Accepted by J. Nekola: 22 Sep. 2014; published: 3 Oct. 2014 ZOOTAXA ISSN 1175-5326 (print edition) ISSN 1175-5334 (online edition) Copyright © 2014 Magnolia Press Zootaxa 3869 (5): 585–593 www.mapress.com/zootaxa/ 585 Correspondence http://dx.doi.org/10.11646/zootaxa.3869.5.9 http://zoobank.org/urn:lsid:zoobank.org:pub:58477926-D081-4692-8378-53AD8DEBF8FF A taxonomic revision of two local endemic Radix spp. (Gastropoda: Lymnaeidae) from Khodutka geothermal area, Kamchatka, Russian Far East IVAN BOLOTOV 1, 3 , YULIA BESPALAYA 1 , OLGA AKSENOVA 1 , ANDREY AKSENOV 1, 2 , NIKITA BOLOTOV 1, 2 , MIKHAIL GOFAROV 1, 2 , ALEXANDER KONDAKOV 1 , INGA PALTSER 1 & ILYA VIKHREV 1 1 Institute of Ecological Problems of the North, the Ural Branch of Russian Academy of Sciences, Severnaya Dvina Emb. 23, 163000 Arkhangelsk, Russian Federation 2 Northern (Arctic) Federal University, Severnaya Dvina Emb. 17, 163002 Arkhangelsk, Russian Federation 3 Corresponding author. E-mail: [email protected] Introduction Khodutka geothermal area is located near Khodutka and Priemysh volcanoes and is one of the largest geothermal areas of the Kamchatka Peninsula. Vakin (2003) described geological, geochemical and geothermic conditions of this geothermal area in detail. The main thermal water sources have temperatures up to 87°C and a discharge of approximately 150 l×sec. -1 are flows out into the warm lake with dimensions of ca. 250 m length and 80 m width. This warm river is ca. 20 m in width beginning from the lake and flows to the Bolshaya Khodutka River basin. Two local endemic Radix species were described from this geothermal area, especially Lymnaea (Radix) hadutkae Kruglov & Starobogatov, 1989 and L. (R.) thermokamtschatica Kruglov & Starobogatov, 1989 (Kruglov & Starobogatov 1989, 1993; Kruglov 2005). These species were separated using proportions of shell and reproductive system (Kruglov & Starobogatov 1989, 1993; Starobogatov et al. 2004). According to the diagnosis, L. (R.) hadutkae differs in the ear-shape shell, a form of the provaginal duct with cylindrical distal part and conical proximal part, and larger value of the index of the copulatory apparatus (ICA: proportion of the preputium to phallotheca is 1.27) from other species within the section Thermoradix Kruglov & Starobogatov, 1989. The last whorl is large, 0.86–0.89 of the shell height; an excess of the last whorl over upper margin of the aperture is 0.15–0.16 of the aperture height. L. (R.) thermokamtschatica has the cylindrical provaginal duct, relatively short bursa duct (1.5X longer than the bursa copulatrix diameter) and very long phallotheca (ICA is 0.77). The last whorl is large, 0.85–0.87 of the shell height; an excess of the last whorl over upper margin of the aperture is 0.2–0.25 of the aperture height. In accordance with an identification key (Starobogatov et al. 2004), the main diagnostic feature is an excess of the last whorl over the shell aperture, which has ≤0.78 and ≥0.80 of the penultimate whorl width in the first and second species, respectively. In the present paper, we revised these taxa using newly collected topotypes and additional Radix spp. specimens from other areas of the Russian Far East. During field expeditions in 2010–2013, lymnaeid snails were collected from eight localities on the Russian Far East, including the Khodutka geothermal area. Sampled specimens were preserved in 96 % ethanol. Materials are deposited in the Biological Museum of the Institute of Ecological Problems of the North (INEP), the Ural Branch of the Russian Academy of Sciences, Arkhangelsk, Russia. Species identification was performed using shell morphology, soft body anatomy and molecular data. Anatomical and morphological details were photographed using a stereomicroscope (Leica M165C, Leica Microsystems, Germany). The type specimens of Lymnaea (Radix) hadutkae Kruglov & Starobogatov, 1989 and L. (R.) thermokamtschatica Kruglov & Starobogatov, 1989 were studied in the collection of the Zoological Institute of the Russian Academy of Sciences, St. Petersburg, Russian Federation (ZIN). Molecular analysis was performed in the Institute of Ecological Problems of the North of the Ural Branch of Russian Academy of Sciences (purification and PCR) and the Engelhardt Institute of Molecular Biology of Russian Academy of Sciences (sequencing on ABI PRISM 3730). We analyzed Radix spp. samples from Khodutka area, as

