, Coniferylalkohol; R1=R2=OCH3, Sinapylalkohol. c) R1=H, p-Cumaroyltyramin; R1=OCH3, Feruloyltyramin. (Bernards 2002). 6
Tabelle 4: Gegenüberstellung der prozentualen Anteile der typischen Cutin- und Suberin-Monomere. (Pollard et al. 2008). 10
Tabelle 5: Übersicht lipolytische Enzyme I, Esterasen (Gruppe II-VIII) mit ausgewählten Beispielen (Hausmann und Jaeger 2010). 18
Tabelle 6: Übersicht lipolytische Enzyme II, Lipasen (Grup. I.1-I.8) mit ausgewählten Beispielen (Hausmann und Jaeger 2010). 21
Tabelle 7: PET Biofunktionalisierung und analysierte Effekte (Zimmermann und Billig 2011). 33 Tabelle 8: Vergleich der PET-Hydrolasen von F. solani (Carvalho et al. 1998, Carvalho et al. 1999, Ronkvist et al. 2009),
T. insolens (Sandal et al. 1996, Ronkvist et al. 2009), P. mendocina (Bott et al. 2003, Ronkvist et al. 2009) und T. fusca (Kleeberg et al. 2005, Chen S et al. 2008). 39
Tabelle 9: Das Kalibrierungskit für die Größenausschlusschromatographie. 52 Tabelle 10: Zusammensetzung des Trenn- und Sammelgels. 70 Tabelle 11: Zeitlicher Ablauf der Silberfärbung. 71 Tabelle 12: Esteraseproduktion durch T. fusca KW3 im MSV-Medium mit verschiedenen Additiven über einen Zeitraum von
10 Tagen. 80 Tabelle 13: Bildung von PET-Hydrolasen bei Bakterien. 93 Tabelle 14: Bildung von PET-Hydrolasen bei Pilzen. 94 Tabelle 15: Aufreinigungstabelle der TfCa aus T. fusca KW3 (Billig et al. 2010). 98 Tabelle 16: Aufreinigungstabelle der TfH aus T. fusca DSM 43793 (Kleeberg et al. 2005). 99

XIII
Tabelle 17: Aufreinigungstabelle der Cutin-induzierten p-NPB Hydrolase aus T. fusca (Chen S et al. 2008). 100 Tabelle 18: Aufreinigungstabelle der Cutinase aus P. mendocina ATCC 55613 (Sebastian et al. 1988). 100 Tabelle 19: Aufreinigungstabelle der Cutinasen und p-NPP Hydrolase aus F. solani f. pisi (Purdy und Kolattukudy 1975a). 102 Tabelle 20: Aufreinigungstabelle der extrazellulären Cutinasen (Lin und Kolattukudy 1980a). 103 Tabelle 21: Aufreinigungstabelle der PET-Hydrolase aus P. citrinum (Liebminger et al. 2007). 104 Tabelle 22: Aufreinigungstabelle der rekombinanten TfCa. 108 Tabelle 23: Aufreinigungstabelle der rekombinanten TfCut2. 108 Tabelle 24: Aufreinigungstabelle der rekombinanten periplasmatischen (pp-rTfH) und zytoplasmatischen (zp-rTfH) TfH
(Dresler et al. 2006). 109 Tabelle 25: Expression der rekombinanten TfH in E. coli und B. megaterium (Yang et al. 2007). 109 Tabelle 26: Übersicht zur Affinitätsaufreinigung der His6-Tag rTfH produziert durch B. megaterium WH323-pYYBm9
gewachsen in einer LB-Batch Kultivierung und Hochzelldichten-Kultivierung (HZDK) mit A5 Medium. I:ohne Vorhbehandlung; II: 50°C für 10 min; III: 18 Stunden Dialyse bei 4°C (Yang et al. 2007). 110
Tabelle 27: Übersicht über die verbleibenden Esteraseaktivitäten nach der Inkubation der TfCa mit verschiedenen Inhibitoren. 117
Tabelle 28: Kinetische Parameter der Hydrolyse von p-NP-Estern und PET-Trimeren durch die TfCa aus T. fusca KW3. 118 Tabelle 29: Identifizierte N-terminale Sequenz der TfCa aus T. fusca KW3. 122 Tabelle 30: Vergleich der Eigenschaften der TfCa mit Cutinasen aus T. fusca. 128 Tabelle 31: Vergleich der PET-Hydrolasen aus F. solani FsC (Carvalho et al. 1998, Carvalho et al. 1999, Ronkvist et al. 2009),
T. insolens HiC (Sandal et al. 1996, Ronkvist et al. 2009), P. mendocina PmC (Bott et al. 2003, Ronkvist et al. 2009), T. fusca TfH/Cutinase 2 (Kleeberg et al. 2005, Chen S et al. 2008, Chen J et al. 2008) und TfCa (Zimmermann und Billig 2011). 133
Tabelle 32: Kinetische Parameter Km und Vmax bestimmt für verschiedene p-NP Ester für die Hydrolasen TfCa, TfH 1/2 (Chen et al. 2010), PmC
a (Sebastian et al. 1988), PmC
b (Gray et al. 1995), FsC (Purdy und Kolattukudy 1973), FsC 1, FsC 2 und
FsE (Purdy und Kolattukudy 1975b). Die geringsten Km Werte je Enzym sind hervorgehoben, ebenso die höchsten Vmax Werte. 137
Tabelle 33: Vergleich der freigesetzten Abbauprodukte der PET-Hydrolasen mit APET-Film. 142 Tabelle 34: Vergleich der freigesetzten Abbauprodukte der PET-Hydrolasen mit PET-Fasern. 150 Tabelle 35: Vergleich der freigesetzten Abbauprodukte der PET-Hydrolasen mit PET-Trimeren. 156 Tabelle 36: Vergleich der Abbauprodukte der PET Hydrolasen mit den drei PET Substraten. Die beiden Hauptprodukte des
Enzyms sind blau unterlegt. 158 Tabelle 37: Proteinkonzentration der eingesetzten Enzyme und die dazugehörigen p-NPB-Aktivitäten in den Ansätzen des
Abbauversuchs. 159

XIV
Einheiten
% Prozent
°C Grad Celsius
µg Mikrogramm
µm Mikrometer
bp Basenpaar
cm Zentimeter
cm2 Quadratzentimeter
d Tag
g/L Gramm / Liter
g/ml Gramm / Milliliter
kDa Kilodalton
LU/ml Lipase Units / Milliliter
m/min Meter / Minute
mg Milligramm
mg/ml Milligramm / Milliliter
mm Millimeter
mM Millimolar
nm Nanometer
S Svendberg
U/ml Units / Milliliter
Abkürzungen
Abkürzungen, die dem allgemeinen deutschen Sprachgebrauch entsprechen, werden nicht
aufgeführt.
2D-Elektrophorese 2-Dimensionale Elektrophorese
3PET Bis(Benzoyloxyethyl)terephthalat
AFM Atomkraftmikroskopie
AIEX Anionenaustauschchromatographie
APET amorphes Polyethylenterephthalat
AS Aminosäure
BEB Ethylenglycol-dibenzylester

XV
BETEB Bis(Benzoyloxyethyl)terephthalat
BHET Bis(2-Hydroxyethylterephthalat)
BHPT Bis(3-Hydroxypropylterephthalat)
BTA Copolyester aus 1,4-Butanediol, Dimethylterephthalat und
Adipinsäure
C. antarctica Candida antarctica
C. cladosporioides Cladosporium cladosporioides
CIEX Kationenaustauschchromatographie
CTR Cyclo-tris-Ethylenterephthalat
DEPA N,N-Diethyl-2-Phenylacetamid
DET Diethylterephthalat
DMSO Dimethylsulfoxid
DMT Dimethylterephthalat
DNA Deoxyribonukleinsäure
DP Diethyl-p-phthalat
DSC Differentialrasterkalorimetrie
DTT Dithiothreitol
EBT 1,2-Ethylen-bis-Terephthalat
EDTA Ethylendiamintetraacetat
EMT 1,2-Ethylen-mono-Terephthalat-mono(2-Hydroxyethylterephthalat)
ESCA Elektronenspektroskopie für chemische Analyse
ESEM Umgebungsrasterelektronenmikroskopie
et al. und andere (lat.)
ETE Diethylterephthalat
F. solani Fusarium solani
FsC Fusarium solani Cutinase
FTIR Fourier-Transformations-Infrarotspektrometrie
GC Gaschromatographie
GC-Gehalt Guanin-Cytosin-Gehalt
GC-MS Gaschromatographie-Massenspektrometrie
GPET Polyethylenterephthalat Granulat
gram (-) gram negativ
gram (+) gram positiv
G. stearothermophilus Geobacillus stearothermophilus
HIC Hydrophobe Interaktionschromatographie

XVI
HPLC Hochleistungsflüssigkeitschromatographie
HTA Hydroxyterephthalat
K/S Verhältnis Farbintensitätsverhältnis, bestimmt mit der Kubelka-Munk Gleichung
LB-Medium Lysogenie Lösung (lysogeny broth)
LC-MS Flüssigkeitschromatographie-Massenspektrometrie
LC-MS/MS Flüssigkeitschromatographie-Massenspektrometrie/
Massenspektrometrie
MG Molekulargewicht
MHET Mono(2-Hydroxyethyl)Terephthalat
MHPT Mono(3-Hydroxypropyl)Terephthalat
mRNA Boten-Ribonukleinsäure
MSTFA N-Methyl-N-trimethylsilyl-trifluoracetamid
MSV-Medium Mineralsalz-Spurenelemente-Vitamin Medium
NaOH Natriumhydroxid
OPET höher kristallines biaxial orientiertes Polyethylenterephthalat
P. mendocina Pseudomonas mendocina
PBT 1,2-Propylen-bis-Terephthalat
PCL Poly-ε-caprolacton
PET Dimer Bis-(p-Methylbenzoesäure)Ethylenglycolester
PET Trimer Bis-(Benzoyloxyethyl)Terephthalat
PET Polyethylenterephthalat
PET-B Polyethylenterephthalat-Pellets
p-NPA para-Nitrophenylacetat
p-NPB para-Nitrophenylbutyrat
p-NPC para-Nitrophenylcaprylat
p-NPP para-Nitrophenylpalmitat
PmC Pseudomonas mendocina Cutinase
PMSF Phenylmethylsulfonylfluorid
PTT Dimer Cyclo-Di-Trimethylenterephthalat
PTT Polytrimethylenterephthalat
PVDF Polyvinylidenfluorid
rDNA ribosomale Deoxyribonukleinsäure
RP-HPLC Umkehrphasen-Hochleistungsflüssigkeitschromatographie
Rt Retentionszeit
SDS-PAGE Natriumdodecylsulfat-Polyacrylamid-Elektrophorese

XVII
SEC Größenausschlusschromatographie
SeL Streptomyces exfoliates Lipase
SEM Rasterelektronenmikroskopie
T Referenzstamm
T. fusca Thermobifida fusca
T. insolens Thermomyces insolens
T. lanuginosus Thermomyces lanuginosus
Td Denaturierungstemperatur
TDOC Natrium-Taurodeoxycholat
TFA Tetrahyrofuran
TfCa Thermobifida fusca Carboxylesterase
TfCut Thermobifida fusca Cutinase
TfH Thermobifida fusca Hydrolase
Tfu Thermobifida fusca
Tg Glasübergangstemperatur
TiC Thermomyces insolens Cutinase
Tm Schmelztemperatur
TPA Terephthalsäure
TPCK Tosyl-L-Phenylalanin-Chloromethylketon
UV Ultraviolett
XPS Röntgenphotoelektronenspektroskopie
Aminosäuren:
A Ala Alanin I Ile Isoleucin R Arg Arginin
C Cys Cystein K Lys Lysin S Ser Serin
D Asp Asparaginsäure L Leu Leucin T Thr Threonin
E Glu Glutaminsäure M Met Methionin V Val Valin
F Phe Phenylalanin N Asn Asparagin W Trp Trytophan
G Gly Glycin P Pro Prolin Y Tyr Tyrosin
H His Histidin Q Gln Glutamin

1 Einführung
1 Einführung
1.1 Actinomyceten
1.1.1 Klassifizierung
Actinomyceten sind filamentöse, Sporen bildende gram (+) Bodenbakterien. Die Angehörigen dieser
Gruppe weisen einen hohen GC-Gehalt von 63-78% auf. Actinomyceten gehören zur Klasse der
Actinobakterien, zu denen auch die Corynebakterien, Arthrobacter, Brevibakterien,
Propionibakterien, Eubakterien, Bifidobakterien, Acetobakterien und Thermoanaerobacter zählen.
Die Actinomyceten werden aufgrund ihrer Eigenschaften in 8 Gruppen unterteilt (Tabelle 1).
Abbildung 1: Phylogenetischer Stammbaum des Lebens auf Grundlage der 16S rDNA Untersuchungen. Die Gruppe der Actinobakterien ist im linken oberen Bereich angeordnet (www.lisci.kitasato-u.ac.jp/me/images/phylogenetic.jpg).
Actinobacteria

2 Einführung
Tabelle 1: Vertreter der Actinomyceten-Gruppe (Madigan et al. 2009).
Bezeichnung, Eigenschaften und Beispiele GC-Gehalt
(DNA)
Actinomyceten: nicht säurefest, fakultativ aerob, keine Myzelbildung, verzweigt;
Actinomyces (anaerob bis fakultativ aerob, filamentöse Mikrokolonien, Filamente jedoch
transitorisch, fragmentieren zu coryneformen Zellen, können human-/tierpathogen sein,
kommen in Mundhöhle vor),
Arachnia, Bacterionema, Rothia, Agromyces
57-69
Mycobakterien: säurefest, übergangsweise Filamente;
Mycobacterium (pathogene Saprophyten, obligat aerob, wächst langsam, Lipidgehalt der
Zellen und Zellwände hoch, enthält Wachse und Mycolsäure; verursacht Tuberkulose,
Lepra, Granulome, Vogeltuberkulose, auch Bodenmikroben)
62-70
Stickstofffixierende Actinomyceten: Symbionten von Pflanzen, bilden echte Mycelien;
Frankia (bilden Knöllchen an Pflanzenwurzeln, wahrscheinlich mikroaerophil, wächst
langsam, fixiert Stickstoff)
67-72
Actinoplanes: bilden echte Mycelien, bildet Sporen, die in Sporangien liegen
Actinoplanes, Streptosporangium
69-71
Dermatophilus-Gruppe: bilden große Mengen beweglicher, kokkoider Elemente,
Luftmycel fehlt, gelegentlich verantwortlich für Infektionen der Oberhaut (Epidermis)
Dermatophilus, Geodermatophilus
56-75
Nocardien: Mycelfäden brechen häufig unter Bildung kokkoider oder länglicher
Elemente auseinander, gelegentlich Bildung von Luftsporen, manchmal säurefest,
Lipidgehalt der Zellen und Zellwände sehr hoch
Nocardia: verbreiteter Bodenorganismus, obligat aerob, viele
Kohlenwasserstoffverwerter
Rhodococcus: Bodensaprophyten, verbreitet im Darm verschiedener Insekten, verwerten
Kohlenwasserstoffe
61-72
59-69
Streptomyceten: Mycel bleibt intakt, üppiges Luftmycel und lange Sporenketten
Streptomyces: fast 500 gültig beschriebene Arten, viele bilden Antibiotika
Andere Gattungen (morphologisch unterschieden): Streptoverticillium, Sporichthya,
Kitasatoa, Chainia
69-75
67-73
Micromonospora-Gruppe: Mycel intakt, Sporen werden einzeln, in Paaren oder kurzen
Ketten gebildet, einige thermophil, Saprophyten des Erdbodens und verrottender
Pflanzenmasse, eine Art bildet Endosporen
Micromonospora, Microbispora, Thermobispora, Thermoactinomyces,
Thermomonospora
54-79
1.1.2 Thermobifida (Thermomonospora) fusca
Die Kolonien von T. fusca sind farblos, das gebildete Luftmycel hingegen weiß und scharf umgrenzt.
Die Lufthyphen sind geradlinig oder gewunden, einfach oder verzweigt und bilden einzelne
akropetale Luftsporen. T. fusca ist nicht säurefest, eurythermal thermophil und fakultativ anaerob

3 Einführung
(Hensen 1957). Das Temperaturoptimum für das Wachstum liegt bei 45-50°C und der optimale pH
von 7,5 bis 11. T. fusca ist Katalase stark positiv, Oxidase negativ, die Säureproduktion aus D-Glucose
ist schwach positiv und die Nitratreduktion negativ. Der Mikroorganismus kann Casein, Gelatine,
Kreatin und Elastin proteolytisch hydrolysieren. Es erfolgt ebenfalls eine Hydrolyse von Stärke, Agar,
Pektin, Cellulose, Xylan, Arbutin und Aesculin, hingegen keine von Chitin, Xanthin, Hypoxanthin,
Guanin und Tyrosin. T. fusca kann die Kohlenhydrate D-Galaktose und Laktose verwerten, D-Ribose
und D-Xylose hingegen nicht (McCarthy et al. 1984).
Phylogenetische Untersuchungen auf Grundlage der 16S rRNA zeigten keine enge
verwandtschaftliche Verbindung zu den anderen Gruppen des Actionmyceten-Klusters (Embley et al.
1994).
Abbildung 2: T. fusca KW3 mit 1000facher Vergrößerung (links, Färbung mit Safranin) und Makroskopisches Aussehen nach Wachstum auf Czapek-Medium (rechts).
Aufgrund der vielfältigen hydrolytischen Aktivität von T. fusca wurden in der Vergangenheit bereits
verschiedenste Untersuchungen zur Identifikation und Aufreinigung von hydrolytischen Enzymen
durchgeführt. Dabei erfolgte unter anderem eine Charakterisierung der Polygalakturonat-Lyase (EC
4.2.2.2) (Stutzenberger 1987), des Xylan-abbauenden Systems (Endoxylanase, α-
Arabinofuranosidase, β-Xylosidase) (Bachmann et al. 1991), einer Maltotriose produzierende α-
Amylase (Busch et al. 1997), einer Acetylxylanesterase (Yang et al. 2008) und von Cutinasen
(Kleeberg et al. 2005, Cheng et al. 2008).
Durch die lückenlose Sequenzierung der genomischen DNA von T. fusca (Lykidis et al. 2007) können
nun mit Hilfe von molekularbiologischen Methoden weitere interessante Proteine identifiziert und
charakterisiert werden. Die Proteom- und Transkriptom-Analyse der extrazellulären Proteine und des
mRNA Levels mittels 2D-Elektrophorese und LC-MS/MS erfolgte ebenfalls für einen T. fusca Stamm
nach Wachstum auf Cellubiose- und Glucose-haltigem Medium (Chen et al. 2007).

4 Einführung
T. fusca KW3 (DSM 6013) wurde aus Kompost isoliert und es erfolgten bereits Studien zur
thermophilen Xylanase-Produktion mit diesem Stamm (Röthlisberger et al. 1992, Casimir-Schenkel
1992). Das Neuisolat unterscheidet sich durch eine Resistenz gegenüber Kristallviolett, einer oberen
Wachstumsgrenze bei 60°C und einer Alkalitoleranz bis pH 11.
Ein besonderes Interesse bei der Suche nach technisch anwendbaren Enzymen gilt dem Abbau der
Biopolymere Cutin und Suberin, von Pflanzen gebildete natürliche Polyester aus aliphatischen und
aromatischen Regionen. Cutin ist ein Bestandteil der Cuticula, welche die Barriere zwischen der
Pflanze und ihrer Umwelt bildet. Die Cuticula verhindert das Austrocknen der Pflanze, reguliert den
Gasaustausch und schützt vor mechanischen und Strahlungsschäden ebenso wie vor
Pflanzenfressern und pathogenen mikrobiellen Attacken. Suberin hingegen, befindlich unter
Anderem in Korkzellen und im Periderm (äußere Barriere der Rinde und der unterirdischen
Pflanzenteile wie Wurzeln und Knollen), dient als Diffusionsbarriere (Nawrath 2002, Kolattukudy
2002).
T. fusca produziert ebenfalls diese interessanten hydrolytischen Enzyme, auf deren Aktivität und
Bedeutung in den folgenden Kapiteln näher eingegangen wird (Fett et al. 1999, Kleeberg et al. 2005,
Cheng et al. 2008).
1.2 Biopolyester Cutin und Suberin
1.2.1 Bestandteile des Cutins
Die Bezeichnung Cutin wurde von Cutose abgeleitet, was die ursprüngliche Bezeichnung für eine
Gewebsschicht der Epidermis von Blättern war, welche eine Resistenz gegenüber starken Säuren
vorwies. Cutin wurde sowohl in den Blättern von Land-, als auch von Wasserpflanzen gefunden, aber
ebenfalls in Stengeln, Blattstielen, Blütenteilen, Früchten, einige Samenschalen und im Inneren von
Zitronen. Die Stärke der Cutinschicht variiert je nach Spezies und Pflanzenteil. In den Blättern von
höheren Pflanzen beträgt die Dicke der Cutinschicht zwischen 0,5 und 14 µm mit < 20 bis 600 µg
Cutin pro cm2 Oberfläche. In manchen Früchten mit hochentwickelter Cuticula beträgt der
Cutingehalt bis zu 1,5 mg pro cm2. Bei niederen Pflanzen ist die Cutinschicht sehr dünn (~0,1 µm)
(Kolattukudy 2002).

5 Einführung
Abbildung 3: Schematische Darstellung der Einlagerung von Cutin in einem Blatt. Die Stärke, Struktur und Zusammen-setzung der einzelnen Schichten der Cuticula können stark variieren abhängig von Spezies, Organen und Entwicklungs-phase. Die epicuticulären Wachse (EW) bedecken die Cuticula (C) komplett, die aus Cutin und darin eingelagerten intracuticulären Wachsen besteht. Die cuticulären Schichten (CL) bestehen höchstwahrscheinlich aus Cutin und Polysacchariden der Zellwand, könnten aber vielleicht auch intracuticuläre Wachse enthalten. (PW: primäre Zellwand, Cy: Cytoplasma, V: Vakuole) (Pollard et al. 2008).
Typische Monomere des Cutins sind C16 und C18 Fettsäuren, welche auch ein- oder mehrfach
hydroxyliert, ungesättigt oder epoxyliert sein können (Kolattukudy 2002).
Tabelle 2: Hauptmonomere des Cutins: C16: Hexadecansäure, 16-Hydroxyhexadecansäure, Dihydroxyhexadecansäure; C18: Octadeca-9-ensäure, 18-Hydroxyoctadeca-9-ensäure, 18-Hydroxy-cis-9,10-epoxyoctadecansäure, 9,10,18-
Trihydroxyoctadecansäure. (*Δ12 ungesättigte Analoga können auftreten; y = 8,7,6,oder 5; x+y = 13) (Kolattukudy 2002).
C16-Familie C18-Familie *
COOH
OH COOH
COOHOH
OH
x
y
COOH
OH COOH
COOHOOH
OH COOH
OH
OH
1.2.2 Bestandteile des Suberins
Suberin ist abgleitet von dem lateinischen Wort suber, was Kork bedeutet. Der Begriff Suberin wurde
bereits vor 200 Jahren von Michel Eugène Chevreul (1786-1889) verwendet, um den im Kork
enthaltenen wasserunlöslichen Bestandteil zu beschreiben. Dieses mehrteilige Material besteht aus
verschiedenen polymeren Domänen, die kovalent an die hoch komplexe Zellwand gebunden sind.

6 Einführung
Abbildung 4: Schematische Darstellung der Einlagerung von Suberin in der Endodermis einer Wurzel. Abgebildet sind zwei angrenzende Zellen mit Mittellamelle (ML) und Pektinschicht zwischen ihren primären Zellwänden (PW). Die Suberinlamellen befinden (SL) sich auf der Innenseite der primären Zellwand. (SW: sekundäre Zellwand, PM: Plasmamembran, Cy: Cytoplasma, V: Vakuole) (Pollard et al. 2008).
Tabelle 3: Struktur der Monomere Suberins: Aliphatische Monomere: 1-Alkanole; Alkansäuren; ω-Hydroxyalkansäuren; α,ω-Dialkansäuren; 9(10),ω-Dihydroxyalkansäure; 9,10-Dihydroxyalkansäure; 9,10,18-Trihydroxyalkansäure; 9,10-
Dihydroxy-α,ω-Dialkansäure; 9,10-Epoxy-ω-Hydroxyalkansäure; 9,10-Epoxy-α,ω-Dialkansäure; Ferulasäure-Ester (mit 1-Alkanole); Glycerin. Aromatische Monomere: a) R1=R2=H, p-Cumarsäure; R1=OH, R2=H, Kaffeesäure; R1=OCH3, R2=H,
Ferulasäure; R1=R2=OCH3, Sinapinsäure. b) R1=R2=H, p-Cumarylalkohol; R1=OCH3, R2=H, Coniferylalkohol; R1=R2=OCH3, Sinapylalkohol. c) R1=H, p-Cumaroyltyramin; R1=OCH3, Feruloyltyramin. (Bernards 2002).
Aliphatische Monomere Aromatische Monomere OH
COOH
COOHOH
COOHHOOC
COOHOH
OH
COOH
OH
OH
COOH
OH
OH
HOOC
COOH
OOH
COOHO
HOOC
OO
OH
OMe
OHOH
OH
a)
OHO
OH
R1R2
b)
CH2OH
OH
R1R2
c) OH
R1R2
O N
OH
H

7 Einführung
Suberin besteht aus polyaliphatischen Bereichen mit langkettigen Hydroxyfettsäuren und
Dicarboxylsäuren und geringeren Mengen an sehr langen Fettsäuren und Fettalkoholen. Desweiteren
gibt es noch polyaromatische Domänen mit verschiedenen aromatischen Carbonsäuren und
Alkoholen, deren genaue Struktur bisher noch nicht eindeutig bekannt ist. Diese Domänen können
auch aufgrund ihres Aussehens unterschieden werden. Die aromatischen Domänen zeigen eine
dunklere Färbung gegenüber den aliphatischen Domänen. (Bernards 2002, Kolattukudy 2002).
1.2.3 Struktur der Biopolymere
Bisher wurden die verschiedensten Modelle zur möglichen Struktur von Cutin und Suberin
veröffentlicht, allerdings gibt es bisher keine genaue Strukturaufklärung. Die nachfolgenden
Ausführungen verdeutlichen die Probleme der Strukturidentifikation aufgrund der Vielzahl an
Monomeren und möglichen Bindungsstellen.
Abbildung 5: Modell der möglichen Bindungsstrukturen im Biopolyester Cutin (Kolattukudy 2002).
Da Cutin ein unlösliches und kompaktes Polymer ist, gibt es nur wenige Möglichkeiten für eine
Strukturanalyse. Bei einer Transmissions-Fourier-Transformations-Infrarot-Spektrum Analyse (FTIR)
konnten Absorptionen in Bereichen für Hydroxylgruppen (-OH), für aliphatische C-H Bindungen, für
Carbonylester-Bindungen (C=O) und C-O Ester identifiziert werden. Des Weiteren wurden
verschiedene Analysen zur Identifikation von freien funktionellen Gruppen im Polymer durchgeführt.

8 Einführung
Als Schlussfolgerung aus den durchgeführten Untersuchungen liegen die primären Alkoholgruppen
im Cutin als Esterbindungen vor, das bedeutet dass das Polymer, ein Polyester, aus hauptsächlich
primären Alkoholesterverbindungen besteht. Etwa die Hälfte der sekundären Alkohol-Gruppen ist
ebenfalls verestert, was bedeutet, dass Verzweigungen und/oder Quervernetzungen vorliegen.
Basierend auf diesem Wissen wurde eine Modelstruktur des Cutins erstellt (Abbildung 5).
Weitere Analysen ergaben, dass Cutin ein moderat bewegliches Geflecht darstellt mit
Bewegungseinschränkungen wahrscheinlich auf Grund der quervernetzten Seitenketten. Cutin
besteht größtenteils aus mittelständigen sekundären Hydroxylgruppen, die dadurch eine große
Bedeutung bei der Quervernetzung des Polymers spielen. Für weitere Untersuchungen der Struktur
des Cutins wurden auch enzymatisch freigesetzte Cutin-Oligomere analysiert. Dabei erfolgte der
Nachweis der vorausgesagten sekundären Alkoholester der mittelständigen 10-Hydroxy-Gruppe der
Hauptkomponente des Cutins, der 10,16-Dihydroxyhexadacansäure (Kolattukudy 2002).
Bisher ist nicht bekannt, ob Cutin als eigenständiges Polymer-Molekül mit unbekanntem
Molekulargewicht existiert, verankert an die Zellwand an verschiedenen Stellen oder ob Cutin nur ein
hoch verzweigter Bestandteil ist (Pollard et al. 2008).
Eine Besonderheit in der Cutinstruktur stellt ein nichtabbaubarer Kern, der nach hydrolytischen
Behandlungen zurück bleibt, dar. Er besteht aus einem dreidimensionalen Netzwerk aus Polyethylen-
Molekülen mit Doppelbindungen und freien Carboxyl-Gruppen, und enthält zusätzliche
Etherbindungen (Kolattukudy 2002). Diese Struktur, bestehend aus aliphatischen Bestandteilen, wird
als Cutan bezeichnet und gewonnen durch Delipidation und Depolymerisation der Esterbindungen
der Cuticula. Cutan kann aber auch Zellwand-Kohlenhydrate und Aromaten enthalten. Dieses
Polymer besteht aus C-O- und C-C- verbundenen aliphatischen Ketten. Die Linolensäure scheint im
Vergleich zu Palmitin- oder Ölsäure das bevorzugte Substrat für die Biosynthese des Cutans zu sein.
Für das allgemeine Verständnis des Cutins ist es wichtig, auch die Struktur und Besonderheiten des
Cutans zu verstehen. Ist dieses Polymer mit Cutin verbunden oder ist es ein Bestandteil davon
(Pollard et al. 2008)?
Bei dem Biopolyester Suberin ist aufgrund der beiden vorherrschenden Domänen eine Struktur-
identifikation noch schwieriger. Eine besondere Rolle im Suberin hat das Glycerin: zum Einen dient es
als Verbindungsstruktur zwischen den Acyl-Monomeren und bewahrt somit ihre lineare Anordnung
und zum anderen ist es auch die Vernetzungsstruktur der beiden Domänen.
Die polyaromatische Schicht ist kovalent mit Kohlenhydraten der primären Zellwand verbunden
(Abbildung 6). Diese Bindung erfolgt nicht ausschließlich über Esterbindungen. Das Glycerin dient
hier ebenfalls als Verbindungsstruktur zwischen den beiden Schichten des Suberins. Die Darstellung
zeigt einen Ausschnitt einer suberinisierten Kartoffelzelle bestehend aus 2 aliphatischen Domänen
(heller Bereich) und einer aromatischen Domäne (dunkler Bereich), welche kovalent an die primäre

9 Einführung
Zellwand gebunden sind. Die hervorgehobenen aromatischen Monomere konnten mittels
verschiedener analytischer Methoden nachgewiesen werden. Die eingelagerten Wachse und extrem
langkettigen Fettsäuren in der Suberinschicht sind in der Abbildung nicht berücksichtigt (Bernards
2002).
Primäre Zellwand Suberin-Lamellen
Abbildung 6: Modell der hypotetischen Struktur der Suberinschicht der Kartoffel. P: phenolisches Monomer, C:
Kohlenhydrat, S: Suberin (aromatisch oder aliphatisch) (Bernards 2002).
Ein bedeutender Unterschied zwischen Suberin und Cutin ist, dass Suberin sich auf der inneren Seite
der primären Zellwände befindet.
Das aliphatische Monomerenprofil von Suberin ist ähnlich dem von Cutin, es gibt aber auch
Unterschiede im prozentualen Anteil und der Kettenlänge der Monomere (Tabelle 4). Abgesehen von
den Fettsäuren, ω-Hydroxy-Fettsäuren und Glycerin, besteht Suberin zusätzlich auch aus hohen
Mengen von α,ω-Dicarbonsäuren, Hydroxyzimtsäuren (besonders Ferulasäure) und Fettalkoholen.
Unterschiedlich ist auch, dass in Suberin eine große Menge an gesättigten Aliphaten gefunden
wurden mit einer Kettenlänge >C20. Es wurden ebenfalls Esterbindungen zwischen ω-Hydroxy-
Fettsäuren und zwischen ω-Hydroxy-Fettsäuren und α,ω-Dicarbonsäuren oder Ferulasäure
nachgewiesen (Pollard et al. 2008).

10 Einführung
Tabelle 4: Gegenüberstellung der prozentualen Anteile der typischen Cutin- und Suberin-Monomere. (Pollard et al. 2008).
Monomertyp Cutin Suberin Unsubstituierte Fettsäure
COOH
1-12% C16:0, C18:0, C18:1, C18:2
1-10% C18:0 bis C24:0
ω-Hydroxyfettsäuren COOHOH
1-32% C16:0, C18:1, C18:2
11-43% C18:1, C16:0 bis C26:0
α,ω-Dicarbonsäuren
COOHHOOC
Normal <5% aber >50% in Arabidopsis C16;0, C18:0, C18:1, C18:2
24-45% C18:1, C18:2, C16:0 bis C26:0
Mittelständige funktionelle Gruppen Epoxy-Fettsäuren
COOHO
Polyhydroxy-Fettsäuren
COOHOH
OH
Polyhydroxy-α,ω-Dicarbonsäuren
COOH
OH
OH
HOOC
0-34% C18:0 (9,10-epoxy), C18:1 (9,10-epoxy) 16-92% C16:0 (10,16-dihydroxy), C18:0 (9,10,18-trihydroxy) Spuren
0-30% C18:1 (9,10-epoxy-18-hydroxy), C18:0 (9,10-epoxy-1,18-disäure) 0-2% C18:0 (9,10,18-trihydroxy) 0-8% C18:0 (9,10-dihydroxy)
Fettalkohole 1-Alkanole und 1-Alkenole
OH
α,ω-Alkandiole und α,ω-Alkendiole OHOH
0-8% C16:0, C18:1 0-5% C18:1
1-6% C18:0 bis C22:0 0-3% C22:0
Glycerin
OHOH
OH
1-14%
14-26%
Phenole OHO
OH
OMe
0-1% Ferulasäure
0-10% Ferulasäure, geringere Mengen von Cumarsäure, Sinapinsäure und Kaffeesäure
Bei ersten Untersuchungen zur Aufklärung der Bindungen in Suberinpolymeren wurden lineare
Dimere der ω-Hydroxy-Fettsäuren verestert mit α,ω-Dicarbonsäuren und quervernetzt mit ω-
Hydroxy-Fettsäuren nachgewiesen. Glycerin konnte in verschiedenen Bindungsstrukturen detektiert
werden: verestert mit ω-Hydroxy-Fettsäuren oder α,ω-Dicarbonsäuren, verestert in beiden
Positionen mit α,ω-Dicarbonsäuren oder als Verbinder zwischen zwei α,ω-Dicarbonsäuren. Ebenfalls

11 Einführung
identifiziert wurde ein trimerer Diester des Glycerins verbunden mit einer ω-Hydroxy-Fettsäure und
Ferulasäure. Damit wurde nachgewiesen, dass die Ferulasäure kovalent an die aliphatische Suberin-
Domäne gebunden ist.
Schlussfolgerungen der Studien sind, dass Suberin langkettige α,ω-Dicarbonsäuren, an beiden Enden
mit Glycerin verestert, als Hauptbestandteil des Polymers enthält. Vom Glycerin aus sind 2- und 3-
dimensionale Esterbindungen zu α,ω-Dicarbonsäuren und ω-Hydroxy-Fettsäuren ausgebildet. Am
Rand des Glycerin-basierenden Polymers ist die Ferulasäure an die aliphatische Domäne als
polyaromatische Domäne gebunden und diese ist wiederum gebunden an die
Zellwandkohlenhydrate (Franke et al. 2007).
Mit 2-dimensionalen Kugel-Stab-Modellen werden mögliche Monomerverbindungen lokaler
Polyesterdomänenstrukturen dargestellt (Abbildung 7 b-d). Nur mittig stehende Hydroxyl-Gruppen
wurden in die Betrachtung einbezogen und andere mittelständige funktionelle Gruppen wurden
nicht berücksichtigt, ebenso wie mögliche Nicht-Ester-Verbindungen und Veresterungen mit den
Zellwänden. Es ist bisher nicht bekannt, ob solche klar definierten Domänen aus Hydroxyfettsäure-,
α,ω-Dicarbonsäure- und Glycerin-Monomeren gebildet werden oder ob die Strukturen vermischt
vorliegen (Pollard et al. 2008).
Abbildung 7: Darstellung der möglichen Verbindungen der Monomere der Biopolyester Cutin und Suberin. a) Hauptsächlich treten dabei primäre Esterbindungen auf (roter Pfeil), wodurch lange Ketten gebildet werden. Die sekundären Esterbindungen (blauer Pfeil) stellen zusätzliche Verzweigungspunkte dar. b) Anordnung der Fettsäure- und ω-Hydroxyfettsäure-Monomere unter Bildung eines Dendrimers. c) Anordnung der α,ω-Dicarbonsäure- (DCS) und Glycerin-Monomere unter Bildung einer Dendrimer Struktur, angeordnet um freie OH Gruppen um eine Bindung an einige Glycerin Monomere zu ermöglichen. d) Anordnung der DCS- und Glycerin-Monomere unter Bildung einer quervernetzten Domäne. (Pollard et al. 2008).
d)
c)
a)
b)
Fettsäure
ω-Hydroxyfettsäure
Substituierte ω-Hydroxyfettsäure
Dicarboxylsäure
-OH
Glycerin
-COOH
Ester

12 Einführung
In den letzten Jahren wurden ebenfalls Studien zur synthetischen Herstellung von Cutin durchgeführt
und die erhaltenen Polymere mit dem natürlichen Biopolyester verglichen. Die gebildeten Strukturen
der Hauptkomponenten stimmten sehr gut überein (Benίtez et al. 2004). Mittlerweile werden auch
die Enzyme des Cutin und Suberin Biosyntheseweges als interessante Biokatalysatoren für die
Produktion von Chemikalien und Polymeren studiert (Li et al. 2009).
1.3 Hydrolasen
1.3.1 Struktur und katalytischer Mechanismus
Die Enzymklasse der Hydrolasen ist eine große Gruppe an strukturell ähnlichen Enzymen mit
verschiedensten katalytischen Funktionen. Diese Enzyme katalysieren unter Einbau eines
Wassermoleküls die reversible Spaltung von Biomolekülen (Hydrolyse), aber auch unter Abspaltung
eines Wassermoleküls die Synthese dieser Strukturen. Die Gemeinsamkeit dieser Klasse ist die α/β
Hydrolase Faltungsstruktur, die aus 8 meist parallel angeordneten β-Faltblättern (β1-β8) und auf
beiden Seiten umgebende stabilisierende α-Helices (αA-αF) aufgebaut ist. Eine gewisse Variabilität in
der Anzahl der Strukturen ist bei den verschiedenen hydrolytischen Enzymen identifiziert worden,
allerdings ist die zentrale Anordnung und Form des aktiven Zentrums als definierte Bindungstasche
bei allen Enzymen gleich. Diese dreidimensionale Struktur der Hydrolasen bildet das Gerüst für die
enzymatische Funktion und Substratspezifität der Enzyme. Die hydrolytischen Katalysatoren sind ein
Beispiel für die divergierende Evolution von Enzymen, die aus einer Struktur durch den Austausch
von Aminosäuren oder durch die Insertion/Deletion von Schleifen über einen zeitlichen Abstand
entstehen und unterschiedliche katalytische Aufgaben erfüllen. Die Enzyme besitzen dadurch zwar
geringe Sequenzhomologien aber eine vergleichbare Raumstruktur und eine identische Anordnung
am katalytischen Zentrum. Die Aminosäuren des aktiven Zentrums sind ein Histidin, eine Säure
(Asparaginsäure oder Glutaminsäure) und ein Nucleophil (Serin, Asparagin oder Cystein) in einer
konservierten Region, welche sich über Wasserstoffbrückenbindungen untereinander stabilisieren
(Abbildung 8) (Ollis et al. 1992).

13 Einführung
Abbildung 8: α/β Hydrolase-Faltungsmotiv der Hydrolasen ist eine „doubly wound α/β superfold“ bestehend aus 8 zentral parallel angeordneten β-Faltblättern (rot) und 6 umgebenden stabilisierenden α-Helices (blau). Im Speziellen kann die Anzahl der Elemente variieren, das allgemeine Gerüst ist aber hochkonserviert. Das aktive Zentrum bilden 3 Aminosäuren (grün), das Nucleophil befindet sich im hoch konservierten Bereich zwischen der αC-Helix und dem β5-Faltblatt, dem „nucleophilic elbow“ (Ollis et al. 1992).
Die katalytische Hydrolyse am aktiven Zentrum erfolgt in mehreren Schritten. Im Grundzustand
stabilisieren sich die Aminosäuren der katalytischen Triade untereinander durch Wasserstoff-
Brückenbindungen (Abbildung 9). Bei Anwesenheit eines Esters im katalytischen Zentrum greift das
Nucleophil, in diesem Fall das Serin, den Ester unter Ausbildung eines tetraedrischen
Zwischenprodukts nucleophil an. Das frei werdende Proton des Nucleophils übernimmt der
Imidazolring des Histidin und dieser wird unterstützt durch das Carboxylatanion der Säure, in diesem
Fall der Asparaginsäure. Das gebildete, negativ geladene Intermediat wird durch mehrere
Aminosäure-Reste, welche sich in der Region des aktiven Zentrums befinden, stabilisiert. Diese
Struktur wird als Oxyanionen-Loch bezeichnet. Im nächsten Schritt erfolgt die Abspaltung des
Alkohols und die Bildung eines Enzym-Substrat Komplexes, dem Acylenzym. Durch den Eintritt eines
Wassermoleküls in das aktive Zentrum und dessen nucleophilen Angriff am Acylenzym bildet sich
wiederum ein tetraetrisches Intermediat, welches durch Wasserstoff-Brückenbindungen im
Oxyanionen-Loch stabilisiert wird. Durch die Abspaltung der Säure und ihrer Freigabe erfolgt die
Rückführung des aktiven Zentrums in seinen Ausgangszustand und steht somit direkt für die Bindung
eines weiteren Esters bereit.
NH2
β8
β7
β6
β5
β4
β3
β2
β1
αA
αB
αC
αD
αE
αF
His
Säure
Nuc
COOH

14 Einführung
Abbildung 9: Der katalytische Reaktionsmechanismus der Hydrolasen am Beispiel der Spaltung eines Esters in die dazugehörige Säure und Alkohol. Die Reaktion erfolgt ohne Verwendung eines Cofaktors und ist somit sehr einfach und effizient. Die Bildung des Acylenzyms ist ein schneller Reaktionsschritt, die Freisetzung der Säure und des Enzyms ist der geschwindigkeitsbestimmende Reaktionsschritt. (Holmquist 2000).
Eine weitere Besonderheit der Hydrolasen ist das hochkonservierte Pentapeptid G-X-S-X-G, in das das
katalytische Serin eingebettet ist. Bei den beiden flankierenden Aminosäuren handelt es sich um
Glycin, für X können verschiedene Aminosäuren stehen.
Die Enzymklasse der Hydrolasen beinhaltet unter Anderem die Carboxylesterasen (EC 3.1.1.1,
Esterase), die Triacylglycerol Lipasen (EC 3.1.1.3, Lipase) und die Cutin Hydrolasen (EC 3.1.1.74,
Cutinase). Sie hydrolysieren alle Ester-Verbindungen, variieren aber bezüglich ihres
Substratspektrums. Die Carboxylesterase, auch unspezifische Esterase, spaltet unter Einbau eine
Wassermoleküls Carboxylester in seine Bestandteile Alkohol und Carboxylsäure. Die Lipasen finden
Anwendung bei dem hydrolytischen Abbau von Fetten (Triacylglyceride) in Glycerol und Fettsäuren.
Die Cutinasen zeichnen sich durch die Besonderheit der katalytischen Spaltung des Biopolymers
Cutin in dessen Monomere (u.a. Fettsäuren, Hydroxyfettsäuren, aliphatische Alkohole, aromatische
Säuren) aus. Alle dargestellten Reaktionen laufen im wässrigen Milieu ab.
Säure O
O
NHN
His Ser
OH
Säure O
OH
NNH
His Ser
O
N
H
O
N
H
O
RO
O
R´
Säure O
OH
NNH
His Ser
O
N
H
O
N
H
O
R
O
R´ O
Säure O
OH
NNH
His Ser
O
O
R
N
H
O
N
H
O
Säure O
OH
NNH
His Ser
O
N
H
O
N
H
O
R
O
H O
RO
O
R´
ROH
O
Freies Enzym
R´OH
OH2
tetrahedraler Übergangszustand I
tetrahedraler Übergangszustand II
Acyl-Enzym-Komplex

15 Einführung
Abbildung 10: Übersicht über die katalysierten hydrolytischen Spaltungen der Enzymgruppen der Carboxylesterase, Triacylglycerol Hydrolasen und der Cutin Hydrolasen im wässrigen Milieu.
Die Carboxylesterase zeichnet sich durch die Hydrolyse von kurz- und mittelkettigen, wasserlöslichen
Estern aus und ihre Reaktionskinetik kann durch die Michaelis-Menten-Gleichung beschrieben
werden (Abbildung 11). Im Gegensatz dazu hydrolysieren die Lipasen bevorzugt langkettige
Triglyceride, welche meist wasserunlöslich sind. Die Reaktionskinetik der Lipasen zeichnet sich durch
einen sigmoiden Verlauf aus, da diese Enzyme eine „Grenzflächenaktivierung“ aufweisen
(Abbildung 11). Das bedeutet, dass bei geringen Substratkonzentrationen die Lipase inaktiv bleibt
bzw. nur geringe Aktivitäten detektiert werden. Sobald aber die Substratkonzentration die
spezifische Konzentration (kritische micellare Konzentration) erreicht, ab der eine Agglomeration des
Substrats erfolgt, bildet sich eine Grenzfläche zwischen den hydrophoben Substratmicellen und der
hydrophilen Wasserphase mit der Lipase aus. Ab dieser Substratmenge steigt die hydrolytische
Aktivität der Lipasen rapide an.
OR2
O
R1 OH
O
R1
O
O
O
O
O
O
R3
R2
R1 OH
OH
OH+ H2O
+ R1COOH
+ R2COOH
+ R3COOH
+ H2O + R2OH
+ (H2O)n
COOH
OH COOH
COOHOH
OH
x
y
COOH
OH COOH
COOHOOH
OH COOH
OH
OH
Carboxylesterase
Triacylglycerol Hydrolase
Cutin Hydrolase
n

16 Einführung
Abbildung 11: Reaktionskinetik der Carboxylesterasen (links) nach Michaelis-Menten und der sigmoide Kurvenverlauf der Reaktionskinetik der Lipasen (rechts) (Pütz 2006).
Der Grund für dieses Verhalten lässt sich anhand der Struktur der Lipasen erklären. Das aktive
Zentrum des Enzyms wird durch eine bewegliche Proteinschlaufe, eine α-Helix, abgedeckt. Dieser
Deckel (engl.: lid) ist der Grund für die geringen Enzymaktivitäten unterhalb der kritischen mizellaren
Konzentration. Sobald diese Proteinschlaufe mit der Grenzfläche der Substratmizelle in Berührung
kommt, erfolgt eine Konformationsänderung und der Deckel gibt das aktive Zentrum frei (Abbildung
12). Gleichzeitig wird dadurch die hydrophobe Oberfläche, die das aktive Zentrum umgibt, frei
zugänglich und unterstützt die Interaktion zwischen dem Enzym und dem hydrophoben Substrat.
In neueren Studien wurden weitere Lipasen identifiziert und biochemisch charakterisiert, die keine
Grenzflächenaktivierung zeigten. Die Lipase LipA aus B. subtilis besitzt zum Beispiel keinen Deckel
und die Lipase aus P. aeruginosa LipA besitzt zwar einen Deckel, zeigt allerdings keine
Grenzflächenaktivierung. Somit ist die Anwesenheit oder Abwesenheit des Deckels kein struktureller
Hinweis zur Unterscheidung zwischen den Carboxylesterasen und Lipasen (Hausmann und Jaeger
2010).
1.3.2 Carboxylesterasen
Die Carboxylesterasen werden aufgrund ihrer primären Struktur und der biochemischen
Eigenschaften in 7 Gruppen (II-VIII) unterteilt (Hausmann und Jaeger 2010). Die meisten Esterasen
gehören zu der α/β-Hydrolase-Faltung Superfamilie. Ausnahmen enthält die Gruppe VIII, die
Esterasen mit einem β-Lactamase Faltungsmotiv beinhaltet. Im Jahre 2009 waren 696 bakterielle
Esterasen in der Enzymdatenbank BRENDA (Schomburg et al. 2004) enthalten und viele von ihnen
zeigen keine signifikante Übereinstimmung mit einer der bisher bekannten Esterasefamilien. Einige
Vertreter der einzelnen Gruppen sowie einige ihrer Merkmale sind in Tabelle 5 gegenübergestellt.

17 Einführung
Bakterielle Esterasen können im Cytoplasma (Droege et al. 2005), im Periplasma (Weadge et al.
2005) oder in der Membran vorhanden sein (von Tigerstrom und Stelmaschuk 1989, Wilhelm et al.
1999) oder sekretiert werden (von Tigerstrom und Stelmaschuk 1989, Xiang et al. 2006). Eine
spezifische Funktion der Esterase ist bisher nicht bekannt, sie sind unter normalen Standard-
Laborbedingungen nicht überlebenswichtig für die Bakterien (Berger et al. 1998). Sie sind involviert
in Prozessen, die eine C-Quelle zugänglich machen, wie die pathogene Infektion, der
Pflanzenzellwandabbau oder die Entgiftung (Khalameyzer et al. 1999). Es kommt zum Beispiel zur
Esteraseproduktion beim probiotischen Lactobacillus reuteri bei Anwesenheit von Gallensaft und es
wird vermutet, dass die Esterase bei der Erkennung von Zellmembranen beteiligt ist (Wall et al. 2007,
Whitehead et al. 2008). Andere Esterasen werden durch die Anwesenheit von Wasserstoffperoxid
gebildet und helfen dem Bakterium bei der Bewältigung von oxidativem Stress (Baatout et al. 2006).
Esterasen sind ebenfalls eingebunden in die Regulation der Kommunikation und Interaktion zwischen
den Spezies, sie bauen Signalmoleküle ab (Shinohara et al. 2007, Khalameyzer et al. 1999) oder
hydrolysieren Triacylglyceride als Ausgangsstoffe für die Antibiotika-Synthese (Olukoshi und Packter
1994).
Esterasen sind aufgrund ihrer Fähigkeit, Esterbindungen zwischen der Hydroxyzimtsäure und Zuckern
zu hydrolysieren, wichtige Enzyme beim Pflanzenzellwandabbau durch Clostridium und Lactobacillus
(Fonaghy et al. 2000, Prates et al. 2001, Wang et al. 2004, Garcia-Conesa et al. 1999). Die einzige
identifizierte Esterase im humanpathogenen Helicobacter pylori (Ruiz et al. 2007) ist direkt beteiligt
an der Bildung von Magen- und Zwölffingerdarmgeschwüren und Magenkrebs (Kuster et al. 2006)
durch den Abbau der Fette des Magenschleims (Slomiany et al. 1989). Das oft in Krankenhäusern
übertragene pathogene Corynebacterium jeikeium besitzt keine eigene Fettsäure-Synthase und
spaltet deshalb mit einer Cholesterin-Esterase das Cholesterin im Blut und an extrazellulären
Matrixproteinen (Hansmeier et al. 2007, Tauch et al. 2005). Es ist Verursacher von Blutvergiftungen
und Herzinnenhautentzündungen. Die LipF aus dem pathogenen Mycobacterium tuberculosis,
gehörend zur Gruppe IV der Esterasen, ist ebenfalls ein wichtiger Faktor bei der Infektion des
Menschen durch das Bakterium. Als Autotransporter ist die Esterase EstA aus
Pseudomonas aeruginosa ebenfalls an der Infektion beteiligt (Camacho et al. 1999, Zhang et al.
2005).

18 Einführung
Tabelle 5: Übersicht lipolytische Enzyme I, Esterasen (Gruppe II-VIII) mit ausgewählten Beispielen (Hausmann und Jaeger 2010).
Gruppe Herkunft der Esterase (Proteinnr.) Merkmale
II
(GDSL)
Aeromonas hydrophila (P10480)
Pseudomonas aeruginosa
(O33407)
Bacillus cereus (Q81AY4)
Xanthomonas vesicatoria
(Q7X4K7)
Enthält die GDSL/SGNH-Esterasen, besitzen kein
GXSXG-Motiv am aktiven Zentrum, dafür ein GDS(L)-
Tetrapeptid im N-terminalen Bereich; Esterasen
enthalten fünf konservierte Sequenzbereiche, GDS(L)
mit Ser Block I, Gly in Block II, Asp in Block III und His in
Block V
III Streptomyces exfoliatus (Q56008)
Kineococcus radiotolerans
(A6WEQ4)
Clavibacter michiganensis
(B0RCM8, B0RFW0)
Thermobifida fusca (Q47RJ6,
Q47RJ7)
Extrazelluläre Enzyme, molekulare Masse von 32-35
kDa, besitzen Ähnlichkeit mit dem menschlichen
Blutblättchen-Aktivierungsfaktor Acetylhydrolase
(PAF-AH)
In dieser Gruppe sind Lipasen (definitionsgemäß) ohne
Deckel enthalten z.B. Tfu_0882 und Tfu_0883 aus T.
fusca
IV
(HSL)
Alicyclobacillus acidocaldarius
(Q7SIG1)
Burkholderia sp. 383 (u.a.
Q39FH4)
Pseudomonas sp. B11-1 (O52270)
Bacillus subtilis (O68884)
Ähnlich zur Gruppe der Hormonsensitiven
Säugetierlipasen, besitzen 3 konservierte
Sequenzblöcke, Bock II und III mit den katalytischen
AS, Block I mit HGGG Consensus-Sequenz, bevorzugen
kurzkettige Substrate und zeigen Esterase-
Enzymkinetik, besitzen am aktiven Zentrum eine „cap“
aus 2 separaten helikalen Regionen, spezifische
Inhibition
V Moraxella sp. (P24640)
Salmonella typhimurium (Q8ZNL5)
Serratia proteamaculans (A8GC20)
Rhodobacter sphaeroides
(A4WX93, A3PMP7)
Enzyme zeigen auch Homologie zu anderen
Enzymgruppen mit dem typischen α/β-Faltungsmotiv
(Epoxidhydrolase, Dehalogenase, Haloperoxidase),
besitzen 3 konservierte Sequenzblöcke, katalytisches
Ser in Bock II, Asp und His in Block III, Esterase-Kinetik,
kurzkettige Substrate bevorzugt
VI Spirulina platensis (Q53415)
Anabaena variabilis (Q3M6G8)
Shewanella amazonensis (A1S771)
Rhodoferax ferrireducens
(Q21XU9)
Kleine Enzyme mit 23-26 kDa, bisher nur eine mit 31,6
kDa, 40% Sequenzähnlichkeit zu eukaryotischen
Lysophospholipasen, 3 konservierte Sequenzblöcke
analog zu Gruppe IV und V, bevorzugt kurzkettige
Substrate

19 Einführung
Fortführung Tabelle 5
Gruppe Herkunft der Esterase (Proteinnr.) Merkmale
VII Arthrobacter oxydans (Q01470)
Streptomyces coelicolor (Q9Z545)
Burkholderia sp. 383 (Q398P3)
Bacillus pumilus (Q66M67)
Molekulare Masse von ca. 55 kDa, hohe Ähnlichkeit zu
Acetylcholinesterase und Darm/Leber
Carboxylesterase, 4 konservierte Sequenzblöcke,
hydrolysieren kurzkettige p-NP Ester und Triglyceride,
aber auch biotechnologisch interessante Substrate
VIII Arthrobacter aurescens (A1RB78)
Saccharopolyspora erythraea
(A4F8E6)
Pseudomonas gingeri (B0M0H4)
Streptomyces anulatus (O87861)
Unterscheiden sich in der Struktur deutlich von den
anderen Gruppen, zeigen Ähnlichkeiten zu β-
Lactamasen und DD-Peptidasen, Größe ca. 42 kDa,
Multimerbildung kann auftreten, hohe
Lösungsmitteltoleranz, katalytisch aktives Serin
befindet sich in SXXK Motiv, Ser und Lys im N-
terminalen Block und Tyr bilden aktives Zentrum,
zusätzlich wurde eine GXSXG Sequenz am C-Terminus
identifiziert
1.3.3 Lipasen
Überraschenderweise ist die genaue Funktion der Lipase bei Bakterien noch nicht bekannt, da ihre
Produktion einer komplexen Regulation unterliegt und sie somit wahrscheinlich nicht nur für die
Hydrolyse von Fetten als Energiequelle verantwortlich sind (Hausmann und Jaeger 2010). Die Art der
Regulation lässt die Schlussfolgerung zu, dass die Lipasen auch als Virulenzfaktoren auftreten
(Degrassi et al. 2008, Devescovi et al 2007, Heurlier et al. 2004, Lewenza et al. 1999, Ulrich et al.
2004) oder ähnlich den Xylan-Esterasen durch die Hydrolyse von Xylan die Schutzbarriere der
Wirtszelle zerstören (Khalameyzer et al. 1999).
Der die Atemwege infizierende, opportunistisch pathogene Pseudomonas aeruginosa produziert eine
Lipase, welche zusammen mit der Phospholipase C Substanzen abbaut, die zur Reduktion der
Oberflächenspannung an den Lungenbläschenmembranen und somit zur Unterstützung der Atmung
wichtig sind (Beatty et al. 2005, Schmidt et al. 2007). Isolierte Stämme von P. aeruginosa und
Salmonella zeigten untypische Zusammensetzungen der Lipid A Schicht, was sie für die mikrobiellen
Abwehrmechanismen des Menschen nicht erkennbar machten und es somit zu häufigeren
Erkrankungen kam (Ernst et al. 1999, Guo et al. 1998, Tanamoto und Azumi 2000). Die normale Lipid

20 Einführung
A Schicht besteht aus Capron-, Laurin-, Myristin-, Palmitin- und Stearinsäuren, bei den Isolaten wurde
allerdings eine Palmitinsäure-Konzentration von bis zu 33% gefunden (Ernst et al. 1999).
Ebenso können freie Fettsäuren die normalen Abläufe, wie die Hinderung der Lymphozyten-
Proliferation und die Chemotaxis von neutrophilen Granulozyten, im menschlichen Organismus
stören (Buttke 1984, Buttke und Cuchens 1984, Hawley und Gordon 1976, Jaeger et al. 1991,
Nordstrom et al. 1991, Pourbohloul et al. 1985).
Die Anwesenheit einer Lipase bei Propionibacterium acnes unterstützt die Ansiedlung des
Organismus in den Talgdrüsenfollikeln der Haut. Durch die Hydrolyse von Glyceriden aus dem Serum
werden Fettsäuren freigesetzt, die die Ansiedlung des Bakteriums unterstützen (Burkhart und
Burkhart 2003, Gribbon et al. 1993).
Eine weitere wichtige Funktion der Lipasen ist der Abbau von lipolytischen Substraten zur
Verstoffwechselung. P. aeruginosa YS-7 kann mit Ölgemischen mit einem Wassergehalt von nur 1%
zum Wachsen gebracht werden (Shabtai et al. 1991). Microthrix parvicella kann Lipide und
Fettsäuren in hohen Konzentrationen in Abwässern abbauen, sowohl aerob als auch anaerob
(Nielsen et al. 2002).
Die Produktion von Lipasen kann durch die Anwesenheit von Alkanen angeregt werden, allerdings ist
der Abbau dieser Substrate noch nicht vollständig aufgeklärt. Selbst wenn die Lipase durch die Alkane
induziert wurde, kann sie nicht immer ohne weitere enzymatische Mithilfe die Alkane abbauen
(Breuil et al. 1978). Nur sehr wenige Stämme können diese Energiequelle verwerten, Hefestämme
noch eher als Bakterienstämme (Margesin et al. 2003).
Abbildung 12: Struktur der offenen und geschlossenen Lipase von Burkholderia cepacia BcL. Die offene Kristallstruktur ist blau dargestellt und das geschlossene Homologie-Modell grün, die gelbe Kugel stellt ein Ca
2+-Ion dar. Die katalytische
Triade besteht aus Serin, Histidin und Asparaginsäure (Trodler et al. 2009).

