Ecological Aspects of the Marine Planctomycete Rhodopirellula baltica SH1 T - a Whole Genome Array Approach - by Dipl. Biol. Patricia Wecker A thesis submitted in partial fulfilment of the requirements for the degree of Doctor of Philosophy in Microbiology Approved, Thesis Committee Prof. Dr. Frank Oliver Glöckner (Chair) Prof. Dr. Matthias Ullrich (2 nd Reviewer) Prof. Dr. Rudolph Amann (3 rd Reviewer, External) Prof. Dr. Georgi Muskhelisvili (4 th Reviewer) Date of Defence: May 20, 2009 SCHOOL OF ENGINEERING AND SCIENCE

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Ecological Aspects
of the Marine Planctomycete Rhodopirellula baltica SH1T
- a Whole Genome Array Approach -
by
Dipl. Biol. Patricia Wecker
A thesis submitted in partial fulfilment
of the requirements for the degree of
Doctor of Philosophy in Microbiology
Approved, Thesis Committee
Prof. Dr. Frank Oliver Glöckner (Chair)
Prof. Dr. Matthias Ullrich (2nd Reviewer)
Prof. Dr. Rudolph Amann (3rd Reviewer, External)
Prof. Dr. Georgi Muskhelisvili (4th Reviewer)
Date of Defence: May 20, 2009
SCHOOL OF ENGINEERING AND SCIENCE


2
I looked down. Descent was totally unappetizing……
Too much labour, too many sleepless nights,
and too many dreams had been invested to bring us this far.
We couldn’t come back for another try next weekend.
To go down now, even if we could have,
would be descending to a future marked by one huge question:
what might have been?
Thomas F. Hornbein Everest: The West Ridge

Background
3

4
THESIS ABSTRACT
Since the first microbial genome has been finished in the year 1995 more than 800 sequenced
microorganisms are publicly available and many more are on their way. Sophisticated
bioinformatic (in silico) analysis of the sequence information is required to process this huge
amount of data and to generate sound hypotheses in terms of physiology and ecology of an
organism as well as to efficiently screen for candidate genes for biotechnological relevant
enzymes. Current success to directly obtain sequence information from the environment
without former cultivation (the metagenomic approach) even speeds up the process. The most
prominent in the marine field being the Venter Sargasso Sea expedition in 2004 delivering
more than 1.2 million new genes. The introduction of new sequencing technologies, such as
pyrosequencing will soon enable biologists to perform sequencing on a daily basis.
However, high-throughput lab-methods, implemented by the DNA microarray technology, are
more than ever required for the subsequent verification of initial bioinformatic findings and
hypotheses. Post-genomic approaches like DNA microarrays have revolutionised molecular
biology by enabling a new dimension of parallelism in expression profiling.
Eight years ago, under the light of the up-coming cutting-edge technologies of the genomic
era, the Max Planck Institute for Marine Microbiology in Bremen has initiated a genomics
project for the complete genome sequencing and analysis of environmental relevant marine
bacteria. Rhodopirellula baltica SH 1T was selected as a marine representative of the globally
distributed and environmentally important bacterial phylum Planctomycetales. It is a model
organism for aerobic carbohydrate degradation in marine systems, where polysaccharides
represent the dominant components of biomass.
The complete genome sequence of R. baltica has been determined and functional annotation
was performed. However, pure in silico predictions left about 55% of the genes without
functional assignment. The availability of the whole genome sequence has already stimulated
a set of follow-up studies ranging from proteomics to protein crystallisation as well as
biotechnological applications.
This work presents the establishment of, and the results gained with, the first whole genome
microarray for a Planctomycete. It shows that massive expression profiling under defined
environmental conditions can take the challenge to bridge the gap between classical studies on
the cell biology and physiology of Planctomycetes and their active genetic repertoire.

Background
5
TABLE OF CONTENT
1 BACKGROUND ............................................................................................................................. 1
1.1 MARINE MICROBIAL ECOLOGY AND THE GENOMIC REVOLUTION............................................ 1 1.2 PLANCTOMYCETES......................................................................................................................... 2 1.2.1 RHODOPIRELLULA BALTICA .......................................................................................................... 4 1.3 FURTHER OUTCOME OF THE GENOME ANNOTATION AND ITS LIMITATIONS............................ 9 1.4 GENE EXPRESSION PROFILING BY DNA MICROARRAY ANALYSIS .......................................... 10 1.5 PRINCIPLE OF DNA MICROARRAY ANALYSIS........................................................................... 11 1.6 MICROARRAY DATA ANALYSIS .................................................................................................. 14 1.7 MICROARRAY DATA MANAGEMENT.......................................................................................... 16
2 RESEARCH AIMS....................................................................................................................... 17
2.1 THE RHODOPIRELLULA BALTICA LIFE-CYCLE: GROWTH-DEPENDENT EXPRESSION PROFILING
OF THE COMPLETE GENOME WITH A CDNA-BASED MICROARRAY .................................................. 17 2.2 EXPRESSION PROFILING OF THE SULPHATASES GENES OF THE PLANCTOMYCETE
RHODOPIRELLULA BALTICA GROWN ON DIFFERENT SULPHATED POLYSACCHARIDES .................... 18 2.3 DESIGN OF THE WHOLE GENOME ARRAY OF RHODOPIRELLULA BALTICA .............................. 20
3 RESULTS AND DISCUSSION ................................................................................................... 21
3.1 TRANSCRIPTIONAL RESPONSE OF THE MODEL PLANCTOMYCETE RHODOPIRELLULA BALTICA
SH1T TO CHANGING ENVIRONMENTAL CONDITIONS......................................................................... 23 3.2 THE RHODOPIRELLULA BALTICA LIFE-CYCLE: GROWTH-DEPENDENT EXPRESSION PROFILING
OF THE COMPLETE GENOME WITH MICROARRAYS ........................................................................... 55 3.3 HIGHLY ENANTIOSELECTIVE SEC-ALKYL SULFATASE ACTIVITY OF THE MARINE
PLANCTOMYCETE RHODOPIRELLULA BALTICA SHOWS RETENTION OF CONFIGURATION............... 89 3.4 FOSMIDS OF NOVEL MARINE PLANCTOMYCETES FROM THE NAMIBIAN AND OREGON COAST
UPWELLING SYSTEMS AND THEIR CROSS-COMPARISON WITH PLANCTOMYCETE GENOMES ......... 90
4 SUMMARY ................................................................................................................................... 91
4.1 TRANSCRIPTIONAL RESPONSE OF THE MODEL PLANCTOMYCETE RHODOPIRELLULA BALTICA
SH1T TO CHANGING ENVIRONMENTAL CONDITIONS......................................................................... 92

6
4.2 THE RHODOPIRELLULA BALTICA LIFE-CYCLE: GROWTH-DEPENDENT EXPRESSION PROFILING
OF THE COMPLETE GENOME WITH MICROARRAYS ........................................................................... 93 4.3 HIGHYL ENANTIOSELECTIVE SEC-ALKYL SULFATASE ACTIVITY OF THE MARINE
PLANCTOMYCETE RHODOPIRELLULA BALTICA SHOWS RETENTION OF CONFIGURATION............... 94 4.4 FOSMIDS OF NOVEL MARINE PLANCTOMYCETES FROM THE NAMIBIAN AND OREGON COAST
UPWELLING SYSTEMS AND THEIR CROSS-COMPARISON AND THEIR CROSS-COMPARISON WITH
PLANCTOMYCETE GENOMES ............................................................................................................... 95
5 ESTABLISHING THE MICROARRAY PIPELINE................................................................ 96
6 CONTRIBUTION TO THE RESEARCH ON PLANCTOMYCETES..................................... 98
7 OUTLOOK.................................................................................................................................. 100
8 REFERENCE.............................................................................................................................. 102
9 ACKNOWLEDGEMENT.......................................................................................................... 109
10 ANNEX ...................................................................................................................................... 111


Background
1
1 BACKGROUND
1.1 Marine microbial ecology and the genomic revolution
Modern marine microbial ecology was born in the 1970`s, when it was shown that bacteria
were very abundant in the oceans and that most marine respiration was in the bacterial size
fractions (Pedros-Alio 2006). Before then microbes were not even considered in the ecology
of the oceans. Today we know that marine microbes are present at billions of cells per litre in
seawater and that they play a key role in ocean ecosystems (DeLong and Karl 2005). It is
therefore of crucial importance to understand microbial roles in oceanic primary production,
the global carbon cycle and functioning of the biosphere.
Microbiologists have been advancing their knowledge over the last 30 years through a
progression of approaches that include taxonomic and physiological studies of cultivatable
isolates, as well as cultivation-independent, rRNA-based phylogenetic survey techniques and
genetic approaches. The invention of genome-based technologies (genomics) in the 1990´s
revolutionised microbial ecology. The technological power of high-throughput sequencing
and functional genomics enabled the study of genetic complexity of organisms at the genome
level, while genetics had before focused on limited numbers of genes and gene products
(Dupont et al. 2007). Microbiologists are now facing a genomic “data flood” with more than
900 completed bacterial or archaeal genome sequences available and over 3000 more in
progress (Thomas et al. 2007).
With this, a new scientific discipline was created in marine microbial ecology, “marine
ecological genomics” that studies the structure and functioning of a genome with the aim to
understand the relationship between the organism and its biotic and abiotic environments.
Approaches include (1) whole genome sequencing of key organisms and (2) genomic analysis
of natural communities to understand how biodiversity supports ecosystem function
(metagenomics). However, even though a given genome represents the blueprint of life for an
organism, there is a need for functional analysis at the transcriptional and proteomic levels in
order to define which of (1) the predicted genes can be expressed in principle, and (2) which
physiological condition(s) induce their expression. In sharp contrast to the numerous
publications of microbial genome sequences, several transcriptome and proteome studies have
been reported so far (Zhou 2003).
Already in the year 2000, the Department of Molecular Ecology at the Max Planck Institute
for Marine Microbiology has initiated a marine environmental genomics project – the real

Background
2
environmental genomics-project (REGX). REGX aims at understanding the adaptations of
marine bacteria to changing environmental conditions. Two sulphate reducing bacteria
(i.e.: Desulfotalea psychrophila and Desulfobacterium autotrophicum) and one Planctomycete
(Rhodopirellula baltica SH1T) were selected for whole genome sequencing, annotation and
functional analysis. The successful sequencing and annotation of the whole genome of the
Planctomycete R. baltica in 2003 (Glöckner et al. 2003) has established the framework for
further functional genomic analysis.
1.2 Planctomycetes
Planctomycetes are a group of budding, peptidoglycan-less bacteria with increasing
significance for microbial evolution, ecology, cell biology and genomics (Fuerst 1995). The
phylum consists of a single family (Planctomycetaceae) with six accepted (Rhodopirellula,
Blastopirellula, Pirellula, Planctomyces, Isosphaera, Gemmata) (Schlesner et al. 2004) and
four candidate genera (Kuenenia, Brocadia, Scalindua, Anammoxoglobus). Latest studies
group Planctomycetes with the PVC super phylum. This monophyletic clade is formed
besides the Planctomycetes by the phyla Chlamydiae, Verrrucomicrobia, Lentisphaerae and
the candidate phyla ‘Poribacter’and OP3 (Wagner and Horn 2006).
Most cultured strains so far have been isolated from terrestrial, freshwater and marine habitats
(Schlesner 1994; Fuerst 1995; Wang et al. 2002). In addition, studies based on cultivation-
independent molecular methods, e.g. Fluorescence in situ Hybridisation (Amann and Ludwig
2000), repeatedly confirmed the occurrence of Planctomycetes in terrestrial and aquatic
habitats (DeLong and Franks 1993; Chatzinotas et al. 1998; Neef et al. 1998; Vergin et al.
1998; Crump et al. 1999) and revealed an even broader distribution of the Planctomycetes in
the environment, e.g. in hot springs (Giovannoni et al. 1987), marine sponges (Fuerst et al.
1998; Fuerst et al. 1999; Pimentel-Elardo et al. 2003) as well as in hepatopancreas of the
crustacean Panaeus monodon (Fuerst et al. 1991; Fuerst et al. 1997). This shows the
successful colonisation of a broad spectrum of ecological niches by members of this phylum.
Further surveys reported the presence of Planctomycetes in marine-snow particles (Rath
1998) as well as in shallow and deep sea sediments of marine systems (Llobet-Brossa et al.
1998; Inagaki et al. 2006; Kirkpatrick et al. 2006; Musat et al. 2006). Morris and coworkers
showed that Planctomycetes appear and disappear with the blooming of diatoms (Morris et al.
2006). They postulate a direct interaction and carbon flow between the algae and
Planctomycetes. In addition, growth studies with R. baltica and other group members revealed

Background
3
their nutritional specialisation in aerobic carbohydrate utilisation (Schlesner et al. 2004),
which points an important role in the global carbon cycle (Glöckner et al. 2003).
In terms of cell biology all members of this group share several morphologically unique
properties, such as a peptidoglycan-free proteinaceous cell wall (König et al. 1984; Liesack
et al. 1986), intracellular compartmentalisation (Lindsay et al. 1997; Lindsay et al. 2001) and
a mode of reproduction via budding which results in a cell-cycle that is characterised by
motile and sessile morphotypes and resembles that known from Caulobacter crescentus
(Tekniepe et al. 1981; Fuerst 1995; Glöckner et al. 2003; Jacobs-Wagner 2004).
Glöckner et al. (Glöckner et al. 2003) published the first complete genome sequence of the
marine planctomycete Rhodopirellula baltica SH1T. With that, the “genomic revolution”
reached the Planctomycetes. Several genome projects have been initiated in the last year in
order to get the complete picture of the potential of this phylum. An early draft of the genome
of the freshwater isolate of Gemmata obscuriglobus UQM 2246T (Franzmann and Skerman
1984) from the Institute for Genomic Research (TIGR) project is now available. Draft
sequences of the two marine planctomycetes Blastopirellula marina DSM 3645T (Schlesner
et al. 2004) and Planctomyces maris DSM 8797T (Bauld and Staley 1976) are funded by the
Gordon and Betty Moore foundation on behalf of the Max Planck Institute, Bremen and are
now almost completely annotated (Woebken et al. 2007). Finally, the anammox
planctomycete Candidatus Kuenenia stuttgartiensis was investigated by a metagenomic
project (Strous et al. 2006).

Background
4
1.2.1 Rhodopirellula baltica Pirellula sp. strain 1, which now has been validated as "Rhodopirellula baltica
SH1T"(Schlesner et al. 2004), is a marine aerobic, heterotrophic representative of the phylum
Planctomycetes. R. baltica was isolated from the
water column in the Kiel fjord which is part of the
Baltic Sea. The whole genome sequence of
R. baltica has been completed and published
(Glöckner et al. 2003) by the REGX project research
group at the Max Planck Institute for Marine
Microbiology, Germany (www.regx.de). With 7,145
Mb and 7,325 open reading frames, the genome of
R. baltica represented one of the largest bacterial
genomes sequenced at that time. Fig 1 Rhodopirellula baltica EM (Schlesner et al. 2004)
1.2.1.1 Division and cell-cycle
The pear-shaped adult cells of Rhodopirellula baltica are non-motile and display a polar
organisation. From the smaller cell pole a holdfast substance of yet unknown chemical
composition is excreted. It mediates attachment of cells to surfaces or to each other leading to
the formation of rosettes (Fig 2). The broader cell pole is the site of budding. The bud is a
mirror image of the mother cell, except that it is smaller in size and has a single flagellum
inserted in a sub polar position at the proximate pole. At a certain developmental stage the
bud (daughter cell) is released as a motile swarmer. The swarmer cells abort their flagellum,
when differentiating into adult cells. Only these adult cells possess fimbriae that originate
from so-called crateriform structures at the broader (reproductive) cell pole (Olga Lang,
personal. communication). While these crateriform structures are restricted to the broader cell
pole in adult cells, they are observed across the complete cell surface of buds (Gade et al.
2005).
Gade and coworkers (Gade et al. 2005) showed that there is a correlation between distinct
morphotypes and different growth phases of R. baltica cultures based on microscopic
examination and analysis. As illustrated in the growth curve in Fig 2, during early stages of
growth numerous budding mother cells and swarmer cells are present. As growth proceeds,
the number of swarmers decreases, and single cells begin to attach to each other forming

Background
5
rosettes. The latter are the dominating morphotype during transition to and in the stationary
phase, where swarmer cells are rarely observed.
Fig 2 Schematic drawing of the life-cycle of Rhodopirellula baltica. See text for details (Gade et al. 2005)
The cellular mechanisms regulating these morphological changes during cell-cycle of
R. baltica are not currently understood. For example during the unequal cell division nothing
is known about how the cell compartments are distributed between the daughter cells. In
general, the cell division machinery of Bacteria consists of different cell division genes, such
as the filamentation temperature sensitive genes (fts) named ftsZ, ftsA, ftsI, ftsL, ftsQ, ftsN,
ftsK, ftsW and the FtsZ interacting proteinA (zipA). The ftsZ-gene is the major constituent of
the septal replication ring. Surprisingly, all of these genes, except ftsK, are absent from the
genome of R. baltica. Interestingly, also not present in the genome of the planctomycete
Gemmata obscuriglobus UQM2246T. Teeling (Teeling 2004) concluded that the cell division
in Planctomycetes, like in Chlamydia, and must go through a mechanism totally unknown at
present.
As noted earlier, morphological changes, during the life-cycle of R. baltica seem to be similar
to C. crescentus CB15. Generally the key gene of the cell-cycle is the control protein CtrA. Its
task as the essential master regulator is overseeing several important cellular processes
(Stephens 2001). Within the R. baltica genome no homologue to the protein CtrA has been
found. However some patterns (e.g. TTAAN7AAAC), were found which are similar to CtrA
and CzcR – binding site patterns (TTAAN7TTAA).

Background
6
1.2.1.2 Substrate spectrum
Physiological studies on R. baltica showed optimal growth between 28 and 30°C. The
bacterium is strictly aerobic, glucose will not be fermented and nitrate cannot serve as an
electron acceptor. Growth experiments on liquid yielded that R. baltica can utilise a wide
range of mono- and disaccharides (Tab 1) (Schlesner et al. 2004).
Tab 1 Substrates tested on Rhodopirellula baltica modified after Schlesner et al. (Schlesner et al. 2004) Substrates used as carbon source Substrates not used as carbon source
Monosaccharides
C6: glucose, fructose, mannose, galactose, trehalose,
C5: lyxose, ribose, xylose
Monosaccharides
C6: sorbose
C5: -
Modified monosaccarides
methylated: rhamnose
N-acetylated: N-acetylglucosamine
Others: esculin, salicin
Modified monosaccarides
methylated: fucose
N-acetylated: -
Others: -
Disaccharides
cellobiose, lactose, maltose, sucrose, melibiose,
amygdalin
Disaccharides
-
Trisaccharides
melezitoase, raffinose
Trisaccharides
-
Polysaccarides
chondroitin sulphate, gelatine, starch, dextrin
Polysaccarides
cellulose, chitin, alginate
C1
-
C1
methylamine, methylsulfonate, methanol
Others
glycerol, gluconate, glucuronate
Others
methanol, ethanol, erythritol, adonitol, arabitol,
dulcitol, inositol, mannitol, sorbitol, acetate,
adipate, benzoate, caproate, citrate, formiate,
fumarate, glutarate, lactate, malate, 2-
oxoglutarate, phthalate, propionate, pyruvate,
succinate, tartrate, norleucine, ornithine, urea,
indole, inulin, pectin, casein, tween 80, all 20
amino acids
N-acetylglucosamine (NAG) can be used as the only energy and nitrogen source (Rabus et al.
2002). NAG is a main structural component in chitin and a substantial constituent of the

Background
7
bacterial peptidoglycan cell wall. It is one of the main amino sugars in marine habitats
(Riemann and Azam 2002). Additional growth studies show that R. baltica can grow on
chondroitin sulphate (component of fish cartilage) (Schlesner et al. 2004) and iota-carrageen-
plates (algae) (Gurvan Michel pers. communication).
The annotation process identified the standard pathways for heterotrophic bacteria including
glycolysis, citrate cycle and oxidative phosphorylation. R. baltica SH1T lacks the glyoxylate
bypass and the Entner-Doudoroff pathway but exhibits the pentose phosphate cycle.
Unexpected for an aerobic heterotrophic bacterium was the presence of all genes for
heterolactic acid fermentation, though the organism has not yet been successfully cultured
under anoxic conditions. Interestingly, key genes for the inter-conversion of C1-compounds
were also found. Previously these were believed to occur only in methanogenic Archaea and
methylotrophic Alphaproteobacteria (Glöckner et al. 2003; Bauer et al. 2004).
1.2.1.3 Sulphatases and their role
The annotation results showed that the R. baltica genome harbours 110 genes encoding
proteins with significant similarity to prokaryotic (82 genes; 75%) and eukaryotic (28 genes;
25%) sulphatases. For instance, similarities were found to alkylsulphatase of Pseudomonas
aeruginosa, to arylsulphatases of Pseudomonas sp., to mucin-desulfating sulphatase of
Prevotella sp., and to archaeal arylsulphatase, as well as to mammalian iduronate-2-
sulphatase and arylsulphatases A and B.
The annotation also revealed that in R. baltica the sulphatase genes are distributed across the
genome in 22 clusters containing two to five genes. 59 sulphatases out of the set of 110
predicted sulphatases contain signal peptides prediction, indicating that most of these proteins
are probably secreted. 11 predicted sulphatases have a high expression level prediction (PHX)
(Lombardot 2004).
In comparison, analysis of 70 published prokaryotic genomes with a specific Pfam profile
revealed a maximum of only 6 sulphatases found in the Pseudomonas aeruginosa PAO1
genome and 30 in Bacteriodes thetaiotaomicron (Lombardot 2004). However, recent studies
conclude that the number of genes and bacterial species encoding sulphatase enzymes is
currently underestimated (Berteau et al. 2006). Until now, the 110 sulphatases found in
R. baltica (15.3 per Mb) represents the highest number in any sequenced bacterial genome. In
other planctomycetal genomes, genes encoding sulphatases were found in various numbers.
Planctomyces maris DSM8797T draft genome codes for 87 sulphatases (10.7/Mb), in
Blastopirellula marina DSM3645T 41 were found (6.16/Mb) and in the freshwater

Background
8
planctomycete Gemmata obscuriglobus UQM2246T only 11 sulphatases are present
(1.37 Mb) (Woebken et al. 2007). Another marine organism, Zobellia galactanivorans,
belonging to the CFB group has around 70 genes annotated as sulphatases (Gurvan Michel,
personal communication).
Sulphatases are widespread enzymes found in Bacteria, Archaea and Eukarya. They are
involved in various metabolic processes, ranging from a sulphate starvation response in
bacteria to hormone biosynthesis and the modulation of developmental cell signalling in
mammals (Parenti 1997). In humans, their biological relevance is particularly underlined by
their involvement in several inherited diseases, such a mucopolysaccaridoses. It is known that
sulphatases act on a broad diversity of substrates, which leads to their classification by the
IUBMB into 17 classes (from EC 3.1.6.1 to EC 3.1.6.18) (Berteau et al. 2006). The role of the
sulphatases in R. baltica is not yet clearly understood.
Investigations of the 110 sulphatases in R. baltica based on the transcriptome and proteome
level revealed that at least a fraction of them are expressed (Gade et al. 2005; Gade et al.
2005; Würdemann 2006). Sulphatase activity studies performed at the University of Graz
(Wallner et al. 2004), show a high enantioselective sec-alkyl sulphatase activity with retention
of configuration when several substrates where tested on resting whole cells of R. baltica
In summary, the data on the nature of substrates utilised as carbon sources by R. baltica and
the high numbers of sulphatase genes found in the genome and the proven activity of at least
some of them suggest that sulphatases are metabolically important in R. baltica and could
play a role in the efficient degradation of sulphated glycopolymers. Such compounds (e.g.
carrageen) are abundant in marine environments in the form of phytodetrital macroaggregates
(“marine snow”), and Planctomycetes have been shown to be components of the microbial
communities on such aggregates (Glöckner et al. 2003; Woebken et al. 2007).

Background
9
1.3 Further outcome of the genome annotation and its limitations
The availability of different planctomycetal genomes has led to an enormous flood of new
information. This has led to the popular phrase that the pre-genome era was the “dark age”
(Dunham 2000). However, it is well known that, in any newly sequenced bacterial genome
30-40% of the genes do not have an assigned function. This number is even higher for
archaeal and eukaryotic genomes and for the relatively large genomes of bacteria with a
complex life-style, such as Anabaena and Streptomyces (Galperin and Koonin 2004). It is also
valid for R. baltica. In 2003, of the 7325 potential proteins in R. baltica only 3380 (46%) had
a significant hit when compared with public databases. This means that over 54% of proteins
in the genome remain uncharacterised and will be referred to as “hypothetical proteins” or
with the affix “conserved” in case of wider phyletic distribution (Galperin and Koonin 2004).
So far, their function can only be subject to speculation. Some of these hypothetical proteins
are planctomycete-specific or even unique to R. baltica. The former means that they only
occur in a maximum of all four planctomycetal genomes. It seems likely that some of these
genes code for the unique cellular planctomycetal characteristics and for planctomycete-
specific metabolic traits. A large proportion of these planctomycete-specific genes carry one
or more specific domains of as yet unknown function (DUF) (Woebken et al. 2007). Often
they appear in groups and/ or before operon-like arrangements, indicating a potential
regulatory function. This may be supported by the fact, that an unusually low number (2.4%)
of transcriptional regulators from known families were found in the genome (Gade et al.
2005).

Background
10
1.4 Gene expression profiling by DNA microarray analysis
The genome annotation of R. baltica combined with the experimental work that has been
carried out on this organism and on other members of the Planctomycetes provided new
information and generated hypotheses about the metabolic capabilities and life-style of these
organisms. However, in order to turn these mostly computationally derived hypotheses into
real biological knowledge there is an urgent need to verify the predicted gene functions by
experiments using different molecular techniques, for example gene expression profiling.
Gene expression is a highly complex and tightly regulated process that allows a cell to
respond dynamically both to environmental stimuli and to its own changing needs. This
mechanism acts as both an “on/off” switch to control which genes are expressed in a cell, and
as a ”volume control” that increases or decreases the level of expression of particular genes as
necessary (Dupont et al. 2007). Gene expression analysis examines the composition of
cellular messenger RNA populations.
Traditional gene expression analysis has used techniques such as Northern blotting, RT-PCR
and nuclease protection assays. More advanced methods – some of these include differential
display, subtractive hybridisation, representational difference analysis, expressed sequence
tags, cDNA fragment fingerprinting, and serial analysis of gene expression – have enabled the
discovery of novel differentially expressed genes. However, the technical challenges of these
methods still limit their use to the analysis of just a few samples/genes at a time. Microarray
analysis, in contrast, allows the analysis of thousands of genes in multiple samples with
relative ease (Duggan et al. 1999).
Gene expression analysis using DNA microarrays has been applied to numerous mammalian
tissues, yeast (Alizadeh and Staudt 2000), and bacteria alike (Hu et al. 2005; Steglich et al.
2006). These studies examined the effects of different chemicals on cells, the consequences of
over-expression of regulatory factors in transfected cells, and compared mutant strains with
parental strains to delineate functional pathways. In cancer research, microarrays have been
used to find gene expression changes in transformed cells and metastases, to identify
diagnostic markers, and to classify tumours based on their gene expression profile (Amersham
2002). Lately, transcriptional profiling is even applied to study environmental questions
(Zhou 2003; Parro et al. 2007). Dupont and co-workers expect this to be the major activity of
ecological genomics in the near future (Dupont et al. 2007).

Background
11
1.5 Principle of DNA microarray analysis
DNA microarrays contain thousands of DNA samples or oligonucleotide sequences named
probes (Fig 3). The probes are printed or synthesised in a precise and known pattern onto
nylon membrane filter or microscope glass slide. In the latter case the slides are coated with
chemically reactive groups typically aldehydes or primary amines to help to stabilise the DNA
(probes) onto the slide, either by covalent bonds or electrostatic
interactions. Applied onto the printed chip surface will be the labelled
hybridisation partner, which is complementary to the probe and referred
to as target (Fig 3). The basic principle of complementary base pairing
underlies the specific hybridisation of these two molecule types, and is
common to all microarray approaches.
Fig 3 Nomenclature: tethered nucleic acid as “probe” (spotted oligos) and free nucleic acid as “target” (labelled cDNA) (Phimister 1999)
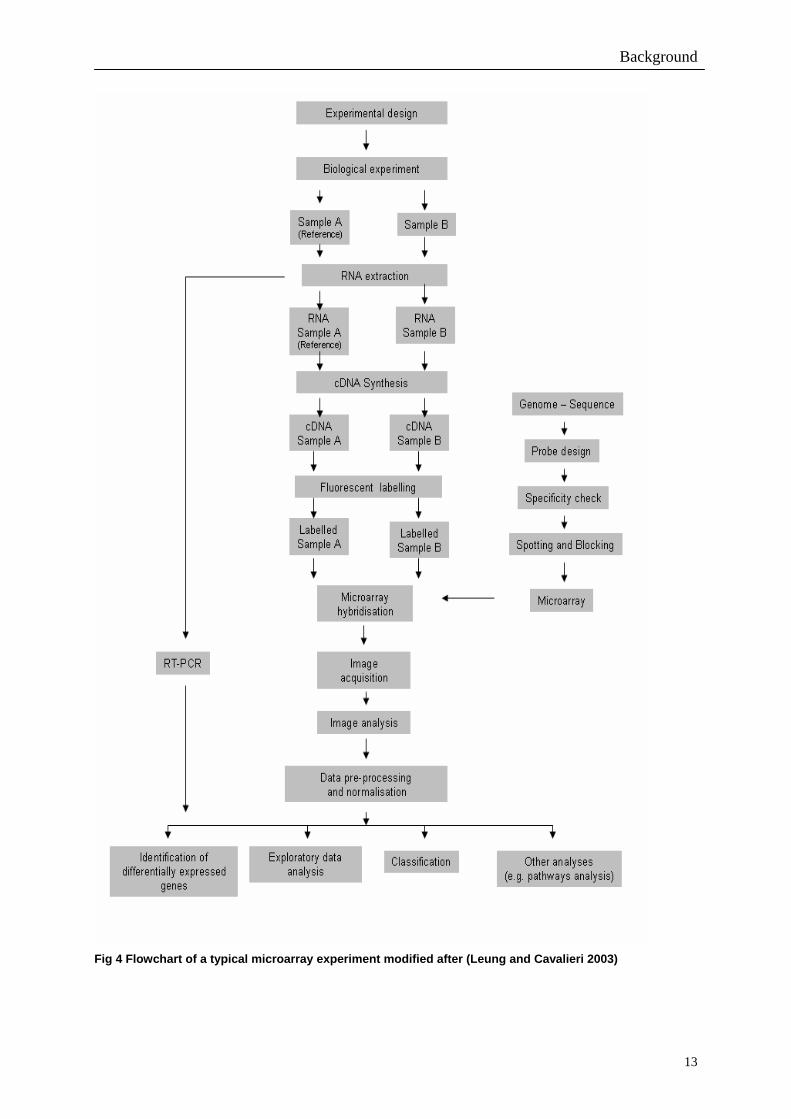
A typical workflow of a cDNA microarray experiment used in gene expression analysis is
summarised in Fig 4. First, the probes have to be designed by the use of a bioinformatic
pipeline. This includes screening oligos for their thermodynamic properties as well as
secondary structures, calculation of oligo Tm for uniform Tm value and BLAST searches
against the organism to guaranty specificity. Meanwhile, immobilisation parameters like
humidity and temperature used for slide production have to be optimised according to the
chemical features of the printing tip and the slide surface chemistry.
Once the probes have been synthesised and printed on the chip, focus is shifted to the sample.
Our flowchart experiment (Fig. 4) compares the relative expression level of specific
transcripts in two samples. One of these samples is the control, named reference (A) and the
other is derived from cells whose response or status is being investigated (B). The sampling is
followed by RNA extraction. Both steps are important components of the target preparation
process since successful microarray studies depend on the consistent extraction of high
quality RNA. The extracted sample and reference RNA are reverse-transcribed into cDNA
and labelled with one of two fluorochromes: one fluorochrome for the reference and one
fluorochrome for the sample (e.g. Cyanine- or Alexa- dyes). Different techniques of labelling
are used. The label can be either introduced by direct enzymatic incorporation of fluorescently
labelled nucleotides or by the direct labelling approach (Zhang et al. 2001; Gupta and
Cherkassky 2003) and also indirectly by incorporation of aminoally-dUTP and subsequent

Background
12
coupling of fluorescent dyes (Beier et al. 2004). Equal amounts of the labelled target cDNA
(reference + sample) are combined, co-hybridised under stringent conditions onto a
microarray replicate and washed in a defined procedure. Afterwards the slide is scanned using
a specialised laser scanner to record the emission of the two fluorochromes for each spot on
the microarray after excitation at a given wavelength. The intensity of the fluorescence
emission signals on each spot is proportional to transcript levels in the biological samples and
the fluorescent signal from each gene probe is quantified by using specialised image
processing software. The output can then be analysed and normalised using different analysis
software. A standard method to evaluate the results is quantitative Real-Time (RT)-PCR.
Background
13
Fig 4 Flowchart of a typical microarray experiment modified after (Leung and Cavalieri 2003)

Background
14
1.6 Microarray data analysis
Data analysis is an essential process in DNA microarray experiments, since these experiments
normally result in a large amount of information. These data must be adequately processed to
find statistically significant correlations – e.g. co regulation of genes - within and between
different arrays (FIG 4).
First, the hybridisation signal intensities must be filtered and normalised. This transformation
is done to minimise the bias arising from unequal quantities of starting RNA, differences in
labelling or detection efficiencies of the fluorescent dyes applied, and other systematic biases
(Quackenbush 2002). For an overview on different normalisation methods see Foster and
Ghazal 2003(Foster and Ghazal 2003). In the next step, data mining techniques are required to
answer the biological question behind the experiment. Normally, microarray experiments are
conducted to identify genes which are either under- or over-expressed after a shift in the
experimental conditions. For example, we might be interested in genes that have an elevated
expression because of a drug treatment. Such genes are most easily found by simple filtering.
If the log-transformed data (method) is used for filtering, differentially expressed genes are
inferred by a fixed threshold cut off method (i.e. a two-fold increase or decrease). Filtering by
absolute expression change can even be used for experiments, where there are no replicates.
However, there are also ranking-methods available [t-test (Pan 2002), ANOVA (Kerr et al.
2000), Bayesian method or Mann-Whitney test]. All these methods produce errors (false-
positive and false-negatives), therefore differential gene expression is usually confirmed by
RT-PCR or northern blots (Leung and Cavalieri 2003). In case of interest for co-regulation of
genes (or related arrays), various cluster techniques should be considered. The basic concept
in clustering is to try to identify and group together similarly expressed genes and to correlate
the observations to biology. The idea is that co-regulated and functionally related genes are
grouped into clusters. Some often used grouping techniques are hierarchical clustering (Eisen
et al. 1998), k-means clustering (Soukas et al. 2000), self-organising maps (SOMs) (Kohonen
1992) and principal component analysis (PCA) (Raychaudhuri 2000) (Methods reviews
Quackenbush 2002; Gollub and Sherlock 2006). There is no clustering method that can be
applied for all kinds of experiments. Different cluster methods used on the same data set can
reveal unique aspects of the data (Leung and Cavalieri 2003). It is therefore advisable to
analyse the data using several methods rather than just one (Leung 2002).

Background
15
Finally, cluster data can be mined for e.g. promoter elements involved in the regulation
observed (Brazma and Vilo 2000), for novel functions of orphan proteins, and for novel
regulatory relationships between genes under study (Knudsen 2004). Then, gene expression
patterns can be correlated with physiology through the functional networks (pathways) and
regulatory mechanisms in which the relevant genes are involved. A wide variety of resources
and tools exists to display pathway information, enabling presentation of microarray data in
the framework of documented biological pathways (Imbeaud and Auffray 2005).

Background
16
1.7 Microarray data management
Data warehousing for microarray experiments is an informatic challenge due to the thousands
of elements a typical microarray experiment will produce. There are two types of storage
systems, local for in-house data management and public array databases for meta-mining.
Raw data in the form of images carry the most information but are difficult to share
effectively through the internet due to their large size. Analysed data, however, can be
disclosed easily but a lot of information essential for quality assessment is lost during the
analysis process. Attempts to compare data from different databases without realising their
underlying incomparability are devastating to the future development of the microarray field.
Consequently, there is an urgent need to set up a standardised definition for experimental
information to guide the development of a comparable and integrated experimental design and
database system (www.mged.org/Workgroups/MIAMI/miami.html) (Leung 2002). A solution
has been presented in the Minimum Information About a Microarray Experiment (MIAMI)
document offered by the Microarray Gene Expression Data Society (MGED) working group.
The document describes the minimum information required to ensure that microarray data can
be easily interpreted and that results derived from its analysis can be independently verified
(Brazma et al. 2001).
The aphorism “Well begun is half done” is especially true for microarray analysis since
adequate experimental design is extremely important for these kinds of investigations. Leung
and Cavalieri (Leung and Cavalieri 2003) summarise in their review what a good microarray
experimental design should comprise. (i) A clearly defined biological question and/or
hypothesis; (ii) treatment, perturbation and observation of the biological materials, as well as
the microarray experimental protocols should be as little affected by systematic and
experimental errors as possible; (iii) a simple, sensible and sound microarray experimental
arrangement that will yield the maximal amount of information given in the cost structure and
complexity of the study (review by (Churchill 2002)) and (iv) compliance with the MIAMI
(Leung and Cavalieri 2003).

Research Aims
17
2 RESEARCH AIMS
2.1 The Rhodopirellula baltica life-cycle: Growth-dependent expression profiling of the complete genome with a cDNA-based microarray
As in any other bacterium, regulatory networks must coordinate cell-cycle progression with
cell growth, including temporal and spatial control of DNA replication and cytokinesis, to
ensure equal partitioning of chromosomes into daughter cells. The completion and annotation
of the R. baltica genome sequence has allowed a first estimate of the genetic potential.
However, nothing is known so far about the genetic network controlling development, the
cell-cycle and the cellular mechanisms regulating these morphological changes in R. baltica.
It is anticipated that the genes functioning in these networks should show varying mRNA
levels over the life-cycle. Therefore growth-dependent expression patterns could give hints
about genes that might be required in physiological processes, morphotypes and cellular
activity typical for R. baltica. The approach of microarray technology is particularly useful
when applied to complex processes such as the cell-cycle, where many genes are expected to
be involved (Spellman et al. 1998; Laub et al. 2000). Global assays of gene expression during
the Caulobacter crescentus cell-cycle revealed that surprisingly 19% of the genome is
affected as cells grow and divide (Jenal and Stephens 2002).
The whole genome array will be a means to explore the regulation of all genes during the life-
cycle. Clustering of all genes could help to discriminate the function of potential hypothetical
proteins through co-regulation with proteins of known function and also to assign a function
to clustered hypothetical proteins because of their specific cell-cycle pattern. Furthermore,
analysing the set of cell-cycle regulated genes for potential promoter elements could give
some hint about unknown regulatory mechanisms for genes in R. baltica. Fortunately, cell-
cycle proteome data are also available for comparison with obtained transcriptome analysis
data (Gade et al. 2005).
At the same time, fluorescence imaging of membrane stained (Lindsay et al. 2001) dividing
R. baltica cells could help to understand how the division process of the compartments in the
three-dimensional space is taking place. Combining the “omic” knowledge and with the
fluorescence imaging results contribute to our understanding of the underlying molecular cell
biology of the cell compartmentalisation of the phylum Planctomycetales.

Research Aims
18
2.2 Expression profiling of the sulphatases genes of the planctomycete Rhodopirellula baltica grown on different sulphated polysaccharides
The carbon cycle in natural environments depends on the remineralisation of biomass. In
marine systems biomass is mainly produced by phototrophic microorganisms in the upper
layer of the water column (annual production of 20 − 30 × 109 tons of carbon). Dead biomass
settles to the sediment, often aggregating as particles known as marine snow. During this
process, organic carbon is already decomposed by aerobic microorganisms (Rullkötter 1999).
Since polysaccharides are major components of biomass, carbohydrate degradation is
particularly relevant for carbon turnover. R. baltica and other members of the Planctomycetes
are considered key players in carbohydrate metabolism in marine systems, because of their
nutritional specialisation, their high abundance in the marine water column and their
association with marine snow, and also the high number of sulphatases genes found in their
genome.
The role of the sulphatases however, is still unclear. The presence of 110 sulphatase genes of
unknown physiological role in the R. baltica genome represents an ideal starting point for
further functional analysis. Expression profiling of the 110 sulphatase genes and enzymes
from associated pathways (ECF-sigma factor, sugar degradation) in R. baltica provides a
deeper view of the regulation and expression of bacterial sulphatases and their role in the
natural habitat of the organism. First expression profiling experiments on a partial genome
array were carried out with chemically labelled cDNA extracted from R. baltica cultures
grown on glucose and N-acetylglucosamine, ribose and chrondroitinsulphate A and C.
Although some differences in the expression patterns could be detected it could also be seen
that most of the sulphatases under investigation were already expressed during cultivation
with glucose (reference) (Würdemann 2006). The reason for their obvious constitutive
expression is unclear. Further microarray experiments with R. baltica cultures grown on
different oligosaccharides allow specification of the substrate preference of different
sulphatases. The different polysaccharides derived from different marine macroalgae are
provided by cooperation with the CNRS – Laboratoires GOËMAR – UPMC of Prof.
B. Kloareg in Roscoff: iota- and kappa carrageenan (Rhodophyta) (Barbeyron et al. 2000),
alginate and fucane (Phaeophyta) (Descamps et al. 2005).
Bioinformatic analysis based on the similarity comparison of the sulphatase genes found in all
available Planctomycete genomes and in the genome of the marine bacteroidetes

Research Aims
19
Zobellia galactanivorans are carried out to identify the number of sulphatase genes
responsible for the degradation of polysaccharides.
If expression or regulation of some sulphatases under special conditions is detectable, an in
depth characterisation of the catalysed reaction would be interesting also for industrial use,
e.g. for the food industry where sulphatases are needed that selectively remove part of the
sulphate groups, thereby stabilising the consistency of the chemical network. Such approaches
are used to thicken cream, aerate mousse and suspend the herbs in salad dressing
(www.degussa.com).

Research Aims
20
2.3 Design of the whole genome array of Rhodopirellula baltica
In 1999, Lander called DNA microarrays 'arrays of hope' because for the first time it was
possible to take “global views” of biological processes at the genome level (Lander 1999).
This statement nicely matches the expectations linked to the whole genome array of the
marine planctomycete Rhodopirellula baltica. The design of the array was driven by the
ambition to understand more about the role of R. baltica, representing the phylum
Planctomycetes as one of the key players in the carbon cycle and about its adaptation
mechanisms to changing environmental conditions and cell biology. Microarray-mediated
expression profiling should also to help reveal regulation-patterns of the large number of
genes encoding hypothetical proteins.
A conventional whole genome array targeting all 7325 genes annotated within the R. baltica
genome (Glöckner et al. 2003) is set up within the framework of the Network of Excellence
Marine Genomics Europe (MGE). So far, no universal microarray hybridisation protocol was
available (Li et al. 2002). Therefore, every slide type or chemical surface, every new target
preparation process and labelling method, hybridisation and washing buffer composition as
well as hybridisation time and conditions have to be optimised corresponding to the new
application. Moreover, the specific proteinaceous cell wall of R. baltica makes it difficult to
apply common RNA extraction, cDNA-transcription and labelling methods, and thus requires
substantial methodological adaptations.
In parallel, a pipeline for microarray data processing and data storage has to be implemented.
Different microarray data analysis software tools are reviewed with the focus on usability and
calculation transparency. Moreover, a database for storing the data in-house needs to be
initiated. The validation/quality of the resulting data during the optimisation process needs to
be checked by the spotted positive and negative controls and consistency with published
results of proteome studies (Gade et al. 2003; Gade et al. 2005).

Results and Discussion
21
3 RESULTS AND DISCUSSION Manuscripts and Publications
3.1 Transcriptional response of the marine planctomycete Rhodopirellula baltica SH1T to
changing environmental conditions
Manuscript submitted to BMC Genomics
Authors: Patricia Wecker, Christine Klockow, Christian Quast, Andreas Ellrott, Phillip
Langhammer, Jens Harder and Frank Oliver Glöckner
Contribution: Idea and concept, development of the manuscript under editorial cooperation of
F.O.G
3.2 The Rhodopirellula baltica life-cycle: Growth-dependent expression profiling of the
complete genome with microarrays
Manuscript in preparation
Authors: Patricia Wecker, Marga Schüler, Andreas Ellrott, and Frank Oliver Glöckner
Contribution: Idea and concept, development of the manuscript under editorial cooperation of
M.S. and F.O.G

Results and Discussion
22
3.3 Highly enantioselective sec-alkyl sulfatase activity of the marine planctomycete
Rhodopirellula baltica shows retention of configuration
Published in: Angewandte Chemie International Edition 44: 2 –4 (2005)
Authors: Sabine R. Wallner, Marga Bauer, Chris Würdemann, Patricia Wecker, Frank Oliver
Glöckner, and Kurt Faber
Contribution: Selection of the media composition, Cell culturing and growth, Cell harvesting
and storage
3.4 Fosmids of novel marine Planctomycetes from the Namibian and Oregon coast upwelling
systems and their cross-comparison with planctomycete genomes
Published in: The ISME Journal 1, 419-435 (2007)
Authors: Dagmar Woebken, Hanno Teeling, Patricia Wecker, Alexandra Dumitriu, Ivaylo
Kostadinov, Rudolf Amann, Edward F DeLong and Frank Oliver Glöckner
Contribution: Microarray experiments and Data analysis

Results and Discussion
23
3.1 Transcriptional response of the model planctomycete Rhodopirellula baltica SH1T to changing environmental conditions
Authors: Patricia Wecker, Christine Klockow, Christian Quast, Andreas Ellrott, Phillip
Langhammer, Jens Harder, and Frank Oliver Glöckner
Manuscript submitted to BMC Genomics

Results and Discussion
24
Transcriptional response of the model planctomycete Rhodopirellula baltica SH1T to changing environmental conditions
Patricia Wecker1,3, Christine Klockow1,3, Andreas Ellrott1, Christian Quast1, Philipp
Langhammer2, Jens Harder2, Frank Oliver Glöckner1,3§ 1 Microbial Genomics Group, Max Planck Institute for Marine Microbiology, Microbial
Genomics Group, Celsiusstr. 1, 28359 Bremen, Germany 2 Department of Microbiology, Max Planck Institute for Marine Microbiology, Microbiology,
Celsiusstr. 1, 28359 Bremen, Germany 3 Jacobs University Bremen gGmbH, Campusring 1, 28759 Bremen, Germany §Corresponding author
Keywords: Planctomycetes, Rhodopirellula baltica, marine, expression profiling,
hypothetical genes, shift experiments, whole genome array, microarray, heat shock, cold
shock, high salinity
Email addresses:
P. Wecker: [email protected]
C. Klockow: [email protected]
A. Ellrott: [email protected]
C. Quast: [email protected]
P. Langhammer: [email protected]
J. Harder: [email protected]
F.O. Glöckner: [email protected]

Results and Discussion
25
Abstract
Background: The marine model organism Rhodopirellula baltica SH1T was the first
Planctomycete to be completely sequenced. The genome analysis predicted a complex life
style and a variety of genetic opportunities to adapt to the marine environment. Its adaptation
to environmental stressors was studied by transcriptional profiling using a whole genome
microarray.
Results: Stress responses to salinity and temperature shifts were monitored in time series
experiments. Chemostat cultures grown on mineral medium at 28°C were compared to
cultures that were shifted to either elevated (37°C) or reduced (6°C) temperatures as well as
high salinity (59.5‰) and observed over 300 min. Heat shock showed the induction of
several known chaperone genes. Cold shock altered the expression of genes in lipid
metabolism and stress proteins. High salinity resulted in the modulation of genes coding for
compatible solutes, ion transporter and morphology. In summary, over 3000 of the 7325
genes were affected by temperature and/or salinity changes.
Conclusions: Transcriptional profiling confirmed that R. baltica is highly responsive to
its environment. The distinct responses gave new insights into the complex adaptation
machinery of an environmentally relevant marine bacterium. Our transcriptome study and
previous proteome data suggest a set of genes of unknown functions that are most probably
involved in the global stress response. This work lays the foundation for further bioinformatic
and genetic studies which leads to a comprehensive understanding of the biology of a marine
Planctomycete.

Results and Discussion
26
Background
Marine ecosystems, covering approximately 71% of the Earth’s surface, host the majority of
biomass and contribute significantly to global cycles of matters and energy. Microorganisms
are known to be the ’gatekeepers‘ of these processes, and any insights into their lifestyle and
fitness enhances our ability to monitor, model and predict the effect of global changes.
Nevertheless, the specific knowledge about their functions is still sparse. The ‘genomic
revolution’ [1] has opened the door to investigate their genetic potential and activity on the
molecular level.
A particularly interesting representative of the marine picoplankton community is
Rhodopirellula baltica, a free-living bacterium which was isolated from the water column of
the Kiel Fjord (Baltic Sea) [2]. R. baltica belongs to the phylum Planctomycetes, a broadly
distributed group of bacteria, whose members can be found in terrestrial, marine and
freshwater habitats [3-7], but also in extreme environments like hot springs [8], marine
sponges [9] and the hepatopancreas of crustaceans [10].
In terms of cell biology all Planctomycetes share several morphologically unique properties,
such as a peptidoglycan-lacking proteinaceous cell wall [11, 12], intracellular
compartmentalization [13] and a mode of reproduction via budding. The latter results in a
cell cycle that is characterized by motile and sessile morphotypes similar to Caulobacter
crescentus [14-17]. A specific holdfast substance produced by sessile cells allows R. baltica
to attach to macroscopic detrital aggregates (marine snow) [3, 18].
At present, four planctomycete genomes are currently available [19], of these the genome of
R. baltica is the only one completely closed [16]. The genome was found to be 7,145,576
bases in size and codes for 7325 open reading frames (ORFs) plus 72 RNA genes. Originally
for only 45% of the ORFs particular functions could be assigned. This means that over 55%
of all proteins in the genome remain functionally uncharacterized. They were referred to as
’hypothetical proteins‘ or with the affix ’conserved‘ in case of a wider phylogenetic
distribution [20]. A subset of these conserved hypothetical proteins is specific for
Planctomycetes [19]. It seems likely that some of these genes code for the unique
planctomycetal cellular characteristics and metabolic traits.
The availability of the genome information triggered several key post-genomic studies
including studies of the proteome [21-26] enzyme activity [27] and protein crystallization
[28].

Results and Discussion
27
In summary, these studies confirmed the hypothesis of Glöckner et al. [16] that R. baltica is a
polysaccharide degrader. It seems that R. baltica is gaining carbon and energy from the
decomposition of complex heteropolysaccharides originally produced by algae in the photic
zone while slowly sedimenting with the marine snow.
Marine microorganisms like R. baltica are exposed to rapidly changing environmental
conditions, e.g. varying temperature, salinity, irradiance and oxygen concentration. Typically,
sudden changes of these environmental conditions induce a stress response in the exposed
planktonic community characterized by a distinct change in their gene expression pattern.
This stress response enables the organisms to protect vital processes and to adapt to the new
condition, which has been shown for a set of organisms in different environments, e.g. for
Shewanella oneidensis [29, 30], Pseudomonas aeruginosa [31],
Desulfovibrio vulgaris Hildenborough [32], Xylella fastidiosa [33], Synechocystis sp. [34]
and Yeast [35].
To gain insights into the stress responses of R. baltica with respect to salinity and
temperature the first whole genome array for R. baltica, which is also the first planctomycete
microarray, was established and applied. The reported data will serve as a resource to expand
our understanding of the physiological and transcriptional response of R. baltica to the wide
range of changing environmental conditions, a free-living marine bacterium is exposed to.
Results and Discussion
Overview
54 different total RNA samples were analyzed by whole-genome microarray hybridization. In
summary, for heat shock 2372 genes, for cold shock 922 genes and for salt stress 1127 genes
exhibited significant differential expression at one or more of the five time points compared
to the reference samples (FIGURE 1 i; ii & iii). With only 45% of functionally annotated
genes in the genome it is not surprising that most of the differentially expressed genes were
hypothetical or conserved hypothetical proteins. The complete list of the differentially
expressed genes for each shift experiment and time point is available in the ADDITIONAL
FILE 1.
A COG function could be assigned to only 32% of the regulated genes in the heat and cold
shock experiments (FIGURE 2 i & ii) compared to 37% in the salt stress (FIGURE 2 iii).
This is in line with the 36% (2661 genes) COG functional class designations in R. baltica. A

Results and Discussion
28
striking feature of the expression profiles displayed is that a large fraction of the genome
responded in a stereotypical manner to all three stressful conditions tested. In summary 152
genes are up- or down-regulated at any time point for all stressors. Out of the 152 genes 62
are induced and 90 are repressed (TABLE 2 & TABLE 3). 49% of the induced and 61% of
the repressed genes were annotated as hypothetical proteins. The Venn diagrams shown in
FIGURE 3 provide an overview of the specific and common genes of the three stress-specific
responses. To identify co-regulated patterns of gene expression, we classified all
differentially expressed genes of all three stress expressions into 30 k-means clusters based
on their expression log ratio. The cluster data are available in the ADDITIONAL FILE 2.
Cluster 1, 3 and 4 show a similar response to the specific environmental changes, called
environmental stress response (ERS) over all experiments. Cluster 2, 4, 5, 7, 15 and 22
revealed genes that are reacting only to one or inverse to a specific environmental factor.
Experimental design and assessment of array data quality
The experimental conditions were chosen to mimic the natural environment of R. baltica with
the constraint that the stress response must be detectable by the microarray technology and
should therefore be significantly pronounced. In contrast to steady-state or single-time-point
studies, time series experiments can provide useful data for generating computational models
of stress response pathways [29].
The negative, positive and stringency controls printed on the array gave no indications for
unspecific hybridizations. Co-hybridizations of two cDNA samples prepared from the same
total cellular RNA (self-self hybridization) suggested that genes with an expression log ratio
value greater than 1.5 and smaller than -1.0 for heat and cold shock, respectively, could be
regarded as differentially expressed. For the salt stress log ratio values over 1.2 and
below -1.0 were considered as significant.
Effect of stress on Rhodopirellula baltica
No growth was detectable during stress conditions nor was any obvious morphology change
visible under the microscope. Under optimal conditions R. baltica has a doubling time of 10-
12 hours [22], indicating that physiological effects are not measurable during the short time
period of max. 5 hours the cells were exposed to stress.

Results and Discussion
29
Specific results of the shift experiments
Heat shock
In their natural environment R. baltica cells can be regularly exposed to higher temperatures
e.g. at the water surface. Therefore, R. baltica cells were rapidly shifted from 28°C to 37°C
and observed over a period of 300 min in the first experiment. This is approximately 9°C
above the optimal growth temperature reported by Schlesner et al. [2]. A higher temperature
would have killed the cells. The time series reveals a quick response of R. baltica to sudden
temperature up-shifts. In total 2372 genes are regulated out of which 1140 genes encode
hypothetical proteins. 390 genes (5%) were already regulated after 10 min. This number
increased to 750 genes (10%) after 300 min (FIGURE 1 i). The COG classes containing the
translation [J] and amino acid transport and metabolism [E] were the largest classes down-
regulated. Up-regulated genes were assigned to the COG classes replication, recombination
and repair [L], post-translation modification, protein turnover and chaperons [O],
transcription [K], secondary metabolites biosynthesis, transport and catabolism [Q], cell
envelope biogenesis, outer membrane [M] and general function prediction [R] (FIGURE 2 i).
Taking a closer look at the response of R. baltica to thermal stress revealed the induction of
many known heat shock proteins (Hsp): ClpB (RB6751), GroEL (RB8970), DnaJ (RB8972),
GrpE (RB8974), Hsp20 (RB10279, RB10283), dnaK (RB9105), as well as the ATP-
dependent protease ClpP (RB9103). Also up-regulated are the chaperonins Cpn10 (RB10627
and RB8969) and Cpn60 (RB8966) as well as the cell division protein FtsH (RB2966)
(Cluster 4 in Additional files 2). Previous proteomic studies found the proteins of these genes
as well, except FtsH, DnaJ and Hsp20 [25, 26].
The regulation of the heat shock response in R. baltica involves many transcriptional
regulators. TetR (RB838) and GntR (RB1862, RB8695) showed an up-regulation, which
confirmed their important role in early heat shock response [36]. A gene encoding for GntR
was also found in the environment on the planctomycete fosmid 3FN from the metagenome
study of the Namibian coast [19]. In E. coli the induction of the majority of heat shock genes
results from a rapid and transient increase in the cellular level of an alternative 32-kDa sigma
factor (sigma32) encoded by rpoH and by alternative sigma factors E and 54, encoded by
rpoE (RB2302) and rpoN (RB6491), respectively [29]. Although, all genes are present in the
R. baltica genome, they were not regulated, suggesting a significantly different response
cascade.

Results and Discussion
30
R. baltica also showed an extracyctoplasmic stress response. The gene coding for SecA
(RB11690), belonging to the Sec system, was induced. This indicated an activation of the
protein translocation most probably from the riboplasma to the paryphoplasm or maybe also
to the medium. Proton channels were induced and R. baltica inhibited the motility. The
flagellar motor switch protein (FliG - RB12502) was already down-regulated after 20 min.
Followed by the type 4 fimbrial assembly protein (pilC - RB11597) after 40 min.
Cold Shock
During sedimentation, e.g. attached to marine snow flocks, R. baltica needs to cope with
decreasing temperatures. To investigate the response to cold shock, R. baltica cells were
shifted from 28°C to 6°C and observed for a period of 300 min. 6°C was chosen for this
study because this is a common temperature in the Baltic Sea. Also, the temperature
difference of 22°C is generally regarded as standard for cold shock studies with bacteria [29,
37]. Compared to heat shock only one third (922) of the regulated genes were differentially
expressed. Out of these 922 regulated proteins, 391 genes (42%) encode for hypothetical
proteins. With 419 differentially expressed genes (6%) the cold shock response reached its
peak after 20 min and was decreasing afterwards (FIGURE 1 ii). In contrary to the heat shock
experiment it seemed that R. baltica needed approximately one hour to adapt to cold
conditions. Like other bacteria R. baltica responded to cold conditions with the up-regulation
of genes coding for stress response [COG class O], cell envelope and transport [M],
transcription factors and solute uptake. Genes for amino acid biosynthesis [E], protein fate
and synthesis [J] were down-regulated (FIGURE 2 ii) [30].
The transcriptional activity was regulated by the up-regulation of diverse RNA polymerase
sigma factors, like rpoD (RB6780) and sigK (RB1392). A homolog of rpoD (RB6780) was
also found on the planctomycete fosmid 13FN [19]. 20 min after the exposure of R. baltica to
cold conditions it started to express genes that change the cytoplasmic membrane
composition and fluidity as well as its morphology. The alteration of the lipid composition in
the cold has been previously reported from other microorganisms [38]. In R. baltica genes
coding for cell envelope (RB6114 and RB6895), transport (RB4870), lipid metabolism
(RB316) and 18 genes coding for membrane proteins were repressed after 20 min.
Furthermore, R. baltica repressed genes involved in sporulation oppB (RB12861) and O-
antigen flippase (RB2503), flaA (RB4454) and pilus assembly (RB4061 and RB5478),
leading to a reduced motility and budding ability. Genes associated to the amino acid

Results and Discussion
31
biosynthesis, especially to the synthesis and fate of glutamine (RB4269) and glutamate
(RB5653) were also affected. Latter are shown to be translated [25, 26]. A
glycosyltransferase (RB12831) and glycosidases (RB2988, RB2990 and RB2991) are up-
regulated at 300 min probably for remodeling of the cell wall.
Although incorrect protein folding at low temperature is less expected, chaperons and
proteases are required to deal with intracellular protein perturbations [30, 39]. This was
confirmed by the induction of GroEL (RB8970) [25, 26] and htrA-protease (RB12752) in
R. baltica as well. One of the most prominent responses of microorganisms to cold shock is
the induction of cold shock proteins. However, the two annotated cold shock proteins of class
I (CspA - RB4681 and Cspl - RB10009) were not regulated in the genome of R. baltica [40,
41]. Consequently, the stabilization of RNA seems to employ a different set of proteins than
in E. coli.
High salinity
As a marine organism, R. baltica needs to adjust to the haline stratification of the Baltic Sea
[42, 43]. While wandering through the water column R. baltica cells are exposed to high, as
well as changing concentrations of dissolved salts. In general, an osmotic up-shift forces
bacteria to change their physiology by activating or deactivating specific enzymes or
transporters, in order to maintain water balance [44]. To gain an understanding of the genetic
events that occur during the early stages of salt adaptation, R. baltica cells were subjected to
salt up-shock from 17.5‰ salinity (Baltic Sea) to 59.5‰ (hyper saline environment). Former
experiments showed that R. baltica is able to grow between 4.2‰ and 59.5‰ salinity [2] and
does not grow at the next tested salinity over 90‰ (Wohlrab, unpublished data).
In total, 1127 genes showed differences in gene expression over the whole time series. 656 of
these genes (58%) were annotated as hypothetical proteins. The salt up-shock results
indicated an increase in the number of regulated genes over time. After 10 min 61 genes (1%)
were regulated. The largest number (543 – 8%) was observed at 300 min (FIGURE 1 iii).
R. baltica cells seem to adapt slowly to high salt concentration. This might be a result of the
cell compartmentalization and the ability of R. baltica to temporarily resist higher salt
concentration without significant adaptations.
The response of R. baltica to salt stress includes repression of genes associated with the COG
classes: induction of amino acid transport and metabolism [E], lipid metabolism [I],
transcription [K], translation process [J]. Induced were genes involved in the already

Results and Discussion
32
mentioned classes of the heat shock experiment: [O], [M] and [L], but also in the energy
production [C] and cell division and chromosome partitioning [D] classes (FIGURE 2 iii).
Similar to other bacteria, R. baltica accumulated glutamate and trehalose as cytoplasmic
osmoprotectants in response to osmotic stress [45]. Glutamate dehydrogenase (RB6930)
showed an up-regulation already after 10 min and was also present in the proteome [26].
Trehalose synthetase (treS - RB519) was induced after 60 min. Cysteine, as a general
protective component, was only needed in the first hour in elevated concentrations. The
synthesis was repressed after 1 hour (RB4386).
The accumulation of compatible solutes is a widely distributed mechanism coping with
environmental conditions [45, 46]. In R. baltica 74 planctomycetes-group-specific genes are
annotated as hypothetical proteins carrying a Domain of Unknown Function (DUF) named
1559 [19]. This domain belongs to a new family of solute binding proteins (PF07596) [47],
and was also found on the planctomycete fosmid 8FN [19]. Nine of these genes were up-
regulated during the first hour in the cold and salt shock experiments, as opposed to the heat
shock experiment where 16 of these genes were down-regulated. In vitro experiments have
shown that some of these compatible solutes also possess general protein stabilizing
properties beside their osmoprotective property [48]. These homologue proteins do not play
an integral role in the transport process per se, but probably serve as receptors to trigger or
initiate translocation of the solute through the membrane by binding to external sites of the
integral membrane proteins of the efflux system. In addition, some solute-binding proteins
function in the initiation of sensory transduction pathways [47].
R. baltica up-regulated an efflux pump (RB7603) and a Na+/H+ antiporter (RB1433) 300 min
after salt shift. Both may play a role in the active export of salt ions out of the cells. Quinone
oxidoreductase-like protein (RB10967), induced after 40 min, had been implicated in
respiration-coupled Na+ efflux as also shown in D. vulgaris [32]. Regulatory proteins like
sigma-54 factor rpoN (RB6491), rpoA (RB12626) and rfaY (12251) were down-regulated.
rpoN and rpoA were found to be translated [25, 26]. R. baltica inhibited the genes for cell
division (soj - RB2291) and chromosome segregation (SMC - RB6065) after 60 min salt
stress, as well as diverse transferases (RB12080, RB8898, RB12690, RB2498, RB8222,
RB9617) involved in the cell envelope modification. Interestingly, the pilin transport
apparatus and the thin-pilus basal body (pilM -RB2860 and pilT - RB12773) were induced
after one hour, and principle pilus associated adhesion (pilC - RB12781) and pilB
(RB12774). Also induced were genes coding for biopolymer transport proteins (exbB -
RB12053 and exbD - RB12055). A homolog to exbD was annotated on the planctomycete

Results and Discussion
33
fosmid 3FN [19]. It is known from other organisms that genes encoding the flagellar and
chemotaxis systems are up-regulated to move away from the stressful cations [32]. However
none of the flagellar genes were regulated and the genome does not harbor any essential
chemotaxis genes except cheY [16]. Notably, the survival protein (SurE -RB10258) and two
genes coding for the mechanosensitive ion channel (MscS - RB12279 and RB10255) are
induced. Latter provide protection against hypo-osmotic shock, responding both to stretching
of the cell membrane and to membrane depolarization [49]. Genes in Cluster 22
(ADDITIONAL FILE 2) seemed to be significantly affected by the salt stress.
Common stress response
R. baltica showed a common stress response to all three tested environmental factors. Several
known general stress genes were induced, like genes coding for the manganese-containing
catalase (RB10727), which is also present on the proteome [23, 25, 26]. Ferritin and Dps
(RB4433) or pyridoxamine 5'-phosphate oxidase (RB4438) belong to a general stress cluster
(RB4432-4438) and were initially described by Hieu et al. [26]. Thioredoxin (RB10378)
could serve as an electron donor for the up-regulated methionine sulfoxide reductase gene
(msrB - RB2268) [50, 51]. The genes could be regulated via rpoN based of the proposed
upstream sigma 54-dependent promoter (RB10378) [52].
To eventually cope with reactive oxygen species (ROS), typically present under stressful
conditions [51], the nitrogen fixation protein (nifU - RB3596) was induced. NifU is involved
in the biosynthesis and repair of ROS scavenging iron-sulfur clusters. Finally, the induced
peptidase M50 (RB6092) could have been transcripted to regulate stress responses,
sporulation, cell division, and cell differentiation [53].
Genes involved in the fatty acid metabolism were repressed under all conditions in R. baltica.
Examples are genes like CoA-acyl carrier protein transacylase (fabD - RB314), the acyl
carrier protein (acpP - RB318) and the fabB (RB320) gene.
Interestingly, the machinery for rearrangement and the interchange of the genetic material
was induced under all three stressful conditions. It seems to play an important role in the
organism’s long-term adaptation. R. baltica harbors 81 non-randomly distributed
transposases in its genome. Noteworthy, under heat stress three times more transposase genes
was up-regulated than under cold stress and twice as many as under salt stress. Shared
induction shows five IS3/IS911, three ISXo8, two putative transposases (RB170, RB5888,
RB11749, RB11802, RB12940, RB2186, RB9907, RB12239, RB934 and RB7389), and one

Results and Discussion
34
integrase (RB11750). Rearranging the genome to select the most efficient gene combination
has been described as a common way to adapt quickly to extreme environments [36].
Relaxed DNA may also be required to get better access to the gene regions for increased
expression. This is indicated by the repressed histone-like DNA-binding protein (RB6276).
In line with an alternative global sensing and regulation system initially proposed by
Glöckner et al. [16], a common sensing and regulation was found. R. baltica contains 37
genes belonging to the extracytoplasmic function (ECF) subfamily of sigma 70 [54]. The
genes RB138, RB13241 and RB10049 are up-regulated under all three stressful conditions.
Studholme et al. [47] suggests that ECF-factor RB10049 is the regulator for the conserved
hypothetical protein RB10051. The conserved domain belongs to a new group of proteins
that share novel domains referred to as planctomycete-specific (PSD) or planctomycete-
specific cytochrome C (PSC). RB10051 contains the PSD1 (DUF1553 - PF07587) and PSC2
(DUF1549 - PF07583) domains suggestion a function in redox reactions [47]. Each domain is
represented 41 times in the whole genome of R. baltica [19].
Additionally, at 300 min the ECF-sigma factor RB138 was up-regulated together with
serine/threonine protein kinase (RB140). Protein kinases are believed to be involved in the
stress response [38, 55]. The serine/threonine protein kinase (RB12942) and two histidine-
kinases (RB4511 and RB10330) were up-regulated during heat shock. Whereas, under cold
shock only one serine/threonine kinase (RB8505) was induced. Under salt stress a histidine-
kinase (RB13122) and three two-component systems (RB5780, RB12952 and RB13118)
were induced.
Finally, the ECF-sigma factor RB1790 was up-regulated, but only under high salinity
conditions. In summary, the results confirmed that ECF sigma factors, as well as two-
component systems, are heavily involved in stress sensing and regulation of R. baltica. The
importance of these genes in the natural environment is indicated by the presence of a
homolog to RB12952 on the planctomycete fosmid 6N14 [19].
The down-regulation of genes associated with the ribosomal machinery (55%) was observed.
During heat shock and high salinity they were permanently repressed, whereas under cold
shock they were only repressed within the first hour. Out of 51 ribosomal proteins in the
whole genome, 18 genes encoding proteins of the small- and large subunit (RB1233,
RB12821, RB12824, RB12839, RB7117, RB7837 - RB7841, RB7849, RB7850, RB7852,
RB7854, RB7856, RB7857, RB7859 and RB7899) were repressed. Additionally, a set of
genes involved in RNA metabolism, protein synthesis, as well as the only translation
elongation factor (EF-Tu - RB7894) in R. baltica were repressed. The genes for the

Results and Discussion
35
conserved hypothetical protein RB12818 and the hypothetical protein RB12837 were co-
regulated which indicates an association with the translation machinery. The repression of the
ribosomal genes, along with a large set of genes involved in RNA metabolism, protein
synthesis, cell growth (Cluster 1 ADDITIONAL FILE 2), has been reported as a general
feature of the environmental stress responses (ESR) [35]. It is assumed that they are acting as
stress sensors [56]. This coincides nicely to the induction of the ribosomal proteins at
300 min under cold shock conditions. Recovery and ongoing adaptation of R. baltica was
further supported by the up-regulation of the ribosomal-binding factor rbfA (RB5503), which
is, besides csdA, required for optimal growth at low temperatures [57].
Hypothetical proteins
Approximately 50% of the regulated genes have no known function in each of the three
environmental stress experiments. Some of them even share a similar expression profile
(TABLE 2 and TABLE 3). We propose that some of these genes are involved in cell
morphology changes, stress sensing and regulation. The low number of known transcriptional
regulators (2.4%) in the genome of R. baltica [54], and the fact that most of the essential
pathways are not organized in operon structures [16] support the hypothesis of novel kinds of
global regulation mechanisms. Candidates are hypothetical proteins that carry regulatory
domains, like the FHA domain in RB1789 or a putative transcriptional regulatory domain in
RB9999. RB11766 might regulate the gene next to it, which is a so called giant gene
(RB11769) [58]. This giant gene carries a novel peptide motif that is most likely involved in
cell morphology changes [47]. The importance of the hypothetical proteins RB11505,
RB10954, RB10956 and RB10958 was further confirmed by their presence on the proteome
gels of Hieu et al. as well as Gade et al. [23, 25, 26]. The latter three of these genes were
claimed to be one of the most abundant proteins when R. baltica cultures are grown on
mineral medium.
Planctomycete special feature: Genes encoding sulfatases
The genome of R. baltica contains no less than 110 sulfatases. It is assumed that they are
involved in the recycling of carbon from complex sulfated heteropolysaccharides. Although,
the mineral medium does not contain any sulfated polysaccharide, we found 11 sulphatase
genes up- or down-regulated (TABLE 1) including one choline sulphatase (RB1205), seven
arylsulfatases (RB13148, RB1477, RB3403, RB406, RB5146, RB684 and RB9498), two

Results and Discussion
36
sulphatase genes without specificity (RB3956, RB5294), and one alkylsulfatase (RB11502)
within the different stress experiments. Furthermore, during life cycle experiments
(unpublished data) we found evidence that certain sulfatases are only regulated in specific
growth stages which indicates their involvement in the remodeling of the distinct
morphological features of R. baltica. RB1477, RB5294, RB9498 and RB11502 were
induced. We propose a RB9498 and RB11502 have an extracellular function and may be
involved in the formation of a protective substance.
Six sulfatase genes (RB406, RB684, RB1205, RB3403, RB5146 and RB13145) were
repressed after 300 min of heat shock. They could have been involved in the rearrangement
of the cell wall formation, which consists of a protein sacculus with disulfide bonds [12]. In
summary, these results show again how diverse the role of the sulfatases might be and that
only a variety of different experiments will lead to more knowledge on the sulfatases.
Conclusion
This work presents the first transcriptome study of the environmental stress response of a
marine, free-living Planctomycete. Although R. baltica is an unusual organism in many
aspects, the stress responses to heat and cold shock as well to changing salinity were in line
with earlier results reported for other model organisms. Heat shock induced a set of
chaperons, likely to protect cellular proteins from denaturation and breakdown. Growth in the
cold can be followed by the induction of genes altering lipid metabolism. Salinity shift
resulted in the activation of a planctomycete-specific groups of genes, genes involved in
morphology change and the induction of an extracytoplasmatic stress response. All stressors
triggered the down-regulation of the ribosomal machinery, the up-regulation of transposases
to enhance the genetic activity and the induction of several ECF-sigma factors and two-
component systems. This confirms the hypothesis that R. baltica is regulating its gene
activity on a global- rather than on the operon level. Besides well characterized stress
response genes about 1000 genes of unknown function, constituting 13 % of the genes
predicted in the genome, were affected. Among them a set of genes most likely involved in
global regulation could be proposed. This in combination with proteome studies and the
presence of some of the genes in fosmid libraries confirmed the essential role of the vast
amount of genes with so far unknown functions in the adaptation of environmental organisms
to changing conditions. Noteworthy is the regulation of 11 sulfatases during stressful
conditions, which indicates that these genes are heavily involved in the core functions

Results and Discussion
37
exposed by R. baltica. The data presented lead to the conclusion that R. baltica’s rich
repertoire of genes is combined with a fine tuned regulation mechanism to best adapt to the
permanent changing conditions in its habitat. This work lays the foundation to get a deeper
understanding of the biology and behavior of Planctomycetes, a fascinating and diverse
phylum.
Methods
Bacterial growth conditions
For all experiments Rhodopirellula baltica SH 1T cells were grown as chemostat cultures in a
mineral medium containing 10 mM glucose as sole carbon and 1 mM ammonium chloride as
nitrogen source at 28°C [21]. Chemostate (Ø 13.5 cm x 25 cm, 1 l, Schott, modified by Ochs,
Bovenden) parameters were: pH 7.4, average dilution rate 0.75 ml/min and pO2 around
100%. The cultures had an OD600nm of 0.5 – 0.6 (corresponding to log phase). The cells were
harvested after 5 dwell times.
Sample collection, cell lyses, RNA Isolation and cDNA synthesis
After harvesting an aliquot was collected to serve as the time-zero reference. The culture
broth was collected in 500 ml tubes and swirled briefly in an ethanol-dry ice bath to rapidly
cool the cultures and prevent shifts in the RNA profile. Subsequently, the broth was
centrifuged at 6000 rpm for 20 min at 4°C (Beckman CoulterTM AvantiTM626 J-20XP, JA10
Rotor). The pellets were re-suspended in 0.1 M Tris-HCL and then re-centrifuged to cell
pellets that were shock-frozen in liquid nitrogen and stored at -80°C. Total RNA was isolated
using the protocol of the TRI Reagent® Kit by Ambion (Austin, USA). The purity and quality
of the extracted total RNA was checked with an Agilent 2100 Bioanalyzer (Agilent
Technologies, Palto Alto, USA) and gel electrophoresis. cDNA synthesis was performed
using the SuperScript direct cDNA labeling kit by Invitrogen (Karlsruhe, Germany)
according to the manufacturer´s instructions with random hexamers and unlabeled
dCTP/dUTP, followed by an three hours reverse transcription incubation step at 46°C and 3
min at 95°C to stop the RT reaction. To hydrolyze the RNA 0.1 M NaOH was added,
incubated at 65°C for 15 min and neutralized with 0.1 M HCL. The remaining cDNA was
precipitated over night at -20°C and the pellet was washed with 70% Ethanol.

Results and Discussion
38
cDNA was directly labeled using the PlatinumBrightTM nucleic acid labeling kit based on
KREATECH´s patented Universal Linkage System (ULS) (Biocat, Heidelberg, Germany)
according to the manufacture’s protocol.
Concentrations of RNA and cDNA were measured, and incorporation of the dyes Alexa 546
and Alexa 647 were checked, using a Nanodrop ND-1000 spectrophotometer (NanoDrop
Technologies, Wilmington, USA).
Experimental design and sample preparation
In three independent hybridizations for each experiments and time point the expression
profiles of cells that had undergone stress were compared with those of cells at time zero.
This means that for the array analysis each Alexa 647 labeled sample was compared with
Alexa 546 labeled time-zero samples. The data shown are based on the analysis of all three
replicates performed for each of the conditions.
For expression profiling and microscopic analysis samples were collected at 10, 20, 40, 60
and 300 min for all three stress experiments.
Heat shock from 28°C to 37°C
Cells grown continuously at 28°C were collected by centrifugation. An aliquot was removed
for RNA extraction and taken as the time zero reference for the heat, cold and salt stress
experiments. Aliquots were re-suspended in an equal volume of 37°C medium and returned
to 37°C for cultivation.
Cold shock from 28°C to 6°C
Cells grown continuously at 28°C were collected by centrifugation, re-suspended in an equal
volume of 6°C medium and returned to 6°C for cultivation.
Salt stress from 17.5‰ to 59.5‰ salinity
Similar to the heat and cold shock experiments, a R. baltica culture was grown in mineral
media with 17.5‰ salinity. Cells were harvested and aliquots were taken up in a mineral
media with a salinity of 59.5‰.

Results and Discussion
39
Whole Genome Array construction, hybridization and image analysis
The whole-genome oligonucleotides for R. baltica SH 1T (Pirellula AROS 630 Version 1.0)
were purchased from Operon (Cologne, Germany) and diluted to 20 μM concentration in
Micro Spotting Solution Plus spotting buffer (Telechem, Sunnyvale, USA). Spotting was
done with three replicates per gene and slide onto GAPS II aminosilane slides (Corning,
Schiphol-Rijk, Netherlands) using a SpotArray 24 spotting device (Perkin Elmer, Wellesley,
USA) together with 48 Telechem Stealth Pins (Telechem, Sunnyvale, USA). The arrays were
subsequently exposed at 245 nm and 360 mJ in the GS Gene Linker (Bio-Rad, München,
Germany), followed by incubation at 80°C for at least 3 h. Slides were stored at room
temperature in the dark until use.
Blocking, denaturing, hybridization, washing and drying of the slides with N2 were carried
out in an automated hybridization station HS400 (Tecan, Crailsheim, Germany). The spotted
arrays were blocked in prehybridization solution containing 250 mM NaCl, 5 mM Tris/HCl
at pH 8.0, 50% formamide, 0.5x SSC, 0.05% BSA, and 1% blocking reagent from Roche
Diagnostics, Mannheim, Germany for 45 min at 52°C. For hybridization at least 2 μg of
Alexa 546 dye-labeled and 2 μg of Alexa 647 dye-labeled total cDNA were combined and
taken up in a final volume of 100 µl DIG Easy Hyb hybridization solution (Roche
Diagnostics, Mannheim, Germany). After the blocking step the sample solution was applied
to the arrays, denaturized at 95°C for 3 min and hybridized under stringent conditions at
52°C for over 12 hours. After hybridization slides were washed at room temperature in
ULTRArray Low Stringency Wash Buffer (Ambion, Austin, USA) and dried by N2.
Signal detection and data analysis
Slides were scanned at a resolution of 5 μm using a ScanArray Express Microarray scanner
(Perkin Elmer, Wellesley, USA) with varied laser power and sensitivity level of the
photomultiplier tube (PMT) for each slide. The provided image analysis software ScanArray
Express Version 4.0 was used for automatic spot detection and signal quantification of both
fluorophores. Raw data were automatically processed using the in-house developed
microarray data analysis software tool MADA (www.megx.net/mada). First of all the spot
intensities were local background corrected (mean spot intensity minus mean spot
background intensity). Then signals were only assessed as positive if mean spot pixel
intensity was higher than the mean local background intensity plus two times the standard
deviation of the mean local background pixel intensity. Each gene is spotted in three

Results and Discussion
40
replicates. Spot replicates with poor quality were removed from the data set according to the
outlier test of MADA. For this test, first the standard deviation of all replicates is computed.
Second, one replicate is omitted and the standard deviation is recalculated, if the deviation
differs more than 50% from the previous deviation, the omitted replicate is regarded as an
outlier. This procedure is alternately repeated for all replicates
The expression is described through the ratio and intensity, where R is the fluorescence log
ratio of the experiment time point relative to the control condition (e.g. R = log2 (result of
channel 10min / result of channel control/reference)) and I is the log mean fluorescence
intensity (e.g. I = log10 (result of channel 10min x result of channel control/reference)).
Each data point represents a regulation factor (ratio) in a logarithmic scale for one gene
calculated from the positive replicates for a particular probe coming from two RNA pools
(reference and sample). Normalization was carried out by LOWESS fitting on an R-versus-I
plot with a smoothing factor of 0.5. Each time point of the time-series experiment was
hybridized independently three times. The expression data (ratio) of the three hybridizations
were combined to one expression data point (ratio) by average and the standard deviation was
calculated. Only ratios with a standard deviation less than 25% were taken as regulated.
Differentially expressed genes are determinate by a fixed threshold cut off method (i.e. a two-
fold increase or decrease) based on the self-self hybridization. Taking the same biological
sample the reference is labeled twice, once with Alexa 546 and once with Alexa 647 and the
variability between the two sets of measurements are calculated to estimate the experimental
noise. Ideally there should not be any variability and all expression points should have a ratio
close to zero. In reality this is never the case and thresholds on the basis of the distribution of
these data along the y-axis were defined for the further experiments.
Consequently, R. baltica genes detected with intensities resulting in ratios above or below
these thresholds can be regarded as up- or down-regulated.
Cluster analysis
Differentially expressed genes presenting the complete time course profile (10, 20, 40, 60
und 300 min) for all three experiments were clustered using the k-means clustering approach
(Euclidean distance metric, k = 30 clusters and 49 (max. 500) iterations) [59] with the
software tool Multiexperiment Viewer MeV Version 4.0.2 from the TM4 microarray
software suite [60]. Briefly, the clustering algorithm arranges genes into a given number of
clusters k according to their similarity in expression profiles across all of the array

Results and Discussion
41
experiments, such that genes with similar expression patterns are clustered together. The data
are graphically displayed in tabular format in which each row of colored boxes represents the
variation on transcript abundance for each gene and each column represents the variation in
transcript levels of every gene in a given mRNA sample, as detected on one array. The
variations in transcript abundance for each gene are depicted by means of a color scale, in
which shades of red represents increases and shades of green represent decrease in mRNA
levels, relative to the unstressed culture, and the saturation of the color corresponds to the
magnitude of the differences. A black color indicates no change in transcript level, and a grey
color represents missing data.
Genome tools
The genome of Rhodopirellula baltica was automatically re-annotated based on updated
homology searches (June 2005 - MicHanThi [61]). The updated annotation including all tool
results are publicly available [62]. JCoast [63] was used as a tool for the visualization,
interpretation, COG-assignment statistics and comparison of genomic data stored in GenDB
V2.2 [64]. The Venn diagrams were generated by BioVenn [65].
Microarray Datasets
Each microarray used in this study contained 7325 known or predicted R. baltica genes
according to Glöckner et al. [16]. A detailed description of the array can be found at the
NCBI´s Gene Expression Omnibus (GEO) database under accession number GPL7654. The
complete microarray datasets covering the expression of R. baltica cultures exposed to heat,
cold and high salinity, are public available in the GEO repository
(http://www.ncbi.nlm.nih.gov/geo/) under accession numbers GSE13769, GSE13856 and
GSE14075 [66].
List of abbreviations
COG Cluster of Orthologous Group of Genes
DUF Domain of Unknown Function
ECF Extra Cytoplasmic Function
ERS Environmental Stress Response
FHA Forkhead-associated
GEO Gene Expression Omnibus

Results and Discussion
42
R. Rhodopirellula
RB Rhodopirellula baltica
ROS Reactive Oxygen Species
ORF Open Reading Frame
Competing interests
The authors declare that they have no competing interests.
Authors contribution
PW conceived the study, initiated, conducted the experimental analysis, validated microarray
and optimized experimental steps, wrote the manuscript, did the statistical analysis and
analyzed the data.
CK was involved in the chemostat cultivation of R. baltica and statistical analysis of the data
AE wrote MADA the microarray analysis tool and designed the microarrays.
CQ was responsible for the automatic reannotation of the genome and set up the web access
PL and JH established the chemostat cultivation of R. baltica. JH supervised the chemostat
cultivation.
FOG contributed background information and was involved in writing and finishing the
manuscript.
All authors read and approved the final manuscript.
Acknowledgements
We thank Sylke Wohlrab for preliminary work and Matthias Gottschall for excellent
technical assistance in the chemostat cultivation.
The setup of the microarray and transcriptomic analysis of Rhodopirellula baltica was
supported by European Commission contract GOCE-CT-2004-505403.

Results and Discussion
43
References
1. Field D, Kyrpides N: The positive role of the ecological community in the genomic revolution. Microb Ecology 2007, 53(3):507-511.
2. Schlesner H, Rensmann C, Tindall BJ, Gade D, Rabus R, Pfeiffer S, Hirsch P: Taxonomic heterogeneity within the Planctomycetales as derived by DNA-DNA hybridization, description of Rhodopirellula baltica gen. nov., sp. nov., transfer of Pirellula marina to the genus Blastopirellula gen. nov. as Blastopirellula marina comb. nov. and emended description of the genus Pirellula. Int J Syst Evol Microbiol 2004, 54(Pt 5):1567-1580.
3. DeLong E, Franks D: Phylogenetic diversity of aggregate-attached vs. free-living marine bacterial assemblages. Limnol Oceanogr 1993, 38:924-934.
4. Chatzinotas A, Sandaa R, W S, Amann R, FL D, Torsvik V, Zeyer J, Hahn D: Analysis of broad-scale differences in microbial community composition of two pristine forest soils. System Appl Microbiol 1998, 21(4):579-587.
5. Neef A, Amann R, Schlesner H, Schleifer K: Monitoring a widespread bacterial group: in situ detection of Planctomycetes with 16S rRNA-targeted probes. Microbiology 1998, 144(12):3257-3266.
6. Vergin KL, Urbach E, Stein JL, DeLong EF, Lanoil BD, Giovannoni SJ: Screening of a fosmid library of marine environmental genomic DNA fragments reveals four clones related to members of the order Planctomycetales. Appl Environ Microbiol 1998, 64:3075-3078.
7. Crump BC, Armbrust EV, Baross JA: Phylogenetic analysis of particle-attached and free-living bacterial communities in the Columbia River, its estuary, and the adjacent coastal ocean. Appl Environ Microbiol 1999, 65(7):3192-3204.
8. Giovannoni SJ, Scabtach E, Castenholz RW: Isosphaera pallida, gen. and comb. nov., a gliding, budding eubacterium from hot springs. Arch Microbiol 1987, 147:276-284.
9. Pimentel-Elardo S, Wehrl M, Friederich AB, Jensen PR, Henschel U: Isolation of Planctomycetes from aplysina sponges. Aqua Micro Ecol 2003, 33:239-245.
10. Fuerst JA, Gwilliam HG, Lindsay M, Lichanska A, Belcher C, Vickers JE, Hugenholtz P: Isolation and molecular identification of planctomycete bacteria from postlarvae of the giant tiger prawn, Penaeus monodon. Appl Environ Microbiol 1997, 63(1):254-262.
11. König E, Schlesner H, Hirsch P: Cell wall studies on budding bacteria of the Planctomyces/Pasteuria group and on a Prosthecomicrobium sp. Arch Microbiol 1984, 138:200-205.
12. Liesack W, König H, Schlesner H, Hirsch P: Chemical composition of the peptidoglycan-free cell envelopes of budding bacteria of the Pirella/Planctomyces group. Arch Microbiol 1986, 145:361-366.
13. Fuerst AJ: Intracellular compartmentation in Planctomycetes. Annu Rev Microbiol 2005, 59:299-328.
14. Tekniepe BL, Schmidt JM, Starr MP: Life cycle of a budding and appendaged bacterium belonging to morphotype IV of the Blastocaulis-Planctomyces group. Curr Microbiol 1981, 5:1-6.
15. Fuerst JA: The Planctomycetes: emerging models for microbial ecology, evolution and cell biology. Microbiology 1995, 141 (Pt 7):1493-1506.

Results and Discussion
44
16. Glöckner FO, Kube M, Bauer M, Teeling H, Lombardot T, Ludwig W, Gade D, Beck A, Borzym K, Heitmann K et al: Complete genome sequence of the marine planctomycete Pirellula sp. strain 1. Proc Natl Acad Sci U S A 2003, 100(14):8298-8303.
17. Jacobs-Wagner C: Regulatory proteins with a sense of direction: cell cycle signalling network in Caulobacter. Mol Microbiol 2004, 51(1):7-13.
18. Crump B, Armbrust E, Baross J: Phylogenetic analysis of particle-attached and free-living marine bacterial assemblages. Appl Environ Microbiol 1999, 65:3192-3204.
19. Woebken D, Teeling H, Wecker P, Dumitriu A, Kostadinov I, DeLong EF, Amann R, Glöckner FO: Fosmids of novel marine Planctomycetes from the Namibian and Oregon coast upwelling systems and their cross-comparison with planctomycete genomes. ISME Journal 2007, 1(5):419-435.
20. Galperin MY, Koonin EV: 'Conserved hypothetical' proteins: prioritization of targets for experiemental study. Nucl Acids Res 2004, 32(18):5452-5463.
21. Rabus R, Gade D, Helbig R, Bauer M, Glöckner FO, Kube M, Schlesner H, Reinhardt R, Amann R: Analysis of N-acetylglucosamine metabolism in the marine bacterium Pirellula sp. strain 1 by a proteomic approach. Proteomics 2002, 2(6):649-655.
22. Gade D, Stührmann T, Reinhardt R, Rabus R: Growth phase dependent regulation of protein composition in Rhodopirellula baltica. Environ Microbiol 2005, 7(8):1074-1084.
23. Gade D, Gobom J, Rabus R: Proteomic analysis of carbohydrate catabolism and regulation in the marine bacterium Rhodopirellula baltica. Proteomics 2005, 5(14):3672-3683.
24. Gade D, Thiermann J, Markowsky D, Rabus R: Evaluation of two-dimensional difference gel electrophoresis for protein profiling. Soluble proteins of the marine bacterium Pirellula sp. strain 1. J Mol Microbiol Biotechnol 2003, 5(4):240-251.
25. Gade D, Theiss D, Lange D, Mirgorodskaya E, Lombardot T, Glöckner FO, Kube M, Reinhardt R, Amann R, Lehrach H et al: Towards the proteome of the marine bacterium Rhodopirellula baltica: Mapping the soluble proteins. Proteomics 2005, 5(14):3654-3671.
26. Hieu CX, Voigt B, Albrecht D, Becher D, Lombardot T, Glöckner FO, Amann R, Hecker M, Schweder T: Detailed proteome analysis of growing cells of the planctomycete Rhodopirellula baltica SH1T. Proteomics 2008, 8(8):1608-1623.
27. Wallner S, Bauer M, Würdemann C, Wecker P, Glöckner FO, Faber K: Highly enantioselective sec-alkyl sulfatase activity of the marine planctomycete Rhodopirellula baltica shows retention of configuration. Angew Chem Intern Edition English 2004, 44(39):6381-6384.
28. Dabin J, Jam M, Czjzek M, Michel G: Expression, purification, crystallization and preliminary X-ray analysis of the polysaccharide lyase RB5312 from the marine planctomycete Rhodopirellula baltica. Acta Crystallogr Sect F Struct Biol Cryst Commun 2008, 1(64):224-227.
29. Gao H, Wang Y, Liu X, Yan T, Wu L, Alm E, Arkin A, Thompson DK, Zhou J: Global transcriptome analysis of the heat shock response of Shewanella oneidensis. J Bacteriol 2004, 186(22):7796-7803.
30. Gao H, Yang ZK, Wu L, Thompson DK, Zhou J: Global transcriptome analysis of the cold shock response of Shewanella oneidensis MR-1 and mutational analysis of its classical cold shock proteins. J Bacteriol 2006, 188(12):4560-4569.

Results and Discussion
45
31. Aspedon A, Palmer K, Whiteley M: Microarray analysis of the osmotic stress response in Pseudomonas aeruginosa. J Bacteriol 2006, 188(7):2721-2725.
32. Mukhopadhyay A, He Z, Alm EJ, Arkin AP, Baidoo EE, Borglin SC, Chen W, Hazen TC, He Q, Holman H-Y et al: Salt stress in Desulfovibrio vulgaris Hildenborough: an integrated genomics approach. J Bacteriol 2006, 188(11):4068-4078.
33. Koide T, Vencio RZN, Gomes SL: Global gene expression analysis of the heat shock response in the phytopathogen Xylella fastidiosa. J Bacteriol 2006, 188(16):5821-5830.
34. Kanesaki Y, Suzuki I, Allakhverdiev SI, Mikami K, Murata N: Salt stress and hyperosmotic stress regulate the expression of different sets of genes in Synechocystis sp. PCC 6803. Biochem and Biophys Res Comm 2002, 290(1):339-348.
35. Gasch AP, Spellman PT, Kao CM, Carmel-Harel O, Eisen MB, Storz G, Botstein D, Brown PO: Genomic expression programs in the response of yeast cells to environmental changes. Mol Biol Cell 2000, 11(12):4241-4257.
36. Tachdjian S, Kelly RM: Dynamic metabolic adjustments and genome plasticity are implicated in the heat shock response of the extremely thermoacidophilic Archaeon Sulfolobus solfataricus. J Bacteriol 2006, 188(12):4553-4559.
37. Jones PG, Cashel M, Glaser G, Neidhardt FC: Function of a relaxed-like state following temperature downshifts in Escherichia coli. J Bacteriol 1992, 174:3903-3914.
38. Coker J, DasDarma P, Kumar J, Muller JA, DasSarma S: Transcriptional profiling of the model Archaeon Halobacterium sp. NRC-1: Responses to changes in salinity and temperature. Sal Systems 2007, 3(6).
39. Sowell SM, Norberck AD, Lipton MS, Nicora CD, Callister SJ, Smith RD, Barofsky DF, Giovannoni SJ: Proteomic Analysis of stationary phase in the marine Bacterium "Candidatus Pelagibacter ubique". Appl Environ Microbiol 2008, 74(13):4091-4100.
40. Derzelle S, Hallet B, Ferain T, Delcour J, Hols P: Improved adaptation to cold-shock, stationary phase, and freezing stresses in Lactobacillus plantarum overproducing cold-shock proteins. Appl Environ Microbiol 2003, 69(7).
41. Phadtare S: Recent developments in bacterial cold-shock response. Curr Issues Mol Biol 2004, 6:125-136.
42. Ott J: Meereskunde. Stuttgart: Ulmer Verlag; 1996. 43. Rheinheimer G: Meereskunde der Ostsee. Berlin: Springer Verlag; 1996. 44. Kempf B, Bremer E: Uptake and synthesis of compatible solutes as microbial
stress response to high-osmolarity environments. Arch Microbiol 1998, 170:319-330.
45. Roeßler M, Müller V: Osmoadaptation in bacteria and archaea: common principles and differences. Environ Microbiol 2001, 3(12):743-754.
46. Kunte: Osmoregulation in Bacteria: Compatible solute accumulation and osmosensing. Environ Chem 2006, 3:94-99.
47. Studholme DJ, Fuerst JA, Bateman A: Novel protein domains and motifs in the marine planctomycete Rhodopirellula baltica. FEMS Microbiol Lett 2004, 236:333-340.
48. Vargas C, Argandoña M, Reina-Bueno M, Rodriguez-Moya J, Fernández-Aunión C, Nieto JJ: Unravelling the adaptation responses to osmotic and temperature stress in Chromohalobacter salexigens, a bacterium with broad salinity tolerance. Saline Systems 2008, 4(14).
49. Pivetti CD, Yen M-R, Miller S, Busch W, Tseng Y-H, Booth IR, Saier MH, Jr.: Two families of mechanosensitive channel proteins. MMBR 2003, 67(1):66-85.

Results and Discussion
46
50. Zeller T, Klug G: Thioredoxins in bacteria: functions in oxidative stress response and regulation of thioredoxin genes. Naturwissenschaften 2006, 93(6):259-266.
51. Pereira PM, He Q, Xavier AV, Zhou J, Pereira IAC, Louro RO: Transcriptional response of Desulfovibrio vulgaris Hildenborough to oxidative stress mimicking environmental conditons. Arch Microbiol 2008, 189:451-461.
52. Studholme DJ, Dixon R: In silico analysis of the [sigma] 54-dependent enhancer-binding proteins in Pirellula species strain 1. FEMS Microbiol Lett 2004, 230(2):215-225.
53. Marchler-Bauer A, Anderson J, Derbyshire M, DeWeese-Scott C, Gonzales N, Gwadz M, Hao L, He S, Hurwitz D, Jackson J et al: CDD: a conserved domain database for interactive domain family analysis. Nucl Acids Res 2007, 35:D237-240.
54. Lombardot T, Bauer M, Teeling H, Amann R, Glöckner FO: The transcriptional regulator pool of the marine bacterium Rhodopirellula baltica SH 1T as revealed by whole genome comparisons. FEMS Microbiol Lett 2005, 242(1):137-145.
55. Mikami K, Kanesaki Y, Suzuki I, Murata N: The histidine kinase Hik33 perceives osmotic stress and cold stress in Synechocystis sp. PCC 6803. Mol Microbiol 2002, 46(4):905-915.
56. RA V, FC N: Ribosomes as sensors of heat and cold shock in Escherichia coli. Proc Natl Acad Sci U S A 1990, 87:5589-5593.
57. Thieringer HA, Jones PG, Inouye M: Cold shock and adaptation. Bioessays 1998, 20(1):49-57.
58. Reva O, Tümmler B: Think big -- giant genes in bacteria. Environ Microbiol 2008, 10(3):768-777.
59. Soukas A, Cohen P, Socci ND, Friedman JM: Leptin-specific patterns of gene expression in white adipose tissue. Genes Dev 2000, 14(8):963-980.
60. Saeed AI, Sharov V, White J, Li J, Liang W, Bhagabati N, Braisted J, Klapa M, Currier T, Thiagarajan M et al: TM4: a free, open-source system for microarray data management and analysis. Biotechniques 2003, 34(2):374-378.
61. Quast C: MicHanThi - Design and Implementation of a System for the Prediction of Gene Functions in Genome Annotation Projects. In: Technical Report. Edited by Informatik T-Z. Bremen: TZI-Bremen; 2007.
62. The reannotated Rhodopirellula baltica genome [http://gendb.mpi-bremen.de/gendb/BX119912]
63. Richter M, Lombardot T, Kostadinov I, Kottmann R, Duhaime M, Peplies J, Glöckner F: JCoast - A biologist-centric software tool for data mining and comparison of prokaryotic (meta)genomes. BMC Bioinformatics 2008, 9(1):177.
64. Meyer F, Goesmann A, McHardy AC, Bartels D, Bekel T, Clausen J, Kalinowski J, Linke B, Rupp O, Giegerich R et al: GenDB - an open source genome annotation system for prokaryote genomes. Nucl Acids Res 2003, 31(8):2187-2195.
65. Hulsen T, de Vlieg J, Alkema W: BioVenn - a web application for the comparison and visualization of biological lists using area-proportional Venn diagrams. BMC Genomics 2008, 9(1):488.
66. Barrett T, Troup DB, Wilhite SE, Ledoux P, Rudnev D, Evangelista C, Kim IF, Soboleva A, Tomashevsky M, Edgar R: NCBI GEO: mining tens of millions of expression profiles - database and tools update. Nucl Acids Res 2006, 35.
67. Wecker P, Schueler M, Ellrott A, Glöckner FO: The Rhodopirellula baltica life cycle: Growth-dependent expression profiling of the complete genome with microarrays. in prep 2009.
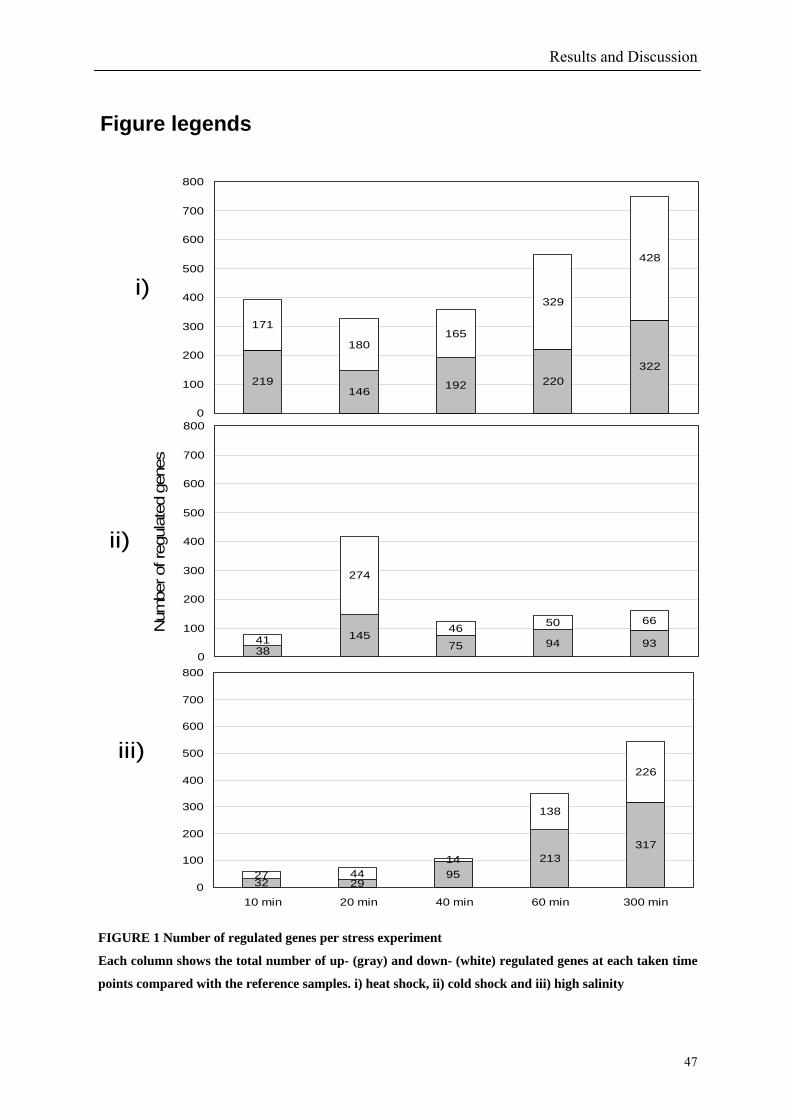
Results and Discussion
47
Figure legends
219146 192 220
322
171
180165
329
428
0
100
200
300
400
500
600
700
800
38145
75 94 9341
274
46 50 66
0
100
200
300
400
500
600
700
800
32 2995
213317
27 4414
138
226
0
100
200
300
400
500
600
700
800
10 min 20 min 40 min 60 min 300 min
i)
ii)
iii)
Num
ber o
f reg
ulat
ed g
enes
FIGURE 1 Number of regulated genes per stress experiment
Each column shows the total number of up- (gray) and down- (white) regulated genes at each taken time
points compared with the reference samples. i) heat shock, ii) cold shock and iii) high salinity
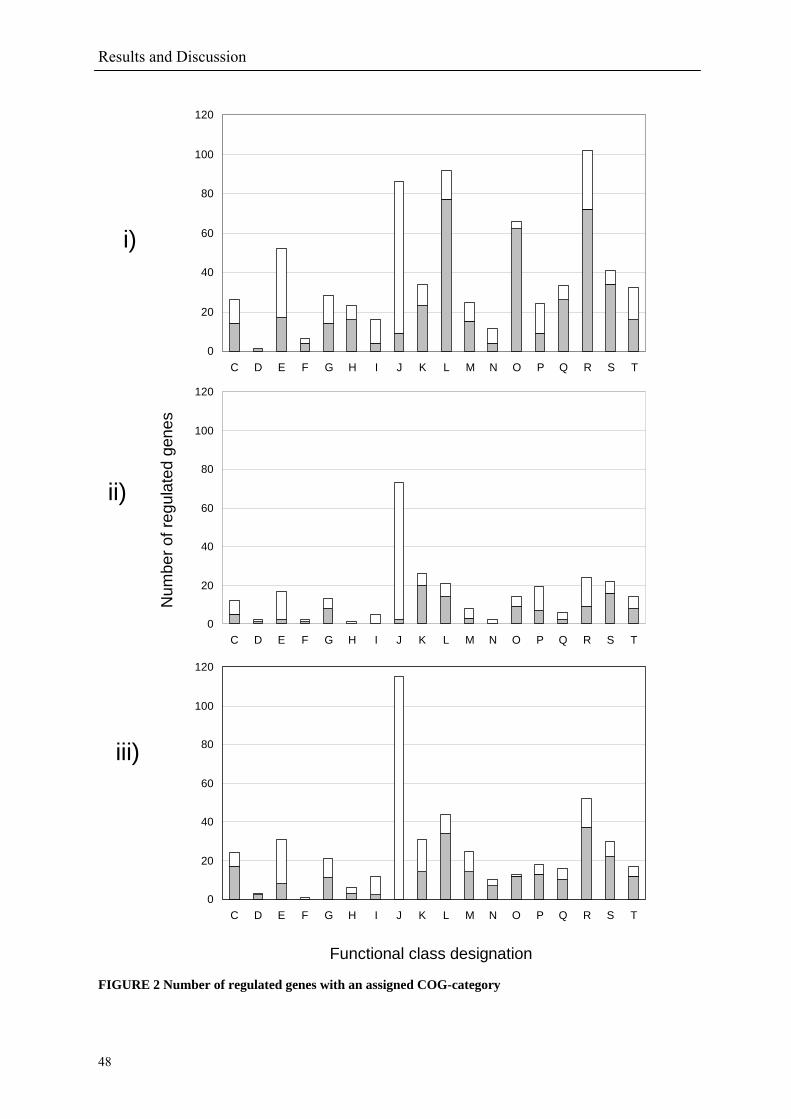
Results and Discussion
48
0
20
40
60
80
100
120
C D E F G H I J K L M N O P Q R S T
i)
ii)
iii)
0
20
40
60
80
100
120
C D E F G H I J K L M N O P Q R S T
0
20
40
60
80
100
120
C D E F G H I J K L M N O P Q R S T
Num
ber o
f reg
ulat
ed g
enes
Functional class designation
FIGURE 2 Number of regulated genes with an assigned COG-category
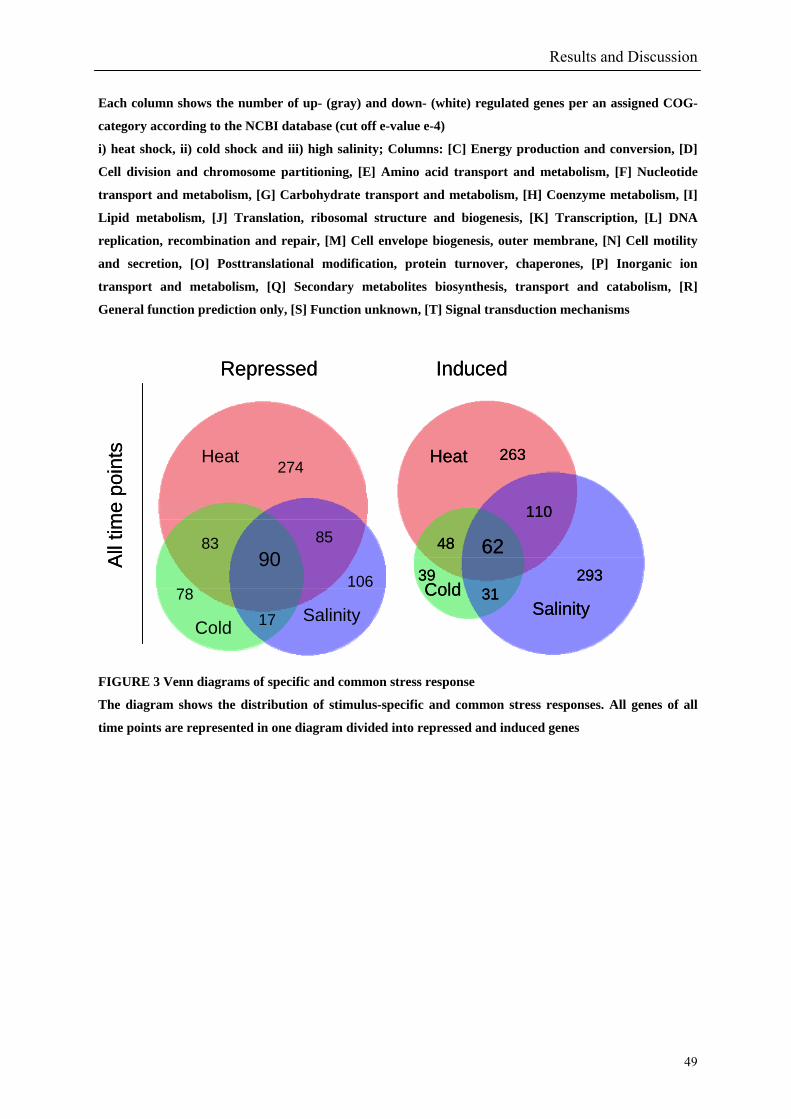
Results and Discussion
49
Each column shows the number of up- (gray) and down- (white) regulated genes per an assigned COG-
category according to the NCBI database (cut off e-value e-4)
i) heat shock, ii) cold shock and iii) high salinity; Columns: [C] Energy production and conversion, [D]
Cell division and chromosome partitioning, [E] Amino acid transport and metabolism, [F] Nucleotide
transport and metabolism, [G] Carbohydrate transport and metabolism, [H] Coenzyme metabolism, [I]
Lipid metabolism, [J] Translation, ribosomal structure and biogenesis, [K] Transcription, [L] DNA
replication, recombination and repair, [M] Cell envelope biogenesis, outer membrane, [N] Cell motility
and secretion, [O] Posttranslational modification, protein turnover, chaperones, [P] Inorganic ion
transport and metabolism, [Q] Secondary metabolites biosynthesis, transport and catabolism, [R]
General function prediction only, [S] Function unknown, [T] Signal transduction mechanisms
Heat
ColdSalinity
Repressed
90
274
78106
17
8583
Heat
ColdSalinity
Induced
62293
263
39
48
110
31
All
time
poin
ts Heat
ColdSalinity
Repressed
90
274
78106
17
8583
Heat
ColdSalinity
Induced
62293
263
39
48
110
31
All
time
poin
ts
FIGURE 3 Venn diagrams of specific and common stress response
The diagram shows the distribution of stimulus-specific and common stress responses. All genes of all
time points are represented in one diagram divided into repressed and induced genes
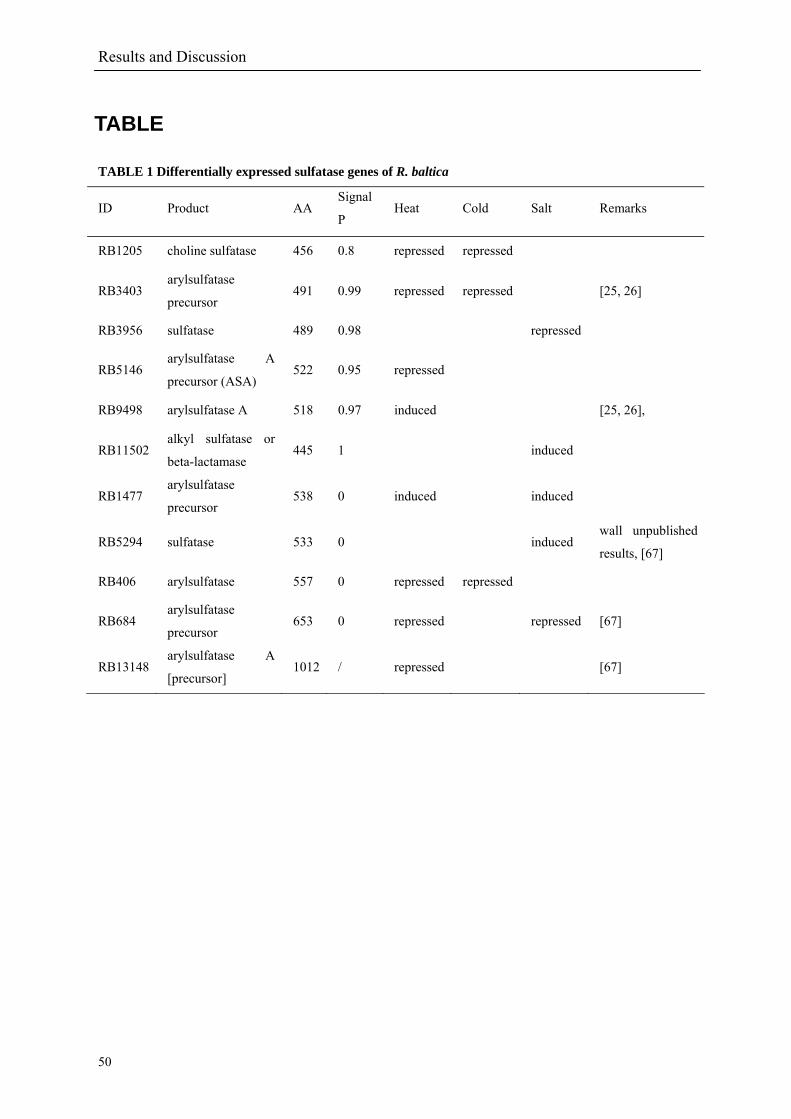
Results and Discussion
50
TABLE
TABLE 1 Differentially expressed sulfatase genes of R. baltica
ID Product AA Signal
P Heat Cold Salt Remarks
RB1205 choline sulfatase 456 0.8 repressed repressed
RB3403 arylsulfatase
precursor 491 0.99 repressed repressed [25, 26]
RB3956 sulfatase 489 0.98 repressed
RB5146 arylsulfatase A
precursor (ASA) 522 0.95 repressed
RB9498 arylsulfatase A 518 0.97 induced [25, 26],
RB11502 alkyl sulfatase or
beta-lactamase 445 1 induced
RB1477 arylsulfatase
precursor 538 0 induced induced
RB5294 sulfatase 533 0 induced wall unpublished
results, [67]
RB406 arylsulfatase 557 0 repressed repressed
RB684 arylsulfatase
precursor 653 0 repressed repressed [67]
RB13148 arylsulfatase A
[precursor] 1012 / repressed [67]

Results and Discussion
51
TABLE 2 Shared stress response: Selection of the induced genes
ID AA Product IEP Strand Potentially involved in / Comments
RB170 96 Transposase IS3/IS911 10.1 + RB370 553 nitrate transporter substrate-binding
protein 4.6 +
RB521 63 hypothetical protein 10.7 - [25, 26] RB723 60 hypothetical protein 7.3 + RB934 375 Putative transposase 9.9 - RB1394 78 hypothetical protein 10.1 - regulatory mechanism RB1395 319 secreted protein similar to DNA-
binding protein 5.5 + regulatory mechanism
RB1789 243 conserved hypothetical protein 9.4 + regulatory mechanism RB1872 38 hypothetical protein 12.3 + RB2186 433 ISXo8 transposase 9.4 - RB2268 282 peptide methionine sulfoxide reductase 9.9 - [23] RB3596 144 nitrogen fixation protein (NifU protein) 4.3 + RB4299 96 Transposase IS3/IS911 9.9 + RB4347 156 conserved hypothetical protein 4.7 + [25, 26] RB4397 55 protein containing DUF1560 10.5 + RB4429 89 conserved hypothetical protein 4.9 + stress response RB4433 162 Ferritin and Dps 4.3 + RB4438 160 Pyridoxamine 5'-phosphate oxidase- 4.4 + RB4510 49 hypothetical protein 9 - RB5238 73 hypothetical protein 10.6 + RB5551 663 hypothetical protein 5.7 + PFAM:DVL RB5888 96 Transposase IS3/IS911 10.1 - RB5938 370 hypothetical protein- 5.6 - [25] RB6928 160 hypothetical protein 4.1 + RB7389 375 Putative transposase 9.9 + RB8409 97 hypothetical protein 8.9 + RB8527 330 protein containing DUF1559 6.6 + stress response RB8987 48 hypothetical protein 9.1 - RB9230 107 hypothetical protein 9.9 + next to transposase RB9907 433 ISXo8 transposase 9.4 + RB9955 452 secreted protein containing DUF1552 5.6 - regulatory mechanism RB9999 281 conserved hypothetical protein- 4.6 - regulatory mechanism RB10049 217 RNA polymerase ECF-type sigma
factor 10.1 +
RB10378 144 Thioredoxin 4.6 - RB10727 276 manganese-containing catalase 5 + [25, 26] RB10728 132 secreted protein 9.9 + stress response RB10896 161 secreted protein 10 - stress response RB10954 143 hypothetical protein 10.4 - RB10956 117 hypothetical protein 4.8 + [26] RB10957 99 conserved hypothetical protein 5.6 + regulatory mechanism RB10958 158 hypothetical protein 5.4 + RB11176 153 protein containing DUF442 4.8 - [26] stress response RB11260 121 dnaK suppressor protein, 5.2 - RB11475 57 conserved hypothetical protein 4.7 + next acyltranscferase, short
protein RB11504 72 conserved hypothetical protein 10.7 - short protein, RB11505 199 conserved hypothetical protein, secreted 7.5 - RB11515 74 conserved hypothetical protein 11.7 + RB11566 195 hypothetical protein 10.8 + RB11749 96 Transposase IS3/IS911 10.1 + RB11750 292 integrase 10 + RB11802 96 Transposase IS3/IS911 10.1 + RB11855 101 conserved hypothetical protein 12.5 - RB11918 134 protein containing DUF971 6.1 -

Results and Discussion
52
RB11977 196 conserved hypothetical protein 9.5 + RB12066 135 hypothetical protein 10 + RB12239 433 ISXo8 transposase 9.4 + RB12247 74 conserved hypothetical protein 6.3 - RB12936 580 conserved hypothetical protein 5.3 - DUF 444 RB12940 96 Transposase IS3/IS911 10.1 + RB13222 208 SOUL heme-binding protein 8.7 - RB13241 167 RNA polymerase ECF-type sigma
factor 8.8 -

Results and Discussion
53
TABLE 3 Shared stress response: Selection of the repressed genes
ID AA Product IEP Strand Potentially involved in / Comments
RB61 58 hypothetical protein 9.6 +
RB314 309 malonyl CoA-acyl carrier protein transacylase 4.4 +
RB318 81 Acyl carrier protein 3.7 + RB319 95 hypothetical protein 10.2 + fatty acid process RB767 311 conserved hypothetical protein, secreted 5.7 + RB825 117 hypothetical protein 7.8 - RB951 234 protein containing DUF1596 12.3 -
RB1129 895 conserved hypothetical protein 5.8 - (S) bombinin, defense response
RB1233 206 30S ribosomal protein S4 11.2 - RB2105 470 membrane protein 9.6 - RB2306 41 hypothetical protein 9 + RB2479 273 conserved hypothetical protein 5.5 - RB3277 221 hypothetical protein 10.4 - RB3362 87 hypothetical protein 11.9 - RB3366 78 hypothetical protein 5.6 + RB3394 36 hypothetical protein 10.5 + RB3399 65 hypothetical protein 9.7 - RB3575 152 membrane protein 10.1 + [22] RB3603 344 secreted protein 4.6 - RB3675 742 secreted protein 8.4 + RB3688 53 hypothetical protein 9.3 - RB3880 82 hypothetical protein 10.7 - RB3953 857 hypothetical protein 5.2 - RB3981 161 hypothetical protein 4.1 + RB3994 191 hypothetical protein 4.1 + RB4097 733 conserved hypothetical protein 6.2 + RB4145 90 hypothetical protein 12 -
RB4194 53 hypothetical protein 11.4 - next to a seronine-threonine kinase
RB4269 282 glutamic acid specific endopeptidase 5.6 - RB4358 123 hypothetical protein 6.5 - [25, 26] RB4373 109 hypothetical protein 4.8 - RB4657 123 hypothetical protein 12.2 + RB4951 95 hypothetical protein 12.1 + RB5262 95 membrane protein 6.3 + RB5409 97 hypothetical protein 12.7 + RB5415 62 hypothetical protein 12.3 +
RB5745 130 hypothetical protein 10.7 - genetic information processing
RB6092 361 Peptidase M50 9.6 + RB6158 142 hypothetical protein 6 - RB6174 69 hypothetical protein 10.6 -
RB6276 105 Histone-like bacterial DNA-binding protein 10.4 -
RB6634 365 protein containing DUF1559 5.3 + RB6699 47 hypothetical protein 11.1 + RB6766 55 hypothetical protein 12 + RB6849 101 hypothetical protein 12.8 - RB7042 91 hypothetical protein 10.4 + RB7116 59 hypothetical protein 11.7 + ribosomal machinery RB7117 181 Ribosomal protein L35 11.4 -
RB7557 327 von Willebrand factor type A domain protein 4.9 +
RB7646 62 hypothetical protein 10.5 + RB7647 73 hypothetical protein 7.4 +

Results and Discussion
54
RB7837 286 Ribosomal protein L2 11.8 + RB7838 89 Ribosomal protein S19/S15 10.8 + RB7839 119 Ribosomal protein L22/L17 11 + RB7840 236 30S ribosomal protein S3 10.4 + RB7841 138 Ribosomal protein L16 11.1 + RB7849 108 Ribosomal protein S17 10 + RB7850 122 Ribosomal protein L14b/L23e 11 + RB7852 196 50S ribosomal protein L5 10.4 + RB7854 61 Ribosomal protein S14 11.8 + RB7856 181 50S ribosomal protein L6 10 + RB7857 149 Ribosomal protein L18P/L5E 11.6 + RB7859 177 Ribosomal protein S5 10.6 + RB7894 398 translation elongation factor EF-Tu 5.2 + [25, 26] RB7899 141 50S ribosomal protein L11 9.6 + RB8119 142 hypothetical protein 10.1 + RB8457 113 hypothetical protein 11.5 - RB8594 41 hypothetical protein 9.2 + RB8669 37 hypothetical protein 11.5 + ribosomal machinery RB9343 59 hypothetical protein 11.4 + RB9417 103 hypothetical protein 10.5 + RB9460 79 hypothetical protein 10.3 - RB9872 67 hypothetical protein 5.4 + cell division related RB10581 384 secreted protein containing DUF1559 6.2 + [26] RB11287 75 hypothetical protein 9.1 + RB11392 148 conserved hypothetical protein 5 +
RB11490 181 conserved hypothetical protein, membrane 10.3 +
RB11707 83 conserved hypothetical protein 9.8 + stress function RB11766 129 hypothetical protein 10.4 + [47]
RB12193 36 hypothetical protein 7.5 - overlapping with asnB (RB12191)
RB12251 567 RNA polymerase specialized sigma factor 9.4 -
RB12327 686 TGF-beta receptor, type I/II extracellular region 4.5 -
RB12329 110 conserved hypothetical protein, membrane 4 +
RB12396 57 hypothetical protein 11.3 - RB12454 199 hypothetical protein 10.3 - RB12818 163 conserved hypothetical protein 11.7 - ribosomal machinery RB12821 117 Ribosomal protein L19 11.1 - RB12824 146 Ribosomal protein S16 5.3 - RB12837 65 hypothetical protein 9.8 + ribosomal machinery RB12839 225 Ribosomal protein L1 9.8 +
Additional files
ADDITIONAL FILE 1 - List of differentially expressed genes for all three stress experiments
ADDITIONAL FILE 2 - List of selected clusters of the k-means clustering

Results and Discussion
55
3.2 The Rhodopirellula baltica life-cycle: Growth-dependent expression profiling of the complete genome with microarrays
Authors: Patricia Wecker, Marga Schüler, Andreas Ellrott, and Frank Oliver Glöckner
Manuscript in preparation

Results and Discussion
56
The Rhodopirellula baltica life cycle: Growth-dependent expression profiling of the complete genome with microarrays
Patricia Wecker1,2, Margarete Schüler3, Andreas Ellrott1 and Frank Oliver Glöckner1,2§
1 Max Planck Institute for Marine Microbiology, Microbial Genomics Group, Celsiusstr. 1,
28359 Bremen, Germany
2 Jacobs University Bremen gGmbH, Campusring 1, 28759 Bremen, Germany
3 Max Planck Institute for Biochemistry, Dept. Molecular Structural Biology, Am
Klopferspitz 18, 82152 Martinsried, Germany
Keywords: Bacteria, Planctomycetes, Rhodopirellula baltica, expression profiling, whole
genome array, microarray, hypothetical genes, cell cycle, cell compartmentalisation, FM-dye
Author´s e-mail addresses:
P. Wecker: [email protected]
M. Schüler: [email protected]
A. Ellrott: [email protected]
F.O. Glöckner: [email protected]
§ Corresponding author

Results and Discussion
57
Abstract
Background: Rhodopirellula baltica is a marine representative of the globally distributed
phylum Planctomycetes. As typical for Planctomycetes, R. baltica has several unique
properties, like a peptidoglycan-free proteinaceous cell wall, intracellular
compartmentalisation, and it undergoes morphological changes during its life cycle, such as
budding and cell aggregations. The aim of this transcriptomic study was to get first hints on
genes involved in the replication and life cycle of R. baltica which are mostly unknown. Here
we report the results of the first growth dependent expression profiling.
Results: Four different growth phases of a R. baltica culture were analysed using a whole
genome microarray. The number of regulated genes increased from 90 in the exponential to
863 in the late stationary phase when compared to the transition phase. Results indicate a
remodelling of the cell wall and membrane in the exponential phase, sensing of environmental
changes in the transition phase, followed by a stationary-phase specific response. More than
half of the regulated genes per growth stage were annotated as hypothetical proteins. These
genes are probable candidates for functions associated with the planctomycete specific
morphotype-differentiation during the life cycle. Interesting was the growth-dependent
regulation of 12 sulphatases and the up-regulation of genes involved in a predicted C1-
metabolism pathway after 240 h of cultivation.
Conclusion: The whole genome array of R. baltica gave initial insights into the regulation of
all genes during the life cycle. We were able to show activity of genes that might be involved
in the formation of budding and swarmer cells. The transcriptional profiling showed that a
large number of hypothetical proteins function within the cell cycle and in the formation of
the different cell morphologies. The transcriptomic results are contributing to a better
understanding of the so far unknown molecular cell biology underlying the cell
compartmentalisation of R. baltica and, more general, of the phylum Planctomycetales and
possibly even of the PVC superphylum.

Results and Discussion
58
Background
Rhodopirellula baltica SH1T was isolated from the water column in the Kiel Fjord (Baltic
Sea) and taxonomically grouped within the bacterial phylum Planctomycetes [1, 2]. Members
of this group are abundant in aquatic habitats [3, 4] and considered to play a significant role in
carbon recycling [5]. Planctomycetes share several unique properties, such as peptidoglycan-
free proteinaceous cell walls [6, 7], intracellular compartmentalisation [8] and a mode of
reproduction via budding. The latter results in a life cycle which is characterised by motile
and sessile morphotypes and resembles that of Caulobacter crescentus [9: Gade, 2005 #186].
During early stages of growth, numerous budding mother cells and swarmer cells are present,
as illustrated in FIG 1 (redrawn from Gade et al. [10]). As growth proceeds, the number of
swarmer cells decreases, and single cells form rosettes by attaching to each other. These
aggregates dominate during transition- and stationary phase.
The annotation of the completely sequenced genome of R. baltica allowed an initial
assessment of its genetic potential and revealed several interesting and surprising traits such
as high number of sulphatase genes or a conspicuous C1-metabolism pathway [5 and
references therein]. However, not much information could be inferred from this genome
content survey with respect to genes functioning in the development of the different
morphologies during the cell cycle and its regulation. In general, the cell division machinery
of Bacteria consists of different cell division genes, like the filamentation temperature
sensitive genes (fts) named ftsZ, ftsA, ftsI, ftsL, ftsQ, ftsN, ftsW, ftsK, and the FtsZ interacting
proteinA (zipA) [11]. The ftsZ-gene is the major constituent of the septal replication ring.
Surprisingly, all of these genes, except ftsK, are absent from the genome of R. baltica as well
as from the genomes of other Planctomycetes such as Gemmata obscuriglobus UQM 2246T,
Planctomycetes maris DSM 8797T, and Blastopirellula marina DSM 3645T, ‘Candidatus
Kuenenia stuttgartiensis’ harbours genes coding for ftsI and ftsW in addition to the ftsK gene.
Teeling [12] concluded that the cell division in Planctomycetes, like in Chlamydia, must take
place by a totally unknown mechanism.
Regarding the morphological changes, the life cycle of R. baltica seems to be similar to
Caulobacter crescentus CB15. In C. crescentus, the key gene of the cell-cycle is the control
protein CtrA. It acts as the master regulator by overseeing several important cellular processes
[13]. Within the R. baltica genome no homologue to the protein CtrA has been found.
However, some base patterns (e.g. TTAAN7AAAC), were found which are similar to CtrA

Results and Discussion
59
and CzcR – binding site patterns (TTAAN7TTAA). CzcR is a control gene and responsible for
induction of resistance.
It was anticipated that genes involved in the cellular changes during the life cycle show
growth-dependent changes on the mRNA level. To monitor complex processes such as the
cell cycle, whole genome expression profiling has been shown to be particularly useful [14,
15]. Investigations of the gene expression during the C. crescentus cell cycle revealed that,
surprisingly, around 19% of the genes in the genome are affected [16]. For R. baltica, growth-
dependent expression patterns were expected to reveal genes that might be required for
morphotype differentiation and for physiological functions, and cellular processes typical of
this group of organisms. In addition, morphology studies based on fluorescence imaging of
the stained membrane [8] of dividing R. baltica cells were undertaken that to aid in
understanding of how the cell compartments are distributed to the daughter cells during the
unequal cell division.
Results and discussion
Gene expression studies
R. baltica was grown in a defined mineral medium with glucose as carbon source to study
changes on the gene expression level that reflect changes due to different life cycle phases.
Cultures of R. baltica were harvested at the onset of the exponential phase (after 44 h of
incubation), during the exponential phase (62 h), in the transition phase between exponential
and stationary phase (82 h), and in the early and late stationary phase (96 h and 240 h,
respectively). Gade and coworkers [10] have shown that there is a correlation between distinct
morphotypes and different phases of the growth curve of R. baltica cultures based on
microscopic examination. Since the R. baltica cultures used in the present study were not
synchronised, we applied Fluorescence Activated Cell Sorting (FACS) to examine the
morphological distribution in the cultures at each of the time points of interest. Looking at
FIG 3 a correlation can be seen between the scatter plot and the microscopic pictures from
this study with the microscopic observations of Gade et al. [10]. In the early exponential
growth phase the culture was dominated by swarmer and budding cells (FIG. 3 A), in the
transition phase we found single and budding cells as well as rosette formations (FIG. 3 B)
and the stationary phase was dominated by rosette formations (FIG. 3 C).

Results and Discussion
60
Global differential gene expression results
An expression ratio of higher 1.5 or below -1.5 was used as a threshold for significant
changes. The threshold was determined in a self-self-hybridisation experiment.
The total number of genes detected as differentially regulated for each investigated growth
stage is shown in TABLE 1. A complete list of genes can be found in the ADDITIONAL
FILE 1. The comparison of samples taken from the early log-phase (44 h) and the exponential
phase (62 h) showed that 149 genes were regulated, of which 64% are annotated as
hypothetical proteins. The comparison between exponential (62 h) and transition phase (82 h)
resulted in only 90 regulated genes with more than 50% having an unknown function. The
first pronounced change of the transcriptome with 235 regulated genes (63% hypothetical
proteins) was observed when comparing the transition phase (82 h) with the stationary phase
(96 h). A comparison of cultures from 82 h with the late stationary phase (240 h) revealed the
most pronounced changes of the transcriptome in our study: 863 genes were differentially
expressed, of which 61% are annotated as hypothetical proteins (FIG 4).
The assignment of the differentially expressed genes to functional classes according to COG
delivered first hints about the activity in the R. baltica cells at the different growth stages
(TABLE 2). The comparison of the samples from within the exponential phase (44 h and 82 h
with 62 h) showed that a maximum fraction of 2% of the total number of genes was regulated.
Gade et al. [10] observed the same effect on the proteome level and proposed that it might
reflect the favourable nutritional conditions encountered throughout these growth stages.
Genes associated with the metabolism of amino acids [E] and carbohydrates [G], genes for the
energy production and conservation [C], as well as genes for the DNA replication and
recombination [L] were up-regulated in the early exponential phase (44 h) compared to 62 h.
The induced oxidases (RB4416 and RB5701), peptidases (RB4269 and RB6443), lipases and
esterases (RB7072 and RB3413) reflect a higher metabolic activity in the cells.
Interestingly, genes for the “cell envelope biogenesis, outer membrane” [M] and “cell motility
and secretion” [N] were induced at 44 h. Genes coding for flagellin A (flaA - RB4454), for
bacterial type II and III secretion system protein (RB5174) and for diverse transferases
(RB6305 and RB6624) were induced. Thus, it seems that at the onset of the exponential phase
(44 h) compared to the mid-exponential phase (62 h) a remodelling of the cell wall and the
morphology was ongoing, perhaps triggered by an enhancement of the swarmer cell
production which is likely very costly in energy and would explain the observed higher
expression level of genes from COG category C [17]. Although R. baltica was in the middle

Results and Discussion
61
of the exponential phase (62 h), fewer genes are regulated than in the stationary phase. This
indicates that all genes necessary for exponential growth under favourable conditions were
already expressed in early log phase. R. baltica increased the glutamate dehydrogenase
(RB6930) level. This enzyme is involved in the biosynthesis of arginine, glutamate and
proline. Latter is one of the major components of the cell wall of R. baltica.
The cell density has reached the highest point in the R. baltica flask culture at the transition
phase. Cells seem stressed as genes coding for glutathione peroxidase (RB2244), thioredoxin
(RB121260) and bacterioferritin comigratory protein (RB12362) were up-regulated, which
corresponds to proteomic data [10]. General stress protectants like trehalose synthase
(RB5196), universal stress protein (uspE - RB4742) and chaperones (e.g. RB8966) were up-
regulated. The observation that diverse dehydrogenasese, hydrolases and reductases were
regulated, suggests a metabolic adaptation of R. baltica to the nutrient limitation in the
culture. The transition from the logarithmic phase of growth into the stationary phase is not a
passive process; and the expression levels of a number of gene products underwent substantial
changes during this period [18]. The transition phase is usually characterised by the onset of
nutrient depletion and starvation stress; cells need to prepare for long-term survival under
suboptimal or even unfavourable conditions. Earlier experiments have shown that R. baltica
survives for at least 14 days in the stationary phase at 28°C [10]. Apparently, R. baltica is
well adapted for survival under nutrient limiting conditions which are often encountered in its
natural environment. We compared expression profiles of the transition phase (82 h) with that
of the early (96 h) and the late stationary phase (240 h). The intersection of both sample sets
(96 h vs. 82 h and 240 h vs. 82 h), visualised by a Venn diagram (FIG 5), revealed 103 genes
with similar regulation behaviour. 71 out of the 103 genes are annotated as hypothetical
proteins and most of the remaining 32 genes are stress-related as expected. The general
response of cells to the conditions in stationary phase consists of the induction of genes
associated with energy production, amino acid biosynthesis, signal transduction,
transcriptional regulation, stress response and protein folding. Genes coding for enzymes of
the carbon metabolism, translation control, energy production and amino acid biosynthesis
were repressed.
R. baltica decreased the expression level of genes belonging to the ribosomal machinery
(RB12197, RB2543, RB264, RB5801, RB5804, RB7022, RB7818, RB8253, RB8725 and
RB9304), to transcription regulation (RB10339, RB10458, RB11223, RB11242, RB12372
and RB1392) and to DNA replication and recombination (RB11863 and RB3281) in the late
stationary phase, suggesting a rearrangement of the genome. In addition, the organism

Results and Discussion
62
expressed many genes coding for transposases, integrases and recombinases (RB10096,
RB11303, RB11750, RB1190, RB3144, RB4826, RB5887, RB7388, RB12239, RB2186,
RB6736, RB9907, RB6167, RB7389 and RB934), suggesting that under stress conditions
genome arrangements take place. Six out of those 15 genes (RB11750, RB12239, RB2186,
RB9907, RB7389, and RB934) have been found active under abiotic stress factors, like
temperature and salinity ([19] in prep), which indicates that expression of these genes
constitutes a response to general stress. Noteworthy is the up-regulation of genes for the
phenylalanine, tyrosine and tryptophane biosynthesis (RB6822 and RB6147). The latter have
been found expressed on the proteome level as well [10] but the physiological meaning is
presently unknown. Also genes involved in the biosynthesis of serine, threonine and glycine
(RB6932) and of lysine (RB4382) were up-regulated. Interesting is the induction of genes for
the ubiquinone biosynthesis (RB2748, RB2749 and RB2750). Because it is known from
A. tumefaciens and R. sphaeroides, that when oxygen is lacking, the production of ubiquinone
increases [20]. In addition, it seems that the composition of the cell wall responded to changes
in the physiological state of the R. baltica cell during the late stationary phase. The organism
exported more polysaccharides. And diverse genes belonging to the cell membrane class [M]
were up-regulated. R. baltica activated the production of many outer membrane transporter
and biopolymer exporters (RB12053, RB12055, RB9308, RB1120, RB754, RB12279 and
RB10255), as well as several transferases (RB12690, RB2507, RB2985 and RB5846) for
alteration of the lipopolysaccharides that belong to the cell membrane. In addition, we found
19 genes annotated as membrane proteins regulated, as well as 13 secreted proteins. Two
genes coding for bacterial micro-compartment proteins (RB2585 and RB2586) were up-
regulated. They are supposed to form primitive organelles in a variety of bacteria [21].
Perhaps the cell wall became adapted to maintain the cell shape, cell division and genome
segregation and/or is acting as an intermediary between the cell and its environment. It could
be that after entering the stationary phase, R. baltica cells, like the marine bacterium
’Candidatus Pelagibacter ubique‘, undergo morphological changes (cell shape), enabling
them to return to exponential growth again after being transferred to fresh medium or to a
nutrient rich environment respectively [22]. We also found hints as to the formation of the
storage compound glycogen in the expression profile of the stationary phase. So far glgB
(RB548) was only found to be expressed when R. baltica cultures were grown on agar plate
[10]. In the present study, we found two genes induced; glgB (RB548) at 96 h and glgX
(RB4894) at 240 h.

Results and Discussion
63
In the following sections, regulation of selected genes will be described and discussed
according to their functional classification.
Sulphatases
The genome of R. baltica is harbouring 110 sulphatases. It has been proposed on the basis of
their sequence similarity that they are involved in the utilisation of carbon from complex
sulphated heteropolysaccharides [5].
During transcriptional analysis of the different R. baltica growth stages we found 12
sulphatases up- or down-regulated including four putative choline sulphatases (RB10161,
RB110, RB2254 and RB5282), six different arylsulphatases (RB13148, RB4815, RB5195,
RB684, RB686 and RB7875) and two other sulphatases (RB5294 and RB5424) (TABLE 3).
Up-regulation of sulphatase genes cannot have been caused by substrate induction, since the
mineral medium used in this study did not contain any sulphated carbon compounds. Five of
the sulphatase genes mentioned above (RB110, RB2254, RB4815, RB5294 and RB686) were
also found up-regulated when the expression profile of R. baltica cells grown on a solid
surface was compared with that of cells grown in liquid culture (data not shown). The
observation that certain sulphatases were only expressed in specific growth phases suggests
that degrading polymers is not the only role of sulphatase genes in R. baltica. Based on the
differential expression pattern of these sulphatase genes we propose an involvement in
structural remodelling during the morphological differentiation of R. baltica cells, such as the
formation of the holdfast substance, which is needed to attach to surfaces, the production of
slime polysaccharides [23], or in the cell wall formation, which consists of a protein sacculus
with disulphide bonds [7]. More experimental work is needed to clarify the potentially diverse
role of the many sulphatase genes in R. baltica.
Lipids
In contrast to Gade et al. [10], who found indications that the biosynthesis of sulphated
saturated fatty acids occurs specifically during the stationary phase, we only found that delta 9
acyl-lipid fatty acid desaturase (RB4876), belonging to the unsaturated fatty acid pathway,
was up-regulated in the transition phase (82 h). Furthermore, malonyl CoA-acyl carrier
protein transacylase (RB314), belonging to the fatty acid pathway, was up-regulated in the
late stationary phase (240 h). Fatty acids appear also in the membrane of diverse groups of
Planctomycetes [24]. RB314 together with RB318 was found up-regulated under stress

Results and Discussion
64
conditions [19], leading to the assumption of a general stress response to enhance the stability
of the membrane to protect the cell.
C1-metabolism
R. baltica has the genetic potential to degrade C1-compounds and the functional prediction
revealed all enzymes necessary for the oxidation of formaldehyde to formate [5]. Out of the
13 proposed genes encoding enzymes of C1-metabolism [25] four genes were regulated in our
experiment. The following genes were induced in the late stationary phase: fae (RB9338),
mtdA (EC 1.5.1.5 - RB10295) and citG (Orf7 - RB10246), whereas ATP-dependent
carboligase (OrfY -RB10299) was found to be induced in the exponential and repressed in the
early stationary phase. Although in silico studies have revealed the presence of this C1-
pathway also in other Planctomycetes [26, 27] the specific functions of the genes in
Planctomycetes are still unknown. The expression of some of the genes observed in this study
suggests at least that this pathway has physiological relevance in R. baltica and probably in
other Planctomycetes.
Cell division and cell cycle
Not much is known about the regulation of the replication and the process of cell division in
Planctomycetes. None of the 20 R. baltica genes assigned to the COG category “cell division
and chromosome partitioning” [D] were found to be regulated during this study, nor any of
the homologues of genes that are generally known to be involved in the cell division, but
without specific category assignment (ADDITIONAL FILE 2). This suggests that a number
of so far hypothetical R. baltica proteins must play a role in the regulation of replication and
cell divison. R. baltica exhibits a life cycle similar to C. crescentus. DnaK/DnaJ and
GroES/GroEL are the best-characterised chaperone systems in bacteria and induced in cell
cycle regulation and under heat shock in C. crescentus [28]. Those chaperones have an
important role in chromosome replication and partition and/or cell division. The R. baltica
genome harbours genes coding for groEL (RB10629 and RB8970), dnaK (RB5754 and
RB5755) and dnaJ (RB8972) and groES (RB) in the genome. However, only the expression
of groEL (RB10629) is up-regulated at 82 h in the life cycle experiment, which could also be
a general response to stress [19].
R. baltica also possesses genes encoding dnaA (RB11579 and RB1706) which initiates not
only the DNA replication but also promotes expression of the components necessary for

Results and Discussion
65
successful chromosome duplication in C. crescentus [28]. However, dnaA was not found to be
regulated during our experiment.
Alternative sigma factors and two-component regulators
The R. baltica genome is harbouring 33 genes coding for RNA polymerase sigma factors with
extra cytoplasmic function (ECF). 5 out of 33 genes were found to be regulated in this study.
ECF-sigma factors are known to be involved in the regulation of bacterial interaction with the
extracellular environment, e.g. sensing of stationary-phase conditions and initiation of the
global response. This should lead to an increased survivability of R. baltica [22].
Furthermore, it is known that ECF sigma factors are involved in the cell envelope response of
B. licheniformis [29] and in the cell division of B. subtilis [30] The gene coding RNA
polymerase sigma-70 factor ECF (RB11242) was induced in the transition phase (82 h) and
repressed together with three other genes coding ECF-sigma factors (RB12372, RB138 and
RB8027) in the stationary phase. RB9852 was up-regulated in the early stationary phase.
In addition, two sigma factors were regulated in the late stationary phase; RB7924 was up-
and RB1392 down-regulated. RB1392 could be responsible for the regulation of the cell
division inhibitor (RB1391). A sigma-54 dependent regulator (RB7579) was also induced.
Two out of four genes coding for the RNA polymerase sigma factor rpoD (RB10339 and
RB6780) were down-regulated. They are also found to be reduced in expression during the S-
phase of C. crecsentus [14]. The authors of this study suggest a transcriptional regulation of
the central metabolism. Although R. baltica possesses the stationary-phase sigma factor rpoS
(RB3727), which is known to lead to a global stationary-phase response, it was not regulated
in our experiment.
Two-component-system response regulators allow bacteria to respond to changes in the
environment. 68 genes in the R. baltica genome were annotated as response regulators, of
which four were found to be differentially expressed in this study (RB4484, RB10308,
RB10491 and RB6403). Response regulators RB10308 and RB4487 were detected in the
stationary phase proteome [10]. In our case, RB4487 was induced at 96 h and repressed at
240 h. The response regulator RB6403 was also repressed in the late stationary phase, which
was as well shown for the proteome [10]. The gene coding for the response regulator
(RB10491) was not found to be differentially expressed on the transcriptome level, but it was
induced in the stationary phase on the proteome level [10]. It seems that these regulator
proteins are involved in the stress response of R. baltica during stationary phase conditions.

Results and Discussion
66
Taken, for example, the two stress sensor genes encoding carbon storage regulator (CrsA -
RB10261 and RB8149); only RB10261 was induced at 96 h but both were repressed in the
late stationary phase (240h). Noteworthy is the fact that no histidine kinase was found to be
differentially expressed during our experiment, whereas sixteen histidine kinases are found to
be expressed in a cell-cycle-dependent manner in C. crescentus [14]. Those transmembrane
sensor proteins plus response regulator are supposed to be involved in the monitoring of
environmental conditions and respond to perturbations harmful to the cell envelope [29].
Hypothetical proteins
It was not surprising that a large number of genes found to be differentially expressed in this
study encoded (conserved) hypothetical proteins, given the limited functional knowledge on
cellular processes in Planctomycetes. Some of these regulated hypothetical proteins were
planctomycete-specific. They are supposed to code for the unique cellular planctomycetal
characteristics or for specific metabolic traits, or are proposed to be involved in regulation
[26]. Among the regulated hypothetical proteins 51 ORFs with a Domain of Unknown
Function (DUF) were found. The R. baltica genome encodes 74 planctomycetes-group-
specific proteins that are annotated as hypothetical proteins containing DUF1559 (PF07596)
and comprise a new family of “solute-binding” proteins. These are high affinity transport
systems which could have an additional function in signal transduction systems [31]. Ten of
these proteins were found to be expressed, in this study, and of these nine were especially up-
regulated in the stationary phase. RB2556 and RB4325 encoding for proteins of the family
“planctomycete PGAMP” (PF07644) and RB560, coding for a protein containing the DGPF
domain (PF04946) were up-regulated in the late stationary phase (240 h). Their function is
unknown and so far no further data are available to support our speculation that they are in
involved in morphological changes in Planctomycetes. In proteome studies [10], the products
of 14 genes annotated as hypothetical proteins are assumed to be involved in the cell cycle
and/or morphotype differentiation rather than stress or nutrient limitation response. In part,
the results of the present transcriptome analysis support the data gained from an earlier
proteome study [10]. The authors of the latter propose for the three planctomycete-specific
proteins (RB5240, RB6221 and RB6941) an involvement in proliferation or swarmer cell
differentiation. On the transcriptome level RB6221 and RB6941 were down-regulated in the
late stationary phase. RB5240 was not differentially expressed at all. None of the three were
regulated under stress [19]. Gade et al. [10] also propose that the five unique proteins

Results and Discussion
67
RB3577, RB5268, RB7235, RB8559 and RB12461, whose expression has been found to be
increased in the stationary phase, might be involved in the rosette formation or surface
attachment. Neither of them were found to be regulated in our growth experiments. However,
since RB7235 was induced 20 min after cold shock and 10 min after salt shock and RB12461
was induced 20 min after heat shock [19] and at least additional role in stress response should
be considered for these genes (ADDITIONAL FILE 3).
Cluster-overview
In order to get some more information on the potential function of those differentially
expressed genes that encode hypothetical proteins we delineated groups of genes with similar
expression pattern during culture growth. Information on the genes populating each cluster
can be found in the ADDITIONAL FILE 4. TABLE 5 shows the average expression pattern
of the clusters that are discussed in the following. Cluster 4, 12 and 17 group genes that were
only regulated in the exponential phase where nutrient limitation or stress conditions are
unlikely. Therefore, these genes can be supposed to have a function in the morphology
differentiation to swarmer cells or in the budding process. Genes belonging to cluster 11, 13,
19, 20, 21, 22 and 23 were up-regulated during the transition phase. Cluster 21 is interesting,
with genes induced only in the transition phase (82 h) and repressed at 240h. They might be
involved in stress monitoring and handling. Genes annotated as hypothetical proteins
belonging to the early stationary phase (96 h) were grouped in cluster 18, 24 and 25. Cluster
18 and 24 consist of hypothetical proteins that might be involved in the morphology changes,
e.g. cell shape and wall. Cluster 25 consists mainly of genes belonging to the energy
production plus hypothetical proteins. Genes only induced in the late stationary phase (240 h)
belong to the cluster 3 and 16. Finally, cluster 14 genes were up-regulated in the exponential
(62 h) and down-regulated in the stationary (96 h) phase in comparison to the transition phase.
This cluster contains one sulphatase gene and 14 genes encoding hypothetical proteins. It can
be assumed that these have a function in the morphotype differentiation.
Cell biology studies
The whole genome array for the marine planctomycete R. baltica [19] was taken as a tool to
explore the regulation of all genes during the life cycle. The expression data provided hints as
to which genes might be involved in the cell division of R. baltica. In order to get a better
understanding of how the division process of the compartments takes place in three-

Results and Discussion
68
dimensional space, for instance how the budding mechanism works, which membranes break
down in the course of cell division or how the nucleoid is segregated to daughter cells, we
conducted cell biology experiments using specific fluorescent DNA- and membrane-markers
on cells from the transition phase of the R. baltica culture. Fluorescence membrane staining
and 3D-reconstruction using cryosections has previously been done on the planctomycete
Gemmata obscuriglobus [8, 32] to understand more about the cell compartmentalisation. We
stained the R. baltica cells with two different permanent membrane dyes, a cytoplasmic
membrane specific dye (FM4-64), and a general endomembrane dye (DiOC6 (3)). The DNA-
specific fluorescent dye DAPI was used for the nucleoid. Results are shown in FIG 6 and
FIG 7 A, B and C. FIG 7 shows dividing and sessile R. baltica cells stained with DAPI and A)
DiOC6(3) or B) FM4-64. FIG 7 C) shows a composite picture of R. baltica cells forming
rosettes stained with all three dyes (DAPI, DiOC6(3) andFM4-64).
To get a different resolution on the whole single cell level we applied the new nanoSIMS-
technology, which is available at the University of Mainz, to membrane-stained R. baltica
cells from the transition phase of the culture. Both membrane dyes contain halogen atoms
(FM4-64 bromine, DiOC6(3) iodine). The incorporated membrane dye halogens can be
detected and quantified by the nanoSIMS microprobe and displayed by a graphical
reconstruction. Initial results show that the method is successful. The shape of the R. baltica
cell was recognisable (FIG 8). The background was satisfying and, despite the comparative
large cell size, the first pictures look promising and encourage further studies.

Results and Discussion
69
Conclusions
Our global transcriptome analysis, for example, documents the morphological changes during
the exponential growth. The lack of nutrients and a high cell density in the stationary phase
imposes the activation of general stress response genes, genes for the reorganisation of the
genome and the cell wall. The ability to sense and adapt to a changing environment can be
monitored by the up-regulation of ECF-sigma factors and response regulators. The results
provide also hints that a subset of the 110 sulphatases might be involved in the formation of
planctomycete-specific morphological features (e.g. cell wall components or holdfast
substance), beyond sulphated carbon compound utilisation.
Proteins of unknown function constitute a major part of the regulated genes in all growth
stages of R. baltica. Their correlation to morphotype and their expression only at specific
growth stages provides first hints about their function in the life cycle and the underlying
cellular processes in R. baltica. Follow-up nanoSIMS studies will help to understand how the
division process of the compartments in three-dimensional space is taking place. Our results
form the basis for further studies on the transcriptome level to reveal the unknown mechanism
behind the cell cycle and cell division of R. baltica, as a representative of the phylum
Planctomycetes.
Material and Methods
Cultivation of Rhodopirellula baltica cells
Cells of R. baltica were grown in batch cultures (500ml) with mineral medium containing
10 mM glucose as sole carbon and 1 mM ammonium chloride as nitrogen source [33]. The
R. baltica cultures were incubated at 28°C on a rotary shaker. Three independent batch
cultures (biological replicates) per incubation time point were inoculated, totalling 18 flasks at
the beginning of the experiment.
Harvesting of cells and preparation for microarray hybridisations
Cultures were harvested by centrifugation (Beckman CoulterTM AvantiTM J-20XP, JA10
Rotor, 20 min, 6000 rpm, 4°C), after different incubation times (44, 68, 82, 96 and 240 h).
The culture broth was collected in 500 ml tubes and swirled briefly in an ethanol/dry ice bath
to rapidly cool down the cultures and "freeze" the RNA profile. Subsequently, the broth was

Results and Discussion
70
centrifuged at 6000 rpm for 20 min at 4°C (Beckman CoulterTM AvantiTM J-20XP, JA10
Rotor). The pellets were re-suspended in 0.1 M Tris-HCl and then re-centrifuged to obtain
cell pellets that were shock-frozen in liquid nitrogen and stored at -80°C. Total RNA
isolation, cDNA synthesis, array construction, hybridisation and washing were performed as
previously described [19]. A detailed description of the array can be found at the NCBI´s
Gene Expression Omnibus (GEO) database [34] under accession number GPL7654 and [19].
Experimental design and sample preparation
For expression profiling and microscopic analysis samples were collected at 44, 62, 82, 96
and 240 h. In two independent hybridisations for each sample, cDNA samples were
hybridised according to FIG 2. At each growth time point the sample was labelled either with
Alexa 647 or Alexa 546 for the array analysis. The data shown are based on the analysis of
the two replicates performed for each of the conditions. Dots and arrowheads represent
samples labelled with Alexa 546 and Alexa 647, respectively.
Signal detection and data analysis
Slides were scanned at a resolution of 5 µm using a ScanArray Express Microarray scanner
(Perkin Elmer). The image analysis software provided with this scanner was used for
automatic spot detection and signal quantification. Raw data were automatically processed
using the microarray data analysis software tool MADA [35]. LOWESS Normalisation was
applied on an R-versus-I plot with a smoothing factor of 0.5. If replicates were available, the
expression data (ratio) of all hybridisations were averaged and combined to one expression
data point (ratio).
Cluster analysis and Database
Differentially expressed genes displaying the complete loop profile were clustered using the
k-means clustering approach (Euclidean distance metric, k = 25 clusters and 9 iterations (max.
500)) [36] with the software tool Multi-experiment Viewer MeV Version 4.0.2 from the TM4
microarray software suite [37]. The genome of Rhodopirellula baltica was automatically re-
annotated based on updated homology searches (June 2005 - MicHanThi [38]). The updated
annotation including all tool results are publicly available [39]. JCoast [40] was used as a tool
for the visualisation, interpretation, statistic (COG-assignment) and comparison of genomic
data stored in GenDB V2.2 [41]. Venn diagrams were generated by BioVenn [42].

Results and Discussion
71
A detailed description of the life cycle data will be made publicly available at the GEO
database.
Fluorescence microscopy modified after [8]
R. baltica cells were harvested into Milli-Q-grade deionised water. After centrifuging, the
pellet was re-suspended in dye solution for 5 to 60 min before pipetting 5 µl onto an agarose
slab on a glass microscope slide, allowed to soak in and a coverslip was placed on top. The
solution of dyes used contained final concentrations of 3.3 μg ml-1 – DiOC6(3) (3,3′-
dihexyloxacarbocyanine iodide) (Molecular Probes), 200 μg ml-1 – DAPI (4′-6-diamidino-2-
phenylindole) (Sigma) and 3.3 μg ml-1 – FM4–64 (N- (3 triethylammoniumpropyl)-4-(p-
diethylaminophenyl-hexatrienyl) pyridimium dibromide) (Molecular Probes) or 3.3 μg ml-1 –
FM4–64FX (molecular probes) [43]. Specimens were viewed using a Zeiss Axioskop2
motplus epifluorescence microscope (Carl Zeiss, Jena, Germany) using a ×100 objective in
conjunction with AxioVision software and a AxioCam MRm digital camera. Images were
acquired separately for each excitation-emission filter block combination appropriate to each
dye used (filter set for 365 nm excitation of DAPI, filter set for 450–490 nm excitation of
DiOC6(3) and filter set for 546 nm excitation of FM4–64). The resulting image files were
merged for final image production including brightness and contrast adjustment using the
indicated software.
Nano-Scale secondary-ion mass spectrometry (nanoSIMS)
A 1 ml mix containing dye-stock-solution plus R. baltica cells were incubated for 30 min.
R. baltica cells plus a) DiOC6(3); b) FM4–64; c) DiOC6(3) and FM4–64; were analysed. 5 µl
aliquot of each sample was taken for microscopic method control. Stained cells were collected
onto a gold-palladium (20:80) coated filter and washed with 20 ml Milli-Q-grade deionised
water. Samples were stored in the darkness and at room temperature until analysis. The filters
were analysed using a nanoSIMS 50 manufactured by CAMECA [44] at the University of
Mainz, Germany. Secondary ion images of the halogens bromide and iodine were recorded
simultaneously using 5 electron multipliers. Images and data were processed using the
proprietary CAMECA WinImage processing software.

Results and Discussion
72
Flow cytometry analysis modified after [45]
Samples used for flow cytometry and for the microscopy were taken from the same batch
cultures of R. baltica at different growth stages: early exponential (OD600 0.3), exponential
(OD600 0.5) and stationary phase (OD600 0.8). For flow cytometry 1%-PFA-fixed R. baltica
cells, stained with SYBR Green I DNA dye, were flow-sorted using a FACSort instrument
(Becton Dickinson) with standard laser and detection conFIGuration. Yellow-green 0.5 µm
bead standards (Fluoresbrite Microparticles) were used in all analyses as an internal standard.
Triplicates per sample were measured 1 min and displayed as a dotplot diagramm
(CellQuest), which is representing DNA-SYBR Green I fluorescence versus 90° light scatter
(SSC). The dotplot diagrams were used for defining gates. 5 gates (regions) were selected: R1
(beads), R2 (swarmer, single cells), R3 (budding cells), R4 (rosette formations), R5 (total cell
count). The assignment of the gates R2 to R4 arose from the corresponding microscopy
pictures.
Competing interests
The authors declare that they have no competing interests
Author’s contribution
PW conceived the study, initiated and conducted the experimental analysis, validated
microarray and optimised experimental steps, wrote the manuscript, did the statistical analysis
and analysed the data.
MS contributed background information and was involved in the writing and finishing of the
manuscript.
AE wrote MADA the microarray analysis tool and designed the microarrays.
FOG contributed background information and was involved in the writing of the manuscript.
All authors read and approved the final manuscript.
Acknowledgements
We would like to Bernhard Fuchs for the introduction of FACS-technology and Sebastian
Rokitta for excellent technical assistance. We also thank Niculina Musat for the introduction
into nanoSIMS-technology and her for the attendance to take the samples for analysing to the
University of Mainz.

Results and Discussion
73
The setup of the microarray and transcriptomic analysis of Rhodopirellula baltica was
supported by the European Commission contract GOCE-CT-2004-505403.
Abbreviations
COG Cluster of Orthologous Group
RB Rhodopirellula baltica
R. Rhodopirellula
GEO Gene Expression Omnibus
ORF Open Reading Frame
ECF Extra cytoplasmic Function
Fts filamentation temperature sensitive
FACS Fluorescence Activated Cell Sorting

Results and Discussion
74
Reference
1. Schlesner H: The development of media suitable for the microorganisms morphologically resembling Planctomyces spp., Pirellula spp., and other Planctomycetales from various aquatic habitats using dilute media. System Appl Microbiol 1994, 17:135-145.
2. Schlesner H, Rensmann C, Tindall BJ, Gade D, Rabus R, Pfeiffer S, Hirsch P: Taxonomic heterogeneity within the Planctomycetales as derived by DNA-DNA hybridization, description of Rhodopirellula baltica gen. nov., sp. nov., transfer of Pirellula marina to the genus Blastopirellula gen. nov. as Blastopirellula marina comb. nov. and emended description of the genus Pirellula. Int J Syst Evol Microbiol 2004, 54(Pt 5):1567-1580.
3. Ravenschlag K, Sahm K, Knoblauch C, Jorgensen BB, Amann R: Community structure, cellular rRNA content, and activity of sulfate-reducing bacteria in marine arctic sediments. Appl Environ Microbiol 2000, 66(8):3592-3602.
4. Gade D, Schlesner H, Glöckner FO, Amann R, Pfeiffer S, Thomm M: Identification of planctomycetes with order-, genus-, and strain-specific 16S rRNA-targeted probes. Microb Ecol 2004, 47(3):243-251.
5. Glöckner FO, Kube M, Bauer M, Teeling H, Lombardot T, Ludwig W, Gade D, Beck A, Borzym K, Heitmann K et al: Complete genome sequence of the marine planctomycete Pirellula sp. strain 1. Proc Natl Acad Sci U S A 2003, 100(14):8298-8303.
6. König E, Schlesner H, Hirsch P: Cell wall studies on budding bacteria of the Planctomyces/Pasteuria group and on a Prosthecomicrobium sp. Arch Microbiol 1984, 138:200-205.
7. Liesack W, König H, Schlesner H, Hirsch P: Chemical composition of the peptidoglycan-free cell envelopes of budding bacteria of the Pirella/Planctomyces group. Arch Microbiol 1986, 145:361-366.
8. Lindsay MR, Webb RI, Strous M, Jetten MS, Butler MK, Forde RJ, Fuerst JA: Cell compartmentalisation in planctomycetes: novel types of structural organisation for the bacterial cell. Arch Microbiol 2001, 175(6):413-429.
9. Jacobs-Wagner C: Regulatory proteins with a sense of direction: cell cycle signalling network in Caulobacter. Mol Microbiol 2004, 51(1):7-13.
10. Gade D, Stührmann T, Reinhardt R, Rabus R: Growth phase dependent regulation of protein composition in Rhodopirellula baltica. Environ Microbiol 2005, 7(8):1074-1084.
11. Margolin W: Themes and variations in prokaryotic cell division. FEMS Microbiology Reviews 2000, 24(4):531-548.
12. Teeling H: Aspekte der bioinformatischen Analyse und Annotation des Genoms von Rhodopirellula baltica. Dr. rer. nat. Bremen: Bremen; 2004.
13. Stephens C: Bacterial cell cycle: Seeing the big picture with microarrays. Current Biology 2001, 11(6):R222-R225.
14. Laub MT, McAdams HH, Feldblyum T, Fraser CM, Shapiro L: Global analysis of the genetic network controlling a bacterial cell cycle. Science 2000, 290(5499):2144-2148.
15. Spellman P, Sherlock G, Zhang Y-L, Vishwanath R, Anders K, Eisen MB, Brown PO, Botstein D, Futcher B: Comprehensive Identification of cell cycle-regulated genes of the yeast Saccharomyces cerevisiae by microarray hybridization. MBC 1998, 9:3273-3297.
16. Jenal U, Stephens C: The Caulobacter cell cycle: timing, spatial organization and checkpoints. Curr Opin Microbiol 2002, 5(6):558-563.
17. Jolly ER, Chin C-S, Miller S, Bahgat MM, Lim K, DeRisi J, McKerrow J: Gene expression patterns during adaptation of a helominth parasite to different environmental niches. Genome Biol 2007, 8.
18. Dougherty TJ, Pucci MJ: Penicillin-binding proteins are regulated by rpoS during transitions in growth states of Escherichia coli. Antimicrobial agents and chemotherapy 1994, 38(2):205-210.
19. Wecker P, Klockow C, Quast C, Langhammer P, Harder J, Glöckner F: Transcriptional response of the model planctomycete Rhodopirellula baltica SH1T to changing environmental conditions. in prep 2009.
20. Cluis CP, Burja AM, Martin VJJ: Current prospects for the production of coenzyme Q10 in microbes. Trends in Biotechnology 2007, 25(11):514-521.
21. Kerfeld CA, Sawaya MR, Tanaka S, Nguyen CV, Phillips M, Beeby M, Yeates TO: Protein structures forming the shell of primitive bacterial organelles. Science 2005, 309(5736):936-938.
22. Sowell SM, Norberck AD, Lipton MS, Nicora CD, Callister SJ, Smith RD, Barofsky DF, Giovannoni SJ: Proteomic Analysis of stationary phase in the marine Bacterium "Candidatus Pelagibacter ubique". Appl Environ Microbiol 2008, 74(13):4091-4100.