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ZOOTAXA
ISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2014 Magnolia Press
Zootaxa 3869 (5): 585–593
www.mapress.com/zootaxa/Correspondence
http://dx.doi.org/10.11646/zootaxa.3869.5.9
http://zoobank.org/urn:lsid:zoobank.org:pub:58477926-D081-4692-8378-53AD8DEBF8FF
A taxonomic revision of two local endemic Radix spp. (Gastropoda: Lymnaeidae)
from Khodutka geothermal area, Kamchatka, Russian Far East
IVAN BOLOTOV1, 3, YULIA BESPALAYA1, OLGA AKSENOVA1, ANDREY AKSENOV1, 2,
NIKITA BOLOTOV1, 2, MIKHAIL GOFAROV1, 2, ALEXANDER KONDAKOV1,
INGA PALTSER1 & ILYA VIKHREV1
1Institute of Ecological Problems of the North, the Ural Branch of Russian Academy of Sciences, Severnaya Dvina Emb. 23, 163000
Arkhangelsk, Russian Federation2Northern (Arctic) Federal University, Severnaya Dvina Emb. 17, 163002 Arkhangelsk, Russian Federation3Corresponding author. E-mail: [email protected]
Introduction
Khodutka geothermal area is located near Khodutka and Priemysh volcanoes and is one of the largest geothermal
areas of the Kamchatka Peninsula. Vakin (2003) described geological, geochemical and geothermic conditions of
this geothermal area in detail. The main thermal water sources have temperatures up to 87°C and a discharge of
approximately 150 l×sec.-1 are flows out into the warm lake with dimensions of ca. 250 m length and 80 m width.
This warm river is ca. 20 m in width beginning from the lake and flows to the Bolshaya Khodutka River basin. Two
local endemic Radix species were described from this geothermal area, especially Lymnaea (Radix) hadutkae
Kruglov & Starobogatov, 1989 and L. (R.) thermokamtschatica Kruglov & Starobogatov, 1989 (Kruglov &
Starobogatov 1989, 1993; Kruglov 2005). These species were separated using proportions of shell and reproductive
system (Kruglov & Starobogatov 1989, 1993; Starobogatov et al. 2004). According to the diagnosis, L. (R.)
hadutkae differs in the ear-shape shell, a form of the provaginal duct with cylindrical distal part and conical
proximal part, and larger value of the index of the copulatory apparatus (ICA: proportion of the preputium to
phallotheca is 1.27) from other species within the section Thermoradix Kruglov & Starobogatov, 1989. The last
whorl is large, 0.86–0.89 of the shell height; an excess of the last whorl over upper margin of the aperture is
0.15–0.16 of the aperture height. L. (R.) thermokamtschatica has the cylindrical provaginal duct, relatively short
bursa duct (1.5X longer than the bursa copulatrix diameter) and very long phallotheca (ICA is 0.77). The last whorl
is large, 0.85–0.87 of the shell height; an excess of the last whorl over upper margin of the aperture is 0.2–0.25 of
the aperture height. In accordance with an identification key (Starobogatov et al. 2004), the main diagnostic feature
is an excess of the last whorl over the shell aperture, which has ≤0.78 and ≥0.80 of the penultimate whorl width in
the first and second species, respectively. In the present paper, we revised these taxa using newly collected
topotypes and additional Radix spp. specimens from other areas of the Russian Far East.
During field expeditions in 2010–2013, lymnaeid snails were collected from eight localities on the Russian Far
East, including the Khodutka geothermal area. Sampled specimens were preserved in 96 % ethanol. Materials are
deposited in the Biological Museum of the Institute of Ecological Problems of the North (INEP), the Ural Branch
of the Russian Academy of Sciences, Arkhangelsk, Russia. Species identification was performed using shell
morphology, soft body anatomy and molecular data. Anatomical and morphological details were photographed
using a stereomicroscope (Leica M165C, Leica Microsystems, Germany). The type specimens of Lymnaea (Radix)
hadutkae Kruglov & Starobogatov, 1989 and L. (R.) thermokamtschatica Kruglov & Starobogatov, 1989 were
studied in the collection of the Zoological Institute of the Russian Academy of Sciences, St. Petersburg, Russian
Federation (ZIN).