21 Einführung
Tabelle 6: Übersicht lipolytische Enzyme II, Lipasen (Grup. I.1-I.8) mit ausgewählten Beispielen (Hausmann und Jaeger 2010).
Gruppe Herkunft der Esterase (Proteinnr.) Merkmale
I.1 Pseudomonas mendocina
(Q8RKT7)
Rhodoferax ferrireducens
(Q21T36)
Vibrio cholerae (u.a. P15493)
Chromobacterium violaceum
(Q7NUI4)
Starke Ähnlichkeit zu P. aeruginosa Lipase, molekulare
Masse 30-32 kDa, Typ II Sekretionsweg, werden ATP-
abhängig ins Periplasma transportiert, in die aktive
Konformation umgefaltet und in den extrazellulären
Raum sekretiert, einige benötigen Faltungsproteine, 2
Asp binden ein Ca2+-Ion, das für die katalytisch aktive
Struktur benötigt wird, katalytisches His
I.2 Burkholderia glumae (Q05489)
Pseudomonas KWI-56 (P25275)
Burkholderia cepacia (u.a. P22088)
Pseudomonas luteola (O68551)
Starke Ähnlichkeit zu B. glumae Lipase, eine etwas
größere molekulare Masse als I.1 aufgrund 2 zusätzlicher
β-Faltblätter, Typ II Sekretionsweg, werden ATP-abhängig
ins Periplasma transportiert, in die aktive Konformation
umgefaltet und in den extrazellulären Raum sekretiert,
einige benötigen Faltungsproteine, 2 Asp binden ein Ca2+-
Ion, das für die katalytisch aktive Struktur benötigt wird,
katalytisches His
I.3 Pseudomonas fluorescens PfO1
(Q3KCS9)
Pseudomonas sp. 7323 (Q2KTB3)
Serratia marcescens (u.a. Q09KJ5)
Uncultered bacterium (A7J993)
Molekulare Masse 50-65 kDa, Typ I Sekretionsweg, Ca2+-
Ion enthalten für katalytische Funktion, näher verwandt
mit eukaryotischen Enzymen als mit bakteriellen, das C-
terminale Sekretionssignal mit Glycin-reichen Wieder-
holungen (GGXGXDX(U)X)n kann an das Ca2+-Ion binden,
welches als Chaperon bei hohen Kalzium-
Konzentrationen agiert
I.4 Bacillus subtilis (P37957)
Bacillus sp. NK13 (B0LW76)
Bacillus megaterium (Q8RJP5)
Bacillus clausii (Q5WDN0)
Molekulare Masse 20 kDa, kleinste Lipasen, häufig aus
Bacillus sp., enthalten konserviertes AXSXG Pentapeptid,
maximale Enzymaktivität bei pH 10,0-11,5, Lipase ist
Kalzium-unabhängig und enthält kein Cystein, weisen
eine kompakte minimale Hydrolasefaltung auf ohne
Deckel und Ca2+-Ionen
I.5 Geobacillus zalihae (Q842J9)
Bacillus sp. L2 (Q51413)
Geobacillus sp. SF1 (Q1L776)
Bacillus stearothermophilus
(u.a. A0MTM1)
Bacillus Lipasen zeigen nur 15% Ähnlichkeit mit Gruppe
I.4, pH Optimum variiert (7,5-9,5), Molekulargewichte ca.
46 kDa, wahrscheinlich aufgrund von Insertionen, die
eine Zink-Bindungsstelle enthalten, weisen deshalb
höhere thermale Stabilität auf

22 Einführung
Fortführung Tabelle 6
Gruppe Herkunft der Esterase (Proteinnr.) Merkmale
I.6 Staphylococcus hyicus (P04635)
Staphylococcus xylosus (Q2TPV1)
Staphylococcus warneri (Q5DWE2)
Staphylococcus aureus (u.a.
P10335)
Ca. 75 kDa großes Präproprotein mit ca. 200 AS am N-
terminalen Ende für die effiziente Translokation und
mit möglicher Funktion als intramolekulare Chaperone,
aktives Enzym ca. 46 kDa groß mit breiter
Substratspezifität, pH Stabilität von pH 4 bis 9, einige
Lipasen zeigen Aktivitätssteigerung mit Ca2+-Ionen und
Hemmung durch EDTA, können Polymere bilden,
können als Virulenzfaktoren agieren
I.7 Streptomyces cinnamoneus
(O33969)
Propionibacterium acnes
(u.a. Q59644)
Corynebacterium glutamicum
(u.a. Q8NU60)
Zeigen Ähnlichkeit zu Lipasen der Gruppe I.2 in der
Region von AS 50-150, Lipase S. cinnamoneus 29,2 kDa
und P. acnes 36,4 kDa, Lipase aus P. acnes besitzt
weites Substratspektrum Triacylglyceride und p-NP
Ester C2-C16
I.8 Pseudoalteromonas haloplanktis
(Q3IF07)
Hahella chejuensis (Q2SGZ8)
Colwellia psychrerythraea
(Q48AN1)
Pseudoalteromonas tunicata
(A4CF12)
Neue Gruppe, identifiziert und charakterisiert anhand
Lip1 aus P. haloplanktis, 51 kDa, gebunden an äußere
Bakterienmembran, höhere Aktivität mit langkettigen
Estern, Sequenz zeigt keine nähere Übereinstimmung
mit anderen lipolytischen Enzymen, eher zu marinen
psychrophilen Stämmen, kein Deckel, keine Ca2+-Ionen
Bindungsstelle, neues Motiv LGG(F/L/Y)STG mit Ser
anstatt GXSXG
1.3.4 Cutinasen
Cutinasen sind die einzigen Enzyme, die das Biopolyester Cutin und das Modellsubstrat Poly-ε-
caprolacton abbauen können. Sie zeigen einige Eigenschaften von Lipasen (Hydrolyse unlöslicher
Substrate) und Esterasen (Hydrolyse von löslichen Substraten). Diese Enzyme werden hauptsächlich
von pflanzenpathogenen Pilzen gebildet, es sind aber auch bereits einige Bakterien mit
Cutinaseaktivität identifiziert worden. Sie sind die kleinsten Hydrolasen in der α/β-Hydrolase Familie
(Dutta et al. 2009). Die Enzymklasse der Cutinasen ist eine bisher noch umstrittene Gruppe bei den
Hydrolasen. Es gibt einige bekannte und charakterisierte Cutinasen, z.B. die Cutinase aus

23 Einführung
Fusarium solani, Thermomyces insolens, Pseudomonas mendocina, Aspergillus oryzae und
Thermobifida fusca. Bei der Einteilung der lipolytischen Enzyme wurden die Cutinasen nicht als
selbstständige Gruppe deklariert, sondern es erfolgte z. B. aufgrund der Sequenz die Zuordnung der
Cutinasen aus T. fusca zur Gruppe III der Esterasen (Hausmann und Jaeger 2010). Bei einem
Sequenzalignment zeigten diese Cutinasen Ähnlichkeiten zu Triacylglycerol Lipasen aus
Streptomyces albus G und Streptomyces sp. M1 (Kleeberg et al. 2005). Es sollten weitere
Untersuchungen erfolgen, um die Zuordnung der Cutinasen bei den lipolytischen Enzymen noch
genauer zu versehen.
1.4 Polyethylenterephthalat
Synthetische Polymere und die daraus hergestellten Produkte sind aus unserem heutigen Leben nicht
mehr wegzudenken. Auch in der Textilindustrie haben die künstlichen Fasern eine große Bedeutung.
Speziell Polyesterfasern erlangten in den letzten Jahrzehnten immer mehr Einfluss. Die Produktion
dieser Fasern ist in den vergangenen 50 Jahren konstant angestiegen. Im Jahr 2007 waren bereits
76% aller synthetischen Fasern Polyesterfasern. Bei einer jährlichen Produktion von 30,7 Millionen
Tonnen waren davon 12,4 Millionen Tonnen Stapelfasern und 18,3 Millionen Tonnen Filamentgarn
(Saurer Management AG 2008). Für 2011 wird eine weltweite Produktion von 36 Millionen Tonnen
prognostiziert (Chemical Market Associates Inc. 2010). Die PET-Fasern werden weit verbreitet
genutzt, sowohl einzeln als auch in Mischungen mit anderen Fasern. Einige ihrer vielen Vorteile, die
sie für die Textilindustrie so unverzichtbar machen, sind ihre starke Reißfestigkeit, ihre Beständigkeit
und die Widerstandsfähigkeit gegenüber einer Vielzahl von Chemikalien. Doch besitzen
Polyesterfasern auch einige ungünstige Eigenschaften, wie zum Beispiel ihre starke Hydrophobie,
eine daraus resultierende geringe Wasseradsorption und sie neigen zum Pilling.
Polyethylenterephthalatfasern (PET), die bedeutendsten Polyesterfasern, werden durch
Polykondensation aus Terephthalsäure und Ethylenglycol hergestellt (Abbildung 13).
Abbildung 13: Polykondensationsreaktion von Terephthalsäure und Ethylenglycol zu Polyethylenterephthalat.
COOH
COOH
Terephthalsäure
+
OH
OH
Ethylenglycol
C
O
OR
C
O
O
R
PET
n

24 Einführung
Doch dabei entstehen ebenfalls einige unerwünschte Nebenprodukte, die die negativen
Eigenschaften der Polyesterfasern noch verstärken. Dabei handelt es sich um lineare und zyklische
Oligomere der Ausgangsstoffe. Besonders die Anreicherung von zyklischen Trimeren (Cyclo-tris-
Ethylenterephthalat, CTR), welche extrem wasserunlöslich sind, hat die Bildung von Agglomeraten
auf den Fasern und eine daraus resultierende Störung bei der Weiterverarbeitung (u.a. beim Färben)
zur Folge. Ebenfalls zeigen diese zyklischen Trimere eine Tendenz, sich an metallischen Oberflächen
anzulagern und haben somit auch negative Auswirkungen auf die Färbe- und
Veredelungsmaschinerie. Bisher angewandte Methoden zur Entfernung der zyklischen Trimere, wie
zum Beispiel der Abbau unter hohen Temperaturen, mit NaOH und mittels kationischer Tenside,
haben ebenfalls sehr viele negative Auswirkungen auf die Fasern und die Maschinen. Zum Einen
vermindert die aggressive Behandlung die Qualität der Polyesterfasern und zum anderen wird ein
hoher Gewichtsverlust beobachtet. Weiterhin kommt es neben dem hohen Energieeinsatz durch die
Anwendung der Chemikalien zu einer Anhäufung umweltschädigender Abfälle und zu Störungen an
den Maschinen (Valk et al. 1970; Senner 1973; Hempel 1979; Grosse et al. 1994; Höhn 2003).
Abbildung 14: Struktur eines zyklischen PET-Trimers.
1.4.1 Konventionelle PET-Faserbehandlung
Die bei der Polykondensation entstehenden niedermolekularen Nebenprodukte sind vielfältig und
können sowohl zyklische als auch lineare Verbindungen mit einem Polymerisationsgrad von 2 bis 10
Einheiten sein, wobei nicht alle die weitere Bearbeitung der Textilfaser im gleichen Maße
beeinflussen. Die wichtigsten Problemverursacher sind die zyklischen Trimere und im geringen
Ausmaß die Tetramere, Pentamere und Hexamere (Valk et al. 1970, Senner 1973, Hempel 1979,
Grosse et al. 1994, Höhn 2003). Zyklische und lineare Oligomere mit höheren Polymerisationsgraden
entstehen nur in kleinen Mengen. In Abhängigkeit vom Faserherstellungsprozess beträgt die Menge
O
O
O
O
O
O
O O
O
O O
O
Zyklischer PET-Trimer

25 Einführung
an an der Faser lokalisierten Oligomer zwischen 1 bis 3% des Fasergewichtes. Während des Prozesses
der Hochtemperatur-Färbung erfolgt ein Austritt von bis zu 70% der PET-Oligomer aus der Faser.
Durch weitere Färbe- und Abkühlungsprozesse bilden sich Oligomerablagerungen auf den
Färbemaschinen, welche mit jeder Färbung zunehmen. Die überwiegende Menge der als
Ablagerungen vorliegenden Oligomere (ca. 50-80%) bilden eine grobdisperse Aufschwemmung, die
von den PET-Produkten mittels Filtration abgetrennt werden kann. Diese so gewonnenen Oligomere
werden auch als Oberflächenoligomere bezeichnet und bestehen zu 97 bis 99% aus zyklischen
Trimeren. Sie verursachen eine Vielzahl von negativen Beeinträchtigungen bei der
Weiterverarbeitung der PET-Ware, eine Verringerung der Qualität des Produktes als auch technische
Probleme, wie unter anderem eine Leistungsverringerung der Maschinen, einen höheren
Energieverbrauch und längere Stillstandzeiten. Um diese Probleme auf ein Minimum zu reduzieren,
wurden in der Vergangenheit bereits einige Verfahren entwickelt (Dugal et al. 1973, Kusch 1974,
Baker 1974, Werkes 1975, Keller et al. 1981, Yang und Li 2000, Höhn 2003).
alkalische Vorbehandlung bei hohen Temperaturen
Die Nachteile dabei sind erhöhte Kosten durch den zeit- und energieaufwendigen
Arbeitsschritt, ein Einsatz von umweltschädlichen Chemikalien ebenso wie eine Beschädigung
der Textilfasern.
Vorbehandlung zur PET-Oligomerentfernung mit chlorierten (Methylenchlorid,
Perchlorethylen) oder chlorfreiem Lösungsmittel (Dioxan)
Dies bietet sich aus umwelttoxikologischen und sicherheitstechnischen Gründen nicht als
Alternative an.
Einsatz von Dispergatoren (Tenside) zur Verhinderung der Ablagerung von austretenden
Oligomer
Diese Methode führt zu einem höheren Gehalt an organischen Verbindungen im Abwasser.
alkalisches Färben
Positiv ist dabei eine gleichzeitige Verseifung der Oligomere, diese Methode eignet sich aber
nicht für alle Textilfärbungen.
Nachwäsche als Lösemittelwäsche mit Tetrachlorethen oder Dispergatorwäsche
Auch dieses Verfahren führt zu einem höheren Gehalt an organischen Verbindungen in den
Abwässern.
Nachreinigung als alkalische Verseifung mit Tetraalkylammoniumverbindungen als
Katalysator
Auch hier treffen die bereits oben genannten Nachteile zu.

26 Einführung
Diese Verfahren und auch alle nach dem Oligomermobilisierenden Färbeprozess angewandten und
beschriebenen Methoden gegen die PET-Oligomerdeposition weisen ähnliche wirtschaftliche,
technische und ökologische Nachteile, wie ein hoher zusätzlicher Energieeinsatz, die Verwendung
von produkt-, umwelt- und personenschädigenden Chemikalien und besonders deren Entsorgung,
auf.
Sowohl aus diesen Gründen, aber auch aufgrund des drastischen Gewichtsverlusts der PET-Fasern
und der Verschlechterung ihrer Qualität während der Bearbeitung (Zeronian und Collins 1989, Shukla
et al. 1997), wurde in den letzten Jahren zunehmend nach einem biotechnologischen Prozess zur
Oligomerbearbeitung und -entfernung geforscht, um die traditionellen Faserverarbeitungs-
methoden mit ihren Nachteilen zu ersetzen (Gübitz und Cavaco-Paulo 2001).
1.4.2 Charakterisierung von PET
Bei der PET-Herstellung erfolgen anschließend verschiedenste Modifikationsschritte, welche sich auf
die Eigenschaften des PET-Textiles oder Films auswirken. Während des Schmelzspinnverfahrens
(Abbildung 14) wird die Spinnmasse geschmolzen und anschließend durch eine beheizte Spinndüse
mit einer Geschwindigkeit von 4000 m/min gepresst. Diese Spinndüsen weisen eine Größe zwischen
1-10 cm auf und bestehen aus bis zu 60000 Spinndüsenlöchern mit einem Durchmesser zwischen
0,05 und 1 mm (Abbildung 15). Durch die anschließende Luftkühlung erstarren die Fibrillen und es
erfolgt das Verstrecken, was eine Neuanordnung der PET-Ketten in der Fibrille zur Folge hat.
Abbildung 15: Schematische Darstellung des Schmelzspinnverfahrens von PET-Fibrillen (links) und Darstellung einer
beheizten Schmelzspinndüse (Jahreiß 2005).

27 Einführung
Das Verstrecken erfolgt entweder unter sehr hohen Geschwindigkeiten beim Abziehen von der
Schmelzspinndüse oder indem das gesponnene Filament vor dem Aufrollen über mehrere
verschieden große Rollen geführt wird. Beim Verstrecken werden die zuvor wirr durcheinander
liegenden PET-Ketten in Faserrichtung und auch zueinander ausgerichtet (Abbildung 16). Dadurch
bilden sich auch Wechselwirkungen zwischen den Ketten aus, wie die Wasserstoffbrückenbindungen
(Abbildung 17), und es entstehen teilkristalline Bereiche, welche die Festigkeit der Fasern deutlich
erhöhen. Diese Bindungen sind zu diesem Zeitpunkt noch nicht stabil und würden beim Waschen
zerstört werden, deshalb erfolgt anschließend eine Hitzefixierung.
Abbildung 16: Anordnung der PET-Ketten vor (links) und nach (rechts) dem Verstrecken unter Ausbildung von geordneten kristallinen Bereichen (Jahreiß 2005).
Abbildung 17: Darstellung der Wasserstoffbrückenbindungen zwischen den durch Verstrecken entstandenen geordneten PET-Ketten (grün).
Verstrecken
O
O
O
O
O
O
O
O
O
O
O
O
H
H
H
HH
H
H
H
H
H
H
H
O
O
O
O
O
O
O
O
O
O
O
O
H
H
HH
H
H
HH
H
H
HH
O
O
O
O
O
O
O
O
O
O
O
O
H
H
H
HH
H
H
H
H
H
H
H
O
O
O
O
O
O
O
O
O
O
O
O
H
H
HH
H
H
HH
H
H
HH

28 Einführung
Die Eigenschaften von PET-Fasern, wie Glanz, Transparenz, Farbeindruck (dunkler/heller),
Schmutzsichtbarkeit, Griff (weicher/härter), eine Volumenerhöhung und Wärmeisolierung, werden
ebenfalls durch ihren Faserquerschnitt bzw. der Faseroberfläche beeinflusst (Abbildung 18). Eine
weitere Modifikation zur Optimierung der Volumenzunahme, der elastischen Dehnung, besserer
Wärmeisolierung (durch Lufteinschlüsse), des flauschigen Griffs und zur besseren
Feuchtigkeitsaufnahme für die Herstellung von elastischen Bekleidungswaren ist die Texturierung.
Dabei werden die zuvor glatten Synthesefasern mechanisch gekräuselt und somit liegen nun
Faserschlingen zwischen der Haut und dem Kleidungsstück, was zur Verbesserung der
Trageeigenschaften führt (Abbildung 18). Weiterhin hat die Modifikation eine höheren
Feuchtigkeitsaufnahme und einen erhöhten Lufteinschluss zur Folge, hingegen wird Glanz und Pilling
vermindert (Jahreiß 2005).
Abbildung 18: Darstellung möglicher Faserquerschnitte und Oberflächen (links) und die PET-Fasern vor und nach dem Texturieren (rechts) (Jahreiß 2005).
Die Textilveredelung, ein Hauptgebiet der Textilchemie, umfasst sämtliche Arbeitsprozesse, die die
äußeren Eigenschaften der Textilien verschönern, wie Färben, Bleichen, Bedrucken, sowie
Maßnahmen, die ihren Gebrauchswert steigern (Pflegeleichtausrüstung, Knitterfestmachen), sowie
Imprägnierung und Silikonisieren (weicher und geschmeidiger, durch Minimierung der Reibung der
Fasern untereinander). Diese Maßnahmen werden im unversponnenen Zustand der Textilfaser („in
der Flocke“) durchgeführt. Dazu zählt ebenfalls das Finish oder auch Ausrüstung genannt, die letzte
Bearbeitung von Textilien. Es dient zur Verschönerung, Hervorhebung oder Unterdrückung
bestimmter Eigenschaften. Dazu zählen unter anderem Waschen, Bleichen, Färben, Bedrucken,
Prägen, Imprägnieren und Hydrophobieren (d.h. wasserabweisend machen, z.B. durch
Aluminiumsalze oder Paraffinprodukte), knitterfest machen, Mattieren (z.B. durch Einschlüsse von
Titandioxid in die Faser), antimikrobiell machen (durch Zusätze von z.B. Bisphenolen oder

29 Einführung
Hexachlorophen; wodurch es zu weniger Hautpilzen und Körpergeruch kommt) und
schwerentflammbar machen (z.B. durch Borax oder Titankomplexe) (Jahreiß 2005).
PET besteht aus kristallinen und amorphen Bereichen und weist somit sowohl eine
Glasübergangstemperatur (für die amorphen Bereiche) als auch eine Schmelztemperatur (für die
kristallinen Bereiche) auf. Die Glasübergangstemperatur (Tg) ist die Temperatur, bei der ganz oder
teilweise amorphe Polymere vom glasigen oder hartelastischen, spröden Zustand in den
gummielastischen, flexiblen Zustand übergehen. Tg wird daher auch "Erweichungstemperatur"
genannt und ist für jeden Kunststoff spezifisch (Abbildung 19). Da Wärme nichts anderes als
kinetische Energie ist, bewegen sich unterhalb der Glasübergangstemperatur die Molekülketten
kaum. Bei der Erwärmung des Kunststoffes bewegen sie sich immer mehr, halten aber noch
zusammen, bis schließlich mit dem Erreichen der Glasübergangstemperatur sich längere Abschnitte
der Molekülketten frei bewegen können. Die bei Temperaturen oberhalb der
Glasumwandlungstemperatur (ca. 80°C) beweglichen Molekülketten kristallisieren bei ca. 160°C aus
(Abbildung 19, Kristallisationspeak), bevor bei ca. 255°C der endotherm ablaufende Schmelzvorgang
beginnt (Abbildung 19, Schmelzpeak).
PET kann mit Hilfe der Differentialrasterkalorimetrie (DSC) bezüglich der
Glasumwandlungstemperatur, dem Schmelzverhalten, der Kristallinität und des
Kristallisationsverhaltens charakterisiert werden (Abbildung 19). Dabei erfolgt mit der Probe eine
lineare Erhitzung und anhand der Enthalpieänderungen bezüglich einer Referenz kann ein wie in
Abbildung 19 dargestelltes Diagramm erstellt werden. Durch die Integration des Schmelzpeaks kann
die Kristallinität der vorliegenden PET Probe berechnet werden.
Abbildung 19: DSC-Kurve einer PET-Folie (http://web.utk.edu/~mse/Textiles/Polymer%20Crystallinity.htm).
T g Kristallistionspeak
Schmelzpeak

30 Einführung
Die Glasübergangstemperatur hängt im Wesentlichen von der Struktur der Polymere ab. Die
Flexibilität der Hauptkette ist einer der wichtigsten Faktoren. Ein Kunststoff hat eine niedrige
Glasübergangstemperatur, wenn sich die Molekülketten bei geringen Temperaturen schon von sich
aus gut bewegen können. Das bedeutet, dass Polymere, die aus aliphatischen Bausteinen aufgebaut
sind, eine geringere Glasübergangstemperatur besitzen als Polymere, welche aus aromatischen
Monomeren, wie der Terephthalsäure, zusammengesetzt sind. Bei PET handelt es sich um ein
Polymer, das sowohl aliphatische als auch aromatische Bestandteile besitzt und eine
Glasübergangstemperatur von ca. 80°C.
1.4.3 Biofunktionalisierung von PET
In den letzten Jahrzehnten erfolgten verschiedenste Studien über die Biofunktionalisierung von
Polyethylenterephthalat, eine Übersicht darüber gibt Zimmermann und Billig (2011). Zunächst wurde
noch angenommen, dass aromatische Polyester, also auch PET, resistent gegenüber einem
hydrolytischen Abbau sind (Kint und Munoz-Guerra 1999, Müller et al. 2001). Ferner erfolgten
Veröffentlichungen von verschiedenen Autoren, die keine enzymatischen Effekte auf den PET-Fasern
nachweisen konnten (Tokiwa und Suzuki 1977, Levefre et al. 1999). Weitere Untersuchungen
hingegen dokumentieren die Möglichkeit der biokatalytischen Modifikation der PET-Fasern.
Veränderungen in den physikalischen Eigenschaften wie Reißfestigkeit, Viskosität und
Dehnungsverhalten von PET-Film und Fasern wurde nach der Inkubation mit einer Enzym-
Präparation aus Cladosporium cladosporioides festgestellt (Sato 1983). Ferner wurde von Oda et al.
(1999) beschrieben, dass die Inkubation mit Arthrobacter und Trichosporon Spezies ebenfalls
Modifikationen der PET-Fasern zur Folge hat. Ebenfalls konnte eine Verbesserung der
Wasserbenetzbarkeit und der Absorptionseigenschaften sowie eine Verbesserung der Anfärbbarkeit
von PET-Textilien und Fasern nach einer enzymatischen Hydrolyse mit bakteriellen Cutinasen aus
Pseudomonas mendocina (Hsieh und Cram 1998, Hsieh et al. 2003, Bott et al. 2003, Kellis et al. 2001,
Yoon et al. 2002, Dyson et al. 2005), aus Thermobifida fusca (Alisch et al. 2004, Alisch-Mark et al.
2006, Heumann et al. 2006, Feuerhack et al. 2008, Eberl et al. 2008, Eberl et al. 2009, Brueckner
et al. 2008) und mit eukaryotischen Cutinasen aus Fusarium solani (O´Neill und Cavaco-Paulo 2004,
Silva et al. 2005, Alisch-Mark et al. 2006, Heumann et al. 2006, Araujo et al. 2007, O´Neill et al. 2007,
Nimchua et al. 2007, Nimchua et al. 2008) und Thermomyces insolens (Abo et al. 2004, Svendsen
et al. 2005, McCloskey und Jump 2005, Liu et al. 2008) nachgewiesen werden (Tabelle 7). Die Enzyme
bewirken dabei durch die partielle Hydrolyse von Esterbindungen und die dadurch entstehenden
Hydroxyl- und Carboxylgruppen eine Zunahme der hydrophilen Eigenschaften auf der Oberfläche der
PET-Fasern. Mit Hilfe einer solchen Behandlung erhält man bei Polyestertextilien verbesserte

31 Einführung
Eigenschaften, wie zum Beispiel größere Bindung von kationischen Farbstoffen, einen Depilling
Effekt, die Entfernung von Polyesterschlichte und eine bessere Entfernbarkeit von Ölflecken. In
weiteren Untersuchungen wurden Textilien und Fasern mit eukaryotischen Lipasen aus
Aspergillus sp. (Fischer-Colbrie et al. 2004, Heumann et al. 2006, Wang et al. 2008), aus
Thermomyces lanuginosus (Almansa et al. 2008, Brueckner et al. 2008, Eberl et al. 2008, Eberl et al.
2009), aus Candida antarctica (Anderson et al. 1999, Khoddami et al. 2001, Lee und Song 2010, Kim
und Song 2006), mit der bakteriellen Lipase aus Burkholderia cepacia (Heumann et al. 2006, Kim und
Song 2006) und einer tierischen Lipase (Kim und Song 2006, Kim und Song 2008, Hsieh und Cram
1998) behandelt und dabei eine Erhöhung der Hydrophilie erzielt. Weiterhin konnte eine
Verbesserung der Eigenschaften der PET-Fasern durch die Inkubation mit Esterasen und Proteasen
bewiesen werden (Kontkanen et al. 2006, Zhang et al. 2004, Zhang et al. 2006, Liebminger et al.
2007, Kim und Song 2010).
Abbildung 20: Darstellung der chemischen Struktur der Polyethylenterephthalat Polymerkette, der zyklischen Trimere und der enzymatisch freigesetzten Hydrolyseprodukte (Zimmermann und Billig 2011).
Neben den PET-Textilien wurden in den letzten Jahren auch Studien zur Modifikation und zum
Bioabbau von PET-Folien, welche unter anderem in der Verpackungsindustrie Anwendung finden,
durchgeführt. Dabei erfolgte mit den bereits benannten Cutinasen aus F. solani (Vertommen et al.
2005, Ronkvist et al. 2009), P. mendocina (Ronkvist et al. 2009), T. fusca (Müller et al. 2005, Eberl
et al. 2008, Eberl et al. 2009) und T. insolens (Abo et al. 2004, Ronkvist et al. 2009) die partielle
Hydrolyse der Folien unter verschiedenen Bedingungen (Tabelle 5). Ebenfalls waren die Lipase aus
T. lanuginosus (Almansa et al. 2008, Brueckner et al. 2008, Eberl et al. 2008, Eberl et al. 2009) und die
Esterase aus C. cladosporioides (Sato 1983) in der Lage, den PET-Film zu modifizieren. Diese
hydrophilen PET-Filme werden ferner für medizinische Anwendung genutzt.

32 Einführung
Abbildung 21: Darstellung der chemischen Struktur der Polytrimethylenterephthalat Polymerkette, der zyklischen Dimere und der enzymatisch freigesetzten Hydrolyseprodukte (Zimmermann und Billig 2011).
Der direkte Abbau von zyklischen PET-Trimeren erfolgte bereits mit den Cutinasen aus T. fusca (Chen
et al. 2010) und T. insolens (Andersen et al. 1999, Riegels et al. 2001, Hooker et al. 2003, Abo et al.
2004, Jump und McCloskey 2004, Svendsen et al. 2005, Figueroa et al. 2006). Für Screening-Studien,
die Optimierung der Hydrolysebedingungen und kinetische Studien wurden in den letzten Jahren
ebenfalls diverse kurzkettige Modellsubstrate eingesetzt (Abbildung 20-22) (Andersen et al. 1999,
Kellis et al. 2001, Riegels et al. 2001, Hooker et al. 2003, Jump et al. 2004, Abo et al. 2004, Zhang
et al. 2004, Dyson et al. 2005, Svendsen et al. 2005, Figueroa et al. 2006, Heumann et al. 2006,
Michels et al. 2006, Pütz 2006, Zhang et al. 2006, Liebminger et al. 2007, Eberl et al. 2008, Wang
et al. 2008, Eberl et al. 2009, Chen et al. 2010).
Nach der Behandlung der PET-Textilien oder -Filme kommen verschiedenste analytische Methoden
zur Charakterisierung der modifizierten Oberfläche zur Anwendung. Dabei wird zwischen der
direkten und der indirekten Oberflächenanalytik unterschieden.
Abbildung 22: Darstellung der chemischen Struktur der angewandten Polyethylenterephthalat Modellsubstrate (Zimmermann und Billig 2011).

33 Einführung
Tab
elle
7:
PET
Bio
fun
ktio
nal
isie
run
g u
nd
an
alys
iert
e Ef
fekt
e (
Zim
mer
man
n u
nd
Bill
ig 2
01
1).
Lite
ratu
r
O´N
eill
un
d C
avac
o-P
aulo
20
04
, Silv
a et
al.
20
05
, Alis
ch-M
ark
et a
l. 2
00
6, H
eum
ann
et
al.
20
06
, Ara
ujo
et
al.
20
07
, O´N
eill
et a
l. 2
00
7,
Nim
chu
a et
al.
20
07
, Nim
chu
a et
al.
20
08
Ver
tom
men
et
al.
20
05
, Ro
nkv
ist
et a
l. 2
00
9
Heu
man
n e
t a
l. 2
00
6, E
ber
l et
al.
20
09
Hsi
eh u
nd
Cra
m 1
99
8, H
sieh
et
al.
20
03
, Bo
tt
et a
l. 2
00
3, K
elli
s et
al.
20
01
, Yo
on
et
al.
20
02
, D
yso
n e
t a
l. 2
00
5
Ke
llis
et a
l. 2
00
1, D
yso
n e
t a
l. 2
00
5
Ro
nkv
ist
et a
l. 2
00
9
Alis
ch e
t a
l. 2
00
4, A
lisch
-Mar
k et
al.
20
06
, H
eum
ann
et
al.
20
06
, Feu
erh
ack
et a
l. 2
00
8,
Eber
l et
al.
20
08
, Eb
erl e
t a
l. 2
00
9, B
ruec
kner
et
al.
20
08
Mü
ller
et a
l. 2
00
5, E
ber
l et
al.
20
08
, Eb
erl e
t a
l.
20
09
Heu
man
n e
t a
l. 2
00
6, E
ber
l et
al.
20
08
, Eb
erl
et a
l. 2
00
9, C
hen
et
al.
20
10
Effe
kt b
ei P
ET
Hyd
roly
se, e
rhö
hte
Hyd
rop
hili
e,
erh
öh
te A
nfä
rbb
arke
it
Hyd
roly
se, A
bb
au, G
ewic
hts
verl
ust
, Ä
nd
eru
ng
der
O
ber
fläc
hen
eige
nsc
haf
ten
Hyd
roly
se
erh
öh
te H
ydro
ph
ilie,
Gew
ich
tsve
rlu
st,
erh
öh
te A
nfä
rbb
arke
it, Ä
nd
eru
ng
der
O
ber
fläc
hen
eige
nsc
haf
ten
Hyd
roly
se
Hyd
roly
se, Ä
nd
eru
ng
der
O
ber
fläc
hen
eige
nsc
haf
ten
, G
ewic
hts
verl
ust
Hyd
roly
se, e
rhö
hte
Hyd
rop
hili
e,
erh
öh
te A
nfä
rbb
arke
it,
Gew
ich
tsve
rlu
st
Gew
ich
tsve
rlu
st, e
rhö
hte
Hyd
rop
hili
e
Hyd
roly
se
Sub
stra
t
Text
ilien
Film
PET
-Tri
mer
Text
ilien
, Fas
ern
DET
Film
Text
ilien
, Fas
ern
Film
(P
ET; P
TT)
CTR
, BET
EB, B
HP
T,
zykl
isch
e P
TT
Dim
er, B
HET
, PET
Tr
imer
Vo
rko
mm
en
Fusa
riu
m s
ola
ni
Pse
ud
om
on
as
men
do
cin
a
Ther
mo
bif
ida
fu
sca
Enzy
m
Cu
tin
ase

34 Einführung
Fo
rtfü
hru
ng
Tab
elle
7
Lite
ratu
r
Ab
o e
t a
l. 2
00
4, S
ven
dse
n e
t a
l. 2
00
5,
McC
losk
ey u
nd
Ju
mp
20
05
, Liu
et
al.
20
08
An
der
sen
et
al.
19
99
, Rie
gels
et
al.
20
01
, H
oo
ker
et a
l. 2
00
3, A
bo
et
al.
20
04
, Ju
mp
un
d
McC
losk
ey 2
00
4, S
ven
dse
n e
t a
l. 2
00
5, F
igu
ero
a
et a
l. 2
00
6
Ab
o e
t a
l. 2
00
4, R
on
kvis
t et
al.
20
09
Kim
un
d S
on
g 2
00
6, K
im u
nd
So
ng
20
08
, Hsi
eh
un
d C
ram
19
98
Heu
man
n e
t a
l. 2
00
6, K
im u
nd
So
ng
20
06
An
der
son
et
al.
19
99
, Kh
od
dam
i et
al.
20
01
, Lee
un
d S
on
g 2
01
0, K
im u
nd
So
ng
20
06
Alm
ansa
et
al.
20
08
, Bru
eckn
er e
t a
l. 2
00
8,
Eber
l et
al.
20
08
, Eb
erl e
t a
l. 2
00
9
Eber
l et
al.
20
08
, Eb
erl e
t a
l. 2
00
9
Fisc
her
-Co
lbri
e et
al.
20
04
, Heu
man
n e
t a
l. 2
00
6
Effe
kt b
ei P
ET
Hyd
roly
se, e
rhö
hte
Hyd
rop
hili
e,
erh
öh
te A
nfä
rbb
arke
it,
Gew
ich
tsve
rlu
st
Hyd
roly
se
Hyd
roly
se, Ä
nd
eru
ng
der
O
ber
fläc
hen
eige
nsc
haf
ten
, G
ewic
hts
verl
ust
erh
öh
te H
ydro
ph
ilie,
erh
öh
te
An
färb
bar
keit
, Än
der
un
g d
er
Ob
erfl
äch
enei
gen
sch
afte
n
Hyd
roly
se, e
rhö
hte
Hyd
rop
hili
e
Hyd
roly
se, e
rhö
hte
Hyd
rop
hili
e,
Än
der
un
g d
er
Ob
erfl
äch
enei
gen
sch
afte
n
Hyd
roly
se, e
rhö
hte
Hyd
rop
hili
e
Hyd
roly
se
Hyd
roly
se, e
rhö
hte
Hyd
rop
hili
e
Sub
stra
t
Text
ilien
, Fas
ern
DET
, BEB
, PET
Tr
imer
, CTR
Film
Text
ilien
Text
ilien
, PET
Tr
imer
Text
ilien
, Fas
ern
BEB
, DET
Text
ilien
, Fas
ern
, Fi
lm (
PET
; PTT
)
BH
PT,
BH
ET, P
ET
Trim
er
Text
ilien
, PET
Trim
er
Vo
rko
mm
en
Ther
mo
myc
es in
sole
ns
Sch
we
ine
pan
krea
s
Bu
rkh
old
eria
cep
aci
a
Ca
nd
ida
an
tarc
tica
Ther
mo
myc
es la
nu
gin
osu
s
Asp
erg
illu
s sp
.
Enzy
m
Lip
ase

35 Einführung
Fo
rtfü
hru
ng
Tab
elle
7
Lite
ratu
r
Wan
g et
al.
20
08
Pü
tz 2
00
6, M
ich
els
et a
l. 2
00
6
Sato
et
al.
19
83
Oe
ser
et a
l. 2
01
0, B
illig
et
al. 2
01
0
Ko
ntk
anen
et
al.
20
06
Zhan
g et
al.
20
04
, Zh
ang
et a
l.
20
06
Lieb
min
ger
et a
l. 2
00
7
Kim
un
d S
on
g 2
01
0
Effe
kt b
ei P
ET
Hyd
roly
se, e
rhö
hte
Hyd
rop
hili
e,
erh
öh
te A
nfä
rbb
arke
it,
Gew
ich
tsve
rlu
st
Hyd
roly
se
Hyd
roly
se, Ä
nd
eru
ng
der
p
hys
ikal
isch
en E
igen
sch
afte
n
Hyd
roly
se
erh
öh
te H
ydro
ph
ilie,
erh
öh
te
An
färb
bar
keit
Hyd
roly
se, Ä
nd
eru
ng
der
Fase
rob
erfl
äch
en
Hyd
roly
se, e
rhö
hte
Hyd
rop
hili
e
Hyd
roly
se, e
rhö
hte
Hyd
rop
hili
e
Sub
stra
t
Text
ilien
, DP
PET
Dim
er, P
ET
Nan
op
arti
kle,
D
ialk
ylp
hth
alat
e
Film
, Fas
ern
CTR
, PET
Nan
op
arti
kle
Text
ilien
Fase
rn, D
ET
Text
ilien
, PET
Tr
imer
Text
ilien
Vo
rko
mm
en
Asp
erg
illu
s or
yza
e
Ba
cillu
s su
bti
lis,
Ba
cillu
s lic
hen
ifo
rmis
Cla
do
spo
riu
m c
lad
osp
ori
oid
es
Ther
mo
bif
ida
fu
sca
Mel
an
carp
us
alb
om
yces
Sch
we
inel
eber
Pen
icill
ium
cit
rin
um
Pap
ain
Enzy
m
Este
rase
Pro
teas
e

36 Einführung
Zur direkten Oberflächenanalytik zählen unter anderem XPS (Röntgenphotoelektronen-
spektroskopie), bekannt auch als ESCA (Elektronenspektroskopie für chemische Analyse), FTIR
(Fourier-Transformations-Infrarotspektrometrie), SEM (Rasterelektronenmikroskopie) bzw. ESEM
(Umgebungsrasterelektronenmikroskopie) und AFM (Atomkraftmikroskopie). Während der XPS
Analyse wird die elementare Zusammensetzung der PET Oberfläche in einem Bereich von 10 nm
detektiert und die spezifischen funktionellen Gruppen nachgewiesen. Enzymatische Modifikationen
durch Hydrolasen wurden bereits mehrfach bestätigt (Vertommen et al. 2005, Feuerhack et al. 2008,
Brueckner et al. 2008, Eberl et al. 2009), allerdings ist eine vorherige Reinigung der PET-Filme
notwendig, um verfälschende Proteinverunreinigungen zunächst zu entfernen (Vertommen et al.
2005, Feuerhack et al. 2008). Mit der FTIR Analyse werden ebenfalls chemische Veränderungen auf
der PET-Oberfläche nachgewiesen. Mit Hilfe infraroter Strahlung werden die Moleküle in ein
energetisch höheres Niveau versetzt und die bei der Rückkehr in den nicht angeregten Zustand
freiwerdende Strahlung wird analysiert und gibt Rückschlüsse über die Zusammensetzung der Probe
(Goddard und Hotchkiss 2007). Änderungen in der chemischen Struktur von PET-Filmen nach einer
enzymatischen Behandlung wurden mittels FTIR bereits von Donelli et al. 2009 nachgewiesen. Das
SEM nutzt einen Elektronenstrahl, um die Oberfläche abzutasten und somit ein dreidimensionales
Abbild zu erschaffen. Damit können Veränderungen wie Ablagerungen oder Löcher nachgewiesen
werden. Fasermodifikationen wurden bereits bestätigt durch eine Schweineleber Esterase (Zhang
et al. 2004, Zhang et al. 2006), Lipasen von C. antarctica und aus Schweineleber (Lee und Song 2010,
Kim und Song 2006) und Cutinasen aus P. mendocina und T. insolens (Kellis et al. 2001, Yoon et al.
2004). Die Cutinase aus T. insolens bildete während der Enzymbehandlung von PET-Filmen viele
Löcher und somit eine rauere Oberfläche (Ronkvist et al. 2009). Nach der Inkubation von PTT-
(Polytrimethylenterephthalat) Fasern mit der Cutinase aus T. fusca zeigte sich hingegen keine
Oberflächenveränderung per ESEM (Eberl et al. 2008, Brueckner et al. 2008). Die Abtastung der
Oberfläche beim AFM erfolgt mit einer dünnen Nadel, welche einen Laserstrahl aussendet. Der von
der Oberfläche reflektierte Laserstrahl wird von einem Photodetektor aufgenommen und in eine
Oberflächenaufnahme umgewandelt. Bisher wurde per AFM die Oberflächenveränderung von PET-
Fasern nach der Behandlung mit der Cutinase aus T. fusca nachgewiesen (Feuerhack et al. 2008).
Bei der indirekten Oberflächenanalyse wird die Modifikation mit Hilfe weiterer Agenzien detektiert.
Die gesteigerte Hydrophilie der Textilien wird mittels Kontaktwinkelmessung, Steighöhen-
bestimmung, Tropfentest, Flüssigkeitsaufnahmekapazität, Feuchtigkeitsaufnahme und Färbetests mit
anschließender Farbintensitätsmessung nachgewiesen. Für diese Analysemethoden ist es ebenfalls
wichtig, dass das noch verbliebene Enzym an der Oberfläche vor dem Nachweis entfernt wird, da
sonst verfälsche Ergebnisse auftreten (Vertommen et al. 2005, Brueckner et al. 2008, Donelli et al.
2009). Bei der Kontaktwinkelmessung wird der auftretende Winkel eines Wassertropfens auf einer

37 Einführung
festen Oberfläche bestimmt. Je kleiner der Winkel ist, desto hydrophiler ist die Oberfläche (ca. 0-
90°), ab 90° gilt der Feststoff als hydrophob und ab ca. 160° als superhydrophob, was auch als
Lotuseffekt bekannt ist. Enzymatisch behandelte PET-Oberflächen wurden unter anderem von Hsieh
und Yu 1992, Hsieh 1995, Yoon et al. 2002, Heumann et al. 2006, Donelli et al. 2009 und Lee und
Song 2010 mit dieser Methode analysiert. Bei der Steighöhenbestimmung wird die Wasseraufnahme
eines Textilstreifens, der mit einem Ende in eine Flüssigkeit getaucht ist, nach definierten
Zeitabständen in Zentimetern bestimmt. Eine hohe Steighöhe geht einher mit einer hydrophilen
Oberfläche des PET-Textils (Fischer-Colbrie et al. 2004, Alisch-Mark et al. 2006, Müller 2006). Eine
ähnliche Methode zum Nachweis der Hydrophilie ist der Tropfentest, bei dem der Zeitabstand von
der Aufbringung eines definierten Tropfens Flüssigkeit auf ein PET-Textil bis zur vollständigen
Aufnahme des Tropfen in das Textil gemessen wird (Heumann et al. 2006, Brueckner et al. 2008, Liu
et al. 2008). Das Verhältnis an Flüssigkeit, das ein trockenes PET-Textil maximal aufnehmen kann,
zum Textil wird als Flüssigkeitsaufnahmekapazität bezeichnet. Die Flüssigkeitsaufnahme hingegen ist
die Menge an Wasser, welche eine trockene Faser oder Textil aus der Luft mit definierter
Feuchtigkeit absorbiert kann. Beide Methoden verdeutlichen Veränderungen in den
Absorptionseigenschaften von modifizierten PET-Textilien (Hsieh et al. 1996, Hsieh und Cram 1998,
Kim und Song 2006, Lee und Song 2010). Aufgrund der eingefügten hydrophilen funktionellen
Gruppen werden die Textilien verstärkt durch kationische oder reaktive Farbstoffe angefärbt. Die
Brillianz der Farbe kann durch eine Farbintensitätsmessung bestimmt werden und somit indirekt die
Hydrophilie nachweisen (Yoon et al. 2001, Alisch et al. 2004, O´Neill und Cavaco-Paulo 2004,
Brueckner et al. 2008, Eberl et al. 2009).
Die enzymatische Hydrolyse kann nicht nur durch die Modifikationen auf der Oberfläche
nachgewiesen werden, sondern auch freigesetzte Hydrolyseprodukte können mit verschiedensten
Methoden analysiert werden. Da diese wasserlöslichen Produkte Derivate der Terephthalsäure bzw.
Terephthalsäure direkt sind, können sie einfach im ultravioletten Wellenlängenbereich von 240-
255 nm spektrophotometrisch detektiert werden (Kellis et al. 2001, Alisch et al. 2004, O´Neill und
Cavaco-Paulo 2004). Da enthaltene Proteine die UV Messung beeinflussen, dient dieser Nachweis nur
zum Screening nach PET Aktivität. Terephthalsäure kann ebenfalls mittels Dünnschich-
chromatographie nachgewiesen werden (Fischer-Colbrie et al. 2004, Pütz 2006, Liebminger et al.
2007). Die Bestimmung der Konzentrationen der verschiedenen Hydrolyseprodukte; u. a. TPA, MHET
und BHET; freigesetzt aus PET-Textilien, Filmen, zyklischen PET-Trimeren und weiteren
Modellsubstraten kann per RP-HPLC (Umkehrphasen-Hochleistungsflüssigkeitschromatographie)
erfolgen (Riegels et al. 2001, Hooker et al. 2003, Fischer-Colbrie et al. 2004, Vertommen et al. 2005,
Figueroa et al. 2006, Heumann et al. 2006, Eberl et al. 2008, Eberl et al. 2009, Ronkvist et al. 2009,
Chen et al. 2010). Terephthalsäure einzeln kann auch mittels Fluoreszenz-Spektroskopie detektiert

38 Einführung
werden, dazu erfolgt zunächst die Bildung von Hydroxyterephthalsäure (HTA), welche bei 329 nm
angeregt und bei 425 nm die freigesetzte Emissionsstrahlung gemessen wird (O´Neill und Cavaco-
Paulo 2004, Nimchua et al. 2007). Die freigesetzte TPA kann ebenfalls mittels Titration mit NaOH
nach der enzymatischen Hydrolyse nachgewiesen werden (Kim und Song 2008, Kim und Song 2010).
Für kinetische Studien zur Charakterisierung verschiedener PET-Hydrolasen erfolgte auch schon die
Anwendung eines pH-Staten um die Säurefreisetzung über einen längeren Zeitraum zu detektieren
(Ronkvist et al. 2009).
Bei den Untersuchungen mit den verschiedenen PET-Hydrolasen wurde eine Korrelation der
enzymatischen Abbaurate mit dem Temperaturunterschied zwischen der Glasübergangstemperatur
Tg (amorphe Bereiche) bzw. der Schmelztemperatur Tm (kristalline Bereiche) und der
Hydrolysetemperatur identifiziert. Da die Polymerketten in der Nähe dieser charakteristischen
Temperaturen deutlich mobiler sind, können sie schneller erreicht und hydrolysiert werden durch die
PET-Hydrolasen. Deshalb sollten die PET Hydrolysen nahe der Tg von amorphen PET erfolgen (Witt
et al. 1997, Marten et al. 2003, Marten et al. 2005).
1.4.4 PET-Hydrolasen
In den letzten Jahren wurden verschiedene Enzymgruppen für eine enzymatische Hydrolyse von PET
Substraten eingesetzt, mit unterschiedlichen Abbauraten als Folge. Die am häufigsten verwendeten
Enzyme gehören zur Gruppe der Cutin Hydrolasen (EC 3.1.1.74.), bekannt auch als Cutinasen
(Tabelle 5).
Die am häufigsten eingesetzten und somit am besten charakterisierten PET-Cutinasen sind die
rekombinant produzierten Wildformen oder bereits optimierten Mutanten isoliert aus den
Bakterienstämmen Pseudomonas mendocina (zuvor Pseudomonas putita), Thermobifida fusca und
den eukaryotischen Stämmen Fusarium solani und Thermomyces insolens. Bei den drei zuerst
genannten Enzymen handelt es sich definitiv um Cutinasen, da diese Enzyme nachgewiesene
hydrolytische Aktivität gegenüber dem Biopolyester Cutin zeigten (Sebastian et al. 1987, Kleeberg
et al. 2005, Purdy und Kolattukudy 1973, Purdy und Kolattukudy 1975). Die Cutinase aus T. insolens
wird von der ersten literarischen Erwähnung an als Cutinase bezeichnet, einen eindeutigen Nachweis
anhand von Daten sind die Autoren aber bisher schuldig geblieben (Sandal et al.1995, Andersen et al.
1999, Riegels et al. 2001, Hooker et al. 2003, Abo et al. 2004, Svendsen et al. 2005, Figueroa et al.
2006, Ronkvist et al. 2009). Trotz dieses fehlenden Nachweises wird für die nachfolgende Darstellung
in Übereinstimmung mit der Literatur weiterhin die Hydrolase als Cutinase bezeichnet.

39 Einführung
Tabelle 8: Vergleich der PET-Hydrolasen von F. solani (Carvalho et al. 1998, Carvalho et al. 1999, Ronkvist et al. 2009), T. insolens (Sandal et al. 1996, Ronkvist et al. 2009), P. mendocina (Bott et al. 2003, Ronkvist et al. 2009) und T. fusca (Kleeberg et al. 2005, Chen S et al. 2008).
F. solani T. insolens P. mendocina T. fusca
Anzahl der Aminosäuren 197 194 272 261
Molekulargewicht 22 kDa 20-21 kDa 30 kDa 28 kDa
Isoelektrischer Punkt 7,2 8 nb 6,43
pH Optimum 6,5-8,5 8,5-9,5 9,0-9,5 6,5-8,0
Temperatur Optimum 50°C 75-80°C 50°C 60-70°C
Katalytische Triade Ser120
Asp175
His188
Ser140
Asp195
His208
Ser126
Asp176
His206
Ser132
Asp176
His210
GXSXG Motiv G-Y-S-Q-G G-Y-S-Q-G G-H-S-Q-G G-H-S-M-G
α-Helices / β-Faltblätter 5 / 5 Nb 7 / 9 Nb
nb: nicht bestimmt
Die wichtigsten Informationen über die vier Cutinasen sind in Tabelle 8 zusammengefasst. Es ist
deutlich ersichtlich, dass alle Enzyme eine ähnlich kleine Größe zwischen 20 und 30 kDa aufweisen,
ein neutrales bis leicht alkalischen pH Optimum und ein thermophiles Temperatur Optimum
besitzen. Die katalytische Triade besteht bei allen vier Cutinasen aus den Aminosäuren (AS) Serin,
Asparaginsäure und Histidin, natürlich an verschiedenen Positionen im Protein, aber dennoch in
einem Abstand von maximal 20-22 AS. Das spezifische Hydrolase-Motiv ist bei den beiden
eukaryotischen Cutinasen identisch, bei den bakteriellen Cutinasen unterscheidet es sich nur in einer
flankierenden Aminosäure.
Die Cutinase aus F. solani ist das am besten untersuchte Enzym für den PET-Abbau. Es wurden bereits
mehrere Reviews veröffentlicht, die eine Übersicht über die wichtigsten katalytischen und
strukturellen Eigenschaften der Cutinase beinhalten. Die angewandten Methoden für die
Cutinaseinduktion, die rekombinante Produktion und Aufreinigung wurden in dem Übersichtsartikel
von Carvalho et al. 1999 beschrieben. Die F. solani Cutinase ist aufgebaut aus 197 Aminosäuren,
besitzt ein Molekulargewicht von 22 kDa und vier nichtvariable Cysteine bilden zwei Disulfidbrücken.
Die Cutinase gehört zur Klasse der Serin-Esterasen und zeigt die typische α/β-Hydrolase Faltung. Die
katalytische Triade ist zusammengesetzt aus Ser120, Asp175 und His188. Mit einer ortsspezifischen
Mutagenese wurden Studien über die funktionellen Interaktionen mit verschiedenen Substraten und
zur Identifikation der Struktur und Bindungen am aktiven Zentrum durchgeführt. Es sind viele Daten
über die thermale Stabilität der Cutinase in eingekapselten Präparationen und in kovalent

40 Einführung
gebundenen oder absorbierten Strukturen beschrieben worden. Hingegen gibt es bisher keine
Informationen über die thermale Stabilität des frei gelösten Enzyms (Carvalho et al. 1999).
Eine Cutinase Präparation von Genencore International, Inc. (GCI 2002/1410) wurde zum ersten Mal
für die Hydrolyse von Polyester eingesetzt. Freigesetzte Hydrolyseprodukte von PET-Textilien wurden
durch die Fluoreszenzdetektion von Hydroxyterephthalat nachgewiesen, ebenso die Steigerung der
Hydrophilie durch die Anfärbung und Farbintensitätsbestimmung (Andre und Charmoille 1999,
O´Neill und Cavaco-Paulo 2004). 2005 wurde eine granulierte Cutinase von Unilever Research and
Development Laboratory, Vlaardingen, gelöst in Puffer, eingesetzt und war in der Lage aromatische
Hydrolyseprodukte vom einem amorphen PET-Film (APET; 4,1% Kristallinität) und von PET Granulat
(GPET; 13,7% Kristallinität) während einer 5-tägigen Inkubation bei 30°C freizusetzen, detektiert
durch RP-HPLC. Die Cutinase setzte mehr Hydrolyseprodukte vom geringer kristallinen APET und nur
Spuren an Produkten vom höher kristallinen biaxial orientierten PET (OPET; 48,2% Kristallinität) frei.
Die Hydrolyseprodukte wurden identifiziert als TPA (Rt: 11,0 min), MHET (Rt: 12,3 min) und BHET (Rt:
12,9 min). Das Haupthydrolyseprodukt war MHET, aber das Verhältnis von MHET zu TPA war
abhängig vom Verhältnis von Enzym zum Substrat. Zusätzlich wurden höher molekulare
Hydrolyseprodukte während der Inkubation freigesetzt, allerding konnten die Produkte zu diesem
Zeitpunkt nicht strukturell identifiziert werden. Veränderungen auf der Polyesteroberfläche wurde
detektiert mittels einer ESCA-Analyse und zeigten Modifikationen auf dem PET-Film (Vertommen
et al. 2005).
Aus der Kultivierung eines neuen Fusarium sp. wurde eine Rohenzymlösung gewonnen, die
hydrolytische Aktivität gegenüber PET-Fasern bei 30°C besaß und freigesetzte aromatische Produkte
wurden mittels Fluoreszenz-und UV-Absorbtionsmessung nachgewiesen (Nimchua et al. 2007,
Nimchua et al. 2008).
Die genetisch modifizierte F. solani Cutinase, erhalten durch die Substitutionen L81A und L182A,
zeigte eine gesteigerte Polyesteraktivität gegenüber PET-Textilien. Die mit Hilfe von ortsspezifischen
Mutationen eingeführten Strukturen am aktiven Zentrum unterstützten die Stabilisierung des
Enzyms und erlauben eine bessere Einbettung des Substrates (Araújo et al. 2007). Mit der L182A
Substitutionsmutante und dem Wildtyp Enzym erfolgten mehrere Studien über den Einfluss der
Durchmischung auf die hydrolytische Effizienz. Die Inkubation der PET-Textilien erfolgte bei 37°C und
die Hydrolyseprodukte wurden mittels Fluoreszenzdetektion nachgewiesen. Die Detektion der
Modifikationen an den Textilien erfolgte durch eine Anfärbung mit Reaktivfarbstoffen und der
Bestimmung der Farbintensität. In der Studie wurde gezeigt, dass eine geringere Durchmischung
höhere Funktionalisierungsraten zu Folge hatte (O´Neill et al. 2007).
Intensivere Studien zur enzymatischen Oberflächenmodifikation und Funktionalisierung wurden mit
TEXAZYM EM durchgeführt (InoTEX Ltd., Tschechische Republik). Die Enzympräparation zeigte eine

41 Einführung
intensive Proteinbande in der SDS-PAGE bei etwa 22 kDa, womit es sich bei dem hydrolytischen
Enzym um eine Cutinase von F. solani handelt. Die Analyse der PET-Filme, erfolgt per FTIR, brachte
Informationen über die chemischen und strukturellen Eigenschaften sowie über den enzymatischen
Effekt auf der Oberfläche des Films. Abschließende Untersuchungen zeigten, dass der enzymatisch
behandelte Film effizient mit einem fluorszierenden Alkylbromid (2-Bromomethylnaphthalen (BrNP))
verestert werden konnten, was eines der neuen Anwendungsgebiete des biofunktionalisierten PET
ist (Donelli et al. 2009).
Bis zum jetzigen Zeitpunkt sind keine Informationen über eine Steigerung der Thermostabilität der
löslichen Cutinase von F. solani veröffentlicht. Das Enzym ist instabil oberhalb von 45°C und verliert
mehr als 80% seiner Aktivität über einen Zeitraum von 60 Minuten bei 60°C (Dutta et al. 2009).
In neuesten Studien wurde die hydrolytische Aktivität der eukaryotischen Cutinase aus F. solani
(40°C) und der bakteriellen Cutinase aus T. fusca (60°C) verglichen. Dabei wurde gezeigt, dass die
eukaryotische Cutinase mehr Hydrolyseprodukte in Abhängigkeit zu der eingesetzten Menge an
Protein (mg) freisetzte im Vergleich zur bakteriellen Cutinase (Chen et al. 2010).
Die Cutinase aus F. solani ist eine höchst aktive Hydrolase für PET-Modifikationen und von Interesse
für verschiedene Hydrolysestudien. In den nächsten Jahren sollte die Steigerung der Thermostabilität
durch eine Substitutionsmutagenese stärker untersucht werden, um das Interesse an dieser Cutinase
für die Anwendung zur PET Modifikation noch zu steigern.
Thermomyces (Humicola) insolens ist ein thermophiler Pilz. Ein lipolytische Enzym wurde in einer
E. coli Klonbibliothek, hergestellt aus der genomischen DNA aus H. insolens DSM 1800, identifiziert.
Die Cutinase wurde rekombinant produziert, chromatographisch aufgereinigt und charakterisiert. Die
Bestimmung des Molekulargewichts ergab 20-21 kDa, der isoelektrische Punkt des Enzyms betrug 8.
Das pH Optimum lag bei pH 8, aber auch mit einer hohen Aktivität bis pH 10. Das aktive Zentrum der
eukaryotischen Cutinase besteht aus den Aminosäuren Ser140, Asp195 und His208. Bei der
Untersuchung der lipolytischen Aktivität während eines Waschganges zeigte sich, dass das Enzym
eine hervorragende Fettentfernung bei den Textilien erzielte und eine hohe Affinität gegenüber
Olivenöl zeigte (Sandal et al. 1996).
Die Cutinase wurde eingesetzt für den Abbau von ETE und BEB, für die Bestimmung ihres Depilling
Effekts und die Farbauffrischungsfähigkeit bei Polyestertextilien bei 50°C und zeigte bei allen
Untersuchungen ausgezeichnete Ergebnisse (Andersen et al. 1999).
Die hydrolytische Aktivität der Cutinase gegenüber zyklischen PET-Trimeren (CTR) wurde ebenfalls
untersucht. Während der Inkubation von 16 Stunden bei 30°C wurden durch das Enzym 56% der
zyklischen Oligomere hydrolysiert. Das Haupthydrolyseprodukt war MHET und nur geringe Mengen
an TPA und BHET konnten detektiert werden (Riegels et al. 2001).

42 Einführung
Durch eine lokale zufällige Mutagenese konnten neue Varianten der Cutinase hergestellt werden und
die positiven Klone wurden durch die Inkubation bei 60°C selektiert. Drei positive Varianten wurden
isoliert und durch eine Sequenzierung erfolgte die Identifizierung der Modifikationen A14P + E47K,
E47K und E179Q verglichen zur Wildtyp Cutinase. Durch eine ortsspezifische Mutation wurden
zusätzliche Modifikationen eingefügt. Für sieben Mutanten erfolgte die Bestimmung der
Thermostabilität mittels DSC. Die Wildtyp Cutinase besaß eine Td (thermale Denaturierungs-
temperatur) von 61°C bei pH 4,5. Fünf der Varianten besaßen eine Td von 64-66°C, was eine
Erhöhung von 3-5°C bedeutete. Eine Variante (E6Q, E10Q, A14P, E47K, R51P, E179Q) wies eine
erhöhte Thermostabilität während der CTR Hydrolyse auf und eine optimale Oligomerentfernung bei
pH 9 und 75°C. Das Temperaturoptimum dieser Mutante lag bei 65°C (Abo et al. 2004).
Weitere Mutanten wurden durch eine ortsspezifische Mutagenese mit dotierten Oligonukleotiden
erzeugt. Die erstellten Bibliotheken wurden nach Varianten mit einer höheren Thermostabilität
mittels Hofbildung auf BETEB Agarplatten bei 68°C durchsucht. Alle erzeugten Mutanten bildeten
Höfe bei 68°C, der Wildtyp jedoch nicht, seine bevorzugte Wachstumstemperatur lag bei 60°C. Einige
Varianten waren auch noch aktiv während der Inkubation bei 76°C. Bei der DSC
Thermostabilitätsbestimmung bei pH 10 zeigte der Wildtyp eine Td von 68°C, hingegen zeigten drei
Varianten eine Td von 71-72°C. Somit konnte eine deutliche Steigerung der Thermostabilität erzielt
werden. Mit einer dieser Varianten wurden zusätzliche Textilienbehandlungen durchgeführt
(Waschgänge, Behandlung der Polyestertextilien und Gewichtsverlustberechnungen,
Reinigungseffekt gegenüber Fettflecken, Depillingeffekt und Effekt auf die Wasserabsorption), die
meisten Behandlungen erfolgten bei 75°C und die Mutante zeigte exzellente Resultate (Svendsen
et al. 2005).
2003 veröffentlichten Hooker et al. Abbauergebnisse unter Anwendung einer Cutinase von Novozym
North Amerika, Inc. (Franklinton, NC). Die Cutinase wurde für die Hydrolyse von CTR bei 60°C mit
verschiedenen CTR- und Enzymkonzentrationen, unter Zugabe von Lösungsmitteln und für eine
Langzeitinkubation eingesetzt. Durch die Variation der Reaktionsparameter konnte eine annähernd
komplette Hydrolyse der CTR erreicht werden. Die nachgewiesenen Abbauprodukte waren nur TPA,
MHET und BHET, es konnten keine linearen Dimere oder Trimere per RP-HPLC detektiert werden. Das
bedeutet, dass nach der Ringöffnung eine schnelle Hydrolyse der CTR erfolgte und somit fast alle
Esterbindungen gespalten wurden. Die Durchmischung konnte als ein wichtiger Faktor für die
Hydrolyserate identifiziert werden. Eine höhere Durchmischung resultierte in einer höheren
prozentualen Hydrolyse. Zusätzlich zeigten höhere Enzymkonzentrationen eine komplettere
Hydrolyse der Esterbindungen und das Hauptprodukt verschob sich von MHET zu TPA (Hooker et al.
2003).