Results and Discussion
75
23. Kumar A: Bacterial exopolysaccharides - a perception. J Bas Microbiol 2007, 47:103-117. 24. Lindsay MR, Webb RI, Fuerst J: Pirellulosomes: a new type pf membranebounded cell
compartment in planctomycete bacteria of the genus Pirellula. Microbiology 1997, 143:739-748. 25. Bauer M, Lombardot T, Teeling H, Ward N, Amann R, Glöckner FO: Archaea-like genes for C1-
transfer enzymes in Planctomycetes: Phylogenetic implications of their unexpected presence in this phylum. J Mol Evol 2004, 59:571-586.
26. Woebken D, Teeling H, Wecker P, Dumitriu A, Kostadinov I, DeLong EF, Amann R, Glöckner FO: Fosmids of novel marine Planctomycetes from the Namibian and Oregon coast upwelling systems and their cross-comparison with planctomycete genomes. ISME Journal 2007, 1(5):419-435.
27. Chistoserdova L, Jenkins C, Kalyuzhnaya M, Marx C, Lapidus A, Vorholt J: The enigmatic Planctomycetes may hold a key to the origins of methanogenesis and methylotrophy. Mol Biol Evol 2004(21):1234-1241.
28. Susin MF, Baldini RL, Gueiros-Filho F, Gomes SL: GroES/GroEL and DnaK/DnaJ have distinct roles in stress responses and during cell cycle progression in Caulobacter crescentus. J Bacteriol 2006, 188(23):8044-8053.
29. Wecke T, Veith B, Ehrenreich A, Mascher T: Cell envelope stress response in Bacillus licheniformis: Integrating comparative genomics, transcriptional profiling, and regulon mining to decipher a complex regulatory network. In., vol. 188; 2006: 7500-7511.
30. Eiamphungporn W, Helmann JD: The Bacillus subtilis sigma M regulon and its contribution to cell envelope stress responses. 2008, 67(4):830-848.
31. Studholme DJ, Fuerst JA, Bateman A: Novel protein domains and motifs in the marine planctomycete Rhodopirellula baltica. FEMS Microbiol Lett 2004, 236:333-340.
32. Lee K-C, Webb RI, Fuerst AJ: The cell cycle of the planctomycete Gemmata obscuriglobus with respect to cell compartmentalization. BMC Cell Biology 2009, 10(4).
33. Rabus R, Gade D, Helbig R, Bauer M, Glöckner FO, Kube M, Schlesner H, Reinhardt R, Amann R: Analysis of N-acetylglucosamine metabolism in the marine bacterium Pirellula sp. strain 1 by a proteomic approach. Proteomics 2002, 2(6):649-655.
34. Gene Expression Omnibus (GEO) database [http://www.ncbi.nlm.nih.gov/geo/] 35. Microarray data analysis software tool MADA [http://www.megx.net/mada] 36. Soukas A, Cohen P, Socci ND, Friedman JM: Leptin-specific patterns of gene expression in white
adipose tissue. Genes Dev 2000, 14(8):963-980. 37. Saeed AI, Sharov V, White J, Li J, Liang W, Bhagabati N, Braisted J, Klapa M, Currier T, Thiagarajan
M et al: TM4: a free, open-source system for microarray data management and analysis. Biotechniques 2003, 34(2):374-378.
38. Quast C: MicHanThi - Design and Implementation of a System for the Prediction of Gene Functions in Genome Annotation Projects. In: Technical Report. Edited by Informatik T-Z. Bremen: TZI-Bremen; 2007.
39. The reannotated Rhodopirellula baltica genome [http://gendb.mpi-bremen.de/gendb/BX119912] 40. Richter M, Lombardot T, Kostadinov I, Kottmann R, Duhaime M, Peplies J, Glöckner F: JCoast - A
biologist-centric software tool for data mining and comparison of prokaryotic (meta)genomes. BMC Bioinformatics 2008, 9(1):177.
41. Meyer F, Goesmann A, McHardy AC, Bartels D, Bekel T, Clausen J, Kalinowski J, Linke B, Rupp O, Giegerich R et al: GenDB - an open source genome annotation system for prokaryote genomes. Nucl Acids Res 2003, 31(8):2187-2195.
42. Hulsen T, de Vlieg J, Alkema W: BioVenn - a web application for the comparison and visualization of biological lists using area-proportional Venn diagrams. BMC Genomics 2008, 9(1):488.
43. Bolte S, Talbot C, Boutte Y, Catrice O, Read ND, Satiat-Jeunemaitre B: FM-dyes as experimental probes for dissecting vesicle trafficking in living plant cells. Journal of Microscopy 2004, 214:159-173.
44. Musat N, Halm H, Winterholler B, Hoppe P, Peduzzi S, Hillion F, Horreard F, Amann R, Jörgensen BB, Kuypers MMM: A single-cell view on the ecophysiology of anaerobic phototrophic bacteria. In., vol. 105; 2008: 17861-17866.
45. Mary I, Heywood JL, Fuchs BM, Amann R, Tarran GA, Burkill PH, Zubkov MV: SAR11 dominace among metabolically active low nucleic acid bacterioplankton in surface waters along the Atlantica meridional transcet. Aquatic Microbial Ecology 2006, 45:107-113.
46. Fuerst AJ: Intracellular compartmentation in Planctomycetes. Annu Rev Microbiol 2005, 59:299-328.
47. Gade D, Theiss D, Lange D, Mirgorodskaya E, Lombardot T, Glöckner FO, Kube M, Reinhardt R, Amann R, Lehrach H et al: Towards the proteome of the marine bacterium Rhodopirellula baltica: Mapping the soluble proteins. Proteomics 2005, 5(14):3654-3671.
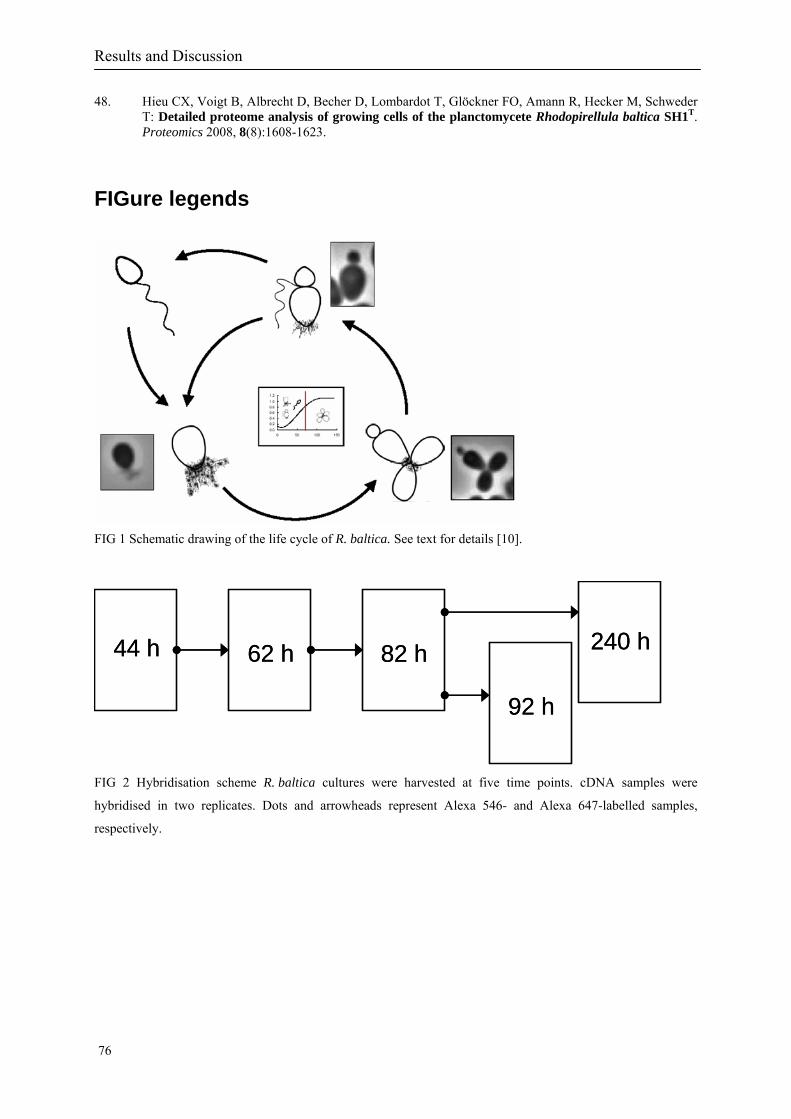
Results and Discussion
76
48. Hieu CX, Voigt B, Albrecht D, Becher D, Lombardot T, Glöckner FO, Amann R, Hecker M, Schweder T: Detailed proteome analysis of growing cells of the planctomycete Rhodopirellula baltica SH1T. Proteomics 2008, 8(8):1608-1623.
FIGure legends
FIG 1 Schematic drawing of the life cycle of R. baltica. See text for details [10].
44 h 62 h 82 h
92 h
240 h44 h 62 h 82 h
92 h
240 h
FIG 2 Hybridisation scheme R. baltica cultures were harvested at five time points. cDNA samples were
hybridised in two replicates. Dots and arrowheads represent Alexa 546- and Alexa 647-labelled samples,
respectively.
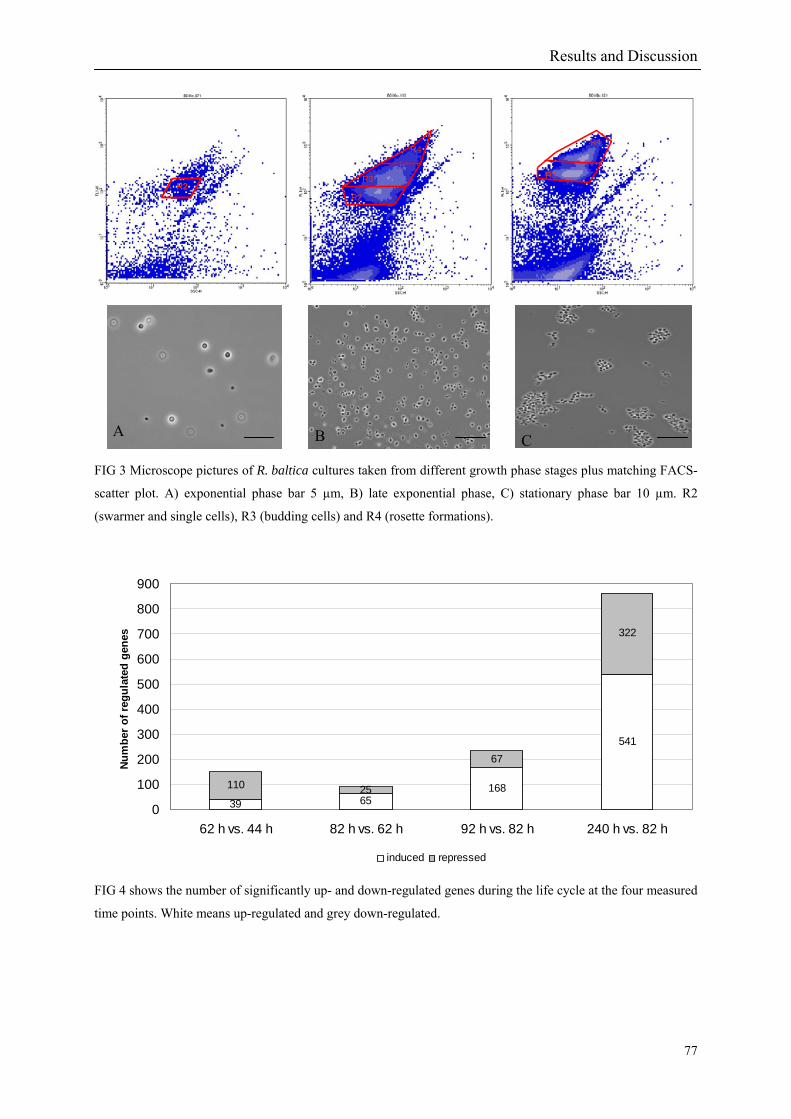
Results and Discussion
77
FIG 3 Microscope pictures of R. baltica cultures taken from different growth phase stages plus matching FACS-
scatter plot. A) exponential phase bar 5 µm, B) late exponential phase, C) stationary phase bar 10 µm. R2
(swarmer and single cells), R3 (budding cells) and R4 (rosette formations).
39 65168
541
110 25
67
322
0
100
200
300
400
500
600
700
800
900
62 h vs. 44 h 82 h vs. 62 h 92 h vs. 82 h 240 h vs. 82 h
Num
ber o
f reg
ulat
ed g
enes
induced repressed
FIG 4 shows the number of significantly up- and down-regulated genes during the life cycle at the four measured
time points. White means up-regulated and grey down-regulated.
R3
R2
R4
R3
R2
R4
R3
R4
R3
R4
R2R2R2
A B C
Results and Discussion
78
210
66
132
4
76
1492 h vs. 82 h
82 h vs. 62 h
44 h vs. 62 h
210
66
132
4
76
1492 h vs. 82 h
82 h vs. 62 h
44 h vs. 62 h
103634
12 114
720
38
92 h vs. 82 h82 h vs. 62 h
240 h vs. 82 h
103634
12 114
720
38
92 h vs. 82 h82 h vs. 62 h
240 h vs. 82 h
FIG 5 Venn diagrams showing the distribution of all regulated genes.
Cytoplasmic membraneFM4-64
Intracytoplasmic membraneDiOC6 (3)
Nucleoid DAPI
Cell wallPirellulosome Paryphoplasma
Cytoplasmic membraneFM4-64
Intracytoplasmic membraneDiOC6 (3)
Nucleoid DAPI
Cell wallPirellulosome Paryphoplasma
Cytoplasmic membraneFM4-64
Intracytoplasmic membraneDiOC6 (3)
Nucleoid DAPI
Cell wallPirellulosome Paryphoplasma
Intracytoplasmic membraneDiOC6 (3)
Nucleoid DAPI
Cell wallPirellulosome Paryphoplasma
FIG 6 Diagrams of cell organisation and compartmentalisation in R. baltica modified after Fuerst et al. [46].
DNA-specific fluorescence dye DAPI (blue) stains nucleoid, membrane-specific dye FM4-64 (red) stains the
external cytoplasm membrane and membrane-specific dye DiOC6(3) (green) passes through the external
membrane and stains the internal membranes.
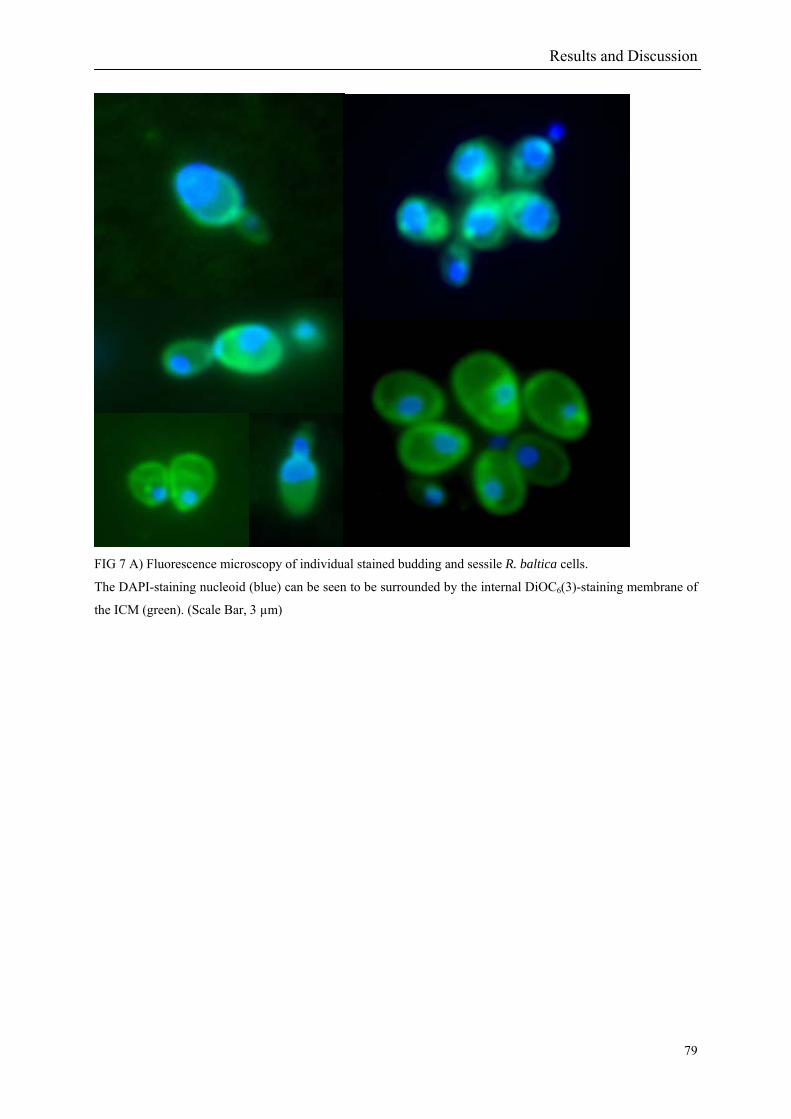
Results and Discussion
79
FIG 7 A) Fluorescence microscopy of individual stained budding and sessile R. baltica cells.
The DAPI-staining nucleoid (blue) can be seen to be surrounded by the internal DiOC6(3)-staining membrane of
the ICM (green). (Scale Bar, 3 µm)
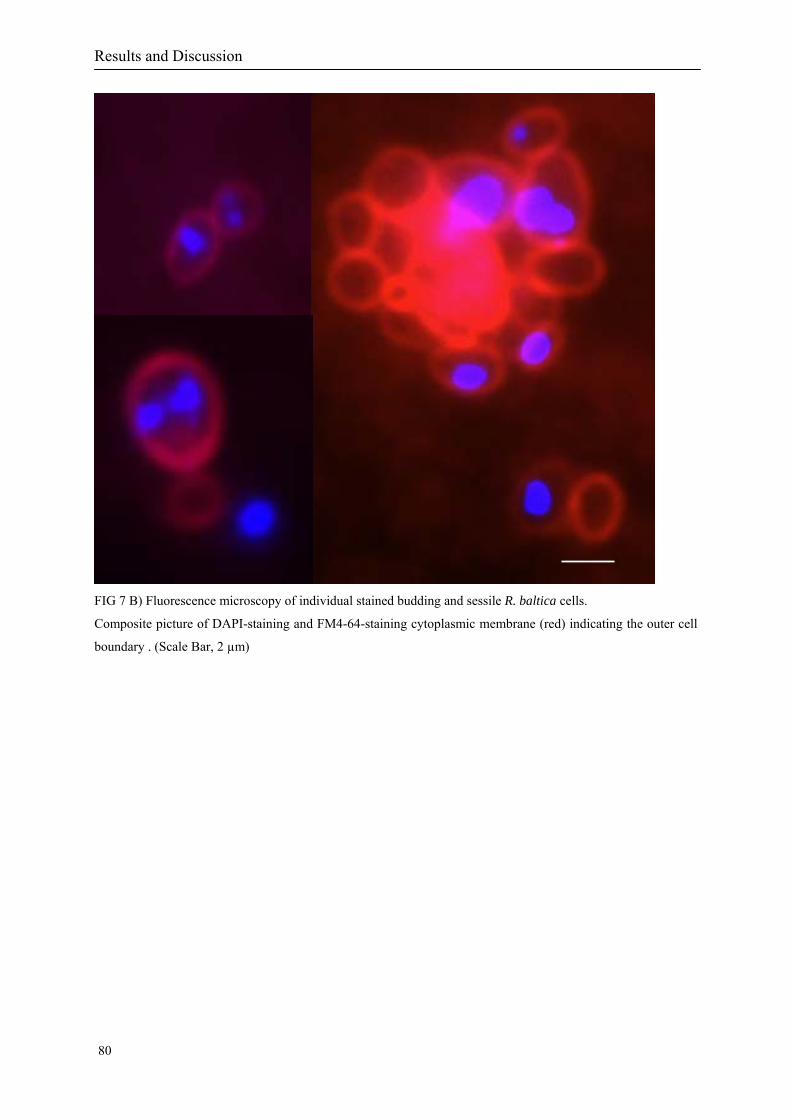
Results and Discussion
80
FIG 7 B) Fluorescence microscopy of individual stained budding and sessile R. baltica cells.
Composite picture of DAPI-staining and FM4-64-staining cytoplasmic membrane (red) indicating the outer cell
boundary . (Scale Bar, 2 µm)
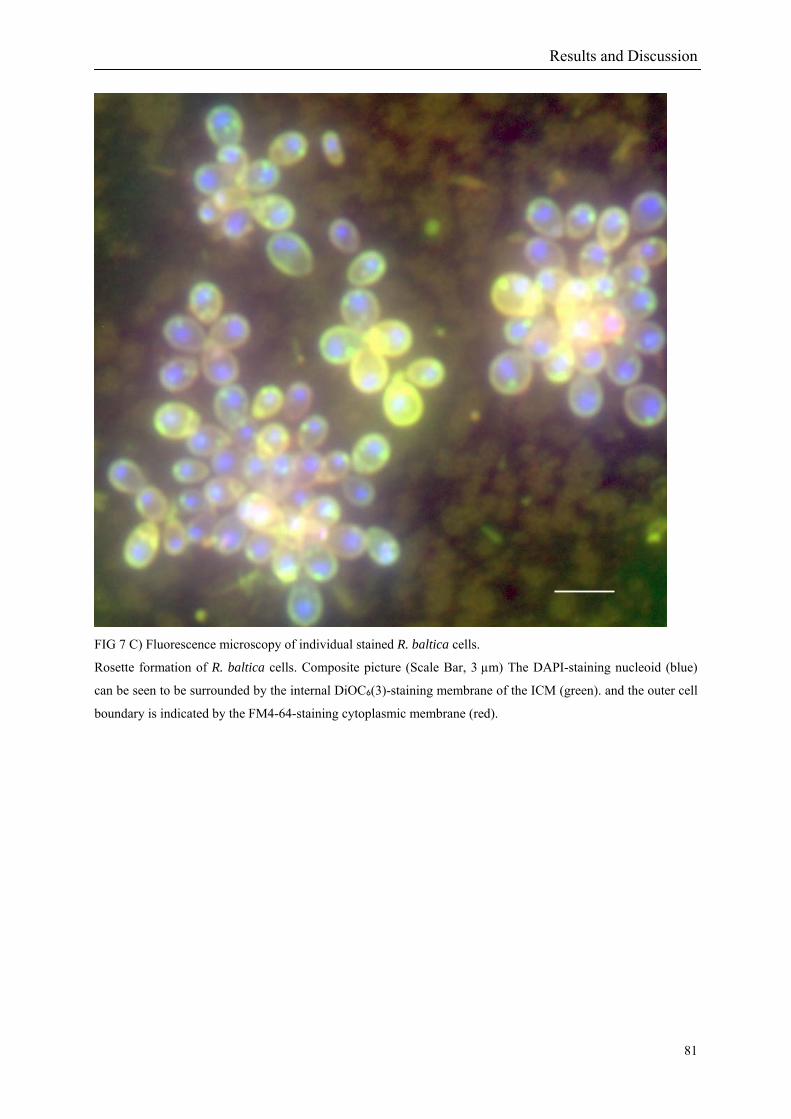
Results and Discussion
81
FIG 7 C) Fluorescence microscopy of individual stained R. baltica cells.
Rosette formation of R. baltica cells. Composite picture (Scale Bar, 3 µm) The DAPI-staining nucleoid (blue)
can be seen to be surrounded by the internal DiOC6(3)-staining membrane of the ICM (green). and the outer cell
boundary is indicated by the FM4-64-staining cytoplasmic membrane (red).
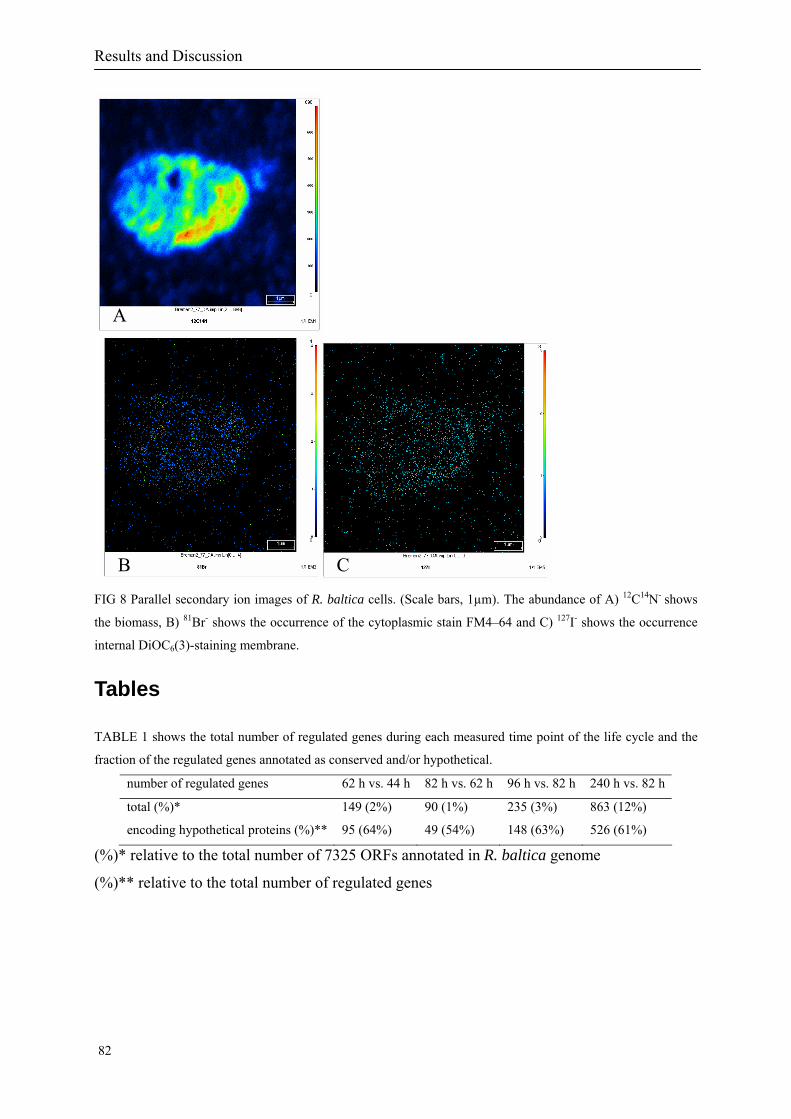
Results and Discussion
82
FIG 8 Parallel secondary ion images of R. baltica cells. (Scale bars, 1µm). The abundance of A) 12C14N- shows
the biomass, B) 81Br- shows the occurrence of the cytoplasmic stain FM4–64 and C) 127I- shows the occurrence
internal DiOC6(3)-staining membrane.
Tables
TABLE 1 shows the total number of regulated genes during each measured time point of the life cycle and the
fraction of the regulated genes annotated as conserved and/or hypothetical.
number of regulated genes 62 h vs. 44 h 82 h vs. 62 h 96 h vs. 82 h 240 h vs. 82 h
total (%)* 149 (2%) 90 (1%) 235 (3%) 863 (12%)
encoding hypothetical proteins (%)** 95 (64%) 49 (54%) 148 (63%) 526 (61%)
(%)* relative to the total number of 7325 ORFs annotated in R. baltica genome
(%)** relative to the total number of regulated genes
A
B C
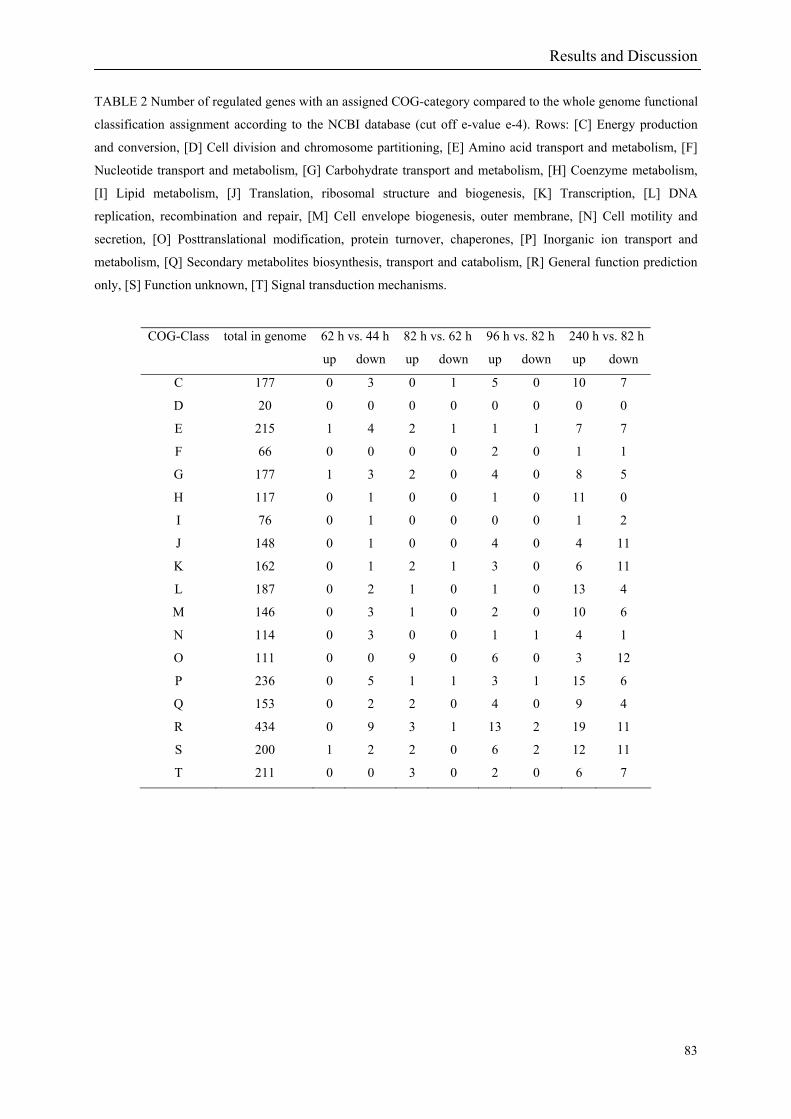
Results and Discussion
83
TABLE 2 Number of regulated genes with an assigned COG-category compared to the whole genome functional
classification assignment according to the NCBI database (cut off e-value e-4). Rows: [C] Energy production
and conversion, [D] Cell division and chromosome partitioning, [E] Amino acid transport and metabolism, [F]
Nucleotide transport and metabolism, [G] Carbohydrate transport and metabolism, [H] Coenzyme metabolism,
[I] Lipid metabolism, [J] Translation, ribosomal structure and biogenesis, [K] Transcription, [L] DNA
replication, recombination and repair, [M] Cell envelope biogenesis, outer membrane, [N] Cell motility and
secretion, [O] Posttranslational modification, protein turnover, chaperones, [P] Inorganic ion transport and
metabolism, [Q] Secondary metabolites biosynthesis, transport and catabolism, [R] General function prediction
only, [S] Function unknown, [T] Signal transduction mechanisms.
COG-Class total in genome 62 h vs. 44 h 82 h vs. 62 h 96 h vs. 82 h 240 h vs. 82 h
up down up down up down up down
C 177 0 3 0 1 5 0 10 7
D 20 0 0 0 0 0 0 0 0
E 215 1 4 2 1 1 1 7 7
F 66 0 0 0 0 2 0 1 1
G 177 1 3 2 0 4 0 8 5
H 117 0 1 0 0 1 0 11 0
I 76 0 1 0 0 0 0 1 2
J 148 0 1 0 0 4 0 4 11
K 162 0 1 2 1 3 0 6 11
L 187 0 2 1 0 1 0 13 4
M 146 0 3 1 0 2 0 10 6
N 114 0 3 0 0 1 1 4 1
O 111 0 0 9 0 6 0 3 12
P 236 0 5 1 1 3 1 15 6
Q 153 0 2 2 0 4 0 9 4
R 434 0 9 3 1 13 2 19 11
S 200 1 2 2 0 6 2 12 11
T 211 0 0 3 0 2 0 6 7
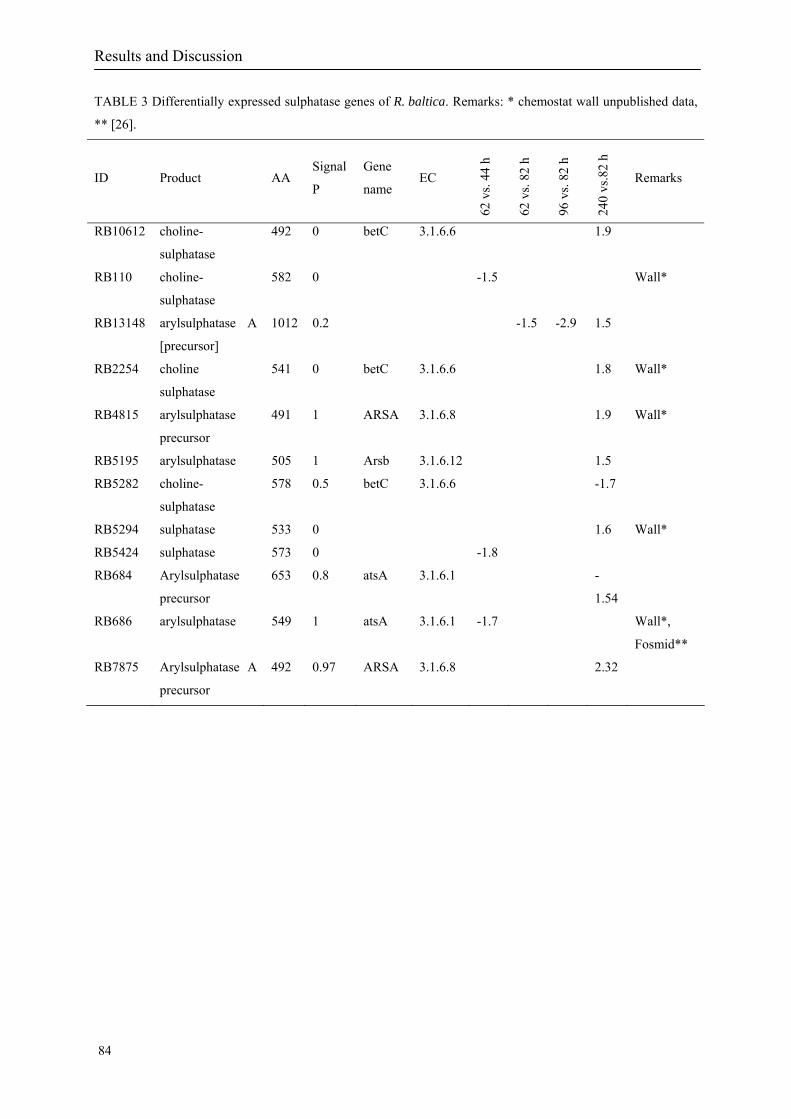
Results and Discussion
84
TABLE 3 Differentially expressed sulphatase genes of R. baltica. Remarks: * chemostat wall unpublished data,
** [26].
ID Product AA Signal
P
Gene
name EC
62
vs. 4
4 h
62
vs. 8
2 h
96
vs. 8
2 h
240
vs.8
2 h
Remarks
RB10612 choline-
sulphatase
492 0 betC 3.1.6.6 1.9
RB110 choline-
sulphatase
582 0 -1.5 Wall*
RB13148 arylsulphatase A
[precursor]
1012 0.2 -1.5 -2.9 1.5
RB2254 choline
sulphatase
541 0 betC 3.1.6.6 1.8 Wall*
RB4815 arylsulphatase
precursor
491 1 ARSA 3.1.6.8 1.9 Wall*
RB5195 arylsulphatase 505 1 Arsb 3.1.6.12 1.5
RB5282 choline-
sulphatase
578 0.5 betC 3.1.6.6 -1.7
RB5294 sulphatase 533 0 1.6 Wall*
RB5424 sulphatase 573 0 -1.8
RB684 Arylsulphatase
precursor
653 0.8 atsA 3.1.6.1 -
1.54
RB686 arylsulphatase 549 1 atsA 3.1.6.1 -1.7 Wall*,
Fosmid**
RB7875 Arylsulphatase A
precursor
492 0.97 ARSA 3.1.6.8 2.32
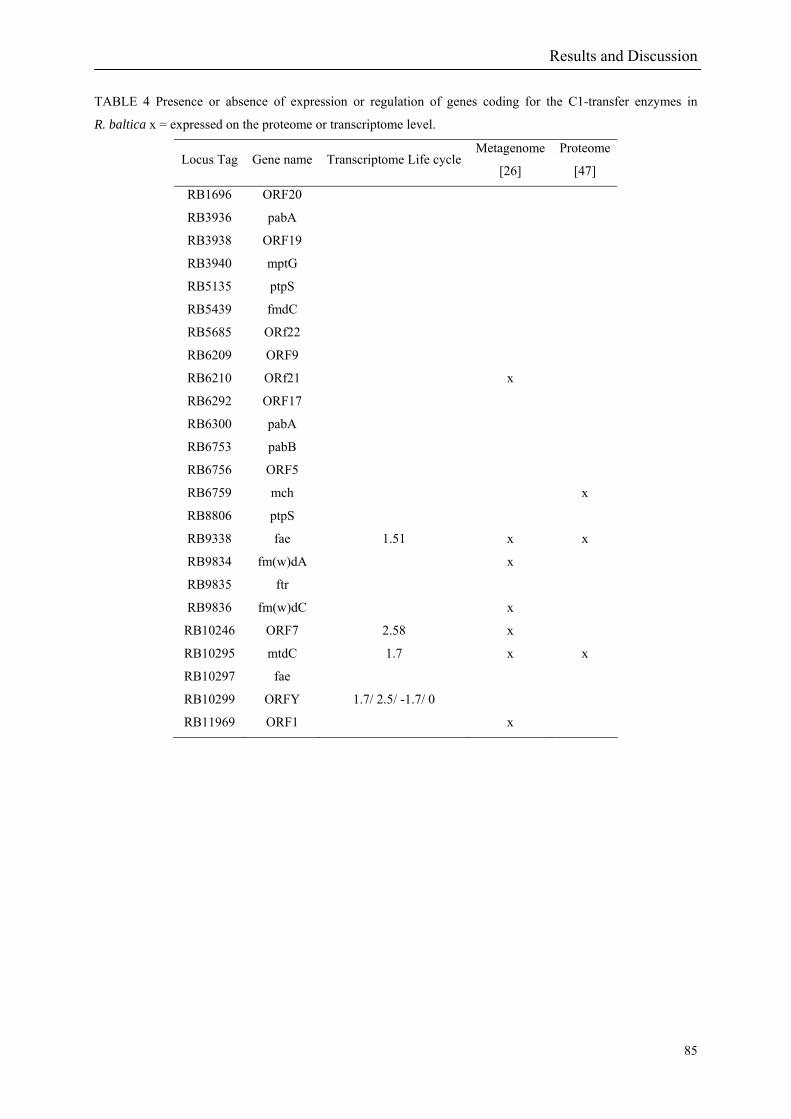
Results and Discussion
85
TABLE 4 Presence or absence of expression or regulation of genes coding for the C1-transfer enzymes in
R. baltica x = expressed on the proteome or transcriptome level.
Locus Tag Gene name Transcriptome Life cycle Metagenome
[26]
Proteome
[47]
RB1696 ORF20
RB3936 pabA
RB3938 ORF19
RB3940 mptG
RB5135 ptpS
RB5439 fmdC
RB5685 ORf22
RB6209 ORF9
RB6210 ORf21 x
RB6292 ORF17
RB6300 pabA
RB6753 pabB
RB6756 ORF5
RB6759 mch x
RB8806 ptpS
RB9338 fae 1.51 x x
RB9834 fm(w)dA x
RB9835 ftr
RB9836 fm(w)dC x
RB10246 ORF7 2.58 x
RB10295 mtdC 1.7 x x
RB10297 fae
RB10299 ORFY 1.7/ 2.5/ -1.7/ 0
RB11969 ORF1 x
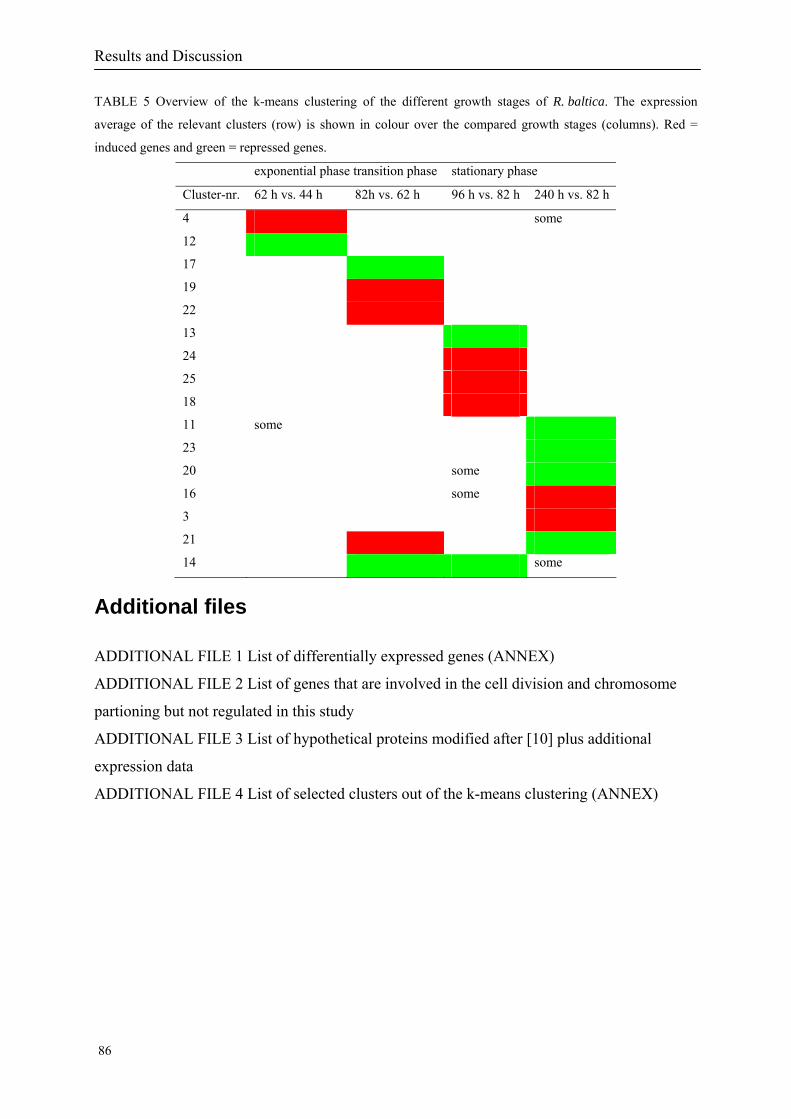
Results and Discussion
86
TABLE 5 Overview of the k-means clustering of the different growth stages of R. baltica. The expression
average of the relevant clusters (row) is shown in colour over the compared growth stages (columns). Red =
induced genes and green = repressed genes.
exponential phase transition phase stationary phase
Cluster-nr. 62 h vs. 44 h 82h vs. 62 h 96 h vs. 82 h 240 h vs. 82 h
4 some
12
17
19
22
13
24
25
18
11 some
23
20 some
16 some
3
21
14 some
Additional files
ADDITIONAL FILE 1 List of differentially expressed genes (ANNEX)
ADDITIONAL FILE 2 List of genes that are involved in the cell division and chromosome
partioning but not regulated in this study
ADDITIONAL FILE 3 List of hypothetical proteins modified after [10] plus additional
expression data
ADDITIONAL FILE 4 List of selected clusters out of the k-means clustering (ANNEX)

Results and Discussion
87
Additional file 2 ID Locus AA Product Gene EC COG
526 RB965 156 membrane protein D
882 RB1641 339 sodium/calcium exchanger protein, D
1145 RB2176 125 hypothetical protein D
1195 RB2264 168 Large-conductance mechanosensitive channel mscL D
1303 RB2434 81 protein containing RNA-binding S4 domain S4 D
1531 RB2833 462 dehydrogenase D
2077 RB3771 222 Putative cyclase D
2241 RB4054 87 hypothetical protein D
2903 RB5151 415 threonine dehydratase, biosynthetic, ilvA 4.3.1.19 D
3162 RB5613 360 protein containing DUF1006 D
3408 RB6045 627 membrane protein D
3628 RB6438 485 secreted protein D
3856 RB6827 966 serine/threonine-protein kinase 2.7.1.37 D
4297 RB7620 49 hypothetical protein D
4966 RB8859 332 D-lactate dehydrogenase ldhA 1.1.1.28 D
5165 RB9239 231 conserved hypothetical protein D
5329 RB9546 276 conserved hypothetical protein, D
5793 RB10397 314 hypothetical protein D
6585 RB11813 80 conserved hypothetical protein D
7263 RB13068 499 conserved hypothetical protein D
745 RB1391 538 cell division inhibitor-
2302 RB4173 337 signal recognition particle-docking protein FtsY
3138 RB5566 728 cell division protein FtsH 3.4.24.-
1607 RB2966 672 cell division protein FtsH 3.4.24.-
3306 RB5871 485 Cell division protein
4490 RB7945 231 glucose-inhibited division protein
5170 RB9255 667 glucose-inhibited division protein gidA
6587 RB11817 937 DNA translocase FtsK
2091 RB3794 1412 FtsK/SpoIIIE family protein
5504 RB9874 267 segregation and condensation gene scpA
3926 RB6962 297 segregation and condensation gene scpB
3418 RB6065 1234 chromosome segregation protein
6463 RB11587 540 segregation and condensation gene scpB
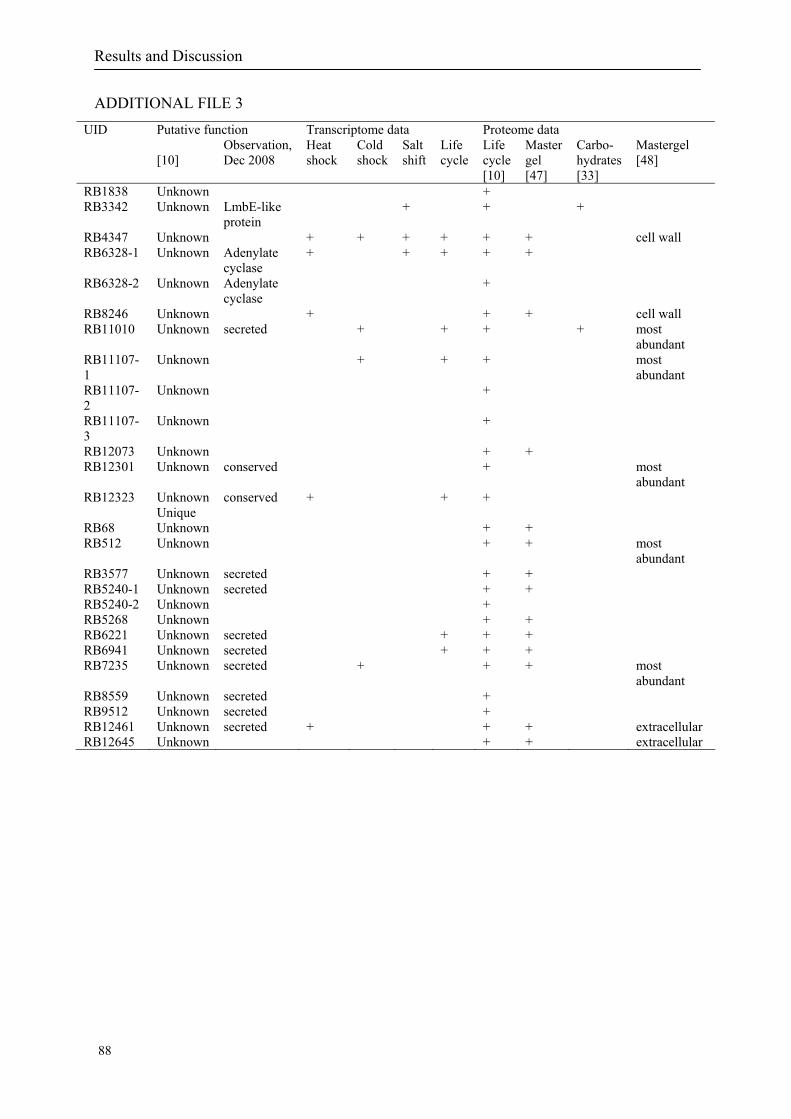
Results and Discussion
88
ADDITIONAL FILE 3 UID Putative function Transcriptome data Proteome data
[10] Observation, Dec 2008
Heat shock
Cold shock
Salt shift
Life cycle
Life cycle [10]
Master gel [47]
Carbo-hydrates [33]
Mastergel [48]
RB1838 Unknown + RB3342 Unknown LmbE-like
protein + + +
RB4347 Unknown + + + + + + cell wall RB6328-1 Unknown Adenylate
cyclase + + + + +
RB6328-2 Unknown Adenylate cyclase
+
RB8246 Unknown + + + cell wall RB11010 Unknown secreted + + + + most
abundant RB11107-1
Unknown + + + most abundant
RB11107-2
Unknown +
RB11107-3
Unknown +
RB12073 Unknown + + RB12301 Unknown conserved + most
abundant RB12323 Unknown conserved + + + Unique RB68 Unknown + + RB512 Unknown + + most
abundant RB3577 Unknown secreted + + RB5240-1 Unknown secreted + + RB5240-2 Unknown + RB5268 Unknown + + RB6221 Unknown secreted + + + RB6941 Unknown secreted + + + RB7235 Unknown secreted + + + most
abundant RB8559 Unknown secreted + RB9512 Unknown secreted + RB12461 Unknown secreted + + + extracellular RB12645 Unknown + + extracellular


Results and Discussion
89
3.3 Highly enantioselective sec-alkyl sulfatase activity of the marine planctomycete Rhodopirellula baltica shows retention of configuration
Sabine R. Wallner, Marga Bauer, Chris Würdemann, Patricia Wecker, Frank O. Glöckner,
and Kurt Faber
Published in Angewandte Chemie International Edition 44: 2 –4 (2005)

Results and Discussion
90
3.4 Fosmids of novel marine Planctomycetes from the Namibian and Oregon coast upwelling systems and their cross-comparison with planctomycete genomes
Authors: Dagmar Woebken, Hanno Teeling, Patricia Wecker, Alexandra Dumitriu, Ivaylo
Kostadinov, Rudolf Amann, Edward F Delong, and Frank Oliver Glöckner 2007
Published in The ISME Journal 1, 419-435 (2007)

Summary
91
4 SUMMARY In 2003, the completely sequenced and annotated genome of the first planctomycete
Rhodopirellula baltica was published (Glöckner et al. 2003). Besides common genomic
features including Glycolysis, Citrate–cycle or ribosomal RNAs the annotation also revealed
unexpected genetic potential such as a high number of sulphatase genes or genes encoding
enzymes of the C1-metabolism pathway. In conclusion, bioinformatic analysis allowed new
hypotheses to be built about the capabilities and the ecological niche of R. baltica. However,
bioinformatics could not resolve open questions about many features, for example, the
interesting cell-cycle or character and the formation of the holdfast substance. In addition,
bioinformatics could not assign a function to 55% of the predicted ORFs. The annotation
contributed to the understanding of the phylum Planctomycetes but also made clear how poor
our knowledge remains concerning the genetic potential of many environmental organisms.
To turn the information about the genetic potential of an organism into sound biological
knowledge, functional post-genomics using transcriptomics and proteomics are the next
logical step. To further substantiate the predicted functions and get hints for some of the
unknown gene functions, microarray technology was selected. Whole genome expression
profiling allows the investigation of thousands of genes in parallel at the mRNA level in a
single experiment. This is a clear advantage compared to other techniques like Real-Time
PCR, which allows investigation of only a rather small set of genes.
During my thesis I set up a pipeline (2.3) for the first planctomycete-specific whole genome
array. This included testing and selection of the slide format and its chemistry. Protocols for
RNA isolation, cDNA synthesis and labelling, required adaptation due to the specific
proteinaceous cell wall of R. baltica. Hybridisation and washing buffer composition as well as
hybridisation time and conditions were adjusted for an optimal signal to background ratio
based on an in-house automatic hybridisation machine (Tecan HS400). The quality of the
array hybridisation was validated by checking for the negative, positive and stringency
controls printed on the array. The pipeline also included the set up of three chemostats for
improving the R. baltica culturing compared to the initially used batch culture systems.
The processing of the resulting expression data included reviewing different data
normalisation approaches and selection of the one which fits the experimental design. For the
analysis of the normalised and filtered expression data, various microarray data analysis tools
were screened. Finally, the open source software program TIGR Multiexperiment Viewer

Summary
92
(MeV) was chosen. This incorporates sophisticated algorithms for clustering, visualisation,
classification, statistical analysis and biological interpretation.
The successful establishment of microarray technology led to the second part of the thesis
dealing with whole genome array expression profiling to address specific questions. The
whole genome array was applied to R. baltica cells exposed to different temperatures and high
salinity in order to learn more about the common and specific stress responses of this
organism (Wecker et al. 2009in prep). The whole genome array was also applied to R. baltica
cells in different growth stages with the aim to investigate and analyse unknown mechanisms
during the cell-cycle (Wecker et al. 2009 in prep).
The following section provides a brief summary of the (main) results obtained in this thesis.
For details, please refer to the original publications which can be found in chapter 3.
4.1 Transcriptional response of the model planctomycete Rhodopirellula baltica SH1T to changing environmental conditions
The first manuscript introduces the first whole genome array for any member of the phylum
Planctomycetes, specifically Rhodopirellula baltica. The successful establishment of a
complete microarray pipeline is demonstrated by a model system based on comparisons of the
behaviour of R. baltica cells at different environmental stress factors at the transcriptome
level.
The first experiments focused on the better understanding of the adaptations of R. baltica
grown under changing environmental conditions, e.g. variations in temperature and high
salinity. The rationale behind these experiments is the observation that R. baltica can be
isolated from various marine and brackish habitats while bioinformatic analyses of the
genome support the hypothesis that R. baltica passes through a halocline while passing from
the surface to the sediment. Stress responses to salinity and temperature shifts were monitored
in time series experiments. Cultures grown on mineral media at 28°C were compared to
cultures that were shifted to either elevated (37°C) or reduced (6°C) temperatures or high
salinity (59.5‰) over 300 min.
Transcriptional profiling confirmed that R. baltica is highly responsive to its environment.
Heat shock showed the induction of several known chaperone genes while cold shock altered
the expression of genes involved in lipid metabolism and stress protein production. High
salinity resulted in the modulation of genes coding for compatible solutes, ion transporters
and morphology. We were also able to detect a shared environmental stress response of

Summary
93
R. baltica. Results show the induction of known general stress response genes. The ribosomal
machinery is repressed and is probably functioning as a general stress sensor. Transposase
genes are induced to enhance this genetic activity through rearrangement of the genome.
Noteworthy is the shared induction of three ECF- sigma factors over all three experiments,
which led to the assumption that they are part of the environmental stress signalling pathway.
Another highlight is the regulation of several sulphatase genes in all experiments. We propose
a role for these in the remodelling of the cell wall of R. baltica and point out the diverse role
of the sulphatases. Nearly 50% of all regulated genes have no known function. The
combination of transcriptomics and proteomics points to a set of genes of unknown functions
most probably involved in the global stress response. In summary this work gave new insights
into the complex adaptation machinery of an environmentally relevant marine bacterium. This
work lays the foundation for further bioinformatic and genetic studies, which leads to a
comprehensive understanding of the biology of a model marine Planctomycete.
The corresponding paper is submitted to BMC Genomics.
4.2 The Rhodopirellula baltica life-cycle: Growth-dependent expression profiling of the complete genome with microarrays
The focus of the second manuscript is on the expression profiling of different growth stages of
the marine planctomycete Rhodopirellula baltica. Although synchronised cell cultures of
R. baltica were not available, it was possible to get first idea about the genes involved in the
characteristic morphological stage of Planctomycetes.
The marine planctomycete R. baltica has a life-cycle consisting of motile swarmer cells and
sessile adult cells. During the genome annotation none of the known genes involved in the
cell-cycle or cell division were found. The life-cycle must therefore have a so far unknown
control mechanism. To find out more about the genes involved in the life-cycle, a whole
genome array for R. baltica was used for the first time. R. baltica cell samples were taken at
different growth stages from a R. baltica culture and growth dependent expression profiling
was carried out. The number of regulated genes increased from 90 exponentially to 863 in the
late stationary phase when compared to the transition phase. Results indicate a remodelling of
the cell wall and membrane in the exponential phase, sensing of environmental changes in the
transition phase, followed by a stationary-phase specific response. 12 out of 110 sulphatases
genes showed a growth-dependent expression pattern. This indicates that some of them might
also be involved in basic cell functionalities such as the remodelling of cell wall components
during the life-cycle. It is also noteworthy that genes associated with the C1-metabolism

Summary
94
appear to be regulated. More than half of the regulated genes per growth stage were annotated
as hypothetical proteins. These genes are likely candidates for functions associated with
planctomycete-specific morphology and/or morphotype-differentiation during the life-cycle.
In summary, the whole genome array of R. baltica gave initial insights into the regulation of
genes during the life-cycle. The transcriptomic results are contributing to a better
understanding of the so far unknown molecular cell biology underlying the cell
compartmentalisation of the phylum Planctomycetales and maybe even of the PVC
superphylum.
The corresponding paper is in preparation.
4.3 Highyl enantioselective sec-alkyl sulfatase activity of the marine planctomycete Rhodopirellula baltica shows retention of configuration
The preliminary results of the partial genome array targeting sulphatase genes (C.
Würdemann unpublished results) and the parallel investigations concerning the sulphatase
activity in R. baltica were the fundamental basis for research aim 2.2.
The annotation process of the complete genome sequence of the marine planctomycete
Rhodopirellula baltica revealed that the genome includes 109 genes assigned as sulphatases.
A subset of 85 sulphatases showed the conserved “core” of the sulphatase consensus motif,
which is required to render activated sulphatase proteins. Each of these 85 sequences
contained the adjacent “auxiliary” part of the consensus motif, either fully conserved or only
slightly modified. The sulphatases are active in resting cells of R. baltica and act through
retention of configuration (shown in collaboration with Prof. Dr. Faber University of Graz,
Austria). In general, the enantio preference was shown to be “R”, while “S” enantiomers
remained untouched. R. baltica displayed high activities on linear (w-1)-sulphate esters,
which were readily accepted as substrates. Even sterically demanding sulcatyl sulphate was
converted with good rates, which was neither accepted by an inverting alkyl sulphatase
(termed “RS2”) nor by Sulfolobus spp.. Not surprisingly was the quick conversion of the
primary sulphate ester by one of the large set of sulphatases genes. However, no loss of
sulphatase activity in the protein extract of R. baltica cells grown on various C-sources:
glucose, chondroitin sulphate A and condroitin sulphate C could be seen.
The corresponding paper has been published in Angewandte Chemie International Edition 44:
2 –4 (2005).

Summary
95
4.4 Fosmids of novel marine Planctomycetes from the Namibian and Oregon coast upwelling systems and their cross-comparison and their cross-comparison with planctomycete genomes
This metagenomic study of fosmids of novel marine Planctomycetes is a good example of the
knowledge transfer of transcriptome data from model organisms to the ecosystem with the
purpose to understand the role of Planctomycetes in the environment.
The objective of the study was to broaden our knowledge about the genetic potential of
marine Planctomycetes using a cultivation independent approach. Therefore, six fosmids from
two different marine upwelling systems were investigated from metagenome libraries. The
fosmids were compared with all available planctomycete genomic sequences (Rhodopirellula
baltica SH 1T, Candidatus Kuenenia stuttgartiensis and Gemmata obscuriglobus DSM 2246T)
including the as yet unpublished near-complete genomes of Blastopirellula marina DSM
3645(T) and Planctomyces maris DSM 8797(T). Comparative sequence analysis revealed the
following: A high number of genes coding for sulphatases in all genomes from marine
Planctomycetes. R. baltica harbours the highest number (109), while the freshwater
planctomycete Gemmata obscuriglobus and Candidatus Kueninia stuttgartiensis included
only 12 and 3 sulphatases, respectively. Additionally two fosmids (6FN and 5H12) contained
each one sulphatase. The high number of sulfatase genes in all genomes and their presence on
the fosmids underlined again the specialication of Planctomycetes in carbohydrate
degradation.
Genes involved in the C1-metabolism were found on all Planctomycete genomes, except of
Candidatus Kuenenia stuttgartiensis, and on two fosmids. Their role in the life-style of the
Planctomycetes is so far unknown. However expression profiling revealed that even in the
absence of C1 compounds seven of the respective genes are expressed, which indicates an
active role in the central metabolism of R. baltica.
Genes that are only present in planctomycete genomes and lack similarities with other
bacterial or archaeal genes were present in all five sequenced genomes and on all six fosmids.
It can be speculated that they might be involved in the unique cellular features or adaptations
of Planctomycetes.
In summary, the comparison of genomic information from cultivated and uncultivated
Planctomycetes offered new insights in their physiological capabilities with a focus on
planctomycete-specific features and striking metabolic pathways.
The corresponding paper has been published in the ISME Journal 1, 419-435 (2007).

Establishing of the Microarray Pipeline
96
5 ESTABLISHING THE MICROARRAY PIPELINE Comparing the results obtained and the initial outline of in my proposal, some changes
become obvious. During the initiation of this PhD thesis it was assumed that only small
methodological adaptations were needed to use whole genome arrays for expression profiling
in Rhodopirellula baltica. However, first tests showed unexpected challenges, like sample
processing or increased background. Optimisations also included the use of chemostat
reactors for the cultivation of R. baltica. This was triggered by two main requirements.
Firstly, the amount of cells needed for microarray experiments could not be obtained by
standard (batch) cultivation approaches. At least three biological independent replicates are
needed to obtain consistent regulation data. Second, synchronised cells are required to assure
reproducibility of the experiments.
After the microarray pipeline was finally running in-house, the reliability needed to be
optimised. Due to the unique character of the Planctomycetes there were no existing
transcriptome studies of other Planctomycetes available for validating the results. Therefore, I
performed expression profiling on the response of R. baltica to different stressors
(temperature and high salinity) to check if the results matched the expectations and to provide
new information on the environmental adaptation of R. baltica. Consequently research aim
2.2 was postponed. In addition, sharing cultures to conduct complementary proteome studies
with our collaboration partner at the Institute of Marine Biotechnology in Greifswald was
initiated. The integration of transcriptome and proteome data is expected to deliver a better
picture about the existence, annotation, regulation and expression of the genes in the
R. baltica.
Our aim was and is to combine our transcriptome results with all publicly available data of
regulated genes in R. baltica. Unfortunately, all results so far were not consistently stored in a
computer readable electronic format. Therefore a beta-version of an R. baltica database was
set up where information on the organism and genes can be stored. The software tool
JCOAST (Richter et al. 2008), which was already used in-house for comparative genomics on
the four planctomycetes genomes and their statistical analysis, was extended to host additional
information about each gene and soon it will be able to show expression data of all
transcriptome studies. All published results on each gene have been integrated into this
internal database. This also includes the latest results of Jerome Dabin from the marine station
in Roscoff (France). They are currently evaluating the catalytic potential of R. baltica with
respect to degradation of carbohydrates. This also includes the re-annotation of 128

Establishing of the Microarray Pipeline
97
polysaccharidases in the genome of R. baltica done by experts. Four out of these 128 proteins
were chosen for further functional and structural studies, identified by potential biotechnical
applications and originality. Recently, Dabin et al. (Dabin et al. 2008) published the results of
the crystallisation and preliminary results off x-ray analysis of a pectin lyase RB5312. The
expert re-annotation demonstrated that any genome annotation is only as good as the
knowledge of the annotator. The annotation sets the basis for all further knowledge
extrapolation. Therefore, part of the whole genome pipeline should include curation of the
annotation and a regular automatic update of the annotation against the current databases. At
least the awareness of partially incorrect annotations should be present while generating
hypothesis out of the significantly expressed genes.
In summary, more time than expected had to be invested in method optimisation, resulting in
a reduction of the time available to generate results on the ecology and biology of R. baltica.

Contribution to the Research on Planctomycetes
98
6 CONTRIBUTION TO THE RESEARCH ON PLANCTOMYCETES Planctomycetes are a group of budding, peptidoglycan-free bacteria, which attract more and
more attention within the fields of microbial ecology, cell biology and genomics. Studies of
cultured isolates and clone library sequences from natural communities contribute to this
development. The phylogeny of Planctomycetes has been under debate for several years now
(Stackebrandt et al. 1984; Woese 1987; Fuerst 1995; Brochier and Phillippe 2002; Teeling et
al. 2004). Based on genomic information they have been successfully allocated to the PVC
superphylum (Teeling et al. 2004; Fieseler et al. 2006; Wagner and Horn 2006). Their rather
uncommon features such as compartmentalised cell organisation (Fuerst 2005), the ability of
some species to grow chemolithoheterotroph via oxidation of ammonium (Jetten et al. 1997),
their possession of large genomes with partly unexpected genetic potential (Glöckner et al.
2003; Woebken et al. 2007), combined with their wide distribution in a variety of habitats,
has further reinforced a steadily increasing scientific and industrial interest.
Since the start of my thesis many more scientific studies on different Planctomycetes were
carried out highlighting even more how fascinating this phylum is. However none of these
have revealed details of underlying genetic mechanisms:
First of all more planctomycete isolates were published (Kulichevskaya et al. 2007;
Kulichevskaya et al. 2008). The latest diversity studies show that Planctomycetes can be
found in the intertidal sediments of the North Sea (Musat et al. 2006), in the north American
coast diatom bloom (Morris et al. 2006), in the anoxic laxer of Sphagnum peat bog (Ivanova
and Dedysh 2006) and in the alkaline gut of soil-feeding termites (Köhler et al. 2008). Shu et
al. (Shu and Jiao 2008) showed for the first time the systematic assessment of Planctomycetes
in latitudinal surface seawater of the open sea and comparative diversity patterns between
surface seawater and sediments. Latter Planctomycetes show a higher diversity. Looking at
the environmental factors, Shu proposes an influence of high organic carbon and oxygen on
the planctomycete diversity. Even a new metabolism for Planctomycete living in anaerobic
habitats has been proposed (Elshahed et al. 2007). Elshahed et al. (Elshahed et al. 2007)
assumes that the organisms either reduce sulphur via the annotated polysulfide reductase,
although the reducing of elementar sulphur has not been reported so far, or through sugar
fermentation. This is supported by the annotated genes for the lactate acid fermentation
pathway.

Contribution to the Research on Planctomycetes
99
Lately, new techniques were applied to get a better understanding of the planctomycete
morphology. Compartment organisation and chromatin structure of Gemmata obscuriglobus
were explored by cyro-electron tomography (Lieber et al. 2008). Lee et al. (Lee et al. 2009)
showed that the compartmentalised cell plan of members of the Planctomycetes can also be
found in the phylum Verrucomicrobia, belonging to the PVC superphylum. Interesting is also
a recent microscopic study where the morphological changes of the planctomycete Gemmata
obscuriglobus during the cell-cycle was investigated (Lee et al. 2009). This study might be a
good basis for further genetic studies to elucidate the responsible genes. The entire mentioned
studied have provided a wealth of new information about Planctomycetes.
However this work is the first to address the distinct features of Planctomycetes at the
genomic level. It is intended to bridge the gap between classical studies on the cell biology
and physiology of Planctomycetes and their active genetic repertoire.
The genomic information of uncultured Planctomycetes was addressed by the metagenomics
approach to get a clue of their environmental role. The fosmid libraries supported the
hypothesis that Planctomyces play an active role as a carbohydrate degrader (Woebken et al.
2007). Highlight of the comparison of the four complete closed planctomycete genomes were
the high amount of shared planctomycete-specific genes without known functions (Woebken
et al. 2007).
Within this work the first whole genome microarray for a cultured Planctomycete has been
established. This pipeline comprises reproducible and controlled cultivation of R. baltica´s
cultures, a protocol for expression profiling on the transcriptome level, data analysis and
evaluation up to data integration and comparison. Taken as a tool it helps to get hints on the
huge amount of genes in R. baltica currently lacking functional assignments, it elucidates the
function of annotated genes and suggests genes of interest for further functional analysis or
biotechnological applications.
In this work we gained results on certain genes with unknown function that probably involved
in the morphological changes or in the regulation of the cell-cycle and stress response of
R. baltica. For the first time we obtained information about the function of diverse
planctomycete-specific genes that are active under stress. We were able to highlight the
diverse functions of sulphatase genes in the genome of R. baltica. Results showed activity of
genes associated with the C1-metabolism. Finally, we gained knowledge about the behaviour
of R. baltica in its natural habitat. The possibility of transferring this knowledge to the
phylum Planctomycetes has a major impact for our understanding of the potential ecological
roles and adaptations of Planctomycetes in marine environments.

Outlook
100
7 OUTLOOK Expression profiling: microarray vs. sequencing technology
Recently, a manuscript by Shendure (Shendure 2008) with the controversial title “The
beginning of the end for microarrays” was published in Nature. The authors claimed that
microarrays will be replaced by the faster and cheaper sequencing technologies. The era of
“next generation sequencing technologies” has been heralded with the introduction of the 454
pyrosequencing system in 2005 (Margulies et al. 2005). Since then, four technology platforms
have entered the market (Roche 454, Illumina Genome Analyzer, ABI SOLiD and Helicos
HeliScope) and especially the Roche 454 GS FLX sequencing platform is now heavily used
for bacterial genome and small RNA sequencing, metagenomics, genomic structure variation
and human whole genome sequencing (Rothberg and Leamon 2008). Massively parallel 454
pyrosequencing has become a feasible method for increasing sequencing depth and coverage
while reducing time, labour, and cost (Margulies et al. 2005; Moore et al. 2006; Wicker et al.
2006; Huse et al. 2007). Typical output from a 4.5 h run of the original Roche 454 GS 20
sequencer are 20–30 Mb, comprising roughly 300,000 sequence reads averaging 100 bp. With
state of the art machines read length of 250 bp (FLX) and 400 bp (FLX Titanium) can be
routinely reached, increasing the amount of basepairs per run drastically.
Lately, also transcriptomes were analysed by tag sequencing approaches (Cristobal-Vera et al.
2008; Marioni et al. 2008) the former exclusive field of DNA microarrays. In contrast to
microarrays, new sequencing technologies do not rely on the availability of a sequenced
genome and also do not suffer from severe methodological limitations as mentioned already
in this thesis. In general, for tag sequencing a population of RNA (total or only mRNA) is
converted to a library of cDNA fragments with adaptors attached to one or both ends. Each
molecule, with or without amplification, is then sequenced in a high-throughput manner to
obtain short sequences from one end (single-end sequencing) or both ends (pair-end
sequencing). The reads are typically 30–400 bp, depending on the sequencing technology
used (Wang et al. 2009). A major advantage is that this approach is quantitative, resolving
one of the major limitations of the current microarray based technologies.
Sequencing costs are continuously decreasing and the technique has already become a fast
and cost-effective option for functional genomics, especially in case of mixed microbial
populations (meta-transcriptomics). This development narrows the gap between approaches
based on model organisms offering a genetic model and so far barely investigated organisms,
which are most tractable for ecological and evolutionary studies (Cristobal-Vera et al. 2008).

Outlook
101
However, there will still be important applications for microarrays in the future as pointed out
by Shendure et al. (Shendure 2008). Most likely not for large-scale expression profiling due
to the effectiveness of the new sequencing technologies but for other applications such as
custom genotyping and detection of copy number variation. This is because microarrays allow
us to address such questions in a more targeted way. Also in case of expression profiling there
will be niches for microarrays in the future. In particular to answer specific questions about
specific groups of genes of interest (“partial” genome microarrays), or as a screening tool to
get first information on the gene regulation in model organisms (whole genome array).
Another unique feature might be RNA-targeted microarrays. No cDNA synthesis is required
like for sequencing technology and a huge bias can be avoided (Würdemann et al. 2006).
Interesting is also the comparison of the expanses between one Solexa and one established
R. baltica cDNA-microarray run. A Solexa run with a sequence yield of 30-70 bp costs about
3000 € with up to six different experiments per run (500 € per experiment) plus costs for the
instrument and RNA-isolation. An established R. baltica cDNA microarray run costs around
125 Euro per experiment (per slide) plus 160 Euros per slide for the instruments. Calculations
do not include cost of labour and of computer hardware for data analysis and storage. An
established cDNA microarray is still cheaper than a Solexa run at the moment. However,
sequencing technology is already a serious competitor, concerning the cost, for new
expression profiling projects, meaning which are not established yet.
Finally, there is one major limitation correlated with the new sequencing technologies, which
now produce a mass of raw sequencing data in shortest time. Appropriate bioinformatic
infrastructure is required to handle this flood of data. At the moment, only very few working
groups in the world are really prepared for this task. A complete rethinking is required in this
regard. Despite the complexity of establishing a platform for microarray experiments, this
might be a reason for researchers to stick to microarray technology within the next years.

Reference
102
8 REFERENCE Alizadeh, A. A. and L. M. Staudt (2000). "Genomic-scale gene expression profiling of
normal and malignant immune cells." Curr Opinion Immun 12(2): 219-225. Amann, R. and W. Ludwig (2000). "Ribosomal RNA-targeted nucleic acid probes for
studies in microbial ecology." FEMS Microbiol Reviews 24(5): 555-565. Amersham (2002). Microarray Handbook, Amersham Biosciences. Barbeyron, T., G. Michel, P. Potin, B. Henrissat and B. Kloareg (2000). "Iota-
carrageenases constitute a novel family of glycoside hydrolase, unrelated to that of kappa-carragenenases." J Bacteriol 275(10): 35499-35505.
Bauer, M., T. Lombardot, H. Teeling, N. Ward, R. Amann and F. O. Glöckner (2004). "Archaea-like genes for C1-transfer enzymes in Planctomycetes: Phylogenetic implications of their unexpected presence in this phylum." J Mol Evol 59: 571-586.
Bauld, J. and J. Staley (1976). "Planctomyces maris sp. nov.: A marine isolate of the Planctomycetes-Blastocaulis group of budding bacteria." J. Gen. Microbiol 97(45-55).
Beier, V., M. Baum and J. Hoheisel (2004). "Flourescent sample labeling for DNA microarray analyses." Methods Mol Biol 4(2): 57-64.
Berteau, O., A. Guillot, A. Benjdia and S. Rabot (2006). "A new type of bacterial sulfatase reveals a novel maturation pathway in prokaryotes." J. Biol. Chem. 281(32): 22464-22470.
Brazma, A., P. Hingamp, J. Quackenbush, G. Sherlock, P. Spellman, C. Stoeckert, J. Aach, W. Ansorge, C. A. Ball, H. C. Causton, T. Gaasterland, P. Glenisson, F. C. P. Holstege, I. F. Kim, V. Markowitz, J. C. Matese, H. Parkinson, A. Robinson, U. Sarkans, S. Schulze-Kremer, J. Stewart, R. Taylor, J. Vilo and M. Vingron (2001). "Minimum information about a microarray experiment (MIAME) - toward standards for microarray data." Nat Genet 29(4): 365-371.
Brazma, A. and J. Vilo (2000). "Gene expression data analysis." FEBS J Funct Genom 480(1): 17-24.
Brochier, C. and H. Phillippe (2002). "Phylogeny: a non-hyperthermophilic ancestor for bacteria." Nature 417(244).
Chatzinotas, A., R. Sandaa, S. W, R. Amann, D. FL, V. Torsvik, J. Zeyer and D. Hahn (1998). "Analysis of broad-scale differences in microbial community composition of two pristine forest soils." System. Appl. Microbiol. 21(4): 579-87.
Churchill, G. A. (2002). "Fundamentals of experimental design for cDNA microarrays." Nature Genetics 32: 490-495.
Cristobal-Vera, J., C. W. Wheat, H. W. Fescemyer, M. J. Frilander, D. L. Crawford, I. Hanski and J. H. Marden (2008). "Rapid transcriptome characterization for a nonmodel organism using 454 pyrosequencing." 17(7): 1636-1647.
Crump, B. C., E. V. Armbrust and J. A. Baross (1999). "Phylogenetic analysis of particle-attached and free-living bacterial communities in the Columbia River, its estuary, and the adjacent coastal ocean." Appl. Environ. Microbiol. 65(7): 3192-3204.
Dabin, J., M. Jam, M. Czjzek and G. Michel (2008). "Expression, purification, crystallization and preliminary X-ray analysis of the polysaccharide lyase RB5312 from the marine planctomycete Rhodopirellula baltica." Acta Crystallogr Sect F Struct Biol Cryst Commun. 1(64): 224-7.
DeLong, E. and D. Franks (1993). "Phylogenetic diversity of aggregate-attached vs. free-living marine bacterial assemblages." Limnol Oceanogr 38: 924-934.
DeLong, E. F. and D. M. Karl (2005). "Genomic perspectives in microbial oceanography." 437(7057): 336-342.

Reference
103
Descamps, V., S. Colin, M. Lahaye, M. Jam, C. Richard, P. Potin, T. Barbeyron, J. C. Yvin and B. Kloareg (2005). "Isolation and culture of a marine bacterium degrading the sulfates fucans from marine brown algae." Marine Biotechnology 8: 27-39.
Duggan, D. J., M. Bittner, Y. Chen, P. Meltzer and J. M. Trent (1999). "Expression profiling using cDNA microarrays." Nature genetics supplements 21: 10-14.
Dunham, I. (2000). "Genomics - the new rock and roll?" Trends Genet 16: 456-461. Dupont, S., K. Wilson, M. Obst, H. Sköld, H. Nakano and M. C. Thorndyke (2007).
"Marine ecological genomics: when genomics meets marine ecology." Mar Ecol Prog Ser 332: 257-273.
Eisen, M. B., P. T. Spellman, P. O. Brown and D. Botstein (1998). "Cluster analysis and display of genome-wide expression patterns." PNAS 95(25): 14863-14868.
Elshahed, M. S., N. H. Youssef, Q. Luo, F. Z. Najar, B. A. Roe, T. M. Sisk, S. I. Buhring, K.-U. Hinrichs and L. R. Krumholz (2007). "Phylogenetic and Metabolic Diversity of Planctomycetes from Anaerobic, Sulfide- and Sulfur-Rich Zodletone Spring, Oklahoma." 73: 4707-4716.
Fieseler, L., A. Quaiser, C. Schleper and U. Hentschel (2006). "Analysis of the first genome fragment from the marine sponge-associated, novel candidate phylum Poribacteria by environmental genomics." Environ Microbiol. 8(4): 612-624.
Foster, T. and P. Ghazal (2003). "Experiments using microarray technology: limitations and standard operatin procedures." J Endocrinol 178: 195-204.
Franzmann, P. D. and B. Skerman (1984). "Gemmata obscuriglobus, a new genus and species of the budding bacteria." Antonie van Leeuwenhoek 50: 261-268.
Fuerst, A. J. (2005). "Intracellular compartmentation in Planctomycetes." Annu. Rev. Microbiol. 59: 299-328.
Fuerst, J. A. (1995). "The Planctomycetes: emerging models for microbial ecology, evolution and cell biology." Microbiology 141 (Pt 7): 1493-506.
Fuerst, J. A., H. G. Gwilliam, M. Lindsay, A. Lichanska, C. Belcher, J. E. Vickers and P. Hugenholtz (1997). "Isolation and molecular identification of planctomycete bacteria from postlarvae of the giant tiger prawn, Penaeus monodon." Appl Environ Microbiol 63(1): 254-62.
Fuerst, J. A., S. K. Sambhi, J. L. Paynter, J. A. Hawkins and J. G. Atherton (1991). "Isolation of a bacterium resembling Pirellula species from primary tissue culture of the giant tiger prawn (Penaeus monodon)." Appl Environ Microbiol 57(11): 3127-34.
Fuerst, J. A., R. I. Webb, M. J. Garson, L. Hardy and H. M. Reiswig (1998). "Membrane-bounded nucleoids in microbial symbionts of marine sponges." FEMS Microbiol Lett 166: 29-34.
Fuerst, J. A., R. I. Webb, M. J. Garson, L. Hardy and H. M. Reiswig (1999). "Membrane-bounded nuclear bodies in a diverse range of symbionts of Great Barrier Reef sponges." Mem Queensl Mus 44: 193-203.
Gade, D., J. Gobom and R. Rabus (2005). "Proteomic analysis of carbohydrate catabolism and regulation in the marine bacterium Rhodopirellula baltica." Proteomics 5(14): 3672-3683.
Gade, D., T. Stührmann, R. Reinhardt and R. Rabus (2005). "Growth phase dependent regulation of protein composition in Rhodopirellula baltica." Environ Microbiol 7(8): 1074-1084.
Gade, D., D. Theiss, D. Lange, E. Mirgorodskaya, T. Lombardot, F. O. Glöckner, M. Kube, R. Reinhardt, R. Amann, H. Lehrach, R. Rabus and J. Gobom (2005). "Towards the proteome of the marine bacterium Rhodopirellula baltica: Mapping the soluble proteins." Proteomics 5(14): 3654-3671.

Reference
104
Gade, D., J. Thiermann, D. Markowsky and R. Rabus (2003). "Evaluation of two-dimensional difference gel electrophoresis for protein profiling. Soluble proteins of the marine bacterium Pirellula sp. strain 1." J Mol Microbiol Biotechnol 5(4): 240-251.
Galperin, M. Y. and E. V. Koonin (2004). "'Conserved hypothetical' proteins: prioritization of targets for experiemental study." Nucl. Acids Res. 32(18): 5452-5463.
Giovannoni, S. J., E. Scabtach and R. W. Castenholz (1987). "Isosphaera pallida, gen. and comb. nov., a gliding, budding eubacterium from hot springs." Arch Microbiol 147: 276-284.
Glöckner, F. O., M. Kube, M. Bauer, H. Teeling, T. Lombardot, W. Ludwig, D. Gade, A. Beck, K. Borzym, K. Heitmann, R. Rabus, H. Schlesner, R. Amann and R. Reinhardt (2003). "Complete genome sequence of the marine planctomycete Pirellula sp. strain 1." Proc Natl Acad Sci U S A 100(14): 8298-303.
Gollub, J. and G. Sherlock (2006). Clustering Microarray Data Methods in Enzymology. DNA Microarrays, Part B: Databases and Statistics. A. Kimmel and B. Oliver, Academic Press: 194-213.
Gupta, V. and A. Cherkassky (2003). "Directly labeled mRNA produces highly precise and unbiased differential gene expression data." Nucl. Acids Res. 31(2).
Hu, P., E. L. Brodie, Y. Suzuki, H. H. McAdams and G. L. Andersen (2005). "Whole-genome transcriptional analysis of heavy metal stresses in Caulobacter crescentus." J Bacteriol 187(24): 8437-49.
Huse, S., J. Huber, H. Morrison, M. Sogin and D. Welch (2007). "Accuracy and quality of massively parallel DNA pyrosequencing." Genome Biol 8(7): R143.
Imbeaud, S. and C. Auffray (2005). "`The 39 steps' in gene expression profiling: critical issues and proposed best practices for microarray experiments." Drug Discovery Today 10(17): 1175-1182.
Inagaki, F., T. Nunoura, S. Nakagawa, A. Teske, M. Lever, A. Lauer, M. Suzuki, K. Takai, M. Delwiche, F. Colwell, K. H. Nealson, K. Horikoshi and B. B. Jorgensen (2006). "Biogeographical distribution and diversity of microbes in methane hydrate-bearing deep marine sediments on the Pacific Ocean Margin." Proc Natl Acad Sci U S A 103: 2815-2820.
Ivanova, A. and S. Dedysh (2006). "High abundance of planctomycetes in anoxic lavers of a Sphagnum peat bog." Mikrobiologiia 75(6): 823-7.
Jacobs-Wagner, C. (2004). "Regulatory proteins with a sense of direction: cell-cycle signalling network in Caulobacter." Mol. Microbiol 51(1): 7-13.
Jenal, U. and C. Stephens (2002). "The Caulobacter cell-cycle: timing, spatial organization and checkpoints." Curr Opin Microbiol 5(6): 558-563.
Jetten, M. S., S. J. Horn and M. C. M. Van Loosdrecht (1997). "Towards a more sustainable wastewater treatment system." Water Sci Technol 35: 171-180.
Kerr, M. K., M. Martin and G. A. Churchill (2000). "Analysis of variance for gene expression microarray data." Journal Computer Biology 7: 819-837.
Kirkpatrick, J., B. Oakley, C. Fuchsman, S. Srinivasan, J. T. Staley and J. W. Murray (2006). "Diversity and distribution of Planctomycetes and related bacteria in the suboxic zone of the black sea." Appl. Environ. Microbiol. 72(4): 3079-3083.
Knudsen, S. (2004). Guide to analysis of DNA microarray data, John Wiley & Sons. Köhler, T., U. Stingl, K. Meuser and A. Brune (2008). "Novel lineages of Planctomycetes
densely colonize the alkaline gut of soil-feeding termites (Cubitermes spp.)." 10(5): 1260-1270.
Kohonen, T. (1992). "Self-organized formation of topologically correct feature maps." Biol. Cybernetics 43: 59-69.

Reference
105
König, E., H. Schlesner and P. Hirsch (1984). "Cell wall studies on budding bacteria of the Planctomyces/Pasteuria group and on a Prosthecomicrobium sp." Arch Microbiol 138: 200-205.
Kulichevskaya, I. S., A. O. Ivanova, O. I. Baulina, P. L. E. Bodelier, J. S. S. Damste and S. N. Dedysh (2008). "Singulisphaera acidiphila gen. nov., sp. nov., a non-filamentous, Isosphaera-like Planctomycete from acidic northern wetlands." Int J Syst Evol Microbiol. 58(5): 1186-1193.
Kulichevskaya, I. S., A. O. Ivanova, S. E. Belova, O. I. Baulina, P. L. E. Bodelier, W. I. C. Rijpstra, J. S. Sinninghe Damste, G. A. Zavarzin and S. N. Dedysh (2007). "Schlesneria paludicola gen. nov., sp. nov., the first acidophilic member of the order Planctomycetales, from Sphagnum-dominated boreal wetlands." Int J Syst Evol Microbiol 57(11): 2680-2687.
Lander, E. S. (1999). "Array of hope." Nature genetics 21: 3-4. Laub, M. T., H. H. McAdams, T. Feldblyum, C. M. Fraser and L. Shapiro (2000).
"Global analysis of the genetic network controlling a bacterial cell-cycle." Science 290(5499): 2144-2148.
Lee, K.-C., R. I. Webb and A. J. Fuerst (2009). "The cell-cycle of the planctomycete Gemmata obscuriglobus with respect to cell compartmentalization." BMC Cell Biology 10(4).
Lee, K.-C., R. I. Webb, P. H. Janssen, P. Sangwan, T. Romeo, J. T. Staley and A. J. Fuerst (2009). "Phylum Verrucomicrobia representatives share a compartmentalized cell plan with members of bacterial phylum Planctomycetes." BMC Microbiology 9(5).
Leung, Y. F. (2002). "Unravelling the mystery of microarray data analysis." Trends in Biotechnology 20(9): 366-368.
Leung, Y. F. and D. Cavalieri (2003). "Fundamentals of cDNA microarray data analysis." Trends in Genetics 19(11): 649-659.
Li, X., W. Gu, S. Mohan and D. Baylink (2002). "DNA microarrays: their use and misuse." Microcirculation 9(1): 13-22.
Lieber, A., A. Leis, A. Kushmaro, A. Minsky and O. Medalia (2008). "Chromatin organization and radio-resistance in the bacterium Gemmata obscuriglobus." J Bacteriol. 191(5): 1439-45.
Liesack, W., H. König, H. Schlesner and P. Hirsch (1986). "Chemical composition of the peptidoglycan-free cell envelopes of budding bacteria of the Pirella/Planctomyces group." Arch Microbiol 145: 361-366.
Lindsay, M. R., R. I. Webb and J. Fuerst (1997). "Pirellulosomes: a new type pf membranebounded cell compartment in planctomycete bacteria of the genus Pirellula." Microbiology 143: 739-748.
Lindsay, M. R., R. I. Webb, M. Strous, M. S. Jetten, M. K. Butler, R. J. Forde and J. A. Fuerst (2001). "Cell compartmentalisation in planctomycetes: novel types of structural organisation for the bacterial cell." Arch Microbiol 175(6): 413-29.
Llobet-Brossa, E., R. Rossello-Mora and R. Amann (1998). "Microbial community composition of Wadden Sea sediments as revealed by Fluorescence In Situ Hybridization." Appl. Environ. Microbiol. 64(7): 2691-2696.
Lombardot, T. (2004). The genome of free living, marine planctomycete Pirellula sp. strain 1. Faculty of Biology/ Chemistry. Bremen, IUniversität Bremen: 154.
Margulies, M., M. Egholm, W. E. Altman, S. Attiya, J. S. Bader, L. A. Bemben, J. Berka, M. S. Braverman, Y. J. Chen, Z. Chen, S. B. Dewell, L. Du, J. M. Fierro, X. V. Gomes, B. C. Godwin, W. He, S. Helgesen, C. H. Ho, G. P. Irzyk, S. C. Jando, M. L. Alenquer, T. P. Jarvie, K. B. Jirage, J. B. Kim, J. R. Knight, J. R.

Reference
106
Lanza, J. H. Leamon, S. M. Lefkowitz, M. Lei, J. Li, K. L. Lohman, H. Lu, V. B. Makhijani, K. E. McDade, M. P. McKenna, E. W. Myers, E. Nickerson, J. R. Nobile, R. Plant, B. P. Puc, M. T. Ronan, G. T. Roth, G. J. Sarkis, J. F. Simons, J. W. Simpson, M. Srinivasan, K. R. Tartaro, A. Tomasz, K. A. Vogt, G. A. Volkmer, S. H. Wang, Y. Wang, M. P. Weiner, P. Yu, R. F. Begley and J. M. Rothberg (2005). "Genome sequencing in microfabricated high-density picolitre reactors." Nature genetics 437(7057): 376 - 380.
Marioni, J. C., C. E. Mason and S. M. Mane (2008). "RNA-seq: An assessment of technical reproducibility and comparison with gene expression arrays." Genome Research 18: 1509-1517.
Moore, M., A. Dhingra, P. Soltis, R. Shaw, W. Farmerie, K. Folta and D. Soltis (2006). Rapid and accurate pyrosequencing of angiosperm plastid genomes. 6: 17.
Morris, R. M., K. Longnecker and S. J. Giovannoni (2006). "Pirellula and OM43 are among the dominant lineages identified in an Oregon coast diatom bloom." Environ Microbiol. 8(8): 1361-1370.
Musat, N., U. Werner, K. Knittel, S. Kolb, T. Dodenhof, J. E. E. van Beusekom, D. de Beer, N. Dubilier and R. Amann (2006). "Microbial community structure of sandy intertidal sediments in the North Sea, Sylt-Romo Basin, Wadden Sea." Syst. Appl. Microbiol. 29(4): 333-348.
Neef, A., R. Amann, H. Schlesner and K. Schleifer (1998). "Monitoring a widespread bacterial group: in situ detection of Planctomycetes with 16S rRNA-targeted probes." Microbiology 144(12): 3257-3266.
Pan, W. (2002). "A comparative review of statistical methods for discovering differentially expressed genes in replicated microarray experiments." Bioinformatics 18(4): 546-554.
Parenti, G. (1997). "The sulfatase gene family." Curr Opin Genet Dev 7(3): 386-91. Parro, V., M. Moreno-Paz and E. Gonzalez-Toril (2007). "Analysis of environmental
transcriptomes by DNA microarrays." Environ Microbiol. 9(2): 453-464. Pedros-Alio, C. (2006). "Genomics and marine microbial ecology." Intern Microbiol 9: 191-
197. Phimister, B. (1999). "A note on nomenclature." Nature Genetics 21: 1. Pimentel-Elardo, S., M. Wehrl, A. B. Friederich, P. R. Jensen and U. Henschel (2003).
"Isolation of Planctomycetes from aplysina sponges." Aqua Micro Ecol 33: 239-245. Quackenbush, J. (2002). "Microarray data normalization and transformation." Nature
Genetics 32: 495-501. Rabus, R., D. Gade, R. Helbig, M. Bauer, F. O. Glöckner, M. Kube, H. Schlesner, R.
Reinhardt and R. Amann (2002). "Analysis of N-acetylglucosamine metabolism in the marine bacterium Pirellula sp. strain 1 by a proteomic approach." Proteomics 2(6): 649-55.
Rath (1998). "High phylogenetic diversity in a marine-snow-associated bacterial assemblage." Aqua Micro Ecol 14(3): 261-269.
Raychaudhuri, S. (2000). "Pattern recognition of genomic features with microarrays: site typing of Mycobacterium tuberculosis strains." Proc Int Conf Intell Syst Mol Biol 8: 286-295.
Richter, M., T. Lombardot, I. Kostadinov, R. Kottmann, M. Duhaime, J. Peplies and F. Glöckner (2008). "JCoast - A biologist-centric software tool for data mining and comparison of prokaryotic (meta)genomes." BMC Bioinformatics 9(1): 177.
Riemann, L. and F. Azam (2002). "Widespread N-acetyl-D-glucosamine uptake among pelagic marine bacteria and its ecological implications." Appl. Environ. Microbiol. 68(11): 5554-5562.

Reference
107
Rothberg, J. M. and J. H. Leamon (2008). "The development and impact of 454 sequencing." Nat Biotech 26(10): 1117-1124.
Rullkötter (1999). Marine Geochemistry. H. D. Schulz and M. Zabel. Berlin, Springer-Verlag: 129-172.
Schlesner, H. (1994). "The development of media suitable for the microorganisms morphologically resembling Planctomyces spp., Pirellula spp., and other Planctomycetales from various aquatic habitats using dilute media." System. Appl. Microbiol. 17: 135-145.
Schlesner, H., C. Rensmann, B. J. Tindall, D. Gade, R. Rabus, S. Pfeiffer and P. Hirsch (2004). "Taxonomic heterogeneity within the Planctomycetales as derived by DNA-DNA hybridization, description of Rhodopirellula baltica gen. nov., sp. nov., transfer of Pirellula marina to the genus Blastopirellula gen. nov. as Blastopirellula marina comb. nov. and emended description of the genus Pirellula." Int J Syst Evol Microbiol 54(Pt 5): 1567-80.
Shendure, J. (2008). "The beginning of the end for microarrays?" Nature Methods 5(7): 585-587.
Shu, Q. and N. Jiao (2008). "Diffferent planctomycetes diversity patterns in latitudinal surface seawater of the open sea and in sediment." J Microbiol 46(2): 154-159.
Soukas, A., P. Cohen, N. D. Socci and J. M. Friedman (2000). "Leptin-specific patterns of gene expression in white adipose tissue." Genes Dev. 14(8): 963-980.
Spellman, P., G. Sherlock, Y.-L. Zhang, R. Vishwanath, K. Anders, M. B. Eisen, P. O. Brown, D. Botstein and B. Futcher (1998). "Comprehensive Identification of cell-cycle-regulated genes of the yeast Saccharomyces cerevisiae by microarray hybridization." MBC 9: 3273-3297.
Stackebrandt, E., W. Ludwig, W. Schubert, F. Klink, H. Schlesner, T. Roggentin and P. Hirsch (1984). "Molecular genetic evidence for early evolutionary origin of budding peptidoglycan-less eubacteria." Nature 307: 735-737.
Steglich, C., M. Futschik, M. Rector, R. Steen and S. Chrisholm (2006). "Genome-wide analysis of light sensing in Prochlorococcus." J Bacteriol 188(22): 7796-7806.
Stephens, C. (2001). "Bacterial cell-cycle: Seeing the big picture with microarrays." Current Biology 11(6): R222-R225.
Strous, M., E. Pelletier, S. Mangenot, T. Rattei, A. Lehner, M. Taylor, M. Horn, H. Daims, D. Bartol-Mavel, P. Wincker, V. Barbe, N. Fonknechten, D. Vallenet, C. Schenowitz-Truong, C. Medigue, A. Collingro, B. Snel, B. Dutilh, H. Op den Camp, C. van der Drift, I. Cirpus, K. van de Pas-Schoonen, H. Harhangi, L. van Niftrik, M. Schmid, J. Keltjens, J. van de Vossenberg, B. Kartal, H. Meier, D. Frishman, M. Huynen, H. Mewes, J. Weissenbach, M. Jetten, M. Wagner, . and D. Le Paslier (2006). "Deciphering the evolution and metabolism of an anammox bacterium from a community genome." Nature 440 790-794.
Teeling, H. (2004). Aspekte der bioinformatischen Analyse und Annotation des Genoms von Rhodopirellula baltica. Faculty Biology/Chemistry. Bremen, Bremen: 129.
Teeling, H., T. Lombardot, M. Bauer, W. Ludwig and F. O. Glockner (2004). "Evaluation of the phylogenetic position of the planctomycete Rhodopirellula baltica SH 1T by means of concatenated ribosomal protein sequences, DNA-directed RNA polymerase subunit sequences and whole genome trees." Int J Syst Evol Microbiol 54(Pt 3): 791-801.
Tekniepe, B. L., J. M. Schmidt and M. P. Starr (1981). "Life-cycle of a budding and appendaged bacterium belonging to morphotype IV of the Blastocaulis-Planctomyces group." Curr Microbiol 5: 1-6.

Reference
108
Thomas, T., S. Egan, D. Burg, C. Ng, L. Ting and R. Caviccioli (2007). "Integration of genomics and proteomics into marine microbial ecology." Mar Ecol Prog Ser 332: 291-299.
Vergin, K. L., E. Urbach, J. L. Stein, E. F. DeLong, B. D. Lanoil and S. J. Giovannoni (1998). "Screening of a fosmid library of marine environmental genomic DNA fragments reveals four clones related to members of the order Planctomycetales." Appl Environ Microbiol 64: 3075-3078.
Wagner, M. and M. Horn (2006). "The Planctomycetes, Verrucomicrobia, Chlamydiae and sister phyla compris a superphylum with biotechnological and medical relevance." Curr Opin Biotechn 17(3): 241-249.
Wallner, S., M. Bauer, C. Würdemann, P. Wecker, F. O. Glöckner and K. Faber (2004). "Highly enantioselective sec-alkyl sulfatase activity of the marine planctomycete Rhodopirellula baltica shows retention of configuration." Angew Chem Intern Edition English 44(39): 6381-4.
Wang, J., C. Jenkins, R. I. Webb and J. A. Fuerst (2002). "Isolation of Gemmata-like and Isosphaera-like planctomycete bacteria from soil and freshwater." Appl Environ Microbiol 68(1): 417-22.
Wang, Z., M. Gerstein and M. Snyder (2009). "RNA-Seq: a revolutionary tool for transcriptomics." Nat Rev Genet 10(1): 57-63.
Wecker, P., C. Klockow, C. Quast, P. Langhammer, J. Harder and F. Glöckner (2009). "Transcriptional response of the model planctomycete Rhodopirellula baltica SH1T to changing environmental conditions." submitted.
Wecker, P., M. Schueler, A. Ellrott and F. O. Glöckner (2009). "The Rhodopirellula baltica life-cycle: Growth-dependent expression profiling of the complete genome with microarrays." in prep.
Wicker, T., E. Schlagenhauf, A. Graner, T. Close, B. Keller and N. Stein (2006). 454 sequencing put to the test using the complex genome of barley. 7: 275.
Woebken, D., H. Teeling, P. Wecker, A. Dumitriu, I. Kostadinov, E. F. DeLong, R. Amann and F. O. Glöckner (2007). "Fosmids of novel marine Planctomycetes from the Namibian and Oregon coast upwelling systems and their cross-comparison with planctomycete genomes." ISME Journal 1(5): 419-435.
Woese, C. R. (1987). "Bacterial evolution." Mircobiol. Rev. 51: 221-271. Würdemann, C. (2006). Method development for expression profiling in prokaryotes and the
application in marine environments. School of Engineering and Science. Bremen, International University Bremen: 159.
Würdemann, C., J. Peplies, S. Schübbe, A. Ellrott, D. Schüler and F. O. Glöckner (2006). "Evaluation of gene expression analysis using RNA-targeted partial genome arrays." System Appl Microbiol 29(5): 349-357.
Zhang, Y.-L., B. D. Price, S. Tetradis, G. Chakrabarti, G. Maulik and G. M. Makrigiorigos (2001). "Reproducible and inexprensive probe preparation for oligonucleotide arrays." Nucl. Acids Res. 29(13): E66-6.
Zhou, J. (2003). "Microarrays for bacterial detection and microbial community analysis." Curr Biol 6(3): 288-294.

Acknowledgement
109
9 ACKNOWLEDGEMENT ……………………….because you never make it on your own……………………….
I wish to express my sincere gratitude to my supervisor Prof. Dr. Frank Oliver Glöckner, who
guided this work and gave me support and scientific freedom. He was always there for me
when I needed advice.
I wish to thank Prof. Dr. Matthias Ullrich and Prof. Dr. Georgi Muskhelishvili for being
members of my thesis and dissertation committee. I would also like to thank Prof. Dr. Rudolf
Amann for taking part in the evaluation of my work and for his constant support in the
background.
I am grateful to Dr. Christine Klockow and Dr. Marga Bauer for their help, advice and
guidance during my PhD.
I am indebted to Andreas Ellrott, Chris Würdemann and Jörg Peplies for providing
inspiration, friendship and wise words.
I would like to thank all members of the Microbial Genomics Group who helped me during
my PhD work and made science enjoyable. Special thanks to the Michael Richter, Christian
Quast, Jost Waldmann and Elmar Prüsse.
I would like to thank all former and present members of the Department of Molecular
Ecology for their support, discussions, experience, their comradeship and the refreshing
evenings outside of the institute, especially Julia Arnds, Katharina Kohls, Anke Meyerdierks,
Jill Peterson, Caroline Rühland, Martha Schattenhofer, Regina Schauer, Silke Wetzel,
Dagmar Wöbken, Jörg Wulf, and my “Findorff-WG-colleagues” Elke Allers and Claudia
Bergin for just being there.
Special thanks to Jörg Wulf, Silke Wetzel and Nicole Rödiger for their excellent support and
patience in the laboratory.

Acknowledgement
110
Pour cette fructueuse collaboration et les bons moments partagés, je voudrai remercier Gurvan
Michel et Tristan Barbeyron de la Station Biologique de Roscoff.
The Network Marine Genomics Europe with all these fascinating members and meetings that
broaden my horizon deserves special mentioning. I would especially like to thank Mike
Thorndyke, Ulrika Hjelm and Michèle Barbier for supporting my skills. Thank you! Tack så
mycket! Merci beaucoup!
This work would not have been possible without the financial support of the Max-Planck-
Society, the Marine Genomics Europe Network and the Jacobs University Bremen. This work
was done between June 2005 and March 2009 at the Microbial Genomics Group of the Max
Planck Institute for Marine Microbiology in Bremen.
Zum Schluß möchte ich mich bei meinen Brüder Olaf und Axel, meinem Neffen Johannes
und meinen Eltern Elke und Josef für Ihre Unterstützung bedanken. Worte sind Blätter und
Taten sind Früchte.
Ludovic, merci de m’avoir aider à aller de l’avant et de n’avoir jamais arrêter de croire
fermement en moi. Hey Dude, ca fart!
This work is dedicated to all the wonderful, supportive and helpful people I met in the last six
years!

Annex
111
10 ANNEX

Transcriptional response of the model planctomycete Rhodopirellula baltica SH1T to changing
environmental conditions
ADDITIONAL FILE 1 – List of differentially expressed genes for all three stress experiments ADDITIONAL FILE 2 – List of selected clusters of the k-means clustering

Heat Induced10min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand Intensity Ratio Ratio Stdev %
4296 RB7619 278 837 conserved hypothetical protein 4097010 4096174 5.6 30607 - 5.93 1.68 0.11 6.426201 RB11150 385 1158 NADH:flavin oxidoreductase/NADH oxidase 6046862 6048019 4.5 42128 + 6.50 1.51 0.13 8.656831 RB12276 110 333 conserved hypothetical protein 6605112 6605444 4.7 11911 + 5.89 3.04 0.21 7.074149 RB7375 412 1239 Peptidase T pepT 3.4.11.4 3948831 3947593 4.4 44404 - 5.80 1.56 0.21 13.786638 RB11919 671 2016 Dihydroxy-acid and 6-phosphogluconate dehydratase 6417713 6415698 5.3 70685 - 5.33 2.10 0.31 14.885972 RB10733 389 1170 Glutathione-dependent formaldehyde dehydrogenase fdh 1.2.1.1 5832679 5833848 4.8 42303 + 6.24 2.55 0.26 10.292891 RB5134 208 627 Ham1-like protein 2639251 2639877 5.7 22783 + 5.57 1.85 0.09 5.105648 RB10127 468 1407 secreted glucose dehydrogenase 5466726 5465320 4.8 51430 - 5.86 2.19 0.06 2.786604 RB11853 229 690 GTP cyclohydrolase I folE 3.5.4.16 6382501 6383190 6.2 25392 + 5.99 2.04 0.13 6.396263 RB11253 226 681 CDP-diacylglycerol--glycerol-3-phosphate 3-phosphatidyltransferase 6105171 6104491 6.2 24053 - 5.52 1.51 0.05 3.045194 RB9305 360 1083 peptide chain release factor 1 prfA 4974620 4975702 4.9 40794 + 5.92 1.82 0.04 2.035195 RB9306 296 891 HemK protein hemK 2.1.1.- 4975706 4976596 4.7 32925 + 5.90 1.51 0.14 9.146837 RB12285 224 675 phospholipid N-methyltransferase 2.1.1.17 6609391 6610065 6.8 24262 + 6.10 2.07 0.16 7.705606 RB10049 217 654 RNA polymerase ECF-type sigma factor 5419507 5420160 10.1 25205 + 5.93 3.14 0.32 10.246090 RB10940 183 552 acetyltransferase 5934856 5935407 6.1 20469 + 5.96 2.16 0.24 11.28
98 RB170 96 291 Transposase IS3/IS911 89333 89623 10.1 11124 + 5.44 2.16 0.08 3.51382 RB704 96 291 Transposase IS3/IS911 378137 378427 10.1 11124 + 5.62 1.75 0.07 4.12628 RB1189 96 291 Transposase IS3/IS911 606838 607128 10.1 11124 + 5.45 1.78 0.27 15.14842 RB1571 564 1695 DNA ligase:ATP-dependent DNA ligase lig 6.5.1.1 803159 804853 5.4 63326 + 5.63 1.97 0.13 6.82
2381 RB4299 96 291 Transposase IS3/IS911 2214533 2214823 9.9 11182 + 5.65 2.37 0.32 13.672463 RB4433 162 489 Ferritin and Dps 2277208 2277696 4.3 17757 + 6.65 2.04 0.07 3.383320 RB5888 96 291 Transposase IS3/IS911 3087636 3087346 10.1 11124 - 5.61 2.09 0.17 8.124555 RB8076 173 522 Holliday junction resolvase YqgF 3.1.-.- 4323454 4323975 5.1 18918 + 5.57 1.50 0.24 15.684721 RB8397 708 2127 UvrABC system protein B uvrB 4489596 4487470 6.1 80350 - 5.59 1.75 0.32 18.156052 RB10876 839 2520 excinuclease ABC subunit A 5903770 5901251 6.4 90046 - 5.80 1.51 0.11 7.016558 RB11749 96 291 Transposase IS3/IS911 6328064 6328354 10.1 11124 + 5.42 2.08 0.28 13.366559 RB11750 292 879 integrase 6328357 6329235 10 33354 + 5.75 1.85 0.13 6.806690 RB12007 526 1581 deoxyribodipyrimidine photo-lyase phrB 4.1.99.3 6478047 6476467 6.2 61173 - 5.52 1.54 0.08 5.057098 RB12746 294 885 Formamidopyrimidine-DNA glycolase mutM 3.2.2.23 6869843 6870727 10.3 32447 + 5.59 3.07 0.18 5.807203 RB12940 96 291 Transposase IS3/IS911 6966246 6966536 10.1 11124 + 5.46 1.92 0.23 12.147353 RB13225 560 1683 ATP-dependent DNA helicase RecQ 7096300 7097982 9.8 61698 + 6.34 1.89 0.09 4.903620 RB6428 164 495 conserved hypothetical protein, secreted 3382816 3382322 6.5 17292 - 7.41 2.96 0.13 4.556653 RB11941 427 1284 Glycosyl transferase, family 2 6429975 6428692 7.8 47063 - 5.32 1.73 0.34 19.716759 RB12148 708 2127 Tail-specific protease precursor (Protease Re) prc 3.4.21.102 6556800 6554674 4.6 79196 - 5.88 2.20 0.01 0.657021 RB12606 428 1287 Bacterial sugar transferase 2.7.8.6 6791866 6790580 10.7 48185 - 5.94 1.90 0.10 5.337150 RB12831 264 795 Glycosyltransferase 6921326 6920532 4.6 29501 - 6.09 2.57 0.05 1.866425 RB11505 199 600 conserved hypothetical protein, secreted 6221829 6221230 7.5 21660 - 6.13 2.37 0.03 1.456603 RB11851 346 1041 Bacterial type II secretion system protein 6381415 6382455 9.3 37954 + 5.93 1.54 0.27 17.37731 RB1359 629 1890 Protease do precursor degP 3.4.21.- 693488 695377 6.1 66729 + 5.96 2.83 0.08 2.98
3812 RB6751 859 2580 chaperone ClpB clpB 3586111 3588690 5.2 95764 + 6.34 1.94 0.04 2.285026 RB8966 660 1983 Chaperonin Cpn60/TCP-1 4777493 4779475 5.3 69819 + 6.56 2.86 0.23 7.895027 RB8969 101 306 Chaperonin Cpn10 4779554 4779859 4.9 10628 + 6.39 3.13 0.15 4.685028 RB8970 540 1623 GroEL 4779905 4781527 4.8 57516 + 6.74 2.95 0.22 7.565029 RB8972 391 1176 Chaperone protein dnaJ dnaJ 4781697 4782872 5.7 42391 + 6.25 1.78 0.11 6.265030 RB8974 200 603 GrpE protein grpE 4782869 4783471 4.2 21927 + 5.96 3.18 0.22 6.795092 RB9103 899 2700 ATP-dependent Clp protease, ATP-binding subunit ClpB 4866549 4863850 4.8 99439 - 6.05 2.99 0.05 1.695094 RB9105 704 2115 Heat shock protein Hsp70 dnaK 4868834 4866720 4.6 76038 - 6.67 3.47 0.36 10.355713 RB10242 330 993 Curved DNA-binding protein cbpA 5536118 5535126 6.7 35177 - 5.86 2.74 0.17 6.375733 RB10279 151 456 heat shock protein, Hsp20 family GrpE 5557118 5557573 4.9 17072 + 6.40 2.84 0.21 7.335734 RB10283 107 324 small heat shock protein 5557737 5558060 4.7 12267 + 6.97 4.58 0.38 8.235911 RB10627 93 282 Chaperonin Cpn10 5769148 5769429 3.9 9966 + 7.13 2.27 0.07 3.155912 RB10629 539 1620 GroEL 5769500 5771119 4.9 57511 + 6.93 2.99 0.15 4.956601 RB11847 243 732 Peptide methionine sulfoxide reductase msrA msrA 1.8.4.6 6379337 6380068 6.5 26839 + 5.98 1.91 0.21 11.146619 RB11878 168 507 peptide methionine sulfoxide reductase msrA msrA3 1.8.4.6 6396598 6397104 4.8 19100 + 5.75 2.32 0.26 11.036727 RB12080 492 1479 Protein-L-isoaspartate(D-aspartate) O-methyltransferase 2.1.1.77 6517231 6515753 8.9 53385 - 5.57 1.79 0.09 4.764754 RB8465 184 555 Ferric-uptake regulator fur 4520236 4519682 9.7 21381 - 5.83 1.92 0.12 6.035967 RB10727 276 831 manganese-containing catalase 5830775 5831605 5 30055 + 6.08 2.69 0.16 6.056102 RB10965 204 615 conserved hypothetical protein, membrane 5942116 5942730 4.9 21483 + 5.61 1.52 0.05 3.51708 RB1323 429 1290 Protein of unknown function DUF894, DitE 672945 671656 11.8 45426 - 5.96 2.01 0.03 1.52
3648 RB6469 252 759 ABC transporter ATP-binding protein 3419515 3420273 5.6 27684 + 6.01 2.01 0.08 4.194868 RB8679 477 1434 oxidoreductase 4620013 4618580 5.1 51903 - 5.56 1.66 0.14 8.675260 RB9420 663 1992 Lipid A export ATP-binding/permease protein msbA 5055368 5053377 9.3 73442 - 6.00 2.11 0.30 14.065727 RB10270 537 1614 phytoene dehydrogenase carC 1.14.99.- 5551969 5550356 5.5 60117 - 5.43 2.81 0.23 8.186655 RB11943 534 1605 phytoene desaturase 6432376 6430772 5.8 57855 - 5.58 1.64 0.33 19.906414 RB11488 265 798 oxidoreductase, short-chain dehydrogenase/reductase family 6213893 6214690 5.1 27687 + 5.54 1.92 0.08 4.38557 RB1044 269 810 TatD-related deoxyribonuclease 543422 544231 5.2 30726 + 5.84 1.55 0.18 11.86745 RB1391 538 1617 cell division inhibitor- 710780 709164 7.8 59400 - 6.07 1.84 0.15 8.25846 RB1577 860 2583 ATP dependent DNA helicase 806289 808871 9.9 95417 + 5.78 2.12 0.21 10.05952 RB1798 364 1095 methanol dehydrogenase regulatory protein 930673 931767 4.6 40611 + 5.60 1.87 0.24 12.86
3602 RB6395 778 2337 conserved hypothetical protein 3364879 3367215 4.6 85714 + 5.80 1.71 0.04 2.313936 RB6982 286 861 ABC transporter ATP-binding protein 3715019 3715879 5 31150 + 5.61 1.73 0.14 7.994596 RB8157 364 1095 conserved hypothetical protein 4360483 4359389 5.1 40859 - 5.49 1.59 0.21 13.144603 RB8173 339 1020 MoxR protein 4366881 4367900 5.1 37556 + 5.64 2.12 0.45 21.294622 RB8214 589 1770 membrane protein 4392948 4391179 4.5 63301 - 5.92 2.04 0.21 10.466104 RB10967 341 1026 quinone oxidoreductase-like protein 5944040 5943015 4.5 37393 - 6.36 1.71 0.12 6.976106 RB10971 303 912 Short-chain dehydrogenase/reductase SDR 5946406 5945495 9.8 32799 - 6.32 2.05 0.08 3.956267 RB11262 331 996 diacylglycerol kinase 6108302 6107307 5 35708 - 6.02 2.26 0.09 3.826641 RB11923 231 696 Electron transport protein SCO1/SenC 6419736 6419041 4.3 25792 - 5.60 1.60 0.17 10.767369 RB13257 286 861 2-hydroxy-6-ketonona-2,4-dienedioic acid hydrolase bphD 3.7.1.- 7105590 7106450 4.6 32540 + 5.50 2.31 0.13 5.503490 RB6207 436 1311 conserved hypothetical protein, secreted 3252449 3253759 4.8 48106 + 6.34 1.65 0.33 20.044294 RB7616 221 666 protein containing DUF162 4094752 4094087 4.7 23976 - 5.54 2.27 0.11 4.714641 RB8246 266 801 conserved hypothetical protein 4409241 4408441 6 29589 - 5.84 2.50 0.06 2.254914 RB8767 291 876 conserved hypothetical protein 4660451 4659576 10.1 33956 - 5.59 1.81 0.06 3.376217 RB11176 153 462 protein containing DUF442 6065605 6065144 4.8 17161 - 6.25 1.86 0.14 7.536265 RB11258 237 714 conserved hypothetical protein, membrane 6106840 6106127 10.3 26175 - 6.11 1.80 0.15 8.466348 RB11399 132 399 Putative thiol-disulphide oxidoreductase DCC 6179167 6179565 6.5 15047 + 6.70 3.28 0.09 2.616422 RB11498 446 1341 conserved hypothetical protein 6218361 6219701 5.4 50914 + 6.18 1.66 0.08 4.603294 RB5848 140 423 dnaK suppressor protein, 3066328 3065906 4.6 15633 - 6.33 1.75 0.16 8.986266 RB11260 121 366 dnaK suppressor protein, 6107202 6106837 5.2 13639 - 5.99 2.55 0.08 3.28
79 RB138 214 645 Sigma factor, ECF-like 60872 61516 4.8 24057 + 5.40 2.67 0.23 8.52300 RB520 375 1128 conserved hypothetical protein 284989 286116 4.8 41300 + 6.06 1.73 0.12 6.85301 RB521 63 192 hypothetical protein 286399 286208 10.7 7040 - 6.02 2.31 0.49 21.24316 RB546 485 1458 secreted protein containing DUF1501 302834 304291 7.8 53385 + 6.08 1.86 0.17 9.14319 RB550 137 414 secreted protein 306709 306296 12.2 15112 - 5.75 1.65 0.20 12.43358 RB655 346 1041 conserved hypothetical protein, UCP012641 family 354993 353953 5.2 38936 - 6.54 1.74 0.18 10.28362 RB663 79 240 hypothetical protein 357608 357369 11.4 8792 - 6.39 1.71 0.20 11.59366 RB671 122 369 hypothetical protein 359218 359586 10.7 13383 + 6.22 1.80 0.31 17.13580 RB1095 525 1578 hypothetical protein 567469 569046 4.5 57051 + 5.80 2.27 0.11 4.78662 RB1239 155 468 hypothetical protein 632500 632033 9.7 17106 - 5.80 2.22 0.10 4.34707 RB1322 35 108 hypothetical protein 671526 671633 12.8 3914 + 5.82 2.45 0.17 6.83709 RB1324 77 234 hypothetical protein 673169 672936 10.9 8459 - 5.72 1.69 0.08 4.76747 RB1394 78 237 hypothetical protein 711994 711758 10.1 8765 - 5.64 1.68 0.20 11.85748 RB1395 319 960 secreted protein 712097 713056 5.5 35212 + 6.35 2.10 0.09 4.31869 RB1618 97 294 hypothetical protein 833743 834036 5.3 11108 + 5.64 1.87 0.28 14.76915 RB1728 285 858 membrane protein containing DUF1206 899954 900811 9.9 30055 + 6.10 2.71 0.19 7.20916 RB1729 120 363 hypothetical protein 901146 900784 7.7 13145 - 5.58 2.76 0.08 2.88947 RB1789 243 732 conserved hypothetical protein 925861 926592 9.4 27028 + 5.76 1.60 0.18 10.95
1711 RB3159 73 222 protein containing DUF1590 1629337 1629116 12.1 8119 - 6.16 1.62 0.19 11.682351 RB4256 358 1077 conserved hypothetical protein, secreted 2180303 2181379 4.5 37091 + 7.60 1.65 0.06 3.592388 RB4306 457 1374 transposase and inactivated derivative 2221207 2219834 8.4 52492 - 5.68 1.74 0.15 8.712407 RB4347 156 471 conserved hypothetical protein 2235735 2236205 4.7 17459 + 7.56 1.57 0.08 4.782440 RB4397 55 168 protein containing DUF1560 2263976 2264143 10.5 5816 + 5.95 2.96 0.11 3.722458 RB4426 60 183 hypothetical protein 2275786 2275604 6.3 6516 - 5.18 1.87 0.30 16.152461 RB4429 89 270 conserved hypothetical protein 2275996 2276265 4.9 10034 + 7.01 1.62 0.07 4.072465 RB4438 160 483 Pyridoxamine 5'-phosphate oxidase- 2278247 2278729 4.4 18043 + 6.37 1.77 0.07 3.882609 RB4679 175 528 secreted protein 2391214 2391741 4.2 18596 + 6.73 1.56 0.17 11.032951 RB5238 73 222 hypothetical protein 2697775 2697996 10.6 7866 + 5.78 2.22 0.24 10.713010 RB5348 457 1374 transposase and inactivated derivative 2765787 2764414 8.4 52492 - 5.86 1.52 0.06 4.263022 RB5370 309 930 Rhodopirellula transposase 2774826 2773897 5.9 34530 - 5.74 2.09 0.18 8.773023 RB5371 109 330 hypothetical protein 2774829 2775158 7.5 12033 + 5.15 1.52 0.15 10.153118 RB5523 91 276 hypothetical protein 2860984 2860709 6.3 9939 - 5.87 1.79 0.14 7.58
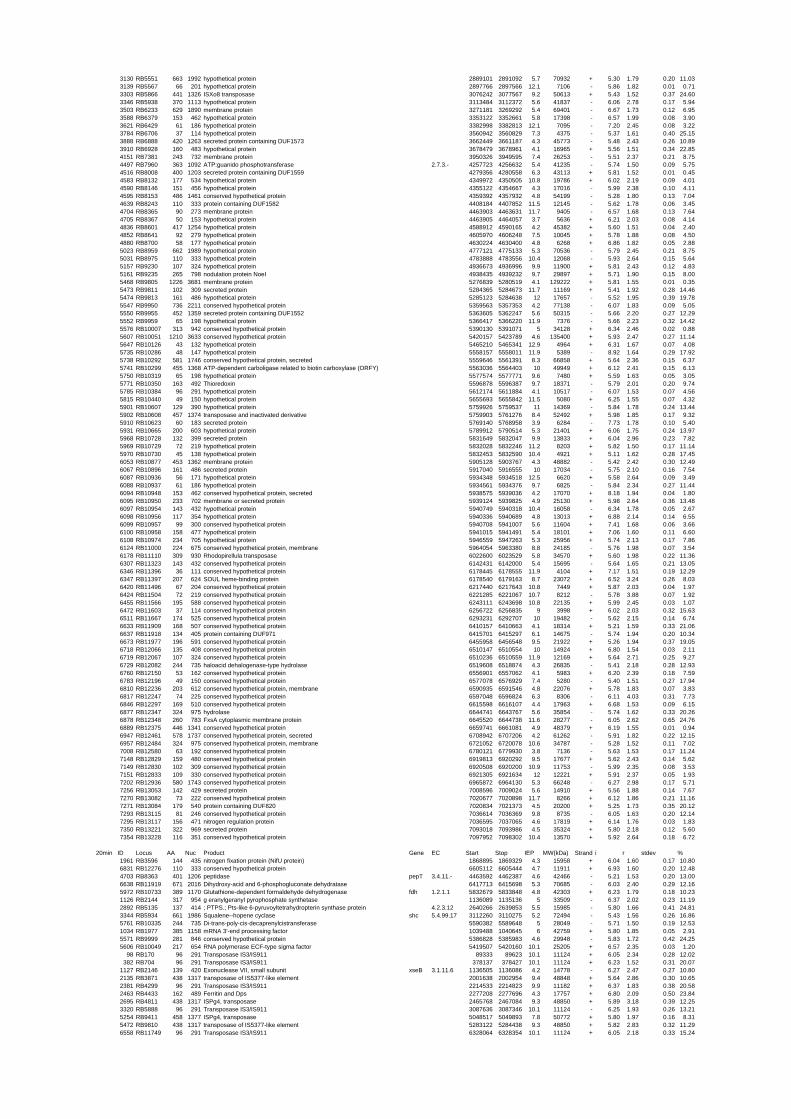
3130 RB5551 663 1992 hypothetical protein 2889101 2891092 5.7 70932 + 5.30 1.79 0.20 11.033139 RB5567 66 201 hypothetical protein 2897766 2897566 12.1 7106 - 5.86 1.82 0.01 0.713303 RB5866 441 1326 ISXo8 transposase 3076242 3077567 9.2 50613 + 5.43 1.52 0.37 24.603346 RB5938 370 1113 hypothetical protein 3113484 3112372 5.6 41837 - 6.06 2.78 0.17 5.943503 RB6233 629 1890 membrane protein 3271181 3269292 5.4 69401 - 6.67 1.73 0.12 6.953588 RB6379 153 462 hypothetical protein 3353122 3352661 5.8 17398 - 6.57 1.99 0.08 3.903621 RB6429 61 186 hypothetical protein 3382998 3382813 12.1 7095 - 7.20 2.45 0.08 3.223784 RB6706 37 114 hypothetical protein 3560942 3560829 7.3 4375 - 5.37 1.61 0.40 25.153888 RB6888 420 1263 secreted protein containing DUF1573 3662449 3661187 4.3 45773 - 5.48 2.43 0.26 10.893910 RB6928 160 483 hypothetical protein 3678479 3678961 4.1 16965 + 5.56 1.51 0.34 22.854151 RB7381 243 732 membrane protein 3950326 3949595 7.4 26253 - 5.51 2.37 0.21 8.754497 RB7960 363 1092 ATP:guanido phosphotransferase 2.7.3.- 4257723 4256632 5.4 41235 - 5.74 1.50 0.09 5.754516 RB8008 400 1203 secreted protein containing DUF1559 4279356 4280558 6.3 43113 + 5.81 1.52 0.01 0.454583 RB8132 177 534 hypothetical protein 4349972 4350505 10.8 19786 + 6.02 2.19 0.09 4.014590 RB8146 151 456 hypothetical protein 4355122 4354667 4.3 17016 - 5.99 2.38 0.10 4.114595 RB8153 486 1461 conserved hypothetical protein 4359392 4357932 4.8 54199 - 5.28 1.80 0.13 7.044639 RB8243 110 333 protein containing DUF1582 4408184 4407852 11.5 12145 - 5.62 1.78 0.06 3.454704 RB8365 90 273 membrane protein 4463903 4463631 11.7 9405 - 6.57 1.68 0.13 7.644705 RB8367 50 153 hypothetical protein 4463905 4464057 3.7 5636 + 6.21 2.03 0.08 4.144836 RB8601 417 1254 hypothetical protein 4588912 4590165 4.2 45382 + 5.60 1.51 0.04 2.404852 RB8641 92 279 hypothetical protein 4605970 4606248 7.5 10045 + 5.78 1.88 0.08 4.504880 RB8700 58 177 hypothetical protein 4630224 4630400 4.8 6268 + 6.86 1.82 0.05 2.885023 RB8959 662 1989 hypothetical protein 4777121 4775133 5.3 70536 - 5.79 2.45 0.21 8.755031 RB8975 110 333 hypothetical protein 4783888 4783556 10.4 12068 - 5.93 2.64 0.15 5.645157 RB9230 107 324 hypothetical protein 4936673 4936996 9.9 11900 + 5.81 2.43 0.12 4.835161 RB9235 265 798 nodulation protein NoeI 4938435 4939232 9.7 29897 + 5.71 1.90 0.15 8.005468 RB9805 1226 3681 membrane protein 5276839 5280519 4.1 129222 + 5.81 1.55 0.01 0.355473 RB9811 102 309 secreted protein 5284365 5284673 11.7 11169 + 5.41 1.92 0.28 14.465474 RB9813 161 486 hypothetical protein 5285123 5284638 12 17657 - 5.52 1.95 0.39 19.785547 RB9950 736 2211 conserved hypothetical protein 5359563 5357353 4.2 77138 - 6.07 1.83 0.09 5.055550 RB9955 452 1359 secreted protein containing DUF1552 5363605 5362247 5.6 50315 - 5.66 2.20 0.27 12.295552 RB9959 65 198 hypothetical protein 5366417 5366220 11.9 7376 - 5.66 2.23 0.32 14.425576 RB10007 313 942 conserved hypothetical protein 5390130 5391071 5 34128 + 6.34 2.46 0.02 0.885607 RB10051 1210 3633 conserved hypothetical protein 5420157 5423789 4.6 135400 + 5.93 2.47 0.27 11.145647 RB10126 43 132 hypothetical protein 5465210 5465341 12.9 4964 + 6.31 1.67 0.07 4.085735 RB10286 48 147 hypothetical protein 5558157 5558011 11.9 5389 - 8.92 1.64 0.29 17.925738 RB10292 581 1746 conserved hypothetical protein, secreted 5559646 5561391 8.3 66858 + 5.64 2.36 0.15 6.375741 RB10299 455 1368 ATP-dependent carboligase related to biotin carboxylase (ORFY) 5563036 5564403 10 49949 + 6.12 2.41 0.15 6.135750 RB10319 65 198 hypothetical protein 5577574 5577771 9.6 7480 + 5.59 1.63 0.05 3.055771 RB10350 163 492 Thioredoxin 5596878 5596387 9.7 18371 - 5.79 2.01 0.20 9.745785 RB10384 96 291 hypothetical protein 5612174 5611884 4.1 10517 - 6.07 1.53 0.07 4.565815 RB10440 49 150 hypothetical protein 5655693 5655842 11.5 5080 + 6.25 1.55 0.07 4.325901 RB10607 129 390 hypothetical protein 5759926 5759537 11 14369 - 5.84 1.78 0.24 13.445902 RB10608 457 1374 transposase and inactivated derivative 5759903 5761276 8.4 52492 + 5.98 1.85 0.17 9.325910 RB10623 60 183 secreted protein 5769140 5768958 3.9 6284 - 7.73 1.78 0.10 5.405931 RB10665 200 603 hypothetical protein 5789912 5790514 5.3 21401 + 6.06 1.75 0.24 13.975968 RB10728 132 399 secreted protein 5831649 5832047 9.9 13833 + 6.04 2.96 0.23 7.825969 RB10729 72 219 hypothetical protein 5832028 5832246 11.2 8203 + 5.82 1.50 0.17 11.145970 RB10730 45 138 hypothetical protein 5832453 5832590 10.4 4921 + 5.11 1.62 0.28 17.456053 RB10877 453 1362 membrane protein 5905128 5903767 4.3 48882 - 5.42 2.42 0.30 12.496067 RB10896 161 486 secreted protein 5917040 5916555 10 17034 - 5.75 2.10 0.16 7.546087 RB10936 56 171 hypothetical protein 5934348 5934518 12.5 6620 + 5.58 2.64 0.09 3.496088 RB10937 61 186 hypothetical protein 5934561 5934376 9.7 6825 - 5.84 2.34 0.27 11.446094 RB10948 153 462 conserved hypothetical protein, secreted 5938575 5939036 4.2 17070 + 8.18 1.94 0.04 1.806095 RB10950 233 702 membrane or secreted protein 5939124 5939825 4.9 25130 + 5.98 2.64 0.36 13.486097 RB10954 143 432 hypothetical protein 5940749 5940318 10.4 16058 - 6.34 1.78 0.05 2.676098 RB10956 117 354 hypothetical protein 5940336 5940689 4.8 13013 + 6.88 2.14 0.14 6.556099 RB10957 99 300 conserved hypothetical protein 5940708 5941007 5.6 11604 + 7.41 1.68 0.06 3.666100 RB10958 158 477 hypothetical protein 5941015 5941491 5.4 18101 + 7.06 1.60 0.11 6.606108 RB10974 234 705 hypothetical protein 5946559 5947263 5.3 25956 + 5.74 2.13 0.17 7.866124 RB11000 224 675 conserved hypothetical protein, membrane 5964054 5963380 8.8 24185 - 5.76 1.98 0.07 3.546178 RB11110 309 930 Rhodopirellula transposase 6022600 6023529 5.8 34570 + 5.60 1.98 0.22 11.366307 RB11323 143 432 conserved hypothetical protein 6142431 6142000 5.4 15695 - 5.64 1.65 0.21 13.056346 RB11396 36 111 conserved hypothetical protein 6178445 6178555 11.9 4104 + 7.17 1.51 0.19 12.296347 RB11397 207 624 SOUL heme-binding protein 6178540 6179163 8.7 23072 + 6.52 3.24 0.26 8.036420 RB11496 67 204 conserved hypothetical protein 6217440 6217643 10.8 7449 + 5.87 2.03 0.04 1.976424 RB11504 72 219 conserved hypothetical protein 6221285 6221067 10.7 8212 - 5.78 3.88 0.07 1.926455 RB11566 195 588 conserved hypothetical protein 6243111 6243698 10.8 22135 + 5.99 2.45 0.03 1.076472 RB11603 37 114 conserved hypothetical protein 6256722 6256835 9 3998 + 6.02 2.03 0.32 15.636511 RB11667 174 525 conserved hypothetical protein 6293231 6292707 10 19482 - 5.62 2.15 0.14 6.746633 RB11909 168 507 conserved hypothetical protein 6410157 6410663 4.1 18314 + 5.21 1.59 0.33 21.066637 RB11918 134 405 protein containing DUF971 6415701 6415297 6.1 14675 - 5.74 1.94 0.20 10.346673 RB11977 196 591 conserved hypothetical protein 6455958 6456548 9.5 21922 + 5.26 1.94 0.37 19.056718 RB12066 135 408 conserved hypothetical protein 6510147 6510554 10 14924 + 6.80 1.54 0.03 2.116719 RB12067 107 324 conserved hypothetical protein 6510236 6510559 11.9 12169 + 5.64 2.71 0.25 9.276729 RB12082 244 735 haloacid dehalogenase-type hydrolase 6519608 6518874 4.3 26835 - 5.41 2.18 0.28 12.936760 RB12150 53 162 conserved hypothetical protein 6556901 6557062 4.1 5983 + 6.20 2.39 0.18 7.596783 RB12196 49 150 conserved hypothetical protein 6577078 6576929 7.4 5280 - 5.40 1.51 0.27 17.946810 RB12236 203 612 conserved hypothetical protein, membrane 6590935 6591546 4.8 22076 + 5.78 1.83 0.07 3.836817 RB12247 74 225 conserved hypothetical protein 6597048 6596824 6.3 8306 - 6.11 4.03 0.31 7.736846 RB12297 169 510 conserved hypothetical protein 6615598 6616107 4.4 17963 + 6.68 1.53 0.09 6.156877 RB12347 324 975 hydrolase 6644741 6643767 5.6 35854 - 5.74 1.62 0.33 20.266878 RB12348 260 783 FxsA cytoplasmic membrane protein 6645520 6644738 11.6 28277 - 6.05 2.62 0.65 24.766889 RB12375 446 1341 conserved hypothetical protein 6659741 6661081 4.9 48379 + 6.19 1.55 0.01 0.946947 RB12461 578 1737 conserved hypothetical protein, secreted 6708942 6707206 4.2 61262 - 5.91 1.82 0.22 12.156957 RB12484 324 975 conserved hypothetical protein, membrane 6721052 6720078 10.6 34787 - 5.28 1.52 0.11 7.027008 RB12580 63 192 conserved hypothetical protein 6780121 6779930 3.8 7136 - 5.63 1.53 0.17 11.247148 RB12829 159 480 conserved hypothetical protein 6919813 6920292 9.5 17677 + 5.62 2.43 0.14 5.627149 RB12830 102 309 conserved hypothetical protein 6920508 6920200 10.9 11753 - 5.99 2.35 0.08 3.537151 RB12833 109 330 conserved hypothetical protein 6921305 6921634 12 12221 + 5.91 2.37 0.05 1.937202 RB12936 580 1743 conserved hypothetical protein 6965872 6964130 5.3 66248 - 6.27 2.98 0.17 5.717256 RB13053 142 429 secreted protein 7008596 7009024 5.6 14910 + 5.56 1.88 0.14 7.677270 RB13082 73 222 conserved hypothetical protein 7020677 7020898 11.7 8266 + 6.12 1.86 0.21 11.167271 RB13084 179 540 protein containing DUF820 7020834 7021373 4.5 20200 + 5.25 1.73 0.35 20.127293 RB13115 81 246 conserved hypothetical protein 7036614 7036369 9.8 8735 - 6.05 1.63 0.20 12.147295 RB13117 156 471 nitrogen regulation protein 7036595 7037065 4.6 17819 + 6.14 1.76 0.03 1.837350 RB13221 322 969 secreted protein 7093018 7093986 4.5 35324 + 5.80 2.18 0.12 5.607354 RB13228 116 351 conserved hypothetical protein 7097952 7098302 10.4 13570 + 5.92 2.64 0.18 6.72
20min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand i r stdev %1961 RB3596 144 435 nitrogen fixation protein (NifU protein) 1868895 1869329 4.3 15958 + 6.04 1.60 0.17 10.806831 RB12276 110 333 conserved hypothetical protein 6605112 6605444 4.7 11911 + 6.93 1.60 0.20 12.484703 RB8363 401 1206 peptidase pepT 3.4.11.- 4463592 4462387 4.6 42466 - 5.21 1.53 0.20 13.006638 RB11919 671 2016 Dihydroxy-acid and 6-phosphogluconate dehydratase 6417713 6415698 5.3 70685 - 6.03 2.40 0.29 12.165972 RB10733 389 1170 Glutathione-dependent formaldehyde dehydrogenase fdh 1.2.1.1 5832679 5833848 4.8 42303 + 6.23 1.79 0.18 10.231126 RB2144 317 954 g eranylgeranyl pyrophosphate synthetase 1136089 1135136 5 33509 - 6.37 2.02 0.23 11.192892 RB5135 137 414 ; PTPS.; Pts-like 6-pyruvoyltetrahydropterin synthase protein 4.2.3.12 2640266 2639853 5.5 15985 - 5.80 1.66 0.41 24.813344 RB5934 661 1986 Squalene--hopene cyclase shc 5.4.99.17 3112260 3110275 5.2 72494 - 5.43 1.56 0.26 16.865761 RB10335 244 735 Di-trans-poly-cis-decaprenylcistransferase 5590382 5589648 5 28049 - 5.71 1.50 0.19 12.531034 RB1977 385 1158 mRNA 3'-end processing factor 1039488 1040645 6 42759 + 5.80 1.85 0.05 2.915571 RB9999 281 846 conserved hypothetical protein 5386828 5385983 4.6 29948 - 5.83 1.72 0.42 24.255606 RB10049 217 654 RNA polymerase ECF-type sigma factor 5419507 5420160 10.1 25205 + 6.57 2.35 0.03 1.20
98 RB170 96 291 Transposase IS3/IS911 89333 89623 10.1 11124 + 6.05 2.34 0.28 12.02382 RB704 96 291 Transposase IS3/IS911 378137 378427 10.1 11124 + 6.23 1.52 0.31 20.07
1127 RB2146 139 420 Exonuclease VII, small subunit xseB 3.1.11.6 1136505 1136086 4.2 14778 - 6.27 2.47 0.27 10.802135 RB3871 438 1317 transposase of IS5377-like element 2001638 2002954 9.4 48848 + 5.64 2.86 0.30 10.652381 RB4299 96 291 Transposase IS3/IS911 2214533 2214823 9.9 11182 + 6.37 1.83 0.38 20.582463 RB4433 162 489 Ferritin and Dps 2277208 2277696 4.3 17757 + 6.80 2.09 0.50 23.842695 RB4811 438 1317 ISPg4, transposase 2465768 2467084 9.3 48850 + 5.89 3.18 0.39 12.253320 RB5888 96 291 Transposase IS3/IS911 3087636 3087346 10.1 11124 - 6.25 1.93 0.26 13.215254 RB9411 458 1377 ISPg4, transposase 5048517 5049893 7.8 50772 + 5.80 1.97 0.16 8.315472 RB9810 438 1317 transposase of IS5377-like element 5283122 5284438 9.3 48850 + 5.82 2.83 0.32 11.296558 RB11749 96 291 Transposase IS3/IS911 6328064 6328354 10.1 11124 + 6.05 2.18 0.33 15.24
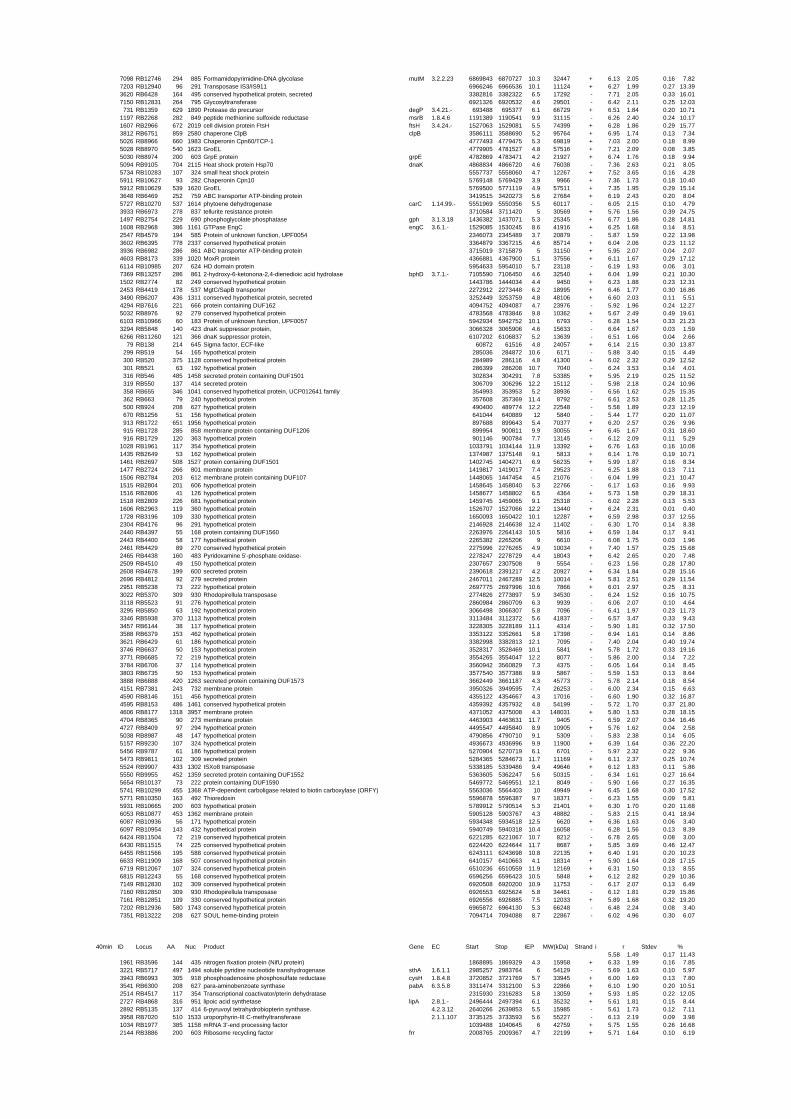
7098 RB12746 294 885 Formamidopyrimidine-DNA glycolase mutM 3.2.2.23 6869843 6870727 10.3 32447 + 6.13 2.05 0.16 7.827203 RB12940 96 291 Transposase IS3/IS911 6966246 6966536 10.1 11124 + 6.27 1.99 0.27 13.393620 RB6428 164 495 conserved hypothetical protein, secreted 3382816 3382322 6.5 17292 - 7.71 2.05 0.33 16.017150 RB12831 264 795 Glycosyltransferase 6921326 6920532 4.6 29501 - 6.42 2.11 0.25 12.03731 RB1359 629 1890 Protease do precursor degP 3.4.21.- 693488 695377 6.1 66729 + 6.51 1.84 0.20 10.71
1197 RB2268 282 849 peptide methionine sulfoxide reductase msrB 1.8.4.6 1191389 1190541 9.9 31115 - 6.26 2.40 0.24 10.171607 RB2966 672 2019 cell division protein FtsH ftsH 3.4.24.- 1527063 1529081 5.5 74399 + 6.28 1.86 0.29 15.773812 RB6751 859 2580 chaperone ClpB clpB 3586111 3588690 5.2 95764 + 6.95 1.74 0.13 7.345026 RB8966 660 1983 Chaperonin Cpn60/TCP-1 4777493 4779475 5.3 69819 + 7.03 2.00 0.18 8.995028 RB8970 540 1623 GroEL 4779905 4781527 4.8 57516 + 7.21 2.09 0.08 3.855030 RB8974 200 603 GrpE protein grpE 4782869 4783471 4.2 21927 + 6.74 1.76 0.18 9.945094 RB9105 704 2115 Heat shock protein Hsp70 dnaK 4868834 4866720 4.6 76038 - 7.36 2.63 0.21 8.055734 RB10283 107 324 small heat shock protein 5557737 5558060 4.7 12267 + 7.52 3.65 0.16 4.285911 RB10627 93 282 Chaperonin Cpn10 5769148 5769429 3.9 9966 + 7.36 1.73 0.18 10.405912 RB10629 539 1620 GroEL 5769500 5771119 4.9 57511 + 7.35 1.95 0.29 15.143648 RB6469 252 759 ABC transporter ATP-binding protein 3419515 3420273 5.6 27684 + 6.19 2.43 0.20 8.045727 RB10270 537 1614 phytoene dehydrogenase carC 1.14.99.- 5551969 5550356 5.5 60117 - 6.05 2.15 0.10 4.793933 RB6973 278 837 tellurite resistance protein 3710584 3711420 5 30569 + 5.76 1.56 0.39 24.751497 RB2754 229 690 phosphoglycolate phosphatase gph 3.1.3.18 1436382 1437071 5.3 25345 + 6.77 1.86 0.28 14.811608 RB2968 386 1161 GTPase EngC engC 3.6.1.- 1529085 1530245 8.6 41916 + 6.25 1.68 0.14 8.512547 RB4579 194 585 Protein of unknown function, UPF0054 2346073 2345489 3.7 20879 - 5.87 1.59 0.22 13.983602 RB6395 778 2337 conserved hypothetical protein 3364879 3367215 4.6 85714 + 6.04 2.06 0.23 11.123936 RB6982 286 861 ABC transporter ATP-binding protein 3715019 3715879 5 31150 + 5.95 2.07 0.04 2.074603 RB8173 339 1020 MoxR protein 4366881 4367900 5.1 37556 + 6.11 1.67 0.29 17.126114 RB10985 207 624 HD domain protein 5954633 5954010 5.7 23118 - 6.19 1.93 0.06 3.017369 RB13257 286 861 2-hydroxy-6-ketonona-2,4-dienedioic acid hydrolase bphD 3.7.1.- 7105590 7106450 4.6 32540 + 6.04 1.99 0.21 10.301502 RB2774 82 249 conserved hypothetical protein 1443786 1444034 4.4 9450 + 6.23 1.88 0.23 12.312453 RB4419 178 537 MgtC/SapB transporter 2272912 2273448 6.2 18995 + 6.46 1.77 0.30 16.863490 RB6207 436 1311 conserved hypothetical protein, secreted 3252449 3253759 4.8 48106 + 6.60 2.03 0.11 5.514294 RB7616 221 666 protein containing DUF162 4094752 4094087 4.7 23976 - 5.92 1.96 0.24 12.275032 RB8976 92 279 conserved hypothetical protein 4783568 4783846 9.8 10362 + 5.67 2.49 0.49 19.616103 RB10966 60 183 Protein of unknown function, UPF0057 5942934 5942752 10.1 6793 - 6.28 1.54 0.33 21.233294 RB5848 140 423 dnaK suppressor protein, 3066328 3065906 4.6 15633 - 6.64 1.67 0.03 1.596266 RB11260 121 366 dnaK suppressor protein, 6107202 6106837 5.2 13639 - 6.51 1.66 0.04 2.66
79 RB138 214 645 Sigma factor, ECF-like 60872 61516 4.8 24057 + 6.14 2.15 0.30 13.87299 RB519 54 165 hypothetical protein 285036 284872 10.6 6171 - 5.88 3.40 0.15 4.49300 RB520 375 1128 conserved hypothetical protein 284989 286116 4.8 41300 + 6.02 2.32 0.29 12.52301 RB521 63 192 hypothetical protein 286399 286208 10.7 7040 - 6.24 3.53 0.14 4.01316 RB546 485 1458 secreted protein containing DUF1501 302834 304291 7.8 53385 + 5.95 2.19 0.25 11.52319 RB550 137 414 secreted protein 306709 306296 12.2 15112 - 5.98 2.18 0.24 10.96358 RB655 346 1041 conserved hypothetical protein, UCP012641 family 354993 353953 5.2 38936 - 6.56 1.62 0.25 15.35362 RB663 79 240 hypothetical protein 357608 357369 11.4 8792 - 6.61 2.53 0.28 11.25500 RB924 208 627 hypothetical protein 490400 489774 12.2 22548 - 5.58 1.89 0.23 12.19670 RB1256 51 156 hypothetical protein 641044 640889 12 5840 - 5.44 1.77 0.20 11.07913 RB1722 651 1956 hypothetical protein 897688 899643 5.4 70377 + 6.20 2.57 0.26 9.96915 RB1728 285 858 membrane protein containing DUF1206 899954 900811 9.9 30055 + 6.45 1.67 0.31 18.60916 RB1729 120 363 hypothetical protein 901146 900784 7.7 13145 - 6.12 2.09 0.11 5.29
1028 RB1961 117 354 hypothetical protein 1033791 1034144 11.9 13392 + 6.76 1.63 0.16 10.081435 RB2649 53 162 hypothetical protein 1374987 1375148 9.1 5813 + 6.14 1.76 0.19 10.711461 RB2697 508 1527 protein containing DUF1501 1402745 1404271 6.9 56235 + 5.99 1.87 0.16 8.341477 RB2724 266 801 membrane protein 1419817 1419017 7.4 29523 - 6.25 1.88 0.13 7.111506 RB2784 203 612 membrane protein containing DUF107 1448065 1447454 4.5 21076 - 6.04 1.99 0.21 10.471515 RB2804 201 606 hypothetical protein 1458645 1458040 5.3 22766 - 6.17 1.63 0.16 9.931516 RB2806 41 126 hypothetical protein 1458677 1458802 6.5 4364 + 5.73 1.58 0.29 18.311518 RB2809 226 681 hypothetical protein 1459745 1459065 9.1 25318 - 6.02 2.28 0.13 5.531606 RB2963 119 360 hypothetical protein 1526707 1527066 12.2 13440 + 6.24 2.31 0.01 0.401728 RB3196 109 330 hypothetical protein 1650093 1650422 10.1 12287 + 6.59 2.98 0.37 12.552304 RB4176 96 291 hypothetical protein 2146928 2146638 12.4 11402 - 6.30 1.70 0.14 8.382440 RB4397 55 168 protein containing DUF1560 2263976 2264143 10.5 5816 + 6.59 1.84 0.17 9.412443 RB4400 58 177 hypothetical protein 2265382 2265206 9 6610 - 6.08 1.75 0.03 1.962461 RB4429 89 270 conserved hypothetical protein 2275996 2276265 4.9 10034 + 7.40 1.57 0.25 15.682465 RB4438 160 483 Pyridoxamine 5'-phosphate oxidase- 2278247 2278729 4.4 18043 + 6.42 2.65 0.20 7.482509 RB4510 49 150 hypothetical protein 2307657 2307508 9 5554 - 6.23 1.56 0.28 17.802608 RB4678 199 600 secreted protein 2390618 2391217 4.2 20927 + 6.34 1.84 0.28 15.162696 RB4812 92 279 secreted protein 2467011 2467289 12.5 10014 + 5.81 2.51 0.29 11.542951 RB5238 73 222 hypothetical protein 2697775 2697996 10.6 7866 + 6.01 2.97 0.25 8.313022 RB5370 309 930 Rhodopirellula transposase 2774826 2773897 5.9 34530 - 6.24 1.52 0.16 10.753118 RB5523 91 276 hypothetical protein 2860984 2860709 6.3 9939 - 6.06 2.07 0.10 4.643295 RB5850 63 192 hypothetical protein 3066498 3066307 5.8 7096 - 6.41 1.97 0.23 11.733346 RB5938 370 1113 hypothetical protein 3113484 3112372 5.6 41837 - 6.57 3.47 0.33 9.433457 RB6144 38 117 hypothetical protein 3228305 3228189 11.1 4314 - 5.90 1.81 0.32 17.503588 RB6379 153 462 hypothetical protein 3353122 3352661 5.8 17398 - 6.94 1.61 0.14 8.863621 RB6429 61 186 hypothetical protein 3382998 3382813 12.1 7095 - 7.40 2.04 0.40 19.743746 RB6637 50 153 hypothetical protein 3528317 3528469 10.1 5841 + 5.78 1.72 0.33 19.163771 RB6685 72 219 hypothetical protein 3554265 3554047 12.2 8077 - 5.86 2.00 0.14 7.223784 RB6706 37 114 hypothetical protein 3560942 3560829 7.3 4375 - 6.05 1.64 0.14 8.453803 RB6735 50 153 hypothetical protein 3577540 3577388 9.9 5867 - 5.59 1.53 0.13 8.643888 RB6888 420 1263 secreted protein containing DUF1573 3662449 3661187 4.3 45773 - 5.78 2.14 0.18 8.544151 RB7381 243 732 membrane protein 3950326 3949595 7.4 26253 - 6.00 2.34 0.15 6.634590 RB8146 151 456 hypothetical protein 4355122 4354667 4.3 17016 - 6.60 1.90 0.32 16.874595 RB8153 486 1461 conserved hypothetical protein 4359392 4357932 4.8 54199 - 5.72 1.70 0.37 21.804606 RB8177 1318 3957 membrane protein 4371052 4375008 4.3 148031 + 5.80 1.53 0.28 18.154704 RB8365 90 273 membrane protein 4463903 4463631 11.7 9405 - 6.59 2.07 0.34 16.464727 RB8409 97 294 hypothetical protein 4495547 4495840 8.9 10905 + 5.76 1.62 0.04 2.585038 RB8987 48 147 hypothetical protein 4790856 4790710 9.1 5309 - 5.83 2.38 0.14 6.055157 RB9230 107 324 hypothetical protein 4936673 4936996 9.9 11900 + 6.39 1.64 0.36 22.205456 RB9787 61 186 hypothetical protein 5270904 5270719 6.1 6701 - 5.97 2.32 0.22 9.365473 RB9811 102 309 secreted protein 5284365 5284673 11.7 11169 + 6.11 2.37 0.25 10.745524 RB9907 433 1302 ISXo8 transposase 5338185 5339486 9.4 49646 + 6.12 1.83 0.11 5.865550 RB9955 452 1359 secreted protein containing DUF1552 5363605 5362247 5.6 50315 - 6.34 1.61 0.27 16.645654 RB10137 73 222 protein containing DUF1590 5469772 5469551 12.1 8049 - 5.90 1.66 0.27 16.355741 RB10299 455 1368 ATP-dependent carboligase related to biotin carboxylase (ORFY) 5563036 5564403 10 49949 + 6.45 1.68 0.30 17.525771 RB10350 163 492 Thioredoxin 5596878 5596387 9.7 18371 - 6.23 1.55 0.09 5.815931 RB10665 200 603 hypothetical protein 5789912 5790514 5.3 21401 + 6.30 1.70 0.20 11.686053 RB10877 453 1362 membrane protein 5905128 5903767 4.3 48882 - 5.83 2.15 0.41 18.946087 RB10936 56 171 hypothetical protein 5934348 5934518 12.5 6620 + 6.36 1.63 0.06 3.406097 RB10954 143 432 hypothetical protein 5940749 5940318 10.4 16058 - 6.28 1.56 0.13 8.396424 RB11504 72 219 conserved hypothetical protein 6221285 6221067 10.7 8212 - 6.78 2.65 0.08 3.006430 RB11515 74 225 conserved hypothetical protein 6224420 6224644 11.7 8687 + 5.85 3.69 0.46 12.476455 RB11566 195 588 conserved hypothetical protein 6243111 6243698 10.8 22135 + 6.40 1.91 0.20 10.236633 RB11909 168 507 conserved hypothetical protein 6410157 6410663 4.1 18314 + 5.90 1.64 0.28 17.156719 RB12067 107 324 conserved hypothetical protein 6510236 6510559 11.9 12169 + 6.31 1.50 0.13 8.556815 RB12243 55 168 conserved hypothetical protein 6596256 6596423 10.5 5848 + 6.12 2.82 0.29 10.367149 RB12830 102 309 conserved hypothetical protein 6920508 6920200 10.9 11753 - 6.17 2.07 0.13 6.497160 RB12850 309 930 Rhodopirellula transposase 6926553 6925624 5.8 34461 - 6.12 1.81 0.29 15.867161 RB12851 109 330 conserved hypothetical protein 6926556 6926885 7.5 12033 + 5.89 1.68 0.32 19.207202 RB12936 580 1743 conserved hypothetical protein 6965872 6964130 5.3 66248 - 6.48 2.24 0.08 3.407351 RB13222 208 627 SOUL heme-binding protein 7094714 7094088 8.7 22867 - 6.02 4.96 0.30 6.07
40min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand i r Stdev %5.58 1.49 0.17 11.43
1961 RB3596 144 435 nitrogen fixation protein (NifU protein) 1868895 1869329 4.3 15958 + 6.33 1.99 0.16 7.853221 RB5717 497 1494 soluble pyridine nucleotide transhydrogenase sthA 1.6.1.1 2985257 2983764 6 54129 - 5.69 1.63 0.10 5.973943 RB6993 305 918 phosphoadenosine phosphosulfate reductase cysH 1.8.4.8 3720852 3721769 5.7 33945 + 6.00 1.69 0.13 7.803541 RB6300 208 627 para-aminobenzoate synthase pabA 6.3.5.8 3311474 3312100 5.3 22866 + 6.10 1.90 0.20 10.512514 RB4517 117 354 Transcriptional coactivator/pterin dehydratase 2315930 2316283 5.8 13059 + 5.93 1.85 0.22 12.052727 RB4868 316 951 lipoic acid synthetase lipA 2.8.1.- 2496444 2497394 6.1 35232 + 5.61 1.81 0.15 8.442892 RB5135 137 414 6-pyruvoyl tetrahydrobiopterin synthase. 4.2.3.12 2640266 2639853 5.5 15985 - 5.61 1.73 0.12 7.113958 RB7020 510 1533 uroporphyrin-III C-methyltransferase 2.1.1.107 3735125 3733593 5.6 55227 - 6.13 2.19 0.09 3.981034 RB1977 385 1158 mRNA 3'-end processing factor 1039488 1040645 6 42759 + 5.75 1.55 0.26 16.682144 RB3886 200 603 Ribosome recycling factor frr 2008765 2009367 4.7 22199 + 5.71 1.64 0.10 6.19