Molecular analysis was performed in the Institute of Ecological Problems of the North of the Ural Branch of
Russian Academy of Sciences (purification and PCR) and the Engelhardt Institute of Molecular Biology of Russian
Academy of Sciences (sequencing on ABI PRISM 3730). We analyzed Radix spp. samples from Khodutka area, as
Accepted by J. Nekola: 22 Sep. 2014; published: 3 Oct. 2014 585

well as from other Far Eastern localities (Appendix 1). We extracted genomic DNA from individual snails using
the DiatomTM DNA Prep 200 reagents kit (IsoGene Lab., Moscow, Russia). In the present study, we used data
inferred from the mitochondrial cytochrome oxidase subunit I (COI) gene, because DNA barcoding provides an
effective method for species-level identification and is widely accepted for an integrative taxonomy of Radix spp.
(Albrecht et al. 2008; Glöer & Pešić 2008). However, the molecular analyses of the nuclear and mitochondrial
markers are often incongruent (Schniebs et al. 2012) which reveals a limited application of a single mtDNA
marker, which may lead to over-estimation of species diversity. A total 657 bp of the COI gene were amplified and
sequenced using primers LCO1490 and HCO 2198 (Folmer et al. 1994). Additional sequences were obtained from
NCBI’s Genbank (Appendix 2). The alignment of COI sequences was performed using the CrustalW algorithm
implemented in MEGA6 (Tamura et al. 2013). For phylogenetic analyses, each sequence was trimmed, leaving a
588 bp-long COI fragment, according to a minimal length of a sequence in the aligned dataset. The analysis was
performed using 22 unique COI haplotypes, including Galba truncatula and Lymnaea palustris as outgroups. The
sequence dataset was collapsed into haplotypes using an online fasta sequence toolbox (FaBox v1.41, http://users-
birc.au.dk/biopv/php/fabox). The best model of sequence evolution is HKY+G as suggested with respect to the
corrected Akaike Information Criterion of MEGA6 (Tamura et al. 2013). Phylogenetic relationships were
reconstructed based on Bayesian inference implemented in MrBayes version 3.2.2 (Ronquist et al. 2012) using the
following parameters: mcmc ngen = 10,000,000, nchains = 4, samplefreq = 100, temp = 0.1; 25% of the sampled
trees were discarded as burn-in (pre-convergence part). Convergence of the MCMC chains to the stationary
distribution was checked visually based on the plotted posterior estimates using a MCMC trace analysis tool
(Tracer v1.6, http://beast.bio.ed.ac.uk/software/tracer). The effective sample size (ESS) value for each parameter
sampled from the MCMC analysis was always recorded >300. The combined set of trees showed a smooth
frequency plot. We conducted two independent runs of a Bayesian analysis using each outgroup in order to test
their influence on tree topology.
The diagnoses of two endemics from the Khodutka area are not clear, because all used diagnostic features
show a relatively high interspecific variability within the genus Radix (Glöer & Pešić 2008). The difference
between values of an excess of the last whorl over the shell aperture (Kruglov & Starobogatov 1989; Starobogatov
et al. 2004; Kruglov 2005) is very small and cannot be used as a diagnostic feature because of a high intraspecific
shell variability among R. auricularia populations due to environmental conditions (Stift et al. 2004). Studied
specimens from the Khodutka area belong to R. auricularia based on morphological and anatomical characters
(Table 1 and Figs. 1–3) and molecular data (Appendix 1 and Fig. 5).
Bayesian analysis reveals that R. auricularia is a monophyletic taxon across Eurasia (Bayesian posterior
probabilities (BPP) 1.00), which includes several highly divergent mitochondrial clades (Fig. 5). Two haplotypes
from the Khodutka geothermal area belong to a separate clade, which is well supported (BPP 0.96), and includes
haplotypes from other areas of the Russian Far East, as well as from the Middle East (Tajikistan). The topology of
R. auricularia clade and position of the Khodutka haplotypes remain stable and robust regardless of outgroup. The
mean COI p-distance among haplotypes that belong to this clade, calculated using MEGA6 software (Tamura et al.
2013), is 1.1±0.3 % (0.2–2.0 %).
Taxonomic account
Genus Radix Montfort, 1810
Radix auricularia (Linnaeus, 1758)
(Figs. 1–7, Table 1)
Lymnaea (Radix) hadutkae Kruglov & Starobogatov, 1989 syn. n.: 22, figs. 1.12 & 2.7; Kruglov & Starobogatov, 1993: 85, fig.