43 Einführung
Weitere Studien mit der entwickelten Cutinase wurden 2006 von der gleichen Arbeitsgruppe
durchgeführt. Der Einfluss der Partikelgröße der CTR auf die Enzymkinetik wurde bei 60°C
untersucht. Grob dispergierte CTR zeigten eine größere Partikelgröße als aus einer Dioxanlösung
ausgefällte CTR. Als Konsequenz für die größere Partikelgröße wurde eine heterogene Kinetik 0.
Ordnung detektiert, hingegen zeigten die kleineren Partikel eine homogene Kinetik 1. Ordnung.
Deshalb wurde geschlussfolgert, dass weitere Studien für einen optimaleren Abbau mit Trimeren der
kleineren Partikelgröße durchgeführt werden sollten (Figueroa et al. 2006).
Zwei Mutanten aus dem Patent von Svendsen et al. (2005) wurden ausgewählt für die
Charakterisierung der neuen Varianten. Die Variante A hatte folgende Substitutionen: E6Q, G8D,
A14P, N15D, T29M, E47K, S48E, R51P, A88H, N91H, A130V, T166I, L167P, E179Q, R189V und die
Variante B: E6Q, G8D, A14P, N15D, E47K, S48E, R51P, A88H, N91H, A130V, E179Q, R189V. Die
entwickelten Enzyme wurden angewandt für die Bestimmung ihrer Depilling Eigenschaften bei 70°C
mit und ohne der Zugabe von Triton X-100 als Lösungsvermittler und ihrer Fähigkeit zum Biopolishing
von 100% Polyester und 50%/50% Polyester/Baumwolle Mischgarn bei 70°C. Die Mutante B zeigte
eine höhere hydrolytische Aktivität während der Polyesterbehandlung und exzellente Ergebnisse bei
der Anwendungstemperatur von 70°C (Jump und McCloskey 2004).
Novozym China veröffentlichte 2008 Ergebnisse von enzymatischen Hydrolysestudien von PET-
Textilien zur Steigerung der Hydrophilizität unter Anwendung der Cutinasevariante B von T. insolens.
Der optimale pH Bereich für die Enzympräparation lag bei pH 7-9 und der optimalen Temperatur
Bereich von 65 bis 80°C. In Verbindung mit Triton X-100 konnte bereits bei einer
Cutinasekonzentration von 5 LU/ml eine Durchfeuchtungszeit von 5 Sekunden erreicht werden,
hingegen ohne Lösungsvermittler und mit 200 LU/ml Cutinase wurde eine Durchfeuchtungszeit von
180 Sekunden plus x bestimmt. Die Hydrophilizität und Feuchtigkeitseigenschaften wurden analysiert
durch den Tropfentest (Liu et al. 2008).
Zusätzliche Abbauergebnisse zur PET-Behandlung mit der T. insolens Cutinase wurden 2009
veröffentlicht. Es erfolgte ein Vergleich zwischen der T. insolens Cutinase und zwei weiteren
Cutinasen, einer bakteriellen und einer weiteren eukaryotischen. Die PET-Film Hydrolyse erfolgte bei
70 bis 80°C mit der Cutinase aus T. insolens. Der PET-Film mit der geringen Kristallinität wies eine Tg
von 75°C auf, das bedeutet, dass die Abbaustudien sowohl 5°C unterhalb als auch oberhalb der
Glasübergangstemperatur des PET-Films erfolgten und somit unter optimalen Bedingungen. Mit
dieser Gewissheit war es nicht verwunderlich, dass die entwickelte Cutinase die besten
Abbauergebnisse erzielte mit 100% Gewichtsverlust nach 96 Stunden Inkubation bei 70°C. Ein
adäquates Kinetik-Modell für den Filmabbau wurde ebenfalls bestimmt und mittels RP-HPLC wurde
TPA als Haupthydrolyseprodukt identifiziert (Ronkvist et al. 2009).

44 Einführung
Zu diesem Zeitpunkt ist die entwickelte Cutinase aus T. insolens, erhalten durch ortsspezifische
Mutagenese, das wohl beste PET-hydrolysierende Enzym.
Die Cutinase aus dem gram (-) Pseudomonas putita (später umbenannt in Pseudomonas mendocina)
wurde 1987 aus einer Bakterienkultur aus einer Phyllosphäre identifiziert und zeigte die höchste
cutinolytische Aktivität zwischen pH 9 und 11. Die Wachstumstemperatur war für diesen Stamm mit
41°C relativ hoch und bei der Temperaturstabilitätbestimmung zeigte die Cutinase im extrazellulären
Überstand nach 1 Stunde Inkubation bei 60°C 100% Restaktivität und bei 70°C 85% Restaktivität
(Sebastian et al. 1987). Nach der Aufreinigung der Cutinase wurde das Molekulargewicht mittels SDS-
PAGE und Größenausschlusschromatographie auf 30 kDa bestimmt und eine monomerische Struktur
wurde geschlussfolgert. Das pH Optimum für die Cutinhydrolyse des aufgereinigten Enzyms lag
zwischen pH 8,5 und 10,5 (Sebastian et al. 1988).
Die Cutinase zeigte keine Grenzflächenaktivierung, aber die typische α/β-Hydrolase Faltungsstruktur
und ein aktives Zentrum mit Ser126, Asp176 und His206. Es erfolgte ein positionsspezifischer
Aminosäurenaustausch, um die Cutinase für die Fetthydrolyse und die Detergenzienanwendung zu
verbessern. Die Cutinase aus P. mendocina wurde aufgrund der Triglyceridhydrolyse im alkalischen
Bereich ausgewählt. Der Austausch an Position Y202L/R203L steigerte die Acylkettenbindung der
Triglyceride und damit die Aktivität gegenüber längeren Triglyceriden mit C18 Acylseitenketten
(Boston et al. 1997).
Die ersten enzymatischen Behandlungen von PET-Textilien wurden 1988 mit der Cutinase aus
P. mendocina, zur Verfügung gestellt von Genencore International, durchgeführt. Die Cutinase
steigerte die Durchfeuchtungs- und Absorbtionseigenschaften von Dacron 54 (PET-Homopolymer),
Dacron 64 (sulfoniertes PET, SPET) und MicromattiqueR (Mikrodenier PET). Die optimalen
Reaktionsbedingungen lagen mit 0,12 g/L Cutinase bei 35°C für 10 Minuten. Im Vergleich zur
alkalischen Hydrolyse zeigte die Cutinasebehandlung den Vorteil, dass sie eine stärkere Steigerung
der Durchfeuchtung bei einer geringeren Temperatur (Hsieh und Cram 1998, Hsieh et al. 2003)
erzielte. In weiteren Studien wurde die Cutinase eingesetzt, um die Farbstoffaufnahme und
Absorption von kationischen Farbstoffen bei Dacron 54 und Dacron 64 nach einer Behandlung für 24
Stunden bei 40°C zu untersuchen. Vor der Färbung wurden die Textilien mit einer Protease und
Triton X-100 behandelt und gründlich gewaschen, um die anheftenden Proteinverunreinigungen zu
entfernen. In einer anderen Untersuchung wurde nachgewiesen, dass die Cutinasebehandlung eine
Steigerung des Depillingeffekts zur Folge hatte, zusätzlich wurde ein Gewichtsverlust im Vergleich zur
Kontrolle detektiert und die Oberflächenmodifikationen der PET-Fasern per SEM beobachtet. Die
Cutinase modifizierte ebenso PTT Fasern, ebenfalls nachgewiesen durch die Farbstoffaufnahme. Die
während der enzymatischen Inkubation freigesetzten Hydrolyseprodukte wurden mittels UV

45 Einführung
Absorbtionsmessung und RP-HPLC nachgewiesen. Das detektierte Produkt nach der
Natronlaugebehandlung war nur TPA, hingegen bei der Cutinasebehandlung wurde ein unbekanntes
Produkt analysiert. Mit der Cutinase aus P. mendocina wurde zum ersten Mal gezeigt, dass nur
Textilien mit einer geringen Orientierung/Kristallinität effektiv hydrolysiert werden im Gegensatz zu
hoch orientierten/kristallinen Textilien (Yoon et al. 2002, Dyson et al. 2005).
Eine ortsspezifische gesättigte Mutagenese erfolgte mit der Cutinase aus P. mendocina, um die
hydrolytische Aktivität gegenüber Polyethylenterephthalat Textilien und/oder die Enzymstabilität zu
steigern. Die Mutanten wurden bezüglich ihre Freisetzung von löslichen TPA-enthaltenden Derivaten
während der enzymatischen Hydrolyse von PET mittels UV Absorptionsmessung im Plattenleser
getestet. Die Stabilitätsuntersuchungen wurden ebenfalls im Plattenleser anhand der Spaltung von p-
NPB ermittelt. Die ortsspezifische abgesättigte Mutagenese an den Positionen 63-65, 178-182, 205,
209 und 211 hatte Varianten zur Folge, von denen mehr als 2% eine höhere Aktivität besaßen als die
Kontrolle. Mutationen an den Positionen 59, 64, 150, 177, 180 und 182 brachten Varianten hervor,
von denen mehr als 2% der Klone eine größere Stabilität aufwiesen gegenüber der Kontrolle.
Varianten die sowohl eine erhöhte Aktivität und Stabilität zeigten, wiesen Substitutionen an den
Positionen 64, 180, 182 und 211 auf. Zusätzliche Mutationen wurden durchgeführt und der
Austausch des Serins an den Stellen 20 und 85 kreierten neue Bibliotheken mit neuen Positionen.
Mutanten mit einer Substitution von Phenylalanin an der Position 180 durch Isoleucin, Leucin,
Asparagin oder Prolin brachten Varianten hervor mit einer erhöhten Stabilität gegenüber dem
Wildtyp. Ebenso zeigte ein Austausch des Glycins an der Position 59 mit Phenylalanin, Lysin, Leucin
oder Valin eine Erhöhung der Stabilität im Vergleich zum Wildtyp. Die Kombination der
Substitutionen von Phenylalanin an der Position 180 und Serin an der Position 205 ergaben Varianten
mit einer signifikanten Steigerung der relativen Stabilität. Zusätzliche Austausche an einer bis drei
Positionen hatten Mutanten zur Folge mit gesteigerter PET-Hydrolaseaktivität (Bott et al. 2003).
Nicht überraschend zeigte die Cutinase von P. mendocina in den Untersuchungen von Ronkvist et al
(2009) ein Temperaturoptimum von 50°C. Verglichen zu den Cutinasen von T. insolens und F. solani
zeigte die Cutinase von P. mendocina die stärkste Affinität gegenüber PET. Während einer 96
stündigen Abbaustudie bei 50°C setzte die Cutinase nur TPA und Ethylenglycol als lösliche Produkte
vom gering kristallinen PET-Film (7% Kristallinität) frei, detektiert durch die RP-HPLC. Der
Gewichtsverlust des Films war nur 5% aufgrund der Temperaturdifferenz von etwa 25°C zwischen der
Inkubationstemperatur und der Glasübergangstemperatur des PET (Ronkvist et al. 2009).
Zusammenfassend ist aufgrund der Tatsache, dass PET besser abbaubar ist bei Temperaturen um 75
bis 80°C, die Temperaturstabilität und die optimale Reaktionstemperatur für eine industrielle
Anwendung immer noch zu gering.

46 Einführung
Thermobifida (zuvor Thermomonospora) fusca ist ein gram (+), thermophiler, aerober,
sporenbildender Actinomycet mit einem Temperaturoptimum zwischen 50 und 55°C. Sein
natürliches Habitat sind Dung, Kompost und überhitztes Heu (McCarthy 1989).
Vier verschiedene Stämme von T. fusca sind bisher bekannt und untersucht, T. fusca DSM 43792
(ATCC 27730) und seine Mutanten, T. fusca DSM 43793, T. fusca YX und T. fusca KW3.
Der erste untersuchte Stamm war der Referenzstamm T. fusca DSM 43792, welcher auf die Bildung
von cutinolytischer Aktivität getestet wurde. Die Rohenzymlösung zeigte eine optimale Temperatur
von 50 bis 70°C und einen optimalen pH von 11. Die Enzympräparation besaß eine Halbwertszeit bei
70°C von 60 Minuten (Fett et al 1999).
Weiterführend wurde eine Mutante des Referenzstamms mittels DES und UV-Licht Behandlung
erzeugt. Diese Mutante, WSH04, zeigte eine höhere Cutinaseproduktion gegenüber dem Wildtyp.
Mit diesem Bakterienstamm erfolgte die Optimierung der Cutinaseproduktion durch eine zwei-
stufige pH Kontrolle und eine maximale Cutinaseaktivität von 19,8 U/ml konnte erreicht werden nach
50 Stunden Inkubation (Du et al. 2007).
Eine Enzympräparation von T. fusca KW3 mit Esteraseaktivität, induziert während des Wachstums
mit Suberin, zeigte nach der Inkubation mit PET-Fasern, PBT-Fasern und recyceltem PET-Granulat
eine Freisetzung von aromatischen Bestandteilen bei 50°C und 70°C. Mit der höheren Temperatur
konnten mehr Hydrolyseprodukte nachgewiesen werden. Die Effekte der Fasermodifikation wurden
nach der Färbung mit Reaktivfarbstoffen durch die Bestimmung der Farbintensität nachgewiesen,
ebenso wie mit der Steighöhenbestimmung und Flüssigkeitsaufnahmekapazitätsmessung (Alisch
et al. 2004, Alisch-Mark et al. 2006, Feuerhack et al. 2008).
Ein weiteres Enzym mit cutinolytischer Aktivität wurde 2005 aus T. fusca DSM 43793 isoliert: BTA1
(TfH). Dieses Enzym wurde induziert durch den aliphatisch-aromatischen Copolyester (BTA40:60) und
dessen Oligomere, aber nicht mit dem Monomer TPA oder mit Poly-ε-caprolacton (PCL) produziert.
Die molekulare Masse des Enzyms war 28,2 kDa, der pI lag bei 6,43. TfH ist eine aus 261
Aminosäuren bestehende Serin-Hydrolase. Die katalytische Triade besteht aus den Aminosäuren
Ser132, Asp176 und His210. Das Enzym konnte Cutin aus Apfelschalen abbauen und das aktive
Zentrum ist frei zugänglich, das heißt, dass kein schließender Deckel das aktive Zentrum bedeckt
(Kleeberg et al. 2005). Zusätzlich baute die TfH käufliche PET-Flaschen und PET-B Pellets bei 55°C ab
(Müller et al. 2006). In weiteren Arbeiten wurde die Cutinase rekombinant in E. coli und nach einer
Codonoptimierung in B. megaterium mit einem N-terminalen Signalpeptid für die B. megaterium
Lipase A und einem C-terminalen His6-Anhang unter der Kontrolle eines Xylose-induzierten
Promotors produziert (Dresler et al. 2006, Yang et al. 2007). Alle diese Studien über die Cutinase TfH
aus T. fusca DSM 43793 basieren auf einem Patent von Deckwer et al. aus dem Jahr 2000, in dem die
Lokalisation der Gene der BTA Hydrolasen BTA1 und BTA2 beschrieben wurde. Beide Enzyme liegen

47 Einführung
nacheinander im Genom vor und nach ihrer Transkription und Prozessierung besitzen sie fast das
gleiche Molekulargewicht (beide bestehen aus 261 Aminosäuren, BTA1 mit 28,2 kDa und BTA2 mit
28,4 kDa). Bei einem Sequenzvergleich wurde festgestellt, dass 20 der 261 Aminosäuren sich in den
beiden Cutinasen unterscheiden, woraus sich eine 92% Übereinstimmung der beiden Cutinasen
ableiten lässt (Deckwer et al. 2000). Bisher wurden nur Studien über eine PET-hydrolysierende
Aktivität mit BTA1 (TfH) durchgeführt, da die hydrolytische Aktivität der BTA2 geringer war. Die
rekombinante TfH baute das PET Modellsubstrat BETEB ab und steigerte die Hydrophilität von PET-
Textilien während der enzymatischen Behandlung (die Inkubationstemperatur wurde nicht genannt).
Der Nachweis der Modifikation erfolgte mittels Steighöhenbestimmung und Tropfentest (Heumann
et al. 2006). Die Hydrolyse von BHPT, der bis(3-Hydroxypropyl)Ester der Terephthalsäure, erfolgte bei
60°C über einen Zeitraum von 24 Stunden. Die TfH hydrolysierte zunächst das BHPT zu MHPT und
später den Monoester komplett zu Terephthalsäure. Hingegen wurde beim Abbau von zyklischen
PPT-Dimeren bei 60°C durch die TfH kein BHPT nachgewiesen. Zu Beginn der Hydrolyse war das
Hauptprodukt MHPT (bis etwa 48 Stunden Inkubation), aber nach 96 Stunden wurde TPA in höheren
Konzentrationen detektiert. Die PTT-Film-Modifikationen durch die TfH bei 60°C wurden
nachgewiesen durch die Erniedrigung des Kontaktwinkels nach der Behandlung und durch den
fehlenden Kristallisationspeak bei 72°C, analysiert per DSC. Dabei betrug der Temperaturunterschied
zwischen der Inkubationstemperatur und der Glasübergangstemperatur (Tg) 12°C. Die Behandlung
von PTT-Textilien mit der Cutinase aus T. fusca bei 60°C zeigte ebenfalls eine hydrolytische Aktivität
durch die Freisetzung von MHPT und TPA, den bestimmten Gewichtsverlust und eines deutlichen
Anstiegs des K/S Verhältnisses (Eberl et al. 2008). Weiterhin wurde die enzymatische Behandlung von
PET-Textilien mit der TfH bei 60°C untersucht. Die Modifikationen wurden nachgewiesen durch die
Detektion der Hydrolyseprodukte per HPLC, von den amorphen Textilien wurden TPA, MHET und
BHET freigesetzt und von den semi-kristallinen Textilien TPA und MHET. Veränderungen in der
elementaren Zusammensetzung der Textilien nach der enzymatischen Behandlung wurden durch
eine XPS-Analyse nachgewiesen. Zusätzlich erfolgte der Nachweis der Oberflächenmodifikationen
durch einen Tropfentest, durch die Bestimmung der Durchfeuchtungszeit und eine K/S-
Verhältnisbestimmung (Brueckner et al. 2008). Überraschenderweise ergab die Zugabe von Triton X-
100 zum BETEB (3PET)-Abbau keine Steigerung der hydrolytischen Aktivität der Cutinase während
der Inkubation bei 60°C. Hingegen zeigte die Addition von N,N-Diethyl-2-Phenylacetamid (DEPA),
einem Weichmacher, eine Steigerung der Abbaurate bei den semi-kristallinen PET-Filmen und
Textilien. Daraus wurde geschlussfolgert, dass für die Cutinase aus T. fusca die Zugabe eines
Weichmachers für eine Steigerung der Hydrolyserate angewandt werden sollte (Eberl et al. 2009).
Basierend auf der vollständigen Sequenzierung des Genoms von T. fusca YX im Jahre 2008 (Lykidis
et al. 2008) wurden zwei ähnliche Cutinasen aus T. fusca WSH03-11 identifiziert (Chen S et al. 2008,

48 Einführung
Chen J et al. 2008). Beide Enzyme, kloniert und produziert in E. coli, wiesen ähnliche enzymatische
Eigenschaften auf und beide hydrolysierten Apfelcutin unter Freisetzung von Cutinmonomeren. Die
optimale Temperatur für die Hydrolyse von Triolein und p-NPB lag bei beiden Enzymen bei 60°C und
der optimale pH Wert lag bei 8. Eine verbleibende Restaktivität von 80% nach 160 Stunden
Inkubation bei 40°C und eine Restaktivität von 50% nach 40 Stunden Inkubation bei 60°C wurde
nachgewiesen (Chen S et al. 2008). Die identifizierten Cutinasen zeigten eine 93% Übereinstimmung
ihrer Proteinsequenz. Interessanterweise konnte während der Inkubation der Enzyme mit zyklischen
PET-Trimeren (CTR) bei 60°C nur eine Cutinase lösliche Hydrolyseprodukte freisetzen. Die löslichen
Hydrolyseprodukte wurden mittels RP-HPLC analysiert und per LC-MS verifiziert (Chen et al. 2010).
Zusammenfassend ist bekannt, dass der thermophile Actinomycet T. fusca zwei Isoenzyme mit mehr
als 90% Übereinstimmung produziert, welche sich in ihrer Hydrolyserate gegenüber PET deutlich
unterscheiden. Nur mit einer der Cutinasen konnten PET-Textilien effektiv modifizieren werden. Die
Untersuchung der Unterschiede der beiden Enzyme durch die Bestimmung der Kristallstrukturen,
durch Sequenzvergleiche und Mutationsanalysen und die Bestimmung ihres Einflusses auf die
katalytische Aktivität gibt einen interessanten Einblick in die PET-Hydrolysereaktion. Zusätzlich sollte
auch für diese Enzyme eine Steigerung der Thermostabilität und Aktivität durch eine ortsspezifische
Mutagenese oder einen Austausch von Aminosäuren im Hinblick auf eine industrielle Anwendung
erfolgen.

49 Einführung
1.5 Zielsetzung
Besonders thermophile Actinomyceten sind außerordentlich leistungsfähig beim Abbau von
Kunststoffen sind (Kleeberg et al. 1998). Ferner spielen Actinomyceten, darunter auch
Thermomonospora spp., eine wichtige Rolle beim Abbau von Pflanzenpolymermaterial und sind
ebenfalls in der Lage, eine große Vielfalt an hydrolytischen extrazellulären Enzymen zu bilden (Wilson
1988; Zimmermann 1989; Röthlisberger et al. 1992; Hilge et al. 1998). Bei Untersuchungen mit
Streptomyces spp. (Lin und Kolattukudy 1980; Fett et al. 1992) und Thermomonospora fusca (Fett
et al. 1999) konnte eine Cutinaseaktivität nachgewiesen werden, welche die Hydrolyse des
aliphatischen Pflanzenpolyesters Cutin verursachte. Von Zimmermann et al. (1985) und Bernards
et al. (1995) ist bekannt, dass auch der pflanzliche Polyester Suberin sowohl aliphatische als auch
aromatische Regionen enthält und aufgrund seiner Struktur zur Induktion von Polyester
hydrolysierenden Enzymen in Actinomycetenisolaten eingesetzt werden könnte.
Diese hydrolytischen Enzyme können mit ihren günstigen Eigenschaften, wie zum Beispiel
Temperaturstabilität oder Substratspezifität, bei der Entwicklung eines biotechnologischen
Verfahrens zum Abbau von PET-Oligomer eingesetzt werden.
Dadurch eröffnet sich die Chance, durch den Einsatz von spezifischen Biokatalysatoren zur
Elimination von synthetischen PET-Oligomer ein alternatives umweltfreundliches biotechnologisches
Verfahren zur Textilveredelung von Chemiefasern zu erhalten.
Das Ziel dieser Arbeit war die Untersuchung eines biokatalytischen Abbaus von PET-Oligomeren,
welcher umweltfreundlich und faserschonend zugleich ist und somit eine optimale Alternative zur
problembehafteten industriellen Textilveredelung darstellt.
Mit dem aus Kompost gewonnenen Actinomyceten Thermobifida fusca KW3 wurde ein Enzym (TfCa)
produziert, welches in der Lage war, die PET-Oligomere auf der Faseroberfläche abzubauen, ohne die
Faser zu beschädigen. Nach der Optimierung der Induktion der Hydrolase erfolgte die Produktion des
Enzyms in größeren Mengen und es wurde eine Methode zur Aufreinigung entwickelt. Anschließend
wurde die TfCa biochemisch charakterisiert und verglichen mit bereits bekannten PET-Hydrolasen.
Desweiteren erfolgte die Aufreinigung der beiden rekombinant produzierten Cutinasen TfCut1 und
TfCut2 aus T.°fusca KW3 parallel zu rekombinant produzierten Carboxylesterase TfCa, welche alle
drei in der Lage waren, PET mit unterschiedlicher Effektivität zu modifizieren.
Mit den aufgereinigten Enzymen und weiteren bekannten PET-Hydrolasen aus
Fusarium solani sp. pisi, T. fusca sp. und einer eukaryotischen Cutinasepräparation von Novozymes
erfolgten anschließend Abbauversuche, um den Abbaumechanismus auch im Vergleich zueinander
zu untersuchen. Dabei kamen verschiedene PET-Substrate in Form von PET-Trimeren, PET-Fasern
und PET-Film zum Einsatz.

Material und Methoden 50
2 Material und Methoden
2.1 Materialien
2.1.1 Verwendete Mikroorganismen
Thermobifida fusca KW3 (Thermomonospora), ein Isolat aus Komposterde, ist ein gram positives
aerobes thermophiles und alkalophiles Bakterium, das den Actinomyceten zugeordnet wurde.
Hinterlegt ist der Stamm unter der Stammbezeichnung DSM 6013 bei der Deutschen
Stammsammlung für Mikroorganismen und Zellkulturen (Braunschweig, Deutschland). Die optimalen
Wachstumsbedingungen für T. fusca KW3 sind 50-55°C und pH 7,3 (Casimir-Schenkel 1992). Die
Kultivierung von T. fusca KW3 erfolgte auf Czapek-Medium.
Des Weiteren wurden T. fusca DSM 43792 und T. fusca DSM 43793, bezogen von der Deutschen
Stammsammlung für Mikroorganismen und Zellkulturen (Braunschweig, Deutschland), in die
Arbeiten einbezogen. Die Kultivierung von T. fusca DSM 43792 erfolgte ebenfalls mit Czapek-
Medium, die Kultivierung von T. fusca DSM 43793 mit einem modifizierten Czapek-Medium, welches
6,1 g/L Casein Hydrolysat anstatt 5 g /L Pepton enthielt.
2.1.2 Verwendete Enzyme
Die aufgereinigte Carboxylesterase (EC 3.1.1.1) TfCa aus T. fusca KW3 wurde mit einer spezifischen
Aktivität von 11,6 U/mg bei den Abbauuntersuchungen eingesetzt, dazu wurde die erhaltene
vereinigte Enzymlösung gefriergetrocknet und bei –20°C gelagert.
Für die Abbauuntersuchungen wurde ebenfalls eine eukaryotische Cutinase (EC 3.1.1.74) aus
Fusarium solani pisi, erhalten von Unilever Research and Development Laboratory in Vlaardingen,
eingesetzt. Die Cutinase lag in immobilisierter Form vor und wurde, wie von Vertommen et al. (2005)
beschrieben, vorbereitet. Weiterhin wurde eine Cutinase, vorliegend als flüssiges Konzentrat von
Novozymes (NS-29061, LN05012), für die Abbauuntersuchungen verwendet.
Die Cutinasen 1 und 2 aus dem Isolat T. fusca WSH03-11 wurden rekombinant produziert von Chen S
et al. (2008) erhalten und lagen lyophilisiert vor (Chen J et al. 2008).
Die rekombinant produzierten Cutinasen TfCu1 und TfCu2 und die Carboxylesterase TfCa aus T. fusca
KW3, bereitgestellt von Herrn Ren Wei und Herrn Thorsten Oeser, lagen aufgereinigt mit einer
spezifischen Aktivität von 230 U/mg, 155 U/mg und 204 U/mg in Lösung vor.

Material und Methoden 51
2.1.3 Größenstandards
2.1.3.1 Low Molecular Weight Marker (GE Healthcare)
Der Proteinstandard diente zum Abschätzen der molekularen Größe der Proteine im SDS Gel. Die
enthaltenen Proteine besaßen eine Größe von 97 000 g/mol, 66 000 g/mol, 45 000 g/mol,
30 000 g/mol, 20 100 g/mol und 14 400 g/mol.
2.1.3.2 Roti®-Mark Standard (Fa. Carl Roth GmbH)
Der Roti®-Mark SDS Proteinstandard wurde für die Berechnung des Molekulargewichtes der Esterase
verwendet. Die enthaltenen Proteine besaßen eine Größe von 200 000 g/mol, 119 000 g/mol,
66 000 g/mol, 43 000 g/mol, 29 000 g/mol, 20 000 g/mol und 14 500 g/mol.
2.1.3.3 SpectraTM Multicolor Broad range Protein Ladder (Fermentas)
Der Proteinstandard diente zum Abschätzen der molekularen Größe der Proteine im SDS Gel. Die
enhaltenen Proteine besaßen eine Größe von 260 000 g/mol, 135 000 g/mol, 95 000 g/mol,
72 000 g/mol, 52 000 g/mol, 42 000 g/mol, 34 000 g/mol, 26 000 g/mol, 17 000 g/mol und
10 000 g/mol.
2.1.3.4 PageRulerTM plus prestained protein ladder (Fermentas)
Der Proteinstandard diente zum Abschätzen der molekularen Größe der Proteine im SDS Gel. Die
enhaltenen Proteine besaßen eine Größe von 250 000 g/mol, 130 000 g/mol, 95 000 g/mol,
72 000 g/mol, 55 000 g/mol, 36 000 g/mol, 28 000 g/mol, 17 000 g/mol und 11 000 g/mol.

Material und Methoden 52
2.1.3.5 Kalibrierungskit für pI Bestimmung (pH 3-10, GE Healthcare)
Das Kalibrierungskit für die pI Bestimmung wurde für die Berechnung des pI der Esterase mittels
Isoelektrischer Fokusierung verwendet.
Abbildung 23: Kalibrierungskit für die pI Bestimmung, Breiter pI Bereich (pH 3-10, rechte Spur, GE Healthcare).
2.1.3.6 Kalibrierungskit für die Größenausschlusschromatographie (LMW, GE Healthcare)
Das Kalibrierungskit für die Größenausschlusschromatographie wurde für die Kalibrierung der
Größenausschlusschromatographiesäule verwendet.
Tabelle 9: Das Kalibrierungskit für die Größenausschlusschromatographie.
Protein Molekulargewicht (g/mol) Herkunft Position
Blue dextran 2000 2 000 000
Conalbumin 75 000 Hühnerei a
Ovalbumin 43 000 Hühnerei b
Carboanhydrase 29 000 Schweineerythrozyten c
Ribonuclease A 13 700 Schweinepankreas d
Aprotinin 6 500 Schweinelunge e

Material und Methoden 53
0
100
200
300
400
500
600
700
800
900
0 50 100 150 200 250 300 350 400
UV
(m
AU
)
Elutionsvolumen (ml)
y = -0.03x + 17.239R² = 0.9891
0
5
10
15
200 250 300ln M
ole
kula
rge
wic
ht
Elutionsvolumen (ml)
Abbildung 24: Elutionsprofil des Kalibrierungskit für die Größenausschlusschromatographie eluiert von einer XK 26/60
Säule, Superdex 200 prep grade, 320 ml SV, 50 mM Tris HCl pH7,2 150 mM NaCl, 2,5 ml/min, 280 nm und dazugehörige Kalibrierkurve (kleine Abbildung).
2.1.4 Chemikalien
Die Chemikalien wurden, wenn nicht weiter erwähnt, mit der Reinheit p.a. eingesetzt.
Acros Organics (Geel, Belgium) Diethylterephthalat (DET), 2-Naphtylacetat,
Phenylmethyl-sulfonylfluorid (PMSF),
Thiamindichlorid
Aldrich Chemical Company (St. Louis, UAS) p-Aminobenzoat, (NH4)6Mo7O24 x 4 H2O,
Terephthalsäure (TPA)
Fluka Chemie (Buchs, Schweiz) Coomassie Brilliant Blue R250, Jodacetamid, N,N´-
Methylenbisacrylamid, Poly(ε-caprolacton) (PCL)
Grüssing GmbH (Filsum, Deutschland) Essigsäure, KH2PO4, NaH2PO4 x 2 H2O, Na2HPO4,
(NH4)2SO4, NH4OH
Macherey-Nagel GmbH & Co. KG (Düren, N-Methyl-N-trimethylsilyl-trifluoracetamid (MSTFA)
Deutschland)
a
b
c
d
e

Material und Methoden 54
Merck (Darmstadt, Deutschland) Acrylamid, AgNO3, Biotin, CuCl2 x 2 H2O, Eisessig,
Ethanol absolut, FeCl3 x 6 H2O, FeSO4 x 7 H2O,
Glycerin, konz. HCl, Hefeextrakt, Hexansäure
(Caproinsäure), Isopropanol, KCl, K2HPO4, MgSO4 x 7
H2O, MnCl2 x 4 H2O, NaNO3, Natriumsulfit, NH4Cl,
Niacinamid, Triton-X-100, Vitamin B12, ZnCl2
Riedel de Haën AG (Seelze, Deutschland) n-Caprylsäure, Malachitgrün-Oxalat, Safranin T
Carl Roth GmbH (Karlsruhe, Deutschland) Acetonitril HPLC rein, Agar, Buttersäure, CHAPS,
Glycin, β-Mercaptoethanol, Methanol HPLC rein,
NaCl, NaOH, NH4HCO3, Pepton, Roti-Quant,
Saccharose, SDS, Tributyrin, Tricaprylin,
Trifluoressigsäure (TFA), TRIS
Scharlau Chemie S.A. (Barcelona, Spanien) 1,4-Dioxan
Serva Feinbiochemika GmbH Bromphenolblau, BSA, EDTA
(Heidelberg, Deutschland)
Sigma Chemical Company (St. Louis, UAS) Ammoniumpersulfat, bis (2-hydroxyethyl)
Terephthalat (BHET), Chymotrypsin, D,L-6,8-
Dithiooctansäure, Dithiothreitol (DTT), Ethylenglycol,
Fast Red TR Salt, Folsäure, Formaldehyd, Gum
arabicum, Methyloleat, Na2B4O7 x 10 H2O, para-
Nitrophenylacetat (p-NPA), para-Nitrophenylbutyrat
(p-NPB), para-Nitrophenylcaprylat (p-NPC), para-
Nitrophenylpalmitat (p-NPP), D-Panthotensäure,
Pyridoxalhydrochlorid, Riboflavin, N,N,N´,N´-
Tetramethylethyldiamin, Tosyl-L-phenylalanine-
chloromethylketon (TPCK), Tricaproin, Trypsin,
Zitronensäure
VEB Laborchemie Apolda (Deutschland) Ammoniumoxalat, CaCO3, Oxalsäure

Material und Methoden 55
2.1.5 Geräte und Materialien
Chromatographie-Säulen XK 16/20 (GE Healthcare, München, Deutschland)
XK 16/40 (GE Healthcare, München, Deutschland)
XK 16/100 (GE Healthcare, München, Deutschland)
Octyl Sepharose 4 Fast Flow (GE Healthcare,
München, Deutschland)
Dialyse Spectra/Por® Membrane, MWCO 6-8.000 (Spectrum
Laboratories Inc., Rancho Dominguez, USA)
Elektrophorese Typ P8DS, Owl Separation Systems Inc. (Portsmouth,
USA)
Protean®II xi Cell (Bio-Rad Laboratories, Hercules,
USA)
PhastSystemTM (GE Healthcare, Schweden)
ESCA PHI Quantera XPS (Physical Electronics Inc.,
Chanhassen, USA)
0,45 µm Filter Zapcap-CR (Schleicher & Schuell MicroScience
GmbH, Dassel, Deutschland)
GC CP-3380 (Varian Inc., Palo Alto, USA)
GC Säule Roti Cap-5-0,25 µm, 30 m x 0,25 mm ID (Carl Roth
GmbH, Karlsruhe, Deutschland)
GC-MS Finnigan Trace GC ultra, Finnigan MAT 95XP (Thermo
Electron, Waltham, USA)
GC-MS Säule Thermo TR-5MS, 30 m x 0,25 mm ID 0,25 µm, C12
(Thermo Electron, Waltham, USA)

Material und Methoden 56
HPLC Äkta Basic 10 (Pumpe P-903, Detektor UV-900,
Fraktionssammler Frac-900, Motor Valve PV-908,
Injektionsventil Inv-907, Mischkammer M-925 0.6ml)
(GE Healthcare, München, Deutschland)
Elite LaChrom (Pump L-2130, Diode Array Detector L-
2455, Autosampler L-2200, Column oven L-2350)
(VWR Hitachi, Darmstadt, Deutschland)
Inkubationsschüttler Innova 4400 (New Brunswick Scientific Co., Edison,
USA)
Heraeus T12 (Kendro Laboratory Products GmbH,
Wien, Österreich) Schüttler Typ 3006 (GFL -
Gesellschaft für Labortechnik mbH, Burgwedel,
Deutschland)
Isoelektrische Fokusierung PhastSystemTM, PhastGel® IEF 3-9 und PhastGel® IEF 4-
6,5 (GE Healthcare, München, Deutschland)
LC-MS HP 1100 Serie HPLC Technologie (Agilent
Technologies, Santa Clara, USA), UV Detektor
(Cohesive Technologies, Thermo Fisher Scientific Inc.,
Franklin, USA), Massenspektrometer Esquire 3000
plus mit Elektrospray-Ionisation und Ionenfalle
(Bruker Daltonik GmbH, Bremen, Deutschland)
LC-MS Säule Gemini Säule 150 x 2,00 mm 5 µ C18 110A
(Phenomenex, Aschaffenburg, Deutschland)
MALDI Tof/Tof Bruker
Photometer Cary 50 Bio UV-Visible Spectrophotometer (Varian
Inc., Palo Alto, USA)
Plattenleser PowerWave XS (Bio-Tek®, Instruments, Bad
Friedrichshall, Deutschland)

Material und Methoden 57
Power supply Typ E844 (Consort nv, Turnhout, Belgium)
Rundfilter Durchmesser 90mm (Schleicher & Schuell
MicroScience GmbH, Dassel, Deutschland)
Säulenmaterial Phenyl Sepharose 6 Fast Flow (low sub) (GE
Healthcare, München, Deutschland)
Toyopearl DEAE-650M (Tosoh Bioscience GmbH,
Stuttgart, Germany)
Superdex 200 prep grade (GE Healthcare, München,
Deutschland)
Säule RP Chromsep HPLC Colum SS 250 x 4,6 mm Microsorb
300-5 C18 (Varian Inc., Palo Alto, USA)
Schüttler HTM 130L (HLC Biotech, Bovenden, Deutschland)
Sequenzer Procise® cLC Sequencing System, Model 492cLC
(Applied Biosystems, Foster City, USA)
Thermoblock für Derivatisierung Typ BBA2 (Grant-Boekel, Grant Instruments,
Cambridge, UK)
Zentrifuge Rotina 46R (Hettich Zentrifugen, Tuttlingen,
Deutschland)
Micro R22 (Hettich Zentrifugen, Tuttlingen,
Deutschland)

Material und Methoden 58
2.1.6 Software
Folgende Software wurde für Analysen oder die Auswertung von Analysen oder Datensätzen
verwendet:
Cary WinUV Kinetics (Varian Inc., Palo Alto, USA)
Unicorn 3.21 (GE Healthcare, Uppsala, Schweden)
PyMOL (DeLano Scientific, San Carlos, USA)
Varian Star 5.5 (Varian Inc., Palo Alto, USA)
EZChrom Elite 3.3.1 (VWR, Darmstadt, Deutschland)
Xcalibur 1.4 (Thermo Electron, Waltham, USA)
KC4 v3.3 (Bio-Tek®, Instruments, Bad Friedrichshall, Deutschland)
2.1.7 Nährmedien
Die Medien wurden 20 min bei 121°C und 1 bar Überdruck autoklaviert. Temperaturlabile Lösungen
wurden sterilfiltriert und nach dem Autoklavieren und Abkühlen steril dem Medium zugeführt.
Soweit nicht anders angegeben wurden die Medienbestandteile in deionisiertem Wasser gelöst. Für
die Herstellung von festen Medienplatten wurden der Kulturlösung vor dem Autoklavieren 15 g/L
Agar zugesetzt.
Czapek-Medium (Vollmedium) Saccharose 30 g/L
NaNO3 3,0 g/L
K2HPO4 1,0 g/L
MgSO4 x 7 H2O 0,5 g/L
KCl 0,5 g/L
FeSO4 x 7 H2O 0,001 g/L
Hefeextrakt 2,0 g/L
Pepton 5,0 g/L
pH-Wert 7,3
MSV-Medium (Mineralsalz-Spurenelement-Vitamin-Medium) (Produktionsmedium)
Pepton 5 g/L
Mineralsalzlösung1 12,5 mL/L
Spurenelementlösung2 1 mL/L

Material und Methoden 59
Additiv 2 g/L
Gelöst in 50 mM Phosphatpuffer pH 8
Nach dem Autoklavieren steril zugeben
Vitaminlösung3 1 mL/L
Zusammensetzung der Lösungen:
1 Mineralsalzlösung: 0,4 g/L CaCO3; 6 g/L KH2PO4; 1 g/L MgSO4 x 7 H2O; 1 g/L NaCl; 10 g/L
NH4Cl; 3 g/L (NH4)2SO4
2 Spurenelementlösung: 0,01 g/L CuCl2 x 2 H2O; 0,2 g/L FeCl3 x 6 H2O; 0,01 g/L MnCl2 x 4 H2O;
0,01 g/L Na2B4O7 x 10 H2O; 0,01 g/L (NH4)6Mo7O24 x 4 H2O; 0,04 g/L
ZnCl2
3 Vitaminlösung: 20 mg/L p-Aminobenzoat, 6 mg/L Biotin, 50 mg/L D,L-6,8-Dithiooctansäure,
20 mg/L Folsäure, 20 mg/L Niacinamid, 10 mg/L D-Panthotensäure, 100 mg/L
Pyridoxalhydrochlorid, 50 mg/L Riboflavin, 10 mg/L Thiamindichlorid,
50 mg/L Vitamin B12, Sterilfiltrieren und nach dem Autoklavieren Medium
zusetzen
Als Additive im MSV Medium wurde Polyethylenterephthalat und Polyethylenbutyrat in folgender
Form eingesetzt:
unbehandeltes, vorgewaschenes Polyethylenterephthalat-Garn (Dr. Th. Böhme KG,
Geretsried, Deutschland);
zyklische PET-Oligomerpräparation (~90%, 9/03, Dr. Th. Böhme KG, Geretsried,
Deutschland);
lineare und zyklische PET-Oligomerpräparation (Prof. Andreaus, FURB Universität in
Blumenau, Brasilien);
PET-Granulat (TEXPET A P800, particle size 4-8 mm, Texplast GmbH Wolfen,
Deutschland);
Polybutylenterephthalat-Garn (AMTEX Ltd., 78/34-1 CS, Barcelona, Spanien).

Material und Methoden 60
2.1.8 Suberin- und Cutin-Präparationen
Die Suberinpräparation wurde durch die Behandlung von Kartoffelschalen mit einem
Ammoniumoxalat-Oxalsäure-Puffer bei 100°C und 24 Stunden Inkubation unter Rühren hergestellt.
Die Schalen wurden mit dest. Wasser gespült, getrocknet und zerkleinert (Fernando et al. 1984).
Die Cutinpräparation wurde wie oben beschrieben aus Apfelschalen gewonnen. Nach dem Waschen
der Schalen mit Wasser erfolgte die Behandlung mit Cellulase (Sigma, 5 g/L), Amylase (Sigma, 1 g/L)
und Pektinase (Aspergilus niger, Fluka, 1 g/L) in Natriumacetat Lösung (pH 5, 50 mM) für 30°C und 16
Stunden. Die Reste wurden getrocknet und im Soxhlet mit Chloroform für 24-48 Stunden extrahiert.
Die Enzymbehandlung und Extraktion wurde noch zweimal wiederholt und die Reste wurden
getrocknet und zerkleinert (Fernando et al. 1984).
2.1.9 PET-Substrate für die Abbauuntersuchungen
Die Abbauuntersuchungen erfolgten an drei verschiedenen PET Substraten, welche im gleichen
Gewichtsverhältnis zum Einsatz kamen. Zu den Substraten gehörte eine APET Folie (Vertommen et al.
2005), welche 3% (w/w) Isophthalat enthielt, um amorphe Regionen in der Folie zu erzeugen. Diese
wurde bereitgestellt von Herrn Dr. V.A. Nierstrasz, Universität Gent. Das zweite Substrat waren
unbehandelte PET-Fasern, erhalten von der Firma Dr. Th. Böhme KG Geretsried, welche für die
Inkubation in etwa 1 cm große Stücke geschnitten wurden. Als drittes Substrat wurde eine zyklische
PET Trimerpräparation eingesetzt, welche mittels Soxlett-Extraktion mit Dichlormethan und 1,4-
Dioxan aus PET Stoffen gewonnen wurden. Die PET Stoffe wurden ebenfalls von der Firma Dr. Th.
Böhme KG Geretsried bezogen.
2.1.10 Puffer und Lösungen
Soweit nicht anders angegeben wurden die Substanzen in deionisiertem Wasser gelöst.
50% Acetonitril Acetonitril 500 ml/L
47.5%iges Acetonitril / 5%ige TFA Lösung Acetonitril 475 ml/L
TFA 50 ml/L
Acrylamidstammlösung Acrylamid 292 g/L
N,N´-Methylenbisacrylamid 8 g/L
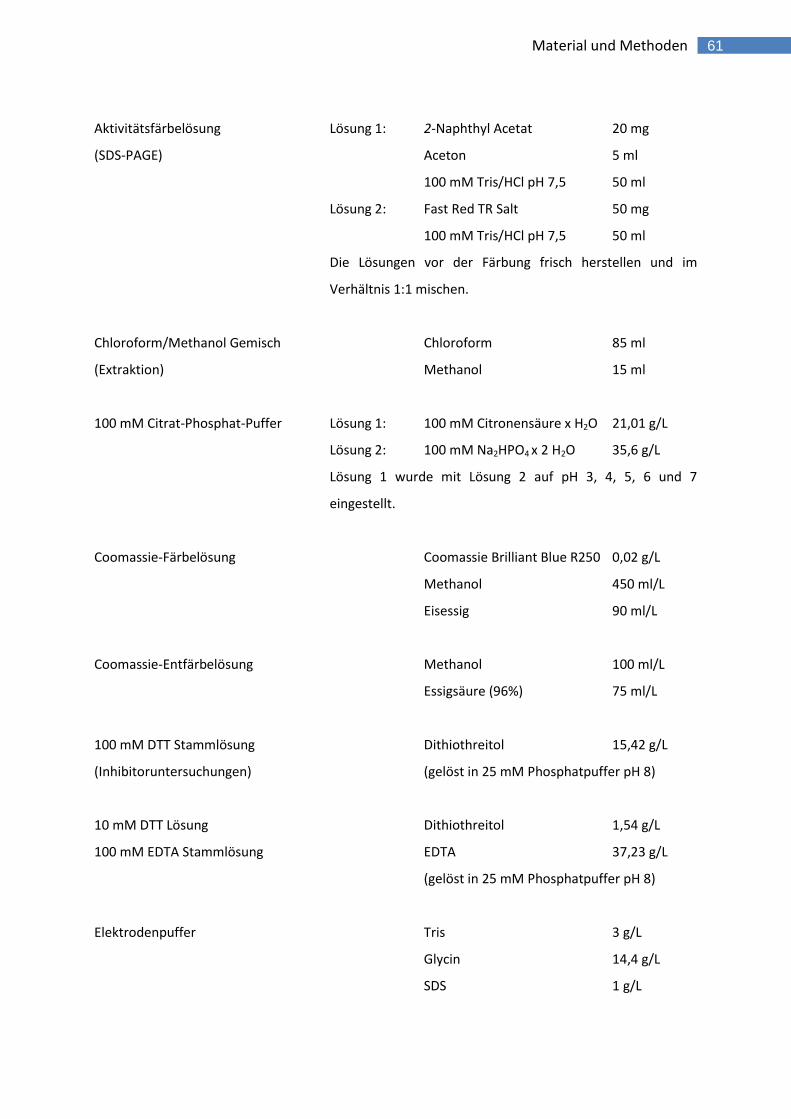
Material und Methoden 61
Aktivitätsfärbelösung Lösung 1: 2-Naphthyl Acetat 20 mg
(SDS-PAGE) Aceton 5 ml
100 mM Tris/HCl pH 7,5 50 ml
Lösung 2: Fast Red TR Salt 50 mg
100 mM Tris/HCl pH 7,5 50 ml
Die Lösungen vor der Färbung frisch herstellen und im
Verhältnis 1:1 mischen.
Chloroform/Methanol Gemisch Chloroform 85 ml
(Extraktion) Methanol 15 ml
100 mM Citrat-Phosphat-Puffer Lösung 1: 100 mM Citronensäure x H2O 21,01 g/L
Lösung 2: 100 mM Na2HPO4 x 2 H2O 35,6 g/L
Lösung 1 wurde mit Lösung 2 auf pH 3, 4, 5, 6 und 7
eingestellt.
Coomassie-Färbelösung Coomassie Brilliant Blue R250 0,02 g/L
Methanol 450 ml/L
Eisessig 90 ml/L
Coomassie-Entfärbelösung Methanol 100 ml/L
Essigsäure (96%) 75 ml/L
100 mM DTT Stammlösung Dithiothreitol 15,42 g/L
(Inhibitoruntersuchungen) (gelöst in 25 mM Phosphatpuffer pH 8)
10 mM DTT Lösung Dithiothreitol 1,54 g/L
100 mM EDTA Stammlösung EDTA 37,23 g/L
(gelöst in 25 mM Phosphatpuffer pH 8)
Elektrodenpuffer Tris 3 g/L
Glycin 14,4 g/L
SDS 1 g/L

Material und Methoden 62
100 mM Glycin-NaOH Puffer Lösung 1: 100 mM Glycin 7,51 g/L
100 mM NaCl 5,84 g/L
Lösung 2: 0,1 M NaOH
Lösung 1 wurde mit Lösung 2 auf pH 9, 10 und 11 eingestellt.
50 mM Jodacetamidlösung Jodacetamid 9,25 g/L
50 mM Natriumacetat Natriumacetat 4,1 g/L
Mit konzentrierter HCl wurde pH 5 eingestellt.
100 mM NH4HCO3 NH4HCO3 7,9 g/L
10 mM p-NP Acetat (gelöst in Ethanol absolut) p-NPA 1,811 g/L
(Lagerung bei –20°C)
10 mM p-NP Butyrat (gelöst in Ethanol absolut) p-NPB 2,09 g/L
(Lagerung bei –20°C)
10 mM p-NP Caprylat (gelöst in Ethanol absolut) p-NPC 2,65 g/L
(Lagerung bei –20°C)
10 mM p-NP Palmitat (gelöst in Ethanol absolut) p-NPP 3,78 g/L
(Lagerung bei –20°C)
100 mM Phosphatpuffer Lösung 1: NaH2PO4 x 2 H2O 15,6 g/L
Lösung 2: Na2HPO4 14,2 g/L
Lösung 2 mit Lösung 1 auf pH 8 eingestellt.
Andere Konzentrationen des Puffers wurden durch
Verdünnen der Stammlösung hergestellt.
20 mM Phosphatpuffer 1 M NaCl NaCl 58,44 g/L
20 mM Phosphatpuffer, 30% Isopropanol Isopropanol 300 ml/L
100 mM PMSF Stammlösung Phenylmethylsulfonylfluorid 17,42 g/L
(Lagerung bei 4°C) (gelöst in Isopropanol)

Material und Methoden 63
Proteinbestimmung Roti®-Quant, 5fach Konzentrat zur Proteinbestimmung
Puffer für die Kartoffelschalenbehandlung Ammoniumoxalat 16 g/L
Oxalsäure 4 g/L
Puffer für p-NPC und p-NPP Gum arabicum 1 g/L
Deoxycholat 2 g/L
(gelöst in 100 mM Phosphatpuffer pH 7,5)
Renaturierungslösung Triton X-100 0,5 ml
100 mM Tris/HCl pH 7,5 100 ml
Sammelgelpuffer Tris 60,46 g/L
SDS 4 g/L
HCl pH 6,8
Silberfärbelösungen Lösung 1: Ethanol 400 ml/L
Essigsäure (96%) 100 ml/L
Lösung 2: Ethanol 400 ml/L
Essigsäure (96%) 100 ml/L
Formaldehyd 10 ml/L
Lösung 3: Silbernitrat-Färbelösung (frisch ansetzen)
NH4OH 2 ml
0,1 N NaOH 28 ml
AgNO3 (20%) 5 ml
(tropfenweise unter ständigem Rühren
zugeben, die Lösung muss klar bleiben)
dest. Wasser 115 ml
Lösung 4: Entwickler-Lösung (frisch ansetzen)
Formaldehyd 1 ml/L
Zitronensäure 0,25 g/L
Mit NaOH den pH-Wert 7,3 einstellen.
Lösung 5: Essigsäure (96%) 50 ml/L
Lösung 6: Essigsäure (96%) 10 ml/L

Material und Methoden 64
SDS-Probenpuffer (50 ml, 5-fach) SDS 1,25 g
EDTA 18,6 mg
Bromphenolblau 2 mg
β-Mercaptoethanol 2,5 ml
Glycerin 20 ml
100 mM Tris/HCl pH 6,8 25 ml
Sporenfärbung nach Wirtz Lösung 1: Malachitgrün-Oxalat 50 g/L
Lösung 2: Safranin 5 g/L
100 mM TPCK Stammlösung Tosyl-L-phenylalanine- 35,18 g/L
(gelöst in Ethanol absolut, Lagerung bei 4°C) chloromethylketon
Trenngelpuffer Tris 181,5 g/L
SDS 4 g/L
HCl pH 6,8
0,1% Trifluoressigsäure TFA 1 ml/L
100 mM Tris/HCl Tris 12,1 g/L
Mit konzentrierter HCl wurde der gewünschte pH-Wert
eingestellt. Andere Konzentrationen des Puffers wurden
durch Verdünnen der Stammlösung hergestellt.
20 mM Tris/HCl pH 8; 0,15 M NaCl NaCl 8,77 g/L
Trypsinlösung bzw. Chymotrypsinlösung 0,5 µg/µl Protein in 1 mM HCl 2 µl
50 mM NH4HCO3 18 µl

Material und Methoden 65
2.2 Mikrobiologische Methoden
2.2.1 Stammhaltung und Kultivierung
Die Kultivierung von T. fusca KW3 erfolgte in einem 300 ml Schüttelkolben mit 50 ml Czapek-Medium
für 2 Tage bei 50°C und 100 rpm. Zur mittelfristigen Aufbewahrung bzw. Vereinzelung wurde die
Kultur auf Czapek-Agarplatten ausplattiert und ebenfalls 2 Tage bei 50°C inkubiert.
2.2.2 Mikroskopische Untersuchungen
Zellen von T. fusca wurden auf einem Objektträger in physiologischer Kochsalzlösung ausgestrichen.
Nach dem Lufttrocknen erfolgte eine Färbung der Zellen mit Malachitgrün für 10 Minuten. Nach
Entfernung der Färbelösung wurden die Objektträger einmal hitzefixiert und dann gründlich mit
demineralisiertem Wasser gespült. Die Gegenfärbung erfolgte mit Safraninlösung 15 Sekunden lang.
Nach dem Spülen mit demineralisiertem Wasser und Lufttrocknen des Objektträgers konnten die
Zellen bei einer 1000fachen Vergrößerung mit Immersionsöl mikroskopisch betrachten werden.
2.2.3 Trockengewichtsbestimmung
Für die Trockengewichtsbestimmung wurden Rundfilter verwendet, die zunächst bis zur
Gewichtskonstanz 24 Stunden im Trockenschrank bei 105°C getrocknet, im Exsikkator abgekühlt und
bis zum weiteren Gebrauch dort aufbewahrt wurden. Das Ausgangsgewicht der Filter wurde dann an
einer Feinwaage bestimmt.
3 x 250 ml Czapek-Medium wurden mit T. fusca KW3 angeimpft und 7 Tage in einem Schüttler bei
50°C und 150 rpm kultiviert. Täglich wurden 10 ml Probe entnommen. Die Zellen wurden über eine
Filternutsche und einen Rundfilter durch Anlegen eines Vakuums von der Nährlösung getrennt. Der
Rundfilter wurde anschließend mit 10 ml demineralisiertem Wasser gespült, um Reste der
Nährlösung zu entfernen. Nachdem die Flüssigkeit vollständig abgesaugt wurde, erfolgte die
Trocknung des Filters im Trockenschrank bei 105°C über Nacht. Nach dem Abkühlen im Exsikkator
wurde das Gewicht des Filters bestimmt.
Die Trockenmasse wurde aus der Differenz der Filtergewichte und dem Volumen der Probe
berechnet.

Material und Methoden 66
2.3 Proteinchemische Methoden
T. fusca KW3 wurde in 500 ml MSV-Medium mit unbehandelten PET-Fasern bei 50°C und 150 rpm
kultiviert, um eine maximale Ausbeute an Esterase zu erhalten. Aller 24 h wurden 200 µl Proben für
die Esteraseaktivitätsbestimmung entnommen (siehe 2.4.1). Die Proben wurden abzentrifugiert
(10 min, 10 000 rpm, RT) und die Überstände für den Nachweis eingesetzt. Nach 10 Tagen Inkubation
wurden die Zellen durch Zentrifugation (4000 rpm, 20 min, 4°C) entfernt. 200 µl Überstand wurden
erneut für die Esteraseaktivitätsbestimmung (siehe 2.4.1) entnommen. Der zellfreie Überstand
wurde im Gefriertrockner aufkonzentriert und bei –21°C bis zur weiteren Aufreinigung gelagert. Die
Aufreinigung erfolgte bei Raumtemperatur.
2.3.1 Proteinaufreinigung der Wildtyp TfCa
Der gefriergetrocknete Überstand wurde in einem Aliquot demineralisiertem Wasser gelöst und
gegen demineralisiertes Wasser für 5-6 Stunden dialysiert. Die Enzymlösung wurde anschließend
zentrifugiert (15 min bei 11 000 x g, 22°C) und der klare Überstand mit 100 mM Phosphatpuffer (pH
8) auf 20 mM eingestellt.
Die chromatographische Aufreinigung erfolgte mit einem Äkta Basic 10 HPLC-System. Alle
verwendeten Puffer wurden vor Benutzung filtriert und entgast, die Chromatographiesäulen wurden
zunächst mit dem Startpuffer equilibriert. Die Enzymlösung wurde auf eine Phenylsepharose 6 Fast
Flow (high sub) XK 16/20 Säule (Säulenvolumen 25 ml), equilibriert mit 20 mM Phosphatpuffer pH 8,
aufgetragen. Die Elution erfolgte mit einem Gradienten von 0 bis 100% des Elutionspuffers, 30%
Isopropanol in 20 mM Phosphatpuffer pH 8. Die aufgefangenen Fraktionen mit Esteraseaktivität
wurden vereinigt und das Isopropanol wurde durch Evaporation entfernt.
Die vereinigten Fraktionen wurden auf eine DEAE-650M XK 16/40 Säule (Säulenvolumen 54 ml),
equilibriert mit 20 mM Phosphatpuffer pH 8, aufgetragen und eluiert mit 1 M NaCl in 20 mM
Phosphatpuffer pH 8. Die Fraktionen wurden auf Esteraseaktivität getestet. Fraktionen mit
Esteraseaktivität wurden vereinigt und dialysiert gegen 20 mM Phosphatpuffer pH 8.
Die vereinigten Fraktionen wurden auf eine HiTrap™ Octyl Fast Flow Säule (Säulenvolumen 1 ml)
aufgetragen und eluiert mit 30% Isopropanol in 20 mM Phosphatpuffer pH 8. Die Fraktionen mit
Esteraseaktivität wurden vereinigt und das Isopropanol wurde durch Evaporation entfernt.
Die vereinigten Fraktionen wurden auf eine Superdex 200 prep grade XK 16/70 Säule
(Säulenvolumen 110 ml) equilibriert mit 0,15 M NaCl in 20 mM Phosphatpuffer pH 8 aufgetragen und
eluiert. Die Fraktionen wurden auf Esteraseaktivität getestet.

Material und Methoden 67
Nach jedem Reinigungsschritt wurde ein Aliquot der vereinigten Fraktionen entnommen und für die
SDS-PAGE Analyse aufbereitet.
2.3.2 Proteinaufreinigung der rekombinanten TfCa, TfCut1 und TfCut2
Von 3 PET-Hydrolasen aus T. fusca KW3 wurde das jeweilige Gen isoliert, in den Vektor pET21B(+)
eingeschleust und in E. coli BL21(DE3) transformiert. Die Proteine besaßen ein His6-Tag für die
spätere Aufreinigung. Die Produktion der rekombinanten Cutinase TfCut1 und TfCut2 und der
Carboxylesterase TfCa erfolgte bei 18°C im LB-Medium sowohl im Schüttelkolben als auch im 5 L
Fermenter. Nach Beendigung der Kultivierung wurden die Zellen mit Ultraschall aufgeschlossen und
der partikelfreie Überstand für die chromatographische Aufreinigung verwendet. Alle beschriebenen
Vorarbeiten wurden von Herrn Thorsten Oeser und Herrn Ren Wei im Rahmen ihrer Dissertationen
entwickelt und beschrieben (Oeser et al. 2010, Chen S et al. 2008).
Die Aufreinigung der rekombinanten Hydrolasen erfolgte mittels eines Äkta Basic 10 HPLC-System.
Die HisTrap™ FF Crude Säule (Säulenvolumen 2 x 5 ml) wurde zunächst mit einem 50 mM
Phosphatpuffer mit 300 mM NaCl pH 8 equilibriert und anschließend der partikelfreie Überstand
aufgetragen. Es erfolgte eine Gradientenelution (10 Säulenvolumen) bis 100% 50 mM
Phosphatpuffer mit 300 mM NaCl und 250 mM Imidazol pH 8. Die Fraktionen wurden auf
Esteraseaktivität getestet. Fraktionen mit Esteraseaktivität wurden vereinigt und auf eine Superdex
200 prep grade XK 26/60 Säule (Säulenvolumen 320 ml) equilibriert mit 0,15 M NaCl in 20 mM
Phosphatpuffer pH 8 aufgetragen und eluiert. Die Fraktionen wurden auf Esteraseaktivität getestet.
Nach jedem Reinigungsschritt wurde ein Aliquot der vereinigten Fraktionen entnommen und für die
SDS-PAGE Analyse aufbereitet.
2.4 Analytische Methoden
2.4.1 Esterase-Aktivitätsbestimmung
Bei diesem Nachweis wird das nach enzymatisch katalysierter Spaltung von chromogenen para-
Nitrophenyl-Acylestern freigesetzte gelbe para-Nitrophenol bei 410 nm spektrophotometrisch
bestimmt (Purdy und Kolattukudy 1973). Ein Unit (U) ist definiert als die Menge an Enzym, die 1 µmol
p-NPB pro Minute freisetzt.