5194 RB9305 360 1083 peptide chain release factor 1 prfA 4974620 4975702 4.9 40794 + 6.48 1.55 0.12 7.72456 RB838 253 762 transcriptional regulator, TetR family 441593 440832 5.7 28095 - 5.90 1.91 0.06 2.96981 RB1862 135 408 transcriptional regulator, GntR family 960630 961037 10.2 14884 + 5.63 1.85 0.19 10.13
4878 RB8695 131 396 transcriptional regulator, GntR family 4630087 4629692 9.9 14673 - 5.87 1.55 0.14 9.065571 RB9999 281 846 conserved hypothetical protein 5386828 5385983 4.6 29948 - 6.23 2.72 0.14 5.147362 RB13241 167 504 RNA polymerase ECF-type sigma factor 7103064 7102561 8.8 19488 - 5.89 1.79 0.18 9.994187 RB7437 239 720 LexA repressor lexA 3.4.21.88 3987953 3988672 9.8 25951 + 5.68 1.67 0.26 15.67382 RB704 96 291 Transposase IS3/IS911 378137 378427 10.1 11124 + 6.15 1.89 0.22 11.57522 RB957 195 588 Tyrosine recombinase xerD xerD 505476 504889 10.2 21971 - 5.31 1.64 0.09 5.63628 RB1189 96 291 Transposase IS3/IS911 606838 607128 10.1 11124 + 6.33 1.58 0.28 17.48
2381 RB4299 96 291 Transposase IS3/IS911 2214533 2214823 9.9 11182 + 6.38 2.19 0.23 10.262463 RB4433 162 489 Ferritin and Dps 2277208 2277696 4.3 17757 + 6.49 2.15 0.18 8.273320 RB5888 96 291 Transposase IS3/IS911 3087636 3087346 10.1 11124 - 6.23 2.22 0.14 6.363420 RB6070 201 606 HNH endonuclease family protein 3186811 3187416 9.9 23317 + 6.02 1.62 0.18 11.235728 RB10271 158 477 ribonuclease H rnhA 3.1.26.4 5552547 5552071 9.1 17665 - 5.88 1.97 0.22 11.086481 RB11618 552 1659 type I restriction-modification system, M subunit, 6263708 6265366 6.7 62726 + 5.65 2.25 0.15 6.746558 RB11749 96 291 Transposase IS3/IS911 6328064 6328354 10.1 11124 + 6.10 2.92 0.26 8.916559 RB11750 292 879 integrase 6328357 6329235 10 33354 + 6.29 1.62 0.12 7.516577 RB11802 96 291 Transposase IS3/IS911 6361950 6362240 10.1 11124 + 5.81 1.73 0.19 11.047098 RB12746 294 885 Formamidopyrimidine-DNA glycolase mutM 3.2.2.23 6869843 6870727 10.3 32447 + 5.93 1.91 0.03 1.607203 RB12940 96 291 Transposase IS3/IS911 6966246 6966536 10.1 11124 + 6.21 2.44 0.20 8.207150 RB12831 264 795 Glycosyltransferase 6921326 6920532 4.6 29501 - 6.47 2.10 0.21 9.88731 RB1359 629 1890 Protease do precursor degP 3.4.21.- 693488 695377 6.1 66729 + 6.61 1.81 0.21 11.43
1197 RB2268 282 849 peptide methionine sulfoxide reductase msrB 1.8.4.6 1191389 1190541 9.9 31115 - 6.60 2.34 0.46 19.652726 RB4867 210 633 SmpB RNA-binding protein SmpB 2496354 2495722 10.8 23762 - 6.20 2.52 0.04 1.503812 RB6751 859 2580 chaperone ClpB clpB 3586111 3588690 5.2 95764 + 6.31 1.51 0.29 19.425094 RB9105 704 2115 Heat shock protein Hsp70 dnaK 4868834 4866720 4.6 76038 - 7.04 1.82 0.16 8.895713 RB10242 330 993 Curved DNA-binding protein cbpA 5536118 5535126 6.7 35177 - 6.69 1.59 0.33 20.425911 RB10627 93 282 Chaperonin Cpn10 5769148 5769429 3.9 9966 + 7.37 1.51 0.20 13.523648 RB6469 252 759 ABC transporter ATP-binding protein 3419515 3420273 5.6 27684 + 6.04 2.30 0.12 5.124528 RB8029 245 738 oxidoreductase, short-chain dehydrogenase/reductase family 4292625 4291888 4.5 25817 - 5.59 1.54 0.22 14.455616 RB10074 273 822 conserved hypothetical protein 5435071 5434250 8.6 29283 - 5.96 1.62 0.29 17.873933 RB6973 278 837 tellurite resistance protein 3710584 3711420 5 30569 + 5.67 1.79 0.33 18.672571 RB4619 481 1446 dehydrogenase 2363086 2364531 5.1 52511 + 6.43 1.73 0.29 16.592896 RB5140 613 1842 hypothetical protein 2641937 2643778 5.5 67565 + 5.94 2.08 0.36 17.373602 RB6395 778 2337 conserved hypothetical protein 3364879 3367215 4.6 85714 + 5.85 1.92 0.10 5.154796 RB8533 196 591 protease I pfpI 3.2.-.- 4552172 4552762 4.8 21046 + 6.22 1.55 0.20 12.806370 RB11438 54 165 conserved hypothetical protein 6197926 6197762 11.3 6209 - 6.44 2.55 0.43 17.087369 RB13257 286 861 2-hydroxy-6-ketonona-2,4-dienedioic acid hydrolase bphD 3.7.1.- 7105590 7106450 4.6 32540 + 6.47 2.15 0.06 2.631502 RB2774 82 249 conserved hypothetical protein 1443786 1444034 4.4 9450 + 6.06 1.94 0.19 9.961985 RB3630 313 942 conserved hypothetical protein 1888468 1889409 6.3 34702 + 5.45 1.73 0.33 18.882453 RB4419 178 537 MgtC/SapB transporter 2272912 2273448 6.2 18995 + 6.60 1.79 0.18 9.824776 RB8505 439 1320 serine/threonine protein kinase related protein 4538446 4537127 4.7 48224 - 6.00 1.53 0.10 6.563294 RB5848 140 423 dnaK suppressor protein, 3066328 3065906 4.6 15633 - 6.87 1.84 0.24 13.316266 RB11260 121 366 dnaK suppressor protein, 6107202 6106837 5.2 13639 - 6.53 1.53 0.08 5.19
10 RB17 164 495 protein containing DUF1589 8251 7757 11.5 18498 - 5.72 2.49 0.25 10.0979 RB138 214 645 Sigma factor, ECF-like 60872 61516 4.8 24057 + 6.37 2.18 0.26 12.06
173 RB303 53 162 hypothetical protein 151630 151791 11.5 5832 + 6.21 1.54 0.23 15.15234 RB402 151 456 hypothetical protein 212820 213275 4.4 17306 + 5.62 2.33 0.23 9.74299 RB519 54 165 hypothetical protein 285036 284872 10.6 6171 - 5.20 1.73 0.28 16.00301 RB521 63 192 hypothetical protein 286399 286208 10.7 7040 - 5.45 1.70 0.11 6.37319 RB550 137 414 secreted protein 306709 306296 12.2 15112 - 5.89 2.32 0.12 5.13362 RB663 79 240 hypothetical protein 357608 357369 11.4 8792 - 6.14 1.69 0.23 13.44475 RB873 74 225 hypothetical protein 463836 464060 9.6 8292 + 5.82 1.77 0.15 8.66506 RB933 208 627 hypothetical protein 493451 492825 12.2 22575 - 5.22 2.10 0.27 12.89507 RB934 375 1128 Putative transposase 494203 493076 9.9 41881 - 5.76 2.21 0.22 10.05520 RB955 208 627 hypothetical protein 504140 503514 12.2 22575 - 5.72 2.49 0.07 2.95556 RB1043 89 270 conserved hypothetical protein 543083 543352 4.7 10039 + 6.69 1.52 0.21 13.97624 RB1182 88 267 hypothetical protein 604706 604972 11.6 9996 + 5.97 1.55 0.14 9.24669 RB1255 51 156 hypothetical protein 640910 640755 12.3 5933 - 5.36 1.96 0.01 0.75670 RB1256 51 156 hypothetical protein 641044 640889 12 5840 - 5.23 1.64 0.36 22.15774 RB1449 65 198 hypothetical protein 736500 736697 10 7487 + 5.56 1.66 0.16 9.65947 RB1789 243 732 conserved hypothetical protein 925861 926592 9.4 27028 + 5.99 1.52 0.06 4.06985 RB1872 38 117 hypothetical protein 966053 966169 12.3 4345 + 5.97 1.69 0.15 9.14
1044 RB1999 37 114 hypothetical protein 1053955 1054068 12.2 4228 + 6.14 1.78 0.42 23.361150 RB2185 50 153 hypothetical protein 1152681 1152529 10.1 5841 - 5.64 1.92 0.31 16.111435 RB2649 53 162 hypothetical protein 1374987 1375148 9.1 5813 + 6.48 1.85 0.26 13.861477 RB2724 266 801 membrane protein 1419817 1419017 7.4 29523 - 6.37 2.10 0.21 10.241490 RB2745 41 126 hypothetical protein 1431275 1431400 9.9 4387 + 5.99 1.58 0.35 22.341518 RB2809 226 681 hypothetical protein 1459745 1459065 9.1 25318 - 5.91 2.59 0.36 13.931566 RB2900 90 273 hypothetical protein 1506927 1507199 9.1 9939 + 6.46 1.64 0.31 18.941606 RB2963 119 360 hypothetical protein 1526707 1527066 12.2 13440 + 6.00 1.96 0.10 5.181640 RB3020 99 300 membrane protein 1566234 1565935 9.8 10462 - 6.15 1.57 0.18 11.581711 RB3159 73 222 protein containing DUF1590 1629337 1629116 12.1 8119 - 6.78 1.53 0.08 5.541728 RB3196 109 330 hypothetical protein 1650093 1650422 10.1 12287 + 6.48 2.87 0.20 7.091868 RB3430 248 747 conserved hypothetical protein, membrane 1772646 1773392 7.5 26767 + 6.31 1.70 0.31 18.331957 RB3587 113 342 hypothetical protein 1864796 1864455 11.8 13003 - 5.83 1.55 0.31 19.872169 RB3929 54 165 hypothetical protein 2034407 2034243 10.7 5796 - 5.28 2.41 0.32 13.162170 RB3930 54 165 hypothetical protein 2034417 2034253 11.6 6445 - 5.43 1.50 0.27 18.302330 RB4219 74 225 hypothetical protein 2163305 2163081 6.8 8050 - 5.59 2.05 0.33 16.132440 RB4397 55 168 protein containing DUF1560 2263976 2264143 10.5 5816 + 6.24 1.69 0.39 23.182443 RB4400 58 177 hypothetical protein 2265382 2265206 9 6610 - 6.12 2.05 0.16 7.772458 RB4426 60 183 hypothetical protein 2275786 2275604 6.3 6516 - 5.71 2.12 0.23 10.702465 RB4438 160 483 Pyridoxamine 5'-phosphate oxidase- 2278247 2278729 4.4 18043 + 6.45 2.52 0.07 2.792584 RB4642 85 258 conserved hypothetical protein 2377301 2377558 4.4 9855 + 6.35 2.35 0.09 3.662585 RB4643 96 291 conserved hypothetical protein 2377555 2377845 8.9 11064 + 5.92 2.37 0.14 6.002608 RB4678 199 600 secreted protein 2390618 2391217 4.2 20927 + 6.64 1.91 0.10 5.112609 RB4679 175 528 secreted protein 2391214 2391741 4.2 18596 + 6.74 1.60 0.36 22.492854 RB5069 65 198 hypothetical protein 2609575 2609378 10.3 7107 - 5.36 2.02 0.39 19.402866 RB5092 59 180 secreted protein 2618771 2618950 3.7 6013 + 5.58 1.73 0.08 4.402881 RB5114 38 117 hypothetical protein 2625052 2624936 10.8 4310 - 5.54 1.92 0.17 8.872951 RB5238 73 222 hypothetical protein 2697775 2697996 10.6 7866 + 5.92 2.66 0.11 4.113022 RB5370 309 930 Rhodopirellula transposase 2774826 2773897 5.9 34530 - 5.98 1.60 0.21 13.313023 RB5371 109 330 hypothetical protein 2774829 2775158 7.5 12033 + 5.45 1.62 0.10 6.153047 RB5412 72 219 hypothetical protein 2794838 2794620 12.3 8592 - 6.09 1.58 0.05 2.953118 RB5523 91 276 hypothetical protein 2860984 2860709 6.3 9939 - 5.98 2.19 0.13 5.813130 RB5551 663 1992 hypothetical protein 2889101 2891092 5.7 70932 + 5.94 2.31 0.03 1.363139 RB5567 66 201 hypothetical protein 2897766 2897566 12.1 7106 - 6.03 2.84 0.42 14.693295 RB5850 63 192 hypothetical protein 3066498 3066307 5.8 7096 - 6.34 1.70 0.42 24.803346 RB5938 370 1113 hypothetical protein 3113484 3112372 5.6 41837 - 5.85 2.57 0.20 7.783355 RB5955 177 534 hypothetical protein 3121488 3122021 4.5 19364 + 6.25 1.67 0.18 10.963363 RB5968 207 624 hypothetical protein 3128602 3129225 9.7 23009 + 5.33 1.68 0.31 18.293390 RB6011 50 153 hypothetical protein 3150235 3150083 10.1 5841 - 5.74 1.96 0.28 14.073391 RB6012 433 1302 ISXo8 transposase 3151547 3150246 9.4 49646 - 5.11 2.31 0.34 14.723457 RB6144 38 117 hypothetical protein 3228305 3228189 11.1 4314 - 6.17 2.94 0.42 14.163471 RB6167 375 1128 Putative transposase 3235937 3237064 9.9 41895 + 5.92 1.74 0.16 9.003472 RB6168 208 627 hypothetical protein 3236689 3237315 12.2 22575 + 6.05 2.17 0.31 14.323482 RB6188 68 207 hypothetical protein 3244384 3244178 9.6 6986 - 6.17 1.73 0.30 17.513531 RB6282 200 603 conserved hypothetical protein 3302795 3303397 4.8 22451 + 6.19 2.22 0.42 19.023588 RB6379 153 462 hypothetical protein 3353122 3352661 5.8 17398 - 6.75 1.50 0.11 7.163739 RB6627 176 531 hypothetical protein 3522169 3522699 6 19706 + 6.14 1.63 0.26 15.983741 RB6629 708 2127 secreted protein 3522859 3524985 5.1 78383 + 5.59 1.69 0.18 10.493745 RB6636 433 1302 ISXo8 transposase 3527005 3528306 9.4 49646 + 5.60 1.65 0.29 17.803746 RB6637 50 153 hypothetical protein 3528317 3528469 10.1 5841 + 5.45 1.63 0.20 12.233771 RB6685 72 219 hypothetical protein 3554265 3554047 12.2 8077 - 5.85 1.84 0.23 12.453784 RB6706 37 114 hypothetical protein 3560942 3560829 7.3 4375 - 5.79 1.94 0.14 6.993804 RB6736 433 1302 ISXo8 transposase 3578852 3577551 9.4 49646 - 5.92 2.13 0.34 16.103888 RB6888 420 1263 secreted protein containing DUF1573 3662449 3661187 4.3 45773 - 6.09 2.92 0.49 16.853969 RB7039 202 609 hypothetical protein 3743573 3742965 7.9 23105 - 5.43 1.87 0.37 20.043981 RB7063 148 447 hypothetical protein 3751250 3751696 10.5 17004 + 6.38 2.98 0.19 6.254157 RB7389 375 1128 Putative transposase 3953590 3954717 9.9 41895 + 5.70 2.58 0.35 13.594161 RB7394 136 411 hypothetical protein 3955937 3955527 4.2 14568 - 6.19 1.52 0.18 11.974291 RB7612 136 411 hypothetical protein 4092907 4093317 11.4 15607 + 5.91 2.82 0.16 5.50
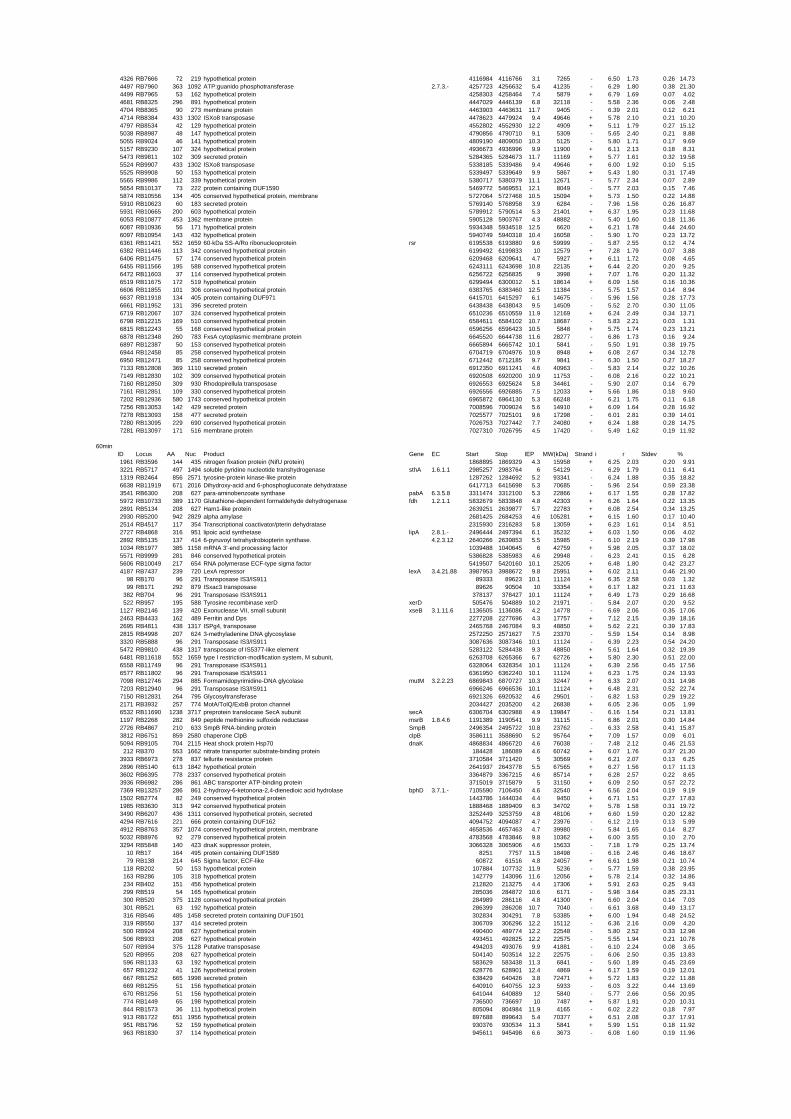
4326 RB7666 72 219 hypothetical protein 4116984 4116766 3.1 7265 - 6.50 1.73 0.26 14.734497 RB7960 363 1092 ATP:guanido phosphotransferase 2.7.3.- 4257723 4256632 5.4 41235 - 6.29 1.80 0.38 21.304499 RB7965 53 162 hypothetical protein 4258303 4258464 7.4 5879 + 6.79 1.69 0.07 4.024681 RB8325 296 891 hypothetical protein 4447029 4446139 6.8 32118 - 5.58 2.36 0.06 2.484704 RB8365 90 273 membrane protein 4463903 4463631 11.7 9405 - 6.39 2.01 0.12 6.214714 RB8384 433 1302 ISXo8 transposase 4478623 4479924 9.4 49646 + 5.78 2.10 0.21 10.204797 RB8534 42 129 hypothetical protein 4552802 4552930 12.2 4909 + 5.11 1.79 0.27 15.125038 RB8987 48 147 hypothetical protein 4790856 4790710 9.1 5309 - 5.65 2.40 0.21 8.885055 RB9024 46 141 hypothetical protein 4809190 4809050 10.3 5125 - 5.80 1.71 0.17 9.695157 RB9230 107 324 hypothetical protein 4936673 4936996 9.9 11900 + 6.11 2.13 0.18 8.315473 RB9811 102 309 secreted protein 5284365 5284673 11.7 11169 + 5.77 1.61 0.32 19.585524 RB9907 433 1302 ISXo8 transposase 5338185 5339486 9.4 49646 + 6.00 1.92 0.10 5.155525 RB9908 50 153 hypothetical protein 5339497 5339649 9.9 5867 + 5.43 1.80 0.31 17.495565 RB9986 112 339 hypothetical protein 5380717 5380379 11.1 12671 - 5.77 2.34 0.07 2.895654 RB10137 73 222 protein containing DUF1590 5469772 5469551 12.1 8049 - 5.77 2.03 0.15 7.465874 RB10556 134 405 conserved hypothetical protein, membrane 5727064 5727468 10.5 15094 + 5.73 1.50 0.22 14.885910 RB10623 60 183 secreted protein 5769140 5768958 3.9 6284 - 7.96 1.56 0.26 16.875931 RB10665 200 603 hypothetical protein 5789912 5790514 5.3 21401 + 6.37 1.95 0.23 11.686053 RB10877 453 1362 membrane protein 5905128 5903767 4.3 48882 - 5.40 1.60 0.18 11.366087 RB10936 56 171 hypothetical protein 5934348 5934518 12.5 6620 + 6.21 1.78 0.44 24.606097 RB10954 143 432 hypothetical protein 5940749 5940318 10.4 16058 - 5.90 1.70 0.23 13.726361 RB11421 552 1659 60-kDa SS-A/Ro ribonucleoprotein rsr 6195538 6193880 9.6 59999 - 5.87 2.55 0.12 4.746382 RB11446 113 342 conserved hypothetical protein 6199492 6199833 10 12579 + 7.28 1.79 0.07 3.886406 RB11475 57 174 conserved hypothetical protein 6209468 6209641 4.7 5927 + 6.11 1.72 0.08 4.656455 RB11566 195 588 conserved hypothetical protein 6243111 6243698 10.8 22135 + 6.44 2.20 0.20 9.256472 RB11603 37 114 conserved hypothetical protein 6256722 6256835 9 3998 + 7.07 1.76 0.20 11.326519 RB11675 172 519 hypothetical protein 6299494 6300012 5.1 18614 + 6.09 1.56 0.16 10.366606 RB11855 101 306 conserved hypothetical protein 6383765 6383460 12.5 11384 - 5.75 1.57 0.14 8.946637 RB11918 134 405 protein containing DUF971 6415701 6415297 6.1 14675 - 5.96 1.56 0.28 17.736661 RB11952 131 396 secreted protein 6438438 6438043 9.5 14509 - 5.52 2.70 0.30 11.056719 RB12067 107 324 conserved hypothetical protein 6510236 6510559 11.9 12169 + 6.24 2.49 0.34 13.716798 RB12215 169 510 conserved hypothetical protein 6584611 6584102 10.7 18687 - 5.83 2.21 0.03 1.316815 RB12243 55 168 conserved hypothetical protein 6596256 6596423 10.5 5848 + 5.75 1.74 0.23 13.216878 RB12348 260 783 FxsA cytoplasmic membrane protein 6645520 6644738 11.6 28277 - 6.86 1.73 0.16 9.246897 RB12387 50 153 conserved hypothetical protein 6665894 6665742 10.1 5841 - 5.50 1.91 0.38 19.756944 RB12458 85 258 conserved hypothetical protein 6704719 6704976 10.9 8948 + 6.08 2.67 0.34 12.786950 RB12471 85 258 conserved hypothetical protein 6712442 6712185 9.7 9841 - 6.30 1.50 0.27 18.277133 RB12808 369 1110 secreted protein 6912350 6911241 4.6 40963 - 5.83 2.14 0.22 10.267149 RB12830 102 309 conserved hypothetical protein 6920508 6920200 10.9 11753 - 6.08 2.16 0.22 10.217160 RB12850 309 930 Rhodopirellula transposase 6926553 6925624 5.8 34461 - 5.90 2.07 0.14 6.797161 RB12851 109 330 conserved hypothetical protein 6926556 6926885 7.5 12033 + 5.66 1.86 0.18 9.607202 RB12936 580 1743 conserved hypothetical protein 6965872 6964130 5.3 66248 - 6.21 1.75 0.11 6.187256 RB13053 142 429 secreted protein 7008596 7009024 5.6 14910 + 6.09 1.64 0.28 16.927278 RB13093 158 477 secreted protein 7025577 7025101 9.6 17298 - 6.01 2.81 0.39 14.017280 RB13095 229 690 conserved hypothetical protein 7026753 7027442 7.7 24080 + 6.24 1.88 0.28 14.757281 RB13097 171 516 membrane protein 7027310 7026795 4.5 17420 - 5.49 1.62 0.19 11.92
60minID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand i r Stdev %1961 RB3596 144 435 nitrogen fixation protein (NifU protein) 1868895 1869329 4.3 15958 + 6.25 2.03 0.20 9.913221 RB5717 497 1494 soluble pyridine nucleotide transhydrogenase sthA 1.6.1.1 2985257 2983764 6 54129 - 6.29 1.79 0.11 6.411319 RB2464 856 2571 tyrosine-protein kinase-like protein 1287262 1284692 5.2 93341 - 6.24 1.88 0.35 18.826638 RB11919 671 2016 Dihydroxy-acid and 6-phosphogluconate dehydratase 6417713 6415698 5.3 70685 - 5.96 2.54 0.59 23.383541 RB6300 208 627 para-aminobenzoate synthase pabA 6.3.5.8 3311474 3312100 5.3 22866 + 6.17 1.55 0.28 17.825972 RB10733 389 1170 Glutathione-dependent formaldehyde dehydrogenase fdh 1.2.1.1 5832679 5833848 4.8 42303 + 6.26 1.64 0.22 13.352891 RB5134 208 627 Ham1-like protein 2639251 2639877 5.7 22783 + 6.08 2.54 0.34 13.252930 RB5200 942 2829 alpha amylase 2681425 2684253 4.6 105281 + 6.15 1.60 0.17 10.402514 RB4517 117 354 Transcriptional coactivator/pterin dehydratase 2315930 2316283 5.8 13059 + 6.23 1.61 0.14 8.512727 RB4868 316 951 lipoic acid synthetase lipA 2.8.1.- 2496444 2497394 6.1 35232 + 6.03 1.50 0.06 4.022892 RB5135 137 414 6-pyruvoyl tetrahydrobiopterin synthase. 4.2.3.12 2640266 2639853 5.5 15985 - 6.10 2.19 0.39 17.981034 RB1977 385 1158 mRNA 3'-end processing factor 1039488 1040645 6 42759 + 5.98 2.05 0.37 18.025571 RB9999 281 846 conserved hypothetical protein 5386828 5385983 4.6 29948 - 6.23 2.41 0.15 6.285606 RB10049 217 654 RNA polymerase ECF-type sigma factor 5419507 5420160 10.1 25205 + 6.48 1.80 0.42 23.274187 RB7437 239 720 LexA repressor lexA 3.4.21.88 3987953 3988672 9.8 25951 + 6.02 2.11 0.46 21.90
98 RB170 96 291 Transposase IS3/IS911 89333 89623 10.1 11124 + 6.35 2.58 0.03 1.3299 RB171 292 879 ISxac3 transposase 89626 90504 10 33354 + 6.17 1.82 0.21 11.63
382 RB704 96 291 Transposase IS3/IS911 378137 378427 10.1 11124 + 6.49 1.73 0.29 16.68522 RB957 195 588 Tyrosine recombinase xerD xerD 505476 504889 10.2 21971 - 5.84 2.07 0.20 9.52
1127 RB2146 139 420 Exonuclease VII, small subunit xseB 3.1.11.6 1136505 1136086 4.2 14778 - 6.69 2.06 0.35 17.062463 RB4433 162 489 Ferritin and Dps 2277208 2277696 4.3 17757 + 7.12 2.15 0.39 18.162695 RB4811 438 1317 ISPg4, transposase 2465768 2467084 9.3 48850 + 5.62 2.21 0.39 17.832815 RB4998 207 624 3-methyladenine DNA glycosylase 2572250 2571627 7.5 23370 - 5.59 1.54 0.14 8.983320 RB5888 96 291 Transposase IS3/IS911 3087636 3087346 10.1 11124 - 6.39 2.23 0.54 24.205472 RB9810 438 1317 transposase of IS5377-like element 5283122 5284438 9.3 48850 + 5.61 1.64 0.32 19.396481 RB11618 552 1659 type I restriction-modification system, M subunit, 6263708 6265366 6.7 62726 + 5.80 2.30 0.51 22.006558 RB11749 96 291 Transposase IS3/IS911 6328064 6328354 10.1 11124 + 6.39 2.56 0.45 17.566577 RB11802 96 291 Transposase IS3/IS911 6361950 6362240 10.1 11124 + 6.23 1.75 0.24 13.937098 RB12746 294 885 Formamidopyrimidine-DNA glycolase mutM 3.2.2.23 6869843 6870727 10.3 32447 + 6.33 2.07 0.31 14.987203 RB12940 96 291 Transposase IS3/IS911 6966246 6966536 10.1 11124 + 6.48 2.31 0.52 22.747150 RB12831 264 795 Glycosyltransferase 6921326 6920532 4.6 29501 - 6.82 1.53 0.29 19.222171 RB3932 257 774 MotA/TolQ/ExbB proton channel 2034427 2035200 4.2 26838 + 6.05 2.36 0.05 1.996532 RB11690 1238 3717 preprotein translocase SecA subunit secA 6306704 6302988 4.9 139847 - 6.16 1.54 0.21 13.811197 RB2268 282 849 peptide methionine sulfoxide reductase msrB 1.8.4.6 1191389 1190541 9.9 31115 - 6.86 2.01 0.30 14.842726 RB4867 210 633 SmpB RNA-binding protein SmpB 2496354 2495722 10.8 23762 - 6.33 2.58 0.41 15.873812 RB6751 859 2580 chaperone ClpB clpB 3586111 3588690 5.2 95764 + 7.09 1.57 0.09 6.015094 RB9105 704 2115 Heat shock protein Hsp70 dnaK 4868834 4866720 4.6 76038 - 7.48 2.12 0.46 21.53212 RB370 553 1662 nitrate transporter substrate-binding protein 184428 186089 4.6 60742 + 6.07 1.76 0.37 21.30
3933 RB6973 278 837 tellurite resistance protein 3710584 3711420 5 30569 + 6.21 2.07 0.13 6.252896 RB5140 613 1842 hypothetical protein 2641937 2643778 5.5 67565 + 6.27 1.56 0.17 11.133602 RB6395 778 2337 conserved hypothetical protein 3364879 3367215 4.6 85714 + 6.28 2.57 0.22 8.653936 RB6982 286 861 ABC transporter ATP-binding protein 3715019 3715879 5 31150 + 6.09 2.50 0.57 22.727369 RB13257 286 861 2-hydroxy-6-ketonona-2,4-dienedioic acid hydrolase bphD 3.7.1.- 7105590 7106450 4.6 32540 + 6.56 2.04 0.19 9.191502 RB2774 82 249 conserved hypothetical protein 1443786 1444034 4.4 9450 + 6.71 1.51 0.27 17.831985 RB3630 313 942 conserved hypothetical protein 1888468 1889409 6.3 34702 + 5.78 1.58 0.31 19.723490 RB6207 436 1311 conserved hypothetical protein, secreted 3252449 3253759 4.8 48106 + 6.60 1.59 0.20 12.824294 RB7616 221 666 protein containing DUF162 4094752 4094087 4.7 23976 - 6.12 2.19 0.13 5.994912 RB8763 357 1074 conserved hypothetical protein, membrane 4658536 4657463 4.7 39980 - 5.84 1.65 0.14 8.275032 RB8976 92 279 conserved hypothetical protein 4783568 4783846 9.8 10362 + 6.00 3.55 0.10 2.703294 RB5848 140 423 dnaK suppressor protein, 3066328 3065906 4.6 15633 - 7.18 1.79 0.25 13.74
10 RB17 164 495 protein containing DUF1589 8251 7757 11.5 18498 - 6.16 2.46 0.46 18.6779 RB138 214 645 Sigma factor, ECF-like 60872 61516 4.8 24057 + 6.61 1.98 0.21 10.74
118 RB202 50 153 hypothetical protein 107884 107732 11.9 5236 - 5.77 1.59 0.38 23.95163 RB286 105 318 hypothetical protein 142779 143096 11.6 12056 + 5.78 2.14 0.32 14.86234 RB402 151 456 hypothetical protein 212820 213275 4.4 17306 + 5.91 2.63 0.25 9.43299 RB519 54 165 hypothetical protein 285036 284872 10.6 6171 - 5.98 3.64 0.85 23.31300 RB520 375 1128 conserved hypothetical protein 284989 286116 4.8 41300 + 6.60 2.04 0.14 7.03301 RB521 63 192 hypothetical protein 286399 286208 10.7 7040 - 6.61 3.68 0.49 13.17316 RB546 485 1458 secreted protein containing DUF1501 302834 304291 7.8 53385 + 6.00 1.94 0.48 24.52319 RB550 137 414 secreted protein 306709 306296 12.2 15112 - 6.36 2.16 0.09 4.20500 RB924 208 627 hypothetical protein 490400 489774 12.2 22548 - 5.80 2.52 0.33 12.98506 RB933 208 627 hypothetical protein 493451 492825 12.2 22575 - 5.55 1.94 0.21 10.78507 RB934 375 1128 Putative transposase 494203 493076 9.9 41881 - 6.10 2.24 0.08 3.65520 RB955 208 627 hypothetical protein 504140 503514 12.2 22575 - 6.06 2.50 0.35 13.83596 RB1133 63 192 hypothetical protein 583629 583438 11.3 6841 - 5.60 1.89 0.45 23.69657 RB1232 41 126 hypothetical protein 628776 628901 12.4 4869 + 6.17 1.59 0.19 12.01667 RB1252 665 1998 secreted protein 638429 640426 3.8 72471 + 5.72 1.83 0.22 11.88669 RB1255 51 156 hypothetical protein 640910 640755 12.3 5933 - 6.03 3.22 0.44 13.69670 RB1256 51 156 hypothetical protein 641044 640889 12 5840 - 5.77 2.66 0.56 20.95774 RB1449 65 198 hypothetical protein 736500 736697 10 7487 + 5.87 1.91 0.20 10.31844 RB1573 36 111 hypothetical protein 805094 804984 11.9 4165 - 6.02 2.22 0.18 7.97913 RB1722 651 1956 hypothetical protein 897688 899643 5.4 70377 + 6.51 2.08 0.37 17.91951 RB1796 52 159 hypothetical protein 930376 930534 11.3 5841 + 5.99 1.51 0.18 11.92963 RB1830 37 114 hypothetical protein 945611 945498 6.6 3673 - 6.08 1.60 0.19 11.96
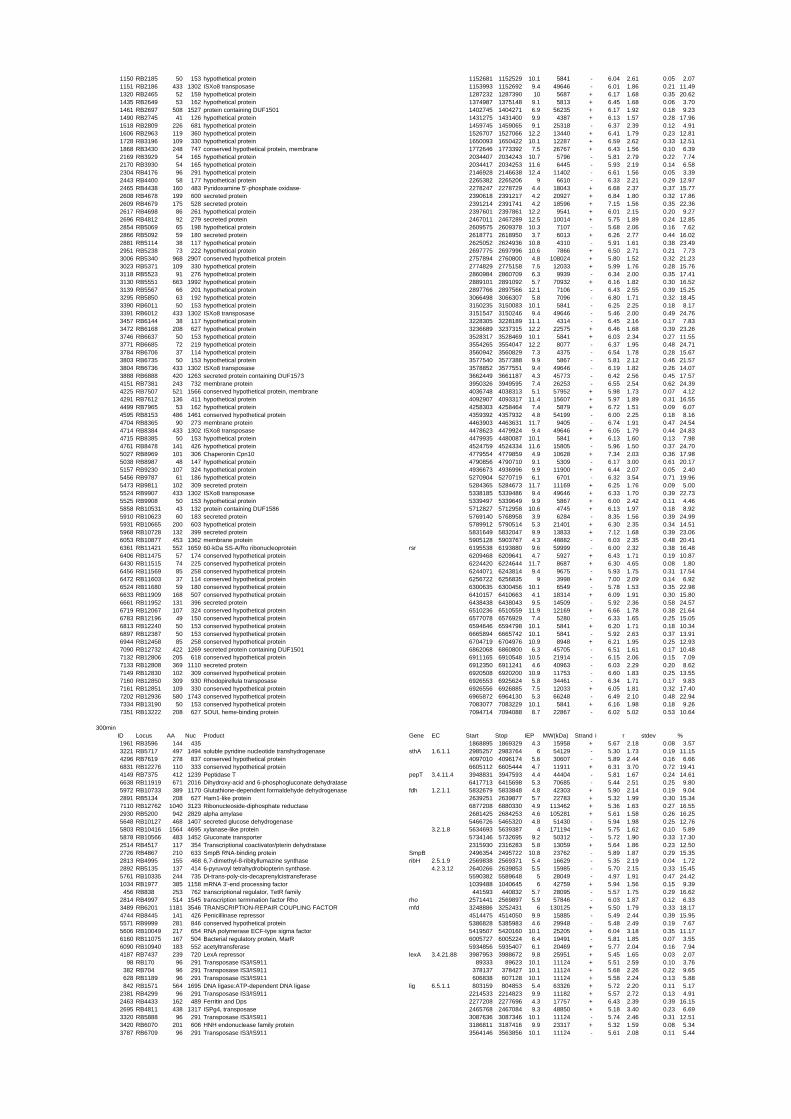
1150 RB2185 50 153 hypothetical protein 1152681 1152529 10.1 5841 - 6.04 2.61 0.05 2.071151 RB2186 433 1302 ISXo8 transposase 1153993 1152692 9.4 49646 - 6.01 1.86 0.21 11.491320 RB2465 52 159 hypothetical protein 1287232 1287390 10 5687 + 6.17 1.68 0.35 20.621435 RB2649 53 162 hypothetical protein 1374987 1375148 9.1 5813 + 6.45 1.68 0.06 3.701461 RB2697 508 1527 protein containing DUF1501 1402745 1404271 6.9 56235 + 6.17 1.92 0.18 9.231490 RB2745 41 126 hypothetical protein 1431275 1431400 9.9 4387 + 6.13 1.57 0.28 17.961518 RB2809 226 681 hypothetical protein 1459745 1459065 9.1 25318 - 6.37 2.39 0.12 4.911606 RB2963 119 360 hypothetical protein 1526707 1527066 12.2 13440 + 6.41 1.79 0.23 12.811728 RB3196 109 330 hypothetical protein 1650093 1650422 10.1 12287 + 6.59 2.62 0.33 12.511868 RB3430 248 747 conserved hypothetical protein, membrane 1772646 1773392 7.5 26767 + 6.43 1.56 0.10 6.392169 RB3929 54 165 hypothetical protein 2034407 2034243 10.7 5796 - 5.81 2.79 0.22 7.742170 RB3930 54 165 hypothetical protein 2034417 2034253 11.6 6445 - 5.93 2.19 0.14 6.582304 RB4176 96 291 hypothetical protein 2146928 2146638 12.4 11402 - 6.61 1.56 0.05 3.392443 RB4400 58 177 hypothetical protein 2265382 2265206 9 6610 - 6.33 2.21 0.29 12.972465 RB4438 160 483 Pyridoxamine 5'-phosphate oxidase- 2278247 2278729 4.4 18043 + 6.68 2.37 0.37 15.772608 RB4678 199 600 secreted protein 2390618 2391217 4.2 20927 + 6.84 1.80 0.32 17.862609 RB4679 175 528 secreted protein 2391214 2391741 4.2 18596 + 7.15 1.56 0.35 22.362617 RB4698 86 261 hypothetical protein 2397601 2397861 12.2 9541 + 6.01 2.15 0.20 9.272696 RB4812 92 279 secreted protein 2467011 2467289 12.5 10014 + 5.75 1.89 0.24 12.852854 RB5069 65 198 hypothetical protein 2609575 2609378 10.3 7107 - 5.68 2.06 0.16 7.622866 RB5092 59 180 secreted protein 2618771 2618950 3.7 6013 + 6.26 2.77 0.44 16.022881 RB5114 38 117 hypothetical protein 2625052 2624936 10.8 4310 - 5.91 1.61 0.38 23.492951 RB5238 73 222 hypothetical protein 2697775 2697996 10.6 7866 + 6.50 2.71 0.21 7.733006 RB5340 968 2907 conserved hypothetical protein 2757894 2760800 4.8 108024 + 5.80 1.52 0.32 21.233023 RB5371 109 330 hypothetical protein 2774829 2775158 7.5 12033 + 5.99 1.76 0.28 15.763118 RB5523 91 276 hypothetical protein 2860984 2860709 6.3 9939 - 6.34 2.00 0.35 17.413130 RB5551 663 1992 hypothetical protein 2889101 2891092 5.7 70932 + 6.16 1.82 0.30 16.523139 RB5567 66 201 hypothetical protein 2897766 2897566 12.1 7106 - 6.43 2.55 0.39 15.253295 RB5850 63 192 hypothetical protein 3066498 3066307 5.8 7096 - 6.80 1.71 0.32 18.453390 RB6011 50 153 hypothetical protein 3150235 3150083 10.1 5841 - 6.25 2.25 0.18 8.173391 RB6012 433 1302 ISXo8 transposase 3151547 3150246 9.4 49646 - 5.46 2.00 0.49 24.763457 RB6144 38 117 hypothetical protein 3228305 3228189 11.1 4314 - 6.45 2.16 0.17 7.833472 RB6168 208 627 hypothetical protein 3236689 3237315 12.2 22575 + 6.46 1.68 0.39 23.263746 RB6637 50 153 hypothetical protein 3528317 3528469 10.1 5841 + 6.03 2.34 0.27 11.553771 RB6685 72 219 hypothetical protein 3554265 3554047 12.2 8077 - 6.37 1.95 0.48 24.713784 RB6706 37 114 hypothetical protein 3560942 3560829 7.3 4375 - 6.54 1.78 0.28 15.673803 RB6735 50 153 hypothetical protein 3577540 3577388 9.9 5867 - 5.81 2.12 0.46 21.573804 RB6736 433 1302 ISXo8 transposase 3578852 3577551 9.4 49646 - 6.19 1.82 0.26 14.073888 RB6888 420 1263 secreted protein containing DUF1573 3662449 3661187 4.3 45773 - 6.42 2.56 0.45 17.574151 RB7381 243 732 membrane protein 3950326 3949595 7.4 26253 - 6.55 2.54 0.62 24.394225 RB7507 521 1566 conserved hypothetical protein, membrane 4036748 4038313 5.1 57952 + 5.98 1.73 0.07 4.124291 RB7612 136 411 hypothetical protein 4092907 4093317 11.4 15607 + 5.97 1.89 0.31 16.554499 RB7965 53 162 hypothetical protein 4258303 4258464 7.4 5879 + 6.72 1.51 0.09 6.074595 RB8153 486 1461 conserved hypothetical protein 4359392 4357932 4.8 54199 - 6.00 2.25 0.18 8.164704 RB8365 90 273 membrane protein 4463903 4463631 11.7 9405 - 6.74 1.91 0.47 24.544714 RB8384 433 1302 ISXo8 transposase 4478623 4479924 9.4 49646 + 6.05 1.79 0.44 24.834715 RB8385 50 153 hypothetical protein 4479935 4480087 10.1 5841 + 6.13 1.60 0.13 7.984761 RB8478 141 426 hypothetical protein 4524759 4524334 11.6 15805 - 5.96 1.50 0.37 24.705027 RB8969 101 306 Chaperonin Cpn10 4779554 4779859 4.9 10628 + 7.34 2.03 0.36 17.985038 RB8987 48 147 hypothetical protein 4790856 4790710 9.1 5309 - 6.17 3.00 0.61 20.175157 RB9230 107 324 hypothetical protein 4936673 4936996 9.9 11900 + 6.44 2.07 0.05 2.405456 RB9787 61 186 hypothetical protein 5270904 5270719 6.1 6701 - 6.32 3.54 0.71 19.965473 RB9811 102 309 secreted protein 5284365 5284673 11.7 11169 + 6.25 1.76 0.09 5.005524 RB9907 433 1302 ISXo8 transposase 5338185 5339486 9.4 49646 + 6.33 1.70 0.39 22.735525 RB9908 50 153 hypothetical protein 5339497 5339649 9.9 5867 + 6.00 2.42 0.11 4.465858 RB10531 43 132 protein containing DUF1586 5712827 5712958 10.6 4745 + 6.13 1.97 0.18 8.925910 RB10623 60 183 secreted protein 5769140 5768958 3.9 6284 - 8.35 1.56 0.39 24.995931 RB10665 200 603 hypothetical protein 5789912 5790514 5.3 21401 + 6.30 2.35 0.34 14.515968 RB10728 132 399 secreted protein 5831649 5832047 9.9 13833 + 7.12 1.68 0.39 23.066053 RB10877 453 1362 membrane protein 5905128 5903767 4.3 48882 - 6.03 2.35 0.48 20.416361 RB11421 552 1659 60-kDa SS-A/Ro ribonucleoprotein rsr 6195538 6193880 9.6 59999 - 6.00 2.32 0.38 16.486406 RB11475 57 174 conserved hypothetical protein 6209468 6209641 4.7 5927 + 6.43 1.71 0.19 10.876430 RB11515 74 225 conserved hypothetical protein 6224420 6224644 11.7 8687 + 6.30 4.65 0.08 1.806456 RB11569 85 258 conserved hypothetical protein 6244071 6243814 9.4 9675 - 5.93 1.75 0.31 17.546472 RB11603 37 114 conserved hypothetical protein 6256722 6256835 9 3998 + 7.00 2.09 0.14 6.926524 RB11680 59 180 conserved hypothetical protein 6300635 6300456 10.1 6549 - 5.78 1.53 0.35 22.986633 RB11909 168 507 conserved hypothetical protein 6410157 6410663 4.1 18314 + 6.09 1.91 0.30 15.806661 RB11952 131 396 secreted protein 6438438 6438043 9.5 14509 - 5.92 2.36 0.58 24.576719 RB12067 107 324 conserved hypothetical protein 6510236 6510559 11.9 12169 + 6.66 1.78 0.38 21.646783 RB12196 49 150 conserved hypothetical protein 6577078 6576929 7.4 5280 - 6.33 1.65 0.25 15.056813 RB12240 50 153 conserved hypothetical protein 6594646 6594798 10.1 5841 + 6.20 1.71 0.18 10.346897 RB12387 50 153 conserved hypothetical protein 6665894 6665742 10.1 5841 - 5.92 2.63 0.37 13.916944 RB12458 85 258 conserved hypothetical protein 6704719 6704976 10.9 8948 + 6.21 1.95 0.25 12.937090 RB12732 422 1269 secreted protein containing DUF1501 6862068 6860800 6.3 45705 - 6.51 1.61 0.17 10.487132 RB12806 205 618 conserved hypothetical protein 6911165 6910548 10.5 21914 - 6.15 2.06 0.15 7.097133 RB12808 369 1110 secreted protein 6912350 6911241 4.6 40963 - 6.03 2.29 0.20 8.627149 RB12830 102 309 conserved hypothetical protein 6920508 6920200 10.9 11753 - 6.60 1.83 0.25 13.557160 RB12850 309 930 Rhodopirellula transposase 6926553 6925624 5.8 34461 - 6.34 1.71 0.17 9.837161 RB12851 109 330 conserved hypothetical protein 6926556 6926885 7.5 12033 + 6.05 1.81 0.32 17.407202 RB12936 580 1743 conserved hypothetical protein 6965872 6964130 5.3 66248 - 6.49 2.10 0.48 22.947334 RB13190 50 153 conserved hypothetical protein 7083077 7083229 10.1 5841 + 6.16 1.98 0.18 9.267351 RB13222 208 627 SOUL heme-binding protein 7094714 7094088 8.7 22867 - 6.02 5.02 0.53 10.64
300minID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand i r stdev %1961 RB3596 144 435 1868895 1869329 4.3 15958 + 5.67 2.18 0.08 3.573221 RB5717 497 1494 soluble pyridine nucleotide transhydrogenase sthA 1.6.1.1 2985257 2983764 6 54129 - 5.30 1.73 0.19 11.154296 RB7619 278 837 conserved hypothetical protein 4097010 4096174 5.6 30607 - 5.89 2.44 0.16 6.666831 RB12276 110 333 conserved hypothetical protein 6605112 6605444 4.7 11911 + 6.31 3.70 0.72 19.414149 RB7375 412 1239 Peptidase T pepT 3.4.11.4 3948831 3947593 4.4 44404 - 5.81 1.67 0.24 14.616638 RB11919 671 2016 Dihydroxy-acid and 6-phosphogluconate dehydratase 6417713 6415698 5.3 70685 - 5.44 2.51 0.25 9.805972 RB10733 389 1170 Glutathione-dependent formaldehyde dehydrogenase fdh 1.2.1.1 5832679 5833848 4.8 42303 + 5.90 2.14 0.19 9.042891 RB5134 208 627 Ham1-like protein 2639251 2639877 5.7 22783 + 5.32 1.99 0.30 15.347110 RB12762 1040 3123 Ribonucleoside-diphosphate reductase 6877208 6880330 4.9 113462 + 5.36 1.63 0.27 16.552930 RB5200 942 2829 alpha amylase 2681425 2684253 4.6 105281 + 5.61 1.58 0.26 16.255648 RB10127 468 1407 secreted glucose dehydrogenase 5466726 5465320 4.8 51430 - 5.94 1.98 0.25 12.765803 RB10416 1564 4695 xylanase-like protein 3.2.1.8 5634693 5639387 4 171194 + 5.75 1.62 0.10 5.895878 RB10566 483 1452 Gluconate transporter 5734146 5732695 9.2 50312 - 5.72 1.90 0.33 17.302514 RB4517 117 354 Transcriptional coactivator/pterin dehydratase 2315930 2316283 5.8 13059 + 5.64 1.86 0.23 12.502726 RB4867 210 633 SmpB RNA-binding protein SmpB 2496354 2495722 10.8 23762 - 5.89 1.87 0.29 15.352813 RB4995 155 468 6,7-dimethyl-8-ribityllumazine synthase ribH 2.5.1.9 2569838 2569371 5.4 16629 - 5.35 2.19 0.04 1.722892 RB5135 137 414 6-pyruvoyl tetrahydrobiopterin synthase. 4.2.3.12 2640266 2639853 5.5 15985 - 5.70 2.15 0.33 15.455761 RB10335 244 735 Di-trans-poly-cis-decaprenylcistransferase 5590382 5589648 5 28049 - 4.97 1.91 0.47 24.421034 RB1977 385 1158 mRNA 3'-end processing factor 1039488 1040645 6 42759 + 5.94 1.56 0.15 9.39456 RB838 253 762 transcriptional regulator, TetR family 441593 440832 5.7 28095 - 5.57 1.75 0.29 16.62
2814 RB4997 514 1545 transcription termination factor Rho rho 2571441 2569897 5.9 57846 - 6.03 1.87 0.12 6.333489 RB6201 1181 3546 TRANSCRIPTION-REPAIR COUPLING FACTOR mfd 3248886 3252431 6 130125 + 5.50 1.79 0.33 18.174744 RB8445 141 426 Penicillinase repressor 4514475 4514050 9.9 15885 - 5.49 2.44 0.39 15.955571 RB9999 281 846 conserved hypothetical protein 5386828 5385983 4.6 29948 - 5.48 2.49 0.19 7.675606 RB10049 217 654 RNA polymerase ECF-type sigma factor 5419507 5420160 10.1 25205 + 6.04 3.18 0.35 11.176160 RB11075 167 504 Bacterial regulatory protein, MarR 6005727 6005224 6.4 19491 - 5.81 1.85 0.07 3.556090 RB10940 183 552 acetyltransferase 5934856 5935407 6.1 20469 + 5.77 2.04 0.16 7.944187 RB7437 239 720 LexA repressor lexA 3.4.21.88 3987953 3988672 9.8 25951 + 5.45 1.65 0.03 2.07
98 RB170 96 291 Transposase IS3/IS911 89333 89623 10.1 11124 + 5.51 2.59 0.10 3.76382 RB704 96 291 Transposase IS3/IS911 378137 378427 10.1 11124 + 5.68 2.26 0.22 9.65628 RB1189 96 291 Transposase IS3/IS911 606838 607128 10.1 11124 + 5.58 2.24 0.13 5.88842 RB1571 564 1695 DNA ligase:ATP-dependent DNA ligase lig 6.5.1.1 803159 804853 5.4 63326 + 5.72 2.20 0.11 5.17
2381 RB4299 96 291 Transposase IS3/IS911 2214533 2214823 9.9 11182 + 5.57 2.72 0.13 4.912463 RB4433 162 489 Ferritin and Dps 2277208 2277696 4.3 17757 + 6.43 2.39 0.39 16.152695 RB4811 438 1317 ISPg4, transposase 2465768 2467084 9.3 48850 + 5.18 3.40 0.23 6.693320 RB5888 96 291 Transposase IS3/IS911 3087636 3087346 10.1 11124 - 5.74 2.46 0.31 12.513420 RB6070 201 606 HNH endonuclease family protein 3186811 3187416 9.9 23317 + 5.32 1.59 0.08 5.343787 RB6709 96 291 Transposase IS3/IS911 3564146 3563856 10.1 11124 - 5.61 2.08 0.11 5.44

4555 RB8076 173 522 Holliday junction resolvase YqgF 3.1.-.- 4323454 4323975 5.1 18918 + 5.70 1.96 0.18 9.114721 RB8397 708 2127 UvrABC system protein B uvrB 4489596 4487470 6.1 80350 - 5.69 2.01 0.33 16.306481 RB11618 552 1659 type I restriction-modification system, M subunit, 6263708 6265366 6.7 62726 + 5.11 2.42 0.03 1.346558 RB11749 96 291 Transposase IS3/IS911 6328064 6328354 10.1 11124 + 5.66 2.61 0.18 6.926577 RB11802 96 291 Transposase IS3/IS911 6361950 6362240 10.1 11124 + 5.37 2.02 0.27 13.437098 RB12746 294 885 Formamidopyrimidine-DNA glycolase mutM 3.2.2.23 6869843 6870727 10.3 32447 + 5.66 3.87 0.26 6.817203 RB12940 96 291 Transposase IS3/IS911 6966246 6966536 10.1 11124 + 5.45 2.50 0.26 10.24944 RB1784 477 1434 UDP-glucose/GDP-mannose dehydrogenase 924070 925503 4.7 52259 + 5.57 1.72 0.08 4.72
3620 RB6428 164 495 conserved hypothetical protein, secreted 3382816 3382322 6.5 17292 - 7.40 2.37 0.06 2.596653 RB11941 427 1284 Glycosyl transferase, family 2 6429975 6428692 7.8 47063 - 5.65 2.30 0.32 13.687150 RB12831 264 795 Glycosyltransferase 6921326 6920532 4.6 29501 - 6.14 2.55 0.56 21.746425 RB11505 199 600 conserved hypothetical protein, secreted 6221829 6221230 7.5 21660 - 6.31 1.99 0.36 18.076532 RB11690 1238 3717 preprotein translocase SecA subunit secA 6306704 6302988 4.9 139847 - 5.36 1.84 0.23 12.33731 RB1359 629 1890 Protease do precursor degP 3.4.21.- 693488 695377 6.1 66729 + 6.41 3.06 0.11 3.74
1197 RB2268 282 849 peptide methionine sulfoxide reductase msrB 1.8.4.6 1191389 1190541 9.9 31115 - 6.32 1.59 0.25 15.783138 RB5566 728 2187 cell division protein FtsH ftsH 3.4.24.- 2897569 2895383 5 79839 - 5.68 1.99 0.34 17.093812 RB6751 859 2580 chaperone ClpB clpB 3586111 3588690 5.2 95764 + 6.51 2.12 0.45 21.354636 RB8238 198 597 alkyl hydroperoxide reducatase 4403997 4404593 4.5 22069 + 5.35 1.87 0.11 6.114837 RB8604 1227 3684 Cytochrome c assembly protein 4590203 4593886 5.2 134894 + 5.79 1.97 0.18 8.945026 RB8966 660 1983 Chaperonin Cpn60/TCP-1 4777493 4779475 5.3 69819 + 6.77 2.08 0.04 2.125028 RB8970 540 1623 GroEL 4779905 4781527 4.8 57516 + 6.76 2.31 0.47 20.335030 RB8974 200 603 GrpE protein grpE 4782869 4783471 4.2 21927 + 6.55 2.49 0.23 9.395092 RB9103 899 2700 ATP-dependent Clp protease, ATP-binding subunit ClpB 4866549 4863850 4.8 99439 - 6.44 2.64 0.28 10.575094 RB9105 704 2115 Heat shock protein Hsp70 dnaK 4868834 4866720 4.6 76038 - 6.84 3.28 0.09 2.795713 RB10242 330 993 Curved DNA-binding protein cbpA 5536118 5535126 6.7 35177 - 6.16 2.29 0.39 16.935733 RB10279 151 456 heat shock protein, Hsp20 family 5557118 5557573 4.9 17072 + 7.33 4.02 0.42 10.405734 RB10283 107 324 small heat shock protein 5557737 5558060 4.7 12267 + 7.46 4.53 0.12 2.595911 RB10627 93 282 Chaperonin Cpn10 5769148 5769429 3.9 9966 + 7.24 1.70 0.21 12.186601 RB11847 243 732 Peptide methionine sulfoxide reductase msrA msrA 1.8.4.6 6379337 6380068 6.5 26839 + 6.15 1.90 0.19 9.816619 RB11878 168 507 peptide methionine sulfoxide reductase msrA msrA3 1.8.4.6 6396598 6397104 4.8 19100 + 5.86 2.06 0.45 21.626727 RB12080 492 1479 Protein-L-isoaspartate(D-aspartate) O-methyltransferase 2.1.1.77 6517231 6515753 8.9 53385 - 5.77 1.71 0.14 7.925783 RB10378 144 435 Thioredoxin trxA 5610497 5610063 4.6 15274 - 5.81 1.70 0.29 16.96791 RB1477 538 1617 arylsulfatase precursor ARSA 3.1.6.8 755327 756943 5.5 59805 + 5.26 1.59 0.14 8.78
5301 RB9498 518 1557 Arylsulfatase A Arsa 3.1.6.8 5099139 5100695 6 58375 + 5.27 2.04 0.29 14.125967 RB10727 276 831 manganese-containing catalase 5830775 5831605 5 30055 + 6.91 2.59 0.07 2.77708 RB1323 429 1290 Protein of unknown function DUF894, DitE 672945 671656 11.8 45426 - 5.98 1.67 0.20 11.94592 RB1122 449 1350 secretion protein lktD 578202 576853 5.3 50153 - 5.50 1.54 0.20 12.81
1759 RB3253 357 1074 ABC transporter, ATP-binding protein yhcG 1675049 1673976 5.7 39316 - 5.81 1.59 0.20 12.833648 RB6469 252 759 ABC transporter ATP-binding protein 3419515 3420273 5.6 27684 + 5.81 2.60 0.19 7.184039 RB7166 312 939 ABC-type multidrug transport system, ATPase component 3800705 3801643 5.4 35063 + 5.47 1.78 0.14 8.125260 RB9420 663 1992 Lipid A export ATP-binding/permease protein msbA 5055368 5053377 9.3 73442 - 5.96 1.98 0.12 6.285727 RB10270 537 1614 phytoene dehydrogenase carC 1.14.99.- 5551969 5550356 5.5 60117 - 5.72 2.82 0.23 8.176112 RB10981 600 1803 MULTIDRUG EFFLUX SYSTEM PROTEIN 5948851 5950653 4.5 66407 + 6.01 1.60 0.21 13.326113 RB10984 1115 3348 Acriflavin resistance protein 5950650 5953997 4.6 122096 + 5.81 1.62 0.12 7.156655 RB11943 534 1605 phytoene desaturase 6432376 6430772 5.8 57855 - 5.75 1.56 0.28 17.693933 RB6973 278 837 tellurite resistance protein 3710584 3711420 5 30569 + 5.48 2.67 0.45 16.793950 RB7004 271 816 Short-chain dehydrogenase/reductase SDR 3727471 3726656 9.7 29211 - 5.80 1.68 0.16 9.806414 RB11488 265 798 oxidoreductase, short-chain dehydrogenase/reductase family 6213893 6214690 5.1 27687 + 6.07 2.04 0.15 7.12158 RB282 342 1029 conserved hypothetical protein, membrane 141075 140047 7.4 37446 - 5.97 1.93 0.08 4.07228 RB392 218 657 hydrolase 204269 204925 4.3 24145 + 5.82 1.96 0.04 1.99745 RB1391 538 1617 cell division inhibitor- 710780 709164 7.8 59400 - 6.36 2.02 0.26 12.71846 RB1577 860 2583 ATP dependent DNA helicase 806289 808871 9.9 95417 + 5.93 1.65 0.19 11.59952 RB1798 364 1095 methanol dehydrogenase regulatory protein 930673 931767 4.6 40611 + 5.50 2.12 0.20 9.63
1608 RB2968 386 1161 GTPase EngC engC 3.6.1.- 1529085 1530245 8.6 41916 + 5.88 2.14 0.22 10.202547 RB4579 194 585 Protein of unknown function, UPF0054 2346073 2345489 3.7 20879 - 5.50 2.07 0.39 18.612548 RB4580 794 2385 metal-dependent phosphohydrolase 2348454 2346070 8.7 86766 - 5.91 1.50 0.31 20.832571 RB4619 481 1446 dehydrogenase 2363086 2364531 5.1 52511 + 6.21 1.64 0.29 17.812896 RB5140 613 1842 hypothetical protein 2641937 2643778 5.5 67565 + 5.49 1.99 0.32 15.873602 RB6395 778 2337 conserved hypothetical protein 3364879 3367215 4.6 85714 + 6.02 2.44 0.04 1.653936 RB6982 286 861 ABC transporter ATP-binding protein 3715019 3715879 5 31150 + 5.54 2.80 0.12 4.434037 RB7161 106 321 protein containing pyrophosphohydrolase domain 3798000 3798320 4.3 11777 + 5.35 1.72 0.43 24.864603 RB8173 339 1020 MoxR protein 4366881 4367900 5.1 37556 + 5.91 2.05 0.07 3.314622 RB8214 589 1770 membrane protein 4392948 4391179 4.5 63301 - 6.10 1.76 0.15 8.454915 RB8768 347 1044 methanol dehydrogenase regulatory protein (MoxR protein) 4661498 4660455 4.8 38221 - 5.38 1.59 0.26 16.626114 RB10985 207 624 HD domain protein 5954633 5954010 5.7 23118 - 5.13 2.61 0.29 11.006267 RB11262 331 996 diacylglycerol kinase 6108302 6107307 5 35708 - 5.90 1.93 0.14 7.246370 RB11438 54 165 conserved hypothetical protein 6197926 6197762 11.3 6209 - 5.39 2.12 0.18 8.396641 RB11923 231 696 Electron transport protein SCO1/SenC 6419736 6419041 4.3 25792 - 5.80 1.77 0.22 12.187201 RB12934 253 762 conserved hypothetical protein 6964029 6963268 4.8 27657 - 5.51 1.55 0.27 17.587369 RB13257 286 861 2-hydroxy-6-ketonona-2,4-dienedioic acid hydrolase bphD 3.7.1.- 7105590 7106450 4.6 32540 + 5.89 2.67 0.26 9.671502 RB2774 82 249 conserved hypothetical protein 1443786 1444034 4.4 9450 + 6.21 1.89 0.27 14.442438 RB4395 170 513 carboxymuconolactone decarboxylase 4.1.1.44 2262023 2262535 4.7 18791 + 6.05 1.70 0.28 16.412453 RB4419 178 537 Ferritin and Dps 2272912 2273448 6.2 18995 + 6.20 2.23 0.37 16.373454 RB6139 187 564 Protein of unknown function, UPF0157 3226050 3226613 5.7 21081 + 5.29 2.02 0.26 12.724294 RB7616 221 666 protein containing DUF162 4094752 4094087 4.7 23976 - 5.73 2.44 0.16 6.554604 RB8174 369 1110 conserved hypothetical protein 4367851 4368960 9.9 40726 + 6.05 1.50 0.18 11.944911 RB8759 747 2244 conserved hypothetical protein, membrane 4657568 4655325 4.4 82203 - 5.73 1.59 0.36 22.676131 RB11013 218 657 conserved hypothetical protein, membrane 5971802 5971146 4.9 24214 - 5.28 1.56 0.25 16.036217 RB11176 153 462 protein containing DUF442 6065605 6065144 4.8 17161 - 6.36 1.68 0.29 17.396348 RB11399 132 399 Putative thiol-disulphide oxidoreductase DCC 6179167 6179565 6.5 15047 + 6.96 2.58 0.16 6.22
80 RB140 987 2964 serine/threonine protein kinase pkn3 2.7.1.37 61578 64541 5.2 106565 + 5.82 1.90 0.08 4.462510 RB4511 1637 4914 two-component sensor histidine kinase 2307911 2312824 4.8 183570 + 5.44 1.64 0.29 17.683294 RB5848 140 423 dnaK suppressor protein, 3066328 3065906 4.6 15633 - 6.35 2.39 0.18 7.463615 RB6417 249 750 conserved hypothetical protein 3378045 3378794 5.9 27679 + 6.55 1.84 0.26 14.245758 RB10330 860 2583 sensory box sensor histidine kinase/response regulator 5587514 5584932 4.9 94952 - 5.69 2.87 0.22 7.636266 RB11260 121 366 dnaK suppressor protein, 6107202 6106837 5.2 13639 - 6.02 1.85 0.23 12.65
79 RB138 214 645 Sigma factor, ECF-like 60872 61516 4.8 24057 + 5.71 3.59 0.24 6.55234 RB402 151 456 hypothetical protein 212820 213275 4.4 17306 + 5.21 2.14 0.35 16.43300 RB520 375 1128 conserved hypothetical protein 284989 286116 4.8 41300 + 5.86 2.31 0.41 17.82301 RB521 63 192 hypothetical protein 286399 286208 10.7 7040 - 6.10 3.40 0.26 7.64319 RB550 137 414 secreted protein 306709 306296 12.2 15112 - 5.83 2.04 0.08 3.90362 RB663 79 240 hypothetical protein 357608 357369 11.4 8792 - 6.43 2.32 0.26 11.18366 RB671 122 369 hypothetical protein 359218 359586 10.7 13383 + 6.09 2.03 0.43 21.10392 RB723 60 183 hypothetical protein 388051 388233 7.3 6624 + 5.71 1.53 0.21 13.60507 RB934 375 1128 Putative transposase 494203 493076 9.9 41881 - 5.26 1.63 0.18 11.31517 RB952 258 777 hypothetical protein 502128 502904 10.3 28841 + 5.39 1.79 0.31 17.11662 RB1239 155 468 hypothetical protein 632500 632033 9.7 17106 - 5.63 1.54 0.17 10.98667 RB1252 665 1998 secreted protein 638429 640426 3.8 72471 + 5.23 2.11 0.46 21.60669 RB1255 51 156 hypothetical protein 640910 640755 12.3 5933 - 5.21 2.13 0.20 9.59670 RB1256 51 156 hypothetical protein 641044 640889 12 5840 - 4.75 1.57 0.27 17.19707 RB1322 35 108 hypothetical protein 671526 671633 12.8 3914 + 5.95 2.46 0.45 18.31709 RB1324 77 234 hypothetical protein 673169 672936 10.9 8459 - 5.85 1.84 0.26 13.90742 RB1386 350 1053 inosine-uridine preferring nucleoside hydrolase 708536 707484 4.8 38419 - 5.08 1.66 0.39 23.23743 RB1389 152 459 hypothetical protein 708892 708434 11 16822 - 5.90 1.69 0.18 10.62744 RB1390 149 450 membrane protein 709167 708718 9.9 16437 - 6.06 1.61 0.03 2.11747 RB1394 78 237 hypothetical protein 711994 711758 10.1 8765 - 6.13 2.54 0.30 11.99748 RB1395 319 960 secreted protein 712097 713056 5.5 35212 + 6.62 2.42 0.20 8.39804 RB1501 50 153 hypothetical protein 769419 769571 7.3 5710 + 5.58 3.20 0.17 5.19844 RB1573 36 111 hypothetical protein 805094 804984 11.9 4165 - 5.27 2.19 0.35 15.95913 RB1722 651 1956 hypothetical protein 897688 899643 5.4 70377 + 5.84 4.18 0.54 12.84915 RB1728 285 858 membrane protein containing DUF1206 899954 900811 9.9 30055 + 5.98 2.42 0.43 17.89916 RB1729 120 363 hypothetical protein 901146 900784 7.7 13145 - 5.51 2.95 0.15 5.25933 RB1760 395 1188 membrane protein 915210 914023 7.8 43386 - 5.45 1.82 0.22 11.96936 RB1767 907 2724 60-kDa cysteine-rich outer membrane protein precursor omcB 919059 916336 4.2 96071 - 5.36 1.80 0.24 13.53947 RB1789 243 732 conserved hypothetical protein 925861 926592 9.4 27028 + 5.96 1.85 0.43 23.23
1371 RB2525 76 231 hypothetical protein 1320284 1320054 6.1 8326 - 5.84 1.56 0.28 18.031477 RB2724 266 801 membrane protein 1419817 1419017 7.4 29523 - 6.30 1.64 0.19 11.771515 RB2804 201 606 hypothetical protein 1458645 1458040 5.3 22766 - 5.82 2.36 0.12 5.101518 RB2809 226 681 hypothetical protein 1459745 1459065 9.1 25318 - 6.19 1.77 0.04 2.461548 RB2864 596 1791 hypothetical protein 1490057 1491847 3.9 62722 + 4.87 2.02 0.02 0.971549 RB2867 697 2094 hypothetical protein 1491909 1494002 4.7 78102 + 5.41 1.50 0.20 13.181606 RB2963 119 360 hypothetical protein 1526707 1527066 12.2 13440 + 6.41 1.89 0.08 4.171728 RB3196 109 330 hypothetical protein 1650093 1650422 10.1 12287 + 6.88 2.20 0.10 4.67

1744 RB3228 962 2889 protein containing DUF1585 1656635 1659523 4.6 107564 + 5.43 2.87 0.52 18.121957 RB3587 113 342 hypothetical protein 1864796 1864455 11.8 13003 - 5.57 1.54 0.16 10.562169 RB3929 54 165 hypothetical protein 2034407 2034243 10.7 5796 - 4.94 1.83 0.36 19.732396 RB4322 457 1374 transposase and inactivated derivative 2226866 2228239 8.4 52492 + 5.32 2.02 0.13 6.512398 RB4325 48 147 Planctomycete PGAMP 2228477 2228623 12.5 5403 + 5.16 2.30 0.24 10.462405 RB4341 109 330 hypothetical protein 2234849 2235178 7.5 12018 + 4.91 1.98 0.09 4.702429 RB4381 59 180 hypothetical protein 2255621 2255442 10.7 7087 - 6.92 1.79 0.08 4.322431 RB4383 545 1638 decarboxylase 2258768 2257131 5.4 59839 - 6.53 1.57 0.32 20.352440 RB4397 55 168 protein containing DUF1560 2263976 2264143 10.5 5816 + 6.11 2.80 0.44 15.712446 RB4405 119 360 conserved hypothetical protein 2266894 2266535 4.3 13746 - 5.85 1.71 0.39 22.652458 RB4426 60 183 hypothetical protein 2275786 2275604 6.3 6516 - 5.16 2.65 0.21 7.872461 RB4429 89 270 conserved hypothetical protein 2275996 2276265 4.9 10034 + 6.86 1.77 0.19 10.842465 RB4438 160 483 Pyridoxamine 5'-phosphate oxidase- 2278247 2278729 4.4 18043 + 6.15 2.88 0.68 23.582475 RB4457 98 297 hypothetical protein 2285628 2285924 10 10915 + 5.62 1.53 0.21 13.862478 RB4460 422 1269 conserved hypothetical protein, membrane 2289696 2288428 4.8 48179 - 6.07 1.74 0.10 6.022585 RB4643 96 291 conserved hypothetical protein 2377555 2377845 8.9 11064 + 5.57 1.91 0.15 7.852607 RB4676 221 666 hypothetical protein 2391079 2390414 10.4 23751 - 5.64 1.65 0.18 11.212608 RB4678 199 600 secreted protein 2390618 2391217 4.2 20927 + 6.41 2.11 0.27 12.992609 RB4679 175 528 secreted protein 2391214 2391741 4.2 18596 + 6.52 1.84 0.39 20.982617 RB4698 86 261 hypothetical protein 2397601 2397861 12.2 9541 + 5.38 2.83 0.20 7.202696 RB4812 92 279 secreted protein 2467011 2467289 12.5 10014 + 5.31 2.73 0.02 0.752850 RB5064 648 1947 secreted protein containing DUF1585 2605707 2607653 4.6 72145 + 5.43 1.64 0.27 16.392951 RB5238 73 222 hypothetical protein 2697775 2697996 10.6 7866 + 5.53 3.64 0.27 7.493022 RB5370 309 930 Rhodopirellula transposase 2774826 2773897 5.9 34530 - 5.69 2.23 0.22 9.703023 RB5371 109 330 hypothetical protein 2774829 2775158 7.5 12033 + 5.11 2.23 0.28 12.783118 RB5523 91 276 hypothetical protein 2860984 2860709 6.3 9939 - 5.71 1.98 0.32 16.223130 RB5551 663 1992 hypothetical protein 2889101 2891092 5.7 70932 + 5.70 2.59 0.40 15.523139 RB5567 66 201 hypothetical protein 2897766 2897566 12.1 7106 - 5.83 2.64 0.29 11.153303 RB5866 441 1326 ISXo8 transposase 3076242 3077567 9.2 50613 + 5.36 1.52 0.19 12.733346 RB5938 370 1113 hypothetical protein 3113484 3112372 5.6 41837 - 6.14 4.02 0.25 6.183347 RB5939 39 120 hypothetical protein 3113462 3113581 10.9 4404 + 5.22 2.50 0.28 11.013457 RB6144 38 117 hypothetical protein 3228305 3228189 11.1 4314 - 5.63 2.46 0.33 13.223472 RB6168 208 627 hypothetical protein 3236689 3237315 12.2 22575 + 5.45 1.74 0.28 16.333482 RB6188 68 207 hypothetical protein 3244384 3244178 9.6 6986 - 5.38 1.54 0.07 4.633503 RB6233 629 1890 membrane protein 3271181 3269292 5.4 69401 - 6.84 1.67 0.05 2.853531 RB6282 200 603 conserved hypothetical protein 3302795 3303397 4.8 22451 + 5.37 1.93 0.09 4.673596 RB6388 144 435 hypothetical protein 3360906 3360472 9.1 16558 - 5.60 2.43 0.19 8.013621 RB6429 61 186 hypothetical protein 3382998 3382813 12.1 7095 - 7.35 1.79 0.15 8.363771 RB6685 72 219 hypothetical protein 3554265 3554047 12.2 8077 - 5.54 1.85 0.07 3.803784 RB6706 37 114 hypothetical protein 3560942 3560829 7.3 4375 - 5.45 1.96 0.22 11.093888 RB6888 420 1263 secreted protein containing DUF1573 3662449 3661187 4.3 45773 - 5.64 3.35 0.27 7.973910 RB6928 160 483 hypothetical protein 3678479 3678961 4.1 16965 + 5.87 1.80 0.43 24.133969 RB7039 202 609 hypothetical protein 3743573 3742965 7.9 23105 - 5.41 1.85 0.22 11.714021 RB7128 85 258 hypothetical protein 3782627 3782370 4.8 9275 - 5.44 1.52 0.13 8.484151 RB7381 243 732 membrane protein 3950326 3949595 7.4 26253 - 5.78 3.08 0.42 13.724157 RB7389 375 1128 Putative transposase 3953590 3954717 9.9 41895 + 5.42 1.82 0.06 3.254583 RB8132 177 534 hypothetical protein 4349972 4350505 10.8 19786 + 6.21 1.86 0.14 7.394595 RB8153 486 1461 conserved hypothetical protein 4359392 4357932 4.8 54199 - 5.45 2.53 0.14 5.484681 RB8325 296 891 hypothetical protein 4447029 4446139 6.8 32118 - 5.39 2.14 0.19 8.844704 RB8365 90 273 membrane protein 4463903 4463631 11.7 9405 - 6.60 1.58 0.27 16.934727 RB8409 97 294 hypothetical protein 4495547 4495840 8.9 10905 + 5.57 1.50 0.06 4.214791 RB8527 330 993 protein containing DUF1559 4549097 4550089 6.6 35645 + 5.70 1.54 0.12 8.034836 RB8601 417 1254 hypothetical protein 4588912 4590165 4.2 45382 + 5.86 1.78 0.29 16.465023 RB8959 662 1989 hypothetical protein 4777121 4775133 5.3 70536 - 6.46 1.58 0.39 24.365027 RB8969 101 306 Chaperonin Cpn10 4779554 4779859 4.9 10628 + 6.67 2.80 0.43 15.295031 RB8975 110 333 hypothetical protein 4783888 4783556 10.4 12068 - 6.28 2.44 0.30 12.305038 RB8987 48 147 hypothetical protein 4790856 4790710 9.1 5309 - 5.10 2.86 0.10 3.465093 RB9104 45 138 hypothetical protein 4866696 4866559 9 4979 - 6.56 1.79 0.24 13.615157 RB9230 107 324 hypothetical protein 4936673 4936996 9.9 11900 + 5.98 2.10 0.45 21.365158 RB9231 109 330 hypothetical protein 4937008 4936679 7.5 12033 - 4.93 2.14 0.39 18.405159 RB9232 309 930 Rhodopirellula transposase 4937011 4937940 5.9 34530 + 5.32 1.69 0.39 23.065346 RB9577 51 156 hypothetical protein 5150501 5150346 9.3 5646 - 5.31 1.57 0.16 10.085456 RB9787 61 186 hypothetical protein 5270904 5270719 6.1 6701 - 5.29 2.73 0.24 8.935473 RB9811 102 309 secreted protein 5284365 5284673 11.7 11169 + 5.65 2.44 0.23 9.355524 RB9907 433 1302 ISXo8 transposase 5338185 5339486 9.4 49646 + 5.45 1.83 0.09 5.125550 RB9955 452 1359 secreted protein containing DUF1552 5363605 5362247 5.6 50315 - 5.95 2.42 0.23 9.475552 RB9959 65 198 hypothetical protein 5366417 5366220 11.9 7376 - 6.11 2.01 0.12 6.185576 RB10007 313 942 conserved hypothetical protein 5390130 5391071 5 34128 + 6.69 2.43 0.11 4.705607 RB10051 1210 3633 conserved hypothetical protein 5420157 5423789 4.6 135400 + 5.83 1.92 0.16 8.485654 RB10137 73 222 protein containing DUF1590 5469772 5469551 12.1 8049 - 5.46 2.03 0.15 7.225738 RB10292 581 1746 conserved hypothetical protein, secreted 5559646 5561391 8.3 66858 + 5.91 2.65 0.23 8.735741 RB10299 455 1368 ATP-dependent carboligase related to biotin carboxylase (ORFY) c1 5563036 5564403 10 49949 + 7.02 2.65 0.07 2.655771 RB10350 163 492 Thioredoxin 5596878 5596387 9.7 18371 - 6.04 2.03 0.44 21.645806 RB10423 2079 6240 hemolysin 5640680 5646919 3.6 219600 + 5.56 2.28 0.11 4.855815 RB10440 49 150 hypothetical protein 5655693 5655842 11.5 5080 + 6.84 1.68 0.31 18.485874 RB10556 134 405 conserved hypothetical protein, membrane 5727064 5727468 10.5 15094 + 5.23 2.12 0.30 14.065876 RB10560 934 2805 hypothetical protein 5731867 5729063 4.6 100880 - 5.72 2.23 0.22 9.865910 RB10623 60 183 secreted protein 5769140 5768958 3.9 6284 - 7.86 1.74 0.14 8.155912 RB10629 539 1620 GroEL 5769500 5771119 4.9 57511 + 7.17 2.19 0.25 11.435931 RB10665 200 603 hypothetical protein 5789912 5790514 5.3 21401 + 6.32 1.67 0.05 2.775968 RB10728 132 399 secreted protein 5831649 5832047 9.9 13833 + 6.38 3.22 0.30 9.266053 RB10877 453 1362 membrane protein 5905128 5903767 4.3 48882 - 5.38 2.78 0.30 10.806067 RB10896 161 486 secreted protein 5917040 5916555 10 17034 - 5.85 2.06 0.16 7.996087 RB10936 56 171 hypothetical protein 5934348 5934518 12.5 6620 + 5.59 2.91 0.28 9.466094 RB10948 153 462 conserved hypothetical protein, secreted 5938575 5939036 4.2 17070 + 8.45 1.55 0.15 9.896095 RB10950 233 702 membrane or secreted protein 5939124 5939825 4.9 25130 + 5.72 1.55 0.15 9.876108 RB10974 234 705 hypothetical protein 5946559 5947263 5.3 25956 + 5.50 1.50 0.16 10.646124 RB11000 224 675 conserved hypothetical protein, membrane 5964054 5963380 8.8 24185 - 5.95 1.80 0.32 17.896177 RB11109 109 330 hypothetical protein 6022597 6022268 7.5 12033 - 5.34 1.58 0.35 22.466178 RB11110 309 930 Rhodopirellula transposase 6022600 6023529 5.8 34570 + 5.76 1.76 0.20 11.396347 RB11397 207 624 SOUL heme-binding protein 6178540 6179163 8.7 23072 + 6.86 2.66 0.12 4.516424 RB11504 72 219 conserved hypothetical protein 6221285 6221067 10.7 8212 - 6.02 3.56 0.41 11.666430 RB11515 74 225 conserved hypothetical protein 6224420 6224644 11.7 8687 + 5.45 3.72 0.40 10.736455 RB11566 195 588 conserved hypothetical protein 6243111 6243698 10.8 22135 + 5.97 2.51 0.27 10.956472 RB11603 37 114 conserved hypothetical protein 6256722 6256835 9 3998 + 6.32 2.41 0.15 6.286475 RB11612 81 246 conserved hypothetical protein 6260715 6260470 12 9246 - 5.42 1.98 0.21 10.836511 RB11667 174 525 conserved hypothetical protein 6293231 6292707 10 19482 - 5.66 2.24 0.08 3.456556 RB11746 503 1512 protein containing DUF1501 6326162 6327673 5.8 55474 + 5.66 2.00 0.18 8.986606 RB11855 101 306 conserved hypothetical protein 6383765 6383460 12.5 11384 - 5.20 1.59 0.10 6.476633 RB11909 168 507 conserved hypothetical protein 6410157 6410663 4.1 18314 + 5.22 2.35 0.32 13.556673 RB11977 196 591 conserved hypothetical protein 6455958 6456548 9.5 21922 + 5.55 2.45 0.37 15.236691 RB12009 514 1545 conserved hypothetical protein 6479495 6477951 4.8 54935 - 5.39 1.92 0.09 4.916719 RB12067 107 324 conserved hypothetical protein 6510236 6510559 11.9 12169 + 5.96 2.08 0.37 17.946729 RB12082 244 735 haloacid dehalogenase-type hydrolase 6519608 6518874 4.3 26835 - 5.49 2.38 0.30 12.506760 RB12150 53 162 conserved hypothetical protein 6556901 6557062 4.1 5983 + 6.37 2.50 0.42 16.916783 RB12196 49 150 conserved hypothetical protein 6577078 6576929 7.4 5280 - 5.56 1.97 0.10 4.826812 RB12239 433 1302 ISXo8 transposase 6593334 6594635 9.4 49646 + 5.33 1.65 0.11 6.546815 RB12243 55 168 conserved hypothetical protein 6596256 6596423 10.5 5848 + 5.81 3.19 0.47 14.736817 RB12247 74 225 conserved hypothetical protein 6597048 6596824 6.3 8306 - 6.10 3.47 0.81 23.266861 RB12323 248 747 conserved hypothetical protein 6627001 6627747 4.4 27447 + 5.21 1.72 0.31 17.986878 RB12348 260 783 FxsA cytoplasmic membrane protein 6645520 6644738 11.6 28277 - 6.31 1.72 0.40 23.406889 RB12375 446 1341 conserved hypothetical protein 6659741 6661081 4.9 48379 + 6.30 1.72 0.07 3.786950 RB12471 85 258 conserved hypothetical protein 6712442 6712185 9.7 9841 - 5.62 1.53 0.11 6.947088 RB12727 680 2043 conserved hypothetical protein 6860301 6858259 5.7 73171 - 5.94 2.45 0.52 21.087090 RB12732 422 1269 secreted protein containing DUF1501 6862068 6860800 6.3 45705 - 5.50 3.01 0.13 4.217092 RB12737 65 198 conserved hypothetical protein 6863917 6864114 9.1 6753 + 6.77 1.87 0.12 6.407095 RB12741 905 2718 conserved hypothetical protein 6868329 6865612 4.2 97067 - 5.17 2.68 0.08 3.117132 RB12806 205 618 conserved hypothetical protein 6911165 6910548 10.5 21914 - 5.10 2.30 0.12 5.357133 RB12808 369 1110 secreted protein 6912350 6911241 4.6 40963 - 5.12 2.46 0.48 19.427148 RB12829 159 480 conserved hypothetical protein 6919813 6920292 9.5 17677 + 5.79 2.23 0.20 8.917149 RB12830 102 309 conserved hypothetical protein 6920508 6920200 10.9 11753 - 5.97 2.50 0.43 17.057160 RB12850 309 930 Rhodopirellula transposase 6926553 6925624 5.8 34461 - 5.35 2.16 0.21 9.557161 RB12851 109 330 conserved hypothetical protein 6926556 6926885 7.5 12033 + 5.21 2.36 0.23 9.73
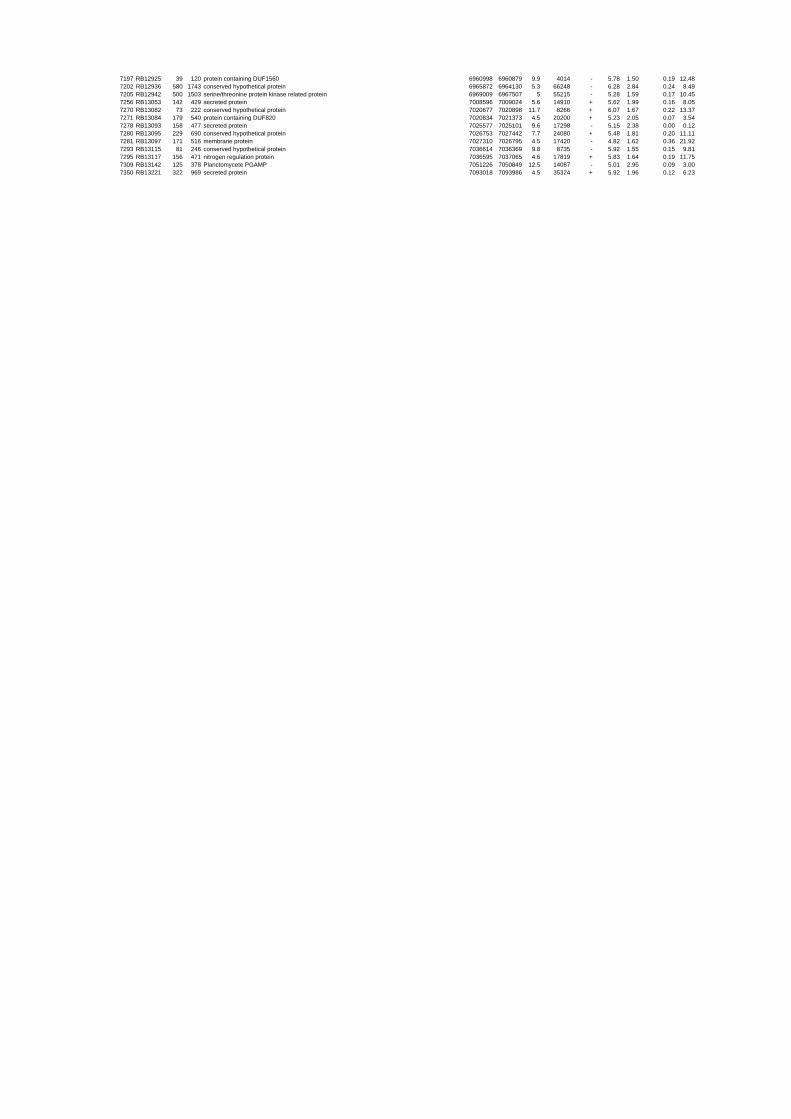
7197 RB12925 39 120 protein containing DUF1560 6960998 6960879 9.9 4014 - 5.78 1.50 0.19 12.487202 RB12936 580 1743 conserved hypothetical protein 6965872 6964130 5.3 66248 - 6.28 2.84 0.24 8.497205 RB12942 500 1503 serine/threonine protein kinase related protein 6969009 6967507 5 55215 - 5.28 1.59 0.17 10.457256 RB13053 142 429 secreted protein 7008596 7009024 5.6 14910 + 5.62 1.99 0.16 8.057270 RB13082 73 222 conserved hypothetical protein 7020677 7020898 11.7 8266 + 6.07 1.67 0.22 13.377271 RB13084 179 540 protein containing DUF820 7020834 7021373 4.5 20200 + 5.23 2.05 0.07 3.547278 RB13093 158 477 secreted protein 7025577 7025101 9.6 17298 - 5.15 2.38 0.00 0.127280 RB13095 229 690 conserved hypothetical protein 7026753 7027442 7.7 24080 + 5.48 1.81 0.20 11.117281 RB13097 171 516 membrane protein 7027310 7026795 4.5 17420 - 4.82 1.62 0.36 21.927293 RB13115 81 246 conserved hypothetical protein 7036614 7036369 9.8 8735 - 5.92 1.55 0.15 9.817295 RB13117 156 471 nitrogen regulation protein 7036595 7037065 4.6 17819 + 5.83 1.64 0.19 11.757309 RB13142 125 378 Planctomycete PGAMP 7051226 7050849 12.5 14087 - 5.01 2.95 0.09 3.007350 RB13221 322 969 secreted protein 7093018 7093986 4.5 35324 + 5.92 1.96 0.12 6.23

Heat repressed10min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand Intensity Ratio Stdev %
238 RB410 342 1029 anthranilate phosphoribosyltransferase trpD 2.4.2.18 219986 218958 5.1 35653 - 6.19 -1.13 0.10 8.471967 RB3603 344 1035 secreted protein 1875461 1874427 4.6 37999 - 6.12 -1.59 0.22 13.962359 RB4269 282 849 glutamic acid specific endopeptidase, GSE=serine endopeptidase 2185049 2184201 5.6 31283 - 6.14 -1.55 0.18 11.386865 RB12327 686 2061 TGF-beta receptor, type I/II extracellular region 6633918 6631858 4.5 75645 - 6.02 -1.29 0.04 3.462192 RB3967 544 1635 sodium:proline/pantothenate symporter (proline/pantothenate permease) 2053237 2054871 9.4 59256 + 6.13 -1.17 0.12 9.931423 RB2627 342 1029 Glyceraldehyde 3-phosphate dehydrogenase gap 1.2.1.12 1364551 1365579 5.4 36716 + 6.92 -1.44 0.07 4.53179 RB314 309 930 malonyl CoA-acyl carrier protein transacylase fabD 2.3.1.39 156968 157897 4.4 32349 + 7.17 -1.91 0.12 6.29181 RB318 81 246 Acyl carrier protein acpP 158928 159173 3.7 9018 + 6.63 -1.25 0.09 6.78
2733 RB4876 397 1194 delta 9 acyl-lipid fatty acid desaturase 1.14.19.- 2506214 2505021 9.5 44741 - 7.98 -1.19 0.09 7.29145 RB264 64 195 Ribosomal protein S21 132677 132871 12.3 8074 + 6.59 -1.03 0.17 16.90
2286 RB4143 499 1500 Amidase 2130113 2131612 4.5 52330 + 6.09 -1.02 0.13 12.544014 RB7115 118 357 Ribosomal protein L20 3779618 3779262 12.2 13584 - 6.49 -1.14 0.09 7.834427 RB7836 107 324 Ribosomal L23 protein 4194060 4194383 10.8 12651 + 7.68 -1.02 0.05 5.104429 RB7838 89 270 Ribosomal protein S19/S15 4195376 4195645 10.8 9943 + 6.86 -1.05 0.24 22.734435 RB7850 122 369 Ribosomal protein L14b/L23e 4198081 4198449 11 13526 + 6.91 -1.05 0.11 10.824440 RB7856 181 546 50S ribosomal protein L6 rplF 4200306 4200851 10 19733 + 6.71 -1.13 0.19 16.524750 RB8459 55 168 30S ribosomal protein S18 rpsR 4518496 4518663 10.5 6203 + 6.82 -1.39 0.19 13.644824 RB8580 113 342 sigma-54 modulation protein 4577393 4577734 5.7 12706 + 7.03 -1.03 0.06 5.665832 RB10471 151 456 Ribosomal protein L13 5678759 5678304 10 17369 - 7.27 -1.15 0.08 6.977154 RB12839 225 678 Ribosomal protein L1 6921934 6922611 9.8 24159 + 7.34 -1.82 0.07 4.054459 RB7894 398 1197 translation elongation factor EF-Tu 4217484 4218680 5.2 43184 + 7.70 -1.75 0.09 4.951919 RB3521 108 327 ArsR family transcriptional regulatory protein 1823417 1823743 9.6 11920 + 6.23 -1.02 0.16 15.492610 RB4681 65 198 cold shock protein cspA 2391934 2392131 4.7 6982 + 7.32 -1.93 0.02 1.065823 RB10458 206 621 Rnpc2-prov protein 5671130 5670510 10.3 21043 - 6.71 -1.47 0.12 8.293528 RB6276 105 318 Histone-like bacterial DNA-binding protein 3297504 3297187 10.4 11299 - 7.66 -2.04 0.06 3.141615 RB2978 152 459 nudix (nucleoside diphosphate linked moiety X)-type motif 2 1536161 1536619 5.9 17438 + 6.10 -1.00 0.12 12.151349 RB2500 306 921 NAD-dependent epimerase/dehydratase 1307899 1306979 8.5 33992 - 6.22 -1.06 0.08 7.672244 RB4061 488 1467 Peptidase A24A, prepilin type IV:Peptidase A24A, N-terminal 2095498 2096964 9.7 54251 + 6.05 -1.07 0.03 3.061850 RB3403 491 1476 arylsulfatase precursor arsA 3.1.6.8 1756892 1755417 5.5 55139 - 6.01 -1.02 0.19 18.891644 RB3032 477 1434 Major facilitator superfamily MFS_1 1567813 1569246 9.8 52109 + 6.04 -1.07 0.06 5.401332 RB2479 273 822 conserved hypothetical protein 1296683 1295862 5.5 30436 - 6.09 -1.25 0.04 2.822476 RB4458 342 1029 Protein of unknown function, UPF0118 2287089 2286061 6.4 37001 - 6.31 -1.04 0.18 17.78
2 RB2 59 180 secreted protein 182 3 3.7 6489 - 6.80 -2.66 0.14 5.097 RB13 130 393 hypothetical protein 5850 5458 11 14731 - 6.36 -1.07 0.11 10.71
22 RB37 160 483 membrane protein 18328 17846 5.6 17283 - 6.08 -1.02 0.17 16.2734 RB56 89 270 hypothetical protein 24833 24564 4.5 9645 - 5.93 -1.08 0.11 10.1437 RB61 58 177 hypothetical protein 25378 25554 9.6 6492 + 6.04 -1.13 0.10 8.49
130 RB229 585 1758 secreted protein containing DUF1551 122782 121025 4.5 62454 - 6.09 -1.09 0.06 5.70182 RB319 95 288 hypothetical protein 159334 159621 10.2 10505 + 6.75 -1.56 0.11 7.20193 RB337 97 294 hypothetical protein 169241 169534 12.5 11059 + 6.03 -1.00 0.12 11.56202 RB354 51 156 hypothetical protein 177535 177380 12 5703 - 6.09 -1.15 0.13 11.71209 RB367 67 204 hypothetical protein 183924 183721 10.4 7521 - 5.89 -1.02 0.10 9.43250 RB434 65 198 hypothetical protein 231791 231988 7.4 7385 + 6.41 -1.41 0.12 8.33409 RB767 311 936 conserved hypothetical protein, secreted 411420 412355 5.7 34002 + 5.78 -1.00 0.16 16.12516 RB951 234 705 protein containing DUF1596 502459 501755 12.3 25583 - 6.30 -1.47 0.13 8.81622 RB1179 128 387 hypothetical protein 603998 604384 12 13863 + 7.02 -1.09 0.01 1.26
1067 RB2034 155 468 hypothetical protein 1071018 1071485 10.5 17610 + 6.82 -1.01 0.01 0.841088 RB2080 46 141 hypothetical protein 1091549 1091409 9.9 5071 - 6.76 -1.56 0.04 2.411105 RB2105 470 1413 membrane protein 1114012 1112600 9.6 51853 - 6.06 -1.14 0.12 10.131130 RB2150 93 282 hypothetical protein 1138311 1138030 4.4 10755 - 7.08 -2.06 0.14 6.601183 RB2247 579 1740 conserved hypothetical protein containing YTV domains 1181238 1179499 9.8 65013 - 7.18 -1.67 0.06 3.291184 RB2251 75 228 hypothetical protein 1181714 1181487 4.5 8027 - 7.30 -1.84 0.07 3.971188 RB2255 628 1887 hypothetical protein 1186018 1184132 4.1 66330 - 6.72 -1.58 0.06 3.721217 RB2302 46 141 hypothetical protein 1204388 1204528 5.6 5335 + 6.85 -1.29 0.12 9.671277 RB2384 65 198 hypothetical protein 1242697 1242500 12 7263 - 6.41 -1.27 0.08 6.521352 RB2503 515 1548 O-antigen flippase wzx 1311056 1309509 10 55740 - 5.93 -1.07 0.25 22.971353 RB2505 82 249 hypothetical protein 1311059 1311307 10 9658 + 6.06 -1.06 0.20 18.981424 RB2628 43 132 hypothetical protein 1365707 1365576 10 4777 - 6.35 -1.39 0.05 3.721428 RB2636 116 351 hypothetical protein 1368450 1368100 11.7 12761 - 6.88 -1.54 0.02 1.111446 RB2667 236 711 membrane protein 1387198 1386488 10.7 25698 - 6.10 -1.11 0.20 17.741479 RB2726 56 171 hypothetical protein 1421143 1421313 10.5 6492 + 6.25 -1.02 0.07 7.341557 RB2883 120 363 hypothetical protein 1499497 1499859 11.8 13089 + 6.09 -1.09 0.23 21.411560 RB2886 118 357 membrane protein 1501522 1501878 7.6 13062 + 6.20 -1.02 0.12 11.941597 RB2951 174 525 membrane protein 1523616 1523092 9.3 20061 - 6.02 -1.02 0.16 15.541684 RB3117 89 270 hypothetical protein 1615433 1615164 11.2 9951 - 6.20 -1.28 0.27 20.891692 RB3132 112 339 membrane protein 1619400 1619062 10.3 12831 - 5.98 -1.01 0.13 12.471705 RB3152 68 207 hypothetical protein 1626667 1626461 12.2 7261 - 6.51 -1.21 0.04 3.211771 RB3268 231 696 hypothetical protein 1683003 1682308 12 25730 - 6.12 -1.11 0.16 14.691776 RB3277 221 666 hypothetical protein 1686552 1685887 10.4 24222 - 6.05 -1.26 0.14 11.341789 RB3301 50 153 hypothetical protein 1694208 1694056 10.5 5470 - 6.22 -1.25 0.11 8.641819 RB3353 409 1230 Sialidase 1 Neu1 3.2.1.18 1725519 1724290 5.9 45043 - 5.98 -1.07 0.11 10.221825 RB3362 87 264 hypothetical protein 1728546 1728283 11.9 10017 - 6.17 -1.29 0.14 10.531827 RB3365 43 132 hypothetical protein 1730124 1730255 11.4 4886 + 8.16 -1.80 0.12 6.581828 RB3366 78 237 hypothetical protein 1730380 1730616 5.6 9481 + 6.11 -1.24 0.03 2.581833 RB3372 115 348 hypothetical protein 1733666 1734013 10 12916 + 6.02 -1.06 0.12 11.381843 RB3393 60 183 secreted protein 1751865 1752047 12.3 7156 + 6.78 -1.29 0.06 4.381844 RB3394 36 111 hypothetical protein 1752044 1752154 10.5 4239 + 6.34 -1.41 0.10 7.061845 RB3395 398 1197 secreted protein containing DUF1559 1752151 1753347 7.8 43136 + 6.62 -1.09 0.10 8.851847 RB3399 65 198 hypothetical protein 1753650 1753453 9.7 7418 - 6.22 -1.51 0.16 10.331943 RB3566 64 195 hypothetical protein 1854080 1854274 9.4 7400 + 6.08 -1.18 0.23 19.641950 RB3575 152 459 membrane protein 1858779 1859237 10.1 17195 + 6.12 -1.43 0.15 10.391992 RB3644 127 384 hypothetical protein 1895184 1894801 4.9 14294 - 7.24 -1.78 0.10 5.752006 RB3660 94 285 hypothetical protein 1900854 1900570 9.4 10411 - 6.33 -1.07 0.11 10.222011 RB3665 119 360 hypothetical protein 1903076 1902717 5.9 12608 - 6.05 -1.09 0.17 15.752017 RB3675 742 2229 secreted protein 1904552 1906780 8.4 82951 + 6.11 -1.11 0.10 9.302025 RB3688 53 162 hypothetical protein 1910629 1910468 9.3 5981 - 6.19 -1.24 0.09 7.172048 RB3719 141 426 hypothetical protein 1927500 1927075 10.8 16330 - 6.54 -1.52 0.15 10.132113 RB3835 50 153 hypothetical protein 1989687 1989839 11.3 5789 + 6.12 -1.04 0.18 17.532124 RB3851 75 228 hypothetical protein 1996660 1996433 7.6 8884 - 6.15 -1.02 0.17 16.902146 RB3890 125 378 hypothetical protein 2011554 2011177 4.4 14580 - 6.17 -1.17 0.05 3.932154 RB3905 38 117 hypothetical protein 2018601 2018717 5.1 4131 + 6.23 -1.19 0.06 4.952174 RB3935 55 168 hypothetical protein 2036241 2036074 9.3 5982 - 6.25 -1.01 0.11 11.092184 RB3953 857 2574 hypothetical protein 2048784 2046211 5.2 93881 - 6.11 -1.29 0.27 21.352198 RB3975 204 615 hypothetical protein 2056726 2057340 12.3 22602 + 6.10 -1.27 0.07 5.132200 RB3981 161 486 hypothetical protein 2058000 2058485 4.1 18283 + 6.04 -1.09 0.04 4.052210 RB3994 191 576 hypothetical protein 2061291 2061866 4.1 20739 + 6.02 -1.15 0.22 18.972258 RB4093 45 138 hypothetical protein 2107600 2107737 10 5285 + 6.08 -1.09 0.14 12.532260 RB4097 733 2202 conserved hypothetical protein 2108244 2110445 6.2 83188 + 6.12 -1.23 0.09 7.052285 RB4142 43 132 hypothetical protein 2130009 2130140 12 4964 + 6.28 -1.00 0.10 10.122287 RB4145 90 273 hypothetical protein 2131933 2131661 12 10727 - 6.49 -1.09 0.11 10.212312 RB4194 53 162 hypothetical protein 2156685 2156524 11.4 5728 - 7.23 -1.35 0.09 6.792323 RB4210 148 447 hypothetical protein 2160917 2161363 6.5 16112 + 6.18 -1.15 0.09 8.112343 RB4243 58 177 hypothetical protein 2174155 2174331 4.4 6442 + 6.01 -1.00 0.12 11.832356 RB4264 120 363 membrane protein 2183157 2182795 10.7 13335 - 6.28 -1.27 0.16 12.532368 RB4282 291 876 conserved hypothetical protein 2199894 2199019 5.3 33373 - 6.08 -1.08 0.19 17.492369 RB4283 499 1500 PvdD/PvdJ 2201462 2199963 5.7 56411 - 6.04 -1.26 0.08 6.052371 RB4285 841 2526 membrane protein 2204136 2201611 5.9 93852 - 6.03 -1.07 0.18 17.102378 RB4296 240 723 hypothetical protein 2213931 2213209 5.8 27191 - 6.11 -1.23 0.14 11.242416 RB4358 123 372 hypothetical protein 2239474 2239103 6.5 13459 - 6.19 -1.22 0.09 7.782417 RB4360 156 471 hypothetical protein 2240075 2239605 4.7 17592 - 6.16 -1.09 0.12 11.002424 RB4373 109 330 hypothetical protein 2242863 2242534 4.8 12263 - 6.12 -1.07 0.18 17.172593 RB4652 52 159 hypothetical protein 2380494 2380652 12.6 5628 + 6.91 -1.20 0.13 10.542596 RB4657 123 372 hypothetical protein 2381865 2382236 12.2 13398 + 6.29 -1.87 0.18 9.702647 RB4740 50 153 hypothetical protein 2431814 2431966 11.4 5598 + 6.17 -1.15 0.08 6.722746 RB4897 184 555 hypothetical protein 2519505 2518951 10.9 20446 - 5.85 -1.00 0.14 13.882777 RB4941 173 522 hypothetical protein 2541619 2542140 11.2 19306 + 6.00 -1.51 0.04 2.762781 RB4951 95 288 hypothetical protein 2547498 2547785 12.1 10953 + 6.73 -1.38 0.09 6.532964 RB5262 95 288 membrane protein 2713742 2714029 6.3 10993 + 6.04 -1.27 0.10 8.243045 RB5409 97 294 hypothetical protein 2794195 2794488 12.7 10898 + 8.04 -1.98 0.08 3.833049 RB5415 62 189 hypothetical protein 2798737 2798925 12.3 7098 + 6.86 -1.51 0.11 7.263052 RB5418 73 222 secreted protein 2805185 2804964 11 8406 - 5.85 -1.06 0.09 8.57

3106 RB5505 73 222 hypothetical protein 2850866 2851087 12.9 8233 + 6.40 -1.06 0.13 12.233132 RB5555 54 165 hypothetical protein 2891274 2891438 6.2 5971 + 6.10 -1.11 0.07 6.003239 RB5745 130 393 hypothetical protein 3001234 3000842 10.7 13522 - 7.50 -1.89 0.07 3.463271 RB5802 156 471 hypothetical protein 3039864 3040334 10.1 17001 + 7.00 -1.34 0.13 9.543466 RB6158 142 429 hypothetical protein 3233532 3233104 6 15746 - 6.20 -1.51 0.19 12.533476 RB6174 69 210 hypothetical protein 3239734 3239525 10.6 7870 - 5.95 -1.14 0.19 16.983485 RB6193 62 189 hypothetical protein 3246096 3245908 11.4 6970 - 6.39 -1.51 0.17 11.023547 RB6314 52 159 hypothetical protein 3319320 3319478 6.3 6180 + 6.96 -2.29 0.17 7.473617 RB6423 45 138 hypothetical protein 3380603 3380740 5.5 5402 + 6.58 -1.29 0.06 4.693701 RB6553 74 225 hypothetical protein 3479627 3479403 12.1 8987 - 6.37 -1.18 0.03 2.273731 RB6609 45 138 hypothetical protein 3511456 3511593 10.4 5004 + 6.66 -1.10 0.17 15.593744 RB6634 365 1098 protein containing DUF1559 3525785 3526882 5.3 40217 + 5.88 -1.17 0.14 11.553836 RB6789 180 543 secreted protein 3608487 3607945 5 20191 - 6.06 -1.32 0.09 6.713867 RB6849 101 306 hypothetical protein 3642537 3642232 12.8 12002 - 6.08 -1.12 0.12 11.063971 RB7042 91 276 hypothetical protein 3743971 3744246 10.4 10835 + 5.77 -1.08 0.05 5.104015 RB7116 59 180 hypothetical protein 3779586 3779765 11.7 6458 + 7.14 -1.37 0.06 4.614016 RB7117 181 546 Ribosomal protein L35 3780322 3779777 11.4 19816 - 7.00 -1.48 0.05 3.634049 RB7178 45 138 hypothetical protein 3806924 3807061 12.3 5320 + 7.17 -1.64 0.11 6.854050 RB7180 104 315 hypothetical protein 3807345 3807031 10.8 11535 - 6.84 -1.50 0.04 2.524051 RB7184 190 573 conserved hypothetical protein 3807517 3808089 9.2 21306 + 6.51 -1.58 0.05 3.364092 RB7246 50 153 hypothetical protein 3850393 3850545 9.7 5973 + 6.82 -1.40 0.02 1.514131 RB7313 51 156 hypothetical protein 3892097 3891942 10.5 5558 - 7.82 -1.61 0.10 6.104200 RB7455 838 2517 conserved hypothetical protein containing YTV domains 3999277 3996761 9 92217 - 6.93 -1.24 0.12 9.324237 RB7530 97 294 hypothetical protein 4049298 4049005 12.4 11656 - 6.39 -2.97 0.12 4.054241 RB7535 136 411 hypothetical protein 4050024 4050434 10.9 15058 + 6.90 -1.48 0.03 2.204278 RB7589 86 261 hypothetical protein 4080238 4079978 12.1 9472 - 5.65 -1.15 0.08 6.984314 RB7646 62 189 hypothetical protein 4111735 4111923 10.5 6950 + 6.91 -1.63 0.05 2.974315 RB7647 73 222 hypothetical protein 4111931 4112152 7.4 8269 + 9.28 -1.65 0.07 4.034327 RB7667 38 117 hypothetical protein 4117097 4116981 6.7 4315 - 7.05 -1.41 0.05 3.684424 RB7832 50 153 hypothetical protein 4192499 4192651 13 5566 + 7.71 -1.16 0.04 3.144486 RB7939 105 318 hypothetical protein 4238296 4238613 11 12019 + 6.22 -1.07 0.20 18.694531 RB8034 147 444 membrane protein 4295344 4295787 10.2 16546 + 5.87 -1.31 0.25 19.234568 RB8098 71 216 hypothetical protein 4337334 4337119 10.8 7950 - 9.46 -1.09 0.21 19.104696 RB8353 82 249 hypothetical protein 4459374 4459622 10.8 9402 + 6.33 -2.57 0.19 7.304861 RB8669 37 114 hypothetical protein 4614557 4614670 11.5 4395 + 9.56 -1.08 0.21 19.045052 RB9020 381 1146 secreted protein 4807486 4808631 5.9 42557 + 6.74 -1.05 0.14 13.655068 RB9057 95 288 hypothetical protein 4834849 4834562 10.9 10885 - 7.16 -1.08 0.09 7.965072 RB9067 57 174 secreted protein 4839661 4839834 3.4 6172 + 6.09 -1.04 0.08 7.665205 RB9324 267 804 secreted protein 4992474 4993277 6.7 29463 + 5.61 -1.01 0.09 9.165214 RB9343 59 180 hypothetical protein 5003789 5003968 11.4 6521 + 6.09 -1.03 0.18 17.375280 RB9460 79 240 hypothetical protein 5078860 5078621 10.3 8991 - 9.22 -1.24 0.10 8.445694 RB10209 110 333 H+-transporting two-sector ATPase, C subunit 3.6.3.14 5515135 5515467 10.2 11565 + 6.92 -1.07 0.05 5.045838 RB10483 133 402 conserved hypothetical protein 5685437 5685838 5.7 14433 + 6.30 -1.38 0.20 14.145855 RB10521 53 162 hypothetical protein 5708182 5708343 9.1 6074 + 6.72 -1.29 0.06 4.685887 RB10581 384 1155 secreted protein containing DUF1559 5739353 5740507 6.2 41431 + 7.21 -1.07 0.06 5.426364 RB11429 37 114 conserved hypothetical protein 6196478 6196591 12.4 4245 + 6.94 -1.40 0.10 6.986371 RB11439 93 282 conserved hypothetical protein 6197993 6198274 10.6 10906 + 8.15 -2.53 0.05 1.906385 RB11450 60 183 conserved hypothetical protein 6200046 6200228 11.8 6517 + 8.71 -1.58 0.09 5.526697 RB12021 82 249 conserved hypothetical protein 6485018 6484770 11.9 9305 - 6.37 -1.06 0.15 14.506867 RB12329 110 333 conserved hypothetical protein, membrane 6634234 6634566 4 11862 + 7.08 -1.18 0.04 3.057046 RB12649 112 339 conserved hypothetical protein 6810876 6810538 10.3 12615 - 5.53 -1.01 0.03 3.067141 RB12818 163 492 conserved hypothetical protein 6915989 6915498 11.7 18010 - 7.62 -1.82 0.09 4.79
20min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand Intensity Ratio Stdev %4829 RB8592 233 702 conserved hypothetical protein 4580947 4581648 6 26322 + 6.52 -1.14 0.03 3.056656 RB11945 556 1671 aldehyde dehydrogenase family protein 6434111 6432441 10.9 59626 - 6.66 -1.28 0.04 3.31515 RB950 442 1329 Cationic amino acid transporter 501765 500437 9.7 46180 - 6.31 -1.01 0.02 2.34
1967 RB3603 344 1035 secreted protein 1875461 1874427 4.6 37999 - 6.21 -1.02 0.05 4.862359 RB4269 282 849 glutamic acid specific endopeptidase, GSE=serine endopeptidase 2185049 2184201 5.6 31283 - 6.27 -1.09 0.10 9.056865 RB12327 686 2061 TGF-beta receptor, type I/II extracellular region 6633918 6631858 4.5 75645 - 6.39 -1.69 0.08 4.894927 RB8785 542 1629 L-fucose isomerase, 4672793 4671165 4.8 59809 - 6.31 -1.16 0.14 11.696891 RB12379 240 723 riboflavin synthase, alpha subunit 6662808 6662086 4.6 25728 - 6.56 -1.28 0.24 18.56179 RB314 309 930 malonyl CoA-acyl carrier protein transacylase fabD 2.3.1.39 156968 157897 4.4 32349 + 7.20 -2.10 0.36 16.94578 RB1092 396 1191 3-hydroxybutyryl-CoA dehydrogenase 565978 567168 5.2 43249 + 6.20 -1.01 0.09 8.93
2733 RB4876 397 1194 delta 9 acyl-lipid fatty acid desaturase 1.14.19.- 2506214 2505021 9.5 44741 - 7.98 -1.17 0.13 10.86658 RB1233 206 621 30S ribosomal protein S4 rpsD 629490 628870 11.2 23193 - 7.15 -1.31 0.10 7.41
4423 RB7829 108 327 Ribosomal protein S10 4192031 4192357 10.3 11931 + 7.67 -1.07 0.08 7.474427 RB7836 107 324 Ribosomal L23 protein 4194060 4194383 10.8 12651 + 7.53 -1.27 0.10 7.674428 RB7837 286 861 Ribosomal protein L2 4194481 4195341 11.8 31108 + 7.08 -1.25 0.27 21.404750 RB8459 55 168 30S ribosomal protein S18 rpsR 4518496 4518663 10.5 6203 + 7.33 -1.37 0.26 19.174824 RB8580 113 342 sigma-54 modulation protein 4577393 4577734 5.7 12706 + 7.05 -1.09 0.18 16.815832 RB10471 151 456 Ribosomal protein L13 5678759 5678304 10 17369 - 7.61 -1.45 0.09 6.147029 RB12623 127 384 Ribosomal protein S13 6796904 6797287 10.8 14481 + 7.38 -1.35 0.14 10.107030 RB12625 127 384 Ribosomal protein S11 6797354 6797737 11.4 13570 + 7.24 -1.04 0.15 14.017143 RB12821 117 354 Ribosomal protein L19 6916558 6916205 11.1 13347 - 6.83 -1.55 0.31 19.787145 RB12824 146 441 Ribosomal protein S16 6917743 6917303 5.3 15885 - 7.14 -1.54 0.28 18.094459 RB7894 398 1197 translation elongation factor EF-Tu 4217484 4218680 5.2 43184 + 7.57 -2.56 0.09 3.642610 RB4681 65 198 cold shock protein cspA 2391934 2392131 4.7 6982 + 7.32 -2.75 0.32 11.596819 RB12251 567 1704 DNA-directed RNA polymerase specialized sigma factor 6599262 6597559 9.4 59396 - 6.38 -1.06 0.16 14.823525 RB6271 325 978 Tyrosine recombinase xerD xerD 3294814 3295791 10 36623 + 6.38 -1.34 0.11 8.163528 RB6276 105 318 Histone-like bacterial DNA-binding protein 3297504 3297187 10.4 11299 - 8.02 -2.23 0.13 5.825531 RB9917 169 510 Single-strand binding protein/Primosomal replication protein n ssb 5344370 5343861 5.3 18108 - 7.18 -1.36 0.26 19.076885 RB12363 787 2364 UvrD/REP helicase 6656202 6653839 5.3 88835 - 6.49 -1.10 0.13 11.873524 RB6269 405 1218 UDP-glucose 4-epimerase galE 5.1.3.2 3294798 3293581 5.3 44117 - 6.21 -1.21 0.17 14.136968 RB12502 435 1308 flagellar motor switch protein FliG fliG 6732844 6731537 7 47011 - 6.35 -1.12 0.24 21.205282 RB9463 628 1887 ATPase with chaperone activity 5079098 5080984 9.4 69732 + 6.25 -1.07 0.18 16.75642 RB1205 456 1371 choline sulfatase betC 3.1.6.6 615037 613667 6.6 51848 - 6.31 -1.26 0.14 10.88
7313 RB13148 1012 3039 arylsulfatase A [precursor] 7058006 7054968 6.2 111097 - 6.37 -1.37 0.13 9.126663 RB11958 302 909 protein containing DUF191 6443018 6442110 4.5 32855 - 6.58 -1.41 0.16 11.596983 RB12532 405 1218 metallophosphoesterase, secreted 6749031 6750248 5.8 45205 + 6.34 -1.32 0.21 15.866686 RB11998 412 1239 secreted protein containing DUF323 6474041 6472803 4.1 45898 - 6.66 -1.18 0.14 12.184269 RB7578 189 570 Carbon storage regulator csrA 4070466 4071035 8.8 20421 + 7.23 -1.77 0.12 7.025722 RB10261 81 246 Carbon storage regulator csrA 5545964 5545719 5.1 8704 - 7.91 -1.11 0.07 6.276680 RB11988 880 2643 serine/threonine-protein kinase spk1 2.7.1.37 6468163 6465521 5.4 97920 - 6.34 -1.04 0.09 8.906776 RB12183 585 1758 GGDEF family protein 6573556 6571799 4.7 63291 - 6.28 -1.02 0.23 22.47
2 RB2 59 180 secreted protein 182 3 3.7 6489 - 7.11 -3.11 0.58 18.4837 RB61 58 177 hypothetical protein 25378 25554 9.6 6492 + 6.26 -1.18 0.08 7.01
449 RB825 117 354 hypothetical protein 434184 433831 7.8 13302 - 6.17 -1.01 0.18 17.58516 RB951 234 705 protein containing DUF1596 502459 501755 12.3 25583 - 6.53 -1.37 0.22 16.44622 RB1179 128 387 hypothetical protein 603998 604384 12 13863 + 7.35 -1.12 0.14 12.47655 RB1230 117 354 hypothetical protein 627328 626975 11.7 13093 - 6.73 -1.38 0.21 15.60686 RB1281 230 693 hypothetical protein 653595 654287 10.5 25350 + 6.33 -1.09 0.09 8.05691 RB1291 97 294 hypothetical protein 656684 656977 10.4 10290 + 6.24 -1.16 0.17 14.49980 RB1860 110 333 hypothetical protein 960361 960693 10.9 11827 + 6.37 -1.26 0.02 1.63
1130 RB2150 93 282 hypothetical protein 1138311 1138030 4.4 10755 - 7.18 -1.92 0.39 20.421183 RB2247 579 1740 conserved hypothetical protein containing YTV domains 1181238 1179499 9.8 65013 - 7.17 -1.77 0.32 17.871184 RB2251 75 228 hypothetical protein 1181714 1181487 4.5 8027 - 7.28 -1.64 0.35 21.141827 RB3365 43 132 hypothetical protein 1730124 1730255 11.4 4886 + 8.48 -2.19 0.25 11.342312 RB4194 53 162 hypothetical protein 2156685 2156524 11.4 5728 - 7.53 -1.11 0.12 10.862593 RB4652 52 159 hypothetical protein 2380494 2380652 12.6 5628 + 7.03 -1.34 0.11 8.022596 RB4657 123 372 hypothetical protein 2381865 2382236 12.2 13398 + 6.52 -1.59 0.14 8.883045 RB5409 97 294 hypothetical protein 2794195 2794488 12.7 10898 + 7.61 -2.73 0.08 2.753239 RB5745 130 393 hypothetical protein 3001234 3000842 10.7 13522 - 7.29 -2.96 0.24 8.023356 RB5957 154 465 secreted protein 3122093 3122557 3.3 15514 + 7.51 -1.32 0.12 8.863476 RB6174 69 210 hypothetical protein 3239734 3239525 10.6 7870 - 6.31 -1.18 0.14 12.013485 RB6193 62 189 hypothetical protein 3246096 3245908 11.4 6970 - 6.43 -1.21 0.15 12.233527 RB6274 59 180 hypothetical protein 3297130 3296951 12 7254 - 6.31 -1.14 0.12 10.583725 RB6594 295 888 hypothetical protein 3505472 3506359 10.2 32951 + 6.33 -1.42 0.13 9.463731 RB6609 45 138 hypothetical protein 3511456 3511593 10.4 5004 + 6.73 -1.12 0.15 13.003733 RB6615 230 693 secreted protein containing DUF1080 3515430 3516122 5.1 25770 + 6.54 -1.24 0.13 10.833826 RB6777 137 414 hypothetical protein 3602786 3603199 11.5 15512 + 7.41 -1.07 0.10 9.243836 RB6789 180 543 secreted protein 3608487 3607945 5 20191 - 6.43 -1.36 0.09 6.623901 RB6911 430 1293 protein containing DUF1559 3671445 3672737 6 45752 + 6.46 -1.25 0.21 17.02
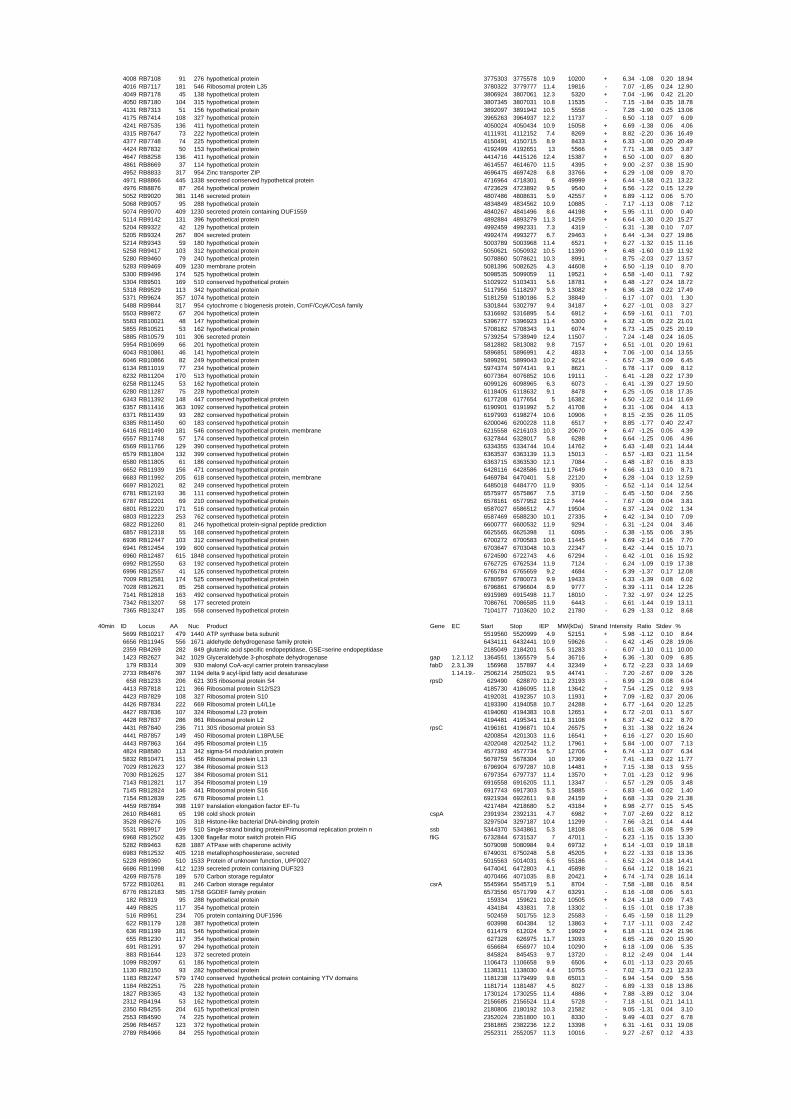
4008 RB7108 91 276 hypothetical protein 3775303 3775578 10.9 10200 + 6.34 -1.08 0.20 18.944016 RB7117 181 546 Ribosomal protein L35 3780322 3779777 11.4 19816 - 7.07 -1.85 0.24 12.904049 RB7178 45 138 hypothetical protein 3806924 3807061 12.3 5320 + 7.04 -1.96 0.42 21.204050 RB7180 104 315 hypothetical protein 3807345 3807031 10.8 11535 - 7.15 -1.84 0.35 18.784131 RB7313 51 156 hypothetical protein 3892097 3891942 10.5 5558 - 7.28 -1.90 0.25 13.084175 RB7414 108 327 hypothetical protein 3965263 3964937 12.2 11737 - 6.50 -1.18 0.07 6.094241 RB7535 136 411 hypothetical protein 4050024 4050434 10.9 15058 + 6.69 -1.38 0.06 4.064315 RB7647 73 222 hypothetical protein 4111931 4112152 7.4 8269 + 8.82 -2.20 0.36 16.494377 RB7748 74 225 hypothetical protein 4150491 4150715 8.9 8433 + 6.33 -1.00 0.20 20.494424 RB7832 50 153 hypothetical protein 4192499 4192651 13 5566 + 7.71 -1.38 0.05 3.874647 RB8258 136 411 hypothetical protein 4414716 4415126 12.4 15387 + 6.50 -1.00 0.07 6.804861 RB8669 37 114 hypothetical protein 4614557 4614670 11.5 4395 + 9.00 -2.37 0.38 15.904952 RB8833 317 954 Zinc transporter ZIP 4696475 4697428 6.8 33766 + 6.29 -1.08 0.09 8.704971 RB8866 445 1338 secreted conserved hypothetical protein 4716964 4718301 6 49999 + 6.44 -1.58 0.21 13.224976 RB8876 87 264 hypothetical protein 4723629 4723892 9.5 9540 + 6.56 -1.22 0.15 12.295052 RB9020 381 1146 secreted protein 4807486 4808631 5.9 42557 + 6.89 -1.12 0.06 5.705068 RB9057 95 288 hypothetical protein 4834849 4834562 10.9 10885 - 7.17 -1.13 0.08 7.125074 RB9070 409 1230 secreted protein containing DUF1559 4840267 4841496 8.6 44198 + 5.95 -1.11 0.00 0.405114 RB9142 131 396 hypothetical protein 4892884 4893279 11.3 14259 + 6.64 -1.30 0.20 15.275204 RB9322 42 129 hypothetical protein 4992459 4992331 7.3 4319 - 6.31 -1.38 0.10 7.075205 RB9324 267 804 secreted protein 4992474 4993277 6.7 29463 + 6.44 -1.34 0.27 19.865214 RB9343 59 180 hypothetical protein 5003789 5003968 11.4 6521 + 6.27 -1.32 0.15 11.165258 RB9417 103 312 hypothetical protein 5050621 5050932 10.5 11390 + 6.48 -1.60 0.19 11.925280 RB9460 79 240 hypothetical protein 5078860 5078621 10.3 8991 - 8.75 -2.03 0.27 13.575283 RB9469 409 1230 membrane protein 5081396 5082625 4.3 44608 + 6.50 -1.19 0.10 8.705300 RB9496 174 525 hypothetical protein 5098535 5099059 11 19521 + 6.58 -1.40 0.11 7.925304 RB9501 169 510 conserved hypothetical protein 5102922 5103431 5.6 18781 + 6.48 -1.27 0.24 18.725318 RB9529 113 342 hypothetical protein 5117956 5118297 9.3 13082 + 6.36 -1.28 0.22 17.495371 RB9624 357 1074 hypothetical protein 5181259 5180186 5.2 38849 - 6.17 -1.07 0.01 1.305488 RB9844 317 954 cytochrome c biogenesis protein, CcmF/CcyK/CcsA family 5301844 5302797 9.4 34187 + 6.27 -1.01 0.03 3.275503 RB9872 67 204 hypothetical protein 5316692 5316895 5.4 6912 + 6.59 -1.61 0.11 7.015583 RB10021 48 147 hypothetical protein 5396777 5396923 11.4 5300 + 6.32 -1.05 0.22 21.015855 RB10521 53 162 hypothetical protein 5708182 5708343 9.1 6074 + 6.73 -1.25 0.25 20.195885 RB10579 101 306 secreted protein 5739254 5738949 12.4 11507 - 7.24 -1.48 0.24 16.055954 RB10699 66 201 hypothetical protein 5812882 5813082 9.8 7157 + 6.51 -1.01 0.20 19.616043 RB10861 46 141 hypothetical protein 5896851 5896991 4.2 4833 + 7.06 -1.00 0.14 13.556046 RB10866 82 249 hypothetical protein 5899291 5899043 10.2 9214 - 6.57 -1.39 0.09 6.456134 RB11019 77 234 hypothetical protein 5974374 5974141 9.1 8621 - 6.78 -1.17 0.09 8.126232 RB11204 170 513 hypothetical protein 6077364 6076852 10.6 19111 - 6.41 -1.28 0.22 17.396258 RB11245 53 162 hypothetical protein 6099126 6098965 6.3 6073 - 6.41 -1.39 0.27 19.506280 RB11287 75 228 hypothetical protein 6118405 6118632 9.1 8478 + 6.25 -1.05 0.18 17.356343 RB11392 148 447 conserved hypothetical protein 6177208 6177654 5 16382 + 6.50 -1.22 0.14 11.696357 RB11416 363 1092 conserved hypothetical protein 6190901 6191992 5.2 41708 + 6.31 -1.06 0.04 4.136371 RB11439 93 282 conserved hypothetical protein 6197993 6198274 10.6 10906 + 8.15 -2.35 0.26 11.056385 RB11450 60 183 conserved hypothetical protein 6200046 6200228 11.8 6517 + 8.85 -1.77 0.40 22.476416 RB11490 181 546 conserved hypothetical protein, membrane 6215558 6216103 10.3 20670 + 6.47 -1.25 0.05 4.396557 RB11748 57 174 conserved hypothetical protein 6327844 6328017 5.8 6288 + 6.64 -1.25 0.06 4.966569 RB11766 129 390 conserved hypothetical protein 6334355 6334744 10.4 14762 + 6.43 -1.48 0.21 14.446579 RB11804 132 399 conserved hypothetical protein 6363537 6363139 11.3 15013 - 6.57 -1.83 0.21 11.546580 RB11805 61 186 conserved hypothetical protein 6363715 6363530 12.1 7084 - 6.48 -1.87 0.16 8.336652 RB11939 156 471 conserved hypothetical protein 6428116 6428586 11.9 17649 + 6.66 -1.13 0.10 8.716683 RB11992 205 618 conserved hypothetical protein, membrane 6469784 6470401 5.8 22120 + 6.28 -1.04 0.13 12.596697 RB12021 82 249 conserved hypothetical protein 6485018 6484770 11.9 9305 - 6.52 -1.14 0.14 12.546781 RB12193 36 111 conserved hypothetical protein 6575977 6575867 7.5 3719 - 6.45 -1.50 0.04 2.566787 RB12201 69 210 conserved hypothetical protein 6578161 6577952 12.5 7444 - 7.67 -1.09 0.04 3.816801 RB12220 171 516 conserved hypothetical protein 6587027 6586512 4.7 19504 - 6.37 -1.24 0.02 1.346803 RB12223 253 762 conserved hypothetical protein 6587469 6588230 10.1 27335 + 6.42 -1.34 0.10 7.096822 RB12260 81 246 hypothetical protein-signal peptide prediction 6600777 6600532 11.9 9294 - 6.31 -1.24 0.04 3.466857 RB12318 55 168 conserved hypothetical protein 6625565 6625398 11 6095 - 6.38 -1.55 0.06 3.956936 RB12447 103 312 conserved hypothetical protein 6700272 6700583 10.6 11445 + 6.69 -2.14 0.16 7.706941 RB12454 199 600 conserved hypothetical protein 6703647 6703048 10.3 22347 - 6.42 -1.44 0.15 10.716960 RB12487 615 1848 conserved hypothetical protein 6724590 6722743 4.6 67294 - 6.42 -1.01 0.16 15.926992 RB12550 63 192 conserved hypothetical protein 6762725 6762534 11.9 7124 - 6.24 -1.09 0.19 17.386996 RB12557 41 126 conserved hypothetical protein 6765784 6765659 9.2 4684 - 6.39 -1.37 0.17 12.087009 RB12581 174 525 conserved hypothetical protein 6780597 6780073 9.9 19433 - 6.33 -1.39 0.08 6.027028 RB12621 85 258 conserved hypothetical protein 6796861 6796604 8.9 9777 - 6.39 -1.11 0.14 12.267141 RB12818 163 492 conserved hypothetical protein 6915989 6915498 11.7 18010 - 7.32 -1.97 0.24 12.257342 RB13207 58 177 secreted protein 7086761 7086585 11.9 6443 - 6.61 -1.44 0.19 13.117365 RB13247 185 558 conserved hypothetical protein 7104177 7103620 10.2 21780 - 6.29 -1.33 0.12 8.68
40min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand Intensity Ratio Stdev %5699 RB10217 479 1440 ATP synthase beta subunit 5519560 5520999 4.9 52151 + 5.98 -1.12 0.10 8.646656 RB11945 556 1671 aldehyde dehydrogenase family protein 6434111 6432441 10.9 59626 - 6.42 -1.45 0.28 19.062359 RB4269 282 849 glutamic acid specific endopeptidase, GSE=serine endopeptidase 2185049 2184201 5.6 31283 - 6.07 -1.10 0.11 10.001423 RB2627 342 1029 Glyceraldehyde 3-phosphate dehydrogenase gap 1.2.1.12 1364551 1365579 5.4 36716 + 6.36 -1.30 0.09 6.85179 RB314 309 930 malonyl CoA-acyl carrier protein transacylase fabD 2.3.1.39 156968 157897 4.4 32349 + 6.72 -2.23 0.33 14.69
2733 RB4876 397 1194 delta 9 acyl-lipid fatty acid desaturase 1.14.19.- 2506214 2505021 9.5 44741 - 7.20 -2.67 0.09 3.26658 RB1233 206 621 30S ribosomal protein S4 rpsD 629490 628870 11.2 23193 - 6.99 -1.29 0.08 6.04
4413 RB7818 121 366 Ribosomal protein S12/S23 4185730 4186095 11.8 13642 + 7.54 -1.25 0.12 9.934423 RB7829 108 327 Ribosomal protein S10 4192031 4192357 10.3 11931 + 7.09 -1.82 0.37 20.064426 RB7834 222 669 Ribosomal protein L4/L1e 4193390 4194058 10.7 24288 + 6.77 -1.64 0.20 12.254427 RB7836 107 324 Ribosomal L23 protein 4194060 4194383 10.8 12651 + 6.72 -2.01 0.11 5.674428 RB7837 286 861 Ribosomal protein L2 4194481 4195341 11.8 31108 + 6.37 -1.42 0.12 8.704431 RB7840 236 711 30S ribosomal protein S3 rpsC 4196161 4196871 10.4 26575 + 6.31 -1.38 0.22 16.244441 RB7857 149 450 Ribosomal protein L18P/L5E 4200854 4201303 11.6 16541 + 6.16 -1.27 0.20 15.604443 RB7863 164 495 Ribosomal protein L15 4202048 4202542 11.2 17961 + 5.84 -1.00 0.07 7.134824 RB8580 113 342 sigma-54 modulation protein 4577393 4577734 5.7 12706 + 6.74 -1.13 0.07 6.345832 RB10471 151 456 Ribosomal protein L13 5678759 5678304 10 17369 - 7.41 -1.83 0.22 11.777029 RB12623 127 384 Ribosomal protein S13 6796904 6797287 10.8 14481 + 7.15 -1.38 0.13 9.557030 RB12625 127 384 Ribosomal protein S11 6797354 6797737 11.4 13570 + 7.01 -1.23 0.12 9.967143 RB12821 117 354 Ribosomal protein L19 6916558 6916205 11.1 13347 - 6.57 -1.29 0.05 3.487145 RB12824 146 441 Ribosomal protein S16 6917743 6917303 5.3 15885 - 6.83 -1.46 0.02 1.407154 RB12839 225 678 Ribosomal protein L1 6921934 6922611 9.8 24159 + 6.68 -1.33 0.29 21.384459 RB7894 398 1197 translation elongation factor EF-Tu 4217484 4218680 5.2 43184 + 6.98 -2.77 0.15 5.452610 RB4681 65 198 cold shock protein cspA 2391934 2392131 4.7 6982 + 7.07 -2.69 0.22 8.123528 RB6276 105 318 Histone-like bacterial DNA-binding protein 3297504 3297187 10.4 11299 - 7.66 -3.21 0.14 4.445531 RB9917 169 510 Single-strand binding protein/Primosomal replication protein n ssb 5344370 5343861 5.3 18108 - 6.81 -1.36 0.08 5.996968 RB12502 435 1308 flagellar motor switch protein FliG fliG 6732844 6731537 7 47011 - 6.23 -1.15 0.15 13.305282 RB9463 628 1887 ATPase with chaperone activity 5079098 5080984 9.4 69732 + 6.14 -1.03 0.19 18.186983 RB12532 405 1218 metallophosphoesterase, secreted 6749031 6750248 5.8 45205 + 6.22 -1.33 0.18 13.365228 RB9360 510 1533 Protein of unknown function, UPF0027 5015563 5014031 6.5 55186 - 6.52 -1.24 0.18 14.416686 RB11998 412 1239 secreted protein containing DUF323 6474041 6472803 4.1 45898 - 6.64 -1.12 0.18 16.214269 RB7578 189 570 Carbon storage regulator 4070466 4071035 8.8 20421 + 6.74 -1.74 0.28 16.145722 RB10261 81 246 Carbon storage regulator csrA 5545964 5545719 5.1 8704 - 7.58 -1.88 0.16 8.546776 RB12183 585 1758 GGDEF family protein 6573556 6571799 4.7 63291 - 6.16 -1.08 0.06 5.61182 RB319 95 288 hypothetical protein 159334 159621 10.2 10505 + 6.24 -1.18 0.09 7.43449 RB825 117 354 hypothetical protein 434184 433831 7.8 13302 - 6.15 -1.01 0.18 17.38516 RB951 234 705 protein containing DUF1596 502459 501755 12.3 25583 - 6.45 -1.59 0.18 11.29622 RB1179 128 387 hypothetical protein 603998 604384 12 13863 + 7.17 -1.11 0.03 2.42636 RB1199 181 546 hypothetical protein 611479 612024 5.7 19929 + 6.18 -1.11 0.24 21.96655 RB1230 117 354 hypothetical protein 627328 626975 11.7 13093 - 6.65 -1.26 0.20 15.90691 RB1291 97 294 hypothetical protein 656684 656977 10.4 10290 + 6.18 -1.09 0.06 5.35883 RB1644 123 372 secreted protein 845824 845453 9.7 13720 - 8.12 -2.49 0.04 1.44
1099 RB2097 61 186 hypothetical protein 1106473 1106658 9.9 6506 + 6.01 -1.13 0.23 20.651130 RB2150 93 282 hypothetical protein 1138311 1138030 4.4 10755 - 7.02 -1.73 0.21 12.331183 RB2247 579 1740 conserved hypothetical protein containing YTV domains 1181238 1179499 9.8 65013 - 6.94 -1.54 0.09 5.561184 RB2251 75 228 hypothetical protein 1181714 1181487 4.5 8027 - 6.89 -1.33 0.18 13.861827 RB3365 43 132 hypothetical protein 1730124 1730255 11.4 4886 + 7.88 -3.89 0.12 3.042312 RB4194 53 162 hypothetical protein 2156685 2156524 11.4 5728 - 7.18 -1.51 0.21 14.112350 RB4255 204 615 hypothetical protein 2180806 2180192 10.3 21582 - 9.05 -1.31 0.04 3.102553 RB4590 74 225 hypothetical protein 2352024 2351800 10.1 8330 - 9.49 -4.03 0.27 6.782596 RB4657 123 372 hypothetical protein 2381865 2382236 12.2 13398 + 6.31 -1.61 0.31 19.082789 RB4966 84 255 hypothetical protein 2552311 2552057 11.3 10016 - 9.27 -2.67 0.12 4.33
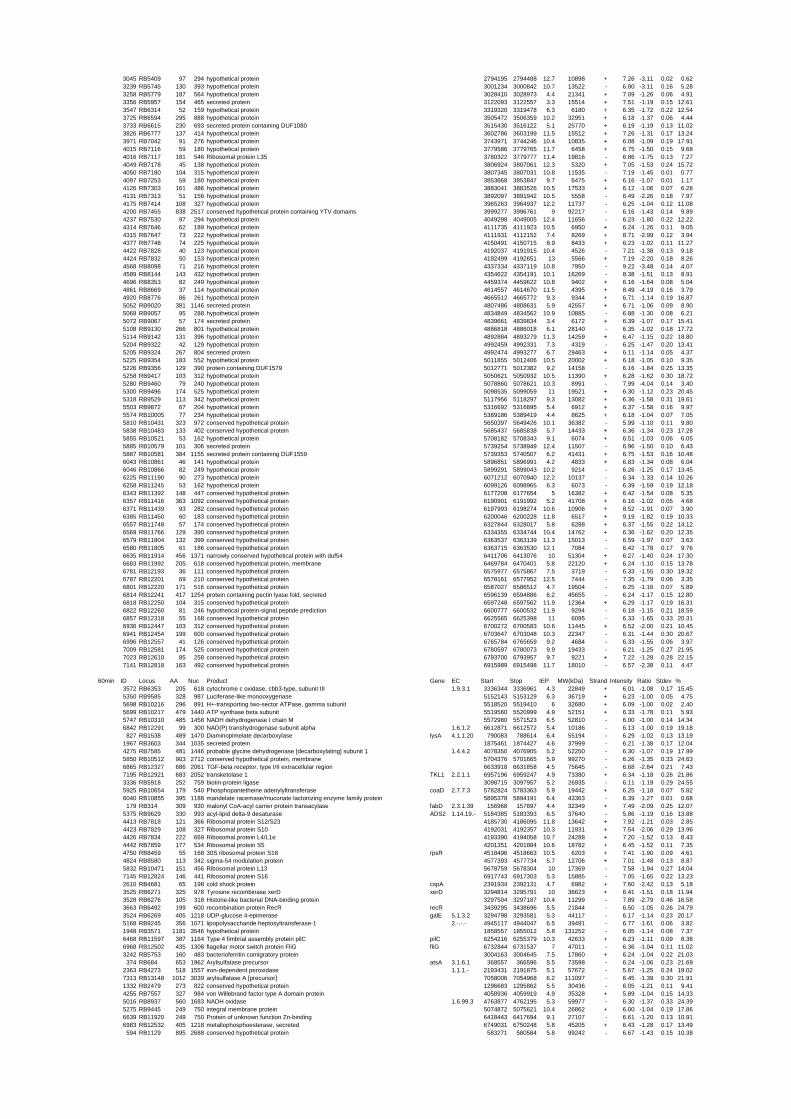
3045 RB5409 97 294 hypothetical protein 2794195 2794488 12.7 10898 + 7.26 -3.11 0.02 0.623239 RB5745 130 393 hypothetical protein 3001234 3000842 10.7 13522 - 6.80 -3.11 0.16 5.283258 RB5779 187 564 hypothetical protein 3028410 3028973 4.4 21341 + 7.09 -1.26 0.06 4.913356 RB5957 154 465 secreted protein 3122093 3122557 3.3 15514 + 7.51 -1.19 0.15 12.613547 RB6314 52 159 hypothetical protein 3319320 3319478 6.3 6180 + 6.35 -1.72 0.22 12.543725 RB6594 295 888 hypothetical protein 3505472 3506359 10.2 32951 + 6.18 -1.37 0.06 4.443733 RB6615 230 693 secreted protein containing DUF1080 3515430 3516122 5.1 25770 + 6.19 -1.19 0.13 11.023826 RB6777 137 414 hypothetical protein 3602786 3603199 11.5 15512 + 7.26 -1.31 0.17 13.243971 RB7042 91 276 hypothetical protein 3743971 3744246 10.4 10835 + 6.08 -1.09 0.19 17.914015 RB7116 59 180 hypothetical protein 3779586 3779765 11.7 6458 + 6.75 -1.50 0.15 9.684016 RB7117 181 546 Ribosomal protein L35 3780322 3779777 11.4 19816 - 6.86 -1.75 0.13 7.274049 RB7178 45 138 hypothetical protein 3806924 3807061 12.3 5320 + 7.05 -1.53 0.24 15.724050 RB7180 104 315 hypothetical protein 3807345 3807031 10.8 11535 - 7.19 -1.45 0.01 0.774097 RB7253 59 180 hypothetical protein 3853668 3853847 9.7 6475 + 6.16 -1.07 0.01 1.174126 RB7303 161 486 hypothetical protein 3883041 3883526 10.5 17533 + 6.12 -1.06 0.07 6.284131 RB7313 51 156 hypothetical protein 3892097 3891942 10.5 5558 - 6.49 -2.26 0.18 7.974175 RB7414 108 327 hypothetical protein 3965263 3964937 12.2 11737 - 6.25 -1.04 0.12 11.084200 RB7455 838 2517 conserved hypothetical protein containing YTV domains 3999277 3996761 9 92217 - 6.16 -1.43 0.14 9.894237 RB7530 97 294 hypothetical protein 4049298 4049005 12.4 11656 - 6.23 -1.80 0.22 12.224314 RB7646 62 189 hypothetical protein 4111735 4111923 10.5 6950 + 6.24 -1.26 0.11 9.054315 RB7647 73 222 hypothetical protein 4111931 4112152 7.4 8269 + 8.71 -2.99 0.12 3.944377 RB7748 74 225 hypothetical protein 4150491 4150715 8.9 8433 + 6.23 -1.02 0.11 11.274422 RB7828 40 123 hypothetical protein 4192037 4191915 10.4 4526 - 7.21 -1.38 0.13 9.184424 RB7832 50 153 hypothetical protein 4192499 4192651 13 5566 + 7.19 -2.20 0.18 8.264568 RB8098 71 216 hypothetical protein 4337334 4337119 10.8 7950 - 9.22 -3.48 0.14 4.074589 RB8144 143 432 hypothetical protein 4354622 4354191 10.1 16269 - 8.38 -1.51 0.13 8.914696 RB8353 82 249 hypothetical protein 4459374 4459622 10.8 9402 + 6.16 -1.64 0.08 5.044861 RB8669 37 114 hypothetical protein 4614557 4614670 11.5 4395 + 8.49 -4.19 0.16 3.794920 RB8776 86 261 hypothetical protein 4665512 4665772 9.3 9344 + 6.71 -1.14 0.19 16.875052 RB9020 381 1146 secreted protein 4807486 4808631 5.9 42557 + 6.71 -1.06 0.09 8.905068 RB9057 95 288 hypothetical protein 4834849 4834562 10.9 10885 - 6.88 -1.30 0.08 6.215072 RB9067 57 174 secreted protein 4839661 4839834 3.4 6172 + 6.39 -1.07 0.17 15.415108 RB9130 266 801 hypothetical protein 4886818 4886018 6.1 28140 - 6.35 -1.02 0.18 17.725114 RB9142 131 396 hypothetical protein 4892884 4893279 11.3 14259 + 6.47 -1.15 0.22 18.805204 RB9322 42 129 hypothetical protein 4992459 4992331 7.3 4319 - 6.25 -1.47 0.20 13.415205 RB9324 267 804 secreted protein 4992474 4993277 6.7 29463 + 6.11 -1.14 0.05 4.375225 RB9354 183 552 hypothetical protein 5011855 5012406 10.5 20002 + 6.18 -1.05 0.10 9.355226 RB9356 129 390 protein containing DUF1579 5012771 5012382 9.2 14158 - 6.16 -1.84 0.25 13.355258 RB9417 103 312 hypothetical protein 5050621 5050932 10.5 11390 + 6.28 -1.62 0.30 18.725280 RB9460 79 240 hypothetical protein 5078860 5078621 10.3 8991 - 7.99 -4.04 0.14 3.405300 RB9496 174 525 hypothetical protein 5098535 5099059 11 19521 + 6.30 -1.12 0.23 20.455318 RB9529 113 342 hypothetical protein 5117956 5118297 9.3 13082 + 6.36 -1.58 0.31 19.615503 RB9872 67 204 hypothetical protein 5316692 5316895 5.4 6912 + 6.37 -1.58 0.16 9.975574 RB10005 77 234 hypothetical protein 5389186 5389419 4.4 8625 + 6.18 -1.04 0.07 7.055810 RB10431 323 972 conserved hypothetical protein 5650397 5649426 10.1 36382 - 5.99 -1.10 0.11 9.805838 RB10483 133 402 conserved hypothetical protein 5685437 5685838 5.7 14433 + 6.36 -1.34 0.23 17.285855 RB10521 53 162 hypothetical protein 5708182 5708343 9.1 6074 + 6.51 -1.03 0.06 6.055885 RB10579 101 306 secreted protein 5739254 5738949 12.4 11507 - 6.96 -1.50 0.10 6.435887 RB10581 384 1155 secreted protein containing DUF1559 5739353 5740507 6.2 41431 + 6.75 -1.53 0.16 10.486043 RB10861 46 141 hypothetical protein 5896851 5896991 4.2 4833 + 6.83 -1.34 0.08 6.046046 RB10866 82 249 hypothetical protein 5899291 5899043 10.2 9214 - 6.26 -1.25 0.17 13.456225 RB11190 90 273 hypothetical protein 6071212 6070940 12.2 10137 - 6.34 -1.33 0.14 10.266258 RB11245 53 162 hypothetical protein 6099126 6098965 6.3 6073 - 6.39 -1.59 0.19 12.186343 RB11392 148 447 conserved hypothetical protein 6177208 6177654 5 16382 + 6.42 -1.54 0.08 5.356357 RB11416 363 1092 conserved hypothetical protein 6190901 6191992 5.2 41708 + 6.16 -1.02 0.05 4.686371 RB11439 93 282 conserved hypothetical protein 6197993 6198274 10.6 10906 + 8.52 -1.91 0.07 3.906385 RB11450 60 183 conserved hypothetical protein 6200046 6200228 11.8 6517 + 9.19 -1.82 0.19 10.336557 RB11748 57 174 conserved hypothetical protein 6327844 6328017 5.8 6288 + 6.37 -1.55 0.22 14.126569 RB11766 129 390 conserved hypothetical protein 6334355 6334744 10.4 14762 + 6.36 -1.62 0.20 12.356579 RB11804 132 399 conserved hypothetical protein 6363537 6363139 11.3 15013 - 6.59 -1.97 0.07 3.636580 RB11805 61 186 conserved hypothetical protein 6363715 6363530 12.1 7084 - 6.42 -1.78 0.17 9.766635 RB11914 456 1371 narrowly conserved hypothetical protein with duf54 6411706 6413076 10 51304 + 6.27 -1.40 0.24 17.306683 RB11992 205 618 conserved hypothetical protein, membrane 6469784 6470401 5.8 22120 + 6.24 -1.10 0.15 13.786781 RB12193 36 111 conserved hypothetical protein 6575977 6575867 7.5 3719 - 6.33 -1.55 0.30 19.326787 RB12201 69 210 conserved hypothetical protein 6578161 6577952 12.5 7444 - 7.35 -1.79 0.06 3.356801 RB12220 171 516 conserved hypothetical protein 6587027 6586512 4.7 19504 - 6.25 -1.16 0.07 5.896814 RB12241 417 1254 protein containing pectin lyase fold, secreted 6596139 6594886 6.2 45655 - 6.24 -1.17 0.15 12.806818 RB12250 104 315 conserved hypothetical protein 6597248 6597562 11.9 12364 + 6.29 -1.17 0.19 16.316822 RB12260 81 246 hypothetical protein-signal peptide prediction 6600777 6600532 11.9 9294 - 6.18 -1.15 0.21 18.596857 RB12318 55 168 conserved hypothetical protein 6625565 6625398 11 6095 - 6.33 -1.65 0.33 20.316936 RB12447 103 312 conserved hypothetical protein 6700272 6700583 10.6 11445 + 6.52 -2.00 0.21 10.456941 RB12454 199 600 conserved hypothetical protein 6703647 6703048 10.3 22347 - 6.31 -1.44 0.30 20.676996 RB12557 41 126 conserved hypothetical protein 6765784 6765659 9.2 4684 - 6.33 -1.55 0.06 3.977009 RB12581 174 525 conserved hypothetical protein 6780597 6780073 9.9 19433 - 6.21 -1.25 0.27 21.957023 RB12610 85 258 conserved hypothetical protein 6793700 6793957 9.7 9221 + 7.22 -1.28 0.28 22.157141 RB12818 163 492 conserved hypothetical protein 6915989 6915498 11.7 18010 - 6.57 -2.38 0.11 4.47
60min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand Intensity Ratio Stdev %3572 RB6353 205 618 cytochrome c oxidase, cbb3-type, subunit III 1.9.3.1 3336344 3336961 4.3 22849 + 6.01 -1.08 0.17 15.455350 RB9585 328 987 Luciferase-like monooxygenase 5152143 5153129 6.3 36719 + 6.23 -1.00 0.05 4.755698 RB10216 296 891 H+-transporting two-sector ATPase, gamma subunit 5518520 5519410 6 32680 + 6.09 -1.00 0.02 2.405699 RB10217 479 1440 ATP synthase beta subunit 5519560 5520999 4.9 52151 + 6.33 -1.78 0.11 5.935747 RB10310 485 1458 NADH dehydrogenase I chain M 5572980 5571523 6.5 52810 - 6.00 -1.00 0.14 14.346842 RB12291 99 300 NAD(P) transhydrogenase subunit alpha 1.6.1.2 6612871 6612572 5.4 10186 - 6.13 -1.00 0.19 19.18827 RB1538 489 1470 Diaminopimelate decarboxylase lysA 4.1.1.20 790083 788614 6.4 55194 - 6.29 -1.02 0.13 13.19
1967 RB3603 344 1035 secreted protein 1875461 1874427 4.6 37999 - 6.21 -1.38 0.17 12.044275 RB7585 481 1446 probable glycine dehydrogenase [decarboxylating] subunit 1 1.4.4.2 4078350 4076905 5.2 52250 - 6.30 -1.07 0.19 17.995850 RB10512 903 2712 conserved hypothetical protein, membrane 5704376 5701665 5.9 99270 - 6.26 -1.35 0.33 24.636865 RB12327 686 2061 TGF-beta receptor, type I/II extracellular region 6633918 6631858 4.5 75645 - 6.68 -2.84 0.21 7.437195 RB12921 683 2052 transketolase 1 TKL1 2.2.1.1 6957196 6959247 4.9 73380 + 6.34 -1.18 0.26 21.863336 RB5918 252 759 biotin-protein ligase 3098715 3097957 5.2 26935 - 6.11 -1.19 0.29 24.555925 RB10654 179 540 Phosphopantetheine adenylyltransferase coaD 2.7.7.3 5782824 5783363 5.9 19442 + 6.25 -1.18 0.07 5.826040 RB10855 395 1188 mandelate racemase/muconate lactonizing enzyme family protein 5895378 5894191 6.4 43363 - 6.39 -1.27 0.01 0.68179 RB314 309 930 malonyl CoA-acyl carrier protein transacylase fabD 2.3.1.39 156968 157897 4.4 32349 + 7.49 -2.09 0.25 12.07
5375 RB9629 330 993 acyl-lipid delta-9 desaturase ADS2 1.14.19.- 5184385 5183393 6.5 37640 - 5.86 -1.19 0.16 13.884413 RB7818 121 366 Ribosomal protein S12/S23 4185730 4186095 11.8 13642 + 7.92 -1.21 0.03 2.854423 RB7829 108 327 Ribosomal protein S10 4192031 4192357 10.3 11931 + 7.54 -2.06 0.29 13.964426 RB7834 222 669 Ribosomal protein L4/L1e 4193390 4194058 10.7 24288 + 7.20 -1.52 0.13 8.434442 RB7859 177 534 Ribosomal protein S5 4201351 4201884 10.6 18782 + 6.45 -1.52 0.11 7.354750 RB8459 55 168 30S ribosomal protein S18 rpsR 4518496 4518663 10.5 6203 + 7.41 -1.90 0.09 4.614824 RB8580 113 342 sigma-54 modulation protein 4577393 4577734 5.7 12706 + 7.01 -1.48 0.13 8.875832 RB10471 151 456 Ribosomal protein L13 5678759 5678304 10 17369 - 7.58 -1.94 0.27 14.047145 RB12824 146 441 Ribosomal protein S16 6917743 6917303 5.3 15885 - 7.05 -1.65 0.22 13.232610 RB4681 65 198 cold shock protein cspA 2391934 2392131 4.7 6982 + 7.60 -2.42 0.13 5.183525 RB6271 325 978 Tyrosine recombinase xerD xerD 3294814 3295791 10 36623 + 6.41 -1.51 0.18 11.943528 RB6276 105 318 Histone-like bacterial DNA-binding protein 3297504 3297187 10.4 11299 - 7.89 -2.79 0.46 16.583663 RB6492 199 600 recombination protein RecR recR 3439295 3438696 5.5 21844 - 6.50 -1.05 0.26 24.793524 RB6269 405 1218 UDP-glucose 4-epimerase galE 5.1.3.2 3294798 3293581 5.3 44117 - 6.17 -1.14 0.23 20.175168 RB9245 356 1071 lipopolysaccharide heptosyltransferase-1 2.-.-.- 4945117 4944047 6.5 39491 - 6.77 -1.61 0.06 3.821948 RB3571 1181 3546 hypothetical protein 1858557 1855012 5.8 131252 - 6.05 -1.14 0.08 7.376468 RB11597 387 1164 Type 4 fimbrial assembly protein pilC pilC 6254216 6255379 10.3 42633 + 6.23 -1.11 0.09 8.386968 RB12502 435 1308 flagellar motor switch protein FliG fliG 6732844 6731537 7 47011 - 6.36 -1.04 0.11 11.023242 RB5753 160 483 bacterioferritin comigratory protein 3004163 3004645 7.5 17860 + 6.24 -1.04 0.22 21.03374 RB684 653 1962 Arylsulfatase precursor atsA 3.1.6.1 368557 366596 5.5 73598 - 6.24 -1.06 0.23 21.69
2363 RB4273 518 1557 iron-dependent peroxidase 1.1.1.- 2193431 2191875 5.1 57672 - 5.67 -1.25 0.24 19.027313 RB13148 1012 3039 arylsulfatase A [precursor] 7058006 7054968 6.2 111097 - 6.45 -1.39 0.30 21.911332 RB2479 273 822 conserved hypothetical protein 1296683 1295862 5.5 30436 - 6.05 -1.21 0.11 9.414255 RB7557 327 984 von Willebrand factor type A domain protein 4058936 4059919 4.9 35328 + 5.89 -1.04 0.15 14.335016 RB8937 560 1683 NADH oxidase 1.6.99.3 4763877 4762195 5.3 59977 - 6.30 -1.37 0.33 24.395275 RB9445 249 750 integral membrane protein 5074872 5075621 10.4 26862 + 6.00 -1.04 0.19 17.866639 RB11920 249 750 Protein of unknown function Zn-binding 6418443 6417694 9.1 27107 - 6.61 -1.20 0.13 10.916983 RB12532 405 1218 metallophosphoesterase, secreted 6749031 6750248 5.8 45205 + 6.43 -1.28 0.17 13.49594 RB1129 895 2688 conserved hypothetical protein 583271 580584 5.8 99242 - 6.67 -1.43 0.15 10.38