11D
Lymnaea (Radix) thermokamtschatica Kruglov & Starobogatov, 1989 syn. n.: 22, figs. 1.9 & 2.9; Kruglov & Starobogatov,
1993: 85, fig. 11E
BOLOTOV ET AL.586 · Zootaxa 3869 (5) © 2014 Magnolia Press

FIGURES 1–7. Radix auricularia (Linnaeus, 1758) from Russian Far East. Figs. 1-4. Shell and proximal part of the female
genitalia. Scale bar 2 mm. Fig. 1. Holotype of Lymnaea (R.) hadutkae Kruglov & Starobogatov, 1989 syn. n. (shell photo: M.V.
Vinarski; genitalia redrawn from the protologue, Fig. 2.7). Fig. 2. Holotype of L. (R.) thermokamtschatica Kruglov &
Starobogatov, 1989 syn. n. (shell photo: M.V. Vinarski; genitalia redrawn from the protologue, Fig. 2.9). Fig. 3. Newly
collected specimen from the Khodutka geothermal area, Kamchatka (photos: O.V. Aksenova). Fig. 4. Specimen from
Malkinskie hot springs, Kamchatka (photos: O.V. Aksenova). Fig. 5. Bayesian phylogram of haplotypes based on
mitochondrial COI gene dataset (Appendixes 1 & 2). The scale bar indicates the branch length. Asterisks: Posterior
probabilities ≥0.95; other significant node support values are mentioned in the figure. Haplotypes exclusively from NCBI’s
Genbank are marked with circumflex accents (˄). The well-supported clade with two haplotypes from the Khodutka
geothermal area is highlighted in blue. Fig. 6. General view of the Khodutka warm lake. Fig. 7. Map of observed localities on
the Russian Far East. The red circles represent species records. Locality codes are given in Appendix 1. The digital elevation
model and other layers of the map were added from Esri Data & Maps 10 dataset (Map: M.Yu. Gofarov).
Zootaxa 3869 (5) © 2014 Magnolia Press · 587RADIX SPP. FROM KHODUTKA GEOTHERMAL AREA
Material examined. Holotype of Lymnaea (Radix) hadutkae (ZIN), Kamchatka: “Khodutkinskiy hot spring,
gradient t° = 15–41°[C], opt. ~ 25–27°[C], 8.ix.1979, N.N. Khmeleva leg.” (no. 510–1982). Holotype of Lymnaea
(Radix) thermokamtschatica (ZIN), Kamchatka: “Khodutkinskiy hot spring, gradient t° = 15–41°[C], opt. ~
25–27°[C], 8.ix.1979, N.N. Khmeleva leg.” (no. 510–1982). Topotypes (INEP), 31 specimens, Kamchatka:
Khodutka geothermal area, warm lake and warm river source, 6th June 2013, Aksenova, Bespalaya & Bolotov leg.
(INEP). Kamchatka: 6 specimens, Paratunskie hot springs, 8th June 2013, Aksenova, Bespalaya & Bolotov leg.; 7
specimens, Malkinskie hot springs, 3rd July 2012, Bespalaya, Bolotov & Vikhrev leg.; 4 specimens, Plotnikova
River (Bolshaya River Basin), 3rd July 2012, Bespalaya, Bolotov & Vikhrev leg. (INEP). Kurile Archipelago: 9
specimens, Kunashir Island, Aliger Lake, 27th July 2011, Bespalaya & Aksenova leg. (INEP). Sakhalin Island: 2
specimens, Tym River, 14th June 2012, Bespalaya, Bolotov & Vikhrev leg. (INEP). Primorye: 3 specimens, a small
pond near Razdolnaya River, 25th June 2012, Bespalaya, Bolotov & Vikhrev leg; 4 specimens, Ussuri River (Amur
River Basin), 20th June 2012, Bespalaya, Bolotov & Vikhrev leg. (INEP). Locality coordinates are given in
Appendix 1.