Material und Methoden 68
2.4.1.1 Bestimmung mittels Spektrophotometer
Zu 940 µl 100 mM Phosphatpuffer pH 7,5 erfolgte die Zugabe von 10 µl 10 mM p-Nitrophenylbutyrat
als Substrat. Die Reaktion wurde durch Zugabe von 50 µl Enzymlösung gestartet, wobei die Messung
30 Sekunden danach im Cary 50 Bio UV-Visible Spektrophotometer bei 410 nm für eine Minute
erfolgte. Die Enzymaktivität wurde in U/ml angegeben.
2.4.1.2 Bestimmung mittels Plattenleser
Zu 80 µl 100 mM Phosphatpuffer pH 7,5 erfolgte die Zugabe von 10 µl 5 mM p-Nitrophenylbutyrat als
Substrat. Die Reaktion wurde durch Zugabe von 10 µl Enzymlösung gestartet, wobei die Messung 10
Sekunden danach im Plattenleser PowerWave XS bei 410 nm für fünf Minuten erfolgte. Die
Enzymaktivität wurde in U/ml angegeben. Die Messung des Blindwerts erfolgte mit 10 µl Puffer
anstatt der Enzymlösung.
2.4.2 Cutinase-Aktivitätsbestimmung
Bei diesem Nachweis werden die durch die enzymatisch katalysierte Spaltung des Biopolyesters Cutin
entstehenden Monomere, meist C16 und C18 Hydroxyfettsäuren, durch eine
Gaschromatographische Analyse nachgewiesen (Chen S et al. 2008). Das Enzym wurde mit 10 mg
Cutinpräparation in 500 µl Phosphatpuffer (25 mM, pH 8) bei 50°C 72 Stunden inkubiert. 500 µl
Eisessig wurden dem Reaktionsgemisch zugesetzt und die freigesetzten Cutinmonomere mit 3 ml
Chloroform nach kräftigem Schütteln und Zentrifugation (2000 rpm, 10 min, RT) extrahiert und unter
Stickstoffbegasung eingedampft. Die Abbauprodukte wurden in 1 ml Chloroform/Methanol-Gemisch
(85:15) aufgenommen, in GC Vials überführt und unter Stickstoff erneut getrocknet. Die Produkte
wurden mit MSTFA syliliert und analysiert per GC-MS. Zum Vergleich wurde eine Negativkontrolle
der Reaktion ohne Enzymzugabe mit inkubiert und detektiert.
Als zusätzlicher Vergleich wurde Cutin chemisch hydrolysiert und die entstandenen Monomere
ebenfalls wie oben beschrieben aufgenommen, derivatisiert und per GC-MS analysiert. Die
chemische Hydrolyse erfolgte zum einen von 50 mg Cutin in 30 ml Tetrahydrofuran mit 2,5-fachem
Gewichtsüberschuss von LiAlH4. Die Lösung wurde für 72 Stunden bei Raumtemperatur schüttelnd
inkubiert und anschließend mit 100 ml dest. Wasser versetzt. Die erhaltene Lösung wurde mit 4 ml
HCl angesäuert und mit Di-Ethylether die entstandenen Monomere extrahiert (5 x 150 ml). Die
organische Phase wurde mit wasserfreiem Natriumsulfat von verbleibenden Wasserresten befreit
und unter Vakuum vollständig getrocknet (Walton und Kolattukudy 1972). Eine weitere Hydrolyse

Material und Methoden 69
von 50 mg Cutin erfolgte in 30 ml 95% Ethanol alkalisch mit 3 g Kaliumhydroxid. Die Lösung wurde
unter Stickstoffatmosphäre 72 Stunden bei Raumtemperatur schüttelnd inkubiert. Das
Reaktionsgemisch wurde mit 4 ml HCl angesäuert und die entstandenen Cutinmonomere mit Di-
Ethylether extrahiert (5 x 100 ml). Die organische Phase wurde mit wasserfreiem Natriumsulfat von
verbleibenden Wasserresten befreit und unter Vakuum vollständig getrocknet (Gerard et al. 1992).
2.4.3 PCL-Abbauuntersuchung
Poly(ε-caprolacton) gilt als Modellsubstrat für Cutinasen und wurde zur genaueren Spezifikation der
untersuchten Enzyme eingesetzt. Die Untersuchung des PCL Abbaus erfolgte als Screening Methode
auf Agarplatten und zur Bestimmung von Abbauraten mittels Plattenleser.
2.4.3.1 Bestimmung mittels Hofbildung
Zunächst wurden in die Agarplatten (18 g/L Agar in 25 mM Phosphatpuffer pH 8, nach dem
Autoklavieren wurden 50 ml/L 1%ige PCL Stammlösung in Aceton zugegeben und die Platten
gegossen) ein Loch gestanzt, in das die Enzymlösung gegeben wurde. Die Platten wurden bei
optimaler Temperatur 24 Stunden inkubiert und die Hofbildung anschließend ausgewertet.
2.4.3.2 Bestimmung mittels Plattenleser
Zunächst wurden 0,5 ml PCL Stammlösung (0,27 g PCL in 50 ml Aceton) in 9,5 ml 100 mM
Phosphatpuffer pH 7,5 frisch durch mehrfaches Durchmischen suspendiert. Die Reaktion wurde
durch Zugabe von 10 µl Enzymlösung gestartet, wobei die Messung der Klärung 10 Sekunden danach
im Plattenleser PowerWave XS bei 600 nm für 10 Minuten erfolgte. Die Enzymaktivität wurde als
Abbaurate in mg min-1 ml-1 angegeben. Die Messung des Blindwerts erfolgte mit 10 µl Puffer anstatt
der Enzymlösung.
2.4.4 Quantitative Protein-Bestimmung nach Bradford (1976)
Die Bestimmung der Proteinkonzentration erfolgte mit einer Roti-Quant® Lösung. 200 µl der Roti-
Quant® Lösung wurden auf 800 µl der zu bestimmenden Proteinlösung pipettiert, gevortext und fünf
Minuten inkubiert. Die Analyse erfolgte spektrophotometrisch bei 595 nm und die Konzentration mit
einer zuvor erstellten Kalibrierkurve mit BSA als Protein berechnet.
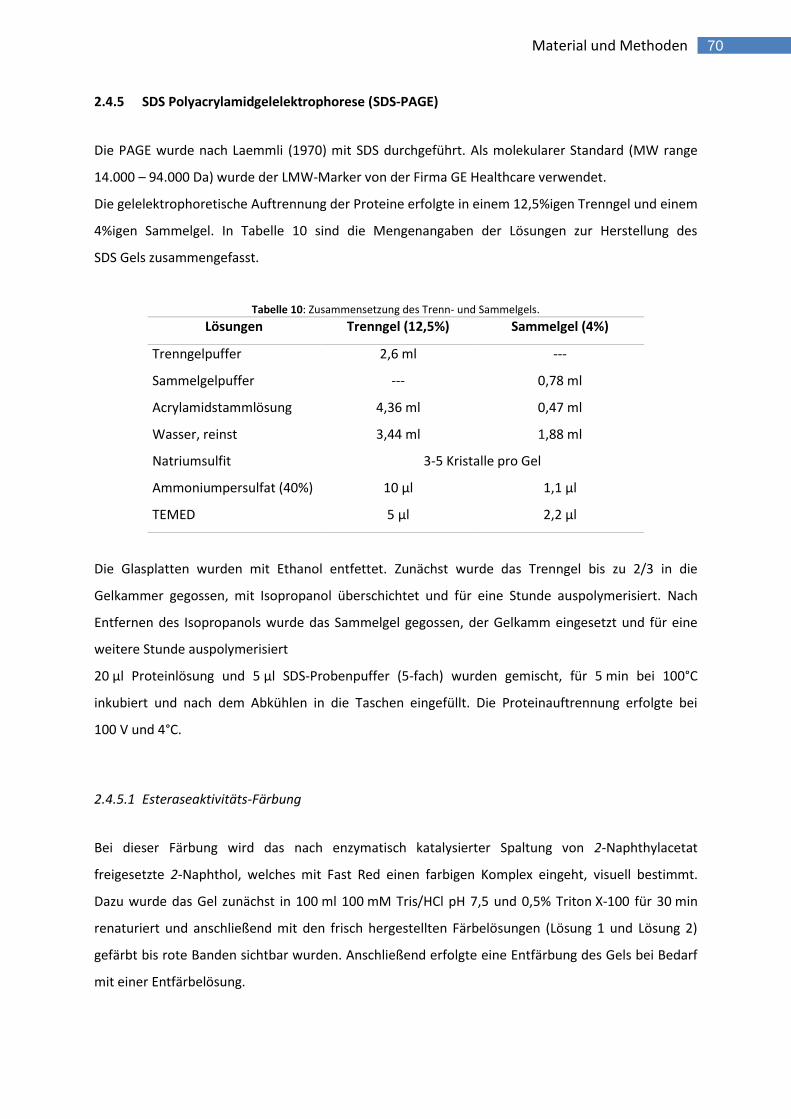
Material und Methoden 70
2.4.5 SDS Polyacrylamidgelelektrophorese (SDS-PAGE)
Die PAGE wurde nach Laemmli (1970) mit SDS durchgeführt. Als molekularer Standard (MW range
14.000 – 94.000 Da) wurde der LMW-Marker von der Firma GE Healthcare verwendet.
Die gelelektrophoretische Auftrennung der Proteine erfolgte in einem 12,5%igen Trenngel und einem
4%igen Sammelgel. In Tabelle 10 sind die Mengenangaben der Lösungen zur Herstellung des
SDS Gels zusammengefasst.
Tabelle 10: Zusammensetzung des Trenn- und Sammelgels.
Lösungen Trenngel (12,5%) Sammelgel (4%)
Trenngelpuffer 2,6 ml ---
Sammelgelpuffer --- 0,78 ml
Acrylamidstammlösung 4,36 ml 0,47 ml
Wasser, reinst 3,44 ml 1,88 ml
Natriumsulfit 3-5 Kristalle pro Gel
Ammoniumpersulfat (40%) 10 µl 1,1 µl
TEMED 5 µl 2,2 µl
Die Glasplatten wurden mit Ethanol entfettet. Zunächst wurde das Trenngel bis zu 2/3 in die
Gelkammer gegossen, mit Isopropanol überschichtet und für eine Stunde auspolymerisiert. Nach
Entfernen des Isopropanols wurde das Sammelgel gegossen, der Gelkamm eingesetzt und für eine
weitere Stunde auspolymerisiert
20 µl Proteinlösung und 5 µl SDS-Probenpuffer (5-fach) wurden gemischt, für 5 min bei 100°C
inkubiert und nach dem Abkühlen in die Taschen eingefüllt. Die Proteinauftrennung erfolgte bei
100 V und 4°C.
2.4.5.1 Esteraseaktivitäts-Färbung
Bei dieser Färbung wird das nach enzymatisch katalysierter Spaltung von 2-Naphthylacetat
freigesetzte 2-Naphthol, welches mit Fast Red einen farbigen Komplex eingeht, visuell bestimmt.
Dazu wurde das Gel zunächst in 100 ml 100 mM Tris/HCl pH 7,5 und 0,5% Triton X-100 für 30 min
renaturiert und anschließend mit den frisch hergestellten Färbelösungen (Lösung 1 und Lösung 2)
gefärbt bis rote Banden sichtbar wurden. Anschließend erfolgte eine Entfärbung des Gels bei Bedarf
mit einer Entfärbelösung.

Material und Methoden 71
2.4.5.2 Coomassie-Färbung
Proteine im Gel wurden mit Coomassie Brilliant Blau R-250 gefärbt (Weber und Osborn 1969) und die
Entfärbung erfolgte mit einer frisch hergestellten Entfärbelösung.
2.4.5.3 Silberfärbung der Proteine
Die Silberfärbung der Proteine wurde nach Kleber, Schlee und Schöpp (1997) durchgeführt. Der
Zeitliche Ablauf der Färbung ist in Tabelle 11 dargestellt.
Tabelle 11: Zeitlicher Ablauf der Silberfärbung.
Schritt Färbezeit
Gel mit Lösung 1 schwenken 2 x 1 Stunde
Gel mit Lösung 2 schwenken 1 Stunde
Gel mit demineralisiertem Wasser schwenken 3 x 40 Minuten
Gel mit Lösung 3 schwenken 15 Minuten
Gel mit demineralisiertem Wasser schwenken 3 x 10 Minuten
Entwicklung in Lösung 4 bis zur Bildung
von braunen Flecken auf klarem Grund
Abstoppen mit Lösung 5
Lagerung in Lösung 6
2.4.6 Bestimmung des pI
Die Bestimmung des isoelektrischen Punktes der gereinigten Carboxylesterase erfolgte mit dem
PhastSystemTM Gelelektrophorese-System mit einem PhastGel® IEF 3-9 Gel und IEF 4-6,5 und dem
Kalibrierkit für pI Bestimmung durch Isoelektrische Fokusierung, Broad range pI (pH 3-10).
Das Protein wurde in verschiedenen Konzentrationen auf das Gel aufgetragen und fokusiert (IEF 3-9:
2000 V; 2,5 A; 3,5 W; 15°C; 410 Vh; IEF 4-6,5: 2000 V; 5,0 mA; 3,5 W; 15°C; 410 Vh; PhastSystemTM).
Nach der Ausrichtung des Proteins wurden die Gele mittels Silberfärbung angefärbt. Die Standard-
Proteine zur Berechnung des pI waren Amyloglucosidase (pI 3,5), Trypsininhibitor (pI 4,55), β-
Lactoglobulin A (pI 5,20), Bovine carbonic anhydrase B (pI 5,85), Human carbonic anhydrase B (pI
6,55), Myoglobin-acid band (pI 6,85), Myoglobin-base band (pI 7,35), Lentil lectinacidic band (pI
8,15), Lentil lectin-middle band (pI 8,45), Lentil lectin-basic band (pI 8,65) und Trypsinogen (pI 9,3).

Material und Methoden 72
2.4.7 Bestimmung der Molaren Masse
Für die Bestimmung des Molekulargewichtes der extrazellularen Esterase wurden die Enzymlösung
und der SDS Elektrophorese Standard Roti®-Mark mit einer SDS-PAGE aufgetrennt und mit dem Gel
erfolgte eine Aktivitäts- und Coomassie-Färbung. Die Laufstrecken der Proteine des Standards und
der Esterase wurden bestimmt. Die bestimmten Laufstrecken der Standards wurden gegen den
Logarithmus des Molekulargewichts der Proteine aufgetragen und somit eine Formel zur Berechnung
des Molekulargewichts erhalten. Damit konnte das Molekulargewicht der Esterase berechnet
werden.
2.4.8 Bestimmung der Temperatur und pH-Wert Stabilität
Zur Bestimmung der pH Stabilität erfolgte die Inkubation der Esterase bei 23°C für 7 Tage in Puffern
von pH 3 bis 11. Citrat-Phosphat-Puffer wurde für pH 3 bis 7 verwendet, Phosphatpuffer für pH 8 und
Glycin-NaOH Puffer für pH 9 bis 11. Die Esteraseaktivität wurde mit dem Esteraseaktivitätsmessung
bestimmt. Die Bestimmung der Temperaturstabilität der Esterase erfolgte bei Temperaturen von 30
bis 55°C in einem 25 mM Phosphatpuffer für 7 Tage. Die Esteraseaktivität wurde mit dem
Esteraseaktivitätsmessung bestimmt.
2.4.9 Bestimmung der Stabilität gegenüber Inhibitoren
Die Esterase wurde mit den Inhibitoren EDTA, Phenylmethylsulfonylfluoride (PMSF), Tosyl-L-
phenylalanine-chloromethylketon (TPCK) und Dithiothreitol (DTT) bei 30°C für 15 min inkubiert. EDTA
wurden in den Konzentrationen 1 mM und 10 mM und DTT mit 0,1 mM; 1 mM und 10 mM in 25 mM
Phosphatpuffer pH 8 eingesetzt. PMSF wurde, in Methanol gelöst, mit einer Endkonzentration von
0,1 mM; 1 mM und 10 mM eingesetzt. TPCK, gelöst in Isopropanol, wurde mit den
Endkonzentrationen 0,1 mM; 1 mM und 10 mM verwendet. Die Esteraseaktivität wurde mit dem
Esteraseaktivitätsnachweis mit p-NPB als Substrat bei 410 nm spektrophotometrisch bestimmt.
2.4.10 Bestimmung der kinetischen Konstanten für die Hydrolyse von p-NP Ester
Die Substratspezifität gegenüber p-NP Estern und Triacylglyceride mit verschiedener Kettenlänge
wurde bestimmt. Die Aktivität gegenüber den p-NP Estern wurde spektrophotometrisch bei 410 nm
mittels Esteraseaktivitäts-Assay determiniert.

Material und Methoden 73
2.4.11 Bestimmung der kinetische Konstanten für die Hydrolyse von CTR
Die kinetischen Konstanten für den Abbau von zyklischen PET-Trimeren wurden bestimmt. Die
Trimere, gelöst in 1,4-Dioxan, wurden in 25 mM Phosphatpuffer pH 8 gelöst und es erfolgte eine
Inkubation mit der Esterase bei 50°C für einer Stunde. Die entstandenen Abbauprodukte wurden
mittels Reversed Phase HPLC aufgetrennt und bei 241 nm spektrometrisch detektiert.
2.4.12 Bestimmung optimaler Temperatur und pH-Wert für die Hydrolyse von CTR
Der optimale pH-Wert wurde bestimmt mit einem Reaktionsgemisch gelöst in Citrat-Phosphat-Puffer
für pH 3 bis 7, Phosphatpuffer für pH 8 oder Glycin-NaOH Puffer für pH 9 bis 11. Alle Puffer besaßen
eine Ionenstärke von 100 mM und der Abbau erfolgte bei 50°C. Der Puffer wurde vortemperiert bei
50°C, das Substrat, zyklische PET Trimere gelöst in 1,4-Dioxan, wurde vor Beginn der Reaktion in den
Puffer gegeben und gemischt. Die Reaktion wurde durch Zugabe der Esterase gestartet. Die
Inkubation erfolgte für eine Stunde und die gebildeten Produkte wurden per Reversed Phase HPLC
aufgetrennt und bei 241 nm detektiert.
Die optimale Temperatur wurde bestimmt mit zyklischen Trimeren, gelöst in 1,4-Dioxan, als Substrat
und dem optimalen pH-Wert, welcher zuvor bestimmt wurde. Das Reaktionsgemisch wurde in
25 mM Phosphatpuffer pH 8 suspendiert und der Abbau erfolgte bei 30 bis 55°C. Der Puffer wurde
vortemperiert, das Substrat zugegeben und gemischt. Durch die Zugabe der Esterase erfolgte der
Start des Abbaus, welcher eine Stunde inkubiert wurde. Die gebildeten Produkte wurden per
Reversed Phase HPLC aufgetrennt und bei 241 nm detektiert.
2.4.13 N-terminale Sequenzierung
Die N-terminale Sequenzierung der Esterase erfolgte durch die Arbeitsgruppe Biochemie und
Bioorganische Chemie am Institut für Biochemie, Fakultät für Biowissenschaften, Pharmazie und
Psychologie der Universität Leipzig.
Dafür wurde die Esterase in einem 12,5% SDS Gel aufgetrennt, auf eine PVDF Membran geblottet
und mit Coomassie Brilliant Blue angefärbt. Die entsprechende Bande wurde ausgeschnitten, halbiert
und in den Sequenzer gegeben. Es wurden die ersten 10 Aminosäuren des Proteins sequenziert.

Material und Methoden 74
2.4.14 MALDI-TOF Sequenzierung
Die Esterase wurde im 12,5%igen SDS-Gel aufgetrennt und mit Coomassie Brilliant Blue angefärbt.
Die entsprechende Proteinbande wurde aus dem Gel ausgeschnitten, im 1 mm große Stücke geteilt
und mit 100 µl Aqua bidest. 10 min bei Raumtemperatur geschüttelt. Nach Entfernen des Wassers
wurde 100 µl 50% Acetonitril zugegeben und der Ansatz nochmals 10 min bei Raumtemperatur
geschüttelt. Dieser Schritt wurde einmal wiederholt. Der Überstand wurde abgenommen und es
erfolgte eine Inkubation der Gelstücke in 80 µl 100% Acetonitril bei Raumtemperatur für 5 min unter
Schütteln. Danach erfolgte die Zugabe von 80 µl 100 mM NH4HCO3. Nach 10 min Schütteln bei
Raumtemperatur wurde der Überstand komplett entfernt und die Gelstücke im einem
Vakuumkonzentrator getrocknet. Die getrockneten Gelstücke wurden zur Reduktion und Alkylierung
in 80 µl frisch hergestellter 10 mM DTT Lösung für 45 min bei 56°C inkubiert. Nach dem Abkühlen
wurde der Überstand abgenommen und sofort 80 µl 50 mM Jodacetamidlösung zugeben (Inkubation
im Dunkeln für 30 min). Der Überstand wurde abgenommen, 80 µl 100 mM NH4HCO3 Lösung
zugegeben und 5 min bei Raumtemperatur geschüttelt. Anschließend wurden 80 µl 100%iges
Acetonitril zugegeben und 10 min bei Raumtemperatur geschüttelt. Der Überstand wurde entfernt
und die Gelstücke im Vakuumkonzentrator komplett getrocknet. Zu den Gelbanden wurde 6,6 µl
Trypsinlösung bzw. Chymotrypsinlösung gegeben und anschließend mit 50 mM NH4HCO3 Lösung
überschichtet. Der enzymatische Verdau erfolgte bei 37°C über Nacht. Um die verdauten
Proteinbruchstücke aus dem Gel zu eluieren, wurde 50 µl 47,5%iges Acetonitril / 5%ige TFA Lösung
zugegeben (Schütteln bei RT für 10 min). Der erhaltene Überstand wurde aufgehoben und der Schritt
noch zweimal wiederholt. Die erhaltenen Überstände wurden vereinigt, eingefroren und mit dem
Vakuumkonzentrator auf etwa 5 bis 10 µl aufkonzentriert. Die Lösung wurde anschließend entsalzt
und für die Auftrennung der Bruchstücke auf eine Reversed Phase HPLC mit Nanopumpe
aufgetragen. Die Auftrennung und die anschließende MALDI-TOF Analyse erfolgte in der
Arbeitsgruppe Biochemie und Bioorganische Chemie am Institut für Biochemie, Fakultät für
Biowissenschaften, Pharmazie und Psychologie der Universität Leipzig.
Die durch die LC aufgetrennten Proteinfragmente wurden auf Trägerplatten gespottet und jeder
einzelne per MALDI-TOF analysiert und anschließend elektronisch ausgewertet.
2.4.15 Abbaustudien
Für die Abbauuntersuchungen wurden die gleichen Mengen an APET Folie, unbehandelten PET-
Fasern und zyklischen PET-Trimeren in 10 ml Phosphatpuffer suspendiert und je 40 U der PET-
Hydrolasen bzw. 25 mg der rekombinanten PET-Hydrolasen aus T. fusca KW3 zugegeben. Die

Material und Methoden 75
Enzymaktivität wurde mit p-NPB als Substrat spektrophotometrisch bestimmt. Die Kolben wurden
verschlossen um einer Evaporation der Flüssigkeit vorzubeugen und die Inkubation erfolgte bei 30°C
für die Cutinasen aus F. solani pisi (Vertommen et al. 2005, Chen et al. 2010); 50°C für die TfCa,
rekombinante TfCa, Cutinase 1 aus T. fusca WSH03-11, Cutinase 2 aus T. fusca WSH03-11 und die
Cutinase-Präparation von Novozymes; 60°C für die rekombinanten TfCut1 und TfCut2 für 74 Stunden
(150 rpm). Es wurden zu verschiedenen Zeiten Proben genommen und per Umkehrphasen HPLC
analysiert. Nur die Proben vom Trimer-Abbau wurden nach der Probennahme zentrifugiert
(9000 rpm, 15 min) und anschließend wurden bei allen Proben der pH-Wert mit 1 M HCl auf 5 bis 6
eingestellt.
Die APET-Folie wurde nach der Inkubation mit der TfCa und der Cutinase aus F. solani pisi
(Vertommen et al. 2005) aus der Pufferlösung entnommen, mit demineralisiertem Wasser
gewaschen, mit einer 1%igen CHAPS-Lösung in Phosphatpuffer zur Entfernung der verbliebenen
Proteinreste von den Oberflächen der Substrate bei 50°C und 150 rpm für 2 Stunden inkubiert und
mit demineralisiertem Wasser gewaschen. Die Modifikationen der Oberflächen der Substrate durch
die Inkubation mit den Enzymen wurden mit SEM analysiert.
2.4.16 Analytik der PET Abbauprodukte
Die Analyse der Proben der Abbauuntersuchungen erfolgte wie bei Vertommen et al. (2005)
beschrieben, jedoch mit folgenden Änderungen: Die Eluenten der polaren mobile Phase waren 0,1%
TFA in Wasser (A) und Acetonitril (B) und das Injektionsvolumen der Proben war bei dem
Trimerabbau 10 µl und bei den anderen Abbauexperimenten 30 µl.
Es wurden Kalibrierkurven für die Standards Terephthalsäure und Bis(2-hydroxyethyl)terephthalat
(BHET) angefertigt.
Die detektierten Produktpeaks wurden ebenfalls per LC-MS analysiert. Diese Messungen wurden von
der AG Massenspektrometrie am Institut für Analytische Chemie, Universität Leipzig durchgeführt.
2.4.17 Konzentrationabhängige CTR-Abbaustudien
Die Enzyme wurden mit den verschiedenen Konzentrationen für jeweils 4 und 8 Stunden mit der
gleichen Menge an CTR (15 mg/0,5 ml CTR; entspricht einer Konzentration von 30 mg/ml) bei 50°C
(rTfCa) und 60°C (rTfCut) bei 150 rpm inkubiert. Nach der Inkubation wurden die Proben wie bereits
beschrieben für die RP-HPLC Analyse vorbereitet und analysiert (Billig et al. 2010, Vertommen et al.
2005).

Material und Methoden 76
Bei den eingesetzten Enzymlösungen wurde zusätzlich die Esterase-Aktivität mit p-NPB als Substrat
bestimmt.
2.5 Homologie-Modelling der TfCa und weitere PET-Hydrolasen
Die Erstellung des Homologie-Modells der TfCa erfolgte mit dem Programm PHYRE unter der
Verwendung der thermostabilen Carboxylesterase aus G. stearothermophilus (PDB code 2OGS resp.
2OGT) als Vorlage (Bennett-Lovsey et al. 2008, Liu et al. 2007). Eine Plausibilitätsprüfung des
endgültigen Modells durchgeführt mit dem Programm VERIFY3D zeigte bei 83% der Aminosäuren
einen Messwert über 0,2, was die Übereinstimmung des Modells bestätigte (Eisenberg et al. 1997).
Die modellierte Struktur der Cutinase Tfu_0882 aus T. fusca YX basierte auf der
Streptomyces exfoliates Lipase (SeL) und wurde mittels dem SWISS-Model homology modeling web
server erstellt (Chen S et al. 2008, Schwede et al. 2003). Das Modell der Tfu_0883 Cutinase aus
T. fusca YX wurde mittels PHYRE erstellt mit der SeL als Vorlage (Bennett-Lovsey et al. 2008, Chen S
et al. 2008, Liu et al. 2007).
Die Strukturen der Cutinasen aus F. solani sp. pisi und P. mendocina wurden aus der NCBI
Proteindatenbank entnommen (http://www.ncbi.nlm.nih.gov, Longhi et al. 1997, Bott et al. 2003).
Die Modellierung der Cutinase aus H. insolens erfolgte mit dem WURST protein threading server
(Sandal et al. 1996, Torda et al. 2004).
Die dargestellten Abbildungen der Proteinstrukturen wurden mittels PyMOL erstellt (DeLano 2002).

Ergebnisse und Diskussion 77
3 Ergebnisse und Diskussion
3.1 Screening nach PET-Hydrolasen aus T. fusca
3.1.1 Wachstum von T. fusca KW3 im Czapek-Medium
Für den Einsatz von T. fusca KW3 zur Esteraseproduktion mussten zunächst Zellen in einem
Vollmedium, dem Czapek-Medium, angezogen werden, welche dann anschließend im
Enzymproduktionsmedium, dem Mineralsalzmedium (MSV-Medium), das gewünschte Enzym in
größeren Mengen synthetisierten. Damit die Zellen sich in der exponentiellen Phase beim
Überimpfen befanden, erfolgte die Aufnahme einer Wachstumskinetik des Actinomyceten
T. fusca KW3 im Komplexmedium. Dafür wurden drei 250 ml Czapek-Medien mit dem Actinomyceten
angeimpft und für 7 Tage bei 50°C und 150 rpm kultiviert. Täglich wurden 10 ml Probe genommen,
die Zellen von der Nährlösung abgetrennt, gewaschen und bei 105°C über Nacht getrocknet. Nach
dem Abkühlen der Filter im Exsikkator wurde ihr Gewicht bestimmt und die Trockenmasse aus der
Differenz der Filtergewichte und dem Volumen der Probe berechnet.
Abbildung 25: Wachstumskurve von T. fusca KW3 im Czapek-Medium bei 50°C und 150 rpm.
Die Wachstumskurve von T. fusca KW3 im Czapek-Medium wurde als Biotrockenmasse (mg/ml) über
die Zeit (Tage) dargestellt (Abbildung 25). In den ersten beiden Tagen erfolgte maximales
Zellwachstum in der exponentiellen Phase. Vom zweiten bis vierten Tag folgte die statische Phase,
0
0,5
1
1,5
2
2,5
3
3,5
4
0 1 2 3 4 5 6 7
Bio
tro
ck
en
ma
ss
e (
mg
/ml)
Kultivierungsdauer (d)

Ergebnisse und Diskussion 78
bei der die Neubildungsrate gleich der Absterberate aufgrund von gebildeten toxischen
Nebenprodukten und mangelnden Nährstoffen ist. In dieser Phase werden Sekundärprodukte, wie
unter anderem Antibiotika, gebildet. Als letzte Phase schloss sich die Absterbephase an, bei der
durch Zelllyse eine deutliche Abnahme der Biomasse erfolgt. Bei T. fusca KW3 im Czapek-Medium
konnte hingegen nur ein leichter Abfall der Biotrockenmasse registriert werden. Die Kultivierung der
Vorkultur für die spätere Enzymproduktion erfolgte, abgeleitet aus der oben dargestellten
Wachstumskurve, in 2 Tagen.
Für die Enzymproduktion im Minimalmedium ist es wichtig, möglichst wenig Saccharose aus dem
Czapek-Medium mit dem Inoculum in das Produktionsmedium einzutragen, da Glucose ein Hemmer
der Esteraseproduktion ist. Deshalb wurden die Zellen nach der sterilen Ernte und erfolgter
Zentrifugation in sterilem 25 mM Phosphatpuffer (pH 8) des gleichen Volumens aufgenommen.
3.1.2 Wachstum von T. fusca KW3 im MSV-Medium
3.1.2.1 Esterasebildung mit verschiedenen synthetischen und natürlichen Polyestern
Als Mediumzusatz für eine Esteraseproduktion wurden folgende Polyester-Oligomere und Polymere
im MSV-Medium eingesetzt (je 2 g/L): recyceltes PET-Granulat, eine PET-Oligomerpräparation, eine
zyklische PET-Oligomerpräparation, BHET, PBT-Fasern und Präparationen der natürlichen
Biopolyester Cutin und Suberin. Dazu wurden die MSV-Medien mit den im Czapek-Medium 2 Tage
vorkultivierten Zellen angeimpft. Die Kultivierung erfolgte über einen Zeitraum von 10 Tagen.
Anschließend wurden die Kulturüberstände gewonnen und Proben davon auf ein SDS Gel
aufgetragen und elektrophoretisch aufgetrennt. Darüber hinaus wurde eine Aktivitätsfärbung mit
Fast Red und 2-Naphtylacetat durchgeführt.
Sowohl unter Einsatz von PET-Fasern, recyceltem PET-Granulat, der PET-Oligomerpräparation und
PBT-Fasern konnte die Bildung einer ca. 52 kDa großen Esterase gelelektrophoretisch nachgewiesen
werden (Abbildung 26, Spur 1, 2, 3 und 6). Die zellfreien Überstände unter Verwendung der
zyklischen PET Oligomerpräparation bzw. BHET zeigten nur eine schwache bzw. keine Bande im SDS-
PAGE (Abbildung 26, Spur 4 und 5). Mit diesen Substraten wurde nur wenig bzw. keine Esterase
produziert. Hingegen konnte mit Cutin und Suberin als Substrat keine 52 kDa Esterase identifiziert
werden, aber dafür eine kleinere, ca. 29 kDa große Esterase, allerdings in verhältnismäßig großen
Mengen (Abbildung 26, Spur 7,8).

Ergebnisse und Diskussion 79
Abbildung 26: SDS-PAGE und Aktivitätsfärbung mit den Esterasen in den zellfreien Überständen des MSV-Mediums nach 10 tägiger Inkubation bei 50°C mit PET-Fasern (Spur 1), recyceltes PET-Granulat (Spur 2), einer PET-Oligomerpräparation (Spur 3), einer zyklischen PET-Oligomerpräparation (Spur 4), BHET (Spur 5), PBT-Fasern (Spur 6), einer Suberinpräparation (Spur 7) und einer Cutinpräparation (Spur 8).
Ausgehend von der Zielstellung der Arbeit, eine PET-Trimere abbauende Esterase zu identifizieren,
aufzureinigen und zu charakterisieren, stand die durch Kultivierung unter Verwendung der Substrate
PET-Fasern, recyceltes PET-Granulat und PET-Oligomerpräparation identifizierte 52 kDa große
Esterase zunächst im Mittelpunkt der nachfolgenden Untersuchung. Inwieweit die durch Induktion
mit der Suberin bzw. Cutinpräparation identifizierte 29 kDa große Esterase auch PET abbauen kann,
wird zu einem späteren Zeitpunkt untersucht.
Im nächsten Schritt wurden die gebildeten Esteraseaktivitäten mittels p-NPB Aktivitätsbestimmung
während 10-tägiger Kultivierung im MSV-Medium mit PBT-Fasern, PET-Fasern, recyceltem PET-
Granulat, einer PET-Oligomerpräparation, einer zyklischer PET-Oligomerpräparation bzw. BHET
untersucht. Die Entwicklung der bestimmten Aktivitäten ist in Abbildung 27 graphisch dargestellt und
die maximal produzierten Aktivitäten sind in Tabelle 12 zusammengefasst. Die höchste
Esteraseaktivität von 24 mU/ml wurde mit PET-Fasern als Additiv erreicht, wobei während der
10 Tage Kultivierung die Enzymaktivität stetig zunahm. Unter Einsatz von recyceltem PET-Granulat,
PBT-Fasern, einer PET-Oligomerpräparation, einer zyklischer PET-Oligomerpräparation und BHET
konnten ebenfalls Esteraseaktivitäten gemessen werden, aber diese waren im Vergleich zur
Enzymaktivität aus dem MSV-Medium mit PET-Fasern deutlich geringer.
Obwohl beim Einsatz von BHET als Mediumzusatz in der SDS-PAGE (Abbildung 26) keine 52 kDa
große Bande im zellfreien Überstand nachzuweisen war, wurde auch hier nach 3 Tagen Kultivierung
eine Esteraseaktivität gemessen, die mit 21 mU/ml nur etwas geringer war als bei Verwendung von
PET-Fasern als Induktor (24 mU/ml). Eine Erklärung, warum die Bande in der SDS-PAGE nach
10 Tagen Kultivierung mit BHET nicht nachzuweisen war, ist, dass zu diesem Zeitpunkt bereits keine
52 kDa Esterase
29 kDa Esterase
1 2 3 4 5 6 7 8

Ergebnisse und Diskussion 80
detektierbare Esterasemenge mehr vorlag. Wahrscheinlich erfolgte mit BHET nur eine Produktion bis
zum 3. Tag der Kultivierung und anschließend wurde das Enzym inaktiviert oder abgebaut, was auch
mit dem Kurvenverlauf in Abbildung 27 übereinstimmen würde.
Abbildung 27: Esteraseproduktion durch T. fusca KW3 mit recyceltem PET-Granulat, PET-Oligomerpräparation, zyklische PET-Oligomerpräparation, BHET, PBT und PET-Fasern über einen Zeitraum von 10 Tagen im MSV-Medium.
Tabelle 12: Esteraseproduktion durch T. fusca KW3 im MSV-Medium mit verschiedenen Additiven über einen Zeitraum von 10 Tagen.
Substrat
(2 g/L)
Kultivierungstag
mit der höchsten
Enzymaktivität
Enzymaktivität
(U/ml)
Proteinkonzentration
(mg/ml)
Spezifische
Enzymaktivität
(U/mg)
BHET 3 0,021 0,25 0,084
PET-Oligomer-
präparation
3 0,007 0,14 0,047
zyklische PET-
Oligomer-
präparation
3 0,003 0,18 0,018
PET-Fasern 10 0,024 0,17 0,142
PBT-Fasern 10 0,014 0,06 0,240
Für die Proteinaufreinigung ist ebenfalls die gebildete Gesamtproteinmenge im Vergleich zur
Esterasemenge von Interesse, um einen optimalen Zeitpunkt für die Ernte der Rohenzymlösung
0
0,005
0,01
0,015
0,02
0,025
0 1 2 3 4 5 6 7
En
zym
ak
tivit
ät
(U/m
l)
Kultivierungsdauer (d)
PET Oligomerpräparation zyklische PET Oligomerpräparation BHET
PBT Fasern PET Fasern recyceltes PET Granulat

Ergebnisse und Diskussion 81
festzulegen. Um diese Entwicklung genauer zu untersuchen, erfolgte während der Kultivierung von
T. fusca KW3 im MSV-Medium mit PET-Fasern zu verschiedenen Zeitpunkten eine Probenahme.
Diese Proben wurden anschließend per SDS-PAGE gelelektrophoretisch aufgetrennt und unspezifisch
mit Coomassie angefärbt. Zum Vergleich wurde diese Untersuchung ebenfalls mit einer Suberin-
präparation im MSV-Medium durchgeführt, um nochmals die Unterschiede zwischen den beiden
verwendeten Additiven und der daraus folgenden Enzymproduktion zu verdeutlichen.
3.1.2.2 Esterasebildung mit einer Suberinpräparation
T. fusca KW3 wurde in 5 Kolben mit 250 ml MSV-Medium mit Suberin als Additiv über einen Zeitraum
von 10 Tagen bei 50°C kultiviert. Der zellfreie Überstand von je einem Kolben wurde nach 0, 2, 4, 7
und 10 Tagen Kultivierung durch Zentrifugation gewonnen, die Esteraseaktivität und der
Proteingehalt mit den Standardbestimmungen gemessen und der Überstand gefriergetrocknet. In
Abbildung 28 sind die Esteraseaktivitäten (U/ml) und die maximalen spezifischen Esteraseaktivitäten
(U/mg) der zellfreien Kulturüberstände während des Zeitraums der Kultivierung graphisch
dargestellt.
Die maximale Esteraseaktivität (8,1 U/ml) und spezifische Esteraseaktivität (50,8 U/mg) wurden nach
48 Stunden Kultivierung gemessen, danach erfolgte eine Aktivitätsabnahme. Bis zum 7. Tag blieb die
Esteraseaktivität konstant auf einem Stand von 30 U/mg Protein, danach sank die Aktivität auf
0,03 U/ml und 0,289 U/mg ab.
Darüber hinaus wurden Proben der zellfeien Überstände mittels SDS-PAGE gelelektrophoretisch
aufgetrennt (Abbildung 29). Dafür wurden die Proben mit gleichen Volumenmengen auf das Gel
aufgetragen. Anhand des Gelphotos wird sichtbar, dass während der Kultivierung ein Protein mit ca.
29 kDa Größe gebildet wurde, dessen Konzentration bis zum 2. Tag anstieg und anschließend
während der weiteren Kultivierung stetig sank.

Ergebnisse und Diskussion 82
Abbildung 28: Bestimmung der Esteraseaktivitäten (◊) und der spezifischen Esteraseaktivitäten () während des
Wachstums im MSV-Medium unter Zugabe von Suberin.
Abbildung 29: SDS-PAGE der zellfreien Überstände nach Inkubation mit Suberin im MSV-Medium, links Coomassie-gefärbtes Gel; rechts Gel mit Aktivitätsfärbung mit Fast Red. Spur 1, 7: Roti
®-Mark SDS Proteinstandard; Spur 2:
Nullwert; Spur 3, 8: zellfreier Überstand nach 2 Tagen Kultivierung; Spur 4: zellfreier Überstand nach 4 Tagen Kultivierung; Spur 5: zellfreier Überstand nach 7 Tagen Kultivierung; Spur 6: zellfreier Überstand nach 10 Tagen Kultivierung.
Zusätzlich erfolgte die Kultivierung von T. fusca KW3 für die Bestimmung der Biotrockenmasse-
bildung im MSV-Medium mit Suberin in 2 L Erlenmeyerkolben mit 750 ml Medium. Die Kultivierung
wurde gestartet durch die Zugabe von 42 ml Inoculum und erfolgte bei 50°C mit 150 rpm im
Schüttelinkubator mit täglicher Probenahme von je 35 ml und anschließender Filtration. Das Filtrat
0
20
40
60
80
0
2
4
6
8
10
0 2 4 6 8 10
Sp
ezif
isch
e E
nzym
akti
vit
ät
(U/m
g)
En
zym
akti
vit
ät
(U/m
l)
Kultivierungsdauer (d)
66 kDa
43 kDa
29 kDa
20 kDa
14.5 kDa
200 kDa
119 kDa
1 2 3 4 5 6 7 8

Ergebnisse und Diskussion 83
wurde für die Esteraseaktivitätsbestimmung eingesetzt und der Filter mit demineralisiertem Wasser
gewaschen, über Nacht getrocknet und nach dem Abkühlen im Exikkator mit der Feinwaage
ausgewogen. Die von T. fusca KW3 gebildete Biotrockenmasse in mg/ml und die Esteraseaktivität in
U/ml wurden gegeneinander aufgetragen (Abbildung 30). Da das MSV-Medium mit den Zellen aus
der Vorkultur angeimpft wurde, lag zum Zeitpunkt 0 ein Trockengewicht an Zellen von 0,7 mg/ml vor.
Bis zum vierten Tag gab es eine Zunahme sowohl der Biomasse als auch der Esteraseaktivität. Ab
dem vierten bis zum neunten Tag nahm die Biotrockenmasse nicht weiter zu. Nach dem neunten Tag
war dann eine Abnahme der Biotrockenmasse zu verzeichnen. Bezüglich der Esteraseaktivität konnte
festgestellt werden, dass die Aktivität vom ersten bis vierten Tag stark anstieg und danach auf ein
Drittel der maximalen Aktivität abfiel. Vom fünften bis achten Tag blieb die Esteraseaktivität konstant
und sank bis zum elften Tag auf 1,3 U/ml ab.
Die Unterschiede im Verlauf der Enzymaktivitäten in Abbildung 28 und 30 sind zum Einen auf die
verschiedenen Größen der Kultivierungsansätze (250 und 750 ml) und zum Anderen auf die
natürlichen Unterschiede der Suberinpräparation zurückzuführen.
Abbildung 30: Biotrockenmasseproduktion () und Esteraseaktivität (◊) von T. fusca KW3 im MSV-Medium.
3.1.2.3 Esterasebildung mit PET-Fasern
Um den optimalen Zeitpunkt für die Gewinnung der 52 kDa Esterase Rohenzymlösung in Form des
zellfreien Überstandes zu ermitteln, wurde T. fusca KW3 in 5 Erlenmeyerkolben mit je 250 mL MSV-
Medium mit PET-Fasern bei 50°C kultiviert. Eine Kontrolle wurde sofort nach der Inokulation
abfiltriert und abzentrifugiert, um den zellfreien Überstand zu gewinnen. Diese Probe diente als
0
0,3
0,6
0,9
1,2
1,5
1,8
2,1
0
1
2
3
4
5
6
7
0 1 2 3 4 5 6 7 8 9 10 11
Bio
tro
ck
en
ma
ss
e (
mg
/ml)
En
zym
ak
tivit
ät
(U/m
l)
Kultivierungsdauer (d)

Ergebnisse und Diskussion 84
Nullwert. Nach 2, 4, 8 und 10 Tagen wurden die zellfreien Überstände von je einem weiteren Kolben
geerntet und die Esteraseaktivität und der Proteingehalt bestimmt. Die Esteraseaktivität (mU/ml)
wurde gegen die Kultivierungsdauer (Tage) aufgetragen (Abbildung 31). Ab der Kultivierungsdauer
von 2 Tagen stieg die Esteraseaktivität bis zum 10. Tag auf 35 mU/ml bzw. die spezifische Aktivität
auf 673 mU/mg an.
Abbildung 31: Graphische Darstellung der Esteraseaktivität (◊) und der spezifischen Esteraseaktivität () gebildet während
der 10-tägigen Kultivierung im MSV-Medium mit PET-Fasern.
Darüber hinaus wurden Proben der zellfeien Überstände mittels SDS-PAGE gelelektrophoretisch
aufgetrennt (Abbildung 32).
Abbildung 32: SDS-PAGE der zellfreien Überstände nach Inkubation mit PET-Fasern im MSV-Medium, links Coomassie-gefärbtes Gel; rechts Gel mit Aktivitätsfärbung mit Fast Red. Spur 1 und 7: Roti
®-Mark SDS Proteinstandard; Spur 2:
Nullwert; Spur 3: zellfreier Überstand nach 2 Tagen Kultivierung; Spur 4: zellfreier Überstand nach 4 Tagen Kultivierung; Spur 5: zellfreier Überstand nach 8 Tagen Kultivierung; Spur 6, 8: zellfreier Überstand nach 10 Tagen Kultivierung.
0
100
200
300
400
500
600
700
800
0
5
10
15
20
25
30
35
40
0 2 4 6 8 10
Sp
ezif
isc
he
En
zym
ak
tivit
ät
(mU
/mg
)
En
zym
ak
tivit
ät
(mU
/ml)
Kultivierungsdauer (d)
1 2 3 4 5 6 7 8
200 kDa
119 kDa
66 kDa
43 kDa
29 kDa
20 kDa
14.5 kDa

Ergebnisse und Diskussion 85
Dafür wurden die Proben mit gleichen Volumenmengen auf das Gel aufgetragen. Anhand des
Gelphotos wurde sichtbar, dass bereits nach 2 Tagen Kultivierung ein Protein mit ca. 52 kDa Größe
gebildet wurde, dessen Konzentration bis zum 10. Tag stetig anstieg.
Der optimale Zeitpunkt für die Gewinnung des zellfreien Überstandes für die Aufreinigung der 52 kDa
PET-Esterase war demnach nach einer 10-tägigen Kultivierung im MSV-Medium mit PET-Fasern.
Die Biotrockenmassebestimmung zur Untersuchung der Wachstumskurve von T. fusca KW3 mit PET-
Fasern konnte nicht durchgeführt werden, da ein Teil der Biomasse an der Faseroberfläche anhaftete
und somit mit den Fasern bei der Filtration von den restlichen Zellen abgetrennt wurde. Somit
konnte keine Gesamtbiotrockenmasse bestimmt werden.
Anhand der bisherigen Versuche konnte folgendes festgestellt werden: Bei der Kultivierung von
T. fusca KW3 im MSV-Medium mit PET-Fasern wurde die Bildung einer 52 kDa großen Esterase
induziert (Abbildung 32). Erfolgte stattdessen die Kultivierung in Gegenwart von Suberin, wurde eine
29 kDa Esterase produziert (Abbildung 29).
Da die 52 kDa Esterase unter Zusatz von synthetischen Polyestern produziert wurde, war sie von
größerem Interesse für die weiteren Untersuchungen. Aus diesem Grund erfolgten alle
nachfolgenden Arbeiten mit der 52 kDa große Esterase, die mit den PET-Fasern als Substrat gebildet
wurde. Allerdings ergab sich daraus das Problem, dass die Produktion der Esterase mit den PET-
Fasern einen hohen Zeitaufwand für die Kultivierung beanspruchte, um genügend Rohenzymlösung
für eine Aufreinigung des Enzyms bereitzustellen. Aus diesem Grund wurde im Anschluss ein
weiteres Terephthalat-enthaltendes Substrat als möglicher Zusatz für die Produktion der gesuchten
Esterase getestet. Ebenfalls wurden zwei weitere Spezies der Gattung Thermobifida fusca in das
Esterase-Screening mit einbezogen, um zu testen, ob sich die Esteraseproduktion deutlich
unterscheidet von der von T. fusca KW3.
3.1.3 Esterasebildung bei T. fusca KW3 DSM 6013, T. fusca DSM 43792 und DSM 43793 mit PET-Fasern und Diethylterephthalat
Das Ziel dieser Untersuchung war es, einen Stamm und einen Zusatzstoff zu finden, mit dem eine
maximale Ausbeute an PET abbauender Esterase in der Rohenzymlösung erhalten wird. Neben PET-
Fasern wurde als weiteres Additiv Diethylterephthalat (DET), ebenfalls wie BHET ein Derivat der
Terephthalsäure, eingesetzt. Diese Substanz wurde dem MSV-Medium wie alle anderen bereits
getesteten Additive in einer Konzentration von 2 g/L zugesetzt. Neben T. fusca KW3 wurden zwei
weitere, vom DSMZ bezogene Stämme unter den gleichen Bedingungen kultiviert (T. fusca DSM
43792 und T. fusca DSM 43793). Bei der Kultivierung sollte die Esterasebildung von T. fusca KW3 mit

Ergebnisse und Diskussion 86
PET-Fasern und DET im MSV-Medium verglichen werden mit der Esterasebildung von
T. fusca DSM 43792 und DSM 43793 unter gleichen Bedingungen.
Für T. fusca KW3 erfolgte während der Kultivierung eine Probenahme zur Bestimmung der
Esteraseaktivitäten im Überstand (Abbildung 34). Darüber hinaus wurden Proben für die SDS-PAGE
und die Aktivitätsfärbung entnommen (Abbildung 33). Es ist ersichtlich, dass nach der Kultivierung
mit PET-Fasern sowohl bei Stamm T. fusca KW3 als auch bei den Stämmen T. fusca DSM 43792 und
DSM 43793 eine 52 kDa große Esterase gebildet wurde, die über die Aktivitätsfärbung mit Fast Red
genau auf dem Gel identifiziert werden konnte (rote Banden Spuren 2, 4, 6). Desweiteren konnte
gezeigt werden, dass nach Kultivierung mit DET bei allen drei untersuchten Stämmen eine 29 kDa
große Esterase produziert wurde, die ebenfalls über Aktivitätsfärbung mit Fast Red genau
identifiziert werden konnte (rote Banden Spuren 3, 5, 7). Damit konnte festgestellt werden, dass die
beiden Additive die Produktion zweier unterschiedlicher Esterasen induzieren: die PET-Fasern eine
52 kDa Esterase und DET eine 29 kDa Esterase. Somit waren die Stämme bei Anwesendheit von DET,
ebenso wie von Suberin und Cutin, nicht in der Lage, die 52 kDa Esterase zu synthetisieren.
Da sich die produzierten Mengen der 52 kDa Esterase bei allen drei Stämmen nicht unterscheiden,
wurde bei den weiteren Untersuchungen das Isolat aus der Stammsammlung, T. fusca KW3,
verwendet.
Abbildung 33: SDS-PAGE der Rohenzymlösungen nach Kultivierung mit unbehandelten PET-Fasern bzw. DET im MSV-
Medium, Gel gefärbt mit Coomassie- und mit Aktivitätsfärbung. Spur 1, 8: Low Molecular Weight Längenstandard; Spur 2: T. fusca DSM 43792 mit PET-Fasern kultiviert; Spur 3: T. fusca DSM 43792 mit DET kultiviert; Spur 4: T. fusca DSM 43793 mit PET-Fasern kultiviert; Spur 5: T. fusca DSM 43793 mit DET kultiviert; Spur 6: T. fusca KW3 DSM 6013 mit PET-Fasern kultiviert; Spur 7: T. fusca KW3 DSM 6013 mit DET kultiviert.
97 kDa 66 kDa 45 kDa 30 kDa 20.1 kDa 14.4 kDa
52 kDa
29 kDa
1 2 3 4 5 6 7 8

Ergebnisse und Diskussion 87
Abbildung 34: Esteraseproduktion durch T. fusca KW3 mit DET (◊) und PET-Fasern () über einen Zeitraum von 13 Tagen im MSV-Medium.
Beim Screening nach PET-Hydrolasen aus T. fusca zeigte sich, dass zunächst die Vorkultur im Czapek-
Medium nach zwei Tagen Wachstum die maximale Zelldichte mit noch exponentiellem Wachstum
vorwies. Die Zellen wurden steril zentrifugiert, in Puffer resuspendiert, um alle Reste an Glucose zu
entfernen, und damit das MSV-Medium angeimpft. Die Kultur im MSV-Medium wurde mit
verschiedenen Zusätzen kultiviert, um hydrolytische Enzyme zu produzieren. Während des
Wachstums war zu beobachten, dass mit Cutin, Suberin und DET die Produktion eines anderen
Enzyms erfolgte, als mit den synthetischen PET- und PBT-Fasern, PET-Granulat und der PET-
Oligomerpräparation. Mit den Monomeren TPA und Ethylenglycol erfolgte keine Enzymproduktion.
Die mit den Biopolyestern und erstaunlicherweise auch mit DET induzierte Hydrolase besaß p-NPB
Aktivität und wies im SDS-PAGE Gel eine molekulare Größe von 29 kDa auf. Desweiteren war die
maximale Produktion des Enzyms bei 50°C nach 2 Tagen mit Suberin erreicht und betrug 8,1 U/ml
und eine spezifische Aktivität von 50,8 U/mg. Mit den synthetischen Additiven bildete T. fusca KW3
hingegen eine andere, größere Hydrolase von ca. 52 kDa und mit p-NPB Aktivität. Für dieses Enzym
wurde die maximale Produktionsrate nach mindestens 10 Tagen Kultivierung mit 35 mU/ml bzw. die
spezifische Aktivität auf 673 mU/mg erreicht. Die Produktion erfolgte ebenfalls bei 50°C, jedoch
wurden deutlich geringere Mengen Enzym produziert im Vergleich zur 29 kDa Hydrolase (Verhältnis
1:230) mit einem 5fach höheren Zeitaufwand.
Diese Produktion zweier verschiedener Esterasen wurde nicht nur bei T. fusca KW3 festgestellt,
sondern auch mit den Stämmen aus der Stammsammlung des DSMZ, T. fusca DSM 43792 (ATCC
27730) und DSM 43793.
Fett et al. testeten bereits 1999 verschiedene Actinomyceten auf ihre Produktion von Cutin-
depolymerisierenden Enzymen (Fett et al. 1999). Von Thermomonospora alba NRRL B-16963T
0
5
10
15
20
25
30
35
40
45
0 2 4 6 8 10 12 14
Es
tera
se
ak
tivit
ät
(mU
/ml)
Kultivierungsdauer (d)

Ergebnisse und Diskussion 88
(NCIMB 10169); T. chromogena ATCC 43196T; T. curvata ATCC 19995T; T. formosensis ATCC 49059T;
T. fusca ATCC 27730T (DSM 43792), NRRL B-11456 und YX-5p; T. mesophila ATCC 27303T;
T. mesouviformis ATCC 27644T und T. sp. NRRL B-16962 produzierte nur der Referenzstamm
T. fusca ATCC 27730 im Trypton-Hefeextrakt-Medium Cutinase-Aktivität. Ebenso produzierte der
verwendete Stamm T. fusca KW3 im MSV-Medium mit Pepton bei Anwesenheit einer Suberin- oder
Cutin-Präparation Esteraseaktivität.
Mit verschiedenen Substanzen und cutin- bzw. lipidhaltigen Naturprodukten (1-Hexadecanol, ein
Cutinhydrolysat, Palmitinsäure, Hexadecan-1-ol, Rizinolsäure, Maisfasern (unbehandelt), Maiskleie,
Rizinusöl, Maiskeimöl und Olivenöl) zeigte der Referenzstamm T. fusca ATCC 27730 keine Anregung
der Cutinaseproduktion und bei höheren Konzentrationen sogar eine Hemmung des
Bakterienwachstums (Fett et al. 1999). T. fusca KW3 produzierte mit den Monomeren
Terephthalsäure und Ethylenglycol ebenfalls keine Esteraseaktivität.
Hingegen konnte bei T. fusca ATCC 27730 mit Apfelcutin, Apfeltrester, Tomatenschalen, Kork und
Suberin aus Kartoffeln Cutinase-Aktivitäten nachgewiesen werden, welche mit den Tomatenschalen
am höchsten waren. T. fusca KW3 produzierte mit Apfelcutin und Suberin die höchsten
Esteraseaktivitäten, wobei die Aktivitäten mit Apfelcutin am Höchsten waren (Daten nicht
dargestellt). Geringere Esteraseaktivitäten wurden mit synthetischen PET- und PBT-Fasern, PET-
Granulat, der PET-Oligomerpräparation und DET bestimmt. Bei Untersuchungen zum biologischen
Abbau von aromatisch-aliphatischen Copolyestern mit den neu isolierten T. fusca K13g und K7a-3
erfolgten die Induktion und das Screening nach hydrolytischer Esteraseaktivität mit Polymerfilmen
aus 1,4-Butandiol, Terephthalsäure und Adipinsäure (Kleeberg et al. 1998). T. fusca K13g und K7a-3
zeigten bei 16S rDNA Sequenzvergleichen die höchsten Übereinstimmungen mit T. fusca DSM 43792T
(99,8% und 100%). Nachfolgend wurden weitere Studien zur Produktion dieser Copolyester-
abbauenden Hydrolase aus T. fusca DSM 43793 mit Ecoflex™-Pulver (BTA Copolyester, Partikelgröße
< 0,8 mm) als Induktor durchgeführt (Gouda et al. 2002). Die höchsten Aktivitäten der Hydrolase
wurden aufgrund des Austauschs von 5 g/L Pepton im MSV-Medium durch 1,87 g/L NH4Cl und 5 g/L
Pektin erzielt (von 1,5 UPCL-Film/mg und 0,8 U/mgProtein auf 1,8 UPCL-Film/mg und 6,0 U/mgProtein). Die
Actinomycetenisolate M5, M9 und Thermobifida fusca KW3b (jetzt KW3), alle isoliert aus Kompost,
wurden in einem Minimalmedium mit PET-Garn, DET oder einer Suberinpräparation aus
Kartoffelschalen (je 2 g/L) zur Enzymproduktion 8 Tage bei 45°C kultiviert (Alisch et al. 2004). Die
produzierte Enzymaktivität, bestimmt mit p-NPB, lag für alle Isolate mit PET-Fasern und DET als
Induktor während der 8 Tage bei maximal 40 mU/ml, ebenso für M5 und M9 mit der
Suberinpräparation. Die einzige Ausnahme war T. fusca KW3b, der nach 8 Tagen mit der
Suberinpräparation 120 mU/ml aufweisen konnte (Alisch et al. 2004).

Ergebnisse und Diskussion 89
Weiterhin konnte festgestellt werden, dass die Zugabe von Glucose zum Produktionsmedium bei
T. fusca ATCC 27730 und T. fusca KW3 eine stark hemmende Auswirkung auf die Enzymproduktion
besaß (Fett et al. 1999). Auch bei Gouda et al. 2002 erfolgte beim Einsatz von Glucose, Saccharose
oder Stärke anstatt Pektin keine Produktion der Hydrolase.
Weiterhin konnte kein signifikanter Einfluss des eingesetzten Inokulationsvolumens (1 bis 15 ml auf
80 ml Medium) auf die maximal produzierte Enzymaktivität (1,3 bis 1,7 UPCL-Film/mL) identifiziert
werden (Gouda et al. 2002). Bei der Untersuchung des Einflusses der freigesetzten BTA-Monomere
auf die Esteraseproduktion wurden die Natriumsalze der Adipinsäure und der Terephthalsäure und
1,4-Butandiol dem Kulturmedium vor der Inokulation beigesetzt. Bei einer Konzentration von
50,1 mg 1,4-Butandiol oder 68,0 mg Adipinsäure oder 50,4 mg Terephthalsäure zusätzlich zu 250 mg
BTA-Pulver wurden keine hydrolytischen Aktivitäten im Medium detektiert. Das bedeutet, dass diese
Konzentrationen sich auf eine weitere Enzymproduktion hemmend auswirken, nicht aber auf bereits
produziertes Enzym, welches die Monomere während eines Abbaus freisetzt. Beim Vergleich von
BTA-Filmen, -Pulver und -Nanopartikeln als Induktor zeigte sich, dass die Polymeroberfläche keinen
bedeutenden Einfluss auf die Enzymproduktion nach 48 Stunden hatte. Hingegen zeigte die Variation
der BTA-Pulver Konzentration im Produktionsmedium einen interessanten Einfluss auf die
produzierte Menge an hydrolytischem Enzym. Beim Scale-up der Produktion vom Schüttelkolben
zum 100 L Fermenter wurden stets Enzymaktivitäten um die 1,5 UPCL-Film/mL und spezifische
Aktivitäten von 4 bis 6 U/mgProtein nach 48 Stunden Inkubation erreicht. Die untersuchte Hydrolase
aus T. fusca DSM 43793 besaß ein Molekulargewicht von ca. 30 kDa (Gouda et al. 2002).
Weiterführend zu Fett et al. (1999) erfolgte die Erzeugung von Mutanten des Referenzstamms
T. fusca ATCC 27730 (T. fusca WSH04) mit einer höheren Cutinaseproduktion mittels Behandlung mit
Diethylsulfat (Du et al. 2007). Weiterhin wurde der Einfluss von Glucose, Saccharose, löslicher Stärke,
Ethanol und Natriumacetat auf die Cutinaseproduktion untersucht und es zeigte sich, dass mit dem
Zusatz von Ethanol und Natriumacetat ähnliche Zelltrockengewichte und Cutinaseaktivitäten erreicht
werden konnten, die spezifische Cutinaseaktivität aber höher beim Einsatz von Natriumacetat war.
Bei der Untersuchung des Einflusses der Natriumacetatkonzentration auf die gebildete Zellmasse und
Cutinaseaktivität zeigte sich, dass mit einer Konzentration von 7,5 g/L die höchsten Ergebnisse erzielt
wurden. Das beste Zellwachstum wurde mit pH 7,3 und die höchste Cutinaseproduktion mit pH 7,6
erreicht. Deshalb entwickelten Du et al. (2007) eine 2-Phasen pH-kontrollierte Kultivierung mit 20
Stunden bei pH 7,3 für eine maximale Biomasseproduktion und anschließend bei pH 7,6 nach 50
Stunden Kultivierung für eine maximale Cutinaseproduktion (Du et al. 2007).
Hu et al. (2010) isolierten von kompostierten Polyesterfilmen zwei Gruppen von Mikroorganismen,
Actinomyceten (vier Gattungen) und Bacillen (drei Gattungen). Von diesen Isolaten war nur
Thermobifida alba AHK119 in der Lage, den aliphatisch-aromatischen Copolyesterfilm (Apexa®, zuvor

Ergebnisse und Diskussion 90
Biomax® 4026 und 4027, ein Polyethylenterephthalat Copolymer aus Terephthalsäure, Ethylenglycol
und einer ungenannten Komponente) abzubauen, ebenso die Partikelgröße von Polymeren (PCL,
Bionolle™ EM-301 (Polybutylensuccinat-co-adipat (PBSA)), Ecoflex™) zu verringern und dabei
Terephthalsäure freizusetzen. Verantwortlich für den Abbau war eine Hydrolase von ca. 30 kDa
Größe (Hu et al. 2010). Bisher wurden allerdings nicht nur aus Thermobifida cutinolytische oder PET-
abbauende Enzyme identifiziert, sondern ebenfalls aus anderen Bakterienstämmen und auch Pilzen.
Aus einer phyllospherischen Bakterienkultur wurde 1987 von Sebastian et al. ein fluoreszierender
Pseudomonas putita ATCC 55613 (wurde umbenannt zu P. mendocina) Stamm isoliert. Das Isolat
produzierte eine extrazelluläre Cutinase während des Wachstums mit Cutin, aber nicht mit Cutin
Hydrolysaten im Vergleich zur eukaryotischen Produktion von Fusarium solani sp. pisi. Die während
der Hydrolyse freigesetzten Cutinmonomere wurden mittels Dünnschichtchromatographie
nachgewiesen (Sebastian et al. 1987). Bei der SDS-PAGE und der Gelfiltration zeigte das Enzym ein
Molekulargewicht von 30 kDa und eine monomere Form (Sebastian et al. 1988).
El-Bendary et al. haben 2010 die Oberflächenhydrolyse von PET-Textilien durch Lipasen aus den
Bacillus Isolaten 5W und 6C untersucht. Die Lipase aus dem Bacillus 5W Isolat wurde in größeren
Mengen als PET-oberflächenmodifizierendes Enzym mit 6% Baumwollsamenmehl und pH 7,5
produziert (90U/ml). Die beste Ausbeute wurde mit 25 ml Medium, 25 x 106 CFU/ml Inoculum und
24 Inkubation erzielt. Die Zugabe von Pepton zum Medium steigerte die Lipaseproduktion um 22,5%
(130,7 U/ml) (El-Bendary et al. 2010).
F. solani f. pisi produzierte während des Wachstums mit Cutin extrazelluläre Cutin-
depolymerisierende Enzyme, welche alle Gruppen von Cutinmonomeren freisetzten (Purdy und
Kolattukudy 1973). Aus dem extrazellulären Überstand wurden zwei Isoenzyme, eine Cutinase und
eine unspezifische Esterase (p-Nitrophenylpalmitat Hydrolase) mit einem Molekulargewicht von
22 kDa bzw. 52 kDa isoliert (Purdy und Kolattukudy 1975a).
Cutinhydrolysate im Medium hatten eine Produktion einer extrazellulären Cutinase bei einem in
Glucose-Medium gewachsenen F. solani f. sp. pisi zur Folge (Lin und Kolattukudy 1978). Die
Produktionsrate war abhängig von der Menge des Cutinhydrolysates, welches bis zu einer
Konzentration von 80 µg/ml zugegeben wurde, und bei dieser Konzentration erfolgte kein weiterer
Anstieg der Produktion. Bei Anwesenheit von Glucose erfolgte keine Cutinaseproduktion. Die
Induktion der Cutinase durch Cutinhydrolysate wurde nicht durch Actinomycin D (zytotoxisches
Antibiotika) inhibiert und durch Cordycepin (Antitumor, antifungales und antivirales Reagenz) sogar
noch erhöht. Durch die Zugabe von Cycloheximid mit dem Induktor oder bis zu 12 Stunden nach der
Zugabe des Induktors resultierte ein fast sofortiger Stillstand der Cutinaseproduktion. Ebenso
unterdrückte Deoxyglucose, ein Hemmer der Proteinglykosylierung, die Produktion der Cutinase mit
Cutinhydrolysaten. Die ω-Hydroxyfettsäuren waren deutlich effizienter bei der Induktion der

Ergebnisse und Diskussion 91
Cutinase als alle anderen noch polareren Säuren des Cutins. Experimente mit Derivaten und Analogas
der ω-Hydroxy-C16-Fettsäuren zeigten, dass eine freie Hydroxylgruppe an der ω-Position als ein
wichtiger Faktor für die Cutinaseinduzierende Aktivität zu sehen ist. Ebenfalls erfolgte mit n-
aliphatischen primären Alkoholen (C14 oder mehr) die Produktion der Cutinase, wobei n-C16 am
effizientesten induzierte. Das bedeutet, dass die Monomere als das chemische Signal agieren,
welches die extrazelluläre Hydrolase induziert (Lin und Kolattukudy 1978).
Ebenfalls wurden Cutinasen aus fünf verschiedenen phytopathogenen Pilzen und Bakterien,
F. roseum culmorum, F. roseum sambucinum, Ulocladium consortiale, Streptomyces scabies und
Helminthosporium sativum, mit einem Molekulargewicht von ca. 25 kDa isoliert. Eine Suspension mit
Sporen oder Mycelien einer 14 Tage alten Kultur wurde dafür in 100 ml eines Mineralmediums mit
pH 7,2 und 0,5 g Cutin gegeben (Lin und Kolattukudy 1980a).
Das F. solani pisi Isolat T-8 war in der Lage, mit Suberin aus Kartoffelschalen als einzige
Kohlenstoffquelle zu wachsen. Der extrazelluläre Überstand dieser Kulturen hydrolysierte p-
Nitrophenylbutyrat und radioaktiv markiertes Apfelcutin. Der Großteil der Esterase war an die
Suberin enthaltende Mycelmatte gebunden und konnte mit einer 0,4% Triton X-100 Lösung
ausgewaschen werden. Die Enzymlösung enthielt zwei Isoenzyme mit einem Molekulargewicht von
23 kDa. Desweiteren wurde festgestellt, dass F. solani pisi sehr ähnliche oder gleiche Polyesterasen
bildet für den Abbau von Cutin und Suberin, wenn diese als einzige Kohlenstoffquellen im Medium
vorhanden sind (Fernando et al. 1984).
Nimchua et al. (2007) verwendete für die Isolation von Mikroorganismen mit PET-hydrolysierenden
Enzymen sowohl Pflanzenproben, wie Blätter, Blüten, unterirdische Speicherorgane und Früchte, als
auch Bodenproben gesammelt an verschiedenen Stellen in Nord-Thailand. Die PCL abbauenden
Isolate und der F. solani f. sp. pisi wurden anschließend mit Mineralmedium kultiviert (Nimchua et al.
2007). Verschiedene Microfungi wurden von Pflanzenoberflächen und Bodenproben mittels PCL-
Mineralmedium-Platten isoliert. 22 der 115 Isolate zeigten Hofbildung, was auf eine Cutinaseaktivität
schließen ließ. Die Produktion der Cutinase im Mineralmedium mit Suberin aus Kartoffeln oder PET-
Fasern wurde mittels p-NPB Aktivitätsbestimmung nachgewiesen, wobei alle Isolate p-NPB Aktivität
zeigten. Das Isolat PBURU-B5 besaß die höchste p-NPB Aktivität mit PET-Fasern und wurde
identifiziert als F. solani (Nimchua et al. 2008).
Wang et al. (2008) untersuchten die Optimierung der Kulturbedingungen der Lipaseproduktion und
die Bereitstellung einer spezifischen Lipase für die PET-Hydrolyse aus Aspergillus oryzae CCUG. Die
unter Zusatz von Olivenöl produzierte Lipase konnte die Eigenschaften der PET-Textilien nicht
deutlich ändern, deshalb wurden Diethyl-p-phthalat (DP), BHET und PET-Fasern als Zusätze
verwendet. Mit BHET wurden die besten Produktionsraten erreicht und die extrazelluläre Lipase
hydrolysierte das PET Modellsubstrat DP (Wang et al. 2008).