5740 RB10297 196 591 formaldehyde activating enzyme fae 5563081 5562491 5.5 20967 - 6.44 -1.24 0.28 22.534269 RB7578 189 570 Carbon storage regulator 4070466 4071035 8.8 20421 + 7.06 -1.94 0.30 15.584477 RB7919 793 2382 serine/threonine protein kinase 4231818 4229437 5.4 87667 - 6.23 -1.44 0.22 15.425722 RB10261 81 246 Carbon storage regulator csrA 5545964 5545719 5.1 8704 - 7.78 -1.69 0.42 24.786071 RB10907 252 759 conserved hypothetical protein 5920466 5919708 4.5 28547 - 6.21 -1.00 0.16 16.076223 RB11185 303 912 universal stress protein family 6069342 6068431 5.4 33738 - 6.39 -1.57 0.17 10.69
2 RB2 59 180 secreted protein 182 3 3.7 6489 - 7.28 -3.23 0.43 13.2156 RB95 157 474 conserved hypothetical protein, membrane 40090 39617 9.6 17913 - 6.13 -1.06 0.14 12.8770 RB125 297 894 membrane protein 54571 53678 6 32981 - 6.25 -1.15 0.02 1.55
182 RB319 95 288 hypothetical protein 159334 159621 10.2 10505 + 6.49 -1.37 0.23 16.42250 RB434 65 198 hypothetical protein 231791 231988 7.4 7385 + 6.40 -1.08 0.23 21.37472 RB869 71 216 hypothetical protein 460848 461063 4.4 8070 + 6.27 -1.60 0.37 23.29553 RB1036 480 1443 protein conserved in bacteria 539939 541381 5.8 51719 + 6.23 -1.39 0.30 21.50636 RB1199 181 546 hypothetical protein 611479 612024 5.7 19929 + 6.30 -1.19 0.20 17.02852 RB1588 151 456 ACP-like protein 817362 817817 5 17451 + 6.57 -1.14 0.22 19.59883 RB1644 123 372 secreted protein 845824 845453 9.7 13720 - 8.37 -2.54 0.17 6.67980 RB1860 110 333 hypothetical protein 960361 960693 10.9 11827 + 6.28 -1.37 0.24 17.17
1130 RB2150 93 282 hypothetical protein 1138311 1138030 4.4 10755 - 7.50 -2.08 0.26 12.511183 RB2247 579 1740 conserved hypothetical protein containing YTV domains 1181238 1179499 9.8 65013 - 7.62 -1.37 0.13 9.501472 RB2714 302 909 protein containing DUF541 1415817 1414909 9.5 33087 - 6.35 -1.17 0.12 10.161660 RB3057 72 219 hypothetical protein 1579920 1580138 10.8 8044 + 6.25 -1.07 0.20 18.821676 RB3097 117 354 hypothetical protein 1606459 1606106 11.3 12852 - 7.20 -1.09 0.02 1.991701 RB3147 168 507 membrane protein 1624840 1624334 8.7 18759 - 6.07 -1.02 0.07 7.201825 RB3362 87 264 hypothetical protein 1728546 1728283 11.9 10017 - 6.14 -1.08 0.18 16.461827 RB3365 43 132 hypothetical protein 1730124 1730255 11.4 4886 + 8.51 -2.76 0.21 7.651844 RB3394 36 111 hypothetical protein 1752044 1752154 10.5 4239 + 5.92 -1.10 0.22 19.881845 RB3395 398 1197 secreted protein containing DUF1559 1752151 1753347 7.8 43136 + 6.19 -1.25 0.21 16.571980 RB3623 60 183 hypothetical protein 1886670 1886488 6.1 6661 - 5.83 -1.12 0.18 16.072200 RB3981 161 486 hypothetical protein 2058000 2058485 4.1 18283 + 6.14 -1.02 0.19 18.252312 RB4194 53 162 hypothetical protein 2156685 2156524 11.4 5728 - 7.47 -1.86 0.40 21.412350 RB4255 204 615 hypothetical protein 2180806 2180192 10.3 21582 - 9.17 -1.16 0.16 14.062354 RB4260 262 789 membrane protein 2181703 2182491 10 27432 + 6.24 -1.06 0.19 17.672369 RB4283 499 1500 PvdD/PvdJ 2201462 2199963 5.7 56411 - 6.09 -1.30 0.19 14.972528 RB4547 422 1269 hypothetical protein 2330116 2328848 7.6 46274 - 5.66 -1.05 0.14 13.382553 RB4590 74 225 hypothetical protein 2352024 2351800 10.1 8330 - 9.62 -3.10 0.58 18.732593 RB4652 52 159 hypothetical protein 2380494 2380652 12.6 5628 + 7.17 -1.16 0.09 7.692596 RB4657 123 372 hypothetical protein 2381865 2382236 12.2 13398 + 6.34 -1.64 0.08 5.142746 RB4897 184 555 hypothetical protein 2519505 2518951 10.9 20446 - 5.83 -1.12 0.16 14.492789 RB4966 84 255 hypothetical protein 2552311 2552057 11.3 10016 - 9.22 -2.29 0.41 17.732964 RB5262 95 288 membrane protein 2713742 2714029 6.3 10993 + 6.09 -1.18 0.27 23.233239 RB5745 130 393 hypothetical protein 3001234 3000842 10.7 13522 - 7.22 -3.11 0.28 8.903258 RB5779 187 564 hypothetical protein 3028410 3028973 4.4 21341 + 7.41 -1.29 0.12 9.083271 RB5802 156 471 hypothetical protein 3039864 3040334 10.1 17001 + 6.68 -1.07 0.26 24.263356 RB5957 154 465 secreted protein 3122093 3122557 3.3 15514 + 7.79 -1.21 0.28 23.573430 RB6092 361 1086 Peptidase M50 3.4.24.- 3192387 3193472 9.6 39370 + 6.06 -1.09 0.13 11.783466 RB6158 142 429 hypothetical protein 3233532 3233104 6 15746 - 6.27 -1.19 0.04 3.443476 RB6174 69 210 hypothetical protein 3239734 3239525 10.6 7870 - 6.24 -1.16 0.03 2.253725 RB6594 295 888 hypothetical protein 3505472 3506359 10.2 32951 + 6.23 -1.06 0.20 19.213780 RB6699 47 144 hypothetical protein 3559781 3559924 11.1 5290 + 5.94 -1.03 0.15 14.123802 RB6734 51 156 hypothetical protein 3577353 3577198 9 5639 - 6.35 -1.60 0.25 15.503827 RB6778 59 180 hypothetical protein 3603142 3603321 5.5 6533 + 6.60 -1.00 0.21 21.133836 RB6789 180 543 secreted protein 3608487 3607945 5 20191 - 6.53 -1.67 0.16 9.383921 RB6951 1120 3363 hypothetical protein 3693377 3690015 4.6 123187 - 6.38 -1.11 0.21 18.973971 RB7042 91 276 hypothetical protein 3743971 3744246 10.4 10835 + 6.23 -1.19 0.22 18.534008 RB7108 91 276 hypothetical protein 3775303 3775578 10.9 10200 + 6.40 -1.39 0.32 22.894016 RB7117 181 546 Ribosomal protein L35 3780322 3779777 11.4 19816 - 7.18 -1.88 0.04 2.264049 RB7178 45 138 hypothetical protein 3806924 3807061 12.3 5320 + 7.21 -1.78 0.44 24.624050 RB7180 104 315 hypothetical protein 3807345 3807031 10.8 11535 - 7.34 -1.58 0.35 22.404175 RB7414 108 327 hypothetical protein 3965263 3964937 12.2 11737 - 6.41 -1.90 0.35 18.714315 RB7647 73 222 hypothetical protein 4111931 4112152 7.4 8269 + 9.15 -2.21 0.18 8.354377 RB7748 74 225 hypothetical protein 4150491 4150715 8.9 8433 + 6.44 -1.25 0.07 5.334424 RB7832 50 153 hypothetical protein 4192499 4192651 13 5566 + 7.75 -1.99 0.41 20.424568 RB8098 71 216 hypothetical protein 4337334 4337119 10.8 7950 - 9.48 -2.79 0.29 10.414589 RB8144 143 432 hypothetical protein 4354622 4354191 10.1 16269 - 8.80 -1.44 0.17 11.634669 RB8301 342 1029 membrane protein 4436412 4435384 9.7 37525 - 5.72 -1.13 0.23 20.524759 RB8474 180 543 hypothetical protein 4524096 4523554 9.3 20951 - 6.24 -1.21 0.21 17.064785 RB8518 370 1113 hypothetical protein 4543984 4545096 11.5 41176 + 6.24 -1.06 0.04 3.524830 RB8594 41 126 hypothetical protein 4581645 4581770 9.2 4509 + 6.17 -1.40 0.11 7.514861 RB8669 37 114 hypothetical protein 4614557 4614670 11.5 4395 + 8.68 -3.89 0.39 10.154920 RB8776 86 261 hypothetical protein 4665512 4665772 9.3 9344 + 7.06 -1.50 0.34 22.514952 RB8833 317 954 Zinc transporter ZIP 4696475 4697428 6.8 33766 + 6.39 -1.88 0.38 20.335044 RB9009 257 774 secreted protein 4799393 4800166 6 29870 + 6.33 -1.33 0.15 11.035068 RB9057 95 288 hypothetical protein 4834849 4834562 10.9 10885 - 7.36 -1.46 0.13 9.205114 RB9142 131 396 hypothetical protein 4892884 4893279 11.3 14259 + 6.78 -1.48 0.34 23.075117 RB9146 552 1659 conserved hypothetical protein, secreted 4895339 4893681 5.8 60781 - 6.11 -1.15 0.05 3.945156 RB9228 637 1914 hypothetical protein 4934679 4936592 7.8 73174 + 6.34 -1.00 0.11 10.845205 RB9324 267 804 secreted protein 4992474 4993277 6.7 29463 + 6.25 -1.94 0.32 16.215225 RB9354 183 552 hypothetical protein 5011855 5012406 10.5 20002 + 6.42 -1.03 0.23 22.225226 RB9356 129 390 protein containing DUF1579 5012771 5012382 9.2 14158 - 7.08 -1.73 0.26 15.035236 RB9376 1827 5484 protein containing cadherin domain 5029102 5023619 3.8 197136 - 6.18 -1.03 0.18 17.395280 RB9460 79 240 hypothetical protein 5078860 5078621 10.3 8991 - 8.38 -3.67 0.28 7.615313 RB9520 532 1599 conserved hypothetical protein 5114495 5112897 4.4 59231 - 6.14 -1.08 0.04 4.105446 RB9768 52 159 hypothetical protein 5264970 5264812 5.6 6062 - 6.12 -1.20 0.19 16.065503 RB9872 67 204 hypothetical protein 5316692 5316895 5.4 6912 + 6.53 -1.96 0.22 11.155574 RB10005 77 234 hypothetical protein 5389186 5389419 4.4 8625 + 6.38 -1.18 0.10 8.635579 RB10014 534 1605 protein containing DUF1552 5395577 5393973 7.2 58534 - 6.25 -1.06 0.20 18.505855 RB10521 53 162 hypothetical protein 5708182 5708343 9.1 6074 + 6.76 -1.45 0.16 10.735885 RB10579 101 306 secreted protein 5739254 5738949 12.4 11507 - 7.41 -1.36 0.25 18.215887 RB10581 384 1155 secreted protein containing DUF1559 5739353 5740507 6.2 41431 + 7.35 -1.29 0.23 17.636042 RB10860 96 291 hypothetical protein 5896828 5896538 10 10535 - 6.23 -1.08 0.12 10.936043 RB10861 46 141 hypothetical protein 5896851 5896991 4.2 4833 + 7.18 -1.15 0.10 8.566045 RB10864 314 945 hypothetical protein 5898139 5899083 4.7 34608 + 6.34 -1.11 0.13 11.666232 RB11204 170 513 hypothetical protein 6077364 6076852 10.6 19111 - 6.50 -1.57 0.35 22.596258 RB11245 53 162 hypothetical protein 6099126 6098965 6.3 6073 - 6.41 -1.29 0.28 21.596280 RB11287 75 228 hypothetical protein 6118405 6118632 9.1 8478 + 6.21 -1.10 0.26 23.516304 RB11319 551 1656 hypothetical protein 6136312 6134657 6.1 63215 - 6.00 -1.09 0.03 2.336336 RB11383 573 1722 conserved hypothetical protein 6174244 6175965 4.5 66716 + 6.42 -1.05 0.03 2.396343 RB11392 148 447 conserved hypothetical protein 6177208 6177654 5 16382 + 6.51 -1.50 0.21 14.306371 RB11439 93 282 conserved hypothetical protein 6197993 6198274 10.6 10906 + 8.85 -1.73 0.06 3.476385 RB11450 60 183 conserved hypothetical protein 6200046 6200228 11.8 6517 + 9.28 -1.87 0.24 12.916536 RB11703 52 159 conserved hypothetical protein 6311093 6311251 12 6046 + 6.38 -1.38 0.15 10.986569 RB11766 129 390 conserved hypothetical protein 6334355 6334744 10.4 14762 + 6.41 -1.77 0.23 12.926579 RB11804 132 399 conserved hypothetical protein 6363537 6363139 11.3 15013 - 6.63 -2.17 0.34 15.736580 RB11805 61 186 conserved hypothetical protein 6363715 6363530 12.1 7084 - 6.63 -2.06 0.38 18.596642 RB11925 76 231 conserved hypothetical protein 6419855 6420085 12.2 8248 + 6.07 -1.50 0.33 21.696683 RB11992 205 618 conserved hypothetical protein, membrane 6469784 6470401 5.8 22120 + 6.41 -1.23 0.05 3.856730 RB12084 285 858 conserved hypothetical protein, secreted 6520429 6519572 4.7 31843 - 6.44 -1.04 0.06 5.856744 RB12114 110 333 conserved hypothetical protein 6540259 6539927 11.8 12716 - 6.29 -1.04 0.17 16.376781 RB12193 36 111 conserved hypothetical protein 6575977 6575867 7.5 3719 - 6.57 -2.25 0.24 10.526787 RB12201 69 210 conserved hypothetical protein 6578161 6577952 12.5 7444 - 7.70 -1.58 0.06 4.096801 RB12220 171 516 conserved hypothetical protein 6587027 6586512 4.7 19504 - 6.39 -1.71 0.37 21.376857 RB12318 55 168 conserved hypothetical protein 6625565 6625398 11 6095 - 6.43 -1.65 0.20 11.966907 RB12403 59 180 conserved hypothetical protein 6673741 6673562 7.3 6301 - 6.48 -1.46 0.03 2.186920 RB12426 137 414 conserved hypothetical protein, membrane 6684059 6684472 10.6 15166 + 6.05 -1.37 0.16 11.526936 RB12447 103 312 conserved hypothetical protein 6700272 6700583 10.6 11445 + 6.59 -1.39 0.28 20.436953 RB12477 354 1065 conserved hypothetical protein, secreted 6716008 6717072 7.6 39728 + 6.61 -2.10 0.03 1.456992 RB12550 63 192 conserved hypothetical protein 6762725 6762534 11.9 7124 - 6.50 -2.30 0.18 7.726996 RB12557 41 126 conserved hypothetical protein 6765784 6765659 9.2 4684 - 6.28 -1.33 0.28 21.317009 RB12581 174 525 conserved hypothetical protein 6780597 6780073 9.9 19433 - 6.32 -1.71 0.11 6.467365 RB13247 185 558 conserved hypothetical protein 7104177 7103620 10.2 21780 - 6.31 -1.40 0.23 16.26
300min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand Intensity Ratio Stdev %5699 RB10217 479 1440 ATP synthase beta subunit 5519560 5520999 4.9 52151 + 5.86 -1.32 0.26 19.56
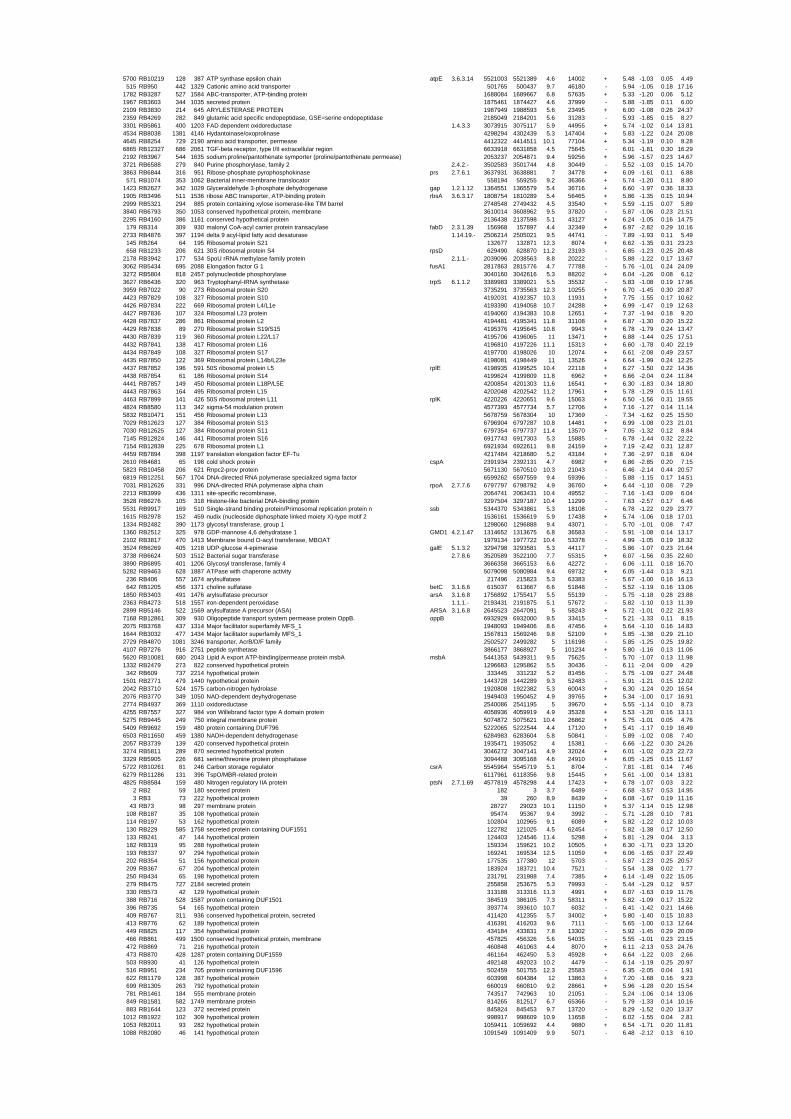
5700 RB10219 128 387 ATP synthase epsilon chain atpE 3.6.3.14 5521003 5521389 4.6 14002 + 5.48 -1.03 0.05 4.49515 RB950 442 1329 Cationic amino acid transporter 501765 500437 9.7 46180 - 5.94 -1.05 0.18 17.16
1782 RB3287 527 1584 ABC-transporter, ATP-binding protein 1688084 1689667 6.8 57635 + 5.33 -1.20 0.06 5.121967 RB3603 344 1035 secreted protein 1875461 1874427 4.6 37999 - 5.88 -1.85 0.11 6.002109 RB3830 214 645 ARYLESTERASE PROTEIN 1987949 1988593 5.6 23495 + 6.00 -1.08 0.26 24.372359 RB4269 282 849 glutamic acid specific endopeptidase, GSE=serine endopeptidase 2185049 2184201 5.6 31283 - 5.93 -1.85 0.15 8.273301 RB5861 400 1203 FAD dependent oxidoreductase 1.4.3.3 3073915 3075117 5.9 44955 + 5.74 -1.02 0.14 13.814534 RB8038 1381 4146 Hydantoinase/oxoprolinase 4298294 4302439 5.3 147404 + 5.83 -1.22 0.24 20.084645 RB8254 729 2190 amino acid transporter, permease 4412322 4414511 10.1 77104 + 5.34 -1.19 0.10 8.286865 RB12327 686 2061 TGF-beta receptor, type I/II extracellular region 6633918 6631858 4.5 75645 - 6.01 -1.81 0.30 16.292192 RB3967 544 1635 sodium:proline/pantothenate symporter (proline/pantothenate permease) 2053237 2054871 9.4 59256 + 5.96 -1.57 0.23 14.673721 RB6588 279 840 Purine phosphorylase, family 2 2.4.2.- 3502583 3501744 4.8 30449 - 5.52 -1.03 0.15 14.703863 RB6844 316 951 Ribose-phosphate pyrophosphokinase prs 2.7.6.1 3637931 3638881 7 34778 + 6.09 -1.61 0.11 6.88571 RB1074 353 1062 Bacterial inner-membrane translocator 558194 559255 9.2 36366 + 5.74 -1.20 0.11 8.80
1423 RB2627 342 1029 Glyceraldehyde 3-phosphate dehydrogenase gap 1.2.1.12 1364551 1365579 5.4 36716 + 6.60 -1.97 0.36 18.331905 RB3496 511 1536 ribose ABC transporter, ATP-binding protein rbsA 3.6.3.17 1808754 1810289 5.4 56465 + 5.86 -1.35 0.15 10.942999 RB5321 294 885 protein containing xylose isomerase-like TIM barrel 2748548 2749432 4.5 33540 + 5.59 -1.15 0.07 5.893840 RB6793 350 1053 conserved hypothetical protein, membrane 3610014 3608962 9.5 37820 - 5.87 -1.06 0.23 21.512295 RB4160 386 1161 conserved hypothetical protein 2136438 2137598 5.1 43127 + 6.24 -1.05 0.16 14.75179 RB314 309 930 malonyl CoA-acyl carrier protein transacylase fabD 2.3.1.39 156968 157897 4.4 32349 + 6.97 -2.82 0.29 10.16
2733 RB4876 397 1194 delta 9 acyl-lipid fatty acid desaturase 1.14.19.- 2506214 2505021 9.5 44741 - 7.89 -1.93 0.11 5.49145 RB264 64 195 Ribosomal protein S21 132677 132871 12.3 8074 + 6.62 -1.35 0.31 23.23658 RB1233 206 621 30S ribosomal protein S4 rpsD 629490 628870 11.2 23193 - 6.85 -1.23 0.25 20.48
2178 RB3942 177 534 SpoU rRNA methylase family protein 2.1.1.- 2039096 2038563 8.8 20222 - 5.88 -1.22 0.17 13.673062 RB5434 695 2088 Elongation factor G 1 fusA1 2817863 2815776 4.7 77788 - 5.76 -1.01 0.24 24.093272 RB5804 818 2457 polynucleotide phosphorylase 3040160 3042616 5.3 88202 + 6.04 -1.26 0.08 6.123627 RB6436 320 963 Tryptophanyl-tRNA synthetase trpS 6.1.1.2 3389983 3389021 5.5 35532 - 5.83 -1.08 0.19 17.963959 RB7022 90 273 Ribosomal protein S20 3735291 3735563 12.3 10255 + 6.70 -1.45 0.30 20.874423 RB7829 108 327 Ribosomal protein S10 4192031 4192357 10.3 11931 + 7.75 -1.55 0.17 10.624426 RB7834 222 669 Ribosomal protein L4/L1e 4193390 4194058 10.7 24288 + 6.99 -1.47 0.19 12.634427 RB7836 107 324 Ribosomal L23 protein 4194060 4194383 10.8 12651 + 7.37 -1.94 0.18 9.204428 RB7837 286 861 Ribosomal protein L2 4194481 4195341 11.8 31108 + 6.87 -1.30 0.20 15.224429 RB7838 89 270 Ribosomal protein S19/S15 4195376 4195645 10.8 9943 + 6.78 -1.79 0.24 13.474430 RB7839 119 360 Ribosomal protein L22/L17 4195706 4196065 11 13471 + 6.88 -1.44 0.25 17.514432 RB7841 138 417 Ribosomal protein L16 4196810 4197226 11.1 15313 + 6.60 -1.78 0.40 22.194434 RB7849 108 327 Ribosomal protein S17 4197700 4198026 10 12074 + 6.61 -2.08 0.49 23.574435 RB7850 122 369 Ribosomal protein L14b/L23e 4198081 4198449 11 13526 + 6.64 -1.99 0.24 12.254437 RB7852 196 591 50S ribosomal protein L5 rplE 4198935 4199525 10.4 22118 + 6.27 -1.50 0.22 14.364438 RB7854 61 186 Ribosomal protein S14 4199624 4199809 11.8 6962 + 6.66 -2.04 0.24 11.844441 RB7857 149 450 Ribosomal protein L18P/L5E 4200854 4201303 11.6 16541 + 6.30 -1.83 0.34 18.804443 RB7863 164 495 Ribosomal protein L15 4202048 4202542 11.2 17961 + 5.78 -1.29 0.15 11.614463 RB7899 141 426 50S ribosomal protein L11 rplK 4220226 4220651 9.6 15063 + 6.50 -1.56 0.31 19.554824 RB8580 113 342 sigma-54 modulation protein 4577393 4577734 5.7 12706 + 7.16 -1.27 0.14 11.145832 RB10471 151 456 Ribosomal protein L13 5678759 5678304 10 17369 - 7.34 -1.62 0.25 15.507029 RB12623 127 384 Ribosomal protein S13 6796904 6797287 10.8 14481 + 6.99 -1.08 0.23 21.017030 RB12625 127 384 Ribosomal protein S11 6797354 6797737 11.4 13570 + 7.05 -1.32 0.12 8.847145 RB12824 146 441 Ribosomal protein S16 6917743 6917303 5.3 15885 - 6.78 -1.44 0.32 22.227154 RB12839 225 678 Ribosomal protein L1 6921934 6922611 9.8 24159 + 7.19 -2.42 0.31 12.874459 RB7894 398 1197 translation elongation factor EF-Tu 4217484 4218680 5.2 43184 + 7.36 -2.97 0.18 6.042610 RB4681 65 198 cold shock protein cspA 2391934 2392131 4.7 6982 + 6.86 -2.85 0.20 7.155823 RB10458 206 621 Rnpc2-prov protein 5671130 5670510 10.3 21043 - 6.46 -2.14 0.44 20.576819 RB12251 567 1704 DNA-directed RNA polymerase specialized sigma factor 6599262 6597559 9.4 59396 - 5.88 -1.15 0.17 14.517031 RB12626 331 996 DNA-directed RNA polymerase alpha chain rpoA 2.7.7.6 6797797 6798792 4.9 36760 + 6.44 -1.10 0.08 7.292213 RB3999 436 1311 site-specific recombinase, 2064741 2063431 10.4 49552 - 7.16 -1.43 0.09 6.043528 RB6276 105 318 Histone-like bacterial DNA-binding protein 3297504 3297187 10.4 11299 - 7.63 -2.57 0.17 6.465531 RB9917 169 510 Single-strand binding protein/Primosomal replication protein n ssb 5344370 5343861 5.3 18108 - 6.78 -1.22 0.29 23.771615 RB2978 152 459 nudix (nucleoside diphosphate linked moiety X)-type motif 2 1536161 1536619 5.9 17438 + 5.74 -1.06 0.18 17.011334 RB2482 390 1173 glycosyl transferase, group 1 1298060 1296888 9.4 43071 - 5.70 -1.01 0.08 7.471360 RB2512 325 978 GDP-mannose 4,6 dehydratase 1 GMD1 4.2.1.47 1314652 1313675 6.8 36583 - 5.91 -1.08 0.14 13.172102 RB3817 470 1413 Membrane bound O-acyl transferase, MBOAT 1979134 1977722 10.4 53378 - 4.99 -1.05 0.19 18.323524 RB6269 405 1218 UDP-glucose 4-epimerase galE 5.1.3.2 3294798 3293581 5.3 44117 - 5.86 -1.07 0.23 21.643738 RB6624 503 1512 Bacterial sugar transferase 2.7.8.6 3520589 3522100 7.7 55315 + 6.07 -1.56 0.35 22.603890 RB6895 401 1206 Glycosyl transferase, family 4 3666358 3665153 6.6 42272 - 6.06 -1.11 0.18 16.705282 RB9463 628 1887 ATPase with chaperone activity 5079098 5080984 9.4 69732 + 6.05 -1.44 0.13 9.21236 RB406 557 1674 arylsulfatase 217496 215823 5.3 63383 - 5.67 -1.00 0.16 16.13642 RB1205 456 1371 choline sulfatase betC 3.1.6.6 615037 613667 6.6 51848 - 5.52 -1.19 0.16 13.06
1850 RB3403 491 1476 arylsulfatase precursor arsA 3.1.6.8 1756892 1755417 5.5 55139 - 5.75 -1.18 0.28 23.882363 RB4273 518 1557 iron-dependent peroxidase 1.1.1.- 2193431 2191875 5.1 57672 - 5.82 -1.10 0.13 11.392899 RB5146 522 1569 arylsulfatase A precursor (ASA) ARSA 3.1.6.8 2645523 2647091 5 58243 + 5.72 -1.01 0.22 21.937168 RB12861 309 930 Oligopeptide transport system permease protein OppB. oppB 6932929 6932000 9.5 33415 - 5.21 -1.33 0.11 8.152075 RB3768 437 1314 Major facilitator superfamily MFS_1 1948093 1949406 8.6 47456 + 5.64 -1.10 0.16 14.831644 RB3032 477 1434 Major facilitator superfamily MFS_1 1567813 1569246 9.8 52109 + 5.85 -1.38 0.29 21.102729 RB4870 1081 3246 transporter, AcrB/D/F family 2502527 2499282 5 116198 - 5.85 -1.25 0.25 19.824107 RB7276 916 2751 peptide synthetase 3866177 3868927 5 101234 + 5.80 -1.16 0.13 11.065620 RB10081 680 2043 Lipid A export ATP-binding/permease protein msbA msbA 5441353 5439311 9.5 75625 - 5.70 -1.07 0.13 11.981332 RB2479 273 822 conserved hypothetical protein 1296683 1295862 5.5 30436 - 6.11 -2.04 0.09 4.29342 RB609 737 2214 hypothetical protein 333445 331232 5.2 81456 - 5.75 -1.09 0.27 24.48
1501 RB2771 479 1440 hypothetical protein 1443728 1442289 9.3 52483 - 5.91 -1.21 0.15 12.022042 RB3710 524 1575 carbon-nitrogen hydrolase 1920808 1922382 5.3 60043 + 6.30 -1.24 0.20 16.542076 RB3770 349 1050 NAD-dependent deyhydrogenase 1949403 1950452 4.9 39765 + 5.34 -1.00 0.17 16.912774 RB4937 369 1110 oxidoreductase 2540086 2541195 5 39670 + 5.55 -1.14 0.10 8.734255 RB7557 327 984 von Willebrand factor type A domain protein 4058936 4059919 4.9 35328 + 5.53 -1.20 0.16 13.115275 RB9445 249 750 integral membrane protein 5074872 5075621 10.4 26862 + 5.75 -1.01 0.05 4.765409 RB9692 159 480 protein containing DUF796 5222065 5222544 4.4 17120 + 5.41 -1.17 0.19 16.496503 RB11650 459 1380 NADH-dependent dehydrogenase 6284983 6283604 5.8 50841 - 5.89 -1.02 0.08 7.402057 RB3739 139 420 conserved hypothetical protein 1935471 1935052 4 15381 - 6.66 -1.22 0.30 24.263274 RB5811 289 870 secreted hypothetical protein 3046272 3047141 4.9 32024 + 6.01 -1.02 0.23 22.733329 RB5905 226 681 serine/threonine protein phosphatase 3094488 3095168 4.6 24910 + 6.05 -1.25 0.15 11.675722 RB10261 81 246 Carbon storage regulator csrA 5545964 5545719 5.1 8704 - 7.81 -1.81 0.14 7.466279 RB11286 131 396 TspO/MBR-related protein 6117961 6118356 9.8 15445 + 5.61 -1.00 0.14 13.814825 RB8584 159 480 Nitrogen regulatory IIA protein ptsN 2.7.1.69 4577819 4578298 4.4 17423 + 6.78 -1.07 0.03 3.22
2 RB2 59 180 secreted protein 182 3 3.7 6489 - 6.68 -3.57 0.53 14.953 RB3 73 222 hypothetical protein 39 260 8.9 8439 + 6.08 -1.67 0.19 11.16
43 RB73 98 297 membrane protein 28727 29023 10.1 11150 + 5.37 -1.14 0.15 12.98108 RB187 35 108 hypothetical protein 95474 95367 9.4 3992 - 5.71 -1.28 0.10 7.81114 RB197 53 162 hypothetical protein 102804 102965 9.1 6089 + 5.82 -1.22 0.12 10.03130 RB229 585 1758 secreted protein containing DUF1551 122782 121025 4.5 62454 - 5.82 -1.38 0.17 12.50133 RB241 47 144 hypothetical protein 124403 124546 11.4 5298 + 5.81 -1.29 0.04 3.13182 RB319 95 288 hypothetical protein 159334 159621 10.2 10505 + 6.30 -1.71 0.23 13.20193 RB337 97 294 hypothetical protein 169241 169534 12.5 11059 + 6.06 -1.65 0.37 22.49202 RB354 51 156 hypothetical protein 177535 177380 12 5703 - 5.87 -1.23 0.25 20.57209 RB367 67 204 hypothetical protein 183924 183721 10.4 7521 - 5.54 -1.38 0.02 1.77250 RB434 65 198 hypothetical protein 231791 231988 7.4 7385 + 6.14 -1.49 0.22 15.05279 RB475 727 2184 secreted protein 255858 253675 5.3 79993 - 5.44 -1.29 0.12 9.57330 RB573 42 129 hypothetical protein 313188 313316 11.3 4991 + 6.07 -1.63 0.19 11.76388 RB716 528 1587 protein containing DUF1501 384519 386105 7.3 58311 + 5.82 -1.09 0.17 15.22396 RB735 54 165 hypothetical protein 393774 393610 10.7 6032 - 6.41 -1.42 0.21 14.66409 RB767 311 936 conserved hypothetical protein, secreted 411420 412355 5.7 34002 + 5.80 -1.40 0.15 10.83413 RB776 62 189 hypothetical protein 416391 416203 9.6 7111 - 5.65 -1.00 0.13 12.64449 RB825 117 354 hypothetical protein 434184 433831 7.8 13302 - 5.92 -1.45 0.29 20.09466 RB861 499 1500 conserved hypothetical protein, membrane 457825 456326 5.6 54035 - 5.55 -1.01 0.23 23.15472 RB869 71 216 hypothetical protein 460848 461063 4.4 8070 + 6.11 -2.13 0.53 24.76473 RB870 428 1287 protein containing DUF1559 461164 462450 5.3 45928 + 6.64 -1.22 0.03 2.66503 RB930 41 126 hypothetical protein 492148 492023 10.2 4479 - 6.14 -1.19 0.25 20.97516 RB951 234 705 protein containing DUF1596 502459 501755 12.3 25583 - 6.35 -2.05 0.04 1.91622 RB1179 128 387 hypothetical protein 603998 604384 12 13863 + 7.20 -1.68 0.16 9.23699 RB1305 263 792 hypothetical protein 660019 660810 9.2 28661 + 5.96 -1.28 0.20 15.54781 RB1461 184 555 membrane protein 743517 742963 10 21051 - 5.24 -1.06 0.14 13.06849 RB1581 582 1749 membrane protein 814265 812517 6.7 65366 - 5.79 -1.33 0.14 10.16883 RB1644 123 372 secreted protein 845824 845453 9.7 13720 - 8.29 -1.52 0.20 13.37
1012 RB1922 102 309 hypothetical protein 998917 998609 10.9 11658 - 6.02 -1.55 0.04 2.811053 RB2011 93 282 hypothetical protein 1059411 1059692 4.4 9880 + 6.54 -1.71 0.20 11.811088 RB2080 46 141 hypothetical protein 1091549 1091409 9.9 5071 - 6.48 -2.12 0.13 6.10

1099 RB2097 61 186 hypothetical protein 1106473 1106658 9.9 6506 + 6.00 -1.65 0.38 22.971105 RB2105 470 1413 membrane protein 1114012 1112600 9.6 51853 - 6.19 -1.85 0.14 7.551106 RB2107 93 282 hypothetical protein 1113972 1114253 10.1 10253 + 6.10 -1.45 0.32 22.321130 RB2150 93 282 hypothetical protein 1138311 1138030 4.4 10755 - 7.13 -2.17 0.27 12.241144 RB2175 98 297 membrane protein 1148511 1148807 10.3 10772 + 6.01 -1.07 0.10 9.691183 RB2247 579 1740 conserved hypothetical protein containing YTV domains 1181238 1179499 9.8 65013 - 6.80 -2.30 0.26 11.451184 RB2251 75 228 hypothetical protein 1181714 1181487 4.5 8027 - 7.17 -2.64 0.22 8.411188 RB2255 628 1887 hypothetical protein 1186018 1184132 4.1 66330 - 6.50 -1.78 0.17 9.491205 RB2281 310 933 hypothetical protein 1196032 1196964 11.2 33398 + 6.49 -1.21 0.07 5.751217 RB2302 46 141 hypothetical protein 1204388 1204528 5.6 5335 + 6.73 -1.69 0.08 4.751220 RB2306 41 126 hypothetical protein 1206782 1206907 9 4759 + 6.12 -1.51 0.26 17.121236 RB2331 133 402 hypothetical protein 1215088 1214687 10.3 15081 - 6.01 -1.03 0.10 9.211253 RB2353 124 375 membrane protein 1221696 1221322 10.4 13329 - 5.93 -1.18 0.25 21.131265 RB2370 54 165 hypothetical protein 1229187 1229023 6.3 6204 - 5.63 -1.01 0.12 11.861277 RB2384 65 198 hypothetical protein 1242697 1242500 12 7263 - 6.06 -1.73 0.36 20.911285 RB2400 66 201 hypothetical protein 1248699 1248499 12.1 7112 - 5.71 -1.04 0.05 4.801295 RB2420 547 1644 hypothetical protein 1258838 1260481 9.3 59939 + 5.75 -1.12 0.25 21.961297 RB2425 102 309 hypothetical protein 1260766 1261074 11.1 11189 + 5.93 -1.44 0.30 20.991310 RB2451 90 273 hypothetical protein 1278610 1278882 10.7 9921 + 5.89 -1.16 0.11 9.411322 RB2467 66 201 hypothetical protein 1287454 1287654 11.3 7649 + 5.77 -1.00 0.05 4.521338 RB2489 66 201 hypothetical protein 1301658 1301458 9 7521 - 5.09 -1.22 0.06 4.701339 RB2490 43 132 hypothetical protein 1301758 1301627 7.4 4782 - 5.81 -1.03 0.20 19.301352 RB2503 515 1548 O-antigen flippase wzx 1311056 1309509 10 55740 - 5.50 -1.19 0.22 18.781424 RB2628 43 132 hypothetical protein 1365707 1365576 10 4777 - 5.98 -1.34 0.18 13.071428 RB2636 116 351 hypothetical protein 1368450 1368100 11.7 12761 - 6.72 -1.94 0.09 4.591446 RB2667 236 711 membrane protein 1387198 1386488 10.7 25698 - 5.73 -1.35 0.25 18.341472 RB2714 302 909 protein containing DUF541 1415817 1414909 9.5 33087 - 5.90 -1.22 0.15 12.641505 RB2782 589 1770 protein containing DUF1559 1445576 1447345 7.2 63826 + 7.37 -1.03 0.13 12.231542 RB2852 240 723 conserved hypothetical protein, membrane 1482871 1482149 9.4 26727 - 5.59 -1.20 0.08 6.321594 RB2948 148 447 membrane protein 1522349 1521903 9.5 16181 - 5.54 -1.00 0.14 13.811597 RB2951 174 525 membrane protein 1523616 1523092 9.3 20061 - 5.57 -1.20 0.14 11.851668 RB3067 153 462 membrane protein 1582850 1582389 7.6 17200 - 5.50 -1.00 0.05 5.031672 RB3073 51 156 hypothetical protein 1585180 1585025 9.1 5384 - 5.80 -1.06 0.21 19.861686 RB3119 195 588 membrane protein 1616510 1615923 9.3 22301 - 5.63 -1.09 0.08 7.381688 RB3125 135 408 hypothetical protein 1618379 1617972 10.5 14861 - 6.90 -1.66 0.25 14.831689 RB3127 68 207 hypothetical protein 1618378 1618584 12.3 7939 + 5.98 -1.14 0.21 18.581692 RB3132 112 339 membrane protein 1619400 1619062 10.3 12831 - 5.72 -1.22 0.19 15.331705 RB3152 68 207 hypothetical protein 1626667 1626461 12.2 7261 - 6.35 -1.44 0.25 17.461767 RB3262 128 387 hypothetical protein 1678132 1678518 4.2 14073 + 5.93 -1.08 0.22 20.121771 RB3268 231 696 hypothetical protein 1683003 1682308 12 25730 - 6.07 -1.60 0.26 16.361776 RB3277 221 666 hypothetical protein 1686552 1685887 10.4 24222 - 6.12 -1.92 0.03 1.771789 RB3301 50 153 hypothetical protein 1694208 1694056 10.5 5470 - 6.05 -1.19 0.06 5.391827 RB3365 43 132 hypothetical protein 1730124 1730255 11.4 4886 + 8.08 -3.20 0.17 5.301828 RB3366 78 237 hypothetical protein 1730380 1730616 5.6 9481 + 5.63 -1.02 0.23 23.121833 RB3372 115 348 hypothetical protein 1733666 1734013 10 12916 + 5.56 -1.14 0.20 17.531842 RB3391 79 240 hypothetical protein 1751875 1751636 12.4 9331 - 6.29 -1.47 0.21 14.231843 RB3393 60 183 secreted protein 1751865 1752047 12.3 7156 + 6.21 -1.78 0.21 11.601844 RB3394 36 111 hypothetical protein 1752044 1752154 10.5 4239 + 6.08 -1.87 0.32 17.391845 RB3395 398 1197 secreted protein containing DUF1559 1752151 1753347 7.8 43136 + 6.09 -1.67 0.27 16.231846 RB3398 67 204 hypothetical protein 1753293 1753496 11.4 7537 + 5.62 -1.04 0.17 16.081847 RB3399 65 198 hypothetical protein 1753650 1753453 9.7 7418 - 6.36 -2.35 0.17 7.091882 RB3455 57 174 hypothetical protein 1785458 1785631 10.2 6258 + 6.01 -1.08 0.26 24.061892 RB3473 443 1332 membrane protein 1794413 1795744 4.8 47669 + 5.92 -1.32 0.23 17.481904 RB3495 218 657 conserved hypothetical protein 1808101 1808757 5 23739 + 5.09 -1.03 0.16 16.011942 RB3565 137 414 membrane protein 1853670 1854083 10.1 15653 + 5.53 -1.07 0.22 20.781943 RB3566 64 195 hypothetical protein 1854080 1854274 9.4 7400 + 5.84 -1.32 0.13 9.671946 RB3569 123 372 conserved hypothetical protein 1854728 1854357 7.5 13875 - 5.63 -1.04 0.18 17.731950 RB3575 152 459 membrane protein 1858779 1859237 10.1 17195 + 5.72 -1.46 0.10 6.921992 RB3644 127 384 hypothetical protein 1895184 1894801 4.9 14294 - 7.30 -1.99 0.26 12.982025 RB3688 53 162 hypothetical protein 1910629 1910468 9.3 5981 - 6.07 -1.32 0.10 7.562047 RB3718 535 1608 Major facilitator superfamily MFS_1 1927114 1925507 4.6 57372 - 5.63 -1.27 0.15 11.712048 RB3719 141 426 hypothetical protein 1927500 1927075 10.8 16330 - 6.21 -2.10 0.25 12.062113 RB3835 50 153 hypothetical protein 1989687 1989839 11.3 5789 + 5.88 -1.06 0.26 24.922132 RB3867 284 855 Endonuclease/exonuclease/phosphatase 2000435 2001289 6.1 31682 + 6.13 -1.22 0.15 12.082133 RB3868 125 378 hypothetical protein 2001663 2001286 11.9 14088 - 5.81 -1.03 0.13 12.992139 RB3880 82 249 hypothetical protein 2006953 2006705 10.7 9535 - 6.05 -1.56 0.28 17.822154 RB3905 38 117 hypothetical protein 2018601 2018717 5.1 4131 + 6.40 -1.54 0.14 8.812166 RB3925 50 153 hypothetical protein 2033534 2033382 10.2 5399 - 5.89 -1.01 0.10 9.742184 RB3953 857 2574 hypothetical protein 2048784 2046211 5.2 93881 - 5.87 -1.43 0.17 11.622198 RB3975 204 615 hypothetical protein 2056726 2057340 12.3 22602 + 5.88 -1.04 0.02 2.362200 RB3981 161 486 hypothetical protein 2058000 2058485 4.1 18283 + 5.83 -1.25 0.20 16.262206 RB3990 55 168 hypothetical protein 2060721 2060888 11.2 6203 + 6.08 -1.56 0.12 7.832210 RB3994 191 576 hypothetical protein 2061291 2061866 4.1 20739 + 5.86 -1.15 0.20 17.322216 RB4002 483 1452 hypothetical protein 2065341 2066792 4.6 53759 + 5.75 -1.14 0.23 20.462244 RB4061 488 1467 Peptidase A24A, prepilin type IV:Peptidase A24A, N-terminal 2095498 2096964 9.7 54251 + 5.76 -1.01 0.12 11.542254 RB4084 119 360 hypothetical protein 2104677 2105036 10 13521 + 6.13 -1.61 0.13 7.832258 RB4093 45 138 hypothetical protein 2107600 2107737 10 5285 + 6.10 -1.76 0.16 9.072260 RB4097 733 2202 conserved hypothetical protein 2108244 2110445 6.2 83188 + 5.88 -1.23 0.21 16.862280 RB4129 426 1281 conserved hypothetical protein 2125088 2126368 4.6 46461 + 5.93 -1.07 0.10 9.682285 RB4142 43 132 hypothetical protein 2130009 2130140 12 4964 + 6.22 -1.43 0.31 21.462287 RB4145 90 273 hypothetical protein 2131933 2131661 12 10727 - 6.35 -1.00 0.17 16.762312 RB4194 53 162 hypothetical protein 2156685 2156524 11.4 5728 - 7.19 -1.98 0.19 9.442343 RB4243 58 177 hypothetical protein 2174155 2174331 4.4 6442 + 5.72 -1.05 0.17 16.112369 RB4283 499 1500 PvdD/PvdJ 2201462 2199963 5.7 56411 - 6.02 -1.75 0.06 3.712371 RB4285 841 2526 membrane protein 2204136 2201611 5.9 93852 - 5.74 -1.02 0.06 5.752416 RB4358 123 372 hypothetical protein 2239474 2239103 6.5 13459 - 6.13 -1.79 0.12 6.722417 RB4360 156 471 hypothetical protein 2240075 2239605 4.7 17592 - 6.04 -1.43 0.02 1.232424 RB4373 109 330 hypothetical protein 2242863 2242534 4.8 12263 - 6.15 -1.99 0.27 13.352528 RB4547 422 1269 hypothetical protein 2330116 2328848 7.6 46274 - 5.57 -1.13 0.18 16.322553 RB4590 74 225 hypothetical protein 2352024 2351800 10.1 8330 - 9.54 -2.32 0.27 11.422593 RB4652 52 159 hypothetical protein 2380494 2380652 12.6 5628 + 6.73 -1.61 0.15 9.442596 RB4657 123 372 hypothetical protein 2381865 2382236 12.2 13398 + 6.26 -1.93 0.16 8.082631 RB4719 54 165 hypothetical protein 2419276 2419112 12.3 6311 - 5.48 -1.02 0.24 23.382647 RB4740 50 153 hypothetical protein 2431814 2431966 11.4 5598 + 6.08 -1.87 0.30 15.912664 RB4766 130 393 membrane protein 2442991 2442599 9.7 14452 - 5.33 -1.01 0.13 12.502687 RB4801 96 291 hypothetical protein 2459502 2459212 9.8 10561 - 5.73 -1.07 0.13 12.032698 RB4814 87 264 hypothetical protein 2467537 2467800 11.3 9301 + 6.40 -1.50 0.34 22.372746 RB4897 184 555 hypothetical protein 2519505 2518951 10.9 20446 - 5.50 -1.11 0.16 14.102777 RB4941 173 522 hypothetical protein 2541619 2542140 11.2 19306 + 5.74 -1.95 0.34 17.442781 RB4951 95 288 hypothetical protein 2547498 2547785 12.1 10953 + 6.65 -1.71 0.33 19.122789 RB4966 84 255 hypothetical protein 2552311 2552057 11.3 10016 - 9.52 -2.05 0.27 13.282794 RB4972 56 171 hypothetical protein 2557220 2557390 9 6197 + 7.05 -1.33 0.19 14.532825 RB5016 235 708 hypothetical protein 2579933 2579226 4.3 27492 - 5.13 -1.11 0.05 4.382858 RB5076 113 342 hypothetical protein 2612505 2612164 10.5 12883 - 5.03 -1.13 0.24 21.162936 RB5211 159 480 conserved hypothetical protein 2688867 2689346 7.4 17871 + 5.93 -1.15 0.12 10.392964 RB5262 95 288 membrane protein 2713742 2714029 6.3 10993 + 5.76 -1.11 0.24 21.843045 RB5409 97 294 hypothetical protein 2794195 2794488 12.7 10898 + 7.53 -3.71 0.16 4.403049 RB5415 62 189 hypothetical protein 2798737 2798925 12.3 7098 + 6.53 -1.69 0.31 18.403052 RB5418 73 222 secreted protein 2805185 2804964 11 8406 - 5.41 -1.10 0.24 21.923239 RB5745 130 393 hypothetical protein 3001234 3000842 10.7 13522 - 6.75 -2.90 0.44 15.143258 RB5779 187 564 hypothetical protein 3028410 3028973 4.4 21341 + 7.19 -1.11 0.18 15.933271 RB5802 156 471 hypothetical protein 3039864 3040334 10.1 17001 + 6.76 -1.57 0.08 5.343291 RB5841 670 2013 serine/threonine protein phosphatase family 3061924 3063936 5.4 73664 + 5.57 -1.05 0.09 8.833356 RB5957 154 465 secreted protein 3122093 3122557 3.3 15514 + 7.13 -1.83 0.30 16.443368 RB5976 54 165 hypothetical protein 3135318 3135482 11.8 6532 + 5.19 -1.04 0.19 17.813388 RB6009 122 369 hypothetical protein 3149941 3149573 4.3 14124 - 5.86 -1.31 0.28 21.183430 RB6092 361 1086 Peptidase M50 3.4.24.- 3192387 3193472 9.6 39370 + 5.85 -1.13 0.01 0.943466 RB6158 142 429 hypothetical protein 3233532 3233104 6 15746 - 6.19 -1.66 0.30 17.973476 RB6174 69 210 hypothetical protein 3239734 3239525 10.6 7870 - 5.70 -1.07 0.07 7.003485 RB6193 62 189 hypothetical protein 3246096 3245908 11.4 6970 - 6.34 -2.04 0.21 10.193513 RB6245 107 324 hypothetical protein 3278762 3279085 12.4 12397 + 6.25 -1.35 0.27 20.063547 RB6314 52 159 hypothetical protein 3319320 3319478 6.3 6180 + 6.84 -3.31 0.23 6.953559 RB6332 250 753 conserved hypothetical protein, secreted 3325222 3325974 9.7 27162 + 5.33 -1.07 0.09 8.81

3643 RB6462 90 273 hypothetical protein 3414908 3415180 9.9 10005 + 6.14 -1.08 0.01 0.523644 RB6463 189 570 conserved hypothetical protein, membrane 3415155 3415724 10.2 19818 + 6.25 -1.10 0.14 12.863652 RB6477 369 1110 hypothetical protein 3424543 3425652 5.7 39787 + 5.98 -1.48 0.14 9.423666 RB6495 39 120 hypothetical protein 3441961 3441842 12.7 4765 - 5.88 -1.09 0.11 10.363701 RB6553 74 225 hypothetical protein 3479627 3479403 12.1 8987 - 6.46 -1.94 0.05 2.743709 RB6567 37 114 hypothetical protein 3489298 3489185 7.6 4468 - 5.99 -1.23 0.08 6.203718 RB6583 996 2991 secreted protein 3498012 3501002 4.5 112232 + 6.01 -1.56 0.38 24.043725 RB6594 295 888 hypothetical protein 3505472 3506359 10.2 32951 + 5.96 -1.67 0.22 12.983731 RB6609 45 138 hypothetical protein 3511456 3511593 10.4 5004 + 6.73 -2.24 0.18 8.193744 RB6634 365 1098 protein containing DUF1559 3525785 3526882 5.3 40217 + 6.03 -1.84 0.12 6.613780 RB6699 47 144 hypothetical protein 3559781 3559924 11.1 5290 + 5.82 -1.08 0.22 20.493820 RB6766 55 168 hypothetical protein 3597367 3597534 12 6133 + 6.04 -1.44 0.21 14.413826 RB6777 137 414 hypothetical protein 3602786 3603199 11.5 15512 + 7.26 -1.59 0.13 8.213827 RB6778 59 180 hypothetical protein 3603142 3603321 5.5 6533 + 6.65 -1.40 0.13 8.993836 RB6789 180 543 secreted protein 3608487 3607945 5 20191 - 5.94 -1.48 0.03 2.363867 RB6849 101 306 hypothetical protein 3642537 3642232 12.8 12002 - 6.06 -1.30 0.09 6.753971 RB7042 91 276 hypothetical protein 3743971 3744246 10.4 10835 + 5.72 -1.22 0.13 10.763977 RB7056 95 288 hypothetical protein 3746488 3746775 4.7 11164 + 5.91 -1.28 0.22 16.914009 RB7109 53 162 hypothetical protein 3775707 3775546 6.7 5850 - 5.51 -1.06 0.14 12.864015 RB7116 59 180 hypothetical protein 3779586 3779765 11.7 6458 + 6.93 -2.16 0.20 9.114016 RB7117 181 546 Ribosomal protein L35 3780322 3779777 11.4 19816 - 6.82 -1.77 0.23 12.784048 RB7177 35 108 hypothetical protein 3806891 3806784 12 4191 - 6.18 -1.18 0.02 1.554049 RB7178 45 138 hypothetical protein 3806924 3807061 12.3 5320 + 6.74 -2.24 0.41 18.364050 RB7180 104 315 hypothetical protein 3807345 3807031 10.8 11535 - 6.85 -2.34 0.21 9.154051 RB7184 190 573 conserved hypothetical protein 3807517 3808089 9.2 21306 + 6.49 -1.92 0.26 13.784092 RB7246 50 153 hypothetical protein 3850393 3850545 9.7 5973 + 6.80 -1.23 0.29 24.044131 RB7313 51 156 hypothetical protein 3892097 3891942 10.5 5558 - 7.27 -3.17 0.11 3.444163 RB7397 37 114 hypothetical protein 3956551 3956438 9.4 3746 - 5.60 -1.00 0.20 19.824200 RB7455 838 2517 conserved hypothetical protein containing YTV domains 3999277 3996761 9 92217 - 6.71 -1.93 0.31 16.034236 RB7525 281 846 secreted protein 4048930 4048085 4.2 27759 - 5.67 -2.67 0.02 0.594237 RB7530 97 294 hypothetical protein 4049298 4049005 12.4 11656 - 6.16 -3.24 0.46 14.244241 RB7535 136 411 hypothetical protein 4050024 4050434 10.9 15058 + 6.94 -1.20 0.11 9.394251 RB7549 60 183 hypothetical protein 4057545 4057727 11.8 6684 + 5.61 -1.22 0.06 5.074278 RB7589 86 261 hypothetical protein 4080238 4079978 12.1 9472 - 5.73 -1.26 0.04 3.504314 RB7646 62 189 hypothetical protein 4111735 4111923 10.5 6950 + 6.71 -2.38 0.13 5.294315 RB7647 73 222 hypothetical protein 4111931 4112152 7.4 8269 + 8.97 -2.89 0.15 5.184327 RB7667 38 117 hypothetical protein 4117097 4116981 6.7 4315 - 7.56 -1.51 0.20 13.144348 RB7709 134 405 protein containing DUF1582 4134747 4134343 11.9 15682 - 5.33 -1.02 0.23 22.894384 RB7756 74 225 hypothetical protein 4154712 4154488 12.1 8781 - 6.28 -1.13 0.19 16.494401 RB7791 533 1602 conserved hypothetical protein, membrane 4173444 4175045 6.3 57958 + 5.26 -1.04 0.03 2.984422 RB7828 40 123 hypothetical protein 4192037 4191915 10.4 4526 - 7.62 -1.23 0.09 7.374424 RB7832 50 153 hypothetical protein 4192499 4192651 13 5566 + 7.51 -1.85 0.23 12.274481 RB7929 75 228 hypothetical protein 4236583 4236810 11.9 8270 + 5.56 -1.16 0.24 20.464484 RB7937 41 126 hypothetical protein 4237809 4237684 10.7 4936 - 5.69 -1.08 0.13 11.674486 RB7939 105 318 hypothetical protein 4238296 4238613 11 12019 + 6.22 -1.41 0.16 11.104531 RB8034 147 444 membrane protein 4295344 4295787 10.2 16546 + 5.89 -1.37 0.11 8.264552 RB8070 59 180 hypothetical protein 4319538 4319717 4.5 6478 + 5.78 -1.46 0.06 4.334568 RB8098 71 216 hypothetical protein 4337334 4337119 10.8 7950 - 9.38 -2.59 0.23 8.964571 RB8102 387 1164 protein containing DUF1559 4339048 4337885 4.5 42114 - 5.37 -1.27 0.18 13.824578 RB8119 142 429 hypothetical protein 4346427 4346855 10.1 15659 + 5.67 -1.36 0.23 16.674589 RB8144 143 432 hypothetical protein 4354622 4354191 10.1 16269 - 8.53 -1.02 0.16 15.554613 RB8193 51 156 hypothetical protein 4382501 4382656 10.5 5994 + 5.88 -1.02 0.22 21.704629 RB8227 59 180 hypothetical protein 4400365 4400186 9.3 6533 - 5.84 -1.03 0.14 13.114677 RB8319 37 114 hypothetical protein 4442421 4442308 4.4 4328 - 5.74 -1.10 0.12 10.454696 RB8353 82 249 hypothetical protein 4459374 4459622 10.8 9402 + 6.17 -3.27 0.44 13.454718 RB8389 939 2820 membrane protein 4484734 4481915 10.1 105627 - 5.49 -1.11 0.11 9.874749 RB8457 113 342 hypothetical protein 4518458 4518117 11.5 12525 - 7.08 -1.02 0.22 21.964755 RB8468 88 267 hypothetical protein 4520799 4520533 11.9 9719 - 6.60 -1.20 0.15 12.674830 RB8594 41 126 hypothetical protein 4581645 4581770 9.2 4509 + 6.06 -1.36 0.26 19.034861 RB8669 37 114 hypothetical protein 4614557 4614670 11.5 4395 + 9.22 -2.87 0.27 9.244971 RB8866 445 1338 secreted conserved hypothetical protein 4716964 4718301 6 49999 + 5.86 -1.27 0.05 4.085052 RB9020 381 1146 secreted protein 4807486 4808631 5.9 42557 + 6.78 -1.76 0.42 23.895066 RB9050 56 171 hypothetical protein 4829846 4829676 9 6404 - 5.81 -1.01 0.11 11.045068 RB9057 95 288 hypothetical protein 4834849 4834562 10.9 10885 - 6.98 -1.70 0.20 11.665074 RB9070 409 1230 secreted protein containing DUF1559 4840267 4841496 8.6 44198 + 5.38 -2.02 0.24 11.995124 RB9160 118 357 hypothetical protein 4900491 4900847 10.8 12876 + 6.23 -1.47 0.00 0.345204 RB9322 42 129 hypothetical protein 4992459 4992331 7.3 4319 - 5.71 -1.24 0.10 8.015205 RB9324 267 804 secreted protein 4992474 4993277 6.7 29463 + 5.69 -1.32 0.18 13.555214 RB9343 59 180 hypothetical protein 5003789 5003968 11.4 6521 + 6.01 -1.31 0.02 1.695225 RB9354 183 552 hypothetical protein 5011855 5012406 10.5 20002 + 6.49 -1.04 0.15 14.625258 RB9417 103 312 hypothetical protein 5050621 5050932 10.5 11390 + 5.90 -1.38 0.17 12.165280 RB9460 79 240 hypothetical protein 5078860 5078621 10.3 8991 - 8.85 -2.89 0.23 8.115313 RB9520 532 1599 conserved hypothetical protein 5114495 5112897 4.4 59231 - 5.87 -1.36 0.19 14.275371 RB9624 357 1074 hypothetical protein 5181259 5180186 5.2 38849 - 5.76 -1.14 0.12 10.325380 RB9641 73 222 hypothetical protein 5189877 5190098 12.1 7925 + 5.89 -1.04 0.20 19.505419 RB9713 1082 3249 large, multifunctional secreted protein - Streptomyces coelicolor (fragment) 5243297 5240049 4.9 120210 - 5.83 -1.02 0.08 7.655469 RB9806 165 498 membrane protein 5280590 5281087 10.2 18166 + 5.83 -1.04 0.07 6.445488 RB9844 317 954 cytochrome c biogenesis protein, CcmF/CcyK/CcsA family 5301844 5302797 9.4 34187 + 6.03 -1.17 0.11 9.485498 RB9861 98 297 hypothetical protein 5313829 5314125 10.7 10762 + 5.83 -1.13 0.15 13.005503 RB9872 67 204 hypothetical protein 5316692 5316895 5.4 6912 + 5.99 -1.50 0.19 12.715644 RB10123 72 219 hypothetical protein 5463443 5463661 10.8 8181 + 5.72 -1.19 0.14 11.465662 RB10146 105 318 hypothetical protein 5476874 5476557 6.1 11800 - 5.33 -1.04 0.01 0.685694 RB10209 110 333 H+-transporting two-sector ATPase, C subunit 3.6.3.14 5515135 5515467 10.2 11565 + 6.44 -1.58 0.28 17.805837 RB10481 82 249 hypothetical protein 5685184 5685432 10.6 9492 + 5.77 -1.00 0.09 8.555855 RB10521 53 162 hypothetical protein 5708182 5708343 9.1 6074 + 6.54 -1.30 0.27 20.405885 RB10579 101 306 secreted protein 5739254 5738949 12.4 11507 - 7.16 -1.63 0.17 10.425887 RB10581 384 1155 secreted protein containing DUF1559 5739353 5740507 6.2 41431 + 7.14 -1.58 0.21 13.045908 RB10620 49 150 hypothetical protein 5768561 5768412 10.6 5707 - 6.80 -1.10 0.26 23.375933 RB10668 58 177 hypothetical protein 5796528 5796352 9.3 6645 - 5.93 -1.17 0.20 16.795937 RB10674 58 177 hypothetical protein 5800080 5799904 11.9 6706 - 5.59 -1.10 0.02 2.185962 RB10712 84 255 hypothetical protein 5824976 5825230 10.2 9185 + 5.83 -1.34 0.28 21.126048 RB10868 136 411 hypothetical protein 5900330 5899920 11.3 15100 - 6.78 -1.09 0.20 18.436280 RB11287 75 228 hypothetical protein 6118405 6118632 9.1 8478 + 5.42 -1.10 0.27 24.706343 RB11392 148 447 conserved hypothetical protein 6177208 6177654 5 16382 + 6.29 -1.37 0.05 3.616364 RB11429 37 114 conserved hypothetical protein 6196478 6196591 12.4 4245 + 6.85 -2.38 0.14 5.986367 RB11434 69 210 conserved hypothetical protein 6197442 6197233 9.8 7670 - 6.02 -1.16 0.20 16.896371 RB11439 93 282 conserved hypothetical protein 6197993 6198274 10.6 10906 + 8.25 -3.09 0.16 5.226385 RB11450 60 183 conserved hypothetical protein 6200046 6200228 11.8 6517 + 8.52 -2.44 0.17 6.896539 RB11707 83 252 conserved hypothetical protein 6312937 6313188 9.8 9109 + 5.60 -1.34 0.19 14.466569 RB11766 129 390 conserved hypothetical protein 6334355 6334744 10.4 14762 + 5.82 -1.20 0.26 21.356579 RB11804 132 399 conserved hypothetical protein 6363537 6363139 11.3 15013 - 5.94 -1.05 0.18 17.536623 RB11886 933 2802 protein containing DUF1585 6403682 6400881 5.1 105437 - 5.90 -1.00 0.19 18.576626 RB11897 253 762 conserved hypothetical protein 6407046 6407807 11.3 27903 + 5.67 -1.01 0.02 1.896697 RB12021 82 249 conserved hypothetical protein 6485018 6484770 11.9 9305 - 6.50 -1.93 0.23 11.746744 RB12114 110 333 conserved hypothetical protein 6540259 6539927 11.8 12716 - 5.85 -1.09 0.04 3.386781 RB12193 36 111 conserved hypothetical protein 6575977 6575867 7.5 3719 - 6.19 -1.89 0.19 10.316787 RB12201 69 210 conserved hypothetical protein 6578161 6577952 12.5 7444 - 7.03 -1.25 0.20 16.256822 RB12260 81 246 hypothetical protein-signal peptide prediction 6600777 6600532 11.9 9294 - 5.85 -1.18 0.11 9.436858 RB12319 187 564 conserved hypothetical protein 6626077 6625514 11.2 21283 - 5.86 -1.16 0.16 13.986867 RB12329 110 333 conserved hypothetical protein, membrane 6634234 6634566 4 11862 + 6.86 -2.13 0.23 10.846904 RB12396 57 174 conserved hypothetical protein 6670870 6670697 11.3 6680 - 5.99 -1.54 0.22 14.346920 RB12426 137 414 conserved hypothetical protein, membrane 6684059 6684472 10.6 15166 + 5.80 -1.06 0.09 8.886936 RB12447 103 312 conserved hypothetical protein 6700272 6700583 10.6 11445 + 6.03 -1.21 0.23 18.727009 RB12581 174 525 conserved hypothetical protein 6780597 6780073 9.9 19433 - 5.86 -1.27 0.17 13.187141 RB12818 163 492 conserved hypothetical protein 6915989 6915498 11.7 18010 - 7.56 -3.62 0.19 5.247153 RB12837 65 198 conserved hypothetical protein 6921732 6921929 9.8 7318 + 7.18 -1.45 0.16 10.887323 RB13171 43 132 conserved hypothetical protein 7069906 7069775 4.4 4834 - 5.69 -1.02 0.16 16.067342 RB13207 58 177 secreted protein 7086761 7086585 11.9 6443 - 6.10 -1.15 0.23 20.487403 RB13327 421 1266 protein containing DUF1559 7143601 7142336 6.9 45140 - 5.17 -1.28 0.06 5.02
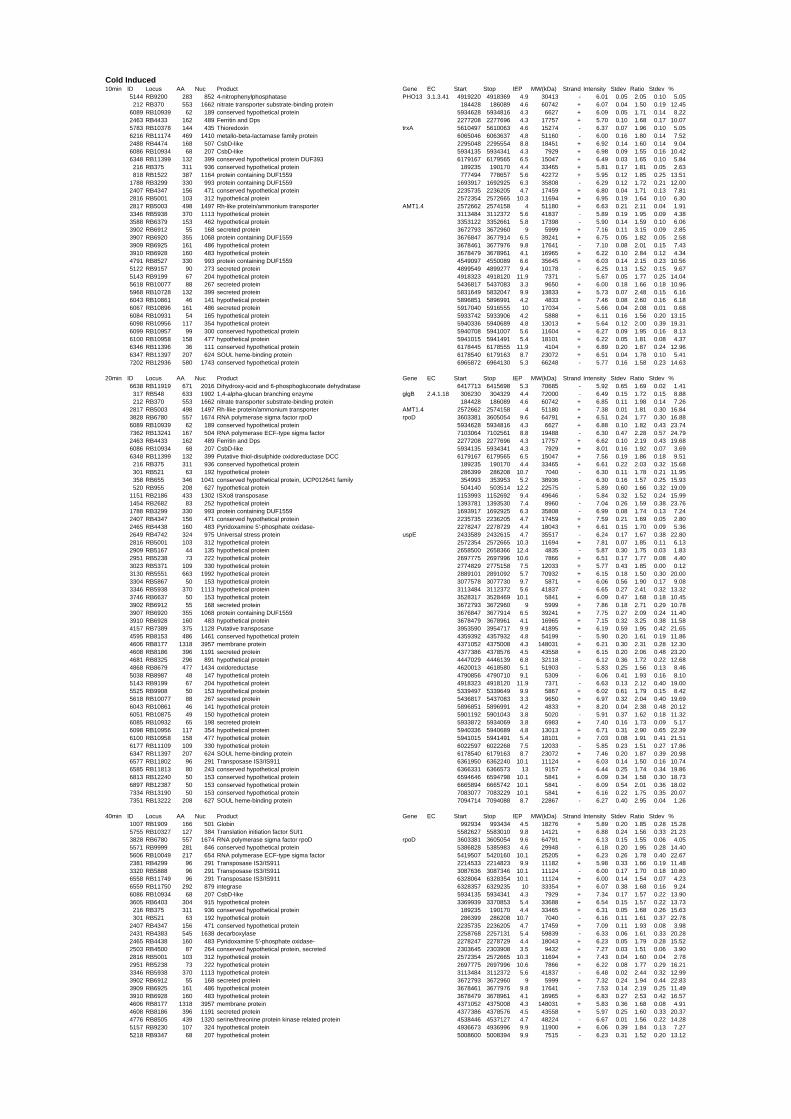
Cold Induced10min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand Intensity Stdev Ratio Stdev %
5144 RB9200 283 852 4-nitrophenylphosphatase PHO13 3.1.3.41 4919220 4918369 4.9 30413 - 6.01 0.05 2.05 0.10 5.05212 RB370 553 1662 nitrate transporter substrate-binding protein 184428 186089 4.6 60742 + 6.07 0.04 1.50 0.19 12.45
6089 RB10939 62 189 conserved hypothetical protein 5934628 5934816 4.3 6627 + 6.09 0.05 1.71 0.14 8.222463 RB4433 162 489 Ferritin and Dps 2277208 2277696 4.3 17757 + 5.70 0.10 1.68 0.17 10.075783 RB10378 144 435 Thioredoxin trxA 5610497 5610063 4.6 15274 - 6.37 0.07 1.96 0.10 5.056216 RB11174 469 1410 metallo-beta-lactamase family protein 6065046 6063637 4.8 51160 - 6.00 0.16 1.80 0.14 7.522488 RB4474 168 507 CsbD-like 2295048 2295554 8.8 18451 + 6.92 0.14 1.60 0.14 9.046086 RB10934 68 207 CsbD-like 5934135 5934341 4.3 7929 + 6.98 0.09 1.55 0.16 10.426348 RB11399 132 399 conserved hypothetical protein DUF393 6179167 6179565 6.5 15047 + 6.49 0.03 1.65 0.10 5.84216 RB375 311 936 conserved hypothetical protein 189235 190170 4.4 33465 + 5.81 0.17 1.81 0.05 2.63818 RB1522 387 1164 protein containing DUF1559 777494 778657 5.6 42272 + 5.95 0.12 1.85 0.25 13.51
1788 RB3299 330 993 protein containing DUF1559 1693917 1692925 6.3 35808 - 6.29 0.12 1.72 0.21 12.002407 RB4347 156 471 conserved hypothetical protein 2235735 2236205 4.7 17459 + 6.80 0.04 1.71 0.13 7.812816 RB5001 103 312 hypothetical protein 2572354 2572665 10.3 11694 + 6.95 0.19 1.64 0.10 6.302817 RB5003 498 1497 Rh-like protein/ammonium transporter AMT1.4 2572662 2574158 4 51180 + 6.63 0.21 2.11 0.04 1.913346 RB5938 370 1113 hypothetical protein 3113484 3112372 5.6 41837 - 5.89 0.19 1.95 0.09 4.383588 RB6379 153 462 hypothetical protein 3353122 3352661 5.8 17398 - 5.90 0.14 1.59 0.10 6.063902 RB6912 55 168 secreted protein 3672793 3672960 9 5999 + 7.16 0.11 3.15 0.09 2.853907 RB6920 355 1068 protein containing DUF1559 3676847 3677914 6.5 39241 + 6.75 0.05 1.82 0.05 2.583909 RB6925 161 486 hypothetical protein 3678461 3677976 9.8 17641 - 7.10 0.08 2.01 0.15 7.433910 RB6928 160 483 hypothetical protein 3678479 3678961 4.1 16965 + 6.22 0.10 2.84 0.12 4.344791 RB8527 330 993 protein containing DUF1559 4549097 4550089 6.6 35645 + 6.03 0.14 2.15 0.23 10.565122 RB9157 90 273 secreted protein 4899549 4899277 9.4 10178 - 6.25 0.13 1.52 0.15 9.675143 RB9199 67 204 hypothetical protein 4918323 4918120 11.9 7371 - 5.67 0.05 1.77 0.25 14.045618 RB10077 88 267 secreted protein 5436817 5437083 3.3 9650 + 6.00 0.18 1.66 0.18 10.965968 RB10728 132 399 secreted protein 5831649 5832047 9.9 13833 + 5.73 0.07 2.48 0.15 6.166043 RB10861 46 141 hypothetical protein 5896851 5896991 4.2 4833 + 7.46 0.08 2.60 0.16 6.186067 RB10896 161 486 secreted protein 5917040 5916555 10 17034 - 5.66 0.04 2.08 0.01 0.686084 RB10931 54 165 hypothetical protein 5933742 5933906 4.2 5888 + 6.11 0.16 1.56 0.20 13.156098 RB10956 117 354 hypothetical protein 5940336 5940689 4.8 13013 + 5.64 0.12 2.00 0.39 19.316099 RB10957 99 300 conserved hypothetical protein 5940708 5941007 5.6 11604 + 6.27 0.09 1.95 0.16 8.136100 RB10958 158 477 hypothetical protein 5941015 5941491 5.4 18101 + 6.22 0.05 1.81 0.08 4.376346 RB11396 36 111 conserved hypothetical protein 6178445 6178555 11.9 4104 + 6.89 0.20 1.87 0.24 12.966347 RB11397 207 624 SOUL heme-binding protein 6178540 6179163 8.7 23072 + 6.51 0.04 1.78 0.10 5.417202 RB12936 580 1743 conserved hypothetical protein 6965872 6964130 5.3 66248 - 5.77 0.16 1.58 0.23 14.63
20min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand Intensity Stdev Ratio Stdev %6638 RB11919 671 2016 Dihydroxy-acid and 6-phosphogluconate dehydratase 6417713 6415698 5.3 70685 - 5.92 0.65 1.69 0.02 1.41317 RB548 633 1902 1,4-alpha-glucan branching enzyme glgB 2.4.1.18 306230 304329 4.4 72000 - 6.49 0.15 1.72 0.15 8.88212 RB370 553 1662 nitrate transporter substrate-binding protein 184428 186089 4.6 60742 + 6.85 0.11 1.98 0.14 7.26
2817 RB5003 498 1497 Rh-like protein/ammonium transporter AMT1.4 2572662 2574158 4 51180 + 7.38 0.01 1.81 0.30 16.843828 RB6780 557 1674 RNA polymerase sigma factor rpoD rpoD 3603381 3605054 9.6 64791 + 6.51 0.24 1.77 0.30 16.886089 RB10939 62 189 conserved hypothetical protein 5934628 5934816 4.3 6627 + 6.88 0.10 1.82 0.43 23.747362 RB13241 167 504 RNA polymerase ECF-type sigma factor 7103064 7102561 8.8 19488 - 6.30 0.47 2.28 0.57 24.792463 RB4433 162 489 Ferritin and Dps 2277208 2277696 4.3 17757 + 6.62 0.10 2.19 0.43 19.686086 RB10934 68 207 CsbD-like 5934135 5934341 4.3 7929 + 8.01 0.16 1.92 0.07 3.696348 RB11399 132 399 Putative thiol-disulphide oxidoreductase DCC 6179167 6179565 6.5 15047 + 7.56 0.19 1.86 0.18 9.51216 RB375 311 936 conserved hypothetical protein 189235 190170 4.4 33465 + 6.61 0.22 2.03 0.32 15.68301 RB521 63 192 hypothetical protein 286399 286208 10.7 7040 - 6.30 0.11 1.78 0.21 11.95358 RB655 346 1041 conserved hypothetical protein, UCP012641 family 354993 353953 5.2 38936 - 6.30 0.16 1.57 0.25 15.93520 RB955 208 627 hypothetical protein 504140 503514 12.2 22575 - 5.89 0.60 1.66 0.32 19.09
1151 RB2186 433 1302 ISXo8 transposase 1153993 1152692 9.4 49646 - 5.84 0.32 1.52 0.24 15.991454 RB2682 83 252 hypothetical protein 1393781 1393530 7.4 8960 - 7.04 0.26 1.59 0.38 23.761788 RB3299 330 993 protein containing DUF1559 1693917 1692925 6.3 35808 - 6.99 0.08 1.74 0.13 7.242407 RB4347 156 471 conserved hypothetical protein 2235735 2236205 4.7 17459 + 7.59 0.21 1.69 0.05 2.802465 RB4438 160 483 Pyridoxamine 5'-phosphate oxidase- 2278247 2278729 4.4 18043 + 6.61 0.15 1.70 0.09 5.362649 RB4742 324 975 Universal stress protein uspE 2433589 2432615 4.7 35517 - 6.24 0.17 1.67 0.38 22.802816 RB5001 103 312 hypothetical protein 2572354 2572665 10.3 11694 + 7.81 0.07 1.85 0.11 6.132909 RB5167 44 135 hypothetical protein 2658500 2658366 12.4 4835 - 5.87 0.30 1.75 0.03 1.832951 RB5238 73 222 hypothetical protein 2697775 2697996 10.6 7866 + 6.51 0.17 1.77 0.08 4.403023 RB5371 109 330 hypothetical protein 2774829 2775158 7.5 12033 + 5.77 0.43 1.85 0.00 0.123130 RB5551 663 1992 hypothetical protein 2889101 2891092 5.7 70932 + 6.15 0.18 1.50 0.30 20.003304 RB5867 50 153 hypothetical protein 3077578 3077730 9.7 5871 + 6.06 0.56 1.90 0.17 9.083346 RB5938 370 1113 hypothetical protein 3113484 3112372 5.6 41837 - 6.65 0.27 2.41 0.32 13.323746 RB6637 50 153 hypothetical protein 3528317 3528469 10.1 5841 + 6.09 0.47 1.68 0.18 10.453902 RB6912 55 168 secreted protein 3672793 3672960 9 5999 + 7.86 0.18 2.71 0.29 10.783907 RB6920 355 1068 protein containing DUF1559 3676847 3677914 6.5 39241 + 7.75 0.27 2.09 0.24 11.403910 RB6928 160 483 hypothetical protein 3678479 3678961 4.1 16965 + 7.15 0.32 3.25 0.38 11.584157 RB7389 375 1128 Putative transposase 3953590 3954717 9.9 41895 + 6.19 0.59 1.95 0.42 21.654595 RB8153 486 1461 conserved hypothetical protein 4359392 4357932 4.8 54199 - 5.90 0.20 1.61 0.19 11.864606 RB8177 1318 3957 membrane protein 4371052 4375008 4.3 148031 + 6.21 0.30 2.31 0.28 12.304608 RB8186 396 1191 secreted protein 4377386 4378576 4.5 43558 + 6.15 0.20 2.06 0.48 23.204681 RB8325 296 891 hypothetical protein 4447029 4446139 6.8 32118 - 6.12 0.36 1.72 0.22 12.684868 RB8679 477 1434 oxidoreductase 4620013 4618580 5.1 51903 - 5.83 0.25 1.56 0.13 8.465038 RB8987 48 147 hypothetical protein 4790856 4790710 9.1 5309 - 6.06 0.41 1.93 0.16 8.105143 RB9199 67 204 hypothetical protein 4918323 4918120 11.9 7371 - 6.63 0.13 2.12 0.40 19.005525 RB9908 50 153 hypothetical protein 5339497 5339649 9.9 5867 + 6.02 0.61 1.79 0.15 8.425618 RB10077 88 267 secreted protein 5436817 5437083 3.3 9650 + 6.97 0.32 2.04 0.40 19.696043 RB10861 46 141 hypothetical protein 5896851 5896991 4.2 4833 + 8.20 0.04 2.38 0.48 20.126051 RB10875 49 150 hypothetical protein 5901192 5901043 3.8 5020 - 5.91 0.37 1.62 0.18 11.326085 RB10932 65 198 secreted protein 5933872 5934069 3.8 6983 + 7.40 0.16 1.73 0.09 5.176098 RB10956 117 354 hypothetical protein 5940336 5940689 4.8 13013 + 6.71 0.31 2.90 0.65 22.396100 RB10958 158 477 hypothetical protein 5941015 5941491 5.4 18101 + 7.03 0.08 1.91 0.41 21.516177 RB11109 109 330 hypothetical protein 6022597 6022268 7.5 12033 - 5.85 0.23 1.51 0.27 17.866347 RB11397 207 624 SOUL heme-binding protein 6178540 6179163 8.7 23072 + 7.46 0.20 1.87 0.39 20.986577 RB11802 96 291 Transposase IS3/IS911 6361950 6362240 10.1 11124 + 6.03 0.14 1.50 0.16 10.746585 RB11813 80 243 conserved hypothetical protein 6366331 6366573 13 9157 + 6.44 0.25 1.74 0.34 19.866813 RB12240 50 153 conserved hypothetical protein 6594646 6594798 10.1 5841 + 6.09 0.34 1.58 0.30 18.736897 RB12387 50 153 conserved hypothetical protein 6665894 6665742 10.1 5841 - 6.09 0.54 2.01 0.36 18.027334 RB13190 50 153 conserved hypothetical protein 7083077 7083229 10.1 5841 + 6.16 0.22 1.75 0.35 20.077351 RB13222 208 627 SOUL heme-binding protein 7094714 7094088 8.7 22867 - 6.27 0.40 2.95 0.04 1.26
40min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand Intensity Stdev Ratio Stdev %1007 RB1909 166 501 Globin 992934 993434 4.5 18276 + 5.89 0.20 1.85 0.28 15.285755 RB10327 127 384 Translation initiation factor SUI1 5582627 5583010 9.8 14121 + 6.88 0.24 1.56 0.33 21.233828 RB6780 557 1674 RNA polymerase sigma factor rpoD rpoD 3603381 3605054 9.6 64791 + 6.13 0.15 1.55 0.06 4.055571 RB9999 281 846 conserved hypothetical protein 5386828 5385983 4.6 29948 - 6.18 0.20 1.95 0.28 14.405606 RB10049 217 654 RNA polymerase ECF-type sigma factor 5419507 5420160 10.1 25205 + 6.23 0.26 1.78 0.40 22.672381 RB4299 96 291 Transposase IS3/IS911 2214533 2214823 9.9 11182 + 5.98 0.33 1.66 0.19 11.483320 RB5888 96 291 Transposase IS3/IS911 3087636 3087346 10.1 11124 - 6.00 0.17 1.70 0.18 10.806558 RB11749 96 291 Transposase IS3/IS911 6328064 6328354 10.1 11124 + 6.00 0.14 1.54 0.07 4.236559 RB11750 292 879 integrase 6328357 6329235 10 33354 + 6.07 0.38 1.68 0.16 9.246086 RB10934 68 207 CsbD-like 5934135 5934341 4.3 7929 + 7.34 0.17 1.57 0.22 13.903605 RB6403 304 915 hypothetical protein 3369939 3370853 5.4 33688 + 6.54 0.15 1.57 0.22 13.73216 RB375 311 936 conserved hypothetical protein 189235 190170 4.4 33465 + 6.31 0.05 1.68 0.26 15.63301 RB521 63 192 hypothetical protein 286399 286208 10.7 7040 - 6.16 0.11 1.61 0.37 22.78
2407 RB4347 156 471 conserved hypothetical protein 2235735 2236205 4.7 17459 + 7.09 0.11 1.93 0.08 3.982431 RB4383 545 1638 decarboxylase 2258768 2257131 5.4 59839 - 6.33 0.06 1.61 0.33 20.282465 RB4438 160 483 Pyridoxamine 5'-phosphate oxidase- 2278247 2278729 4.4 18043 + 6.23 0.05 1.79 0.28 15.522503 RB4500 87 264 conserved hypothetical protein, secreted 2303645 2303908 3.5 9432 + 7.27 0.03 1.51 0.06 3.902816 RB5001 103 312 hypothetical protein 2572354 2572665 10.3 11694 + 7.43 0.04 1.60 0.04 2.782951 RB5238 73 222 hypothetical protein 2697775 2697996 10.6 7866 + 6.22 0.08 1.77 0.29 16.213346 RB5938 370 1113 hypothetical protein 3113484 3112372 5.6 41837 - 6.48 0.02 2.44 0.32 12.993902 RB6912 55 168 secreted protein 3672793 3672960 9 5999 + 7.32 0.24 1.94 0.44 22.833909 RB6925 161 486 hypothetical protein 3678461 3677976 9.8 17641 - 7.53 0.14 2.19 0.25 11.493910 RB6928 160 483 hypothetical protein 3678479 3678961 4.1 16965 + 6.83 0.27 2.53 0.42 16.574606 RB8177 1318 3957 membrane protein 4371052 4375008 4.3 148031 + 5.83 0.36 1.68 0.08 4.914608 RB8186 396 1191 secreted protein 4377386 4378576 4.5 43558 + 5.97 0.25 1.60 0.33 20.374776 RB8505 439 1320 serine/threonine protein kinase related protein 4538446 4537127 4.7 48224 - 6.67 0.01 1.56 0.22 14.285157 RB9230 107 324 hypothetical protein 4936673 4936996 9.9 11900 + 6.06 0.39 1.84 0.13 7.275218 RB9347 68 207 hypothetical protein 5008600 5008394 9.9 7515 - 6.23 0.31 1.52 0.20 13.12
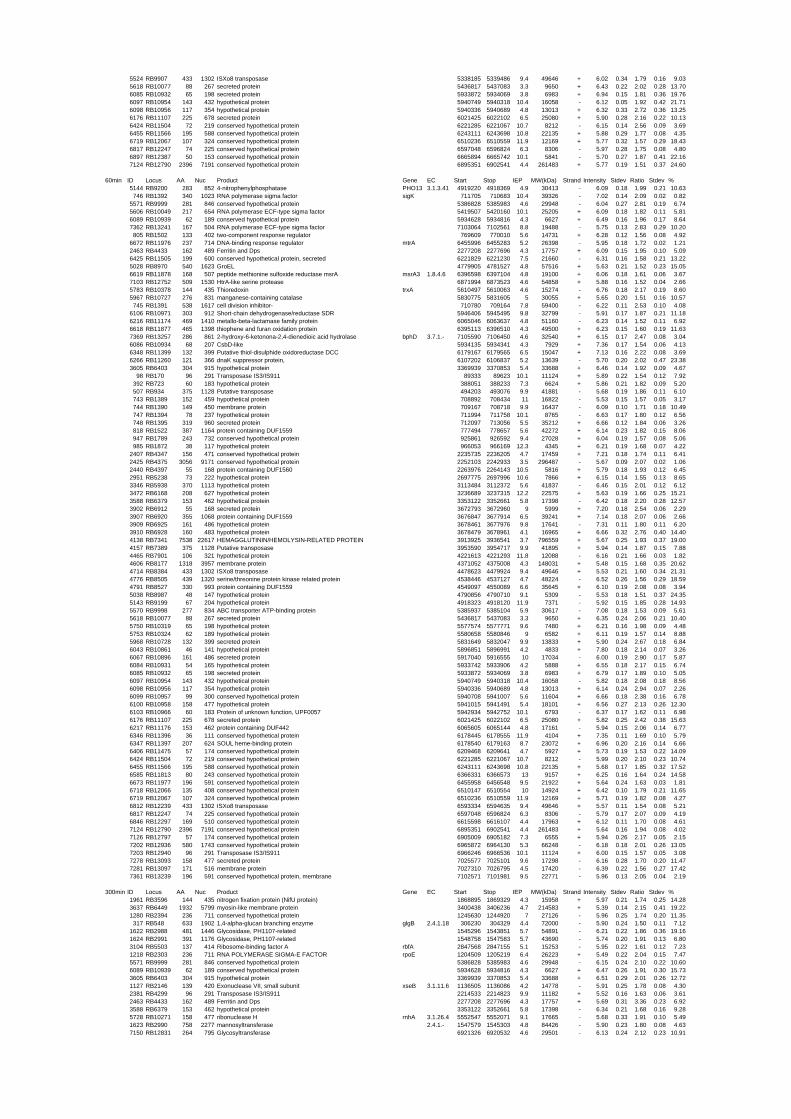
5524 RB9907 433 1302 ISXo8 transposase 5338185 5339486 9.4 49646 + 6.02 0.34 1.79 0.16 9.035618 RB10077 88 267 secreted protein 5436817 5437083 3.3 9650 + 6.43 0.22 2.02 0.28 13.706085 RB10932 65 198 secreted protein 5933872 5934069 3.8 6983 + 6.94 0.15 1.81 0.36 19.766097 RB10954 143 432 hypothetical protein 5940749 5940318 10.4 16058 - 6.12 0.05 1.92 0.42 21.716098 RB10956 117 354 hypothetical protein 5940336 5940689 4.8 13013 + 6.32 0.33 2.72 0.36 13.256176 RB11107 225 678 secreted protein 6021425 6022102 6.5 25080 + 5.90 0.28 2.16 0.22 10.136424 RB11504 72 219 conserved hypothetical protein 6221285 6221067 10.7 8212 - 6.15 0.14 2.56 0.09 3.696455 RB11566 195 588 conserved hypothetical protein 6243111 6243698 10.8 22135 + 5.88 0.29 1.77 0.08 4.356719 RB12067 107 324 conserved hypothetical protein 6510236 6510559 11.9 12169 + 5.77 0.32 1.57 0.29 18.436817 RB12247 74 225 conserved hypothetical protein 6597048 6596824 6.3 8306 - 5.97 0.28 1.75 0.08 4.806897 RB12387 50 153 conserved hypothetical protein 6665894 6665742 10.1 5841 - 5.70 0.27 1.87 0.41 22.167124 RB12790 2396 7191 conserved hypothetical protein 6895351 6902541 4.4 261483 + 5.77 0.19 1.51 0.37 24.60
60min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand Intensity Stdev Ratio Stdev %5144 RB9200 283 852 4-nitrophenylphosphatase PHO13 3.1.3.41 4919220 4918369 4.9 30413 - 6.09 0.18 1.99 0.21 10.63746 RB1392 340 1023 RNA polymerase sigma factor sigK 711705 710683 10.4 39326 - 7.02 0.14 2.09 0.02 0.82
5571 RB9999 281 846 conserved hypothetical protein 5386828 5385983 4.6 29948 - 6.04 0.27 2.81 0.19 6.745606 RB10049 217 654 RNA polymerase ECF-type sigma factor 5419507 5420160 10.1 25205 + 6.09 0.18 1.82 0.11 5.816089 RB10939 62 189 conserved hypothetical protein 5934628 5934816 4.3 6627 + 6.49 0.16 1.96 0.17 8.647362 RB13241 167 504 RNA polymerase ECF-type sigma factor 7103064 7102561 8.8 19488 - 5.75 0.13 2.83 0.29 10.20805 RB1502 133 402 two-component response regulator 769609 770010 5.6 14731 + 6.28 0.12 1.56 0.08 4.92
6672 RB11976 237 714 DNA-binding response regulator mtrA 6455996 6455283 5.2 26398 - 5.95 0.18 1.72 0.02 1.212463 RB4433 162 489 Ferritin and Dps 2277208 2277696 4.3 17757 + 6.09 0.15 1.95 0.10 5.096425 RB11505 199 600 conserved hypothetical protein, secreted 6221829 6221230 7.5 21660 - 6.31 0.16 1.58 0.21 13.225028 RB8970 540 1623 GroEL 4779905 4781527 4.8 57516 + 5.63 0.21 1.52 0.23 15.056619 RB11878 168 507 peptide methionine sulfoxide reductase msrA msrA3 1.8.4.6 6396598 6397104 4.8 19100 + 6.06 0.18 1.61 0.06 3.677103 RB12752 509 1530 HtrA-like serine protease 6871994 6873523 4.6 54858 + 5.88 0.16 1.52 0.04 2.665783 RB10378 144 435 Thioredoxin trxA 5610497 5610063 4.6 15274 - 6.76 0.18 2.17 0.19 8.605967 RB10727 276 831 manganese-containing catalase 5830775 5831605 5 30055 + 5.65 0.20 1.51 0.16 10.57745 RB1391 538 1617 cell division inhibitor- 710780 709164 7.8 59400 - 6.22 0.11 2.53 0.10 4.08
6106 RB10971 303 912 Short-chain dehydrogenase/reductase SDR 5946406 5945495 9.8 32799 - 5.91 0.17 1.87 0.21 11.186216 RB11174 469 1410 metallo-beta-lactamase family protein 6065046 6063637 4.8 51160 - 6.23 0.14 1.52 0.11 6.926618 RB11877 465 1398 thiophene and furan oxidation protein 6395113 6396510 4.3 49500 + 6.23 0.15 1.60 0.19 11.637369 RB13257 286 861 2-hydroxy-6-ketonona-2,4-dienedioic acid hydrolase bphD 3.7.1.- 7105590 7106450 4.6 32540 + 6.15 0.17 2.47 0.08 3.046086 RB10934 68 207 CsbD-like 5934135 5934341 4.3 7929 + 7.36 0.17 1.54 0.06 4.136348 RB11399 132 399 Putative thiol-disulphide oxidoreductase DCC 6179167 6179565 6.5 15047 + 7.13 0.16 2.22 0.08 3.696266 RB11260 121 366 dnaK suppressor protein, 6107202 6106837 5.2 13639 - 5.70 0.20 2.02 0.47 23.383605 RB6403 304 915 hypothetical protein 3369939 3370853 5.4 33688 + 6.46 0.14 1.92 0.09 4.67
98 RB170 96 291 Transposase IS3/IS911 89333 89623 10.1 11124 + 5.89 0.22 1.54 0.12 7.92392 RB723 60 183 hypothetical protein 388051 388233 7.3 6624 + 5.86 0.21 1.82 0.09 5.20507 RB934 375 1128 Putative transposase 494203 493076 9.9 41881 - 5.68 0.19 1.86 0.11 6.10743 RB1389 152 459 hypothetical protein 708892 708434 11 16822 - 5.53 0.15 1.57 0.05 3.17744 RB1390 149 450 membrane protein 709167 708718 9.9 16437 - 6.09 0.10 1.71 0.18 10.49747 RB1394 78 237 hypothetical protein 711994 711758 10.1 8765 - 6.63 0.17 1.80 0.12 6.56748 RB1395 319 960 secreted protein 712097 713056 5.5 35212 + 6.66 0.12 1.84 0.06 3.26818 RB1522 387 1164 protein containing DUF1559 777494 778657 5.6 42272 + 6.14 0.23 1.82 0.15 8.06947 RB1789 243 732 conserved hypothetical protein 925861 926592 9.4 27028 + 6.04 0.19 1.57 0.08 5.06985 RB1872 38 117 hypothetical protein 966053 966169 12.3 4345 + 6.21 0.19 1.68 0.07 4.22
2407 RB4347 156 471 conserved hypothetical protein 2235735 2236205 4.7 17459 + 7.21 0.18 1.74 0.11 6.412425 RB4375 3056 9171 conserved hypothetical protein 2252103 2242933 3.5 296487 - 5.67 0.09 2.07 0.02 1.062440 RB4397 55 168 protein containing DUF1560 2263976 2264143 10.5 5816 + 5.79 0.18 1.93 0.12 6.452951 RB5238 73 222 hypothetical protein 2697775 2697996 10.6 7866 + 6.15 0.14 1.55 0.13 8.653346 RB5938 370 1113 hypothetical protein 3113484 3112372 5.6 41837 - 6.46 0.15 2.01 0.12 6.123472 RB6168 208 627 hypothetical protein 3236689 3237315 12.2 22575 + 5.63 0.19 1.66 0.25 15.213588 RB6379 153 462 hypothetical protein 3353122 3352661 5.8 17398 - 6.42 0.18 2.20 0.28 12.573902 RB6912 55 168 secreted protein 3672793 3672960 9 5999 + 7.20 0.18 2.54 0.06 2.293907 RB6920 355 1068 protein containing DUF1559 3676847 3677914 6.5 39241 + 7.14 0.18 2.07 0.06 2.663909 RB6925 161 486 hypothetical protein 3678461 3677976 9.8 17641 - 7.31 0.11 1.80 0.11 6.203910 RB6928 160 483 hypothetical protein 3678479 3678961 4.1 16965 + 6.66 0.32 2.76 0.40 14.404138 RB7341 7538 22617 HEMAGGLUTININ/HEMOLYSIN-RELATED PROTEIN 3913925 3936541 3.7 796559 + 5.67 0.25 1.93 0.37 19.004157 RB7389 375 1128 Putative transposase 3953590 3954717 9.9 41895 + 5.94 0.14 1.87 0.15 7.884465 RB7901 106 321 hypothetical protein 4221613 4221293 11.8 12088 - 6.16 0.21 1.66 0.03 1.824606 RB8177 1318 3957 membrane protein 4371052 4375008 4.3 148031 + 5.48 0.15 1.68 0.35 20.624714 RB8384 433 1302 ISXo8 transposase 4478623 4479924 9.4 49646 + 5.53 0.21 1.60 0.34 21.314776 RB8505 439 1320 serine/threonine protein kinase related protein 4538446 4537127 4.7 48224 - 6.52 0.26 1.56 0.29 18.594791 RB8527 330 993 protein containing DUF1559 4549097 4550089 6.6 35645 + 6.10 0.19 2.08 0.08 3.945038 RB8987 48 147 hypothetical protein 4790856 4790710 9.1 5309 - 5.53 0.18 1.51 0.37 24.355143 RB9199 67 204 hypothetical protein 4918323 4918120 11.9 7371 - 5.92 0.15 1.85 0.28 14.935570 RB9998 277 834 ABC transporter ATP-binding protein 5385937 5385104 5.9 30617 - 7.08 0.18 1.53 0.09 5.615618 RB10077 88 267 secreted protein 5436817 5437083 3.3 9650 + 6.35 0.24 2.06 0.21 10.405750 RB10319 65 198 hypothetical protein 5577574 5577771 9.6 7480 + 6.21 0.16 1.98 0.09 4.485753 RB10324 62 189 hypothetical protein 5580658 5580846 9 6582 + 6.11 0.19 1.57 0.14 8.885968 RB10728 132 399 secreted protein 5831649 5832047 9.9 13833 + 5.90 0.24 2.67 0.18 6.846043 RB10861 46 141 hypothetical protein 5896851 5896991 4.2 4833 + 7.80 0.18 2.14 0.07 3.266067 RB10896 161 486 secreted protein 5917040 5916555 10 17034 - 6.00 0.19 2.90 0.17 5.876084 RB10931 54 165 hypothetical protein 5933742 5933906 4.2 5888 + 6.55 0.18 2.17 0.15 6.746085 RB10932 65 198 secreted protein 5933872 5934069 3.8 6983 + 6.79 0.17 1.89 0.10 5.056097 RB10954 143 432 hypothetical protein 5940749 5940318 10.4 16058 - 5.82 0.18 2.08 0.18 8.566098 RB10956 117 354 hypothetical protein 5940336 5940689 4.8 13013 + 6.14 0.24 2.94 0.07 2.266099 RB10957 99 300 conserved hypothetical protein 5940708 5941007 5.6 11604 + 6.66 0.18 2.38 0.16 6.786100 RB10958 158 477 hypothetical protein 5941015 5941491 5.4 18101 + 6.56 0.27 2.13 0.26 12.306103 RB10966 60 183 Protein of unknown function, UPF0057 5942934 5942752 10.1 6793 - 6.37 0.17 1.62 0.11 6.986176 RB11107 225 678 secreted protein 6021425 6022102 6.5 25080 + 5.82 0.25 2.42 0.38 15.636217 RB11176 153 462 protein containing DUF442 6065605 6065144 4.8 17161 - 5.94 0.15 2.06 0.14 6.776346 RB11396 36 111 conserved hypothetical protein 6178445 6178555 11.9 4104 + 7.35 0.11 1.69 0.10 5.796347 RB11397 207 624 SOUL heme-binding protein 6178540 6179163 8.7 23072 + 6.96 0.20 2.16 0.14 6.666406 RB11475 57 174 conserved hypothetical protein 6209468 6209641 4.7 5927 + 5.73 0.19 1.53 0.22 14.096424 RB11504 72 219 conserved hypothetical protein 6221285 6221067 10.7 8212 - 5.99 0.20 2.10 0.23 10.746455 RB11566 195 588 conserved hypothetical protein 6243111 6243698 10.8 22135 + 5.68 0.17 1.85 0.32 17.526585 RB11813 80 243 conserved hypothetical protein 6366331 6366573 13 9157 + 6.25 0.16 1.64 0.24 14.586673 RB11977 196 591 conserved hypothetical protein 6455958 6456548 9.5 21922 + 5.64 0.24 1.63 0.03 1.816718 RB12066 135 408 conserved hypothetical protein 6510147 6510554 10 14924 + 6.42 0.10 1.79 0.21 11.656719 RB12067 107 324 conserved hypothetical protein 6510236 6510559 11.9 12169 + 5.71 0.19 1.82 0.08 4.276812 RB12239 433 1302 ISXo8 transposase 6593334 6594635 9.4 49646 + 5.57 0.11 1.54 0.08 5.216817 RB12247 74 225 conserved hypothetical protein 6597048 6596824 6.3 8306 - 5.79 0.17 2.07 0.09 4.196846 RB12297 169 510 conserved hypothetical protein 6615598 6616107 4.4 17963 + 6.12 0.11 1.70 0.08 4.617124 RB12790 2396 7191 conserved hypothetical protein 6895351 6902541 4.4 261483 + 5.64 0.16 1.94 0.08 4.027126 RB12797 57 174 conserved hypothetical protein 6905009 6905182 7.3 6555 + 5.94 0.26 2.17 0.05 2.157202 RB12936 580 1743 conserved hypothetical protein 6965872 6964130 5.3 66248 - 6.18 0.18 2.01 0.26 13.057203 RB12940 96 291 Transposase IS3/IS911 6966246 6966536 10.1 11124 + 6.00 0.15 1.57 0.05 3.087278 RB13093 158 477 secreted protein 7025577 7025101 9.6 17298 - 6.16 0.28 1.70 0.20 11.477281 RB13097 171 516 membrane protein 7027310 7026795 4.5 17420 - 6.39 0.22 1.56 0.27 17.427361 RB13239 196 591 conserved hypothetical protein, membrane 7102571 7101981 9.5 22771 - 5.96 0.13 2.05 0.04 2.19
300min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand Intensity Stdev Ratio Stdev %1961 RB3596 144 435 nitrogen fixation protein (NifU protein) 1868895 1869329 4.3 15958 + 5.97 0.21 1.74 0.25 14.283637 RB6449 1932 5799 myosin-like membrane protein 3400438 3406236 4.7 214583 + 5.39 0.14 2.15 0.41 19.221280 RB2394 236 711 conserved hypothetical protein 1245630 1244920 7 27126 - 5.96 0.25 1.74 0.20 11.35317 RB548 633 1902 1,4-alpha-glucan branching enzyme glgB 2.4.1.18 306230 304329 4.4 72000 - 5.90 0.24 1.50 0.11 7.12
1622 RB2988 481 1446 Glycosidase, PH1107-related 1545296 1543851 5.7 54891 - 6.21 0.22 1.86 0.36 19.161624 RB2991 391 1176 Glycosidase, PH1107-related 1548758 1547583 5.7 43690 - 5.74 0.20 1.91 0.13 6.803104 RB5503 137 414 Ribosome-binding factor A rbfA 2847568 2847155 5.1 15253 - 5.95 0.22 1.61 0.12 7.231218 RB2303 236 711 RNA POLYMERASE SIGMA-E FACTOR rpoE 1204509 1205219 6.4 26223 + 5.49 0.22 2.04 0.15 7.475571 RB9999 281 846 conserved hypothetical protein 5386828 5385983 4.6 29948 - 6.15 0.24 2.10 0.22 10.606089 RB10939 62 189 conserved hypothetical protein 5934628 5934816 4.3 6627 + 6.47 0.26 1.91 0.30 15.733605 RB6403 304 915 hypothetical protein 3369939 3370853 5.4 33688 + 6.51 0.29 2.01 0.26 12.721127 RB2146 139 420 Exonuclease VII, small subunit xseB 3.1.11.6 1136505 1136086 4.2 14778 - 5.91 0.25 1.78 0.08 4.302381 RB4299 96 291 Transposase IS3/IS911 2214533 2214823 9.9 11182 + 5.52 0.16 1.63 0.06 3.612463 RB4433 162 489 Ferritin and Dps 2277208 2277696 4.3 17757 + 5.69 0.31 3.36 0.23 6.923588 RB6379 153 462 hypothetical protein 3353122 3352661 5.8 17398 - 6.34 0.21 1.68 0.16 9.285728 RB10271 158 477 ribonuclease H rnhA 3.1.26.4 5552547 5552071 9.1 17665 - 5.68 0.33 1.91 0.10 5.491623 RB2990 758 2277 mannosyltransferase 2.4.1.- 1547579 1545303 4.8 84426 - 5.90 0.23 1.80 0.08 4.637150 RB12831 264 795 Glycosyltransferase 6921326 6920532 4.6 29501 - 6.13 0.24 2.12 0.23 10.91
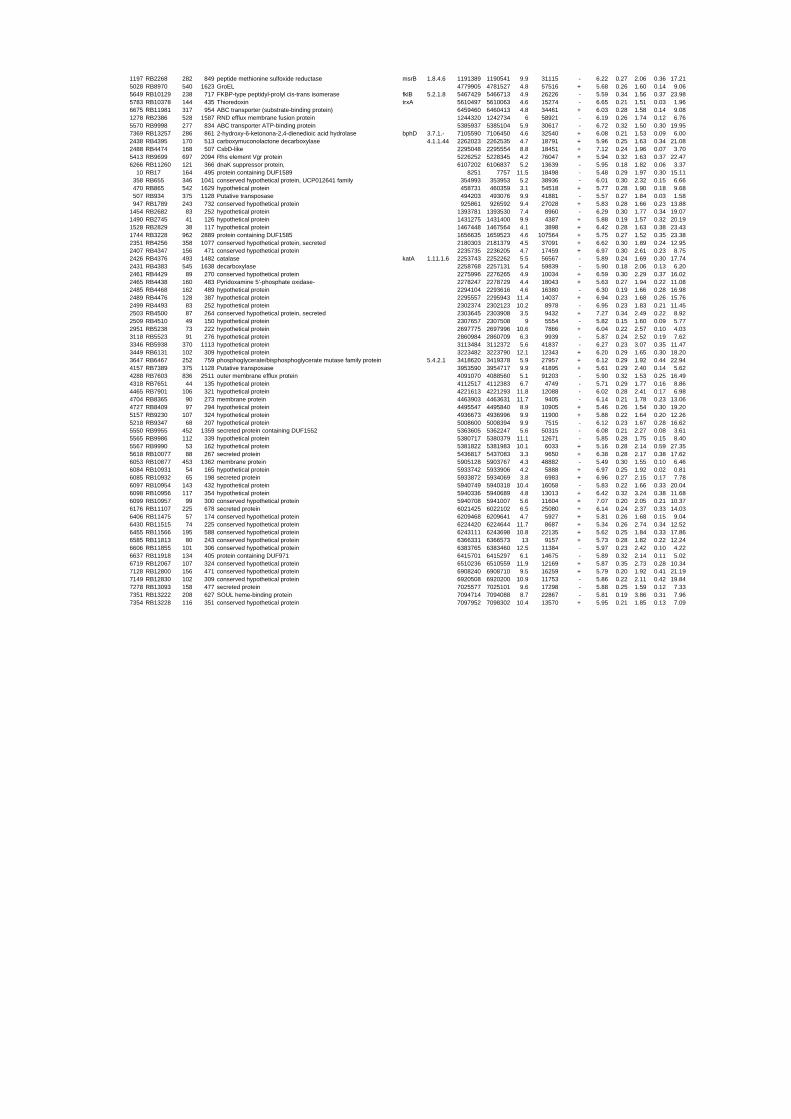
1197 RB2268 282 849 peptide methionine sulfoxide reductase msrB 1.8.4.6 1191389 1190541 9.9 31115 - 6.22 0.27 2.06 0.36 17.215028 RB8970 540 1623 GroEL 4779905 4781527 4.8 57516 + 5.68 0.26 1.60 0.14 9.065649 RB10129 238 717 FKBP-type peptidyl-prolyl cis-trans isomerase fklB 5.2.1.8 5467429 5466713 4.9 26226 - 5.59 0.34 1.56 0.37 23.985783 RB10378 144 435 Thioredoxin trxA 5610497 5610063 4.6 15274 - 6.65 0.21 1.51 0.03 1.966675 RB11981 317 954 ABC transporter (substrate-binding protein) 6459460 6460413 4.8 34461 + 6.03 0.28 1.58 0.14 9.081278 RB2386 528 1587 RND efflux membrane fusion protein 1244320 1242734 6 58921 - 6.19 0.26 1.74 0.12 6.765570 RB9998 277 834 ABC transporter ATP-binding protein 5385937 5385104 5.9 30617 - 6.72 0.32 1.50 0.30 19.957369 RB13257 286 861 2-hydroxy-6-ketonona-2,4-dienedioic acid hydrolase bphD 3.7.1.- 7105590 7106450 4.6 32540 + 6.08 0.21 1.53 0.09 6.002438 RB4395 170 513 carboxymuconolactone decarboxylase 4.1.1.44 2262023 2262535 4.7 18791 + 5.96 0.25 1.63 0.34 21.082488 RB4474 168 507 CsbD-like 2295048 2295554 8.8 18451 + 7.12 0.24 1.96 0.07 3.705413 RB9699 697 2094 Rhs element Vgr protein 5226252 5228345 4.2 76047 + 5.94 0.32 1.63 0.37 22.476266 RB11260 121 366 dnaK suppressor protein, 6107202 6106837 5.2 13639 - 5.95 0.18 1.82 0.06 3.37
10 RB17 164 495 protein containing DUF1589 8251 7757 11.5 18498 - 5.48 0.29 1.97 0.30 15.11358 RB655 346 1041 conserved hypothetical protein, UCP012641 family 354993 353953 5.2 38936 - 6.01 0.30 2.32 0.15 6.66470 RB865 542 1629 hypothetical protein 458731 460359 3.1 54518 + 5.77 0.28 1.90 0.18 9.68507 RB934 375 1128 Putative transposase 494203 493076 9.9 41881 - 5.57 0.27 1.84 0.03 1.58947 RB1789 243 732 conserved hypothetical protein 925861 926592 9.4 27028 + 5.83 0.28 1.66 0.23 13.88
1454 RB2682 83 252 hypothetical protein 1393781 1393530 7.4 8960 - 6.29 0.30 1.77 0.34 19.071490 RB2745 41 126 hypothetical protein 1431275 1431400 9.9 4387 + 5.88 0.19 1.57 0.32 20.191528 RB2829 38 117 hypothetical protein 1467448 1467564 4.1 3898 + 6.42 0.28 1.63 0.38 23.431744 RB3228 962 2889 protein containing DUF1585 1656635 1659523 4.6 107564 + 5.75 0.27 1.52 0.35 23.382351 RB4256 358 1077 conserved hypothetical protein, secreted 2180303 2181379 4.5 37091 + 6.62 0.30 1.89 0.24 12.952407 RB4347 156 471 conserved hypothetical protein 2235735 2236205 4.7 17459 + 6.97 0.30 2.61 0.23 8.752426 RB4376 493 1482 catalase katA 1.11.1.6 2253743 2252262 5.5 56567 - 5.89 0.24 1.69 0.30 17.742431 RB4383 545 1638 decarboxylase 2258768 2257131 5.4 59839 - 5.90 0.18 2.06 0.13 6.202461 RB4429 89 270 conserved hypothetical protein 2275996 2276265 4.9 10034 + 6.59 0.30 2.29 0.37 16.022465 RB4438 160 483 Pyridoxamine 5'-phosphate oxidase- 2278247 2278729 4.4 18043 + 5.63 0.27 1.94 0.22 11.082485 RB4468 162 489 hypothetical protein 2294104 2293616 4.6 16380 - 6.30 0.19 1.66 0.28 16.982489 RB4476 128 387 hypothetical protein 2295557 2295943 11.4 14037 + 6.94 0.23 1.68 0.26 15.762499 RB4493 83 252 hypothetical protein 2302374 2302123 10.2 8978 - 6.95 0.23 1.83 0.21 11.452503 RB4500 87 264 conserved hypothetical protein, secreted 2303645 2303908 3.5 9432 + 7.27 0.34 2.49 0.22 8.922509 RB4510 49 150 hypothetical protein 2307657 2307508 9 5554 - 5.82 0.15 1.60 0.09 5.772951 RB5238 73 222 hypothetical protein 2697775 2697996 10.6 7866 + 6.04 0.22 2.57 0.10 4.033118 RB5523 91 276 hypothetical protein 2860984 2860709 6.3 9939 - 5.87 0.24 2.52 0.19 7.623346 RB5938 370 1113 hypothetical protein 3113484 3112372 5.6 41837 - 6.27 0.23 3.07 0.35 11.473449 RB6131 102 309 hypothetical protein 3223482 3223790 12.1 12343 + 6.20 0.29 1.65 0.30 18.203647 RB6467 252 759 phosphoglycerate/bisphosphoglycerate mutase family protein 5.4.2.1 3418620 3419378 5.9 27957 + 6.12 0.29 1.92 0.44 22.944157 RB7389 375 1128 Putative transposase 3953590 3954717 9.9 41895 + 5.61 0.29 2.40 0.14 5.624288 RB7603 836 2511 outer membrane efflux protein 4091070 4088560 5.1 91203 - 5.90 0.32 1.53 0.25 16.494318 RB7651 44 135 hypothetical protein 4112517 4112383 6.7 4749 - 5.71 0.29 1.77 0.16 8.864465 RB7901 106 321 hypothetical protein 4221613 4221293 11.8 12088 - 6.02 0.28 2.41 0.17 6.984704 RB8365 90 273 membrane protein 4463903 4463631 11.7 9405 - 6.14 0.21 1.78 0.23 13.064727 RB8409 97 294 hypothetical protein 4495547 4495840 8.9 10905 + 5.46 0.26 1.54 0.30 19.205157 RB9230 107 324 hypothetical protein 4936673 4936996 9.9 11900 + 5.88 0.22 1.64 0.20 12.265218 RB9347 68 207 hypothetical protein 5008600 5008394 9.9 7515 - 6.12 0.23 1.67 0.28 16.625550 RB9955 452 1359 secreted protein containing DUF1552 5363605 5362247 5.6 50315 - 6.08 0.21 2.27 0.08 3.615565 RB9986 112 339 hypothetical protein 5380717 5380379 11.1 12671 - 5.85 0.28 1.75 0.15 8.405567 RB9990 53 162 hypothetical protein 5381822 5381983 10.1 6033 + 5.16 0.28 2.14 0.59 27.355618 RB10077 88 267 secreted protein 5436817 5437083 3.3 9650 + 6.38 0.28 2.17 0.38 17.626053 RB10877 453 1362 membrane protein 5905128 5903767 4.3 48882 - 5.49 0.30 1.55 0.10 6.466084 RB10931 54 165 hypothetical protein 5933742 5933906 4.2 5888 + 6.97 0.25 1.92 0.02 0.816085 RB10932 65 198 secreted protein 5933872 5934069 3.8 6983 + 6.96 0.27 2.15 0.17 7.786097 RB10954 143 432 hypothetical protein 5940749 5940318 10.4 16058 - 5.83 0.22 1.66 0.33 20.046098 RB10956 117 354 hypothetical protein 5940336 5940689 4.8 13013 + 6.42 0.32 3.24 0.38 11.686099 RB10957 99 300 conserved hypothetical protein 5940708 5941007 5.6 11604 + 7.07 0.20 2.05 0.21 10.376176 RB11107 225 678 secreted protein 6021425 6022102 6.5 25080 + 6.14 0.24 2.37 0.33 14.036406 RB11475 57 174 conserved hypothetical protein 6209468 6209641 4.7 5927 + 5.81 0.26 1.68 0.15 9.046430 RB11515 74 225 conserved hypothetical protein 6224420 6224644 11.7 8687 + 5.34 0.26 2.74 0.34 12.526455 RB11566 195 588 conserved hypothetical protein 6243111 6243698 10.8 22135 + 5.62 0.25 1.84 0.33 17.866585 RB11813 80 243 conserved hypothetical protein 6366331 6366573 13 9157 + 5.73 0.28 1.82 0.22 12.246606 RB11855 101 306 conserved hypothetical protein 6383765 6383460 12.5 11384 - 5.97 0.23 2.42 0.10 4.226637 RB11918 134 405 protein containing DUF971 6415701 6415297 6.1 14675 - 5.89 0.32 2.14 0.11 5.026719 RB12067 107 324 conserved hypothetical protein 6510236 6510559 11.9 12169 + 5.87 0.35 2.73 0.28 10.347128 RB12800 156 471 conserved hypothetical protein 6908240 6908710 9.5 16259 + 5.79 0.20 1.92 0.41 21.197149 RB12830 102 309 conserved hypothetical protein 6920508 6920200 10.9 11753 - 5.86 0.22 2.11 0.42 19.847278 RB13093 158 477 secreted protein 7025577 7025101 9.6 17298 - 5.88 0.25 1.59 0.12 7.337351 RB13222 208 627 SOUL heme-binding protein 7094714 7094088 8.7 22867 - 5.81 0.19 3.86 0.31 7.967354 RB13228 116 351 conserved hypothetical protein 7097952 7098302 10.4 13570 + 5.95 0.21 1.85 0.13 7.09
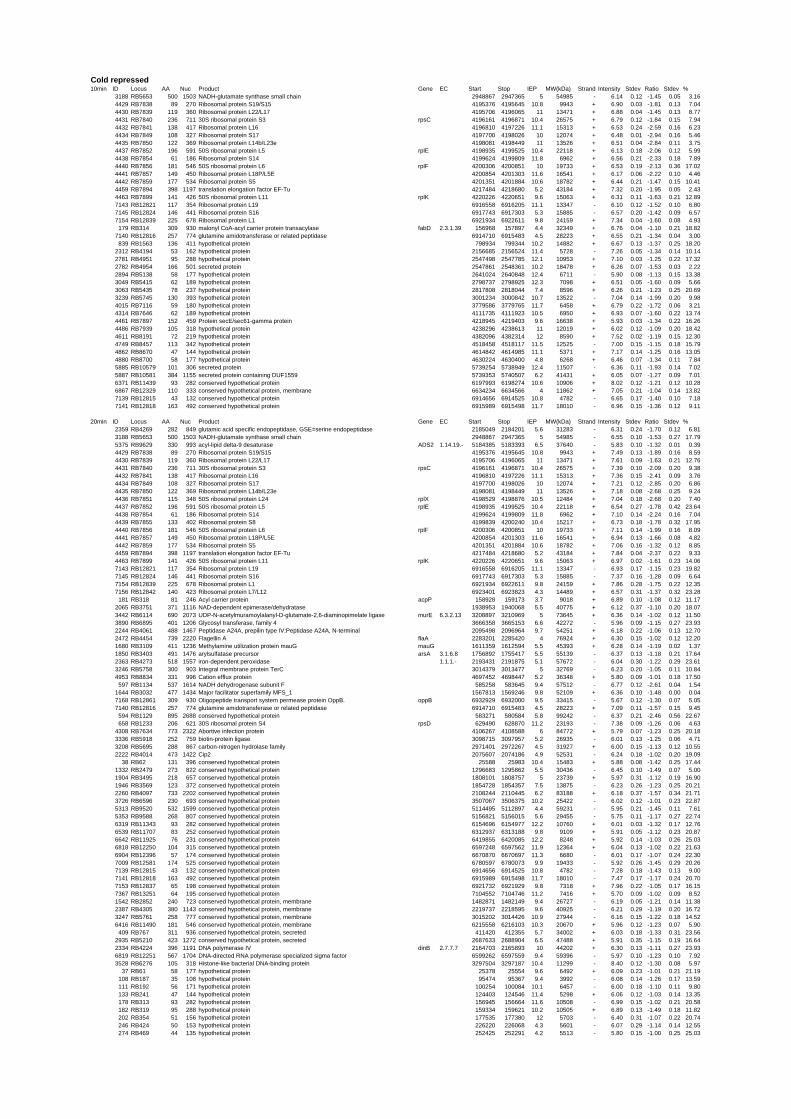
Cold repressed10min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand Intensity Stdev Ratio Stdev %
3188 RB5653 500 1503 NADH-glutamate synthase small chain 2948867 2947365 5 54985 - 6.14 0.12 -1.45 0.05 3.164429 RB7838 89 270 Ribosomal protein S19/S15 4195376 4195645 10.8 9943 + 6.90 0.03 -1.81 0.13 7.044430 RB7839 119 360 Ribosomal protein L22/L17 4195706 4196065 11 13471 + 6.88 0.04 -1.45 0.13 8.774431 RB7840 236 711 30S ribosomal protein S3 rpsC 4196161 4196871 10.4 26575 + 6.79 0.12 -1.84 0.15 7.944432 RB7841 138 417 Ribosomal protein L16 4196810 4197226 11.1 15313 + 6.53 0.24 -2.59 0.16 6.234434 RB7849 108 327 Ribosomal protein S17 4197700 4198026 10 12074 + 6.48 0.01 -2.94 0.16 5.464435 RB7850 122 369 Ribosomal protein L14b/L23e 4198081 4198449 11 13526 + 6.51 0.04 -2.84 0.11 3.754437 RB7852 196 591 50S ribosomal protein L5 rplE 4198935 4199525 10.4 22118 + 6.13 0.18 -2.06 0.12 5.994438 RB7854 61 186 Ribosomal protein S14 4199624 4199809 11.8 6962 + 6.56 0.21 -2.33 0.18 7.894440 RB7856 181 546 50S ribosomal protein L6 rplF 4200306 4200851 10 19733 + 6.53 0.19 -2.13 0.36 17.024441 RB7857 149 450 Ribosomal protein L18P/L5E 4200854 4201303 11.6 16541 + 6.17 0.06 -2.22 0.10 4.464442 RB7859 177 534 Ribosomal protein S5 4201351 4201884 10.6 18782 + 6.44 0.21 -1.47 0.15 10.414459 RB7894 398 1197 translation elongation factor EF-Tu 4217484 4218680 5.2 43184 + 7.32 0.20 -1.95 0.05 2.434463 RB7899 141 426 50S ribosomal protein L11 rplK 4220226 4220651 9.6 15063 + 6.31 0.11 -1.63 0.21 12.897143 RB12821 117 354 Ribosomal protein L19 6916558 6916205 11.1 13347 - 6.10 0.12 -1.52 0.10 6.807145 RB12824 146 441 Ribosomal protein S16 6917743 6917303 5.3 15885 - 6.57 0.20 -1.42 0.09 6.577154 RB12839 225 678 Ribosomal protein L1 6921934 6922611 9.8 24159 + 7.34 0.04 -1.60 0.08 4.93
179 RB314 309 930 malonyl CoA-acyl carrier protein transacylase fabD 2.3.1.39 156968 157897 4.4 32349 + 6.76 0.04 -1.10 0.21 18.827140 RB12816 257 774 glutamine amidotransferase or related peptidase 6914710 6915483 4.5 28223 + 6.55 0.21 -1.34 0.04 3.00
839 RB1563 136 411 hypothetical protein 798934 799344 10.2 14882 + 6.67 0.13 -1.37 0.25 18.202312 RB4194 53 162 hypothetical protein 2156685 2156524 11.4 5728 - 7.26 0.05 -1.34 0.14 10.142781 RB4951 95 288 hypothetical protein 2547498 2547785 12.1 10953 + 7.10 0.03 -1.25 0.22 17.322782 RB4954 166 501 secreted protein 2547861 2548361 10.2 18478 + 6.26 0.07 -1.53 0.03 2.222894 RB5138 58 177 hypothetical protein 2641024 2640848 12.4 6711 - 5.90 0.08 -1.13 0.15 13.383049 RB5415 62 189 hypothetical protein 2798737 2798925 12.3 7098 + 6.51 0.05 -1.60 0.09 5.663063 RB5435 78 237 hypothetical protein 2817808 2818044 7.4 8596 + 6.26 0.21 -1.23 0.25 20.693239 RB5745 130 393 hypothetical protein 3001234 3000842 10.7 13522 - 7.04 0.14 -1.99 0.20 9.984015 RB7116 59 180 hypothetical protein 3779586 3779765 11.7 6458 + 6.79 0.22 -1.72 0.06 3.214314 RB7646 62 189 hypothetical protein 4111735 4111923 10.5 6950 + 6.93 0.07 -1.60 0.22 13.744461 RB7897 152 459 Protein secE/sec61-gamma protein 4218945 4219403 9.6 16638 + 5.93 0.03 -1.34 0.22 16.264486 RB7939 105 318 hypothetical protein 4238296 4238613 11 12019 + 6.02 0.12 -1.09 0.20 18.424611 RB8191 72 219 hypothetical protein 4382096 4382314 12 8590 + 7.52 0.02 -1.19 0.15 12.304749 RB8457 113 342 hypothetical protein 4518458 4518117 11.5 12525 - 7.00 0.15 -1.15 0.18 15.794862 RB8670 47 144 hypothetical protein 4614842 4614985 11.1 5371 + 7.17 0.14 -1.25 0.16 13.054880 RB8700 58 177 hypothetical protein 4630224 4630400 4.8 6268 + 6.46 0.07 -1.34 0.11 7.845885 RB10579 101 306 secreted protein 5739254 5738949 12.4 11507 - 6.36 0.11 -1.93 0.14 7.025887 RB10581 384 1155 secreted protein containing DUF1559 5739353 5740507 6.2 41431 + 6.05 0.07 -1.27 0.09 7.016371 RB11439 93 282 conserved hypothetical protein 6197993 6198274 10.6 10906 + 8.02 0.12 -1.21 0.12 10.286867 RB12329 110 333 conserved hypothetical protein, membrane 6634234 6634566 4 11862 + 7.05 0.21 -1.04 0.14 13.827139 RB12815 43 132 conserved hypothetical protein 6914656 6914525 10.8 4782 - 6.65 0.17 -1.40 0.10 7.187141 RB12818 163 492 conserved hypothetical protein 6915989 6915498 11.7 18010 - 6.96 0.15 -1.36 0.12 9.11
20min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand Intensity Stdev Ratio Stdev %2359 RB4269 282 849 glutamic acid specific endopeptidase, GSE=serine endopeptidase 2185049 2184201 5.6 31283 - 6.31 0.24 -1.70 0.12 6.813188 RB5653 500 1503 NADH-glutamate synthase small chain 2948867 2947365 5 54985 - 6.55 0.10 -1.53 0.27 17.795375 RB9629 330 993 acyl-lipid delta-9 desaturase ADS2 1.14.19.- 5184385 5183393 6.5 37640 - 5.83 0.10 -1.32 0.01 0.394429 RB7838 89 270 Ribosomal protein S19/S15 4195376 4195645 10.8 9943 + 7.49 0.13 -1.89 0.16 8.594430 RB7839 119 360 Ribosomal protein L22/L17 4195706 4196065 11 13471 + 7.61 0.09 -1.63 0.21 12.764431 RB7840 236 711 30S ribosomal protein S3 rpsC 4196161 4196871 10.4 26575 + 7.39 0.10 -2.09 0.20 9.384432 RB7841 138 417 Ribosomal protein L16 4196810 4197226 11.1 15313 + 7.36 0.15 -2.41 0.09 3.764434 RB7849 108 327 Ribosomal protein S17 4197700 4198026 10 12074 + 7.21 0.12 -2.85 0.20 6.864435 RB7850 122 369 Ribosomal protein L14b/L23e 4198081 4198449 11 13526 + 7.18 0.08 -2.68 0.25 9.244436 RB7851 115 348 50S ribosomal protein L24 rplX 4198529 4198876 10.5 12484 + 7.04 0.18 -2.68 0.20 7.404437 RB7852 196 591 50S ribosomal protein L5 rplE 4198935 4199525 10.4 22118 + 6.54 0.27 -1.78 0.42 23.644438 RB7854 61 186 Ribosomal protein S14 4199624 4199809 11.8 6962 + 7.10 0.14 -2.24 0.16 7.044439 RB7855 133 402 Ribosomal protein S8 4199839 4200240 10.4 15217 + 6.73 0.18 -1.78 0.32 17.954440 RB7856 181 546 50S ribosomal protein L6 rplF 4200306 4200851 10 19733 + 7.11 0.14 -1.99 0.16 8.094441 RB7857 149 450 Ribosomal protein L18P/L5E 4200854 4201303 11.6 16541 + 6.94 0.13 -1.66 0.08 4.824442 RB7859 177 534 Ribosomal protein S5 4201351 4201884 10.6 18782 + 7.06 0.16 -1.32 0.12 8.854459 RB7894 398 1197 translation elongation factor EF-Tu 4217484 4218680 5.2 43184 + 7.84 0.04 -2.37 0.22 9.334463 RB7899 141 426 50S ribosomal protein L11 rplK 4220226 4220651 9.6 15063 + 6.97 0.02 -1.61 0.23 14.067143 RB12821 117 354 Ribosomal protein L19 6916558 6916205 11.1 13347 - 6.93 0.17 -1.15 0.23 19.827145 RB12824 146 441 Ribosomal protein S16 6917743 6917303 5.3 15885 - 7.37 0.16 -1.28 0.09 6.647154 RB12839 225 678 Ribosomal protein L1 6921934 6922611 9.8 24159 + 7.86 0.28 -1.75 0.22 12.357156 RB12842 140 423 Ribosomal protein L7/L12 6923401 6923823 4.3 14489 + 6.57 0.31 -1.37 0.32 23.28
181 RB318 81 246 Acyl carrier protein acpP 158928 159173 3.7 9018 + 6.89 0.10 -1.08 0.12 11.172065 RB3751 371 1116 NAD-dependent epimerase/dehydratase 1938953 1940068 5.5 40775 + 6.12 0.37 -1.10 0.20 18.073442 RB6114 690 2073 UDP-N-acetylmuramoylalanyl-D-glutamate-2,6-diaminopimelate ligase murE 6.3.2.13 3208897 3210969 5 73645 + 6.36 0.14 -1.02 0.12 11.503890 RB6895 401 1206 Glycosyl transferase, family 4 3666358 3665153 6.6 42272 - 5.96 0.09 -1.15 0.27 23.932244 RB4061 488 1467 Peptidase A24A, prepilin type IV:Peptidase A24A, N-terminal 2095498 2096964 9.7 54251 + 6.18 0.22 -1.06 0.13 12.702472 RB4454 739 2220 Flagellin A flaA 2283201 2285420 4 76924 + 6.30 0.15 -1.02 0.12 12.201680 RB3109 411 1236 Methylamine utilization protein mauG mauG 1611359 1612594 5.5 45393 + 6.28 0.14 -1.19 0.02 1.371850 RB3403 491 1476 arylsulfatase precursor arsA 3.1.6.8 1756892 1755417 5.5 55139 - 6.37 0.13 -1.18 0.21 17.642363 RB4273 518 1557 iron-dependent peroxidase 1.1.1.- 2193431 2191875 5.1 57672 - 6.04 0.30 -1.22 0.29 23.613246 RB5758 300 903 Integral membrane protein TerC 3014379 3013477 5 32769 - 6.23 0.20 -1.05 0.11 10.844953 RB8834 331 996 Cation efflux protein 4697452 4698447 5.2 36348 + 5.80 0.09 -1.01 0.18 17.50
597 RB1134 537 1614 NADH dehydrogenase subunit F 585258 583645 9.4 57512 - 6.77 0.12 -2.61 0.04 1.541644 RB3032 477 1434 Major facilitator superfamily MFS_1 1567813 1569246 9.8 52109 + 6.36 0.10 -1.48 0.00 0.047168 RB12861 309 930 Oligopeptide transport system permease protein OppB. oppB 6932929 6932000 9.5 33415 - 5.67 0.12 -1.30 0.07 5.057140 RB12816 257 774 glutamine amidotransferase or related peptidase 6914710 6915483 4.5 28223 + 7.09 0.11 -1.57 0.15 9.45
594 RB1129 895 2688 conserved hypothetical protein 583271 580584 5.8 99242 - 6.37 0.21 -2.46 0.56 22.67658 RB1233 206 621 30S ribosomal protein S4 rpsD 629490 628870 11.2 23193 - 7.38 0.09 -1.26 0.06 4.63
4308 RB7634 773 2322 Abortive infection protein 4106267 4108588 6 84772 + 5.79 0.07 -1.23 0.25 20.183336 RB5918 252 759 biotin-protein ligase 3098715 3097957 5.2 26935 - 6.01 0.13 -1.25 0.06 4.713208 RB5695 288 867 carbon-nitrogen hydrolase family 2971401 2972267 4.5 31927 + 6.00 0.15 -1.13 0.12 10.552222 RB4014 473 1422 Cip2 2075607 2074186 4.9 52531 - 6.24 0.18 -1.02 0.20 19.09
38 RB62 131 396 conserved hypothetical protein 25588 25983 10.4 15483 + 5.88 0.08 -1.42 0.25 17.441332 RB2479 273 822 conserved hypothetical protein 1296683 1295862 5.5 30436 - 6.45 0.10 -1.49 0.07 5.001904 RB3495 218 657 conserved hypothetical protein 1808101 1808757 5 23739 + 5.97 0.31 -1.12 0.19 16.901946 RB3569 123 372 conserved hypothetical protein 1854728 1854357 7.5 13875 - 6.23 0.26 -1.23 0.25 20.212260 RB4097 733 2202 conserved hypothetical protein 2108244 2110445 6.2 83188 + 6.18 0.37 -1.57 0.34 21.713726 RB6596 230 693 conserved hypothetical protein 3507067 3506375 10.2 25422 - 6.02 0.12 -1.01 0.23 22.875313 RB9520 532 1599 conserved hypothetical protein 5114495 5112897 4.4 59231 - 5.95 0.21 -1.45 0.11 7.615353 RB9588 268 807 conserved hypothetical protein 5156821 5156015 5.6 29455 - 5.75 0.11 -1.17 0.27 22.746319 RB11343 93 282 conserved hypothetical protein 6154696 6154977 12.2 10760 + 6.01 0.03 -1.32 0.17 12.766539 RB11707 83 252 conserved hypothetical protein 6312937 6313188 9.8 9109 + 5.91 0.05 -1.12 0.23 20.876642 RB11925 76 231 conserved hypothetical protein 6419855 6420085 12.2 8248 + 5.92 0.14 -1.03 0.26 25.036818 RB12250 104 315 conserved hypothetical protein 6597248 6597562 11.9 12364 + 6.04 0.13 -1.02 0.22 21.636904 RB12396 57 174 conserved hypothetical protein 6670870 6670697 11.3 6680 - 6.01 0.17 -1.07 0.24 22.307009 RB12581 174 525 conserved hypothetical protein 6780597 6780073 9.9 19433 - 5.92 0.26 -1.45 0.29 20.267139 RB12815 43 132 conserved hypothetical protein 6914656 6914525 10.8 4782 - 7.28 0.18 -1.43 0.13 9.007141 RB12818 163 492 conserved hypothetical protein 6915989 6915498 11.7 18010 - 7.47 0.17 -1.17 0.24 20.707153 RB12837 65 198 conserved hypothetical protein 6921732 6921929 9.8 7318 + 7.96 0.22 -1.05 0.17 16.157367 RB13251 64 195 conserved hypothetical protein 7104552 7104746 11.2 7416 + 5.70 0.09 -1.02 0.09 8.521542 RB2852 240 723 conserved hypothetical protein, membrane 1482871 1482149 9.4 26727 - 6.19 0.05 -1.21 0.14 11.382387 RB4305 380 1143 conserved hypothetical protein, membrane 2219737 2218595 9.6 40925 - 6.21 0.29 -1.19 0.20 16.723247 RB5761 258 777 conserved hypothetical protein, membrane 3015202 3014426 10.9 27944 - 6.16 0.15 -1.22 0.18 14.526416 RB11490 181 546 conserved hypothetical protein, membrane 6215558 6216103 10.3 20670 + 5.96 0.12 -1.23 0.07 5.90
409 RB767 311 936 conserved hypothetical protein, secreted 411420 412355 5.7 34002 + 6.03 0.18 -1.33 0.31 23.562935 RB5210 423 1272 conserved hypothetical protein, secreted 2687633 2688904 6.5 47488 + 5.91 0.35 -1.15 0.19 16.642334 RB4224 396 1191 DNA polymerase IV dinB 2.7.7.7 2164703 2165893 10 44202 + 6.30 0.13 -1.11 0.27 23.936819 RB12251 567 1704 DNA-directed RNA polymerase specialized sigma factor 6599262 6597559 9.4 59396 - 5.97 0.10 -1.23 0.10 7.923528 RB6276 105 318 Histone-like bacterial DNA-binding protein 3297504 3297187 10.4 11299 - 8.40 0.12 -1.30 0.08 5.97
37 RB61 58 177 hypothetical protein 25378 25554 9.6 6492 + 6.09 0.23 -1.01 0.21 21.19108 RB187 35 108 hypothetical protein 95474 95367 9.4 3992 - 6.08 0.14 -1.26 0.17 13.59111 RB192 56 171 hypothetical protein 100254 100084 10.1 6457 - 6.00 0.18 -1.10 0.11 9.80133 RB241 47 144 hypothetical protein 124403 124546 11.4 5298 + 6.06 0.12 -1.03 0.14 13.35178 RB313 93 282 hypothetical protein 156945 156664 11.6 10508 - 6.99 0.15 -1.02 0.21 20.58182 RB319 95 288 hypothetical protein 159334 159621 10.2 10505 + 6.89 0.13 -1.49 0.18 11.82202 RB354 51 156 hypothetical protein 177535 177380 12 5703 - 6.40 0.31 -1.07 0.22 20.74246 RB424 50 153 hypothetical protein 226220 226068 4.3 5601 - 6.07 0.29 -1.14 0.14 12.55274 RB469 44 135 hypothetical protein 252425 252291 4.2 5513 - 5.80 0.15 -1.00 0.25 25.03

276 RB471 181 546 hypothetical protein 253264 252719 10.8 20224 - 5.98 0.13 -1.25 0.05 4.08283 RB484 58 177 hypothetical protein 262633 262809 10.6 6245 + 5.78 0.09 -1.21 0.19 15.87348 RB625 68 207 hypothetical protein 339703 339497 5 7713 - 5.64 0.04 -1.09 0.28 25.80416 RB779 86 261 hypothetical protein 419045 418785 4.5 9321 - 5.78 0.05 -1.09 0.00 0.39449 RB825 117 354 hypothetical protein 434184 433831 7.8 13302 - 6.30 0.23 -1.05 0.11 10.84691 RB1291 97 294 hypothetical protein 656684 656977 10.4 10290 + 5.98 0.16 -1.38 0.14 10.21839 RB1563 136 411 hypothetical protein 798934 799344 10.2 14882 + 7.47 0.05 -1.09 0.07 6.19
1012 RB1922 102 309 hypothetical protein 998917 998609 10.9 11658 - 6.23 0.06 -1.04 0.25 24.351016 RB1931 68 207 hypothetical protein 1013831 1013625 11.4 7493 - 6.10 0.37 -1.33 0.10 7.311200 RB2274 130 393 hypothetical protein 1193331 1192939 10 14362 - 6.27 0.13 -1.18 0.08 6.831220 RB2306 41 126 hypothetical protein 1206782 1206907 9 4759 + 6.35 0.23 -1.36 0.05 3.431227 RB2317 162 489 hypothetical protein 1211131 1210643 7.7 18020 - 6.06 0.14 -1.00 0.11 10.841251 RB2350 37 114 hypothetical protein 1220890 1221003 11.9 4181 + 6.08 0.21 -1.18 0.09 7.531252 RB2351 133 402 hypothetical protein 1220991 1221392 10.4 15178 + 6.04 0.28 -1.12 0.28 24.951297 RB2425 102 309 hypothetical protein 1260766 1261074 11.1 11189 + 6.26 0.09 -1.06 0.14 13.591344 RB2495 43 132 hypothetical protein 1303912 1304043 11.7 4921 + 6.26 0.27 -1.03 0.19 18.801543 RB2853 42 129 hypothetical protein 1482846 1482974 11.8 5043 + 6.12 0.15 -1.09 0.16 14.701734 RB3205 76 231 hypothetical protein 1652079 1651849 11.3 9082 - 5.91 0.29 -1.00 0.12 11.711761 RB3256 140 423 hypothetical protein 1676574 1676996 4.5 16074 + 6.45 0.14 -1.29 0.24 18.251776 RB3277 221 666 hypothetical protein 1686552 1685887 10.4 24222 - 6.21 0.17 -1.23 0.10 8.231825 RB3362 87 264 hypothetical protein 1728546 1728283 11.9 10017 - 6.44 0.33 -1.29 0.23 17.841828 RB3366 78 237 hypothetical protein 1730380 1730616 5.6 9481 + 6.05 0.33 -1.11 0.07 6.751833 RB3372 115 348 hypothetical protein 1733666 1734013 10 12916 + 6.09 0.22 -1.17 0.06 5.181844 RB3394 36 111 hypothetical protein 1752044 1752154 10.5 4239 + 6.37 0.21 -1.94 0.04 2.071846 RB3398 67 204 hypothetical protein 1753293 1753496 11.4 7537 + 5.97 0.14 -1.08 0.13 12.461847 RB3399 65 198 hypothetical protein 1753650 1753453 9.7 7418 - 6.27 0.19 -1.19 0.15 12.441854 RB3407 89 270 hypothetical protein 1759935 1759666 10.5 10136 - 6.24 0.33 -1.34 0.29 21.531943 RB3566 64 195 hypothetical protein 1854080 1854274 9.4 7400 + 6.13 0.18 -1.24 0.13 10.172006 RB3660 94 285 hypothetical protein 1900854 1900570 9.4 10411 - 6.51 0.47 -1.23 0.29 23.382025 RB3688 53 162 hypothetical protein 1910629 1910468 9.3 5981 - 6.24 0.28 -1.21 0.10 8.422048 RB3719 141 426 hypothetical protein 1927500 1927075 10.8 16330 - 6.49 0.20 -1.25 0.21 16.692098 RB3813 75 228 hypothetical protein 1975188 1975415 4.5 8522 + 6.18 0.32 -1.21 0.26 21.522139 RB3880 82 249 hypothetical protein 2006953 2006705 10.7 9535 - 6.13 0.11 -1.13 0.06 5.022184 RB3953 857 2574 hypothetical protein 2048784 2046211 5.2 93881 - 6.29 0.26 -1.40 0.36 25.652200 RB3981 161 486 hypothetical protein 2058000 2058485 4.1 18283 + 6.29 0.17 -1.17 0.11 9.032216 RB4002 483 1452 hypothetical protein 2065341 2066792 4.6 53759 + 6.28 0.18 -1.01 0.23 23.322258 RB4093 45 138 hypothetical protein 2107600 2107737 10 5285 + 6.21 0.15 -1.04 0.08 7.242287 RB4145 90 273 hypothetical protein 2131933 2131661 12 10727 - 6.91 0.26 -1.07 0.19 17.632298 RB4165 721 2166 hypothetical protein 2139632 2141797 4.5 77994 + 6.19 0.22 -1.07 0.21 20.092312 RB4194 53 162 hypothetical protein 2156685 2156524 11.4 5728 - 8.07 0.12 -1.09 0.03 2.672350 RB4255 204 615 hypothetical protein 2180806 2180192 10.3 21582 - 9.54 0.03 -1.63 0.20 12.542416 RB4358 123 372 hypothetical protein 2239474 2239103 6.5 13459 - 6.33 0.30 -1.49 0.27 17.982424 RB4373 109 330 hypothetical protein 2242863 2242534 4.8 12263 - 6.41 0.14 -1.15 0.10 8.362444 RB4402 53 162 hypothetical protein 2265340 2265501 10.4 5980 + 6.17 0.27 -1.36 0.26 19.152528 RB4547 422 1269 hypothetical protein 2330116 2328848 7.6 46274 - 6.07 0.21 -1.20 0.29 23.862552 RB4589 62 189 hypothetical protein 2351649 2351461 9.4 7014 - 5.90 0.12 -1.17 0.14 12.252553 RB4590 74 225 hypothetical protein 2352024 2351800 10.1 8330 - 9.62 0.01 -1.89 0.26 13.472596 RB4657 123 372 hypothetical protein 2381865 2382236 12.2 13398 + 6.50 0.24 -1.69 0.17 10.052647 RB4740 50 153 hypothetical protein 2431814 2431966 11.4 5598 + 6.35 0.24 -1.03 0.04 4.022687 RB4801 96 291 hypothetical protein 2459502 2459212 9.8 10561 - 6.11 0.34 -1.00 0.05 4.692746 RB4897 184 555 hypothetical protein 2519505 2518951 10.9 20446 - 6.03 0.32 -1.33 0.19 14.602781 RB4951 95 288 hypothetical protein 2547498 2547785 12.1 10953 + 7.46 0.34 -1.52 0.24 16.102864 RB5088 57 174 hypothetical protein 2617615 2617442 6.7 6093 - 5.91 0.24 -1.04 0.15 14.512963 RB5261 126 381 hypothetical protein 2713712 2713332 7.4 13761 - 6.00 0.28 -1.00 0.23 23.413049 RB5415 62 189 hypothetical protein 2798737 2798925 12.3 7098 + 6.88 0.37 -1.87 0.32 17.333063 RB5435 78 237 hypothetical protein 2817808 2818044 7.4 8596 + 6.96 0.20 -1.10 0.05 4.923132 RB5555 54 165 hypothetical protein 2891274 2891438 6.2 5971 + 6.30 0.34 -1.16 0.22 18.563239 RB5745 130 393 hypothetical protein 3001234 3000842 10.7 13522 - 7.72 0.11 -2.40 0.06 2.623389 RB6010 53 162 hypothetical protein 3149911 3150072 11.9 5744 + 6.04 0.27 -1.25 0.11 8.983466 RB6158 142 429 hypothetical protein 3233532 3233104 6 15746 - 6.57 0.19 -1.29 0.10 7.703476 RB6174 69 210 hypothetical protein 3239734 3239525 10.6 7870 - 6.11 0.12 -1.06 0.21 19.643485 RB6193 62 189 hypothetical protein 3246096 3245908 11.4 6970 - 6.81 0.25 -1.45 0.09 5.963720 RB6587 50 153 hypothetical protein 3501747 3501595 7.5 5579 - 6.06 0.12 -1.08 0.10 8.843742 RB6632 55 168 hypothetical protein 3524963 3525130 5.6 6193 + 5.97 0.24 -1.11 0.03 2.613780 RB6699 47 144 hypothetical protein 3559781 3559924 11.1 5290 + 5.99 0.22 -1.09 0.16 14.693820 RB6766 55 168 hypothetical protein 3597367 3597534 12 6133 + 6.21 0.23 -1.23 0.29 23.683860 RB6838 55 168 hypothetical protein 3636563 3636730 4.8 5823 + 5.76 0.19 -1.16 0.18 15.703867 RB6849 101 306 hypothetical protein 3642537 3642232 12.8 12002 - 6.25 0.37 -1.17 0.08 6.773871 RB6856 62 189 hypothetical protein 3645613 3645425 9.9 6382 - 5.75 0.12 -1.19 0.15 12.193921 RB6951 1120 3363 hypothetical protein 3693377 3690015 4.6 123187 - 6.22 0.14 -1.03 0.26 25.583971 RB7042 91 276 hypothetical protein 3743971 3744246 10.4 10835 + 6.19 0.11 -1.22 0.08 6.264009 RB7109 53 162 hypothetical protein 3775707 3775546 6.7 5850 - 5.86 0.15 -1.54 0.21 13.934015 RB7116 59 180 hypothetical protein 3779586 3779765 11.7 6458 + 7.48 0.04 -1.60 0.16 10.274278 RB7589 86 261 hypothetical protein 4080238 4079978 12.1 9472 - 5.85 0.13 -1.00 0.18 18.084313 RB7645 77 234 hypothetical protein 4111716 4111483 10.3 8725 - 5.73 0.14 -1.45 0.25 17.054314 RB7646 62 189 hypothetical protein 4111735 4111923 10.5 6950 + 7.38 0.33 -1.81 0.19 10.354552 RB8070 59 180 hypothetical protein 4319538 4319717 4.5 6478 + 5.84 0.21 -1.27 0.30 23.314568 RB8098 71 216 hypothetical protein 4337334 4337119 10.8 7950 - 9.57 0.02 -1.70 0.24 14.334578 RB8119 142 429 hypothetical protein 4346427 4346855 10.1 15659 + 5.95 0.11 -1.21 0.24 20.074677 RB8319 37 114 hypothetical protein 4442421 4442308 4.4 4328 - 5.79 0.08 -1.58 0.20 12.944755 RB8468 88 267 hypothetical protein 4520799 4520533 11.9 9719 - 6.80 0.13 -1.00 0.09 8.754830 RB8594 41 126 hypothetical protein 4581645 4581770 9.2 4509 + 6.31 0.04 -1.23 0.14 11.584861 RB8669 37 114 hypothetical protein 4614557 4614670 11.5 4395 + 9.53 0.16 -2.04 0.37 18.165245 RB9392 175 528 hypothetical protein 5038107 5037580 4.9 19499 - 5.92 0.16 -1.16 0.07 5.935253 RB9410 138 417 hypothetical protein 5048582 5048166 10 15146 - 5.78 0.13 -1.01 0.25 25.135280 RB9460 79 240 hypothetical protein 5078860 5078621 10.3 8991 - 9.45 0.13 -1.41 0.33 23.245445 RB9767 97 294 hypothetical protein 5264348 5264641 8.9 10547 + 5.89 0.07 -1.14 0.18 15.875988 RB10755 41 126 hypothetical protein 5839576 5839451 12.7 5083 - 5.93 0.17 -1.05 0.08 7.516280 RB11287 75 228 hypothetical protein 6118405 6118632 9.1 8478 + 6.04 0.07 -1.17 0.11 9.692047 RB3718 535 1608 Major facilitator superfamily MFS_1 1927114 1925507 4.6 57372 - 6.04 0.20 -1.16 0.12 10.66
22 RB37 160 483 membrane protein 18328 17846 5.6 17283 - 6.16 0.41 -1.28 0.16 12.7743 RB73 98 297 membrane protein 28727 29023 10.1 11150 + 5.79 0.09 -1.16 0.05 3.9770 RB125 297 894 membrane protein 54571 53678 6 32981 - 5.82 0.11 -1.12 0.08 7.05
547 RB1015 222 669 membrane protein 531723 532391 5.9 24319 + 5.88 0.08 -1.09 0.13 11.531105 RB2105 470 1413 membrane protein 1114012 1112600 9.6 51853 - 6.47 0.07 -1.76 0.22 12.291147 RB2179 199 600 membrane protein 1149988 1150587 10.5 22376 + 6.14 0.25 -1.03 0.22 21.091446 RB2667 236 711 membrane protein 1387198 1386488 10.7 25698 - 6.30 0.23 -1.08 0.16 15.031597 RB2951 174 525 membrane protein 1523616 1523092 9.3 20061 - 6.13 0.40 -1.38 0.20 14.801668 RB3067 153 462 membrane protein 1582850 1582389 7.6 17200 - 6.06 0.19 -1.05 0.22 20.901701 RB3147 168 507 membrane protein 1624840 1624334 8.7 18759 - 6.31 0.18 -1.05 0.14 13.511950 RB3575 152 459 membrane protein 1858779 1859237 10.1 17195 + 6.22 0.21 -1.33 0.05 3.642664 RB4766 130 393 membrane protein 2442991 2442599 9.7 14452 - 6.05 0.21 -1.13 0.08 6.782884 RB5118 140 423 membrane protein 2630572 2630150 9.1 16117 - 6.08 0.21 -1.24 0.28 22.542964 RB5262 95 288 membrane protein 2713742 2714029 6.3 10993 + 6.16 0.21 -1.48 0.30 20.543169 RB5622 175 528 membrane protein 2930538 2930011 4.3 19919 - 5.94 0.23 -1.13 0.07 6.444282 RB7596 175 528 membrane protein 4084008 4084535 7.5 19347 + 5.76 0.07 -1.01 0.22 21.894718 RB8389 939 2820 membrane protein 4484734 4481915 10.1 105627 - 5.93 0.14 -1.07 0.08 7.335469 RB9806 165 498 membrane protein 5280590 5281087 10.2 18166 + 5.74 0.01 -1.03 0.18 17.852083 RB3779 1030 3093 membrane-bound sensor and histidine kinase response regulator hybrid 1957809 1954717 4.8 114806 - 6.10 0.25 -1.14 0.16 13.931352 RB2503 515 1548 O-antigen flippase wzx 1311056 1309509 10 55740 - 6.13 0.32 -1.17 0.12 10.302774 RB4937 369 1110 oxidoreductase 2540086 2541195 5 39670 + 6.14 0.31 -1.06 0.09 8.683278 RB5820 355 1068 oxidoreductase 3050638 3051705 4.9 39347 + 5.97 0.27 -1.07 0.27 25.633430 RB6092 361 1086 Peptidase M50 3.4.24.- 3192387 3193472 9.6 39370 + 6.19 0.25 -1.28 0.30 23.116979 RB12521 726 2181 Peptidase M50 3.4.24.- 6744279 6746459 10.7 80320 + 5.80 0.27 -1.15 0.11 9.373183 RB5645 556 1671 periplasmic serine protease 2944822 2943152 5.2 60661 - 5.73 0.18 -1.13 0.10 8.953092 RB5478 131 396 pilus protein 2834459 2834064 4.2 14264 - 5.75 0.16 -1.01 0.06 6.24
562 RB1052 391 1176 protein containing DUF1559 548353 549528 5.5 42194 + 5.71 0.09 -1.05 0.19 18.493744 RB6634 365 1098 protein containing DUF1559 3525785 3526882 5.3 40217 + 6.11 0.12 -1.17 0.14 11.891659 RB3056 90 273 protein containing DUF1560 1579951 1579679 10.3 10144 - 6.19 0.24 -1.05 0.12 11.62
377 RB693 260 783 protein containing DUF1568 371190 370408 6.8 30735 - 5.91 0.16 -1.23 0.28 22.47516 RB951 234 705 protein containing DUF1596 502459 501755 12.3 25583 - 6.67 0.17 -1.51 0.23 15.00
3688 RB6530 237 714 protein containing DUF480 3468520 3467807 5.4 25750 - 5.96 0.12 -1.01 0.17 16.624461 RB7897 152 459 Protein secE/sec61-gamma protein 4218945 4219403 9.6 16638 + 6.69 0.13 -1.06 0.18 16.593151 RB5593 1290 3873 protein serine/threonine kinase 2.7.1.37 2913054 2909182 4.8 144451 - 5.92 0.14 -1.09 0.05 4.662369 RB4283 499 1500 PvdD/PvdJ 2201462 2199963 5.7 56411 - 6.28 0.25 -1.35 0.25 18.22
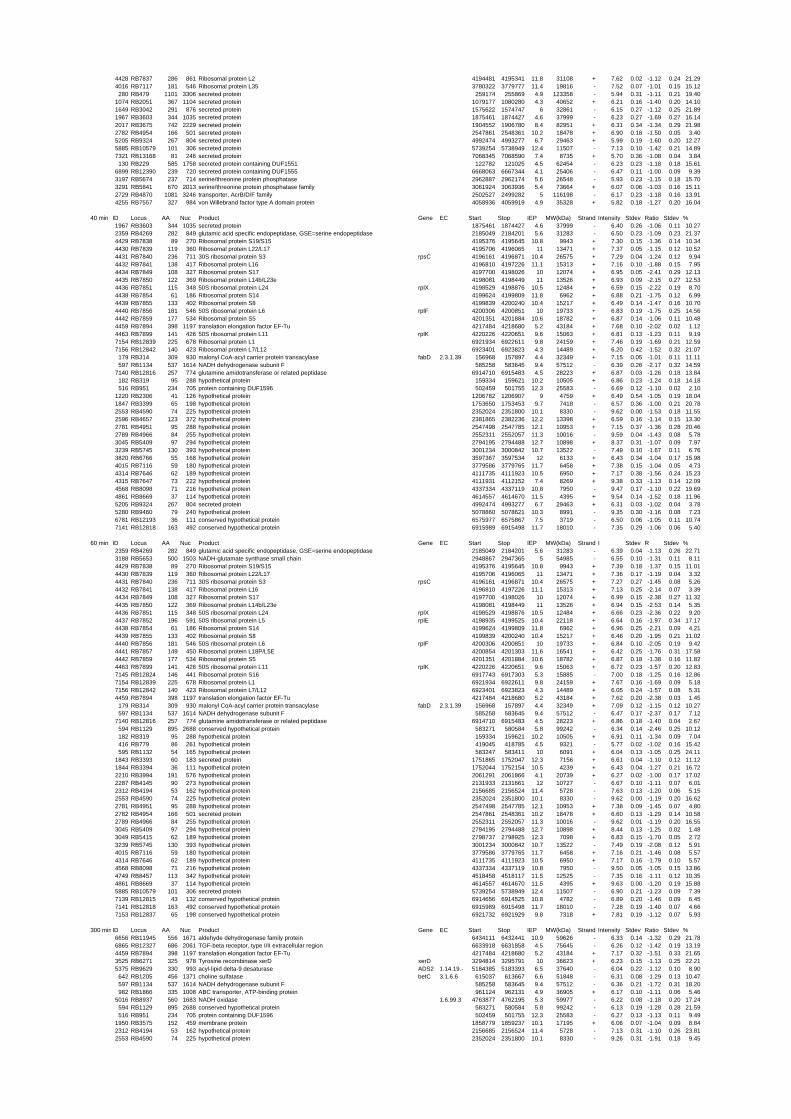
4428 RB7837 286 861 Ribosomal protein L2 4194481 4195341 11.8 31108 + 7.62 0.02 -1.12 0.24 21.294016 RB7117 181 546 Ribosomal protein L35 3780322 3779777 11.4 19816 - 7.52 0.07 -1.01 0.15 15.12
280 RB479 1101 3306 secreted protein 259174 255869 4.9 123358 - 5.94 0.31 -1.11 0.21 19.401074 RB2051 367 1104 secreted protein 1079177 1080280 4.3 40652 + 6.21 0.16 -1.40 0.20 14.101649 RB3042 291 876 secreted protein 1575622 1574747 6 32861 - 6.15 0.27 -1.12 0.25 21.891967 RB3603 344 1035 secreted protein 1875461 1874427 4.6 37999 - 6.23 0.27 -1.69 0.27 16.142017 RB3675 742 2229 secreted protein 1904552 1906780 8.4 82951 + 6.31 0.34 -1.34 0.29 21.982782 RB4954 166 501 secreted protein 2547861 2548361 10.2 18478 + 6.90 0.18 -1.50 0.05 3.405205 RB9324 267 804 secreted protein 4992474 4993277 6.7 29463 + 5.99 0.19 -1.60 0.20 12.275885 RB10579 101 306 secreted protein 5739254 5738949 12.4 11507 - 7.13 0.10 -1.42 0.21 14.897321 RB13168 81 246 secreted protein 7068345 7068590 7.4 8735 + 5.70 0.36 -1.08 0.04 3.84
130 RB229 585 1758 secreted protein containing DUF1551 122782 121025 4.5 62454 - 6.23 0.23 -1.18 0.18 15.616899 RB12390 239 720 secreted protein containing DUF1555 6668063 6667344 4.1 25406 - 6.47 0.11 -1.00 0.09 9.393197 RB5674 237 714 serine/threonine protein phosphatase 2962887 2962174 5.6 26548 - 5.93 0.23 -1.15 0.18 15.703291 RB5841 670 2013 serine/threonine protein phosphatase family 3061924 3063936 5.4 73664 + 6.07 0.06 -1.03 0.16 15.112729 RB4870 1081 3246 transporter, AcrB/D/F family 2502527 2499282 5 116198 - 6.17 0.23 -1.18 0.16 13.914255 RB7557 327 984 von Willebrand factor type A domain protein 4058936 4059919 4.9 35328 + 5.82 0.18 -1.27 0.20 16.04
40 min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand Intensity Stdev Ratio Stdev %1967 RB3603 344 1035 secreted protein 1875461 1874427 4.6 37999 - 6.40 0.26 -1.06 0.11 10.272359 RB4269 282 849 glutamic acid specific endopeptidase, GSE=serine endopeptidase 2185049 2184201 5.6 31283 - 6.50 0.23 -1.09 0.23 21.374429 RB7838 89 270 Ribosomal protein S19/S15 4195376 4195645 10.8 9943 + 7.30 0.15 -1.36 0.14 10.344430 RB7839 119 360 Ribosomal protein L22/L17 4195706 4196065 11 13471 + 7.37 0.05 -1.15 0.12 10.524431 RB7840 236 711 30S ribosomal protein S3 rpsC 4196161 4196871 10.4 26575 + 7.29 0.04 -1.24 0.12 9.944432 RB7841 138 417 Ribosomal protein L16 4196810 4197226 11.1 15313 + 7.16 0.10 -1.88 0.15 7.954434 RB7849 108 327 Ribosomal protein S17 4197700 4198026 10 12074 + 6.95 0.05 -2.41 0.29 12.134435 RB7850 122 369 Ribosomal protein L14b/L23e 4198081 4198449 11 13526 + 6.93 0.09 -2.15 0.27 12.534436 RB7851 115 348 50S ribosomal protein L24 rplX 4198529 4198876 10.5 12484 + 6.59 0.15 -2.22 0.19 8.704438 RB7854 61 186 Ribosomal protein S14 4199624 4199809 11.8 6962 + 6.88 0.21 -1.75 0.12 6.994439 RB7855 133 402 Ribosomal protein S8 4199839 4200240 10.4 15217 + 6.49 0.14 -1.47 0.16 10.704440 RB7856 181 546 50S ribosomal protein L6 rplF 4200306 4200851 10 19733 + 6.83 0.19 -1.75 0.25 14.564442 RB7859 177 534 Ribosomal protein S5 4201351 4201884 10.6 18782 + 6.87 0.14 -1.06 0.11 10.484459 RB7894 398 1197 translation elongation factor EF-Tu 4217484 4218680 5.2 43184 + 7.68 0.10 -2.02 0.02 1.124463 RB7899 141 426 50S ribosomal protein L11 rplK 4220226 4220651 9.6 15063 + 6.81 0.13 -1.23 0.11 9.197154 RB12839 225 678 Ribosomal protein L1 6921934 6922611 9.8 24159 + 7.46 0.19 -1.69 0.21 12.597156 RB12842 140 423 Ribosomal protein L7/L12 6923401 6923823 4.3 14489 + 6.20 0.42 -1.52 0.32 21.07
179 RB314 309 930 malonyl CoA-acyl carrier protein transacylase fabD 2.3.1.39 156968 157897 4.4 32349 + 7.15 0.05 -1.01 0.11 11.11597 RB1134 537 1614 NADH dehydrogenase subunit F 585258 583645 9.4 57512 - 6.39 0.26 -2.17 0.32 14.59
7140 RB12816 257 774 glutamine amidotransferase or related peptidase 6914710 6915483 4.5 28223 + 6.87 0.03 -1.26 0.18 13.84182 RB319 95 288 hypothetical protein 159334 159621 10.2 10505 + 6.86 0.23 -1.24 0.18 14.18516 RB951 234 705 protein containing DUF1596 502459 501755 12.3 25583 - 6.69 0.12 -1.10 0.02 2.10
1220 RB2306 41 126 hypothetical protein 1206782 1206907 9 4759 + 6.49 0.54 -1.05 0.19 18.041847 RB3399 65 198 hypothetical protein 1753650 1753453 9.7 7418 - 6.57 0.36 -1.00 0.21 20.782553 RB4590 74 225 hypothetical protein 2352024 2351800 10.1 8330 - 9.62 0.00 -1.53 0.18 11.552596 RB4657 123 372 hypothetical protein 2381865 2382236 12.2 13398 + 6.59 0.16 -1.14 0.15 13.302781 RB4951 95 288 hypothetical protein 2547498 2547785 12.1 10953 + 7.15 0.37 -1.36 0.28 20.462789 RB4966 84 255 hypothetical protein 2552311 2552057 11.3 10016 - 9.59 0.04 -1.43 0.08 5.783045 RB5409 97 294 hypothetical protein 2794195 2794488 12.7 10898 + 8.37 0.31 -1.07 0.09 7.973239 RB5745 130 393 hypothetical protein 3001234 3000842 10.7 13522 - 7.49 0.10 -1.67 0.11 6.763820 RB6766 55 168 hypothetical protein 3597367 3597534 12 6133 + 6.43 0.34 -1.04 0.17 15.984015 RB7116 59 180 hypothetical protein 3779586 3779765 11.7 6458 + 7.38 0.15 -1.04 0.05 4.734314 RB7646 62 189 hypothetical protein 4111735 4111923 10.5 6950 + 7.17 0.38 -1.56 0.24 15.234315 RB7647 73 222 hypothetical protein 4111931 4112152 7.4 8269 + 9.38 0.33 -1.13 0.14 12.094568 RB8098 71 216 hypothetical protein 4337334 4337119 10.8 7950 - 9.47 0.17 -1.10 0.22 19.694861 RB8669 37 114 hypothetical protein 4614557 4614670 11.5 4395 + 9.54 0.14 -1.52 0.18 11.965205 RB9324 267 804 secreted protein 4992474 4993277 6.7 29463 + 6.31 0.03 -1.02 0.04 3.785280 RB9460 79 240 hypothetical protein 5078860 5078621 10.3 8991 - 9.35 0.30 -1.16 0.08 7.236781 RB12193 36 111 conserved hypothetical protein 6575977 6575867 7.5 3719 - 6.50 0.06 -1.05 0.11 10.747141 RB12818 163 492 conserved hypothetical protein 6915989 6915498 11.7 18010 - 7.35 0.29 -1.06 0.06 5.40
60 min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand I Stdev R Stdev %2359 RB4269 282 849 glutamic acid specific endopeptidase, GSE=serine endopeptidase 2185049 2184201 5.6 31283 - 6.39 0.04 -1.13 0.26 22.713188 RB5653 500 1503 NADH-glutamate synthase small chain 2948867 2947365 5 54985 - 6.55 0.10 -1.31 0.11 8.114429 RB7838 89 270 Ribosomal protein S19/S15 4195376 4195645 10.8 9943 + 7.39 0.18 -1.37 0.15 11.014430 RB7839 119 360 Ribosomal protein L22/L17 4195706 4196065 11 13471 + 7.36 0.17 -1.19 0.04 3.324431 RB7840 236 711 30S ribosomal protein S3 rpsC 4196161 4196871 10.4 26575 + 7.27 0.27 -1.45 0.08 5.264432 RB7841 138 417 Ribosomal protein L16 4196810 4197226 11.1 15313 + 7.13 0.25 -2.14 0.07 3.394434 RB7849 108 327 Ribosomal protein S17 4197700 4198026 10 12074 + 6.99 0.15 -2.38 0.27 11.324435 RB7850 122 369 Ribosomal protein L14b/L23e 4198081 4198449 11 13526 + 6.94 0.15 -2.53 0.14 5.354436 RB7851 115 348 50S ribosomal protein L24 rplX 4198529 4198876 10.5 12484 + 6.66 0.23 -2.36 0.22 9.204437 RB7852 196 591 50S ribosomal protein L5 rplE 4198935 4199525 10.4 22118 + 6.64 0.16 -1.97 0.34 17.174438 RB7854 61 186 Ribosomal protein S14 4199624 4199809 11.8 6962 + 6.96 0.25 -2.21 0.09 4.214439 RB7855 133 402 Ribosomal protein S8 4199839 4200240 10.4 15217 + 6.46 0.20 -1.95 0.21 11.024440 RB7856 181 546 50S ribosomal protein L6 rplF 4200306 4200851 10 19733 + 6.84 0.10 -2.05 0.19 9.424441 RB7857 149 450 Ribosomal protein L18P/L5E 4200854 4201303 11.6 16541 + 6.42 0.25 -1.76 0.31 17.584442 RB7859 177 534 Ribosomal protein S5 4201351 4201884 10.6 18782 + 6.87 0.18 -1.38 0.16 11.824463 RB7899 141 426 50S ribosomal protein L11 rplK 4220226 4220651 9.6 15063 + 6.72 0.23 -1.57 0.20 12.837145 RB12824 146 441 Ribosomal protein S16 6917743 6917303 5.3 15885 - 7.00 0.18 -1.25 0.16 12.867154 RB12839 225 678 Ribosomal protein L1 6921934 6922611 9.8 24159 + 7.67 0.16 -1.69 0.09 5.187156 RB12842 140 423 Ribosomal protein L7/L12 6923401 6923823 4.3 14489 + 6.05 0.24 -1.57 0.08 5.314459 RB7894 398 1197 translation elongation factor EF-Tu 4217484 4218680 5.2 43184 + 7.62 0.20 -2.38 0.03 1.45
179 RB314 309 930 malonyl CoA-acyl carrier protein transacylase fabD 2.3.1.39 156968 157897 4.4 32349 + 7.09 0.12 -1.15 0.12 10.27597 RB1134 537 1614 NADH dehydrogenase subunit F 585258 583645 9.4 57512 - 6.47 0.17 -2.37 0.17 7.12
7140 RB12816 257 774 glutamine amidotransferase or related peptidase 6914710 6915483 4.5 28223 + 6.86 0.18 -1.40 0.04 2.67594 RB1129 895 2688 conserved hypothetical protein 583271 580584 5.8 99242 - 6.34 0.14 -2.46 0.25 10.12182 RB319 95 288 hypothetical protein 159334 159621 10.2 10505 + 6.91 0.11 -1.34 0.09 7.04416 RB779 86 261 hypothetical protein 419045 418785 4.5 9321 - 5.77 0.02 -1.02 0.16 15.42595 RB1132 54 165 hypothetical protein 583247 583411 10 6091 + 6.04 0.13 -1.05 0.25 24.11
1843 RB3393 60 183 secreted protein 1751865 1752047 12.3 7156 + 6.61 0.04 -1.10 0.12 11.121844 RB3394 36 111 hypothetical protein 1752044 1752154 10.5 4239 + 6.43 0.04 -1.27 0.21 16.722210 RB3994 191 576 hypothetical protein 2061291 2061866 4.1 20739 + 6.27 0.02 -1.00 0.17 17.022287 RB4145 90 273 hypothetical protein 2131933 2131661 12 10727 - 6.67 0.10 -1.11 0.07 6.012312 RB4194 53 162 hypothetical protein 2156685 2156524 11.4 5728 - 7.63 0.13 -1.20 0.06 5.152553 RB4590 74 225 hypothetical protein 2352024 2351800 10.1 8330 - 9.62 0.00 -1.19 0.20 16.622781 RB4951 95 288 hypothetical protein 2547498 2547785 12.1 10953 + 7.38 0.09 -1.45 0.07 4.802782 RB4954 166 501 secreted protein 2547861 2548361 10.2 18478 + 6.60 0.13 -1.29 0.14 10.582789 RB4966 84 255 hypothetical protein 2552311 2552057 11.3 10016 - 9.62 0.01 -1.19 0.20 16.553045 RB5409 97 294 hypothetical protein 2794195 2794488 12.7 10898 + 8.44 0.13 -1.25 0.02 1.483049 RB5415 62 189 hypothetical protein 2798737 2798925 12.3 7098 + 6.83 0.15 -1.70 0.05 2.723239 RB5745 130 393 hypothetical protein 3001234 3000842 10.7 13522 - 7.49 0.19 -2.08 0.12 5.914015 RB7116 59 180 hypothetical protein 3779586 3779765 11.7 6458 + 7.16 0.21 -1.46 0.08 5.574314 RB7646 62 189 hypothetical protein 4111735 4111923 10.5 6950 + 7.17 0.16 -1.79 0.10 5.574568 RB8098 71 216 hypothetical protein 4337334 4337119 10.8 7950 - 9.50 0.05 -1.05 0.15 13.864749 RB8457 113 342 hypothetical protein 4518458 4518117 11.5 12525 - 7.35 0.16 -1.11 0.12 10.354861 RB8669 37 114 hypothetical protein 4614557 4614670 11.5 4395 + 9.63 0.00 -1.20 0.19 15.885885 RB10579 101 306 secreted protein 5739254 5738949 12.4 11507 - 6.90 0.21 -1.23 0.09 7.397139 RB12815 43 132 conserved hypothetical protein 6914656 6914525 10.8 4782 - 6.89 0.20 -1.46 0.09 6.457141 RB12818 163 492 conserved hypothetical protein 6915989 6915498 11.7 18010 - 7.28 0.19 -1.40 0.07 4.667153 RB12837 65 198 conserved hypothetical protein 6921732 6921929 9.8 7318 + 7.81 0.19 -1.12 0.07 5.93
300 min ID Locus AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand Intensity Stdev Ratio Stdev %6656 RB11945 556 1671 aldehyde dehydrogenase family protein 6434111 6432441 10.9 59626 - 6.33 0.14 -1.32 0.29 21.786865 RB12327 686 2061 TGF-beta receptor, type I/II extracellular region 6633918 6631858 4.5 75645 - 6.26 0.12 -1.42 0.19 13.194459 RB7894 398 1197 translation elongation factor EF-Tu 4217484 4218680 5.2 43184 + 7.17 0.32 -1.51 0.33 21.653525 RB6271 325 978 Tyrosine recombinase xerD xerD 3294814 3295791 10 36623 + 6.23 0.15 -1.13 0.25 22.215375 RB9629 330 993 acyl-lipid delta-9 desaturase ADS2 1.14.19.- 5184385 5183393 6.5 37640 - 6.04 0.22 -1.12 0.10 8.90
642 RB1205 456 1371 choline sulfatase betC 3.1.6.6 615037 613667 6.6 51848 - 6.31 0.08 -1.29 0.13 10.47597 RB1134 537 1614 NADH dehydrogenase subunit F 585258 583645 9.4 57512 - 6.36 0.21 -1.72 0.31 18.20982 RB1866 335 1008 ABC transporter, ATP-binding protein 961124 962131 4.9 36905 + 6.17 0.10 -1.11 0.06 5.46
5016 RB8937 560 1683 NADH oxidase 1.6.99.3 4763877 4762195 5.3 59977 - 6.22 0.08 -1.18 0.20 17.24594 RB1129 895 2688 conserved hypothetical protein 583271 580584 5.8 99242 - 6.13 0.19 -1.28 0.28 21.59516 RB951 234 705 protein containing DUF1596 502459 501755 12.3 25583 - 6.27 0.13 -1.13 0.11 9.49
1950 RB3575 152 459 membrane protein 1858779 1859237 10.1 17195 + 6.06 0.07 -1.04 0.09 8.842312 RB4194 53 162 hypothetical protein 2156685 2156524 11.4 5728 - 7.13 0.31 -1.10 0.26 23.812553 RB4590 74 225 hypothetical protein 2352024 2351800 10.1 8330 - 9.26 0.31 -1.91 0.18 9.45
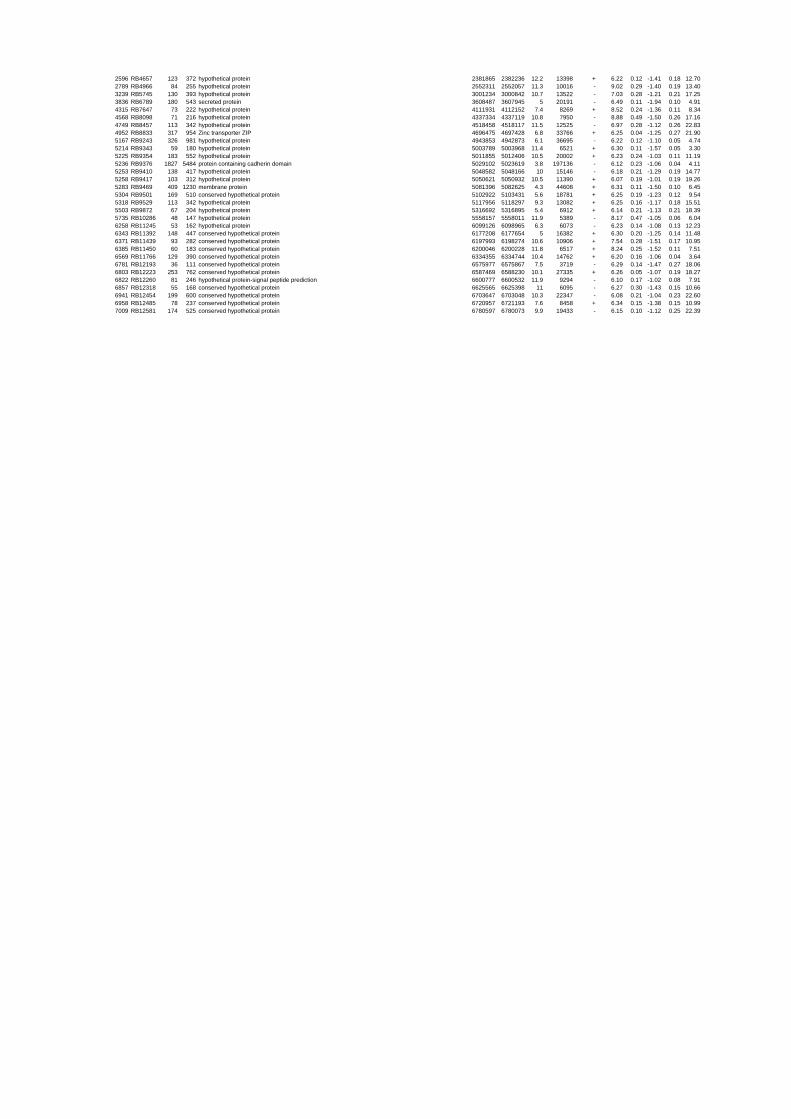
2596 RB4657 123 372 hypothetical protein 2381865 2382236 12.2 13398 + 6.22 0.12 -1.41 0.18 12.702789 RB4966 84 255 hypothetical protein 2552311 2552057 11.3 10016 - 9.02 0.29 -1.40 0.19 13.403239 RB5745 130 393 hypothetical protein 3001234 3000842 10.7 13522 - 7.03 0.28 -1.21 0.21 17.253836 RB6789 180 543 secreted protein 3608487 3607945 5 20191 - 6.49 0.11 -1.94 0.10 4.914315 RB7647 73 222 hypothetical protein 4111931 4112152 7.4 8269 + 8.52 0.24 -1.36 0.11 8.344568 RB8098 71 216 hypothetical protein 4337334 4337119 10.8 7950 - 8.88 0.49 -1.50 0.26 17.164749 RB8457 113 342 hypothetical protein 4518458 4518117 11.5 12525 - 6.97 0.28 -1.12 0.26 22.834952 RB8833 317 954 Zinc transporter ZIP 4696475 4697428 6.8 33766 + 6.25 0.04 -1.25 0.27 21.905167 RB9243 326 981 hypothetical protein 4943853 4942873 6.1 36695 - 6.22 0.12 -1.10 0.05 4.745214 RB9343 59 180 hypothetical protein 5003789 5003968 11.4 6521 + 6.30 0.11 -1.57 0.05 3.305225 RB9354 183 552 hypothetical protein 5011855 5012406 10.5 20002 + 6.23 0.24 -1.03 0.11 11.195236 RB9376 1827 5484 protein containing cadherin domain 5029102 5023619 3.8 197136 - 6.12 0.23 -1.06 0.04 4.115253 RB9410 138 417 hypothetical protein 5048582 5048166 10 15146 - 6.18 0.21 -1.29 0.19 14.775258 RB9417 103 312 hypothetical protein 5050621 5050932 10.5 11390 + 6.07 0.19 -1.01 0.19 19.265283 RB9469 409 1230 membrane protein 5081396 5082625 4.3 44608 + 6.31 0.11 -1.50 0.10 6.455304 RB9501 169 510 conserved hypothetical protein 5102922 5103431 5.6 18781 + 6.25 0.19 -1.23 0.12 9.545318 RB9529 113 342 hypothetical protein 5117956 5118297 9.3 13082 + 6.25 0.16 -1.17 0.18 15.515503 RB9872 67 204 hypothetical protein 5316692 5316895 5.4 6912 + 6.14 0.21 -1.13 0.21 18.395735 RB10286 48 147 hypothetical protein 5558157 5558011 11.9 5389 - 8.17 0.47 -1.05 0.06 6.046258 RB11245 53 162 hypothetical protein 6099126 6098965 6.3 6073 - 6.23 0.14 -1.08 0.13 12.236343 RB11392 148 447 conserved hypothetical protein 6177208 6177654 5 16382 + 6.30 0.20 -1.25 0.14 11.486371 RB11439 93 282 conserved hypothetical protein 6197993 6198274 10.6 10906 + 7.54 0.28 -1.51 0.17 10.956385 RB11450 60 183 conserved hypothetical protein 6200046 6200228 11.8 6517 + 8.24 0.25 -1.52 0.11 7.516569 RB11766 129 390 conserved hypothetical protein 6334355 6334744 10.4 14762 + 6.20 0.16 -1.06 0.04 3.646781 RB12193 36 111 conserved hypothetical protein 6575977 6575867 7.5 3719 - 6.29 0.14 -1.47 0.27 18.066803 RB12223 253 762 conserved hypothetical protein 6587469 6588230 10.1 27335 + 6.26 0.05 -1.07 0.19 18.276822 RB12260 81 246 hypothetical protein-signal peptide prediction 6600777 6600532 11.9 9294 - 6.10 0.17 -1.02 0.08 7.916857 RB12318 55 168 conserved hypothetical protein 6625565 6625398 11 6095 - 6.27 0.30 -1.43 0.15 10.666941 RB12454 199 600 conserved hypothetical protein 6703647 6703048 10.3 22347 - 6.08 0.21 -1.04 0.23 22.606958 RB12485 78 237 conserved hypothetical protein 6720957 6721193 7.6 8458 + 6.34 0.15 -1.38 0.15 10.997009 RB12581 174 525 conserved hypothetical protein 6780597 6780073 9.9 19433 - 6.15 0.10 -1.12 0.25 22.39
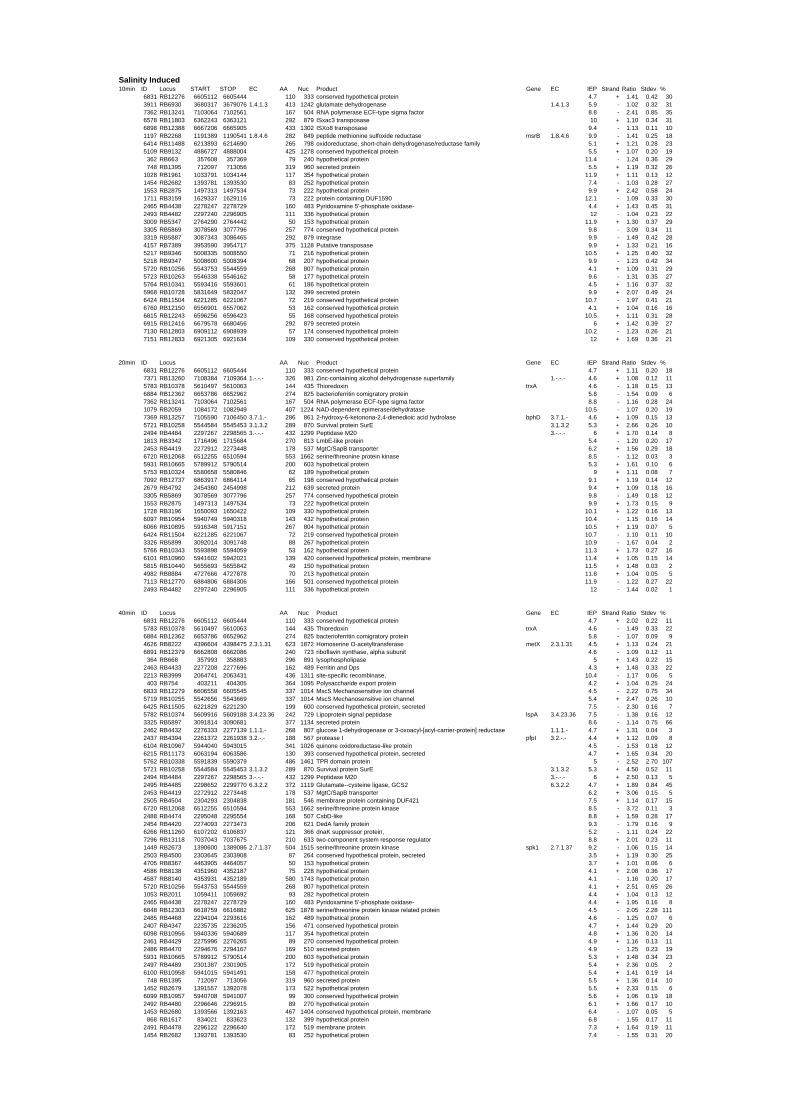
Salinity Induced10min ID Locus START STOP EC AA Nuc Product Gene EC IEP Strand Ratio Stdev %
6831 RB12276 6605112 6605444 110 333 conserved hypothetical protein 4.7 + 1.41 0.42 303911 RB6930 3680317 3679076 1.4.1.3 413 1242 glutamate dehydrogenase 1.4.1.3 5.9 - 1.02 0.32 317362 RB13241 7103064 7102561 167 504 RNA polymerase ECF-type sigma factor 8.8 - 2.41 0.85 356578 RB11803 6362243 6363121 292 879 ISxac3 transposase 10 + 1.10 0.34 316898 RB12388 6667206 6665905 433 1302 ISXo8 transposase 9.4 - 1.13 0.11 101197 RB2268 1191389 1190541 1.8.4.6 282 849 peptide methionine sulfoxide reductase msrB 1.8.4.6 9.9 - 1.41 0.25 186414 RB11488 6213893 6214690 265 798 oxidoreductase, short-chain dehydrogenase/reductase family 5.1 + 1.21 0.28 235109 RB9132 4886727 4888004 425 1278 conserved hypothetical protein 5.5 + 1.07 0.20 19362 RB663 357608 357369 79 240 hypothetical protein 11.4 - 1.24 0.36 29748 RB1395 712097 713056 319 960 secreted protein 5.5 + 1.19 0.32 26
1028 RB1961 1033791 1034144 117 354 hypothetical protein 11.9 + 1.11 0.13 121454 RB2682 1393781 1393530 83 252 hypothetical protein 7.4 - 1.03 0.28 271553 RB2875 1497313 1497534 73 222 hypothetical protein 9.9 + 2.42 0.58 241711 RB3159 1629337 1629116 73 222 protein containing DUF1590 12.1 - 1.09 0.33 302465 RB4438 2278247 2278729 160 483 Pyridoxamine 5'-phosphate oxidase- 4.4 + 1.43 0.45 312493 RB4482 2297240 2296905 111 336 hypothetical protein 12 - 1.04 0.23 223009 RB5347 2764290 2764442 50 153 hypothetical protein 11.9 + 1.30 0.37 293305 RB5869 3078569 3077796 257 774 conserved hypothetical protein 9.8 - 3.09 0.34 113319 RB5887 3087343 3086465 292 879 integrase 9.9 - 1.49 0.42 284157 RB7389 3953590 3954717 375 1128 Putative transposase 9.9 + 1.33 0.21 165217 RB9346 5008335 5008550 71 216 hypothetical protein 10.5 + 1.25 0.40 325218 RB9347 5008600 5008394 68 207 hypothetical protein 9.9 - 1.23 0.42 345720 RB10256 5543753 5544559 268 807 hypothetical protein 4.1 + 1.09 0.31 295723 RB10263 5546338 5546162 58 177 hypothetical protein 9.6 - 1.31 0.35 275764 RB10341 5593416 5593601 61 186 hypothetical protein 4.5 + 1.16 0.37 325968 RB10728 5831649 5832047 132 399 secreted protein 9.9 + 2.07 0.49 246424 RB11504 6221285 6221067 72 219 conserved hypothetical protein 10.7 - 1.97 0.41 216760 RB12150 6556901 6557062 53 162 conserved hypothetical protein 4.1 + 1.04 0.16 166815 RB12243 6596256 6596423 55 168 conserved hypothetical protein 10.5 + 1.11 0.31 286915 RB12416 6679578 6680456 292 879 secreted protein 6 + 1.42 0.39 277130 RB12803 6909112 6908939 57 174 conserved hypothetical protein 10.2 - 1.23 0.26 217151 RB12833 6921305 6921634 109 330 conserved hypothetical protein 12 + 1.69 0.36 21
20min ID Locus AA Nuc Product Gene EC IEP Strand Ratio Stdev %6831 RB12276 6605112 6605444 110 333 conserved hypothetical protein 4.7 + 1.11 0.20 187371 RB13260 7108384 7109364 1.-.-.- 326 981 Zinc-containing alcohol dehydrogenase superfamily 1.-.-.- 4.6 + 1.08 0.12 115783 RB10378 5610497 5610063 144 435 Thioredoxin trxA 4.6 - 1.18 0.15 136884 RB12362 6653786 6652962 274 825 bacterioferritin comigratory protein 5.8 - 1.54 0.09 67362 RB13241 7103064 7102561 167 504 RNA polymerase ECF-type sigma factor 8.8 - 1.16 0.28 241079 RB2059 1084172 1082949 407 1224 NAD-dependent epimerase/dehydratase 10.5 - 1.07 0.20 197369 RB13257 7105590 7106450 3.7.1.- 286 861 2-hydroxy-6-ketonona-2,4-dienedioic acid hydrolase bphD 3.7.1.- 4.6 + 1.09 0.15 135721 RB10258 5544584 5545453 3.1.3.2 289 870 Survival protein SurE 3.1.3.2 5.3 + 2.66 0.26 102494 RB4484 2297267 2298565 3.-.-.- 432 1299 Peptidase M20 3.-.-.- 6 + 1.70 0.14 81813 RB3342 1716496 1715684 270 813 LmbE-like protein 5.4 - 1.20 0.20 172453 RB4419 2272912 2273448 178 537 MgtC/SapB transporter 6.2 + 1.56 0.29 186720 RB12068 6512255 6510594 553 1662 serine/threonine protein kinase 8.5 - 1.12 0.03 35931 RB10665 5789912 5790514 200 603 hypothetical protein 5.3 + 1.61 0.10 65753 RB10324 5580658 5580846 62 189 hypothetical protein 9 + 1.11 0.08 77092 RB12737 6863917 6864114 65 198 conserved hypothetical protein 9.1 + 1.19 0.14 122679 RB4792 2454360 2454998 212 639 secreted protein 9.4 + 1.09 0.18 163305 RB5869 3078569 3077796 257 774 conserved hypothetical protein 9.8 - 1.49 0.18 121553 RB2875 1497313 1497534 73 222 hypothetical protein 9.9 + 1.73 0.15 91728 RB3196 1650093 1650422 109 330 hypothetical protein 10.1 + 1.22 0.16 136097 RB10954 5940749 5940318 143 432 hypothetical protein 10.4 - 1.15 0.16 146066 RB10895 5916348 5917151 267 804 hypothetical protein 10.5 + 1.19 0.07 56424 RB11504 6221285 6221067 72 219 conserved hypothetical protein 10.7 - 1.10 0.11 103326 RB5899 3092014 3091748 88 267 hypothetical protein 10.9 - 1.67 0.04 25766 RB10343 5593898 5594059 53 162 hypothetical protein 11.3 + 1.73 0.27 166101 RB10960 5941602 5942021 139 420 conserved hypothetical protein, membrane 11.4 + 1.05 0.15 145815 RB10440 5655693 5655842 49 150 hypothetical protein 11.5 + 1.48 0.03 24982 RB8884 4727666 4727878 70 213 hypothetical protein 11.8 + 1.04 0.05 57113 RB12770 6884806 6884306 166 501 conserved hypothetical protein 11.9 - 1.22 0.27 222493 RB4482 2297240 2296905 111 336 hypothetical protein 12 - 1.44 0.02 1
40min ID Locus AA Nuc Product Gene EC IEP Strand Ratio Stdev %6831 RB12276 6605112 6605444 110 333 conserved hypothetical protein 4.7 + 2.02 0.22 115783 RB10378 5610497 5610063 144 435 Thioredoxin trxA 4.6 - 1.49 0.33 226884 RB12362 6653786 6652962 274 825 bacterioferritin comigratory protein 5.8 - 1.07 0.09 94626 RB8222 4396604 4398475 2.3.1.31 623 1872 Homoserine O-acetyltransferase metX 2.3.1.31 4.5 + 1.13 0.24 216891 RB12379 6662808 6662086 240 723 riboflavin synthase, alpha subunit 4.6 - 1.09 0.12 11364 RB668 357993 358883 296 891 lysophospholipase 5 + 1.43 0.22 15
2463 RB4433 2277208 2277696 162 489 Ferritin and Dps 4.3 + 1.48 0.33 222213 RB3999 2064741 2063431 436 1311 site-specific recombinase, 10.4 - 1.17 0.06 5403 RB754 403211 404305 364 1095 Polysaccharide export protein 4.2 + 1.04 0.25 24
6833 RB12279 6606558 6605545 337 1014 MscS Mechanosensitive ion channel 4.5 - 2.22 0.75 345719 RB10255 5542656 5543669 337 1014 MscS Mechanosensitive ion channel 5.4 + 2.47 0.26 106425 RB11505 6221829 6221230 199 600 conserved hypothetical protein, secreted 7.5 - 2.30 0.16 75782 RB10374 5609916 5609188 3.4.23.36 242 729 Lipoprotein signal peptidase lspA 3.4.23.36 7.5 - 1.38 0.16 123325 RB5897 3091814 3090681 377 1134 secreted protein 8.6 - 1.14 0.75 662462 RB4432 2276333 2277139 1.1.1.- 268 807 glucose 1-dehydrogenase or 3-oxoacyl-[acyl-carrier-protein] reductase 1.1.1.- 4.7 + 1.31 0.04 32437 RB4394 2261372 2261938 3.2.-.- 188 567 protease I pfpI 3.2.-.- 4.4 + 1.12 0.09 86104 RB10967 5944040 5943015 341 1026 quinone oxidoreductase-like protein 4.5 - 1.53 0.18 126215 RB11173 6063194 6063586 130 393 conserved hypothetical protein, secreted 4.7 + 1.65 0.34 205762 RB10338 5591839 5590379 486 1461 TPR domain protein 5 - 2.52 2.70 1075721 RB10258 5544584 5545453 3.1.3.2 289 870 Survival protein SurE 3.1.3.2 5.3 + 4.50 0.52 112494 RB4484 2297267 2298565 3.-.-.- 432 1299 Peptidase M20 3.-.-.- 6 + 2.50 0.13 52495 RB4485 2298652 2299770 6.3.2.2 372 1119 Glutamate--cysteine ligase, GCS2 6.3.2.2 4.7 + 1.89 0.84 452453 RB4419 2272912 2273448 178 537 MgtC/SapB transporter 6.2 + 3.06 0.15 52505 RB4504 2304293 2304838 181 546 membrane protein containing DUF421 7.5 + 1.14 0.17 156720 RB12068 6512255 6510594 553 1662 serine/threonine protein kinase 8.5 - 3.72 0.11 32488 RB4474 2295048 2295554 168 507 CsbD-like 8.8 + 1.59 0.28 172454 RB4420 2274093 2273473 206 621 DedA family protein 9.3 - 1.79 0.16 96266 RB11260 6107202 6106837 121 366 dnaK suppressor protein, 5.2 - 1.11 0.24 227296 RB13118 7037043 7037675 210 633 two-component system response regulator 8.8 + 2.01 0.23 111449 RB2673 1390600 1389086 2.7.1.37 504 1515 serine/threonine protein kinase spk1 2.7.1.37 9.2 - 1.06 0.15 142503 RB4500 2303645 2303908 87 264 conserved hypothetical protein, secreted 3.5 + 1.19 0.30 254705 RB8367 4463905 4464057 50 153 hypothetical protein 3.7 + 1.01 0.06 64586 RB8138 4351960 4352187 75 228 hypothetical protein 4.1 + 2.08 0.36 174587 RB8140 4353931 4352189 580 1743 hypothetical protein 4.1 - 1.16 0.20 175720 RB10256 5543753 5544559 268 807 hypothetical protein 4.1 + 2.51 0.65 261053 RB2011 1059411 1059692 93 282 hypothetical protein 4.4 + 1.04 0.13 122465 RB4438 2278247 2278729 160 483 Pyridoxamine 5'-phosphate oxidase- 4.4 + 1.95 0.16 86848 RB12303 6618759 6616882 625 1878 serine/threonine protein kinase related protein 4.5 - 2.05 2.28 1112485 RB4468 2294104 2293616 162 489 hypothetical protein 4.6 - 1.25 0.07 62407 RB4347 2235735 2236205 156 471 conserved hypothetical protein 4.7 + 1.44 0.29 206098 RB10956 5940336 5940689 117 354 hypothetical protein 4.8 + 1.36 0.20 142461 RB4429 2275996 2276265 89 270 conserved hypothetical protein 4.9 + 1.16 0.13 112486 RB4470 2294676 2294167 169 510 secreted protein 4.9 - 1.25 0.23 195931 RB10665 5789912 5790514 200 603 hypothetical protein 5.3 + 1.48 0.34 232497 RB4489 2301387 2301905 172 519 hypothetical protein 5.4 + 2.36 0.05 26100 RB10958 5941015 5941491 158 477 hypothetical protein 5.4 + 1.41 0.19 14748 RB1395 712097 713056 319 960 secreted protein 5.5 + 1.36 0.14 10
1452 RB2679 1391557 1392078 173 522 hypothetical protein 5.5 + 2.33 0.15 66099 RB10957 5940708 5941007 99 300 conserved hypothetical protein 5.6 + 1.06 0.19 182492 RB4480 2296646 2296915 89 270 hypothetical protein 6.1 + 1.66 0.17 101453 RB2680 1393566 1392163 467 1404 conserved hypothetical protein, membrane 6.4 - 1.07 0.05 5868 RB1617 834021 833623 132 399 hypothetical protein 6.8 - 1.55 0.17 11
2491 RB4478 2296122 2296640 172 519 membrane protein 7.3 + 1.64 0.19 111454 RB2682 1393781 1393530 83 252 hypothetical protein 7.4 - 1.55 0.31 20
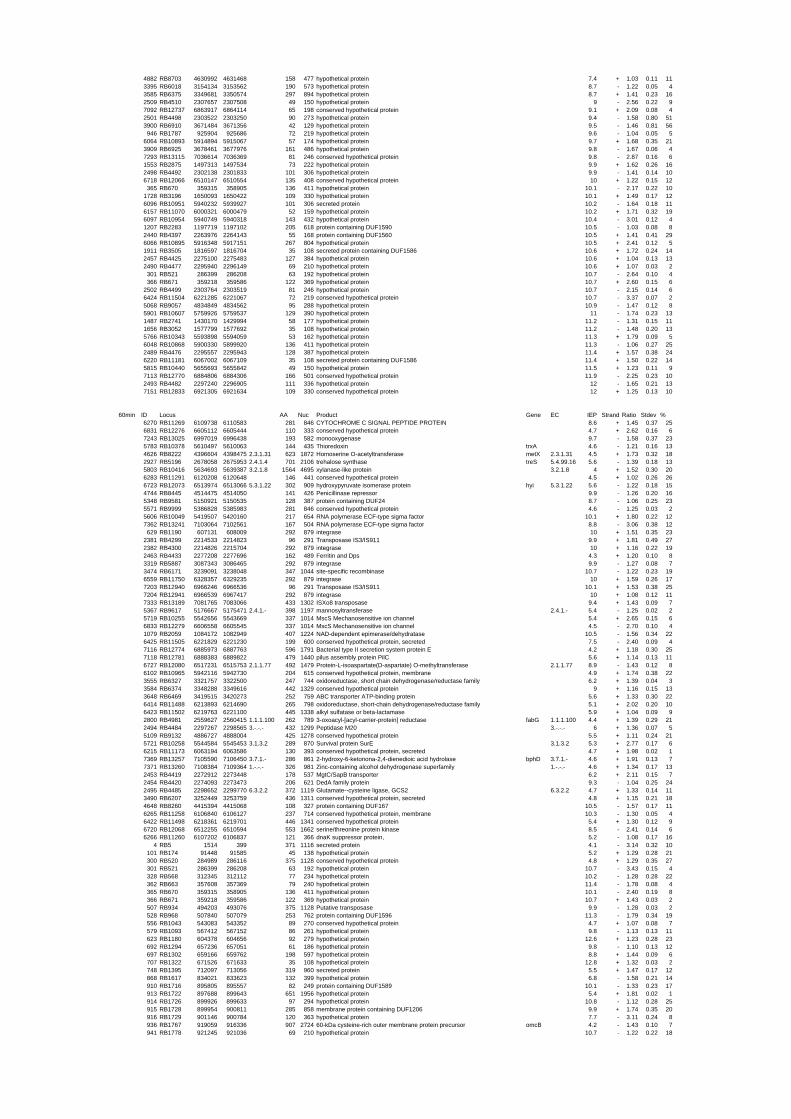
4882 RB8703 4630992 4631468 158 477 hypothetical protein 7.4 + 1.03 0.11 113395 RB6018 3154134 3153562 190 573 hypothetical protein 8.7 - 1.22 0.05 43585 RB6375 3349681 3350574 297 894 hypothetical protein 8.7 + 1.41 0.23 162509 RB4510 2307657 2307508 49 150 hypothetical protein 9 - 2.56 0.22 97092 RB12737 6863917 6864114 65 198 conserved hypothetical protein 9.1 + 2.09 0.08 42501 RB4498 2303522 2303250 90 273 hypothetical protein 9.4 - 1.58 0.80 513900 RB6910 3671484 3671356 42 129 hypothetical protein 9.5 - 1.46 0.81 56946 RB1787 925904 925686 72 219 hypothetical protein 9.6 - 1.04 0.05 5
6064 RB10893 5914894 5915067 57 174 hypothetical protein 9.7 + 1.68 0.35 213909 RB6925 3678461 3677976 161 486 hypothetical protein 9.8 - 1.67 0.06 47293 RB13115 7036614 7036369 81 246 conserved hypothetical protein 9.8 - 2.87 0.16 61553 RB2875 1497313 1497534 73 222 hypothetical protein 9.9 + 1.62 0.26 162498 RB4492 2302138 2301833 101 306 hypothetical protein 9.9 - 1.41 0.14 106718 RB12066 6510147 6510554 135 408 conserved hypothetical protein 10 + 1.22 0.15 12365 RB670 359315 358905 136 411 hypothetical protein 10.1 - 2.17 0.22 10
1728 RB3196 1650093 1650422 109 330 hypothetical protein 10.1 + 1.49 0.17 126096 RB10951 5940232 5939927 101 306 secreted protein 10.2 - 1.64 0.18 116157 RB11070 6000321 6000479 52 159 hypothetical protein 10.2 + 1.71 0.32 196097 RB10954 5940749 5940318 143 432 hypothetical protein 10.4 - 3.01 0.12 41207 RB2283 1197719 1197102 205 618 protein containing DUF1590 10.5 - 1.03 0.08 82440 RB4397 2263976 2264143 55 168 protein containing DUF1560 10.5 + 1.41 0.41 296066 RB10895 5916348 5917151 267 804 hypothetical protein 10.5 + 2.41 0.12 51911 RB3505 1816597 1816704 35 108 secreted protein containing DUF1586 10.6 + 1.72 0.24 142457 RB4425 2275100 2275483 127 384 hypothetical protein 10.6 + 1.04 0.13 132490 RB4477 2295940 2296149 69 210 hypothetical protein 10.6 + 1.07 0.03 2301 RB521 286399 286208 63 192 hypothetical protein 10.7 - 2.64 0.10 4366 RB671 359218 359586 122 369 hypothetical protein 10.7 + 2.60 0.15 6
2502 RB4499 2303764 2303519 81 246 hypothetical protein 10.7 - 2.15 0.14 66424 RB11504 6221285 6221067 72 219 conserved hypothetical protein 10.7 - 3.37 0.07 25068 RB9057 4834849 4834562 95 288 hypothetical protein 10.9 - 1.47 0.12 85901 RB10607 5759926 5759537 129 390 hypothetical protein 11 - 1.74 0.23 131487 RB2741 1430170 1429994 58 177 hypothetical protein 11.2 - 1.31 0.15 111656 RB3052 1577799 1577692 35 108 hypothetical protein 11.2 - 1.48 0.20 135766 RB10343 5593898 5594059 53 162 hypothetical protein 11.3 + 1.79 0.09 56048 RB10868 5900330 5899920 136 411 hypothetical protein 11.3 - 1.06 0.27 252489 RB4476 2295557 2295943 128 387 hypothetical protein 11.4 + 1.57 0.38 246220 RB11181 6067002 6067109 35 108 secreted protein containing DUF1586 11.4 + 1.50 0.22 145815 RB10440 5655693 5655842 49 150 hypothetical protein 11.5 + 1.23 0.11 97113 RB12770 6884806 6884306 166 501 conserved hypothetical protein 11.9 - 2.25 0.23 102493 RB4482 2297240 2296905 111 336 hypothetical protein 12 - 1.65 0.21 137151 RB12833 6921305 6921634 109 330 conserved hypothetical protein 12 + 1.25 0.13 10
60min ID Locus AA Nuc Product Gene EC IEP Strand Ratio Stdev %6270 RB11269 6109738 6110583 281 846 CYTOCHROME C SIGNAL PEPTIDE PROTEIN 8.6 + 1.45 0.37 256831 RB12276 6605112 6605444 110 333 conserved hypothetical protein 4.7 + 2.62 0.16 67243 RB13025 6997019 6996438 193 582 monooxygenase 9.7 - 1.58 0.37 235783 RB10378 5610497 5610063 144 435 Thioredoxin trxA 4.6 - 1.21 0.16 134626 RB8222 4396604 4398475 2.3.1.31 623 1872 Homoserine O-acetyltransferase metX 2.3.1.31 4.5 + 1.73 0.32 182927 RB5196 2678058 2675953 2.4.1.4 701 2106 trehalose synthase treS 5.4.99.16 5.6 - 1.39 0.18 135803 RB10416 5634693 5639387 3.2.1.8 1564 4695 xylanase-like protein 3.2.1.8 4 + 1.52 0.30 206283 RB11291 6120208 6120648 146 441 conserved hypothetical protein 4.5 + 1.02 0.26 266723 RB12073 6513974 6513066 5.3.1.22 302 909 hydroxypyruvate isomerase protein hyi 5.3.1.22 5.6 - 1.22 0.18 154744 RB8445 4514475 4514050 141 426 Penicillinase repressor 9.9 - 1.26 0.20 165348 RB9581 5150921 5150535 128 387 protein containing DUF24 8.7 - 1.06 0.25 235571 RB9999 5386828 5385983 281 846 conserved hypothetical protein 4.6 - 1.25 0.03 25606 RB10049 5419507 5420160 217 654 RNA polymerase ECF-type sigma factor 10.1 + 1.80 0.22 127362 RB13241 7103064 7102561 167 504 RNA polymerase ECF-type sigma factor 8.8 - 3.06 0.38 12629 RB1190 607131 608009 292 879 integrase 10 + 1.51 0.35 23
2381 RB4299 2214533 2214823 96 291 Transposase IS3/IS911 9.9 + 1.81 0.49 272382 RB4300 2214826 2215704 292 879 integrase 10 + 1.16 0.22 192463 RB4433 2277208 2277696 162 489 Ferritin and Dps 4.3 + 1.20 0.10 83319 RB5887 3087343 3086465 292 879 integrase 9.9 - 1.27 0.08 73474 RB6171 3239091 3238048 347 1044 site-specific recombinase 10.7 - 1.22 0.23 196559 RB11750 6328357 6329235 292 879 integrase 10 + 1.59 0.26 177203 RB12940 6966246 6966536 96 291 Transposase IS3/IS911 10.1 + 1.53 0.38 257204 RB12941 6966539 6967417 292 879 integrase 10 + 1.08 0.12 117333 RB13189 7081765 7083066 433 1302 ISXo8 transposase 9.4 + 1.43 0.09 75367 RB9617 5176667 5175471 2.4.1.- 398 1197 mannosyltransferase 2.4.1.- 5.4 - 1.25 0.02 25719 RB10255 5542656 5543669 337 1014 MscS Mechanosensitive ion channel 5.4 + 2.65 0.15 66833 RB12279 6606558 6605545 337 1014 MscS Mechanosensitive ion channel 4.5 - 2.70 0.10 41079 RB2059 1084172 1082949 407 1224 NAD-dependent epimerase/dehydratase 10.5 - 1.56 0.34 226425 RB11505 6221829 6221230 199 600 conserved hypothetical protein, secreted 7.5 - 2.40 0.09 47116 RB12774 6885973 6887763 596 1791 Bacterial type II secretion system protein E 4.2 + 1.18 0.30 257118 RB12781 6888383 6889822 479 1440 pilus assembly protein PilC 5.6 + 1.14 0.13 116727 RB12080 6517231 6515753 2.1.1.77 492 1479 Protein-L-isoaspartate(D-aspartate) O-methyltransferase 2.1.1.77 8.9 - 1.43 0.12 86102 RB10965 5942116 5942730 204 615 conserved hypothetical protein, membrane 4.9 + 1.74 0.38 223555 RB6327 3321757 3322500 247 744 oxidoreductase, short chain dehydrogenase/reductase family 6.2 + 1.39 0.04 33584 RB6374 3348288 3349616 442 1329 conserved hypothetical protein 9 + 1.16 0.15 133648 RB6469 3419515 3420273 252 759 ABC transporter ATP-binding protein 5.6 + 1.33 0.30 226414 RB11488 6213893 6214690 265 798 oxidoreductase, short-chain dehydrogenase/reductase family 5.1 + 2.02 0.20 106423 RB11502 6219763 6221100 445 1338 alkyl sulfatase or beta-lactamase 5.9 + 1.04 0.09 92800 RB4981 2559627 2560415 1.1.1.100 262 789 3-oxoacyl-[acyl-carrier-protein] reductase fabG 1.1.1.100 4.4 + 1.39 0.29 212494 RB4484 2297267 2298565 3.-.-.- 432 1299 Peptidase M20 3.-.-.- 6 + 1.36 0.07 55109 RB9132 4886727 4888004 425 1278 conserved hypothetical protein 5.5 + 1.11 0.24 215721 RB10258 5544584 5545453 3.1.3.2 289 870 Survival protein SurE 3.1.3.2 5.3 + 2.77 0.17 66215 RB11173 6063194 6063586 130 393 conserved hypothetical protein, secreted 4.7 + 1.98 0.02 17369 RB13257 7105590 7106450 3.7.1.- 286 861 2-hydroxy-6-ketonona-2,4-dienedioic acid hydrolase bphD 3.7.1.- 4.6 + 1.91 0.13 77371 RB13260 7108384 7109364 1.-.-.- 326 981 Zinc-containing alcohol dehydrogenase superfamily 1.-.-.- 4.6 + 1.34 0.17 132453 RB4419 2272912 2273448 178 537 MgtC/SapB transporter 6.2 + 2.11 0.15 72454 RB4420 2274093 2273473 206 621 DedA family protein 9.3 - 1.04 0.25 242495 RB4485 2298652 2299770 6.3.2.2 372 1119 Glutamate--cysteine ligase, GCS2 6.3.2.2 4.7 + 1.33 0.14 113490 RB6207 3252449 3253759 436 1311 conserved hypothetical protein, secreted 4.8 + 1.15 0.21 184648 RB8260 4415394 4415068 108 327 protein containing DUF167 10.5 - 1.57 0.17 116265 RB11258 6106840 6106127 237 714 conserved hypothetical protein, membrane 10.3 - 1.30 0.05 46422 RB11498 6218361 6219701 446 1341 conserved hypothetical protein 5.4 + 1.30 0.12 96720 RB12068 6512255 6510594 553 1662 serine/threonine protein kinase 8.5 - 2.41 0.14 66266 RB11260 6107202 6106837 121 366 dnaK suppressor protein, 5.2 - 1.08 0.17 16
4 RB5 1514 399 371 1116 secreted protein 4.1 - 3.14 0.32 10101 RB174 91448 91585 45 138 hypothetical protein 5.2 + 1.29 0.28 21300 RB520 284989 286116 375 1128 conserved hypothetical protein 4.8 + 1.29 0.35 27301 RB521 286399 286208 63 192 hypothetical protein 10.7 - 3.43 0.15 4328 RB568 312345 312112 77 234 hypothetical protein 10.2 - 1.28 0.28 22362 RB663 357608 357369 79 240 hypothetical protein 11.4 - 1.78 0.08 4365 RB670 359315 358905 136 411 hypothetical protein 10.1 - 2.40 0.19 8366 RB671 359218 359586 122 369 hypothetical protein 10.7 + 1.43 0.03 2507 RB934 494203 493076 375 1128 Putative transposase 9.9 - 1.28 0.03 2528 RB968 507840 507079 253 762 protein containing DUF1596 11.3 - 1.79 0.34 19556 RB1043 543083 543352 89 270 conserved hypothetical protein 4.7 + 1.07 0.08 7579 RB1093 567412 567152 86 261 hypothetical protein 9.8 - 1.13 0.13 11623 RB1180 604378 604656 92 279 hypothetical protein 12.6 + 1.23 0.28 23692 RB1294 657236 657051 61 186 hypothetical protein 9.8 - 1.10 0.13 12697 RB1302 659166 659762 198 597 hypothetical protein 8.8 + 1.44 0.09 6707 RB1322 671526 671633 35 108 hypothetical protein 12.8 + 1.32 0.03 2748 RB1395 712097 713056 319 960 secreted protein 5.5 + 1.47 0.17 12868 RB1617 834021 833623 132 399 hypothetical protein 6.8 - 1.58 0.21 14910 RB1716 895805 895557 82 249 protein containing DUF1589 10.1 - 1.33 0.23 17913 RB1722 897688 899643 651 1956 hypothetical protein 5.4 + 1.81 0.02 1914 RB1726 899926 899633 97 294 hypothetical protein 10.8 - 1.12 0.28 25915 RB1728 899954 900811 285 858 membrane protein containing DUF1206 9.9 + 1.74 0.35 20916 RB1729 901146 900784 120 363 hypothetical protein 7.7 - 3.11 0.24 8936 RB1767 919059 916336 907 2724 60-kDa cysteine-rich outer membrane protein precursor omcB 4.2 - 1.43 0.10 7941 RB1778 921245 921036 69 210 hypothetical protein 10.7 - 1.22 0.22 18