Ergebnisse und Diskussion 92
Zur Identifikation neuer PET-modifizierender Mikroorganismen erfolgte ein Screening mit
Bodenproben von verschiedenen Mülldeponien und von einer Abwasserkompostierungsanlage als
Inoculum. Die gesuchten Mikroorganismen sollten mit PET als alleinige C-Quelle wachsen können.
Die Kultivierung erfolgte in einem Mineralsalzmedium mit Spurenelementen bei pH 7 (für Bakterien)
und pH 5 (für Pilze). Das Medium mit PET-Pulver (10 g/L) wurde mit einer Bodenprobe kultiviert.
Nach drei Wochen wurden die Kulturen auf Vollmedium ausgestrichen und Reinkulturen auf ihre
PET-Hydrolyse hin getestet. Es konnten 4 verschiedene bakterielle und 5 verschiedene eukaryotische
Isolate identifiziert werden. Da das Wachstum mit PET sehr langsam erfolgte, wurde zusätzlich Cutin
als Induktor getestet. Alle Pilze und 2 Bakterienisolate wuchsen mit Cutin als einzige C-Quelle und
produzierten eine Esteraseaktivität, hingegen zeigten mit Adipinsäure-bishexylamid als Induktor alle
bakteriellen Isolate extrazelluläre PET-Hydrolyseaktivität. Im Vollmedium mit Pepton, Hefeextrakt
und Glucose besaßen alle Isolate nur eine schwache p-NPA Aktivität (Fischer-Colbrie et al. 2004).
Mit 3PET (BETEB, 1 g/L) als Modellsubstrat isolierten Liebminger et al. (2007) Mikroorganismen, die
PET-abbauende Enzyme produzieren. Dabei wurde ein Penicillium citrinum Isolat identifiziert,
welches eine Polyesterase von 14.1 kDa zum PET Abbau produziert. Die höchsten Enzymausbeuten
konnten unter Zusatz von Cutin (1.2 g/L) erreicht werden (Liebminger et al. 2007).
Der Abbau von BHET/DET (in der Literatur wurde unter Zusatz von Diethylenglycolterephthalat
angegeben, was eine andere Beschreibung für BHET ist; in den Abbildungen ist allerdings DET
dargestellt, was sich durch die beiden endständigen Alkoholgruppen vom BHET unterscheidet; somit
ist keine eindeutige Zuordnung möglich) und PET-Fasern durch Mikroorganismen und Lipasen wurde
von Zhang et al. 2004 untersucht. Die isolierten Mikroorganismen aus Belebtschlamm wurden auf
ihre Fähigkeit hin, PET-Fasern oder BHET/DET zu hydrolysieren, getestet (Zhang et al. 2004, Zhang
et al. 2006).
Zusammenfassend ist aus den Tabellen 13 und 14 ersichtlich, dass bei den Bakterien und Pilzen je
maximal zwei verschiedene Polyesterspaltende Enzyme produziert wurden und diese in einem
Größenbereich von 14 bis 52 kDa lagen. Bei den Bakterien wurden meist hochpolymere natürliche
oder synthetische Verbindungen aus Zusatzstoffe eingesetzt, wohingegen deren Monomere oder
Hydrolysate ebenso wie Glucose oder andere Zucker eher eine hemmende Wirkung auf die
Enzymproduktion zeigten. Zusätzlich zu den Induktoren wurden noch weitere komplexe C-Quellen
für die Bildung der Biokatalysatoren benötigt und die Kultivierungsdauer lag zwischen 1 bis 10 Tagen
bis zum Erreichen der maximalen Enzymaktivität.

Ergebnisse und Diskussion 93
Tab
elle
13
: B
ildu
ng
von
PET
-Hyd
rola
sen
be
i Bak
teri
en.
Re
fere
nz
Fett
et
al.
19
99
Go
ud
a et
al.
20
02
Alis
ch e
t a
l. 2
00
4
Eige
ne
Erge
bn
isse
Du
et
al 2
00
7
Hu
et
al.
20
10
Seb
asti
an
et a
l.
19
87
El-B
end
ary
et a
l.
20
10
Hem
mu
ng
Glu
cose
Glu
cose
,
Sacc
har
ose
,
Stär
ke,
BTA
Mo
no
mer
e
k. A
.
Glu
cose
Glu
cose
k. A
.
Cu
tin
-
hyd
roly
sat
k. A
.
C-Q
uel
le
Tryp
ton
, Hef
eext
rakt
NH
4C
l, P
ekti
n
Pep
ton
Pep
ton
Pep
ton
, Hef
eext
rakt
,
Nat
riu
mac
etat
Hef
eex
trak
t, C
asei
n
Hyd
roly
sat
Hef
eex
trak
t
Pep
ton
, Hef
e-
un
d
Flei
sch
extr
akt
Ind
ukt
or
Ap
felc
uti
n, A
pfe
ltre
ster
,
Tom
aten
sch
alen
, Ko
rk,
Sub
erin
(4
g/L
)
BTA
-Film
e, -
Pu
lver
un
d -
Nan
op
arti
kel (
3,1
25
g/L
)
DET
, PET
-Fas
ern
(2
g/L
)
Sub
erin
(2
g/L
)
PET
-Fas
ern
(2
g/L
)
Sub
erin
(2
g/L
)
Cu
tin
(1
g/L
)
Ap
exa®
Co
po
lyes
ter
Pu
lver
(1 g
/L)
Cu
tin
(4
g/L
)
Bau
mw
olls
amen
meh
l
(60
g/L
)
Inku
bat
ion
7 T
age
2 T
age
8 T
age
10
Tag
e
2 T
age
50
Stu
nd
en
1-5
Tag
e
4 T
age
1 T
ag
Akt
ivit
ät
12
,1 U
/ml
1,8
UP
CL-
Film
/mg
40
mU
/ml
120
mU
/ml
35
mU
/ml
8,1
U/m
l
19
,8 U
/ml
k. A
.
100
-11
0 U
/ml
130
,7 U
/ml
Grö
ße
k. A
.
30
kD
a
k. A
.
52
kD
a
29
kD
a
k. A
.
30
kD
a
30
kD
a
k. A
.
Her
kun
ft
T. f
usc
a D
SM 4
3792
T. f
usc
a D
SM 4
3793
T. f
usc
a K
W3
T. f
usc
a K
W3
T. f
usc
a W
SH04
T. a
lba
AH
K1
19
P. m
end
oci
na
ATC
C 5
561
3
Ba
cillu
s sp
. 5W

Ergebnisse und Diskussion 94
Tab
elle
14
: B
ildu
ng
von
PET
-Hyd
rola
sen
be
i Pilz
en.
Re
fere
nz
Pu
rdy
un
d
Ko
latt
uku
dy
19
75
a
Lin
un
d
Ko
latt
uku
dy
19
78
Lin
un
d
Ko
latt
uku
dy
19
80
a
Fern
and
o e
t a
l.
19
84
Nim
chu
a et
al.
20
07
, Nim
chu
a
et a
l. 2
00
8
Wan
g et
al.
20
08
Lieb
min
ger
et a
l.
20
07
Zhan
g et
al.
20
04
,
Zhan
g et
al.
20
06
Hem
mu
ng
k. A
.
Glu
cose
k. A
.
k. A
.
k. A
.
k. A
.
k. A
.
k. A
.
C-Q
uel
le
kein
e W
eit
eren
Glu
cose
kein
e W
eit
eren
kein
e W
eit
eren
kein
e W
eit
eren
Mal
tose
, Pep
ton
,
Zitr
on
ensä
ure
, Oliv
enö
l,
Po
lyo
xyet
hyl
enal
kyle
ther
oh
ne
(Sc
ree
nin
g); P
ep
ton
,
Hef
eex
trak
t, G
luco
se
(Pro
du
ktio
n)
Hef
eex
trak
t
Ind
ukt
or
Cu
tin
(5
g/L
)
Cu
tin
hyd
roly
sat
(0,0
8 g
/L)
Cu
tin
(5
g/L
))
Sub
erin
(5
g/L
)
PC
L (0
,5 g
/L)
BH
ET (
0,0
25
-
0,5
g/L
)
BET
EB (
1 g
/L),
Cu
tin
(1
,2 g
/L)
BH
ET/D
ET (
0,5
-
1,0
g/L
)
Inku
bat
ion
12
Tag
e
4 T
age
12
Tag
e
21
Tag
e
ein
ige
Tage
ca.
70
Stu
nd
en
10
Tag
e
7 T
age
Akt
ivit
ät
k. A
.
7 Δ
A4
05/
min
/ml
k. A
.
k. A
.
k. A
.
12
-13
LU
/ml
32
nka
t/m
L (p
-
NP
B)
k. A
.
Grö
ße
22
kD
a
52
kD
a
k. A
.
ca.
25
kD
a
23
kD
a
k. A
.
k. A
.
14
kD
a
k. A
.
Her
kun
ft
F. s
ola
ni f
. pis
i
F. s
ola
ni f
. sp
. pis
i
F. r
ose
um
cu
lmo
rum
,
F. r
ose
um
sa
mb
uci
nu
m,
U. c
on
sort
iale
, S. s
cab
ies,
H. s
ati
vum
F. s
ola
ni p
isi T
-8
F. o
xysp
oru
m L
CH
1,
F. s
ola
ni f
. sp
. Pis
i DSM
624
20
A. o
ryza
e C
CU
G
P. c
itri
num
un
bek
ann
te Is
ola
te

Ergebnisse und Diskussion 95
Die verwendeten Induktoren bei den Pilzkultivierungen waren zum Teil Biopolyester, aber auch
Monomere oder Hydrolysate. Einige Pilze konnten sogar nur mit diesen Induktoren als einzige C-
Quelle wachsen. Dieser Unterschied macht deutlich sichtbar, dass die Bakterien nicht nur die Energie
in Form einer C-Quelle benötigen, sondern auch weitere wichtige Bausteine, wie z. B. Vitamine, die
sie selbst nicht synthetisieren können, Pilze hingegen sind dazu in der Lage. Die Hemmung der
Enzymproduktion bei den Bakterien durch die Abbauprodukte der enzymatischen Katalyse stellt
einen wichtigen Regulationsschritt im Stoffwechselweg dar und verhindert die Verschwendung von
Energiereserven. Die Kultivierungsdauer bei den Pilzkulturen war mit 4 bis 21 Tagen deutlich höher
als bei den Bakterien und begründet sich durch das langsamere Wachstum. Ein Vergleich der
gebildeten Aktivitäten ist nicht sehr sinnvoll, da für die Aktivitätsbestimmungen verschiedene
Substrate eingesetzt wurden. Interessanterweise bildete nicht nur T. fusca KW3 zusätzlich zu zwei
isomeren Cutinasen eine 52 kDa große Esterase, dieses Verhalten konnte ebenfalls bei F. solani f. pisi
identifiziert werden. Leider erfolgte keine weitere Studie dieser gebildeten Esterase zu dem
damaligen Zeitpunkt (Purdy und Kolattukudy 1975a, b).
3.2 Charakterisierung der PET-Hydrolasen aus T. fusca KW3
3.2.1 Aufreinigung
3.2.1.1 Aufreinigung der TfCa
Im Vergleich zu intrazellulären Enzymen, die durch Aufschluss mittels Ultraschall oder French Press in
großen Mengen gewonnen werden können und daraus Enzyme von Interesse über verschiedene
chromatographische Verfahren aufgereinigt werden, ist die Aufreinigung extrazellulärer Enzyme
komplizierter.
Die Enzymaufreinigung erfolgte bei Raumtemperatur. Nach Dialyse und Zentrifugation wurde die
Enzympräparation mit 100 mM Phosphatpuffer pH 8 auf 20 mM eingestellt. Dann erfolgte der
Auftrag der Enzymlösung auf eine selbstgepackte equilibrierte Phenylsepharose Säule. Unter den
gewählten Bedingungen band die TfCa an die Säulenmatrix. Durch die Erhöhung der Isopropanol
Konzentration von 0 auf 30% durch den in Abbildung 35 verzeichneten Stufengradienten erfolgte die
Elution des gebundenen aktiven Proteins bei 15 bis 25% Elutionspuffer B (Aufreinigungsschritt 2).
Aktive Fraktionen (p-NPB Aktivität) wurden vereinigt. Das enthaltene Isopropanol in der
Enzympräparation wurde mittels Rotationsverdampfers abgetrennt.
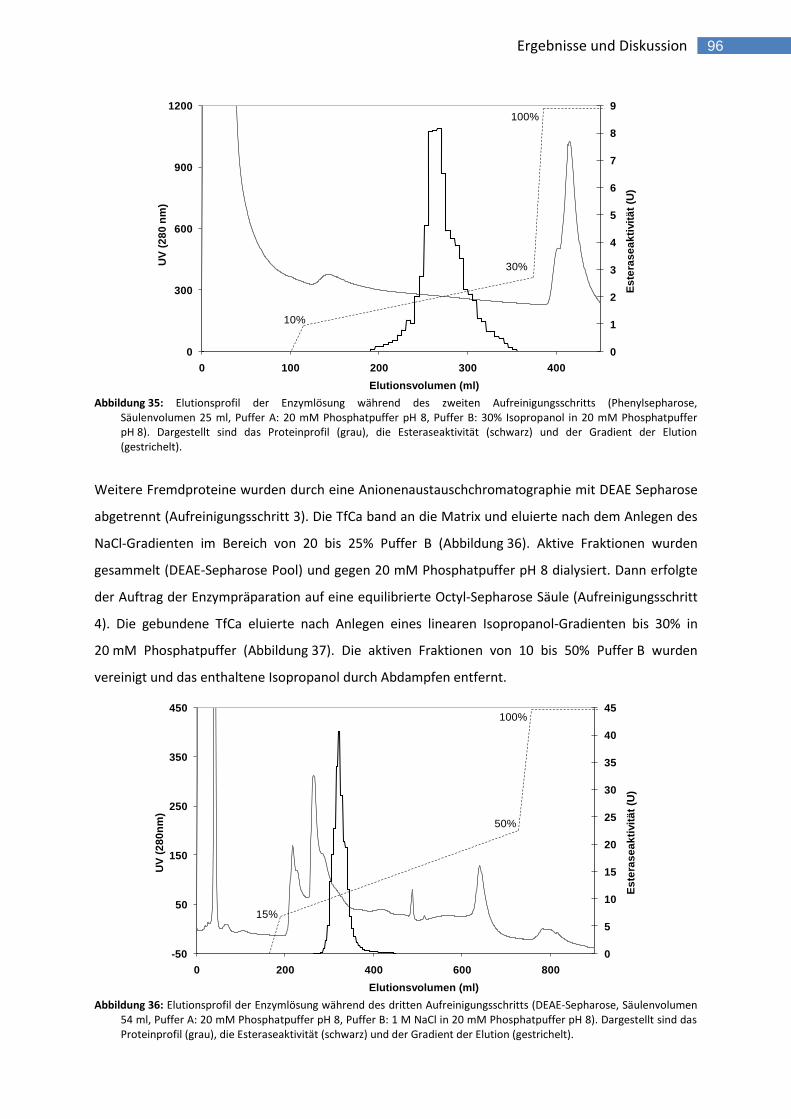
Ergebnisse und Diskussion 96
Abbildung 35: Elutionsprofil der Enzymlösung während des zweiten Aufreinigungsschritts (Phenylsepharose,
Säulenvolumen 25 ml, Puffer A: 20 mM Phosphatpuffer pH 8, Puffer B: 30% Isopropanol in 20 mM Phosphatpuffer pH 8). Dargestellt sind das Proteinprofil (grau), die Esteraseaktivität (schwarz) und der Gradient der Elution (gestrichelt).
Weitere Fremdproteine wurden durch eine Anionenaustauschchromatographie mit DEAE Sepharose
abgetrennt (Aufreinigungsschritt 3). Die TfCa band an die Matrix und eluierte nach dem Anlegen des
NaCl-Gradienten im Bereich von 20 bis 25% Puffer B (Abbildung 36). Aktive Fraktionen wurden
gesammelt (DEAE-Sepharose Pool) und gegen 20 mM Phosphatpuffer pH 8 dialysiert. Dann erfolgte
der Auftrag der Enzympräparation auf eine equilibrierte Octyl-Sepharose Säule (Aufreinigungsschritt
4). Die gebundene TfCa eluierte nach Anlegen eines linearen Isopropanol-Gradienten bis 30% in
20 mM Phosphatpuffer (Abbildung 37). Die aktiven Fraktionen von 10 bis 50% Puffer B wurden
vereinigt und das enthaltene Isopropanol durch Abdampfen entfernt.
Abbildung 36: Elutionsprofil der Enzymlösung während des dritten Aufreinigungsschritts (DEAE-Sepharose, Säulenvolumen
54 ml, Puffer A: 20 mM Phosphatpuffer pH 8, Puffer B: 1 M NaCl in 20 mM Phosphatpuffer pH 8). Dargestellt sind das Proteinprofil (grau), die Esteraseaktivität (schwarz) und der Gradient der Elution (gestrichelt).
0
1
2
3
4
5
6
7
8
9
0
300
600
900
1200
0 100 200 300 400
Es
tera
se
ak
tivit
ät
(U)
UV
(2
80
nm
)
Elutionsvolumen (ml)
10%
100%
30%
0
5
10
15
20
25
30
35
40
45
-50
50
150
250
350
450
0 200 400 600 800
Es
tera
se
ak
tivit
ät
(U)
UV
(2
80
nm
)
Elutionsvolumen (ml)
15%
50%
100%

Ergebnisse und Diskussion 97
Abbildung 37: Elutionsprofil der Enzymlösung während des vierten Aufreinigungsschritts (Octyl-Sepharose, Säulenvolumen
1 ml, Puffer A: 20 mM Phosphatpuffer pH 8, Puffer B: 30% Isopropanol in 20 mM Phosphatpuffer pH 8). Dargestellt sind das Proteinprofil (grau), die Esteraseaktivität (schwarz) und der Gradient der Elution (gestrichelt).
Als letzter chromatographischer Reinigungsschritt erfolgte der Auftrag auf eine selbstgepackte
Sephadex 200 (Größenausschlusschromatographie) und die Elution der Proteine nach ihrer Größe
mit einem 0,15 M NaCl in 20 mM Tris/HCl pH 8 Elutionspuffer (Abbildung 38). Die aktiven Fraktionen
wurden im Bereich von 60-80 ml identifiziert und vereinigt. Die Aufreinigung wurde mit einer SDS-
PAGE mit Coomassie Färbung dokumentiert (Abbildung 39).
Abbildung 38: Elutionsprofil der Enzymlösung während des fünften Aufreinigungsschritts (Sephadex 200, Säulenvolumen
110 ml, Puffer: 0,15 M NaCl in 20 mM Tris/HCl pH 8). Dargestellt sind das Proteinprofil (grau) und die Esteraseaktivität (schwarz).
0
5
10
15
20
25
0
50
100
150
0 5 10 15 20 25 30 35 40
Es
tera
se
ak
tivit
ät
(U)
UV
(2
80
nm
)
Elutionsvolumen (ml)
100%
0
10
20
30
40
50
0
5
10
15
20
25
30
35
0 20 40 60 80 100 120 140 160
Es
tera
se
ak
tivit
ät
(U)
UV
(2
80
nm
)
Elutionsvolumen (ml)

Ergebnisse und Diskussion 98
97 kDa 66 kDa 45 kDa 30 kDa 20.1 kDa 14.4 kDa
1 2 3 4 5 6
Abbildung 39: SDS-PAGE der Aufreinigung der TfCa aus T. fusca KW3. Spur 1: Enzymlösung nach der Dialyse; Spur 2: Enzymlösung nach der HIC 1; Spur 3: Enzymlösung nach der AIEX; Spur 4: Enzymlösung nach der HIC 2; Spur 5: Enzymlösung nach der SEC; Spur 6: Low Molecular Weight Längenstandard (Billig et al. 2010).
Wie aus der Abbildung 39 ersichtlich ist, konnte mit den fünf Reinigungsschritten die TfCa bis zur
elektrophoretischen Reinheit isoliert werden. Es erfolgte eine 7625 fache Aufreinigung der Esterase
mit einer Ausbeute von 7% der Ausgangsenzymlösung mit einer Aktivität von 36,6 U und einer
Proteinkonzentration von 0,012 mg (Tabelle 15). Die spezifische Aktivität konnte von 0,4 U/mg auf
3050 U/mg erhöht werden.
Tabelle 15: Aufreinigungstabelle der TfCa aus T. fusca KW3 (Billig et al. 2010).
Schritt Volumen
(ml)
Aktivität
(U)
Protein
(mg)
Spezifische Aktivität
(U/mg)
Ausbeute
(%)
Kulturüberstand 33 600 526 1212,7 0,4 100
Dialyse 7 655 325,7 808,7 0,4 61,9
HIC 1 844 258 22,2 11,6 49
AIEX 46 187,6 0,24 781,7 35,7
HIC 2 4,4 83,2 0,045 1848,9 15,8
SEC 3,6 36,6 0,012 3050 7
Aufreinigungsprotokolle für einige PET-Hydrolasen wurden bereits in den letzten Jahren
veröffentlicht. Die bereits erwähnte 29 kDa Cutinase (BTA Hydrolase 1, TfH) aus T. fusca DSM 43793
konnte durch Gouda et al. 2002 und Kleeberg et al. 2005 aufgereinigt werden. Zunächst erfolgte die
Konzentrierung und Aufreinigung mittels Ultrafiltration mit einer regenerierten
Zellulosetriacetatmembran (Ausschlussgröße 10 kDa). Dabei stieg die spezifische Aktivität 2,4 fach
mit einer Ausbeute von 60,8%. Die Verwendung einer Membran mit der Ausschlussgröße von 20 kDa

Ergebnisse und Diskussion 99
führte zu einem starken Verlust bei der Ausbeute. Bei der Verwendung einer Polysulfonmembran
(Ausschlussgröße 10 kDa) lag die Rückgewinnung des Proteins bei 99%, allerdings nur mit 34,5%
Ausbeute und somit konnte nur eine geringe Aufreinigung (Faktor 1,2) erzielt werden. Es wurde
spekuliert, dass das Enzym eine starke Hydrophobizität besitzt und somit an die Membran absorbiert
hat (Gouda et al. 2002).
Tabelle 16: Aufreinigungstabelle der TfH aus T. fusca DSM 43793 (Kleeberg et al. 2005).
Schritt Volumen
(ml)
Aktivität
(U)*
Protein
(mg)
Spezifische Aktivität
(U/mg)
Ausbeute
(%)
Kulturüberstand 1600 18133 5520 3.3 100
(NH4)2SO4 Fällung 107 6777 70 97 37
KIEX 42 4584 21 218 25
HIC 9,5 2521 7 360 14
*Bestimmt mit Tributyrin als Substrat, 1 Unit entspricht der Aktivität, die 1 µmol Ester in einer Minute spaltet.
Kleeberg et al. (2005) testeten weitere Konzentrierungsschritte. Mit einer 50%
Ammoniumsulfatfällung konnte die gesamte Menge an BTA-Hydrolase ausgefällt werden, allerdings
nur ein Bruchteil davon konnte durch Waschen mit 20 mM Citratpuffer (pH 4,4) oder 20 mM Tris/HCl
Puffer (pH 9,1) wieder resuspendiert werden. Anschließend wurde die Enzymlösung auf eine S-
Sepharose Säule (Uno, Kationenaustauschchromatographie, pH 4) aufgetragen und mit einem
Salzgradienten von 0 bis 1 M NaCl eluiert. Die noch vorhandenen Verunreinigungen wurden mit einer
Hydrophoben Interaktionschromatographie entfernt. Dazu wurde der aktive Enzympool auf eine
Phenylsepharose Säule, equilibriert mit 0,5 M Ammoniumsulfat, aufgetragen und mit einem
Gradienten von 0,5 auf 0 M Ammoniumsulfat und 0 auf 30% Isopropanol eluiert. Mit dieser 3-Schritt
Aufreinigung konnte eine Steigerung der spezifischen Aktivität um den Faktor 110 mit einer Ausbeute
von 14% der Ausgangsaktivität erreicht werden (Tabelle 16, Kleeberg et al. 2005).
Bei Chen S et al. (2008) erfolgte ebenfalls die Aufreinigung einer mit Cutin induzierten p-NPB
Hydrolase aus T. fusca. Nach der Kultivierung wurden die Zellen entfernt und mit 70% (w/v)
Ammoniumsulfat die Proteine aus der Lösung ausgefällt. Das Präzipitat, gewonnen durch
Zentrifugation, wurde in Puffer gelöst und über Nacht bei 4°C dialysiert. Anschließend erfolgte eine
erneute Ausfällung mit 20% (w/v) Ammoniumsulfat. Der Überstand wurde filtriert (0.22 µm) und auf
eine Phenyl Sepharose Säule geladen. Durch die lineare Erniedrigung der
Ammoniumsulfatkonzentration erfolgte die Elution der Proteine mit p-NBP Aktivität. Nach einer
erneuten Dialyse erfolgte der Auftrag auf eine DEAE-Sepharose Säule, die Elution mit einem linearen
Natriumchlorid Gradienten (0 bis 1 M) und eine weitere Dialyse. Die letztliche Enzympräparation
wurde nochmals konzentriert (Tabelle 17, Chen S et al. 2008).

Ergebnisse und Diskussion 100
Tabelle 17: Aufreinigungstabelle der Cutin-induzierten p-NPB Hydrolase aus T. fusca (Chen S et al. 2008).
Schritt Protein
(mg)
Aktivität
(U)
Spezifische Aktivität
(U/mg)
Aufreinigung
(fach)
Ausbeute
(%)
Kulturüberstand 234,9 12294,0 52,3 1,0 100
(NH4)2SO4 Fällung 46,4 5205,2 112,2 2,1 42,3
HIC 4,7 1521,2 320,3 6,1 12,4
AIEX 2,9 1140,9 398,6 7,6 9,3
Die Aufreinigung der Cutinase aus P. mendocina ATCC 55613 erfolgte zuerst mittels einer
fraktionellen Acetonfällung (75%), um die hochviskosen Verfärbungen vom Enzym abzutrennen. Die
Rückgewinnung des Enzyms lag bei 80%, welche auf eine DEAE Säule aufgetragen wurden. Die
Cutinase band bei diesem Reinigungsschritt nicht an die Säulenmatrix, die Verunreinigung hingegen
schon. Als weiterer Aufreinigungsschritt folgte eine weitere Anionenaustauschchromatographie mit
einer QAE-Sephadex Säule. Dabei konnte ein starker Verlust an hydrolytischer Aktivität während des
Reinigungsschritts verzeichnet werden, eventuell durch die Abtrennung einer weiteren enthaltenen
p-NPB Hydrolase (von 50% Ausbeute auf 9% Ausbeute, Tabelle 17). Die Cutinase zeichnete sich
weiterhin durch nicht vorhandene Bindungseigenschaften an SP-Sephadex, CM-Cellulose, Phenyl-
Sepharose und Octyl-Sepharose aus, womit keine HIC oder Kationische Ionenaustausch-
chromatographie durchgeführt werden konnte. Die letzten Verunreinigungen konnten mittels
Größenausschlusschromatographie abgetrennt werden (Sepharose 6B und Sephadex G-100). Durch
diese 5-Schritt Aufreinigung erfolgte eine 200 fache Aufreinigung der Cutinase mit einer Ausbeute
von 3% verbleibender Enzymaktivität (Tabelle 18, Sebastian et al. 1988)
Tabelle 18: Aufreinigungstabelle der Cutinase aus P. mendocina ATCC 55613 (Sebastian et al. 1988).
Schritt Aktivität
(U)
Protein
(mg)
Spezifische Aktivität
(U/mg)
Ausbeute
(%)
Kulturüberstand 249 450 7360 44 100
Aceton Fällung 182 000 2090 87 78
AIEX 1 121 200 460 262 50
AIEX 2 22 000 11 2650 9
SEC 1 14 500 4 3360 6
SEC 2 4 430 1,1 7030 3
In einem Patent der Firma Genencore (Gray et al. 1995) wurde die Aufreinigung zweier 30 kDa großer
mit Cutin induzierter Hydrolasen beschrieben, produziert durch den Stamm P. mendocina ATCC
53552. Beide Hydrolasen unterschieden sich aufgrund ihrer Fähigkeiten zur Hydrolyse von

Ergebnisse und Diskussion 101
verschiedenen p-NP-Estern. Nach der Kultivierung mit 0,3% Apfelcutin für 12-15 Stunden erfolgte die
Filtration des Überstandes (0,22 µm). Verbleibende Enzymaktivitäten auf den Cutinresten wurden
mittels Zentrifugation gewonnen und es ergab sich eine Ausbeute von 90%. Anschließend erfolgten
die Konzentrierung der Lösung mittels Ultrafiltration und eine Dialyse, wobei eine 80%ige Ausbeute
erreicht wurde. Es erfolgte der Auftrag der Lösung auf eine Octyl-Sepharose Säule und eine
Waschung mit einem Harnstoff enthaltenden Puffer. Die Elution erfolgte mit einem linearen
Gradienten mit 50% Isopropanol und zwei aktive Fraktionen mit unterschiedlichen p-NPB/p-NPC
Verhältnissen wurden identifiziert. In Fraktion 32 besaß die Lipase 1 (mit Cutinase Aktivität) ein p-
NPB/p-NPC Verhältnis von 4,6 und in Fraktion 51 die Lipase 2 (mit Cutinase Aktivität) ein p-NPB/p-
NPC Verhältnis von 1,4. Für eine Sequenzierung erfolgte eine weitere Aufreinigung der getrennten
Hydrolasen mittels Umkehrphasen Chromatographie, wobei Lipase 1 bei 35% des Elutionspuffers
eluierte und Lipase 2 bei 39% (Gray et al. 1995).
Die erste Aufreinigung von Polyesterasen aus Pilzen erfolgte 1975 durch die Gruppe von Purdy et al.
(Purdy und Kolattukudy 1975a). Dabei wurde das extrazelluläre Protein einer 12 Tage alten Kultur
von F. solani f. pisi, gewachsen mit Apfelcutin aus Golden Delicious als einzige C-Quelle, mit 40-50%
Ammoniumsulfat ausgefällt. Im Gegensatz dazu erfolgte 1973 bereits eine Kultivierung von F. solani
f. pisi mit Cutin, isoliert aus Red Delicious Äpfeln, als einzige C-Quelle und nach 12 Tagen erfolgte die
Fällung der aktiven Proteine mit 30% Ammoniumsulfat (Purdy und Kolattukudy 1973). Das
unterschiedliche Fällungsverhalten wurde mit den enthaltenen Phenol-Verunreinigungen beim Cutin
aus dem Golden Delicious begründet, welche mit dem aktiven Protein interagierten. Dabei hatten die
extrazellulären Überstände die gleiche Proteinzusammensetzung bei beiden Kultivierungen. Es
erfolgte eine Größenausschlusschromatographie mit Sephadex G-100, wobei die p-NPP Aktivität bei
einem MG Bereich von 50-60 kDa eluierte. In diesem Bereich war auch eine Cutinaseaktivität
vorhanden, die mit der p-NPP Aktivität überlappte. Da die Aktivitäten zu diesem Zeitpunkt nicht
sauber getrennt werden konnten, wurden sie vereinigt und auf eine QAE-Sephadex Säule
aufgetragen. Bei dem pH Wert von 9 banden Fremdproteine und auch das p-NPP aktive Enzym, die
Cutinase konnte damit von den anderen abgetrennt werden. Die Elution der Fremdproteine erfolgte
bei pH 8,3 und der p-NPP Aktivität bei pH 7. Die 2. SEC mit Sephadex G-100 zeigte für den
Cutinasepeak von der QAE-Sephadex ebenfalls nur einen Proteinpeak mit Cutinaseaktivität und eine
Verschiebung des Molekulargewichts um 40-60 kDa zu 22 kDa aufgrund der Abtrennung der
phenolischen Verunreinigungen. Bei der 2. SEC des Proteinpeaks mit p-NPP Aktiviät zeigten sich ein
Hauptpeak mit Aktivität und zwei kleine Peaks ohne Aktivität. Das MG der p-NPP Hydrolase war das
gleiche, wie bereits bei der ersten SEC, was bedeutet, dass die Phenole keinen Einfluss auf die Elution
der Hydrolase hatten. Bei der anschließenden Kontrolle der Reinheit der Proteine konnten bei der

Ergebnisse und Diskussion 102
Cutinase zwei sehr eng liegende, aber trennbare Banden identifiziert werden, welche nach dem
Ausschneiden beide Cutinaseaktivität besaßen. Somit handelte es sich bei der Cutinase um zwei sehr
ähnliche Isoenzyme, welche mittels Ionenaustauschchromatographie mit einer SE-Sephadex Säule
und einem linearen NaCl Gradienten getrennt werden konnten (Cutinase I und Cutinase II).
Die p-NPP Hydrolase hatte während der 34fachen Aufreinigung die meiste Aktivität bei dem QAE-
Sephadex Schritt verloren. Lediglich eine 6,5 fache Aufreinigung vom Überstand hatte die
Homogenität der Cutinasen zur Folge, da sie die direkte Antwort auf das Wachstum des Pilzes mit
Cutin als einzige C-Quelle in hoher Konzentration waren. Das Ausgangsmedium enthielt 33 g Cutin
und produziert wurden 48 mg Cutinase I und 20 mg Cutinase II nach der Aufreinigung (Purdy und
Kolattukudy 1975a).
Analog dazu wurden aus F. roseum culmorum mit derselben Kultivierung und Aufreinigung eine
Cutinase (MG 24,3 kDa) und eine p-NPP Hydrolase isoliert. Diese Cutinase ähnelte aufgrund ihrer
Aminosäurezusammensetzung eher der Cutinase I aus F. solani f. pisi (Soliday und Kolattukudy 1976).
Bei der Kultivierung der fünf Pilz- bzw. Bakterienkulturen F. roseum culmorum,
F. roseum sambucinum, Ulocladium consortiale, S. scabies und Helminthosporium sativum war die
spezifische Aktivität von F. roseum sambucinum 50fach höher im extrazellulären Überstand
gegenüber den anderen Kulturen. Die zuvor von Purdy und Kolattukudy 1975 entwickelte
Aufreinigungsmethode wurde erfolgreich auch für diese extrazellulären Enzyme angewandt. Nach
der Ammoniumsulfat-Fällung und der SEC mit Sephadex G-100 konnten auch zwei Peaks identifiziert
werden, wovon der vordere cutinolytische Aktivität besaß und der hintere p-NPP Aktivität. Diese
Beobachtungen wurden auch von Purdy und Kollatukudy 1975 und von Soliday und Kolattukudy 1976
gemacht.
Tabelle 19: Aufreinigungstabelle der Cutinasen und p-NPP Hydrolase aus F. solani f. pisi (Purdy und Kolattukudy 1975a).
Schritt Aktivität (%)
Cutinase
Aktivität (%)
p-NPP Hydrolase
Spezifische Aktivität
(104 dpm/µg)
Cutinase1
Spezifische Aktivität
(µmol/mg)
p-NPP Hydrolase2
Kulturüberstand 100 100 1.2 11
(NH4)2SO4 Fällung 67 40 2.0 12
SEC 1 61 26 4,1 17
AIEX 1 36 5 7,6 123
SEC 2 5 340
AIEX 2 30(I)
13(II)
7,8
1 Bestimmt mit Cutin als Substrat.
2 Bestimmt mit p-NPP als Substrat.

Ergebnisse und Diskussion 103
Die Mikroorganismen produzierten alle nur eine Cutinase mit einem Molekulargewicht von 24,3-
26,3 kDa im Vergleich zu F. solani f. pisi. Für die fünf Cutinasen ergab sich eine 6-16 fache
Aufreinigung mit 16-28% Ausbeute (Tabelle 20, Lin und Kolattukudy 1980a).
Tabelle 20: Aufreinigungstabelle der extrazellulären Cutinasen (Lin und Kolattukudy 1980a).
F. roseum sambucinum U. consortiale H. sativum S. scabies
Schritt Protein
(mg)
Spez. Aktivität
(103dpm/µg)
Protein
(mg)
Spez. Aktivität
(103dpm/µg)
Protein
(mg)
Spez. Aktivität
(103dpm/µg)
Protein
(mg)
Spez. Aktivität
(103dpm/µg)
Kulturüberstand 660 5 410 0,11 210 0,11 310 0,1
(NH4)2SO4 Fällung 449 6 148 0,18 69 0,13 102 0,13
SEC 1 233 7 77 0,3 46 0,4 60 0,35
AIEX 1 25 28 10 1,7 4 1 6 1,2
AIEX 2 20 31 7 1,8 3 1,2 4 1,2
Die spezifische Aktivität wurde mit Cutin als Substrat bestimmt.
Fernando et al. (1984) kultivierte einen F. solani pisi Stamm im Mineralmedium mit Suberin als
einzige C-Quelle. Im Mediumüberstand konnte nur eine geringe Menge an p-NPB Aktivität detektiert
werden. Daraufhin wurde die Myzelmatte mit 0,4% Triton X-100 in 0,2 M Natriumphosphatpuffer
gewaschen und es konnte eine 13 fach höhere Aktivität nachgewiesen werden. Das bedeutet, dass
nach der Kultivierung 93% der Gesamtaktivität am Myzel oder am Suberin gebunden vorlag. Beim
Auftrag auf die QAE-Sepharose wurden die farblichen Verunreinigungen gebunden, das aktive Enzym
jedoch eluierte direkt wieder. Bei pH 4,8 band das aktive Enzym anschließend an die SP-Sephadex C-
25 Säule, die vorhandenen Verunreinigungen wanderten durch die Säule durch. Die Elution mit
einem NaCl Gradienten ergab einen Elutionspeak, der nach Auftrag auf eine SP-Sephadex Säule 2
getrennte Peaks mit p-NPB und Cutinaseaktivität hervorbrachte. Dabei handelte es sich um 2
Isoenzyme mit einem Molekulargewicht von 23 kDa. Der Vergleich der Aminosäurezusammen-
setzung der Suberin- und Cutin-induzierten Cutinasen (Purdy und Kolattukudy 1975a) zeigte, dass
diese Isoenzyme sich lediglich aufgrund eines nicht detektierten Methionins unterschieden. Somit
besteht die Möglichkeit, dass die Cutin- und Suberin-induzierten Isoenzyme die gleichen sind
(Fernando et al. 1984).
Die PET-Hydrolase aus P. citrinium wurde aufgrund ihrer Aktivität gegenüber 3PET (BETEB)
aufgereinigt. Zunächst erfolgte eine fraktionelle Ultrafiltration mit den Ausschlussgrößen von 50 und
5 kDa. Die PET-Hydrolase wurde 67,9 fach bis zur elektrophoretischen Reinheit aufgereinigt, die
Ausbeute betrug 6% nach AIEX und SEC. Die spezifische Aktivität betrug am Ende 14,8 nkat/mg und
das Molekulargewicht wurde auf 14,1 kDa bestimmt. Somit ist diese PET-Hydrolase die kleinste
bekannte PET abbauende Hydrolase (Tabelle 21, Liebminger et al. 2007).

Ergebnisse und Diskussion 104
Tabelle 21: Aufreinigungstabelle der PET-Hydrolase aus P. citrinum (Liebminger et al. 2007).
Schritt Volumen
(ml)
Aktivität
(nkat)
Protein
(mg)
Spezifische Aktivität
(nkat/mg)
Ausbeute
(%)
Kulturüberstand 200,0 120,6 552,0 0,2 100
AIEX 40,0 22,4 19,8 1,1 18,6
Entsalzung 30,0 18,0 12,7 1,4 15,0
AIEX 9,0 9,1 2,2 4,1 7,5
SEC 2,0 7,3 0,49 14,8 6,0
Die Rohenzymlösungen mit den hydrolysierenden Enzymen von F. oxysporum LCH 1 und
F. solani f. sp. pisi wurden mittels Zentrifugation und Ausfällung gewonnen und es erfolgte mit ihnen
die Untersuchung der Hydrolyse von PET-Fasern (Nimchua et al. 2007).
Die für die Aufreinigung der PET-Hydrolasen eingesetzten Methoden variierten bei der
Aufkonzentrierung des extrazellulären Überstandes, es kamen Ultrafiltration, verschiedene Fällungen
und die Gefriertrocknung zum Einsatz. Diese Schritte waren meist mit einem höheren Verlust an
aktivem Enzym verbunden, allerdings von Nöten für die weitere Aufreinigung, da das extrazelluläre
Enzym in einer zu hohen Verdünnung vorlag (Purdy und Kolattukudy 1973, Soliday und Kolattukudy
1976, Sebastian et al. 1988, Gouda et al. 2002, Kleeberg et al. 2005). Für die TfCa wurden auch
verschiedene Aufkonzentrierungsmethoden getestet mit dem Ergebnis, dass die
Aufreinigungsverluste mit der Gefriertrocknung am geringsten waren. Bei der anschließenden
chromatographischen Aufreinigung kamen meistens eine IEX oder SEC zum Einsatz, seltener eine HIC
aufgrund der stark hydrophoben Eigenschaften der PET-Hydrolasen und der daraus resultierenden
schlechten Rückgewinnung des Enzyms vom Säulenmaterial. Die Aufreinigung der hydrolytischen
Enzyme konnte mit einer 3 bis 5 Schritt Aufreinigungsmethode bis zur elektrophoretischen Reinheit
bewerkstelligt werden. Dabei lagen die Ausbeuten bei 3-30%, im Vergleich dazu lag die Ausbeute der
TfCa bei 7% (Billig et al. 2010).
Eine interessante Besonderheit konnte bei der Aufreinigung der Cutin-induzierten Hydrolasen aus
den Pilzen F. solani f. pisi, F. roseum culmorum, F. roseum sambucinum, U. consortiale und
H. sativum, aber auch beim Actinomyceten S. scabies identifiziert werden. Parallel zur Cutinase
produzierten diese Stämme mit Cutin als einzige C-Quelle ebenfalls eine Esterase, welche p-NPB und
p-NPP spalten konnte (Purdy und Kolattukudy 1975a, Lin und Kolattukudy 1980a). Ebenfalls besaß
diese p-NPP Esterase ein MG von 52 kDa (Purdy und Kolattukudy 1975a). Der Pilz F. solani f. pisi
unterschied sich zusätzlich von den anderen Kulturen, indem er zwei Isoenzyme während des
Wachstums mit Cutin bildete (Purdy und Kolattukudy 1975a). Bei allen weiteren Aufreinigungen von
PET-Hydrolasen wurde auf die Produktion einer zusätzlichen p-NPB oder p-NPP Esterase nicht
geachtet. Bei der Aufreinigung der Cutinase aus P. mendocina konnte beim ersten

Ergebnisse und Diskussion 105
Anionenaustauschschritt ein starker Verlust der p-NPB Aktivität verzeichnet werden, was mit einer
möglichen Abtrennung einer weiteren Esterase begründet wurde (Sebastian et al. 1988). Aufgrund
der bisherigen Ergebnisse scheint diese Annahme als sehr wahrscheinlich. Die Aufreinigungen der
TfH aus T. fusca DSM 43793 und der PET-Hydrolase aus P. citrinum erfolgte mit BTA bzw. 3PET
induzierten Kulturen und deshalb könnte die Produktion einer mitinduzierten Esterase nicht
stattgefunden haben oder in zu geringen Mengen für einen direkten Nachweis erfolgt sein (Kleeberg
et al. 2005, Liebminger et al. 2007). Fernando et al. (1984) kultivierte den Pilz F. solani f. pisi mit
Suberin als Induktor. Während der anschließenden Aufreinigung der Suberin-hydrolysierenden
Enzyme wurde keine zusätzliche Esterase detektiert bzw. nicht nach ihr gesucht.
Es wäre interessant, die beiden Esterasen TfCa und die eukaryotische Esterase in ihren Sequenzen
und katalytischen Zentren zu vergleichen und herauszufinden, warum die eukaryotische Esterase
unter Zusatz von Cutin produziert wird und ob die Produktion auch unter Zusatz eines synthetischen
Polyesters nachweisbar wäre. Weiterhin ob und wie sich die beiden Esterasen hinsichtlich ihrer PET-
Hydrolyseaktivität unterscheiden würden.
Eine weitere Besonderheit war ebenfalls die beobachtete Hydrophobie der PET-Hydrolasen
begründet durch ihre Affinität zu den abzubauenden hydrophoben Substraten. Diese Eigenschaft
wurde bei der Aufreinigung der TfCa ausgenutzt.
3.2.1.2 Aufreinigung der rekombinanten PET-Hydrolasen
Die TfCa und die Cutinasen TfCut1 und TfCut2 aus T. fusca KW3 wurden bereits erfolgreich kloniert
und mit Hilfe verschiedener E. coli Stämme exprimiert (Oeser et al. 2010, Chen S et al. 2008). Die
Hydrolaseproduktion erfolgte bei einer optimalen Kultivierungstemperatur von 18°C im 5 L
Fermenter. Bei der niedrigeren Temperatur erfolgte eine geringere Bildung an Einschlusskörpern der
produzierten Enzyme und somit stand mehr aktives Protein nach der Kultivierung für die
Aufreinigung zur Verfügung. Mit den optimierten Bedingungen erfolgte die rekombinante Produktion
der TfCa mit einer 4500fach höheren Aktivität gegenüber dem Wildtyp und die spezifische Aktivität
(U/mg) stieg um mehr als das 30fache (Oeser et al. 2010). Die rekombinante TfCa, gewonnen aus
dem löslichen Zellinhalt, wurde mittels Nickel-Affinitätschromatographie (Abbildung 40) und
Größenausschlusschromatographie (Abbildung 41) aufgereinigt. Es erfolgte eine Aufreinigung um das
12fache (spezifische Aktivität 35 U/mg) (Tabelle 22, Abbildung 44A).
Unter den gleichen Kultivierungsbedingungen wurden ebenfalls die beiden Cutinasen TfCut1 und
TfCut2 aus T. fusca KW3 in E. coli produziert und mittels Affinitätschromatographie (Abbildung 42)
und Größenausschlusschromatographie (Abbildung 43) aufgereinigt. Da es sich bei den beiden
Cutinasen um Isoenzyme mit ähnlichen Eigenschaften handelte, sind hier nur die Aufreinigungdaten

Ergebnisse und Diskussion 106
für die Cutinase II exemplarisch dargestellt. Nach der 2-Schritt Aufreinigung der TfCut2 konnte eine
9fach höhere Reinheit nachgewiesen werden (spezifische Aktivität TfCut2 26 U/mg) (Tabelle 23,
Abbildung 44B).
Abbildung 40: Elutionsprofil der TfCa während des ersten Aufreinigungsschritts (HisTrap, Säulenvolumen 10 ml, Puffer A: 50 mM NaH2PO4, 300 mM NaCl; pH 8,0, Puffer B: 50 mM NaH2PO4, 300 mM NaCl, 250mM Imidazol, pH 8). Dargestellt sind das Proteinprofil (grau), die p-NPB Aktivität (schwarz) und der Gradient der Elution (gestrichelt).
Abbildung 41: Elutionsprofil der TfCa während des zweiten Aufreinigungsschritts (Sephadex 200 prep grade, Säulenvolumen 320 ml, Puffer: 0,15 M NaCl, 20 mM Phosphatpuffer pH 8). Dargestellt sind das Proteinprofil (grau), die p-NPB Aktivität (schwarz).

Ergebnisse und Diskussion 107
Abbildung 42: Elutionsprofil der TfCut2 während des ersten Aufreinigungsschritts (HisTrap, Säulenvolumen 10 ml, Puffer A: 50 mM NaH2PO4, 300 mM NaCl; pH 8,0, Puffer B: 50 mM NaH2PO4, 300 mM NaCl, 250 mM Imidazol, pH 8). Dargestellt sind das Proteinprofil (grau), die p-NPB Aktivität (schwarz) und der Gradient der Elution (gestrichelt).
Abbildung 43: Elutionsprofil der TfCut2 während des zweiten Aufreinigungsschritts (Sephadex 200 prep grade, Säulenvolumen 320 ml, Puffer: 0,15 M NaCl, 20 mM Phosphatpuffer pH 8). Dargestellt sind das Proteinprofil (grau), die p-NPB Aktivität (schwarz).

Ergebnisse und Diskussion 108
Abbildung 44: SDS-PAGE der Aufreinigung der rekombinant produzierten TfCa in E. coli (A) und der rekombinant produzierten TfCut2 in E. coli (B). Spur M: Low Molecular Weight Längenstandard; Spur 1, 4: Rohenzymlösung nach Zellaufschluss; Spur 2, 5: Enzymlösung nach Ni-Affinitätschromatographie; Spur 3, 6: Enzymlösung nach SEC.
Tabelle 22: Aufreinigungstabelle der rekombinanten TfCa.
Schritt Volumen
(ml)
Aktivität
(U)
Protein
(mg)
Spezifische Aktivität
(U/mg)
Ausbeute
(%)
Aufreinigung
(fach)
Zellfreier
Extrakt
196 4606 1535 3 100 1
HisTrap 44 2077 112 19 45 6
SEC 75 1538 44 35 33 12
Tabelle 23: Aufreinigungstabelle der rekombinanten TfCut2.
Schritt Volumen
(ml)
Aktivität
(U)
Protein
(mg)
Spezifische Aktivität
(U/mg)
Ausbeute
(%)
Aufreinigung
(fach)
Zellfreier
Extrakt
195 2477 1482 2 100 1
HisTrap 27 1183 54 22 26 7
SEC 50 895 35 26 19 9
Die TfH wurde durch Dresler et al. 2006 auch bereits rekombinant in E. coli TG1 mit dem Plasmid
pCYTEXP1-OmpA-bta1-His6 produziert, sekretiert und aufgereinigt. Im Vergleich mit der Wildtyp
Produktion konnte rekombinant extrazellulär eine 1,5-2fach höhere volumetrische Aktivität in den
Batch Kulturen und eine 20fach höhere in den Fed-Batch Kulturen erzielt werden (unter Glucose
Limitation). Die volumetrische Aktivität intrazellulär konnte sogar um das 50fache gesteigert werden
und somit stieg die Gesamtproduktion um mehr als das 100fache gegenüber dem Wildtyp. Aufgrund
des angehängten His6-Tags erfolgte die einfache Aufreinigung des periplasmatischen und

Ergebnisse und Diskussion 109
zytoplasmatischen (Einschlusskörper) Proteins in zwei Schritten. 57% Ausbeute konnte erzielt
werden und eine 30fache Aufreinigung mit einer spezifischen Aktivität von 445 U/mg wurde erreicht
(Tabelle 24). Zum Vergleich beim Wildtyp konnte nur 14% Ausbeute nachgewiesen werden (Dresler
et al. 2006).
Tabelle 24: Aufreinigungstabelle der rekombinanten periplasmatischen (pp-rTfH) und zytoplasmatischen (zp-rTfH) TfH (Dresler et al. 2006).
Schritt Aktivität
(103 U/L)
Protein
(mg/L)
Spezifische Aktivität
(U/mg)
Ausbeute
(%)
Aufreinigung
(fach)
pp-rTfH Extrakt 8,57 587 15 100 1
HisTrap 6,53 15 436 71 29
SEC 4,89 11 445 57 30
zp-rTfH Extrakt 26,52 2570 10,3 100 1
Hitzebehandlung 22,52 1470 15,3 85 1,5
Ultraschall- und
Hitzebehandlung
28,89 1410 20,5 109* 2
* Rohextrakt nach Ultraschall und Zentrifugation.
In weiterführenden Studien erfolgte nach einer Codon-Optimierung die erfolgreiche rekombinante
Produktion der TfH mit B. megaterium WH323. Bei einer pH-kontrollierten Batchkultivierung konnte
eine weitere Steigerung auf 16,1 mg/L rTfH erzielt werden. Nach der Induktion durch Xylose erfolgte
ein starker Anstieg der rTfH Produktion für 4 Stunden. Beim Vergleich der produzierten Mengen an
rTfH mittels B. megaterium und E. coli (Dresler et al. 2006) konnte für die Batchkultivierung mit LB
Medium und Hochzelldichten-Kultivierung mit synthetischem Medium ähnliche Ergebnisse erzielt
werden (Tabelle 25). Beide zeigten eine 50-100fach höhere Produktion gegenüber dem Wildtyp
(Yang et al. 2007).
Tabelle 25: Expression der rekombinanten TfH in E. coli und B. megaterium (Yang et al. 2007).
Kultivierung Medium
B. megaterium WH323
U/g U/L
E. coli TG 1
U/g U/L
Batch LB 2651 7953 4000 8000
Batch Definiert 116 886 770 7300
Hochzelldichten-
Kultivierung
Definiert 187 6098 76/227* 5500/12000*
* abhängig von der Induktionsphase (Temperaturänderung)

Ergebnisse und Diskussion 110
Die Aufreinigung der rTfH erfolgte ebenfalls mittels Ni-Affinitätschromatographie. Die Protokolle
einiger Proteinaufreinigungen verschiedener Kultivierungsansätze sind in der nachfolgenden Tabelle
zusammengefasst (Yang et al. 2007).
Tabelle 26: Übersicht zur Affinitätsaufreinigung der His6-Tag rTfH produziert durch B. megaterium WH323-pYYBm9 gewachsen in einer LB-Batch Kultivierung und Hochzelldichten-Kultivierung (HZDK) mit A5 Medium. I:ohne
Vorhbehandlung; II: 50°C für 10 min; III: 18 Stunden Dialyse bei 4°C (Yang et al. 2007).
Kultivierung Aktivität
(U)
Protein
(µg)
Spezifische Aktivität
(U/mg)
Ausbeute
(%)
Aufreinigung
(fach)
LB-I Extrakt
HisTrap
37
20
238
41
157
494
100
54
1,0
3,2
LB-II Extrakt
HisTrap
35
17
213
37
162
471
100
50
1,0
2,9
LB-III Extrakt
HisTrap
23
13
147
42
159
310
100
55
1,0
1,9
HZDK-I Extrakt
HisTrap
49
25
203
83
240
307
100
52
1,0
1,3
HZDK-II Extrakt
HisTrap
47
23
221
63
213
367
100
49
1,0
1,7
HZDK-III Extrakt
HisTrap
29
10
161
45
178
215
100
34
1,0
1,2
Die gleichen Cutinasen wurden 2008 von Chen S et al. 2008 als Cutinase 1 und Cutinase 2 aus T. fusca
WSH03-11 nach vorherigen Vermehrungs- und Modifikationsschritten in den pET20b(+) Vektor
kloniert und anschließend in E. coli Rossetta (DE3) pLysS einzeln transformiert (Chen J et al. 2008).
Das Wachstum der transformierten Klone erfolgte unter Zusatz von Ampicillin und Chloramphenicol
und bei einer Zelldichte von 1,5 (OD600) erfolgte die Induktion mit IPTG. Nach 16 Stunden Expression
erfolgte die Zellernte, Konzentrierung des Kulturüberstand, Dialyse und Ni-Affinitätschromatographie
(Chen S et al. 2008).
Die p-NPB Aktivität im Kulturüberstand der transformierten E. coli Zellen mit dem Plasmid pET20b-
Cutinase 2 lag bei 180 U/ml, und damit um das 782fache höher als bei den untransformierten Zellen
und um das 9,5fache höher als bei den Cutin-induzierten Zellen von T. fusca. Die p-NPB Aktivität im
Kulturüberstand der transformierten E. coli Zellen mit dem pET20b-Cutinase 1 Vektor lag bei
69 U/ml, und damit um das 300fache höher als bei den untransformierten Zellen und um das
3,6fache höher als in den Cutin-induzierten T. fusca Zellen. Die rekombinanten Enzyme wurden bis
zur homogenen Reinheit mit einem Schritt mittels Ni-Affinitätschromatographie aufgereinigt und

Ergebnisse und Diskussion 111
besaßen eine spezifische Aktivität von 458 U/mg (Cutinase 2) und 223 U/mg (Cutinase 1) (Chen S
et al. 2008, Chen J et al. 2008).
Sinsereekul et al. (2010) konstruierten einen transkonjuganten S. rimosus R7 mit dem Vektor pIJ-BTA,
der das Gen für eine Ecoflex® Filme-abbauende Hydrolase enthielt. Das Gen wurde aus dem Isolat
Thermobifida sp. BCC23166 gewonnen, isoliert aus einer Probe von einer Deponie. Es erfolgte die
Ligation des Gens in den pIJ8600 Vektor und der Transfer in S. rimosus R7 mittels intergenetischer
Konjugation. Dieses entwickelte Expressionssystem zeichnete sich durch die Produktion des
gesamten nativen Proteins aus dem unmodifizierten Gen ohne Codon Optimierung und Modifikation
durch Fusionsproteine aus. Allerdings war zum damaligen Zeitpunkt die anfängliche Ausbeute aus
der Expression geringer (0,058 mg/L) verglichen mit den Expressionssystemen in E. coli oder
B. megaterium. Das produzierte rekombinante Enzym (mit His6Tag) wurde anschließend bis zu einer
95% Reinheit mittels einer Ein-Schritt Aufreinigung durch eine Ni-Affinitätschromatographie
aufgereinigt. Dabei betrug das Kulturvolumen am Beginn 500 ml und die Ausbeute nach der
Reinigung 82,5% (Sinsereekul et al. 2010).
In neuesten Studien wurde eine thermoaktive Hydrolase Est119 aus Thermobifida alba AHK119
rekombinant ebenfalls in E. coli produziert. Dazu wurde das isolierte Gen der Hydrolase ohne die
Signalsequenz in den Vektor pQE80L kloniert, mit den Vektor pQE80L-est119 als Produkt. Das
rekombinante Protein mit einem N-terminalen His6Tag konnte erfolgreich mit dem E. coli Rosetta-
gami B (DE3) nach der Zugabe von IPTG produziert werden. Die rekombinante 29 kDa Cutinase
konnte um das 2,3fache mittels einer Ein-Schritt Aufreinigung mit der Ni-Affinitätschromatographie
Säule aus dem zellfreien E. coli Extrakt gewonnen werden (Spezifische Aktivität 1.37 U/mg, Hu et al.
2010).
Die Cutinasen aus P. mendocina ATCC 53552 wurden nach der Identifikation der Gene einzeln in das
Plasmid pSNtacII kloniert und rekombinant in E. coli JM101 exprimiert. 74% der Aktivität der
Hydrolase 1 lagen nach der Kultivierung extrazellulär vor, 17% Aktivität in den Zellen, 2% im
periplasmatischen Raum und 7% an der Cytoplasmamembran. Die Aufreinigung der Hydrolasen
erfolgte mittels Octyl-Sepharose analog zur bereits erläuterten Aufreinigung der Wildtyp Enzyme. Die
Cutinase 1 zeigte eine p-NPB Aktivität von 3750 U/mg Protein, bestand aus 259 Aminosäuren ohne
Signalsequenz und zeigte ein p-NPB/p-NPC Verhältnis von 7 (Cutinase 2 besaß ein Verhältnis von 1)
(Gray et al. 1995).
Für die präparative Produktion der F. solani Cutinase erfolgte die Kultivierung von E. coli WK6 Zellen,
welche das Plasmid pMa/c5-CUF enthielten. Bei einer optischen Dichte von 410 erfolgte die
Induktion mit IPTG über 6 bis 12 Stunden (Carvalho et al. 1999). Aufschlussmethoden der Cutinase
waren kombinierte Methoden wie z.B. osmotischer Schock und Präzipitation der Verunreinigungen
mit verdünnter Essigsäure. Anschließend erfolgte die Aufreinigung mittels Kationenaustauscher, wie

Ergebnisse und Diskussion 112
SP-Sephadex oder in zwei Anionenaustauschschritten, z.B. mit DEAE-Cellulose und MonoQ- oder Q-
Sepharose HP. Die endgültige Ausbeute der Aufreinigung lag bei 51% und der Aufreinigungsfaktor bei
10. Die Aufreinigung der Cutinase, produziert durch Hefezellen, verlief ähnlich wie mit den
Bakterienzellen, allerdings folgte am Schluss eine Affinitätschromatographie mit einer Con-A
Sepharose-4B Säule zur Entfernung der glykosylierten Fraktionen (Carvalho et al. 1999). Neuere
Aufreinigungsmethoden sind die Umkehrmyzellen Extraktion und das wässrige Zwei-Phasen System
als Alternative für die Säurepräzipitation, Filtration, Zentrifugation oder Dialyse (Carvalho et al.
1999). Mit der Umkehrmyzellen Extraktion konnten Ausbeuten von 5-50% erreicht werden mit
Aufreinigungsfaktoren von 1,2 bis 10,2. Bei der direkten Extraktion der Cutinase aus dem Zellextrakt
mit Zellbruchstücken der E. coli Zellen lag die Ausbeute bei optimalen Bedingungen bei 54,4%
(Carvalho et al. 1999). Beim wässrigen Zwei-Phasen System lagen die Ausbeuten immer sehr hoch,
meist über 90%. Bei einer Ein-Schritt Aufreinigung der Cutinase aus E. coli Zelltrümmern lagen die
Ausbeute bei 91% und der Aufreinigungsfaktor bei 4,1, bei einem Zeitaufwand von nur 20 min
(Carvalho et al. 1999). Die Cutinase aus F. solani pisi wurde in B. subtilis Zellen mit dem Vektor
pBSMuL3 während des Wachstums im Medium mit Kanamycin produziert. Die Proteine des
Kulturüberstands wurden mit Ammoniumsulfat ausgefällt, zentrifugiert, resuspendiert und dialysiert.
Nach einer erneuten Präzipitation der verunreinigenden Proteine mit Ammoniumsulfat erfolgte die
Filtration der Lösung und der Auftrag auf eine Phenyl HP-Sepharose Fast Flow Säule. Die Elution der
Cutinase erfolgte mit einem linearen Gradienten von 20% auf 0% Ammoniumsulfat im Elutionspuffer.
Die Fraktionen mit p-NPB Aktivität wurden anschließend auf eine CM-Sepharose Säule aufgetragen
und mit einem linearen Gradienten von 0 bis 1 M Natriumchlorid eluiert (Chen S et al. 2008).
Die Cutinase aus T. insolens wurde in A. niger mit dem Vektor pA2L79 rekombinant produziert. Nach
der Produktion erfolgte die Gewinnung der intrazellulären Cutinase mittels Zentrifugation und
Ultrafiltration. Mit dem Überstand erfolgte eine Ammoniumsulfatfällung. Das Präzipitat mit der
Cutinase wurde mittels Zentrifugation gewonnen, in Wasser gelöst und mit Essigsäure auf pH 6
eingestellt. Zur Entfernung der enthaltenen Proteasen passierte die Lösung eine Bacitracin-Agarose
Säule und die eluierten aktiven Fraktionen wurden auf eine Kationenaustausch SP-Sepharose HP
Säule aufgetragen. Die Cutinase wurde mit einem linearen Salzgradienten eluiert (Sandal et al. 1996).
Bisher wurden verschiedenste bekannte Systeme für die rekombinante Expression von Hydrolasen
und deren Aufreinigung zusammengefasst. Bei der Wahl des jeweiligen Expressionssystems besteht
keine Möglichkeit zur Diskussion, da die Auswahl nach dem „try and error“ Verfahren abläuft und
jedes der Systeme seine spezifischen Vor- und Nachteile aufweist. Eines ist allerdings für alle
rekombinanten Produktionen zu empfehlen, die Verwendung eines His6Tags für eine deutlich
einfachere und spezifischere Aufreinigung des rekombinanten Proteins, wie es auch für die beiden in
diesem Abschnitt beschriebenen Hydrolasen aus T. fusca KW3, produziert mittels E. coli, erfolgte.