1151 RB2186 1153993 1152692 433 1302 ISXo8 transposase 9.4 - 1.46 0.09 61207 RB2283 1197719 1197102 205 618 protein containing DUF1590 10.5 - 1.08 0.25 231371 RB2525 1320284 1320054 76 231 hypothetical protein 6.1 - 1.11 0.28 251435 RB2649 1374987 1375148 53 162 hypothetical protein 9.1 + 1.55 0.19 121452 RB2679 1391557 1392078 173 522 hypothetical protein 5.5 + 1.63 0.16 101516 RB2806 1458677 1458802 41 126 hypothetical protein 6.5 + 1.24 0.19 151518 RB2809 1459745 1459065 226 681 hypothetical protein 9.1 - 1.13 0.05 51519 RB2810 1459901 1459752 49 150 hypothetical protein 11.3 - 1.03 0.10 101553 RB2875 1497313 1497534 73 222 hypothetical protein 9.9 + 1.43 0.08 61656 RB3052 1577799 1577692 35 108 hypothetical protein 11.2 - 1.51 0.15 101706 RB3153 1626911 1626681 76 231 hypothetical protein (protein containing DUF1584) 12.4 - 1.40 0.14 101707 RB3154 1627155 1626925 76 231 hypothetical protein 12.7 - 1.02 0.17 171722 RB3189 1646085 1646393 102 309 hypothetical protein 11.1 + 1.53 0.09 61728 RB3196 1650093 1650422 109 330 hypothetical protein 10.1 + 2.28 0.13 61791 RB3303 1695709 1694447 420 1263 conserved hypothetical protein 9.4 - 1.05 0.20 191911 RB3505 1816597 1816704 35 108 secreted protein containing DUF1586 10.6 + 1.50 0.08 61957 RB3587 1864796 1864455 113 342 hypothetical protein 11.8 - 1.84 0.23 132189 RB3963 2052742 2052870 42 129 hypothetical protein 4.1 + 1.14 0.28 252304 RB4176 2146928 2146638 96 291 hypothetical protein 12.4 - 1.28 0.25 202352 RB4258 2181475 2180966 169 510 conserved hypothetical protein 10.3 - 1.15 0.02 22407 RB4347 2235735 2236205 156 471 conserved hypothetical protein 4.7 + 1.22 0.10 82412 RB4353 2237771 2237331 146 441 hypothetical protein 4.1 - 1.08 0.07 62440 RB4397 2263976 2264143 55 168 protein containing DUF1560 10.5 + 2.56 0.21 82457 RB4425 2275100 2275483 127 384 hypothetical protein 10.6 + 2.20 0.45 202458 RB4426 2275786 2275604 60 183 hypothetical protein 6.3 - 1.31 0.20 152465 RB4438 2278247 2278729 160 483 Pyridoxamine 5'-phosphate oxidase- 4.4 + 1.69 0.09 52467 RB4443 2280143 2279973 56 171 hypothetical protein 10.1 - 1.45 0.17 122475 RB4457 2285628 2285924 98 297 hypothetical protein 10 + 1.59 0.37 232478 RB4460 2289696 2288428 422 1269 conserved hypothetical protein, membrane 4.8 - 1.03 0.23 222493 RB4482 2297240 2296905 111 336 hypothetical protein 12 - 1.87 0.12 72497 RB4489 2301387 2301905 172 519 hypothetical protein 5.4 + 1.33 0.15 122509 RB4510 2307657 2307508 49 150 hypothetical protein 9 - 1.37 0.36 262607 RB4676 2391079 2390414 221 666 hypothetical protein 10.4 - 1.37 0.04 32654 RB4750 2435462 2436076 204 615 membrane protein 10 + 1.09 0.27 242722 RB4859 2491989 2491429 186 561 RNA polymerase sigma factor 6.4 - 1.30 0.07 52816 RB5001 2572354 2572665 103 312 hypothetical protein 10.3 + 1.26 0.07 62819 RB5007 2574916 2575812 3.1.1.- 298 897 esterase 6 + 1.12 0.21 192951 RB5238 2697775 2697996 73 222 hypothetical protein 10.6 + 2.06 0.02 13326 RB5899 3092014 3091748 88 267 hypothetical protein 10.9 - 1.09 0.19 173345 RB5937 3112147 3113067 306 921 hypothetical protein 11.7 + 1.02 0.09 93346 RB5938 3113484 3112372 370 1113 hypothetical protein 5.6 - 1.77 0.27 153449 RB6131 3223482 3223790 102 309 hypothetical protein 12.1 + 1.64 0.25 153457 RB6144 3228305 3228189 38 117 hypothetical protein 11.1 - 1.83 0.29 163585 RB6375 3349681 3350574 297 894 hypothetical protein 8.7 + 1.61 0.11 73739 RB6627 3522169 3522699 176 531 hypothetical protein 6 + 1.21 0.11 93770 RB6684 3554034 3553717 105 318 hypothetical protein 11.6 - 1.27 0.07 63900 RB6910 3671484 3671356 42 129 hypothetical protein 9.5 - 2.47 0.07 33909 RB6925 3678461 3677976 161 486 hypothetical protein 9.8 - 1.28 0.02 23969 RB7039 3743573 3742965 202 609 hypothetical protein 7.9 - 2.01 0.47 234077 RB7221 3832041 3831646 131 396 hypothetical protein 10.4 - 1.07 0.16 154109 RB7279 3870019 3870309 96 291 hypothetical protein 6 + 1.02 0.21 214135 RB7318 3895264 3894884 126 381 hypothetical protein 7.3 - 1.04 0.17 164157 RB7389 3953590 3954717 375 1128 Putative transposase 9.9 + 1.18 0.22 194235 RB7523 4047925 4048074 49 150 hypothetical protein 12.4 + 2.73 0.03 14380 RB7752 4153019 4152765 84 255 hypothetical protein 9.8 - 1.20 0.27 224465 RB7901 4221613 4221293 106 321 hypothetical protein 11.8 - 1.55 0.13 84499 RB7965 4258303 4258464 53 162 hypothetical protein 7.4 + 1.20 0.05 44583 RB8132 4349972 4350505 177 534 hypothetical protein 10.8 + 1.81 0.20 114586 RB8138 4351960 4352187 75 228 hypothetical protein 4.1 + 1.73 0.38 224590 RB8146 4355122 4354667 151 456 hypothetical protein 4.3 - 1.71 0.12 74705 RB8367 4463905 4464057 50 153 hypothetical protein 3.7 + 1.44 0.26 184852 RB8641 4605970 4606248 92 279 hypothetical protein 7.5 + 2.61 0.55 214991 RB8898 4737468 4738433 321 966 acetyltransferase 4.5 + 1.65 0.27 164994 RB8902 4740474 4740226 82 249 hypothetical protein 11.2 - 1.10 0.18 165038 RB8987 4790856 4790710 48 147 hypothetical protein 9.1 - 1.13 0.15 135074 RB9070 4840267 4841496 409 1230 secreted protein containing DUF1559 8.6 + 2.40 0.08 35137 RB9191 4914516 4914653 45 138 secreted protein containing DUF1586 9.8 + 1.31 0.05 45217 RB9346 5008335 5008550 71 216 hypothetical protein 10.5 + 1.87 0.18 105347 RB9578 5150392 5150592 66 201 hypothetical protein 5.4 + 1.41 0.05 45377 RB9635 5187031 5185973 352 1059 hypothetical protein 5.9 - 1.34 0.36 275455 RB9786 5270645 5270205 146 441 hypothetical protein 12 - 1.25 0.19 155473 RB9811 5284365 5284673 102 309 secreted protein 11.7 + 2.79 0.37 135524 RB9907 5338185 5339486 433 1302 ISXo8 transposase 9.4 + 1.95 0.42 225552 RB9959 5366417 5366220 65 198 hypothetical protein 11.9 - 1.32 0.29 225723 RB10263 5546338 5546162 58 177 hypothetical protein 9.6 - 1.02 0.23 225753 RB10324 5580658 5580846 62 189 hypothetical protein 9 + 1.24 0.20 165764 RB10341 5593416 5593601 61 186 hypothetical protein 4.5 + 1.71 0.11 65766 RB10343 5593898 5594059 53 162 hypothetical protein 11.3 + 2.10 0.15 75815 RB10440 5655693 5655842 49 150 hypothetical protein 11.5 + 1.81 0.05 35834 RB10473 5679433 5681868 811 2436 conserved hypothetical protein 5.4 + 1.62 0.42 265876 RB10560 5731867 5729063 934 2805 hypothetical protein 4.6 - 1.09 0.09 85901 RB10607 5759926 5759537 129 390 hypothetical protein 11 - 1.36 0.08 65931 RB10665 5789912 5790514 200 603 hypothetical protein 5.3 + 2.55 0.29 116064 RB10893 5914894 5915067 57 174 hypothetical protein 9.7 + 1.43 0.30 216066 RB10895 5916348 5917151 267 804 hypothetical protein 10.5 + 2.03 0.11 66067 RB10896 5917040 5916555 161 486 secreted protein 10 - 1.33 0.03 26097 RB10954 5940749 5940318 143 432 hypothetical protein 10.4 - 1.15 0.04 36101 RB10960 5941602 5942021 139 420 conserved hypothetical protein, membrane 11.4 + 1.15 0.11 96124 RB11000 5964054 5963380 224 675 conserved hypothetical protein, membrane 8.8 - 2.06 0.03 16157 RB11070 6000321 6000479 52 159 hypothetical protein 10.2 + 1.28 0.07 66178 RB11110 6022600 6023529 309 930 Rhodopirellula transposase 5.8 + 1.30 0.10 76220 RB11181 6067002 6067109 35 108 secreted protein containing DUF1586 11.4 + 1.29 0.03 26236 RB11209 6080603 6080914 103 312 hypothetical protein 9.5 + 1.86 0.41 226237 RB11210 6080955 6080653 100 303 hypothetical protein 9.9 - 1.49 0.04 26269 RB11268 6109768 6109475 97 294 hypothetical protein 10.8 - 1.41 0.22 166321 RB11348 6155692 6155237 151 456 conserved hypothetical protein 3.4 - 1.02 0.03 36424 RB11504 6221285 6221067 72 219 conserved hypothetical protein 10.7 - 3.38 0.08 26430 RB11515 6224420 6224644 74 225 conserved hypothetical protein 11.7 + 1.79 0.18 106472 RB11603 6256722 6256835 37 114 conserved hypothetical protein 9 + 1.48 0.29 196571 RB11768 6334808 6334939 43 132 conserved hypothetical protein 9.4 + 1.50 0.33 226606 RB11855 6383765 6383460 101 306 conserved hypothetical protein 12.5 - 1.30 0.09 76673 RB11977 6455958 6456548 196 591 conserved hypothetical protein 9.5 + 1.49 0.14 106691 RB12009 6479495 6477951 514 1545 conserved hypothetical protein 4.8 - 1.17 0.15 126721 RB12071 6512255 6512866 203 612 conserved hypothetical protein 10.7 + 1.15 0.19 176760 RB12150 6556901 6557062 53 162 conserved hypothetical protein 4.1 + 1.83 0.10 56810 RB12236 6590935 6591546 203 612 conserved hypothetical protein, membrane 4.8 + 1.16 0.18 166812 RB12239 6593334 6594635 433 1302 ISXo8 transposase 9.4 + 2.57 0.02 16815 RB12243 6596256 6596423 55 168 conserved hypothetical protein 10.5 + 3.56 0.01 06817 RB12247 6597048 6596824 74 225 conserved hypothetical protein 6.3 - 2.33 0.32 146844 RB12295 6614190 6614450 86 261 conserved hypothetical protein 9.1 + 1.32 0.24 186861 RB12323 6627001 6627747 248 747 conserved hypothetical protein 4.4 + 1.32 0.03 26862 RB12324 6627901 6627713 62 189 conserved hypothetical protein 12.3 - 1.30 0.03 26898 RB12388 6667206 6665905 433 1302 ISXo8 transposase 9.4 - 1.92 0.18 96951 RB12473 6712524 6712706 60 183 conserved hypothetical protein 7.3 + 1.47 0.13 97092 RB12737 6863917 6864114 65 198 conserved hypothetical protein 9.1 + 2.02 0.07 37099 RB12747 6870758 6870919 53 162 conserved hypothetical protein 11.8 + 1.13 0.18 167113 RB12770 6884806 6884306 166 501 conserved hypothetical protein 11.9 - 2.28 0.32 147119 RB12782 6889986 6889819 55 168 conserved hypothetical protein 10.9 - 2.17 0.05 27120 RB12783 6889970 6891277 435 1308 conserved hypothetical protein 4.5 + 1.45 0.28 197130 RB12803 6909112 6908939 57 174 conserved hypothetical protein 10.2 - 1.12 0.14 127148 RB12829 6919813 6920292 159 480 conserved hypothetical protein 9.5 + 1.33 0.04 37151 RB12833 6921305 6921634 109 330 conserved hypothetical protein 12 + 1.14 0.07 67202 RB12936 6965872 6964130 580 1743 conserved hypothetical protein 5.3 - 1.32 0.18 14

7211 RB12959 6974205 6973882 107 324 conserved hypothetical protein 11.4 - 1.20 0.28 237251 RB13038 7005061 7002800 753 2262 conserved hypothetical protein, membrane 6.9 - 1.32 0.09 77256 RB13053 7008596 7009024 142 429 secreted protein 5.6 + 1.05 0.15 157270 RB13082 7020677 7020898 73 222 conserved hypothetical protein 11.7 + 1.17 0.26 227293 RB13115 7036614 7036369 81 246 conserved hypothetical protein 9.8 - 2.00 0.27 147340 RB13202 7086189 7086052 45 138 conserved hypothetical protein 9.4 - 1.44 0.07 57350 RB13221 7093018 7093986 322 969 secreted protein 4.5 + 1.56 0.13 87351 RB13222 7094714 7094088 208 627 SOUL heme-binding protein 8.7 - 1.72 0.28 167360 RB13238 7101410 7101904 164 495 TadE-like 5.3 + 3.52 0.12 37361 RB13239 7102571 7101981 196 591 conserved hypothetical protein, membrane 9.5 - 1.74 0.18 107370 RB13259 7108365 7106461 634 1905 conserved hypothetical protein 5.7 - 1.95 0.41 21
300min ID Locus AA Nuc Product Gene EC IEP Strand Ratio Stdev %1004 RB1902 988719 990776 1.6.99.3 685 2058 NADH oxidase 1.6.99.3 9.3 + 1.20 0.14 91007 RB1909 992934 993434 166 501 Globin 4.5 + 2.10 0.15 81961 RB3596 1868895 1869329 144 435 nitrogen fixation protein (NifU protein) 4.3 + 1.08 0.20 86831 RB12276 6605112 6605444 110 333 conserved hypothetical protein 4.7 + 2.88 0.18 207243 RB13025 6997019 6996438 193 582 monooxygenase 9.7 - 1.20 0.05 91212 RB2291 1199317 1200063 248 747 Sporulation initiation inhibitor protein soj soj 4.5 + 1.06 0.06 61546 RB2860 1487828 1489987 719 2160 type IV pilus assembly protein PilM 4.5 + 1.20 0.08 104149 RB7375 3948831 3947593 3.4.11.4 412 1239 Peptidase T pepT 3.4.11.4 4.4 - 1.18 0.23 185034 RB8979 4785013 4786671 4.3.1.17 552 1659 L-serine dehydratase, iron-sulfur-dependent, alpha subunit 4.3.1.17 4.4 + 1.46 0.06 125144 RB9200 4919220 4918369 3.1.3.41 283 852 4-nitrophenylphosphatase PHO13 3.1.3.41 4.9 - 1.58 0.23 97094 RB12740 6865551 6864364 3.1.1.17 395 1188 gluconolactonase [precursor] gnl 3.1.1.17 4.8 - 1.12 0.15 55878 RB10566 5734146 5732695 483 1452 Gluconate transporter 9.2 - 1.60 0.10 84579 RB8123 4346827 4347360 4.2.1.- 177 534 (3R)-hydroxymyristoyl-[acyl carrier protein] dehydratase fabZ 4.2.1.- 4.9 + 1.05 0.13 19948 RB1790 926593 927168 191 576 RNA polymerase ECF-type sigma factor 10.4 + 1.48 0.13 8
4744 RB8445 4514475 4514050 141 426 Penicillinase repressor 9.9 - 1.95 0.23 25571 RB9999 5386828 5385983 281 846 conserved hypothetical protein 4.6 - 1.35 0.13 105606 RB10049 5419507 5420160 217 654 RNA polymerase ECF-type sigma factor 10.1 + 1.27 0.12 16160 RB11075 6005727 6005224 167 504 Bacterial regulatory protein, MarR 6.4 - 1.03 0.16 46161 RB11077 6006371 6005904 155 468 Protein of unknown function, UPF0074 8.8 - 1.99 0.08 94187 RB7437 3987953 3988672 3.4.21.88 239 720 LexA repressor lexA 3.4.21.88 9.8 + 1.73 0.15 20
98 RB170 89333 89623 96 291 Transposase IS3/IS911 10.1 + 1.34 0.21 1599 RB171 89626 90504 292 879 ISxac3 transposase 10 + 1.29 0.14 11
382 RB704 378137 378427 96 291 Transposase IS3/IS911 10.1 + 1.83 0.08 8383 RB706 378430 379308 292 879 ISxac3 transposase 10 + 1.64 0.13 10628 RB1189 606838 607128 96 291 Transposase IS3/IS911 10.1 + 1.81 0.07 15629 RB1190 607131 608009 292 879 integrase 10 + 1.74 0.26 7
2213 RB3999 2064741 2063431 436 1311 site-specific recombinase, 10.4 - 1.21 0.08 12381 RB4299 2214533 2214823 96 291 Transposase IS3/IS911 9.9 + 1.69 0.18 92382 RB4300 2214826 2215704 292 879 integrase 10 + 1.56 0.14 93319 RB5887 3087343 3086465 292 879 integrase 9.9 - 1.58 0.13 83420 RB6070 3186811 3187416 201 606 HNH endonuclease family protein 9.9 + 1.63 0.13 163474 RB6171 3239091 3238048 347 1044 site-specific recombinase 10.7 - 1.47 0.14 43787 RB6709 3564146 3563856 96 291 Transposase IS3/IS911 10.1 - 1.71 0.08 96558 RB11749 6328064 6328354 96 291 Transposase IS3/IS911 10.1 + 1.64 0.24 36559 RB11750 6328357 6329235 292 879 integrase 10 + 2.02 0.06 26577 RB11802 6361950 6362240 96 291 Transposase IS3/IS911 10.1 + 1.10 0.08 86578 RB11803 6362243 6363121 292 879 ISxac3 transposase 10 + 1.27 0.10 196789 RB12202 6579810 6578185 541 1626 integrase-recombinase protein 9.9 - 1.39 0.29 67203 RB12940 6966246 6966536 96 291 Transposase IS3/IS911 10.1 + 1.81 0.17 117204 RB12941 6966539 6967417 292 879 integrase 10 + 1.15 0.13 221355 RB2507 1313064 1311886 4.2.1.46 392 1179 dTDP-glucose 4,6-dehydratase rfbB 4.2.1.46 5.2 - 1.01 0.13 81727 RB3195 1650068 1648239 609 1830 carboxy-terminal processing protease 6.1 - 1.31 0.18 55719 RB10255 5542656 5543669 337 1014 MscS Mechanosensitive ion channel 5.4 + 1.56 0.10 97070 RB12690 6834088 6833336 2.7.7.38 250 753 3-deoxy-manno-octulosonate cytidylyltransferase kdsB 2.7.7.38 4.7 - 1.54 0.13 146711 RB12053 6502558 6502199 119 360 biopolymer transport protein, ExbD/TolR family 4.2 - 1.08 0.18 66712 RB12055 6503587 6502628 319 960 MotA/TolQ/ExbB proton channel family protein 5.3 - 1.11 0.07 237115 RB12773 6884826 6885932 368 1107 twitching motility protein PilT 6.3 + 1.17 0.20 67118 RB12781 6888383 6889822 479 1440 pilus assembly protein PilC 5.6 + 1.21 0.08 5731 RB1359 693488 695377 3.4.21.- 629 1890 Protease do precursor degP 3.4.21.- 6.1 + 1.16 0.12 11
3529 RB6278 3302303 3297678 1541 4626 Putative Ig 3.6 - 2.20 0.08 43812 RB6751 3586111 3588690 859 2580 chaperone ClpB clpB 5.2 + 1.21 0.10 136727 RB12080 6517231 6515753 2.1.1.77 492 1479 Protein-L-isoaspartate(D-aspartate) O-methyltransferase 2.1.1.77 8.9 - 1.09 0.25 5212 RB370 184428 186089 553 1662 nitrate transporter substrate-binding protein 4.6 + 2.96 0.11 4214 RB373 187285 188283 332 999 ABC transporter, ATP-binding protein 9.9 + 1.50 0.09 6220 RB380 196136 197740 1.8.1.2 534 1605 sulfite reductase [NADPH] flavoprotein cysJ 1.8.1.2 4.4 + 1.54 0.04 3766 RB1433 732848 732411 145 438 Na(+):H(+) antiporter subunit B mrpB 9.2 - 1.03 0.18 5791 RB1477 755327 756943 3.1.6.8 538 1617 arylsulfatase precursor ARSA 3.1.6.8 5.5 + 1.62 0.09 14
2817 RB5003 2572662 2574158 498 1497 Rh-like protein/ammonium transporter AMT1.4 4 + 1.92 0.18 02984 RB5294 2732584 2730983 533 1602 sulfatase 5.6 - 1.20 0.16 95967 RB10727 5830775 5831605 276 831 manganese-containing catalase 5 + 1.41 0.24 136128 RB11009 5969752 5969036 238 717 sulfite reductase-like flavoprotein 3.9 - 1.13 0.06 116675 RB11981 6459460 6460413 317 954 ABC transporter (substrate-binding protein) 4.8 + 1.41 0.26 37165 RB12858 6929710 6928649 353 1062 oligopeptide transport ATP-binding protein OppF oppF 7 - 1.14 0.12 227169 RB12862 6934893 6932926 655 1968 Oligopeptide-binding protein oppA oppA 4.5 - 1.10 0.25 84332 RB7678 4123339 4122659 226 681 protocatechuate 3,4-dioxygenase type II beta subunit 9.4 - 1.25 0.11 101347 RB2498 1305777 1304887 296 891 methyltransferase, 6.2 - 1.10 0.12 13557 RB1044 543422 544231 269 810 TatD-related deoxyribonuclease 5.2 + 1.23 0.19 12835 RB1555 797278 795848 476 1431 dehydrogenases and related proteins 5.8 - 1.02 0.03 4
2896 RB5140 2641937 2643778 613 1842 hypothetical protein 5.5 + 1.29 0.21 144603 RB8173 4366881 4367900 339 1020 MoxR protein 5.1 + 1.51 0.09 54796 RB8533 4552172 4552762 3.2.-.- 196 591 protease I pfpI 3.2.-.- 4.8 + 1.63 0.24 65109 RB9132 4886727 4888004 425 1278 conserved hypothetical protein 5.5 + 1.40 0.08 35568 RB9992 5383545 5382085 486 1461 SufBD 5.9 - 1.07 0.25 105721 RB10258 5544584 5545453 3.1.3.2 289 870 Survival protein SurE 3.1.3.2 5.3 + 1.22 0.11 35815 RB10440 5655693 5655842 49 150 hypothetical protein 11.5 + 1.31 0.05 86215 RB11173 6063194 6063586 130 393 conserved hypothetical protein, secreted 4.7 + 1.56 0.18 46216 RB11174 6065046 6063637 469 1410 metallo-beta-lactamase family protein 4.8 - 1.08 0.05 106641 RB11923 6419736 6419041 231 696 Electron transport protein SCO1/SenC 4.3 - 1.30 0.14 26667 RB11964 6444840 6445160 106 321 conserved hypothetical protein 3.9 + 1.04 0.20 187369 RB13257 7105590 7106450 3.7.1.- 286 861 2-hydroxy-6-ketonona-2,4-dienedioic acid hydrolase bphD 3.7.1.- 4.6 + 1.03 0.07 7324 RB560 309384 308542 280 843 protein containing DGPFAETKE domain DGPF 4.9 - 1.34 0.11 8
2453 RB4419 2272912 2273448 178 537 MgtC/SapB transporter 6.2 + 1.09 0.10 74648 RB8260 4415394 4415068 108 327 protein containing DUF167 10.5 - 1.31 0.06 175413 RB9699 5226252 5228345 697 2094 Rhs element Vgr protein 4.2 + 1.66 0.03 196217 RB11176 6065605 6065144 153 462 protein containing DUF442 4.8 - 1.63 0.16 15
80 RB140 61578 64541 2.7.1.37 987 2964 serine/threonine protein kinase pkn3 2.7.1.37 5.2 + 1.09 0.24 223259 RB5780 3028975 3029472 165 498 two-component response regulator 6.2 + 2.09 0.23 84202 RB7461 3999634 4001478 614 1845 hypothetical protein 4.4 + 1.27 0.25 65788 RB10388 5613357 5615438 2.7.1.37 693 2082 protein kinase KIN11 2.7.1.37 9 + 1.19 0.10 76679 RB11986 6465612 6462064 1182 3549 two-component sensor/response regulator hybrid 6.4 - 1.04 0.03 187209 RB12952 6972002 6972667 221 666 two-component system, regulatory protein 4.7 + 1.53 0.32 27291 RB13112 7036128 7034035 697 2094 Hybrid sensor histidine kinase 5.5 - 1.31 0.11 7
4 RB5 1514 399 371 1116 secreted protein 4.1 - 2.35 0.29 1250 RB85 31507 32739 410 1233 conserved hypothetical protein, secreted 5.1 + 1.10 0.10 979 RB138 60872 61516 214 645 Sigma factor, ECF-like 4.8 + 1.93 0.14 790 RB160 84284 84027 85 258 hypothetical protein 10.7 - 1.62 0.22 14
101 RB174 91448 91585 45 138 hypothetical protein 5.2 + 1.64 0.17 10152 RB275 135819 135370 149 450 hypothetical protein 9.3 - 1.16 0.09 8300 RB520 284989 286116 375 1128 conserved hypothetical protein 4.8 + 1.02 0.12 12301 RB521 286399 286208 63 192 hypothetical protein 10.7 - 2.06 0.11 5392 RB723 388051 388233 60 183 hypothetical protein 7.3 + 2.37 0.24 9469 RB864 458119 458598 159 480 hypothetical protein 6.1 + 1.42 0.13 6470 RB865 458731 460359 542 1629 hypothetical protein 3.1 + 1.69 0.11 17471 RB867 460731 460429 100 303 hypothetical protein 9.7 - 1.47 0.25 4528 RB968 507840 507079 253 762 protein containing DUF1596 11.3 - 1.25 0.05 5530 RB971 509411 508101 436 1311 Patatin-like phospholipase 10.4 - 1.06 0.06 14556 RB1043 543083 543352 89 270 conserved hypothetical protein 4.7 + 1.41 0.20 15599 RB1138 585492 585959 155 468 conserved hypothetical protein 10.5 + 1.54 0.19 9619 RB1174 603031 603498 155 468 secreted protein 9.8 + 1.88 0.17 1

622 RB1179 603998 604384 128 387 hypothetical protein 12 + 2.03 0.01 7623 RB1180 604378 604656 92 279 hypothetical protein 12.6 + 1.69 0.12 4638 RB1201 612754 612629 41 126 hypothetical protein 12.5 - 1.31 0.10 12644 RB1210 617416 616571 281 846 AP endonuclease 2, C-terminal 4.5 - 2.51 0.31 3662 RB1239 632500 632033 155 468 hypothetical protein 9.7 - 1.43 0.05 15668 RB1254 640610 640747 45 138 hypothetical protein 10.6 + 2.44 0.36 11692 RB1294 657236 657051 61 186 hypothetical protein 9.8 - 1.75 0.19 5707 RB1322 671526 671633 35 108 hypothetical protein 12.8 + 1.49 0.07 5725 RB1352 686463 687803 446 1341 hypothetical protein 5.5 + 1.04 0.05 10747 RB1394 711994 711758 78 237 hypothetical protein 10.1 - 1.21 0.13 6748 RB1395 712097 713056 319 960 secreted protein 5.5 + 1.30 0.08 18792 RB1478 756943 758943 666 2001 hypothetical protein 5.3 + 1.88 0.26 6818 RB1522 777494 778657 387 1164 protein containing DUF1559 5.6 + 1.69 0.10 10819 RB1523 778654 779130 158 477 hypothetical protein 7.3 + 1.81 0.18 16823 RB1531 782534 783889 451 1356 membrane protein 4.5 + 1.29 0.21 3836 RB1557 797241 797420 59 180 hypothetical protein 11.8 + 1.60 0.07 5868 RB1617 834021 833623 132 399 hypothetical protein 6.8 - 1.13 0.05 9910 RB1716 895805 895557 82 249 protein containing DUF1589 10.1 - 1.28 0.12 23916 RB1729 901146 900784 120 363 hypothetical protein 7.7 - 1.19 0.28 12927 RB1750 909661 909837 58 177 hypothetical protein 10.1 + 1.25 0.15 13936 RB1767 919059 916336 907 2724 60-kDa cysteine-rich outer membrane protein precursor omcB 4.2 - 1.49 0.19 2937 RB1771 919524 919273 83 252 hypothetical protein 7.6 - 1.39 0.03 13947 RB1789 925861 926592 243 732 conserved hypothetical protein 9.4 + 1.10 0.14 9985 RB1872 966053 966169 38 117 hypothetical protein 12.3 + 1.96 0.15 12
1006 RB1908 992277 992900 207 624 secreted protein 4.6 + 1.42 0.13 71008 RB1910 993764 993393 123 372 hypothetical protein 11.2 - 1.93 0.15 161142 RB2173 1147959 1146895 354 1065 hypothetical protein 6.3 - 2.25 0.37 101185 RB2252 1181701 1181540 53 162 hypothetical protein 9.8 - 1.67 0.17 61207 RB2283 1197719 1197102 205 618 protein containing DUF1590 10.5 - 2.03 0.12 61223 RB2310 1209545 1209222 107 324 hypothetical protein 11.5 - 1.23 0.07 171313 RB2457 1279788 1280144 118 357 hypothetical protein 12.7 + 1.15 0.20 181346 RB2497 1304778 1304906 42 129 hypothetical protein 6.7 + 1.19 0.22 111365 RB2517 1316854 1317012 52 159 hypothetical protein 10.3 + 1.87 0.15 31370 RB2524 1319449 1319796 115 348 hypothetical protein 7.4 + 1.28 0.04 161380 RB2545 1328931 1328773 52 159 hypothetical protein 11.2 - 1.86 0.30 91435 RB2649 1374987 1375148 53 162 hypothetical protein 9.1 + 1.39 0.12 161461 RB2697 1402745 1404271 508 1527 protein containing DUF1501 6.9 + 1.32 0.22 221506 RB2784 1448065 1447454 203 612 membrane protein containing DUF107 4.5 - 1.23 0.28 141515 RB2804 1458645 1458040 201 606 hypothetical protein 5.3 - 1.38 0.19 121518 RB2809 1459745 1459065 226 681 hypothetical protein 9.1 - 1.06 0.13 81524 RB2822 1463434 1464933 499 1500 secreted protein containing DUF1598 6.9 + 1.52 0.13 81528 RB2829 1467448 1467564 38 117 hypothetical protein 4.1 + 1.17 0.09 71549 RB2867 1491909 1494002 697 2094 hypothetical protein 4.7 + 1.91 0.18 61645 RB3034 1572089 1569573 838 2517 secreted protein containing DUF1585 5.7 - 1.11 0.06 91646 RB3036 1573582 1572092 496 1491 protein containing DUF1552 9.1 - 1.52 0.14 51656 RB3052 1577799 1577692 35 108 hypothetical protein 11.2 - 1.57 0.08 91695 RB3140 1620838 1621173 111 336 hypothetical protein 10.2 + 1.06 0.09 101705 RB3152 1626667 1626461 68 207 hypothetical protein 12.2 - 1.21 0.12 41706 RB3153 1626911 1626681 76 231 hypothetical protein (protein containing DUF1584) 12.4 - 2.20 0.09 51707 RB3154 1627155 1626925 76 231 hypothetical protein 12.7 - 1.93 0.10 61708 RB3155 1627430 1627149 93 282 hypothetical protein 12.7 - 2.69 0.16 101711 RB3159 1629337 1629116 73 222 protein containing DUF1590 12.1 - 1.66 0.17 41722 RB3189 1646085 1646393 102 309 hypothetical protein 11.1 + 1.49 0.06 131728 RB3196 1650093 1650422 109 330 hypothetical protein 10.1 + 1.98 0.10 191788 RB3299 1693917 1692925 330 993 protein containing DUF1559 6.3 - 1.68 0.31 71809 RB3333 1713409 1713528 39 120 hypothetical protein 12.2 + 1.04 0.07 151827 RB3365 1730124 1730255 43 132 hypothetical protein 11.4 + 1.03 0.16 61853 RB3406 1759455 1759589 44 135 hypothetical protein 10.5 + 1.01 0.06 61865 RB3426 1771404 1771622 72 219 hypothetical protein 12.4 + 1.86 0.11 171868 RB3430 1772646 1773392 248 747 conserved hypothetical protein, membrane 7.5 + 1.03 0.18 182005 RB3659 1900428 1900607 59 180 hypothetical protein 10.7 + 3.01 0.25 72272 RB4114 2117690 2117971 93 282 protein containing DUF1582 11.1 + 1.76 0.01 62308 RB4185 2151951 2152220 89 270 hypothetical protein 12 + 1.75 0.11 112388 RB4306 2221207 2219834 457 1374 transposase and inactivated derivative 8.4 - 1.20 0.11 52395 RB4320 2226114 2226611 165 498 hypothetical protein 12 + 1.06 0.05 22440 RB4397 2263976 2264143 55 168 protein containing DUF1560 10.5 + 1.92 0.04 92584 RB4642 2377301 2377558 85 258 conserved hypothetical protein 4.4 + 1.37 0.10 132597 RB4658 2382780 2382199 193 582 inosine monophosphate dehydrogenase-related protein 4.8 - 2.03 0.26 112615 RB4694 2396117 2397352 411 1236 protein containing DUF1559 6.5 + 1.81 0.20 112616 RB4697 2397357 2397539 60 183 secreted protein 3.1 + 1.96 0.22 182629 RB4715 2413294 2417169 1291 3876 conserved hypothetical protein 5.3 + 1.09 0.19 62654 RB4750 2435462 2436076 204 615 membrane protein 10 + 1.06 0.07 232694 RB4810 2465774 2465418 118 357 hypothetical protein 10.8 - 1.05 0.24 72783 RB4957 2548380 2548553 57 174 hypothetical protein 12.4 + 1.07 0.07 62792 RB4969 2554443 2555447 334 1005 hypothetical protein 10 + 1.65 0.10 22816 RB5001 2572354 2572665 103 312 hypothetical protein 10.3 + 2.62 0.04 92819 RB5007 2574916 2575812 3.1.1.- 298 897 esterase 6 + 1.59 0.00 102894 RB5138 2641024 2640848 58 177 hypothetical protein 12.4 - 2.31 0.22 163009 RB5347 2764290 2764442 50 153 hypothetical protein 11.9 + 1.32 0.12 133010 RB5348 2765787 2764414 457 1374 transposase and inactivated derivative 8.4 - 1.45 0.18 63022 RB5370 2774826 2773897 309 930 Rhodopirellula transposase 5.9 - 2.11 0.12 213117 RB5522 2860696 2860331 121 366 hypothetical protein 10.3 - 1.35 0.28 113130 RB5551 2889101 2891092 663 1992 hypothetical protein 5.7 + 2.16 0.23 193139 RB5567 2897766 2897566 66 201 hypothetical protein 12.1 - 1.55 0.29 43142 RB5572 2901618 2901499 39 120 hypothetical protein 13 - 2.17 0.09 83233 RB5737 2994415 2994777 120 363 hypothetical protein 11.9 + 1.12 0.09 113320 RB5888 3087636 3087346 96 291 Transposase IS3/IS911 10.1 - 2.02 0.16 163345 RB5937 3112147 3113067 306 921 hypothetical protein 11.7 + 1.10 0.18 153346 RB5938 3113484 3112372 370 1113 hypothetical protein 5.6 - 1.28 0.19 83426 RB6081 3191035 3190877 52 159 hypothetical protein 11.5 - 1.49 0.23 63449 RB6131 3223482 3223790 102 309 hypothetical protein 12.1 + 1.57 0.10 113457 RB6144 3228305 3228189 38 117 hypothetical protein 11.1 - 1.78 0.20 103482 RB6188 3244384 3244178 68 207 hypothetical protein 9.6 - 2.22 0.09 73498 RB6221 3264012 3260995 1005 3018 conserved hypothetical protein, secreted 4.8 - 1.07 0.08 43585 RB6375 3349681 3350574 297 894 hypothetical protein 8.7 + 1.68 0.07 193591 RB6382 3355162 3355043 39 120 hypothetical protein 10.4 - 1.03 0.19 23640 RB6457 3407990 3407787 67 204 hypothetical protein 11.7 - 1.00 0.02 183668 RB6497 3442109 3442804 231 696 conserved hypothetical protein 3.9 + 1.01 0.18 183756 RB6661 3538752 3538486 88 267 hypothetical protein 10.6 - 1.07 0.19 53893 RB6903 3669220 3669059 53 162 hypothetical protein 9.3 - 1.42 0.18 43894 RB6904 3669200 3670501 433 1302 secreted protein containing DUF1552 7.1 + 1.68 0.07 33900 RB6910 3671484 3671356 42 129 hypothetical protein 9.5 - 3.33 0.09 73902 RB6912 3672793 3672960 55 168 secreted protein 9 + 2.38 0.16 33907 RB6920 3676847 3677914 355 1068 protein containing DUF1559 6.5 + 1.92 0.05 173909 RB6925 3678461 3677976 161 486 hypothetical protein 9.8 - 2.17 0.38 83910 RB6928 3678479 3678961 160 483 hypothetical protein 4.1 + 2.64 0.20 153941 RB6990 3719456 3719166 96 291 hypothetical protein 11.9 - 1.62 0.24 33969 RB7039 3743573 3742965 202 609 hypothetical protein 7.9 - 1.18 0.04 194151 RB7381 3950326 3949595 243 732 membrane protein 7.4 - 1.00 0.18 134158 RB7390 3954342 3954968 208 627 hypothetical protein 12.2 + 1.77 0.23 54162 RB7395 3955956 3956351 131 396 hypothetical protein 11.9 + 1.12 0.05 94203 RB7464 4001614 4001480 44 135 hypothetical protein 10.2 - 1.14 0.07 54288 RB7603 4091070 4088560 836 2511 outer membrane efflux protein 5.1 - 1.41 0.07 124291 RB7612 4092907 4093317 136 411 hypothetical protein 11.4 + 1.16 0.14 144318 RB7651 4112517 4112383 44 135 hypothetical protein 6.7 - 1.43 0.20 94398 RB7786 4172318 4171854 154 465 hypothetical protein 10.5 - 1.47 0.14 84465 RB7901 4221613 4221293 106 321 hypothetical protein 11.8 - 1.79 0.14 174497 RB7960 4257723 4256632 2.7.3.- 363 1092 ATP:guanido phosphotransferase 2.7.3.- 5.4 - 1.12 0.19 134586 RB8138 4351960 4352187 75 228 hypothetical protein 4.1 + 1.26 0.24 24590 RB8146 4355122 4354667 151 456 hypothetical protein 4.3 - 1.45 0.03 64705 RB8367 4463905 4464057 50 153 hypothetical protein 3.7 + 1.14 0.19 64715 RB8385 4479935 4480087 50 153 hypothetical protein 10.1 + 2.24 0.14 114727 RB8409 4495547 4495840 97 294 hypothetical protein 8.9 + 1.06 0.12 12
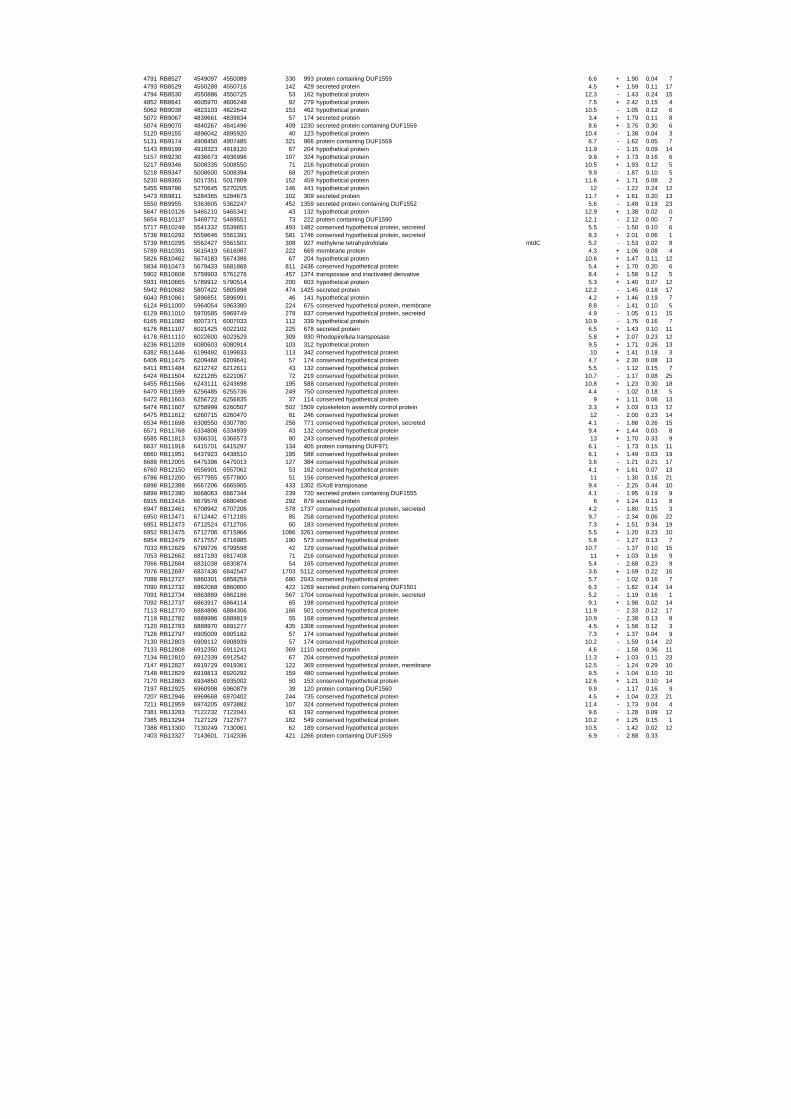
4791 RB8527 4549097 4550089 330 993 protein containing DUF1559 6.6 + 1.90 0.04 74793 RB8529 4550288 4550716 142 429 secreted protein 4.5 + 1.59 0.11 174794 RB8530 4550886 4550725 53 162 hypothetical protein 12.3 - 1.43 0.24 154852 RB8641 4605970 4606248 92 279 hypothetical protein 7.5 + 2.42 0.15 45062 RB9038 4823103 4822642 153 462 hypothetical protein 10.5 - 1.05 0.12 65072 RB9067 4839661 4839834 57 174 secreted protein 3.4 + 1.79 0.11 85074 RB9070 4840267 4841496 409 1230 secreted protein containing DUF1559 8.6 + 3.75 0.30 65120 RB9155 4896042 4895920 40 123 hypothetical protein 10.4 - 1.38 0.04 35131 RB9174 4908450 4907485 321 966 protein containing DUF1559 6.7 - 1.62 0.05 75143 RB9199 4918323 4918120 67 204 hypothetical protein 11.9 - 1.15 0.09 145157 RB9230 4936673 4936996 107 324 hypothetical protein 9.9 + 1.73 0.16 65217 RB9346 5008335 5008550 71 216 hypothetical protein 10.5 + 1.93 0.12 55218 RB9347 5008600 5008394 68 207 hypothetical protein 9.9 - 1.87 0.10 55230 RB9365 5017351 5017809 152 459 hypothetical protein 11.6 + 1.71 0.08 25455 RB9786 5270645 5270205 146 441 hypothetical protein 12 - 1.22 0.24 125473 RB9811 5284365 5284673 102 309 secreted protein 11.7 + 1.61 0.20 135550 RB9955 5363605 5362247 452 1359 secreted protein containing DUF1552 5.6 - 1.48 0.19 235647 RB10126 5465210 5465341 43 132 hypothetical protein 12.9 + 1.38 0.02 05654 RB10137 5469772 5469551 73 222 protein containing DUF1590 12.1 - 2.12 0.00 75717 RB10249 5541332 5539851 493 1482 conserved hypothetical protein, secreted 5.5 - 1.50 0.10 65738 RB10292 5559646 5561391 581 1746 conserved hypothetical protein, secreted 8.3 + 2.01 0.06 15739 RB10295 5562427 5561501 308 927 methylene tetrahydrofolate mtdC 5.2 - 1.53 0.02 85789 RB10391 5615419 5616087 222 669 membrane protein 4.3 + 1.06 0.08 45826 RB10462 5674183 5674386 67 204 hypothetical protein 10.6 + 1.47 0.11 125834 RB10473 5679433 5681868 811 2436 conserved hypothetical protein 5.4 + 1.70 0.20 65902 RB10608 5759903 5761276 457 1374 transposase and inactivated derivative 8.4 + 1.58 0.12 55931 RB10665 5789912 5790514 200 603 hypothetical protein 5.3 + 1.40 0.07 125942 RB10682 5807422 5805998 474 1425 secreted protein 12.2 - 1.45 0.18 176043 RB10861 5896851 5896991 46 141 hypothetical protein 4.2 + 1.46 0.19 76124 RB11000 5964054 5963380 224 675 conserved hypothetical protein, membrane 8.8 - 1.41 0.10 56129 RB11010 5970585 5969749 278 837 conserved hypothetical protein, secreted 4.9 - 1.05 0.11 156165 RB11082 6007371 6007033 112 339 hypothetical protein 10.9 - 1.75 0.16 76176 RB11107 6021425 6022102 225 678 secreted protein 6.5 + 1.43 0.10 116178 RB11110 6022600 6023529 309 930 Rhodopirellula transposase 5.8 + 2.07 0.23 126236 RB11209 6080603 6080914 103 312 hypothetical protein 9.5 + 1.71 0.26 136382 RB11446 6199492 6199833 113 342 conserved hypothetical protein 10 + 1.41 0.18 36406 RB11475 6209468 6209641 57 174 conserved hypothetical protein 4.7 + 2.30 0.08 136411 RB11484 6212742 6212611 43 132 conserved hypothetical protein 5.5 - 1.12 0.15 76424 RB11504 6221285 6221067 72 219 conserved hypothetical protein 10.7 - 1.17 0.08 256455 RB11566 6243111 6243698 195 588 conserved hypothetical protein 10.8 + 1.23 0.30 186470 RB11599 6256485 6255736 249 750 conserved hypothetical protein 4.4 - 1.02 0.18 56472 RB11603 6256722 6256835 37 114 conserved hypothetical protein 9 + 1.11 0.06 136474 RB11607 6258999 6260507 502 1509 cytoskeleton assembly control protein 3.3 + 1.03 0.13 126475 RB11612 6260715 6260470 81 246 conserved hypothetical protein 12 - 2.00 0.23 146534 RB11698 6308550 6307780 256 771 conserved hypothetical protein, secreted 4.1 - 1.88 0.26 156571 RB11768 6334808 6334939 43 132 conserved hypothetical protein 9.4 + 1.44 0.03 86585 RB11813 6366331 6366573 80 243 conserved hypothetical protein 13 + 1.70 0.33 96637 RB11918 6415701 6415297 134 405 protein containing DUF971 6.1 - 1.73 0.15 116660 RB11951 6437923 6438510 195 588 conserved hypothetical protein 6.1 + 1.49 0.03 196688 RB12005 6475396 6475013 127 384 conserved hypothetical protein 3.6 - 1.21 0.21 176760 RB12150 6556901 6557062 53 162 conserved hypothetical protein 4.1 + 1.61 0.07 136786 RB12200 6577955 6577800 51 156 conserved hypothetical protein 11 - 1.30 0.16 216898 RB12388 6667206 6665905 433 1302 ISXo8 transposase 9.4 - 2.25 0.44 106899 RB12390 6668063 6667344 239 720 secreted protein containing DUF1555 4.1 - 1.95 0.19 96915 RB12416 6679578 6680456 292 879 secreted protein 6 + 1.24 0.11 86947 RB12461 6708942 6707206 578 1737 conserved hypothetical protein, secreted 4.2 - 1.80 0.15 36950 RB12471 6712442 6712185 85 258 conserved hypothetical protein 9.7 - 2.34 0.06 226951 RB12473 6712524 6712706 60 183 conserved hypothetical protein 7.3 + 1.51 0.34 196952 RB12475 6712706 6715966 1086 3261 conserved hypothetical protein 5.5 + 1.20 0.23 106954 RB12479 6717557 6716985 190 573 conserved hypothetical protein 5.8 - 1.27 0.13 77033 RB12629 6799726 6799598 42 129 conserved hypothetical protein 10.7 - 1.37 0.10 157053 RB12662 6817193 6817408 71 216 conserved hypothetical protein 11 + 1.03 0.16 97066 RB12684 6831038 6830874 54 165 conserved hypothetical protein 5.4 - 2.68 0.23 97076 RB12697 6837436 6842547 1703 5112 conserved hypothetical protein 3.6 + 1.59 0.22 167088 RB12727 6860301 6858259 680 2043 conserved hypothetical protein 5.7 - 1.02 0.16 77090 RB12732 6862068 6860800 422 1269 secreted protein containing DUF1501 6.3 - 1.82 0.14 147091 RB12734 6863889 6862186 567 1704 conserved hypothetical protein, secreted 5.2 - 1.19 0.16 17092 RB12737 6863917 6864114 65 198 conserved hypothetical protein 9.1 + 1.98 0.02 147113 RB12770 6884806 6884306 166 501 conserved hypothetical protein 11.9 - 2.33 0.12 177119 RB12782 6889986 6889819 55 168 conserved hypothetical protein 10.9 - 2.38 0.13 87120 RB12783 6889970 6891277 435 1308 conserved hypothetical protein 4.5 + 1.58 0.12 37126 RB12797 6905009 6905182 57 174 conserved hypothetical protein 7.3 + 1.37 0.04 97130 RB12803 6909112 6908939 57 174 conserved hypothetical protein 10.2 - 1.59 0.14 227133 RB12808 6912350 6911241 369 1110 secreted protein 4.6 - 1.58 0.36 117134 RB12810 6912339 6912542 67 204 conserved hypothetical protein 11.3 + 1.03 0.11 237147 RB12827 6919729 6919361 122 369 conserved hypothetical protein, membrane 12.5 - 1.24 0.29 107148 RB12829 6919813 6920292 159 480 conserved hypothetical protein 9.5 + 1.04 0.10 107170 RB12863 6934850 6935002 50 153 conserved hypothetical protein 12.6 + 1.21 0.10 147197 RB12925 6960998 6960879 39 120 protein containing DUF1560 9.9 - 1.17 0.16 97207 RB12946 6969668 6970402 244 735 conserved hypothetical protein 4.5 + 1.04 0.23 217211 RB12959 6974205 6973882 107 324 conserved hypothetical protein 11.4 - 1.73 0.04 47381 RB13283 7122232 7122041 63 192 conserved hypothetical protein 9.6 - 1.28 0.09 127385 RB13294 7127129 7127677 182 549 conserved hypothetical protein 10.2 + 1.25 0.15 17388 RB13300 7130249 7130061 62 189 conserved hypothetical protein 10.5 - 1.42 0.02 127403 RB13327 7143601 7142336 421 1266 protein containing DUF1559 6.9 - 2.88 0.33
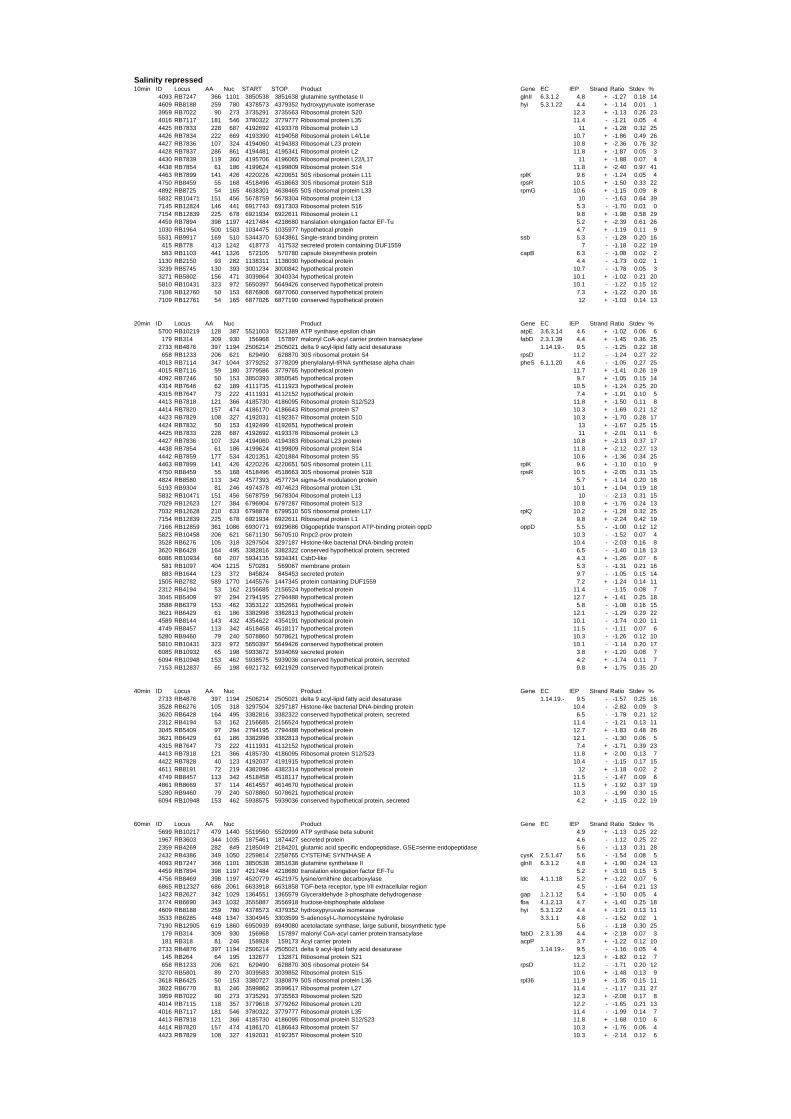
Salinity repressed10min ID Locus AA Nuc START STOP Product Gene EC IEP Strand Ratio Stdev %
4093 RB7247 366 1101 3850538 3851638 glutamine synthetase II glnII 6.3.1.2 4.8 + -1.27 0.18 144609 RB8188 259 780 4378573 4379352 hydroxypyruvate isomerase hyi 5.3.1.22 4.4 + -1.14 0.01 13959 RB7022 90 273 3735291 3735563 Ribosomal protein S20 12.3 + -1.13 0.26 234016 RB7117 181 546 3780322 3779777 Ribosomal protein L35 11.4 - -1.21 0.05 44425 RB7833 228 687 4192692 4193378 Ribosomal protein L3 11 + -1.28 0.32 254426 RB7834 222 669 4193390 4194058 Ribosomal protein L4/L1e 10.7 + -1.86 0.49 264427 RB7836 107 324 4194060 4194383 Ribosomal L23 protein 10.8 + -2.36 0.76 324428 RB7837 286 861 4194481 4195341 Ribosomal protein L2 11.8 + -1.87 0.05 34430 RB7839 119 360 4195706 4196065 Ribosomal protein L22/L17 11 + -1.88 0.07 44438 RB7854 61 186 4199624 4199809 Ribosomal protein S14 11.8 + -2.40 0.97 414463 RB7899 141 426 4220226 4220651 50S ribosomal protein L11 rplK 9.6 + -1.24 0.05 44750 RB8459 55 168 4518496 4518663 30S ribosomal protein S18 rpsR 10.5 + -1.50 0.33 224892 RB8725 54 165 4638301 4638465 50S ribosomal protein L33 rpmG 10.6 + -1.15 0.09 85832 RB10471 151 456 5678759 5678304 Ribosomal protein L13 10 - -1.63 0.64 397145 RB12824 146 441 6917743 6917303 Ribosomal protein S16 5.3 - -1.70 0.01 07154 RB12839 225 678 6921934 6922611 Ribosomal protein L1 9.8 + -1.98 0.58 294459 RB7894 398 1197 4217484 4218680 translation elongation factor EF-Tu 5.2 + -2.39 0.61 261030 RB1964 500 1503 1034475 1035977 hypothetical protein 4.7 + -1.19 0.11 95531 RB9917 169 510 5344370 5343861 Single-strand binding protein ssb 5.3 - -1.28 0.20 16415 RB778 413 1242 418773 417532 secreted protein containing DUF1559 7 - -1.18 0.22 19583 RB1103 441 1326 572105 570780 capsule biosynthesis protein capB 6.3 - -1.08 0.02 2
1130 RB2150 93 282 1138311 1138030 hypothetical protein 4.4 - -1.73 0.02 13239 RB5745 130 393 3001234 3000842 hypothetical protein 10.7 - -1.78 0.05 33271 RB5802 156 471 3039864 3040334 hypothetical protein 10.1 + -1.02 0.21 205810 RB10431 323 972 5650397 5649426 conserved hypothetical protein 10.1 - -1.22 0.15 127108 RB12760 50 153 6876908 6877060 conserved hypothetical protein 7.3 + -1.22 0.20 167109 RB12761 54 165 6877026 6877190 conserved hypothetical protein 12 + -1.03 0.14 13
20min ID Locus AA Nuc Product Gene EC IEP Strand Ratio Stdev %5700 RB10219 128 387 5521003 5521389 ATP synthase epsilon chain atpE 3.6.3.14 4.6 + -1.02 0.06 6179 RB314 309 930 156968 157897 malonyl CoA-acyl carrier protein transacylase fabD 2.3.1.39 4.4 + -1.45 0.36 25
2733 RB4876 397 1194 2506214 2505021 delta 9 acyl-lipid fatty acid desaturase 1.14.19.- 9.5 - -1.25 0.22 18658 RB1233 206 621 629490 628870 30S ribosomal protein S4 rpsD 11.2 - -1.24 0.27 22
4013 RB7114 347 1044 3779252 3778209 phenylalanyl-tRNA synthetase alpha chain pheS 6.1.1.20 4.6 - -1.05 0.27 254015 RB7116 59 180 3779586 3779765 hypothetical protein 11.7 + -1.41 0.26 194092 RB7246 50 153 3850393 3850545 hypothetical protein 9.7 + -1.05 0.15 144314 RB7646 62 189 4111735 4111923 hypothetical protein 10.5 + -1.24 0.25 204315 RB7647 73 222 4111931 4112152 hypothetical protein 7.4 + -1.91 0.10 54413 RB7818 121 366 4185730 4186095 Ribosomal protein S12/S23 11.8 + -1.50 0.11 84414 RB7820 157 474 4186170 4186643 Ribosomal protein S7 10.3 + -1.69 0.21 124423 RB7829 108 327 4192031 4192357 Ribosomal protein S10 10.3 + -1.70 0.28 174424 RB7832 50 153 4192499 4192651 hypothetical protein 13 + -1.67 0.25 154425 RB7833 228 687 4192692 4193378 Ribosomal protein L3 11 + -2.01 0.11 64427 RB7836 107 324 4194060 4194383 Ribosomal L23 protein 10.8 + -2.13 0.37 174438 RB7854 61 186 4199624 4199809 Ribosomal protein S14 11.8 + -2.12 0.27 134442 RB7859 177 534 4201351 4201884 Ribosomal protein S5 10.6 + -1.36 0.34 254463 RB7899 141 426 4220226 4220651 50S ribosomal protein L11 rplK 9.6 + -1.10 0.10 94750 RB8459 55 168 4518496 4518663 30S ribosomal protein S18 rpsR 10.5 + -2.05 0.31 154824 RB8580 113 342 4577393 4577734 sigma-54 modulation protein 5.7 + -1.14 0.20 185193 RB9304 81 246 4974378 4974623 Ribosomal protein L31 10.1 + -1.04 0.19 185832 RB10471 151 456 5678759 5678304 Ribosomal protein L13 10 - -2.13 0.31 157029 RB12623 127 384 6796904 6797287 Ribosomal protein S13 10.8 + -1.76 0.24 137032 RB12628 210 633 6798878 6799510 50S ribosomal protein L17 rplQ 10.2 + -1.28 0.32 257154 RB12839 225 678 6921934 6922611 Ribosomal protein L1 9.8 + -2.24 0.42 197166 RB12859 361 1086 6930771 6929686 Oligopeptide transport ATP-binding protein oppD oppD 5.5 - -1.00 0.12 125823 RB10458 206 621 5671130 5670510 Rnpc2-prov protein 10.3 - -1.52 0.07 43528 RB6276 105 318 3297504 3297187 Histone-like bacterial DNA-binding protein 10.4 - -2.03 0.16 83620 RB6428 164 495 3382816 3382322 conserved hypothetical protein, secreted 6.5 - -1.40 0.18 136086 RB10934 68 207 5934135 5934341 CsbD-like 4.3 + -1.26 0.07 6581 RB1097 404 1215 570281 569067 membrane protein 5.3 - -1.31 0.21 16883 RB1644 123 372 845824 845453 secreted protein 9.7 - -1.05 0.15 14
1505 RB2782 589 1770 1445576 1447345 protein containing DUF1559 7.2 + -1.24 0.14 112312 RB4194 53 162 2156685 2156524 hypothetical protein 11.4 - -1.15 0.08 73045 RB5409 97 294 2794195 2794488 hypothetical protein 12.7 + -1.41 0.25 183588 RB6379 153 462 3353122 3352661 hypothetical protein 5.8 - -1.08 0.16 153621 RB6429 61 186 3382998 3382813 hypothetical protein 12.1 - -1.29 0.29 224589 RB8144 143 432 4354622 4354191 hypothetical protein 10.1 - -1.74 0.20 114749 RB8457 113 342 4518458 4518117 hypothetical protein 11.5 - -1.11 0.07 65280 RB9460 79 240 5078860 5078621 hypothetical protein 10.3 - -1.26 0.12 105810 RB10431 323 972 5650397 5649426 conserved hypothetical protein 10.1 - -1.14 0.20 176085 RB10932 65 198 5933872 5934069 secreted protein 3.8 + -1.20 0.08 76094 RB10948 153 462 5938575 5939036 conserved hypothetical protein, secreted 4.2 + -1.74 0.11 77153 RB12837 65 198 6921732 6921929 conserved hypothetical protein 9.8 + -1.75 0.35 20
40min ID Locus AA Nuc Product Gene EC IEP Strand Ratio Stdev %2733 RB4876 397 1194 2506214 2505021 delta 9 acyl-lipid fatty acid desaturase 1.14.19.- 9.5 - -1.57 0.25 163528 RB6276 105 318 3297504 3297187 Histone-like bacterial DNA-binding protein 10.4 - -2.82 0.09 33620 RB6428 164 495 3382816 3382322 conserved hypothetical protein, secreted 6.5 - -1.78 0.21 122312 RB4194 53 162 2156685 2156524 hypothetical protein 11.4 - -1.21 0.13 113045 RB5409 97 294 2794195 2794488 hypothetical protein 12.7 + -1.83 0.48 263621 RB6429 61 186 3382998 3382813 hypothetical protein 12.1 - -1.30 0.06 54315 RB7647 73 222 4111931 4112152 hypothetical protein 7.4 + -1.71 0.39 234413 RB7818 121 366 4185730 4186095 Ribosomal protein S12/S23 11.8 + -2.00 0.13 74422 RB7828 40 123 4192037 4191915 hypothetical protein 10.4 - -1.15 0.17 154611 RB8191 72 219 4382096 4382314 hypothetical protein 12 + -1.18 0.02 24749 RB8457 113 342 4518458 4518117 hypothetical protein 11.5 - -1.47 0.09 64861 RB8669 37 114 4614557 4614670 hypothetical protein 11.5 + -1.92 0.37 195280 RB9460 79 240 5078860 5078621 hypothetical protein 10.3 - -1.99 0.30 156094 RB10948 153 462 5938575 5939036 conserved hypothetical protein, secreted 4.2 + -1.15 0.22 19
60min ID Locus AA Nuc Product Gene EC IEP Strand Ratio Stdev %5699 RB10217 479 1440 5519560 5520999 ATP synthase beta subunit 4.9 + -1.13 0.25 221967 RB3603 344 1035 1875461 1874427 secreted protein 4.6 - -1.12 0.25 222359 RB4269 282 849 2185049 2184201 glutamic acid specific endopeptidase, GSE=serine endopeptidase 5.6 - -1.13 0.31 282432 RB4386 349 1050 2259814 2258765 CYSTEINE SYNTHASE A cysK 2.5.1.47 5.6 - -1.54 0.08 54093 RB7247 366 1101 3850538 3851638 glutamine synthetase II glnII 6.3.1.2 4.8 + -1.90 0.24 134459 RB7894 398 1197 4217484 4218680 translation elongation factor EF-Tu 5.2 + -3.10 0.15 54756 RB8469 398 1197 4520779 4521975 lysine/ornithine decarboxylase ldc 4.1.1.18 5.2 + -1.22 0.07 66865 RB12327 686 2061 6633918 6631858 TGF-beta receptor, type I/II extracellular region 4.5 - -1.64 0.21 131423 RB2627 342 1029 1364551 1365579 Glyceraldehyde 3-phosphate dehydrogenase gap 1.2.1.12 5.4 + -1.50 0.05 43774 RB6690 343 1032 3555887 3556918 fructose-bisphosphate aldolase fba 4.1.2.13 4.7 + -1.40 0.25 184609 RB8188 259 780 4378573 4379352 hydroxypyruvate isomerase hyi 5.3.1.22 4.4 + -1.21 0.13 113533 RB6285 448 1347 3304945 3303599 S-adenosyl-L-homocysteine hydrolase 3.3.1.1 4.8 - -1.52 0.02 17190 RB12905 619 1860 6950939 6949080 acetolactate synthase, large subunit, biosynthetic type 5.6 - -1.18 0.30 25179 RB314 309 930 156968 157897 malonyl CoA-acyl carrier protein transacylase fabD 2.3.1.39 4.4 + -2.18 0.07 3181 RB318 81 246 158928 159173 Acyl carrier protein acpP 3.7 + -1.22 0.12 10
2733 RB4876 397 1194 2506214 2505021 delta 9 acyl-lipid fatty acid desaturase 1.14.19.- 9.5 - -1.16 0.05 4145 RB264 64 195 132677 132871 Ribosomal protein S21 12.3 + -1.82 0.12 7658 RB1233 206 621 629490 628870 30S ribosomal protein S4 rpsD 11.2 - -1.71 0.20 12
3270 RB5801 89 270 3039583 3039852 Ribosomal protein S15 10.6 + -1.48 0.13 93618 RB6425 50 153 3380727 3380879 50S ribosomal protein L36 rpl36 11.9 + -1.35 0.15 113822 RB6770 81 246 3599862 3599617 Ribosomal protein L27 11.4 - -1.17 0.31 273959 RB7022 90 273 3735291 3735563 Ribosomal protein S20 12.3 + -2.08 0.17 84014 RB7115 118 357 3779618 3779262 Ribosomal protein L20 12.2 - -1.65 0.21 134016 RB7117 181 546 3780322 3779777 Ribosomal protein L35 11.4 - -1.99 0.14 74413 RB7818 121 366 4185730 4186095 Ribosomal protein S12/S23 11.8 + -1.68 0.10 64414 RB7820 157 474 4186170 4186643 Ribosomal protein S7 10.3 + -1.76 0.06 44423 RB7829 108 327 4192031 4192357 Ribosomal protein S10 10.3 + -2.14 0.12 6

4425 RB7833 228 687 4192692 4193378 Ribosomal protein L3 11 + -1.81 0.16 94426 RB7834 222 669 4193390 4194058 Ribosomal protein L4/L1e 10.7 + -2.94 0.06 24427 RB7836 107 324 4194060 4194383 Ribosomal L23 protein 10.8 + -2.78 0.09 34428 RB7837 286 861 4194481 4195341 Ribosomal protein L2 11.8 + -2.68 0.15 64429 RB7838 89 270 4195376 4195645 Ribosomal protein S19/S15 10.8 + -2.48 0.05 24430 RB7839 119 360 4195706 4196065 Ribosomal protein L22/L17 11 + -2.44 0.11 44431 RB7840 236 711 4196161 4196871 30S ribosomal protein S3 rpsC 10.4 + -2.58 0.12 54432 RB7841 138 417 4196810 4197226 Ribosomal protein L16 11.1 + -2.77 0.27 104434 RB7849 108 327 4197700 4198026 Ribosomal protein S17 10 + -3.32 0.16 54435 RB7850 122 369 4198081 4198449 Ribosomal protein L14b/L23e 11 + -3.17 0.12 44437 RB7852 196 591 4198935 4199525 50S ribosomal protein L5 rplE 10.4 + -2.01 0.06 34438 RB7854 61 186 4199624 4199809 Ribosomal protein S14 11.8 + -3.32 0.19 64440 RB7856 181 546 4200306 4200851 50S ribosomal protein L6 rplF 10 + -2.95 0.35 124442 RB7859 177 534 4201351 4201884 Ribosomal protein S5 10.6 + -2.47 0.13 54463 RB7899 141 426 4220226 4220651 50S ribosomal protein L11 rplK 9.6 + -1.61 0.11 74750 RB8459 55 168 4518496 4518663 30S ribosomal protein S18 rpsR 10.5 + -2.53 0.20 84824 RB8580 113 342 4577393 4577734 sigma-54 modulation protein 5.7 + -1.65 0.10 64892 RB8725 54 165 4638301 4638465 50S ribosomal protein L33 rpmG 10.6 + -1.79 0.13 75534 RB9922 190 573 5345441 5344869 Peptidyl-tRNA hydrolase pth 3.1.1.29 8.6 - -1.22 0.18 155651 RB10134 104 315 5469243 5468929 Ribosomal protein L21 10 - -1.39 0.10 75830 RB10468 73 222 5677835 5677614 Translation initiation factor IF-1 infA 10.3 - -1.36 0.18 145831 RB10469 137 414 5678261 5677848 30S ribosomal protein S9 rpsI 11.1 - -1.41 0.16 115832 RB10471 151 456 5678759 5678304 Ribosomal protein L13 10 - -2.27 0.02 15918 RB10640 326 981 5774380 5775360 Elongation factor Ts tsf 4.6 + -1.15 0.14 126055 RB10880 477 1434 5907340 5905907 2-methylthioadenine synthetase 4.4 - -1.20 0.12 107029 RB12623 127 384 6796904 6797287 Ribosomal protein S13 10.8 + -2.12 0.15 77030 RB12625 127 384 6797354 6797737 Ribosomal protein S11 11.4 + -1.56 0.11 77032 RB12628 210 633 6798878 6799510 50S ribosomal protein L17 rplQ 10.2 + -1.03 0.18 177143 RB12821 117 354 6916558 6916205 Ribosomal protein L19 11.1 - -1.41 0.19 137144 RB12823 242 729 6917306 6916578 tRNA (guanine-N1-)-methyltransferase 2.1.1.31 4.9 - -2.21 0.24 117145 RB12824 146 441 6917743 6917303 Ribosomal protein S16 5.3 - -1.07 0.15 147154 RB12839 225 678 6921934 6922611 Ribosomal protein L1 9.8 + -1.36 0.15 114610 RB8189 931 2796 4382134 4379339 ATP-dependent RNA helicase DeaD 5 - -1.64 0.05 31030 RB1964 500 1503 1034475 1035977 hypothetical protein 4.7 + -1.35 0.10 72610 RB4681 65 198 2391934 2392131 cold shock protein cspA 4.7 + -1.86 0.36 193662 RB6491 526 1581 3438676 3437096 RNA polymerase sigma-54 factor rpoN 4.6 - -1.37 0.09 65823 RB10458 206 621 5671130 5670510 Rnpc2-prov protein 10.3 - -1.51 0.10 77031 RB12626 331 996 6797797 6798792 DNA-directed RNA polymerase alpha chain rpoA 2.7.7.6 4.9 + -1.83 0.08 43605 RB6403 304 915 3369939 3370853 hypothetical protein 5.4 + -1.47 0.10 73528 RB6276 105 318 3297504 3297187 Histone-like bacterial DNA-binding protein 10.4 - -2.19 0.25 115531 RB9917 169 510 5344370 5343861 Single-strand binding protein/Primosomal replication protein n ssb 5.3 - -2.32 0.14 62400 RB4333 361 1086 2230615 2231700 glycosyl transferase, group 1 family protein 6.4 + -1.12 0.18 163620 RB6428 164 495 3382816 3382322 conserved hypothetical protein, secreted 6.5 - -1.20 0.14 125812 RB10434 379 1140 5652902 5651763 glycosyl transferase, group 1 family protein 9.9 - -1.32 0.05 45813 RB10436 399 1200 5654064 5652865 glycosyl transferase, group 1 family protein 9.6 - -1.47 0.14 93646 RB6466 640 1923 3416688 3418610 GTP-binding protein TypA/BipA typA 5.1 + -1.13 0.13 117146 RB12825 491 1476 6919309 6917834 signal recognition particle protein ffh 9.8 - -2.03 0.11 51609 RB2970 261 786 1530415 1531200 Phosphatidylethanolamine N-methyltransferase pmtA 2.1.1.17 6.6 + -1.24 0.13 117140 RB12816 257 774 6914710 6915483 glutamine amidotransferase or related peptidase 4.5 + -1.09 0.16 15594 RB1129 895 2688 583271 580584 conserved hypothetical protein 5.8 - -1.27 0.06 5
6086 RB10934 68 207 5934135 5934341 CsbD-like 4.3 + -1.47 0.08 64825 RB8584 159 480 4577819 4578298 Nitrogen regulatory IIA protein ptsN 2.7.1.69 4.4 + -1.15 0.10 9
2 RB2 59 180 182 3 secreted protein 3.7 - -1.15 0.23 20182 RB319 95 288 159334 159621 hypothetical protein 10.2 + -1.42 0.05 3415 RB778 413 1242 418773 417532 secreted protein containing DUF1559 7 - -1.41 0.13 9581 RB1097 404 1215 570281 569067 membrane protein 5.3 - -1.93 0.14 7582 RB1100 166 501 570778 570278 capsule biosynthesis protein CapC capC 4.9 - -2.09 0.19 9583 RB1103 441 1326 572105 570780 capsule biosynthesis protein capB 6.3 - -1.96 0.25 13839 RB1563 136 411 798934 799344 hypothetical protein 10.2 + -1.22 0.07 6852 RB1588 151 456 817362 817817 ACP-like protein 5 + -1.15 0.22 19
1130 RB2150 93 282 1138311 1138030 hypothetical protein 4.4 - -2.47 0.09 41992 RB3644 127 384 1895184 1894801 hypothetical protein 4.9 - -1.09 0.06 62312 RB4194 53 162 2156685 2156524 hypothetical protein 11.4 - -1.10 0.14 132401 RB4336 372 1119 2231697 2232815 conserved hypothetical protein 5.6 + -1.07 0.25 242481 RB4464 223 672 2292075 2291404 secreted protein 5.1 - -1.16 0.03 32500 RB4497 49 150 2302950 2303099 hypothetical protein 10.3 + -1.12 0.03 32596 RB4657 123 372 2381865 2382236 hypothetical protein 12.2 + -1.05 0.28 262781 RB4951 95 288 2547498 2547785 hypothetical protein 12.1 + -1.86 0.14 72782 RB4954 166 501 2547861 2548361 secreted protein 10.2 + -1.91 0.15 83045 RB5409 97 294 2794195 2794488 hypothetical protein 12.7 + -2.26 0.04 23046 RB5411 35 108 2794516 2794623 hypothetical protein 10.5 + -1.24 0.06 53049 RB5415 62 189 2798737 2798925 hypothetical protein 12.3 + -1.04 0.09 93239 RB5745 130 393 3001234 3000842 hypothetical protein 10.7 - -2.34 0.26 113271 RB5802 156 471 3039864 3040334 hypothetical protein 10.1 + -1.57 0.06 43588 RB6379 153 462 3353122 3352661 hypothetical protein 5.8 - -1.37 0.12 83617 RB6423 45 138 3380603 3380740 hypothetical protein 5.5 + -1.08 0.17 163621 RB6429 61 186 3382998 3382813 hypothetical protein 12.1 - -1.03 0.10 103643 RB6462 90 273 3414908 3415180 hypothetical protein 9.9 + -1.06 0.00 03701 RB6553 74 225 3479627 3479403 hypothetical protein 12.1 - -1.22 0.07 64015 RB7116 59 180 3779586 3779765 hypothetical protein 11.7 + -1.64 0.11 74063 RB7203 147 444 3819614 3820057 hypothetical protein 4.6 + -1.24 0.20 164092 RB7246 50 153 3850393 3850545 hypothetical protein 9.7 + -2.04 0.15 74131 RB7313 51 156 3892097 3891942 hypothetical protein 10.5 - -1.00 0.07 74200 RB7455 838 2517 3999277 3996761 conserved hypothetical protein containing YTV domains 9 - -1.72 0.30 184314 RB7646 62 189 4111735 4111923 hypothetical protein 10.5 + -1.89 0.19 104315 RB7647 73 222 4111931 4112152 hypothetical protein 7.4 + -2.13 0.09 44327 RB7667 38 117 4117097 4116981 hypothetical protein 6.7 - -2.00 0.14 74424 RB7832 50 153 4192499 4192651 hypothetical protein 13 + -2.24 0.08 44461 RB7897 152 459 4218945 4219403 Protein secE/sec61-gamma protein 9.6 + -1.67 0.06 34589 RB8144 143 432 4354622 4354191 hypothetical protein 10.1 - -2.22 0.10 44698 RB8356 69 210 4459802 4460011 hypothetical protein 12.3 + -1.49 0.16 114749 RB8457 113 342 4518458 4518117 hypothetical protein 11.5 - -1.04 0.05 54818 RB8565 305 918 4573365 4572448 Chain A, 1.8 A Crystal Structure Of Ioli Protein With A Binding Zinc Atom 4.3 - -1.57 0.14 94827 RB8590 107 324 4578825 4579148 hypothetical protein 10.4 + -1.28 0.17 134830 RB8594 41 126 4581645 4581770 hypothetical protein 9.2 + -1.02 0.19 194861 RB8669 37 114 4614557 4614670 hypothetical protein 11.5 + -1.72 0.26 155226 RB9356 129 390 5012771 5012382 protein containing DUF1579 9.2 - -1.90 0.23 125280 RB9460 79 240 5078860 5078621 hypothetical protein 10.3 - -1.92 0.21 115694 RB10209 110 333 5515135 5515467 H+-transporting two-sector ATPase, C subunit 3.6.3.14 10.2 + -1.35 0.19 145810 RB10431 323 972 5650397 5649426 conserved hypothetical protein 10.1 - -1.90 0.12 65855 RB10521 53 162 5708182 5708343 hypothetical protein 9.1 + -1.01 0.19 195887 RB10581 384 1155 5739353 5740507 secreted protein containing DUF1559 6.2 + -1.11 0.24 216084 RB10931 54 165 5933742 5933906 hypothetical protein 4.2 + -1.35 0.21 166085 RB10932 65 198 5933872 5934069 secreted protein 3.8 + -1.56 0.14 96094 RB10948 153 462 5938575 5939036 conserved hypothetical protein, secreted 4.2 + -1.89 0.09 57055 RB12664 234 705 6817708 6818412 conserved hypothetical protein, secreted 6.7 + -1.02 0.02 27058 RB12669 473 1422 6821952 6823373 membrane protein 9.6 + -1.80 0.05 37108 RB12760 50 153 6876908 6877060 conserved hypothetical protein 7.3 + -1.05 0.00 07109 RB12761 54 165 6877026 6877190 conserved hypothetical protein 12 + -1.23 0.18 157141 RB12818 163 492 6915989 6915498 conserved hypothetical protein 11.7 - -1.89 0.08 47153 RB12837 65 198 6921732 6921929 conserved hypothetical protein 9.8 + -2.31 0.19 8
300min ID Locus AA Nuc Product Gene EC IEP Strand Ratio Stdev %2757 RB4913 109 330 2527319 2527648 ATP synthase C chain atpE 3.6.3.14 6.3 + -1.03 0.19 183570 RB6350 832 2499 3333612 3336110 Cytochrome C oxidase, mono-heme subunit/FixO 6 + -1.20 0.25 213649 RB6470 701 2106 3420270 3422375 Abortive infection protein 5.9 + -1.37 0.09 76105 RB10968 457 1374 5945417 5944044 Succinate-semialdehyde dehydrogenase [NADP+] ssdA 1.2.1.16 4.4 - -1.02 0.17 173418 RB6065 1234 3705 3185756 3182052 chromosome segregation SMC protein 5.5 - -1.05 0.21 20238 RB410 342 1029 219986 218958 anthranilate phosphoribosyltransferase trpD 2.4.2.18 5.1 - -1.06 0.20 19
1967 RB3603 344 1035 1875461 1874427 secreted protein 4.6 - -2.00 0.18 91984 RB3627 280 843 1888482 1887640 Transglutaminase-like domain 4.5 - -1.09 0.04 42359 RB4269 282 849 2185049 2184201 glutamic acid specific endopeptidase, GSE=serine endopeptidase 5.6 - -2.04 0.37 18

2432 RB4386 349 1050 2259814 2258765 CYSTEINE SYNTHASE A cysK 2.5.1.47 5.6 - -1.52 0.09 63188 RB5653 500 1503 2948867 2947365 NADH-glutamate synthase small chain 5 - -1.23 0.10 84459 RB7894 398 1197 4217484 4218680 translation elongation factor EF-Tu 5.2 + -2.68 0.03 14534 RB8038 1381 4146 4298294 4302439 Hydantoinase/oxoprolinase 5.3 + -1.28 0.06 56865 RB12327 686 2061 6633918 6631858 TGF-beta receptor, type I/II extracellular region 4.5 - -1.89 0.20 112192 RB3967 544 1635 2053237 2054871 sodium:proline/pantothenate symporter (proline/pantothenate permease) 9.4 + -1.23 0.20 16571 RB1074 353 1062 558194 559255 Bacterial inner-membrane translocator 9.2 + -1.39 0.05 3
1905 RB3496 511 1536 1808754 1810289 ribose ABC transporter, ATP-binding protein rbsA 3.6.3.17 5.4 + -1.24 0.07 64609 RB8188 259 780 4378573 4379352 hydroxypyruvate isomerase hyi 5.3.1.22 4.4 + -1.16 0.12 11181 RB318 81 246 158928 159173 Acyl carrier protein acpP 3.7 + -1.03 0.09 9
2733 RB4876 397 1194 2506214 2505021 delta 9 acyl-lipid fatty acid desaturase 1.14.19.- 9.5 - -1.51 0.06 45006 RB8921 393 1182 4751930 4750749 conserved hypothetical protein, membrane 9.9 - -1.13 0.19 17145 RB264 64 195 132677 132871 Ribosomal protein S21 12.3 + -1.44 0.08 5658 RB1233 206 621 629490 628870 30S ribosomal protein S4 rpsD 11.2 - -1.09 0.03 3
3959 RB7022 90 273 3735291 3735563 Ribosomal protein S20 12.3 + -1.44 0.14 94016 RB7117 181 546 3780322 3779777 Ribosomal protein L35 11.4 - -1.68 0.05 34413 RB7818 121 366 4185730 4186095 Ribosomal protein S12/S23 11.8 + -1.49 0.07 54414 RB7820 157 474 4186170 4186643 Ribosomal protein S7 10.3 + -1.66 0.14 84423 RB7829 108 327 4192031 4192357 Ribosomal protein S10 10.3 + -1.89 0.18 104424 RB7832 50 153 4192499 4192651 hypothetical protein 13 + -1.92 0.08 44425 RB7833 228 687 4192692 4193378 Ribosomal protein L3 11 + -1.31 0.03 24426 RB7834 222 669 4193390 4194058 Ribosomal protein L4/L1e 10.7 + -2.60 0.03 14427 RB7836 107 324 4194060 4194383 Ribosomal L23 protein 10.8 + -2.24 0.09 44428 RB7837 286 861 4194481 4195341 Ribosomal protein L2 11.8 + -1.84 0.13 74429 RB7838 89 270 4195376 4195645 Ribosomal protein S19/S15 10.8 + -1.79 0.09 54430 RB7839 119 360 4195706 4196065 Ribosomal protein L22/L17 11 + -1.60 0.08 54431 RB7840 236 711 4196161 4196871 30S ribosomal protein S3 rpsC 10.4 + -1.72 0.04 24432 RB7841 138 417 4196810 4197226 Ribosomal protein L16 11.1 + -1.56 0.14 94434 RB7849 108 327 4197700 4198026 Ribosomal protein S17 10 + -2.33 0.09 44435 RB7850 122 369 4198081 4198449 Ribosomal protein L14b/L23e 11 + -2.31 0.01 04436 RB7851 115 348 4198529 4198876 50S ribosomal protein L24 rplX 10.5 + -2.06 0.08 44437 RB7852 196 591 4198935 4199525 50S ribosomal protein L5 rplE 10.4 + -2.06 0.06 34438 RB7854 61 186 4199624 4199809 Ribosomal protein S14 11.8 + -2.29 0.06 34439 RB7855 133 402 4199839 4200240 Ribosomal protein S8 10.4 + -1.91 0.17 94440 RB7856 181 546 4200306 4200851 50S ribosomal protein L6 rplF 10 + -2.10 0.18 94441 RB7857 149 450 4200854 4201303 Ribosomal protein L18P/L5E 11.6 + -1.80 0.03 24442 RB7859 177 534 4201351 4201884 Ribosomal protein S5 10.6 + -1.52 0.10 74443 RB7863 164 495 4202048 4202542 Ribosomal protein L15 11.2 + -1.05 0.22 204750 RB8459 55 168 4518496 4518663 30S ribosomal protein S18 rpsR 10.5 + -1.11 0.07 75534 RB9922 190 573 5345441 5344869 Peptidyl-tRNA hydrolase pth 3.1.1.29 8.6 - -1.29 0.09 75651 RB10134 104 315 5469243 5468929 Ribosomal protein L21 10 - -1.36 0.27 205832 RB10471 151 456 5678759 5678304 Ribosomal protein L13 10 - -1.34 0.08 67029 RB12623 127 384 6796904 6797287 Ribosomal protein S13 10.8 + -1.56 0.09 67030 RB12625 127 384 6797354 6797737 Ribosomal protein S11 11.4 + -1.58 0.08 57032 RB12628 210 633 6798878 6799510 50S ribosomal protein L17 rplQ 10.2 + -1.22 0.03 37145 RB12824 146 441 6917743 6917303 Ribosomal protein S16 5.3 - -1.08 0.03 37154 RB12839 225 678 6921934 6922611 Ribosomal protein L1 9.8 + -1.72 0.06 47156 RB12842 140 423 6923401 6923823 Ribosomal protein L7/L12 4.3 + -1.52 0.08 54610 RB8189 931 2796 4382134 4379339 ATP-dependent RNA helicase DeaD 5 - -1.77 0.16 96709 RB12047 485 1458 6497993 6499450 ATP-dependent RNA helicase rhlE rhlE 10.6 + -1.60 0.15 102610 RB4681 65 198 2391934 2392131 cold shock protein cspA 4.7 + -1.65 0.09 63662 RB6491 526 1581 3438676 3437096 RNA polymerase sigma-54 factor rpoN 4.6 - -1.26 0.11 95823 RB10458 206 621 5671130 5670510 Rnpc2-prov protein 10.3 - -1.47 0.08 56819 RB12251 567 1704 6599262 6597559 DNA-directed RNA polymerase specialized sigma factor 9.4 - -1.57 0.01 17031 RB12626 331 996 6797797 6798792 DNA-directed RNA polymerase alpha chain rpoA 2.7.7.6 4.9 + -1.02 0.11 113605 RB6403 304 915 3369939 3370853 hypothetical protein 5.4 + -1.84 0.06 33528 RB6276 105 318 3297504 3297187 Histone-like bacterial DNA-binding protein 10.4 - -1.10 0.06 52400 RB4333 361 1086 2230615 2231700 glycosyl transferase, group 1 family protein 6.4 + -1.29 0.08 62479 RB4461 473 1422 2291114 2289693 MscS Mechanosensitive ion channel 4.5 - -1.38 0.08 63738 RB6624 503 1512 3520589 3522100 Bacterial sugar transferase 2.7.8.6 7.7 + -1.08 0.09 95813 RB10436 399 1200 5654064 5652865 glycosyl transferase, group 1 family protein 9.6 - -1.09 0.16 145814 RB10439 526 1581 5655714 5654134 Bacterial sugar transferase 2.7.8.6 10.1 - -1.16 0.15 133479 RB6180 100 303 3240284 3240586 Bacterial sec-independent translocation protein mttA/Hcf106 5.5 + -1.01 0.20 206766 RB12160 108 327 6563104 6562778 thioredoxin trxA 4 - -1.12 0.09 8374 RB684 653 1962 368557 366596 Arylsulfatase precursor atsA 3.1.6.1 5.5 - -1.04 0.14 14
2185 RB3956 489 1470 2050223 2048754 SULFATASE 5.5 - -1.06 0.04 45754 RB10326 588 1767 5580833 5582599 sulfate permease family protein 6 + -1.18 0.07 6537 RB990 500 1503 520003 521505 Major facilitator superfamily MFS_1 9.7 + -1.11 0.12 11
4107 RB7276 916 2751 3866177 3868927 peptide synthetase 5 + -1.08 0.14 131332 RB2479 273 822 1296683 1295862 conserved hypothetical protein 5.5 - -1.13 0.17 151609 RB2970 261 786 1530415 1531200 Phosphatidylethanolamine N-methyltransferase pmtA 2.1.1.17 6.6 + -1.30 0.04 3359 RB658 412 1239 356228 354990 transport protein 5.2 - -1.68 0.06 3
2427 RB4377 395 1188 2255017 2253830 transport protein 5.7 - -1.11 0.06 52476 RB4458 342 1029 2287089 2286061 Protein of unknown function, UPF0118 6.4 - -1.13 0.12 104255 RB7557 327 984 4058936 4059919 von Willebrand factor type A domain protein 4.9 + -1.09 0.05 54619 RB8208 412 1239 4387234 4388472 chlorohydrolase family protein 5.9 + -1.15 0.13 125487 RB9840 869 2610 5299200 5301809 DNA internalization-related competence protein ComEC/ 8.6 + -1.27 0.19 156115 RB10987 294 885 5954713 5955597 Phenazine biosynthesis PhzC/PhzF protein 4.6 + -1.04 0.08 86639 RB11920 249 750 6418443 6417694 Protein of unknown function Zn-binding 9.1 - -1.02 0.10 10594 RB1129 895 2688 583271 580584 conserved hypothetical protein 5.8 - -1.53 0.13 9
2337 RB4229 419 1260 2167854 2166595 conserved hypothetical protein 4.6 - -1.22 0.06 55039 RB8988 303 912 4790886 4791797 conserved hypothetical protein 6.1 + -1.02 0.05 55277 RB9448 115 348 5076267 5076614 conserved hypothetical membrane protein 7.4 + -1.01 0.16 153918 RB6945 589 1770 3686749 3688518 ADENYLATE CYCLASE TRANSMEMBRANE PROTEIN 9.8 + -1.08 0.18 166279 RB11286 131 396 6117961 6118356 TspO/MBR-related protein 9.8 + -1.12 0.21 19
37 RB61 58 177 25378 25554 hypothetical protein 9.6 + -1.44 0.09 656 RB95 157 474 40090 39617 conserved hypothetical protein, membrane 9.6 - -1.02 0.13 12
182 RB319 95 288 159334 159621 hypothetical protein 10.2 + -1.26 0.08 6193 RB337 97 294 169241 169534 hypothetical protein 12.5 + -1.10 0.15 14242 RB418 363 1092 222222 223313 conserved hypothetical protein, membrane 10.3 + -1.25 0.09 7250 RB434 65 198 231791 231988 hypothetical protein 7.4 + -1.02 0.20 20402 RB752 1026 3081 399966 403046 hypothetical protein 4.8 + -1.13 0.11 10409 RB767 311 936 411420 412355 conserved hypothetical protein, secreted 5.7 + -1.65 0.12 7415 RB778 413 1242 418773 417532 secreted protein containing DUF1559 7 - -1.95 0.09 5449 RB825 117 354 434184 433831 hypothetical protein 7.8 - -1.12 0.21 19516 RB951 234 705 502459 501755 protein containing DUF1596 12.3 - -1.12 0.27 24581 RB1097 404 1215 570281 569067 membrane protein 5.3 - -1.80 0.07 4582 RB1100 166 501 570778 570278 capsule biosynthesis protein CapC capC 4.9 - -2.91 0.07 3583 RB1103 441 1326 572105 570780 capsule biosynthesis protein capB 6.3 - -2.52 0.33 13849 RB1581 582 1749 814265 812517 membrane protein 6.7 - -1.54 0.15 10883 RB1644 123 372 845824 845453 secreted protein 9.7 - -2.32 0.10 4
1074 RB2051 367 1104 1079177 1080280 secreted protein 4.3 + -1.06 0.15 141105 RB2105 470 1413 1114012 1112600 membrane protein 9.6 - -1.14 0.06 61130 RB2150 93 282 1138311 1138030 hypothetical protein 4.4 - -1.20 0.04 41220 RB2306 41 126 1206782 1206907 hypothetical protein 9 + -1.03 0.08 81285 RB2400 66 201 1248699 1248499 hypothetical protein 12.1 - -1.01 0.09 91626 RB2993 97 294 1550784 1550491 hypothetical protein 5.3 - -1.02 0.15 141627 RB2995 95 288 1551052 1550765 hypothetical protein 6.5 - -1.23 0.15 121649 RB3042 291 876 1575622 1574747 secreted protein 6 - -1.24 0.10 81776 RB3277 221 666 1686552 1685887 hypothetical protein 10.4 - -1.21 0.10 91825 RB3362 87 264 1728546 1728283 hypothetical protein 11.9 - -1.19 0.18 151828 RB3366 78 237 1730380 1730616 hypothetical protein 5.6 + -1.24 0.23 191844 RB3394 36 111 1752044 1752154 hypothetical protein 10.5 + -1.29 0.14 111847 RB3399 65 198 1753650 1753453 hypothetical protein 9.7 - -1.28 0.20 161854 RB3407 89 270 1759935 1759666 hypothetical protein 10.5 - -1.35 0.14 101950 RB3575 152 459 1858779 1859237 membrane protein 10.1 + -1.38 0.07 51992 RB3644 127 384 1895184 1894801 hypothetical protein 4.9 - -1.50 0.03 22017 RB3675 742 2229 1904552 1906780 secreted protein 8.4 + -1.21 0.03 32025 RB3688 53 162 1910629 1910468 hypothetical protein 9.3 - -1.27 0.10 82139 RB3880 82 249 2006953 2006705 hypothetical protein 10.7 - -1.11 0.14 132146 RB3890 125 378 2011554 2011177 hypothetical protein 4.4 - -1.04 0.16 152166 RB3925 50 153 2033534 2033382 hypothetical protein 10.2 - -1.13 0.10 82184 RB3953 857 2574 2048784 2046211 hypothetical protein 5.2 - -1.44 0.06 4
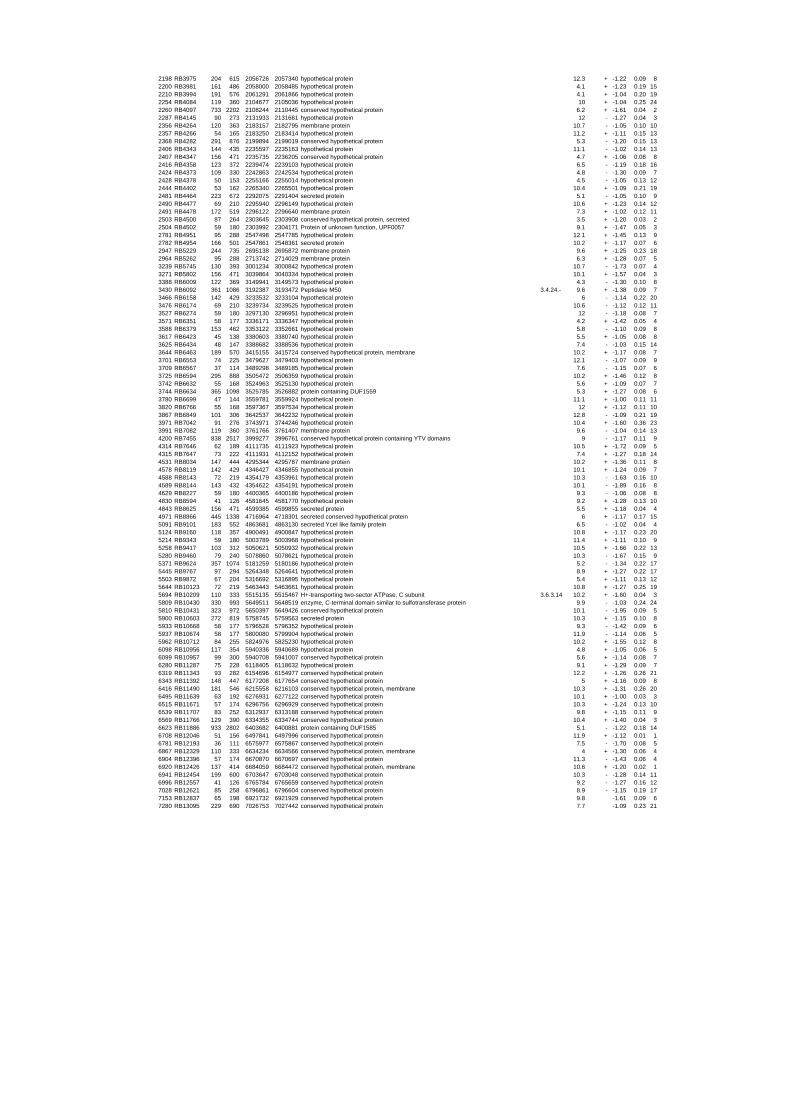
2198 RB3975 204 615 2056726 2057340 hypothetical protein 12.3 + -1.22 0.09 82200 RB3981 161 486 2058000 2058485 hypothetical protein 4.1 + -1.23 0.19 152210 RB3994 191 576 2061291 2061866 hypothetical protein 4.1 + -1.04 0.20 192254 RB4084 119 360 2104677 2105036 hypothetical protein 10 + -1.04 0.25 242260 RB4097 733 2202 2108244 2110445 conserved hypothetical protein 6.2 + -1.61 0.04 22287 RB4145 90 273 2131933 2131661 hypothetical protein 12 - -1.27 0.04 32356 RB4264 120 363 2183157 2182795 membrane protein 10.7 - -1.05 0.10 102357 RB4266 54 165 2183250 2183414 hypothetical protein 11.2 + -1.11 0.15 132368 RB4282 291 876 2199894 2199019 conserved hypothetical protein 5.3 - -1.20 0.15 132406 RB4343 144 435 2235597 2235163 hypothetical protein 11.1 - -1.02 0.14 132407 RB4347 156 471 2235735 2236205 conserved hypothetical protein 4.7 + -1.06 0.08 82416 RB4358 123 372 2239474 2239103 hypothetical protein 6.5 - -1.19 0.18 162424 RB4373 109 330 2242863 2242534 hypothetical protein 4.8 - -1.30 0.09 72428 RB4378 50 153 2255166 2255014 hypothetical protein 4.5 - -1.05 0.13 122444 RB4402 53 162 2265340 2265501 hypothetical protein 10.4 + -1.09 0.21 192481 RB4464 223 672 2292075 2291404 secreted protein 5.1 - -1.05 0.10 92490 RB4477 69 210 2295940 2296149 hypothetical protein 10.6 + -1.23 0.14 122491 RB4478 172 519 2296122 2296640 membrane protein 7.3 + -1.02 0.12 112503 RB4500 87 264 2303645 2303908 conserved hypothetical protein, secreted 3.5 + -1.20 0.03 22504 RB4502 59 180 2303992 2304171 Protein of unknown function, UPF0057 9.1 + -1.47 0.05 32781 RB4951 95 288 2547498 2547785 hypothetical protein 12.1 + -1.45 0.13 92782 RB4954 166 501 2547861 2548361 secreted protein 10.2 + -1.17 0.07 62947 RB5229 244 735 2695138 2695872 membrane protein 9.6 + -1.25 0.23 182964 RB5262 95 288 2713742 2714029 membrane protein 6.3 + -1.28 0.07 53239 RB5745 130 393 3001234 3000842 hypothetical protein 10.7 - -1.73 0.07 43271 RB5802 156 471 3039864 3040334 hypothetical protein 10.1 + -1.57 0.04 33388 RB6009 122 369 3149941 3149573 hypothetical protein 4.3 - -1.30 0.10 83430 RB6092 361 1086 3192387 3193472 Peptidase M50 3.4.24.- 9.6 + -1.38 0.09 73466 RB6158 142 429 3233532 3233104 hypothetical protein 6 - -1.14 0.22 203476 RB6174 69 210 3239734 3239525 hypothetical protein 10.6 - -1.12 0.12 113527 RB6274 59 180 3297130 3296951 hypothetical protein 12 - -1.18 0.08 73571 RB6351 58 177 3336171 3336347 hypothetical protein 4.2 + -1.42 0.05 43588 RB6379 153 462 3353122 3352661 hypothetical protein 5.8 - -1.10 0.09 83617 RB6423 45 138 3380603 3380740 hypothetical protein 5.5 + -1.05 0.08 83625 RB6434 48 147 3388682 3388536 hypothetical protein 7.4 - -1.03 0.15 143644 RB6463 189 570 3415155 3415724 conserved hypothetical protein, membrane 10.2 + -1.17 0.08 73701 RB6553 74 225 3479627 3479403 hypothetical protein 12.1 - -1.07 0.09 93709 RB6567 37 114 3489298 3489185 hypothetical protein 7.6 - -1.15 0.07 63725 RB6594 295 888 3505472 3506359 hypothetical protein 10.2 + -1.46 0.12 83742 RB6632 55 168 3524963 3525130 hypothetical protein 5.6 + -1.09 0.07 73744 RB6634 365 1098 3525785 3526882 protein containing DUF1559 5.3 + -1.27 0.08 63780 RB6699 47 144 3559781 3559924 hypothetical protein 11.1 + -1.00 0.11 113820 RB6766 55 168 3597367 3597534 hypothetical protein 12 + -1.12 0.11 103867 RB6849 101 306 3642537 3642232 hypothetical protein 12.8 - -1.09 0.21 193971 RB7042 91 276 3743971 3744246 hypothetical protein 10.4 + -1.60 0.36 233991 RB7082 119 360 3761766 3761407 membrane protein 9.6 - -1.04 0.14 134200 RB7455 838 2517 3999277 3996761 conserved hypothetical protein containing YTV domains 9 - -1.17 0.11 94314 RB7646 62 189 4111735 4111923 hypothetical protein 10.5 + -1.72 0.09 54315 RB7647 73 222 4111931 4112152 hypothetical protein 7.4 + -1.27 0.18 144531 RB8034 147 444 4295344 4295787 membrane protein 10.2 + -1.36 0.11 84578 RB8119 142 429 4346427 4346855 hypothetical protein 10.1 + -1.24 0.09 74588 RB8143 72 219 4354179 4353961 hypothetical protein 10.3 - -1.63 0.16 104589 RB8144 143 432 4354622 4354191 hypothetical protein 10.1 - -1.89 0.16 84629 RB8227 59 180 4400365 4400186 hypothetical protein 9.3 - -1.06 0.08 84830 RB8594 41 126 4581645 4581770 hypothetical protein 9.2 + -1.28 0.13 104843 RB8625 156 471 4599385 4599855 secreted protein 5.5 + -1.18 0.04 44971 RB8866 445 1338 4716964 4718301 secreted conserved hypothetical protein 6 + -1.17 0.17 155091 RB9101 183 552 4863681 4863130 secreted YceI like family protein 6.5 - -1.02 0.04 45124 RB9160 118 357 4900491 4900847 hypothetical protein 10.8 + -1.17 0.23 205214 RB9343 59 180 5003789 5003968 hypothetical protein 11.4 + -1.11 0.10 95258 RB9417 103 312 5050621 5050932 hypothetical protein 10.5 + -1.66 0.22 135280 RB9460 79 240 5078860 5078621 hypothetical protein 10.3 - -1.67 0.15 95371 RB9624 357 1074 5181259 5180186 hypothetical protein 5.2 - -1.34 0.22 175445 RB9767 97 294 5264348 5264641 hypothetical protein 8.9 + -1.27 0.22 175503 RB9872 67 204 5316692 5316895 hypothetical protein 5.4 + -1.11 0.13 125644 RB10123 72 219 5463443 5463661 hypothetical protein 10.8 + -1.27 0.25 195694 RB10209 110 333 5515135 5515467 H+-transporting two-sector ATPase, C subunit 3.6.3.14 10.2 + -1.60 0.04 35809 RB10430 330 993 5649511 5648519 enzyme, C-terminal domain similar to sulfotransferase protein 9.9 - -1.03 0.24 245810 RB10431 323 972 5650397 5649426 conserved hypothetical protein 10.1 - -1.95 0.09 55900 RB10603 272 819 5758745 5759563 secreted protein 10.3 + -1.15 0.10 85933 RB10668 58 177 5796528 5796352 hypothetical protein 9.3 - -1.42 0.09 65937 RB10674 58 177 5800080 5799904 hypothetical protein 11.9 - -1.14 0.06 55962 RB10712 84 255 5824976 5825230 hypothetical protein 10.2 + -1.55 0.12 86098 RB10956 117 354 5940336 5940689 hypothetical protein 4.8 + -1.05 0.06 56099 RB10957 99 300 5940708 5941007 conserved hypothetical protein 5.6 + -1.14 0.08 76280 RB11287 75 228 6118405 6118632 hypothetical protein 9.1 + -1.29 0.09 76319 RB11343 93 282 6154696 6154977 conserved hypothetical protein 12.2 + -1.26 0.26 216343 RB11392 148 447 6177208 6177654 conserved hypothetical protein 5 + -1.16 0.09 86416 RB11490 181 546 6215558 6216103 conserved hypothetical protein, membrane 10.3 + -1.31 0.26 206495 RB11639 63 192 6276931 6277122 conserved hypothetical protein 10.1 + -1.00 0.03 36515 RB11671 57 174 6296756 6296929 conserved hypothetical protein 10.3 + -1.24 0.13 106539 RB11707 83 252 6312937 6313188 conserved hypothetical protein 9.8 + -1.15 0.11 96569 RB11766 129 390 6334355 6334744 conserved hypothetical protein 10.4 + -1.40 0.04 36623 RB11886 933 2802 6403682 6400881 protein containing DUF1585 5.1 - -1.22 0.18 146708 RB12046 51 156 6497841 6497996 conserved hypothetical protein 11.9 + -1.12 0.01 16781 RB12193 36 111 6575977 6575867 conserved hypothetical protein 7.5 - -1.70 0.08 56867 RB12329 110 333 6634234 6634566 conserved hypothetical protein, membrane 4 + -1.30 0.06 46904 RB12396 57 174 6670870 6670697 conserved hypothetical protein 11.3 - -1.43 0.06 46920 RB12426 137 414 6684059 6684472 conserved hypothetical protein, membrane 10.6 + -1.20 0.02 16941 RB12454 199 600 6703647 6703048 conserved hypothetical protein 10.3 - -1.28 0.14 116996 RB12557 41 126 6765784 6765659 conserved hypothetical protein 9.2 - -1.27 0.16 127028 RB12621 85 258 6796861 6796604 conserved hypothetical protein 8.9 - -1.15 0.19 177153 RB12837 65 198 6921732 6921929 conserved hypothetical protein 9.8 -1.61 0.09 67280 RB13095 229 690 7026753 7027442 conserved hypothetical protein 7.7 -1.09 0.23 21
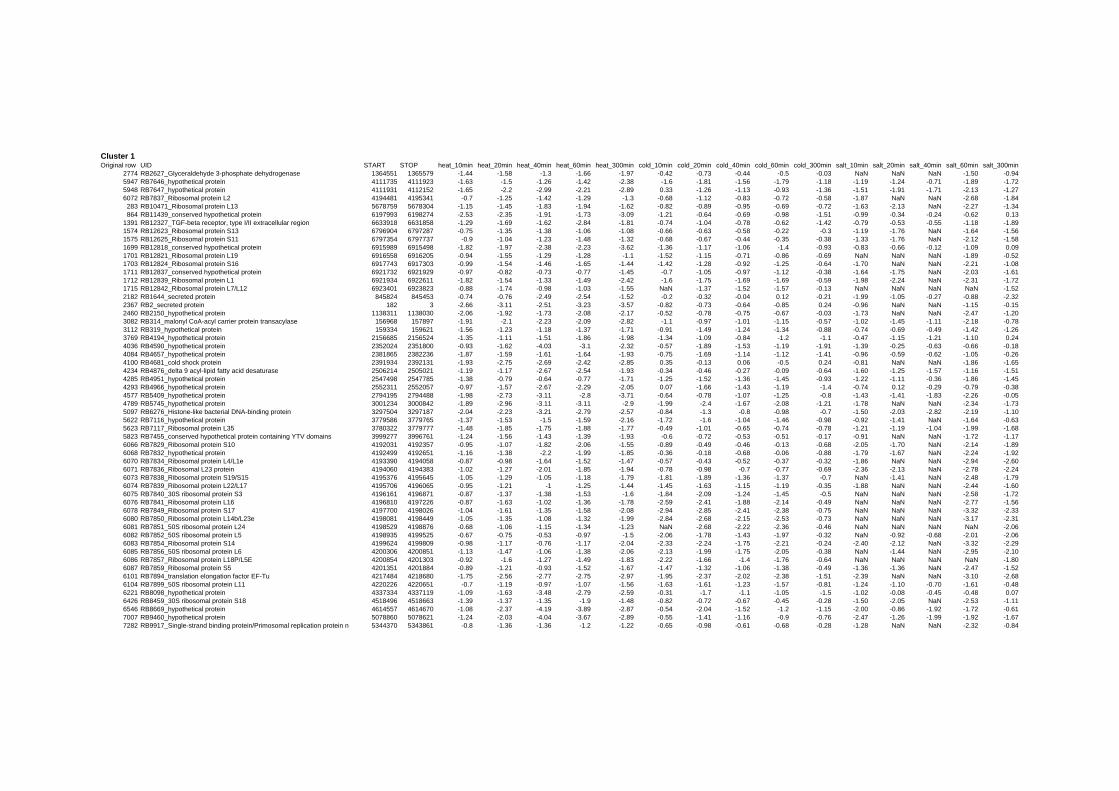
Cluster 1Original row UID START STOP heat_10min heat_20min heat_40min heat_60min heat_300min cold_10min cold_20min cold_40min cold_60min cold_300min salt_10min salt_20min salt_40min salt_60min salt_300min
2774 RB2627_Glyceraldehyde 3-phosphate dehydrogenase 1364551 1365579 -1.44 -1.58 -1.3 -1.66 -1.97 -0.42 -0.73 -0.44 -0.5 -0.03 NaN NaN NaN -1.50 -0.945947 RB7646_hypothetical protein 4111735 4111923 -1.63 -1.5 -1.26 -1.42 -2.38 -1.6 -1.81 -1.56 -1.79 -1.18 -1.19 -1.24 -0.71 -1.89 -1.725948 RB7647_hypothetical protein 4111931 4112152 -1.65 -2.2 -2.99 -2.21 -2.89 0.33 -1.26 -1.13 -0.93 -1.36 -1.51 -1.91 -1.71 -2.13 -1.276072 RB7837_Ribosomal protein L2 4194481 4195341 -0.7 -1.25 -1.42 -1.29 -1.3 -0.68 -1.12 -0.83 -0.72 -0.58 -1.87 NaN NaN -2.68 -1.84283 RB10471_Ribosomal protein L13 5678759 5678304 -1.15 -1.45 -1.83 -1.94 -1.62 -0.82 -0.89 -0.95 -0.69 -0.72 -1.63 -2.13 NaN -2.27 -1.34864 RB11439_conserved hypothetical protein 6197993 6198274 -2.53 -2.35 -1.91 -1.73 -3.09 -1.21 -0.64 -0.69 -0.98 -1.51 -0.99 -0.34 -0.24 -0.62 0.13
1391 RB12327_TGF-beta receptor, type I/II extracellular region 6633918 6631858 -1.29 -1.69 -1.62 -2.84 -1.81 -0.74 -1.04 -0.78 -0.62 -1.42 -0.79 -0.53 -0.55 -1.18 -1.891574 RB12623_Ribosomal protein S13 6796904 6797287 -0.75 -1.35 -1.38 -1.06 -1.08 -0.66 -0.63 -0.58 -0.22 -0.3 -1.19 -1.76 NaN -1.64 -1.561575 RB12625_Ribosomal protein S11 6797354 6797737 -0.9 -1.04 -1.23 -1.48 -1.32 -0.68 -0.67 -0.44 -0.35 -0.38 -1.33 -1.76 NaN -2.12 -1.581699 RB12818_conserved hypothetical protein 6915989 6915498 -1.82 -1.97 -2.38 -2.23 -3.62 -1.36 -1.17 -1.06 -1.4 -0.93 -0.83 -0.66 -0.12 -1.09 0.091701 RB12821_Ribosomal protein L19 6916558 6916205 -0.94 -1.55 -1.29 -1.28 -1.1 -1.52 -1.15 -0.71 -0.86 -0.69 NaN NaN NaN -1.89 -0.521703 RB12824_Ribosomal protein S16 6917743 6917303 -0.99 -1.54 -1.46 -1.65 -1.44 -1.42 -1.28 -0.92 -1.25 -0.64 -1.70 NaN NaN -2.21 -1.081711 RB12837_conserved hypothetical protein 6921732 6921929 -0.97 -0.82 -0.73 -0.77 -1.45 -0.7 -1.05 -0.97 -1.12 -0.38 -1.64 -1.75 NaN -2.03 -1.611712 RB12839_Ribosomal protein L1 6921934 6922611 -1.82 -1.54 -1.33 -1.49 -2.42 -1.6 -1.75 -1.69 -1.69 -0.59 -1.98 -2.24 NaN -2.31 -1.721715 RB12842_Ribosomal protein L7/L12 6923401 6923823 -0.88 -1.74 -0.98 -1.03 -1.55 NaN -1.37 -1.52 -1.57 -0.13 NaN NaN NaN NaN -1.522182 RB1644_secreted protein 845824 845453 -0.74 -0.76 -2.49 -2.54 -1.52 -0.2 -0.32 -0.04 0.12 -0.21 -1.99 -1.05 -0.27 -0.88 -2.322367 RB2_secreted protein 182 3 -2.66 -3.11 -2.51 -3.23 -3.57 -0.82 -0.73 -0.64 -0.85 0.24 -0.96 NaN NaN -1.15 -0.152460 RB2150_hypothetical protein 1138311 1138030 -2.06 -1.92 -1.73 -2.08 -2.17 -0.52 -0.78 -0.75 -0.67 -0.03 -1.73 NaN NaN -2.47 -1.203082 RB314_malonyl CoA-acyl carrier protein transacylase 156968 157897 -1.91 -2.1 -2.23 -2.09 -2.82 -1.1 -0.97 -1.01 -1.15 -0.57 -1.02 -1.45 -1.11 -2.18 -0.783112 RB319_hypothetical protein 159334 159621 -1.56 -1.23 -1.18 -1.37 -1.71 -0.91 -1.49 -1.24 -1.34 -0.88 -0.74 -0.69 -0.49 -1.42 -1.263769 RB4194_hypothetical protein 2156685 2156524 -1.35 -1.11 -1.51 -1.86 -1.98 -1.34 -1.09 -0.84 -1.2 -1.1 -0.47 -1.15 -1.21 -1.10 0.244036 RB4590_hypothetical protein 2352024 2351800 -0.93 -1.62 -4.03 -3.1 -2.32 -0.57 -1.89 -1.53 -1.19 -1.91 -1.39 -0.25 -0.63 -0.66 -0.184084 RB4657_hypothetical protein 2381865 2382236 -1.87 -1.59 -1.61 -1.64 -1.93 -0.75 -1.69 -1.14 -1.12 -1.41 -0.96 -0.59 -0.62 -1.05 -0.264100 RB4681_cold shock protein 2391934 2392131 -1.93 -2.75 -2.69 -2.42 -2.85 0.35 -0.13 0.06 -0.5 0.24 -0.81 NaN NaN -1.86 -1.654234 RB4876_delta 9 acyl-lipid fatty acid desaturase 2506214 2505021 -1.19 -1.17 -2.67 -2.54 -1.93 -0.34 -0.46 -0.27 -0.09 -0.64 -1.60 -1.25 -1.57 -1.16 -1.514285 RB4951_hypothetical protein 2547498 2547785 -1.38 -0.79 -0.64 -0.77 -1.71 -1.25 -1.52 -1.36 -1.45 -0.93 -1.22 -1.11 -0.36 -1.86 -1.454293 RB4966_hypothetical protein 2552311 2552057 -0.97 -1.57 -2.67 -2.29 -2.05 0.07 -1.66 -1.43 -1.19 -1.4 -0.74 0.12 -0.29 -0.79 -0.384577 RB5409_hypothetical protein 2794195 2794488 -1.98 -2.73 -3.11 -2.8 -3.71 -0.64 -0.78 -1.07 -1.25 -0.8 -1.43 -1.41 -1.83 -2.26 -0.054789 RB5745_hypothetical protein 3001234 3000842 -1.89 -2.96 -3.11 -3.11 -2.9 -1.99 -2.4 -1.67 -2.08 -1.21 -1.78 NaN NaN -2.34 -1.735097 RB6276_Histone-like bacterial DNA-binding protein 3297504 3297187 -2.04 -2.23 -3.21 -2.79 -2.57 -0.84 -1.3 -0.8 -0.98 -0.7 -1.50 -2.03 -2.82 -2.19 -1.105622 RB7116_hypothetical protein 3779586 3779765 -1.37 -1.53 -1.5 -1.59 -2.16 -1.72 -1.6 -1.04 -1.46 -0.98 -0.92 -1.41 NaN -1.64 -0.635623 RB7117_Ribosomal protein L35 3780322 3779777 -1.48 -1.85 -1.75 -1.88 -1.77 -0.49 -1.01 -0.65 -0.74 -0.78 -1.21 -1.19 -1.04 -1.99 -1.685823 RB7455_conserved hypothetical protein containing YTV domains 3999277 3996761 -1.24 -1.56 -1.43 -1.39 -1.93 -0.6 -0.72 -0.53 -0.51 -0.17 -0.91 NaN NaN -1.72 -1.176066 RB7829_Ribosomal protein S10 4192031 4192357 -0.95 -1.07 -1.82 -2.06 -1.55 -0.89 -0.49 -0.46 -0.13 -0.68 -2.05 -1.70 NaN -2.14 -1.896068 RB7832_hypothetical protein 4192499 4192651 -1.16 -1.38 -2.2 -1.99 -1.85 -0.36 -0.18 -0.68 -0.06 -0.88 -1.79 -1.67 NaN -2.24 -1.926070 RB7834_Ribosomal protein L4/L1e 4193390 4194058 -0.87 -0.98 -1.64 -1.52 -1.47 -0.57 -0.43 -0.52 -0.37 -0.32 -1.86 NaN NaN -2.94 -2.606071 RB7836_Ribosomal L23 protein 4194060 4194383 -1.02 -1.27 -2.01 -1.85 -1.94 -0.78 -0.98 -0.7 -0.77 -0.69 -2.36 -2.13 NaN -2.78 -2.246073 RB7838_Ribosomal protein S19/S15 4195376 4195645 -1.05 -1.29 -1.05 -1.18 -1.79 -1.81 -1.89 -1.36 -1.37 -0.7 NaN -1.41 NaN -2.48 -1.796074 RB7839_Ribosomal protein L22/L17 4195706 4196065 -0.95 -1.21 -1 -1.25 -1.44 -1.45 -1.63 -1.15 -1.19 -0.35 -1.88 NaN NaN -2.44 -1.606075 RB7840_30S ribosomal protein S3 4196161 4196871 -0.87 -1.37 -1.38 -1.53 -1.6 -1.84 -2.09 -1.24 -1.45 -0.5 NaN NaN NaN -2.58 -1.726076 RB7841_Ribosomal protein L16 4196810 4197226 -0.87 -1.63 -1.02 -1.36 -1.78 -2.59 -2.41 -1.88 -2.14 -0.49 NaN NaN NaN -2.77 -1.566078 RB7849_Ribosomal protein S17 4197700 4198026 -1.04 -1.61 -1.35 -1.58 -2.08 -2.94 -2.85 -2.41 -2.38 -0.75 NaN NaN NaN -3.32 -2.336080 RB7850_Ribosomal protein L14b/L23e 4198081 4198449 -1.05 -1.35 -1.08 -1.32 -1.99 -2.84 -2.68 -2.15 -2.53 -0.73 NaN NaN NaN -3.17 -2.316081 RB7851_50S ribosomal protein L24 4198529 4198876 -0.68 -1.06 -1.15 -1.34 -1.23 NaN -2.68 -2.22 -2.36 -0.46 NaN NaN NaN NaN -2.066082 RB7852_50S ribosomal protein L5 4198935 4199525 -0.67 -0.75 -0.53 -0.97 -1.5 -2.06 -1.78 -1.43 -1.97 -0.32 NaN -0.92 -0.68 -2.01 -2.066083 RB7854_Ribosomal protein S14 4199624 4199809 -0.98 -1.17 -0.76 -1.17 -2.04 -2.33 -2.24 -1.75 -2.21 -0.24 -2.40 -2.12 NaN -3.32 -2.296085 RB7856_50S ribosomal protein L6 4200306 4200851 -1.13 -1.47 -1.06 -1.38 -2.06 -2.13 -1.99 -1.75 -2.05 -0.38 NaN -1.44 NaN -2.95 -2.106086 RB7857_Ribosomal protein L18P/L5E 4200854 4201303 -0.92 -1.6 -1.27 -1.49 -1.83 -2.22 -1.66 -1.4 -1.76 -0.64 NaN NaN NaN NaN -1.806087 RB7859_Ribosomal protein S5 4201351 4201884 -0.89 -1.21 -0.93 -1.52 -1.67 -1.47 -1.32 -1.06 -1.38 -0.49 -1.36 -1.36 NaN -2.47 -1.526101 RB7894_translation elongation factor EF-Tu 4217484 4218680 -1.75 -2.56 -2.77 -2.75 -2.97 -1.95 -2.37 -2.02 -2.38 -1.51 -2.39 NaN NaN -3.10 -2.686104 RB7899_50S ribosomal protein L11 4220226 4220651 -0.7 -1.19 -0.97 -1.07 -1.56 -1.63 -1.61 -1.23 -1.57 -0.81 -1.24 -1.10 -0.70 -1.61 -0.486221 RB8098_hypothetical protein 4337334 4337119 -1.09 -1.63 -3.48 -2.79 -2.59 -0.31 -1.7 -1.1 -1.05 -1.5 -1.02 -0.08 -0.45 -0.48 0.076426 RB8459_30S ribosomal protein S18 4518496 4518663 -1.39 -1.37 -1.35 -1.9 -1.48 -0.82 -0.72 -0.67 -0.45 -0.28 -1.50 -2.05 NaN -2.53 -1.116546 RB8669_hypothetical protein 4614557 4614670 -1.08 -2.37 -4.19 -3.89 -2.87 -0.54 -2.04 -1.52 -1.2 -1.15 -2.00 -0.86 -1.92 -1.72 -0.617007 RB9460_hypothetical protein 5078860 5078621 -1.24 -2.03 -4.04 -3.67 -2.89 -0.55 -1.41 -1.16 -0.9 -0.76 -2.47 -1.26 -1.99 -1.92 -1.677282 RB9917_Single-strand binding protein/Primosomal replication protein n 5344370 5343861 -0.8 -1.36 -1.36 -1.2 -1.22 -0.65 -0.98 -0.61 -0.68 -0.28 -1.28 NaN NaN -2.32 -0.84
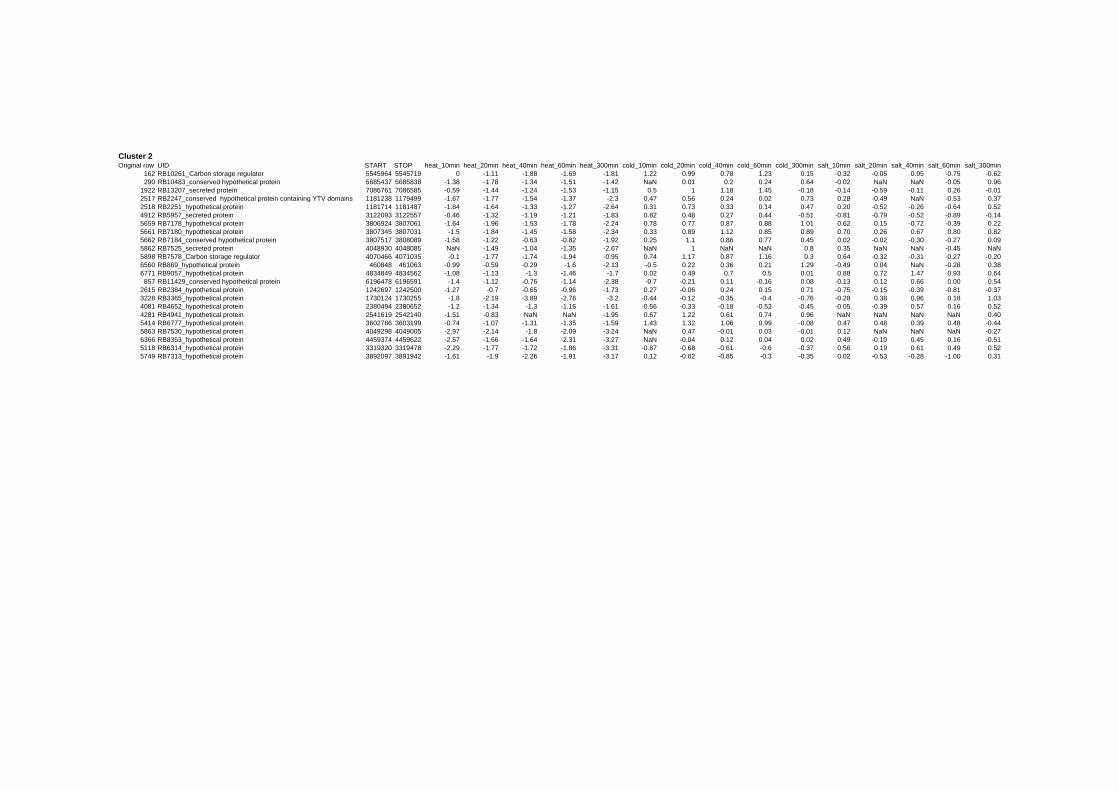
Cluster 2Original row UID START STOP heat_10min heat_20min heat_40min heat_60min heat_300min cold_10min cold_20min cold_40min cold_60min cold_300min salt_10min salt_20min salt_40min salt_60min salt_300min
162 RB10261_Carbon storage regulator 5545964 5545719 0 -1.11 -1.88 -1.69 -1.81 1.22 0.99 0.78 1.23 0.15 -0.32 -0.05 0.95 -0.75 -0.62290 RB10483_conserved hypothetical protein 5685437 5685838 -1.38 -1.78 -1.34 -1.51 -1.42 NaN 0.01 0.2 0.24 0.64 -0.02 NaN NaN -0.05 0.96
1922 RB13207_secreted protein 7086761 7086585 -0.59 -1.44 -1.24 -1.53 -1.15 0.5 1 1.18 1.45 -0.18 -0.14 -0.59 -0.11 0.26 -0.012517 RB2247_conserved hypothetical protein containing YTV domains 1181238 1179499 -1.67 -1.77 -1.54 -1.37 -2.3 0.47 0.56 0.24 0.02 0.73 0.28 -0.49 NaN -0.53 0.372518 RB2251_hypothetical protein 1181714 1181487 -1.84 -1.64 -1.33 -1.27 -2.64 0.31 0.73 0.33 0.14 0.47 0.20 -0.52 -0.26 -0.64 0.524912 RB5957_secreted protein 3122093 3122557 -0.46 -1.32 -1.19 -1.21 -1.83 0.82 0.48 0.27 0.44 -0.51 -0.81 -0.79 -0.52 -0.89 -0.145659 RB7178_hypothetical protein 3806924 3807061 -1.64 -1.96 -1.53 -1.78 -2.24 0.78 0.77 0.87 0.88 1.01 0.62 0.15 -0.72 -0.39 0.225661 RB7180_hypothetical protein 3807345 3807031 -1.5 -1.84 -1.45 -1.58 -2.34 0.33 0.89 1.12 0.85 0.89 0.70 0.26 0.67 0.80 0.825662 RB7184_conserved hypothetical protein 3807517 3808089 -1.58 -1.22 -0.63 -0.82 -1.92 0.25 1.1 0.86 0.77 0.45 0.02 -0.02 -0.30 -0.27 0.095862 RB7525_secreted protein 4048930 4048085 NaN -1.49 -1.04 -1.35 -2.67 NaN 1 NaN NaN 0.8 0.35 NaN NaN -0.45 NaN5898 RB7578_Carbon storage regulator 4070466 4071035 -0.1 -1.77 -1.74 -1.94 -0.95 0.74 1.17 0.87 1.16 0.3 0.64 -0.32 -0.31 -0.27 -0.206560 RB869_hypothetical protein 460848 461063 -0.99 -0.59 -0.29 -1.6 -2.13 -0.5 0.22 0.36 0.21 1.29 -0.49 0.04 NaN -0.28 0.386771 RB9057_hypothetical protein 4834849 4834562 -1.08 -1.13 -1.3 -1.46 -1.7 0.02 0.49 0.7 0.5 0.01 0.88 0.72 1.47 0.93 0.64857 RB11429_conserved hypothetical protein 6196478 6196591 -1.4 -1.12 -0.76 -1.14 -2.38 -0.7 -0.21 0.11 -0.16 0.08 -0.13 0.12 0.66 0.00 0.54
2615 RB2384_hypothetical protein 1242697 1242500 -1.27 -0.7 -0.65 -0.96 -1.73 0.27 -0.06 0.24 0.15 0.71 -0.75 -0.15 -0.39 -0.81 -0.373228 RB3365_hypothetical protein 1730124 1730255 -1.8 -2.19 -3.89 -2.76 -3.2 -0.44 -0.12 -0.35 -0.4 -0.76 -0.28 0.38 0.96 0.18 1.034081 RB4652_hypothetical protein 2380494 2380652 -1.2 -1.34 -1.3 -1.16 -1.61 -0.56 -0.33 -0.18 -0.52 -0.45 -0.05 -0.39 0.57 0.16 0.524281 RB4941_hypothetical protein 2541619 2542140 -1.51 -0.83 NaN NaN -1.95 0.67 1.22 0.61 0.74 0.96 NaN NaN NaN NaN 0.405414 RB6777_hypothetical protein 3602786 3603199 -0.74 -1.07 -1.31 -1.35 -1.59 1.43 1.32 1.06 0.99 -0.08 0.47 0.48 0.39 0.48 -0.445863 RB7530_hypothetical protein 4049298 4049005 -2.97 -2.14 -1.8 -2.09 -3.24 NaN 0.47 -0.01 0.03 -0.01 0.12 NaN NaN NaN -0.276366 RB8353_hypothetical protein 4459374 4459622 -2.57 -1.66 -1.64 -2.31 -3.27 NaN -0.04 0.12 0.04 0.02 0.49 -0.10 0.45 0.16 -0.515118 RB6314_hypothetical protein 3319320 3319478 -2.29 -1.77 -1.72 -1.86 -3.31 -0.87 -0.68 -0.61 -0.6 -0.37 0.56 0.19 0.61 0.49 0.525749 RB7313_hypothetical protein 3892097 3891942 -1.61 -1.9 -2.26 -1.91 -3.17 0.12 -0.02 -0.85 -0.3 -0.35 0.02 -0.53 -0.28 -1.00 0.31
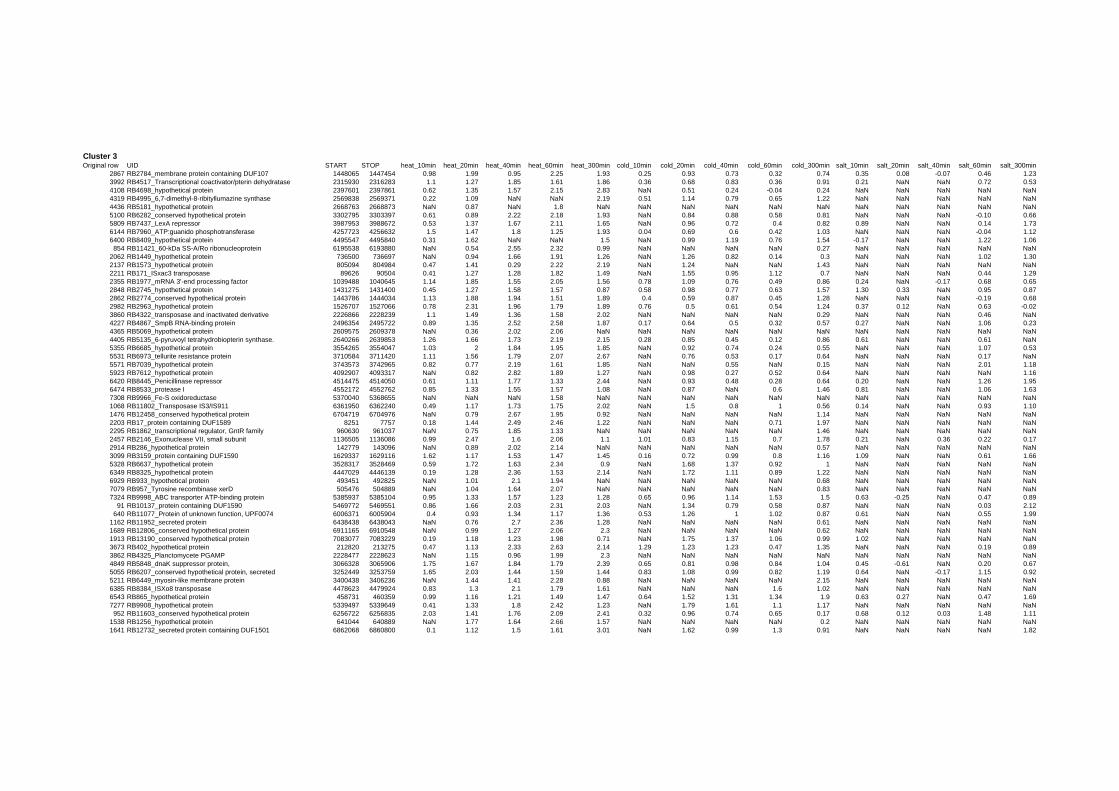
Cluster 3Original row UID START STOP heat_10min heat_20min heat_40min heat_60min heat_300min cold_10min cold_20min cold_40min cold_60min cold_300min salt_10min salt_20min salt_40min salt_60min salt_300min
2867 RB2784_membrane protein containing DUF107 1448065 1447454 0.98 1.99 0.95 2.25 1.93 0.25 0.93 0.73 0.32 0.74 0.35 0.08 -0.07 0.46 1.233992 RB4517_Transcriptional coactivator/pterin dehydratase 2315930 2316283 1.1 1.27 1.85 1.61 1.86 0.36 0.68 0.83 0.36 0.91 0.21 NaN NaN 0.72 0.534108 RB4698_hypothetical protein 2397601 2397861 0.62 1.35 1.57 2.15 2.83 NaN 0.51 0.24 -0.04 0.24 NaN NaN NaN NaN NaN4319 RB4995_6,7-dimethyl-8-ribityllumazine synthase 2569838 2569371 0.22 1.09 NaN NaN 2.19 0.51 1.14 0.79 0.65 1.22 NaN NaN NaN NaN NaN4436 RB5181_hypothetical protein 2668763 2668873 NaN 0.87 NaN 1.8 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN5100 RB6282_conserved hypothetical protein 3302795 3303397 0.61 0.89 2.22 2.18 1.93 NaN 0.84 0.88 0.58 0.81 NaN NaN NaN -0.10 0.665809 RB7437_LexA repressor 3987953 3988672 0.53 1.37 1.67 2.11 1.65 NaN 0.96 0.72 0.4 0.82 0.89 NaN NaN 0.14 1.736144 RB7960_ATP:guanido phosphotransferase 4257723 4256632 1.5 1.47 1.8 1.25 1.93 0.04 0.69 0.6 0.42 1.03 NaN NaN NaN -0.04 1.126400 RB8409_hypothetical protein 4495547 4495840 0.31 1.62 NaN NaN 1.5 NaN 0.99 1.19 0.76 1.54 -0.17 NaN NaN 1.22 1.06854 RB11421_60-kDa SS-A/Ro ribonucleoprotein 6195538 6193880 NaN 0.54 2.55 2.32 0.99 NaN NaN NaN NaN 0.27 NaN NaN NaN NaN NaN
2062 RB1449_hypothetical protein 736500 736697 NaN 0.94 1.66 1.91 1.26 NaN 1.26 0.82 0.14 0.3 NaN NaN NaN 1.02 1.302137 RB1573_hypothetical protein 805094 804984 0.47 1.41 0.29 2.22 2.19 NaN 1.24 NaN NaN 1.43 NaN NaN NaN NaN NaN2211 RB171_ISxac3 transposase 89626 90504 0.41 1.27 1.28 1.82 1.49 NaN 1.55 0.95 1.12 0.7 NaN NaN NaN 0.44 1.292355 RB1977_mRNA 3'-end processing factor 1039488 1040645 1.14 1.85 1.55 2.05 1.56 0.78 1.09 0.76 0.49 0.86 0.24 NaN -0.17 0.68 0.652848 RB2745_hypothetical protein 1431275 1431400 0.45 1.27 1.58 1.57 0.87 0.58 0.98 0.77 0.63 1.57 1.30 0.33 NaN 0.95 0.872862 RB2774_conserved hypothetical protein 1443786 1444034 1.13 1.88 1.94 1.51 1.89 0.4 0.59 0.87 0.45 1.28 NaN NaN NaN -0.19 0.682982 RB2963_hypothetical protein 1526707 1527066 0.78 2.31 1.96 1.79 1.89 0.76 0.5 0.61 0.54 1.24 0.37 0.12 NaN 0.63 -0.023860 RB4322_transposase and inactivated derivative 2226866 2228239 1.1 1.49 1.36 1.58 2.02 NaN NaN NaN NaN 0.29 NaN NaN NaN 0.46 NaN4227 RB4867_SmpB RNA-binding protein 2496354 2495722 0.89 1.35 2.52 2.58 1.87 0.17 0.64 0.5 0.32 0.57 0.27 NaN NaN 1.06 0.234365 RB5069_hypothetical protein 2609575 2609378 NaN 0.36 2.02 2.06 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN4405 RB5135_6-pyruvoyl tetrahydrobiopterin synthase. 2640266 2639853 1.26 1.66 1.73 2.19 2.15 0.28 0.85 0.45 0.12 0.86 0.61 NaN NaN 0.61 NaN5355 RB6685_hypothetical protein 3554265 3554047 1.03 2 1.84 1.95 1.85 NaN 0.92 0.74 0.24 0.55 NaN NaN NaN 1.07 0.535531 RB6973_tellurite resistance protein 3710584 3711420 1.11 1.56 1.79 2.07 2.67 NaN 0.76 0.53 0.17 0.64 NaN NaN NaN 0.17 NaN5571 RB7039_hypothetical protein 3743573 3742965 0.82 0.77 2.19 1.61 1.85 NaN NaN 0.55 NaN 0.15 NaN NaN NaN 2.01 1.185923 RB7612_hypothetical protein 4092907 4093317 NaN 0.82 2.82 1.89 1.27 NaN 0.98 0.27 0.52 0.64 NaN NaN NaN NaN 1.166420 RB8445_Penicillinase repressor 4514475 4514050 0.61 1.11 1.77 1.33 2.44 NaN 0.93 0.48 0.28 0.64 0.20 NaN NaN 1.26 1.956474 RB8533_protease I 4552172 4552762 0.85 1.33 1.55 1.57 1.08 NaN 0.87 NaN 0.6 1.46 0.81 NaN NaN 1.06 1.637308 RB9966_Fe-S oxidoreductase 5370040 5368655 NaN NaN NaN 1.58 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN1068 RB11802_Transposase IS3/IS911 6361950 6362240 0.49 1.17 1.73 1.75 2.02 NaN 1.5 0.8 1 0.56 0.14 NaN NaN 0.93 1.101476 RB12458_conserved hypothetical protein 6704719 6704976 NaN 0.79 2.67 1.95 0.92 NaN NaN NaN NaN 1.14 NaN NaN NaN NaN NaN2203 RB17_protein containing DUF1589 8251 7757 0.18 1.44 2.49 2.46 1.22 NaN NaN NaN 0.71 1.97 NaN NaN NaN NaN NaN2295 RB1862_transcriptional regulator, GntR family 960630 961037 NaN 0.75 1.85 1.33 NaN NaN NaN NaN NaN 1.46 NaN NaN NaN NaN NaN2457 RB2146_Exonuclease VII, small subunit 1136505 1136086 0.99 2.47 1.6 2.06 1.1 1.01 0.83 1.15 0.7 1.78 0.21 NaN 0.36 0.22 0.172914 RB286_hypothetical protein 142779 143096 NaN 0.89 2.02 2.14 NaN NaN NaN NaN NaN 0.57 NaN NaN NaN NaN NaN3099 RB3159_protein containing DUF1590 1629337 1629116 1.62 1.17 1.53 1.47 1.45 0.16 0.72 0.99 0.8 1.16 1.09 NaN NaN 0.61 1.665328 RB6637_hypothetical protein 3528317 3528469 0.59 1.72 1.63 2.34 0.9 NaN 1.68 1.37 0.92 1 NaN NaN NaN NaN NaN6349 RB8325_hypothetical protein 4447029 4446139 0.19 1.28 2.36 1.53 2.14 NaN 1.72 1.11 0.89 1.22 NaN NaN NaN NaN NaN6929 RB933_hypothetical protein 493451 492825 NaN 1.01 2.1 1.94 NaN NaN NaN NaN NaN 0.68 NaN NaN NaN NaN NaN7079 RB957_Tyrosine recombinase xerD 505476 504889 NaN 1.04 1.64 2.07 NaN NaN NaN NaN NaN 0.83 NaN NaN NaN NaN NaN7324 RB9998_ABC transporter ATP-binding protein 5385937 5385104 0.95 1.33 1.57 1.23 1.28 0.65 0.96 1.14 1.53 1.5 0.63 -0.25 NaN 0.47 0.89
91 RB10137_protein containing DUF1590 5469772 5469551 0.86 1.66 2.03 2.31 2.03 NaN 1.34 0.79 0.58 0.87 NaN NaN NaN 0.03 2.12640 RB11077_Protein of unknown function, UPF0074 6006371 6005904 0.4 0.93 1.34 1.17 1.36 0.53 1.26 1 1.02 0.87 0.61 NaN NaN 0.55 1.99
1162 RB11952_secreted protein 6438438 6438043 NaN 0.76 2.7 2.36 1.28 NaN NaN NaN NaN 0.61 NaN NaN NaN NaN NaN1689 RB12806_conserved hypothetical protein 6911165 6910548 NaN 0.99 1.27 2.06 2.3 NaN NaN NaN NaN 0.62 NaN NaN NaN NaN NaN1913 RB13190_conserved hypothetical protein 7083077 7083229 0.19 1.18 1.23 1.98 0.71 NaN 1.75 1.37 1.06 0.99 1.02 NaN NaN NaN NaN3673 RB402_hypothetical protein 212820 213275 0.47 1.13 2.33 2.63 2.14 1.29 1.23 1.23 0.47 1.35 NaN NaN NaN 0.19 0.893862 RB4325_Planctomycete PGAMP 2228477 2228623 NaN 1.15 0.96 1.99 2.3 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN4849 RB5848_dnaK suppressor protein, 3066328 3065906 1.75 1.67 1.84 1.79 2.39 0.65 0.81 0.98 0.84 1.04 0.45 -0.61 NaN 0.20 0.675055 RB6207_conserved hypothetical protein, secreted 3252449 3253759 1.65 2.03 1.44 1.59 1.44 0.83 1.08 0.99 0.82 1.19 0.64 NaN -0.17 1.15 0.925211 RB6449_myosin-like membrane protein 3400438 3406236 NaN 1.44 1.41 2.28 0.88 NaN NaN NaN NaN 2.15 NaN NaN NaN NaN NaN6385 RB8384_ISXo8 transposase 4478623 4479924 0.83 1.3 2.1 1.79 1.61 NaN NaN NaN 1.6 1.02 NaN NaN NaN NaN NaN6543 RB865_hypothetical protein 458731 460359 0.99 1.16 1.21 1.49 1.47 0.64 1.52 1.31 1.34 1.9 0.63 0.27 NaN 0.47 1.697277 RB9908_hypothetical protein 5339497 5339649 0.41 1.33 1.8 2.42 1.23 NaN 1.79 1.61 1.1 1.17 NaN NaN NaN NaN NaN952 RB11603_conserved hypothetical protein 6256722 6256835 2.03 1.41 1.76 2.09 2.41 0.32 0.96 0.74 0.65 0.17 0.68 0.12 0.03 1.48 1.11
1538 RB1256_hypothetical protein 641044 640889 NaN 1.77 1.64 2.66 1.57 NaN NaN NaN NaN 0.2 NaN NaN NaN NaN NaN1641 RB12732_secreted protein containing DUF1501 6862068 6860800 0.1 1.12 1.5 1.61 3.01 NaN 1.62 0.99 1.3 0.91 NaN NaN NaN NaN 1.82
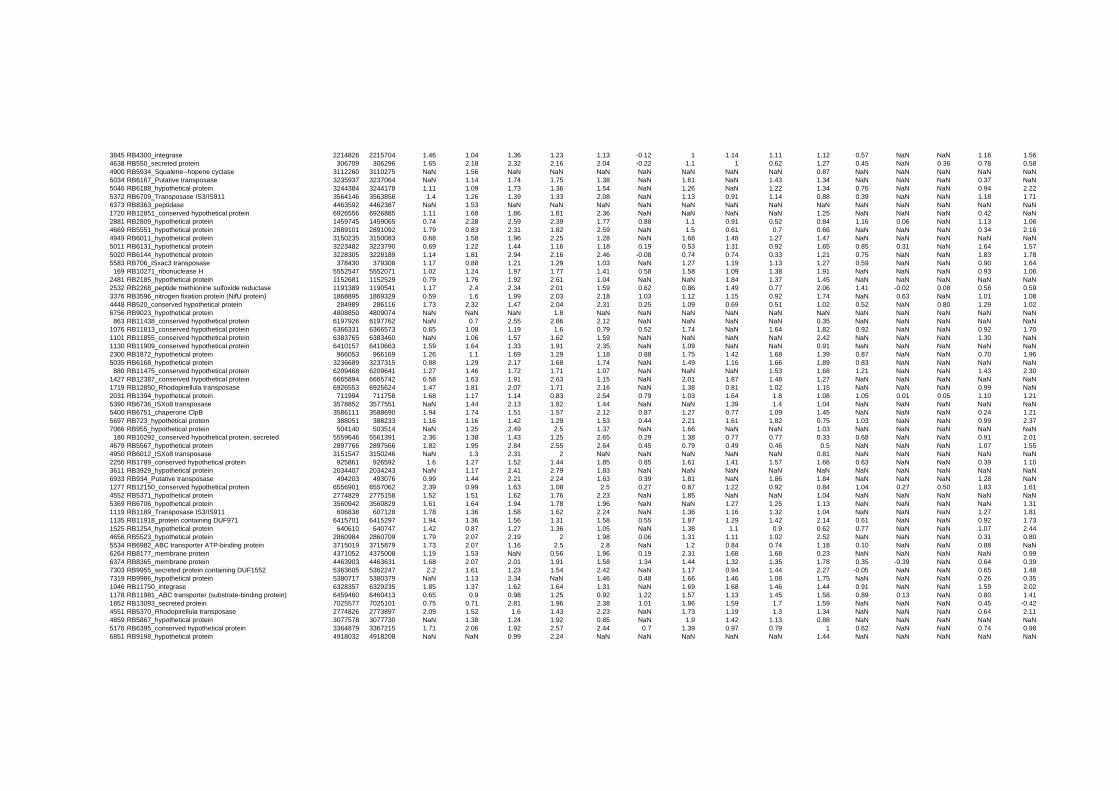
3845 RB4300_integrase 2214826 2215704 1.46 1.04 1.36 1.23 1.13 -0.12 1 1.14 1.11 1.12 0.57 NaN NaN 1.16 1.564638 RB550_secreted protein 306709 306296 1.65 2.18 2.32 2.16 2.04 -0.22 1.1 1 0.62 1.27 0.45 NaN 0.36 0.78 0.584900 RB5934_Squalene--hopene cyclase 3112260 3110275 NaN 1.56 NaN NaN NaN NaN NaN NaN NaN 0.87 NaN NaN NaN NaN NaN5034 RB6167_Putative transposase 3235937 3237064 NaN 1.14 1.74 1.75 1.38 NaN 1.61 NaN 1.43 1.34 NaN NaN NaN 0.37 NaN5046 RB6188_hypothetical protein 3244384 3244178 1.11 1.09 1.73 1.36 1.54 NaN 1.26 NaN 1.22 1.34 0.76 NaN NaN 0.94 2.225372 RB6709_Transposase IS3/IS911 3564146 3563856 1.4 1.26 1.39 1.33 2.08 NaN 1.13 0.91 1.14 0.88 0.39 NaN NaN 1.18 1.716373 RB8363_peptidase 4463592 4462387 NaN 1.53 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN1720 RB12851_conserved hypothetical protein 6926556 6926885 1.11 1.68 1.86 1.81 2.36 NaN NaN NaN NaN 1.25 NaN NaN NaN 0.42 NaN2881 RB2809_hypothetical protein 1459745 1459065 0.74 2.28 2.59 2.39 1.77 0.88 1.1 0.91 0.52 0.84 1.16 0.06 NaN 1.13 1.064669 RB5551_hypothetical protein 2889101 2891092 1.79 0.83 2.31 1.82 2.59 NaN 1.5 0.61 0.7 0.66 NaN NaN NaN 0.34 2.164949 RB6011_hypothetical protein 3150235 3150083 0.68 1.58 1.96 2.25 1.28 NaN 1.68 1.48 1.27 1.47 NaN NaN NaN NaN NaN5011 RB6131_hypothetical protein 3223482 3223790 0.69 1.22 1.44 1.16 1.18 0.19 0.53 1.31 0.92 1.65 0.85 0.31 NaN 1.64 1.575020 RB6144_hypothetical protein 3228305 3228189 1.14 1.81 2.94 2.16 2.46 -0.08 0.74 0.74 0.33 1.21 0.75 NaN NaN 1.83 1.785583 RB706_ISxac3 transposase 378430 379308 1.17 0.88 1.21 1.29 1.03 NaN 1.27 1.19 1.13 1.27 0.59 NaN NaN 0.90 1.64169 RB10271_ribonuclease H 5552547 5552071 1.02 1.24 1.97 1.77 1.41 0.58 1.58 1.09 1.38 1.91 NaN NaN NaN 0.93 1.06
2481 RB2185_hypothetical protein 1152681 1152529 0.79 1.76 1.92 2.61 1.04 NaN NaN 1.84 1.37 1.45 NaN NaN NaN NaN NaN2532 RB2268_peptide methionine sulfoxide reductase 1191389 1190541 1.17 2.4 2.34 2.01 1.59 0.62 0.86 1.49 0.77 2.06 1.41 -0.02 0.08 0.58 0.593376 RB3596_nitrogen fixation protein (NifU protein) 1868895 1869329 0.59 1.6 1.99 2.03 2.18 1.03 1.12 1.15 0.92 1.74 NaN 0.63 NaN 1.01 1.084448 RB520_conserved hypothetical protein 284989 286116 1.73 2.32 1.47 2.04 2.31 0.25 1.09 0.69 0.51 1.02 0.52 NaN 0.80 1.29 1.026756 RB9023_hypothetical protein 4808850 4809074 NaN NaN NaN 1.8 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN863 RB11438_conserved hypothetical protein 6197926 6197762 NaN 0.7 2.55 2.86 2.12 NaN NaN NaN NaN 0.35 NaN NaN NaN NaN NaN
1076 RB11813_conserved hypothetical protein 6366331 6366573 0.65 1.08 1.19 1.6 0.79 0.52 1.74 NaN 1.64 1.82 0.92 NaN NaN 0.92 1.701101 RB11855_conserved hypothetical protein 6383765 6383460 NaN 1.06 1.57 1.62 1.59 NaN NaN NaN NaN 2.42 NaN NaN NaN 1.30 NaN1130 RB11909_conserved hypothetical protein 6410157 6410663 1.59 1.64 1.33 1.91 2.35 NaN 1.09 NaN NaN 0.91 NaN NaN NaN NaN NaN2300 RB1872_hypothetical protein 966053 966169 1.26 1.1 1.69 1.29 1.18 0.88 1.75 1.42 1.68 1.39 0.87 NaN NaN 0.70 1.965035 RB6168_hypothetical protein 3236689 3237315 0.88 1.29 2.17 1.68 1.74 NaN 1.49 1.16 1.66 1.89 0.83 NaN NaN NaN NaN880 RB11475_conserved hypothetical protein 6209468 6209641 1.27 1.46 1.72 1.71 1.07 NaN NaN NaN 1.53 1.68 1.21 NaN NaN 1.43 2.30
1427 RB12387_conserved hypothetical protein 6665894 6665742 0.58 1.63 1.91 2.63 1.15 NaN 2.01 1.87 1.48 1.27 NaN NaN NaN NaN NaN1719 RB12850_Rhodopirellula transposase 6926553 6925624 1.47 1.81 2.07 1.71 2.16 NaN 1.38 0.81 1.02 1.15 NaN NaN NaN 0.99 NaN2031 RB1394_hypothetical protein 711994 711758 1.68 1.17 1.14 0.83 2.54 0.79 1.03 1.64 1.8 1.08 1.05 0.01 0.05 1.10 1.215390 RB6736_ISXo8 transposase 3578852 3577551 NaN 1.44 2.13 1.82 1.44 NaN NaN 1.39 1.4 1.04 NaN NaN NaN NaN NaN5400 RB6751_chaperone ClpB 3586111 3588690 1.94 1.74 1.51 1.57 2.12 0.87 1.27 0.77 1.09 1.45 NaN NaN NaN 0.24 1.215697 RB723_hypothetical protein 388051 388233 1.16 1.16 1.42 1.29 1.53 0.44 2.21 1.61 1.82 0.75 1.03 NaN NaN 0.99 2.377066 RB955_hypothetical protein 504140 503514 NaN 1.25 2.49 2.5 1.37 NaN 1.66 NaN NaN 1.03 NaN NaN NaN NaN NaN180 RB10292_conserved hypothetical protein, secreted 5559646 5561391 2.36 1.38 1.43 1.25 2.65 0.29 1.38 0.77 0.77 0.33 0.68 NaN NaN 0.91 2.01
4679 RB5567_hypothetical protein 2897766 2897566 1.82 1.95 2.84 2.55 2.64 0.45 0.79 0.49 0.46 0.5 NaN NaN NaN 1.07 1.554950 RB6012_ISXo8 transposase 3151547 3150246 NaN 1.3 2.31 2 NaN NaN NaN NaN NaN 0.81 NaN NaN NaN NaN NaN2256 RB1789_conserved hypothetical protein 925861 926592 1.6 1.27 1.52 1.44 1.85 0.85 1.61 1.41 1.57 1.66 0.63 NaN NaN 0.39 1.103611 RB3929_hypothetical protein 2034407 2034243 NaN 1.17 2.41 2.79 1.83 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN6933 RB934_Putative transposase 494203 493076 0.99 1.44 2.21 2.24 1.63 0.39 1.81 NaN 1.86 1.84 NaN NaN NaN 1.28 NaN1277 RB12150_conserved hypothetical protein 6556901 6557062 2.39 0.99 1.63 1.08 2.5 0.27 0.87 1.22 0.92 0.84 1.04 0.27 0.50 1.83 1.614552 RB5371_hypothetical protein 2774829 2775158 1.52 1.51 1.62 1.76 2.23 NaN 1.85 NaN NaN 1.04 NaN NaN NaN NaN NaN5369 RB6706_hypothetical protein 3560942 3560829 1.61 1.64 1.94 1.78 1.96 NaN NaN 1.27 1.25 1.13 NaN NaN NaN NaN 1.311119 RB1189_Transposase IS3/IS911 606838 607128 1.78 1.36 1.58 1.62 2.24 NaN 1.36 1.16 1.32 1.04 NaN NaN NaN 1.27 1.811135 RB11918_protein containing DUF971 6415701 6415297 1.94 1.36 1.56 1.31 1.58 0.55 1.87 1.29 1.42 2.14 0.61 NaN NaN 0.92 1.731525 RB1254_hypothetical protein 640610 640747 1.42 0.87 1.27 1.36 1.05 NaN 1.38 1.1 0.9 0.62 0.77 NaN NaN 1.07 2.444656 RB5523_hypothetical protein 2860984 2860709 1.79 2.07 2.19 2 1.98 0.06 1.31 1.11 1.02 2.52 NaN NaN NaN 0.31 0.805534 RB6982_ABC transporter ATP-binding protein 3715019 3715879 1.73 2.07 1.16 2.5 2.8 NaN 1.2 0.84 0.74 1.18 0.10 NaN NaN 0.88 NaN6264 RB8177_membrane protein 4371052 4375008 1.19 1.53 NaN 0.56 1.96 0.19 2.31 1.68 1.68 0.23 NaN NaN NaN NaN 0.996374 RB8365_membrane protein 4463903 4463631 1.68 2.07 2.01 1.91 1.58 1.34 1.44 1.32 1.35 1.78 0.35 -0.39 NaN 0.64 0.397303 RB9955_secreted protein containing DUF1552 5363605 5362247 2.2 1.61 1.23 1.54 2.42 NaN 1.17 0.94 1.44 2.27 -0.05 NaN NaN 0.65 1.487319 RB9986_hypothetical protein 5380717 5380379 NaN 1.13 2.34 NaN 1.46 0.48 1.66 1.46 1.08 1.75 NaN NaN NaN 0.26 0.351046 RB11750_integrase 6328357 6329235 1.85 1.37 1.62 1.64 1.31 NaN 1.69 1.68 1.46 1.44 0.91 NaN NaN 1.59 2.021178 RB11981_ABC transporter (substrate-binding protein) 6459460 6460413 0.65 0.9 0.98 1.25 0.92 1.22 1.57 1.13 1.45 1.58 0.89 0.13 NaN 0.80 1.411852 RB13093_secreted protein 7025577 7025101 0.75 0.71 2.81 1.96 2.38 1.01 1.86 1.59 1.7 1.59 NaN NaN NaN 0.45 -0.424551 RB5370_Rhodopirellula transposase 2774826 2773897 2.09 1.52 1.6 1.43 2.23 NaN 1.73 1.19 1.3 1.34 NaN NaN NaN 0.64 2.114859 RB5867_hypothetical protein 3077578 3077730 NaN 1.38 1.24 1.92 0.85 NaN 1.9 1.42 1.13 0.88 NaN NaN NaN NaN NaN5176 RB6395_conserved hypothetical protein 3364879 3367215 1.71 2.06 1.92 2.57 2.44 0.7 1.39 0.97 0.79 1 0.62 NaN NaN 0.74 0.986851 RB9198_hypothetical protein 4918032 4918208 NaN NaN 0.99 2.24 NaN NaN NaN NaN NaN 1.44 NaN NaN NaN NaN NaN