Ergebnisse und Diskussion 113
Dadurch sind Ausbeuten von über 50% keine Seltenheit. In dieser Hinsicht sollte die Aufreinigung der
beiden rekombinanten Hydrolasen aus T. fusca KW3 noch weiter optimiert werden, damit eine
Steigerung der Ausbeute erfolgen kann.
3.2.2 pI der TfCa
Der bestimmte pI der TfCa lag bei 4,8 (theoretischer Wert 4,79).
Abbildung 45: PhastGel der Bestimmung des isoelektrischen Punktes der TfCa. (linkes Gel PhastGel IEF 3-9, rechtes Gel PhastGel IEF 4-6,5) Spur 1: 26,7 ng TfCa; Spur 2: 66,7 ng TfCa; Spur 3: 133 ng TfCa; Spur 4: 26,7 ng TfCa; Spur 5: 53,4 ng TfCa; Spur 6: Kalibrierungskit für die pI Bestimmung, Breiter pI Bereich (pH 3-10, GE Healthcare).
3.2.3 Molare Masse der TfCa
Die Molekulargewichtsbestimmung erfolgte mit einer SDS-PAGE mit der partiell aufgereinigten
Enzymlösung und des Roti®-Mark SDS Proteinstandards (Fa. Carl Roth GmbH).
St 1 2 3
9.30
8.65
8.458.15
7.35
6.85
6.55
5.85
5.20
4.55
St 4 5
6.85
6.55
5.85
5.20
4.55

Ergebnisse und Diskussion 114
Abbildung 46: Laufstrecken der Proteine des Roti®-Mark SDS Proteinstandards (Fa. Carl Roth GmbH) aufgetragen gegen den
Logarithmus ihres Molekulargewichtes.
Nach erfolgter elektrophoretischer Auftrennung wurden die Laufstrecken der Proteine des Standards
bestimmt und gegen den Logarithmus ihres Molekulargewichts aufgetragen (Abbildung 46). Für die
untersuchte TfCa ergab sich daraus ein exaktes Molekulargewicht von 52,4 kDa, was mit dem
theoretischen MG von 52,94 kDa gut übereinstimmt.
3.2.4 Temperatur- und pH-Stabilität der TfCa
Für die Bestimmung der Stabilität der TfCa bei verschiedenen pH-Werten wurde die Esterase in
100 mM Puffern mit pH-Werten von 3 bis 11 für 7 Tage inkubiert und die Esteraseaktivitäten zu
verschiedenen Zeiten mit p-NPB als Substrat spektrophotometrisch bestimmt. Ein Citrat-Phosphat-
Puffer wurde für pH 3 bis 7 verwendet, Phosphatpuffer für pH 8 und Glycin-NaOH Puffer für pH 9 bis
11. Die Inkubationen erfolgten bei Raumtemperatur.
Bei der Inkubation der TfCa bei verschiedenen pH-Werten wurde festgestellt, dass die Esterase hohe
Stabilität im alkalischen Bereich auch über einen längeren Zeitraum besitzt (Abbildung 47). Nach 7
Tagen Inkubation konnte bei pH 9 und 10 noch eine relative Aktivität von 85% nachgewiesen werden.
Aber auch bei pH 5 und 11 zeigte die TfCa nach 7 Tagen Inkubation noch eine relative Aktivität von
22% und 16%. Das bedeutet, dass die Esterase bei kürzeren Inkubationen, wie zum Beispiel nach 24
Stunden (Abbildung 48), im pH-Bereich von 5 bis 11 noch eine relative Aktivität von 40% bis 100%
besitzt und somit einen breiten Einsatzbereich vorweisen kann.
y = -0,089x + 2,037
R² = 0,993
1
1,2
1,4
1,6
1,8
2
2 4 6 8 10
lg M
W
Laufstrecke (cm)

Ergebnisse und Diskussion 115
Abbildung 47: Esteraseaktivitäten der TfCa über einen Zeitraum von 7 Tagen bei Inkubation in unterschiedlichen pH-
Werten von 4 bis 11 bestimmt mit p-NPB als Substrat.
Abbildung 48: Esteraseaktivitäten der TfCa nach 24 Stunden Inkubation in verschiedenen pH-Werten von 3 bis 11 bestimmt
mit p-NPB als Substrat.
Für die Stabilitätsuntersuchung der TfCa bei verschiedenen Temperaturen wurde die Esterase in
100 mM Phosphatpuffer pH 8 bei den Temperaturen 40°C, 50°C und 55°C für 7 Tage inkubiert. Die
Esteraseaktivitäten wurden mit p-NPB als Substrat spektrophotometrisch bestimmt.
0
20
40
60
80
100
0 1 2 3 4 5 6 7
Rela
tive A
ktiv
ität (%
)
Inkubationszeit (d)
pH 4 pH 5 pH 6 pH 7 pH 8 pH 9 pH 10 pH 11
0
20
40
60
80
100
3 4 5 6 7 8 9 10 11
Rela
tive A
ktiv
ität (%
)
pH

Ergebnisse und Diskussion 116
Abbildung 49: Esteraseaktivitäten der TfCa über einen Zeitraum von 7 Tagen bei Inkubation bei 40°C, 50°C und 55°C bestimmt mit p-NPB als Substrat (Billig et al. 2010).
Abbildung 50: Esteraseaktivitäten der TfCa nach 24 Stunden Inkubation bei 30°C, 40°C, 50°C und 55°C bestimmt mit p-NPB als Substrat.
Die TfCa besitzt ebenfalls eine hohe Stabilität bei Temperaturen von 40 bis 50°C, (Abbildung 49).
Nach 7 Tagen Inkubation bei diesen Temperaturen war noch eine relative Aktivität zwischen 93 und
100% nachweisbar. Bei 40°C erfolgte über diesen Zeitraum keine Verminderung der Aktivität, die
Aktivität blieb über den gesamten Zeitraum konstant. Bei 55°C hingegen konnte nach 24 Stunden
0
20
40
60
80
100
0 1 2 3 4 5 6 7
Rela
tive A
ktiv
ität (%
)
Inkubationszeit (d)
40°C 50°C 55°C
0
20
40
60
80
100
30 35 40 45 50 55
Rela
tive A
ktiv
ität (%
)
Temperatur (°C)

Ergebnisse und Diskussion 117
Inkubation lediglich 1% Restaktivität detektiert werden (Abbildung 50). Bei höheren Temperaturen
erfolgte die Inaktivierung der Esterase in einem noch kürzeren Zeitraum.
3.2.5 Wirkung von Inhibitoren auf die TfCa
Um eine Einordnung der TfCa in der Enzymgruppe der Hydrolasen durchzuführen, wurde die
Stabilität des Enzyms gegenüber verschiedener spezifischer und unspezifischer Inhibitoren
untersucht. Die Inhibitoren, welche eingesetzt wurden, waren EDTA, PMSF, TPCK und DTT. Sie
wurden nach Anweisung in Puffer, Isopropanol oder Ethanol gelöst. Die eingesetzten
Konzentrationen der Inhibitoren lagen bei 0,1 mM, 1 mM und 10 mM. Für die Untersuchung wurde
die TfCa in 25 mM Phosphatpuffer pH 8 mit dem Inhibitor für 15 min bei 30°C inkubiert und die
verbleibende Esteraseaktivität mit p-NPB als Substrat spektrophotometrisch gemessen (Tabelle 27).
Die Inhibitoren EDTA und DTT zeigten mit verschiedenen Konzentrationen keine Hemmung der
Esteraseaktivität. Hingegen zeigte sich mit den spezifischen Esterasehemmern PMSF und TPCK eine
deutliche Erniedrigung der Aktivität ab einer Konzentration des Inhibitors von 1 mM. Mit 10 mM
Inhibitor war die verbleibende Esteraseaktivität mit PMSF 35% und mit TPCK konnte keine
verbleibende Aktivität bestimmt werden.
Tabelle 27: Übersicht über die verbleibenden Esteraseaktivitäten nach der Inkubation der TfCa mit verschiedenen Inhibitoren.
Inhibitor Konzentration (mM) Aktivität (%)
Ethylendiamintetraacetat (EDTA) 1 100
10 100
Phenylmethylsulfonylfluorid (PMSF) 0,1 100
1 69
10 35
Tosyl-L-Phenylalanin-Chloromethylketon (TPCK) 0,1 100
1 19
10 0
Dithiothreitol (DTT) 0,1 100
1 100
10 100

Ergebnisse und Diskussion 118
3.2.6 Kinetik der Hydrolyse von verschiedenen Esterasesubstraten durch die TfCa
Für die Bestimmung der Substratspezifität der TfCa wurden verschiedene Modellsubstrate und die
zyklischen PET-Trimere (CTR) eingesetzt, um die kinetischen Konstanten für den Umsatz der
Substrate zu bestimmen. Bei den Modelsubstraten handelte es sich um die p-Nitrophenylester der
Essigsäure, der Buttersäure, der Caprylsäure und der Palmitinsäure. Dafür wurde die TfCa in Puffer
mit den Substraten in unterschiedlichen Konzentrationen gegeben und die Umsatzgeschwindigkeit
anhand des entstandenen Produktes, p-Nitrophenol, bestimmt. Mit den Konzentrationen und den
Umsatzgeschwindigkeiten konnten dann die kinetischen Konstanten Km und Vmax für die einzelnen
Substrate per Lineweaver-Burk Darstellung berechnet werden.
Diese Untersuchung wurde abschließend auch für die zyklischen Trimere des PET durchgeführt. Dazu
wurde eine Stammlösung der Trimere in 1,4-Dioxan hergestellt. Für die Bestimmung der kinetischen
Parameter wurden die zyklischen Trimere in verschiedenen Konzentrationen zu einer Pufferlösung
mit der TfCa gegeben und bei 50°C für eine Stunde inkubiert. Anschließend wurden die Proben
zentrifugiert und per Umkehrphasen-HPLC mit 0,1% TFA (Puffer A) und Acetonitril (Puffer B)
analysiert. Die entstandenen Abbauprodukte wurden bei 241 nm mit dem UV Detektor detektiert.
Die kinetischen Konstanten Km und Vmax für die CTR wurden per Lineweaver-Burk Darstellung
berechnet (Abbildung 51).
Tabelle 28: Kinetische Parameter der Hydrolyse von p-NP-Estern und PET-Trimeren durch die TfCa aus T. fusca KW3.
Substrat Acyl-Rest Km (mM) Vmax (µmol/min/mg)
p-Nitrophenylester p-Nitrophenylacetat 2:0 0,27 ± 4% 46,1 ± 1%
p-Nitrophenylbutyrat 4:0 0,16 ± 4% 88,1 ± 1%
p-Nitrophenylcaprylat 8:0 0,20 ± 4% 64,0 ± 1%
p-Nitrophenylpalmitat 16:0 k. A. k. A.
PET Trimere 0,47 ± 9% 9,3 ± 6%
k. A.: keine Aktivität

Ergebnisse und Diskussion 119
Abbildung 51: Lineweaver-Burk Darstellung für die enzymkatalysierte Hydrolyse der CTR durch die TfCa. Mit Hilfe der doppeltreziproken graphischen Darstellung konnten die Geschwindigkeitskonstanten Vmax und Km bestimmt werden.
3.2.7 Optimale Temperatur und optimaler pH-Wert der CTR-Hydrolyse durch die TfCa
Für die Anwendung der Esterase zum Abbau von zyklischen PET-Trimeren wurde die optimale
Temperatur und der optimale pH-Wert der CTR Hydrolse bestimmt. Dazu erfolgte der Ansatz
mehrerer Reaktionen mit verschiedenen pH-Werten des Puffers und die Inkubation bei
verschiedenen Temperaturen, um deren Einfluss auf die Abbaurate zu bestimmen.
Die TfCa besitzt ein Temperaturoptimum für den Abbau von zyklischen PET-Trimeren bei 60°C und
ein pH-Optimum bei pH 6 (Abbildung 52).
Abbildung 52: pH und Temperaturoptimum der TfCa beim Abbau von zyklischen PET-Trimeren, bestimmt per RP HPLC (Billig et al. 2010).

Ergebnisse und Diskussion 120
3.2.8 Cutinaseaktivität der TfCa
Um zu testen, ob die aufgereinigte TfCa auch in der Lage ist, Cutin abzubauen, wurden je 1 U, 2 U
und 4 U der TfCa mit Cutin inkubiert. Die nachfolgenden Chromatogramme zeigen die
gaschromatographischen Analysen der verschiedenen Ansätze (Abbildung 53-56).
Abbildung 53: Gaschromatographische Analyse des Blindwerts (ohne Enzym) der Cutinhydrolyse.
Abbildung 54: Gaschromatographische Analyse des Ansatzes mit 1 U TfCa der Cutinhydrolyse.
0 2 4 6 8 10 12 14 16 18 20 22
Analysezeit (min)
0
10
20
30
40
50
60
70
80
90
100
Rela
tive A
bundan
z
9.73
2.81
8.19
5.27
19.84 12.25 14.08 6.37 16.05 17.63 22.93
0 2 4 6 8 10 12 14 16 18 20 22
Analysezeit (min)
0
10
20
30
40
50
60
70
80
90
100 9.74
7.19 2.76
8.19
5.29 12.86 20.08 16.05 18.34 23.11 21.71
Rela
tive A
bundan
z

Ergebnisse und Diskussion 121
Abbildung 55: Gaschromatographische Analyse des Ansatzes mit 2 U TfCa der Cutinhydrolyse.
Abbildung 56: Gaschromatographische Analyse des Ansatzes mit 4 U TfCa der Cutinhydrolyse.
Die Analysechromatogramme der enzymatischen Cutinhydrolyse zeigen, dass das partiell
aufgereinigte Enzym nicht in der Lage war, das Biopolymer zu hydrolysieren. Damit gehört dieses
Enzym nicht zur Gruppe der Cutinasen.
3.2.9 PCL-Abbau durch die TfCa
Nach 24 Stunden zeigte die Platte mit der TfCa keine Hofbildung, die Platten mit den Enzymlösungen
aus dem Suberin- und Cutin MSV-Medium jedoch eine deutliche Hofbildung (Abbildung 57). Das
bedeutet, dass die TfCa keine Cutinase sondern eine Carboxylesterase ist, da sie weder Cutin noch
0 2 4 6 8 10 12 14 16 18 20 22
Analysezeit (min)
0
10
20
30
40
50
60
70
80
90
100
Rela
tive A
bundan
z
2.75
9.75
14.45
7.20 19.31
5.29 12.25 16.06 23.09 20.64
0 2 4 6 8 10 12 14 16 18 20 22
Analysezeit (min)
0
10
20
30
40
50
60
70
80
90
100
Rela
tive A
bundan
z
9.75
7.20
2.78
8.20
3.67 5.32 12.25
19.81 13.97 16.06 22.67

Ergebnisse und Diskussion 122
PCL hydrolysierte. Die mit den Cutin- und Suberinpräparationen inkubierten PCL-Platten zeigten eine
deutliche Hofbildung, was bedeutet, dass diese Enzyme wahrscheinlich Cutinasen sind.
Abbildung 57: PCL-Agarplatten nach 24 stündiger Inkubation mit TfCa (linkes Photo), mit dem Kulturüberstand des Suberin-MSV-Mediums (mittleres Photo) und mit Kulturüberstand des Cutin-MSV-Mediums (rechtes Photo).
3.2.10 N-terminale Sequenz der TfCa
Die eindeutige Identifizierung der Esterase erfolgte über die Bestimmung der Protein und DNA
Sequenz. Hilfreich dabei war die Publikation von Lykidis et al. (2007), welche die komplett
identifizierte genomische Sequenz von T. fusca XY enthält.
Es wurden die ersten 10 Aminosäure-Reste der TfCa ansequenziert. In Tabelle 29 ist die detektierte
N-terminale Sequenz dargestellt. An Position 3 und 5 können entweder Lysin oder Isoleucin stehen.
Tabelle 29: Identifizierte N-terminale Sequenz der TfCa aus T. fusca KW3.
Aminosäure-Rest 1 2 3 4 5 6 7 8 9 10
Aminosäure M E I/K V I/K R T G S G
Das zu dieser N-terminalen Sequenz mittels Datenbank-Recherche ermittelte identische Protein ist
eine Carboxylesterase (EC 3.1.1.-) mit der Bezeichnung Tfu_2427. Es besitzt eine berechnete Größe
von 52,8 kDa und eine für Hydrolasen typische katalytische Triade aus den Aminosäuren Serin,
Glutamat und Histidin. Ebenfalls ist die Region des Serins hochkonserviert und besitzt das
Sequenzmotiv GESAG. Die Lokalisation des Gens im Genom und die Sequenz einschließlich der
katalytischen Triade sind in den Abbildungen 58 und 59 dargestellt.
Abbildung 58: Lokalisation des Tfu_2427 Gens im Genom von T. fusca YX (Quelle: http://www.ncbi.nlm.nih.gov/).

Ergebnisse und Diskussion 123
1 meivirtgsg dvrgskengi avfrgipyae ppvgahrfta prpprpwdgv rdatefsata
61 prppypeaig allierfipg ddyltlnvwt pdpnavglpv mvwihggaft ngsgsepvyd
121 gaafardgvv fvsfnyrlgi igfadlpdap snrglldqia alewvrdnia rfggdpgnvt
181 vfgesagams vctlmatpra rglfrrailq sgagnmavaa edattiaavi ahrlgvepta
241 aalahvpvaq lldvqqqvaq eiqgapdpav wgeriaggsv llpfapvidg ellsqrpaea
301 iaggaghdvd llfgtttdey rlflaptgll pfitgdyvtt hlaksgldad aakaytaegr
361 geepgdilas iitdqvfrip alriaesrvd apartfgyef awrtpqldgi lgachavelp
421 fvfrtldraa slvgtnppee laetvhnawv rfatsgdpgw pawnpetrsv mrfdhpvsem
481 vtdpypatra lwdgvpl
Abbildung 59: Proteinsequenz von Tfu_2427 aus T. fusca YX, die Aminosäuren des aktiven Zentrums und das GXSXG Motiv sind gelb markiert. Die ersten 10 Aminosäuren am N-Terminus, anhand derer die Identifikation erfolgte, sind unterstrichen (Billig et al. 2010).
3.2.11 MALDI-TOF Sequenzierung der TfCa
Die durch die N-terminale Sequenzierung erhaltene Sequenz stammt von dem Actinomyceten
T. fusca YX, einem sehr nahen Verwandten des T. fusca KW3. Da es aber auch bei Organismen der
gleichen Spezies kleine Unterschiede in einer Proteinsequenz geben kann, wurde mit der isolierten
TfCa eine MALDI-TOF Analyse durchgeführt, um die genaue Proteinsequenz zu bestimmen.
Das Ergebnis war eine 93,5% Übereinstimmung mit der bereits bekannten Sequenz und dem
aufgereinigten Protein. Bis auf zwei Bruchstücke konnte die gesamte Enzymsequenz der TfCa
identifiziert werden. Das bedeutet, dass es sich bei dem aufgereinigten Protein definitiv um eine
Carboxylesterase handelt, welche mindestens zu 93,5% mit der Sequenz des Proteins Tfu_2427 aus
T. fusca YX übereinstimmt.
Für eine vollständige Sequenzidentifikation erfolgte parallel eine Sequenzierung des TfCa Gens
mittels abgeleiteter Primer (Oeser et al. 2010). Bei dieser Sequenzierung konnte die gesamte
Sequenz des TfCa Gens identifiziert werden und somit die gesamte Proteinsequenz abgeleitet
werden. Beim Vergleich der Sequenz der TfCa mit der Tfu_2427 wurde festgestellt, dass die Proteine
bis auf 2 Aminosäuren übereinstimmen. Diese beiden unterschiedlichen Aminosäuren wurden zuvor
mit der MALDI TOF/TOF Analyse nicht identifiziert, da beide Unterschiede sich im gleichen
Proteinfragment befanden und sich somit die Differenzen der Massen ausglichen. An Position 335 bei
der TfCa steht anstelle eines Glycins ein Serin und an Position 340 steht ein Alanin anstelle eines
Threonins (Abbildung 60). Somit ergibt sich eine 99,6% Übereinstimmung zwischen den beiden
Carboxylesterasen.

Ergebnisse und Diskussion 124
1 meivirtgsg dvrgskengi avfrgipyae ppvgahrfta prpprpwdgv rdatefsata
61 prppypeaig allierfipg ddyltlnvwt pdpnavglpv mvwihggaft ngsgsepvyd
121 gaafardgvv fvsfnyrlgi igfadlpdap snrglldqia alewvrdnia rfggdpgnvt
181 vfgesagams vctlmatpra rglfrrailq sgagnmavaa edattiaavi ahrlgvepta
241 aalahvpvaq lldvqqqvaq eiqgapdpav wgeriaggsv llpfapvidg ellsqrpaea
301 iaggaghdvd llfgtttdey rlflaptgll pfitsdyvta hlaksgldad aakaytaegr
361 geepgdilas iitdqvfrip alriaesrvd apartfgyef awrtpqldgi lgachavelp
421 fvfrtldraa slvgtnppee laetvhnawv rfatsgdpgw pawnpetrsv mrfdhpvsem
481 vtdpypatra lwdgvpl
Abbildung 60: Proteinsequenz der TfCa aus T. fusca KW3, die Aminosäuren des aktiven Zentrums und das GXSXG Motiv sind gelb markiert, die identifizierten Unterschiede zu Tfu_2427 aus T. fusca YX sind rot markiert (Billig et al. 2010).
Mit der Sequenz der Carboxylesterase TfCa wurde ein Alignment mit den vier ähnlichsten Enzymen
aus einer Enzymdatenbank (Kyoto Encyclopedia of Genes and Genomes (KEGG) protein database;
http://www.genome.jp/kegg/) erstellt. Es sind alles ebenfalls Hydrolasen mit ähnlichen Sequenzen.
Die Enzyme stammen aus Arthrobacter FB24 (art), Streptomyces coelicolor (sco), Frankia alni (fal)
und Saccharopolyspora erythraea (sen). Es ist ersichtlich, dass besonders um die Bereiche der
Aminosäuren der katalytischen Triade die Enzyme viele Übereinstimmungen besitzen. In der
nachfolgenden Abbildung ist das Alignment der 5 Hydrolasen dargestellt. Die grau unterlegten
Sequenzen treten bei mindestens drei der fünf Enzyme auf. Die Aminosäuren der katalytischen
Triade sind fett gedruckt.

Ergebnisse und Diskussion 125
1 M----------------------------------------------------------- tfu
1 M----------------------------------------------------------- art
1 MRS---------GQT--------------------------------------------- sco
1 MRPAASPVLSRLGQALAALVCLDAPVSFNPWHALVAVLWWSRRQGTVNGWRAFPVLRDDS fal
1 M----------------------------------------------------------- sen
2 -----------------------------------EIVIRTGSGDVRGSKENGIAVFRGI tfu
2 -----------------------------------DTVVKTTHGQLRGSLADGVHCFLGI art
7 -----------------------------------NPVVRTVHGAVGGRYEHDVAVFRGI sco
61 AVRVRVRDRDRGRVRSVRVGGAMTTVGAVPASAGSRPGARVAGGALCGSRESGVAVFRGI fal
2 -----------------------------------DVIVRTGSGRVRGFSAGDVSVFKGI sen
27 PYAEPPVGAHRFTAPRPPRPWDGVRDATEFSATAPRPPYPEAIGALLIERFIPGD--DYL tfu
27 PFAASTAGVNRLRPPQPVQPWSGVRDAMKYGDSPFQLAPPESAGTEW-DTALAGE--DCL art
32 PYAAPPFGPGRFRPPLPPEPWDGVRDAGSFGPTAPKPPYSEAFARYLSDPAIPGD--DCL sco
121 PYAAPPVGGLRFAAPRPAASWDGVRSALSYGPPPPQ---SDVLGRGPSAEGTPGD--DWL fal
27 PYAAPLDGPARFQAPLPAPSWEGVRDSTAFSASVPQ---EAAPMPGQVSPWKPGDSTECL sen
85 TLNVWTPDPNAVG-LPVMVWIHGGAFTNGSGSEPVYDGAAFARD-GVVFVSFNYRLGIIG tfu
84 NLNVWTPDPGNGG-LPVMVWIQGGAFEIGSTAS--YNGRTFARD-GVICVVINWRVGADG art
90 NLNVWTPEPGPGARLPVLVWLHGGALTRGSSAVPVYDGSTFARD-GVVCVSVNYRLGVEG sco
176 TVNVWSPEPDPAAKLPVMVWIYGGAYAIGASSRPEYDGAHLARDGGVVVVTFDYRLGMEG fal
84 TVNVWSPDVGARG-LPVLVWIHGGMFMHGSSASPAYDGAALARE-GVVFVSLNYRVGYEG sen
143 FADLPDAPSNRGLLDQIAALEWVRDNIARFGGDPGNVTVFGESAGAMSVCTLMATPRARG tfu
140 FLDLGDGQANIGLLDQVAALEWVRGNIAAFGEDPANVTVFGQSAGAMSIGVLLSMPRAEG art
149 YGLFPDAPANAGLRDQIAALQWVRDNIAAFGGDPDRVTVAGQSAGAISTGALLAAPRARG sco
236 FAQITGAPPNRGLLDQVAALEWVRDNIAAFGGDPDEVTVFGESAGGGSVAALLAMPRAAG fal
142 FGWVADAPANRGLLDQIAALRWVRDNIAGFGGDPGNVTVAGQSAGGSSVVALVSSPAARG sen
203 LFRRAILQSGAGNMAVAAEDATTIAAVIAHRLGVEPTAAALAHVPVAQLLDVQQQVAQEI tfu
200 LFRRAILQSGAAHHVLPTETAQRIGGFLSEKLGVSPTREAMAAVPVQRFLAAQTELKADL art
209 LFRRAVLQSGAPE-AADRDKVRRMVRRMASRLKIPATAEAFAAADRDQLLRAQAEV---- sco
296 LFRRAIAQSVPGT-FLSPQLAADIAADCADELGLRPTVADLATVDPALLTVAADTVATDT fal
202 LLRRGIAQSVGGL-FVPEAEARAVAEMVTDQLGVSPTAAELGSLPSEAIHGAQSAPSAAM sen
263 QGA---PDPAVWGERIAGGSVLLPFAPVIDGELLSQRPAEAIAGG-AGHDVDLLFGTTTD tfu
260 AAR---PDPARWGPEVVASSML--WQPSVDGDVIPRRPIERIAAG-AGSTVDVMIGSNAE art
264 ---------GRFSSPVLGGPG---FGLVVDGDVLPRDPLEALVDGDAAPGVDLMMGWTRD sco
355 VAATMATRARRWGQAALRSIL---FAPVVDGEVLPATPWQALAAG-AGRDVDLLAGHTRD fal
261 TA-----DPSAWTNSHTP------YAPMFGDDVLERRPWEALRAG-AGREIDLIAGFTRD sen
319 EYRLFLAPTGLLPFITS--------DYVTAHLAKSGLDAD-AAKAYTAEGRGEEPGDILA tfu
314 DWKLFLAITGVIAKVTERDLAESRSVDGFPPIGVYGLPAETALREYRSHYPASSPGELLA art
312 EYRLWLVPGGLVERVDR-----LGAVALAGARARCHCGNE-VVRGYRAARPGAGTAEIVG sco
411 EQRLFTALTGLLGQVTP--------DQAATALQDFAPGRD-GARRYRDAYPDAGPEQLYE fal
309 ECRVFAAGLDLSG------------SDPVAVARGMRLAPE-ALPRYRAAHPGISDAGLHE sen
370 SIITDQVFRIPALRIAESRVD----APARTFGYEFAWRTPQLDGILGACHAVELPFVFRT tfu
374 AVETDWWVRIPALRLADAHAKALSTASARTYMYEFAWPAPGL----GAVHAMEVPFVFDT art
366 QLVTDHLLRIPMHRVADAR-------PGSSHVYEFAWPSNLPD--LGACHALELGFVFDS sco
462 LVHSDWLFRMPTLHLAQAQVA----GGGRVHLYELTWPAPGMGGALGACHGLDVPLVFGN fal
356 LMLTDSLFRMPTLWCAEGHAE----AGGKTYLYEFAWPAPVLDGAYGACHSIDVPFLFGV sen
426 LDRAAS----LVGTNPP---EELAETVHNAWVRFATSGDPGWPAWNPETRSVMRFDHPVS tfu
430 LDTSSRLFGPLLGTDPP---QELADAMHAAWISFASTGDPGWPGYDLERRATMLFNTG-A art
417 GDGPDA--RRLAGEGAP---QELADAMHGAWVRFAETGDPGWQAWDAA-HPVRVFGDGAP sco
518 LTSGQP--ALLIGDSPTPEATALSARMRSAWTTFATRSDPGWPAYDCDHRLTQIFDTS-P fal
412 LDDEIG--RPLLGDPAPAGFEVLSAQLRRAWTSFAASGDPGWPEFGTDWRLARSWNVP-L sen
479 EMVTDPYPATRALWD-GVPL------------ tfu
486 GVVYDPRSWERDLWE-GVR------------- art
471 HTAYGPLDPEYDLWKADVTVPRGTRPRRLVRR sco
575 VVTAYPEERSRLIWQ-DHSFP----PLLLHAP fal
469 EVVSDPAGESRRIWR-EHA------------- sen
Abbildung 61: Alignment der Carboxylesterase TfCa aus T. fusca KW3 (tfu), der Carboxylesterase Arth_3983 aus Arthrobacter FB24 (art), der Carboxylesterase SCO6127 aus Streptomyces coelicolor (sco), der putativen Carboxylesterase FRAAL0556 aus Frankia alni (fal) und der putativen p-Nitrobenzylesterase SACE_2933 aus Saccharopolyspora erythraea (sen) (Billig et al. 2010).

Ergebnisse und Diskussion 126
3.2.12 Homologie-Modeling der TfCa und Vergleich der Struktur mit anderen Hydrolasen
Mit der Proteinsequenz der TfCa wurde mit dem Programm PyMOL eine Modellierung der
Proteinstruktur, unter der Nutzung der thermostabilen Carboxylesterase aus
Geobacillus stearothermophilus (PDB Code 2OGS resp. 2OGT) als Template, vorgenommen
(Abbildung 62, 63) (Liu et al. 2007, Bennett-Lovsey et al 2008). Das aktive Zentrum der
Carboxylesterase mit der katalytischen Triade wurde blau gekennzeichnet.
Abbildung 62: Tertiärstruktur der Carboxylesterase TfCa aus T. fusca KW3. Rot sind die α-Helices gekennzeichnet und gelb die β-Faltblattstrukturen. Die grünen Bereiche sind Sequenzen ohne eine spezielle Sekundärstruktur. Die drei blau gekennzeichneten Aminosäuren sind die katalytische Triade (Billig et al. 2010).
Abbildung 63: Ausschnitt aus der Tertiärstruktur der Carboxylesterase TfCa aus T. fusca KW3. Rot sind die α-Helices gekennzeichnet und gelb die β-Faltblattstrukturen. Die grünen Bereiche sind Sequenzen ohne eine spezielle Sekundärstruktur. Die drei blau gekennzeichneten Aminosäuren sind die katalytische Triade.
Zum Vergleich wurden weitere PET-Hydrolasen modelliert. Die modellierte Struktur der Cutinasen
Tfu_0882 aus T. fusca YX ,basierend auf der Lipase von S. exfoliates (SeL), wurde mit dem SWISS-
Model Homologie Modeling Web Server konstruiert (Abbildung 64d) und Tfu_0883, basierend auf
der Lipase von S. exfoliates (SeL, PDB-ID: 1JFR), wurde mit PHYRE erstellt (Abbildung 64e) (Lykidis
et al. 2007, Chen S et al 2008). Von den Sequenzen der Cutinasen von F. solani sp. pisi und
P. mendocina aus der NCBI Protein Data Base und der Sequenz der Cutinase von H. insolens,
modelliert mit dem Wurst Protein Threading Server, wurde mittels PyMOL ein Strukturbild erstellt
(Abbildung 64c, f, b). Die katalytischen Aminosäure-Reste sind blau dargestellt.

Ergebnisse und Diskussion 127
Abbildung 64: Homologie-Modelle der Carboxylesterase TfCa aus T. fusca KW3 (a) verglichen mit verschiedenen PET-Cutinasen. Die Strukturen der Cutinasen aus H. insolens (b), F. solani sp. pisi (c), der Cutinasen Tfu_0882 (d) und Tfu_0883 (e) aus T. fusca KW3 und der Cutinase aus P. mendocina (f) sind dargestellt (Billig et al. 2010).
c

Ergebnisse und Diskussion 128
3.2.13 Vergleich der TfCa mit bekannten PET-Hydrolasen
Die TfCa unterscheidet sich deutlich von allen anderen bisher bekannten und bereits näher
charakterisierten PET-Hydrolasen nicht nur durch Ihre höhere Größe, sondern auch durch die
Tatsache, dass es sich um eine Carboxylesterase und nicht um eine Cutinase handelt. In der
nachstehenden Tabelle sind die Eigenschaften der TfCa aus T. fusca KW3 verglichen mit den
Cutinasen aus T. fusca DSM 43793 und einem weiteren T. fusca Isolat dargestellt (Kleeberg et al.
2005, Chen S et al. 2008). Diese beiden Cutinasen unterscheiden sich hinsichtlich ihrer Produktion,
die Erstere wurde als Wildtypenzym durch T. fusca DSM 43793 produziert, die Zweite hingegen
rekombinant in E. coli.
Tabelle 30: Vergleich der Eigenschaften der TfCa mit Cutinasen aus T. fusca.
TfCa aus T. fusca KW3 Cutinase aus T. fusca DSM 43793
(Kleeberg et al. 2005)
Cutinase aus einem T. fusca Isolat
(Chen S et al. 2008)
Molekulargewicht
(ermittelt per Gelelektrophorese)
52,4 kDa 27,4 kDa 29 kDa
pI 4,8 6,43 n. b.
pH Stabilität pH 5–11 n. b. n. b.
Temperaturstabilität 30–50°C n. b. 40–60°C
pH Optimum pH 6 (CTR Hydrolyse) pH 6,5–7 (BTA Hydrolyse) pH 8 (Triolein, p-NPB Hydrolyse)
Temperatur Optimum 60°C (CTR Hydrolyse) 65–70°C (BTA Hydrolyse) 60°C (Triolein, p-NPB Hydrolyse)
Inhibitoren PMSF TPCK
n. b. PMSF
Hydrolysierte Substrate p-Nitrophenylacetat p-Nitrophenylbutyrat p-Nitrophenylcaprylat CTR
Triglyceride Aliphatische Polyester Aliphatisch-aromatische Copolyester PET-Filme Cutin
p-Nitrophenylbutyrat Triolein Cutin
Aminosäuren der katalytischen Triade
Ser185, Glu319, His415 G-E-S-A-G Motiv
Ser132, Asp176, His210 G-H-S-M-G Motiv
Ser132, Asp176, His210 G-H-S-M-G Motiv
n. b.: nicht bestimmt
Die optimale Temperatur für die CTR Hydrolyse durch die TfCa lag bei 60°C und der optimale pH Wert
bei 60°C war pH 6. Nach 24 Stunden Inkubation lagen die Restaktivitäten bei pH 5 bis 11 bei 40-100%
und somit stabil in einem breiten pH Bereich. Bei einem Inkubationszeitraum von 7 Tagen bei pH 9

Ergebnisse und Diskussion 129
und 10 lag die Restaktivität bei 85%, bei den pH Werten 5 und 11 lag sie bei 22% und 16%. Bei
erhöhten Temperaturen (40-50°C) war nach 7 Tagen Inkubation noch 93-100% Restaktivität
nachweisbar. Mit 55°C konnte nach 24 Stunden Inkubation nur noch 1% Restaktivität detektiert
werden. Bei der Ermittlung der Substratspezifität mit p-NPEstern zeigte sich, dass die TfCa Aktivität
gegenüber kurz- und mittelkettigen Substraten (C2 bis C8) besaß, hingegen nicht gegenüber
langkettigen (C16) Substraten. Die Inhibitoren PMSF und TPCK reduzierten die hydrolytische Aktivität
der TfCa, was bedeutet, dass ein Serin und ein Histidin Bestandteil der katalytischen Triade sind.
Durch DTT und EDTA erfolgte keine Inhibition der Esterase, somit sind keine essentiellen
Disulfidbrücken im Enzym enthalten und es benötigt auch keine Metallionen als Cofaktoren. Somit
handelt es sich bei dem aufgereinigten Enzym um eine Serin Hydrolase, welche allerdings kein Cutin
oder PCL hydrolysieren kann und somit nicht zur Gruppe der Cutinasen zählt, wie die meisten PET-
Hydrolasen. Bei der Sequenzbestimmung der Esterase stellte sich heraus, dass es sich um eine
Carboxylesterase mit 42% Übereinstimmung mit der Carboxylesterase Arth_3983 aus
Arthrobacter FB24, 42% Übereinstimmung mit der Carboxylesterase SCO6127 aus S. coelicolor, 43%
Übereinstimmung mit der putativen Carboxylesterase FRAAL0556 aus F. alni und 42%
Übereinstimmung mit der putativen p-Nitrobenzylesterase SACE_2933 aus S. erythraea handelt. Das
katalytische Zentrum der TfCa besteht aus Ser185, Glu319, His415, wobei das Serin in das für
Hydrolasen typische G-E-S-A-G Motiv eingebettet ist.
Durch eine Codon Optimierung, der Klonierung in den pET-20b(+) Vektor mit einer N-terminalen pelB
Signalsequenz und einem C-terminalen His6-Tag und einer anschließenden Transformation in E. coli
BL21(DE3) erfolgte die Erstellung eines rekombinanten Wirtes für die Überexpression der
Carboxylesterase TfCa aus T. fusca KW3 (Oeser et al. 2010). Die rekombinant produzierte TfCa war in
der Lage PET-Nanopartikel mit einer Rate von 7 nmol/ml freigesetzte TPA bei 24 Stunden Inkubation
bei 50°C und 0,01 mg TfCa zu hydrolysieren.
Bei der TfH Cutinase aus T. fusca DSM 43793 lag das Temperaturoptimum 10-15°C über der
maximalen Wachstumstemperatur des Mikroorganismus, was untypisch für extrazelluläre Enzyme
ist, hingegen für Lipasen, Xylanasen oder PHB-Depolymerasen schon beschrieben wurde.
Anscheinend gibt es laut Kleeberg et al. (2005) einen Überlagerungseffekt bezüglich der
Enzymaktivität und dem zu hydrolysierenden Substrat. Mit höher molekularen Substraten, wie BTA,
welches erst bei höheren Temperaturen aufgrund der höheren Beweglichkeit der Polymerketten
besser abbaubar ist, konnte ein höheres Temperaturmaximum bestimmt werden als mit
niedermolekularen Substraten, wie Ölen oder Fetten. Der schnelle Verlust an Aktivität bei 70°C ist
auf die Denaturierung der Hydrolase zurückzuführen. Das aufgereinigte Enzym verlor nach 30 min bei
70°C 85% seiner Aktivität, wohingegen die Hydrolase in der Enzymrohlösung 60% Restaktivität nach

Ergebnisse und Diskussion 130
einer Woche Lagerung bei Raumtemperatur aufwies und bei 70°C nach einer Stunde fast die gesamte
hydrolytische Aktivität verloren ging. Das pH Maximum lag bei pH 6-6,5, verlagerte sich aber bei
geringeren Ionenstärken in Richtung der neutralen pH Werte. Die BTA Hydrolase spaltete Triacetin,
Tributyrin und Tricaproin mit einer ähnlichen Aktivität, hingegen mit den längerkettigen Substraten
Tricaprylin, Tricaprin, Trilaurin und Triolein sank die hydrolytische Aktivität mit zunehmender
Kettenlänge. Des Weiteren konnte die TfH auch eine Vielzahl an Polyestern (PCL,
Poly(trimethylenbrassylat) (SP3/13), Bionolle, Polyesteramid Bayer TIR 1874) hydrolysieren, jedoch
nicht das natürliche Polymer Poly(3-hydroxybutyrat) (PHB).
Das Enzym besteht aus 261 Aminosäuren und wurde als Serin Hydrolase klassifiziert, welche
Esterasen, Lipasen und Cutinasen beinhaltet. Sie besaß eine 65% Ähnlichkeit zu einer Triacylglycerin
Lipase aus Streptomyces albus G26 und eine 62% Ähnlichkeit zu einer Triacylglycerin Acylhydrolase
aus Streptomyces sp. M1127. Das Serin des katalytischen Zentrums war in dem hochkonservierten
Motiv G-H-S-M-G, an der Position 132 im Enzym, eingebettet. Der Anteil an hydrophoben
Aminosäuren (Ala, Val, Leu, Ile, Pro, Met, Phe, Trp) war mit 42% hoch im Vergleich zu anderen Serin
Hydrolasen, worauf die Probleme bei der Aufreinigung zurück zu führen waren.
Obwohl die TfH aufgrund der Sequenzübereinstimmungen und der Substratspezifität zu den Lipasen
eingeordnet werden könnten, zeigten sie aber auch deutliche Unterschiede im Vergleich zur
Pseudomonas sp. Lipase (PsL) und Aspergillus oryzae Lipase (AoL) bei der Film und Nanopartikel
Hydrolyse. Die Lipasen hydrolysierten nur 40% der Esterbindungen, da sie nur die Oberfläche des
unlöslichen Substrates angreifen. Auch bei weiterer Enzymzugabe gab es keine weitere Hydrolyse,
was bedeutete, dass das Enzym nicht inaktiviert war. Die Hydrolyse stoppte, als das Substrat
komplett in wasserlösliche Zwischenprodukte umgesetzt war und somit die Lipasen keine
hydrophoben Oberflächen für ihre Konformationsänderung zur Verfügung hatten. Somit wurden die
verbleibenden Esterbindungen der Intermediate nicht hydrolysiert. Die TfH hingegen hydrolysierte
100% der Esterbindungen des Polymerfilms und der Nanopartikeln und zeigte somit keine
Grenzflächenaktivierung. Aufgrund der hydrolytischen Aktivität der TfH gegenüber Apfelcutin und
PCL konnte sie zur Gruppe der Cutinasen gezählt werden (Gouda et al. 2002, Kleeberg et al. 2005).
Die TfH wurde durch Dresler et al. 2006 rekombinant in E. coli TG1 mit dem Plasmid pCYTEXP1-
OmpA-bta1-His6 produziert, sekretiert und aufgrereinigt. Dabei erfolgte allerdings keine weitere
Charakterisierung der Cutinase (Dresler et al. 2006).
Nach einer Codon Optimierung wurde das tfh Gen in einen offenen Leserahmen mit der DNA
Sequenz für das N-terminale Signalpeptid der Bacillus megaterium Lipase A und einem C-terminalen
His6-Tag kloniert und unter einem Xylose induzierbaren Promotor rekombinant in
Bacillus megaterium WH323 produziert. Auch hier erfolgte keine weitere Charakterisierung der
rekombinanten Hydrolase (Yang et al. 2007).

Ergebnisse und Diskussion 131
Die BTA-Hydrolase aus Thermobifida sp. BCC23166, isoliert aus Proben einer Deponie, wurde durch
Transkonjugation in den Produktionsstamm Streptomyces rimosus R7 mit pIJ-BTA verfügbar
(Sinsereekul et al. 2010). Diese aufgereinigte Hydrolase zeigte einen optimalen Temperaturbereich
von 50-55°C und einen optimalen pH im milden bis basischen Bereich, bestimmt mittels p-NPP als
Substrat. Die rTfH besaß die höchste Aktivität mit kurzkettigen p-Nitrophenylestern, aber auch mit
lankettigen Substraten konnten hydrolytische Aktivitäten nachgewiesen werden, was mit den bereits
referierten Aktivitäten übereinstimmt (Kleeberg et al. 2005). Unter Anwendung der
Oberflächenplasmonresonanztechnik zur Untersuchung der Hydrolyse von Polyestern zeigte sich,
dass die rTfH eine unterschiedliche Bevorzugung von aliphatischen und aliphatisch-aromatischen
Polyestern besitzt und zwar in der Reihenfolge PCL>Ecoflex®>PHB. Diese Unterschiede kommen laut
Sinsereekul et al. (2010) aufgrund unterschiedlicher Bindungseigenschaften der Polyester an das
Enzym und aufgrund verschiedener physikalischer Eigenschaften (Kristallinität) zustande. Aufgrund
unterschiedlicher Filmherstellung und Analysetechniken war in diesem Fall auch ein Abbau von PHB
durch die rTfH nachweisbar. Im Vergleich zur Lipase aus T. lanuginosus zeigte die Hydrolase höhere
Spezifität und Abbauraten gegenüber aliphatisch-aromatischen Copolyestern, analog zu den Studien
von Eberl et al. (2008) (Sinsereekul et al. 2010).
Die 29 kDa Cutinase aus T. fusca WSH03-11 (Chen S et al. 2008, Chen J et al. 2008) zeigte deutliche
Übereinstimmungen mit 2 Enzymen, Tfu_0883 und Tfu_0882, identifiziert im T. fusca YX (Lykidis
et al. 2007). Die Enzyme wurden als Triacylglycerin Lipasen in der genomischen Datenbank
bezeichnet, enthalten eine hochkonservierte Sequenz (GXSXG) und gehören der α/β Hydrolase
Familie an. Für eine weitere Charakterisierung erfolgte die rekombinante Expression der beiden Gene
in E. coli BL21 Rossetta (DE3) PlysS mit dem Vektor pET-20b(+) mit einem C-terminalen His6-Tag und
dem PelB Signalpeptid. Die Cutinasen 1 und 2 hydrolysierten Triolein und p-NPB bei einer optimalen
Temperatur von 60°C und einem optimalen pH von 8. Bei den Thermostabilitätsuntersuchungen
zeigten die beiden Hydrolasen Restaktivitäten von 80% nach 160 Stunden bei 40°C und 50% nach 40
Stunden bei 60°C. Die beiden Isoenzyme waren auch in der Lage, Cutin zu hydrolysieren, womit sie
der Gruppe der Cutinasen zuzuordnen sind. Durch den Inhibitor PMSF, der an das aktive Serin der
katalytischen Triade bindet, erfolgte eine Verminderung der katalytischen Aktivität der Cutinasen.
Zusätzliche Mutationstudien bestätigten, dass die beiden Cutinasen Serin Hydrolasen sind, da beim
Austausch des Serins an der Posititon 188 (Cutinase 1) und 170 (Cutinase 2) keine katalytische
Aktivität mehr zu verzeichnen war (Chen S et al. 2008, Chen J et al. 2008).
Die molaren Massen der Isoenzyme, bestimmt mit der SDS-PAGE, waren 29 kDa, die berechneten
Werte lagen bei 29,220 kDa (Cutinase 1) und 28,997 kDa (Cutinase 2). Das native Molekulargewicht
lag bei 32 kDa, bestimmt per Größenausschlusschromatographie, und bestätigte, dass die Cutinasen
in Lösung monomere Enzyme sind. Bei der Studie der Substratspezifität der Cutinasen zeigten beide

Ergebnisse und Diskussion 132
eine höhere Aktivität mit kurzkettigen p-NPEstern, hydrolysieren aber auch langkettige Ester (C16-
C18). Dabei zeichnete sich Cutinase 2 durch eine höhere Hydrolyserate gegenüber Cutinase 1 aus, was
allerdings bei der Hydrolyse von Triolein nicht bestätigt wurde. Somit zeigten sich der 7%-
Sequenzunterschied auch in der hydrolytischen Aktivität der Isoenzyme. Diese Beobachtungen
bestätigten sich auch bei der Untersuchung der CTR-Hydrolyse, bei der die Cutinase 2 deutlich mehr
zyklische Trimere hydrolysierte. Das lässt die Annahme zu, dass die Sequenzunterschiede im Bereich
des aktiven Zentrums oder dessen Umgebung die katalytischen Aktivitäten beeinflussen. Beide
Cutinasen zeigten keine Grenzflächenaktivierung und besitzen keinen Deckel. Ebenfalls benötigen sie
keine Metallionen als Cofaktoren für ihre hydrolytische Aktivität, nachgewiesen mit dem Inhibitor
EDTA. Bei der Untersuchung des Einflusses verschiedener Lösungsvermittler wurde festgestellt, dass
Triton X-100, Tween 20 und TDOC (in Konzentrationen von 1 mM und 10 mM) keine
Aktivitätsänderungen zur Folge hatten, SDS hingegen inhibierte die Cutinasen. In den organischen
Lösungsmitteln Methanol, Ethanol, Aceton, n-Hexan und Dimethylsulfoxid (DMSO) sind die
Isoenzyme stabil, weniger hingegen in Isopropanol und Butanol, was eine breite Anwendung in der
organischen Synthese eröffnet (Chen et al. 2010, Chen J et al. 2008).
Bei der Suche nach Polymer-abbauenden Isolaten wurde eine thermostabile Hydrolase aus
Thermobifida alba AHK119 identifiziert, deren Gensequenz mit spezifischen Primern kompatibel zur
G-X-S-X-G Consensussequenz (903 bp) amplifiziert wurde (Hu et al. 2010). Die Aminosäuresequenz
bestehend aus 300 AS wurde Est119 benannt und gehört zur Familie der Serin Hydrolasen. Est119
besaß 84% Übereinstimmung mit der Triacylglycerin Lipase (TfH) aus T. fusca DSM 43793 (Kleeberg
et al. 2005), 63% Übereinstimmung mit der Lipase aus S. coelicolor A3(2) M145 (Valdez et al. 1999),
62% Übereinstimmung mit der Lipase aus S. albus G (Cruz et al. 1994) und 61% Übereinstimmung mit
der Lipase aus S. exfoliatus M11 (Wei et al. 1988). Die hoch konservierte Sequenz mit dem aktiven
Serin wurde mit G-H-S-M-G in der Hydrolase identifiziert, die katalytischen Aminosäuren sind Ser129,
His207 und Asp175. Eine N-terminale Signalsequenz von 34 Aminosäuren konnte ebenfalls
identifiziert werden, welche zwischen den Aminosäuren 34 und 35 posttranslational abgeschnitten
wird. Somit besaß das aktive verbleibende Enzym 266 Aminosäuren, eine Größe von 30 kDa
(rechnerisch 29,76 kDa) und lag in Suspension als monomeres Enzym vor. Die optimale Temperatur
für die Hydrolyse von p-NPB lag bei 50°C und der optimale pH Wert bei 6. Bei 50°C und pH 6 erfolgte
die Untersuchung der Substratspezifität der Hydrolase mit p-NP Estern. Mit der steigenden
Kettenlänge von C2-C6 stieg auch die Hydrolyserate an, fiel aber ab C8 stark ab. Aufgrund der
Sequenzübereinstimmungen der Est119 mit der Lipase aus S. exfoliatus M11 ordneten Hu et al.
(2010) sie ebenfalls in die Gruppe III der lipolytischen Enzyme ein.

Ergebnisse und Diskussion 133
Tabelle 31: Vergleich der PET-Hydrolasen aus F. solani FsC (Carvalho et al. 1998, Carvalho et al. 1999, Ronkvist et al. 2009), T. insolens HiC (Sandal et al. 1996, Ronkvist et al. 2009), P. mendocina PmC (Bott et al. 2003, Ronkvist et al. 2009), T. fusca
TfH/Cutinase 2 (Kleeberg et al. 2005, Chen S et al. 2008, Chen J et al. 2008) und TfCa (Zimmermann und Billig 2011).
FsC HiC PmC TfH/Cutinase 1 TfCa
Anzahl der
Aminosäuren
197 194 272 261 497
Molekulargewicht 22 kDa 20-21 kDa 30 kDa 28 kDa 52,4 kDa
Isoelektrischer
Punkt
7,2 8 n. b. 6,43 4,8
pH Optimum 6,5-8,5 8,5-9,5 9,0-9,5 6,5-8,0 6,0
Temperatur
Optimum
50°C 75-80°C 50°C 60-70°C 60°C
Katalytische Triade Ser120
Asp175
His188
Ser140
Asp195
His208
Ser126
Asp176
His206
Ser132
Asp176
His210
Ser185 Glu319 His415
GXSXG Motiv G-Y-S-Q-G G-Y-S-Q-G G-H-S-Q-G G-H-S-M-G G-E-S-A-G
Gruppierung Fungale
Cutinase
Fungale
Cutinase
Bakterielle
Cutinase
Bakterielle
Cutinase
Bakterielle Carboxyl-esterase
n. b.: nicht bestimmt
Die Cutinase aus P. mendocina hydrolysierte p-NPB am Besten, hingegen aber keine Triglyceride, was
bedeutete, dass das Enzym mit Cutinaseaktivität keine unspezifische Lipase ist. Die Cutinase zeigte
ein breites pH Optimum zwischen 8,0 und 10,5 mit Apfelcutin als Substrat. Di-
isopropylfluorophosphat (DFP, phosphoryliert Serin) inhibierte das Enzym, was bedeutet, dass ein
aktives Serin bei der katalytischen Hydrolyse beteiligt ist. Die Cutinase war stabil bei 60°C für 1
Stunde und behielt 85% ihrer Aktivität nach 1 Stunde bei 70°C (Sebastian et al. 1987).
Das Molekulargewicht des Proteinmonomers betrug 30 kDa bestimmt mit SDS-PAGE und
Größenausschlusschromatographie. Mit steigender Kettenlänge sank die hydrolytische Aktivität der
Cutinase gegenüber p-NP Estern. Das Enzym wurde nicht durch Thiol-nachweisende Agenzien
inhibiert, womit keine katalytisch aktive -SH Gruppe vorhanden ist, ebenso benötigt die Cutinase
keine Metallionen als Cofaktoren, nachgewiesen mit EDTA (Sebastian et al. 1988).
Nach der Identifikation des PmC-Gens und dessen rekombinanter Expression in E. coli erfolgten
spezifische und unspezifische Mutagenesen zu Steigerung der Thermostabilität und der spezifischen
Aktivität der Cutinase (Boston et al. 1997, Bott et al. 2003).
Bei der Hydrolyse von PET-Filmen zeigte die modifizierte PmC die maximale Aktivität bei 50°C und pH
9,5 und somit eine gute Stabilität im alkalischen Bereich (Ronkvist et al 2009).
Die mit P. menocina ATCC 53552 gewonnene Rohenzymlösung wurde zunächst auf ihre
hydrolytischen Eigenschaften mittels p-NP Ester getestet (Tabelle 32). Nach der Aufreinigung und der
damit verbundenen Identifikation der beiden Hydrolasen Lipase 1 und 2 mit cutinolytischen
Aktivitäten, erfolgte die Bestimmung der hydrolytischen Verhältnisse von p-NPB/p-NPC. Lipase 1

Ergebnisse und Diskussion 134
zeigte ein Verhältnis von 4,6 und Lipase 2 1,4. Beide Enzyme besaßen ein Molekulargewicht von
30 kDa (Gray et al. 1995).
Die Enzympräparation der Fusarium solani Cutinasen hydrolysierte ebenfalls verschiedene
Modellestersubstrate, allerdings nicht Triglyceride und enthielt somit keine unspezifische Lipase. Bei
der Hydrolyse von p-Nitrophenylpalmitat zeigte die Präparation ein deutliches pH Optimum bei 8,5
und mit Triton X-100 erfolgte eine Steigerung der hydrolytischen Aktivität (maximale Aktivität wurde
mit 3,7 mg/ml des Detergenz erreicht). Mit p-Nitrophenylester von C2-C18 ergaben sich vergleichbare
Werte für Km und Vmax, was auf keine vorhandene Spezifität schließen lässt. DFP inhibierte die
Enzympräparation, hingegen Jodacetamid und p-Chlormercuribenzoesäure (reagieren beide mit SH-
Gruppen, Cystein) hatten keinen Einfluss auf die Enzymaktivität, was bedeutet, dass es sich bei den
Fusarium Depolymerasen um Serin Hydrolasen handelt (Purdy und Kolattukudy 1973). Das
Molekulargewicht der Cutinase-Isoenzyme lag bei 22 kDa und das der Esterase bei 52 kDa (Purdy und
Kolattukudy 1975a). Die Cutinasen zeigten unter dem pH-Wert 7 geringe Aktivität, oberhalb von 7
stieg die Aktivität an bis pH 10. Oberhalb von pH 10 fiel die Hydrolyserate wieder ab.
Die beiden Cutinasen zeigten eine sinkende Hydrolyserate mit steigender Kettenlänge der Acylketten
der p-NP Esterer. Nur die berechneten Hydrolysegeschwindigkeiten V für p-Nitrophenyl-Acetat und
Butyrat waren für beide Cutinasen ähnlich, im weiteren Verlauf der Hydrolyse mit längeren p-
NPEstern zeigte die Cutinase I deutlich höhere Werte als die Cutinase II. Somit unterschieden sich die
beiden Cutinasen in ihren Fähigkeiten bei der Hydrolyse von löslichen Substraten, hingegen nicht bei
der Hydrolyse von Cutin. Die unspezifische Esterase hingegen zeigte mit den p-NP Estern deutlich
höhere Hydrolyseraten als die beiden Cutinasen und mit steigender Kettenlänge fiel die
Hydrolyseaktivität der Esterase nicht so stark ab wie bei den Cutinasen. Somit sind die Cutinasen
spezifischer für kurzkettige Ester im Gegensatz zur Esterase mit einem breiteren Hydrolysespektrum.
Bei den Studien zur Identifikation der Aminosäuren am aktiven Zentrum zeigte sich, dass die beiden
Cutinasen durch Dip-F, Paraoxon und Parathion inhibiert wurden, jedoch nicht durch N-
Ethylmaleimid, Jodoacetamid und p-Chloromercuribenzoat (Purdy und Kolattukudy 1975b). Somit
konnte nachgewiesen werden, dass die Cutinasen Serin Hydrolasen sind, jedoch keine Sulfhydryl-
Hydrolasen, wobei auch hier die Cutinase I stärker inhibiert wurde als die Cutinase II. Die
unspezifische Esterase wurde ebenfalls durch Dip-F und Paraoxon inhibiert, wobei Paraoxon das
Enzym stärker inhibierte. Bei den Cutinasen war es genau umgekehrt. Somit ist die unspezifische
Esterase ebenfalls eine Serin Hydrolase. Bei weiteren Untersuchungen stellte sich heraus, dass die
beiden Cutinasen nur ein aktives Serin besitzen. Eine genauere Studie zum Hydrolysemechanismus
der Cutinasen und der unspezifischen Esterase beim Cutinabbau sollte klären, ob diese Enzyme
eventuell parallel als Endo- und Exoenzyme agieren. Zunächst zeigte sich, dass die beiden Cutinasen
keine Unterschiede in ihren Spezifitäten und Art der Hydrolyse besaßen. Die Esterase hydrolysierte

Ergebnisse und Diskussion 135
kein Cutin und nur geringe Mengen an Oligomer im Vergleich zu den Cutinasen. Das bedeutet, dass
die Esterase für den Cutinabbau nicht essentiell ist und somit eine noch unbekannte Funktion besitzt.
Eventuell ist sie beteiligt an der Hydrolyse von cutikulären Wachsestern und wird deshalb mit den
Cutinasen coinduziert (Purdy und Kolattukudy 1975b).
Die 3-dimensionale Struktur einer Cutinase aus F. solani sp. pisi wurde nach der Klonierung und
Expression in E. coli mit einer Auflösung von 1,6 Å und 1,0 Å untersucht. Die Cutinase besteht aus 197
Aminosäuren und ist ein kompaktes Monomer von einer Größe von 45x30x30 Å. Der isoelekrische
Punkt lag bei 7,8. Das aktive Serin 120 ist in die Konsensus Sequenz Gly-Tyr-Ser-Gln-Gly eingebettet
und die beiden weiteren aktiven Aminosäuren sind Asp175 und His188. Das aktive Zentrum ist sehr
gut zugänglich auf der Oberfläche des ellipsoiden Proteins und ist umgeben von den
Aminosäureschleifen 80-87 und 180-188, welche sehr hydrophob sind (Carvalho et al. 1998).
Die Cutinase besaß vier nichtvariable Cysteine, welche zwei Disulfidbrücken ausbilden und somit eine
wichtige Rolle bei der Strukturbildung besitzen. Das aktive Zentrum zeichnete sich durch eine hohe
Flexibilität aus, was die Anpassung an verschiedene Substrate ermöglicht.
Trotz dass die Cutinasen lipolytische Aktivitäten besaßen, unterscheiden sie sich von den klassischen
Lipasen aufgrund der fehlenden Grenzflächenaktivierung. Bei der Cutinase besitzen die das aktive
Zentrum umgebende Proteinschleifen 80–87 und 180–188 hydrophobe Aminosäuren (Leu81, Gly82,
Ala85, Leu86, Pro87, Leu182, Ileu183 und Val184) welche die Interaktion zwischen Substrat und
Bindungstasche fördern. Abgesehen von zwei Seitenkettenbindungen von Aminosäuren, Leu81 und
Val184 bzw. Leu182 und Asn84, ist das katalytische Serin nicht verdeckt durch Proteinschlaufen,
sondern frei zugänglich für Lösungsmittel und Substrat. Somit ist keine Grenzflächenaktivierung nötig
für die Ausbildung der lipolytischen Aktivität, lediglich eine Neuorientierung einiger lipophiler
Seitenketten, z.B. Leu81 und Leu182, was als ein „Mini-Deckel“ bezeichnet werden kann, erfolgt vor
der Hydrolyse. Mit Hilfe der zeitbezogenen Fluoreszenz Analytik konnte die Bildung von Enzym-Lipid
Ansammlungen nachgewiesen werden, was bedeutet, dass sich die Cutinasen eher wie Lipasen ohne
Deckel als wie Esterasen verhalten (Carvalho et al. 1999)
Eine rekombinant produzierte F. solani pisi Cutinase zeigte eine optimale Temperatur von 30°C für
die Triolein Hydrolyse und 40°C für die Hydrolyse von p-NPB, wobei der optimale pH für beide bei 8
lag. Die Thermostabilität betrug mit 50% Restaktivität 85 Stunden bei 40°C bzw. 5 min bei 60°C (Chen
S et al. 2008). Bei der Hydrolyse von PET-Filmen zeigte die rekombinante FsC bei 50°C ihre maximale
Aktivität von 1,8 µmol/mL/h. Bei 60°C hingegen verlor sie ihre gesamte Aktivität. Der Einfluss des pH-
Wertes auf die PET Hydrolyse bei 50°C wurde ebenfalls untersucht und zeigte, dass die FsC Aktivität
sich im Bereich von pH 6,5 bis 8,5 nur gering ändert (1,9-1,6 µmol/mL/h), allerdings bei pH 9 auf
0,2 µmol/mL/h absank. Sie besaß somit eine geringe Stabilität im alkalischen Bereich. Die maximal
erreichte Abbaurate lag bei 0,9 µmol/mL/h für die FsC bei 40°C (Ronkvist et al 2009).