1124 RB1190_integrase 607131 608009 1.49 0.98 0.98 1.36 1.31 0.09 1.29 1.32 1.33 0.84 0.76 NaN NaN 1.51 1.745323 RB663_hypothetical protein 357608 357369 1.71 2.53 1.69 2.53 2.32 0.58 0.86 0.71 0.7 0.84 1.24 0.49 0.32 1.78 0.951175 RB11977_conserved hypothetical protein 6455958 6456548 1.94 1.5 1.06 1.13 2.45 NaN 1.17 1.41 1.63 1.39 1.79 0.35 0.17 1.49 0.391690 RB12808_secreted protein 6912350 6911241 NaN 1.48 2.14 2.29 2.46 NaN 1.57 0.83 NaN 0.71 NaN NaN NaN NaN 1.58697 RB11176_protein containing DUF442 6065605 6065144 1.86 1.3 1.31 1.03 1.68 1.47 2.33 1.86 2.06 1.26 0.54 -0.05 0.84 0.70 1.63960 RB11618_type I restriction-modification system, M subunit, 6263708 6265366 NaN 1.36 2.25 2.3 2.42 NaN NaN NaN NaN 0.62 NaN NaN NaN NaN NaN
5572 RB704_Transposase IS3/IS911 378137 378427 1.75 1.52 1.89 1.73 2.26 1.47 1.49 1.14 1.36 0.87 0.78 NaN NaN 1.33 1.83574 RB10966_Protein of unknown function, UPF0057 5942934 5942752 1.13 1.54 1.07 1.44 1.22 1.43 1.67 1.45 1.62 0.97 0.80 0.53 0.45 0.99 0.15
5778 RB7389_Putative transposase 3953590 3954717 1.02 1.7 2.58 2.39 1.82 0.73 1.95 NaN 1.87 2.4 1.33 NaN NaN 1.18 NaN6940 RB9347_hypothetical protein 5008600 5008394 1 0.64 1.08 0.95 1.24 0.64 1.24 1.52 1.39 1.67 1.23 NaN NaN 0.71 1.876469 RB8527_protein containing DUF1559 4549097 4550089 1.09 0.6 1.26 1.4 1.54 2.15 2.54 2.05 2.08 0.75 0.88 0.11 NaN 0.50 1.901685 RB12800_conserved hypothetical protein 6908240 6908710 0.56 1.08 1.16 1.47 1.3 0.89 1.61 NaN NaN 1.92 NaN NaN NaN NaN NaN6454 RB8505_serine/threonine protein kinase related protein 4538446 4537127 0.89 0.8 1.53 1.13 0.82 1.12 1.57 1.56 1.56 1.12 1.39 0.11 0.55 0.14 0.52
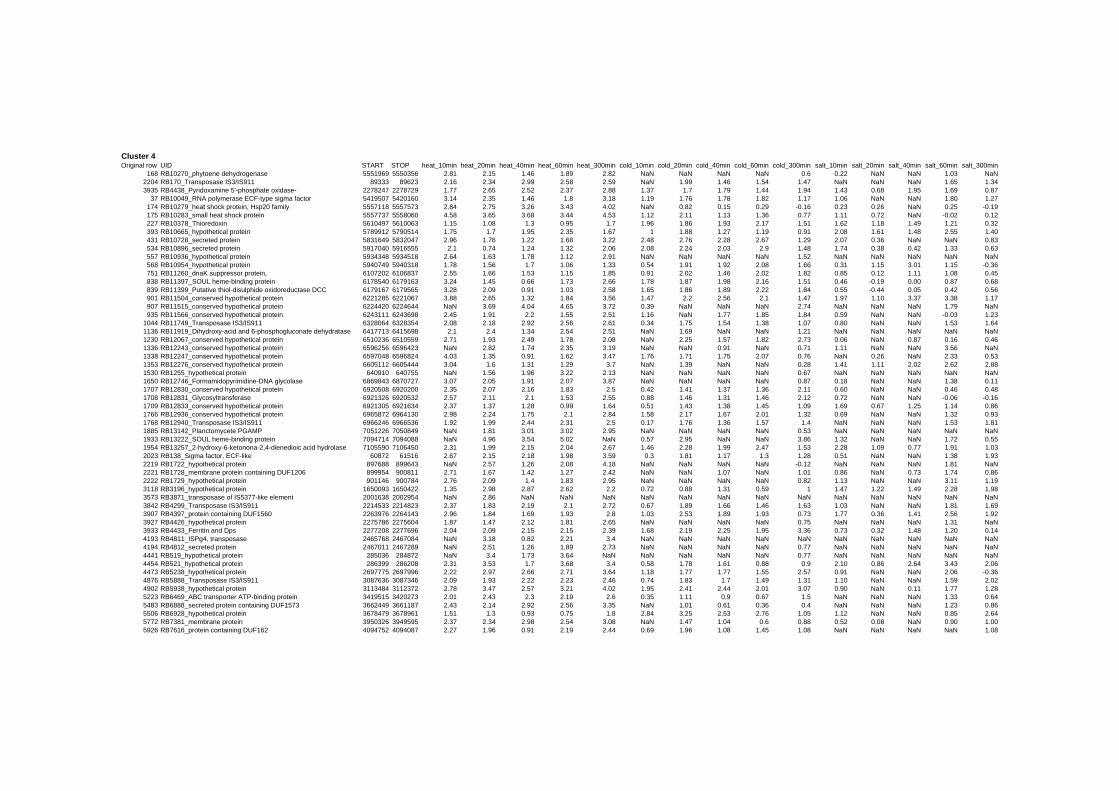
Cluster 4Original row UID START STOP heat_10min heat_20min heat_40min heat_60min heat_300min cold_10min cold_20min cold_40min cold_60min cold_300min salt_10min salt_20min salt_40min salt_60min salt_300min
168 RB10270_phytoene dehydrogenase 5551969 5550356 2.81 2.15 1.46 1.89 2.82 NaN NaN NaN NaN 0.6 0.22 NaN NaN 1.03 NaN2204 RB170_Transposase IS3/IS911 89333 89623 2.16 2.34 2.99 2.58 2.59 NaN 1.99 1.46 1.54 1.47 NaN NaN NaN 1.65 1.343935 RB4438_Pyridoxamine 5'-phosphate oxidase- 2278247 2278729 1.77 2.65 2.52 2.37 2.88 1.37 1.7 1.79 1.44 1.94 1.43 0.68 1.95 1.69 0.87
37 RB10049_RNA polymerase ECF-type sigma factor 5419507 5420160 3.14 2.35 1.46 1.8 3.18 1.19 1.76 1.78 1.82 1.17 1.06 NaN NaN 1.80 1.27174 RB10279_heat shock protein, Hsp20 family 5557118 5557573 2.84 2.75 3.26 3.43 4.02 NaN 0.82 0.15 0.29 -0.16 0.23 0.26 NaN 0.25 -0.19175 RB10283_small heat shock protein 5557737 5558060 4.58 3.65 3.68 3.44 4.53 1.12 2.11 1.13 1.36 0.77 1.11 0.72 NaN -0.02 0.12227 RB10378_Thioredoxin 5610497 5610063 1.15 1.08 1.3 0.95 1.7 1.96 1.86 1.93 2.17 1.51 1.62 1.18 1.49 1.21 0.32393 RB10665_hypothetical protein 5789912 5790514 1.75 1.7 1.95 2.35 1.67 1 1.88 1.27 1.19 0.91 2.08 1.61 1.48 2.55 1.40431 RB10728_secreted protein 5831649 5832047 2.96 1.76 1.22 1.68 3.22 2.48 2.76 2.28 2.67 1.29 2.07 0.36 NaN NaN 0.83534 RB10896_secreted protein 5917040 5916555 2.1 0.74 1.24 1.32 2.06 2.08 2.24 2.03 2.9 1.48 1.74 0.38 0.42 1.33 0.63557 RB10936_hypothetical protein 5934348 5934518 2.64 1.63 1.78 1.12 2.91 NaN NaN NaN NaN 1.52 NaN NaN NaN NaN NaN568 RB10954_hypothetical protein 5940749 5940318 1.78 1.56 1.7 1.06 1.33 0.54 1.91 1.92 2.08 1.66 0.31 1.15 3.01 1.15 -0.36751 RB11260_dnaK suppressor protein, 6107202 6106837 2.55 1.66 1.53 1.15 1.85 0.91 2.02 1.46 2.02 1.82 0.85 0.12 1.11 1.08 0.45838 RB11397_SOUL heme-binding protein 6178540 6179163 3.24 1.45 0.66 1.73 2.66 1.78 1.87 1.98 2.16 1.51 0.46 -0.19 0.00 0.87 0.68839 RB11399_Putative thiol-disulphide oxidoreductase DCC 6179167 6179565 3.28 2.09 0.91 1.03 2.58 1.65 1.86 1.89 2.22 1.84 0.55 -0.44 0.05 0.42 0.56901 RB11504_conserved hypothetical protein 6221285 6221067 3.88 2.65 1.32 1.84 3.56 1.47 2.2 2.56 2.1 1.47 1.97 1.10 3.37 3.38 1.17907 RB11515_conserved hypothetical protein 6224420 6224644 NaN 3.69 4.04 4.65 3.72 0.39 NaN NaN NaN 2.74 NaN NaN NaN 1.79 NaN935 RB11566_conserved hypothetical protein 6243111 6243698 2.45 1.91 2.2 1.55 2.51 1.16 NaN 1.77 1.85 1.84 0.59 NaN NaN -0.03 1.23
1044 RB11749_Transposase IS3/IS911 6328064 6328354 2.08 2.18 2.92 2.56 2.61 0.34 1.75 1.54 1.38 1.07 0.80 NaN NaN 1.53 1.641136 RB11919_Dihydroxy-acid and 6-phosphogluconate dehydratase 6417713 6415698 2.1 2.4 1.34 2.54 2.51 NaN 1.69 NaN NaN 1.21 NaN NaN NaN NaN NaN1230 RB12067_conserved hypothetical protein 6510236 6510559 2.71 1.93 2.49 1.78 2.08 NaN 2.25 1.57 1.82 2.73 0.06 NaN 0.87 0.16 0.461336 RB12243_conserved hypothetical protein 6596256 6596423 NaN 2.82 1.74 2.35 3.19 NaN NaN 0.91 NaN 0.71 1.11 NaN NaN 3.56 NaN1338 RB12247_conserved hypothetical protein 6597048 6596824 4.03 1.35 0.91 1.62 3.47 1.76 1.71 1.75 2.07 0.76 NaN 0.26 NaN 2.33 0.531353 RB12276_conserved hypothetical protein 6605112 6605444 3.04 1.6 1.31 1.29 3.7 NaN 1.39 NaN NaN 0.28 1.41 1.11 2.02 2.62 2.881530 RB1255_hypothetical protein 640910 640755 NaN 1.56 1.96 3.22 2.13 NaN NaN NaN NaN 0.67 NaN NaN NaN NaN NaN1650 RB12746_Formamidopyrimidine-DNA glycolase 6869843 6870727 3.07 2.05 1.91 2.07 3.87 NaN NaN NaN NaN 0.87 0.18 NaN NaN 1.38 0.111707 RB12830_conserved hypothetical protein 6920508 6920200 2.35 2.07 2.16 1.83 2.5 0.42 1.41 1.37 1.36 2.11 0.60 NaN NaN 0.46 0.481708 RB12831_Glycosyltransferase 6921326 6920532 2.57 2.11 2.1 1.53 2.55 0.88 1.46 1.31 1.46 2.12 0.72 NaN NaN -0.06 -0.161709 RB12833_conserved hypothetical protein 6921305 6921634 2.37 1.37 1.28 0.99 1.64 0.51 1.43 1.38 1.45 1.09 1.69 0.67 1.25 1.14 0.861766 RB12936_conserved hypothetical protein 6965872 6964130 2.98 2.24 1.75 2.1 2.84 1.58 2.17 1.67 2.01 1.32 0.69 NaN NaN 1.32 0.931768 RB12940_Transposase IS3/IS911 6966246 6966536 1.92 1.99 2.44 2.31 2.5 0.17 1.76 1.36 1.57 1.4 NaN NaN NaN 1.53 1.811885 RB13142_Planctomycete PGAMP 7051226 7050849 NaN 1.81 3.01 3.02 2.95 NaN NaN NaN NaN 0.53 NaN NaN NaN NaN NaN1933 RB13222_SOUL heme-binding protein 7094714 7094088 NaN 4.96 3.54 5.02 NaN 0.57 2.95 NaN NaN 3.86 1.32 NaN NaN 1.72 0.551954 RB13257_2-hydroxy-6-ketonona-2,4-dienedioic acid hydrolase 7105590 7106450 2.31 1.99 2.15 2.04 2.67 1.46 2.28 1.99 2.47 1.53 2.28 1.09 0.77 1.91 1.032023 RB138_Sigma factor, ECF-like 60872 61516 2.67 2.15 2.18 1.98 3.59 0.3 1.81 1.17 1.3 1.28 0.51 NaN NaN 1.38 1.932219 RB1722_hypothetical protein 897688 899643 NaN 2.57 1.26 2.08 4.18 NaN NaN NaN NaN -0.12 NaN NaN NaN 1.81 NaN2221 RB1728_membrane protein containing DUF1206 899954 900811 2.71 1.67 1.42 1.27 2.42 NaN NaN 1.07 NaN 1.01 0.86 NaN 0.73 1.74 0.862222 RB1729_hypothetical protein 901146 900784 2.76 2.09 1.4 1.83 2.95 NaN NaN NaN NaN 0.82 1.13 NaN NaN 3.11 1.193118 RB3196_hypothetical protein 1650093 1650422 1.35 2.98 2.87 2.62 2.2 0.72 0.88 1.31 0.59 1 1.47 1.22 1.49 2.28 1.983573 RB3871_transposase of IS5377-like element 2001638 2002954 NaN 2.86 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN3842 RB4299_Transposase IS3/IS911 2214533 2214823 2.37 1.83 2.19 2.1 2.72 0.67 1.89 1.66 1.46 1.63 1.03 NaN NaN 1.81 1.693907 RB4397_protein containing DUF1560 2263976 2264143 2.96 1.84 1.69 1.93 2.8 1.03 2.53 1.89 1.93 0.73 1.77 0.36 1.41 2.56 1.923927 RB4426_hypothetical protein 2275786 2275604 1.87 1.47 2.12 1.81 2.65 NaN NaN NaN NaN 0.75 NaN NaN NaN 1.31 NaN3933 RB4433_Ferritin and Dps 2277208 2277696 2.04 2.09 2.15 2.15 2.39 1.68 2.19 2.25 1.95 3.36 0.73 0.32 1.48 1.20 0.144193 RB4811_ISPg4, transposase 2465768 2467084 NaN 3.18 0.82 2.21 3.4 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN4194 RB4812_secreted protein 2467011 2467289 NaN 2.51 1.26 1.89 2.73 NaN NaN NaN NaN 0.77 NaN NaN NaN NaN NaN4441 RB519_hypothetical protein 285036 284872 NaN 3.4 1.73 3.64 NaN NaN NaN NaN NaN 0.77 NaN NaN NaN NaN NaN4454 RB521_hypothetical protein 286399 286208 2.31 3.53 1.7 3.68 3.4 0.58 1.78 1.61 0.88 0.9 2.10 0.86 2.64 3.43 2.064473 RB5238_hypothetical protein 2697775 2697996 2.22 2.97 2.66 2.71 3.64 1.18 1.77 1.77 1.55 2.57 0.91 NaN NaN 2.06 -0.364876 RB5888_Transposase IS3/IS911 3087636 3087346 2.09 1.93 2.22 2.23 2.46 0.74 1.83 1.7 1.49 1.31 1.10 NaN NaN 1.59 2.024902 RB5938_hypothetical protein 3113484 3112372 2.78 3.47 2.57 3.21 4.02 1.95 2.41 2.44 2.01 3.07 0.90 NaN 0.11 1.77 1.285223 RB6469_ABC transporter ATP-binding protein 3419515 3420273 2.01 2.43 2.3 2.19 2.6 0.35 1.11 0.9 0.67 1.5 NaN NaN NaN 1.33 0.645483 RB6888_secreted protein containing DUF1573 3662449 3661187 2.43 2.14 2.92 2.56 3.35 NaN 1.01 0.61 0.36 0.4 NaN NaN NaN 1.23 0.865506 RB6928_hypothetical protein 3678479 3678961 1.51 1.3 0.93 0.75 1.8 2.84 3.25 2.53 2.76 1.05 1.12 NaN NaN 0.85 2.645772 RB7381_membrane protein 3950326 3949595 2.37 2.34 2.98 2.54 3.08 NaN 1.47 1.04 0.6 0.88 0.52 0.08 NaN 0.90 1.005926 RB7616_protein containing DUF162 4094752 4094087 2.27 1.96 0.91 2.19 2.44 0.69 1.96 1.08 1.45 1.08 NaN NaN NaN NaN 1.08

6247 RB8146_hypothetical protein 4355122 4354667 2.38 1.9 1.8 1.46 2.31 0.5 0.95 1.18 1.43 1.21 0.86 0.07 0.98 1.71 1.456475 RB8534_hypothetical protein 4552802 4552930 NaN NaN 1.79 2.24 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN6726 RB8969_Chaperonin Cpn10 4779554 4779859 3.13 1.42 1.02 2.03 2.8 NaN NaN NaN NaN 0 NaN NaN NaN NaN NaN6728 RB8970_GroEL 4779905 4781527 2.95 2.09 1.45 1.92 2.31 0.9 1.7 0.94 1.52 1.6 NaN NaN NaN 0.44 0.596732 RB8976_conserved hypothetical protein 4783568 4783846 NaN 2.49 NaN 3.55 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN6739 RB8987_hypothetical protein 4790856 4790710 NaN 2.38 2.4 3 2.86 NaN 1.93 NaN 1.51 1.27 NaN NaN NaN 1.13 NaN6798 RB9105_Heat shock protein Hsp70 4868834 4866720 3.47 2.63 1.82 2.12 3.28 0.27 1.39 0.44 0.99 0.62 0.54 NaN NaN 0.82 0.196870 RB9230_hypothetical protein 4936673 4936996 2.43 1.64 2.13 2.07 2.1 0.78 2.2 1.84 1.43 1.64 NaN NaN NaN 0.59 1.736879 RB924_hypothetical protein 490400 489774 NaN 1.89 3.08 2.52 NaN NaN NaN NaN NaN 1.11 NaN NaN NaN NaN NaN7203 RB9787_hypothetical protein 5270904 5270719 NaN 2.32 3.29 3.54 2.73 NaN NaN NaN NaN 0.99 NaN NaN NaN NaN NaN7221 RB9810_transposase of IS5377-like element 5283122 5284438 NaN 2.83 NaN 1.64 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN7222 RB9811_secreted protein 5284365 5284673 1.92 2.37 1.61 1.76 2.44 NaN 1.42 1.01 0.75 0.97 0.68 NaN NaN 2.79 1.617276 RB9907_ISXo8 transposase 5338185 5339486 1.44 1.83 1.92 1.7 1.83 NaN 1.74 1.79 1.46 1.23 1.58 NaN NaN 1.95 NaN7325 RB9999_conserved hypothetical protein 5386828 5385983 NaN 1.72 2.72 2.41 2.49 0.6 2.41 1.95 2.81 2.1 0.35 NaN NaN 1.25 1.35519 RB10877_membrane protein 5905128 5903767 2.42 2.15 1.6 2.35 2.78 NaN 1.1 0.58 0.34 1.55 NaN NaN NaN 0.26 0.08
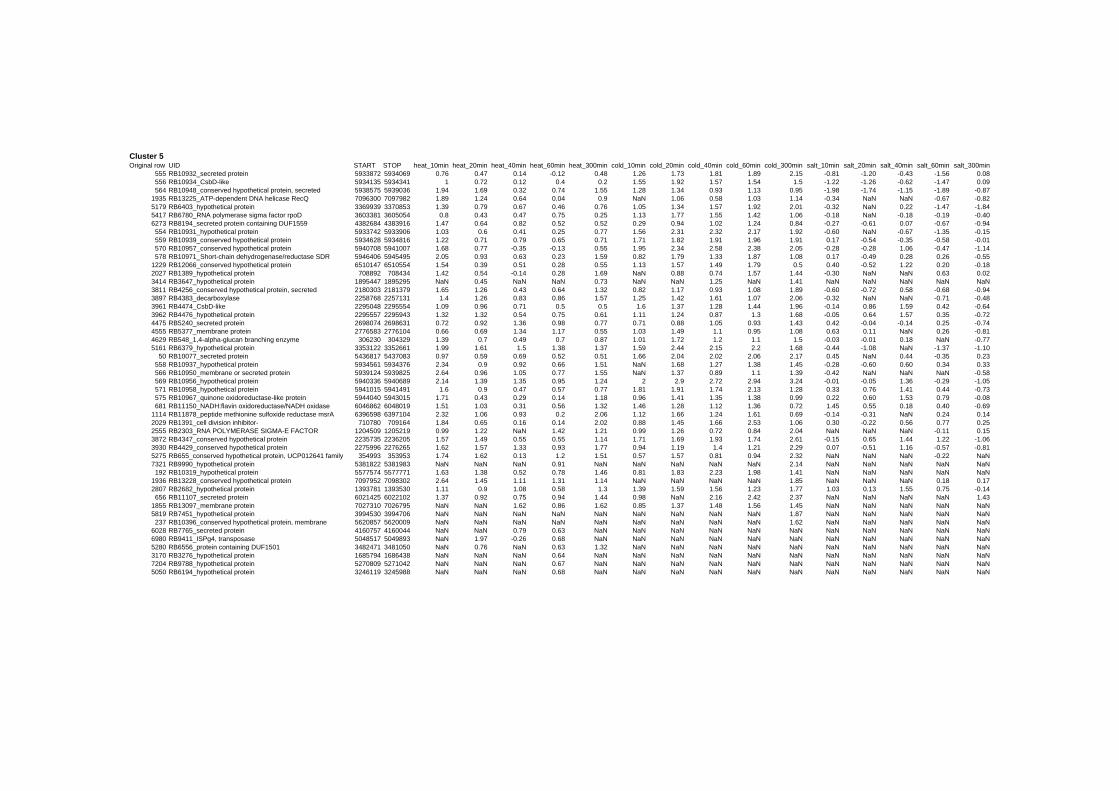
Cluster 5Original row UID START STOP heat_10min heat_20min heat_40min heat_60min heat_300min cold_10min cold_20min cold_40min cold_60min cold_300min salt_10min salt_20min salt_40min salt_60min salt_300min
555 RB10932_secreted protein 5933872 5934069 0.76 0.47 0.14 -0.12 0.48 1.26 1.73 1.81 1.89 2.15 -0.81 -1.20 -0.43 -1.56 0.08556 RB10934_CsbD-like 5934135 5934341 1 0.72 0.12 0.4 0.2 1.55 1.92 1.57 1.54 1.5 -1.22 -1.26 -0.62 -1.47 0.09564 RB10948_conserved hypothetical protein, secreted 5938575 5939036 1.94 1.69 0.32 0.74 1.55 1.28 1.34 0.93 1.13 0.95 -1.98 -1.74 -1.15 -1.89 -0.87
1935 RB13225_ATP-dependent DNA helicase RecQ 7096300 7097982 1.89 1.24 0.64 0.04 0.9 NaN 1.06 0.58 1.03 1.14 -0.34 NaN NaN -0.67 -0.825179 RB6403_hypothetical protein 3369939 3370853 1.39 0.79 0.67 0.46 0.76 1.05 1.34 1.57 1.92 2.01 -0.32 NaN 0.22 -1.47 -1.845417 RB6780_RNA polymerase sigma factor rpoD 3603381 3605054 0.8 0.43 0.47 0.75 0.25 1.13 1.77 1.55 1.42 1.06 -0.18 NaN -0.18 -0.19 -0.406273 RB8194_secreted protein containing DUF1559 4382684 4383916 1.47 0.64 0.82 0.52 0.52 0.29 0.94 1.02 1.24 0.84 -0.27 -0.61 0.07 -0.67 -0.94554 RB10931_hypothetical protein 5933742 5933906 1.03 0.6 0.41 0.25 0.77 1.56 2.31 2.32 2.17 1.92 -0.60 NaN -0.67 -1.35 -0.15559 RB10939_conserved hypothetical protein 5934628 5934816 1.22 0.71 0.79 0.65 0.71 1.71 1.82 1.91 1.96 1.91 0.17 -0.54 -0.35 -0.58 -0.01570 RB10957_conserved hypothetical protein 5940708 5941007 1.68 0.77 -0.35 -0.13 0.55 1.95 2.34 2.58 2.38 2.05 -0.28 -0.28 1.06 -0.47 -1.14578 RB10971_Short-chain dehydrogenase/reductase SDR 5946406 5945495 2.05 0.93 0.63 0.23 1.59 0.82 1.79 1.33 1.87 1.08 0.17 -0.49 0.28 0.26 -0.55
1229 RB12066_conserved hypothetical protein 6510147 6510554 1.54 0.39 0.51 0.28 0.55 1.13 1.57 1.49 1.79 0.5 0.40 -0.52 1.22 0.20 -0.182027 RB1389_hypothetical protein 708892 708434 1.42 0.54 -0.14 0.28 1.69 NaN 0.88 0.74 1.57 1.44 -0.30 NaN NaN 0.63 0.023414 RB3647_hypothetical protein 1895447 1895295 NaN 0.45 NaN NaN 0.73 NaN NaN 1.25 NaN 1.41 NaN NaN NaN NaN NaN3811 RB4256_conserved hypothetical protein, secreted 2180303 2181379 1.65 1.26 0.43 0.64 1.32 0.82 1.17 0.93 1.08 1.89 -0.60 -0.72 0.58 -0.68 -0.943897 RB4383_decarboxylase 2258768 2257131 1.4 1.26 0.83 0.86 1.57 1.25 1.42 1.61 1.07 2.06 -0.32 NaN NaN -0.71 -0.483961 RB4474_CsbD-like 2295048 2295554 1.09 0.96 0.71 0.5 0.5 1.6 1.37 1.28 1.44 1.96 -0.14 0.86 1.59 0.42 -0.643962 RB4476_hypothetical protein 2295557 2295943 1.32 1.32 0.54 0.75 0.61 1.11 1.24 0.87 1.3 1.68 -0.05 0.64 1.57 0.35 -0.724475 RB5240_secreted protein 2698074 2698631 0.72 0.92 1.36 0.98 0.77 0.71 0.88 1.05 0.93 1.43 0.42 -0.04 -0.14 0.25 -0.744555 RB5377_membrane protein 2776583 2776104 0.66 0.69 1.34 1.17 0.55 1.03 1.49 1.1 0.95 1.08 0.63 0.11 NaN 0.26 -0.814629 RB548_1,4-alpha-glucan branching enzyme 306230 304329 1.39 0.7 0.49 0.7 0.87 1.01 1.72 1.2 1.1 1.5 -0.03 -0.01 0.18 NaN -0.775161 RB6379_hypothetical protein 3353122 3352661 1.99 1.61 1.5 1.38 1.37 1.59 2.44 2.15 2.2 1.68 -0.44 -1.08 NaN -1.37 -1.10
50 RB10077_secreted protein 5436817 5437083 0.97 0.59 0.69 0.52 0.51 1.66 2.04 2.02 2.06 2.17 0.45 NaN 0.44 -0.35 0.23558 RB10937_hypothetical protein 5934561 5934376 2.34 0.9 0.92 0.66 1.51 NaN 1.68 1.27 1.38 1.45 -0.28 -0.60 0.60 0.34 0.33566 RB10950_membrane or secreted protein 5939124 5939825 2.64 0.96 1.05 0.77 1.55 NaN 1.37 0.89 1.1 1.39 -0.42 NaN NaN NaN -0.58569 RB10956_hypothetical protein 5940336 5940689 2.14 1.39 1.35 0.95 1.24 2 2.9 2.72 2.94 3.24 -0.01 -0.05 1.36 -0.29 -1.05571 RB10958_hypothetical protein 5941015 5941491 1.6 0.9 0.47 0.57 0.77 1.81 1.91 1.74 2.13 1.28 0.33 0.76 1.41 0.44 -0.73575 RB10967_quinone oxidoreductase-like protein 5944040 5943015 1.71 0.43 0.29 0.14 1.18 0.96 1.41 1.35 1.38 0.99 0.22 0.60 1.53 0.79 -0.08681 RB11150_NADH:flavin oxidoreductase/NADH oxidase 6046862 6048019 1.51 1.03 0.31 0.56 1.32 1.46 1.28 1.12 1.36 0.72 1.45 0.55 0.18 0.40 -0.69
1114 RB11878_peptide methionine sulfoxide reductase msrA 6396598 6397104 2.32 1.06 0.93 0.2 2.06 1.12 1.66 1.24 1.61 0.69 -0.14 -0.31 NaN 0.24 0.142029 RB1391_cell division inhibitor- 710780 709164 1.84 0.65 0.16 0.14 2.02 0.88 1.45 1.66 2.53 1.06 0.30 -0.22 0.56 0.77 0.252555 RB2303_RNA POLYMERASE SIGMA-E FACTOR 1204509 1205219 0.99 1.22 NaN 1.42 1.21 0.99 1.26 0.72 0.84 2.04 NaN NaN NaN -0.11 0.153872 RB4347_conserved hypothetical protein 2235735 2236205 1.57 1.49 0.55 0.55 1.14 1.71 1.69 1.93 1.74 2.61 -0.15 0.65 1.44 1.22 -1.063930 RB4429_conserved hypothetical protein 2275996 2276265 1.62 1.57 1.33 0.93 1.77 0.94 1.19 1.4 1.21 2.29 0.07 -0.51 1.16 -0.57 -0.815275 RB655_conserved hypothetical protein, UCP012641 family 354993 353953 1.74 1.62 0.13 1.2 1.51 0.57 1.57 0.81 0.94 2.32 NaN NaN NaN -0.22 NaN7321 RB9990_hypothetical protein 5381822 5381983 NaN NaN NaN 0.91 NaN NaN NaN NaN NaN 2.14 NaN NaN NaN NaN NaN192 RB10319_hypothetical protein 5577574 5577771 1.63 1.38 0.52 0.78 1.46 0.81 1.83 2.23 1.98 1.41 NaN NaN NaN NaN NaN
1936 RB13228_conserved hypothetical protein 7097952 7098302 2.64 1.45 1.11 1.31 1.14 NaN NaN NaN NaN 1.85 NaN NaN NaN 0.18 0.172807 RB2682_hypothetical protein 1393781 1393530 1.11 0.9 1.08 0.58 1.3 1.39 1.59 1.56 1.23 1.77 1.03 0.13 1.55 0.75 -0.14656 RB11107_secreted protein 6021425 6022102 1.37 0.92 0.75 0.94 1.44 0.98 NaN 2.16 2.42 2.37 NaN NaN NaN NaN 1.43
1855 RB13097_membrane protein 7027310 7026795 NaN NaN 1.62 0.86 1.62 0.85 1.37 1.48 1.56 1.45 NaN NaN NaN NaN NaN5819 RB7451_hypothetical protein 3994530 3994706 NaN NaN NaN NaN NaN NaN NaN NaN NaN 1.87 NaN NaN NaN NaN NaN237 RB10396_conserved hypothetical protein, membrane 5620857 5620009 NaN NaN NaN NaN NaN NaN NaN NaN NaN 1.62 NaN NaN NaN NaN NaN
6028 RB7765_secreted protein 4160757 4160044 NaN NaN 0.79 0.63 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN6980 RB9411_ISPg4, transposase 5048517 5049893 NaN 1.97 -0.26 0.68 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN5280 RB6556_protein containing DUF1501 3482471 3481050 NaN 0.76 NaN 0.63 1.32 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN3170 RB3276_hypothetical protein 1685794 1686438 NaN NaN NaN 0.64 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN7204 RB9788_hypothetical protein 5270809 5271042 NaN NaN NaN 0.67 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN5050 RB6194_hypothetical protein 3246119 3245988 NaN NaN NaN 0.68 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN
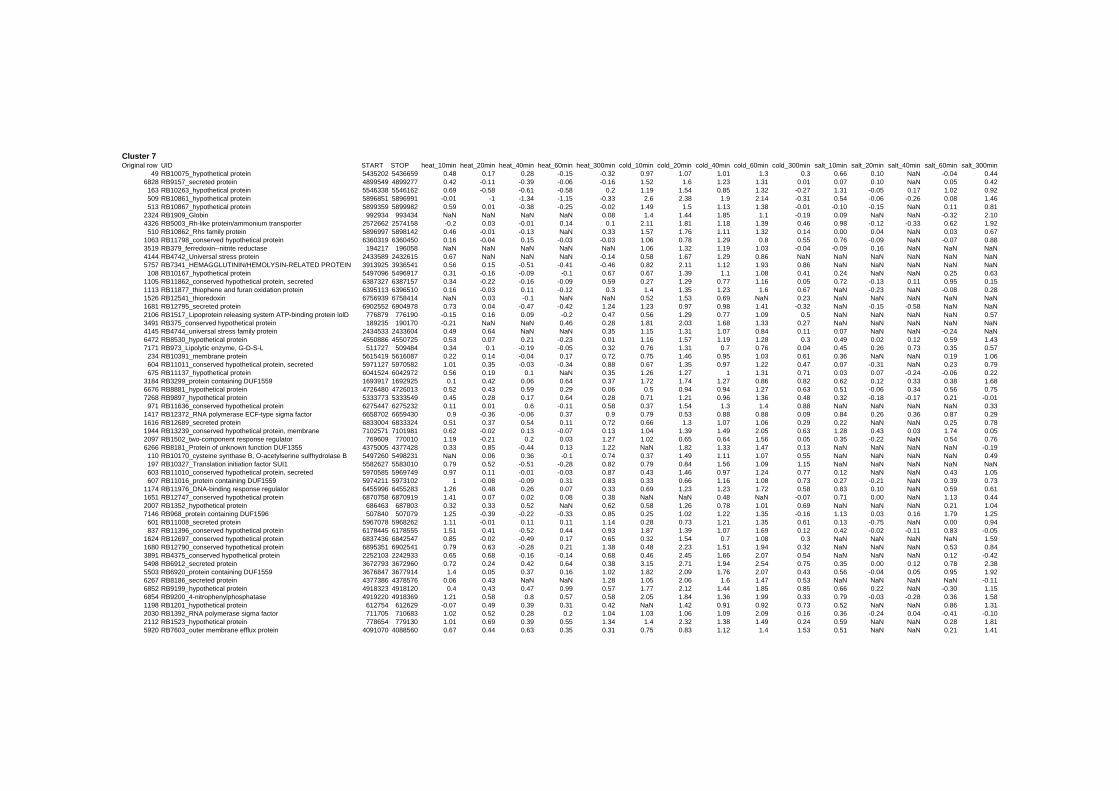
Cluster 7Original row UID START STOP heat_10min heat_20min heat_40min heat_60min heat_300min cold_10min cold_20min cold_40min cold_60min cold_300min salt_10min salt_20min salt_40min salt_60min salt_300min
49 RB10075_hypothetical protein 5435202 5436659 0.48 0.17 0.28 -0.15 -0.32 0.97 1.07 1.01 1.3 0.3 0.66 0.10 NaN -0.04 0.446828 RB9157_secreted protein 4899549 4899277 0.42 -0.11 -0.39 -0.06 -0.16 1.52 1.6 1.23 1.31 0.01 0.07 0.10 NaN 0.05 0.42163 RB10263_hypothetical protein 5546338 5546162 0.69 -0.58 -0.61 -0.58 0.2 1.19 1.54 0.85 1.32 -0.27 1.31 -0.05 0.17 1.02 0.92509 RB10861_hypothetical protein 5896851 5896991 -0.01 -1 -1.34 -1.15 -0.33 2.6 2.38 1.9 2.14 -0.31 0.54 -0.06 -0.26 0.08 1.46513 RB10867_hypothetical protein 5899359 5899982 0.59 0.01 -0.38 -0.25 -0.02 1.49 1.5 1.13 1.38 -0.01 -0.10 -0.15 NaN 0.11 0.81
2324 RB1909_Globin 992934 993434 NaN NaN NaN NaN 0.08 1.4 1.44 1.85 1.1 -0.19 0.09 NaN NaN -0.32 2.104326 RB5003_Rh-like protein/ammonium transporter 2572662 2574158 -0.2 0.03 -0.01 0.14 0.1 2.11 1.81 1.18 1.39 0.46 0.98 -0.12 -0.33 0.62 1.92510 RB10862_Rhs family protein 5896997 5898142 0.46 -0.01 -0.13 NaN 0.33 1.57 1.76 1.11 1.32 0.14 0.00 0.04 NaN 0.03 0.67
1063 RB11798_conserved hypothetical protein 6360319 6360450 0.16 -0.04 0.15 -0.03 -0.03 1.06 0.78 1.29 0.8 0.55 0.76 -0.09 NaN -0.07 0.883519 RB379_ferredoxin--nitrite reductase 194217 196058 NaN NaN NaN NaN NaN 1.06 1.32 1.19 1.03 -0.04 -0.09 0.16 NaN NaN NaN4144 RB4742_Universal stress protein 2433589 2432615 0.67 NaN NaN NaN -0.14 0.58 1.67 1.29 0.86 NaN NaN NaN NaN NaN NaN5757 RB7341_HEMAGGLUTININ/HEMOLYSIN-RELATED PROTEIN 3913925 3936541 0.56 0.15 -0.51 -0.41 -0.46 0.82 2.11 1.12 1.93 0.86 NaN NaN NaN NaN NaN108 RB10167_hypothetical protein 5497096 5496917 0.31 -0.16 -0.09 -0.1 0.67 0.67 1.39 1.1 1.08 0.41 0.24 NaN NaN 0.25 0.63
1105 RB11862_conserved hypothetical protein, secreted 6387327 6387157 0.34 -0.22 -0.16 -0.09 0.59 0.27 1.29 0.77 1.16 0.05 0.72 -0.13 0.11 0.95 0.151113 RB11877_thiophene and furan oxidation protein 6395113 6396510 0.16 -0.03 0.11 -0.12 0.3 1.4 1.35 1.23 1.6 0.67 NaN -0.23 NaN -0.08 0.281526 RB12541_thioredoxin 6756939 6758414 NaN 0.03 -0.1 NaN NaN 0.52 1.53 0.69 NaN 0.23 NaN NaN NaN NaN NaN1681 RB12795_secreted protein 6902552 6904978 0.73 0.04 -0.47 -0.42 1.24 1.23 0.97 0.98 1.41 -0.32 NaN -0.15 -0.58 NaN NaN2106 RB1517_Lipoprotein releasing system ATP-binding protein lolD 776879 776190 -0.15 0.16 0.09 -0.2 0.47 0.56 1.29 0.77 1.09 0.5 NaN NaN NaN NaN 0.573491 RB375_conserved hypothetical protein 189235 190170 -0.21 NaN NaN 0.46 0.28 1.81 2.03 1.68 1.33 0.27 NaN NaN NaN NaN NaN4145 RB4744_universal stress family protein 2434533 2433604 0.49 0.64 NaN NaN 0.35 1.15 1.31 1.07 0.84 0.11 0.07 NaN NaN -0.24 NaN6472 RB8530_hypothetical protein 4550886 4550725 0.53 0.07 0.21 -0.23 0.01 1.16 1.57 1.19 1.28 0.3 0.49 0.02 0.12 0.59 1.437171 RB973_Lipolytic enzyme, G-D-S-L 511727 509484 0.34 0.1 -0.19 -0.05 0.32 0.76 1.31 0.7 0.76 0.04 0.45 0.26 0.73 0.35 0.57234 RB10391_membrane protein 5615419 5616087 0.22 0.14 -0.04 0.17 0.72 0.75 1.46 0.95 1.03 0.61 0.36 NaN NaN 0.19 1.06604 RB11011_conserved hypothetical protein, secreted 5971127 5970582 1.01 0.35 -0.03 -0.34 0.88 0.67 1.35 0.97 1.22 0.47 0.07 -0.31 NaN 0.23 0.79675 RB11137_hypothetical protein 6041524 6042972 0.56 0.19 0.1 NaN 0.35 1.26 1.27 1 1.31 0.71 0.03 0.07 -0.24 -0.06 0.22
3184 RB3299_protein containing DUF1559 1693917 1692925 0.1 0.42 0.06 0.64 0.37 1.72 1.74 1.27 0.86 0.82 0.62 0.12 0.33 0.38 1.686676 RB8881_hypothetical protein 4726480 4726013 0.52 0.43 0.59 0.29 0.06 0.5 0.94 0.94 1.27 0.63 0.51 -0.06 0.34 0.56 0.757268 RB9897_hypothetical protein 5333773 5333549 0.45 0.28 0.17 0.64 0.28 0.71 1.21 0.96 1.36 0.48 0.32 -0.18 -0.17 0.21 -0.01971 RB11636_conserved hypothetical protein 6275447 6275232 0.11 0.01 0.6 -0.11 0.58 0.37 1.54 1.3 1.4 0.88 NaN NaN NaN NaN 0.33
1417 RB12372_RNA polymerase ECF-type sigma factor 6658702 6659430 0.9 -0.36 -0.06 0.37 0.9 0.79 0.53 0.88 0.88 0.09 0.84 0.26 0.36 0.87 0.291616 RB12689_secreted protein 6833004 6833324 0.51 0.37 0.54 0.11 0.72 0.66 1.3 1.07 1.06 0.29 0.22 NaN NaN 0.25 0.781944 RB13239_conserved hypothetical protein, membrane 7102571 7101981 0.62 -0.02 0.13 -0.07 0.13 1.04 1.39 1.49 2.05 0.63 1.28 0.43 0.03 1.74 0.052097 RB1502_two-component response regulator 769609 770010 1.19 -0.21 0.2 0.03 1.27 1.02 0.65 0.64 1.56 0.05 0.35 -0.22 NaN 0.54 0.766266 RB8181_Protein of unknown function DUF1355 4375005 4377428 0.33 0.85 -0.44 0.13 1.22 NaN 1.82 1.33 1.47 0.13 NaN NaN NaN NaN -0.19110 RB10170_cysteine synthase B, O-acetylserine sulfhydrolase B 5497260 5498231 NaN 0.06 0.36 -0.1 0.74 0.37 1.49 1.11 1.07 0.55 NaN NaN NaN NaN 0.49197 RB10327_Translation initiation factor SUI1 5582627 5583010 0.79 0.52 -0.51 -0.28 0.82 0.79 0.84 1.56 1.09 1.15 NaN NaN NaN NaN NaN603 RB11010_conserved hypothetical protein, secreted 5970585 5969749 0.97 0.11 -0.01 -0.03 0.87 0.43 1.46 0.97 1.24 0.77 0.12 NaN NaN 0.43 1.05607 RB11016_protein containing DUF1559 5974211 5973102 1 -0.08 -0.09 0.31 0.83 0.33 0.66 1.16 1.08 0.73 0.27 -0.21 NaN 0.39 0.73
1174 RB11976_DNA-binding response regulator 6455996 6455283 1.26 0.48 0.26 0.07 0.33 0.69 1.23 1.23 1.72 0.58 0.83 0.10 NaN 0.59 0.611651 RB12747_conserved hypothetical protein 6870758 6870919 1.41 0.07 0.02 0.08 0.38 NaN NaN 0.48 NaN -0.07 0.71 0.00 NaN 1.13 0.442007 RB1352_hypothetical protein 686463 687803 0.32 0.33 0.52 NaN 0.62 0.58 1.26 0.78 1.01 0.69 NaN NaN NaN 0.21 1.047146 RB968_protein containing DUF1596 507840 507079 1.25 -0.39 -0.22 -0.33 0.85 0.25 1.02 1.22 1.35 -0.16 1.13 0.03 0.16 1.79 1.25601 RB11008_secreted protein 5967078 5968262 1.11 -0.01 0.11 0.11 1.14 0.28 0.73 1.21 1.35 0.61 0.13 -0.75 NaN 0.00 0.94837 RB11396_conserved hypothetical protein 6178445 6178555 1.51 0.41 -0.52 0.44 0.93 1.87 1.39 1.07 1.69 0.12 0.42 -0.02 -0.11 0.83 -0.05
1624 RB12697_conserved hypothetical protein 6837436 6842547 0.85 -0.02 -0.49 0.17 0.65 0.32 1.54 0.7 1.08 0.3 NaN NaN NaN NaN 1.591680 RB12790_conserved hypothetical protein 6895351 6902541 0.79 0.63 -0.28 0.21 1.38 0.48 2.23 1.51 1.94 0.32 NaN NaN NaN 0.53 0.843891 RB4375_conserved hypothetical protein 2252103 2242933 0.65 0.68 -0.16 -0.14 0.68 0.46 2.45 1.66 2.07 0.54 NaN NaN NaN 0.12 -0.425498 RB6912_secreted protein 3672793 3672960 0.72 0.24 0.42 0.64 0.38 3.15 2.71 1.94 2.54 0.75 0.35 0.00 0.12 0.78 2.385503 RB6920_protein containing DUF1559 3676847 3677914 1.4 0.05 0.37 0.16 1.02 1.82 2.09 1.76 2.07 0.43 0.56 -0.04 0.05 0.95 1.926267 RB8186_secreted protein 4377386 4378576 0.06 0.43 NaN NaN 1.28 1.05 2.06 1.6 1.47 0.53 NaN NaN NaN NaN -0.116852 RB9199_hypothetical protein 4918323 4918120 0.4 0.43 0.47 0.99 0.57 1.77 2.12 1.44 1.85 0.85 0.66 0.22 NaN -0.30 1.156854 RB9200_4-nitrophenylphosphatase 4919220 4918369 1.21 0.58 0.8 0.57 0.58 2.05 1.84 1.36 1.99 0.33 0.79 -0.03 -0.28 0.36 1.581198 RB1201_hypothetical protein 612754 612629 -0.07 0.49 0.39 0.31 0.42 NaN 1.42 0.91 0.92 0.73 0.52 NaN NaN 0.86 1.312030 RB1392_RNA polymerase sigma factor 711705 710683 1.02 0.52 0.28 0.2 1.04 1.03 1.06 1.09 2.09 0.16 0.36 -0.24 0.04 -0.41 -0.102112 RB1523_hypothetical protein 778654 779130 1.01 0.69 0.39 0.55 1.34 1.4 2.32 1.38 1.49 0.24 0.59 NaN NaN 0.28 1.815920 RB7603_outer membrane efflux protein 4091070 4088560 0.67 0.44 0.63 0.35 0.31 0.75 0.83 1.12 1.4 1.53 0.51 NaN NaN 0.21 1.41
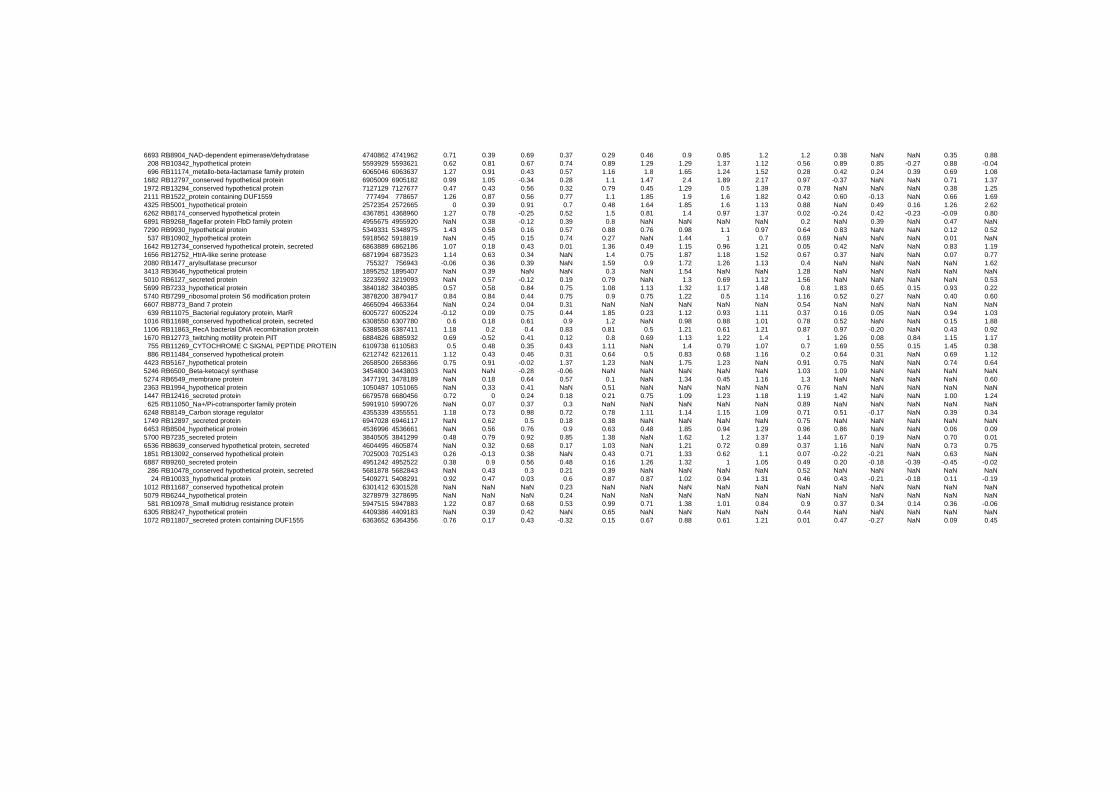
6693 RB8904_NAD-dependent epimerase/dehydratase 4740862 4741962 0.71 0.39 0.69 0.37 0.29 0.46 0.9 0.85 1.2 1.2 0.38 NaN NaN 0.35 0.88208 RB10342_hypothetical protein 5593929 5593621 0.62 0.81 0.67 0.74 0.89 1.29 1.29 1.37 1.12 0.56 0.89 0.85 -0.27 0.88 -0.04696 RB11174_metallo-beta-lactamase family protein 6065046 6063637 1.27 0.91 0.43 0.57 1.16 1.8 1.65 1.24 1.52 0.28 0.42 0.24 0.39 0.69 1.08
1682 RB12797_conserved hypothetical protein 6905009 6905182 0.99 1.05 -0.34 0.28 1.1 1.47 2.4 1.89 2.17 0.97 -0.37 NaN NaN 0.71 1.371972 RB13294_conserved hypothetical protein 7127129 7127677 0.47 0.43 0.56 0.32 0.79 0.45 1.29 0.5 1.39 0.78 NaN NaN NaN 0.38 1.252111 RB1522_protein containing DUF1559 777494 778657 1.26 0.87 0.56 0.77 1.1 1.85 1.9 1.6 1.82 0.42 0.60 -0.13 NaN 0.66 1.694325 RB5001_hypothetical protein 2572354 2572665 0 0.39 0.91 0.7 0.48 1.64 1.85 1.6 1.13 0.88 NaN 0.49 0.16 1.26 2.626262 RB8174_conserved hypothetical protein 4367851 4368960 1.27 0.78 -0.25 0.52 1.5 0.81 1.4 0.97 1.37 0.02 -0.24 0.42 -0.23 -0.09 0.806891 RB9268_flagellar protein FlbD family protein 4955675 4955920 NaN 0.38 -0.12 0.39 0.8 NaN NaN NaN NaN 0.2 NaN 0.39 NaN 0.47 NaN7290 RB9930_hypothetical protein 5349331 5348975 1.43 0.58 0.16 0.57 0.88 0.76 0.98 1.1 0.97 0.64 0.83 NaN NaN 0.12 0.52537 RB10902_hypothetical protein 5918562 5918819 NaN 0.45 0.15 0.74 0.27 NaN 1.44 1 0.7 0.69 NaN NaN NaN 0.01 NaN
1642 RB12734_conserved hypothetical protein, secreted 6863889 6862186 1.07 0.18 0.43 0.01 1.36 0.49 1.15 0.96 1.21 0.05 0.42 NaN NaN 0.83 1.191656 RB12752_HtrA-like serine protease 6871994 6873523 1.14 0.63 0.34 NaN 1.4 0.75 1.87 1.18 1.52 0.67 0.37 NaN NaN 0.07 0.772080 RB1477_arylsulfatase precursor 755327 756943 -0.06 0.36 0.39 NaN 1.59 0.9 1.72 1.26 1.13 0.4 NaN NaN NaN NaN 1.623413 RB3646_hypothetical protein 1895252 1895407 NaN 0.39 NaN NaN 0.3 NaN 1.54 NaN NaN 1.28 NaN NaN NaN NaN NaN5010 RB6127_secreted protein 3223592 3219093 NaN 0.57 -0.12 0.19 0.79 NaN 1.3 0.69 1.12 1.56 NaN NaN NaN NaN 0.535699 RB7233_hypothetical protein 3840182 3840385 0.57 0.58 0.84 0.75 1.08 1.13 1.32 1.17 1.48 0.8 1.83 0.65 0.15 0.93 0.225740 RB7299_ribosomal protein S6 modification protein 3878200 3879417 0.84 0.84 0.44 0.75 0.9 0.75 1.22 0.5 1.14 1.16 0.52 0.27 NaN 0.40 0.606607 RB8773_Band 7 protein 4665094 4663364 NaN 0.24 0.04 0.31 NaN NaN NaN NaN NaN 0.54 NaN NaN NaN NaN NaN639 RB11075_Bacterial regulatory protein, MarR 6005727 6005224 -0.12 0.09 0.75 0.44 1.85 0.23 1.12 0.93 1.11 0.37 0.16 0.05 NaN 0.94 1.03
1016 RB11698_conserved hypothetical protein, secreted 6308550 6307780 0.6 0.18 0.61 0.9 1.2 NaN 0.98 0.88 1.01 0.78 0.52 NaN NaN 0.15 1.881106 RB11863_RecA bacterial DNA recombination protein 6388538 6387411 1.18 0.2 0.4 0.83 0.81 0.5 1.21 0.61 1.21 0.87 0.97 -0.20 NaN 0.43 0.921670 RB12773_twitching motility protein PilT 6884826 6885932 0.69 -0.52 0.41 0.12 0.8 0.69 1.13 1.22 1.4 1 1.26 0.08 0.84 1.15 1.17755 RB11269_CYTOCHROME C SIGNAL PEPTIDE PROTEIN 6109738 6110583 0.5 0.48 0.35 0.43 1.11 NaN 1.4 0.79 1.07 0.7 1.69 0.55 0.15 1.45 0.38886 RB11484_conserved hypothetical protein 6212742 6212611 1.12 0.43 0.46 0.31 0.64 0.5 0.83 0.68 1.16 0.2 0.64 0.31 NaN 0.69 1.12
4423 RB5167_hypothetical protein 2658500 2658366 0.75 0.91 -0.02 1.37 1.23 NaN 1.75 1.23 NaN 0.91 0.75 NaN NaN 0.74 0.645246 RB6500_Beta-ketoacyl synthase 3454800 3443803 NaN NaN -0.28 -0.06 NaN NaN NaN NaN NaN 1.03 1.09 NaN NaN NaN NaN5274 RB6549_membrane protein 3477191 3478189 NaN 0.18 0.64 0.57 0.1 NaN 1.34 0.45 1.16 1.3 NaN NaN NaN NaN 0.602363 RB1994_hypothetical protein 1050487 1051065 NaN 0.33 0.41 NaN 0.51 NaN NaN NaN NaN 0.76 NaN NaN NaN NaN NaN1447 RB12416_secreted protein 6679578 6680456 0.72 0 0.24 0.18 0.21 0.75 1.09 1.23 1.18 1.19 1.42 NaN NaN 1.00 1.24625 RB11050_Na+/Pi-cotransporter family protein 5991910 5990726 NaN 0.07 0.37 0.3 NaN NaN NaN NaN NaN 0.89 NaN NaN NaN NaN NaN
6248 RB8149_Carbon storage regulator 4355339 4355551 1.18 0.73 0.98 0.72 0.78 1.11 1.14 1.15 1.09 0.71 0.51 -0.17 NaN 0.39 0.341749 RB12897_secreted protein 6947028 6946117 NaN 0.62 0.5 0.18 0.38 NaN NaN NaN NaN 0.75 NaN NaN NaN NaN NaN6453 RB8504_hypothetical protein 4536996 4536661 NaN 0.56 0.76 0.9 0.63 0.48 1.85 0.94 1.29 0.96 0.86 NaN NaN 0.06 0.095700 RB7235_secreted protein 3840505 3841299 0.48 0.79 0.92 0.85 1.38 NaN 1.62 1.2 1.37 1.44 1.67 0.19 NaN 0.70 0.016536 RB8639_conserved hypothetical protein, secreted 4604495 4605874 NaN 0.32 0.68 0.17 1.03 NaN 1.21 0.72 0.89 0.37 1.16 NaN NaN 0.73 0.751851 RB13092_conserved hypothetical protein 7025003 7025143 0.26 -0.13 0.38 NaN 0.43 0.71 1.33 0.62 1.1 0.07 -0.22 -0.21 NaN 0.63 NaN6887 RB9260_secreted protein 4951242 4952522 0.38 0.9 0.56 0.48 0.16 1.26 1.32 1 1.05 0.49 0.20 -0.18 -0.39 -0.45 -0.02286 RB10478_conserved hypothetical protein, secreted 5681878 5682843 NaN 0.43 0.3 0.21 0.39 NaN NaN NaN NaN 0.52 NaN NaN NaN NaN NaN24 RB10033_hypothetical protein 5409271 5408291 0.92 0.47 0.03 0.6 0.87 0.87 1.02 0.94 1.31 0.46 0.43 -0.21 -0.18 0.11 -0.19
1012 RB11687_conserved hypothetical protein 6301412 6301528 NaN NaN NaN 0.23 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN5079 RB6244_hypothetical protein 3278979 3278695 NaN NaN NaN 0.24 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN581 RB10978_Small multidrug resistance protein 5947515 5947883 1.22 0.87 0.68 0.53 0.99 0.71 1.38 1.01 0.84 0.9 0.37 0.34 0.14 0.36 -0.06
6305 RB8247_hypothetical protein 4409386 4409183 NaN 0.39 0.42 NaN 0.65 NaN NaN NaN NaN 0.44 NaN NaN NaN NaN NaN1072 RB11807_secreted protein containing DUF1555 6363652 6364356 0.76 0.17 0.43 -0.32 0.15 0.67 0.88 0.61 1.21 0.01 0.47 -0.27 NaN 0.09 0.45
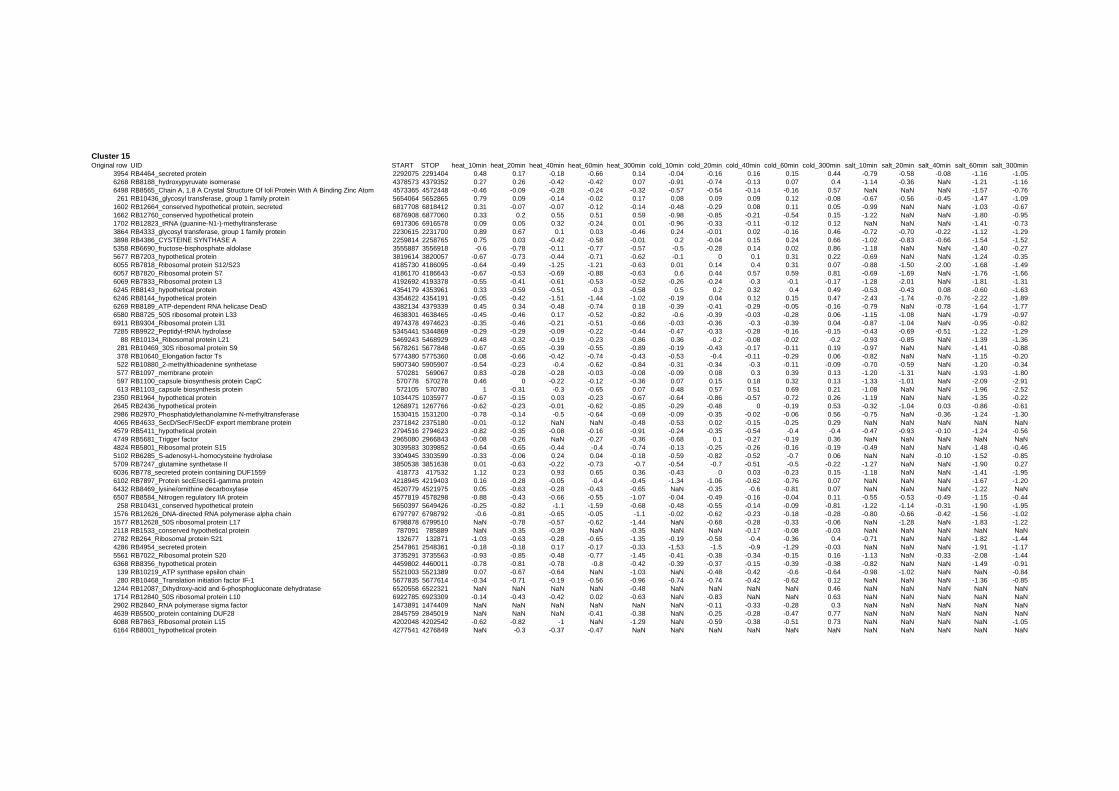
Cluster 15Original row UID START STOP heat_10min heat_20min heat_40min heat_60min heat_300min cold_10min cold_20min cold_40min cold_60min cold_300min salt_10min salt_20min salt_40min salt_60min salt_300min
3954 RB4464_secreted protein 2292075 2291404 0.48 0.17 -0.18 -0.66 0.14 -0.04 -0.16 0.16 0.15 0.44 -0.79 -0.58 -0.08 -1.16 -1.056268 RB8188_hydroxypyruvate isomerase 4378573 4379352 0.27 0.26 -0.42 -0.42 0.07 -0.91 -0.74 -0.13 0.07 0.4 -1.14 -0.36 NaN -1.21 -1.166498 RB8565_Chain A, 1.8 A Crystal Structure Of Ioli Protein With A Binding Zinc Atom 4573365 4572448 -0.46 -0.09 -0.28 -0.24 -0.32 -0.57 -0.54 -0.14 -0.16 0.57 NaN NaN NaN -1.57 -0.76261 RB10436_glycosyl transferase, group 1 family protein 5654064 5652865 0.79 0.09 -0.14 -0.02 0.17 0.08 0.09 0.09 0.12 -0.08 -0.67 -0.56 -0.45 -1.47 -1.09
1602 RB12664_conserved hypothetical protein, secreted 6817708 6818412 0.31 -0.07 -0.07 -0.12 -0.14 -0.48 -0.29 0.08 0.11 0.05 -0.99 NaN NaN -1.03 -0.671662 RB12760_conserved hypothetical protein 6876908 6877060 0.33 0.2 0.55 0.51 0.59 -0.98 -0.85 -0.21 -0.54 0.15 -1.22 NaN NaN -1.80 -0.951702 RB12823_tRNA (guanine-N1-)-methyltransferase 6917306 6916578 0.09 0.05 0.32 -0.24 0.01 -0.96 -0.33 -0.11 -0.12 0.12 NaN NaN NaN -1.41 -0.733864 RB4333_glycosyl transferase, group 1 family protein 2230615 2231700 0.89 0.67 0.1 0.03 -0.46 0.24 -0.01 0.02 -0.16 0.46 -0.72 -0.70 -0.22 -1.12 -1.293898 RB4386_CYSTEINE SYNTHASE A 2259814 2258765 0.75 0.03 -0.42 -0.58 -0.01 0.2 -0.04 0.15 0.24 0.66 -1.02 -0.83 -0.66 -1.54 -1.525358 RB6690_fructose-bisphosphate aldolase 3555887 3556918 -0.6 -0.78 -0.11 -0.77 -0.57 -0.5 -0.28 0.14 0.02 0.86 -1.18 NaN NaN -1.40 -0.275677 RB7203_hypothetical protein 3819614 3820057 -0.67 -0.73 -0.44 -0.71 -0.62 -0.1 0 0.1 0.31 0.22 -0.69 NaN NaN -1.24 -0.356055 RB7818_Ribosomal protein S12/S23 4185730 4186095 -0.64 -0.49 -1.25 -1.21 -0.63 0.01 0.14 0.4 0.31 0.07 -0.88 -1.50 -2.00 -1.68 -1.496057 RB7820_Ribosomal protein S7 4186170 4186643 -0.67 -0.53 -0.69 -0.88 -0.63 0.6 0.44 0.57 0.59 0.81 -0.69 -1.69 NaN -1.76 -1.666069 RB7833_Ribosomal protein L3 4192692 4193378 -0.55 -0.41 -0.61 -0.53 -0.52 -0.26 -0.24 -0.3 -0.1 -0.17 -1.28 -2.01 NaN -1.81 -1.316245 RB8143_hypothetical protein 4354179 4353961 0.33 -0.59 -0.51 -0.3 -0.58 0.5 0.2 0.32 0.4 0.49 -0.53 -0.43 0.08 -0.60 -1.636246 RB8144_hypothetical protein 4354622 4354191 -0.05 -0.42 -1.51 -1.44 -1.02 -0.19 0.04 0.12 0.15 0.47 -2.43 -1.74 -0.76 -2.22 -1.896269 RB8189_ATP-dependent RNA helicase DeaD 4382134 4379339 0.45 0.34 -0.48 -0.74 0.18 -0.39 -0.41 -0.29 -0.05 -0.16 -0.79 NaN -0.78 -1.64 -1.776580 RB8725_50S ribosomal protein L33 4638301 4638465 -0.45 -0.46 0.17 -0.52 -0.82 -0.6 -0.39 -0.03 -0.28 0.06 -1.15 -1.08 NaN -1.79 -0.976911 RB9304_Ribosomal protein L31 4974378 4974623 -0.35 -0.46 -0.21 -0.51 -0.66 -0.03 -0.36 -0.3 -0.39 0.04 -0.87 -1.04 NaN -0.95 -0.827285 RB9922_Peptidyl-tRNA hydrolase 5345441 5344869 -0.29 -0.29 -0.09 -0.22 -0.44 -0.47 -0.33 -0.28 -0.16 -0.15 -0.43 -0.69 -0.51 -1.22 -1.29
88 RB10134_Ribosomal protein L21 5469243 5468929 -0.48 -0.32 -0.19 -0.23 -0.86 0.36 -0.2 -0.08 -0.02 -0.2 -0.93 -0.85 NaN -1.39 -1.36281 RB10469_30S ribosomal protein S9 5678261 5677848 -0.67 -0.65 -0.39 -0.55 -0.89 -0.19 -0.43 -0.17 -0.11 0.19 -0.97 NaN NaN -1.41 -0.88378 RB10640_Elongation factor Ts 5774380 5775360 0.08 -0.66 -0.42 -0.74 -0.43 -0.53 -0.4 -0.11 -0.29 0.06 -0.82 NaN NaN -1.15 -0.20522 RB10880_2-methylthioadenine synthetase 5907340 5905907 -0.54 -0.23 -0.4 -0.62 -0.84 -0.31 -0.34 -0.3 -0.11 -0.09 -0.70 -0.59 NaN -1.20 -0.34577 RB1097_membrane protein 570281 569067 0.83 -0.28 -0.28 -0.03 -0.08 -0.09 0.08 0.3 0.39 0.13 -1.20 -1.31 NaN -1.93 -1.80597 RB1100_capsule biosynthesis protein CapC 570778 570278 0.46 0 -0.22 -0.12 -0.36 0.07 0.15 0.18 0.32 0.13 -1.33 -1.01 NaN -2.09 -2.91613 RB1103_capsule biosynthesis protein 572105 570780 1 -0.31 -0.3 -0.65 0.07 0.48 0.57 0.51 0.69 0.21 -1.08 NaN NaN -1.96 -2.52
2350 RB1964_hypothetical protein 1034475 1035977 -0.67 -0.15 0.03 -0.23 -0.67 -0.64 -0.86 -0.57 -0.72 0.26 -1.19 NaN NaN -1.35 -0.222645 RB2436_hypothetical protein 1268971 1267766 -0.62 -0.23 -0.01 -0.62 -0.85 -0.29 -0.48 0 -0.19 0.53 -0.32 -1.04 0.03 -0.86 -0.612986 RB2970_Phosphatidylethanolamine N-methyltransferase 1530415 1531200 -0.78 -0.14 -0.5 -0.64 -0.69 -0.09 -0.35 -0.02 -0.06 0.56 -0.75 NaN -0.36 -1.24 -1.304065 RB4633_SecD/SecF/SecDF export membrane protein 2371842 2375180 -0.01 -0.12 NaN NaN -0.48 -0.53 0.02 -0.15 -0.25 0.29 NaN NaN NaN NaN NaN4579 RB5411_hypothetical protein 2794516 2794623 -0.82 -0.35 -0.08 -0.16 -0.91 -0.24 -0.35 -0.54 -0.4 -0.4 -0.47 -0.93 -0.10 -1.24 -0.564749 RB5681_Trigger factor 2965080 2966843 -0.08 -0.26 NaN -0.27 -0.36 -0.68 0.1 -0.27 -0.19 0.36 NaN NaN NaN NaN NaN4824 RB5801_Ribosomal protein S15 3039583 3039852 -0.64 -0.65 -0.44 -0.4 -0.74 -0.13 -0.25 -0.26 -0.16 -0.19 -0.49 NaN NaN -1.48 -0.465102 RB6285_S-adenosyl-L-homocysteine hydrolase 3304945 3303599 -0.33 -0.06 0.24 0.04 -0.18 -0.59 -0.82 -0.52 -0.7 0.06 NaN NaN -0.10 -1.52 -0.855709 RB7247_glutamine synthetase II 3850538 3851638 0.01 -0.63 -0.22 -0.73 -0.7 -0.54 -0.7 -0.51 -0.5 -0.22 -1.27 NaN NaN -1.90 0.276036 RB778_secreted protein containing DUF1559 418773 417532 1.12 0.23 0.93 0.65 0.36 -0.43 0 0.03 -0.23 0.15 -1.18 NaN NaN -1.41 -1.956102 RB7897_Protein secE/sec61-gamma protein 4218945 4219403 0.16 -0.28 -0.05 -0.4 -0.45 -1.34 -1.06 -0.62 -0.76 0.07 NaN NaN NaN -1.67 -1.206432 RB8469_lysine/ornithine decarboxylase 4520779 4521975 0.05 -0.63 -0.28 -0.43 -0.65 NaN -0.35 -0.6 -0.81 0.07 NaN NaN NaN -1.22 NaN6507 RB8584_Nitrogen regulatory IIA protein 4577819 4578298 -0.88 -0.43 -0.66 -0.55 -1.07 -0.04 -0.49 -0.16 -0.04 0.11 -0.55 -0.53 -0.49 -1.15 -0.44258 RB10431_conserved hypothetical protein 5650397 5649426 -0.25 -0.82 -1.1 -1.59 -0.68 -0.48 -0.55 -0.14 -0.09 -0.81 -1.22 -1.14 -0.31 -1.90 -1.95
1576 RB12626_DNA-directed RNA polymerase alpha chain 6797797 6798792 -0.6 -0.81 -0.65 -0.05 -1.1 -0.02 -0.62 -0.23 -0.18 -0.28 -0.80 -0.66 -0.42 -1.56 -1.021577 RB12628_50S ribosomal protein L17 6798878 6799510 NaN -0.78 -0.57 -0.62 -1.44 NaN -0.68 -0.28 -0.33 -0.06 NaN -1.28 NaN -1.83 -1.222118 RB1533_conserved hypothetical protein 787091 785889 NaN -0.35 -0.39 NaN -0.35 NaN NaN -0.17 -0.08 -0.03 NaN NaN NaN NaN NaN2782 RB264_Ribosomal protein S21 132677 132871 -1.03 -0.63 -0.28 -0.65 -1.35 -0.19 -0.58 -0.4 -0.36 0.4 -0.71 NaN NaN -1.82 -1.444286 RB4954_secreted protein 2547861 2548361 -0.18 -0.18 0.17 -0.17 -0.33 -1.53 -1.5 -0.9 -1.29 -0.03 NaN NaN NaN -1.91 -1.175561 RB7022_Ribosomal protein S20 3735291 3735563 -0.93 -0.85 -0.48 -0.77 -1.45 -0.41 -0.38 -0.34 -0.15 0.16 -1.13 NaN -0.33 -2.08 -1.446368 RB8356_hypothetical protein 4459802 4460011 -0.78 -0.81 -0.78 -0.8 -0.42 -0.39 -0.37 -0.15 -0.39 -0.38 -0.82 NaN NaN -1.49 -0.91139 RB10219_ATP synthase epsilon chain 5521003 5521389 0.07 -0.67 -0.64 NaN -1.03 NaN -0.48 -0.42 -0.6 -0.64 -0.98 -1.02 NaN NaN -0.84280 RB10468_Translation initiation factor IF-1 5677835 5677614 -0.34 -0.71 -0.19 -0.56 -0.96 -0.74 -0.74 -0.42 -0.62 0.12 NaN NaN NaN -1.36 -0.85
1244 RB12087_Dihydroxy-acid and 6-phosphogluconate dehydratase 6520558 6522321 NaN NaN NaN NaN -0.48 NaN NaN NaN NaN 0.46 NaN NaN NaN NaN NaN1714 RB12840_50S ribosomal protein L10 6922785 6923309 -0.14 -0.43 -0.42 0.02 -0.63 NaN -0.83 NaN NaN 0.63 NaN NaN NaN NaN NaN2902 RB2840_RNA polymerase sigma factor 1473891 1474409 NaN NaN NaN NaN NaN NaN -0.11 -0.33 -0.28 0.3 NaN NaN NaN NaN NaN4639 RB5500_protein containing DUF28 2845759 2845019 NaN NaN NaN -0.41 -0.38 NaN -0.25 -0.28 -0.47 0.77 NaN NaN NaN NaN NaN6088 RB7863_Ribosomal protein L15 4202048 4202542 -0.62 -0.82 -1 NaN -1.29 NaN -0.59 -0.38 -0.51 0.73 NaN NaN NaN NaN -1.056164 RB8001_hypothetical protein 4277541 4276849 NaN -0.3 -0.37 -0.47 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN
6509 RB8590_hypothetical protein 4578825 4579148 -0.44 -0.56 -0.57 -0.73 -0.7 -0.83 -1.07 -0.38 -0.48 -0.19 -0.92 -0.56 0.04 -1.28 -0.494572 RB5403_hypothetical protein 2790360 2790208 NaN NaN NaN NaN NaN NaN -0.32 NaN NaN NaN NaN NaN NaN NaN NaN4658 RB5540_LmbE-like protein 2882665 2883432 NaN NaN NaN NaN NaN NaN -0.34 NaN NaN NaN NaN NaN NaN NaN NaN4755 RB5694_Dethiobiotin synthetase 2971417 2970632 NaN NaN NaN NaN NaN NaN -0.36 -0.26 -0.33 0.1 NaN NaN NaN NaN NaN6948 RB9356_protein containing DUF1579 5012771 5012382 -0.59 -0.58 -1.84 -1.73 -0.57 0.25 0.79 0.7 0.44 0.08 -0.48 NaN NaN -1.90 -0.724041 RB4598_membrane protein 2355806 2356726 NaN NaN NaN NaN NaN NaN -0.11 -0.2 -0.39 0.2 NaN NaN NaN NaN NaN3640 RB3972_hypothetical protein 2056175 2056288 NaN -0.32 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN5250 RB6505_hypothetical protein 3456265 3456110 NaN -0.36 -0.35 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN6941 RB9348_hypothetical protein 5008826 5008536 NaN -0.31 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN136 RB10215_ATP synthase alpha chain 5516975 5518498 0.08 -0.54 -0.41 -0.75 -0.77 NaN -0.61 -0.56 -0.6 -0.39 -0.82 -0.43 -0.41 -0.87 -0.88
7252 RB9871_Acetolactate synthase small subunit 5316629 5316045 0.11 -0.28 0.02 -0.49 -0.34 NaN -0.1 -0.38 -0.1 0 -0.56 NaN -0.45 NaN -0.535162 RB638_PQQ enzyme repeat domain protein 346447 347757 NaN NaN NaN -0.52 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN5218 RB6462_hypothetical protein 3414908 3415180 -0.22 -0.55 -0.52 -0.53 -1.08 -0.69 -0.81 -0.55 -0.47 -0.1 -0.73 -0.44 -0.42 -1.06 -0.996116 RB7913_hypothetical protein 4228083 4227877 NaN -0.27 -0.43 -0.55 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN260 RB10434_glycosyl transferase, group 1 family protein 5652902 5651763 1.07 -0.24 -0.73 -0.67 0.64 0.14 0.09 0.12 0.13 -0.93 -0.87 -0.55 NaN -1.32 -0.86
1284 RB12160_thioredoxin 6563104 6562778 0.18 -0.61 -0.99 -0.76 -0.69 0.07 0.2 -0.14 0.29 0.26 -0.35 -0.54 0.29 -0.88 -1.124355 RB5055_ABC transporter, ATP-binding protein 2601971 2600094 -0.46 -0.48 -0.55 -0.56 -0.61 NaN -0.41 -0.39 -0.49 0.22 NaN NaN NaN NaN NaN3724 RB4113_transposase 2117681 2116503 NaN -0.23 -0.59 -0.24 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN4154 RB4756_conserved hypothetical protein 2437784 2438053 0.05 -0.38 NaN NaN -0.68 NaN 0.05 -0.5 NaN NaN NaN NaN NaN NaN NaN189 RB10310_NADH dehydrogenase I chain M 5572980 5571523 0.14 -0.07 -0.7 -1 -0.65 -0.02 -0.06 0.12 0.34 0.4 -0.32 -0.37 -0.63 NaN -0.78
4607 RB5450_hypothetical protein 2824513 2824635 0.08 -0.12 -0.17 -0.13 -0.26 0.03 -0.29 -0.27 -0.25 -0.19 -0.41 NaN -0.66 -0.67 -1.044598 RB5437_Purine phosphorylase, family 2 2818164 2819081 -0.22 -0.13 -0.13 NaN -0.2 NaN -0.61 -0.34 -0.5 -0.18 -0.70 -0.47 NaN NaN NaN1939 RB13231_conserved hypothetical protein 7098472 7099530 0.54 -0.26 -0.44 -0.6 -0.52 NaN -0.42 -0.61 NaN -0.42 NaN NaN NaN NaN NaN134 RB10211_H+-transporting two-sector ATPase, B/B' subunit 5515421 5516206 0.56 0.23 NaN NaN -0.53 NaN 0.15 -0.45 -0.31 0.01 NaN NaN NaN NaN NaN
1973 RB13297_hypothetical protein 7128034 7128630 NaN NaN NaN NaN NaN NaN NaN NaN NaN 0.11 NaN NaN NaN NaN NaN6360 RB8343_Adenylosuccinate lyase 4454677 4456134 NaN -0.17 -0.19 -0.35 -0.47 NaN -0.26 -0.26 -0.28 0.03 NaN NaN NaN NaN NaN
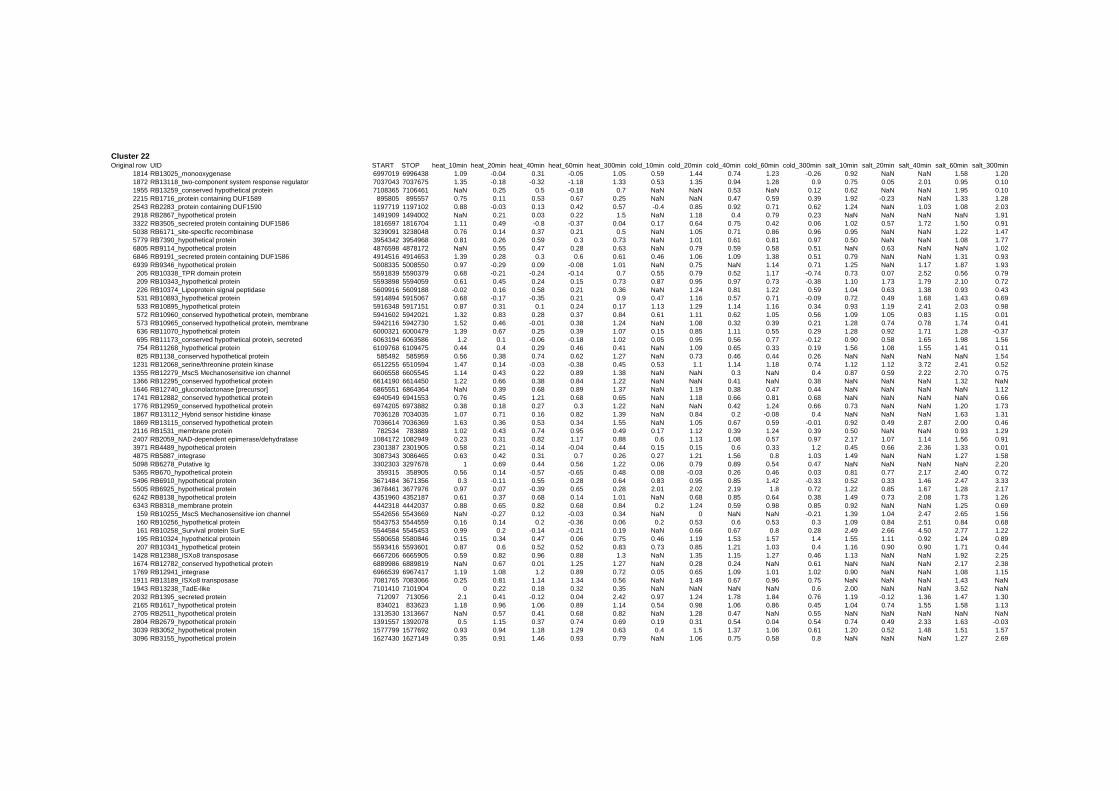
Cluster 22Original row UID START STOP heat_10min heat_20min heat_40min heat_60min heat_300min cold_10min cold_20min cold_40min cold_60min cold_300min salt_10min salt_20min salt_40min salt_60min salt_300min
1814 RB13025_monooxygenase 6997019 6996438 1.09 -0.04 0.31 -0.05 1.05 0.59 1.44 0.74 1.23 -0.26 0.92 NaN NaN 1.58 1.201872 RB13118_two-component system response regulator 7037043 7037675 1.35 -0.18 -0.32 -1.18 1.33 0.53 1.35 0.94 1.28 0.9 0.75 0.05 2.01 0.95 0.101955 RB13259_conserved hypothetical protein 7108365 7106461 NaN 0.25 0.5 -0.18 0.7 NaN NaN 0.53 NaN 0.12 0.62 NaN NaN 1.95 0.102215 RB1716_protein containing DUF1589 895805 895557 0.75 0.11 0.53 0.67 0.25 NaN NaN 0.47 0.59 0.39 1.92 -0.23 NaN 1.33 1.282543 RB2283_protein containing DUF1590 1197719 1197102 0.88 -0.03 0.13 0.42 0.57 -0.4 0.85 0.92 0.71 0.62 1.24 NaN 1.03 1.08 2.032918 RB2867_hypothetical protein 1491909 1494002 NaN 0.21 0.03 0.22 1.5 NaN 1.18 0.4 0.79 0.23 NaN NaN NaN NaN 1.913322 RB3505_secreted protein containing DUF1586 1816597 1816704 1.11 0.49 -0.8 -0.37 0.04 0.17 0.64 0.75 0.42 0.06 1.02 0.57 1.72 1.50 0.915038 RB6171_site-specific recombinase 3239091 3238048 0.76 0.14 0.37 0.21 0.5 NaN 1.05 0.71 0.86 0.96 0.95 NaN NaN 1.22 1.475779 RB7390_hypothetical protein 3954342 3954968 0.81 0.26 0.59 0.3 0.73 NaN 1.01 0.61 0.81 0.97 0.50 NaN NaN 1.08 1.776805 RB9114_hypothetical protein 4876598 4878172 NaN 0.55 0.47 0.28 0.63 NaN 0.79 0.59 0.58 0.51 NaN 0.63 NaN NaN 1.026846 RB9191_secreted protein containing DUF1586 4914516 4914653 1.39 0.28 0.3 0.6 0.61 0.46 1.06 1.09 1.38 0.51 0.79 NaN NaN 1.31 0.936939 RB9346_hypothetical protein 5008335 5008550 0.97 -0.29 0.09 -0.08 1.01 NaN 0.75 NaN 1.14 0.71 1.25 NaN 1.17 1.87 1.93205 RB10338_TPR domain protein 5591839 5590379 0.68 -0.21 -0.24 -0.14 0.7 0.55 0.79 0.52 1.17 -0.74 0.73 0.07 2.52 0.56 0.79209 RB10343_hypothetical protein 5593898 5594059 0.61 0.45 0.24 0.15 0.73 0.87 0.95 0.97 0.73 -0.38 1.10 1.73 1.79 2.10 0.72226 RB10374_Lipoprotein signal peptidase 5609916 5609188 -0.02 0.16 0.58 0.21 0.36 NaN 1.24 0.81 1.22 0.59 1.04 0.63 1.38 0.93 0.43531 RB10893_hypothetical protein 5914894 5915067 0.68 -0.17 -0.35 0.21 0.9 0.47 1.16 0.57 0.71 -0.09 0.72 0.49 1.68 1.43 0.69533 RB10895_hypothetical protein 5916348 5917151 0.87 0.31 0.1 0.24 0.17 1.13 1.29 1.14 1.16 0.34 0.93 1.19 2.41 2.03 0.98572 RB10960_conserved hypothetical protein, membrane 5941602 5942021 1.32 0.83 0.28 0.37 0.84 0.61 1.11 0.62 1.05 0.56 1.09 1.05 0.83 1.15 0.01573 RB10965_conserved hypothetical protein, membrane 5942116 5942730 1.52 0.46 -0.01 0.38 1.24 NaN 1.08 0.32 0.39 0.21 1.28 0.74 0.78 1.74 0.41636 RB11070_hypothetical protein 6000321 6000479 1.39 0.67 0.25 0.39 1.07 0.15 0.85 1.11 0.55 0.29 1.28 0.92 1.71 1.28 -0.37695 RB11173_conserved hypothetical protein, secreted 6063194 6063586 1.2 0.1 -0.06 -0.18 1.02 0.05 0.95 0.56 0.77 -0.12 0.90 0.58 1.65 1.98 1.56754 RB11268_hypothetical protein 6109768 6109475 0.44 0.4 0.29 0.46 0.41 NaN 1.09 0.65 0.33 0.19 1.56 1.08 1.55 1.41 0.11825 RB1138_conserved hypothetical protein 585492 585959 0.56 0.38 0.74 0.62 1.27 NaN 0.73 0.46 0.44 0.26 NaN NaN NaN NaN 1.54
1231 RB12068_serine/threonine protein kinase 6512255 6510594 1.47 0.14 -0.03 -0.38 0.45 0.53 1.1 1.14 1.18 0.74 1.12 1.12 3.72 2.41 0.521355 RB12279_MscS Mechanosensitive ion channel 6606558 6605545 1.14 0.43 0.22 0.89 1.38 NaN NaN 0.3 NaN 0.4 0.87 0.59 2.22 2.70 0.751366 RB12295_conserved hypothetical protein 6614190 6614450 1.22 0.66 0.38 0.84 1.22 NaN NaN 0.41 NaN 0.38 NaN NaN NaN 1.32 NaN1646 RB12740_gluconolactonase [precursor] 6865551 6864364 NaN 0.39 0.68 0.89 1.37 NaN 1.19 0.38 0.47 0.44 NaN NaN NaN NaN 1.121741 RB12882_conserved hypothetical protein 6940549 6941553 0.76 0.45 1.21 0.68 0.65 NaN 1.18 0.66 0.81 0.68 NaN NaN NaN NaN 0.661776 RB12959_conserved hypothetical protein 6974205 6973882 0.38 0.18 0.27 0.3 1.22 NaN NaN 0.42 1.24 0.66 0.73 NaN NaN 1.20 1.731867 RB13112_Hybrid sensor histidine kinase 7036128 7034035 1.07 0.71 0.16 0.82 1.39 NaN 0.84 0.2 -0.08 0.4 NaN NaN NaN 1.63 1.311869 RB13115_conserved hypothetical protein 7036614 7036369 1.63 0.36 0.53 0.34 1.55 NaN 1.05 0.67 0.59 -0.01 0.92 0.49 2.87 2.00 0.462116 RB1531_membrane protein 782534 783889 1.02 0.43 0.74 0.95 0.49 0.17 1.12 0.39 1.24 0.39 0.50 NaN NaN 0.93 1.292407 RB2059_NAD-dependent epimerase/dehydratase 1084172 1082949 0.23 0.31 0.82 1.17 0.88 0.6 1.13 1.08 0.57 0.97 2.17 1.07 1.14 1.56 0.913971 RB4489_hypothetical protein 2301387 2301905 0.58 0.21 -0.14 -0.04 0.44 0.15 0.15 0.6 0.33 1.2 0.45 0.66 2.36 1.33 0.014875 RB5887_integrase 3087343 3086465 0.63 0.42 0.31 0.7 0.26 0.27 1.21 1.56 0.8 1.03 1.49 NaN NaN 1.27 1.585098 RB6278_Putative Ig 3302303 3297678 1 0.69 0.44 0.56 1.22 0.06 0.79 0.89 0.54 0.47 NaN NaN NaN NaN 2.205365 RB670_hypothetical protein 359315 358905 0.56 0.14 -0.57 -0.65 0.48 0.08 -0.03 0.26 0.46 0.03 0.81 0.77 2.17 2.40 0.725496 RB6910_hypothetical protein 3671484 3671356 0.3 -0.11 0.55 0.28 0.64 0.83 0.95 0.85 1.42 -0.33 0.52 0.33 1.46 2.47 3.335505 RB6925_hypothetical protein 3678461 3677976 0.97 0.07 -0.39 0.65 0.28 2.01 2.02 2.19 1.8 0.72 1.22 0.85 1.67 1.28 2.176242 RB8138_hypothetical protein 4351960 4352187 0.61 0.37 0.68 0.14 1.01 NaN 0.68 0.85 0.64 0.38 1.49 0.73 2.08 1.73 1.266343 RB8318_membrane protein 4442318 4442037 0.88 0.65 0.82 0.68 0.84 0.2 1.24 0.59 0.98 0.85 0.92 NaN NaN 1.25 0.69159 RB10255_MscS Mechanosensitive ion channel 5542656 5543669 NaN -0.27 0.12 -0.03 0.34 NaN 0 NaN NaN -0.21 1.39 1.04 2.47 2.65 1.56160 RB10256_hypothetical protein 5543753 5544559 0.16 0.14 0.2 -0.36 0.06 0.2 0.53 0.6 0.53 0.3 1.09 0.84 2.51 0.84 0.68161 RB10258_Survival protein SurE 5544584 5545453 0.99 0.2 -0.14 -0.21 0.19 NaN 0.66 0.67 0.8 0.28 2.49 2.66 4.50 2.77 1.22195 RB10324_hypothetical protein 5580658 5580846 0.15 0.34 0.47 0.06 0.75 0.46 1.19 1.53 1.57 1.4 1.55 1.11 0.92 1.24 0.89207 RB10341_hypothetical protein 5593416 5593601 0.87 0.6 0.52 0.52 0.83 0.73 0.85 1.21 1.03 0.4 1.16 0.90 0.90 1.71 0.44
1428 RB12388_ISXo8 transposase 6667206 6665905 0.59 0.82 0.96 0.88 1.3 NaN 1.35 1.15 1.27 0.46 1.13 NaN NaN 1.92 2.251674 RB12782_conserved hypothetical protein 6889986 6889819 NaN 0.67 0.01 1.25 1.27 NaN 0.28 0.24 NaN 0.61 NaN NaN NaN 2.17 2.381769 RB12941_integrase 6966539 6967417 1.19 1.08 1.2 0.89 0.72 0.05 0.65 1.09 1.01 1.02 0.90 NaN NaN 1.08 1.151911 RB13189_ISXo8 transposase 7081765 7083066 0.25 0.81 1.14 1.34 0.56 NaN 1.49 0.67 0.96 0.75 NaN NaN NaN 1.43 NaN1943 RB13238_TadE-like 7101410 7101904 0 0.22 0.18 0.32 0.35 NaN NaN NaN NaN 0.6 2.00 NaN NaN 3.52 NaN2032 RB1395_secreted protein 712097 713056 2.1 0.41 -0.12 0.04 2.42 0.97 1.24 1.78 1.84 0.76 1.19 -0.12 1.36 1.47 1.302165 RB1617_hypothetical protein 834021 833623 1.18 0.96 1.06 0.89 1.14 0.54 0.98 1.06 0.86 0.45 1.04 0.74 1.55 1.58 1.132705 RB2511_hypothetical protein 1313530 1313667 NaN 0.57 0.41 0.68 0.82 NaN 1.28 0.47 NaN 0.55 NaN NaN NaN NaN NaN2804 RB2679_hypothetical protein 1391557 1392078 0.5 1.15 0.37 0.74 0.69 0.19 0.31 0.54 0.04 0.54 0.74 0.49 2.33 1.63 -0.033039 RB3052_hypothetical protein 1577799 1577692 0.93 0.94 1.18 1.29 0.63 0.4 1.5 1.37 1.06 0.61 1.20 0.52 1.48 1.51 1.573096 RB3155_hypothetical protein 1627430 1627149 0.35 0.91 1.46 0.93 0.79 NaN 1.06 0.75 0.58 0.8 NaN NaN NaN 1.27 2.69
3725 RB4114_protein containing DUF1582 2117690 2117971 1.25 0.91 1.2 1.09 0.4 -0.22 0.54 0.85 0.74 1.05 1.13 NaN NaN 0.95 1.763923 RB4420_DedA family protein 2274093 2273473 0.49 1.27 1.09 0.97 1.43 0.51 0.49 0.56 0.15 1.29 0.92 0.93 1.79 1.04 0.123968 RB4484_Peptidase M20 2297267 2298565 0.61 0.67 0.17 0.5 0.29 0.88 0.54 0.35 0.43 0.63 1.29 1.70 2.50 1.36 0.383969 RB4485_Glutamate--cysteine ligase, GCS2 2298652 2299770 0.23 1.2 0.01 1 0.2 0.72 0.69 0.44 0.12 0.6 1.24 1.31 1.89 1.33 0.543987 RB4510_hypothetical protein 2307657 2307508 0.83 1.56 0.98 1.22 0.82 0.23 0.57 0.72 0.29 1.6 1.23 0.89 2.56 1.37 0.934860 RB5869_conserved hypothetical protein 3078569 3077796 -0.03 0.5 0.96 1.07 0.62 1.13 1.27 1.02 0.87 1.28 3.09 1.49 0.86 0.82 -0.085158 RB6375_hypothetical protein 3349681 3350574 0.94 1.21 0.75 0.54 0.42 0.24 0.38 0.71 0.49 0.7 1.08 0.80 1.41 1.61 1.686107 RB7901_hypothetical protein 4221613 4221293 1.06 0.43 1.13 0.75 0.3 1.24 1.92 1.34 1.66 2.41 1.26 NaN NaN 1.55 1.796178 RB8029_oxidoreductase, short-chain dehydrogenase/reductase family 4292625 4291888 NaN 0.58 1.54 1 0.77 NaN 1.37 0.82 0.54 0.59 2.35 0.84 NaN 0.96 NaN6386 RB8385_hypothetical protein 4479935 4480087 NaN 0.72 0.92 1.6 0.38 NaN 1.46 1.02 0.82 0.81 0.85 NaN NaN 0.93 2.247157 RB9699_Rhs element Vgr protein 5226252 5228345 NaN 0.57 1.05 0.97 0.81 NaN 0.57 NaN 1.02 1.63 0.60 NaN NaN 1.59 1.66264 RB10440_hypothetical protein 5655693 5655842 1.55 1.13 1.02 0.79 1.68 0.53 1.01 0.96 0.67 -0.17 1.42 1.48 1.23 1.81 1.31285 RB10473_conserved hypothetical protein 5679433 5681868 1.23 1 1.06 0.58 1.49 NaN 1.35 0.84 0.91 0.83 0.43 0.20 NaN 1.62 1.70658 RB11110_Rhodopirellula transposase 6022600 6023529 1.98 0.71 0.69 0.48 1.76 NaN 1.41 0.76 0.86 0.76 NaN NaN NaN 1.30 2.07719 RB11209_hypothetical protein 6080603 6080914 0.18 0.76 0.96 1.04 0.78 NaN 1.07 NaN 0.71 0.96 1.94 NaN NaN 1.86 1.71
1058 RB11768_conserved hypothetical protein 6334808 6334939 0.4 0.2 0.97 0.7 0.6 NaN NaN 0.44 0.49 0.4 0.76 0.52 NaN 1.50 1.442923 RB2875_hypothetical protein 1497313 1497534 0.04 0.62 1.03 1.12 0.44 0.53 0.62 0.54 0.16 0.44 2.42 1.73 1.62 1.43 0.823238 RB3375_hypothetical protein 1740198 1740070 NaN 0.8 0.28 0.56 NaN NaN 1.22 NaN NaN 0.46 NaN NaN NaN NaN NaN3458 RB370_nitrate transporter substrate-binding protein 184428 186089 0.32 0.71 0.68 1.76 1.23 1.5 1.98 1.91 1.21 1.19 1.67 NaN NaN 2.39 2.964858 RB5866_ISXo8 transposase 3076242 3077567 1.52 0.82 1.15 0.85 1.52 NaN 1.44 1.26 1.35 0.25 1.05 NaN NaN 1.98 NaN5373 RB671_hypothetical protein 359218 359586 1.8 1.02 0.61 0.89 2.03 NaN 1.24 0.76 0.78 1.02 0.91 0.51 2.60 1.43 0.926416 RB8437_conserved hypothetical protein, secreted 4506842 4509907 NaN 0.29 1.05 1.52 0.55 NaN NaN NaN NaN NaN 2.69 NaN NaN NaN NaN181 RB10295_methylene tetrahydrofolate 5562427 5561501 1.23 0.85 1.45 0.92 0.95 NaN 1.52 0.92 1.24 1.37 1.02 NaN NaN 1.42 1.53491 RB10824_hypothetical protein 5878877 5878999 NaN 0.52 1.08 1.29 0.88 NaN 1.37 0.67 0.81 0.75 NaN NaN NaN NaN NaN
1267 RB12132_conserved hypothetical protein 6548702 6548550 NaN NaN 0.48 NaN NaN NaN NaN NaN NaN 0.82 NaN NaN NaN NaN NaN1479 RB12460_protein containing DUF1571 6707170 6706103 NaN 0.69 1.04 0.8 0.91 NaN 1.25 NaN 0.87 0.71 NaN NaN NaN NaN NaN1643 RB12737_conserved hypothetical protein 6863917 6864114 1.08 0.99 1.45 1.3 1.87 0.35 0.92 1.03 0.81 0.14 1.26 1.19 2.09 2.02 1.981897 RB13165_xylose operon regulatory protein 7067089 7068300 NaN 0.56 0.55 0.79 NaN NaN NaN NaN NaN 0.57 NaN NaN NaN NaN NaN1920 RB13202_conserved hypothetical protein 7086189 7086052 NaN 0.27 1.4 0.71 0.49 NaN NaN NaN NaN -0.15 NaN NaN NaN 1.44 NaN3126 RB3207_hypothetical protein 1652052 1652453 NaN NaN NaN 0.68 NaN NaN NaN NaN NaN 0.55 NaN NaN NaN NaN NaN4322 RB5_secreted protein 1514 399 NaN NaN 0.91 0.81 0.51 NaN -0.06 NaN -0.56 NaN NaN 0.01 NaN 3.14 2.356538 RB8641_hypothetical protein 4605970 4606248 1.88 1.18 1.3 1.24 1.31 NaN NaN 0.25 NaN 0.63 1.52 0.36 0.56 2.61 2.42902 RB11505_conserved hypothetical protein, secreted 6221829 6221230 2.37 1.22 0.49 0.87 1.99 0.96 1.08 1.1 1.58 0.92 0.66 0.46 2.30 2.40 0.76
1764 RB12932_conserved hypothetical protein 6963242 6963015 0.22 1 0.02 1.29 1.37 NaN 1.35 NaN NaN 0.72 NaN NaN NaN NaN NaN1946 RB13241_RNA polymerase ECF-type sigma factor 7103064 7102561 0.73 1.09 1.79 0.85 1.36 NaN 2.28 NaN 2.83 1.27 2.41 1.16 1.09 3.06 0.863869 RB4340_Rhodopirellula transposase 2234846 2233917 0.62 0.83 0.56 1.13 1.59 NaN 1.15 NaN 0.66 0.31 NaN NaN NaN NaN NaN6399 RB8407_Endonuclease/exonuclease/phosphatase 4495545 4494700 NaN 0.41 NaN NaN NaN NaN NaN NaN NaN 0.71 NaN NaN NaN NaN NaN1332 RB12239_ISXo8 transposase 6593334 6594635 1.49 1.15 1.48 1.06 1.65 NaN 1.53 1.49 1.54 0.78 1.32 NaN NaN 2.57 NaN3921 RB4419_MgtC/SapB transporter 2272912 2273448 0.95 1.77 1.79 1.38 2.23 0.82 0.9 1.11 0.77 1.4 1.00 1.56 3.06 2.11 1.092385 RB2019_hypothetical protein 1065619 1065798 NaN NaN NaN 0.59 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN3201 RB3326_hypothetical protein 1711353 1711640 NaN 0.63 0.67 0.88 0.48 NaN NaN NaN NaN 1.09 NaN NaN NaN NaN NaN7307 RB9964_membrane protein 5368651 5367674 NaN NaN NaN NaN 0.81 NaN NaN NaN NaN 0.6 NaN NaN NaN NaN NaN1300 RB12194_SAICAR synthetase 6576972 6575974 NaN 0.84 0.77 NaN 0.48 NaN NaN NaN NaN 0.67 NaN NaN NaN NaN NaN2290 RB1854_protein containing DUF1080 958387 956276 NaN 0.46 0.97 NaN 0.45 NaN NaN NaN NaN 0.82 NaN NaN NaN NaN NaN5729 RB7284_hypothetical protein 3871505 3871639 NaN 0.22 0.83 1.11 0.19 NaN 0.81 0.6 NaN 0.68 NaN NaN NaN NaN NaN6021 RB7755_hypothetical protein 4154446 4154631 0.19 0.4 1.09 0.46 0.45 NaN 1 0.76 0.57 1.06 NaN NaN NaN NaN 0.846372 RB8362_Serine/threonine protein phosphatase 4462144 4460687 0.24 0.74 0.34 NaN 0.68 NaN 1.14 0.69 0.43 0.82 NaN NaN NaN NaN NaN614 RB11030_ABC transporter, ATP-binding protein 5980829 5980074 NaN 0.49 1.32 0.58 0.72 NaN 0.83 NaN NaN 0.53 NaN NaN NaN NaN NaN
4147 RB4748_hypothetical protein 2435328 2435465 0.37 NaN NaN NaN NaN 0.36 1.25 0.86 0.44 NaN NaN NaN NaN NaN NaN360 RB10607_hypothetical protein 5759926 5759537 1.78 0.21 0.38 0.15 0.99 NaN 0.17 NaN NaN 0.17 0.41 -0.04 1.74 1.36 -0.03
1748 RB12895_conserved hypothetical protein 6945636 6946097 NaN 0.72 0.36 NaN 0.66 NaN NaN 0.8 NaN 0.25 NaN NaN NaN NaN NaN6652 RB8843_conserved hypothetical protein 4705018 4706850 NaN NaN NaN 0.8 NaN NaN NaN NaN NaN 0.43 NaN NaN NaN NaN NaN5930 RB7621_hypothetical protein 4097160 4098164 NaN 0.34 0.57 NaN 0.67 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN2173 RB1632_hypothetical protein 839673 840332 NaN 0.39 0.22 0.75 NaN NaN NaN NaN NaN 0.48 NaN NaN NaN NaN NaN3060 RB3077_conserved hypothetical protein 1606106 1588083 NaN NaN NaN NaN NaN NaN NaN NaN NaN 0.64 NaN NaN NaN NaN NaN517 RB10875_hypothetical protein 5901192 5901043 NaN 0.51 -0.11 1.26 0.54 NaN 1.62 0.56 0.08 0.17 NaN NaN NaN NaN NaN
6572 RB871_hypothetical protein 463841 462534 NaN 0.5 NaN NaN 0.72 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN1304 RB1220_Aldose 1-epimerase 622840 623925 0.32 0.46 0.86 0.67 0.71 NaN 0.83 0.39 0.49 0.52 NaN NaN NaN NaN NaN5811 RB7440_hypothetical protein 3990113 3990535 NaN NaN NaN 0.58 NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN NaN

The Rhodopirellula baltica life cycle: Growth-dependent expression profiling of the complete genome with
microarrays
ADDITIONAL FILE 1 - List of differentially expressed genes ADDITIONAL FILE 4 - List of selected clusters out of the k-means clustering

62hvs44h_downID Locus Ratio AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand
547 RB1015 -1.62 222 669 membrane protein 531723 532391 5.9 24319 +5741 RB10299 -1.70 455 1368 conserverd hypothetical protein 5563036 5564403 10 49949 +5910 RB10623 -2.40 60 183 secreted protein 5769140 5768958 3.9 6284 -5921 RB10648 -1.61 310 933 conserved hypothetical protein 5779160 5778228 9.7 34602 -5938 RB10675 -1.82 779 2340 membrane protein 5802507 5800168 7.2 86215 -5939 RB10678 -1.73 116 351 hypothetical protein 5802466 5802816 12 13353 +
63 RB110 -1.50 582 1749 choline-sulfatase 49093 47345 6 65751 -6308 RB11325 -1.92 47 144 conserved hypothetical protein 6142535 6142392 12.2 5354 -598 RB1137 -1.55 94 285 hypothetical protein 585204 585488 11.6 10805 +
6818 RB12250 -1.53 104 315 conserved hypothetical protein 6597248 6597562 11.9 12364 +6870 RB12334 -1.59 266 801 Peptidase M50 3.4.24.- 6636156 6635356 5.4 29077 -6920 RB12426 -1.53 137 414 conserved hypothetical protein, membrane 6684059 6684472 10.6 15166 +6981 RB12526 -1.60 256 771 Rhomboid-like protein 6747132 6747902 6 27269 +7009 RB12581 -1.65 174 525 conserved hypothetical protein 6780597 6780073 9.9 19433 -7113 RB12770 -2.26 166 501 conserved hypothetical protein 6884806 6884306 11.9 18496 -691 RB1291 -1.74 97 294 hypothetical protein 656684 656977 10.4 10290 +
7401 RB13324 -1.74 36 111 hypothetical protein 7141677 7141787 7.3 4100 +1016 RB1931 -1.96 68 207 hypothetical protein 1013831 1013625 11.4 7493 -1105 RB2105 -1.54 470 1413 membrane protein 1114012 1112600 9.6 51853 -1107 RB2110 -1.52 467 1404 multidrug resistance protein norM norM 1114250 1115653 9.6 50647 +1220 RB2306 -1.68 41 126 hypothetical protein 1206782 1206907 9 4759 +1265 RB2370 -1.59 54 165 hypothetical protein 1229187 1229023 6.3 6204 -1332 RB2479 -1.59 273 822 conserved hypothetical protein 1296683 1295862 5.5 30436 -1352 RB2503 -1.61 515 1548 O-antigen flippase wzx 1311056 1309509 10 55740 -1372 RB2529 -1.74 195 588 membrane protein 1320541 1321128 3.9 20867 +1407 RB2592 -1.57 197 594 hypothetical protein 1348926 1348333 4.7 22032 -1701 RB3147 -1.72 168 507 membrane protein 1624840 1624334 8.7 18759 -1742 RB3224 -1.58 148 447 hypothetical protein 1656004 1656450 12.4 16377 +1778 RB3281 -1.53 139 420 Histone-like bacterial DNA-binding protein 1687272 1686853 7.3 15802 -1806 RB3329 -1.66 64 195 hypothetical protein 1711937 1711743 12 6908 -1833 RB3372 -1.72 115 348 hypothetical protein 1733666 1734013 10 12916 +1857 RB3413 -1.66 373 1122 Esterase/lipase/ 1762402 1761281 5.1 40640 -1879 RB3450 -1.52 61 186 hypothetical protein 1782543 1782358 4.7 6530 -1984 RB3627 -1.65 280 843 Transglutaminase-like domain 1888482 1887640 4.5 30412 -2017 RB3675 -1.53 742 2229 secreted protein 1904552 1906780 8.4 82951 +
22 RB37 -1.61 160 483 membrane protein 18328 17846 5.6 17283 -2154 RB3905 -1.89 38 117 hypothetical protein 2018601 2018717 5.1 4131 +2184 RB3953 -2.00 857 2574 hypothetical protein 2048784 2046211 5.2 93881 -2232 RB4032 -1.58 481 1446 secreted protein containing DUF1501 2084885 2083440 6.2 52620 -2260 RB4097 -1.88 733 2202 conserved hypothetical protein 2108244 2110445 6.2 83188 +2263 RB4104 -1.58 72 219 hypothetical protein 2112335 2112553 5.5 7870 +2298 RB4165 -1.57 721 2166 hypothetical protein 2139632 2141797 4.5 77994 +242 RB418 -1.96 363 1092 conserved hypothetical protein, membrane 222222 223313 10.3 39119 +
2357 RB4266 -1.66 54 165 hypothetical protein 2183250 2183414 11.2 6166 +2359 RB4269 -1.61 282 849 glutamic acid specific endopeptidase, GSE=serine endopeptidase 2185049 2184201 5.6 31283 -2363 RB4273 -1.77 518 1557 iron-dependent peroxidase 1.1.1.- 2193431 2191875 5.1 57672 -250 RB434 -1.54 65 198 hypothetical protein 231791 231988 7.4 7385 +
2437 RB4394 -1.70 188 567 protease I pfpI 3.2.-.- 2261372 2261938 4.4 20601 +2444 RB4402 -1.63 53 162 hypothetical protein 2265340 2265501 10.4 5980 +2451 RB4416 -1.85 833 2502 Cytochrome c oxidase, subunit I 2271803 2269302 5.8 92293 -2472 RB4454 -1.54 739 2220 Flagellin A flaA 2283201 2285420 4 76924 +2476 RB4458 -1.77 342 1029 Protein of unknown function, UPF0118 2287089 2286061 6.4 37001 -2477 RB4459 -1.52 462 1389 MscS Mechanosensitive ion channel 2288477 2287089 4.8 51157 -2512 RB4513 -1.54 326 981 serum resistance protein 2315098 2314118 7.4 35351 -2568 RB4613 -1.59 110 333 HesB/YadR/YfhF 2362510 2362178 4.6 12216 -277 RB472 -1.76 47 144 hypothetical protein 253266 253409 10.9 5373 +
2669 RB4773 -1.87 208 627 hypothetical protein 2446723 2447349 10.8 23392 +2746 RB4897 -1.78 184 555 hypothetical protein 2519505 2518951 10.9 20446 -2763 RB4920 -2.34 69 210 hypothetical protein 2531850 2531641 4.4 7587 -2914 RB5174 -1.60 589 1770 Bacterial type II and III secretion system protein 2660936 2662705 4.8 63998 +3055 RB5424 -1.76 573 1722 sulfatase 2807339 2809060 5.7 64433 +3062 RB5434 -1.51 695 2088 Elongation factor G 1 fusA1 2817863 2815776 4.7 77788 -3146 RB5581 -1.68 397 1194 conserved hypothetical protein 2905973 2904780 4.8 44297 -3208 RB5695 -1.69 288 867 carbon-nitrogen hydrolase family 2971401 2972267 4.5 31927 +3212 RB5701 -1.50 1111 3336 oxidase subunit 2975188 2978523 9.1 122095 +3247 RB5761 -1.55 258 777 conserved hypothetical protein, membrane 3015202 3014426 10.9 27944 -3291 RB5841 -1.64 670 2013 serine/threonine protein phosphatase family 3061924 3063936 5.4 73664 +3307 RB5872 -1.51 83 252 hypothetical protein 3080059 3080310 13 9379 +3328 RB5903 -1.54 58 177 hypothetical protein 3094315 3094491 10.1 6651 +3389 RB6010 -1.65 53 162 hypothetical protein 3149911 3150072 11.9 5744 +3396 RB6019 -1.62 161 486 hypothetical protein 3154683 3154198 4.1 18098 -3400 RB6031 -1.55 492 1479 hypothetical protein 3160462 3158984 4.1 54352 -3407 RB6042 -1.55 415 1248 CBS domain protein 3166072 3164825 4.4 45534 -3527 RB6274 -1.71 59 180 hypothetical protein 3297130 3296951 12 7254 -3544 RB6305 -1.52 437 1314 dolichol-phosphate mannosyltransferase 3312842 3314155 8.9 47433 +3617 RB6423 -2.17 45 138 hypothetical protein 3380603 3380740 5.5 5402 +3625 RB6434 -1.77 48 147 hypothetical protein 3388682 3388536 7.4 5104 -3632 RB6443 -1.59 397 1194 Cys/Met metabolism pyridoxal-phosphate-dependent enzymes 3393276 3394469 5.3 42845 +3649 RB6470 -1.63 701 2106 Abortive infection protein 3420270 3422375 5.9 76321 +3667 RB6496 -1.57 57 174 hypothetical protein 3441948 3442121 9.9 6289 +3693 RB6537 -1.57 384 1155 Aldose 1-epimerase 5.1.3.3 3472890 3471736 4.7 42197 -3695 RB6539 -1.75 374 1125 membrane protein 3474598 3475722 10.1 41456 +3713 RB6574 -2.19 723 2172 protein containing DUF1585 3493615 3491444 5.2 81617 -3720 RB6587 -1.92 50 153 hypothetical protein 3501747 3501595 7.5 5579 -3732 RB6610 -1.68 1235 3708 conserved hypothetical protein 3511694 3515401 5.3 137776 +3738 RB6624 -1.63 503 1512 Bacterial sugar transferase 2.7.8.6 3520589 3522100 7.7 55315 +3742 RB6632 -1.68 55 168 hypothetical protein 3524963 3525130 5.6 6193 +3757 RB6663 -1.50 461 1386 Major facilitator superfamily MFS_1 3540255 3538870 9.9 50033 -3780 RB6699 -1.76 47 144 hypothetical protein 3559781 3559924 11.1 5290 +3795 RB6724 -1.54 131 396 hypothetical protein 3569189 3569584 10.5 14916 +3836 RB6789 -1.61 180 543 secreted protein 3608487 3607945 5 20191 -370 RB679 -1.54 60 183 hypothetical protein 362787 362605 10.7 7113 -
3872 RB6858 -1.58 341 1026 dehydrogenases and related proteins 3646807 3645782 5.5 36680 -375 RB686 -1.66 549 1650 arylsulfatase atsA 3.1.6.1 370206 368557 5.4 62276 -
3875 RB6864 -1.65 44 135 hypothetical protein 3648295 3648429 2.8 4573 +3923 RB6958 -1.55 188 567 conserved hypothetical protein 3695752 3695186 9.4 20788 -3971 RB7042 -1.63 91 276 hypothetical protein 3743971 3744246 10.4 10835 +3985 RB7072 -1.53 257 774 lipoate-protein ligase A lplA 6.3.2.- 3754556 3753783 5 29306 -4009 RB7109 -1.72 53 162 hypothetical protein 3775707 3775546 6.7 5850 -4092 RB7246 -1.50 50 153 hypothetical protein 3850393 3850545 9.7 5973 +4303 RB7628 -1.51 122 369 hypothetical protein 4104029 4103661 12.2 13804 -4578 RB8119 -1.60 142 429 hypothetical protein 4346427 4346855 10.1 15659 +4627 RB8223 -1.68 410 1233 protein containing DUF1559 4399759 4398527 6.3 44709 -5060 RB9034 -1.54 1031 3096 conserved hypothetical protein, secreted 4818355 4821450 5.4 113674 +5113 RB9141 -1.50 346 1041 rRNA (guanine-N(2)-)-methyltransferase 4891836 4892876 7.8 38276 +5156 RB9228 -1.51 637 1914 hypothetical protein 4934679 4936592 7.8 73174 +5240 RB9385 -1.56 507 1524 ribose transport ATP-binding protein rbsA rbsA 3.6.3.17 5033732 5032209 4.8 54554 -5275 RB9445 -1.78 249 750 integral membrane protein 5074872 5075621 10.4 26862 +5318 RB9529 -1.76 113 342 hypothetical protein 5117956 5118297 9.3 13082 +5386 RB9651 -2.07 544 1635 sialic acid-specific 9-O-acetylesterase 3.1.1.53 5198277 5196643 5.9 60120 -

62hvs44h_upID Locus Ratio AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand5764 RB10341 2.27 61 186 hypothetical protein 5593416 5593601 4.5 6613 +5766 RB10343 3.62 53 162 hypothetical protein 5593898 5594059 11.3 6302 +5815 RB10440 3.33 49 150 hypothetical protein 5655693 5655842 11.5 5080 +6096 RB10951 3.24 101 306 secreted protein 5940232 5939927 10.2 11754 -
584 RB1104 2.89 59 180 hypothetical protein 572078 572257 10.7 6510 +622 RB1179 1.84 128 387 hypothetical protein 603998 604384 12 13863 +
6720 RB12068 1.63 553 1662 serine/threonine protein kinase 6512255 6510594 8.5 61271 -6785 RB12199 1.58 54 165 conserved hypothetical protein 6577614 6577778 11.3 5569 +6867 RB12329 2.02 110 333 conserved hypothetical protein, membrane 6634234 6634566 4 11862 +7023 RB12610 2.12 85 258 conserved hypothetical protein 6793700 6793957 9.7 9221 +7141 RB12818 1.59 163 492 conserved hypothetical protein 6915989 6915498 11.7 18010 -7151 RB12833 1.85 109 330 conserved hypothetical protein 6921305 6921634 12 12221 +7293 RB13115 2.31 81 246 conserved hypothetical protein 7036614 7036369 9.8 8735 -1028 RB1961 3.07 117 354 hypothetical protein 1033791 1034144 11.9 13392 +1330 RB2477 2.51 123 372 hypothetical protein 1295793 1295422 10.2 13737 -1706 RB3153 1.83 76 231 hypothetical protein (protein containing DUF1584) 1626911 1626681 12.4 8266 -1728 RB3196 1.65 109 330 hypothetical protein 1650093 1650422 10.1 12287 +1835 RB3374 1.50 95 288 hypothetical protein 1740350 1740063 11 11130 -
195 RB341 1.67 90 273 hypothetical protein 169852 170124 11.2 10096 +1992 RB3644 2.98 127 384 hypothetical protein 1895184 1894801 4.9 14294 -2501 RB4498 1.51 90 273 hypothetical protein 2303522 2303250 9.4 9942 -2502 RB4499 2.06 81 246 hypothetical protein 2303764 2303519 10.7 9029 -2553 RB4590 1.51 74 225 hypothetical protein 2352024 2351800 10.1 8330 -3341 RB5929 1.86 74 225 hypothetical protein 3107256 3107480 12.1 8243 +3585 RB6375 1.86 297 894 hypothetical protein 3349681 3350574 8.7 31772 +3587 RB6378 2.75 228 687 hypothetical protein 3352598 3353284 12.3 25698 +3639 RB6454 1.67 93 282 hypothetical protein 3407776 3407495 11.7 10063 -3900 RB6910 3.07 42 129 hypothetical protein 3671484 3671356 9.5 4575 -3909 RB6925 3.02 161 486 hypothetical protein 3678461 3677976 9.8 17641 -3911 RB6930 2.24 413 1242 glutamate dehydrogenase 1.4.1.3 3680317 3679076 5.9 45734 -4422 RB7828 2.96 40 123 hypothetical protein 4192037 4191915 10.4 4526 -4590 RB8146 2.10 151 456 hypothetical protein 4355122 4354667 4.3 17016 -4609 RB8188 2.30 259 780 hydroxypyruvate isomerase hyi 5.3.1.22 4378573 4379352 4.4 28228 +4699 RB8357 1.72 56 171 hypothetical protein 4460174 4460004 7.4 6097 -4861 RB8669 2.61 37 114 hypothetical protein 4614557 4614670 11.5 4395 +4862 RB8670 2.47 47 144 hypothetical protein 4614842 4614985 11.1 5371 +4880 RB8700 1.51 58 177 hypothetical protein 4630224 4630400 4.8 6268 +4919 RB8775 2.49 81 246 hypothetical protein 4665497 4665252 5.3 8745 -5278 RB9450 3.13 52 159 hypothetical protein 5076581 5076739 7.3 5967 +
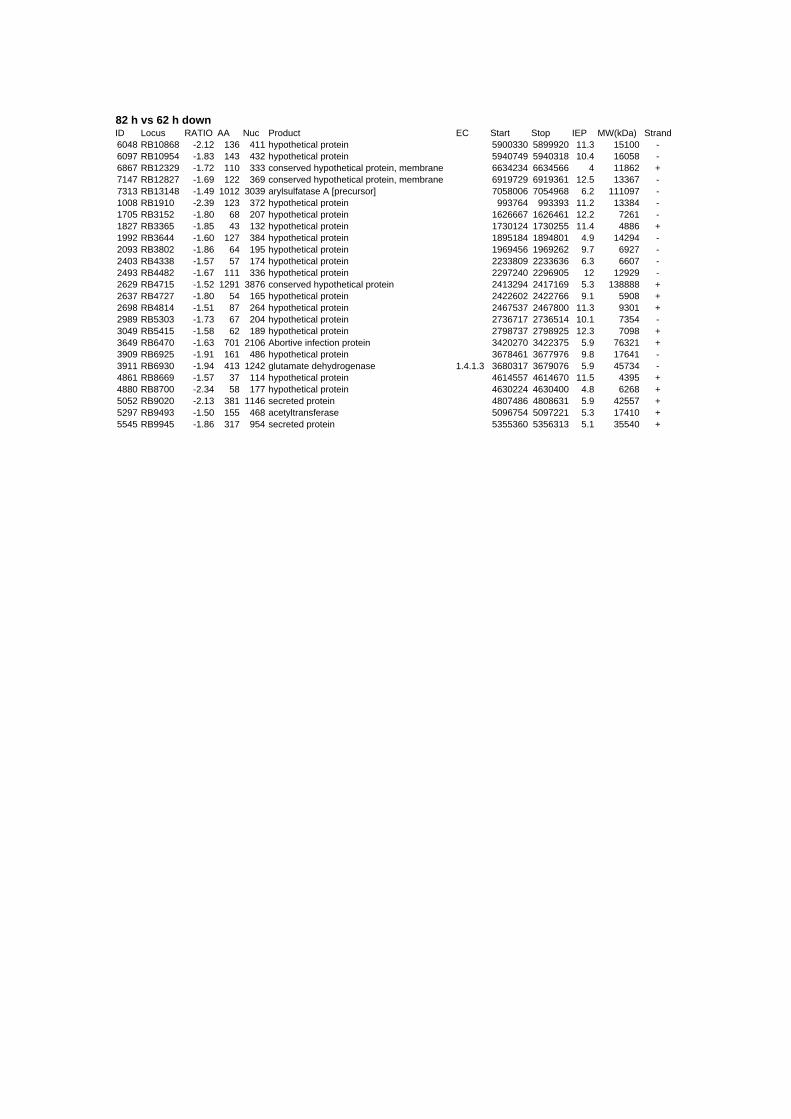
82 h vs 62 h downID Locus RATIO AA Nuc Product EC Start Stop IEP MW(kDa) Strand6048 RB10868 -2.12 136 411 hypothetical protein 5900330 5899920 11.3 15100 -6097 RB10954 -1.83 143 432 hypothetical protein 5940749 5940318 10.4 16058 -6867 RB12329 -1.72 110 333 conserved hypothetical protein, membrane 6634234 6634566 4 11862 +7147 RB12827 -1.69 122 369 conserved hypothetical protein, membrane 6919729 6919361 12.5 13367 -7313 RB13148 -1.49 1012 3039 arylsulfatase A [precursor] 7058006 7054968 6.2 111097 -1008 RB1910 -2.39 123 372 hypothetical protein 993764 993393 11.2 13384 -1705 RB3152 -1.80 68 207 hypothetical protein 1626667 1626461 12.2 7261 -1827 RB3365 -1.85 43 132 hypothetical protein 1730124 1730255 11.4 4886 +1992 RB3644 -1.60 127 384 hypothetical protein 1895184 1894801 4.9 14294 -2093 RB3802 -1.86 64 195 hypothetical protein 1969456 1969262 9.7 6927 -2403 RB4338 -1.57 57 174 hypothetical protein 2233809 2233636 6.3 6607 -2493 RB4482 -1.67 111 336 hypothetical protein 2297240 2296905 12 12929 -2629 RB4715 -1.52 1291 3876 conserved hypothetical protein 2413294 2417169 5.3 138888 +2637 RB4727 -1.80 54 165 hypothetical protein 2422602 2422766 9.1 5908 +2698 RB4814 -1.51 87 264 hypothetical protein 2467537 2467800 11.3 9301 +2989 RB5303 -1.73 67 204 hypothetical protein 2736717 2736514 10.1 7354 -3049 RB5415 -1.58 62 189 hypothetical protein 2798737 2798925 12.3 7098 +3649 RB6470 -1.63 701 2106 Abortive infection protein 3420270 3422375 5.9 76321 +3909 RB6925 -1.91 161 486 hypothetical protein 3678461 3677976 9.8 17641 -3911 RB6930 -1.94 413 1242 glutamate dehydrogenase 1.4.1.3 3680317 3679076 5.9 45734 -4861 RB8669 -1.57 37 114 hypothetical protein 4614557 4614670 11.5 4395 +4880 RB8700 -2.34 58 177 hypothetical protein 4630224 4630400 4.8 6268 +5052 RB9020 -2.13 381 1146 secreted protein 4807486 4808631 5.9 42557 +5297 RB9493 -1.50 155 468 acetyltransferase 5096754 5097221 5.3 17410 +5545 RB9945 -1.86 317 954 secreted protein 5355360 5356313 5.1 35540 +
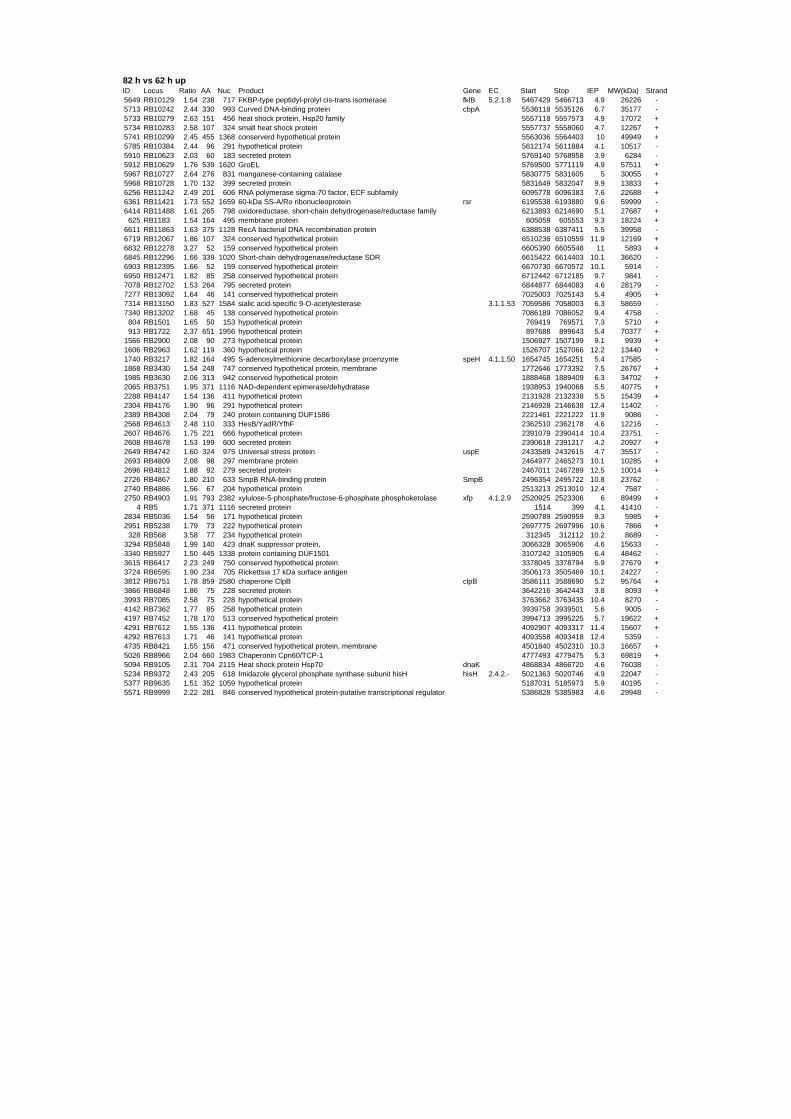
82 h vs 62 h upID Locus Ratio AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand5649 RB10129 1.54 238 717 FKBP-type peptidyl-prolyl cis-trans isomerase fklB 5.2.1.8 5467429 5466713 4.9 26226 -5713 RB10242 2.44 330 993 Curved DNA-binding protein cbpA 5536118 5535126 6.7 35177 -5733 RB10279 2.63 151 456 heat shock protein, Hsp20 family 5557118 5557573 4.9 17072 +5734 RB10283 2.58 107 324 small heat shock protein 5557737 5558060 4.7 12267 +5741 RB10299 2.45 455 1368 conserverd hypothetical protein 5563036 5564403 10 49949 +5785 RB10384 2.44 96 291 hypothetical protein 5612174 5611884 4.1 10517 -5910 RB10623 2.03 60 183 secreted protein 5769140 5768958 3.9 6284 -5912 RB10629 1.76 539 1620 GroEL 5769500 5771119 4.9 57511 +5967 RB10727 2.64 276 831 manganese-containing catalase 5830775 5831605 5 30055 +5968 RB10728 1.70 132 399 secreted protein 5831649 5832047 9.9 13833 +6256 RB11242 2.49 201 606 RNA polymerase sigma-70 factor, ECF subfamily 6095778 6096383 7.6 22688 +6361 RB11421 1.73 552 1659 60-kDa SS-A/Ro ribonucleoprotein rsr 6195538 6193880 9.6 59999 -6414 RB11488 1.61 265 798 oxidoreductase, short-chain dehydrogenase/reductase family 6213893 6214690 5.1 27687 +
625 RB1183 1.54 164 495 membrane protein 605059 605553 9.3 18224 +6611 RB11863 1.63 375 1128 RecA bacterial DNA recombination protein 6388538 6387411 5.5 39958 -6719 RB12067 1.86 107 324 conserved hypothetical protein 6510236 6510559 11.9 12169 +6832 RB12278 3.27 52 159 conserved hypothetical protein 6605390 6605548 11 5893 +6845 RB12296 1.66 339 1020 Short-chain dehydrogenase/reductase SDR 6615422 6614403 10.1 36620 -6903 RB12395 1.66 52 159 conserved hypothetical protein 6670730 6670572 10.1 5914 -6950 RB12471 1.82 85 258 conserved hypothetical protein 6712442 6712185 9.7 9841 -7078 RB12702 1.53 264 795 secreted protein 6844877 6844083 4.6 28179 -7277 RB13092 1.64 46 141 conserved hypothetical protein 7025003 7025143 5.4 4905 +7314 RB13150 1.83 527 1584 sialic acid-specific 9-O-acetylesterase 3.1.1.53 7059586 7058003 6.3 58659 -7340 RB13202 1.68 45 138 conserved hypothetical protein 7086189 7086052 9.4 4758 -
804 RB1501 1.65 50 153 hypothetical protein 769419 769571 7.3 5710 +913 RB1722 2.37 651 1956 hypothetical protein 897688 899643 5.4 70377 +
1566 RB2900 2.08 90 273 hypothetical protein 1506927 1507199 9.1 9939 +1606 RB2963 1.62 119 360 hypothetical protein 1526707 1527066 12.2 13440 +1740 RB3217 1.82 164 495 S-adenosylmethionine decarboxylase proenzyme speH 4.1.1.50 1654745 1654251 5.4 17585 -1868 RB3430 1.54 248 747 conserved hypothetical protein, membrane 1772646 1773392 7.5 26767 +1985 RB3630 2.06 313 942 conserved hypothetical protein 1888468 1889409 6.3 34702 +2065 RB3751 1.95 371 1116 NAD-dependent epimerase/dehydratase 1938953 1940068 5.5 40775 +2288 RB4147 1.54 136 411 hypothetical protein 2131928 2132338 5.5 15439 +2304 RB4176 1.90 96 291 hypothetical protein 2146928 2146638 12.4 11402 -2389 RB4308 2.04 79 240 protein containing DUF1586 2221461 2221222 11.9 9086 -2568 RB4613 2.48 110 333 HesB/YadR/YfhF 2362510 2362178 4.6 12216 -2607 RB4676 1.75 221 666 hypothetical protein 2391079 2390414 10.4 23751 -2608 RB4678 1.53 199 600 secreted protein 2390618 2391217 4.2 20927 +2649 RB4742 1.60 324 975 Universal stress protein uspE 2433589 2432615 4.7 35517 -2693 RB4809 2.08 98 297 membrane protein 2464977 2465273 10.1 10285 +2696 RB4812 1.88 92 279 secreted protein 2467011 2467289 12.5 10014 +2726 RB4867 1.80 210 633 SmpB RNA-binding protein SmpB 2496354 2495722 10.8 23762 -2740 RB4886 1.56 67 204 hypothetical protein 2513213 2513010 12.4 7587 -2750 RB4903 1.91 793 2382 xylulose-5-phosphate/fructose-6-phosphate phosphoketolase xfp 4.1.2.9 2520925 2523306 6 89499 +
4 RB5 1.71 371 1116 secreted protein 1514 399 4.1 41410 -2834 RB5036 1.54 56 171 hypothetical protein 2590789 2590959 9.3 5985 +2951 RB5238 1.79 73 222 hypothetical protein 2697775 2697996 10.6 7866 +
328 RB568 3.58 77 234 hypothetical protein 312345 312112 10.2 8689 -3294 RB5848 1.99 140 423 dnaK suppressor protein, 3066328 3065906 4.6 15633 -3340 RB5927 1.50 445 1338 protein containing DUF1501 3107242 3105905 6.4 48462 -3615 RB6417 2.23 249 750 conserved hypothetical protein 3378045 3378794 5.9 27679 +3724 RB6595 1.90 234 705 Rickettsia 17 kDa surface antigen 3506173 3505469 10.1 24227 -3812 RB6751 1.78 859 2580 chaperone ClpB clpB 3586111 3588690 5.2 95764 +3866 RB6848 1.86 75 228 secreted protein 3642216 3642443 3.8 8093 +3993 RB7085 2.58 75 228 hypothetical protein 3763662 3763435 10.4 8270 -4142 RB7362 1.77 85 258 hypothetical protein 3939758 3939501 5.6 9005 -4197 RB7452 1.78 170 513 conserved hypothetical protein 3994713 3995225 5.7 19622 +4291 RB7612 1.55 136 411 hypothetical protein 4092907 4093317 11.4 15607 +4292 RB7613 1.71 46 141 hypothetical protein 4093558 4093418 12.4 5359 -4735 RB8421 1.55 156 471 conserved hypothetical protein, membrane 4501840 4502310 10.3 16657 +5026 RB8966 2.04 660 1983 Chaperonin Cpn60/TCP-1 4777493 4779475 5.3 69819 +5094 RB9105 2.31 704 2115 Heat shock protein Hsp70 dnaK 4868834 4866720 4.6 76038 -5234 RB9372 2.43 205 618 Imidazole glycerol phosphate synthase subunit hisH hisH 2.4.2.- 5021363 5020746 4.9 22047 -5377 RB9635 1.51 352 1059 hypothetical protein 5187031 5185973 5.9 40195 -5571 RB9999 2.22 281 846 conserved hypothetical protein-putative transcriptional regulator 5386828 5385983 4.6 29948 -
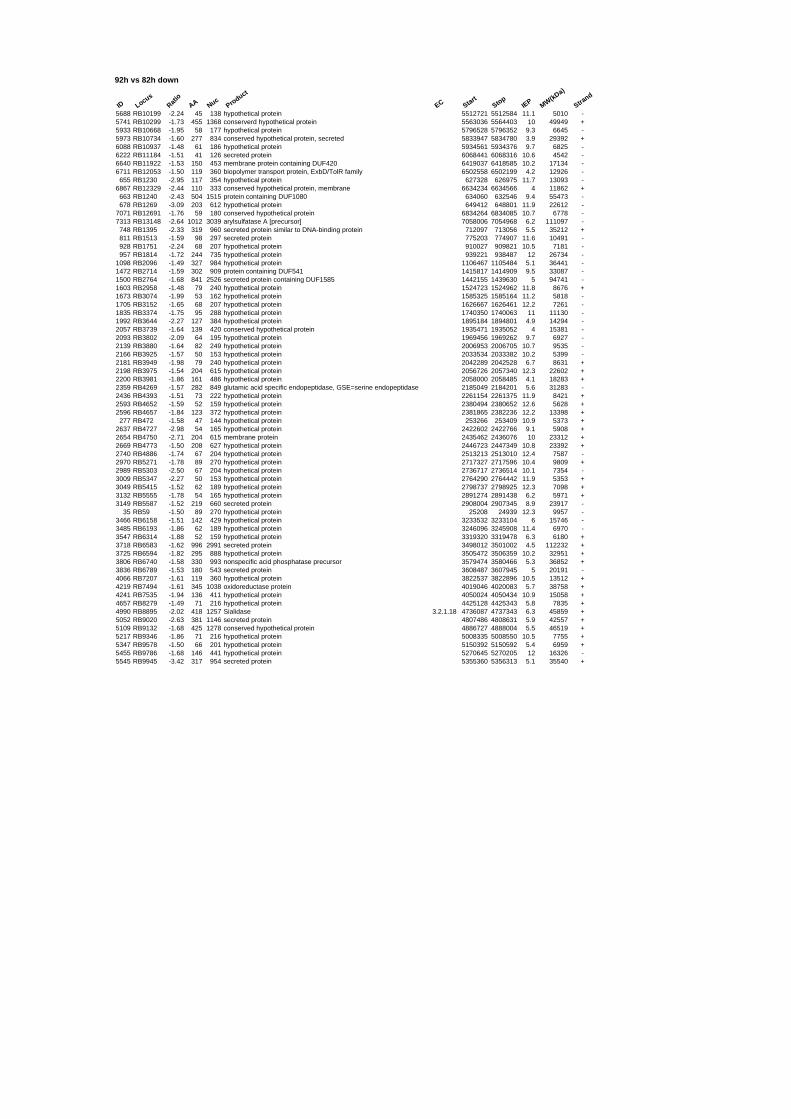
92h vs 82h down
ID LocusRati
oAA Nuc
Product
EC StartStop
IEP MW(kDa)
Strand
5688 RB10199 -2.24 45 138 hypothetical protein 5512721 5512584 11.1 5010 -5741 RB10299 -1.73 455 1368 conserverd hypothetical protein 5563036 5564403 10 49949 +5933 RB10668 -1.95 58 177 hypothetical protein 5796528 5796352 9.3 6645 -5973 RB10734 -1.60 277 834 conserved hypothetical protein, secreted 5833947 5834780 3.9 29392 +6088 RB10937 -1.48 61 186 hypothetical protein 5934561 5934376 9.7 6825 -6222 RB11184 -1.51 41 126 secreted protein 6068441 6068316 10.6 4542 -6640 RB11922 -1.53 150 453 membrane protein containing DUF420 6419037 6418585 10.2 17134 -6711 RB12053 -1.50 119 360 biopolymer transport protein, ExbD/TolR family 6502558 6502199 4.2 12926 -655 RB1230 -2.95 117 354 hypothetical protein 627328 626975 11.7 13093 -
6867 RB12329 -2.44 110 333 conserved hypothetical protein, membrane 6634234 6634566 4 11862 +663 RB1240 -2.43 504 1515 protein containing DUF1080 634060 632546 9.4 55473 -678 RB1269 -3.09 203 612 hypothetical protein 649412 648801 11.9 22612 -
7071 RB12691 -1.76 59 180 conserved hypothetical protein 6834264 6834085 10.7 6778 -7313 RB13148 -2.64 1012 3039 arylsulfatase A [precursor] 7058006 7054968 6.2 111097 -748 RB1395 -2.33 319 960 secreted protein similar to DNA-binding protein 712097 713056 5.5 35212 +811 RB1513 -1.59 98 297 secreted protein 775203 774907 11.6 10491 -928 RB1751 -2.24 68 207 hypothetical protein 910027 909821 10.5 7181 -957 RB1814 -1.72 244 735 hypothetical protein 939221 938487 12 26734 -
1098 RB2096 -1.49 327 984 hypothetical protein 1106467 1105484 5.1 36441 -1472 RB2714 -1.59 302 909 protein containing DUF541 1415817 1414909 9.5 33087 -1500 RB2764 -1.68 841 2526 secreted protein containing DUF1585 1442155 1439630 5 94741 -1603 RB2958 -1.48 79 240 hypothetical protein 1524723 1524962 11.8 8676 +1673 RB3074 -1.99 53 162 hypothetical protein 1585325 1585164 11.2 5818 -1705 RB3152 -1.65 68 207 hypothetical protein 1626667 1626461 12.2 7261 -1835 RB3374 -1.75 95 288 hypothetical protein 1740350 1740063 11 11130 -1992 RB3644 -2.27 127 384 hypothetical protein 1895184 1894801 4.9 14294 -2057 RB3739 -1.64 139 420 conserved hypothetical protein 1935471 1935052 4 15381 -2093 RB3802 -2.09 64 195 hypothetical protein 1969456 1969262 9.7 6927 -2139 RB3880 -1.64 82 249 hypothetical protein 2006953 2006705 10.7 9535 -2166 RB3925 -1.57 50 153 hypothetical protein 2033534 2033382 10.2 5399 -2181 RB3949 -1.98 79 240 hypothetical protein 2042289 2042528 6.7 8631 +2198 RB3975 -1.54 204 615 hypothetical protein 2056726 2057340 12.3 22602 +2200 RB3981 -1.86 161 486 hypothetical protein 2058000 2058485 4.1 18283 +2359 RB4269 -1.57 282 849 glutamic acid specific endopeptidase, GSE=serine endopeptidase 2185049 2184201 5.6 31283 -2436 RB4393 -1.51 73 222 hypothetical protein 2261154 2261375 11.9 8421 +2593 RB4652 -1.59 52 159 hypothetical protein 2380494 2380652 12.6 5628 +2596 RB4657 -1.84 123 372 hypothetical protein 2381865 2382236 12.2 13398 +277 RB472 -1.58 47 144 hypothetical protein 253266 253409 10.9 5373 +
2637 RB4727 -2.98 54 165 hypothetical protein 2422602 2422766 9.1 5908 +2654 RB4750 -2.71 204 615 membrane protein 2435462 2436076 10 23312 +2669 RB4773 -1.50 208 627 hypothetical protein 2446723 2447349 10.8 23392 +2740 RB4886 -1.74 67 204 hypothetical protein 2513213 2513010 12.4 7587 -2970 RB5271 -1.78 89 270 hypothetical protein 2717327 2717596 10.4 9809 +2989 RB5303 -2.50 67 204 hypothetical protein 2736717 2736514 10.1 7354 -3009 RB5347 -2.27 50 153 hypothetical protein 2764290 2764442 11.9 5353 +3049 RB5415 -1.52 62 189 hypothetical protein 2798737 2798925 12.3 7098 +3132 RB5555 -1.78 54 165 hypothetical protein 2891274 2891438 6.2 5971 +3149 RB5587 -1.52 219 660 secreted protein 2908004 2907345 8.9 23917 -
35 RB59 -1.50 89 270 hypothetical protein 25208 24939 12.3 9957 -3466 RB6158 -1.51 142 429 hypothetical protein 3233532 3233104 6 15746 -3485 RB6193 -1.86 62 189 hypothetical protein 3246096 3245908 11.4 6970 -3547 RB6314 -1.88 52 159 hypothetical protein 3319320 3319478 6.3 6180 +3718 RB6583 -1.62 996 2991 secreted protein 3498012 3501002 4.5 112232 +3725 RB6594 -1.82 295 888 hypothetical protein 3505472 3506359 10.2 32951 +3806 RB6740 -1.58 330 993 nonspecific acid phosphatase precursor 3579474 3580466 5.3 36852 +3836 RB6789 -1.53 180 543 secreted protein 3608487 3607945 5 20191 -4066 RB7207 -1.61 119 360 hypothetical protein 3822537 3822896 10.5 13512 +4219 RB7494 -1.61 345 1038 oxidoreductase protein 4019046 4020083 5.7 38758 +4241 RB7535 -1.94 136 411 hypothetical protein 4050024 4050434 10.9 15058 +4657 RB8279 -1.49 71 216 hypothetical protein 4425128 4425343 5.8 7835 +4990 RB8895 -2.02 418 1257 Sialidase 3.2.1.18 4736087 4737343 6.3 45859 +5052 RB9020 -2.63 381 1146 secreted protein 4807486 4808631 5.9 42557 +5109 RB9132 -1.68 425 1278 conserved hypothetical protein 4886727 4888004 5.5 46519 +5217 RB9346 -1.86 71 216 hypothetical protein 5008335 5008550 10.5 7755 +5347 RB9578 -1.50 66 201 hypothetical protein 5150392 5150592 5.4 6959 +5455 RB9786 -1.68 146 441 hypothetical protein 5270645 5270205 12 16326 -5545 RB9945 -3.42 317 954 secreted protein 5355360 5356313 5.1 35540 +