Ergebnisse und Diskussion 136
Die aufgereinigte Humicola (Thermomyces) insolens Cutinase HiC zeigte ein Molekulargewicht von
20-21 kDa und einen pI von 8. Das hydrolytische Enzym wurde durch PMSF inhibiert, somit ist ein
Serin Bestandteil des aktiven Zentrums. Das pH Optimum für die Hydrolyse von Tributyrat wurde
mittels pH-Stat bestimmt und lag bei 8. Die Cutinase zeigte aber auch bei pH 9 und 10 noch hohe
Aktivitäten (Sandal et al. 1996).
Nachdem verschiedene Substitutionen an dieser Cutinase durch molekularbiologische Methoden
erfolgten, wurden von der rekombinant produzierten Hydrolase weitere Eigenschaften bestimmt. Bei
einem pH Wert von 8 stieg die Hydrolyserate eines PET-Films mit steigender Temperatur von 30 bis
80°C um das 30fache auf 10,6 µmol/mL/h. Bei 90°C sank die Aktivität steil ab auf Null, wahrscheinlich
aufgrund von Denaturierung. Weitere Temperatur-abhängige Filmhydrolyse-Studien zeigten, dass ein
drastischer Anstieg der Hydrolyseaktivität oberhalb von 65°C erfolgte, da diese Temperaturen nahe
der Glasübergangstemperatur des PETs lag (75°C). Die maximale initiale Aktivität der HiC lag bei
13,6 µmol/mL/h bei 80°C und pH 8,5. Die maximale Hydrolyserate bei 70°C lag bei 6,1 µmol/mL/h
(Ronkvist et al. 2009).
Beim Vergleich dieser fünf verschiedenen PET-Hydrolasen wird zunächst deutlich, dass sie aufgrund
der Herkunft, Größe und Klasse in drei verschiedene Gruppen eingeteilt werden können, was in
Tabelle 30 auch schon erfolgt ist. Allerdings gibt es auch deutliche Unterschiede zwischen den
Enzymen, z.B. die optimalen Temperaturen und pH Werte oder die Fähigkeit zur Hydrolyse von
Triglyceriden (FsC und PmC hydrolysieren keine TAGs, HiC und TfH hydrolysieren TAGs). Da diese
Eigenschaften aber aufgrund von enzymatischen Assays bestimmt wurden, sind eine Vielzahl von
möglichen Einflüssen (z.B. verwendetes Substrat, Analytik, Puffer, Enzymmenge u.ä.) vorhanden,
welche zur Verschiebung der Ergebnisse beitragen können. Somit würde eine genauere
Untersuchung der Enzyme unter gleichen katalytischen Bedingungen mehr Klarheit über ihre
Einordnung bringen. Interessanterweise zeigten sich zwischen dem Pilz F. solani f. pisi und dem
Bakterium T. fusca einige Parallelen in Bezug auf die Enzymproduktion. Beide bilden zwei Cutinase-
Isoenzyme, welche sich hinsichtlich spezifischer Hydrolyseeigenschaften unterscheiden. Bei beiden
Organismen besitzen die Isoenzyme die gleiche Anzahl an Aminosäuren, unterscheiden sich
allerdings hinsichtlich der Sequenz im geringen Maße (T. fusca 7%, bei F. solani keine Angabe). Von
deutlich höherem Interesse für die vorliegende Studie war allerdings, dass der Pilz ebenfalls eine
Esterase von 52 kDa produzierte. Da zum damaligen Zeitpunkt, als die Studien zu diesen
eukaryotischen Enzymen erfolgten, die beiden Cutinasen von deutlich höherem Interesse waren,
erfolgten keine weiterführenden Untersuchungen, wie z.B. eine Sequenzierung der Esterase. Die
F. solani f. pisi Esterase unterscheidet sich von der TfCa dadurch, dass diese p-NPB und p-NPP
hydrolysiert, die TfCa hingegen nicht (Purdy und Kolattukudy 1975b, Billig et al. 2010).
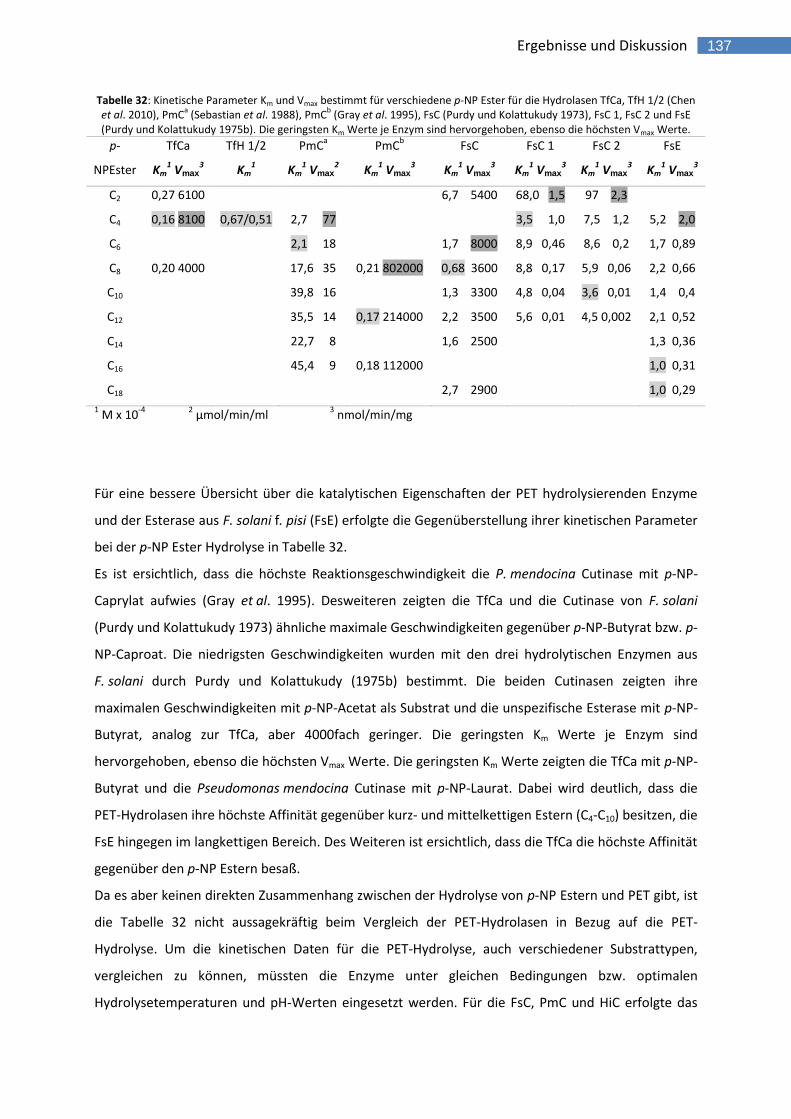
Ergebnisse und Diskussion 137
Tabelle 32: Kinetische Parameter Km und Vmax bestimmt für verschiedene p-NP Ester für die Hydrolasen TfCa, TfH 1/2 (Chen et al. 2010), PmC
a (Sebastian et al. 1988), PmC
b (Gray et al. 1995), FsC (Purdy und Kolattukudy 1973), FsC 1, FsC 2 und FsE
(Purdy und Kolattukudy 1975b). Die geringsten Km Werte je Enzym sind hervorgehoben, ebenso die höchsten Vmax Werte.
p-
NPEster
TfCa
Km1 Vmax
3
TfH 1/2
Km1
PmCa
Km1 Vmax
2
PmCb
Km1 Vmax
3
FsC
Km1 Vmax
3
FsC 1
Km1 Vmax
3
FsC 2
Km1 Vmax
3
FsE
Km1 Vmax
3
C2 0,27 6100 6,7 5400 68,0 1,5 97 2,3
C4 0,16 8100 0,67/0,51 2,7 77 3,5 1,0 7,5 1,2 5,2 2,0
C6 2,1 18 1,7 8000 8,9 0,46 8,6 0,2 1,7 0,89
C8 0,20 4000 17,6 35 0,21 802000 0,68 3600 8,8 0,17 5,9 0,06 2,2 0,66
C10 39,8 16 1,3 3300 4,8 0,04 3,6 0,01 1,4 0,4
C12 35,5 14 0,17 214000 2,2 3500 5,6 0,01 4,5 0,002 2,1 0,52
C14 22,7 8 1,6 2500 1,3 0,36
C16 45,4 9 0,18 112000 1,0 0,31
C18 2,7 2900 1,0 0,29
1 M x 10
-4
2 µmol/min/ml
3 nmol/min/mg
Für eine bessere Übersicht über die katalytischen Eigenschaften der PET hydrolysierenden Enzyme
und der Esterase aus F. solani f. pisi (FsE) erfolgte die Gegenüberstellung ihrer kinetischen Parameter
bei der p-NP Ester Hydrolyse in Tabelle 32.
Es ist ersichtlich, dass die höchste Reaktionsgeschwindigkeit die P. mendocina Cutinase mit p-NP-
Caprylat aufwies (Gray et al. 1995). Desweiteren zeigten die TfCa und die Cutinase von F. solani
(Purdy und Kolattukudy 1973) ähnliche maximale Geschwindigkeiten gegenüber p-NP-Butyrat bzw. p-
NP-Caproat. Die niedrigsten Geschwindigkeiten wurden mit den drei hydrolytischen Enzymen aus
F. solani durch Purdy und Kolattukudy (1975b) bestimmt. Die beiden Cutinasen zeigten ihre
maximalen Geschwindigkeiten mit p-NP-Acetat als Substrat und die unspezifische Esterase mit p-NP-
Butyrat, analog zur TfCa, aber 4000fach geringer. Die geringsten Km Werte je Enzym sind
hervorgehoben, ebenso die höchsten Vmax Werte. Die geringsten Km Werte zeigten die TfCa mit p-NP-
Butyrat und die Pseudomonas mendocina Cutinase mit p-NP-Laurat. Dabei wird deutlich, dass die
PET-Hydrolasen ihre höchste Affinität gegenüber kurz- und mittelkettigen Estern (C4-C10) besitzen, die
FsE hingegen im langkettigen Bereich. Des Weiteren ist ersichtlich, dass die TfCa die höchste Affinität
gegenüber den p-NP Estern besaß.
Da es aber keinen direkten Zusammenhang zwischen der Hydrolyse von p-NP Estern und PET gibt, ist
die Tabelle 32 nicht aussagekräftig beim Vergleich der PET-Hydrolasen in Bezug auf die PET-
Hydrolyse. Um die kinetischen Daten für die PET-Hydrolyse, auch verschiedener Substrattypen,
vergleichen zu können, müssten die Enzyme unter gleichen Bedingungen bzw. optimalen
Hydrolysetemperaturen und pH-Werten eingesetzt werden. Für die FsC, PmC und HiC erfolgte das

Ergebnisse und Diskussion 138
bereits durch Ronkvist et al. (2009), mit der TfCa und den beiden Cutinasen aus T. fusca KW3 wurden
verschiedene Hydrolysestudien durchgeführt, welche im nächsten Kapitel zusammengefasst sind.
3.3 Hydrolyse von PET Substraten durch prokaryotische und eukaryotische Hydrolasen
Es erfolgte die Untersuchung der Hydrolyse von zyklischen PET-Trimeren, unbehandelten PET-Fasern
und einer APET-Folie durch die PET-Hydrolasen aus T. fusca KW3 und weiteren PET-Hydrolasen. Bei
den Enzymen handelte es sich um die rekombinant produzierten Cutinasen 1 und 2 aus T. fusca
WSH03-11 (Chen et al. 2010, Chen J et al. 2008), die Cutinasen aus F. solani sp. pisi (Chen et al. 2010,
Vertommen et al. 2005) und eine Cutinasepräparation von Novozymes (NS-29061, LN05012). Bis zu
diesem Zeitpunkt wurden die meisten Hydrolysestudien mit der Cutinase aus F. solani sp. pisi
durchgeführt, welche deshalb bei den folgenden Untersuchungen als Vergleich eingesetzt wurde.
Zur Abschätzung der Reinheit der Enzympräparationen wurden Aliquote auf ein SDS Gel aufgetrennt.
Abbildung 65: SDS-PAGE der für die Abbauuntersuchungen eingesetzten Hydrolasen, linkes Gel mit Aktivitätsfärbung, rechtes Gel Aktivitäts- und Coomassie-Färbung. Spur 1: Carboxylesterase TfCa aus T. fusca KW3; Spur 2: rekombinante Cutinase 1 von T. fusca WSH03-11 (Chen et al. 2010, Chen J et al. 2008); Spur 3: rekombinante Cutinase 2 von T. fusca WSH03-11 (Chen et al. 2010, Chen J et al. 2008); Spur 4: rekombinante Cutinase von F. solani pisi (Chen et al. 2010); Spur 5: Cutinase aus Fusarium solani sp. pisi (Vertommen et al. 2005); Spur 6: Cutinase von Novozymes. Zum Vergleich die Kulturüberstände verschiedener MSV-Medien, Spur 7: Cutin-induzierte Hydrolase aus T. fusca KW3; Spur 8: Suberin-induzierte Hydrolase aus T. fusca KW3; St
1: Spectra™ Multicolor Broad Range Protein Ladder (Fermentas); St
2:
PageRulerTM
Plus Prestained Protein Ladder (Fermentas).
Die Hydrolasen aus T. fusca (Billig et al. 2010, Chen et al. 2010, Chen J et al. 2008) (Spuren 1-3) sind
aufgrund der Aktivitätsfärbung eindeutig im Gel identifizierbar und die Enzympräparationen zeichnen
sich durch eine hohe Reinheit aus. Die rekombinante Cutinase von F. solani pisi (Chen et al 2010)
(Spur 4) zeigte keine Aktivität und auch keine Proteinbande auf dem Gel. Wahrscheinlich wurde zu
wenig Protein aufgetragen bzw. die Cutinase war nicht aktiv gegenüber dem Substrat der
Aktivitätsfärbung. Die Cutinase aus F. solani sp. pisi (Vertommen et al. 2005) zeigte ebenfalls keine
Aktivität, aber eine Proteinbande (Spur 5), was ebenfalls auf ein Inaktivität des Cutinase gegenüber
St1 1 2 3 4 5 6 St
2 7 8 St
1 1 2 3 4 5 6 St
2 7 8
260 kDa 135 kDa 95 kDa 72 kDa
52 kDa
42 kDa
34 kDa
26 kDa
17 kDa 10 kDa
250 kDa 130 kDa 95 kDa 72 kDa
55 kDa
36 kDa
28 kDa
17 kDa 11 kDa

Ergebnisse und Diskussion 139
2-Naphthylacetat hinweist. Beide Enzyme wiesen aber deutliche Aktivitäten mit p-NPB auf. Die
Cutinasepräparation von Novozymes wies eine deutliche Esteraseaktivität im Gel auf (Spur 6).
Aufgrund der detektierten Größe im Gel (ca. 20 kDa) handelt es sich vermutlich um eine Cutinase aus
T. insolens oder F. solani pisi. Zum Vergleich wurden ebenfalls die zellfreien Überstände aus Kulturen
mit Cutin-MSV und Suberin-MSV in Spur 7 und 8 aufgetragen, welche eine Esterase im Bereich von
28 kDa aufweisen, aber ebenfalls noch weitere Proteine.
Nach Auftrag der verschiedenen Enzympräparationen auf PCL Agarplatten wurde die bereits erfolgte
Einordung der PET-Hydrolasen in Cutinasen und Carboxylesterase bestätigt. Die Cutinasen waren in
der Lage, das Biopolymer Cutin abzubauen und ebenfalls PCL, wohingegen die Carboxylesterase diese
beiden Polymere nicht hydrolysierte. Alle Cutinasen zeigten eine Hofbildung auf den PLC-
Agarplatten, hingegen wies die TfCa, eine Carboxylesterase, keinen PCL-Abbau auf (Abbildung 66).
Die rekombinant produzierten TfCa, TfCut1 und TfCut2 aus T. fusca KW3 zeigten dasselbe
Abbauverhalten (Abbildung 66).
Abbildung 66: PCL Abbautest der für die Abbauuntersuchungen eingesetzten Hydrolasen (obere Reihe: Carboxylesterase TfCa aus T. fusca KW3 (links), rekombinante Cutinase 1 von T. fusca WSH03-11 (Mitte, Chen et al. 2010, Chen J et al. 2008), rekombinante Cutinase 2 von T. fusca WSH03-11 (rechts, Chen et al. 2010, Chen J et al. 2008); mittlere Reihe: rekombinante Cutinase von F. solani pisi (links, Chen et al 2010), Cutinase aus F. solani sp. pisi (Mitte, Vertommen et al. 2005), Cutinase von Novozymes (rechts); untere Reihe : rekombinant produzierten Hydrolasen aus T. fusca KW3: Carboxylesterase TfCa (links), Cutinase TfCut1 (Mitte), Cutinase TfCut2 (rechts).

Ergebnisse und Diskussion 140
3.3.1 Partielle Hydrolyse von APET-Filmen durch die PET-Hydrolasen
In den nachfolgenden Diagrammen ist die Bildung von Hydrolyseprodukten aus APET-Film durch die
PET-Hydrolasen dargestellt.
Abbildung 67: Detektierte Hydrolyseprodukte nach Inkubation des APET-Films mit TfCa aus T. fusca KW3.
Abbildung 68: Detektierte Hydrolyseprodukte nach Inkubation des APET-Films mit der Cutinase 2 aus T. fusca WSH03-11
von Chen et al. 2010 und Chen J et al. 2008.
0
5
10
15
20
25
30
0 10 20 30 40 50 60 70 80
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe
0
2000
4000
6000
8000
10000
12000
14000
16000
0 10 20 30 40 50 60 70 80
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe

Ergebnisse und Diskussion 141
Abbildung 69: Detektierte Hydrolyseprodukte nach Inkubation des APET-Films mit der Cutinase aus F. solani pisi von
Vertommen et al. 2005.
Abbildung 70: Detektierte Hydrolyseprodukte nach Inkubation des APET-Films mit der Cutinase aus F. solani pisi von Chen
et al. 2010.
0
100
200
300
400
500
0 10 20 30 40 50 60 70 80
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe
0
50
100
150
200
250
300
0 10 20 30 40 50 60 70 80
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe

Ergebnisse und Diskussion 142
Abbildung 71: Detektierte Hydrolyseprodukte nach Inkubation des APET-Films mit der Cutinasepräparation von
Novozymes.
Während der drei Tage Inkubation setzte die Cutinase 2 aus T. fusca WSH03-11 die höchste Menge
an löslichen Hydrolyseprodukten vom APET-Film frei (Abbildung 69). Gegenüber der
Carboxylesterase TfCa aus T. fusca KW3 war dies das 475fache an löslichen Hydrolyseprodukten,
gegenüber den F. solani pisi Cutinasen FsC und FspC waren es das 24fache und 40fache und
gegenüber der eukaryotischen Cutinase-Präparation war es das 114fache an löslichen
Hydrolyseprodukten. Alle PET-Hydrolasen wurden mit der Aktivität von 4 U/ml eingesetzt.
Tabelle 33: Vergleich der freigesetzten Abbauprodukte der PET-Hydrolasen mit APET-Film.
Enzym Temperatur Hydrolyseprodukte (%)
TPA MHET BHET EMT EBT
TfCa 50°C 45 10 0,1 0 45
T. fusca Cutinase 22 60°C 57 42 0,5 0,01 0
FsC1 30°C 0 53 2 8 37
FspC2 30°C 21 78 0,4 0,4 0
Novozymes Cutinase Präparation 30°C 18 82 0 0 0
1 Vertommen et al. 2005
2 Chen et al. 2010
Mit 45% waren Terephthalsäure (TPA) und 1,2-Ethylen-bis-terephthalat (EBT) die
Haupthydrolyseprodukte der Carboxylesterase TfCa, bei der Cutinase FsC waren es mit 53% Mono(2-
Hydroxyethyl)Terephthalat (MHET) und 37% EBT. Die Cutinase setzte als einziges Enzym kein TPA
frei, hingegen aber die höchste Menge an 1,2-Ethylen-mono-terephthalat-mono(2-
hydroxyethylterephthalat) (EMT, 8%) verglichen mit den anderen PET-Hydrolasen. Die
0
20
40
60
80
100
120
0 10 20 30 40 50 60 70 80
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe

Ergebnisse und Diskussion 143
Haupthydrolyseprodukte der Cutinase 2 aus T. fusca WSH03-11 waren mit 57% und 42% TPA und
MHET, ebenso bei FspC mit 78% MHET und 21% TPA. Es wurden nur geringste Mengen an Bis(2-
Hydroxyethylterephthalat) (BHET) freigesetzt. Die Hauptabbauprodukte der Cutinase-Präparation
von Novozymes war MHET mit 82% und TPA mit 18%. Es wurden keine weiteren Produkte detektiert
(Tabelle 33). Die Freisetzung der Hydrolyseprodukte erfolgte über den gesamten
Inkubationszeitraum.
Beim Vergleich der verschiedenen F. solani pisi Cutinase-Präparationen (Abbildung 68, 70), bei denen
jeweils 4 U/ml Esteraseaktivität zum APET-Filmabbau eingesetzt wurden, zeigte sich ein ähnliches
Abbauverhalten in Bezug auf Menge und Verteilung der Hauptabbauprodukte. MHET als
Hauptprodukt wurde mit 53% (Vertommen et al. 2005) und 57% (Chen et al. 2010) für die
eukaryotischen Cutinasen identifiziert. Alle weiteren Hydrolyseprodukte unterschieden sich
allerdings in der freigesetzten Menge bei den beiden Cutinasen. Diese Unterschiede könnten durch
Modifikationen bei der rekombinanten Expression entstanden sein, wie zum Beispiel bei der Codon-
Optimierung, beim Entfernen und Anfügen von Signalpeptiden, durch eine fehlende
posttranslationale Modifikation oder durch eine andere Faltung im Expressionswirt (Vertommen
et al. 2005, Chen et al. 2010, Chen S et al. 2008, Chen J et al. 2008). Von den zuvor für den Abbau von
APET-Film eingesetzten Hydrolasen zeigte die Cutinase 2 aus T. fusca WSH03-11 die höchste
Aktivität. Dies könnte auf die höhere Inkubationstemperatur von 60°C und der damit verbundenen
höheren Beweglichkeit der PET-Ketten auf der Filmoberfläche zurückgeführt werden (Marten et al.
2005). Die gleichen Beobachtungen machten auch Ronkvist et al. 2009, die eine modifizierte
Humicola (Thermomyces) insolens Cutinase bei 70°C mit PET-Filmen inkubierten und dabei die
höchste Aktivität verzeichneten im Vergleich zu einer Pseudomonas mendocina Cutinase bei 50°C
und einer Fusarium solani Cutinase bei 40°C (Ronkvist et al. 2009). Des Weiteren begünstigte die
Lage des aktiven Zentrums der Cutinase direkt an der Oberfläche des Enzyms ebenfalls die
Hydrolyseaktivität mit PET-Filmen (Abbildung 64d). Das aktive Zentrum der Carboxylesterase TfCa
liegt hingegen in einer Bindungstasche und ist somit schwerer zugänglich, besonders für größere
Substrate, wie den APET-Film (Abbildung 64a). Des Weiteren lag die Inkubationstemperatur der TfCa
mit APET-Film bei 50°C und somit 10°C geringer gegenüber der Cutinase, was eine niedrigere
Aktivität der Polymerketten zur Folge hat und somit die Hydrolyseaktivität des Enzyms beeinträchtigt
(Marten et al. 2005). Die eukaryotischen Cutinasen wurden aufgrund ihrer geringeren
Thermostabilität bei 30°C inkubiert, wobei die PET-Ketten des APET-Films noch weniger beweglich
waren und dadurch geringer mit dem aktiven Zentrum wechselwirken konnten.
Nach der Hydrolyse der APET-Filme durch die TfCa und die Cutinase FsC wurde die Oberfläche der
Filme rasterelektronenmikroskopisch analysiert (Abbildung 72). Im Vergleich zur unbehandelten
Kontrolle (A) wies die mit der Carboxylesterase behandelte Oberfläche kleine Löcher auf (B). Der

Ergebnisse und Diskussion 144
APET-Film, welcher von der eukaryotischen Cutinase angegriffen wurde, zeigte hingegen einen
großflächigen Abbau (C).
Abbildung 72: Rasterelektronenmikroskopische Analyse der enzymatisch behandelten APET-Film Oberflächen: Unbehandelte Kontrolle (A), Inkubation mit TfCa bei 50°C 74 h (B) und Inkubation mit Cutinase FsC bei 30°C 74 h (C).
Alle bisherigen Studien zur Oberflächenmodifikation durch Mikroorganismen oder Enzyme verglichen
die Oberflächenstrukturen nur mit einer Kontrolle bzw. einer alkalische Oberflächenhydrolyse der
Fasern oder Filme. Diese Hydrolysebehandlungen erfolgten hingegen mit zwei verschiedenen
Enzymen, deren Veränderungen auf der Oberfläche miteinander verglichen werden konnten und
deutliche Unterschiede untereinander, aber auch zu einer alkalischen Hydrolyse aufwiesen
(Abbildung 72, Yoon et al. 2002). Die Ausbildung verschiedener Oberflächenstrukturen, abhängig von
der Behandlung (enzymatisch oder chemisch), wurde bereits in anderen Studien dargelegt. Yoon
et al. wiesen als Erste nach, dass nach der Behandlung von PET-Fasern mit einer PET-Hydrolase die
Faseroberfläche signifikante Oberflächenmodifikationen aufwiesen, welche sich von denen bei einer
NaOH Behandlung deutlich unterschieden (Yoon et al. 2002). Auch Zhang et al. behandelten PET-
Fasern mit Mikroorganismen und einer Lipase und konnten mittels SEM eine deutliche
Oberflächenerosion im Vergleich zu unbehandelten Fasern nachweisen (Zhang et al. 2004). Ähnliche
Ergebnisse wurden auch von Kim und Song 2006 veröffentlicht, die die Oberflächenhydrolyse von
PET-Fasern mit einer Lipase und NaOH verglichen und unterschiedliche Oberflächenstrukturen
nachwiesen. Im Gegensatz dazu konnten bei weiteren Untersuchungen mit PET-Fasern beim
Vergleich der enzymatischen und basischen Hydrolyse bei der enzymatischen Behandlung keine
Veränderung der Oberflächenstruktur im Vergleich zur Kontrolle nachgewiesen werden (Eberl et al.
2008, Brueckner et al 2008). Diese Beobachtungen wurden auf eine besonders faserschonende und
minimale Oberflächenmodifikation zurückgeführt. Es könnte allerdings auch auf eine zu geringe
Inkubationszeit oder Enzymmenge hinweisen, womit die möglichen Auswirkungen der
enzymatischen Behandlung noch nicht für die Oberflächenanalyse sichtbar waren. Feuerhack et al.
wiesen, ähnlich zu allen anderen Autoren, ebenfalls eine leichte Oberflächenveränderung nach der
enzymatischen Hydrolyse der PET-Fasern durch eine Enzympräparation von T. fusca KW3 nach
(Feuerhack et al 2008).
A B C

Ergebnisse und Diskussion 145
Die Untersuchung der veränderten Oberflächenstrukturen von PET-Filmen erfolgte erstmals durch
Donelli et al. 2009 und Ronkvist et al. 2009. Dabei konnten Donelli et al. keine Veränderungen der
Oberflächenstruktur nach der enzymatischen Hydrolyse nachweisen, hingegen aber deutliche
Veränderungen nach der Hydrolyse mit NaOH (Donelli et al. 2009). Dabei handelte es sich allerdings
um einen PET-Film mit einer Kristallinität von 34,8%, welcher sich im Gegensatz zu amorphen PET-
Filmen für enzymatische Hydrolysen nicht eignet. Diese Tatsache bestätigte Ronkvist et al. bei der
enzymatischen Oberflächenhydrolyse eines gering-kristallinen PET-Films mit einer Kristallinität von
7,0% ± 0,5%. Während der verwendete kristalline PET-Film (35,0% ± 0,5%) kaum von den Enzymen
hydrolysiert wurde, konnten mit dem amorphen PET-Film deutliche Oberflächenveränderungen
mittels SEM nachgewiesen werden (Ronkvist et al. 2009). Bei einem zeitlichen Vergleich der
hydrolysierten Oberflächenstrukturen nach 12, 48 und 96 Stunden zeigte sich eine deutliche
Zunahme an porösen Strukturen. Die nachgewiesenen Strukturen gleichen allerdings nicht den
Oberflächenstrukturen aus der Abbildung 72 nach 74 Stunden Inkubation. Mögliche Erklärungen sind
zum Einen ein unterschiedliches Hydrolyseverhalten der verschiedenen Enzyme und zum Anderen
die Variationen bei den Inkubationstemperaturen (HiC bei 70°C (Ronkvist et al. 2009), TfCa bei 50°C
und FsC bei 30°C). Aus diesem Grund waren die Polymerketten des Films unterschiedlich mobil und
nur bei hohen Temperaturen besser hydrolysierbar (Marten et al. 2005).
Bei den Abbauuntersuchungen wurden neben den erwarteten PET Monomeren TPA, MHET und
BHET auch zwei weitere Abbauprodukte mittels RP-LC-MS detektiert. Bei den Produkten handelte es
sich um 1,2-Ethylen-mono-terephthalat-mono(2-hydroxyethylterephthalat) (EMT) und 1,2-Ethylen-
bis-terephthalat (EBT), zwei weitere Dimere des PET. Die Strukturen, die dazugehörigen molaren
Massen und die MS-Spektren der Hydrolyseprodukte sind nachstehend zusammengefasst.
Vertommen et al. hatten 2005 auch zwei unbekannte Hydrolyseprodukte detektiert, denen sie keine
Struktur zuordnen konnten und bezeichneten sie als Produkt A und B (Rt 18,1 und 18,7 min). Sie
vermuteten, dass es sich dabei um Dimere des PET handelte (Vertommen et al. 2005). Aufgrund der
übereinstimmenden Retentionszeiten handelt es sich bei den beiden unbekannten Dimeren
wahrscheinlich um die identifizierten Produkte EMT und EBT.

Ergebnisse und Diskussion 146
Abbildung 73: Chromatogramm der HPLC Analyse des enzymatischen APET Abbaus durch die TfCa.
EMT
EBT
TPA
MHET
BHET
10 15 20
0.00
0.25
0.50
0.75
1.00
1.25
AU
5 25
Retentionszeit (min)
1.50
UV
(2
41
nm
)
(24
1n
m)
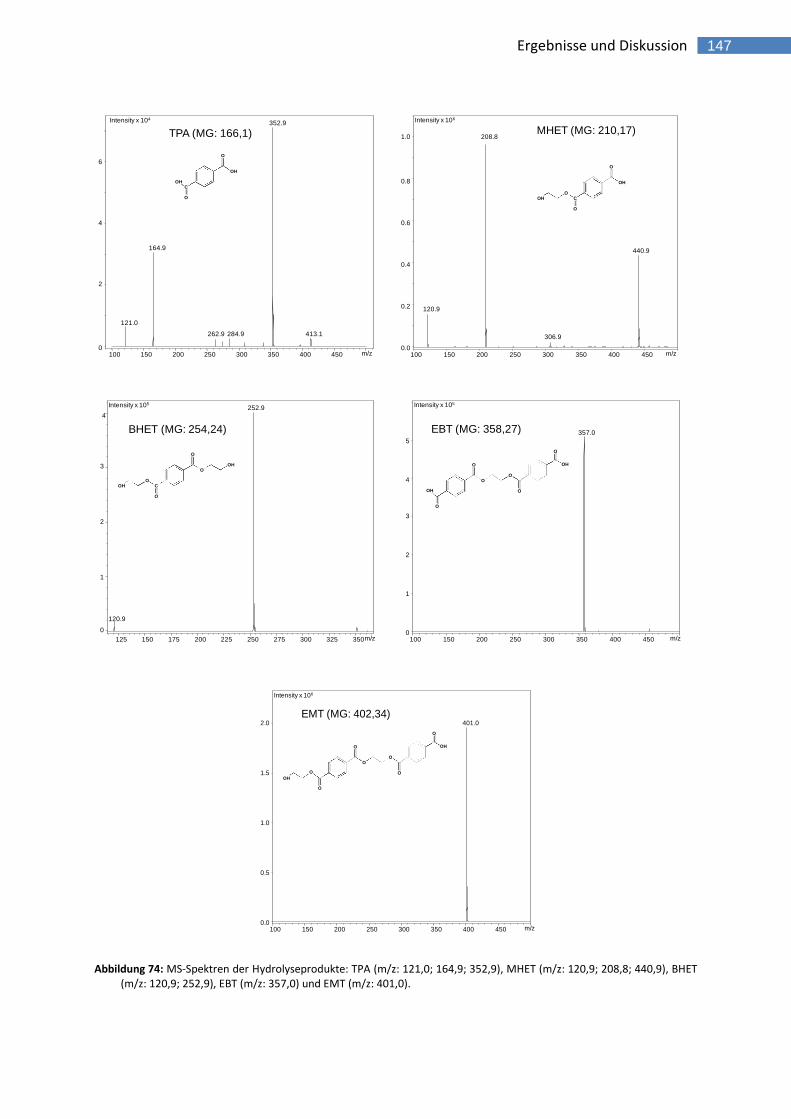
Ergebnisse und Diskussion 147
121.0
164.9
262.9 284.9
352.9
413.1
0
2
4
6
Intensity x 104
100 150 200 250 300 350 400 450 m/z
O
O
COH
OH
TPA (MG: 166,1)
120.9
208.8
306.9
440.9
0.0
0.2
0.4
0.6
0.8
1.0
Intensity x 106
100 150 200 250 300 350 400 450 m/z
O
O
C
OH
OOH
MHET (MG: 210,17)
120.9
252.9
0
1
2
3
4
Intensity x 105
125 150 175 200 225 250 275 300 325 350m/z
O
O
CO
OH
OOH
BHET (MG: 254,24) 357.0
0
1
2
3
4
5
Intensity x 105
100 150 200 250 300 350 400 450 m/z
O
OO
O
O
O
OH
OH
EBT (MG: 358,27)
401.0
0.0
0.5
1.0
1.5
2.0
Intensity x 106
100 150 200 250 300 350 400 450 m/z
O
OH
OO
O
O
O
O
OH
EMT (MG: 402,34)
Abbildung 74: MS-Spektren der Hydrolyseprodukte: TPA (m/z: 121,0; 164,9; 352,9), MHET (m/z: 120,9; 208,8; 440,9), BHET
(m/z: 120,9; 252,9), EBT (m/z: 357,0) und EMT (m/z: 401,0).

Ergebnisse und Diskussion 148
3.3.2 Partielle Hydrolyse von PET-Fasern durch die PET-Hydrolasen
In den nachfolgenden Diagrammen ist die Bildung von Hydrolyseprodukten aus PET-Fasern durch die
PET-Hydrolasen dargestellt.
Abbildung 75: Detektierte Hydrolyseprodukte nach Inkubation der PET-Fasern mit TfCa aus T. fusca KW3.
Abbildung 76: Detektierte Hydrolyseprodukte nach Inkubation der PET-Fasern mit der Cutinase 2 aus T. fusca WSH03-11
von Chen et al. 2010, Chen J et al. 2008.
0
25
50
75
100
125
0 10 20 30 40 50 60 70 80
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe
0
50
100
150
200
250
300
350
400
0 10 20 30 40 50 60 70 80
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe

Ergebnisse und Diskussion 149
Abbildung 77: Detektierte Hydrolyseprodukte nach Inkubation der PET-Fasern mit der Cutinase aus F. solani pisi von
Vertommen et al. 2005.
Abbildung 78: Detektierte Hydrolyseprodukte nach Inkubation der PET-Fasern mit der Cutinase aus F. solani pisi von Chen
et al. 2010.
0
25
50
75
100
125
0 10 20 30 40 50 60 70 80
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe
0
10
20
30
40
50
60
0 10 20 30 40 50 60 70 80
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe

Ergebnisse und Diskussion 150
Abbildung 79: Detektierte Hydrolyseprodukte nach Inkubation der PET-Fasern mit der Cutinasepräparation von Novozymes.
Die Cutinase 2 aus T. fusca WSH03-11 setzte während der Inkubation mit PET-Fasern die höchsten
Mengen an Hydrolyseprodukten frei. Verglichen mit der TfCa war das nach dreitägiger Inkubation das
2,75fache, mit der Cutinase-Präparation von Novozymes das 2,3fache und mit den beiden Cutinasen
aus F. solani pisi FsC und FspC das 3,8fache und 6,5fache.
Tabelle 34: Vergleich der freigesetzten Abbauprodukte der PET-Hydrolasen mit PET-Fasern.
Enzym Temperatur Hydrolyseprodukte (%)
TPA MHET BHET EMT EBT
TfCa 50°C 67 31 0 0,3 2
T. fusca Cutinase 22 60°C 78 22 0 0 0
FsC1 30°C 1 84 2 1 12
FspC2 30°C 14 69 0 18 0
Novozymes Cutinase Präparation 30°C 16 57 4 0 24
1 Vertommen et al. 2005
2 Chen et al. 2010
Dabei produzierte TfCa während der gesamten Inkubation Abbauprodukte, welche durch
Umkehrphasen HPLC nachgewiesen wurden (Tabelle 34). Die Cutinase aus F. solani pisi hingegen
setzte die meisten Hydrolyseprodukte in den ersten 24 Stunden der Inkubation frei. Mit 67% und
31% waren TPA und MHET die Haupthydrolyseprodukte der TfCa, wobei die Menge an TPA während
der gesamten Behandlung stetig anstieg, aber die Menge an MHET nach 24 Stunden bis 72 Stunden
Inkubation konstant blieb. Die Haupthydrolyseprodukte der FsC waren MHET (84%) und EBT (12%).
Im Gegensatz zum APET-Film Abbau wurden durch die FsC von den PET-Fasern geringe Mengen an
0
20
40
60
80
100
120
140
160
180
0 10 20 30 40 50 60 70 80
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe

Ergebnisse und Diskussion 151
TPA freigesetzt. Hingegen wurde das während der ersten 24 Stunden gebildete EBT während der
weiteren Reaktionszeit wieder abgebaut.
Die beiden Cutinasen Cutinase 2 aus T. fusca und FspC aus F. solani zeigten bei der enzymatischen
Hydrolyse der höherkristallinen PET-Fasern unterschiedliche Hauptprodukte und Aktivitäten
gegenüber den Fasern. Dabei produzierten beide Enzyme während der gesamten Reaktionszeit
Abbauprodukte, welche durch Umkehrphasen HPLC nachgewiesen wurden (Tabelle 34). Mit 78% und
22% waren TPA und MHET die Abbauprodukte der Cutinase 2 aus T. fusca WSH03-11, wobei die
Menge an TPA während der gesamten Behandlung stetig anstieg, aber die Menge an MHET nach
24 Stunden bis zum Ende der Inkubation konstant blieb. Die Haupthydrolyseprodukte der
eukaryotischen Cutinase waren MHET (69%) und EBT (18%). Ein ähnliches Verhalten zeigte auch die
F. solani pisi Cutinase FsC mit PET-Fasern und eine Esterase aus Penicillium citrinum, die
hauptsächlich MHET und BHET und nur geringe Mengen an TPA von PET-Textilien freisetzte, was
bedeuten könnte, dass eukaryotische hydrolytische Enzyme ähnliche Abbaumechanismen besitzen
(Liebminger et al. 2007). Vergangene Studien haben ebenfalls bestätigt, dass die eukaryotische
Cutinase aus F. solani pisi nur geringe Aktivitäten gegenüber höher kristallinen PET-Filmen aufweist
(Vertommen et al. 2005). Die eukaryotische Cutinase setzte das 5fache an löslichen Produkten
während der Inkubation mit niederkristallinen APET-Filmen (4%) frei im Vergleich zur Inkubation mit
höherkristallinen PET-Fasern (Abbildung 68, 77), was mit bereits veröffentlichten Studien zum
Abbauverhalten dieser Cutinase in Bezug auf die Kristallinität übereinstimmt (Müller et al. 2005,
Vertommen et al 2005). Der Abbau von aromatischen Polyester wird durch die geringe Beweglichkeit
der Polymerketten im kristallinen Bereich behindert (Marten et al. 2003, 2005).
Die Hauptabbauprodukte der Cutinase-Präparation von Novozymes waren MHET (57%) und EBT
(24%). Beim Vergleich der freigesetzten Hydrolyseprodukte über die Reaktionszeit der Cutinase-
Präparation mit den eukaryotischen Cutinasen (Vertommen et al. 2005, Chen et al. 2010) zeigte sich,
dass die hydrolytische Aktivität für alle drei Enzyme ähnlich war. Aus diesen Beobachtungen kann
geschlussfolgert werden, dass in der Enzympräparation von Novozymes eine eukaryotische Cutinase
enthalten sein könnte.
3.3.3 Hydrolyse von PET-Trimeren durch die PET-Hydrolasen
In den nachfolgenden Diagrammen ist die Bildung von Hydrolyseprodukten aus CTR durch die PET-
Hydrolasen dargestellt.

Ergebnisse und Diskussion 152
Abbildung 80: Detektierte Hydrolyseprodukte nach Inkubation der zyklischen PET-Trimeren mit TfCa aus T. fusca KW3.
Abbildung 81: Detektierte Hydrolyseprodukte nach Inkubation der zyklischen PET-Trimeren mit der Cutinase 1 aus T. fusca
WSH03-11 von Chen et al. 2010, Chen J et al. 2008.
0
5000
10000
15000
20000
25000
0 10 20 30 40 50 60 70 80
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe
0
5000
10000
15000
20000
25000
0 10 20 30 40 50 60 70 80
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe

Ergebnisse und Diskussion 153
Abbildung 82: Detektierte Hydrolyseprodukte nach Inkubation der zyklischen PET-Trimeren mit der Cutinase 2 aus T. fusca
WSH03-11 von Chen et al. 2010, Chen J et al. 2008.
Abbildung 83: Detektierte Hydrolyseprodukte nach Inkubation der zyklischen PET-Trimeren mit rekombinanter TfCut1.
0
5000
10000
15000
20000
25000
30000
0 10 20 30 40 50 60 70 80
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe
0
5000
10000
15000
20000
0 6 12 18 24
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe

Ergebnisse und Diskussion 154
Abbildung 84: Detektierte Hydrolyseprodukte nach Inkubation der zyklischen PET-Trimeren mit rekombinanter TfCut2.
Abbildung 85: Detektierte Hydrolyseprodukte nach Inkubation der zyklischen PET-Trimeren mit der Cutinase aus F. solani pisi von Vertommen et al. 2005.
0
5000
10000
15000
20000
0 6 12 18 24
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe
0
5000
10000
15000
20000
25000
0 10 20 30 40 50 60 70 80
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe
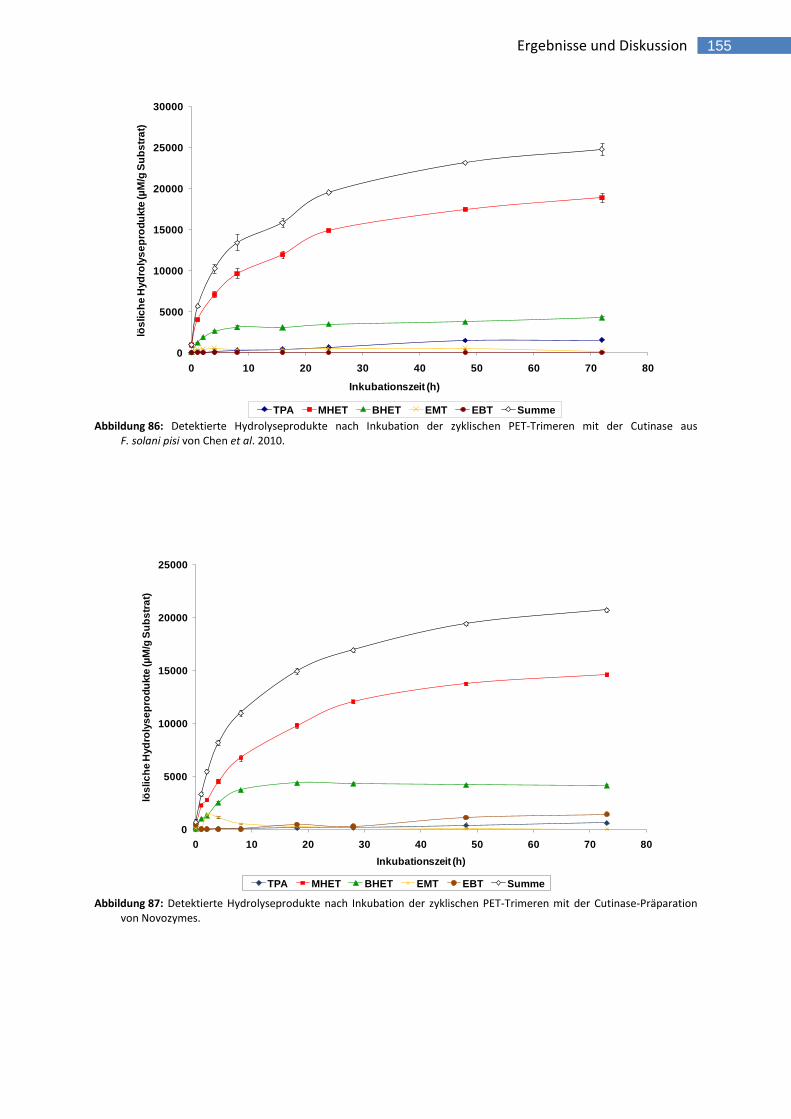
Ergebnisse und Diskussion 155
Abbildung 86: Detektierte Hydrolyseprodukte nach Inkubation der zyklischen PET-Trimeren mit der Cutinase aus
F. solani pisi von Chen et al. 2010.
Abbildung 87: Detektierte Hydrolyseprodukte nach Inkubation der zyklischen PET-Trimeren mit der Cutinase-Präparation
von Novozymes.
0
5000
10000
15000
20000
25000
30000
0 10 20 30 40 50 60 70 80
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe
0
5000
10000
15000
20000
25000
0 10 20 30 40 50 60 70 80
lös
lic
he
Hyd
roly
se
pro
du
kte
(µ
M/g
Su
bs
tra
t)
Inkubationszeit (h)
TPA MHET BHET EMT EBT Summe

Ergebnisse und Diskussion 156
Beim Vergleich wird deutlich, dass die bakterielle Cutinase 2 aus T. fusca WSH03-11 die höchste
Menge an Hydrolyseprodukten freisetzte. Im Vergleich zu den PET-Hydrolasen TfCa, T. fusca Cutinase
1, rTfCut1, rTfCut2, FsC, FspC und der Novozymes Cutinase Präparation war das das bis zu 1.4fache.
Es ist auch ersichtlich, dass alle PET-Hydrolasen mit den zyklischen Trimeren einen ca. 100fach
höheren Abbau erzielten als mit den beiden längerkettigen Substraten.
Tabelle 35: Vergleich der freigesetzten Abbauprodukte der PET-Hydrolasen mit PET-Trimeren.
Enzym Konz. Temperatur Hydrolyseprodukte (%)
TPA MHET BHET EMT EBT
TfCa 40 U
40 U
40 U
50°C 41 36 16 8 0
T. fusca Cutinase 1 60°C 7 68 13 12 0
T. fusca Cutinase 2 60°C 12 68 20 0 0
rTfCut1 39 U
58 U
60°C 11 72 17 0 0
rTfCut2 60°C 18 62 16 4 0
FsC1 40 U
40 U
40 U
30°C 3 61 34 2 0
FspC2 30°C 6 76 17 1 0
Novozymes Cutinase Präparation 30°C 3 70 20 0 7
1 Vertommen et al. 2005
2 Chen et al. 2010
TfCa setzte TPA (41%), MHET (36%) und BHET (16%) als Hauptabbauprodukte frei, hingegen
produzierte die Cutinase FsC 61% MHET, 34% BHET und geringe Mengen von EMT und TPA (Tabelle
35). Der Großteil der Hydrolyseprodukte wurde von den Enzymen nach 24 Stunden Reaktion
freigesetzt. Die eukaryotische Cutinase FspC setzte die Hydrolyseprodukten über 72 Stunden
Reaktionszeit frei. Hingegen produzierte die T. fusca Cutinase 2 die höchste Menge an
Hydrolyseprodukten in 16 Stunden Inkubation und die T. fusca Cutinase 1 bereits in 2 Stunden. Diese
Werte wurden auch für den Vergleich der freigesetzten Hydrolyseprodukte in Tabelle 35 verwendet.
Die Hauptprodukte der T. fusca Cutinase 2 waren MHET (68%) und BHET (20%). Bei der T. fusca
Cutinase 1 waren es 68% MHET und 13% BHET und bei der FspC 76% MHET und 17% BHET. Trotz der
unterschiedlichen Inkubationstemperaturen lagen die Konzentrationen der freigesetzten
Hydrolyseprodukte in einem ähnlichen Bereich, was mit der Struktur der Trimere und der daraus
folgenden Unabhängigkeit gegenüber der Inkubationstemperatur begründet werden kann. Die
Beweglichkeit langer Polymerketten wird beeinflußt durch die Inkubationstemperatur und somit ist
der enzymatische Abbau dieser Polymerketten mit steigender Inkubationstemperatur deutlich höher
(Marten et al. 2005). Die zyklischen PET-Trimere sind Oligomere, welche eine größere Oberfläche
besitzen und damit leichter zugänglich sind für hydrolytische Angriffe. Somit ist der enzymatische
Abbau von CTR abhängig von der zur Verfügung stehenden Oberfläche und weniger von der
Inkubationstemperatur. Weiterhin konnte gezeigt werden, dass die beiden Cutinasen der

Ergebnisse und Diskussion 157
untersuchten T. fusca Stämme in der Lage sind, PET abzubauen, was mit bereits veröffentlichen
Untersuchungen übereinstimmt (Müller et al. 2005, Eberl et al. 2008, Eberl et al. 2009).
Beim Vergleich der Cutinase FsC und FspC aus F. solani pisi (Vertommen et al. 2005, Chen et al. 2010)
zeigte sich, dass auch hier gute Übereinstimmungen bezüglich der Menge der Hydrolyseprodukte
und der Verteilung der Hauptprodukte vorliegen. Die Hauptabbauprodukte der Cutinasepräparation
von Novozymes waren MHET mit 70% und BHET mit 20% (Abbildung 87). Beim Vergleich des Abbaus
der Cutinasepräparation mit den eukaryotischen Cutinasen (Vertommen et al. 2005, Chen et al.
2010) zeigte sich, dass sie ähnliche Produkte auch für alle drei Substrate freisetzten. Diese
Beobachtung bestätigt, dass in der Enzympräparation von Novozymes wahrscheinlich ebenfalls eine
eukaryotische Cutinase enthalten ist.
Die Cutinase aus H. insolens DSM 1800 hydrolysierte zyklische PET-Trimere unter Freisetzung von
BHET, MHET und TPA (Hooker et al. 2003). Bei der Studie des Einflusses der Partikelgröße der CTR auf
ihre Abbaubarkeit zeigte sich, dass kleinere Partikel mit einer größeren Oberfläche schneller
abgebaut wurden als größere. Der Abbau wurde von der zur Verfügung stehenden Oberfläche
stärker beeinflusst als von der Kristallinität des PET (Figueroa et al. 2006). Somit wäre es möglich, die
Hydrolyseraten der beiden verwendeten Enzyme noch zu steigern, indem statt der verwendeten
grobdispersen Partikel kleinere mit einer größeren Oberfläche eingesetzt werden würden.
Die rekombinanten Hydrolasen aus T. fusca KW3 zeigten ähnliche Hydrolyseeigenschaften. Im
Gegensatz zu den anderen Enzymen setzten die Enzyme die Hydrolyseprodukte bereits nach 8
Stunden Inkubation frei. Das Hauptprodukt war MHET mit 72% (rTfCut1) bzw. 62% (rTfCut2) und TPA
mit 11% (rTfCut1) bzw. 18% (rTfCut2). BHET als Produkt wurde mit 17% (rTfCut1) und 16% (rTfCut2)
freigesetzt. Die beiden Cutinasen setzten nur 3% EMT (rTfCut2) oder gar kein EMT (rTfCut1) frei. Die
Hydrolyseaktivität unterschied sich nur in der Freisetzung von TPA, wobei die rTfCut2 während der
gesamten Reaktion TPA freisetzte und die rTfCut1 nur in den ersten 4 Stunden (Abbildung 86, 87). Im
Vergleich dazu zeigten die Cutinase 1 und 2 aus T. fusca WSH03-11 deutlichere Unterschiede bei der
Freisetzung der Hydrolyseprodukte aus CTR (Abbildung 81, 82). Dies könnte auf einen größeren
Unterschied zwischen den beiden Isoenzymen aus T. fusca WSH03-11 hinweisen im Vergleich zu den
beiden Cutinasen aus T. fusca KW3.
Die mit den verschiedenen Hydrolasen erzielten Ergebnisse sind in Tabelle 36 zusammengefasst. Es
zeigte sich, dass alle Enzyme die drei verschiedenen Substrate hydrolysierten. Das
Haupthydrolyseprodukt war in den meisten Fällen MHET, gefolgt von TPA und BHET. Für jedes PET-
Substrat sind in der Tabelle die beiden Hauptprodukte blau unterlegt. Dabei konnte kein direkter
Zusammenhang zwischen den Hydrolasen und den freigesetzten Produkten nachgewiesen werden.
Aufgrund der ähnlichen katalytischen Triade aller Hydrolasen sind ähnliche hydrolytische

Ergebnisse und Diskussion 158
Eigenschaften der Enzyme gegenüber einem identischen Substrat nicht überraschend. Dies bezieht
sich nur auf den Abbau von PET. Die Enzyme hydrolysierten hingegen mit unterschiedlicher Effizienz
den Biopolyester Cutin, wobei nur die Cutinasen aktiv gegenüber dem Biopolyester waren.
Die aktivste Hydrolase für den hydrolytischen Abbau aller drei Substrate war die Cutinase 2 aus
T. fusca WSH03 (Chen S et al. 2008, Chen J et al. 2008, Chen et al. 2010). T. fusca Cutinase 2 setzte
aus den CTR 27,5 mM Produkte/g Substrat frei, aus dem APET-Film 9,5 mM Produkte/g Substrat und
aus den PET-Fasern 0,3 mM Produkte/g Substrat.
Tabelle 36: Vergleich der Abbauprodukte der PET Hydrolasen mit den drei PET Substraten. Die beiden Hauptprodukte des Enzyms sind blau unterlegt.
Enzym Konz. Substrat Temp. Lösliche Hydrolyseprodukte (µM/g Substrat)
TPA MHET BHET EMT EBT Gesamt
TfCa 40 U
CTR
50°C 9924 9189 3988 1664 0 24765
rTfCut1 39 U
58 U
60°C 1810 10932 2746 0 0 15064
rTfCut2 60°C 2864 9889 2594 539 0 15886
T. fusca Cutinase 1 40 U
40 U
40 U
40 U
40 U
60°C 1410 14770 2872 2539 0 21591
T. fusca Cutinase 2 60°C 3175 18820 5488 0 0 27483
FsC1 30°C 604 11890 6557 342 0 19393
FspC2 30°C 1509 18862 4238 163 0 24772
Novozymes Cutinase
Präparation
30°C 588 14590 4139 0 1393 20710
TfCa
40 U APET
50°C 9 2 0.02 0 9 20
Cutinase 2 60°C 5432 4013 47 1 0 9493
FsC1 30°C 0 214 8 33 149 404
FspC2 30°C 50 186 1 1 0 239
Novozymes Cutinase
Präparation
30°C 15 68 0 0 0 83
TfCa
40 U Fasern
50°C 79 38 0 0.3 2 120
Cutinase 2 60°C 258 73 0 0 0 331
FsC1 30°C 1 72 2 1 10 86
FspC2 30°C 7 36 0 9 0 51
Novozymes Cutinase
Präparation
30°C 23 83 6 0 35 146
1 Vertommen et al. 2005
2 Chen et al. 2010
Die Carboxylesterase TfCa zeigte eine hohe Hyrolyserate beim Einsatz zur CTR Hydrolyse, hingegen
nicht bei der Hydrolyse der PET-Fasern oder des PET-Films. Diese Spezifität zeichnet die
Carboxylesterase für die Hydrolyse von kurzkettigen PET Substraten als besser geeignet aus.

Ergebnisse und Diskussion 159
Hooker et al. 2003 zeigten bei ihren hydrolytischen Studien zum CTR Abbau durch eine eukaryotische
Cutinase, dass bei Einsatz einer genügend großen Menge des Enzyms CTR vollständig zu TPA
hydrolysiert wird. Vergleichende Studien wurden dazu mit rTfCa und rTfCut2 mit verschiedenen
Enzymkonzentrationen durchgeführt.
In der nachstehenden Tabelle sind die Enzymaktivitäten mit p-NPB als Substrat der beiden Enzyme
bei den jeweiligen Protein-Konzentrationen zusammengefasst.
Tabelle 37: Proteinkonzentration der eingesetzten Enzyme und die dazugehörigen p-NPB-Aktivitäten in den Ansätzen des Abbauversuchs.
Protein
(µg/ml)
rTfCa
(U/ml)
rTfCut2
(U/ml)
5 4 536 648
10 9 072 1 296
50 45 360 6 481
75 68 040 9 722
100 90 720 12 962
250 226 800 32 405
Abbildung 88: Effekt der rTfCa Konzentration auf die Hydrolyse von PET-Trimeren (Reaktionszeit 4 Stunden).

Ergebnisse und Diskussion 160
Abbildung 89: Effekt der rTfCut2 Konzentration auf die Hydrolyse von PET-Trimeren (Reaktionszeit 4 Stunden).
Die eingesetzten Enzymaktivitäten mit p-NPB der rTfCa und rTfCut2 unterschieden sich um den
Faktor 10.
Beim Vergleich der beiden Diagramme wird sichtbar, dass beide Enzyme mit zunehmender
Enzymkonzentration MHET als Hauptprodukt freisetzten. TfCa bildete während allen Inkubationen
bei einer Reaktionszeit von 4 bzw. 8 Stunden kein EBT und TPA nur in geringen Mengen, die aber mit
zunehmender Enzymkonzentration und Inkubationsdauer zunehmen (Abbildung 88 und 90). Dies
bestätigte die zuvor durchgeführte Hydrolyse der CTR durch die TfCa über 74 Stunden. Die
Freisetzung der TPA erfolgte über den gesamten Reaktionszeitraum und die Konzentration von
9,9 mM TPA/g Substrat wurde erst am Ende der Reaktion nachgewiesen. Als zweites Hauptprodukt
setzte die Esterase bei den Ansätzen mit 5, 10, 50 und 75 µg/ml Enzym EMT frei, bei den Ansätzen
mit 100 und 250 µg/ml BHET und EMT in annähernd gleichen Mengen. Mit zunehmender
Enzymmenge und Inkubationsdauer stieg die freigesetzte Menge an MHET, BHET und TPA,
wohingegen die freigesetzte Menge an EMT mit der zunehmenden Enzymmenge und
Inkubationsdauer sank. Die maximale Freisetzung an Hydrolyseprodukten wurde bei den Ansätzen
mit 50 µg/ml Enzym und 4 Stunden Inkubation mit 54,3 mM/g Substrat detektiert, wobei bei den
Ansätzen mit den höheren Enzymkonzentrationen die Mengen an freigesetzten Hydrolyseprodukten
stagnierten. Die TfCut2 zeigte ähnliche Hydrolyse-Eigenschaften wie die Esterase, obwohl nur 1/10
der p-NPB Aktivität eingesetzt wurde. Ein deutlicher Unterschied zur TfCa war, dass mit 5 und
10 µg/ml auch EBT als Hydrolyse-Produkt freigesetzt wurde, welches bei höheren
Enzymkonzentrationen direkt weiter hydrolysiert wurde (Abbildung 89 und 91). Des Weiteren konnte
für die Cutinase nachgewiesen werden, dass mit zunehmender Enzymkonzentration die Freisetzung
an TPA steigt. Die maximal freigesetzte Menge an Hydrolyse-Produkten mit 49,4 mM/g Substrat
wurde mit 250 µg/ml Enzym und 4 Stunden Inkubationszeit detektiert. Wie bei TfCa war auch bei

Ergebnisse und Diskussion 161
TfCut2 MHET das Haupthydrolyseprodukt bei allen Enzymkonzentrationen. Es erfolgte eine Zunahme
der MHET Menge bei den Enzymkonzentrationen von 5-50 µg/ml und eine gleichbleibend
freigesetzte Menge an MHET bei den höheren Enzymkonzentrationen. Mit steigender
Enzymkonzentration bis 50 µg/ml stieg die freigesetzte Menge an EMT an und sank bei höheren
Enzymkonzentrationen ab. Hingegen stieg die freigesetzte Menge an TPA und BHET mit
zunehmender Enzymkonzentration.
Abbildung 90: Effekt der Reaktionszeit auf die Hydrolyse von PET-Trimeren mit rTfCa.
Abbildung 91: Effekt der Reaktionszeit auf die Hydrolyse von PET-Trimeren mit rTfCut2.
Mit beiden Enzymen, der Carboxylesterase TfCa und der Cutinase TfCut2, konnte bei höheren
Enzymkonzentrationen (≥ 50 µg/ml) keine weitere Freisetzung der Hydrolyseprodukte detektiert
werden. Es wurde jedoch eine Verschiebung der Produktmengen zueinander festgestellt. Bei beiden
Enzymen stieg die Freisetzung an TPA mit steigender Proteinkonzentration, allerdings konnte kein

Ergebnisse und Diskussion 162
Umsatz mit 100% TPA als Produkt erreicht werden, wie es Hooker et al. 2003 beschrieben hat. Dies
könnte an der unterschiedlichen Herstellung der CTR-Suspension liegen. Bei Hooker et al. wurden die
CTR zunächst in Dioxan gelöst und anschließend eine Suspension mit Puffer hergestellt, dagegen
wurden die zyklischen Trimere für die Hydrolyse durch die PET-Hydrolasen direkt in die Pufferlösung
gegeben. Durch die CTR-Stammlösung in Dioxan entstanden beim Verdünnen mit Puffer eine
Suspension mit sehr kleinen Partikeln, was eine große Oberfläche der CTR zur Folge hatte, die für die
Hydrolyse durch die Cutinase von Vorteil war (Figueroa et al. 2006). Des Weiteren war die
eingesetzte CTR Konzentration und die Enzymkonzentration bei Hooker geringer (0,74 mM CTR,
Enzymkonzentration 75-200 LU/ml) im Vergleich zur Trimer Hydrolyse (52,1 mM CTR,
Enzymkonzentration rTfCa 4500-226800 p-NPB-U/ml bzw. rTfCut2 650-32400 p-NPB-U/ml). Sie
wiesen nach, dass nach vollständiger Hydrolyse der CTR das vorhandene MHET weiter zu TPA
hydrolysiert wurde und somit eine vollständige Hydrolyse der CTR zu TPA erfolgte (Hooker et al.
2003). Die PET-Trimere lagen für die enzymatische Hydrolyse hingegen im Überschuss vor. Somit
erfolgte zunächst die Spaltung der Trimere zu Dimeren, jedoch nicht die vollständige Hydrolyse zu
TPA, solang noch genügend Substrat (CTR) zur Verfügung stand.
In den erfolgten Studien konnte gezeigt werden, dass T. fusca neben den beiden PET-
hydrolysierenden Cutinasen Tfu_0882 und Tfu_0883 (BTA1 und BTA2, TfCut1 und TfCut2, Cutinase 1
und 2) auch eine PET-hydrolysierende Carboxylesterase bildet. Somit wurde die Behauptung von Hu
et al. 2010, dass T. fusca nur zwei Enzyme produziert, die synthetische Polyester abbauen, widerlegt.
Da die PET-hydrolysierenden Enzyme verschiedenen EC-Klassen (EC 3.1.1.1 und EC 3.1.1.3)
zugeordnet sind, unterstützt die Entdeckung die Auffassung, dass aufgrund der EC-Klasse kein
Aufschluss über die hydrolytischen Eigenschaften von Enzymen möglich ist (Sinsereekul et al. 2010,
Eberl et al. 2008). Somit ist die Einordnung der Enzyme in Klassen aufgrund ihrer allgemeinen
hydrolytischen Eigenschaften gegenüber einem Substrat nicht aussagekräftig, da meist mehr als ein
Substrat durch das Enzym umgesetzt wird. Vernünftiger wäre es, die Zuordnung der Enzyme
aufgrund struktureller Eigenschaften vorzunehmen, wie es bereits für die lipolytischen Enzyme durch
Hausmann und Jaeger (2010) erfolgte.
In den vergangenen Jahren erfolgten einige Studien zur näheren Untersuchung der Mechanismen bei
der Hydrolyse von PET durch Hydrolasen (Eberl et al. 2008, Sinsereekul et al 2010, Hu et al. 2010,
Donelli et al. 2010). Es ist noch nicht eindeutig geklärt, ob es Unterschiede bezüglich der Hydrolyse
zwischen den Enzymgruppen der Cutinasen, Lipasen oder Carboxylesterasen gibt oder ob die
Hydrolyse bei allen ähnlich abläuft. Bei den in dieser Arbeit durchgeführten Studien baute die TfCa
bevorzugt kleine PET Substrate, in Form von CTR, ab. Die Cutinasen, besonders die Cutinasen aus
T. fusca, wiesen gegenüber allen PET Substraten eine hohe Aktivität auf. Diese Hydrolyse-
eigenschaften sind von den beiden Enzymgruppen bereits gegenüber den p-NP Estern bekannt.

Ergebnisse und Diskussion 163
Grund dafür ist die Konformation bzw. Struktur des aktiven Zentrums der Enzyme. Bei der Cutinase
liegt das aktive Zentrum an der Proteinoberfläche und ist leicht zugänglich für kurz- und langkettige
Substrate. Das aktive Zentrum der Carboxylesterase liegt in einer Bindungstasche bzw. Spalte und ist
zwar zugänglich für kurzkettige Substrate, aber nicht für langkettige. Diese Beobachtungen
bestätigen auch die SEM Aufnahmen der APET-Filme nach der hydrolytischen Behandlung mit den
beiden Enzymen. Der APET-Film, inkubiert mit TfCa, wies nur kleine, lochförmige
Oberflächenveränderungen auf. Nach der Inkubation des APET-Films mit der Cutinase wies dieser
großflächige Abbauspuren auf.
Die Temperaturabhängigkeit des PET Abbaus konnte ebenfalls für langkettige Substrate bestätigt
werden (APET-Film und Fasern). Die Inkubationstemperatur hatte einen Einfluss auf den Abbau des
APET-Films, da aufgrund der höheren Temperatur die Polymerketten mobiler waren und besser mit
dem aktiven Zentrum wechselwirken konnten. Die eukaryotischen Cutinasen aus F. solani pisi,
inkubiert bei 30°C, zeigten eine geringere hydrolytische Aktivität gegenüber dem APET-Film und PET-
Fasern als die Cutinasen aus T. fusca, inkubiert bei 60°C. Bei den kurzkettigen Substraten wie den
PET-Trimeren hatte die Inkubationstemperatur einen geringeren Einfluss auf die Hydrolyse. Bei der
Hydrolyse der CTR ist die für die Hydrolyse zur Verfügung stehende Oberfläche wichtiger.
Zusammenfassend können für den durchgeführten PET Abbau folgende Schlussfolgerungen gezogen
werden. Langkettige PET Substrate werden mit höherer Effizienz bei hohen Inkubationstemperaturen
und geringer Kristallinität und bevorzugt von Cutinasen abgebaut. Bei kurzkettigen Substraten ist für
den hydrolytischen Abbau die Partikelgröße des Substrats von Bedeutung. Die Inkubations-
temperatur ist bei den kurzkettigen Substraten nur abhängig vom hydrolysierenden Enzym. Ein
Abbau der kurzkettigen PET-Substrate kann sowohl durch die Carboxylesterase, als auch durch die
Cutinasen erfolgen.
Ausblickend muss für weitere Untersuchungen der Kinetik der PET Hydrolysen eine PET-spezifische
Aktivitätsbestimmung mit einer schnellen und einfachen Auswertung ausgearbeitet werden, um die
verschiedenen PET-Hydrolasen zu testen. Für die PET-spezifische Aktivitätsbestimmung wird ein
kurzkettiges PET-ähnliches Substrat benötigt, bei dem ein genauer Umsatz bestimmt werden kann,
dass heißt, wenn 1 mol Produkt detektiert wird, muss daraus folgen, dass 1 mol Substrat verbraucht
bzw. umgesetzt wurde. Da die CTR aus mehreren Molekülen TPA bestehen, ist die direkte Zuordnung
der gebildeten Hydrolyseprodukte so nicht möglich. Die Bedingungen für die Anwendung eines
Substrats für die Aktivitätsbestimmung ist entweder
a) es wird während der Hydrolyse nur ein Molekül TPA pro einem Molekül Substrat freigesetzt
(mögliches Substrat BETEB bzw. 3PET)
oder wenn mehrere Moleküle TPA vorhanden sind

Ergebnisse und Diskussion 164
b) ein Molekül TPA pro einem Molekül Substrat muss gesondert markiert sein und somit wird
nur dieses markierte Molekül mit einer geeigneten Methode detektiert (beispielsweise
Isotopenmarkiert (Deuterium), Fluoreszenzmarker oder ähnliches).
Eine andere Möglichkeit der Analytik wäre der Nachweis eines komplexierten Substanzmoleküls nach
der Hydrolyse des CTR-Rings. Durch die Komplexierung dieser Substanz im Inneren des CTR-Rings ist
sie nicht nachweisbar. Erst bei der Hydrolyse wird sie freigesetzt und kann mit einer spezifischen
Nachweismethode detektiert werden. Für diese Methode wird allerdings ein deutlich höherer
Arbeitsaufwand benötigt, da bisher in der Literatur keine Substanz bekannt ist, die mit CTR
komplexiert.

Zusammenfassung 165
4 Zusammenfassung
Der Actinomycet T. fusca KW3, isoliert aus Kompost, bildete während der Kultivierung im
Mineralsalz-Spurenelement-Vitamin-Minimalmedium nach Zusatz von PET-Fasern eine 52 kDa
Carboxylesterase (TfCa), welche effizient zyklische PET-Trimere (CTR) hydrolysiert. Die TfCa besitzt
einen pI von 4.8, eine Substratspezifität gegenüber kurzkettigen p-Nitrophenyl-Estern und wird durch
Phenylmethylsulfonylfluorid (PMSF) und Tosyl-L-Phenylalanin-Chloromethylketon (TPCK) in der
Aktivität gehemmt. Die Carboxylesterase hydrolysiert kein Cutin oder Poly-ε-caprolacton (PCL). CTR
hingegen wurden durch die TfCa mit einem Km von 0,5 mM und einer Vmax von 9,3 µmol/min/mg bei
optimalen Bedingungen (60°C, pH 6) hydrolysiert. Das aktive Zentrum der Carboxylesterase besteht
aus den Aminosäuren Ser185, Glu319 und His415, wobei das Serin in das katalytische Motiv G-E-S-A-
G eingebettet ist.
Während der Reaktion setzte die TfCa auch Hydrolyseprodukte aus PET-Fasern und -Filmen frei. Der
Nachweis der Hydrolyse erfolgte durch Umkehrphasen-Hochleistungsflüssigkeitschromatographie
der Abbauprodukte und bei den PET-Filmen zusätzlich mittels Rasterelektronenmikroskopie. Dabei
zeigte die Carboxylesterase verglichen mit anderen PET-Hydrolasen eine geringere Effizienz, was
durch die Lage des aktiven Zentrums in einer Bindungstasche und der daraus folgenden schlechten
Zugänglichkeit für polymere Substrate begründet werden kann. Bei der Hydrolyse der viel kleineren
CTR war die TfCa deutlich effektiver, was auf eine höhere Spezifität gegenüber kurzkettigen PET
Substraten hinweist.
Beim Abbau von PET Substraten wurde das Produktspektrum der TfCa und anderer PET-Hydrolasen
miteinander verglichen. Dabei zeigten die Hydrolasen ähnliche Abbaumechanismen. Das
Haupthydrolyseprodukt war, außer bei der TfCa aus T. fusca KW3 und der Cutinase 2 aus T. fusca
WSH03-11, unabhängig vom Substrat, MHET, der Monoester der Terephthalsäure. TfCa aus T. fusca
KW3 und die Cutinase 2 aus T. fusca WSH03-11 produzierten als Haupthydrolyseprodukt
Terephthalsäure. Zu Beginn der Hydrolyse wurden bevorzugt die Dimere EMT oder EBT freigesetzt,
welche nach längeren Reaktionszeiten weiter hydrolysiert wurden. Aufgrund der ähnlichen aktiven
Zentren an der Oberfläche der Proteine ist auch ein Abbau von PET-Fasern und Filmen durch diese
Enzyme möglich.

Literaturverzeichnis 166
5 Literaturverzeichnis
Abo M, Fukuyama S, Svendsen A und Matsui T. Cutinase Variants. (2004) US Patent 6,815,190 B1.
Alisch M, Feuerhack A, Müller H, Mensak J, Andreaus J und Zimmermann W. Biocatalytic
modification of polyethylene terephthalate fibres by esterases from actinomycete isolates.
(2004) Biocatalysis and Biotransformation. 22, 347-351.
Alisch-Mark M, Herrmann A und Zimmermann W. Increase of the hydrophilicity of polyethylene
fibres by hydrolases from Thermomonospora fusca and Fusarium solani f. sp. pisi. (2006)
Biotechnology Letters. 28, 681-685.
Almansa E, Heumann S, Eberl A, Fischer-Colbrie G, Martinkova L, Marek J, Cavaco-Paulo A und
Guebitz GM. Enzymatic surface hydrolysis of PET enhances in PVC coating. (2008) Biocatalysis
and Biotransformation. 26, 365-370.
Andersen B, Borch K, Abo M und Damgaard B. A method of treating polyester fabrics. (January
1999) PCT Patent WO 99/01604.
Andre C und Charmoille L. Fusarium isolate and lipases, cutinases and enzyme compositions derived
therefrom. (1999) US Patent 5,990,069.
Araújo R, Casal M und Cavaco-Paulo A. Application of enzymes for textile fibres processing. (2008)
Biocatalysis and Biotransformation. 26, 332-349.
Araújo R, Silva C, O´Neill A, Micaelo N, Guebitz GM, Soares C, Casal M und Cavaco-Paulo A.
Tailoring cutinase activity towards polyethylene terephthalate and polyamide 6,6 fibers. (2007)
Journal of Biotechnology. 128, 849-857.
Baatout S, De Boever P und Mergeay M. Physiological changes induced in four bacterial strains
following oxidative stress. (2006) Prikladnaia Biohimiia I Mikrobiologiia. 42, 418-427.
Bachmann SL und McCarthy AJ. Purification and Cooperative Activity of Enzymes Constituting the
Xylan-Degrading System of Thermomonospora Fusca. (1991) Applied and Environmental
Microbiology. 57, 2121-2130.
Baker RL. How to minimize cyclic trimer when pressure dyeing polyester yarn. (1974) American
Dyestuff Reporter. 63, 34-38.
Beatty AL, Malloy JL und Wright JR. Pseudomonas aeruginosa degrades pulmonary surfactant and
increases conversion in vitro. (2005) American Journal of Respiratory cell and Molecular
Biology. 32, 128-134.
Bennett-Lovsey RM, Herbert AD, Sternberg MJ und Kelley LA. Exploring the extremes of
sequence/structure space with ensemble fold recognition in the program Phyre. (2008)
Proteins. 70, 611-625.

Literaturverzeichnis 167
Benίtez JJ, Garcίa-Segura R und Heredia A. Plant biopolyester cutin: a tough way to its chemical
synthesis. (2004) Biochimica et Biophysica Acta. 1674, 1-3.
Berger R, Hoffmann M und Keller U. Molecular nalysis of a gene encoding a cell-bound esterase from
Streptomyces chrysomallus. (1998) Journal of Bacteriology. 180, 6396-6399.
Bernards MA, Lopez ML, Zajicek J und Lewis NG. Hydroxycinnamic acid-derived polymers constitute
the polyaromatic domain of suberin. (1995) Journal of Biological Chemistry. 270, 7382-7386.
Bernards MA. Demystifying Suberin. (2002) Canadian Journal of Botany. 80, 227-240.
Billig S, Oeser T, Birkemeyer C und Zimmermann W. Hydrolysis of cyclic poly(ethylene
terephthalate) trimers by a carboxylesterase from Thermobifida fusca KW3. (2010) Applied
Microbiology and Biotechnology. 87, 1753-1764.
Boston M, Requadt C, Danko S, Jarnagin A, Ashizawa E, Wu S, Polouse AJ und Bott R. Structure and
function of engineered Pseudomonas mendocina lipase. (1997) Methods in Enzymology 284,
298-317.
Bott R, Kellis JT und Morrison TB. High throughput mutagenesis screening method. (2003) WO
Patent 03/076580 A2.
Breuil C, Shindler DB, Sijher JS und Kushner DJ. Stimulation of lipase production during bacterial
growth on alkanes. (1978) Journal of Bacteriology. 133, 601-606.
Brueckner T, Eberl A, Heumann S, Rabe M und Guebitz GM. Enzymatic and chemical hydrolysis of
poly(ethylene terephthalate) fabrics. (2008) Journal of Polymer Science, Part A: Polymer
Chemistry. 46, 6435-6443.
Burkhart CN und Burkhart CG. Antibiotic-resistant Propionibacteria acnes may not be the major
issue clinically or microbiologically in acne. (2003) British Journal of Dermatology. 148, 365-
366.
Busch JE und Stutzenberger FJ. Amylolytic activity of Thermomonospora fusca. (1997) World Journal
of Microbiology & Biotechnology. 13, 637-642.
Buttke TM und Cuchens MA. Inhibition of lymphocyte proliferation by free fatty acids. II. Toxicity of
stearic acid towards phytohaemagglutinin-acivated T cells. (1984) Immunology. 53, 507-514.
Buttke TM. Inhibition of lymphocyte proliferation by free fatty acids. I. Differential effects on mouse
B and T lymphocytes. (1984) Immunology. 53, 235-242.
Camacho LR, Ensergueix D, Perez E, Gicquel R und Guilhot C. Identification of a virulence gene
cluster of Mycobacterium tuberculosis by signature-tagged transposon mutagenesis. (1999)
Molecular Microbiology. 44, 257-267.
Carvalho CM, Aires-Barros MR und Cabral JMS. Cutinase structure, function and biocatalytic
applications. (1998) Electronical Journal of Biotechnology 1, 160-173.

Literaturverzeichnis 168
Carvalho CM, Aires-Barros MR und Cabral JMS. Cutinase: From molecular level to bioprocess
development. (1999) Biotechnology and Bioengineering 66, 17-34.
Casimir-Schenkel JJ. Thermostabile Xylanasen von Thermomonospora fusca KW3: Produktion,
Isolierung, Charakterisierung und Anwendung zur enzymatischen Zellstoffbleichung. (1992)
Dissertation ETH Nr. 9845.
Chemical Market Asscociates Inc.: 2010 world terephthalate and polyester analysis.
http://www.cmaiglobal.com/WorldAnalysis/xylbook.aspx. Accessed 31.03.2010.
Chen J, Wu J und Chen S. High-temperature cutinase and gene order thereof. (2008) CN Patent
200810020124.0.
Chen S und Wilson DB. Proteomic and Transcriptomic Analysis of Extracellular Proteins and mRNA
Levels in Thermobifida fusca grown on Cellobiose and Glucose. (2007) Journal of Bacteriology.
189, 6260-6265.
Chen S, Su L, Billig S, Zimmermann W, Chen J und Wu J. Biochemical characterization of the
cutinases from Thermobifida fusca. (2010) Journal of Molecular Catalysis B: Enzymatic. 63,
121-127.
Chen S, Tong X, Woodard RW, Du G, Wu J und Chen J. Identification and Characterization of
Bacterial Cutinase. (2008) The Journal of Biological Chemistry. 283, 25854-25862.
Deckwer W-D, Mueller R-J, Kleeberg I und van den Heuvel J. Enzyme which cleaves ester groups and
which is derived from Thermomonospora fusca. (2006) US 6,995,005.
Degassi G, Devescovi G, Kim J, Hwang I und Venturi V. Identification, characterization and regulation
of two secreted polygalacturonases of the emerging rice pathogen Burkholderia glumae.
(2008) FEMS Microbiology and Ecology. 65, 251-262.
DeLano WL. The PyMOL molecular graphics system. (2002) DeLano Scientific, Palo Alto.
Devescovi G, Bigirimana J, Degrassi G, Cabrio L, LiPuma JJ, Kim J, Hwang I und Venturi V.
Involvment of a quorum-sensing-regulated lipase secreted by a clinical isolate of
Burkholderia glumae in severe disease symptoms in rice. (2007) 73, 4950-4958.
Donaghy JA, Bronnenmeier K, Soto-Kelly PF und McKay AM. Purification and characterization of an
extracellular feruloyl ersterase from the thermophilic anaerobe Clostridium stercorarium.
(2000) Journal of Applied Microbiology. 88, 458-466.
Donelli I, Freddi G, Nierstrasz VA und Taddei P. Surface structure and properties of poly-(ethylene
terephthalate) hydrolyzed by alkali and cutinase. (2010) Polymer Degradation and Stability. 95,
1542-1550.
Donelli I, Taddei P, Smet PF, Poelman D, Nierstrasz VA und Freddi G. Enzymatic surface modification
and functionalization of PET: a water contact angle, FTIR, and fluorescence spectroscopy study.
(2009) Biotechnology and Bioengineering. 103, 845-856.

Literaturverzeichnis 169
Dresler K, van den Heuvel J, Müller R-J und Deckwer D-W. Production of a recombinant polyester-
cleaving hydrolase from Thermobifida fusca in Escherichia coli. (2006) Bioprocess and
Biosystems Engineering. 29, 169-183.
Droege MJ, Bos R, Boersma YL und Quax WJ. Comparison and functional characterisation of three
homologous intracellular carboxylesterase of Bacillus subtilis. (2005) Journal of Molecular
Catalysis B: Enzymatic. 32, 261-270.
Du G, Zhang S, Hua Z, Zhu Y und Chen J. Enhanced cutinase production with Thermobifida fusca by
two-stage pH control strategy. (2007) Biotechnology Journal. 2, 365-369.
Dugal S, Jansen K und Stein G. Zum Verhalten der Oligomer beim Färben von Polyester. (1973)
Melliand Textilberichte. 54, 863.
Dutta K, Sen S und Veeranik VD. Production, characterization and applications of microbial
cutinases. (2009) Process Biocemistry. 44, 127-134.
Dyson W, Kellis JT, Poulose AJ und Yoon M-Y. Enzymes useful for changing the properties of
polyester. (2005) US 6,933,140 B1.
Eberl A, Heumann S, Brückner T, Araújo R, Cavaco-Paulo A, Kaufmann F, Kroutil W und Guebitz
GM. Enzymatic surface hydrolysis of poly(ethylene terephthalate) and bis(benzoyloxyethyl)
terephthalate by lipase and cutinase in the presence of surface active molecules. (2009)
Journal of Biotechnology. 143, 207-212.
Eberl A, Heumann S, Kotek R, Kaufmann F, Mitsche S, Cavaco-Paulo A und Gübitz GM. Enzymatic
hydrolysis of PTT polymers and oligomers. (2008) Journal of Biotechnology. 135, 45-51.
Eisenberg D, Lüthy R und Bowie JU. VERIFY3D: assessment of protein models with three-dimensional
profiles. (1997) Methods in Enzymology. 277, 396-404.
El-Bendary MA, El-Ola SMA und Moharam ME. Enzymatic Surface Hydrolysis of
Poly(ethyleneterephthalate) by Lipase Enzyme and its Production. (2010) Canadian Journal of
Pure and Applied Sciences. 4, 1207-1216.
Elend C, Schmeisser C, Leggewie C, Babiak P, Carballeira JD, Steele HL, Reymond JL, Jaeger KE und
Streit WR. Isolation and biochemical characterization of two novel metagenome-derived
esterases. (2006) Applied and Environmental Microbiology. 72, 3637-3645.
Embley TM und Stackebrandt E. The Molecular Phylogeny and Systematics of the Actinomycetes.
(1994) Annual Reviews of Microbiology. 48, 257-289.
Ernst RK, Yi EC, Guo L, Lim KB, Burns JL, Hackett M und Miller SI. Specific lipopolysaccharide found
in cystic fibrosis airway Pseudomonas aeruginosa. (1999) Science. 286, 1561-1565.
Fernandez-Lafuente R. Lipase from Thermomyces lanuginosus. Uses and prospects as an industrial
biocatalyst. (2010) Journal of Molecular Catalysis B: Enzymatic. 62, 197-212.

Literaturverzeichnis 170
Fernando G, Zimmermann W und Kolattukudy PE. Suberin-grown Fusarium solani f. sp. pisi
generates a cutinase like esterase which depolymerizes the aliphatic components of suberin.
(1984) Physiological Plant Pathology. 24, 143-155.
Fett WF, Gérard H, Moreau RA, Osman SF und Jones LE. Cutinase production by Streptomyces spp.
(1992) Current Microbiology 25, 165-171.
Fett WF, Wijey C, Moreau RA und Osman SF. Production of cutinase by Thermomonospora fusca
ATCC 27730. (1999) Journal of Applied Microbiology. 86, 561–568.
Feuerhack A, Alisch-Mark M, Kisner A, Pezzin SH, Zimmermann W und Andreaus J. Biocatalytic
surface modification of knitted fabrics made of poly (ethylene terephthalate) with hydrolytic
enzymes from Thermobifida fusca KW3b. (2008) Biocatalysis and Biotransformation. 26, 357-
364.
Figueroa Y, Hinks D und Montero G. A heterogeneous kinetic model for the cutinase-catalyzed
hydrolysis of cyclo-tris-ethylene terephthalate. (2006) Biotechnology Progress. 22, 1209-1214.
Fischer-Colbrie G, Heumann S, Liebminger S, Almansa E, Cavaco-Paulo A und Guebitz GM. New
enzymes with potential for PET surface modification. (2004) Biocatalysis and
Biotransformation. 22, 341-346.
Franke R und Schreiber L. Suberin – a biopolyester forming apoplastic plant interfaces. (2007)
Current Opinion in Plant Biology. 10, 252-259.
Garcia-Conesa MT, Kroon PA, Ralph J, Mellon FA, Colquhoun IJ, Saulnier L, Thibault JF und
Williamson G. A cinnamoyl esterase from Aspergillus niger can break plant cell wall cross-links
without release of free diferulic acids. (1999) European Journal of Biochemistry. 266, 644-652.
Gerard HC, Osman SF, Fett WF und Moreau RA. Separation, Identification and Quantification of
Monomers from Cutin Polymers by High Performance Liquid Chromatography and Evaporative
Light Scattering Detection. (1992) Phytochemical Analysis. 3, 139-144.
Gouda MK, Kleeberg I, van den Heuvel J, Müller R-J und Deckwer W-D. Production of a polyester
degrading extracellular hydrolase from Thermomonospora fusca. (2002) Biotechnology
Progress. 18, 927-934.
Gray GL, Power SD und Poulose AJ. Lipase from Pseudomonas mendocina having cutinase activity.
(1995) US 5,389,536.
Gribbon EM, Cunliffe WJ und Holland KT. Interaction of Propionibacterium acnes with skin lipids in
vitro. (1993) Journal of General Microbiology. 139, 1745-1751.
Grosse I, Müller U und Schreiber H. Oligomere des Polyethylenterephthalats – Entstehung, Struktur
und Eigenschaften. (1994) Chemiefasern/Textilindustrie. 44, 562-568.
Gübitz GM und Cavaco-Paulo A. New substrates for reliable enzymes: enzymatic modification of
polymers. (2003) Current Opinion in Biotechnology. 14, 577-582.

Literaturverzeichnis 171
Gübitz GM und Cavaco-Paulo A. Biotechnology in the textile industry – perspectives for the new
millennium. (2001) Journal of Biotechnology. 89, 89-90.
Guebitz GM und Cavaco-Paulo A. Enzymes go big: surface hydrolysis and functionalisation of
synthetic polymers. (2008) Trends in Biotechnology. 26, 32-38.
Guo L, Lim KB, Poduje CM, Daniel M, Gunn JS, Hackett M und Miller SI. Lipid A acylation and
bacterial resistance against vertebrate antimicrobial peptides. (1998) Cell. 95, 189-198.
Hansmeier N, Chao TC, Daschkey S, Musken M, Kalinowski J, Puhler A und Tauch A. A
comprehensive proteome map of the lipid-requiring nosocomial pathogen
Corynebacterium jeikeium K411. (2007) Proteomics. 7, 1076-1096.
Hausmann S und JaegerK-E. Lipolytic Enzymes from Bacteria. (2010) In: Timmis KN (ed): Handbook of
Hydrocarbon and Lipid Microbiology. Springer Berlin Heidelberg, 1099-1126.
Hawley HP und Gordon GB. The effects of long chain free fatty acids on human neutrophil function
and structure. (1976) Laboratory Investigation. 34, 216-222.
Hempel WH. Oligomere – ein altes und doch immer wieder neues Färbereiproblem. (1979) Wirkerei-
und Strickereitechnik. 29, 487-490.
Henssen A. Beiträge zur Morphologie und Systematik der thermophilen Actionmyceten. (1957)
Archiv für Mikrobiologie. 26, 373-414.
Herzog K, Müller R-J und Deckwer W-D. Mechanism and kinetics of the enzymatic hydrolysis of
polyester nanoparticles by lipases. (2006) Polymer Degradation and Stability. 91, 2486-2498.
Heumann S, Eberl A, Pobeheim H, Liebminger S, Fischer-Colbrie G, Almansa E, Cavaco-Paulo A und
Gübitz GM. New model substrates for enzymes hydrolyzing polyethyleneterephthalate and
polyamide fibres. (2006) Journal of Biochemical and Biophysical Methods. 39, 89-99.
Heurlier K, Williams F, Heeb S, Dormond C, Pessi G, Singer D, Camara M, Williams P und Haas D.
Positive control of swarming, rhamnolipid synthesis, and lipase production by the
posttranscriptional RsmA/RsmZ system in Pseudomonas aeruginosa PAO1. (2004) Journal of
Bacteriology. 186, 2936-2945.
Hilge M, Gloor SM, Rypniewski W, Sauer O, Heightman TD, Zimmermann W, Winterhalter K und
Piontek K. High-resolution native and complex structure of thermostable beta-mannanase
from Thermomonospora fusca – substrate specifity in glycosyl hydrolase family 5. (1998)
Structure. 6, 1433-1444.
Höhn W. Phenomenon, analysis and minimizing of polyester oligomers. (2003) Melliand
International. 9, 257-260.
Holmquist M. Alpha/Beta-Hydrolase Fold Enzymes: Structures, Functions and Mechanisms. (2000)
Current Protein and Peptide Science. 1, 209-235.

Literaturverzeichnis 172
Hooker J, Hinks D, Montero G und Icherenska M. Enzyme-catalyzed hydrolysis of poly(ethylene
terephthalate) cyclic trimer. (2003) Journal of Applied Polymer Science. 89, 2545-2552.
Hsieh Y-L und Cram LA. Enzymatic hydrolysis to improve wetting and absorbency of polyester fabrics.
(1998) Textile Research Journal. 68, 311-319.
Hsieh Y-L, Hartzell MM, Clarkson KA, Collier KD, Graycar TP und Larenas E. Enzyme treatment to
enhance wettability and absorbency of textiles. (September 1997) PCT Patent WO 97/33001.
Hu X, Thumarat U, Zhang X, Tang M und Kawai F. Diversity of polyester-degrading bacteria in
compost and molecular analysis of a thermoactive esterase from Thermobifida alba AHK119.
(2010) Applied Microbiology and Biotechnology. 87, 771-779.
Jaeger K-E, Kharazmi A und Hoiby N. Extracellular lipase of Pseudomonas aeruginosa: biochemical
characterization and effect on human neutrophil and monocyte function in vitro. (1991)
Microbial Pathogen. 10, 173-182.
Jahreiß K. Synthesefasern im Alltag. (2005) http://daten.didaktikchemie.uni-
bayreuth.de/umat/fasern_syn/synthesefasern.htm
Jump J und McCloskey S. A method of treating polyester fabrics. (2004) WO 2004/059075 A1.
Keller KH, Furer L und Wenger K. Einfluss von Färbereihilfsprodukten und Verfahren auf Oligomere
beim HT-Färben von Polyester. (1981) Textilveredelung. 16, 72-76.
Kellis J, Poulose AJ and Yoon MY. Enzymatic modification of the surface of a polyester fiber or
article. (2001) US 6,254,645 B1.
Khalameyzer V, Fischer I, Bornscheuer UT und Altenbuchner J. Screening, nucleotide sequence, and
biochemical characterization of an esterase from Pseudomonas fluorescens with high activity
towards lactones. (1999) Applied and Environmental Microbiology. 65, 477-482.
Kim HR und Song WS. Lipase treatment of polyester fabrics. (2006) Fibers and Polymers. 7, 339-243.
Kim HR und Song WS. Optimization of enzymatic treatment of polyester fabrics by lipase from
Porcine Pancreas. (2008) Fibers and Polymers. 9, 423-430.
Kint D und Munoz-Guerra M. A review on the potential biodegradability of poly(ethylene
terephthalate). (1999) Polymer International 48, 364-352.
Kleber HP, Schlee D und Schöpp W. Biochemisches Praktikum. Methoden für Studium, Praxis,
Forschung (1997) Gustav Fischer Verlag, Jena.
Kleeberg I, Hetz C, Kroppenstedt RM, Müller R-J und Deckwer W-D. Biodegradation of aliphatic-
aromatic copolyesters by Thermomonospora fusca and other thermophilic compost isolates.
(1998) Applied Environmental Microbiology. 64, 1731-1735.
Kleeberg I, Welzel K, van den Heuvel J, Müller R-J und Deckwer W-D. Characterization of a new
extracellular hydrolase from Thermobifida fusca degrading aliphatic-aromatic copolyesters.
(2005) Biomacromolecules. 6, 262-270.

Literaturverzeichnis 173
Kolattukudy PE. Cutin from Plants. (2002a) Biopolymers, Polyesters I. Wiley-VCH. 1-40.
Kolattukudy PE. Suberin from Plants. (2002b) Biopolymers, Polyesters I. Wiley-VCH. 41-73.
Kontkanen H, Saloheimo M, Pere J, Miettinen-Oinonen A und Reinikainen T. Characterization of
Melanocarpus albomyces steryl esterase produced in Trichoderma reesei and modification of
fibre products with the enzyme. (2006) Applied Microbiology and Biotechnology. 72, 696-704.
Kusch P. Zum Oligomeraustritt aus Polyesterfasern. (1974) Textilveredelung. 9, 597-599.
Kusters JG, van Vliet AH und Kuipers EJ. Pathogenesis of Helicobacter pylori infection. (2006) Clinical
Microbiology Review. 19, 449-490.
Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4.
(1970) Nature. 227, 680–685.
Levefre C, Mathieu C, Tidjani A, Dupret A, van der Wauven C, De Winter W und David C.
Comparative degradation by microorganisms of terephthalic acid, 2,6-naphthalene
dicarboxylic acid, their esters and polyesters. (1999) Polymer Degradation and Stability. 64, 9-
16.
Lewenza S, Conway B, Greenberg EP und Sokol PA. Quorum sensing in Burkholderia cepacia:
identification oft he LuxRI homologs CepRI. (1999) Journal of Bacteriology. 181, 748-756.
Li Y und Beisson F. The biosynthesis of cutin and suberin as an alternative source of enzymes for the
production of bio-based chemicals and materials. (2009) Biochimie. 91, 685-691.
Liebminger S, Eberl A, Sousa F, Heumann S, Fischer-Colbrie G, Cavaco-Paulo A und Guebitz GM.
Hydrolysis of PET and bis-(benzoyloxyethyl) terephthalate with a new polyesterase from
Penicillium citrinum. (2007) Biocatalysis and Biotransformation. 25, 171-177.
Lin TS und Kolattukudy PE. Induction of a Biopolyester Hydrolase (Cutinase) by Low Levels of Cutin
Monomers in Fusarium solani f. sp. pisi. (1978) Journal of Bacteriology. 133, 942-951.
Lin TS und Kolattukudy PE. Isolation and characterization of a cuticular polyester (cutin) hydrolysing
enzyme from phytopathogenic fungi. (1980) Physiological Plant Pathology. 17, 1-15.
Lin TS und Kolattukudy PE. Structural Studies on Cutinase, a Glycoprotein Containing Novel Amino
Acids and Glucuronic Acid Amide at the N Terminus. (1980) European Journal of Biochemistry.
106, 341-351.
Liu P, Ewis HE, Tai PC, Lu CD und Weber IT. Crystal structure of the Geobacillus stearothermophilus
carboxylesterase Est55 and its activation of prodrug CPT-11. (2007) Journal of Molecular
Biology. 367, 212-223.
Liu Y, Wu G und Gu L. Enzymatic Treatment of PET Fabrics for Improved Hydrophilicity. (2008)
AATCC-Review. 8, 44-48.
Liu Z, Gosser Y, Baker PJ, Ravee Y, Lu Z, Alemu G, Li H, Butterfoss GL, Kong XP, Gross R und
Montclare JK. Structural and functional studies of Aspergillus oryzae cutinase: enhanced

Literaturverzeichnis 174
thermostability and hydrolytic activity of synthetic ester and polyester degradation. (2009)
Journal of the American Chemical Society. 131, 15711-15716.
Longhi S, Mannesse M, Verheij HM, Haas GH, Egmond M, Knoops-Mouthuy E und Cambillau C.
Crystal structure of cutinase covalently inhibited by a triglyceride analogue. (1997) Protein
Science. 6, 275-286.
Lykidis A, Mavromatis K, Ivanova N, Anderson I, Land M, DiBartolo G, Martinez M, Lapidus A, Lucas
S, Copeland A, Richardson P, Wilson DB und Kyrpides N. Genome Sequence and Analysis of
the Soil Cellulolytic Actinomycete Thermobifida fusca YX. (2007) Journal of Bacteriology. 189,
2477–2486.
Madigan MT und Martinko JM. Brock Mikrobiologie. Pearson Education, 2009.
Margesin R, Gander S, Zacke G, Gounot AM und Schinner F. Hydrocarbon degradation and enzyme
activities of cold-adapted bacteria and yeasts. (2003) Extremophiles. 7, 451-458.
Marten E, Müller RJ und Deckwer WD. Studies on the enzymatic hydrolysis of polyesters. I. Low
molecular mass model esters and aliphatic polyesters. (2003) Polymer Degradation and
Stability. 80, 485–501.
Marten E, Müller RJ und Deckwer WD. Studies on the enzymatic hydrolysis of polyesters: II.
Aliphatic-aromatic copolyesters. (2005) Polymer Degradation and Stability. 88, 371–381.
McCarthy AJ und Cross T. A Taxonomic Study of Thermomonospora and other Monosporic
Actinomycetes. (1984) Journal of General Microbiology. 130, 5-25.
McCarthy AJ. Thermomonospora and related genera. (1989) In: Williams ST, Sharpe ME, Holt JG (ed)
Bergey's Manual of Systematic Bacteriology, vol. 4, 2552-2572. Williams & Wilkins, Baltimore.
Mueller R-J. Biological degradation of synthetic polyesters - Enzymes as potential catalysts for
polyester recycling. (2006) Process Biochemistry. 41, 2124-2128.
Müller R-J, Kleeberg I und Deckwer W-D. Biodegradation of polyesters containing aromatic
constituents. (2001) Journal of Biotechnology. 86, 87-95.
Müller R-J, Schrader H, Profe J, Dresler K und Deckwer W-D. Enzymatic degradation of poly(ethylene
terephthalate): rapid hydrolyse using a hydrolase from T. fusca. (2005) Macromolecular Rapid
Communications. 26, 1400-1405.
Nawrath C. The Biopolymers Cutin and Suberin. (2002) The Arabidopsis Book.
Nechwatal A, Blokesch A, Nicolai M, Krieg M, Kolbe A, Wolf M und Gerhardt M. A contribution to
the investigation of enzyme-catalysed hydrolysis of poly(ethylene terephthalate) oligomers.
(2006) Macromolecular Materials and Engineering. 291, 1486-1494.
Nielsen PH, Roslev P, Dueholm TE und Nielsen JL. Microthrix parvicella, a specialized lipid consumer
in anaerobic-aerobic activated sludge plants. (2002) Water Science and Technology. 46, 73-80.

Literaturverzeichnis 175
Nimchua T, Eveleigh DE, Sangwatanaroj U und Punnapayak H. Screening of tropical fungi producing
polyethylene terephthalate-hydrolyzing enzyme for fabric modification. (2008) Journal of
Industrial Microbiology & Biotechnology. 35, 843–850.
Nimchua T, Punnapayak H und Zimmermann W. Comparison of the hydrolysis of polyethylene
terephthalate fibres by a hydrolase from Fusarium oxysporum LCH I and
Fusarium solani f. sp. pisi. (2007) Biotechnology Journal. 1, 1-4.
Nordstrom T, Lindqvist C, Stahls A, Mustelin T und Andersson LC. Inhibition of CD3-induced Ca2+
signals in Jurkat T-cells by myristic acid. (1991) Cell Calcium. 12, 449-455.
O´Neill A und Cavaco-Paulo A. Monitoring biotransformations in polyesters. (2004) Biocatalysis and
Biotransformation. 22, 353-356.
O´Neill A, Araújo R, Casal M, Guebitz GM und Cavaco-Paulo A. Effect of the agitation on the
adsorption and hydrolytic efficiency of cutinases on polyethylene terephthalate fibres. (2007)
Enzyme and Microbial Technology. 40, 1801-1805.
Oda K, Izumi O, Kimura Y und Ohmihachiman S. Verfahren zum Zersetzen von aromatischen
Gruppen enthaltenden Polyestern, Verfahren zur Denier-Verringerung von Fasern und
Mikroorganismen, die eine Aktivität zum Zersetzen derartiger Polyester aufweisen. (1999)
Patent DE 199 35 156 A 1.
Oeser T, Wei R, Baumgarten T, Billig S, Föllner C und Zimmermann W. High level expression of a
hydrophobic poly(ethylene terephthalate)-hydrolyzing carboxylesterase from
Thermobifida fusca KW3 in Escherichia coli BL21(DE3). (2010) Journal of Biotechnology. 146,
100-104.
Ollis DL, Cheah E, Cygler M, Dijkstra B, Frolow F, Franken SM, Harel M, Remington SJ, Silman I,
Schrag J, Sussman JL, Verschueren KHG und Goldman A. The α/ß hydrolase fold. (1992)
Protein Engineering. 5, 197-211.
Olukoshi ER und Packter NM. Importance of stored triacylglycerols in Streptomyces: possible carbon
source for antibiotics. (1994) Microbiology 140, 931-943.
Pollard M, Beisson F, Li Y, und Ohlogge JB. Building lipid barriers: biosynthesis of cutin and suberin.
(2008) Trends in Plant Science. 13, 236-246.
Pourbohloul S, Mallett GS und Buttke TM. Inhibition of lymphocyte proliferation by free fatty acids.
III. Modulation of thymus-dependent immune responses. (1985) Immunology. 56, 659-666.
Prates JA, Tarbouriech N, Charnock SJ, Fontes CM, Ferreira LM und Davies GJ. The structure of the
feruloyl esterase module of xylanase 10B from Clostridium thermocellum provides insights into
substrate recognition. Structure. 9, 1183-1190.

Literaturverzeichnis 176
Purdy RE und Kolattukudy PE. Depolymerization of a Hydroxy Fatty Acid Biopolymer, Cutin, by an
Extracellular Enzyme from Fusarium solani f. pisi: Isolation and Some Properties of the Enzyme.
(1973) Archives of Biochemistry and Biophysics. 159, 61-69.
Purdy RE und Kolattukudy PE. Hydrolysis of Plant Cuticle by Plant Pathogens. Purification, Amino
Acid Composition, and Molecular Weight of Two Isozymes of Cutinase and a Nonspecific
Esterase from Fusarium solani f. pisi. (1975) Biochemistry. 14, 2824-2831.
Purdy RE und Kolattukudy PE. Hydrolysis of Plant Cuticle by Plant Pathogens. Properties of
Cutinase I, Cutinase II, and a Nonspecific Esterase Isolated from Fusarium solani f. pisi. (1975)
Biochemistry. 14, 2832-2840.
Pütz A. Isolierung, Identifizierung und biochemische Charakterisierung Dialkylphthalat spaltender
Esterasen. (2006) Dissertation Universität Düsseldorf.
Riegels M, Koch R, Pedersen LS und Lund H. Enzymatic hydrolysis of cyclic oligomers. (February
2001) US Patent 6,184,010 B1.
Ronkvist ÅM, Xie W, Lu W und Gross RA. Cutinase-catalyzed Hydrolysis of poly(ethylene
terephthalate). (2009) Macromolecules. 42, 5128-5138.
Röthlisberger P, Fiechter A und Zimmermann W. Production of thermostable xylanases in batch and
continous culture by Thermomonospora fusca KW 3. (1992) Applied Microbiology and
Biotechnology. 37, 416-419.
Ruiz C, Falcocchio S, Pastor FI, Saso L und Diaz P. Helicobacter pylori EstV: identification, cloning, and
characterization of the first lipase isolated from an epsilon-proteobacterium. (2007) Applied
and Environmental Microbiology. 73, 2423-2431.
Sandal T, Kauppinen S und Kofod LV. An enzyme with lipolytic activity. (1996) WO Patent 96/13580.
Sato M. Deterioration of filaments and films of polyethyleneterephthalate with enzyme of
Cladosporium cladosporioides FERM J-8. (1983) Sen-I Gakkaishi. 39, T209-T219.
Saurer Management AG. The Fiber Year 2006/07. (2008) The Saurer Group.
http://www.google.de/url?sa=t&source=web&cd=2&ved=0CBsQFjAB&url=http%3A%2F%2Fw
ww.oerlikon.com%2FecomaXL%2Fget_blob.php%3Fname%3DLayout_Fibre_Year_en_08.pdf&r
ct=j&q=Saurer%20Management%20AG.%20%20The%20Fiber%20Year%202006%2F07.%20%2
0(2008)%20The%20Saurer%20Group.&ei=fLc2TJfaDYeVOP7l2OAE&usg=AFQjCNGCfkZVVSIIVBC
30tQfByM3D78KPQ
Schmidt R, Markart P, Ruppert C, Wygrecka M, Kuchenbuch T, Walmrath D, Seeger W und
Guenther A. Time-dependent changes in pulmonary suffactant function and composition in
acute respiratory distress syndrom due to pneumonia or aspiration. (2007) Respiratory
Research. 8, 55.

Literaturverzeichnis 177
Schomburg I, Chang A, Ebeling C, Gremse M, Heldt C, Huhn G und Schomburg D. BRENDA, the
enzyme database: updates and major new developments. (2004) Nucleic Acids Research. 32,
D431-433.
Schwede T, Kopp J, Guex N und Peitsch MC. SWISS-MODEL: an automated protein homology-
modeling server. (2003) Nucleic Acids Research. 31, 3381-3385.
Sebastian J und Kolattukudy PE. Purification and characterization of cutinase from a fluorescent
Pseudomonas putida bacterial strain isolated from phyllosphere. (1988) Archives of
Biochemistry and Biophysics. 263, 77-85.
Sebastian J, Chandra AK und Kolattukudy PE. Discovery of a cutinase-producing Pseudomonas sp.
cohabiting with an apparently nitrogen-fixing Corynebacterium sp. in the phyllosphere. (1987)
Journal of Bacteriology. 169, 131-136.
Senner P. Mit Oligomer leben. (1973) Chemiefasern/Textilindustrie. 344-347.
Shabtai Y. Isolation and characterization of a lipolytic bacterium capable of growing in a low-water-
content oil-water emulsion. (1991) Applied and Environmental Microbiology. 57, 1740-1745.
Shinohara M, Nakajima N und Uehara Y. Purification and characterization of a novel esterase (beta
hydroxypalmitate methyl ester hydrolase) and prevention of the expression of virulence by
Ralstonia solanacearum. (2007) Journal of Applied Microbiology. 103, 152-162.
Shukla SR, Mathur MR und Hedaoo VB. Alkaline weight reduction of polyester fibres. (1997)
American Dyestuff Reporter. Oc, 48-56.
Silva C und Cavaco-Paulo A. Biotransformations in synthetic fibres. (2008) Biocatalysis and
Biotransformation. 26, 350-356.
Silva CM, Carneiro F, O´Neill A, Fonseca LP, Cabral JSM, Gübitz GM und Cavaco-Paulo A. Cutinase -
A new tool for biomodification of synthetic fibres. (2005) Journal of Polymer Science. 43, 2448-
2450.
Sinsereekul N, Wangkam T, Thamchaipenet A, Srikhirin T, Eurwilaichitr L und Champreda V.
Recombinant expression of BTA hydrolase in Streptomyces rimosus and catalytic analysis on
polyesters by surface plasmon resonance. (2010) Applied Microbiology and Biotechnology. 86,
1775–1784.
Slomiany BL, Nishikawa H, Piotrowski J, Okazaki K und Slomany A. Lipolytic activity of
Campylobacter pylori: effect of sofalcone. (1989) Digestion. 43, 33-40.
Soliday CL und Kolattukudy PE. lsolation and Characterization of a Cutinase from
Fusarium roseum culmorum and Its Immunological Comparison with Cutinases from
F. solani pisi. (1976) Archives of Biochemistry and Biophysics. 176, 334-343.
Stutzenberger FJ. Inducible Thermoalkalophilic Polygalacturonate Lyase from
Thermomonospora fusca. (1987) Journal of Bacteriology. 169, 2774-2780.

Literaturverzeichnis 178
Svendsen A, Schroeder Gald SO, Fukuyama S und Matsui T. Cutinase Variants. (2005) US 6,960,459
B2.
Tanamoto K und Azumi S. Salmonella-type heptaacylated lipid A is inactive and acts as an antagonist
of lipopolysaccharide action on human line cells. (2000) Journal of Immunology. 164, 3149-
3156.
Tauch A, Kaiser O, Hain T, Goesmann A, Weisshaar B, Albersmeier A, Bekel T, Bischoff N, Brune I,
Chakraborty T, Kalinowski J, Meyer F, Rupp O, Schneiker S, Viehoever P und Puhler A.
Complete genome sequence and analysis of the multiresistant nosocomial pathogen
Corynebacterium jeikeium K411, a lipid-requiring bacterium of the human skin flora. (2005)
Journal of Bacteriology. 187, 4671-4682.
Tokiwa Y und Suzuki T. Hydrolysis of polyester by lipases. (1977) Nature. 270, 76-78.
Torda AE, Procter JB und Huber T. Wurst: a protein threading server with a structural scoring
function, sequence profiles and optimized substitution matrices. (2004) Nucleic Acids
Research. 32, W532-W535.
Trodler P, Schmid RD und Pleiss J. Modeling of solvent-dependent conformational transitions in
Burkholderia cepacia lipase. (2009) BMC Structural Biology. 9, 38-50.
Ulrich RL, Hines HB, Parthasarathy N und Jeddeloh JA. Mutational analysis and biochemical
characterization of the Burkholderia thailandensis DW503 quorum-sensing network. (2004)
Journal of Bacteriology. 186, 4350-4360.
Valk G, Loers E und Küppers P. Die Rolle von Oligomer bei textilen Verarbeitungsprozessen von
Polyesterfasern. (1970) Melliand Textilberichte. 5, 504-508.
Vertommen MAME, Nierstrasz VA, van der Veer M und Warmoeskerken MMCG. Enzymatic surface
modification of poly(ethylene terephthalate). (2005) Journal of Biotechnology. 120, 376-386.
von Tigerstrom RG und Stelmaschuk S. The use of Tween 20 in a sensitive turbidimetic assay of
lipolytic enzymes. (1989) Canadian Journal of Microbiology. 35, 511-514.
Wall T, Bath K, Britton RA, Jonsson H, Versalovic J und Roos S. The early response to acid shock in
Lactobacillus reuteri involves the ClpL chaperone and a putative cell wall-altering esterase.
(2007) Applied and Environmental Microbiology. 73, 3924-3935.
Walton TJ und Kolattukudy PE. Determination of the Structures of Cutin Monomers by a Novel
Depolymerization Procedure and Combined Gas Chromatography and Mass Spectrometry.
(1972) Biochemistry. 11, 1885-1897.
Wang X, Geng X, Egashira Y und Sanada H. Purification and characterization of a feruloyl esterase
from the intestinal bacterium Lactobacillus acidophilus. (2004) Applied and Environmental
Microbiology. 70, 2367-2372.

Literaturverzeichnis 179
Wang X, Lu D, Jönsson LJ und Hong F. Preparation of a PET-hydrolyzing lipase from
Aspergillus oryzae by the addition of bis(2-hydroxyethyl)terephthalate to the culture medium
and enzymatic modification of PET fabrics. (2008) Engineering in Life Sciences. 8, 268-276.
Weadge JT, Pfeffer JM und Clarke AJ. Identification of a new family of enzymes with potential O-
acetylpeptidoglycan esterase activity in both Gram-positive and Gram-negative bacteria.
(2005) BMC Microbiology. 5, 49.
Weber K und Osborne M. The Reliability of Molecular Weight Determinations by Dodecyl Sulfate-
Polyacrylamide Gel Electrophoresis. (1969) The Journal of Biological Chemistry. 244, 4406-
4412.
Welzel K, Müller R-J und Deckwer W-D. Enzymatischer Abbau von Polyester-Nanopartikeln. (2002)
Chemie Ingenieur Technik. 74, 1496-1500.
Werkes H. Der Einfluss der Nachbehandlung auf den Oberflächengehalt an Oligomer bei
Polyesterfasern aus Polyglykolterephthalat. (1975) Diplomarbeit, Institut für Textilchemie und
Makromolekulare Chemie, TH Aachen.
Whitehead K, Versalovic J, Roos S und Britton RA. Genomic and genetic characterization of the bile
stress response of probiotic Lactobacillus reuteri ATCC 55730. (2008) Applied and
Environmental Microbiology. 74, 1812-1819.
Wilhelm S, Tommassen J und Jaeger K-E. A novel lipolytic enzyme located in the outer membrane of
Pseudomonas aeruginosa. (1999) Journal of Bacteriology. 181, 6977-6986.
Wilson DB. Cellulases of Thermomonospora fusca. Methods in Enzymology. (1988) 160, 314-323.
Wu C, Jim TF, Gan Z, Zhao Y und Wang S. A heterogeneous catalytic kinetics for enzymatic
biodegradation of poly(ε-caprolactam) nanoparticles in aqueous solution. (2000) Polymer. 41,
3593–3597.
Xiang H, Takaya N und Hoshino T. Novel cholesterol esterase secreted by Streptomyces persists
during aqueous long-term storage. (2006) Journal of Bioscience and Bioengineering. 101, 19-
25.
Xie J und Hsieh YL. Modification of cellulose solids by enzyme catalysed transesterification with vinyl
esters in anhydrous organic solvents. (2003) ACS Symposium Series. 840, 217-230.
Yang C-H und Liu W-H. Purification and properties of an acetylxylan esterase from
Thermobifida fusca. (2008) Enzyme and Microbial Technology. 42, 181-186.
Yang Y und Li S. Removal of cyclic trimer from the surface of disperse dyed polyester. (2000) Textile
Chemist and Colorist & American Dyestuff Reporter. 32, 42-46.
Yang Y, Malten M, Grote A, Jahn D und Deckwer W-D. Codon optimized Thermobifida fusca
hydrolase secreted by Bacillus megaterium. (2007) Biotechnology and Bioengineering. 96, 780-
794.

Literaturverzeichnis 180
Yoon M-Y, Kellis J und Poulose AJ. Enzymatic modification of polyester. (2002) AATCC – Review. 2,
33-36.
Zeronian SH und Collins MJ. Surface modification of polyester by alkaline treatment. (1989) Textile
Progress. 20, 1-34.
Zhang J, Gong J, Shao S, Qin J und Gu Z. Biodegradability of diethylene glycol terephthalate and
poly(ethylene terephthalate) fiber by crude enzymes extracted from activated sludge. (2006)
Journal of Applied Polymer Science. 100, 3855-3859.
Zhang J, Wang X, Gong J und Gu Z. A study on the biodegradability of polyethylene terephthalate
fiber and diethylene glycol terephthalate. (2004) Journal of Applied Polymer Science. 93, 1089-
1096.
Zhang M, Wang JD, Li ZF, Xie J, Yang YP, Zhong Y und Wang HH. Expression and characterization of
the carboxyl esterase Rv3487c from Mycobacterium tuberculosis. (2005) Protein Expression
and Purification. 42, 59-66.
Zimmermann W und Billig S. Enzymes for the biofunctionalization of poly(ethylene terephthalate).
In: Scheper T (Series Ed) Advances in Biochemical Engineering/Biotechnology, (2011), 1-24,
DOI: 10.1007/10_2010_87.
Zimmermann W, Nimz H und Seemüller E. 1H and 13C NMR spectroscopic study of extracts from
corks of Rubus idaens, Solanum tuberosum and Quercus suber. (1985) Holzforschung. 39, 45-
49.
Zimmermann W. Enzyme Systems for Lignocellulose Degradation (ed. MP Coughlan) In:
Hemicellulolytic enzyme systems from actinomycetes. (1989) Elsevier Applied Science. London
and New York. 167-181.

Publikationen 181
6 Publikationen
Zimmermann W, Billig S. Enzymes for the biofunctionalization of poly(ethylene terephthalate). In:
Biofunctionalization of Polymers and their Applications, Scheper T (Series Ed) Advances in
Biochemical Engineering/Biotechnology, (2011), Volume 125/2011, 97-120, DOI:
10.1007/10_2010_87
Billig S, Oeser T, Birkemeyer C und Zimmermann W. Hydrolysis of cyclic poly(ethylene
terephthalate) trimers by a carboxylesterase from Thermobifida fusca KW3. (2010) Applied
Microbiology and Biotechnology. 87, 1753-1764
Oeser T, Wei R, Baumgarten T, Billig S, Föllner C und Zimmermann W. High level expression of a
poly(ethylene terephthalate)-hydrolyzing carboxylesterase from Thermobifida fusca KW3 in
Escherichia coli BL21(DE3). (2010) Journal of Biotechnology. 146, 100-104
Chen S, Su L, Billig S, Zimmermann W, Chen J und Wu J. Biochemical characterization of the
cutinases from Thermobifida fusca. (2010) Journal of Molecular Catalysis B: Enzymatic. 63, 121-127
Korpecka J, Heumann S, Billig S, Zimmermann W, Zinn M, Ihssen J, Cavaco-Paulo A und Guebitz
GM. Hydrolysis of Cutin by PET-Hydrolases. (2010) Macromolecular Symposium. 296, 342–346

Poster und Vorträge 182
7 Poster und Vorträge
Vorträge bei nationalen und internationalen Veranstaltungen
10/2008 European BioPerspectives in Hannover
Purification and characterization of a carboxyl esterase from
Thermobifida fusca KW3 degrading poly(ethylene terephthalate) trimers
09/2008 Workshop of COST 868 action in Varna
Polyester-cleaving hydrolases from the actinomycete Thermobifida fusca
04/2008 Workshop of COST 868 action in Bratislava
Degradation and modification of poly(ethylene terephthalate) substrates
with an esterase from Thermobifida fusca KW3
04/2007 Workshop of COST 868 action in Sitges / Barcelona
Production of a poly(ethylene terephthalate) hydrolyzing esterase from
Thermobifida fusca KW 3
Postepräsentation bei nationalen Veranstaltungen
09/2010 Workshop of COST 868 action in Analipsi, Crete
Characterization of an extracellular PET-hydrolyzing carboxylesterase
from Thermobifida fusca
05/2009 Biokatalyse: Neue Verfahren, neue Produkte, Bad Schandau
Aufreinigung und Charakterisierung einer Polyethylen-terephthalat-
Trimer spaltenden Carboxylesterase aus Thermobifida fusca KW3
04/2007 Annual Conference of the VAAM, Osnabrück
Degradation of PET (poly ethylene terephthalate) oligomers by an
esterase from Thermobifida fusca
12/2006 5th Leipzig Research Festival for Life Sciences
Degradation of PET (poly (ethylene terephthalate)) Oligomers by an
esterase from Thermobifida fusca
05/2006 5th Biotechnology Symposium of the University of Leipzig
Production of an esterase from Thermomonospora (Thermobifida) fusca
cultivated on cutin, suberin and poly(ethylene terephthalate) substrates
03/2006 Annual Conference of the VAAM, Jena
Production of an esterase from Thermomonospora (Thermobifida) fusca
cultivated on cutin, suberin and poly(ethylene terephthalate) substrates
12/2005 4th Leipzig Research Festival for Life Sciences
Charakterisierung einer Polyesterase aus Thermomonospora fusca für
den biokatalytischen Abbau von PET-Oligomer

Lebenslauf 183
8 Lebenslauf
Promotion
04/2005-03/2008 Promotionsstipendiatin der Deutschen Bundesstiftung Umwelt
(DBU) an der Technischen Universität Chemnitz und der Universität
Leipzig
Professur Mikrobiologie und Bioverfahrenstechnik
„Induktion und Aufreinigung einer Polyesterase aus
Thermobifida fusca KW 3 zur Untersuchung und Optimierung eines
biokatalytischen Abbaus von PET-Oligomer“
http://www.dbu.de/stipendien_20004/730_db.html
Auslandsaufenthalt
11/2009 Proteinanalytik mittels MALDI Tof/Tof
State Key Laboratory of Biochemical Engineering, Institute of Process
Engineering, Chinese Academy of Science, Beijing
09/2007 Short Term Scientific Mission
Unterstützt durch die COST 868 Action (Biotechnical functionalisation
of renewable polymeric materials) durchgeführt an der Universität
Twente, Niederlande
Berufspraxis
04/2008-03/2010 Universität Leipzig
Mitarbeiterin der AG für Mikrobiologie und Bioverfahrenstechnik
03/2004-03/2005 Technische Universität Chemnitz
Mitarbediterin der AG Bioverfahrenstechnik
01/2003-06/2003 Mitarbeiterin des Forschungszentrum Mittweida e. V.
Universitäre Ausbildung
1996 - 2003 Technische Universität Berlin
Dipl.-Ing. der Medizinischen Biotechnologie („gut“; 1,6)
Diplomarbeit mit dem Thema:
„Probiotische Mikroorganismen in Gärgetränken (bzw. fermentierten
Getränken) – Identifizierung und Stabilitätsuntersuchungen“
Schulbildung
1992 - 1996 Städtisches Gymnasium Mittweida
Allgemeine Hochschulreife („gut“; 1,8)
1984 - 1992 Polytechnische Oberschule Mittweida
Related Documents