92h vs 82h upID Locus Ratio Parent ID AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand5576 RB10007 2.48 1 313 942 conserved hypothetical protein 5390130 5391071 5 34128 +5647 RB10126 2.24 1 43 132 hypothetical protein 5465210 5465341 12.9 4964 +5694 RB10209 2.01 1 110 333 H+-transporting two-sector ATPase, C subunit 3.6.3.14 5515135 5515467 10.2 11565 +5722 RB10261 3.21 1 81 246 Carbon storage regulator csrA 5545964 5545719 5.1 8704 -5723 RB10263 2.07 1 58 177 hypothetical protein 5546338 5546162 9.6 6643 -5754 RB10326 1.84 1 588 1767 sulfate permease family protein 5580833 5582599 6 62785 +5765 RB10342 2.17 1 102 309 hypothetical protein 5593929 5593621 12.2 11300 -5823 RB10458 1.54 1 206 621 Rnpc2-prov protein 5671130 5670510 10.3 21043 -5838 RB10483 1.84 1 133 402 conserved hypothetical protein 5685437 5685838 5.7 14433 +5911 RB10627 2.48 1 93 282 Chaperonin Cpn10 5769148 5769429 3.9 9966 +5967 RB10727 3.91 1 276 831 manganese-containing catalase 5830775 5831605 5 30055 +6043 RB10861 3.60 1 46 141 hypothetical protein 5896851 5896991 4.2 4833 +6047 RB10867 1.71 1 207 624 hypothetical protein 5899359 5899982 5.7 23041 +6085 RB10932 3.11 1 65 198 secreted protein 5933872 5934069 3.8 6983 +6089 RB10939 1.69 1 62 189 conserved hypothetical protein 5934628 5934816 4.3 6627 +6096 RB10951 1.51 1 101 306 secreted protein 5940232 5939927 10.2 11754 -6100 RB10958 3.41 1 158 477 hypothetical protein 5941015 5941491 5.4 18101 +6103 RB10966 2.02 1 60 183 Protein of unknown function, UPF0057 5942934 5942752 10.1 6793 -6104 RB10967 2.40 1 341 1026 quinone oxidoreductase-like protein 5944040 5943015 4.5 37393 -6106 RB10971 2.81 1 303 912 Short-chain dehydrogenase/reductase SDR 5946406 5945495 9.8 32799 -6127 RB11008 3.55 1 394 1185 secreted protein 5967078 5968262 5.1 42891 +6133 RB11016 1.59 1 369 1110 protein containing DUF1559 5974211 5973102 5.4 39469 -6135 RB11021 1.60 1 295 888 ATP-binding protein of ABC transporter 5975385 5974498 5 31784 -6140 RB11031 1.66 1 62 189 hypothetical protein 5980865 5981053 12.1 7649 +6144 RB11041 1.79 1 349 1050 secreted protein containing DUF1559 5987234 5988283 6.1 38326 +6157 RB11070 1.71 1 52 159 hypothetical protein 6000321 6000479 10.2 5582 +6201 RB11150 1.94 1 385 1158 NADH:flavin oxidoreductase/NADH oxidase 6046862 6048019 4.5 42128 +6216 RB11174 1.63 1 469 1410 metallo-beta-lactamase family protein 6065046 6063637 4.8 51160 -6265 RB11258 2.39 1 237 714 conserved hypothetical protein, membrane 6106840 6106127 10.3 26175 -6268 RB11266 1.84 1 304 915 3',5'-cyclic-nucleotide phosphodiesterase 6108475 6109389 9.7 33880 +6269 RB11268 1.83 1 97 294 hypothetical protein 6109768 6109475 10.8 11235 -6346 RB11396 1.50 1 36 111 conserved hypothetical protein 6178445 6178555 11.9 4104 +6347 RB11397 3.07 1 207 624 SOUL heme-binding protein 6178540 6179163 8.7 23072 +6348 RB11399 2.49 1 132 399 conserved hypothetical protein 6179167 6179565 6.5 15047 +6362 RB11424 4.52 1 158 477 conserved hypothetical protein 6195656 6196132 11.1 17372 +6363 RB11427 2.45 1 93 282 conserved hypothetical protein 6196136 6196417 9.7 10315 +6364 RB11429 4.88 1 37 114 conserved hypothetical protein 6196478 6196591 12.4 4245 +6367 RB11434 2.50 1 69 210 conserved hypothetical protein 6197442 6197233 9.8 7670 -6371 RB11439 4.95 1 93 282 conserved hypothetical protein 6197993 6198274 10.6 10906 +6382 RB11446 2.40 1 113 342 conserved hypothetical protein 6199492 6199833 10 12579 +6385 RB11450 3.94 1 60 183 conserved hypothetical protein 6200046 6200228 11.8 6517 +6414 RB11488 1.74 1 265 798 oxidoreductase, short-chain dehydrogenase/reductase family 6213893 6214690 5.1 27687 +6417 RB11492 1.64 1 95 288 conserved hypothetical protein, membrane 6216103 6216390 12.3 10176 +6425 RB11505 1.68 1 199 600 conserved hypothetical protein, secreted 6221829 6221230 7.5 21660 -6718 RB12066 2.33 1 135 408 hypothetical protein 6510147 6510554 10 14924 +6766 RB12160 2.53 1 108 327 thioredoxin trxA 6563104 6562778 4 11608 -6836 RB12283 2.92 1 52 159 conserved hypothetical protein 6609220 6609378 7.6 5927 +6837 RB12285 2.19 1 224 675 phospholipid N-methyltransferase 2.1.1.17 6609391 6610065 6.8 24262 +6846 RB12297 2.40 1 169 510 conserved hypothetical protein 6615598 6616107 4.4 17963 +6884 RB12362 2.03 1 274 825 bacterioferritin comigratory protein 6653786 6652962 5.8 30428 -6890 RB12377 1.49 1 284 855 Ribosomal RNA adenine methylase transferase 6661963 6661109 5.7 31560 -673 RB1259 1.79 1 59 180 hypothetical protein 642452 642631 10.5 6484 +
7069 RB12689 2.18 1 106 321 secreted protein 6833004 6833324 9.8 11239 +7149 RB12830 1.83 1 102 309 conserved hypothetical protein 6920508 6920200 10.9 11753 -7153 RB12837 1.92 1 65 198 conserved hypothetical protein 6921732 6921929 9.8 7318 +7295 RB13117 1.68 1 156 471 nitrogen regulation protein 7036595 7037065 4.6 17819 +7341 RB13205 2.60 1 75 228 hypothetical protein 7086564 7086337 12.4 8283 -7342 RB13207 3.98 1 58 177 secreted protein 7086761 7086585 11.9 6443 -707 RB1322 2.28 1 35 108 hypothetical protein 671526 671633 12.8 3914 +
7369 RB13257 2.81 1 286 861 2-hydroxy-6-ketonona-2,4-dienedioic acid hydrolase bphD 3.7.1.- 7105590 7106450 4.6 32540 +733 RB1362 1.85 1 82 249 hypothetical protein 696352 696600 10.4 9054 +804 RB1501 2.68 1 50 153 hypothetical protein 769419 769571 7.3 5710 +839 RB1563 2.09 1 136 411 hypothetical protein 798934 799344 10.2 14882 +883 RB1644 3.27 1 123 372 secreted protein 845824 845453 9.7 13720 -915 RB1728 2.29 1 285 858 membrane protein containing DUF1206 899954 900811 9.9 30055 +946 RB1787 1.88 1 72 219 hypothetical protein 925904 925686 9.6 8167 -
1053 RB2011 2.47 1 93 282 hypothetical protein 1059411 1059692 4.4 9880 +1182 RB2244 1.85 1 192 579 Glutathione peroxidase GPXMC1 1.11.1.12 1178675 1179253 5.2 20717 +1183 RB2247 3.12 1 579 1740 conserved hypothetical protein containing YTV domains 1181238 1179499 9.8 65013 -1275 RB2382 1.61 1 75 228 hypothetical protein 1238921 1239148 9.7 8092 +1278 RB2386 1.98 1 528 1587 RND efflux membrane fusion protein 1244320 1242734 6 58921 -1280 RB2394 3.13 1 236 711 conserved hypothetical protein 1245630 1244920 7 27126 -1341 RB2492 2.12 1 69 210 hypothetical protein 1302190 1302399 9.9 7373 +1364 RB2516 1.76 1 63 192 hypothetical protein 1316669 1316860 11.2 6506 +1428 RB2636 2.94 1 116 351 hypothetical protein 1368450 1368100 11.7 12761 -1452 RB2679 1.53 1 173 522 hypothetical protein 1391557 1392078 5.5 19502 +1453 RB2680 2.91 1 467 1404 conserved hypothetical protein, membrane 1393566 1392163 6.4 49834 -1505 RB2782 3.32 1 589 1770 protein containing DUF1559 1445576 1447345 7.2 63826 +1566 RB2900 1.94 1 90 273 hypothetical protein 1506927 1507199 9.1 9939 +1567 RB2901 2.39 1 392 1179 conserved hypothetical protein, secreted 1507210 1508388 4.8 42805 +177 RB309 1.94 1 482 1449 Magnesium chelatase, ChlI subunit chlI 6.6.1.1 156441 154993 5.1 53979 -
1786 RB3295 2.35 1 140 423 secreted protein 1692681 1692259 3.9 14786 -1787 RB3298 2.24 1 39 120 hypothetical protein 1692797 1692678 12.2 4684 -1882 RB3455 2.38 1 57 174 hypothetical protein 1785458 1785631 10.2 6258 +2024 RB3686 2.12 1 327 984 oxidoreductase, zinc-binding 1910523 1909540 5.5 35045 -2059 RB3742 2.18 1 718 2157 Flagellin, N-terminal 1935693 1937849 3.8 72031 +2239 RB4051 1.68 1 180 543 hypothetical protein 2092752 2092210 10.1 20712 -2243 RB4060 2.42 1 190 573 TadE-like 2095403 2094831 10.2 20868 -2308 RB4185 1.51 1 89 270 hypothetical protein 2151951 2152220 12 9730 +2338 RB4230 2.31 1 331 996 conserved hypothetical protein 2168783 2167788 5.3 37197 -2351 RB4256 3.13 1 358 1077 conserved hypothetical protein, secreted 2180303 2181379 4.5 37091 +2401 RB4336 2.54 1 372 1119 conserved hypothetical protein 2231697 2232815 5.6 41876 +2429 RB4381 2.50 1 59 180 hypothetical protein 2255621 2255442 10.7 7087 -2430 RB4382 1.53 1 555 1668 Diaminopimelate decarboxylase lysA 4.1.1.20 2257285 2255618 7.7 62732 -2437 RB4394 1.61 1 188 567 protease I pfpI 3.2.-.- 2261372 2261938 4.4 20601 +2440 RB4397 2.12 1 55 168 protein containing DUF1560 2263976 2264143 10.5 5816 +2442 RB4399 2.02 1 328 987 zinc-binding oxidoreductase 2265237 2264251 4.6 35121 -2443 RB4400 2.51 1 58 177 hypothetical protein 2265382 2265206 9 6610 -2445 RB4404 1.82 1 302 909 oxidoreductase, short chain dehydrogenase/reductase family 2265539 2266447 4.9 32804 +2453 RB4419 2.71 1 178 537 MgtC/SapB transporter 2272912 2273448 6.2 18995 +2456 RB4423 1.72 1 165 498 cytidine and deoxycytidylate deaminase family protein 2275085 2274588 4.7 17104 -2462 RB4432 2.69 1 268 807 glucose 1-dehydrogenase or 3-oxoacyl-[acyl-carrier-protein] reductase 1.1.1.- 2276333 2277139 4.7 28181 +2464 RB4436 1.84 1 90 273 membrane protein 2277820 2278092 12.5 9479 +2478 RB4460 1.51 1 422 1269 conserved hypothetical protein, membrane 2289696 2288428 4.8 48179 -2481 RB4464 1.87 1 223 672 secreted protein 2292075 2291404 5.1 25669 -2485 RB4468 2.36 1 162 489 hypothetical protein 2294104 2293616 4.6 16380 -2486 RB4470 2.85 1 169 510 secreted protein 2294676 2294167 4.9 19277 -2488 RB4474 3.76 1 168 507 CsbD-like 2295048 2295554 8.8 18451 +2489 RB4476 3.10 1 128 387 hypothetical protein 2295557 2295943 11.4 14037 +2490 RB4477 2.70 1 69 210 hypothetical protein 2295940 2296149 10.6 7470 +2491 RB4478 2.30 1 172 519 membrane protein 2296122 2296640 7.3 18536 +2492 RB4480 1.85 1 89 270 hypothetical protein 2296646 2296915 6.1 9442 +2494 RB4484 2.16 1 432 1299 Peptidase M20 3.-.-.- 2297267 2298565 6 46450 +2496 RB4487 1.93 1 514 1545 two-component system response regulator 2299859 2301403 4.6 56207 +
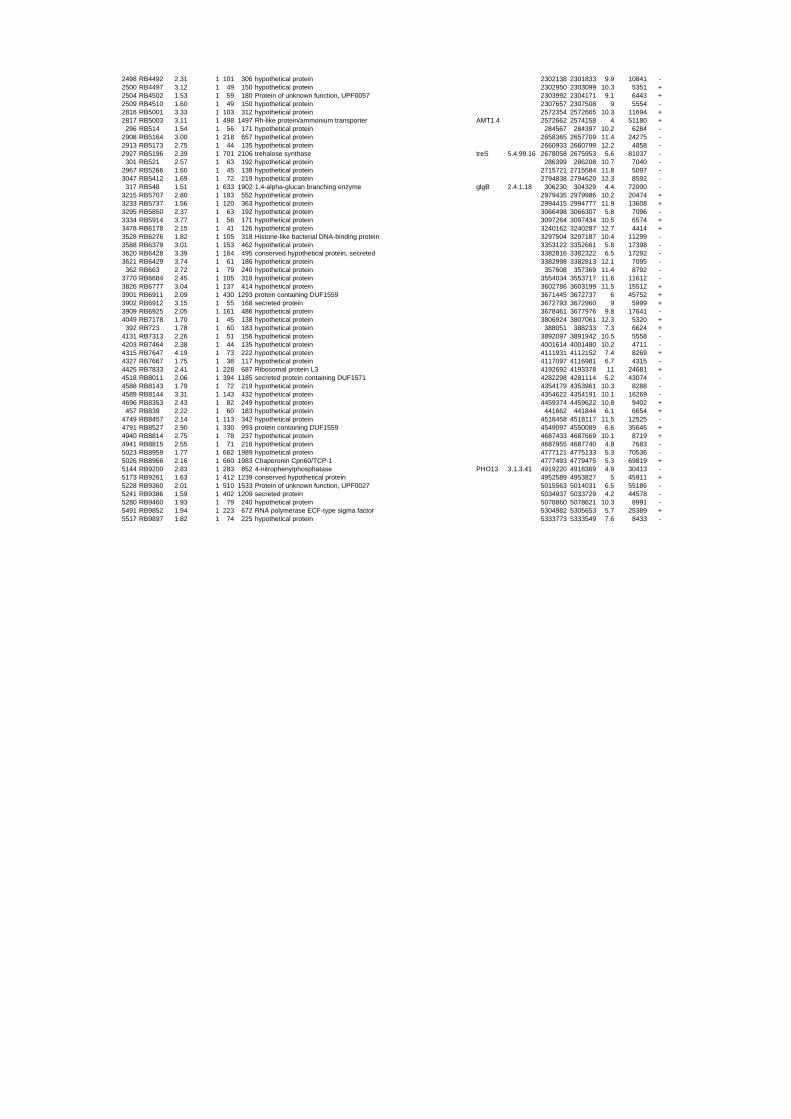
2498 RB4492 2.31 1 101 306 hypothetical protein 2302138 2301833 9.9 10841 -2500 RB4497 3.12 1 49 150 hypothetical protein 2302950 2303099 10.3 5351 +2504 RB4502 1.53 1 59 180 Protein of unknown function, UPF0057 2303992 2304171 9.1 6443 +2509 RB4510 1.60 1 49 150 hypothetical protein 2307657 2307508 9 5554 -2816 RB5001 3.33 1 103 312 hypothetical protein 2572354 2572665 10.3 11694 +2817 RB5003 3.11 1 498 1497 Rh-like protein/ammonium transporter AMT1.4 2572662 2574158 4 51180 +296 RB514 1.54 1 56 171 hypothetical protein 284567 284397 10.2 6284 -
2908 RB5164 3.00 1 218 657 hypothetical protein 2658365 2657709 11.4 24275 -2913 RB5173 2.75 1 44 135 hypothetical protein 2660933 2660799 12.2 4858 -2927 RB5196 2.39 1 701 2106 trehalose synthase treS 5.4.99.16 2678058 2675953 5.6 81037 -301 RB521 2.57 1 63 192 hypothetical protein 286399 286208 10.7 7040 -
2967 RB5266 1.60 1 45 138 hypothetical protein 2715721 2715584 11.8 5097 -3047 RB5412 1.69 1 72 219 hypothetical protein 2794838 2794620 12.3 8592 -317 RB548 1.51 1 633 1902 1,4-alpha-glucan branching enzyme glgB 2.4.1.18 306230 304329 4.4 72000 -
3215 RB5707 2.80 1 183 552 hypothetical protein 2979435 2979986 10.2 20474 +3233 RB5737 1.56 1 120 363 hypothetical protein 2994415 2994777 11.9 13608 +3295 RB5850 2.37 1 63 192 hypothetical protein 3066498 3066307 5.8 7096 -3334 RB5914 3.77 1 56 171 hypothetical protein 3097264 3097434 10.5 6574 +3478 RB6178 2.15 1 41 126 hypothetical protein 3240162 3240287 12.7 4414 +3528 RB6276 1.82 1 105 318 Histone-like bacterial DNA-binding protein 3297504 3297187 10.4 11299 -3588 RB6379 3.01 1 153 462 hypothetical protein 3353122 3352661 5.8 17398 -3620 RB6428 3.39 1 164 495 conserved hypothetical protein, secreted 3382816 3382322 6.5 17292 -3621 RB6429 3.74 1 61 186 hypothetical protein 3382998 3382813 12.1 7095 -362 RB663 2.72 1 79 240 hypothetical protein 357608 357369 11.4 8792 -
3770 RB6684 2.45 1 105 318 hypothetical protein 3554034 3553717 11.6 11612 -3826 RB6777 3.04 1 137 414 hypothetical protein 3602786 3603199 11.5 15512 +3901 RB6911 2.09 1 430 1293 protein containing DUF1559 3671445 3672737 6 45752 +3902 RB6912 3.15 1 55 168 secreted protein 3672793 3672960 9 5999 +3909 RB6925 2.05 1 161 486 hypothetical protein 3678461 3677976 9.8 17641 -4049 RB7178 1.70 1 45 138 hypothetical protein 3806924 3807061 12.3 5320 +392 RB723 1.78 1 60 183 hypothetical protein 388051 388233 7.3 6624 +
4131 RB7313 2.26 1 51 156 hypothetical protein 3892097 3891942 10.5 5558 -4203 RB7464 2.38 1 44 135 hypothetical protein 4001614 4001480 10.2 4711 -4315 RB7647 4.19 1 73 222 hypothetical protein 4111931 4112152 7.4 8269 +4327 RB7667 1.75 1 38 117 hypothetical protein 4117097 4116981 6.7 4315 -4425 RB7833 2.41 1 228 687 Ribosomal protein L3 4192692 4193378 11 24681 +4518 RB8011 2.06 1 394 1185 secreted protein containing DUF1571 4282298 4281114 5.2 43074 -4588 RB8143 1.79 1 72 219 hypothetical protein 4354179 4353961 10.3 8288 -4589 RB8144 3.31 1 143 432 hypothetical protein 4354622 4354191 10.1 16269 -4696 RB8353 2.43 1 82 249 hypothetical protein 4459374 4459622 10.8 9402 +457 RB839 2.22 1 60 183 hypothetical protein 441662 441844 6.1 6654 +
4749 RB8457 2.14 1 113 342 hypothetical protein 4518458 4518117 11.5 12525 -4791 RB8527 2.90 1 330 993 protein containing DUF1559 4549097 4550089 6.6 35645 +4940 RB8814 2.75 1 78 237 hypothetical protein 4687433 4687669 10.1 8719 +4941 RB8815 2.55 1 71 216 hypothetical protein 4687955 4687740 4.8 7683 -5023 RB8959 1.77 1 662 1989 hypothetical protein 4777121 4775133 5.3 70536 -5026 RB8966 2.16 1 660 1983 Chaperonin Cpn60/TCP-1 4777493 4779475 5.3 69819 +5144 RB9200 2.83 1 283 852 4-nitrophenylphosphatase PHO13 3.1.3.41 4919220 4918369 4.9 30413 -5173 RB9261 1.63 1 412 1239 conserved hypothetical protein 4952589 4953827 5 45911 +5228 RB9360 2.01 1 510 1533 Protein of unknown function, UPF0027 5015563 5014031 6.5 55186 -5241 RB9386 1.59 1 402 1209 secreted protein 5034937 5033729 4.2 44578 -5280 RB9460 1.93 1 79 240 hypothetical protein 5078860 5078621 10.3 8991 -5491 RB9852 1.94 1 223 672 RNA polymerase ECF-type sigma factor 5304982 5305653 5.7 25389 +5517 RB9897 1.82 1 74 225 hypothetical protein 5333773 5333549 7.6 8433 -
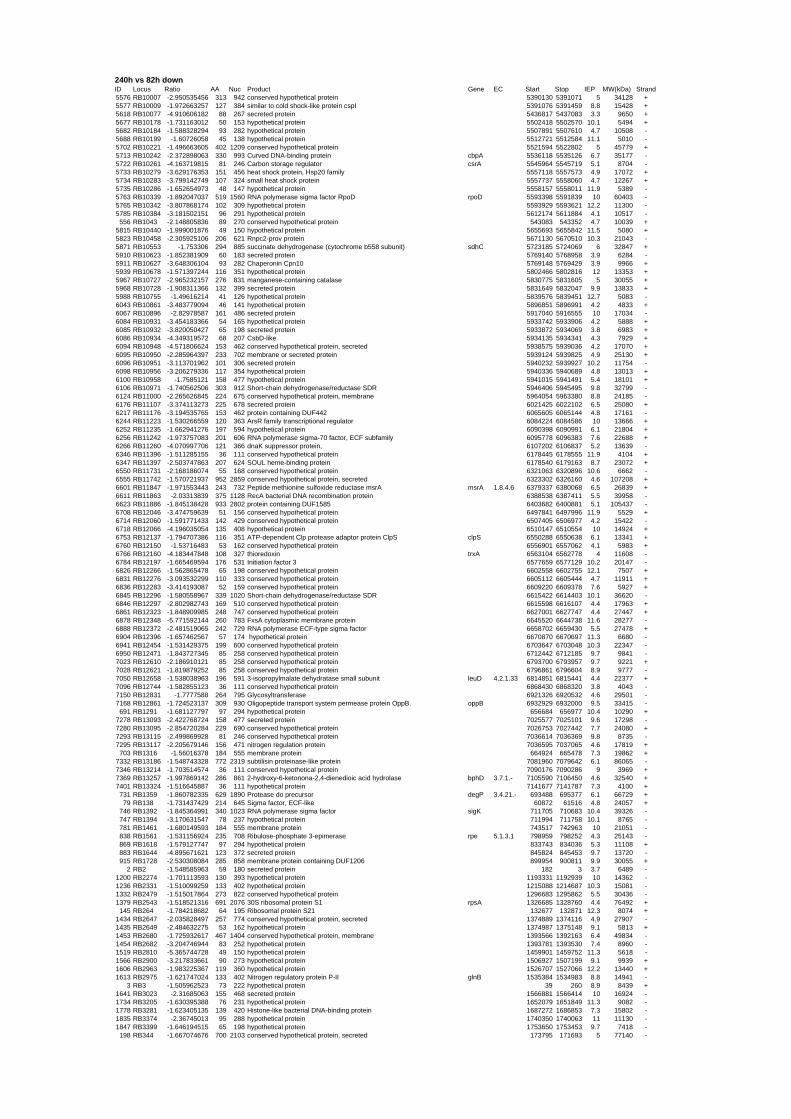
240h vs 82h downID Locus Ratio AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand5576 RB10007 -2.950535456 313 942 conserved hypothetical protein 5390130 5391071 5 34128 +5577 RB10009 -1.972663257 127 384 similar to cold shock-like protein cspI 5391076 5391459 8.8 15428 +5618 RB10077 -4.910606182 88 267 secreted protein 5436817 5437083 3.3 9650 +5677 RB10178 -1.731163012 50 153 hypothetical protein 5502418 5502570 10.1 5494 +5682 RB10184 -1.588328294 93 282 hypothetical protein 5507891 5507610 4.7 10508 -5688 RB10199 -1.60726058 45 138 hypothetical protein 5512721 5512584 11.1 5010 -5702 RB10221 -1.496663605 402 1209 conserved hypothetical protein 5521594 5522802 5 45779 +5713 RB10242 -2.372898063 330 993 Curved DNA-binding protein cbpA 5536118 5535126 6.7 35177 -5722 RB10261 -4.163719815 81 246 Carbon storage regulator csrA 5545964 5545719 5.1 8704 -5733 RB10279 -3.629176353 151 456 heat shock protein, Hsp20 family 5557118 5557573 4.9 17072 +5734 RB10283 -3.799142749 107 324 small heat shock protein 5557737 5558060 4.7 12267 +5735 RB10286 -1.652654973 48 147 hypothetical protein 5558157 5558011 11.9 5389 -5763 RB10339 -1.892047037 519 1560 RNA polymerase sigma factor RpoD rpoD 5593398 5591839 10 60403 -5765 RB10342 -3.807868174 102 309 hypothetical protein 5593929 5593621 12.2 11300 -5785 RB10384 -3.181502151 96 291 hypothetical protein 5612174 5611884 4.1 10517 -556 RB1043 -2.148805836 89 270 conserved hypothetical protein 543083 543352 4.7 10039 +
5815 RB10440 -1.999001876 49 150 hypothetical protein 5655693 5655842 11.5 5080 +5823 RB10458 -2.305925106 206 621 Rnpc2-prov protein 5671130 5670510 10.3 21043 -5871 RB10553 -1.753306 294 885 succinate dehydrogenase (cytochrome b558 subunit) sdhC 5723185 5724069 6 32847 +5910 RB10623 -1.852381909 60 183 secreted protein 5769140 5768958 3.9 6284 -5911 RB10627 -3.648306104 93 282 Chaperonin Cpn10 5769148 5769429 3.9 9966 +5939 RB10678 -1.571397244 116 351 hypothetical protein 5802466 5802816 12 13353 +5967 RB10727 -2.965232157 276 831 manganese-containing catalase 5830775 5831605 5 30055 +5968 RB10728 -1.908311366 132 399 secreted protein 5831649 5832047 9.9 13833 +5988 RB10755 -1.49616214 41 126 hypothetical protein 5839576 5839451 12.7 5083 -6043 RB10861 -3.483779094 46 141 hypothetical protein 5896851 5896991 4.2 4833 +6067 RB10896 -2.82978587 161 486 secreted protein 5917040 5916555 10 17034 -6084 RB10931 -3.454183366 54 165 hypothetical protein 5933742 5933906 4.2 5888 +6085 RB10932 -3.820050427 65 198 secreted protein 5933872 5934069 3.8 6983 +6086 RB10934 -4.349319572 68 207 CsbD-like 5934135 5934341 4.3 7929 +6094 RB10948 -4.571806624 153 462 conserved hypothetical protein, secreted 5938575 5939036 4.2 17070 +6095 RB10950 -2.285964397 233 702 membrane or secreted protein 5939124 5939825 4.9 25130 +6096 RB10951 -3.113701962 101 306 secreted protein 5940232 5939927 10.2 11754 -6098 RB10956 -3.206279336 117 354 hypothetical protein 5940336 5940689 4.8 13013 +6100 RB10958 -1.7585121 158 477 hypothetical protein 5941015 5941491 5.4 18101 +6106 RB10971 -1.740562506 303 912 Short-chain dehydrogenase/reductase SDR 5946406 5945495 9.8 32799 -6124 RB11000 -2.265626845 224 675 conserved hypothetical protein, membrane 5964054 5963380 8.8 24185 -6176 RB11107 -3.374113273 225 678 secreted protein 6021425 6022102 6.5 25080 +6217 RB11176 -3.194535765 153 462 protein containing DUF442 6065605 6065144 4.8 17161 -6244 RB11223 -1.530266559 120 363 ArsR family transcriptional regulator 6084224 6084586 10 13666 +6252 RB11235 -1.662941276 197 594 hypothetical protein 6090398 6090991 6.1 21804 +6256 RB11242 -1.973757083 201 606 RNA polymerase sigma-70 factor, ECF subfamily 6095778 6096383 7.6 22688 +6266 RB11260 -4.070997706 121 366 dnaK suppressor protein, 6107202 6106837 5.2 13639 -6346 RB11396 -1.511285155 36 111 conserved hypothetical protein 6178445 6178555 11.9 4104 +6347 RB11397 -2.503747863 207 624 SOUL heme-binding protein 6178540 6179163 8.7 23072 +6550 RB11731 -2.168186074 55 168 conserved hypothetical protein 6321063 6320896 10.6 6662 -6555 RB11742 -1.570721937 952 2859 conserved hypothetical protein, secreted 6323302 6326160 4.6 107208 +6601 RB11847 -1.971553443 243 732 Peptide methionine sulfoxide reductase msrA msrA 1.8.4.6 6379337 6380068 6.5 26839 +6611 RB11863 -2.03313839 375 1128 RecA bacterial DNA recombination protein 6388538 6387411 5.5 39958 -6623 RB11886 -1.845138428 933 2802 protein containing DUF1585 6403682 6400881 5.1 105437 -6708 RB12046 -3.474759639 51 156 conserved hypothetical protein 6497841 6497996 11.9 5529 +6714 RB12060 -1.591771433 142 429 conserved hypothetical protein 6507405 6506977 4.2 15422 -6718 RB12066 -4.196035054 135 408 hypothetical protein 6510147 6510554 10 14924 +6753 RB12137 -1.794707386 116 351 ATP-dependent Clp protease adaptor protein ClpS clpS 6550288 6550638 6.1 13341 +6760 RB12150 -1.53716483 53 162 conserved hypothetical protein 6556901 6557062 4.1 5983 +6766 RB12160 -4.183447848 108 327 thioredoxin trxA 6563104 6562778 4 11608 -6784 RB12197 -1.665469594 176 531 Initiation factor 3 6577659 6577129 10.2 20147 -6826 RB12266 -1.562865478 65 198 conserved hypothetical protein 6602558 6602755 12.1 7507 +6831 RB12276 -3.093532299 110 333 conserved hypothetical protein 6605112 6605444 4.7 11911 +6836 RB12283 -3.414193087 52 159 conserved hypothetical protein 6609220 6609378 7.6 5927 +6845 RB12296 -1.580558967 339 1020 Short-chain dehydrogenase/reductase SDR 6615422 6614403 10.1 36620 -6846 RB12297 -2.802982743 169 510 conserved hypothetical protein 6615598 6616107 4.4 17963 +6861 RB12323 -1.848909985 248 747 conserved hypothetical protein 6627001 6627747 4.4 27447 +6878 RB12348 -5.771592144 260 783 FxsA cytoplasmic membrane protein 6645520 6644738 11.6 28277 -6888 RB12372 -2.481519065 242 729 RNA polymerase ECF-type sigma factor 6658702 6659430 5.5 27478 +6904 RB12396 -1.657462567 57 174 hypothetical protein 6670870 6670697 11.3 6680 -6941 RB12454 -1.531429375 199 600 conserved hypothetical protein 6703647 6703048 10.3 22347 -6950 RB12471 -1.843727345 85 258 conserved hypothetical protein 6712442 6712185 9.7 9841 -7023 RB12610 -2.186910121 85 258 conserved hypothetical protein 6793700 6793957 9.7 9221 +7028 RB12621 -1.819879252 85 258 conserved hypothetical protein 6796861 6796604 8.9 9777 -7050 RB12658 -1.538038963 196 591 3-isopropylmalate dehydratase small subunit leuD 4.2.1.33 6814851 6815441 4.4 22377 +7096 RB12744 -1.582855123 36 111 conserved hypothetical protein 6868430 6868320 3.8 4043 -7150 RB12831 -1.7777588 264 795 Glycosyltransferase 6921326 6920532 4.6 29501 -7168 RB12861 -1.724523137 309 930 Oligopeptide transport system permease protein OppB. oppB 6932929 6932000 9.5 33415 -691 RB1291 -1.681127797 97 294 hypothetical protein 656684 656977 10.4 10290 +
7278 RB13093 -2.422768724 158 477 secreted protein 7025577 7025101 9.6 17298 -7280 RB13095 -2.854720284 229 690 conserved hypothetical protein 7026753 7027442 7.7 24080 +7293 RB13115 -2.499869928 81 246 conserved hypothetical protein 7036614 7036369 9.8 8735 -7295 RB13117 -2.205679146 156 471 nitrogen regulation protein 7036595 7037065 4.6 17819 +703 RB1316 -1.56016378 184 555 membrane protein 664924 665478 7.3 19862 +
7332 RB13186 -1.548743328 772 2319 subtilisin proteinase-like protein 7081960 7079642 6.1 86065 -7346 RB13214 -1.703514574 36 111 conserved hypothetical protein 7090176 7090286 9 3969 +7369 RB13257 -1.997869142 286 861 2-hydroxy-6-ketonona-2,4-dienedioic acid hydrolase bphD 3.7.1.- 7105590 7106450 4.6 32540 +7401 RB13324 -1.516645887 36 111 hypothetical protein 7141677 7141787 7.3 4100 +731 RB1359 -1.860782335 629 1890 Protease do precursor degP 3.4.21.- 693488 695377 6.1 66729 +79 RB138 -1.731437429 214 645 Sigma factor, ECF-like 60872 61516 4.8 24057 +
746 RB1392 -1.845364991 340 1023 RNA polymerase sigma factor sigK 711705 710683 10.4 39326 -747 RB1394 -3.170631547 78 237 hypothetical protein 711994 711758 10.1 8765 -781 RB1461 -1.680149593 184 555 membrane protein 743517 742963 10 21051 -838 RB1561 -1.531156924 235 708 Ribulose-phosphate 3-epimerase rpe 5.1.3.1 798959 798252 4.3 25143 -869 RB1618 -1.579127747 97 294 hypothetical protein 833743 834036 5.3 11108 +883 RB1644 -4.895671621 123 372 secreted protein 845824 845453 9.7 13720 -915 RB1728 -2.530308084 285 858 membrane protein containing DUF1206 899954 900811 9.9 30055 +
2 RB2 -1.548585963 59 180 secreted protein 182 3 3.7 6489 -1200 RB2274 -1.701113593 130 393 hypothetical protein 1193331 1192939 10 14362 -1236 RB2331 -1.510099259 133 402 hypothetical protein 1215088 1214687 10.3 15081 -1332 RB2479 -1.515017864 273 822 conserved hypothetical protein 1296683 1295862 5.5 30436 -1379 RB2543 -1.518521316 691 2076 30S ribosomal protein S1 rpsA 1326685 1328760 4.4 76492 +145 RB264 -1.784218682 64 195 Ribosomal protein S21 132677 132871 12.3 8074 +
1434 RB2647 -2.035828497 257 774 conserved hypothetical protein, secreted 1374889 1374116 4.9 27907 -1435 RB2649 -2.484632275 53 162 hypothetical protein 1374987 1375148 9.1 5813 +1453 RB2680 -1.725932617 467 1404 conserved hypothetical protein, membrane 1393566 1392163 6.4 49834 -1454 RB2682 -3.204746944 83 252 hypothetical protein 1393781 1393530 7.4 8960 -1519 RB2810 -5.365744728 49 150 hypothetical protein 1459901 1459752 11.3 5618 -1566 RB2900 -3.217833661 90 273 hypothetical protein 1506927 1507199 9.1 9939 +1606 RB2963 -1.983225367 119 360 hypothetical protein 1526707 1527066 12.2 13440 +1613 RB2975 -1.621747024 133 402 Nitrogen regulatory protein P-II glnB 1535384 1534983 8.8 14941 -
3 RB3 -1.505962523 73 222 hypothetical protein 39 260 8.9 8439 +1641 RB3023 -2.31685063 155 468 secreted protein 1566881 1566414 10 16924 -1734 RB3205 -1.630395388 76 231 hypothetical protein 1652079 1651849 11.3 9082 -1778 RB3281 -1.623405135 139 420 Histone-like bacterial DNA-binding protein 1687272 1686853 7.3 15802 -1835 RB3374 -2.36745013 95 288 hypothetical protein 1740350 1740063 11 11130 -1847 RB3399 -1.646194515 65 198 hypothetical protein 1753650 1753453 9.7 7418 -198 RB344 -1.667074676 700 2103 conserved hypothetical protein, secreted 173795 171693 5 77140 -

1967 RB3603 -1.519398413 344 1035 secreted protein 1875461 1874427 4.6 37999 -1984 RB3627 -1.69976521 280 843 Transglutaminase-like domain 1888482 1887640 4.5 30412 -2088 RB3789 -3.05899621 159 480 hypothetical protein 1962878 1962399 6.4 18218 -2098 RB3813 -1.586022583 75 228 hypothetical protein 1975188 1975415 4.5 8522 +2141 RB3882 -1.638277832 50 153 hypothetical protein 2007369 2007217 12.4 5918 -2154 RB3905 -1.769567638 38 117 hypothetical protein 2018601 2018717 5.1 4131 +2164 RB3923 -1.624470467 55 168 hypothetical protein 2032453 2032286 10.5 5956 -2168 RB3928 -1.546926335 173 522 hypothetical protein 2033789 2034310 10 19677 +2184 RB3953 -1.717863646 857 2574 hypothetical protein 2048784 2046211 5.2 93881 -2205 RB3989 -1.571221978 110 333 membrane protein 2060382 2060714 6.1 12440 +2210 RB3994 -1.62263894 191 576 hypothetical protein 2061291 2061866 4.1 20739 +234 RB402 -2.202205745 151 456 hypothetical protein 212820 213275 4.4 17306 +
2247 RB4071 -1.932729384 369 1110 integral membrane protein 2098441 2099550 6.7 40439 +2260 RB4097 -1.760503575 733 2202 conserved hypothetical protein 2108244 2110445 6.2 83188 +2298 RB4165 -1.637930288 721 2166 hypothetical protein 2139632 2141797 4.5 77994 +241 RB417 -2.048581122 314 945 nonspecific nucleoside hydrolase 221324 222268 4.8 33970 +
2304 RB4176 -3.424167784 96 291 hypothetical protein 2146928 2146638 12.4 11402 -242 RB418 -1.961141914 363 1092 conserved hypothetical protein, membrane 222222 223313 10.3 39119 +
2314 RB4196 -1.517887215 114 345 hypothetical protein 2157140 2157484 9 12738 +2350 RB4255 -1.647794393 204 615 hypothetical protein 2180806 2180192 10.3 21582 -2351 RB4256 -1.700309632 358 1077 conserved hypothetical protein, secreted 2180303 2181379 4.5 37091 +248 RB430 -1.514532205 522 1569 protein containing DUF1501 229809 231377 6 58047 +
2407 RB4347 -3.317265914 156 471 conserved hypothetical protein 2235735 2236205 4.7 17459 +2417 RB4360 -1.741651416 156 471 hypothetical protein 2240075 2239605 4.7 17592 -2437 RB4394 -3.867110768 188 567 protease I pfpI 3.2.-.- 2261372 2261938 4.4 20601 +2439 RB4396 -1.561295647 451 1356 glutathione reductase gor 1.8.1.7 2262532 2263887 5 48547 +2441 RB4398 -1.720721935 35 108 hypothetical protein 2264261 2264154 9 3825 -2443 RB4400 -3.078351438 58 177 hypothetical protein 2265382 2265206 9 6610 -2451 RB4416 -1.710492788 833 2502 Cytochrome c oxidase, subunit I 2271803 2269302 5.8 92293 -2461 RB4429 -3.079626574 89 270 conserved hypothetical protein 2275996 2276265 4.9 10034 +2462 RB4432 -2.017757176 268 807 glucose 1-dehydrogenase or 3-oxoacyl-[acyl-carrier-protein] reductase 1.1.1.- 2276333 2277139 4.7 28181 +2465 RB4438 -3.0073471 160 483 Pyridoxamine 5'-phosphate oxidase- 2278247 2278729 4.4 18043 +2472 RB4454 -1.784472046 739 2220 Flagellin A flaA 2283201 2285420 4 76924 +2477 RB4459 -1.720756801 462 1389 MscS Mechanosensitive ion channel 2288477 2287089 4.8 51157 -2481 RB4464 -3.330533712 223 672 secreted protein 2292075 2291404 5.1 25669 -2485 RB4468 -2.265372113 162 489 hypothetical protein 2294104 2293616 4.6 16380 -2486 RB4470 -3.659108285 169 510 secreted protein 2294676 2294167 4.9 19277 -2488 RB4474 -4.231918079 168 507 CsbD-like 2295048 2295554 8.8 18451 +2489 RB4476 -2.194667158 128 387 hypothetical protein 2295557 2295943 11.4 14037 +2490 RB4477 -2.465392744 69 210 hypothetical protein 2295940 2296149 10.6 7470 +2492 RB4480 -2.486728305 89 270 hypothetical protein 2296646 2296915 6.1 9442 +2498 RB4492 -1.813744209 101 306 hypothetical protein 2302138 2301833 9.9 10841 -2499 RB4493 -4.82841864 83 252 hypothetical protein 2302374 2302123 10.2 8978 -2500 RB4497 -2.781127188 49 150 hypothetical protein 2302950 2303099 10.3 5351 +2503 RB4500 -4.108372895 87 264 conserved hypothetical protein, secreted 2303645 2303908 3.5 9432 +2504 RB4502 -1.587856413 59 180 Protein of unknown function, UPF0057 2303992 2304171 9.1 6443 +2505 RB4504 -1.515612594 181 546 membrane protein containing DUF421 2304293 2304838 7.5 19859 +2524 RB4538 -1.598974592 172 519 secreted protein 2326219 2325701 5.4 18500 -2568 RB4613 -3.390322421 110 333 HesB/YadR/YfhF 2362510 2362178 4.6 12216 -2569 RB4614 -3.763789304 44 135 hypothetical protein 2362625 2362491 3.9 4907 -2584 RB4642 -2.739852866 85 258 conserved hypothetical protein 2377301 2377558 4.4 9855 +2607 RB4676 -1.862123889 221 666 hypothetical protein 2391079 2390414 10.4 23751 -2608 RB4678 -1.853751899 199 600 secreted protein 2390618 2391217 4.2 20927 +2657 RB4754 -2.122934062 58 177 hypothetical protein 2437589 2437765 6 6098 +280 RB479 -1.7608127 1101 3306 secreted protein 259174 255869 4.9 123358 -
2693 RB4809 -2.337028238 98 297 membrane protein 2464977 2465273 10.1 10285 +2726 RB4867 -2.169969784 210 633 SmpB RNA-binding protein SmpB 2496354 2495722 10.8 23762 -2733 RB4876 -3.631304332 397 1194 delta 9 acyl-lipid fatty acid desaturase 1.14.19.- 2506214 2505021 9.5 44741 -2747 RB4900 -1.59067028 166 501 hypothetical protein 2520067 2519567 11.4 18298 -2770 RB4931 -1.666717737 217 654 secreted protein 2537481 2538134 5.9 24246 +2773 RB4936 -1.738166802 269 810 O-acyl-[acyl-carrier-protein]-UDP-N-acetylglucosamine O-acyltransferase lpxA 2.3.1.129 2539280 2540089 8.7 29110 +2784 RB4958 -1.77450529 250 753 hypothetical protein 2549283 2548531 9.8 27813 -289 RB497 -1.56741782 368 1107 methanol dehydrogenase regulatory protein; (moxR) 269443 270549 5.4 40725 +
2816 RB5001 -1.64544186 103 312 hypothetical protein 2572354 2572665 10.3 11694 +2850 RB5064 -1.7367901 648 1947 secreted protein containing DUF1585 2605707 2607653 4.6 72145 +2875 RB5103 -1.67343724 352 1059 membrane protein 2622260 2623318 9.1 39152 +296 RB514 -2.436714418 56 171 hypothetical protein 284567 284397 10.2 6284 -
2927 RB5196 -1.84638216 701 2106 trehalose synthase treS 5.4.99.16 2678058 2675953 5.6 81037 -300 RB520 -2.441328044 375 1128 conserved hypothetical protein 284989 286116 4.8 41300 +
2963 RB5261 -1.777215676 126 381 hypothetical protein 2713712 2713332 7.4 13761 -2964 RB5262 -1.987732634 95 288 membrane protein 2713742 2714029 6.3 10993 +2978 RB5282 -1.563469412 578 1737 choline-sulfatase betC 3.1.6.6 2727443 2725707 5 65084 -3024 RB5373 -1.610866446 44 135 hypothetical protein 2775404 2775270 10.4 5046 -3100 RB5499 -1.718253529 447 1344 conserved hypothetical protein c 2844998 2843655 7.1 48835 -3107 RB5506 -1.763298362 51 156 hypothetical protein 2851102 2850947 10.3 6122 -3130 RB5551 -1.726697769 663 1992 hypothetical protein 2889101 2891092 5.7 70932 +3139 RB5567 -2.807682104 66 201 hypothetical protein 2897766 2897566 12.1 7106 -3162 RB5613 -1.718686676 360 1083 protein containing DUF1006 2921546 2922628 9.9 42148 +327 RB566 -2.408254421 294 885 exopolysaccharide synthesis, ExoD family 312007 311123 10 32340 -328 RB568 -1.54389448 77 234 hypothetical protein 312345 312112 10.2 8689 -
3202 RB5681 -1.899201568 587 1764 Trigger factor tig 2965080 2966843 4.5 66546 +3219 RB5715 -1.852875395 210 633 hypothetical protein 2982826 2983458 4.9 24368 +3247 RB5761 -1.678461629 258 777 conserved hypothetical protein, membrane 3015202 3014426 10.9 27944 -3270 RB5801 -1.514537677 89 270 Ribosomal protein S15 3039583 3039852 10.6 10154 +3272 RB5804 -1.667295501 818 2457 polynucleotide phosphorylase 3040160 3042616 5.3 88202 +3294 RB5848 -4.212733889 140 423 dnaK suppressor protein, 3066328 3065906 4.6 15633 -3295 RB5850 -2.107814411 63 192 hypothetical protein 3066498 3066307 5.8 7096 -3316 RB5882 -2.135767975 243 732 hypothetical protein 3084330 3083599 9.8 27773 -3339 RB5924 -1.591642193 847 2544 hypothetical protein 3105828 3103285 5.2 92073 -3355 RB5955 -2.57021459 177 534 hypothetical protein 3121488 3122021 4.5 19364 +3389 RB6010 -1.641562068 53 162 hypothetical protein 3149911 3150072 11.9 5744 +3421 RB6071 -1.658689447 213 642 hypothetical protein 3187463 3188104 5.1 23869 +3465 RB6156 -1.525242339 134 405 membrane protein 3233026 3232622 9.5 14531 -3478 RB6178 -1.844910512 41 126 hypothetical protein 3240162 3240287 12.7 4414 +3547 RB6314 -2.653088765 52 159 hypothetical protein 3319320 3319478 6.3 6180 +3571 RB6351 -1.737859307 58 177 hypothetical protein 3336171 3336347 4.2 6411 +3587 RB6378 -1.986811448 228 687 hypothetical protein 3352598 3353284 12.3 25698 +3588 RB6379 -3.063507449 153 462 hypothetical protein 3353122 3352661 5.8 17398 -354 RB638 -1.810321183 436 1311 PQQ enzyme repeat domain protein 346447 347757 4.5 47685 +
3605 RB6403 -3.371448655 304 915 hypothetical protein 3369939 3370853 5.4 33688 +3615 RB6417 -2.965326241 249 750 conserved hypothetical protein 3378045 3378794 5.9 27679 +3620 RB6428 -1.530633865 164 495 conserved hypothetical protein, secreted 3382816 3382322 6.5 17292 -3621 RB6429 -2.59203864 61 186 hypothetical protein 3382998 3382813 12.1 7095 -3644 RB6463 -1.646018383 189 570 conserved hypothetical protein, membrane 3415155 3415724 10.2 19818 +3674 RB6504 -1.754818069 53 162 hypothetical protein 3456120 3455959 9.6 5968 -3716 RB6578 -1.495239464 311 936 secreted protein 3495957 3495022 4.7 34703 -359 RB658 -1.69573478 412 1239 transport protein 356228 354990 5.2 44434 -
3718 RB6583 -1.91791944 996 2991 secreted protein 3498012 3501002 4.5 112232 +3724 RB6595 -1.552798644 234 705 Rickettsia 17 kDa surface antigen 3506173 3505469 10.1 24227 -3768 RB6681 -1.549810836 139 420 hypothetical protein 3551775 3552194 4.9 15804 +3781 RB6701 -1.632497838 87 264 hypothetical protein 3559944 3560207 12 9863 +3795 RB6724 -1.861775884 131 396 hypothetical protein 3569189 3569584 10.5 14916 +3797 RB6728 -1.553174354 113 342 hypothetical protein 3570581 3570922 9.8 12410 +3820 RB6766 -1.498615661 55 168 hypothetical protein 3597367 3597534 12 6133 +3827 RB6778 -2.494952322 59 180 hypothetical protein 3603142 3603321 5.5 6533 +374 RB684 -1.537572032 653 1962 Arylsulfatase precursor atsA 3.1.6.1 368557 366596 5.5 73598 -
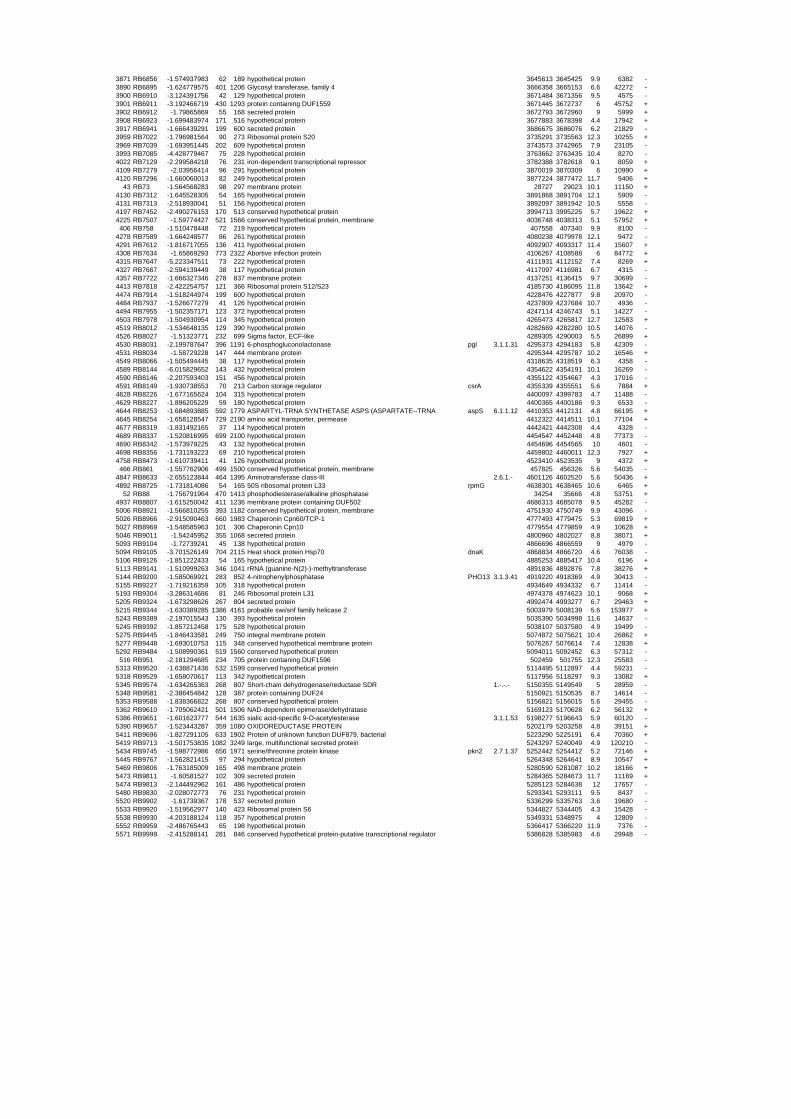
3871 RB6856 -1.574937983 62 189 hypothetical protein 3645613 3645425 9.9 6382 -3890 RB6895 -1.624779575 401 1206 Glycosyl transferase, family 4 3666358 3665153 6.6 42272 -3900 RB6910 -3.124391756 42 129 hypothetical protein 3671484 3671356 9.5 4575 -3901 RB6911 -3.192466719 430 1293 protein containing DUF1559 3671445 3672737 6 45752 +3902 RB6912 -1.79865869 55 168 secreted protein 3672793 3672960 9 5999 +3908 RB6923 -1.699483974 171 516 hypothetical protein 3677883 3678398 4.4 17942 +3917 RB6941 -1.666439291 199 600 secreted protein 3686675 3686076 6.2 21829 -3959 RB7022 -1.796981564 90 273 Ribosomal protein S20 3735291 3735563 12.3 10255 +3969 RB7039 -1.693951445 202 609 hypothetical protein 3743573 3742965 7.9 23105 -3993 RB7085 -4.428779467 75 228 hypothetical protein 3763662 3763435 10.4 8270 -4022 RB7129 -2.299584218 76 231 iron-dependent transcriptional repressor 3782388 3782618 9.1 8059 +4109 RB7279 -2.03956414 96 291 hypothetical protein 3870019 3870309 6 10990 +4120 RB7296 -1.660060013 82 249 hypothetical protein 3877224 3877472 11.7 9406 +
43 RB73 -1.564568283 98 297 membrane protein 28727 29023 10.1 11150 +4130 RB7312 -1.645528305 54 165 hypothetical protein 3891868 3891704 12.1 5909 -4131 RB7313 -2.518930041 51 156 hypothetical protein 3892097 3891942 10.5 5558 -4197 RB7452 -2.490276153 170 513 conserved hypothetical protein 3994713 3995225 5.7 19622 +4225 RB7507 -1.59774427 521 1566 conserved hypothetical protein, membrane 4036748 4038313 5.1 57952 +406 RB758 -1.510478448 72 219 hypothetical protein 407558 407340 9.9 8100 -
4278 RB7589 -1.664248577 86 261 hypothetical protein 4080238 4079978 12.1 9472 -4291 RB7612 -1.816717055 136 411 hypothetical protein 4092907 4093317 11.4 15607 +4308 RB7634 -1.65869293 773 2322 Abortive infection protein 4106267 4108588 6 84772 +4315 RB7647 -5.223347511 73 222 hypothetical protein 4111931 4112152 7.4 8269 +4327 RB7667 -2.594139449 38 117 hypothetical protein 4117097 4116981 6.7 4315 -4357 RB7722 -1.666327346 278 837 membrane protein 4137251 4136415 9.7 30699 -4413 RB7818 -2.422254757 121 366 Ribosomal protein S12/S23 4185730 4186095 11.8 13642 +4474 RB7914 -1.518244974 199 600 hypothetical protein 4228476 4227877 9.8 20970 -4484 RB7937 -1.526677279 41 126 hypothetical protein 4237809 4237684 10.7 4936 -4494 RB7955 -1.502357171 123 372 hypothetical protein 4247114 4246743 5.1 14227 -4503 RB7978 -1.504930954 114 345 hypothetical protein 4265473 4265817 12.7 12583 +4519 RB8012 -1.534648135 129 390 hypothetical protein 4282669 4282280 10.5 14076 -4526 RB8027 -1.51323771 232 699 Sigma factor, ECF-like 4289305 4290003 5.5 26899 +4530 RB8031 -2.199787647 396 1191 6-phosphogluconolactonase pgl 3.1.1.31 4295373 4294183 5.8 42309 -4531 RB8034 -1.58729228 147 444 membrane protein 4295344 4295787 10.2 16546 +4549 RB8066 -1.505494445 38 117 hypothetical protein 4318635 4318519 6.3 4358 -4589 RB8144 -6.015829652 143 432 hypothetical protein 4354622 4354191 10.1 16269 -4590 RB8146 -2.207593403 151 456 hypothetical protein 4355122 4354667 4.3 17016 -4591 RB8149 -1.930738553 70 213 Carbon storage regulator csrA 4355339 4355551 5.6 7884 +4628 RB8226 -1.677165624 104 315 hypothetical protein 4400097 4399783 4.7 11488 -4629 RB8227 -1.896205229 59 180 hypothetical protein 4400365 4400186 9.3 6533 -4644 RB8253 -1.684893885 592 1779 ASPARTYL-TRNA SYNTHETASE ASPS (ASPARTATE--TRNA aspS 6.1.1.12 4410353 4412131 4.8 66195 +4645 RB8254 -1.658128547 729 2190 amino acid transporter, permease 4412322 4414511 10.1 77104 +4677 RB8319 -1.831492165 37 114 hypothetical protein 4442421 4442308 4.4 4328 -4689 RB8337 -1.520816995 699 2100 hypothetical protein 4454547 4452448 4.8 77373 -4690 RB8342 -1.573979225 43 132 hypothetical protein 4454696 4454565 10 4601 -4698 RB8356 -1.731193223 69 210 hypothetical protein 4459802 4460011 12.3 7927 +4758 RB8473 -1.610739411 41 126 hypothetical protein 4523410 4523535 9 4372 +466 RB861 -1.557762906 499 1500 conserved hypothetical protein, membrane 457825 456326 5.6 54035 -
4847 RB8633 -2.655123844 464 1395 Aminotransferase class-III 2.6.1.- 4601126 4602520 5.6 50436 +4892 RB8725 -1.731814086 54 165 50S ribosomal protein L33 rpmG 4638301 4638465 10.6 6465 +
52 RB88 -1.756791964 470 1413 phosphodiesterase/alkaline phosphatase 34254 35666 4.8 53751 +4937 RB8807 -1.615250042 411 1236 membrane protein containing DUF502 4686313 4685078 9.5 45282 -5006 RB8921 -1.566810255 393 1182 conserved hypothetical protein, membrane 4751930 4750749 9.9 43096 -5026 RB8966 -2.915090463 660 1983 Chaperonin Cpn60/TCP-1 4777493 4779475 5.3 69819 +5027 RB8969 -1.548585963 101 306 Chaperonin Cpn10 4779554 4779859 4.9 10628 +5046 RB9011 -1.54245952 355 1068 secreted protein 4800960 4802027 8.8 38071 +5093 RB9104 -1.72739241 45 138 hypothetical protein 4866696 4866559 9 4979 -5094 RB9105 -3.701526149 704 2115 Heat shock protein Hsp70 dnaK 4868834 4866720 4.6 76038 -5106 RB9126 -1.851222433 54 165 hypothetical protein 4885253 4885417 10.4 6196 +5113 RB9141 -1.510999263 346 1041 rRNA (guanine-N(2)-)-methyltransferase 4891836 4892876 7.8 38276 +5144 RB9200 -1.585069921 283 852 4-nitrophenylphosphatase PHO13 3.1.3.41 4919220 4918369 4.9 30413 -5155 RB9227 -1.719216359 105 318 hypothetical protein 4934649 4934332 6.7 11414 -5193 RB9304 -3.286314686 81 246 Ribosomal protein L31 4974378 4974623 10.1 9068 +5205 RB9324 -1.673298626 267 804 secreted protein 4992474 4993277 6.7 29463 +5215 RB9344 -1.630389285 1386 4161 probable swi/snf family helicase 2 5003979 5008139 5.6 153977 +5243 RB9389 -2.197015543 130 393 hypothetical protein 5035390 5034998 11.6 14637 -5245 RB9392 -1.857212458 175 528 hypothetical protein 5038107 5037580 4.9 19499 -5275 RB9445 -1.846433581 249 750 integral membrane protein 5074872 5075621 10.4 26862 +5277 RB9448 -1.693010753 115 348 conserved hypothetical membrane protein 5076267 5076614 7.4 12838 +5292 RB9484 -1.508990361 519 1560 conserved hypothetical protein 5094011 5092452 6.3 57312 -516 RB951 -2.181294685 234 705 protein containing DUF1596 502459 501755 12.3 25583 -
5313 RB9520 -1.638871436 532 1599 conserved hypothetical protein 5114495 5112897 4.4 59231 -5318 RB9529 -1.658070617 113 342 hypothetical protein 5117956 5118297 9.3 13082 +5345 RB9574 -1.634265363 268 807 Short-chain dehydrogenase/reductase SDR 1.-.-.- 5150355 5149549 5 28959 -5348 RB9581 -2.386454842 128 387 protein containing DUF24 5150921 5150535 8.7 14614 -5353 RB9588 -1.838366822 268 807 conserved hypothetical protein 5156821 5156015 5.6 29455 -5362 RB9610 -1.705062421 501 1506 NAD-dependent epimerase/dehydratase 5169123 5170628 6.2 56132 +5386 RB9651 -1.601623777 544 1635 sialic acid-specific 9-O-acetylesterase 3.1.1.53 5198277 5196643 5.9 60120 -5390 RB9657 -1.523443287 359 1080 OXIDOREDUCTASE PROTEIN 5202179 5203258 4.8 39151 +5411 RB9696 -1.827291105 633 1902 Protein of unknown function DUF879, bacterial 5223290 5225191 6.4 70360 +5419 RB9713 -1.501753835 1082 3249 large, multifunctional secreted protein 5243297 5240049 4.9 120210 -5434 RB9745 -1.598772986 656 1971 serine/threonine protein kinase pkn2 2.7.1.37 5252442 5254412 5.2 72146 +5445 RB9767 -1.562821415 97 294 hypothetical protein 5264348 5264641 8.9 10547 +5469 RB9806 -1.763185009 165 498 membrane protein 5280590 5281087 10.2 18166 +5473 RB9811 -1.60581527 102 309 secreted protein 5284365 5284673 11.7 11169 +5474 RB9813 -2.144492962 161 486 hypothetical protein 5285123 5284638 12 17657 -5480 RB9830 -2.028072773 76 231 hypothetical protein 5293341 5293111 9.5 8437 -5520 RB9902 -1.61739367 178 537 secreted protein 5336299 5335763 3.6 19680 -5533 RB9920 -1.519562977 140 423 Ribosomal protein S6 5344827 5344405 4.3 15428 -5538 RB9930 -4.203188124 118 357 hypothetical protein 5349331 5348975 4 12809 -5552 RB9959 -2.486765443 65 198 hypothetical protein 5366417 5366220 11.9 7376 -5571 RB9999 -2.415288141 281 846 conserved hypothetical protein-putative transcriptional regulator 5386828 5385983 4.6 29948 -

240h vs 82h upID Locus Ratio AA Nuc Product Gene EC Start Stop IEP MW(kDa) Strand
58 RB100 1.654514659 123 372 membrane protein 41097 40726 7.5 13930 -5578 RB10010 4.373631074 840 2523 secreted protein containing DUF1585 5393982 5391460 4.9 95249 -5579 RB10014 2.042872232 534 1605 protein containing DUF1552 5395577 5393973 7.2 58534 -5582 RB10020 2.189639697 64 195 hypothetical protein 5396788 5396594 9.1 7272 -5584 RB10022 1.605837782 165 498 membrane protein 5396916 5397413 6.3 17893 +5588 RB10026 2.159526021 883 2652 conserved hypothetical protein 5401902 5399251 6.5 98323 -5608 RB10053 1.579968872 496 1491 secreted protein containing DUF1501 5423854 5425344 6.6 54820 +5625 RB10096 2.67333667 341 1026 Transposase IS116/IS110/IS902 5446891 5445866 10 38283 -5627 RB10098 1.885841688 47 144 hypothetical protein 5447310 5447453 9.7 5129 +5631 RB10105 1.531582074 115 348 protein containing DUF1582 5451202 5450855 8.9 12499 -5633 RB10107 1.628209869 50 153 hypothetical protein 5451876 5451724 9.1 5550 -5642 RB10119 1.627089735 766 2301 serine/threonine protein kinase pknD 2.7.1.37 5462723 5460423 5.3 82693 -5647 RB10126 4.215826536 43 132 hypothetical protein 5465210 5465341 12.9 4964 +5670 RB10167 1.676202415 59 180 hypothetical protein 5497096 5496917 9 6356 -5673 RB10172 1.509922517 489 1470 aldehyde dehydrogenase family protein (Betaine) 1.2.1.36 5499799 5498330 5 53366 -5711 RB10238 1.71830836 577 1734 protein containing DUF1559 5531834 5533567 5.8 62799 +5715 RB10245 2.200657651 328 987 haloalkane dehalogenase dhmA2 3.8.1.5 5537951 5538937 7.7 36310 +5716 RB10246 2.57631347 316 951 Triphosphoribosyl-dephospho-CoA protein (ORF7) citG 5538891 5539841 5.1 34552 +5718 RB10252 2.292741304 386 1161 hypothetical protein 5541386 5542546 4.3 42018 +5719 RB10255 3.701967215 337 1014 MscS Mechanosensitive ion channel 5542656 5543669 5.4 35647 +5720 RB10256 3.225664198 268 807 hypothetical protein 5543753 5544559 4.1 30801 +5721 RB10258 3.223167304 289 870 Survival protein SurE 3.1.3.2 5544584 5545453 5.3 31584 +5732 RB10277 2.90124719 476 1431 Pyruvate kinase pykF 2.7.1.40 5555414 5556844 5.2 52151 +5737 RB10290 2.928019934 142 429 hypothetical protein 5559631 5559203 12.4 16122 -5739 RB10295 1.699754772 308 927 methylene tetrahydrofolate/ mtdC 5562427 5561501 5.2 32122 -5745 RB10308 1.548630277 308 927 response regulator 5569131 5570057 4.9 34171 +5750 RB10319 3.610640028 65 198 hypothetical protein 5577574 5577771 9.6 7480 +5756 RB10328 2.940635557 227 684 protein containing DUF633 5583675 5582992 9.5 25921 -5762 RB10338 2.43243988 486 1461 TPR domain protein 5591839 5590379 5 53945 -5770 RB10347 1.690703459 184 555 hypothetical protein 5596258 5595704 5.5 20536 -5804 RB10419 1.500890259 40 123 hypothetical protein 5639506 5639384 12.1 4388 -5807 RB10427 1.50099123 47 144 hypothetical protein 5647025 5646882 12.1 5635 -5811 RB10433 1.722495307 441 1326 membrane protein 5651746 5650421 9.9 48642 -5880 RB10569 1.581605225 388 1167 protein containing DUF1559 5736174 5735008 5.4 41686 -5902 RB10608 2.057744599 457 1374 transposase and inactivated derivative 5759903 5761276 8.4 52492 +5903 RB10612 1.886271614 492 1479 choline-sulfatase betC 3.1.6.6 5761454 5762932 5.8 54603 +5942 RB10682 1.654794846 474 1425 secreted protein 5807422 5805998 12.2 53138 -5947 RB10689 1.616678827 85 258 hypothetical protein 5808207 5807950 12.2 9609 -5953 RB10698 1.843186951 150 453 hypothetical protein 5813025 5812573 5.8 16611 -5974 RB10735 1.978886422 332 999 hypothetical protein 5834722 5835720 5.3 37343 +5985 RB10752 1.904220821 42 129 hypothetical protein 5838740 5838612 10.7 4817 -5992 RB10764 1.988421087 195 588 hypothetical protein 5845271 5844684 8.6 22760 -6018 RB10816 1.863339078 376 1131 DNA polymerase III tau and gamma subunits 5875071 5873941 8.6 40800 -6040 RB10855 1.989745932 395 1188 mandelate racemase/muconate lactonizing enzyme family protein 5895378 5894191 6.4 43363 -6046 RB10866 3.289082377 82 249 hypothetical protein 5899291 5899043 10.2 9214 -6048 RB10868 2.008192745 136 411 hypothetical protein 5900330 5899920 11.3 15100 -6052 RB10876 1.853661907 839 2520 excinuclease ABC subunit A 5903770 5901251 6.4 90046 -6064 RB10893 2.84725917 57 174 hypothetical protein 5914894 5915067 9.7 6059 +580 RB1095 1.805808757 525 1578 hypothetical protein 567469 569046 4.5 57051 +
6101 RB10960 1.540150809 139 420 conserved hypothetical protein, membrane 5941602 5942021 11.4 15743 +6102 RB10965 1.700480847 204 615 conserved hypothetical protein, membrane 5942116 5942730 4.9 21483 +581 RB1097 1.579125526 404 1215 membrane protein 570281 569067 5.3 42931 -
6109 RB10978 1.591971636 122 369 Small multidrug resistance protein 5947515 5947883 4.6 12506 +6125 RB11001 3.10014494 294 885 conserved hypothetical protein 5964931 5964047 9.6 33860 -6128 RB11009 3.753956045 238 717 sulfite reductase-like flavoprotein 5969752 5969036 3.9 25651 -6129 RB11010 2.909442479 278 837 conserved hypothetical protein, secreted 5970585 5969749 4.9 30852 -6130 RB11011 2.956454785 181 546 conserved hypothetical protein, secreted 5971127 5970582 9.5 20389 -6131 RB11013 2.913287746 218 657 conserved hypothetical protein, membrane 5971802 5971146 4.9 24214 -6136 RB11023 1.809482834 59 180 hypothetical protein 5975603 5975424 3.8 6205 -6140 RB11031 2.682575659 62 189 hypothetical protein 5980865 5981053 12.1 7649 +6142 RB11036 1.911367719 470 1413 two-component system, regulatory protein 5985609 5984197 5.3 51036 -6143 RB11037 1.859310972 493 1482 two-component system sensor protein 5987098 5985617 5.8 55133 -6152 RB11062 1.687148515 214 645 conserved hypothetical protein 5995358 5994714 4.7 23983 -6157 RB11070 2.836648643 52 159 hypothetical protein 6000321 6000479 10.2 5582 +6165 RB11082 2.919691689 112 339 hypothetical protein 6007371 6007033 10.9 12552 -6206 RB11156 1.841373417 350 1053 secreted protein 6056074 6055022 5.8 38283 -6212 RB11168 2.882446961 149 450 thiosulfate sulfurtransferase 6062177 6061728 9.8 17041 -591 RB1120 1.607182316 651 1956 multidrug efflux protein, outer membrane component 576988 575033 4.7 72081 -
6248 RB11228 2.001536123 42 129 hypothetical protein 6087599 6087471 13 5329 -6251 RB11233 2.219312927 295 888 hypothetical protein 6090340 6089453 4.7 33694 -6269 RB11268 1.86099064 97 294 hypothetical protein 6109768 6109475 10.8 11235 -6285 RB11293 1.562813644 97 294 hypothetical protein 6121182 6120889 10.3 11032 -6292 RB11303 1.816331576 464 1395 Transposase IS66 6128599 6127205 10.2 51739 -6321 RB11348 2.306868407 151 456 conserved hypothetical protein 6155692 6155237 3.4 16759 -6324 RB11356 1.621754315 237 714 glycosyltransferase, family 2 6158199 6157486 7.1 27585 -599 RB1138 2.497575401 155 468 conserved hypothetical protein 585492 585959 10.5 17130 +
6362 RB11424 3.968781865 158 477 conserved hypothetical protein 6195656 6196132 11.1 17372 +6363 RB11427 2.669809078 93 282 conserved hypothetical protein 6196136 6196417 9.7 10315 +6364 RB11429 4.836258644 37 114 conserved hypothetical protein 6196478 6196591 12.4 4245 +6371 RB11439 4.771980115 93 282 conserved hypothetical protein 6197993 6198274 10.6 10906 +6382 RB11446 4.373489974 113 342 conserved hypothetical protein 6199492 6199833 10 12579 +6385 RB11450 3.292516777 60 183 conserved hypothetical protein 6200046 6200228 11.8 6517 +6399 RB11462 2.292414397 129 390 CoA-binding protein 6202949 6203338 5.1 13908 +6401 RB11468 1.600332302 239 720 conserved hypothetical protein, secreted 6205637 6204918 4.9 26639 -6411 RB11484 1.731982898 43 132 conserved hypothetical protein 6212742 6212611 5.5 4566 -6479 RB11616 2.050443667 56 171 conserved hypothetical protein 6262622 6262452 5.5 6002 -6480 RB11617 2.653050908 297 894 conserved hypothetical protein 6263689 6262796 5.5 33558 -6497 RB11641 2.148369724 297 894 conserved hypothetical protein 6278514 6277621 5.5 33558 -6508 RB11663 1.781211968 450 1353 sugar transport protein 6290653 6292005 9.6 48415 +6521 RB11677 2.207148141 47 144 conserved hypothetical protein 6300221 6300078 10.9 5108 -6524 RB11680 1.902883532 59 180 conserved hypothetical protein 6300635 6300456 10.1 6549 -6528 RB11685 1.624754627 59 180 conserved hypothetical protein 6301256 6301077 10.2 6734 -
66 RB117 1.845310737 129 390 membrane or secreted protein 51525 51136 11 14217 -6544 RB11720 1.526186688 264 795 conserved hypothetical protein 6316234 6317028 10.7 27475 +6545 RB11722 1.764001072 342 1029 conserved hypothetical protein 6317037 6318065 6.7 37460 +6547 RB11728 1.859522593 295 888 D-Tagatose 3-epimerase 5.3.1.- 6318857 6319744 4.6 32417 +6548 RB11729 2.095879613 85 258 conserved hypothetical protein 6320009 6319752 10.6 9507 -6559 RB11750 2.072392265 292 879 integrase 6328357 6329235 10 33354 +6571 RB11768 2.488324614 43 132 conserved hypothetical protein 6334808 6334939 9.4 4603 +6572 RB11769 1.550502675 8173 24522 domain/calx-beta domain protein 6335006 6359527 3 826151 +622 RB1179 1.836873669 128 387 hypothetical protein 603998 604384 12 13863 +623 RB1180 1.682087161 92 279 hypothetical protein 604378 604656 12.6 9917 +
6589 RB11823 1.936606455 634 1905 conserved hypothetical protein, secreted 6371378 6373282 5 68778 +6590 RB11826 1.664574845 100 303 conserved hypothetical protein 6373298 6373600 12.3 11008 +626 RB1184 1.629663004 98 297 protein containing DUF1556 605971 605675 11.4 10926 -
6598 RB11844 1.69066366 94 285 conserved hypothetical protein 6378510 6378794 6.8 10448 +6605 RB11854 1.570100857 77 234 conserved hypothetical protein 6383404 6383171 13.5 8576 -

6606 RB11855 2.19926923 101 306 conserved hypothetical protein 6383765 6383460 12.5 11384 -6610 RB11862 1.6336847 56 171 conserved hypothetical protein, secreted 6387327 6387157 12 6073 -629 RB1190 1.531303399 292 879 integrase 607131 608009 10 33354 +630 RB1191 2.621249142 126 381 hypothetical transposase 608435 608055 9.6 13925 -
6660 RB11951 3.121435039 195 588 conserved hypothetical protein 6437923 6438510 6.1 21768 +6670 RB11973 1.521276023 273 822 conserved hypothetical protein, membrane 6447543 6446722 10.1 28672 -6689 RB12006 1.721932414 341 1026 regulatory subunit 6475429 6476454 4.4 36562 +6691 RB12009 1.689190216 514 1545 conserved hypothetical protein 6479495 6477951 4.8 54935 -638 RB1201 2.499019821 41 126 hypothetical protein 612754 612629 12.5 4691 -
6694 RB12015 1.614312795 321 966 conserved hypothetical protein 6483388 6482423 5.9 34513 -6701 RB12031 1.954623073 146 441 conserved hypothetical protein 6490799 6490359 9 16668 -6711 RB12053 1.729367164 119 360 biopolymer transport protein, ExbD/TolR family 6502558 6502199 4.2 12926 -6712 RB12055 2.850799464 319 960 MotA/TolQ/ExbB proton channel family protein 6503587 6502628 5.3 34206 -6720 RB12068 2.037532136 553 1662 serine/threonine protein kinase 6512255 6510594 8.5 61271 -6727 RB12080 1.62139257 492 1479 Protein-L-isoaspartate(D-aspartate) O-methyltransferase 2.1.1.77 6517231 6515753 8.9 53385 -644 RB1210 2.291039759 281 846 AP endonuclease 2, C-terminal 617416 616571 4.5 30665 -
6745 RB12116 1.528505705 355 1068 DNA polymerase III epsilon subunit 6541348 6540281 9.2 38668 -6786 RB12200 1.907346776 51 156 conserved hypothetical protein 6577955 6577800 11 5747 -6800 RB12219 1.829206519 472 1419 conserved hypothetical protein 6586411 6584993 6.4 53798 -652 RB1223 2.353571948 73 222 hypothetical protein 624091 624312 10.4 7925 +
6812 RB12239 2.627303086 433 1302 ISXo8 transposase 6593334 6594635 9.4 49646 +6815 RB12243 2.005776569 55 168 conserved hypothetical protein 6596256 6596423 10.5 5848 +654 RB1225 1.555906229 834 2505 dipeptidyl peptidase IV DPP4 3.4.14.5 627017 624513 4.8 93242 -
6825 RB12265 1.506648506 42 129 conserved hypothetical protein 6602530 6602402 10.4 4412 -6833 RB12279 1.845574656 337 1014 MscS Mechanosensitive ion channel 6606558 6605545 4.5 36901 -6838 RB12286 1.697767005 57 174 conserved hypothetical protein 6610293 6610120 10.9 6473 -6844 RB12295 1.549620958 86 261 conserved hypothetical protein 6614190 6614450 9.1 9350 +6890 RB12377 4.034629067 284 855 Ribosomal RNA adenine methylase transferase 6661963 6661109 5.7 31560 -6891 RB12379 3.873999592 240 723 riboflavin synthase, alpha subunit 6662808 6662086 4.6 25728 -6894 RB12383 2.261031645 88 267 conserved hypothetical protein 6664767 6664501 3.9 9433 -6896 RB12386 1.963666988 61 186 conserved hypothetical protein, secreted 6665667 6665852 11.2 6546 +6897 RB12387 2.351753993 50 153 conserved hypothetical protein 6665894 6665742 10.1 5841 -6901 RB12392 2.308597949 35 108 conserved hypothetical protein 6668543 6668436 10.5 3832 -6902 RB12393 2.22524548 668 2007 c-type cytochrome precursor 6670561 6668555 5.9 74121 -6905 RB12398 1.713211441 361 1086 conserved hypothetical protein 6672013 6670928 10.1 39805 -663 RB1240 1.521961995 504 1515 protein containing DUF1080 634060 632546 9.4 55473 -
6907 RB12403 2.082915079 59 180 conserved hypothetical protein 6673741 6673562 7.3 6301 -6908 RB12404 2.145144818 97 294 conserved hypothetical protein 6674021 6673728 12.2 10758 -6915 RB12416 2.659967583 292 879 secreted protein 6679578 6680456 6 31935 +6921 RB12427 2.178493515 56 171 conserved hypothetical protein 6684645 6684475 12.8 6556 -6930 RB12439 1.835694179 44 135 protein containing DUF1586 6695571 6695437 10.7 4885 -6933 RB12442 1.766365352 511 1536 conserved hypothetical protein 6698977 6697442 4.4 55156 -6954 RB12479 2.301385379 190 573 conserved hypothetical protein 6717557 6716985 5.8 21665 -6957 RB12484 1.921747867 324 975 conserved hypothetical protein, membrane 6721052 6720078 10.6 34787 -668 RB1254 1.631322418 45 138 hypothetical protein 640610 640747 10.6 4883 +
6989 RB12543 1.623659414 331 996 conserved hypothetical protein, membrane 6759514 6758519 9.6 37564 -669 RB1255 1.967180069 51 156 hypothetical protein 640910 640755 12.3 5933 -
6994 RB12554 2.448543373 71 216 conserved hypothetical protein 6764163 6764378 10.2 7146 +7045 RB12648 1.928441091 69 210 conserved hypothetical protein 6810287 6810496 11.2 7886 +677 RB1266 1.583975133 99 300 hypothetical protein 648804 648505 11.8 11071 -
7066 RB12684 1.536286714 54 165 conserved hypothetical protein 6831038 6830874 5.4 5628 -7069 RB12689 2.115580227 106 321 secreted protein 6833004 6833324 9.8 11239 +678 RB1269 1.855473031 203 612 hypothetical protein 649412 648801 11.9 22612 -
7070 RB12690 1.932274176 250 753 3-deoxy-manno-octulosonate cytidylyltransferase kdsB 2.7.7.38 6834088 6833336 4.7 27384 -7074 RB12694 1.792575761 343 1032 serine/threonine-protein kinase pknB 2.7.1.37 6837178 6836147 9.6 37011 -7081 RB12710 1.734953496 36 111 conserved hypothetical protein 6846593 6846703 9.2 4110 +7084 RB12716 1.68107695 114 345 conserved hypothetical protein 6849459 6849115 12.4 13128 -7088 RB12727 2.18182508 680 2043 conserved hypothetical protein 6860301 6858259 5.7 73171 -7092 RB12737 2.385652769 65 198 conserved hypothetical protein 6863917 6864114 9.1 6753 +7093 RB12739 2.910582075 56 171 conserved hypothetical protein 6864276 6864106 10.8 6057 -7119 RB12782 3.953832549 55 168 conserved hypothetical protein 6889986 6889819 10.9 5879 -7132 RB12806 1.69219261 205 618 conserved hypothetical protein 6911165 6910548 10.5 21914 -7134 RB12810 2.10152585 67 204 conserved hypothetical protein 6912339 6912542 11.3 7443 +7139 RB12815 2.403965196 43 132 conserved hypothetical protein 6914656 6914525 10.8 4782 -7140 RB12816 1.596657606 257 774 glutamine amidotransferase or related peptidase 6914710 6915483 4.5 28223 +7141 RB12818 4.913335797 163 492 conserved hypothetical protein 6915989 6915498 11.7 18010 -7147 RB12827 2.06699605 122 369 conserved hypothetical protein, membrane 6919729 6919361 12.5 13367 -7148 RB12829 2.28308508 159 480 conserved hypothetical protein 6919813 6920292 9.5 17677 +7152 RB12836 2.071792125 38 117 conserved hypothetical protein 6921619 6921735 10.8 4178 +7161 RB12851 1.900824009 109 330 conserved hypothetical protein 6926556 6926885 7.5 12033 +7167 RB12860 1.592005277 411 1236 Oligopeptide transport system permease protein oppC oppC 6932003 6930768 9.3 45876 -7170 RB12863 2.070343076 50 153 conserved hypothetical protein 6934850 6935002 12.6 5550 +7184 RB12891 1.69510928 258 777 LmbE-like protein 6944411 6945187 4.8 28480 +7186 RB12895 2.394774712 153 462 conserved hypothetical protein 6945636 6946097 10.7 17703 +7211 RB12959 1.765394992 107 324 conserved hypothetical protein 6974205 6973882 11.4 12121 -7221 RB12978 1.519476268 221 666 conserved hypothetical protein 6980722 6980057 4.9 23400 -7243 RB13025 2.580278072 193 582 monooxygenase 6997019 6996438 9.7 21953 -7251 RB13038 2.214420043 753 2262 conserved hypothetical protein, membrane 7005061 7002800 6.9 84132 -7273 RB13088 1.93093716 210 633 conserved hypothetical protein 7022309 7021677 5.6 24052 -7307 RB13139 2.205283961 416 1251 xylose operon regulatory protein xylR 7049875 7048625 9.5 46427 -7324 RB13172 1.940192944 78 237 conserved hypothetical protein 7070198 7069962 10.2 8753 -7330 RB13184 1.558333715 207 624 protein disulphide isomerase 7079314 7078691 6.2 23072 -7338 RB13198 1.638651618 124 375 conserved hypothetical protein 7084995 7085369 11.8 13938 +7348 RB13217 1.632026935 127 384 Penicillinase repressor blaI 7091881 7091498 9.3 14071 -707 RB1322 3.633490801 35 108 hypothetical protein 671526 671633 12.8 3914 +
7352 RB13224 1.638640446 485 1458 nucleoside-diphosphate-sugar epimerase 7096270 7094813 9.5 53929 -7354 RB13228 1.822963597 116 351 conserved hypothetical protein 7097952 7098302 10.4 13570 +7363 RB13244 2.110430508 89 270 conserved hypothetical protein 7103330 7103061 10.3 9849 -7370 RB13259 1.966297351 634 1905 conserved hypothetical protein 7108365 7106461 5.7 72709 -7376 RB13267 1.800702079 239 720 conserved hypothetical protein 7117297 7116578 10.7 24985 -7400 RB13323 1.547636022 45 138 conserved hypothetical protein 7141536 7141673 11.7 5169 +715 RB1336 2.117833351 220 663 hypothetical protein 679357 680019 5.4 24662 +717 RB1339 1.700307676 101 306 hypothetical protein 681689 681384 11.1 11329 -725 RB1352 1.534628777 446 1341 hypothetical protein 686463 687803 5.5 46849 +733 RB1362 3.653375571 82 249 hypothetical protein 696352 696600 10.4 9054 +741 RB1384 1.983097668 177 534 membrane protein 707461 706928 9.8 19787 -759 RB1420 1.635284548 350 1053 inosine-uridine preferring nucleoside hydrolase 723375 724427 4.7 38646 +765 RB1432 1.713195711 199 600 NADH-ubiquinone oxidoreductase, chain 4L 1.6.5.3 732414 731815 4.2 21323 -766 RB1433 1.984728044 145 438 Na(+):H(+) antiporter subunit B (Multiple resistance and pH homeostasis protein B) mrpB 732848 732411 9.2 15860 -82 RB144 1.705177287 467 1404 conserved hypothetical protein 68273 66870 6.4 51000 -
783 RB1463 1.653889741 100 303 hypothetical protein 743584 743886 9.6 10972 +799 RB1491 2.793289876 111 336 hypothetical protein 766600 766265 12.3 12531 -807 RB1507 2.441120945 334 1005 tRNA isopentenyltransferase 2.5.1.8 772202 771198 5.5 37063 -809 RB1509 2.837151983 156 471 hypothetical protein 773379 772909 6.2 17244 -810 RB1511 2.137167157 539 1620 hypothetical protein 774995 773376 5.2 59301 -811 RB1513 1.660911513 98 297 secreted protein 775203 774907 11.6 10491 -816 RB1519 1.508592061 74 225 hypothetical protein 777299 777075 11.3 8617 -823 RB1531 1.772778159 451 1356 membrane protein 782534 783889 4.5 50616 +839 RB1563 4.115142655 136 411 hypothetical protein 798934 799344 10.2 14882 +

840 RB1565 2.454683213 76 231 hypothetical protein 799552 799322 10.4 8648 -859 RB1601 2.37761474 75 228 hypothetical protein 824863 824636 10 8335 -861 RB1603 2.226007927 77 234 hypothetical protein 825177 824944 11.7 8721 -862 RB1604 1.654313812 399 1200 DNA alkylation repair enzyme 826373 825174 10.1 45416 -870 RB1621 2.277332839 350 1053 Phosphoesterase, PA-phosphatase related 835361 834309 6.5 38846 -871 RB1624 1.756863483 97 294 hypothetical protein 835745 835452 10.2 10697 -880 RB1638 1.825309659 65 198 hypothetical protein 842521 842718 6.1 6959 +94 RB164 1.995785075 166 501 hypothetical protein 87060 87560 9.8 19615 +
885 RB1648 1.904541392 101 306 secreted protein 846423 846118 11.1 10776 -10 RB17 1.857901865 164 495 protein containing DUF1589 8251 7757 11.5 18498 -
911 RB1717 1.687655588 541 1626 conserved hypothetical protein, membrane 897501 895876 10.3 60139 -927 RB1750 1.745721227 58 177 hypothetical protein 909661 909837 10.1 6339 +928 RB1751 1.940030528 68 207 hypothetical protein 910027 909821 10.5 7181 -937 RB1771 1.872017804 83 252 hypothetical protein 919524 919273 7.6 9390 -941 RB1778 3.146980452 69 210 hypothetical protein 921245 921036 10.7 7677 -963 RB1830 3.073407605 37 114 hypothetical protein 945611 945498 6.6 3673 -980 RB1860 1.704480171 110 333 hypothetical protein 960361 960693 10.9 11827 +981 RB1862 1.745419592 135 408 transcriptional regulator, GntR family 960630 961037 10.2 14884 +983 RB1868 1.995003137 835 2508 membrane protein 962128 964635 5.4 93195 +997 RB1893 1.700274014 1090 3273 Serine/threonine-protein kinase spk1 2.7.1.37 979639 982911 4.7 120745 +999 RB1896 2.940217947 106 321 membrane protein containing DUF1294 985295 985615 11.2 11976 +
1003 RB1901 1.929260348 134 405 hypothetical protein 988429 988833 10.5 14815 +1008 RB1910 4.066965296 123 372 hypothetical protein 993764 993393 11.2 13384 -1053 RB2011 2.329166223 93 282 hypothetical protein 1059411 1059692 4.4 9880 +1068 RB2038 1.837125761 1553 4662 hypothetical protein 1076265 1071604 6.9 172935 -1069 RB2043 2.364366155 61 186 hypothetical protein 1076233 1076418 10.7 6909 +1122 RB2140 1.713943258 49 150 hypothetical protein 1131458 1131607 11.9 5498 +1126 RB2144 1.568531299 317 954 g eranylgeranyl pyrophosphate synthetase 1136089 1135136 5 33509 -1133 RB2155 1.989149473 156 471 hypothetical protein 1140660 1140190 4.9 17026 -1142 RB2173 2.986620064 354 1065 hypothetical protein 1147959 1146895 6.3 37862 -1150 RB2185 2.599831227 50 153 hypothetical protein 1152681 1152529 10.1 5841 -1151 RB2186 2.366402589 433 1302 ISXo8 transposase 1153993 1152692 9.4 49646 -1185 RB2252 1.718849778 53 162 hypothetical protein 1181701 1181540 9.8 6301 -1187 RB2254 1.811503778 541 1626 choline sulfatase betC 3.1.6.6 1184068 1182443 7.6 60782 -1193 RB2262 1.883608885 62 189 hypothetical protein 1188965 1188777 10.6 7138 -129 RB227 1.542512543 59 180 hypothetical protein 120858 121037 11.3 6741 +
1209 RB2287 1.753647245 63 192 protein containing DUF1586 1199020 1198829 9.3 6890 -1223 RB2310 2.050664594 107 324 hypothetical protein 1209545 1209222 11.5 11581 -1235 RB2330 2.654605928 183 552 secreted protein 1214565 1214014 8.7 20019 -1245 RB2342 1.572016185 72 219 hypothetical protein 1217901 1218119 11.6 7727 +1275 RB2382 1.798186499 75 228 hypothetical protein 1238921 1239148 9.7 8092 +1283 RB2398 1.990644567 294 885 conserved hypothetical protein 1248404 1247520 4.7 31364 -1286 RB2401 2.51191176 706 2121 Phosphoesterase, PA-phosphatase related 1248679 1250799 3.7 76509 +1313 RB2457 2.235239536 118 357 hypothetical protein 1279788 1280144 12.7 13138 +1330 RB2477 1.792540082 123 372 hypothetical protein 1295793 1295422 10.2 13737 -1331 RB2478 2.716028379 38 117 hypothetical protein 1295756 1295872 10.7 4233 +1341 RB2492 3.097118649 69 210 hypothetical protein 1302190 1302399 9.9 7373 +1347 RB2498 1.838717804 296 891 methyltransferase, 1305777 1304887 6.2 33105 -1355 RB2507 2.535623852 392 1179 dTDP-glucose 4,6-dehydratase rfbB 4.2.1.46 1313064 1311886 5.2 42907 -1357 RB2509 1.680878147 46 141 hypothetical protein 1313237 1313377 5.4 4895 +1358 RB2510 1.686900431 55 168 hypothetical protein 1313396 1313563 10.7 5612 +1359 RB2511 2.147696849 45 138 hypothetical protein 1313530 1313667 11.8 5256 +1361 RB2513 1.567537981 46 141 hypothetical protein 1314624 1314764 4.7 4748 +1364 RB2516 2.301111387 63 192 hypothetical protein 1316669 1316860 11.2 6506 +1370 RB2524 2.265273084 115 348 hypothetical protein 1319449 1319796 7.4 11654 +1376 RB2534 2.03212221 100 303 hypothetical protein 1324897 1325199 11.8 11196 +1380 RB2545 3.271000229 52 159 hypothetical protein 1328931 1328773 11.2 5992 -1387 RB2556 2.296831119 60 183 Planctomycete PGAMP 1335620 1335438 13.1 6572 -1396 RB2574 1.570776807 249 750 hypothetical protein 1342282 1341533 10.8 26506 -1401 RB2585 2.259234668 89 270 Bacterial microcompartments protein 1346035 1345766 4.6 8850 -1402 RB2586 1.972115656 98 297 Bacterial microcompartments protein 1346402 1346106 9 9952 -1423 RB2627 1.611677151 342 1029 Glyceraldehyde 3-phosphate dehydrogenase gap 1.2.1.12 1364551 1365579 5.4 36716 +1428 RB2636 2.493759853 116 351 hypothetical protein 1368450 1368100 11.7 12761 -1431 RB2640 1.945623977 349 1050 Sugar phosphate isomerase/epimerase 1372769 1371720 4.6 38081 -1452 RB2679 1.995932725 173 522 hypothetical protein 1391557 1392078 5.5 19502 +1486 RB2740 1.59555329 126 381 hypothetical protein 1430028 1429648 10.5 13639 -1492 RB2747 2.234430805 80 243 hypothetical protein 1432443 1432685 10.1 8485 +1493 RB2748 1.778516188 387 1164 conserved hypothetical protein 1433804 1432641 5.8 44722 -1494 RB2749 1.673075235 305 918 4-hydroxybenzoate octaprenyltransferase ubiA 2.5.1.- 1434783 1433866 9.8 32877 -1495 RB2750 3.030088826 222 669 3-octaprenyl-4-hydroxybenzoate carboxy-lyase ubiX 4.1.1.- 1435465 1434797 6.3 24338 -1500 RB2764 2.655704279 841 2526 secreted protein containing DUF1585 1442155 1439630 5 94741 -1506 RB2784 2.156914688 203 612 membrane protein containing DUF107 1448065 1447454 4.5 21076 -155 RB279 2.278596176 308 927 ABC-type multidrug transport system, ATPase component 138584 139510 5 34067 +
1511 RB2795 1.525548971 214 645 hypothetical protein 1454726 1455370 10.5 24354 +1522 RB2818 1.85753569 54 165 hypothetical protein 1463057 1462893 9.6 5981 -163 RB286 1.869403058 105 318 hypothetical protein 142779 143096 11.6 12056 +
1553 RB2875 1.57863758 73 222 hypothetical protein 1497313 1497534 9.9 8090 +1555 RB2878 1.501456248 219 660 conserved hypothetical protein 1497670 1498329 6.1 24857 +1572 RB2912 1.974925237 313 942 secreted protein 1512238 1511297 4.4 33483 -1578 RB2924 1.531150096 143 432 secreted protein 1515456 1515025 8.9 16141 -1591 RB2943 2.015262346 86 261 hypothetical protein 1521275 1521535 9.5 9549 +1614 RB2976 1.647844896 196 591 ATP:cob(I)alamin adenosyltransferase 1536008 1535418 4.9 21602 -1619 RB2983 2.05395807 293 882 Hydrogen peroxide-inducible genes activator oxyR 1540680 1539799 6 33942 -1620 RB2985 2.358237497 365 1098 glycosyl transferase, group 1 family protein 1542040 1540943 5.1 40609 -1637 RB3013 2.141860925 77 234 hypothetical protein 1563386 1563619 12.3 7957 +1672 RB3073 2.369863432 51 156 hypothetical protein 1585180 1585025 9.1 5384 -179 RB314 2.187367848 309 930 malonyl CoA-acyl carrier protein transacylase fabD 2.3.1.39 156968 157897 4.4 32349 +
1695 RB3140 1.675840507 111 336 hypothetical protein 1620838 1621173 10.2 12286 +1698 RB3144 1.582245667 348 1047 INTEGRASE/RECOMBINASE 1621739 1622785 9.6 39332 +1700 RB3146 2.738232396 98 297 protein containing DUF1556 1623952 1624248 11.4 10926 +1705 RB3152 2.029559749 68 207 hypothetical protein 1626667 1626461 12.2 7261 -1706 RB3153 2.297179262 76 231 hypothetical protein (protein containing DUF1584) 1626911 1626681 12.4 8266 -1708 RB3155 1.737756771 93 282 hypothetical protein 1627430 1627149 12.7 10369 -180 RB316 2.357048696 252 759 3-oxoacyl-[acyl-carrier-protein] reductase fabG 1.1.1.100 158031 158789 8.7 26160 +
1722 RB3189 2.063916051 102 309 hypothetical protein 1646085 1646393 11.1 11570 +1742 RB3224 1.97493902 148 447 hypothetical protein 1656004 1656450 12.4 16377 +186 RB324 2.065626064 49 150 hypothetical protein 162731 162880 10.5 5404 +
1755 RB3247 1.855081086 91 276 hypothetical protein 1668665 1668390 9.8 10018 -1766 RB3261 1.779391508 94 285 hypothetical protein 1678001 1677717 11.6 10565 -1803 RB3326 1.954199744 95 288 hypothetical protein 1711353 1711640 10.7 10480 +1809 RB3333 2.026225922 39 120 hypothetical protein 1713409 1713528 12.2 4095 +191 RB335 2.057101688 85 258 hypothetical protein 168642 168385 12.6 9143 -195 RB341 2.265892107 90 273 hypothetical protein 169852 170124 11.2 10096 +
1878 RB3449 1.601850015 52 159 hypothetical protein 1782475 1782317 11.3 5845 -1915 RB3509 1.521458235 336 1011 glycosyl hydrolase 1818015 1819025 5 37024 +1942 RB3565 1.920719145 137 414 membrane protein 1853670 1854083 10.1 15653 +1964 RB3599 2.279590265 365 1098 KLHL5 protein 1871888 1872985 6.5 40419 +1987 RB3633 1.666390417 90 273 hypothetical protein 1890483 1890211 12 10096 -2005 RB3659 2.633863645 59 180 hypothetical protein 1900428 1900607 10.7 6782 +

2008 RB3662 2.034633667 43 132 hypothetical protein 1902024 1901893 10.6 4816 -2009 RB3663 1.866613594 69 210 secreted protein 1902328 1902119 12.1 7609 -2129 RB3864 2.159302365 130 393 hypothetical protein 1998657 1999049 12.2 14855 +2130 RB3865 1.657844263 178 537 acetyltransferase, GNAT family 1999065 1999601 6.1 20057 +2295 RB4160 2.226402394 386 1161 conserved hypothetical protein 2136438 2137598 5.1 43127 +2300 RB4167 1.933027443 979 2940 secreted cytochrome c-like protein 2144846 2141907 4.8 107275 -2308 RB4185 2.383681463 89 270 hypothetical protein 2151951 2152220 12 9730 +2312 RB4194 2.433488216 53 162 hypothetical protein 2156685 2156524 11.4 5728 -2388 RB4306 1.625468979 457 1374 transposase and inactivated derivative 2221207 2219834 8.4 52492 -2398 RB4325 3.765600993 48 147 Planctomycete PGAMP 2228477 2228623 12.5 5403 +2403 RB4338 2.78496096 57 174 hypothetical protein 2233809 2233636 6.3 6607 -2405 RB4341 1.659604036 109 330 hypothetical protein 2234849 2235178 7.5 12018 +2440 RB4397 2.691758559 55 168 protein containing DUF1560 2263976 2264143 10.5 5816 +2454 RB4420 1.767751346 206 621 DedA family protein 2274093 2273473 9.3 22775 -2457 RB4425 1.812016826 127 384 hypothetical protein 2275100 2275483 10.6 14111 +2466 RB4440 1.634912345 347 1044 Amidohydrolase 2 4.1.1.45 2278882 2279925 6.6 39008 +2475 RB4457 2.385063444 98 297 hypothetical protein 2285628 2285924 10 10915 +2487 RB4473 1.514698444 57 174 hypothetical protein 2294981 2294808 9.1 5884 -2493 RB4482 2.526830138 111 336 hypothetical protein 2297240 2296905 12 12929 -2501 RB4498 2.197458016 90 273 hypothetical protein 2303522 2303250 9.4 9942 -2509 RB4510 3.283537625 49 150 hypothetical protein 2307657 2307508 9 5554 -2521 RB4530 3.168417353 1130 3393 heavy metal efflux pump, CzcA family 2323867 2320475 4.8 123379 -2542 RB4572 2.349259751 90 273 hypothetical protein 2339375 2339103 7.7 9938 -2558 RB4598 1.584206651 306 921 membrane protein 2355806 2356726 4 33125 +2559 RB4601 2.100239027 318 957 protein containing DUF1559 2356778 2357734 5.3 34567 +2587 RB4645 1.641416077 69 210 hypothetical protein 2378266 2378057 10.9 7946 -2593 RB4652 1.897678047 52 159 hypothetical protein 2380494 2380652 12.6 5628 +2601 RB4662 3.347879694 37 114 secreted protein 2383563 2383676 4.4 4069 +2613 RB4691 2.507103018 284 855 hypothetical protein 2395001 2395855 12 32256 +2622 RB4705 1.622136032 56 171 hypothetical protein 2403864 2403694 12.6 6330 -2637 RB4727 2.004479953 54 165 hypothetical protein 2422602 2422766 9.1 5908 +2654 RB4750 1.53101482 204 615 membrane protein 2435462 2436076 10 23312 +2673 RB4781 2.200497845 156 471 HesB/YadR/YfhF 2449655 2450125 4.6 17413 +2699 RB4815 1.865895895 491 1476 arylsulfatase precursor ARSA 3.1.6.8 2469289 2467814 5.7 54617 -2704 RB4826 1.656393726 464 1395 Transposase IS66 2471205 2472599 10.2 51739 +282 RB483 2.063475736 54 165 hypothetical protein 262569 262733 12.2 6454 +
2717 RB4850 1.857021056 150 453 membrane protein 2484018 2484470 7.6 15931 +2745 RB4894 1.639002853 712 2139 glycogen debranching enzyme glgX 3.2.1.- 2518939 2516801 5 81197 -2783 RB4957 2.146895728 57 174 hypothetical protein 2548380 2548553 12.4 6569 +2792 RB4969 2.023603795 334 1005 hypothetical protein 2554443 2555447 10 37084 +2821 RB5010 2.70359212 94 285 hypothetical protein 2577161 2576877 11.9 10778 -2830 RB5029 2.463803039 236 711 conserved hypothetical protein, secreted 2589744 2589034 4.6 26291 -2867 RB5093 1.893127073 67 204 hypothetical protein 2619010 2619213 6.3 7246 +2894 RB5138 2.710550482 58 177 hypothetical protein 2641024 2640848 12.4 6711 -2908 RB5164 2.562994028 218 657 hypothetical protein 2658365 2657709 11.4 24275 -2910 RB5168 1.794347644 60 183 hypothetical protein 2658508 2658690 9.7 6626 +299 RB519 2.066265663 54 165 hypothetical protein 285036 284872 10.6 6171 -
2926 RB5195 1.527558092 505 1518 arylsulfatase Arsb 3.1.6.12 2674328 2675845 5 56413 +301 RB521 4.512236615 63 192 hypothetical protein 286399 286208 10.7 7040 -
2984 RB5294 1.540465479 533 1602 sulfatase 2732584 2730983 5.6 61291 -3023 RB5371 1.76572172 109 330 hypothetical protein 2774829 2775158 7.5 12033 +3047 RB5412 3.524059925 72 219 hypothetical protein 2794838 2794620 12.3 8592 -316 RB546 3.008885687 485 1458 secreted protein containing DUF1501 302834 304291 7.8 53385 +
3103 RB5502 2.242859912 144 435 membrane protein 2845859 2846293 3.1 15161 +321 RB556 1.708513615 47 144 hypothetical protein 308213 308356 12.4 5508 +322 RB557 1.801550469 53 162 hypothetical protein 308223 308384 10.5 5688 +
3142 RB5572 1.937860486 39 120 hypothetical protein 2901618 2901499 13 4731 -3153 RB5597 2.927301342 170 513 conserved hypothetical protein, secreted 2913295 2913807 11.9 17556 +324 RB560 2.240012693 280 843 protein containing DGPFAETKE domain DGPF 309384 308542 4.9 30721 -
3215 RB5707 3.494468009 183 552 hypothetical protein 2979435 2979986 10.2 20474 +3233 RB5737 2.165159487 120 363 hypothetical protein 2994415 2994777 11.9 13608 +3292 RB5846 1.552211655 297 894 glycosyltransferase 3064164 3065057 6.3 32718 +3304 RB5867 1.636075198 50 153 hypothetical protein 3077578 3077730 9.7 5871 +3309 RB5874 1.69375552 47 144 hypothetical protein 3080941 3080798 4.6 5338 -3312 RB5877 1.674709272 35 108 hypothetical protein 3082302 3082409 10.6 3895 +3319 RB5887 2.051189207 292 879 integrase 3087343 3086465 9.9 33335 -3334 RB5914 2.906505812 56 171 hypothetical protein 3097264 3097434 10.5 6574 +3345 RB5937 2.853944961 306 921 hypothetical protein 3112147 3113067 11.7 34734 +3361 RB5966 1.580545054 140 423 hypothetical protein 3128115 3128537 10.1 15593 +3374 RB5988 2.208521161 174 525 secreted protein 3141494 3140970 9.4 18622 -3376 RB5990 2.310006948 113 342 hypothetical protein 3141918 3141577 11.8 12587 -3390 RB6011 1.682557663 50 153 hypothetical protein 3150235 3150083 10.1 5841 -3394 RB6016 1.906776176 188 567 hypothetical protein 3152860 3153426 11.7 20633 +3426 RB6081 1.51616219 52 159 hypothetical protein 3191035 3190877 11.5 5837 -3440 RB6108 1.523955685 1096 3291 secreted protein 3208185 3204895 5.4 123062 -3459 RB6147 1.516334798 394 1185 3-dehydroquinate synthase aroB 4.2.3.4 3229258 3230442 6.2 43267 +3471 RB6167 1.500109148 375 1128 Putative transposase 3235937 3237064 9.9 41895 +3472 RB6168 2.304188414 208 627 hypothetical protein 3236689 3237315 12.2 22575 +3477 RB6175 2.184505189 125 378 hypothetical protein 3240125 3239748 11.9 14247 -3482 RB6188 3.009885374 68 207 hypothetical protein 3244384 3244178 9.6 6986 -3513 RB6245 1.931203785 107 324 hypothetical protein 3278762 3279085 12.4 12397 +3516 RB6250 1.514075438 35 108 hypothetical protein 3282079 3282186 9.4 4025 +3520 RB6257 3.142660037 207 624 secreted protein 3288666 3288043 4.6 22458 -3573 RB6355 1.53314894 505 1518 iron-sulfur cluster-binding protein 3336973 3338490 9.6 55993 +3584 RB6374 2.969521508 442 1329 conserved hypothetical protein 3348288 3349616 9 48323 +3585 RB6375 2.716117152 297 894 hypothetical protein 3349681 3350574 8.7 31772 +3589 RB6380 2.144492471 74 225 hypothetical protein 3353359 3353583 12.4 8433 +3591 RB6382 2.35405938 39 120 hypothetical protein 3355162 3355043 10.4 4208 -3598 RB6390 1.942322628 86 261 hypothetical protein 3361959 3362219 12.8 9768 +3607 RB6405 1.914661933 93 282 hypothetical protein 3371368 3371087 9.6 10148 -3629 RB6440 1.624957167 57 174 hypothetical protein 3391656 3391829 10.4 6422 +3642 RB6459 1.590981738 2028 6087 cell surface protein 3408788 3414874 3.5 214858 +3711 RB6571 1.630308658 154 465 secreted protein 3489550 3490014 9 17657 +
39 RB66 1.62209628 84 255 hypothetical protein 26082 26336 11.1 9333 +3735 RB6619 1.663215867 262 789 hypothetical protein 3518291 3517503 9.7 29227 -362 RB663 2.084810827 79 240 hypothetical protein 357608 357369 11.4 8792 -
3753 RB6655 1.582402313 36 111 hypothetical protein 3535829 3535719 9.4 4005 -3756 RB6661 1.538114086 88 267 hypothetical protein 3538752 3538486 10.6 9093 -3770 RB6684 1.61337283 105 318 hypothetical protein 3554034 3553717 11.6 11612 -3771 RB6685 1.596892111 72 219 hypothetical protein 3554265 3554047 12.2 8077 -365 RB670 1.649686669 136 411 hypothetical protein 359315 358905 10.1 15444 -
3784 RB6706 1.931579477 37 114 hypothetical protein 3560942 3560829 7.3 4375 -366 RB671 2.477782668 122 369 hypothetical protein 359218 359586 10.7 13383 +
3793 RB6720 2.082468911 72 219 hypothetical protein 3567752 3567970 10.3 8140 +3804 RB6736 2.854676533 433 1302 ISXo8 transposase 3578852 3577551 9.4 49646 -3817 RB6761 1.720050338 198 597 bacterioferritin comigratory protein 3594163 3594759 5.7 21350 +3834 RB6786 1.685047733 172 519 hypothetical protein 3607257 3607775 12.3 20026 +3852 RB6822 2.882630068 404 1215 Chorismate synthase aroC 4.2.3.5 3623410 3624624 5.8 42918 +3885 RB6883 2.308615096 255 768 Lipoprotein releasing system ATP-binding protein lolD lolD 3657855 3658622 5.6 28302 +

3893 RB6903 1.641538975 53 162 hypothetical protein 3669220 3669059 9.3 6058 -3904 RB6916 2.497799815 67 204 hypothetical protein 3676188 3676391 12 7266 +3909 RB6925 3.784134297 161 486 hypothetical protein 3678461 3677976 9.8 17641 -3911 RB6930 3.097264296 413 1242 glutamate dehydrogenase 1.4.1.3 3680317 3679076 5.9 45734 -3912 RB6932 2.972180516 319 960 CYSTEINE SYNTHASE A cysK 2.5.1.47 3681308 3680349 5 33739 -3914 RB6935 1.797951989 627 1884 pyruvate:ferredoxin oxidoreductase (alpha subunit) 3684260 3682377 5.5 67263 -3940 RB6989 2.291433415 82 249 hypothetical protein 3719206 3718958 12.5 9143 -3949 RB7002 1.955400985 232 699 secreted protein 3725925 3726623 7.7 25003 +3956 RB7016 2.59004597 241 726 hypothetical protein 3732513 3733238 7.7 26263 +3963 RB7028 1.604036296 545 1638 hypothetical protein 3736857 3738494 5.1 58222 +4013 RB7114 1.673446914 347 1044 phenylalanyl-tRNA synthetase alpha chain pheS 6.1.1.20 3779252 3778209 4.6 38018 -4015 RB7116 3.58641694 59 180 hypothetical protein 3779586 3779765 11.7 6458 +4027 RB7136 1.638258201 68 207 hypothetical protein 3787932 3788138 7.4 7499 +4047 RB7176 1.561644294 46 141 hypothetical protein 3806783 3806643 10.1 4647 -392 RB723 2.617929723 60 183 hypothetical protein 388051 388233 7.3 6624 +
4118 RB7292 2.345087554 256 771 metallo-beta-lactamase 3875577 3876347 5.4 28641 +4135 RB7318 3.003763502 126 381 hypothetical protein 3895264 3894884 7.3 13046 -4136 RB7319 2.55122604 76 231 hypothetical protein 3895491 3895261 12.4 8291 -4156 RB7388 1.795157437 282 849 INTEGRASE/RECOMBINASE 3952745 3953593 10.6 31873 +4157 RB7389 2.206131059 375 1128 Putative transposase 3953590 3954717 9.9 41895 +4158 RB7390 1.5717743 208 627 hypothetical protein 3954342 3954968 12.2 22575 +4160 RB7393 1.768724913 66 201 hypothetical protein 3955353 3955553 11 7563 +4203 RB7464 3.263712295 44 135 hypothetical protein 4001614 4001480 10.2 4711 -4207 RB7471 1.769148263 92 279 hypothetical protein 4006361 4006083 11.9 10550 -4216 RB7486 1.83812896 130 393 hypothetical protein 4017064 4016672 8.8 14588 -403 RB754 2.092237647 364 1095 Polysaccharide export protein 403211 404305 4.2 39246 +
4285 RB7599 1.837320006 39 120 hypothetical protein 4086498 4086379 10.8 4566 -4318 RB7651 1.910539418 44 135 hypothetical protein 4112517 4112383 6.7 4749 -4319 RB7652 2.842117135 315 948 malate dehydrogenase mdh 1.1.1.37 4112505 4113452 4.5 33055 +4332 RB7678 1.89392125 226 681 protocatechuate 3,4-dioxygenase type II beta subunit 4123339 4122659 9.4 25090 -4345 RB7703 2.098931056 98 297 hypothetical protein 4131842 4132138 8.9 10578 +4400 RB7790 1.879833001 89 270 hypothetical protein 4173233 4173502 10.6 9770 +4425 RB7833 1.690086332 228 687 Ribosomal protein L3 4192692 4193378 11 24681 +4449 RB7875 2.32438741 492 1479 Arylsulfatase A precursor ARSA 3.1.6.8 4208176 4209654 5.4 54661 +4470 RB7909 1.671996189 330 993 secreted protein 4225494 4224502 10.6 34211 -4478 RB7924 1.688366017 228 687 RNA polymerase sigma factor 4231813 4232499 6.2 25242 +4548 RB8065 1.999674433 38 117 hypothetical protein 4318540 4318424 10.4 3952 -4561 RB8084 2.15890448 38 117 hypothetical protein 4330006 4329890 12.4 4461 -4573 RB8107 1.84185736 315 948 secreted protein 4339329 4340276 5.4 34412 +4582 RB8127 1.688135503 262 789 hypothetical protein 4349191 4349979 11.7 28414 +4586 RB8138 1.99933795 75 228 hypothetical protein 4351960 4352187 4.1 8440 +4632 RB8231 1.523420874 170 513 ACT domain protein 4401779 4402291 4.4 18932 +4643 RB8248 1.579525345 253 762 dienelactone hydrolase family protein 4409395 4410156 5.7 28139 +4648 RB8260 1.55919706 108 327 protein containing DUF167 4415394 4415068 10.5 11641 -4655 RB8275 1.614651716 45 138 hypothetical protein 4424897 4424760 11.4 5115 -4657 RB8279 2.194205192 71 216 hypothetical protein 4425128 4425343 5.8 7835 +4696 RB8353 1.828073353 82 249 hypothetical protein 4459374 4459622 10.8 9402 +4699 RB8357 1.947454134 56 171 hypothetical protein 4460174 4460004 7.4 6097 -4714 RB8384 1.753384306 433 1302 ISXo8 transposase 4478623 4479924 9.4 49646 +457 RB839 2.182062009 60 183 hypothetical protein 441662 441844 6.1 6654 +
4724 RB8404 1.711268941 239 720 Silent information regulator protein Sir2 4492899 4493618 4.7 26142 +4749 RB8457 3.160312784 113 342 hypothetical protein 4518458 4518117 11.5 12525 -4755 RB8468 1.777768949 88 267 hypothetical protein 4520799 4520533 11.9 9719 -4761 RB8478 1.742151616 141 426 hypothetical protein 4524759 4524334 11.6 15805 -4794 RB8530 1.749276032 53 162 hypothetical protein 4550886 4550725 12.3 5816 -4802 RB8541 1.642385639 357 1074 Peptidase M42 4554758 4555831 4.6 38279 +4861 RB8669 1.687927001 37 114 hypothetical protein 4614557 4614670 11.5 4395 +471 RB867 2.017027094 100 303 hypothetical protein 460731 460429 9.7 11273 -
4862 RB8670 4.214509032 47 144 hypothetical protein 4614842 4614985 11.1 5371 +4879 RB8697 3.261367962 56 171 hypothetical protein 4630281 4630111 9.7 6374 -4880 RB8700 3.773767421 58 177 hypothetical protein 4630224 4630400 4.8 6268 +4891 RB8723 2.487879984 74 225 hypothetical protein 4638077 4638301 12 8520 +4893 RB8726 1.567912793 599 1800 single-stranded-DNA-specific exonuclease RecJ recJ 3.1.-.- 4638505 4640304 5.3 64794 +4905 RB8747 2.083737467 218 657 DNA/pantothenate metabolism flavoprotein 4648152 4648808 5.5 23702 +4906 RB8748 2.332252043 305 918 dihydroorotate dehydrogenase pyrD 1.3.3.1 4648815 4649732 4.6 31813 +4941 RB8815 3.22054004 71 216 hypothetical protein 4687955 4687740 4.8 7683 -4943 RB8818 2.134712008 60 183 hypothetical protein 4689021 4688839 10.4 6695 -4956 RB8840 2.085169881 94 285 acylphosphatases 4701445 4701729 10.2 10806 +4976 RB8876 1.625211468 87 264 hypothetical protein 4723629 4723892 9.5 9540 +4982 RB8884 2.25520575 70 213 hypothetical protein 4727666 4727878 11.8 8000 +4994 RB8902 2.296035668 82 249 hypothetical protein 4740474 4740226 11.2 9412 -5004 RB8917 2.592910345 74 225 hypothetical protein 4747435 4747659 10.4 8075 +5031 RB8975 2.603055071 110 333 hypothetical protein 4783888 4783556 10.4 12068 -5035 RB8980 1.741796604 618 1857 protein containing DUF1559 4788689 4786833 4.8 67763 -5036 RB8983 1.679549934 346 1041 methanol dehydrogenase regulatory protein 4788960 4790000 4.8 37993 +5038 RB8987 1.637584191 48 147 hypothetical protein 4790856 4790710 9.1 5309 -5050 RB9017 1.676962039 193 582 hypthetical protein 4805225 4805806 6.3 21847 +5055 RB9024 1.646716221 46 141 hypothetical protein 4809190 4809050 10.3 5125 -5061 RB9037 1.681174613 344 1035 secreted protein 4821537 4822571 6 36987 +5062 RB9038 2.739410555 153 462 hypothetical protein 4823103 4822642 10.5 17019 -5079 RB9078 1.55341567 969 2910 2-oxoglutarate dehydrogenase E1 component OGDH 1.2.4.2 4846881 4849790 5.7 109319 +5116 RB9144 1.996588404 150 453 hypothetical protein 4893346 4893798 11 16267 +496 RB919 1.772829794 109 330 hypothetical protein 488799 488470 10.2 11958 -
5136 RB9190 2.009547343 132 399 ferredoxin 4914039 4914437 5 14357 +5140 RB9196 2.109875658 446 1341 secreted protein containing DUF1080 4916635 4917975 5.4 50522 +5142 RB9198 5.126650093 58 177 hypothetical protein 4918032 4918208 11.8 6612 +5158 RB9231 2.16301151 109 330 hypothetical protein 4937008 4936679 7.5 12033 -500 RB924 2.860471873 208 627 hypothetical protein 490400 489774 12.2 22548 -
5169 RB9249 1.661379564 810 2433 hypothetical protein 4947571 4945139 5.4 86740 -5181 RB9276 1.574652752 233 702 membrane protein 4958461 4959162 7.5 24790 +5197 RB9308 2.017185123 629 1890 Outer membrane efflux family protein 4978849 4976960 6.9 69055 -504 RB931 2.874769085 39 120 hypothetical protein 492159 492278 9.4 4169 +506 RB933 2.254207171 208 627 hypothetical protein 493451 492825 12.2 22575 -
5211 RB9338 1.506192607 182 549 formaldehyde-activating enzyme fae 5002305 5002853 5.6 19774 +507 RB934 2.171115834 375 1128 Putative transposase 494203 493076 9.9 41881 -
5217 RB9346 1.777973228 71 216 hypothetical protein 5008335 5008550 10.5 7755 +508 RB936 1.589630607 85 258 hypothetical protein 494524 494267 11.1 9715 -
5230 RB9365 3.002822981 152 459 hypothetical protein 5017351 5017809 11.6 16637 +5257 RB9415 2.013265204 36 111 hypothetical protein 5050419 5050529 10.6 4057 +5278 RB9450 1.892134357 52 159 hypothetical protein 5076581 5076739 7.3 5967 +522 RB957 1.975418743 195 588 Tyrosine recombinase xerD xerD 505476 504889 10.2 21971 -
5355 RB9592 1.552934579 269 810 conserved hypothetical protein 5157928 5158737 5.5 30261 +5415 RB9702 2.664021836 96 291 conserved hypothetical protein 5229342 5229632 4.6 9322 +5461 RB9793 1.845305093 63 192 hypothetical protein 5273041 5272850 12.5 6957 -5524 RB9907 1.925591118 433 1302 ISXo8 transposase 5338185 5339486 9.4 49646 +5525 RB9908 2.150644277 50 153 hypothetical protein 5339497 5339649 9.9 5867 +5532 RB9918 2.745466406 167 504 hypothetical protein 5344369 5344872 5.3 18361 +
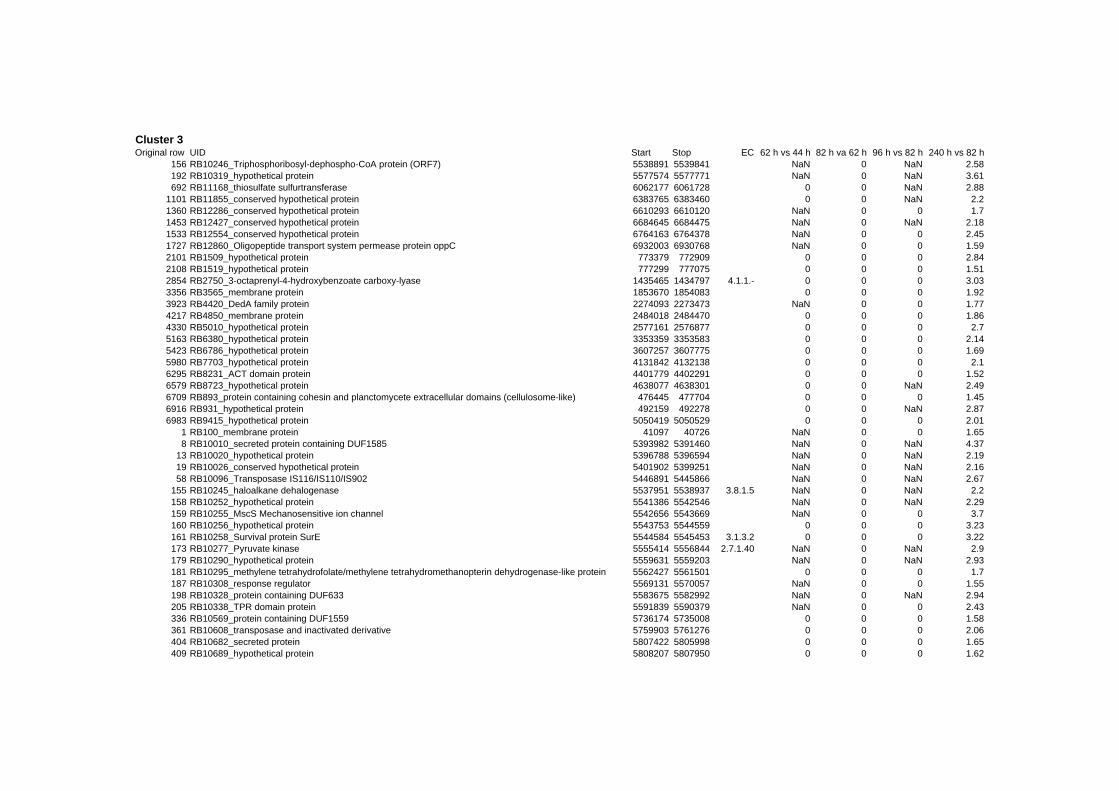
Cluster 3Original row UID Start Stop EC 62 h vs 44 h 82 h va 62 h 96 h vs 82 h 240 h vs 82 h
156 RB10246_Triphosphoribosyl-dephospho-CoA protein (ORF7) 5538891 5539841 NaN 0 NaN 2.58192 RB10319_hypothetical protein 5577574 5577771 NaN 0 NaN 3.61692 RB11168_thiosulfate sulfurtransferase 6062177 6061728 0 0 NaN 2.88
1101 RB11855_conserved hypothetical protein 6383765 6383460 0 0 NaN 2.21360 RB12286_conserved hypothetical protein 6610293 6610120 NaN 0 0 1.71453 RB12427_conserved hypothetical protein 6684645 6684475 NaN 0 NaN 2.181533 RB12554_conserved hypothetical protein 6764163 6764378 NaN 0 0 2.451727 RB12860_Oligopeptide transport system permease protein oppC 6932003 6930768 NaN 0 0 1.592101 RB1509_hypothetical protein 773379 772909 0 0 0 2.842108 RB1519_hypothetical protein 777299 777075 0 0 0 1.512854 RB2750_3-octaprenyl-4-hydroxybenzoate carboxy-lyase 1435465 1434797 4.1.1.- 0 0 0 3.033356 RB3565_membrane protein 1853670 1854083 0 0 0 1.923923 RB4420_DedA family protein 2274093 2273473 NaN 0 0 1.774217 RB4850_membrane protein 2484018 2484470 0 0 0 1.864330 RB5010_hypothetical protein 2577161 2576877 0 0 0 2.75163 RB6380_hypothetical protein 3353359 3353583 0 0 0 2.145423 RB6786_hypothetical protein 3607257 3607775 0 0 0 1.695980 RB7703_hypothetical protein 4131842 4132138 0 0 0 2.16295 RB8231_ACT domain protein 4401779 4402291 0 0 0 1.526579 RB8723_hypothetical protein 4638077 4638301 0 0 NaN 2.496709 RB893_protein containing cohesin and planctomycete extracellular domains (cellulosome-like) 476445 477704 0 0 0 1.456916 RB931_hypothetical protein 492159 492278 0 0 NaN 2.876983 RB9415_hypothetical protein 5050419 5050529 0 0 0 2.01
1 RB100_membrane protein 41097 40726 NaN 0 0 1.658 RB10010_secreted protein containing DUF1585 5393982 5391460 NaN 0 NaN 4.37
13 RB10020_hypothetical protein 5396788 5396594 NaN 0 NaN 2.1919 RB10026_conserved hypothetical protein 5401902 5399251 NaN 0 NaN 2.1658 RB10096_Transposase IS116/IS110/IS902 5446891 5445866 NaN 0 NaN 2.67
155 RB10245_haloalkane dehalogenase 5537951 5538937 3.8.1.5 NaN 0 NaN 2.2158 RB10252_hypothetical protein 5541386 5542546 NaN 0 NaN 2.29159 RB10255_MscS Mechanosensitive ion channel 5542656 5543669 NaN 0 0 3.7160 RB10256_hypothetical protein 5543753 5544559 0 0 0 3.23161 RB10258_Survival protein SurE 5544584 5545453 3.1.3.2 0 0 0 3.22173 RB10277_Pyruvate kinase 5555414 5556844 2.7.1.40 NaN 0 NaN 2.9179 RB10290_hypothetical protein 5559631 5559203 NaN 0 NaN 2.93181 RB10295_methylene tetrahydrofolate/methylene tetrahydromethanopterin dehydrogenase-like protein 5562427 5561501 0 0 0 1.7187 RB10308_response regulator 5569131 5570057 NaN 0 0 1.55198 RB10328_protein containing DUF633 5583675 5582992 NaN 0 NaN 2.94205 RB10338_TPR domain protein 5591839 5590379 NaN 0 0 2.43336 RB10569_protein containing DUF1559 5736174 5735008 0 0 0 1.58361 RB10608_transposase and inactivated derivative 5759903 5761276 0 0 0 2.06404 RB10682_secreted protein 5807422 5805998 0 0 0 1.65409 RB10689_hypothetical protein 5808207 5807950 0 0 0 1.62
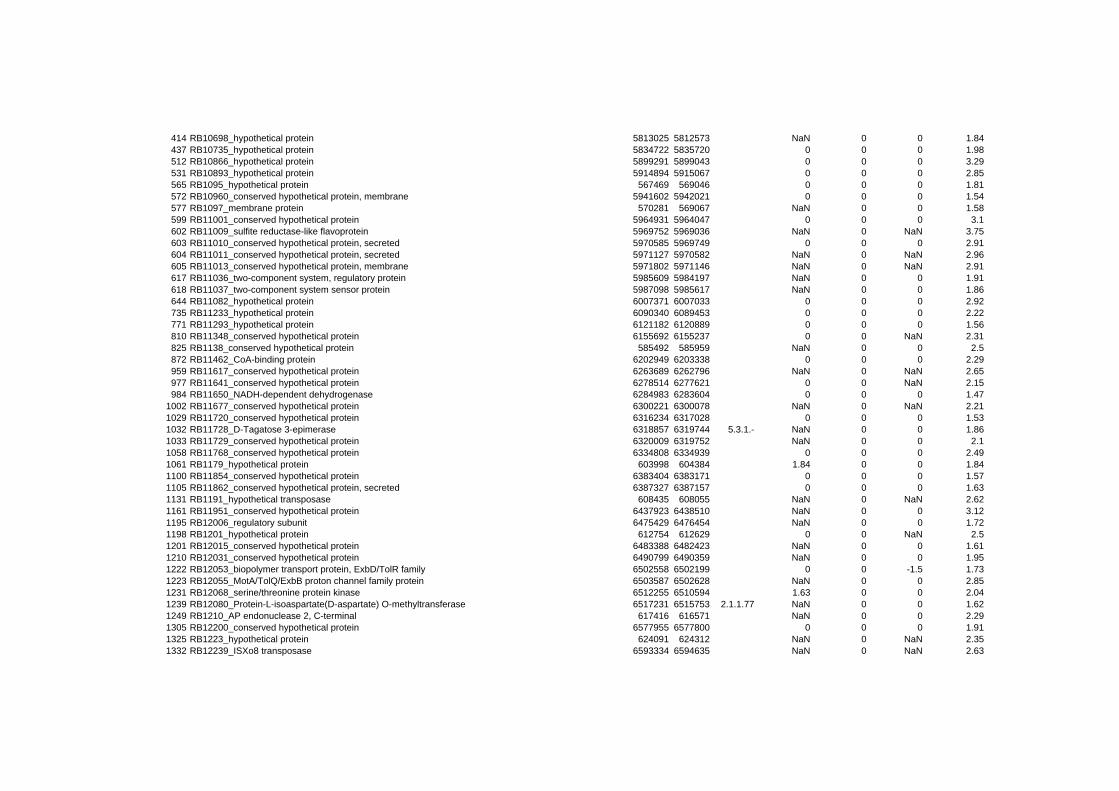
414 RB10698_hypothetical protein 5813025 5812573 NaN 0 0 1.84437 RB10735_hypothetical protein 5834722 5835720 0 0 0 1.98512 RB10866_hypothetical protein 5899291 5899043 0 0 0 3.29531 RB10893_hypothetical protein 5914894 5915067 0 0 0 2.85565 RB1095_hypothetical protein 567469 569046 0 0 0 1.81572 RB10960_conserved hypothetical protein, membrane 5941602 5942021 0 0 0 1.54577 RB1097_membrane protein 570281 569067 NaN 0 0 1.58599 RB11001_conserved hypothetical protein 5964931 5964047 0 0 0 3.1602 RB11009_sulfite reductase-like flavoprotein 5969752 5969036 NaN 0 NaN 3.75603 RB11010_conserved hypothetical protein, secreted 5970585 5969749 0 0 0 2.91604 RB11011_conserved hypothetical protein, secreted 5971127 5970582 NaN 0 NaN 2.96605 RB11013_conserved hypothetical protein, membrane 5971802 5971146 NaN 0 NaN 2.91617 RB11036_two-component system, regulatory protein 5985609 5984197 NaN 0 0 1.91618 RB11037_two-component system sensor protein 5987098 5985617 NaN 0 0 1.86644 RB11082_hypothetical protein 6007371 6007033 0 0 0 2.92735 RB11233_hypothetical protein 6090340 6089453 0 0 0 2.22771 RB11293_hypothetical protein 6121182 6120889 0 0 0 1.56810 RB11348_conserved hypothetical protein 6155692 6155237 0 0 NaN 2.31825 RB1138_conserved hypothetical protein 585492 585959 NaN 0 0 2.5872 RB11462_CoA-binding protein 6202949 6203338 0 0 0 2.29959 RB11617_conserved hypothetical protein 6263689 6262796 NaN 0 NaN 2.65977 RB11641_conserved hypothetical protein 6278514 6277621 0 0 NaN 2.15984 RB11650_NADH-dependent dehydrogenase 6284983 6283604 0 0 0 1.47
1002 RB11677_conserved hypothetical protein 6300221 6300078 NaN 0 NaN 2.211029 RB11720_conserved hypothetical protein 6316234 6317028 0 0 0 1.531032 RB11728_D-Tagatose 3-epimerase 6318857 6319744 5.3.1.- NaN 0 0 1.861033 RB11729_conserved hypothetical protein 6320009 6319752 NaN 0 0 2.11058 RB11768_conserved hypothetical protein 6334808 6334939 0 0 0 2.491061 RB1179_hypothetical protein 603998 604384 1.84 0 0 1.841100 RB11854_conserved hypothetical protein 6383404 6383171 0 0 0 1.571105 RB11862_conserved hypothetical protein, secreted 6387327 6387157 0 0 0 1.631131 RB1191_hypothetical transposase 608435 608055 NaN 0 NaN 2.621161 RB11951_conserved hypothetical protein 6437923 6438510 NaN 0 0 3.121195 RB12006_regulatory subunit 6475429 6476454 NaN 0 0 1.721198 RB1201_hypothetical protein 612754 612629 0 0 NaN 2.51201 RB12015_conserved hypothetical protein 6483388 6482423 NaN 0 0 1.611210 RB12031_conserved hypothetical protein 6490799 6490359 NaN 0 0 1.951222 RB12053_biopolymer transport protein, ExbD/TolR family 6502558 6502199 0 0 -1.5 1.731223 RB12055_MotA/TolQ/ExbB proton channel family protein 6503587 6502628 NaN 0 0 2.851231 RB12068_serine/threonine protein kinase 6512255 6510594 1.63 0 0 2.041239 RB12080_Protein-L-isoaspartate(D-aspartate) O-methyltransferase 6517231 6515753 2.1.1.77 NaN 0 0 1.621249 RB1210_AP endonuclease 2, C-terminal 617416 616571 NaN 0 0 2.291305 RB12200_conserved hypothetical protein 6577955 6577800 0 0 0 1.911325 RB1223_hypothetical protein 624091 624312 NaN 0 NaN 2.351332 RB12239_ISXo8 transposase 6593334 6594635 NaN 0 NaN 2.63
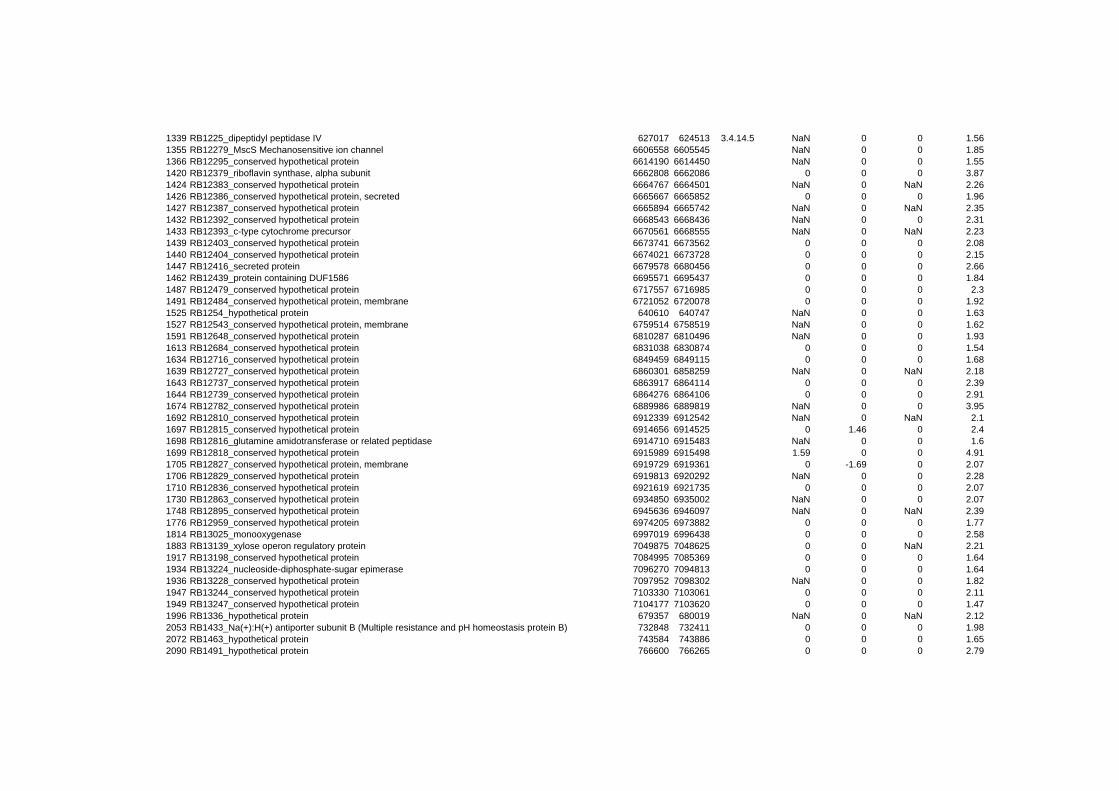
1339 RB1225_dipeptidyl peptidase IV 627017 624513 3.4.14.5 NaN 0 0 1.561355 RB12279_MscS Mechanosensitive ion channel 6606558 6605545 NaN 0 0 1.851366 RB12295_conserved hypothetical protein 6614190 6614450 NaN 0 0 1.551420 RB12379_riboflavin synthase, alpha subunit 6662808 6662086 0 0 0 3.871424 RB12383_conserved hypothetical protein 6664767 6664501 NaN 0 NaN 2.261426 RB12386_conserved hypothetical protein, secreted 6665667 6665852 0 0 0 1.961427 RB12387_conserved hypothetical protein 6665894 6665742 NaN 0 NaN 2.351432 RB12392_conserved hypothetical protein 6668543 6668436 NaN 0 0 2.311433 RB12393_c-type cytochrome precursor 6670561 6668555 NaN 0 NaN 2.231439 RB12403_conserved hypothetical protein 6673741 6673562 0 0 0 2.081440 RB12404_conserved hypothetical protein 6674021 6673728 0 0 0 2.151447 RB12416_secreted protein 6679578 6680456 0 0 0 2.661462 RB12439_protein containing DUF1586 6695571 6695437 0 0 0 1.841487 RB12479_conserved hypothetical protein 6717557 6716985 0 0 0 2.31491 RB12484_conserved hypothetical protein, membrane 6721052 6720078 0 0 0 1.921525 RB1254_hypothetical protein 640610 640747 NaN 0 0 1.631527 RB12543_conserved hypothetical protein, membrane 6759514 6758519 NaN 0 0 1.621591 RB12648_conserved hypothetical protein 6810287 6810496 NaN 0 0 1.931613 RB12684_conserved hypothetical protein 6831038 6830874 0 0 0 1.541634 RB12716_conserved hypothetical protein 6849459 6849115 0 0 0 1.681639 RB12727_conserved hypothetical protein 6860301 6858259 NaN 0 NaN 2.181643 RB12737_conserved hypothetical protein 6863917 6864114 0 0 0 2.391644 RB12739_conserved hypothetical protein 6864276 6864106 0 0 0 2.911674 RB12782_conserved hypothetical protein 6889986 6889819 NaN 0 0 3.951692 RB12810_conserved hypothetical protein 6912339 6912542 NaN 0 NaN 2.11697 RB12815_conserved hypothetical protein 6914656 6914525 0 1.46 0 2.41698 RB12816_glutamine amidotransferase or related peptidase 6914710 6915483 NaN 0 0 1.61699 RB12818_conserved hypothetical protein 6915989 6915498 1.59 0 0 4.911705 RB12827_conserved hypothetical protein, membrane 6919729 6919361 0 -1.69 0 2.071706 RB12829_conserved hypothetical protein 6919813 6920292 NaN 0 0 2.281710 RB12836_conserved hypothetical protein 6921619 6921735 0 0 0 2.071730 RB12863_conserved hypothetical protein 6934850 6935002 NaN 0 0 2.071748 RB12895_conserved hypothetical protein 6945636 6946097 NaN 0 NaN 2.391776 RB12959_conserved hypothetical protein 6974205 6973882 0 0 0 1.771814 RB13025_monooxygenase 6997019 6996438 0 0 0 2.581883 RB13139_xylose operon regulatory protein 7049875 7048625 0 0 NaN 2.211917 RB13198_conserved hypothetical protein 7084995 7085369 0 0 0 1.641934 RB13224_nucleoside-diphosphate-sugar epimerase 7096270 7094813 0 0 0 1.641936 RB13228_conserved hypothetical protein 7097952 7098302 NaN 0 0 1.821947 RB13244_conserved hypothetical protein 7103330 7103061 0 0 0 2.111949 RB13247_conserved hypothetical protein 7104177 7103620 0 0 0 1.471996 RB1336_hypothetical protein 679357 680019 NaN 0 NaN 2.122053 RB1433_Na(+):H(+) antiporter subunit B (Multiple resistance and pH homeostasis protein B) 732848 732411 0 0 0 1.982072 RB1463_hypothetical protein 743584 743886 0 0 0 1.652090 RB1491_hypothetical protein 766600 766265 0 0 0 2.79
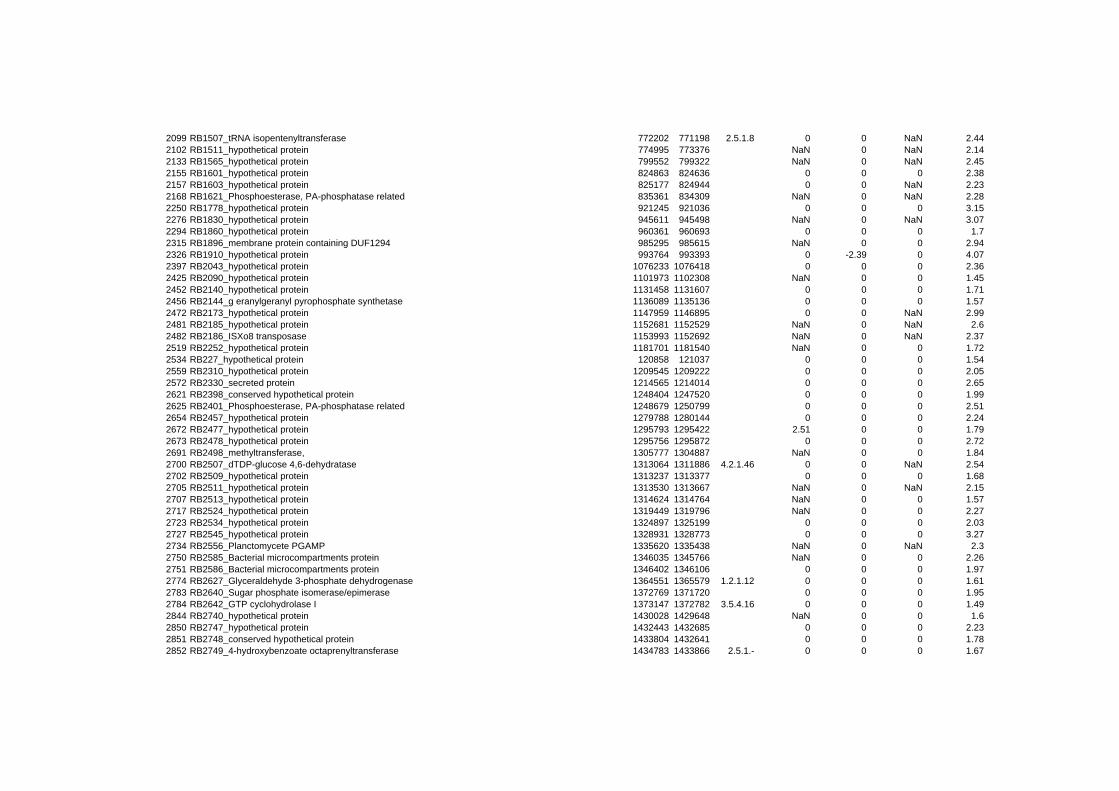
2099 RB1507_tRNA isopentenyltransferase 772202 771198 2.5.1.8 0 0 NaN 2.442102 RB1511_hypothetical protein 774995 773376 NaN 0 NaN 2.142133 RB1565_hypothetical protein 799552 799322 NaN 0 NaN 2.452155 RB1601_hypothetical protein 824863 824636 0 0 0 2.382157 RB1603_hypothetical protein 825177 824944 0 0 NaN 2.232168 RB1621_Phosphoesterase, PA-phosphatase related 835361 834309 NaN 0 NaN 2.282250 RB1778_hypothetical protein 921245 921036 0 0 0 3.152276 RB1830_hypothetical protein 945611 945498 NaN 0 NaN 3.072294 RB1860_hypothetical protein 960361 960693 0 0 0 1.72315 RB1896_membrane protein containing DUF1294 985295 985615 NaN 0 0 2.942326 RB1910_hypothetical protein 993764 993393 0 -2.39 0 4.072397 RB2043_hypothetical protein 1076233 1076418 0 0 0 2.362425 RB2090_hypothetical protein 1101973 1102308 NaN 0 0 1.452452 RB2140_hypothetical protein 1131458 1131607 0 0 0 1.712456 RB2144_g eranylgeranyl pyrophosphate synthetase 1136089 1135136 0 0 0 1.572472 RB2173_hypothetical protein 1147959 1146895 0 0 NaN 2.992481 RB2185_hypothetical protein 1152681 1152529 NaN 0 NaN 2.62482 RB2186_ISXo8 transposase 1153993 1152692 NaN 0 NaN 2.372519 RB2252_hypothetical protein 1181701 1181540 NaN 0 0 1.722534 RB227_hypothetical protein 120858 121037 0 0 0 1.542559 RB2310_hypothetical protein 1209545 1209222 0 0 0 2.052572 RB2330_secreted protein 1214565 1214014 0 0 0 2.652621 RB2398_conserved hypothetical protein 1248404 1247520 0 0 0 1.992625 RB2401_Phosphoesterase, PA-phosphatase related 1248679 1250799 0 0 0 2.512654 RB2457_hypothetical protein 1279788 1280144 0 0 0 2.242672 RB2477_hypothetical protein 1295793 1295422 2.51 0 0 1.792673 RB2478_hypothetical protein 1295756 1295872 0 0 0 2.722691 RB2498_methyltransferase, 1305777 1304887 NaN 0 0 1.842700 RB2507_dTDP-glucose 4,6-dehydratase 1313064 1311886 4.2.1.46 0 0 NaN 2.542702 RB2509_hypothetical protein 1313237 1313377 0 0 0 1.682705 RB2511_hypothetical protein 1313530 1313667 NaN 0 NaN 2.152707 RB2513_hypothetical protein 1314624 1314764 NaN 0 0 1.572717 RB2524_hypothetical protein 1319449 1319796 NaN 0 0 2.272723 RB2534_hypothetical protein 1324897 1325199 0 0 0 2.032727 RB2545_hypothetical protein 1328931 1328773 0 0 0 3.272734 RB2556_Planctomycete PGAMP 1335620 1335438 NaN 0 NaN 2.32750 RB2585_Bacterial microcompartments protein 1346035 1345766 NaN 0 0 2.262751 RB2586_Bacterial microcompartments protein 1346402 1346106 0 0 0 1.972774 RB2627_Glyceraldehyde 3-phosphate dehydrogenase 1364551 1365579 1.2.1.12 0 0 0 1.612783 RB2640_Sugar phosphate isomerase/epimerase 1372769 1371720 0 0 0 1.952784 RB2642_GTP cyclohydrolase I 1373147 1372782 3.5.4.16 0 0 0 1.492844 RB2740_hypothetical protein 1430028 1429648 NaN 0 0 1.62850 RB2747_hypothetical protein 1432443 1432685 0 0 0 2.232851 RB2748_conserved hypothetical protein 1433804 1432641 0 0 0 1.782852 RB2749_4-hydroxybenzoate octaprenyltransferase 1434783 1433866 2.5.1.- 0 0 0 1.67
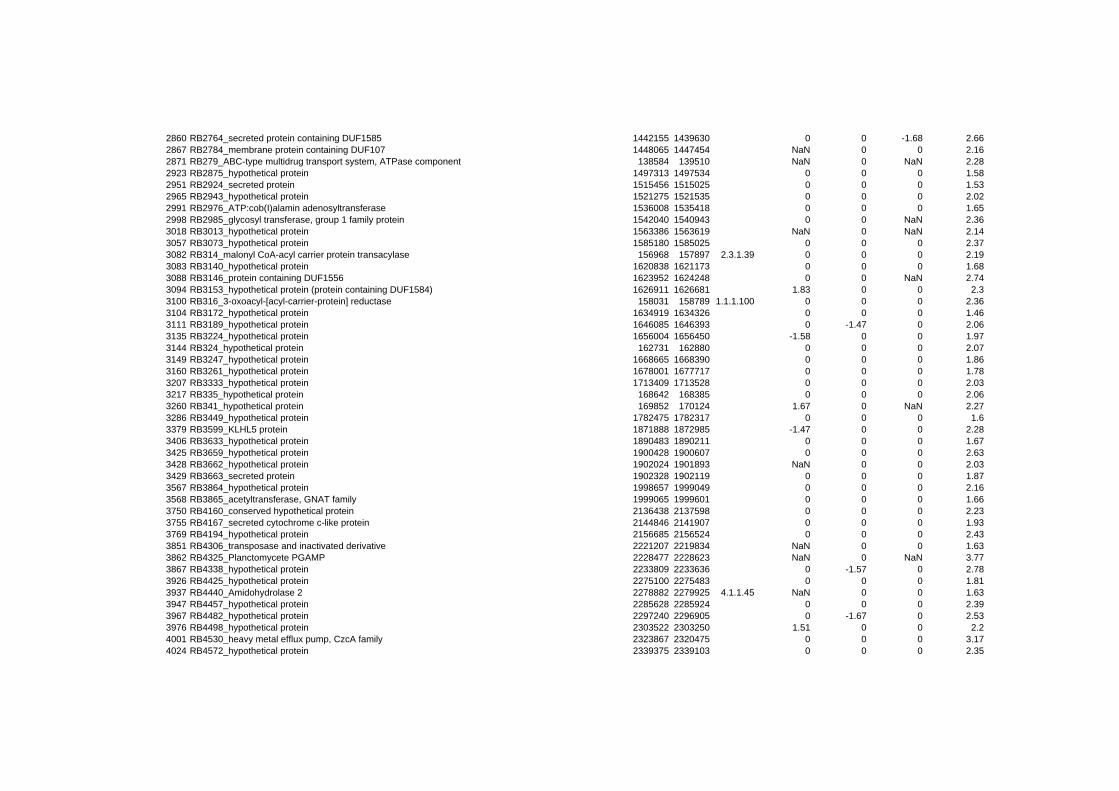
2860 RB2764_secreted protein containing DUF1585 1442155 1439630 0 0 -1.68 2.662867 RB2784_membrane protein containing DUF107 1448065 1447454 NaN 0 0 2.162871 RB279_ABC-type multidrug transport system, ATPase component 138584 139510 NaN 0 NaN 2.282923 RB2875_hypothetical protein 1497313 1497534 0 0 0 1.582951 RB2924_secreted protein 1515456 1515025 0 0 0 1.532965 RB2943_hypothetical protein 1521275 1521535 0 0 0 2.022991 RB2976_ATP:cob(I)alamin adenosyltransferase 1536008 1535418 0 0 0 1.652998 RB2985_glycosyl transferase, group 1 family protein 1542040 1540943 0 0 NaN 2.363018 RB3013_hypothetical protein 1563386 1563619 NaN 0 NaN 2.143057 RB3073_hypothetical protein 1585180 1585025 0 0 0 2.373082 RB314_malonyl CoA-acyl carrier protein transacylase 156968 157897 2.3.1.39 0 0 0 2.193083 RB3140_hypothetical protein 1620838 1621173 0 0 0 1.683088 RB3146_protein containing DUF1556 1623952 1624248 0 0 NaN 2.743094 RB3153_hypothetical protein (protein containing DUF1584) 1626911 1626681 1.83 0 0 2.33100 RB316_3-oxoacyl-[acyl-carrier-protein] reductase 158031 158789 1.1.1.100 0 0 0 2.363104 RB3172_hypothetical protein 1634919 1634326 0 0 0 1.463111 RB3189_hypothetical protein 1646085 1646393 0 -1.47 0 2.063135 RB3224_hypothetical protein 1656004 1656450 -1.58 0 0 1.973144 RB324_hypothetical protein 162731 162880 0 0 0 2.073149 RB3247_hypothetical protein 1668665 1668390 0 0 0 1.863160 RB3261_hypothetical protein 1678001 1677717 0 0 0 1.783207 RB3333_hypothetical protein 1713409 1713528 0 0 0 2.033217 RB335_hypothetical protein 168642 168385 0 0 0 2.063260 RB341_hypothetical protein 169852 170124 1.67 0 NaN 2.273286 RB3449_hypothetical protein 1782475 1782317 0 0 0 1.63379 RB3599_KLHL5 protein 1871888 1872985 -1.47 0 0 2.283406 RB3633_hypothetical protein 1890483 1890211 0 0 0 1.673425 RB3659_hypothetical protein 1900428 1900607 0 0 0 2.633428 RB3662_hypothetical protein 1902024 1901893 NaN 0 0 2.033429 RB3663_secreted protein 1902328 1902119 0 0 0 1.873567 RB3864_hypothetical protein 1998657 1999049 0 0 0 2.163568 RB3865_acetyltransferase, GNAT family 1999065 1999601 0 0 0 1.663750 RB4160_conserved hypothetical protein 2136438 2137598 0 0 0 2.233755 RB4167_secreted cytochrome c-like protein 2144846 2141907 0 0 0 1.933769 RB4194_hypothetical protein 2156685 2156524 0 0 0 2.433851 RB4306_transposase and inactivated derivative 2221207 2219834 NaN 0 0 1.633862 RB4325_Planctomycete PGAMP 2228477 2228623 NaN 0 NaN 3.773867 RB4338_hypothetical protein 2233809 2233636 0 -1.57 0 2.783926 RB4425_hypothetical protein 2275100 2275483 0 0 0 1.813937 RB4440_Amidohydrolase 2 2278882 2279925 4.1.1.45 NaN 0 0 1.633947 RB4457_hypothetical protein 2285628 2285924 0 0 0 2.393967 RB4482_hypothetical protein 2297240 2296905 0 -1.67 0 2.533976 RB4498_hypothetical protein 2303522 2303250 1.51 0 0 2.24001 RB4530_heavy metal efflux pump, CzcA family 2323867 2320475 0 0 0 3.174024 RB4572_hypothetical protein 2339375 2339103 0 0 0 2.35
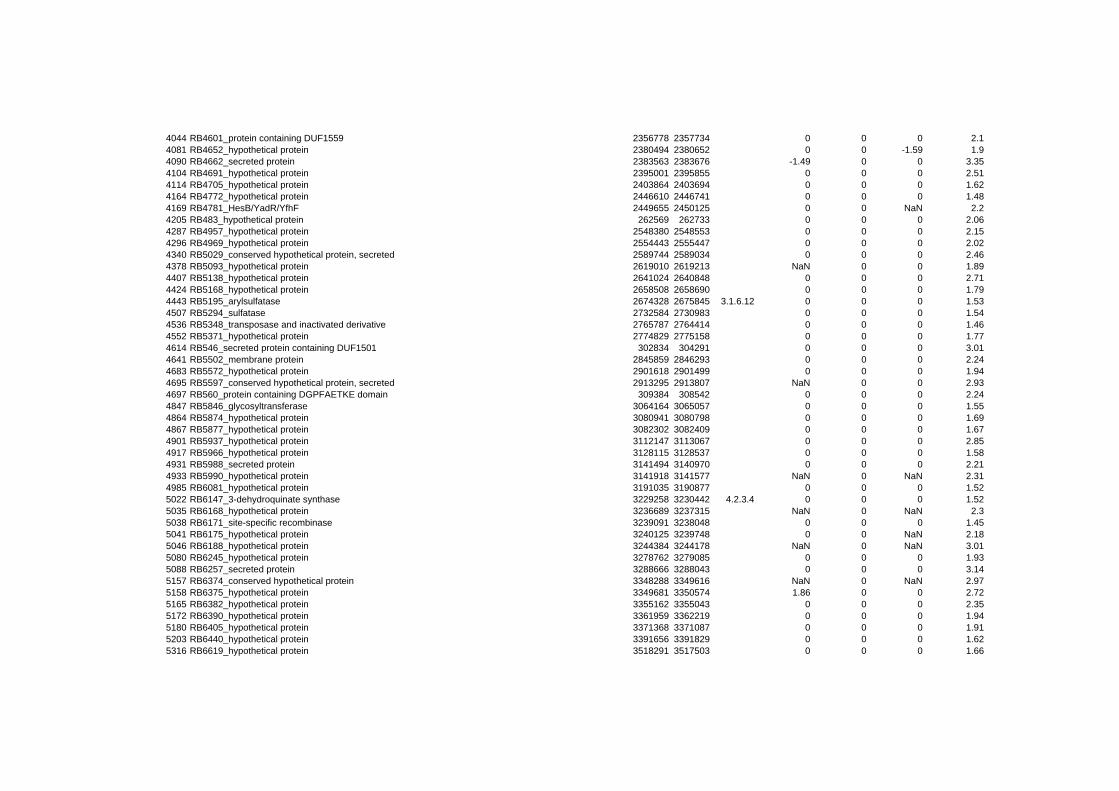
4044 RB4601_protein containing DUF1559 2356778 2357734 0 0 0 2.14081 RB4652_hypothetical protein 2380494 2380652 0 0 -1.59 1.94090 RB4662_secreted protein 2383563 2383676 -1.49 0 0 3.354104 RB4691_hypothetical protein 2395001 2395855 0 0 0 2.514114 RB4705_hypothetical protein 2403864 2403694 0 0 0 1.624164 RB4772_hypothetical protein 2446610 2446741 0 0 0 1.484169 RB4781_HesB/YadR/YfhF 2449655 2450125 0 0 NaN 2.24205 RB483_hypothetical protein 262569 262733 0 0 0 2.064287 RB4957_hypothetical protein 2548380 2548553 0 0 0 2.154296 RB4969_hypothetical protein 2554443 2555447 0 0 0 2.024340 RB5029_conserved hypothetical protein, secreted 2589744 2589034 0 0 0 2.464378 RB5093_hypothetical protein 2619010 2619213 NaN 0 0 1.894407 RB5138_hypothetical protein 2641024 2640848 0 0 0 2.714424 RB5168_hypothetical protein 2658508 2658690 0 0 0 1.794443 RB5195_arylsulfatase 2674328 2675845 3.1.6.12 0 0 0 1.534507 RB5294_sulfatase 2732584 2730983 0 0 0 1.544536 RB5348_transposase and inactivated derivative 2765787 2764414 0 0 0 1.464552 RB5371_hypothetical protein 2774829 2775158 0 0 0 1.774614 RB546_secreted protein containing DUF1501 302834 304291 0 0 0 3.014641 RB5502_membrane protein 2845859 2846293 0 0 0 2.244683 RB5572_hypothetical protein 2901618 2901499 0 0 0 1.944695 RB5597_conserved hypothetical protein, secreted 2913295 2913807 NaN 0 0 2.934697 RB560_protein containing DGPFAETKE domain 309384 308542 0 0 0 2.244847 RB5846_glycosyltransferase 3064164 3065057 0 0 0 1.554864 RB5874_hypothetical protein 3080941 3080798 0 0 0 1.694867 RB5877_hypothetical protein 3082302 3082409 0 0 0 1.674901 RB5937_hypothetical protein 3112147 3113067 0 0 0 2.854917 RB5966_hypothetical protein 3128115 3128537 0 0 0 1.584931 RB5988_secreted protein 3141494 3140970 0 0 0 2.214933 RB5990_hypothetical protein 3141918 3141577 NaN 0 NaN 2.314985 RB6081_hypothetical protein 3191035 3190877 0 0 0 1.525022 RB6147_3-dehydroquinate synthase 3229258 3230442 4.2.3.4 0 0 0 1.525035 RB6168_hypothetical protein 3236689 3237315 NaN 0 NaN 2.35038 RB6171_site-specific recombinase 3239091 3238048 0 0 0 1.455041 RB6175_hypothetical protein 3240125 3239748 0 0 NaN 2.185046 RB6188_hypothetical protein 3244384 3244178 NaN 0 NaN 3.015080 RB6245_hypothetical protein 3278762 3279085 0 0 0 1.935088 RB6257_secreted protein 3288666 3288043 0 0 0 3.145157 RB6374_conserved hypothetical protein 3348288 3349616 NaN 0 NaN 2.975158 RB6375_hypothetical protein 3349681 3350574 1.86 0 0 2.725165 RB6382_hypothetical protein 3355162 3355043 0 0 0 2.355172 RB6390_hypothetical protein 3361959 3362219 0 0 0 1.945180 RB6405_hypothetical protein 3371368 3371087 0 0 0 1.915203 RB6440_hypothetical protein 3391656 3391829 0 0 0 1.625316 RB6619_hypothetical protein 3518291 3517503 0 0 0 1.66
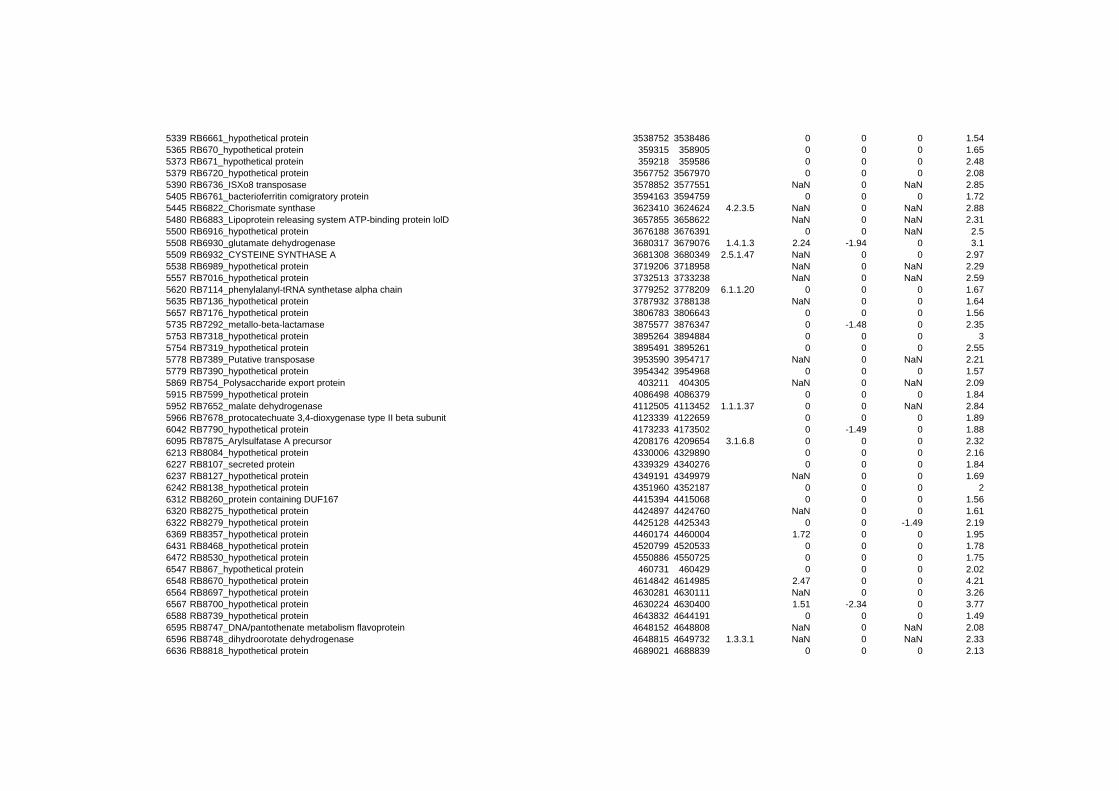
5339 RB6661_hypothetical protein 3538752 3538486 0 0 0 1.545365 RB670_hypothetical protein 359315 358905 0 0 0 1.655373 RB671_hypothetical protein 359218 359586 0 0 0 2.485379 RB6720_hypothetical protein 3567752 3567970 0 0 0 2.085390 RB6736_ISXo8 transposase 3578852 3577551 NaN 0 NaN 2.855405 RB6761_bacterioferritin comigratory protein 3594163 3594759 0 0 0 1.725445 RB6822_Chorismate synthase 3623410 3624624 4.2.3.5 NaN 0 NaN 2.885480 RB6883_Lipoprotein releasing system ATP-binding protein lolD 3657855 3658622 NaN 0 NaN 2.315500 RB6916_hypothetical protein 3676188 3676391 0 0 NaN 2.55508 RB6930_glutamate dehydrogenase 3680317 3679076 1.4.1.3 2.24 -1.94 0 3.15509 RB6932_CYSTEINE SYNTHASE A 3681308 3680349 2.5.1.47 NaN 0 0 2.975538 RB6989_hypothetical protein 3719206 3718958 NaN 0 NaN 2.295557 RB7016_hypothetical protein 3732513 3733238 NaN 0 NaN 2.595620 RB7114_phenylalanyl-tRNA synthetase alpha chain 3779252 3778209 6.1.1.20 0 0 0 1.675635 RB7136_hypothetical protein 3787932 3788138 NaN 0 0 1.645657 RB7176_hypothetical protein 3806783 3806643 0 0 0 1.565735 RB7292_metallo-beta-lactamase 3875577 3876347 0 -1.48 0 2.355753 RB7318_hypothetical protein 3895264 3894884 0 0 0 35754 RB7319_hypothetical protein 3895491 3895261 0 0 0 2.555778 RB7389_Putative transposase 3953590 3954717 NaN 0 NaN 2.215779 RB7390_hypothetical protein 3954342 3954968 0 0 0 1.575869 RB754_Polysaccharide export protein 403211 404305 NaN 0 NaN 2.095915 RB7599_hypothetical protein 4086498 4086379 0 0 0 1.845952 RB7652_malate dehydrogenase 4112505 4113452 1.1.1.37 0 0 NaN 2.845966 RB7678_protocatechuate 3,4-dioxygenase type II beta subunit 4123339 4122659 0 0 0 1.896042 RB7790_hypothetical protein 4173233 4173502 0 -1.49 0 1.886095 RB7875_Arylsulfatase A precursor 4208176 4209654 3.1.6.8 0 0 0 2.326213 RB8084_hypothetical protein 4330006 4329890 0 0 0 2.166227 RB8107_secreted protein 4339329 4340276 0 0 0 1.846237 RB8127_hypothetical protein 4349191 4349979 NaN 0 0 1.696242 RB8138_hypothetical protein 4351960 4352187 0 0 0 26312 RB8260_protein containing DUF167 4415394 4415068 0 0 0 1.566320 RB8275_hypothetical protein 4424897 4424760 NaN 0 0 1.616322 RB8279_hypothetical protein 4425128 4425343 0 0 -1.49 2.196369 RB8357_hypothetical protein 4460174 4460004 1.72 0 0 1.956431 RB8468_hypothetical protein 4520799 4520533 0 0 0 1.786472 RB8530_hypothetical protein 4550886 4550725 0 0 0 1.756547 RB867_hypothetical protein 460731 460429 0 0 0 2.026548 RB8670_hypothetical protein 4614842 4614985 2.47 0 0 4.216564 RB8697_hypothetical protein 4630281 4630111 NaN 0 0 3.266567 RB8700_hypothetical protein 4630224 4630400 1.51 -2.34 0 3.776588 RB8739_hypothetical protein 4643832 4644191 0 0 0 1.496595 RB8747_DNA/pantothenate metabolism flavoprotein 4648152 4648808 NaN 0 NaN 2.086596 RB8748_dihydroorotate dehydrogenase 4648815 4649732 1.3.3.1 NaN 0 NaN 2.336636 RB8818_hypothetical protein 4689021 4688839 0 0 0 2.13

6650 RB8840_acylphosphatases 4701445 4701729 NaN 0 NaN 2.096672 RB8876_hypothetical protein 4723629 4723892 0 0 0 1.636678 RB8884_hypothetical protein 4727666 4727878 0 0 NaN 2.266691 RB8902_hypothetical protein 4740474 4740226 0 0 0 2.36700 RB8917_hypothetical protein 4747435 4747659 0 0 0 2.596731 RB8975_hypothetical protein 4783888 4783556 NaN 0 0 2.66737 RB8983_methanol dehydrogenase regulatory protein 4788960 4790000 0 0 0 1.686763 RB9037_secreted protein 4821537 4822571 NaN 0 0 1.686764 RB9038_hypothetical protein 4823103 4822642 0 0 0 2.746782 RB9078_2-oxoglutarate dehydrogenase E1 component 4846881 4849790 1.2.4.2 NaN 0 0 1.556849 RB9196_secreted protein containing DUF1080 4916635 4917975 0 0 NaN 2.116851 RB9198_hypothetical protein 4918032 4918208 NaN 0 NaN 5.136871 RB9231_hypothetical protein 4937008 4936679 NaN 0 NaN 2.166879 RB924_hypothetical protein 490400 489774 NaN 0 NaN 2.866897 RB9276_membrane protein 4958461 4959162 0 0 0 1.576929 RB933_hypothetical protein 493451 492825 NaN 0 NaN 2.256933 RB934_Putative transposase 494203 493076 NaN 0 NaN 2.176950 RB936_hypothetical protein 494524 494267 0 0 0 1.596953 RB9365_hypothetical protein 5017351 5017809 0 0 0 37157 RB9699_Rhs element Vgr protein 5226252 5228345 0 0 0 1.487159 RB9702_conserved hypothetical protein 5229342 5229632 0 0 NaN 2.667277 RB9908_hypothetical protein 5339497 5339649 NaN 0 NaN 2.157283 RB9918_hypothetical protein 5344369 5344872 NaN 0 NaN 2.757292 RB9936_hypothetical protein 5350464 5350640 0 0 0 1.48
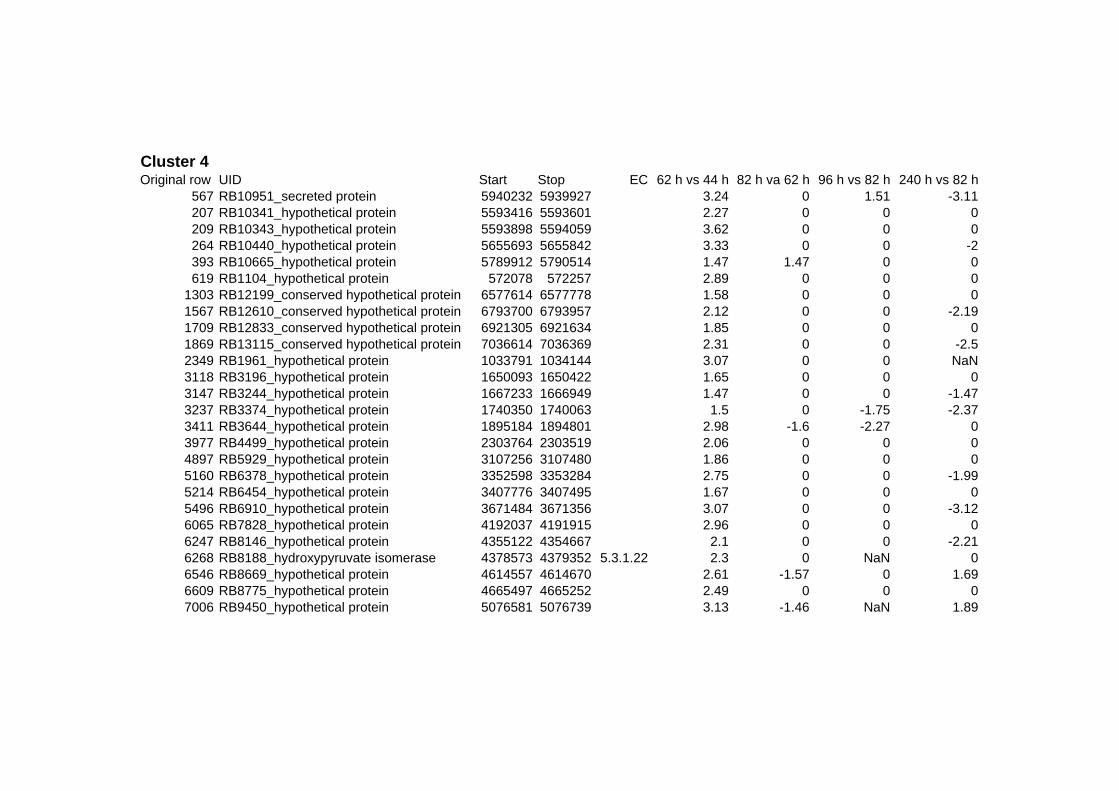
Cluster 4Original row UID Start Stop EC 62 h vs 44 h 82 h va 62 h 96 h vs 82 h 240 h vs 82 h
567 RB10951_secreted protein 5940232 5939927 3.24 0 1.51 -3.11207 RB10341_hypothetical protein 5593416 5593601 2.27 0 0 0209 RB10343_hypothetical protein 5593898 5594059 3.62 0 0 0264 RB10440_hypothetical protein 5655693 5655842 3.33 0 0 -2393 RB10665_hypothetical protein 5789912 5790514 1.47 1.47 0 0619 RB1104_hypothetical protein 572078 572257 2.89 0 0 0
1303 RB12199_conserved hypothetical protein 6577614 6577778 1.58 0 0 01567 RB12610_conserved hypothetical protein 6793700 6793957 2.12 0 0 -2.191709 RB12833_conserved hypothetical protein 6921305 6921634 1.85 0 0 01869 RB13115_conserved hypothetical protein 7036614 7036369 2.31 0 0 -2.52349 RB1961_hypothetical protein 1033791 1034144 3.07 0 0 NaN3118 RB3196_hypothetical protein 1650093 1650422 1.65 0 0 03147 RB3244_hypothetical protein 1667233 1666949 1.47 0 0 -1.473237 RB3374_hypothetical protein 1740350 1740063 1.5 0 -1.75 -2.373411 RB3644_hypothetical protein 1895184 1894801 2.98 -1.6 -2.27 03977 RB4499_hypothetical protein 2303764 2303519 2.06 0 0 04897 RB5929_hypothetical protein 3107256 3107480 1.86 0 0 05160 RB6378_hypothetical protein 3352598 3353284 2.75 0 0 -1.995214 RB6454_hypothetical protein 3407776 3407495 1.67 0 0 05496 RB6910_hypothetical protein 3671484 3671356 3.07 0 0 -3.126065 RB7828_hypothetical protein 4192037 4191915 2.96 0 0 06247 RB8146_hypothetical protein 4355122 4354667 2.1 0 0 -2.216268 RB8188_hydroxypyruvate isomerase 4378573 4379352 5.3.1.22 2.3 0 NaN 06546 RB8669_hypothetical protein 4614557 4614670 2.61 -1.57 0 1.696609 RB8775_hypothetical protein 4665497 4665252 2.49 0 0 07006 RB9450_hypothetical protein 5076581 5076739 3.13 -1.46 NaN 1.89
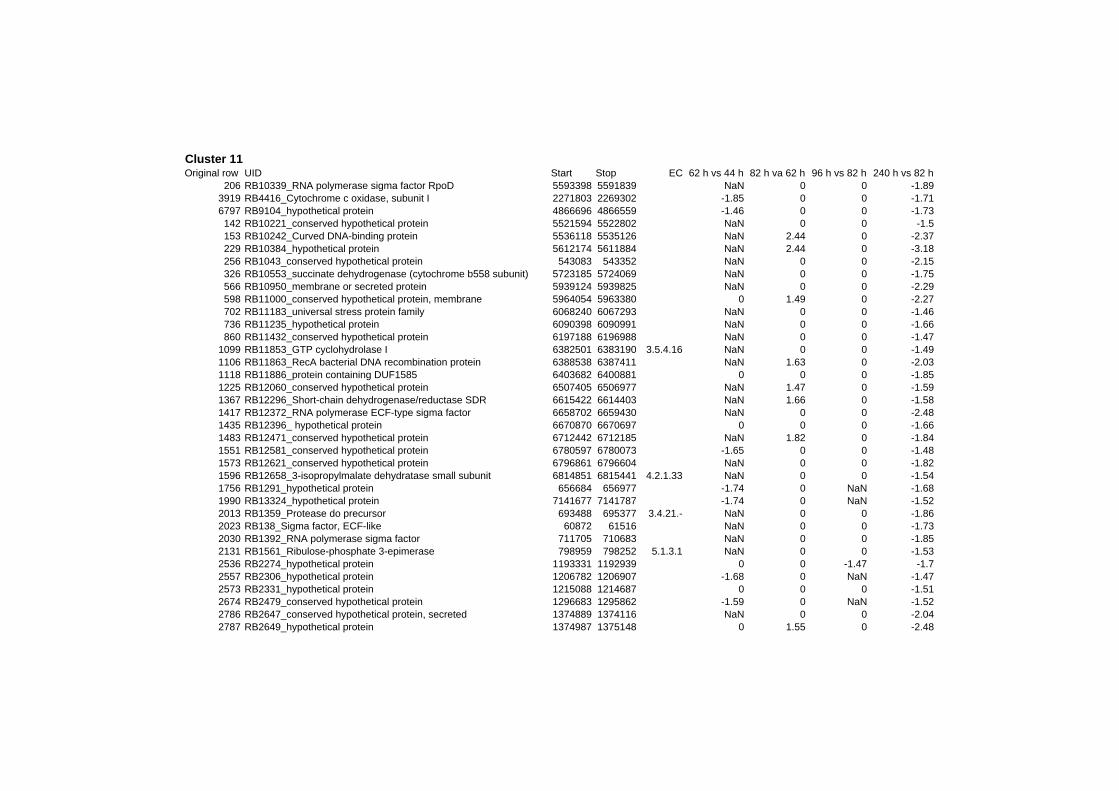
Cluster 11Original row UID Start Stop EC 62 h vs 44 h 82 h va 62 h 96 h vs 82 h 240 h vs 82 h
206 RB10339_RNA polymerase sigma factor RpoD 5593398 5591839 NaN 0 0 -1.893919 RB4416_Cytochrome c oxidase, subunit I 2271803 2269302 -1.85 0 0 -1.716797 RB9104_hypothetical protein 4866696 4866559 -1.46 0 0 -1.73142 RB10221_conserved hypothetical protein 5521594 5522802 NaN 0 0 -1.5153 RB10242_Curved DNA-binding protein 5536118 5535126 NaN 2.44 0 -2.37229 RB10384_hypothetical protein 5612174 5611884 NaN 2.44 0 -3.18256 RB1043_conserved hypothetical protein 543083 543352 NaN 0 0 -2.15326 RB10553_succinate dehydrogenase (cytochrome b558 subunit) 5723185 5724069 NaN 0 0 -1.75566 RB10950_membrane or secreted protein 5939124 5939825 NaN 0 0 -2.29598 RB11000_conserved hypothetical protein, membrane 5964054 5963380 0 1.49 0 -2.27702 RB11183_universal stress protein family 6068240 6067293 NaN 0 0 -1.46736 RB11235_hypothetical protein 6090398 6090991 NaN 0 0 -1.66860 RB11432_conserved hypothetical protein 6197188 6196988 NaN 0 0 -1.47
1099 RB11853_GTP cyclohydrolase I 6382501 6383190 3.5.4.16 NaN 0 0 -1.491106 RB11863_RecA bacterial DNA recombination protein 6388538 6387411 NaN 1.63 0 -2.031118 RB11886_protein containing DUF1585 6403682 6400881 0 0 0 -1.851225 RB12060_conserved hypothetical protein 6507405 6506977 NaN 1.47 0 -1.591367 RB12296_Short-chain dehydrogenase/reductase SDR 6615422 6614403 NaN 1.66 0 -1.581417 RB12372_RNA polymerase ECF-type sigma factor 6658702 6659430 NaN 0 0 -2.481435 RB12396_ hypothetical protein 6670870 6670697 0 0 0 -1.661483 RB12471_conserved hypothetical protein 6712442 6712185 NaN 1.82 0 -1.841551 RB12581_conserved hypothetical protein 6780597 6780073 -1.65 0 0 -1.481573 RB12621_conserved hypothetical protein 6796861 6796604 NaN 0 0 -1.821596 RB12658_3-isopropylmalate dehydratase small subunit 6814851 6815441 4.2.1.33 NaN 0 0 -1.541756 RB1291_hypothetical protein 656684 656977 -1.74 0 NaN -1.681990 RB13324_hypothetical protein 7141677 7141787 -1.74 0 NaN -1.522013 RB1359_Protease do precursor 693488 695377 3.4.21.- NaN 0 0 -1.862023 RB138_Sigma factor, ECF-like 60872 61516 NaN 0 0 -1.732030 RB1392_RNA polymerase sigma factor 711705 710683 NaN 0 0 -1.852131 RB1561_Ribulose-phosphate 3-epimerase 798959 798252 5.1.3.1 NaN 0 0 -1.532536 RB2274_hypothetical protein 1193331 1192939 0 0 -1.47 -1.72557 RB2306_hypothetical protein 1206782 1206907 -1.68 0 NaN -1.472573 RB2331_hypothetical protein 1215088 1214687 0 0 0 -1.512674 RB2479_conserved hypothetical protein 1296683 1295862 -1.59 0 NaN -1.522786 RB2647_conserved hypothetical protein, secreted 1374889 1374116 NaN 0 0 -2.042787 RB2649_hypothetical protein 1374987 1375148 0 1.55 0 -2.48
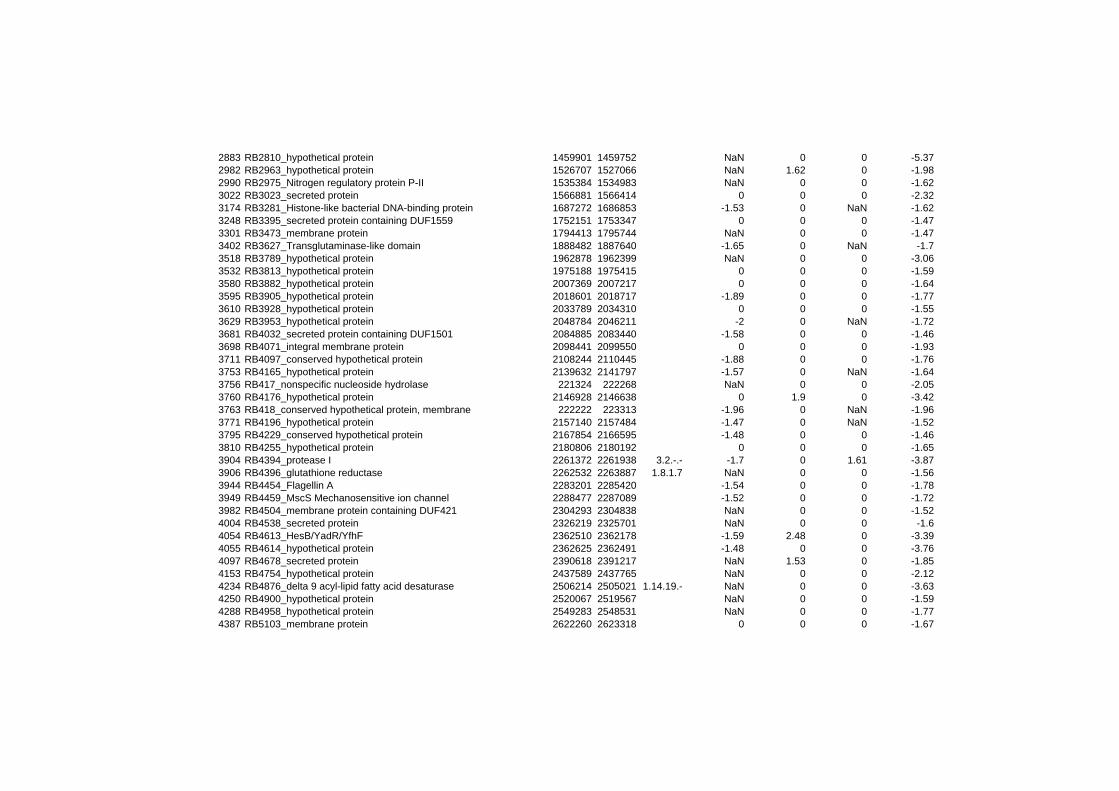
2883 RB2810_hypothetical protein 1459901 1459752 NaN 0 0 -5.372982 RB2963_hypothetical protein 1526707 1527066 NaN 1.62 0 -1.982990 RB2975_Nitrogen regulatory protein P-II 1535384 1534983 NaN 0 0 -1.623022 RB3023_secreted protein 1566881 1566414 0 0 0 -2.323174 RB3281_Histone-like bacterial DNA-binding protein 1687272 1686853 -1.53 0 NaN -1.623248 RB3395_secreted protein containing DUF1559 1752151 1753347 0 0 0 -1.473301 RB3473_membrane protein 1794413 1795744 NaN 0 0 -1.473402 RB3627_Transglutaminase-like domain 1888482 1887640 -1.65 0 NaN -1.73518 RB3789_hypothetical protein 1962878 1962399 NaN 0 0 -3.063532 RB3813_hypothetical protein 1975188 1975415 0 0 0 -1.593580 RB3882_hypothetical protein 2007369 2007217 0 0 0 -1.643595 RB3905_hypothetical protein 2018601 2018717 -1.89 0 0 -1.773610 RB3928_hypothetical protein 2033789 2034310 0 0 0 -1.553629 RB3953_hypothetical protein 2048784 2046211 -2 0 NaN -1.723681 RB4032_secreted protein containing DUF1501 2084885 2083440 -1.58 0 0 -1.463698 RB4071_integral membrane protein 2098441 2099550 0 0 0 -1.933711 RB4097_conserved hypothetical protein 2108244 2110445 -1.88 0 0 -1.763753 RB4165_hypothetical protein 2139632 2141797 -1.57 0 NaN -1.643756 RB417_nonspecific nucleoside hydrolase 221324 222268 NaN 0 0 -2.053760 RB4176_hypothetical protein 2146928 2146638 0 1.9 0 -3.423763 RB418_conserved hypothetical protein, membrane 222222 223313 -1.96 0 NaN -1.963771 RB4196_hypothetical protein 2157140 2157484 -1.47 0 NaN -1.523795 RB4229_conserved hypothetical protein 2167854 2166595 -1.48 0 0 -1.463810 RB4255_hypothetical protein 2180806 2180192 0 0 0 -1.653904 RB4394_protease I 2261372 2261938 3.2.-.- -1.7 0 1.61 -3.873906 RB4396_glutathione reductase 2262532 2263887 1.8.1.7 NaN 0 0 -1.563944 RB4454_Flagellin A 2283201 2285420 -1.54 0 0 -1.783949 RB4459_MscS Mechanosensitive ion channel 2288477 2287089 -1.52 0 0 -1.723982 RB4504_membrane protein containing DUF421 2304293 2304838 NaN 0 0 -1.524004 RB4538_secreted protein 2326219 2325701 NaN 0 0 -1.64054 RB4613_HesB/YadR/YfhF 2362510 2362178 -1.59 2.48 0 -3.394055 RB4614_hypothetical protein 2362625 2362491 -1.48 0 0 -3.764097 RB4678_secreted protein 2390618 2391217 NaN 1.53 0 -1.854153 RB4754_hypothetical protein 2437589 2437765 NaN 0 0 -2.124234 RB4876_delta 9 acyl-lipid fatty acid desaturase 2506214 2505021 1.14.19.- NaN 0 0 -3.634250 RB4900_hypothetical protein 2520067 2519567 NaN 0 0 -1.594288 RB4958_hypothetical protein 2549283 2548531 NaN 0 0 -1.774387 RB5103_membrane protein 2622260 2623318 0 0 0 -1.67
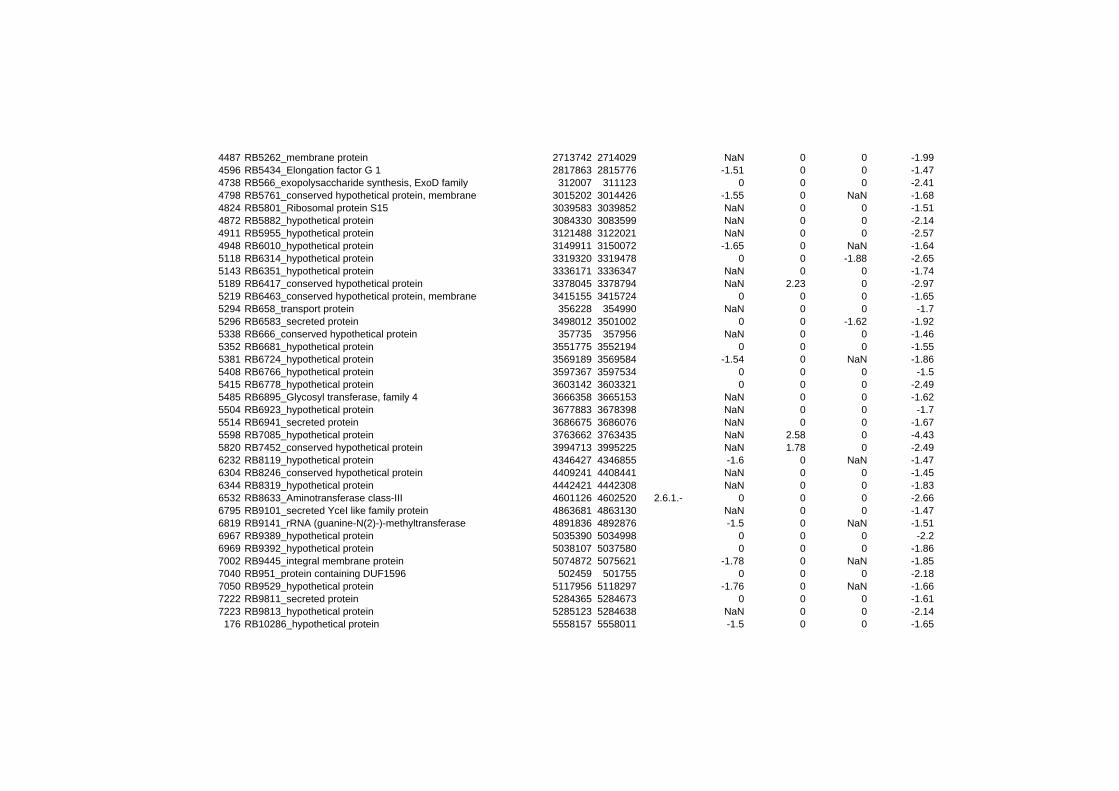
4487 RB5262_membrane protein 2713742 2714029 NaN 0 0 -1.994596 RB5434_Elongation factor G 1 2817863 2815776 -1.51 0 0 -1.474738 RB566_exopolysaccharide synthesis, ExoD family 312007 311123 0 0 0 -2.414798 RB5761_conserved hypothetical protein, membrane 3015202 3014426 -1.55 0 NaN -1.684824 RB5801_Ribosomal protein S15 3039583 3039852 NaN 0 0 -1.514872 RB5882_hypothetical protein 3084330 3083599 NaN 0 0 -2.144911 RB5955_hypothetical protein 3121488 3122021 NaN 0 0 -2.574948 RB6010_hypothetical protein 3149911 3150072 -1.65 0 NaN -1.645118 RB6314_hypothetical protein 3319320 3319478 0 0 -1.88 -2.655143 RB6351_hypothetical protein 3336171 3336347 NaN 0 0 -1.745189 RB6417_conserved hypothetical protein 3378045 3378794 NaN 2.23 0 -2.975219 RB6463_conserved hypothetical protein, membrane 3415155 3415724 0 0 0 -1.655294 RB658_transport protein 356228 354990 NaN 0 0 -1.75296 RB6583_secreted protein 3498012 3501002 0 0 -1.62 -1.925338 RB666_conserved hypothetical protein 357735 357956 NaN 0 0 -1.465352 RB6681_hypothetical protein 3551775 3552194 0 0 0 -1.555381 RB6724_hypothetical protein 3569189 3569584 -1.54 0 NaN -1.865408 RB6766_hypothetical protein 3597367 3597534 0 0 0 -1.55415 RB6778_hypothetical protein 3603142 3603321 0 0 0 -2.495485 RB6895_Glycosyl transferase, family 4 3666358 3665153 NaN 0 0 -1.625504 RB6923_hypothetical protein 3677883 3678398 NaN 0 0 -1.75514 RB6941_secreted protein 3686675 3686076 NaN 0 0 -1.675598 RB7085_hypothetical protein 3763662 3763435 NaN 2.58 0 -4.435820 RB7452_conserved hypothetical protein 3994713 3995225 NaN 1.78 0 -2.496232 RB8119_hypothetical protein 4346427 4346855 -1.6 0 NaN -1.476304 RB8246_conserved hypothetical protein 4409241 4408441 NaN 0 0 -1.456344 RB8319_hypothetical protein 4442421 4442308 NaN 0 0 -1.836532 RB8633_Aminotransferase class-III 4601126 4602520 2.6.1.- 0 0 0 -2.666795 RB9101_secreted YceI like family protein 4863681 4863130 NaN 0 0 -1.476819 RB9141_rRNA (guanine-N(2)-)-methyltransferase 4891836 4892876 -1.5 0 NaN -1.516967 RB9389_hypothetical protein 5035390 5034998 0 0 0 -2.26969 RB9392_hypothetical protein 5038107 5037580 0 0 0 -1.867002 RB9445_integral membrane protein 5074872 5075621 -1.78 0 NaN -1.857040 RB951_protein containing DUF1596 502459 501755 0 0 0 -2.187050 RB9529_hypothetical protein 5117956 5118297 -1.76 0 NaN -1.667222 RB9811_secreted protein 5284365 5284673 0 0 0 -1.617223 RB9813_hypothetical protein 5285123 5284638 NaN 0 0 -2.14176 RB10286_hypothetical protein 5558157 5558011 -1.5 0 0 -1.65

401 RB10678_hypothetical protein 5802466 5802816 -1.73 0 0 -1.577127 RB9651_sialic acid-specific 9-O-acetylesterase 5198277 5196643 3.1.1.53 -2.07 0 NaN -1.6
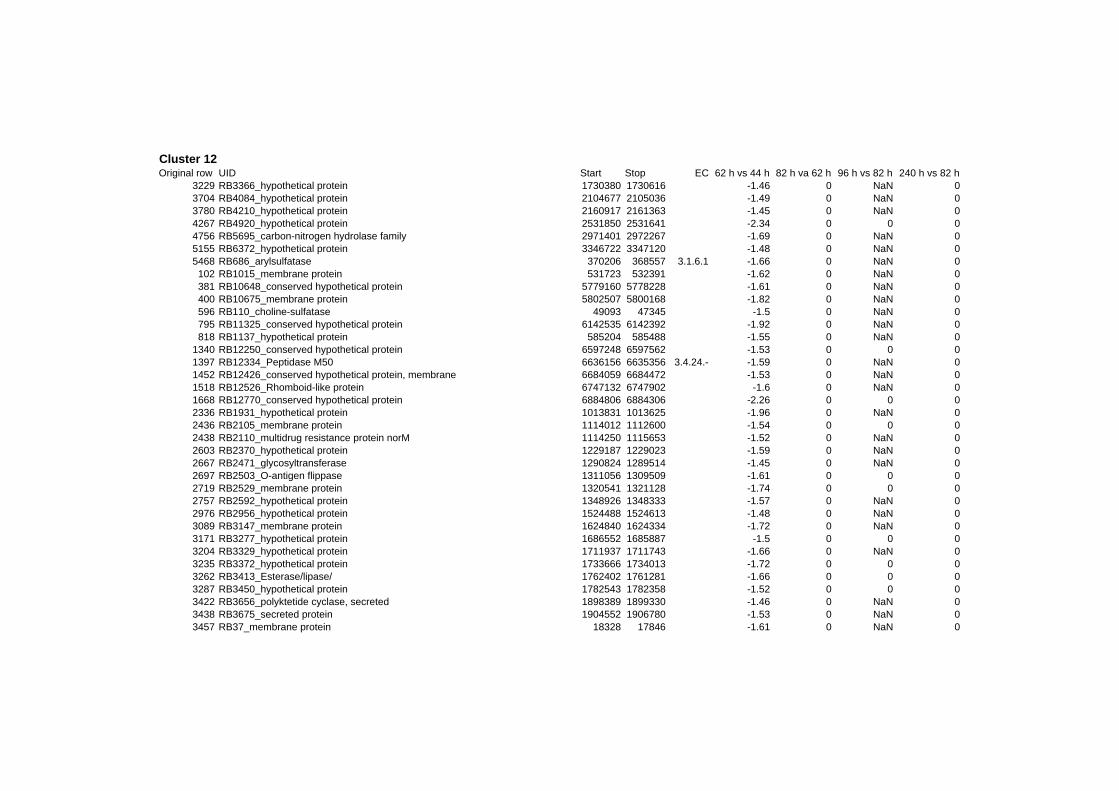
Cluster 12Original row UID Start Stop EC 62 h vs 44 h 82 h va 62 h 96 h vs 82 h 240 h vs 82 h
3229 RB3366_hypothetical protein 1730380 1730616 -1.46 0 NaN 03704 RB4084_hypothetical protein 2104677 2105036 -1.49 0 NaN 03780 RB4210_hypothetical protein 2160917 2161363 -1.45 0 NaN 04267 RB4920_hypothetical protein 2531850 2531641 -2.34 0 0 04756 RB5695_carbon-nitrogen hydrolase family 2971401 2972267 -1.69 0 NaN 05155 RB6372_hypothetical protein 3346722 3347120 -1.48 0 NaN 05468 RB686_arylsulfatase 370206 368557 3.1.6.1 -1.66 0 NaN 0102 RB1015_membrane protein 531723 532391 -1.62 0 NaN 0381 RB10648_conserved hypothetical protein 5779160 5778228 -1.61 0 NaN 0400 RB10675_membrane protein 5802507 5800168 -1.82 0 NaN 0596 RB110_choline-sulfatase 49093 47345 -1.5 0 NaN 0795 RB11325_conserved hypothetical protein 6142535 6142392 -1.92 0 NaN 0818 RB1137_hypothetical protein 585204 585488 -1.55 0 NaN 0
1340 RB12250_conserved hypothetical protein 6597248 6597562 -1.53 0 0 01397 RB12334_Peptidase M50 6636156 6635356 3.4.24.- -1.59 0 NaN 01452 RB12426_conserved hypothetical protein, membrane 6684059 6684472 -1.53 0 NaN 01518 RB12526_Rhomboid-like protein 6747132 6747902 -1.6 0 NaN 01668 RB12770_conserved hypothetical protein 6884806 6884306 -2.26 0 0 02336 RB1931_hypothetical protein 1013831 1013625 -1.96 0 NaN 02436 RB2105_membrane protein 1114012 1112600 -1.54 0 0 02438 RB2110_multidrug resistance protein norM 1114250 1115653 -1.52 0 NaN 02603 RB2370_hypothetical protein 1229187 1229023 -1.59 0 NaN 02667 RB2471_glycosyltransferase 1290824 1289514 -1.45 0 NaN 02697 RB2503_O-antigen flippase 1311056 1309509 -1.61 0 0 02719 RB2529_membrane protein 1320541 1321128 -1.74 0 0 02757 RB2592_hypothetical protein 1348926 1348333 -1.57 0 NaN 02976 RB2956_hypothetical protein 1524488 1524613 -1.48 0 NaN 03089 RB3147_membrane protein 1624840 1624334 -1.72 0 NaN 03171 RB3277_hypothetical protein 1686552 1685887 -1.5 0 0 03204 RB3329_hypothetical protein 1711937 1711743 -1.66 0 NaN 03235 RB3372_hypothetical protein 1733666 1734013 -1.72 0 0 03262 RB3413_Esterase/lipase/ 1762402 1761281 -1.66 0 0 03287 RB3450_hypothetical protein 1782543 1782358 -1.52 0 0 03422 RB3656_polyktetide cyclase, secreted 1898389 1899330 -1.46 0 NaN 03438 RB3675_secreted protein 1904552 1906780 -1.53 0 NaN 03457 RB37_membrane protein 18328 17846 -1.61 0 NaN 0
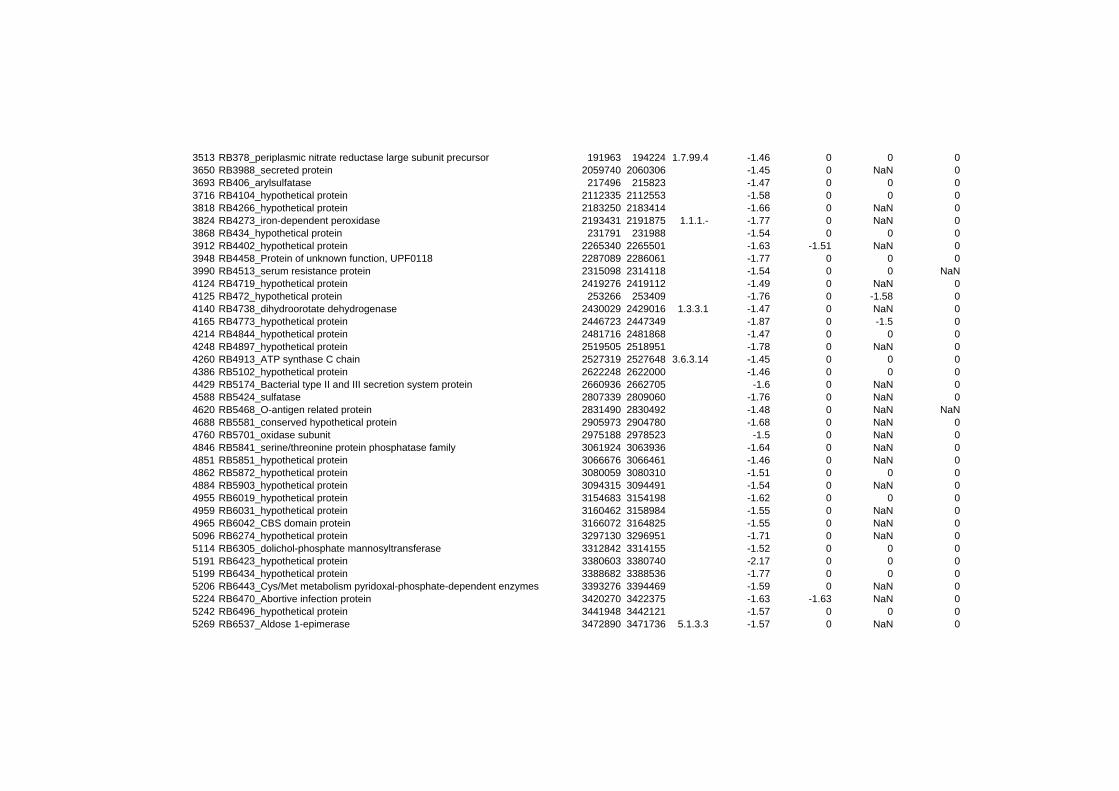
3513 RB378_periplasmic nitrate reductase large subunit precursor 191963 194224 1.7.99.4 -1.46 0 0 03650 RB3988_secreted protein 2059740 2060306 -1.45 0 NaN 03693 RB406_arylsulfatase 217496 215823 -1.47 0 0 03716 RB4104_hypothetical protein 2112335 2112553 -1.58 0 0 03818 RB4266_hypothetical protein 2183250 2183414 -1.66 0 NaN 03824 RB4273_iron-dependent peroxidase 2193431 2191875 1.1.1.- -1.77 0 NaN 03868 RB434_hypothetical protein 231791 231988 -1.54 0 0 03912 RB4402_hypothetical protein 2265340 2265501 -1.63 -1.51 NaN 03948 RB4458_Protein of unknown function, UPF0118 2287089 2286061 -1.77 0 0 03990 RB4513_serum resistance protein 2315098 2314118 -1.54 0 0 NaN4124 RB4719_hypothetical protein 2419276 2419112 -1.49 0 NaN 04125 RB472_hypothetical protein 253266 253409 -1.76 0 -1.58 04140 RB4738_dihydroorotate dehydrogenase 2430029 2429016 1.3.3.1 -1.47 0 NaN 04165 RB4773_hypothetical protein 2446723 2447349 -1.87 0 -1.5 04214 RB4844_hypothetical protein 2481716 2481868 -1.47 0 0 04248 RB4897_hypothetical protein 2519505 2518951 -1.78 0 NaN 04260 RB4913_ATP synthase C chain 2527319 2527648 3.6.3.14 -1.45 0 0 04386 RB5102_hypothetical protein 2622248 2622000 -1.46 0 0 04429 RB5174_Bacterial type II and III secretion system protein 2660936 2662705 -1.6 0 NaN 04588 RB5424_sulfatase 2807339 2809060 -1.76 0 NaN 04620 RB5468_O-antigen related protein 2831490 2830492 -1.48 0 NaN NaN4688 RB5581_conserved hypothetical protein 2905973 2904780 -1.68 0 NaN 04760 RB5701_oxidase subunit 2975188 2978523 -1.5 0 NaN 04846 RB5841_serine/threonine protein phosphatase family 3061924 3063936 -1.64 0 NaN 04851 RB5851_hypothetical protein 3066676 3066461 -1.46 0 NaN 04862 RB5872_hypothetical protein 3080059 3080310 -1.51 0 0 04884 RB5903_hypothetical protein 3094315 3094491 -1.54 0 NaN 04955 RB6019_hypothetical protein 3154683 3154198 -1.62 0 0 04959 RB6031_hypothetical protein 3160462 3158984 -1.55 0 NaN 04965 RB6042_CBS domain protein 3166072 3164825 -1.55 0 NaN 05096 RB6274_hypothetical protein 3297130 3296951 -1.71 0 NaN 05114 RB6305_dolichol-phosphate mannosyltransferase 3312842 3314155 -1.52 0 0 05191 RB6423_hypothetical protein 3380603 3380740 -2.17 0 0 05199 RB6434_hypothetical protein 3388682 3388536 -1.77 0 0 05206 RB6443_Cys/Met metabolism pyridoxal-phosphate-dependent enzymes 3393276 3394469 -1.59 0 NaN 05224 RB6470_Abortive infection protein 3420270 3422375 -1.63 -1.63 NaN 05242 RB6496_hypothetical protein 3441948 3442121 -1.57 0 0 05269 RB6537_Aldose 1-epimerase 3472890 3471736 5.1.3.3 -1.57 0 NaN 0
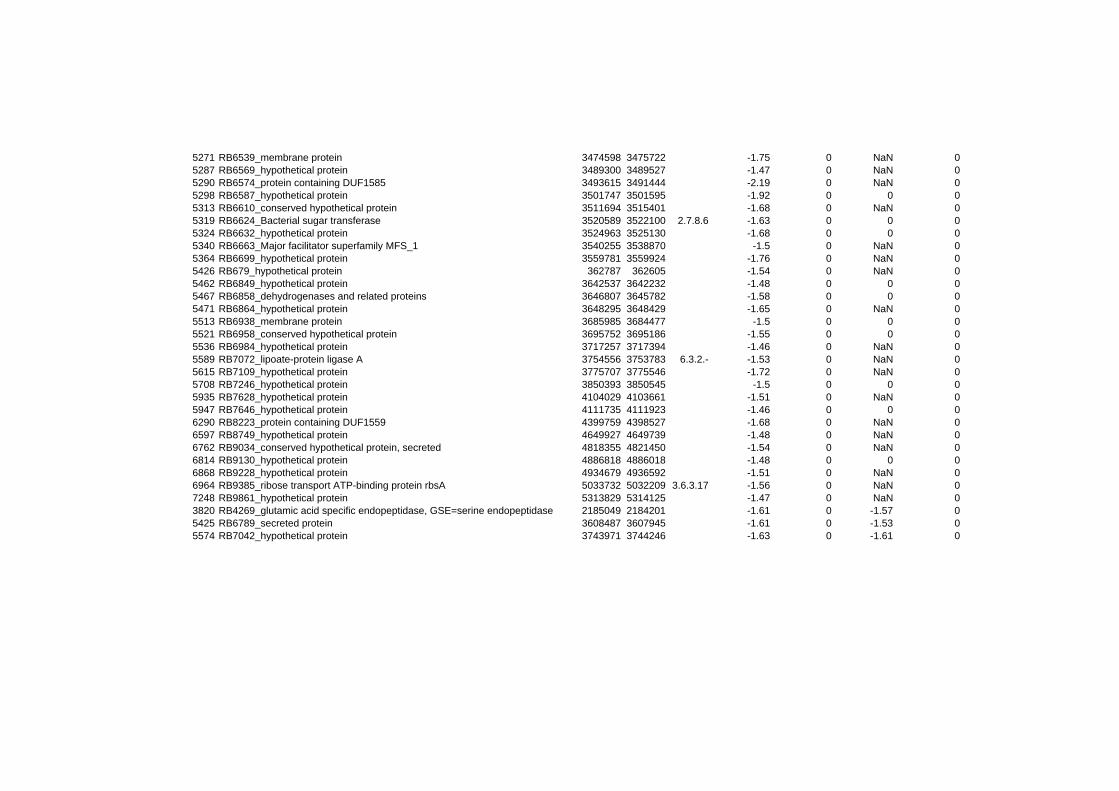
5271 RB6539_membrane protein 3474598 3475722 -1.75 0 NaN 05287 RB6569_hypothetical protein 3489300 3489527 -1.47 0 NaN 05290 RB6574_protein containing DUF1585 3493615 3491444 -2.19 0 NaN 05298 RB6587_hypothetical protein 3501747 3501595 -1.92 0 0 05313 RB6610_conserved hypothetical protein 3511694 3515401 -1.68 0 NaN 05319 RB6624_Bacterial sugar transferase 3520589 3522100 2.7.8.6 -1.63 0 0 05324 RB6632_hypothetical protein 3524963 3525130 -1.68 0 0 05340 RB6663_Major facilitator superfamily MFS_1 3540255 3538870 -1.5 0 NaN 05364 RB6699_hypothetical protein 3559781 3559924 -1.76 0 NaN 05426 RB679_hypothetical protein 362787 362605 -1.54 0 NaN 05462 RB6849_hypothetical protein 3642537 3642232 -1.48 0 0 05467 RB6858_dehydrogenases and related proteins 3646807 3645782 -1.58 0 0 05471 RB6864_hypothetical protein 3648295 3648429 -1.65 0 NaN 05513 RB6938_membrane protein 3685985 3684477 -1.5 0 0 05521 RB6958_conserved hypothetical protein 3695752 3695186 -1.55 0 0 05536 RB6984_hypothetical protein 3717257 3717394 -1.46 0 NaN 05589 RB7072_lipoate-protein ligase A 3754556 3753783 6.3.2.- -1.53 0 NaN 05615 RB7109_hypothetical protein 3775707 3775546 -1.72 0 NaN 05708 RB7246_hypothetical protein 3850393 3850545 -1.5 0 0 05935 RB7628_hypothetical protein 4104029 4103661 -1.51 0 NaN 05947 RB7646_hypothetical protein 4111735 4111923 -1.46 0 0 06290 RB8223_protein containing DUF1559 4399759 4398527 -1.68 0 NaN 06597 RB8749_hypothetical protein 4649927 4649739 -1.48 0 NaN 06762 RB9034_conserved hypothetical protein, secreted 4818355 4821450 -1.54 0 NaN 06814 RB9130_hypothetical protein 4886818 4886018 -1.48 0 0 06868 RB9228_hypothetical protein 4934679 4936592 -1.51 0 NaN 06964 RB9385_ribose transport ATP-binding protein rbsA 5033732 5032209 3.6.3.17 -1.56 0 NaN 07248 RB9861_hypothetical protein 5313829 5314125 -1.47 0 NaN 03820 RB4269_glutamic acid specific endopeptidase, GSE=serine endopeptidase 2185049 2184201 -1.61 0 -1.57 05425 RB6789_secreted protein 3608487 3607945 -1.61 0 -1.53 05574 RB7042_hypothetical protein 3743971 3744246 -1.63 0 -1.61 0
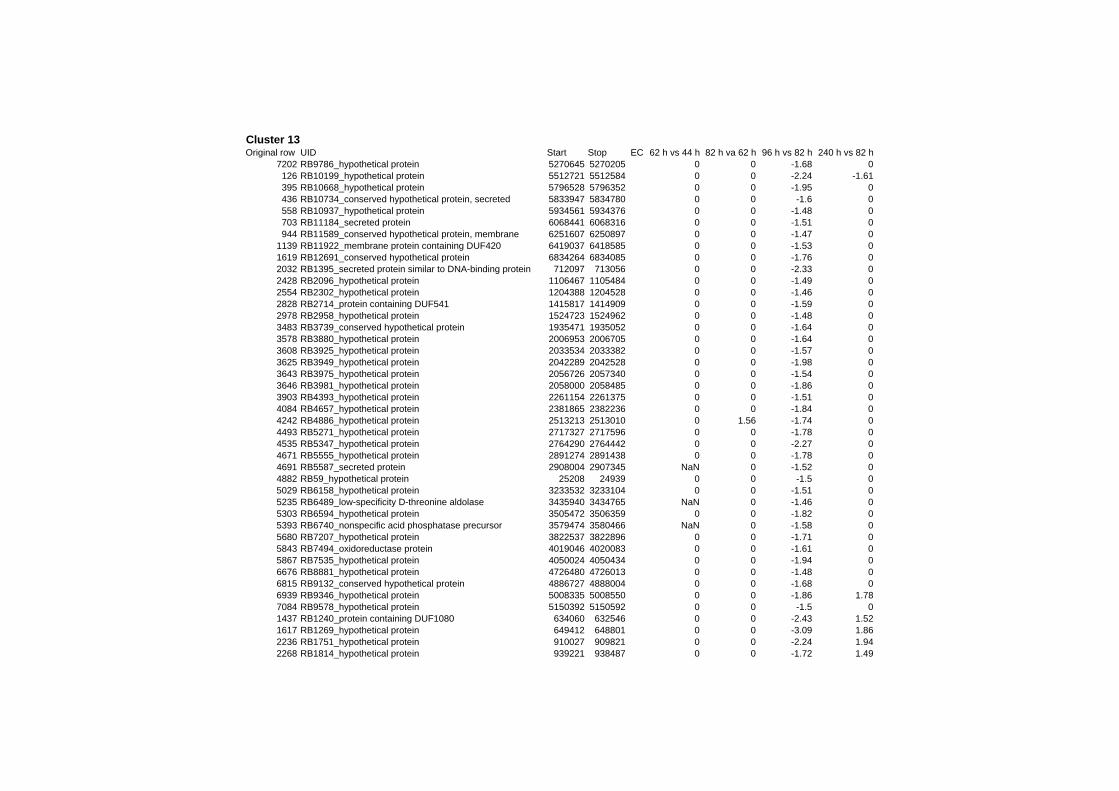
Cluster 13Original row UID Start Stop EC 62 h vs 44 h 82 h va 62 h 96 h vs 82 h 240 h vs 82 h
7202 RB9786_hypothetical protein 5270645 5270205 0 0 -1.68 0126 RB10199_hypothetical protein 5512721 5512584 0 0 -2.24 -1.61395 RB10668_hypothetical protein 5796528 5796352 0 0 -1.95 0436 RB10734_conserved hypothetical protein, secreted 5833947 5834780 0 0 -1.6 0558 RB10937_hypothetical protein 5934561 5934376 0 0 -1.48 0703 RB11184_secreted protein 6068441 6068316 0 0 -1.51 0944 RB11589_conserved hypothetical protein, membrane 6251607 6250897 0 0 -1.47 0
1139 RB11922_membrane protein containing DUF420 6419037 6418585 0 0 -1.53 01619 RB12691_conserved hypothetical protein 6834264 6834085 0 0 -1.76 02032 RB1395_secreted protein similar to DNA-binding protein 712097 713056 0 0 -2.33 02428 RB2096_hypothetical protein 1106467 1105484 0 0 -1.49 02554 RB2302_hypothetical protein 1204388 1204528 0 0 -1.46 02828 RB2714_protein containing DUF541 1415817 1414909 0 0 -1.59 02978 RB2958_hypothetical protein 1524723 1524962 0 0 -1.48 03483 RB3739_conserved hypothetical protein 1935471 1935052 0 0 -1.64 03578 RB3880_hypothetical protein 2006953 2006705 0 0 -1.64 03608 RB3925_hypothetical protein 2033534 2033382 0 0 -1.57 03625 RB3949_hypothetical protein 2042289 2042528 0 0 -1.98 03643 RB3975_hypothetical protein 2056726 2057340 0 0 -1.54 03646 RB3981_hypothetical protein 2058000 2058485 0 0 -1.86 03903 RB4393_hypothetical protein 2261154 2261375 0 0 -1.51 04084 RB4657_hypothetical protein 2381865 2382236 0 0 -1.84 04242 RB4886_hypothetical protein 2513213 2513010 0 1.56 -1.74 04493 RB5271_hypothetical protein 2717327 2717596 0 0 -1.78 04535 RB5347_hypothetical protein 2764290 2764442 0 0 -2.27 04671 RB5555_hypothetical protein 2891274 2891438 0 0 -1.78 04691 RB5587_secreted protein 2908004 2907345 NaN 0 -1.52 04882 RB59_hypothetical protein 25208 24939 0 0 -1.5 05029 RB6158_hypothetical protein 3233532 3233104 0 0 -1.51 05235 RB6489_low-specificity D-threonine aldolase 3435940 3434765 NaN 0 -1.46 05303 RB6594_hypothetical protein 3505472 3506359 0 0 -1.82 05393 RB6740_nonspecific acid phosphatase precursor 3579474 3580466 NaN 0 -1.58 05680 RB7207_hypothetical protein 3822537 3822896 0 0 -1.71 05843 RB7494_oxidoreductase protein 4019046 4020083 0 0 -1.61 05867 RB7535_hypothetical protein 4050024 4050434 0 0 -1.94 06676 RB8881_hypothetical protein 4726480 4726013 0 0 -1.48 06815 RB9132_conserved hypothetical protein 4886727 4888004 0 0 -1.68 06939 RB9346_hypothetical protein 5008335 5008550 0 0 -1.86 1.787084 RB9578_hypothetical protein 5150392 5150592 0 0 -1.5 01437 RB1240_protein containing DUF1080 634060 632546 0 0 -2.43 1.521617 RB1269_hypothetical protein 649412 648801 0 0 -3.09 1.862236 RB1751_hypothetical protein 910027 909821 0 0 -2.24 1.942268 RB1814_hypothetical protein 939221 938487 0 0 -1.72 1.49
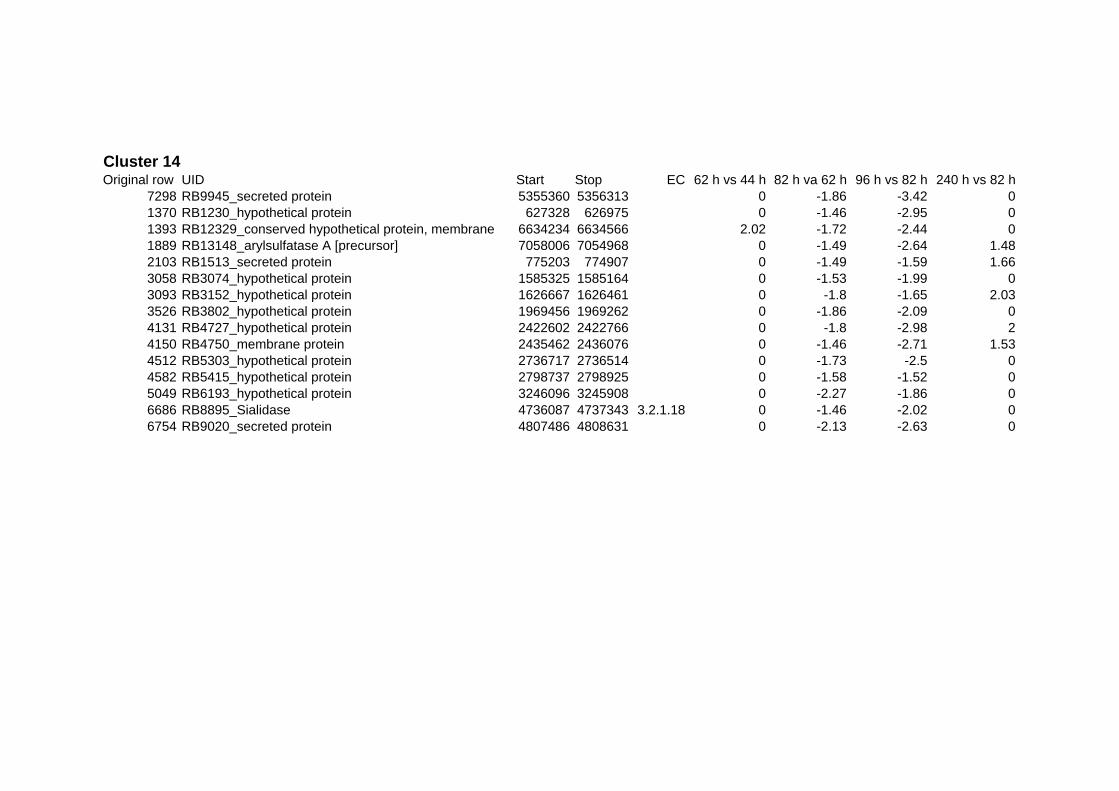
Cluster 14Original row UID Start Stop EC 62 h vs 44 h 82 h va 62 h 96 h vs 82 h 240 h vs 82 h
7298 RB9945_secreted protein 5355360 5356313 0 -1.86 -3.42 01370 RB1230_hypothetical protein 627328 626975 0 -1.46 -2.95 01393 RB12329_conserved hypothetical protein, membrane 6634234 6634566 2.02 -1.72 -2.44 01889 RB13148_arylsulfatase A [precursor] 7058006 7054968 0 -1.49 -2.64 1.482103 RB1513_secreted protein 775203 774907 0 -1.49 -1.59 1.663058 RB3074_hypothetical protein 1585325 1585164 0 -1.53 -1.99 03093 RB3152_hypothetical protein 1626667 1626461 0 -1.8 -1.65 2.033526 RB3802_hypothetical protein 1969456 1969262 0 -1.86 -2.09 04131 RB4727_hypothetical protein 2422602 2422766 0 -1.8 -2.98 24150 RB4750_membrane protein 2435462 2436076 0 -1.46 -2.71 1.534512 RB5303_hypothetical protein 2736717 2736514 0 -1.73 -2.5 04582 RB5415_hypothetical protein 2798737 2798925 0 -1.58 -1.52 05049 RB6193_hypothetical protein 3246096 3245908 0 -2.27 -1.86 06686 RB8895_Sialidase 4736087 4737343 3.2.1.18 0 -1.46 -2.02 06754 RB9020_secreted protein 4807486 4808631 0 -2.13 -2.63 0
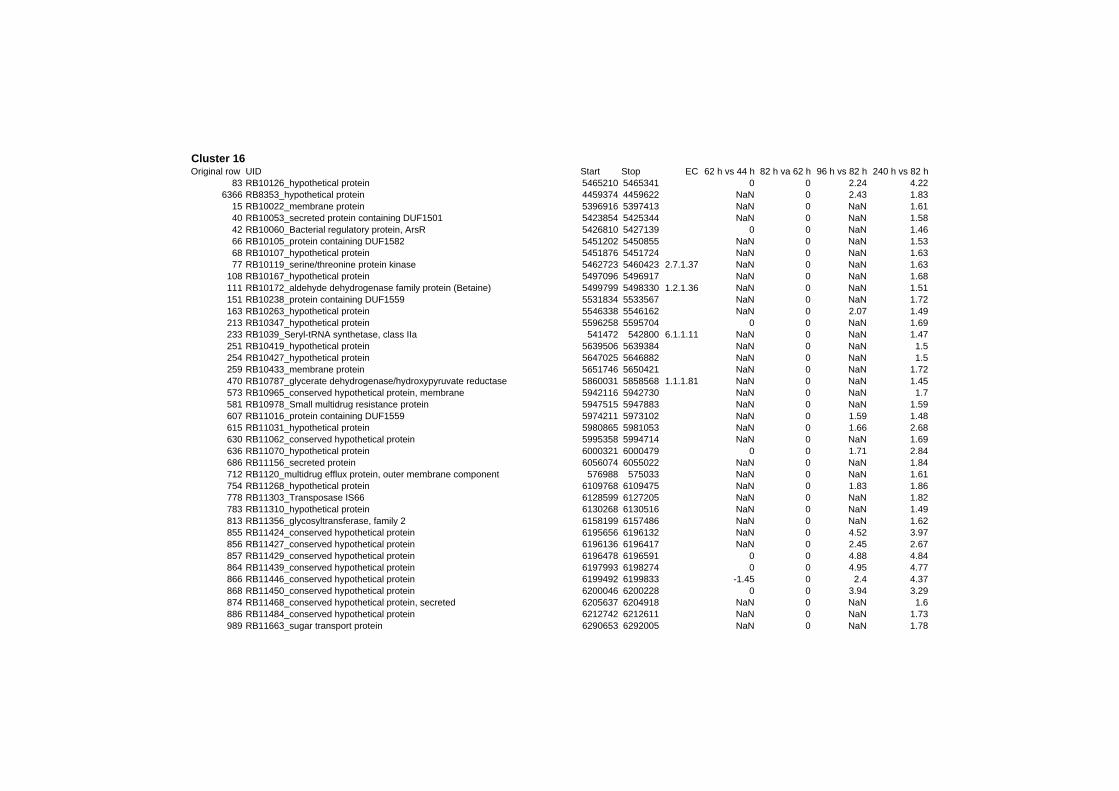
Cluster 16Original row UID Start Stop EC 62 h vs 44 h 82 h va 62 h 96 h vs 82 h 240 h vs 82 h
83 RB10126_hypothetical protein 5465210 5465341 0 0 2.24 4.226366 RB8353_hypothetical protein 4459374 4459622 NaN 0 2.43 1.83
15 RB10022_membrane protein 5396916 5397413 NaN 0 NaN 1.6140 RB10053_secreted protein containing DUF1501 5423854 5425344 NaN 0 NaN 1.5842 RB10060_Bacterial regulatory protein, ArsR 5426810 5427139 0 0 NaN 1.4666 RB10105_protein containing DUF1582 5451202 5450855 NaN 0 NaN 1.5368 RB10107_hypothetical protein 5451876 5451724 NaN 0 NaN 1.6377 RB10119_serine/threonine protein kinase 5462723 5460423 2.7.1.37 NaN 0 NaN 1.63
108 RB10167_hypothetical protein 5497096 5496917 NaN 0 NaN 1.68111 RB10172_aldehyde dehydrogenase family protein (Betaine) 5499799 5498330 1.2.1.36 NaN 0 NaN 1.51151 RB10238_protein containing DUF1559 5531834 5533567 NaN 0 NaN 1.72163 RB10263_hypothetical protein 5546338 5546162 NaN 0 2.07 1.49213 RB10347_hypothetical protein 5596258 5595704 0 0 NaN 1.69233 RB1039_Seryl-tRNA synthetase, class IIa 541472 542800 6.1.1.11 NaN 0 NaN 1.47251 RB10419_hypothetical protein 5639506 5639384 NaN 0 NaN 1.5254 RB10427_hypothetical protein 5647025 5646882 NaN 0 NaN 1.5259 RB10433_membrane protein 5651746 5650421 NaN 0 NaN 1.72470 RB10787_glycerate dehydrogenase/hydroxypyruvate reductase 5860031 5858568 1.1.1.81 NaN 0 NaN 1.45573 RB10965_conserved hypothetical protein, membrane 5942116 5942730 NaN 0 NaN 1.7581 RB10978_Small multidrug resistance protein 5947515 5947883 NaN 0 NaN 1.59607 RB11016_protein containing DUF1559 5974211 5973102 NaN 0 1.59 1.48615 RB11031_hypothetical protein 5980865 5981053 NaN 0 1.66 2.68630 RB11062_conserved hypothetical protein 5995358 5994714 NaN 0 NaN 1.69636 RB11070_hypothetical protein 6000321 6000479 0 0 1.71 2.84686 RB11156_secreted protein 6056074 6055022 NaN 0 NaN 1.84712 RB1120_multidrug efflux protein, outer membrane component 576988 575033 NaN 0 NaN 1.61754 RB11268_hypothetical protein 6109768 6109475 NaN 0 1.83 1.86778 RB11303_Transposase IS66 6128599 6127205 NaN 0 NaN 1.82783 RB11310_hypothetical protein 6130268 6130516 NaN 0 NaN 1.49813 RB11356_glycosyltransferase, family 2 6158199 6157486 NaN 0 NaN 1.62855 RB11424_conserved hypothetical protein 6195656 6196132 NaN 0 4.52 3.97856 RB11427_conserved hypothetical protein 6196136 6196417 NaN 0 2.45 2.67857 RB11429_conserved hypothetical protein 6196478 6196591 0 0 4.88 4.84864 RB11439_conserved hypothetical protein 6197993 6198274 0 0 4.95 4.77866 RB11446_conserved hypothetical protein 6199492 6199833 -1.45 0 2.4 4.37868 RB11450_conserved hypothetical protein 6200046 6200228 0 0 3.94 3.29874 RB11468_conserved hypothetical protein, secreted 6205637 6204918 NaN 0 NaN 1.6886 RB11484_conserved hypothetical protein 6212742 6212611 NaN 0 NaN 1.73989 RB11663_sugar transport protein 6290653 6292005 NaN 0 NaN 1.78
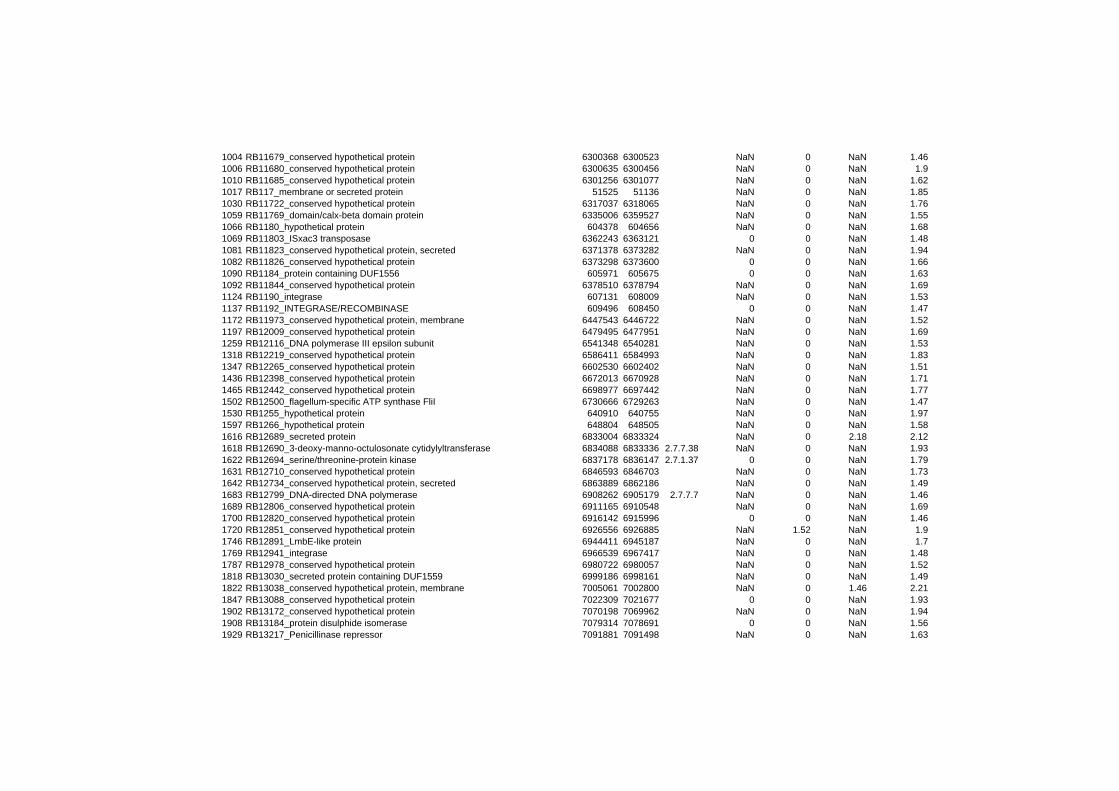
1004 RB11679_conserved hypothetical protein 6300368 6300523 NaN 0 NaN 1.461006 RB11680_conserved hypothetical protein 6300635 6300456 NaN 0 NaN 1.91010 RB11685_conserved hypothetical protein 6301256 6301077 NaN 0 NaN 1.621017 RB117_membrane or secreted protein 51525 51136 NaN 0 NaN 1.851030 RB11722_conserved hypothetical protein 6317037 6318065 NaN 0 NaN 1.761059 RB11769_domain/calx-beta domain protein 6335006 6359527 NaN 0 NaN 1.551066 RB1180_hypothetical protein 604378 604656 NaN 0 NaN 1.681069 RB11803_ISxac3 transposase 6362243 6363121 0 0 NaN 1.481081 RB11823_conserved hypothetical protein, secreted 6371378 6373282 NaN 0 NaN 1.941082 RB11826_conserved hypothetical protein 6373298 6373600 0 0 NaN 1.661090 RB1184_protein containing DUF1556 605971 605675 0 0 NaN 1.631092 RB11844_conserved hypothetical protein 6378510 6378794 NaN 0 NaN 1.691124 RB1190_integrase 607131 608009 NaN 0 NaN 1.531137 RB1192_INTEGRASE/RECOMBINASE 609496 608450 0 0 NaN 1.471172 RB11973_conserved hypothetical protein, membrane 6447543 6446722 NaN 0 NaN 1.521197 RB12009_conserved hypothetical protein 6479495 6477951 NaN 0 NaN 1.691259 RB12116_DNA polymerase III epsilon subunit 6541348 6540281 NaN 0 NaN 1.531318 RB12219_conserved hypothetical protein 6586411 6584993 NaN 0 NaN 1.831347 RB12265_conserved hypothetical protein 6602530 6602402 NaN 0 NaN 1.511436 RB12398_conserved hypothetical protein 6672013 6670928 NaN 0 NaN 1.711465 RB12442_conserved hypothetical protein 6698977 6697442 NaN 0 NaN 1.771502 RB12500_flagellum-specific ATP synthase FliI 6730666 6729263 NaN 0 NaN 1.471530 RB1255_hypothetical protein 640910 640755 NaN 0 NaN 1.971597 RB1266_hypothetical protein 648804 648505 NaN 0 NaN 1.581616 RB12689_secreted protein 6833004 6833324 NaN 0 2.18 2.121618 RB12690_3-deoxy-manno-octulosonate cytidylyltransferase 6834088 6833336 2.7.7.38 NaN 0 NaN 1.931622 RB12694_serine/threonine-protein kinase 6837178 6836147 2.7.1.37 0 0 NaN 1.791631 RB12710_conserved hypothetical protein 6846593 6846703 NaN 0 NaN 1.731642 RB12734_conserved hypothetical protein, secreted 6863889 6862186 NaN 0 NaN 1.491683 RB12799_DNA-directed DNA polymerase 6908262 6905179 2.7.7.7 NaN 0 NaN 1.461689 RB12806_conserved hypothetical protein 6911165 6910548 NaN 0 NaN 1.691700 RB12820_conserved hypothetical protein 6916142 6915996 0 0 NaN 1.461720 RB12851_conserved hypothetical protein 6926556 6926885 NaN 1.52 NaN 1.91746 RB12891_LmbE-like protein 6944411 6945187 NaN 0 NaN 1.71769 RB12941_integrase 6966539 6967417 NaN 0 NaN 1.481787 RB12978_conserved hypothetical protein 6980722 6980057 NaN 0 NaN 1.521818 RB13030_secreted protein containing DUF1559 6999186 6998161 NaN 0 NaN 1.491822 RB13038_conserved hypothetical protein, membrane 7005061 7002800 NaN 0 1.46 2.211847 RB13088_conserved hypothetical protein 7022309 7021677 0 0 NaN 1.931902 RB13172_conserved hypothetical protein 7070198 7069962 NaN 0 NaN 1.941908 RB13184_protein disulphide isomerase 7079314 7078691 0 0 NaN 1.561929 RB13217_Penicillinase repressor 7091881 7091498 NaN 0 NaN 1.63
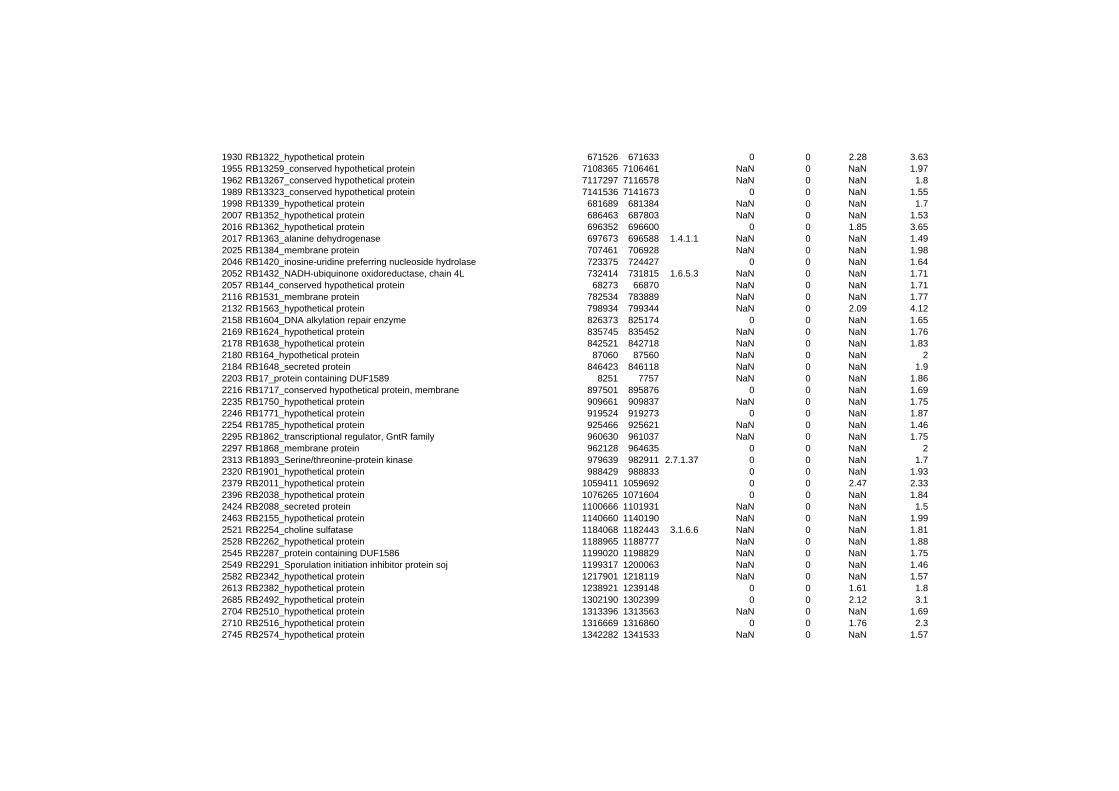
1930 RB1322_hypothetical protein 671526 671633 0 0 2.28 3.631955 RB13259_conserved hypothetical protein 7108365 7106461 NaN 0 NaN 1.971962 RB13267_conserved hypothetical protein 7117297 7116578 NaN 0 NaN 1.81989 RB13323_conserved hypothetical protein 7141536 7141673 0 0 NaN 1.551998 RB1339_hypothetical protein 681689 681384 NaN 0 NaN 1.72007 RB1352_hypothetical protein 686463 687803 NaN 0 NaN 1.532016 RB1362_hypothetical protein 696352 696600 0 0 1.85 3.652017 RB1363_alanine dehydrogenase 697673 696588 1.4.1.1 NaN 0 NaN 1.492025 RB1384_membrane protein 707461 706928 NaN 0 NaN 1.982046 RB1420_inosine-uridine preferring nucleoside hydrolase 723375 724427 0 0 NaN 1.642052 RB1432_NADH-ubiquinone oxidoreductase, chain 4L 732414 731815 1.6.5.3 NaN 0 NaN 1.712057 RB144_conserved hypothetical protein 68273 66870 NaN 0 NaN 1.712116 RB1531_membrane protein 782534 783889 NaN 0 NaN 1.772132 RB1563_hypothetical protein 798934 799344 NaN 0 2.09 4.122158 RB1604_DNA alkylation repair enzyme 826373 825174 0 0 NaN 1.652169 RB1624_hypothetical protein 835745 835452 NaN 0 NaN 1.762178 RB1638_hypothetical protein 842521 842718 NaN 0 NaN 1.832180 RB164_hypothetical protein 87060 87560 NaN 0 NaN 22184 RB1648_secreted protein 846423 846118 NaN 0 NaN 1.92203 RB17_protein containing DUF1589 8251 7757 NaN 0 NaN 1.862216 RB1717_conserved hypothetical protein, membrane 897501 895876 0 0 NaN 1.692235 RB1750_hypothetical protein 909661 909837 NaN 0 NaN 1.752246 RB1771_hypothetical protein 919524 919273 0 0 NaN 1.872254 RB1785_hypothetical protein 925466 925621 NaN 0 NaN 1.462295 RB1862_transcriptional regulator, GntR family 960630 961037 NaN 0 NaN 1.752297 RB1868_membrane protein 962128 964635 0 0 NaN 22313 RB1893_Serine/threonine-protein kinase 979639 982911 2.7.1.37 0 0 NaN 1.72320 RB1901_hypothetical protein 988429 988833 0 0 NaN 1.932379 RB2011_hypothetical protein 1059411 1059692 0 0 2.47 2.332396 RB2038_hypothetical protein 1076265 1071604 0 0 NaN 1.842424 RB2088_secreted protein 1100666 1101931 NaN 0 NaN 1.52463 RB2155_hypothetical protein 1140660 1140190 NaN 0 NaN 1.992521 RB2254_choline sulfatase 1184068 1182443 3.1.6.6 NaN 0 NaN 1.812528 RB2262_hypothetical protein 1188965 1188777 NaN 0 NaN 1.882545 RB2287_protein containing DUF1586 1199020 1198829 NaN 0 NaN 1.752549 RB2291_Sporulation initiation inhibitor protein soj 1199317 1200063 NaN 0 NaN 1.462582 RB2342_hypothetical protein 1217901 1218119 NaN 0 NaN 1.572613 RB2382_hypothetical protein 1238921 1239148 0 0 1.61 1.82685 RB2492_hypothetical protein 1302190 1302399 0 0 2.12 3.12704 RB2510_hypothetical protein 1313396 1313563 NaN 0 NaN 1.692710 RB2516_hypothetical protein 1316669 1316860 0 0 1.76 2.32745 RB2574_hypothetical protein 1342282 1341533 NaN 0 NaN 1.57
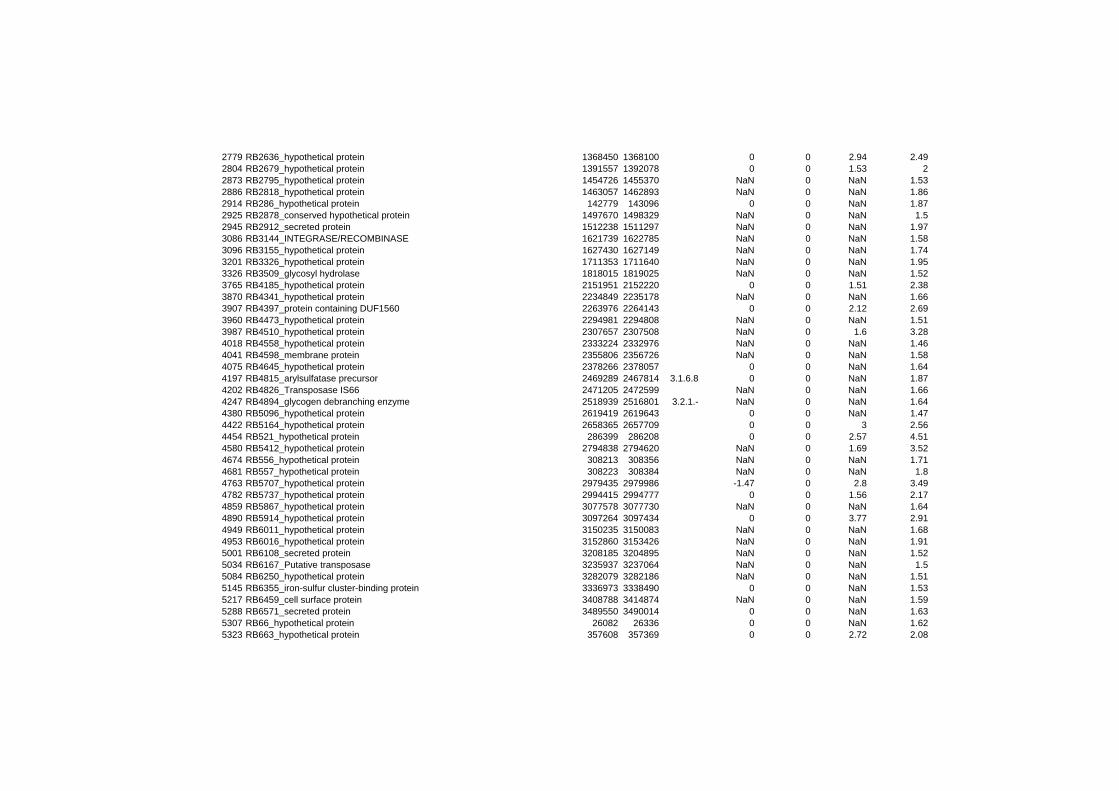
2779 RB2636_hypothetical protein 1368450 1368100 0 0 2.94 2.492804 RB2679_hypothetical protein 1391557 1392078 0 0 1.53 22873 RB2795_hypothetical protein 1454726 1455370 NaN 0 NaN 1.532886 RB2818_hypothetical protein 1463057 1462893 NaN 0 NaN 1.862914 RB286_hypothetical protein 142779 143096 0 0 NaN 1.872925 RB2878_conserved hypothetical protein 1497670 1498329 NaN 0 NaN 1.52945 RB2912_secreted protein 1512238 1511297 NaN 0 NaN 1.973086 RB3144_INTEGRASE/RECOMBINASE 1621739 1622785 NaN 0 NaN 1.583096 RB3155_hypothetical protein 1627430 1627149 NaN 0 NaN 1.743201 RB3326_hypothetical protein 1711353 1711640 NaN 0 NaN 1.953326 RB3509_glycosyl hydrolase 1818015 1819025 NaN 0 NaN 1.523765 RB4185_hypothetical protein 2151951 2152220 0 0 1.51 2.383870 RB4341_hypothetical protein 2234849 2235178 NaN 0 NaN 1.663907 RB4397_protein containing DUF1560 2263976 2264143 0 0 2.12 2.693960 RB4473_hypothetical protein 2294981 2294808 NaN 0 NaN 1.513987 RB4510_hypothetical protein 2307657 2307508 NaN 0 1.6 3.284018 RB4558_hypothetical protein 2333224 2332976 NaN 0 NaN 1.464041 RB4598_membrane protein 2355806 2356726 NaN 0 NaN 1.584075 RB4645_hypothetical protein 2378266 2378057 0 0 NaN 1.644197 RB4815_arylsulfatase precursor 2469289 2467814 3.1.6.8 0 0 NaN 1.874202 RB4826_Transposase IS66 2471205 2472599 NaN 0 NaN 1.664247 RB4894_glycogen debranching enzyme 2518939 2516801 3.2.1.- NaN 0 NaN 1.644380 RB5096_hypothetical protein 2619419 2619643 0 0 NaN 1.474422 RB5164_hypothetical protein 2658365 2657709 0 0 3 2.564454 RB521_hypothetical protein 286399 286208 0 0 2.57 4.514580 RB5412_hypothetical protein 2794838 2794620 NaN 0 1.69 3.524674 RB556_hypothetical protein 308213 308356 NaN 0 NaN 1.714681 RB557_hypothetical protein 308223 308384 NaN 0 NaN 1.84763 RB5707_hypothetical protein 2979435 2979986 -1.47 0 2.8 3.494782 RB5737_hypothetical protein 2994415 2994777 0 0 1.56 2.174859 RB5867_hypothetical protein 3077578 3077730 NaN 0 NaN 1.644890 RB5914_hypothetical protein 3097264 3097434 0 0 3.77 2.914949 RB6011_hypothetical protein 3150235 3150083 NaN 0 NaN 1.684953 RB6016_hypothetical protein 3152860 3153426 NaN 0 NaN 1.915001 RB6108_secreted protein 3208185 3204895 NaN 0 NaN 1.525034 RB6167_Putative transposase 3235937 3237064 NaN 0 NaN 1.55084 RB6250_hypothetical protein 3282079 3282186 NaN 0 NaN 1.515145 RB6355_iron-sulfur cluster-binding protein 3336973 3338490 0 0 NaN 1.535217 RB6459_cell surface protein 3408788 3414874 NaN 0 NaN 1.595288 RB6571_secreted protein 3489550 3490014 0 0 NaN 1.635307 RB66_hypothetical protein 26082 26336 0 0 NaN 1.625323 RB663_hypothetical protein 357608 357369 0 0 2.72 2.08
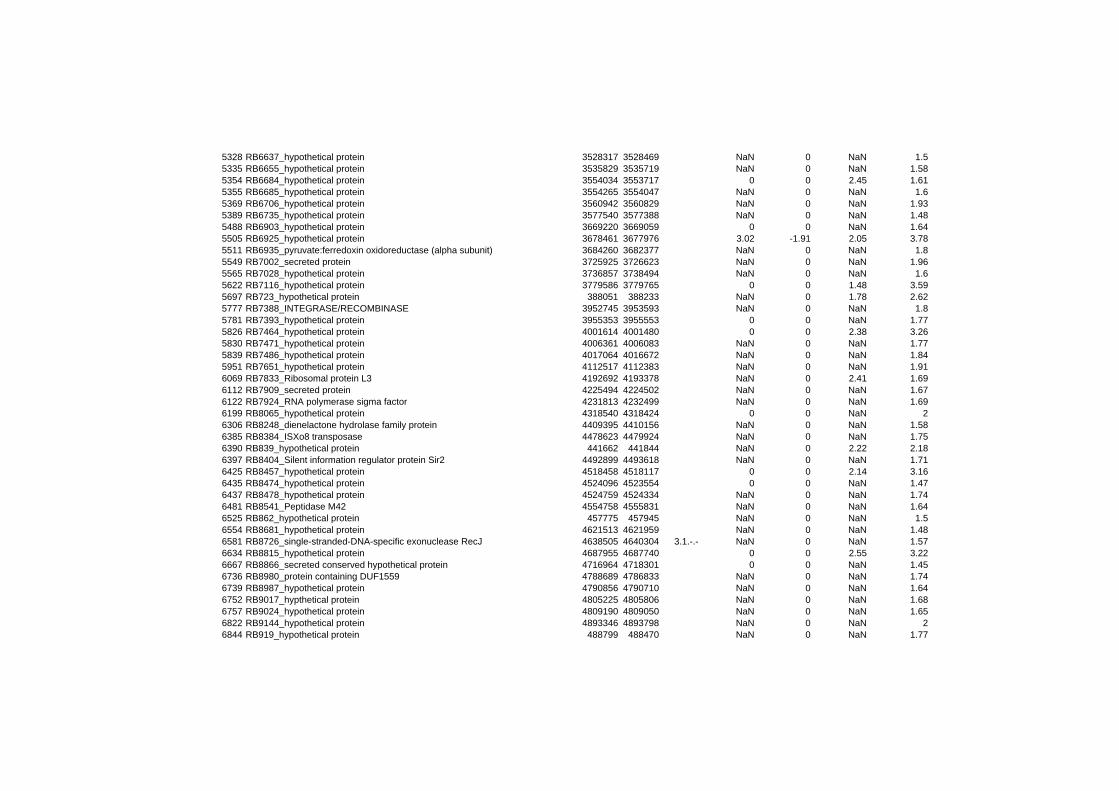
5328 RB6637_hypothetical protein 3528317 3528469 NaN 0 NaN 1.55335 RB6655_hypothetical protein 3535829 3535719 NaN 0 NaN 1.585354 RB6684_hypothetical protein 3554034 3553717 0 0 2.45 1.615355 RB6685_hypothetical protein 3554265 3554047 NaN 0 NaN 1.65369 RB6706_hypothetical protein 3560942 3560829 NaN 0 NaN 1.935389 RB6735_hypothetical protein 3577540 3577388 NaN 0 NaN 1.485488 RB6903_hypothetical protein 3669220 3669059 0 0 NaN 1.645505 RB6925_hypothetical protein 3678461 3677976 3.02 -1.91 2.05 3.785511 RB6935_pyruvate:ferredoxin oxidoreductase (alpha subunit) 3684260 3682377 NaN 0 NaN 1.85549 RB7002_secreted protein 3725925 3726623 NaN 0 NaN 1.965565 RB7028_hypothetical protein 3736857 3738494 NaN 0 NaN 1.65622 RB7116_hypothetical protein 3779586 3779765 0 0 1.48 3.595697 RB723_hypothetical protein 388051 388233 NaN 0 1.78 2.625777 RB7388_INTEGRASE/RECOMBINASE 3952745 3953593 NaN 0 NaN 1.85781 RB7393_hypothetical protein 3955353 3955553 0 0 NaN 1.775826 RB7464_hypothetical protein 4001614 4001480 0 0 2.38 3.265830 RB7471_hypothetical protein 4006361 4006083 NaN 0 NaN 1.775839 RB7486_hypothetical protein 4017064 4016672 NaN 0 NaN 1.845951 RB7651_hypothetical protein 4112517 4112383 NaN 0 NaN 1.916069 RB7833_Ribosomal protein L3 4192692 4193378 NaN 0 2.41 1.696112 RB7909_secreted protein 4225494 4224502 NaN 0 NaN 1.676122 RB7924_RNA polymerase sigma factor 4231813 4232499 NaN 0 NaN 1.696199 RB8065_hypothetical protein 4318540 4318424 0 0 NaN 26306 RB8248_dienelactone hydrolase family protein 4409395 4410156 NaN 0 NaN 1.586385 RB8384_ISXo8 transposase 4478623 4479924 NaN 0 NaN 1.756390 RB839_hypothetical protein 441662 441844 NaN 0 2.22 2.186397 RB8404_Silent information regulator protein Sir2 4492899 4493618 NaN 0 NaN 1.716425 RB8457_hypothetical protein 4518458 4518117 0 0 2.14 3.166435 RB8474_hypothetical protein 4524096 4523554 0 0 NaN 1.476437 RB8478_hypothetical protein 4524759 4524334 NaN 0 NaN 1.746481 RB8541_Peptidase M42 4554758 4555831 NaN 0 NaN 1.646525 RB862_hypothetical protein 457775 457945 NaN 0 NaN 1.56554 RB8681_hypothetical protein 4621513 4621959 NaN 0 NaN 1.486581 RB8726_single-stranded-DNA-specific exonuclease RecJ 4638505 4640304 3.1.-.- NaN 0 NaN 1.576634 RB8815_hypothetical protein 4687955 4687740 0 0 2.55 3.226667 RB8866_secreted conserved hypothetical protein 4716964 4718301 0 0 NaN 1.456736 RB8980_protein containing DUF1559 4788689 4786833 NaN 0 NaN 1.746739 RB8987_hypothetical protein 4790856 4790710 NaN 0 NaN 1.646752 RB9017_hypthetical protein 4805225 4805806 NaN 0 NaN 1.686757 RB9024_hypothetical protein 4809190 4809050 NaN 0 NaN 1.656822 RB9144_hypothetical protein 4893346 4893798 NaN 0 NaN 26844 RB919_hypothetical protein 488799 488470 NaN 0 NaN 1.77
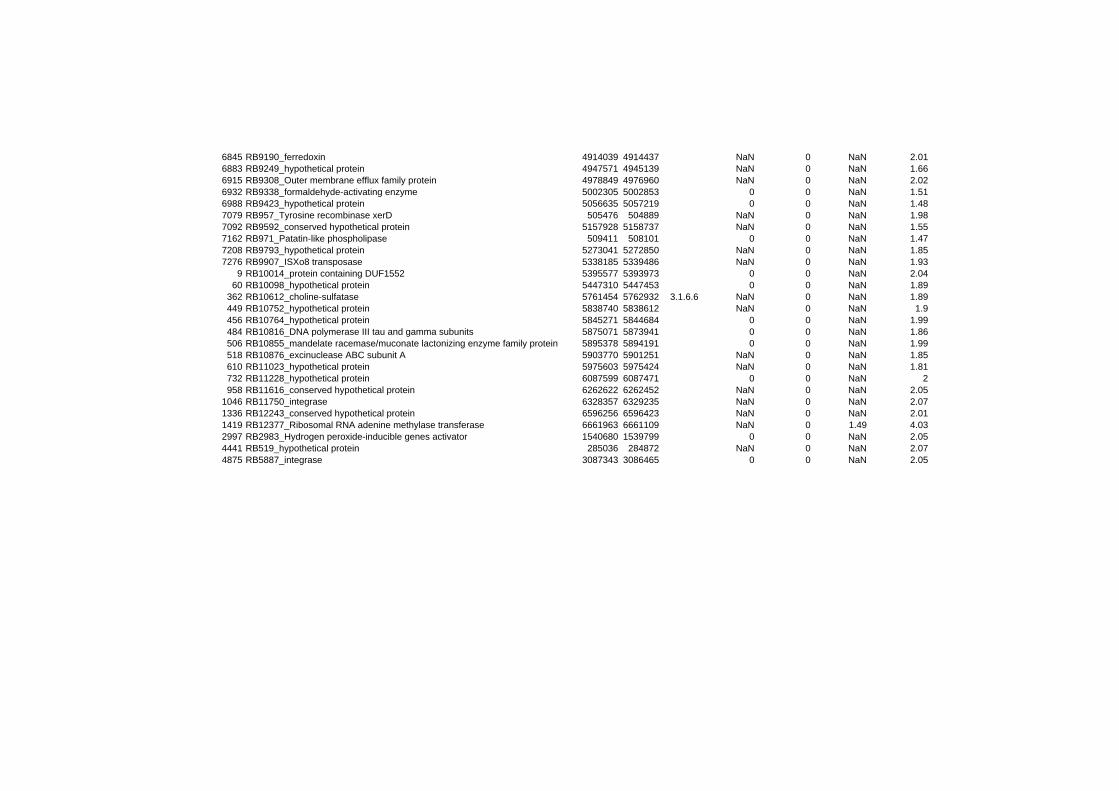
6845 RB9190_ferredoxin 4914039 4914437 NaN 0 NaN 2.016883 RB9249_hypothetical protein 4947571 4945139 NaN 0 NaN 1.666915 RB9308_Outer membrane efflux family protein 4978849 4976960 NaN 0 NaN 2.026932 RB9338_formaldehyde-activating enzyme 5002305 5002853 0 0 NaN 1.516988 RB9423_hypothetical protein 5056635 5057219 0 0 NaN 1.487079 RB957_Tyrosine recombinase xerD 505476 504889 NaN 0 NaN 1.987092 RB9592_conserved hypothetical protein 5157928 5158737 NaN 0 NaN 1.557162 RB971_Patatin-like phospholipase 509411 508101 0 0 NaN 1.477208 RB9793_hypothetical protein 5273041 5272850 NaN 0 NaN 1.857276 RB9907_ISXo8 transposase 5338185 5339486 NaN 0 NaN 1.93
9 RB10014_protein containing DUF1552 5395577 5393973 0 0 NaN 2.0460 RB10098_hypothetical protein 5447310 5447453 0 0 NaN 1.89
362 RB10612_choline-sulfatase 5761454 5762932 3.1.6.6 NaN 0 NaN 1.89449 RB10752_hypothetical protein 5838740 5838612 NaN 0 NaN 1.9456 RB10764_hypothetical protein 5845271 5844684 0 0 NaN 1.99484 RB10816_DNA polymerase III tau and gamma subunits 5875071 5873941 0 0 NaN 1.86506 RB10855_mandelate racemase/muconate lactonizing enzyme family protein 5895378 5894191 0 0 NaN 1.99518 RB10876_excinuclease ABC subunit A 5903770 5901251 NaN 0 NaN 1.85610 RB11023_hypothetical protein 5975603 5975424 NaN 0 NaN 1.81732 RB11228_hypothetical protein 6087599 6087471 0 0 NaN 2958 RB11616_conserved hypothetical protein 6262622 6262452 NaN 0 NaN 2.05
1046 RB11750_integrase 6328357 6329235 NaN 0 NaN 2.071336 RB12243_conserved hypothetical protein 6596256 6596423 NaN 0 NaN 2.011419 RB12377_Ribosomal RNA adenine methylase transferase 6661963 6661109 NaN 0 1.49 4.032997 RB2983_Hydrogen peroxide-inducible genes activator 1540680 1539799 0 0 NaN 2.054441 RB519_hypothetical protein 285036 284872 NaN 0 NaN 2.074875 RB5887_integrase 3087343 3086465 0 0 NaN 2.05
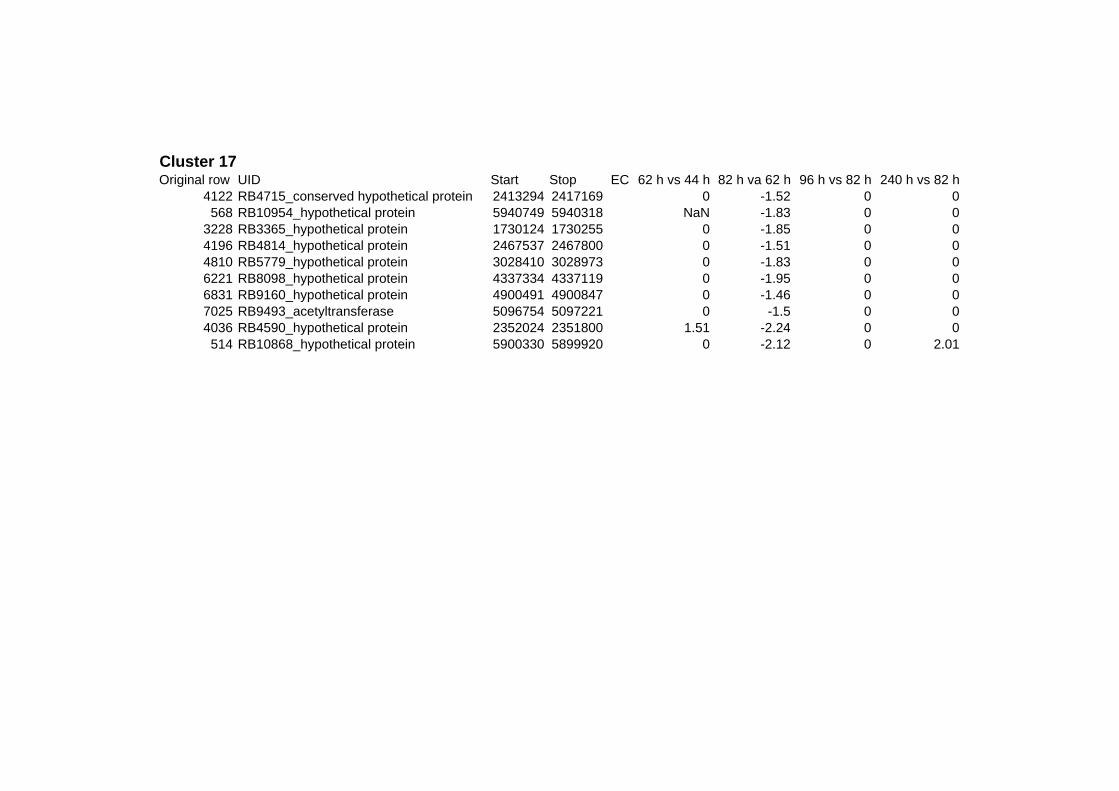
Cluster 17Original row UID Start Stop EC 62 h vs 44 h 82 h va 62 h 96 h vs 82 h 240 h vs 82 h
4122 RB4715_conserved hypothetical protein 2413294 2417169 0 -1.52 0 0568 RB10954_hypothetical protein 5940749 5940318 NaN -1.83 0 0
3228 RB3365_hypothetical protein 1730124 1730255 0 -1.85 0 04196 RB4814_hypothetical protein 2467537 2467800 0 -1.51 0 04810 RB5779_hypothetical protein 3028410 3028973 0 -1.83 0 06221 RB8098_hypothetical protein 4337334 4337119 0 -1.95 0 06831 RB9160_hypothetical protein 4900491 4900847 0 -1.46 0 07025 RB9493_acetyltransferase 5096754 5097221 0 -1.5 0 04036 RB4590_hypothetical protein 2352024 2351800 1.51 -2.24 0 0514 RB10868_hypothetical protein 5900330 5899920 0 -2.12 0 2.01
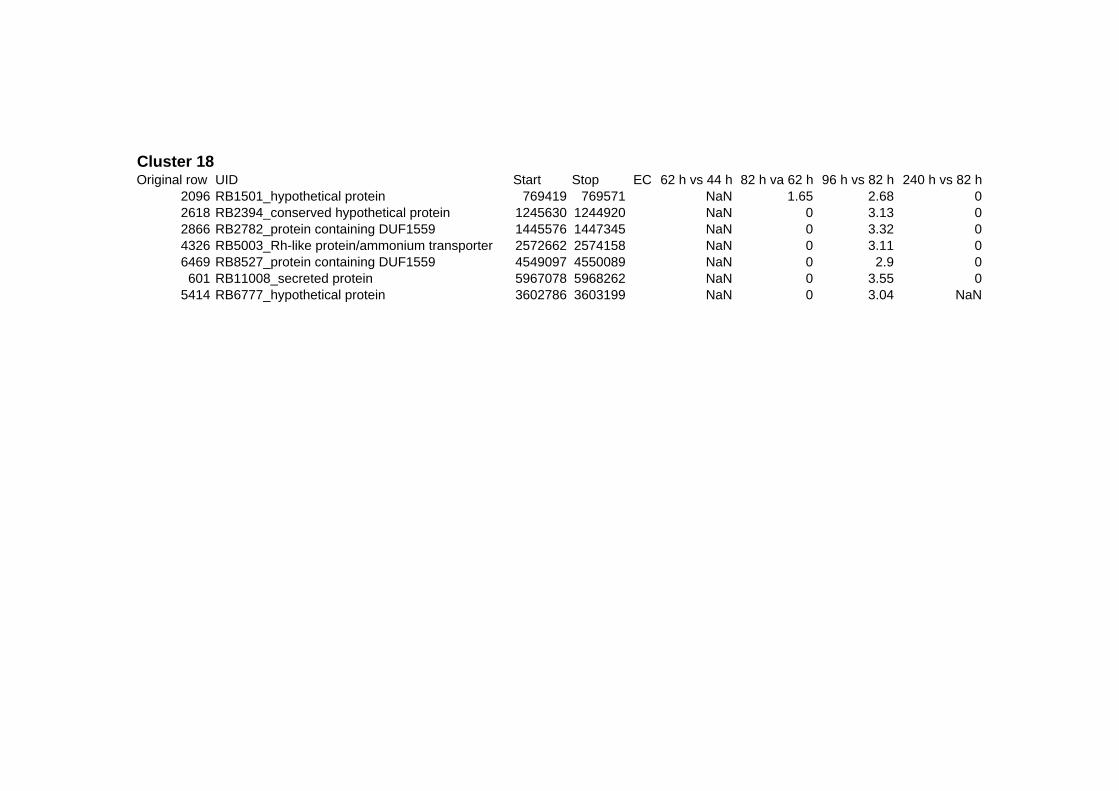
Cluster 18Original row UID Start Stop EC 62 h vs 44 h 82 h va 62 h 96 h vs 82 h 240 h vs 82 h
2096 RB1501_hypothetical protein 769419 769571 NaN 1.65 2.68 02618 RB2394_conserved hypothetical protein 1245630 1244920 NaN 0 3.13 02866 RB2782_protein containing DUF1559 1445576 1447345 NaN 0 3.32 04326 RB5003_Rh-like protein/ammonium transporter 2572662 2574158 NaN 0 3.11 06469 RB8527_protein containing DUF1559 4549097 4550089 NaN 0 2.9 0601 RB11008_secreted protein 5967078 5968262 NaN 0 3.55 0
5414 RB6777_hypothetical protein 3602786 3603199 NaN 0 3.04 NaN
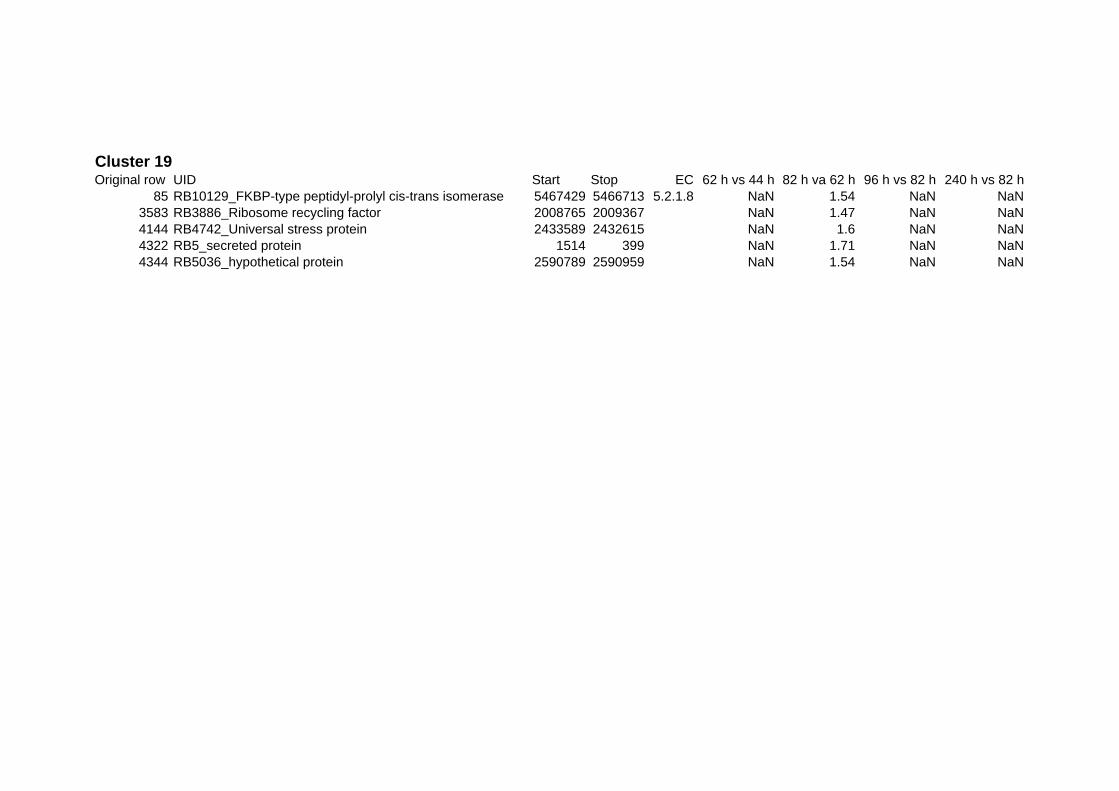
Cluster 19Original row UID Start Stop EC 62 h vs 44 h 82 h va 62 h 96 h vs 82 h 240 h vs 82 h
85 RB10129_FKBP-type peptidyl-prolyl cis-trans isomerase 5467429 5466713 5.2.1.8 NaN 1.54 NaN NaN3583 RB3886_Ribosome recycling factor 2008765 2009367 NaN 1.47 NaN NaN4144 RB4742_Universal stress protein 2433589 2432615 NaN 1.6 NaN NaN4322 RB5_secreted protein 1514 399 NaN 1.71 NaN NaN4344 RB5036_hypothetical protein 2590789 2590959 NaN 1.54 NaN NaN
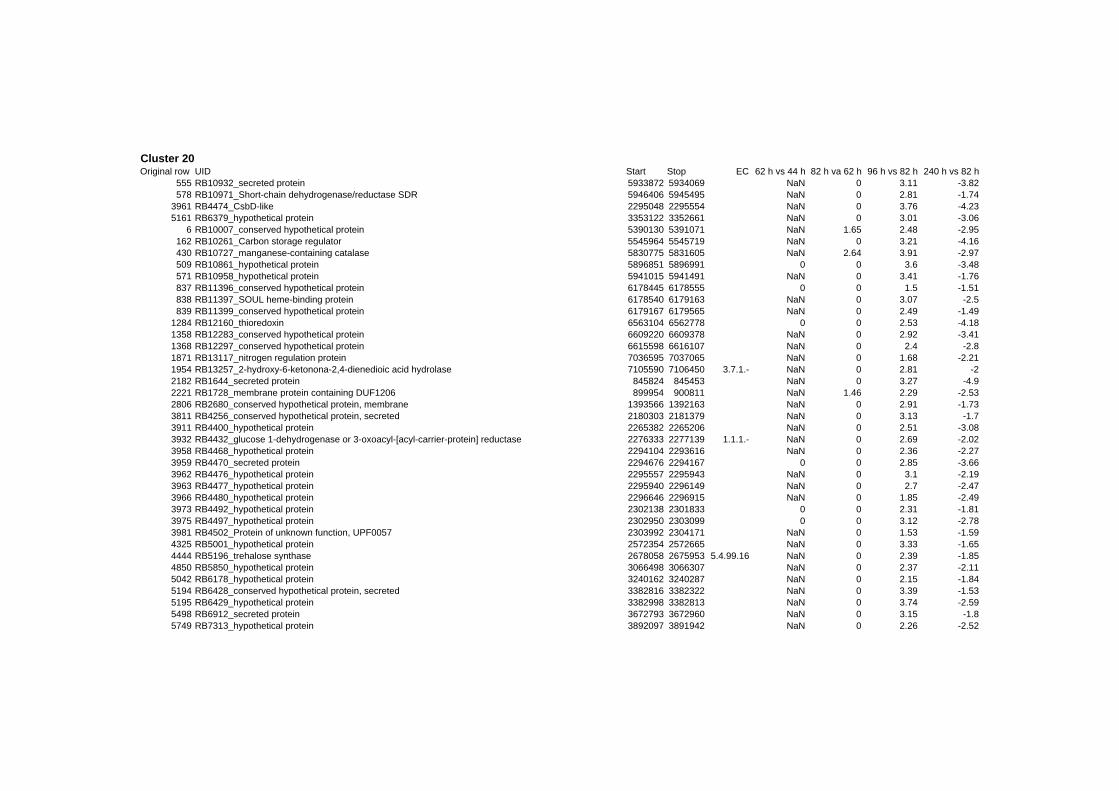
Cluster 20Original row UID Start Stop EC 62 h vs 44 h 82 h va 62 h 96 h vs 82 h 240 h vs 82 h
555 RB10932_secreted protein 5933872 5934069 NaN 0 3.11 -3.82578 RB10971_Short-chain dehydrogenase/reductase SDR 5946406 5945495 NaN 0 2.81 -1.74
3961 RB4474_CsbD-like 2295048 2295554 NaN 0 3.76 -4.235161 RB6379_hypothetical protein 3353122 3352661 NaN 0 3.01 -3.06
6 RB10007_conserved hypothetical protein 5390130 5391071 NaN 1.65 2.48 -2.95162 RB10261_Carbon storage regulator 5545964 5545719 NaN 0 3.21 -4.16430 RB10727_manganese-containing catalase 5830775 5831605 NaN 2.64 3.91 -2.97509 RB10861_hypothetical protein 5896851 5896991 0 0 3.6 -3.48571 RB10958_hypothetical protein 5941015 5941491 NaN 0 3.41 -1.76837 RB11396_conserved hypothetical protein 6178445 6178555 0 0 1.5 -1.51838 RB11397_SOUL heme-binding protein 6178540 6179163 NaN 0 3.07 -2.5839 RB11399_conserved hypothetical protein 6179167 6179565 NaN 0 2.49 -1.49
1284 RB12160_thioredoxin 6563104 6562778 0 0 2.53 -4.181358 RB12283_conserved hypothetical protein 6609220 6609378 NaN 0 2.92 -3.411368 RB12297_conserved hypothetical protein 6615598 6616107 NaN 0 2.4 -2.81871 RB13117_nitrogen regulation protein 7036595 7037065 NaN 0 1.68 -2.211954 RB13257_2-hydroxy-6-ketonona-2,4-dienedioic acid hydrolase 7105590 7106450 3.7.1.- NaN 0 2.81 -22182 RB1644_secreted protein 845824 845453 NaN 0 3.27 -4.92221 RB1728_membrane protein containing DUF1206 899954 900811 NaN 1.46 2.29 -2.532806 RB2680_conserved hypothetical protein, membrane 1393566 1392163 NaN 0 2.91 -1.733811 RB4256_conserved hypothetical protein, secreted 2180303 2181379 NaN 0 3.13 -1.73911 RB4400_hypothetical protein 2265382 2265206 NaN 0 2.51 -3.083932 RB4432_glucose 1-dehydrogenase or 3-oxoacyl-[acyl-carrier-protein] reductase 2276333 2277139 1.1.1.- NaN 0 2.69 -2.023958 RB4468_hypothetical protein 2294104 2293616 NaN 0 2.36 -2.273959 RB4470_secreted protein 2294676 2294167 0 0 2.85 -3.663962 RB4476_hypothetical protein 2295557 2295943 NaN 0 3.1 -2.193963 RB4477_hypothetical protein 2295940 2296149 NaN 0 2.7 -2.473966 RB4480_hypothetical protein 2296646 2296915 NaN 0 1.85 -2.493973 RB4492_hypothetical protein 2302138 2301833 0 0 2.31 -1.813975 RB4497_hypothetical protein 2302950 2303099 0 0 3.12 -2.783981 RB4502_Protein of unknown function, UPF0057 2303992 2304171 NaN 0 1.53 -1.594325 RB5001_hypothetical protein 2572354 2572665 NaN 0 3.33 -1.654444 RB5196_trehalose synthase 2678058 2675953 5.4.99.16 NaN 0 2.39 -1.854850 RB5850_hypothetical protein 3066498 3066307 NaN 0 2.37 -2.115042 RB6178_hypothetical protein 3240162 3240287 NaN 0 2.15 -1.845194 RB6428_conserved hypothetical protein, secreted 3382816 3382322 NaN 0 3.39 -1.535195 RB6429_hypothetical protein 3382998 3382813 NaN 0 3.74 -2.595498 RB6912_secreted protein 3672793 3672960 NaN 0 3.15 -1.85749 RB7313_hypothetical protein 3892097 3891942 NaN 0 2.26 -2.52
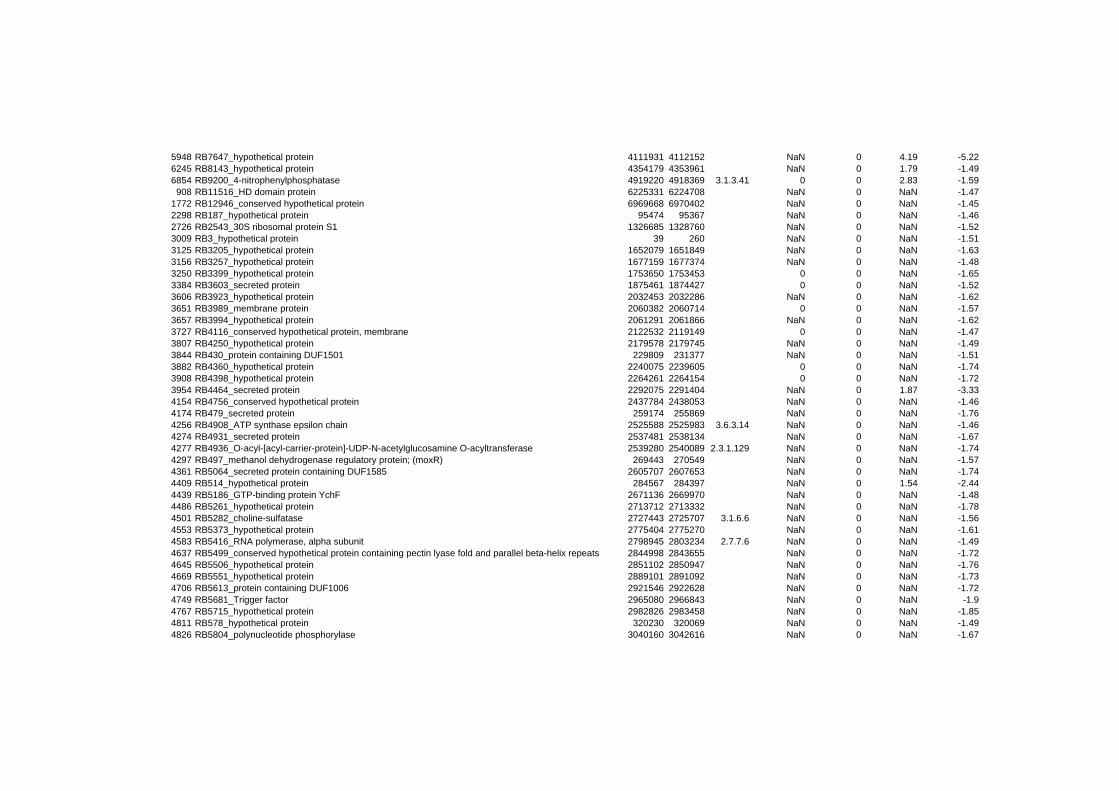
5948 RB7647_hypothetical protein 4111931 4112152 NaN 0 4.19 -5.226245 RB8143_hypothetical protein 4354179 4353961 NaN 0 1.79 -1.496854 RB9200_4-nitrophenylphosphatase 4919220 4918369 3.1.3.41 0 0 2.83 -1.59908 RB11516_HD domain protein 6225331 6224708 NaN 0 NaN -1.47
1772 RB12946_conserved hypothetical protein 6969668 6970402 NaN 0 NaN -1.452298 RB187_hypothetical protein 95474 95367 NaN 0 NaN -1.462726 RB2543_30S ribosomal protein S1 1326685 1328760 NaN 0 NaN -1.523009 RB3_hypothetical protein 39 260 NaN 0 NaN -1.513125 RB3205_hypothetical protein 1652079 1651849 NaN 0 NaN -1.633156 RB3257_hypothetical protein 1677159 1677374 NaN 0 NaN -1.483250 RB3399_hypothetical protein 1753650 1753453 0 0 NaN -1.653384 RB3603_secreted protein 1875461 1874427 0 0 NaN -1.523606 RB3923_hypothetical protein 2032453 2032286 NaN 0 NaN -1.623651 RB3989_membrane protein 2060382 2060714 0 0 NaN -1.573657 RB3994_hypothetical protein 2061291 2061866 NaN 0 NaN -1.623727 RB4116_conserved hypothetical protein, membrane 2122532 2119149 0 0 NaN -1.473807 RB4250_hypothetical protein 2179578 2179745 NaN 0 NaN -1.493844 RB430_protein containing DUF1501 229809 231377 NaN 0 NaN -1.513882 RB4360_hypothetical protein 2240075 2239605 0 0 NaN -1.743908 RB4398_hypothetical protein 2264261 2264154 0 0 NaN -1.723954 RB4464_secreted protein 2292075 2291404 NaN 0 1.87 -3.334154 RB4756_conserved hypothetical protein 2437784 2438053 NaN 0 NaN -1.464174 RB479_secreted protein 259174 255869 NaN 0 NaN -1.764256 RB4908_ATP synthase epsilon chain 2525588 2525983 3.6.3.14 NaN 0 NaN -1.464274 RB4931_secreted protein 2537481 2538134 NaN 0 NaN -1.674277 RB4936_O-acyl-[acyl-carrier-protein]-UDP-N-acetylglucosamine O-acyltransferase 2539280 2540089 2.3.1.129 NaN 0 NaN -1.744297 RB497_methanol dehydrogenase regulatory protein; (moxR) 269443 270549 NaN 0 NaN -1.574361 RB5064_secreted protein containing DUF1585 2605707 2607653 NaN 0 NaN -1.744409 RB514_hypothetical protein 284567 284397 NaN 0 1.54 -2.444439 RB5186_GTP-binding protein YchF 2671136 2669970 NaN 0 NaN -1.484486 RB5261_hypothetical protein 2713712 2713332 NaN 0 NaN -1.784501 RB5282_choline-sulfatase 2727443 2725707 3.1.6.6 NaN 0 NaN -1.564553 RB5373_hypothetical protein 2775404 2775270 NaN 0 NaN -1.614583 RB5416_RNA polymerase, alpha subunit 2798945 2803234 2.7.7.6 NaN 0 NaN -1.494637 RB5499_conserved hypothetical protein containing pectin lyase fold and parallel beta-helix repeats 2844998 2843655 NaN 0 NaN -1.724645 RB5506_hypothetical protein 2851102 2850947 NaN 0 NaN -1.764669 RB5551_hypothetical protein 2889101 2891092 NaN 0 NaN -1.734706 RB5613_protein containing DUF1006 2921546 2922628 NaN 0 NaN -1.724749 RB5681_Trigger factor 2965080 2966843 NaN 0 NaN -1.94767 RB5715_hypothetical protein 2982826 2983458 NaN 0 NaN -1.854811 RB578_hypothetical protein 320230 320069 NaN 0 NaN -1.494826 RB5804_polynucleotide phosphorylase 3040160 3042616 NaN 0 NaN -1.67

4895 RB5924_hypothetical protein 3105828 3103285 0 0 NaN -1.594928 RB5983_Acetylglutamate kinase 3137623 3138486 2.7.2.8 0 0 NaN -1.474981 RB6071_hypothetical protein 3187463 3188104 NaN 0 NaN -1.665028 RB6156_membrane protein 3233026 3232622 NaN 0 NaN -1.535162 RB638_PQQ enzyme repeat domain protein 346447 347757 NaN 0 NaN -1.815249 RB6504_hypothetical protein 3456120 3455959 NaN 0 NaN -1.755293 RB6578_secreted protein 3495957 3495022 NaN 0 NaN -1.55366 RB6701_hypothetical protein 3559944 3560207 NaN 0 NaN -1.635377 RB6716_arsenical pump membrane protein 3567209 3565803 NaN 0 NaN -1.465383 RB6728_hypothetical protein 3570581 3570922 0 0 NaN -1.555392 RB674_membrane protein 359779 359528 0 0 NaN -1.465417 RB6780_RNA polymerase sigma factor rpoD 3603381 3605054 NaN 0 NaN -1.465456 RB684_Arylsulfatase precursor 368557 366596 3.1.6.1 0 0 NaN -1.545466 RB6856_hypothetical protein 3645613 3645425 NaN 0 NaN -1.575484 RB6891_O-antigen polymerase-like protein, membrane 3665156 3662541 0 0 NaN -1.455497 RB6911_protein containing DUF1559 3671445 3672737 NaN 0 2.09 -3.195507 RB693_protein containing DUF1568 371190 370408 NaN 0 NaN -1.455561 RB7022_Ribosomal protein S20 3735291 3735563 NaN 0 NaN -1.85592 RB7078_outer membrane efflux protein 3758295 3760328 NaN 0 NaN -1.475629 RB7129_iron-dependent transcriptional repressor 3782388 3782618 NaN 0 NaN -2.35725 RB7279_hypothetical protein 3870019 3870309 NaN 0 NaN -2.045737 RB7296_hypothetical protein 3877224 3877472 NaN 0 NaN -1.665741 RB73_membrane protein 28727 29023 NaN 0 NaN -1.565748 RB7312_hypothetical protein 3891868 3891704 NaN 0 NaN -1.655849 RB7507_conserved hypothetical protein, membrane 4036748 4038313 NaN 0 NaN -1.65899 RB7579_sigma-54 dependent transcriptional regulator/response regulator 4071162 4072619 NaN 0 NaN -1.495900 RB758_hypothetical protein 407558 407340 NaN 0 NaN -1.515908 RB7589_hypothetical protein 4080238 4079978 NaN 0 NaN -1.665941 RB7634_Abortive infection protein 4106267 4108588 NaN 0 NaN -1.665960 RB7667_hypothetical protein 4117097 4116981 NaN 0 1.75 -2.595964 RB7673_membrane protein containing DUF107 4120808 4119234 NaN 0 NaN -1.485993 RB7722_membrane protein 4137251 4136415 0 0 NaN -1.676117 RB7914_hypothetical protein 4228476 4227877 NaN 0 NaN -1.526128 RB7937_hypothetical protein 4237809 4237684 NaN 0 NaN -1.536140 RB7955_hypothetical protein 4247114 4246743 NaN 0 NaN -1.56150 RB7978_hypothetical protein 4265473 4265817 NaN 0 NaN -1.56169 RB8012_hypothetical protein 4282669 4282280 NaN 0 NaN -1.536176 RB8027_Sigma factor, ECF-like 4289305 4290003 NaN 0 NaN -1.516181 RB8031_6-phosphogluconolactonase 4295373 4294183 3.1.1.31 NaN 0 NaN -2.26182 RB8034_membrane protein 4295344 4295787 NaN 0 NaN -1.596200 RB8066_hypothetical protein 4318635 4318519 NaN 0 NaN -1.516204 RB8070_hypothetical protein 4319538 4319717 NaN 0 NaN -1.48
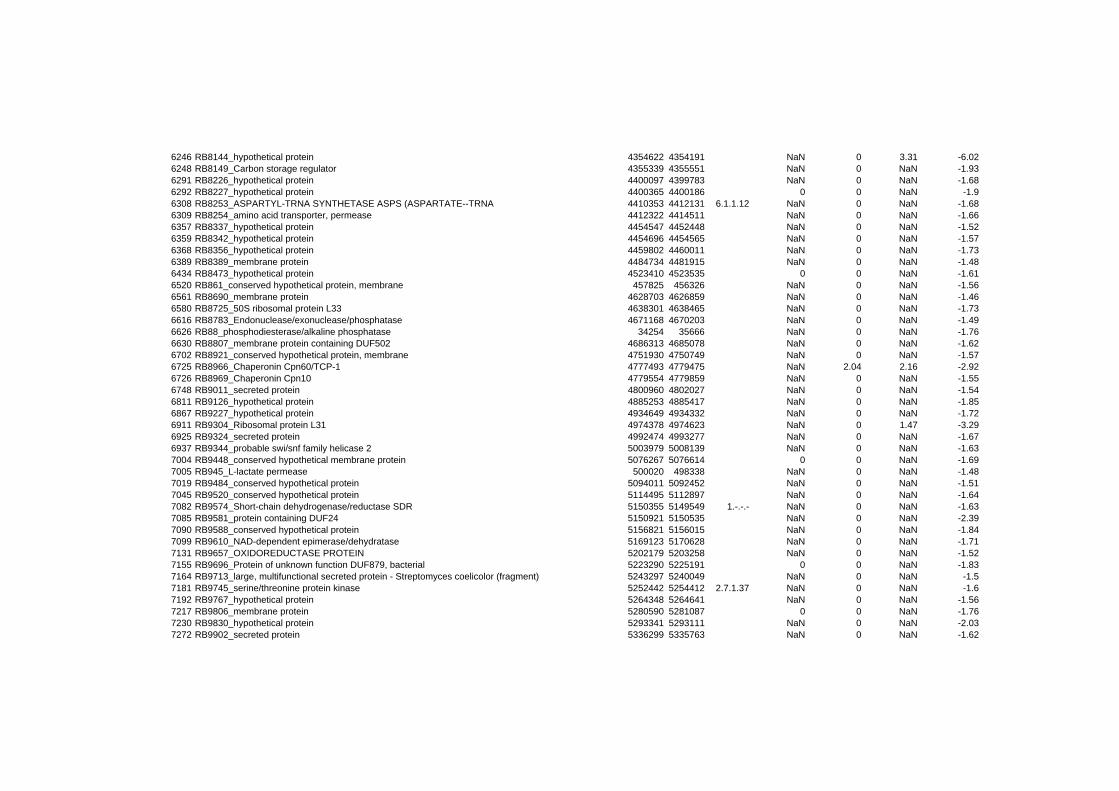
6246 RB8144_hypothetical protein 4354622 4354191 NaN 0 3.31 -6.026248 RB8149_Carbon storage regulator 4355339 4355551 NaN 0 NaN -1.936291 RB8226_hypothetical protein 4400097 4399783 NaN 0 NaN -1.686292 RB8227_hypothetical protein 4400365 4400186 0 0 NaN -1.96308 RB8253_ASPARTYL-TRNA SYNTHETASE ASPS (ASPARTATE--TRNA 4410353 4412131 6.1.1.12 NaN 0 NaN -1.686309 RB8254_amino acid transporter, permease 4412322 4414511 NaN 0 NaN -1.666357 RB8337_hypothetical protein 4454547 4452448 NaN 0 NaN -1.526359 RB8342_hypothetical protein 4454696 4454565 NaN 0 NaN -1.576368 RB8356_hypothetical protein 4459802 4460011 NaN 0 NaN -1.736389 RB8389_membrane protein 4484734 4481915 NaN 0 NaN -1.486434 RB8473_hypothetical protein 4523410 4523535 0 0 NaN -1.616520 RB861_conserved hypothetical protein, membrane 457825 456326 NaN 0 NaN -1.566561 RB8690_membrane protein 4628703 4626859 NaN 0 NaN -1.466580 RB8725_50S ribosomal protein L33 4638301 4638465 NaN 0 NaN -1.736616 RB8783_Endonuclease/exonuclease/phosphatase 4671168 4670203 NaN 0 NaN -1.496626 RB88_phosphodiesterase/alkaline phosphatase 34254 35666 NaN 0 NaN -1.766630 RB8807_membrane protein containing DUF502 4686313 4685078 NaN 0 NaN -1.626702 RB8921_conserved hypothetical protein, membrane 4751930 4750749 NaN 0 NaN -1.576725 RB8966_Chaperonin Cpn60/TCP-1 4777493 4779475 NaN 2.04 2.16 -2.926726 RB8969_Chaperonin Cpn10 4779554 4779859 NaN 0 NaN -1.556748 RB9011_secreted protein 4800960 4802027 NaN 0 NaN -1.546811 RB9126_hypothetical protein 4885253 4885417 NaN 0 NaN -1.856867 RB9227_hypothetical protein 4934649 4934332 NaN 0 NaN -1.726911 RB9304_Ribosomal protein L31 4974378 4974623 NaN 0 1.47 -3.296925 RB9324_secreted protein 4992474 4993277 NaN 0 NaN -1.676937 RB9344_probable swi/snf family helicase 2 5003979 5008139 NaN 0 NaN -1.637004 RB9448_conserved hypothetical membrane protein 5076267 5076614 0 0 NaN -1.697005 RB945_L-lactate permease 500020 498338 NaN 0 NaN -1.487019 RB9484_conserved hypothetical protein 5094011 5092452 NaN 0 NaN -1.517045 RB9520_conserved hypothetical protein 5114495 5112897 NaN 0 NaN -1.647082 RB9574_Short-chain dehydrogenase/reductase SDR 5150355 5149549 1.-.-.- NaN 0 NaN -1.637085 RB9581_protein containing DUF24 5150921 5150535 NaN 0 NaN -2.397090 RB9588_conserved hypothetical protein 5156821 5156015 NaN 0 NaN -1.847099 RB9610_NAD-dependent epimerase/dehydratase 5169123 5170628 NaN 0 NaN -1.717131 RB9657_OXIDOREDUCTASE PROTEIN 5202179 5203258 NaN 0 NaN -1.527155 RB9696_Protein of unknown function DUF879, bacterial 5223290 5225191 0 0 NaN -1.837164 RB9713_large, multifunctional secreted protein - Streptomyces coelicolor (fragment) 5243297 5240049 NaN 0 NaN -1.57181 RB9745_serine/threonine protein kinase 5252442 5254412 2.7.1.37 NaN 0 NaN -1.67192 RB9767_hypothetical protein 5264348 5264641 NaN 0 NaN -1.567217 RB9806_membrane protein 5280590 5281087 0 0 NaN -1.767230 RB9830_hypothetical protein 5293341 5293111 NaN 0 NaN -2.037272 RB9902_secreted protein 5336299 5335763 NaN 0 NaN -1.62
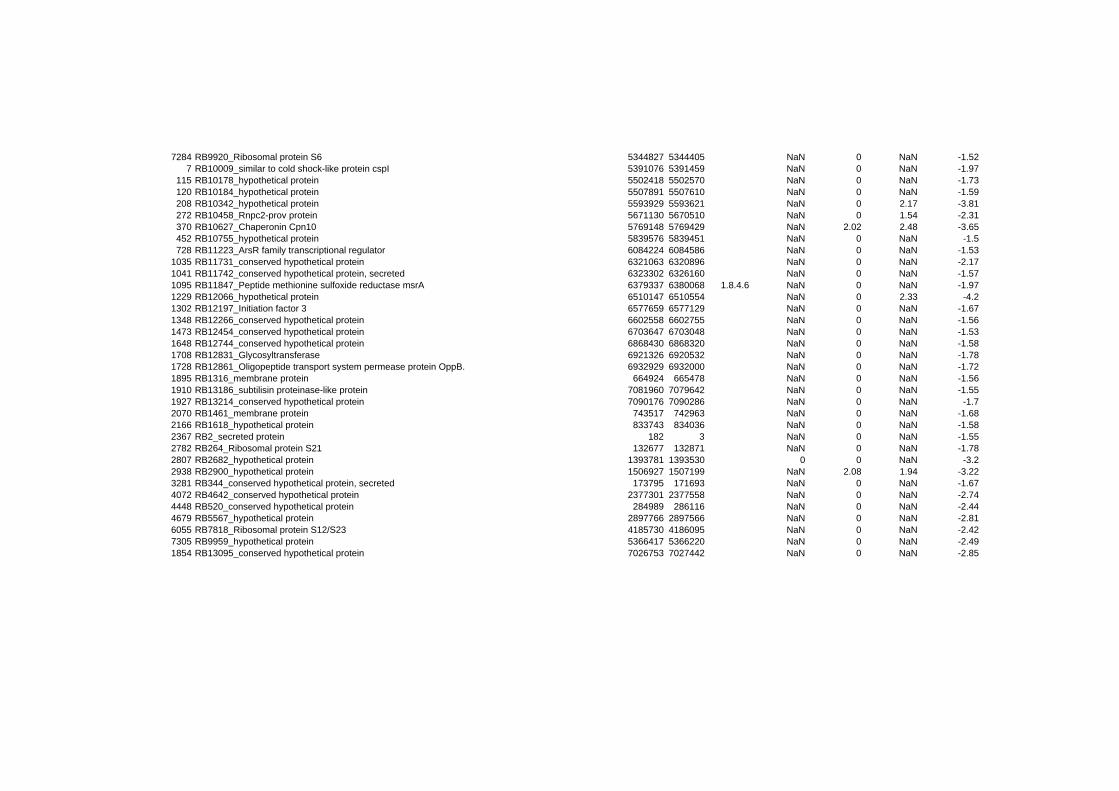
7284 RB9920_Ribosomal protein S6 5344827 5344405 NaN 0 NaN -1.527 RB10009_similar to cold shock-like protein cspI 5391076 5391459 NaN 0 NaN -1.97
115 RB10178_hypothetical protein 5502418 5502570 NaN 0 NaN -1.73120 RB10184_hypothetical protein 5507891 5507610 NaN 0 NaN -1.59208 RB10342_hypothetical protein 5593929 5593621 NaN 0 2.17 -3.81272 RB10458_Rnpc2-prov protein 5671130 5670510 NaN 0 1.54 -2.31370 RB10627_Chaperonin Cpn10 5769148 5769429 NaN 2.02 2.48 -3.65452 RB10755_hypothetical protein 5839576 5839451 NaN 0 NaN -1.5728 RB11223_ArsR family transcriptional regulator 6084224 6084586 NaN 0 NaN -1.53
1035 RB11731_conserved hypothetical protein 6321063 6320896 NaN 0 NaN -2.171041 RB11742_conserved hypothetical protein, secreted 6323302 6326160 NaN 0 NaN -1.571095 RB11847_Peptide methionine sulfoxide reductase msrA 6379337 6380068 1.8.4.6 NaN 0 NaN -1.971229 RB12066_hypothetical protein 6510147 6510554 NaN 0 2.33 -4.21302 RB12197_Initiation factor 3 6577659 6577129 NaN 0 NaN -1.671348 RB12266_conserved hypothetical protein 6602558 6602755 NaN 0 NaN -1.561473 RB12454_conserved hypothetical protein 6703647 6703048 NaN 0 NaN -1.531648 RB12744_conserved hypothetical protein 6868430 6868320 NaN 0 NaN -1.581708 RB12831_Glycosyltransferase 6921326 6920532 NaN 0 NaN -1.781728 RB12861_Oligopeptide transport system permease protein OppB. 6932929 6932000 NaN 0 NaN -1.721895 RB1316_membrane protein 664924 665478 NaN 0 NaN -1.561910 RB13186_subtilisin proteinase-like protein 7081960 7079642 NaN 0 NaN -1.551927 RB13214_conserved hypothetical protein 7090176 7090286 NaN 0 NaN -1.72070 RB1461_membrane protein 743517 742963 NaN 0 NaN -1.682166 RB1618_hypothetical protein 833743 834036 NaN 0 NaN -1.582367 RB2_secreted protein 182 3 NaN 0 NaN -1.552782 RB264_Ribosomal protein S21 132677 132871 NaN 0 NaN -1.782807 RB2682_hypothetical protein 1393781 1393530 0 0 NaN -3.22938 RB2900_hypothetical protein 1506927 1507199 NaN 2.08 1.94 -3.223281 RB344_conserved hypothetical protein, secreted 173795 171693 NaN 0 NaN -1.674072 RB4642_conserved hypothetical protein 2377301 2377558 NaN 0 NaN -2.744448 RB520_conserved hypothetical protein 284989 286116 NaN 0 NaN -2.444679 RB5567_hypothetical protein 2897766 2897566 NaN 0 NaN -2.816055 RB7818_Ribosomal protein S12/S23 4185730 4186095 NaN 0 NaN -2.427305 RB9959_hypothetical protein 5366417 5366220 NaN 0 NaN -2.491854 RB13095_conserved hypothetical protein 7026753 7027442 NaN 0 NaN -2.85
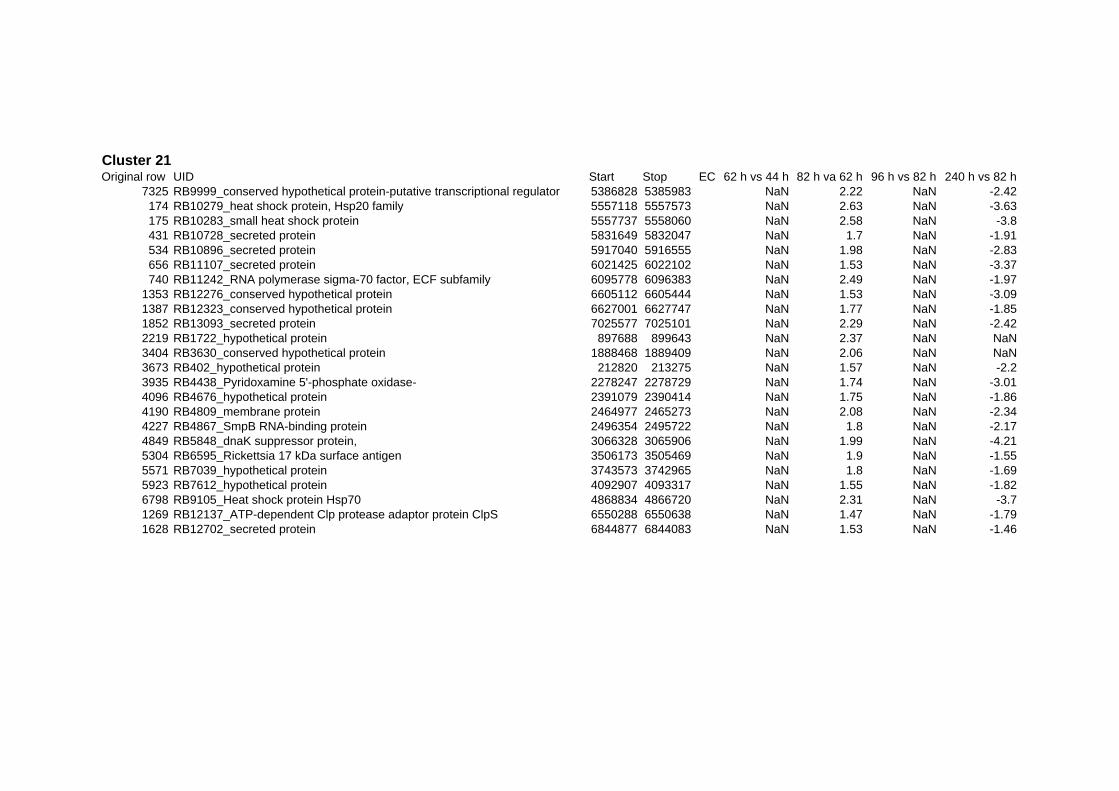
Cluster 21Original row UID Start Stop EC 62 h vs 44 h 82 h va 62 h 96 h vs 82 h 240 h vs 82 h
7325 RB9999_conserved hypothetical protein-putative transcriptional regulator 5386828 5385983 NaN 2.22 NaN -2.42174 RB10279_heat shock protein, Hsp20 family 5557118 5557573 NaN 2.63 NaN -3.63175 RB10283_small heat shock protein 5557737 5558060 NaN 2.58 NaN -3.8431 RB10728_secreted protein 5831649 5832047 NaN 1.7 NaN -1.91534 RB10896_secreted protein 5917040 5916555 NaN 1.98 NaN -2.83656 RB11107_secreted protein 6021425 6022102 NaN 1.53 NaN -3.37740 RB11242_RNA polymerase sigma-70 factor, ECF subfamily 6095778 6096383 NaN 2.49 NaN -1.97
1353 RB12276_conserved hypothetical protein 6605112 6605444 NaN 1.53 NaN -3.091387 RB12323_conserved hypothetical protein 6627001 6627747 NaN 1.77 NaN -1.851852 RB13093_secreted protein 7025577 7025101 NaN 2.29 NaN -2.422219 RB1722_hypothetical protein 897688 899643 NaN 2.37 NaN NaN3404 RB3630_conserved hypothetical protein 1888468 1889409 NaN 2.06 NaN NaN3673 RB402_hypothetical protein 212820 213275 NaN 1.57 NaN -2.23935 RB4438_Pyridoxamine 5'-phosphate oxidase- 2278247 2278729 NaN 1.74 NaN -3.014096 RB4676_hypothetical protein 2391079 2390414 NaN 1.75 NaN -1.864190 RB4809_membrane protein 2464977 2465273 NaN 2.08 NaN -2.344227 RB4867_SmpB RNA-binding protein 2496354 2495722 NaN 1.8 NaN -2.174849 RB5848_dnaK suppressor protein, 3066328 3065906 NaN 1.99 NaN -4.215304 RB6595_Rickettsia 17 kDa surface antigen 3506173 3505469 NaN 1.9 NaN -1.555571 RB7039_hypothetical protein 3743573 3742965 NaN 1.8 NaN -1.695923 RB7612_hypothetical protein 4092907 4093317 NaN 1.55 NaN -1.826798 RB9105_Heat shock protein Hsp70 4868834 4866720 NaN 2.31 NaN -3.71269 RB12137_ATP-dependent Clp protease adaptor protein ClpS 6550288 6550638 NaN 1.47 NaN -1.791628 RB12702_secreted protein 6844877 6844083 NaN 1.53 NaN -1.46

Cluster 22Original row UID Start Stop EC 62 h vs 44 h 82 h va 62 h 96 h vs 82 h 240 h vs 82 h
3743 RB4147_hypothetical protein 2131928 2132338 NaN 1.54 NaN 03992 RB4517_Transcriptional coactivator/pterin dehydratase 2315930 2316283 NaN 1.75 NaN 04194 RB4812_secreted protein 2467011 2467289 NaN 1.88 NaN 04896 RB5927_protein containing DUF1501 3107242 3105905 NaN 1.5 0 05020 RB6144_hypothetical protein 3228305 3228189 NaN 1.46 NaN 05763 RB7362_hypothetical protein 3939758 3939501 NaN 1.77 0 06410 RB8421_conserved hypothetical protein, membrane 4501840 4502310 NaN 1.55 NaN 06957 RB9372_Imidazole glycerol phosphate synthase subunit hisH 5021363 5020746 2.4.2.- NaN 2.43 -1.48 07116 RB9635_hypothetical protein 5187031 5185973 NaN 1.51 NaN 07319 RB9986_hypothetical protein 5380717 5380379 NaN 1.49 NaN 0183 RB10299_ATP-dependent carboligase related to biotin carboxylase (ORFY) 5563036 5564403 -1.7 2.45 -1.73 0369 RB10623_secreted protein 5769140 5768958 -2.4 2.03 0 -1.85371 RB10629_GroEL 5769500 5771119 NaN 1.76 NaN 0854 RB11421_60-kDa SS-A/Ro ribonucleoprotein 6195538 6193880 NaN 1.73 NaN 0960 RB11618_type I restriction-modification system, M subunit, 6263708 6265366 NaN 1.58 NaN 0
1085 RB1183_membrane protein 605059 605553 NaN 1.54 0 01136 RB11919_Dihydroxy-acid and 6-phosphogluconate dehydratase 6417713 6415698 NaN 1.67 NaN 01354 RB12278_conserved hypothetical protein 6605390 6605548 NaN 3.27 0 01434 RB12395_conserved hypothetical protein 6670730 6670572 NaN 1.66 0 01851 RB13092_conserved hypothetical protein 7025003 7025143 NaN 1.64 NaN 01916 RB13194_conserved hypothetical protein 7084723 7084920 NaN 1.94 0 01920 RB13202_conserved hypothetical protein 7086189 7086052 NaN 1.68 NaN 02062 RB1449_hypothetical protein 736500 736697 NaN 2.71 NaN 02655 RB2459_hypothetical protein 1280452 1280204 NaN 1.58 NaN 03275 RB3430_conserved hypothetical protein, membrane 1772646 1773392 NaN 1.54 NaN 03493 RB3751_NAD-dependent epimerase/dehydratase 1938953 1940068 NaN 1.95 NaN 03852 RB4308_protein containing DUF1586 2221461 2221222 NaN 2.04 NaN 04473 RB5238_hypothetical protein 2697775 2697996 NaN 1.79 NaN NaN4748 RB568_hypothetical protein 312345 312112 0 3.58 0 -1.545461 RB6848_secreted protein 3642216 3642443 NaN 1.86 NaN NaN5924 RB7613_hypothetical protein 4093558 4093418 NaN 1.71 0 1.51230 RB12067_conserved hypothetical protein 6510236 6510559 NaN 1.86 NaN NaN1277 RB12150_conserved hypothetical protein 6556901 6557062 NaN 1.75 0 -1.541890 RB13150_sialic acid-specific 9-O-acetylesterase 7059586 7058003 3.1.1.53 NaN 1.83 NaN NaN3132 RB3217_S-adenosylmethionine decarboxylase proenzyme 1654745 1654251 4.1.1.50 0 1.82 NaN 05400 RB6751_chaperone ClpB 3586111 3588690 NaN 1.78 NaN NaN4253 RB4903_xylulose-5-phosphate/fructose-6-phosphate phosphoketolase 2520925 2523306 4.1.2.9 NaN 1.91 NaN NaN
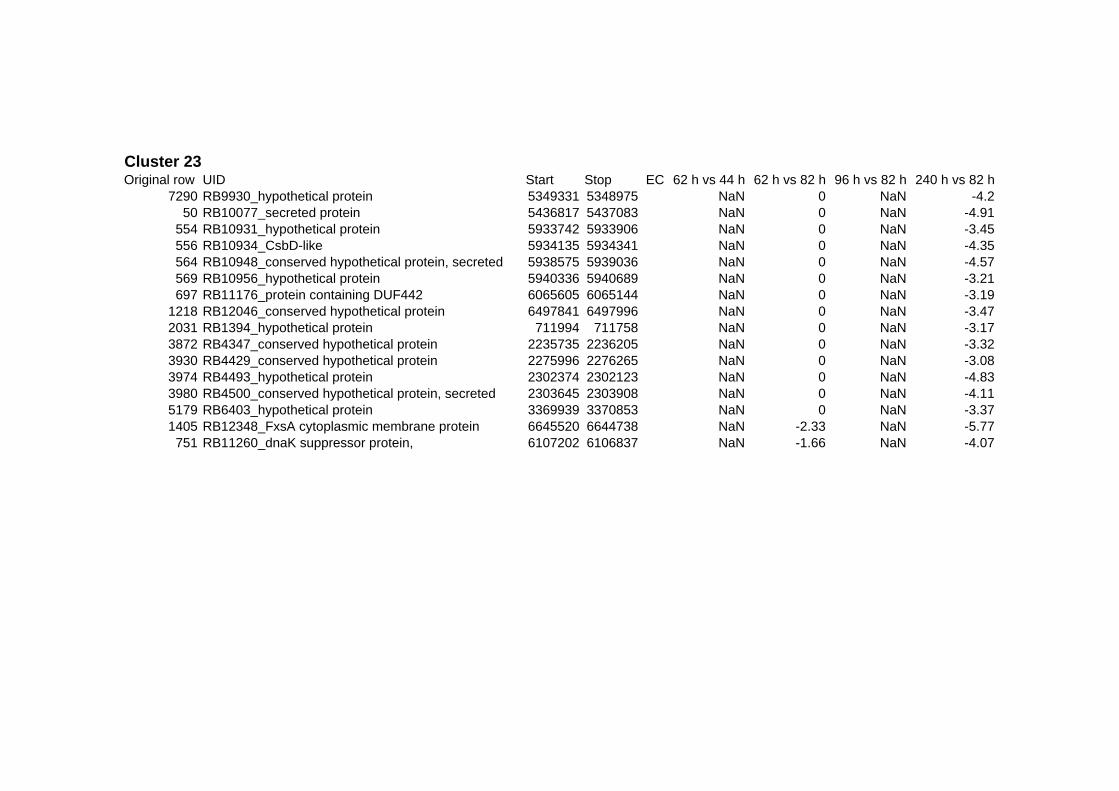
Cluster 23Original row UID Start Stop EC 62 h vs 44 h 62 h vs 82 h 96 h vs 82 h 240 h vs 82 h
7290 RB9930_hypothetical protein 5349331 5348975 NaN 0 NaN -4.250 RB10077_secreted protein 5436817 5437083 NaN 0 NaN -4.91
554 RB10931_hypothetical protein 5933742 5933906 NaN 0 NaN -3.45556 RB10934_CsbD-like 5934135 5934341 NaN 0 NaN -4.35564 RB10948_conserved hypothetical protein, secreted 5938575 5939036 NaN 0 NaN -4.57569 RB10956_hypothetical protein 5940336 5940689 NaN 0 NaN -3.21697 RB11176_protein containing DUF442 6065605 6065144 NaN 0 NaN -3.19
1218 RB12046_conserved hypothetical protein 6497841 6497996 NaN 0 NaN -3.472031 RB1394_hypothetical protein 711994 711758 NaN 0 NaN -3.173872 RB4347_conserved hypothetical protein 2235735 2236205 NaN 0 NaN -3.323930 RB4429_conserved hypothetical protein 2275996 2276265 NaN 0 NaN -3.083974 RB4493_hypothetical protein 2302374 2302123 NaN 0 NaN -4.833980 RB4500_conserved hypothetical protein, secreted 2303645 2303908 NaN 0 NaN -4.115179 RB6403_hypothetical protein 3369939 3370853 NaN 0 NaN -3.371405 RB12348_FxsA cytoplasmic membrane protein 6645520 6644738 NaN -2.33 NaN -5.77751 RB11260_dnaK suppressor protein, 6107202 6106837 NaN -1.66 NaN -4.07
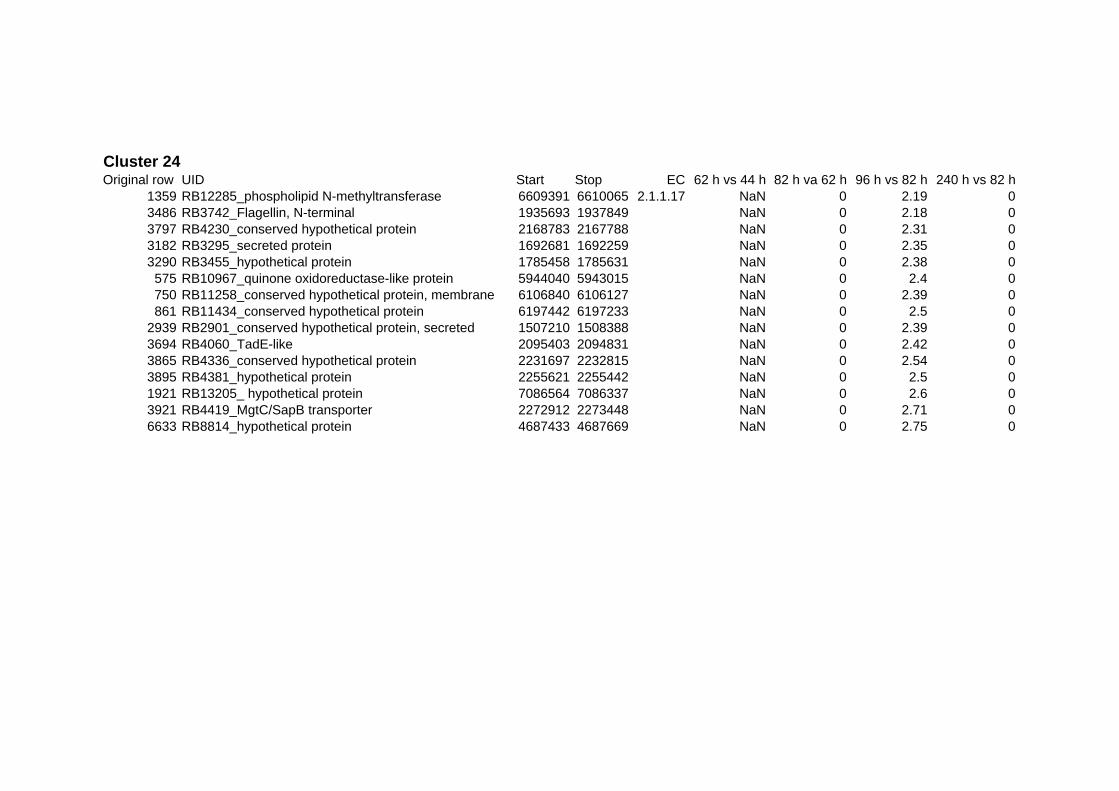
Cluster 24Original row UID Start Stop EC 62 h vs 44 h 82 h va 62 h 96 h vs 82 h 240 h vs 82 h
1359 RB12285_phospholipid N-methyltransferase 6609391 6610065 2.1.1.17 NaN 0 2.19 03486 RB3742_Flagellin, N-terminal 1935693 1937849 NaN 0 2.18 03797 RB4230_conserved hypothetical protein 2168783 2167788 NaN 0 2.31 03182 RB3295_secreted protein 1692681 1692259 NaN 0 2.35 03290 RB3455_hypothetical protein 1785458 1785631 NaN 0 2.38 0575 RB10967_quinone oxidoreductase-like protein 5944040 5943015 NaN 0 2.4 0750 RB11258_conserved hypothetical protein, membrane 6106840 6106127 NaN 0 2.39 0861 RB11434_conserved hypothetical protein 6197442 6197233 NaN 0 2.5 0
2939 RB2901_conserved hypothetical protein, secreted 1507210 1508388 NaN 0 2.39 03694 RB4060_TadE-like 2095403 2094831 NaN 0 2.42 03865 RB4336_conserved hypothetical protein 2231697 2232815 NaN 0 2.54 03895 RB4381_hypothetical protein 2255621 2255442 NaN 0 2.5 01921 RB13205_ hypothetical protein 7086564 7086337 NaN 0 2.6 03921 RB4419_MgtC/SapB transporter 2272912 2273448 NaN 0 2.71 06633 RB8814_hypothetical protein 4687433 4687669 NaN 0 2.75 0

Cluster 25Original row UID Start Stop EC 62 h vs 44 h 82 h va 62 h 96 h vs 82 h 240 h vs 82 h
6721 RB8959_hypothetical protein 4777121 4775133 0 1.54 1.77 0513 RB10867_hypothetical protein 5899359 5899982 0 0 1.71 0559 RB10939_conserved hypothetical protein 5934628 5934816 NaN 0 1.69 0609 RB11021_ATP-binding protein of ABC transporter 5975385 5974498 NaN 0 1.6 0696 RB11174_metallo-beta-lactamase family protein 6065046 6063637 NaN 0 1.63 0889 RB11488_oxidoreductase, short-chain dehydrogenase/reductase family 6213893 6214690 0 1.61 1.74 0893 RB11492_conserved hypothetical protein, membrane 6216103 6216390 NaN 0 1.64 0902 RB11505_conserved hypothetical protein, secreted 6221829 6221230 0 0 1.68 0
1711 RB12837_conserved hypothetical protein 6921732 6921929 0 0 1.92 02516 RB2244_Glutathione peroxidase 1178675 1179253 1.11.1.12 0 0 1.85 03061 RB309_Magnesium chelatase, ChlI subunit 156441 154993 6.6.1.1 0 0 1.94 03183 RB3298_hypothetical protein 1692797 1692678 0 0 2.24 03689 RB4051_hypothetical protein 2092752 2092210 NaN 0 1.68 03896 RB4382_Diaminopimelate decarboxylase 2257285 2255618 4.1.1.20 -1.47 0 1.53 03909 RB4399_zinc-binding oxidoreductase 2265237 2264251 0 0 2.02 03925 RB4423_cytidine and deoxycytidylate deaminase family protein 2275085 2274588 NaN 0 1.72 03934 RB4436_membrane protein 2277820 2278092 0 0 1.84 03951 RB4460_conserved hypothetical protein, membrane 2289696 2288428 NaN 0 1.51 03964 RB4478_membrane protein 2296122 2296640 0 0 2.3 03970 RB4487_two-component system response regulator 2299859 2301403 0 0 1.93 04428 RB5173_hypothetical protein 2660933 2660799 0 0 2.75 04490 RB5266_hypothetical protein 2715721 2715584 0 0 1.6 04629 RB548_1,4-alpha-glucan branching enzyme 306230 304329 2.4.1.18 NaN 0 1.51 05659 RB7178_hypothetical protein 3806924 3807061 NaN 0 1.7 06888 RB9261_conserved hypothetical protein 4952589 4953827 NaN 0 1.63 06965 RB9386_secreted protein 5034937 5033729 NaN 0 1.59 NaN7007 RB9460_hypothetical protein 5078860 5078621 0 0 1.93 01922 RB13207_secreted protein 7086761 7086585 0 0 3.98 02517 RB2247_conserved hypothetical protein containing YTV domains 1181238 1179499 0 0 3.12 0620 RB11041_secreted protein containing DUF1559 5987234 5988283 NaN 0 1.79 0753 RB11266_3',5'-cyclic-nucleotide phosphodiesterase 6108475 6109389 NaN 0 1.84 0
1555 RB1259_hypothetical protein 642452 642631 NaN 0 1.79 01707 RB12830_conserved hypothetical protein 6920508 6920200 NaN 0 1.83 02255 RB1787_hypothetical protein 925904 925686 NaN 0 1.88 03913 RB4404_oxidoreductase, short chain dehydrogenase/reductase family 2265539 2266447 NaN 0 1.82 05097 RB6276_Histone-like bacterial DNA-binding protein 3297504 3297187 NaN 0 1.82 NaN7241 RB9852_RNA polymerase ECF-type sigma factor 5304982 5305653 NaN 0 1.94 07268 RB9897_hypothetical protein 5333773 5333549 NaN 0 1.82 0196 RB10326_sulfate permease family protein 5580833 5582599 NaN 0 1.84 0290 RB10483_conserved hypothetical protein 5685437 5685838 NaN 0 1.84 0681 RB11150_NADH:flavin oxidoreductase/NADH oxidase 6046862 6048019 NaN 0 1.94 0
1412 RB12362_bacterioferritin comigratory protein 6653786 6652962 NaN 0 2.03 02616 RB2386_RND efflux membrane fusion protein 1244320 1242734 NaN 0 1.98 06168 RB8011_secreted protein containing DUF1571 4282298 4281114 NaN 0 2.06 06951 RB9360_Protein of unknown function, UPF0027 5015563 5014031 NaN 0 2.01 0132 RB10209_H+-transporting two-sector ATPase, C subunit 5515135 5515467 3.6.3.14 NaN 0 2.01 0574 RB10966_Protein of unknown function, UPF0057 5942934 5942752 NaN 0 2.02 0
3446 RB3686_oxidoreductase, zinc-binding 1910523 1909540 NaN 0 2.12 03968 RB4484_Peptidase M20 2297267 2298565 3.-.-.- NaN 0 2.16 0
Related Documents