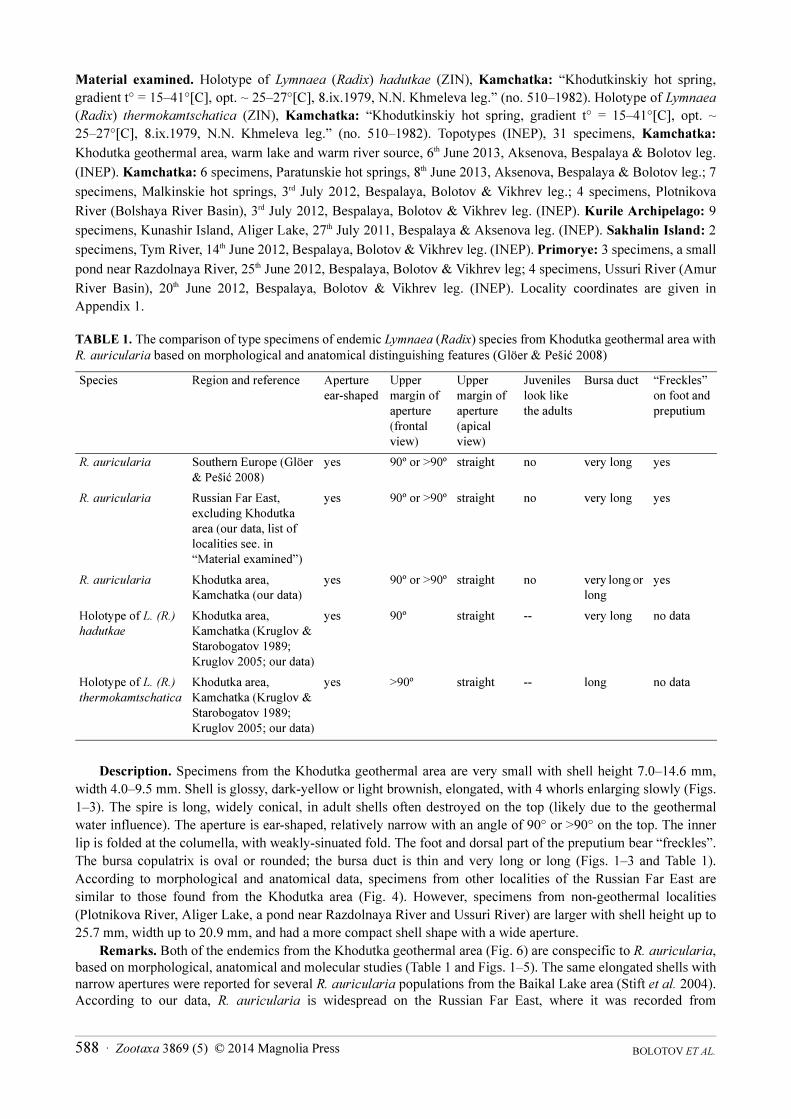
TABLE 1. The comparison of type specimens of endemic Lymnaea (Radix) species from Khodutka geothermal area with
R. auricularia based on morphological and anatomical distinguishing features (Glöer & Pešić 2008)
Description. Specimens from the Khodutka geothermal area are very small with shell height 7.0–14.6 mm,
width 4.0–9.5 mm. Shell is glossy, dark-yellow or light brownish, elongated, with 4 whorls enlarging slowly (Figs.
1–3). The spire is long, widely conical, in adult shells often destroyed on the top (likely due to the geothermal
water influence). The aperture is ear-shaped, relatively narrow with an angle of 90° or >90° on the top. The inner
lip is folded at the columella, with weakly-sinuated fold. The foot and dorsal part of the preputium bear “freckles”.
The bursa copulatrix is oval or rounded; the bursa duct is thin and very long or long (Figs. 1–3 and Table 1).
According to morphological and anatomical data, specimens from other localities of the Russian Far East are
similar to those found from the Khodutka area (Fig. 4). However, specimens from non-geothermal localities
(Plotnikova River, Aliger Lake, a pond near Razdolnaya River and Ussuri River) are larger with shell height up to
25.7 mm, width up to 20.9 mm, and had a more compact shell shape with a wide aperture.
Remarks. Both of the endemics from the Khodutka geothermal area (Fig. 6) are conspecific to R. auricularia,
based on morphological, anatomical and molecular studies (Table 1 and Figs. 1–5). The same elongated shells with
narrow apertures were reported for several R. auricularia populations from the Baikal Lake area (Stift et al. 2004).
According to our data, R. auricularia is widespread on the Russian Far East, where it was recorded from
Species Region and reference Aperture
ear-shaped
Upper
margin of
aperture
(frontal
view)
Upper
margin of
aperture
(apical
view)
Juveniles
look like
the adults
Bursa duct “Freckles”
on foot and
preputium
R. auricularia Southern Europe (Glöer
& Pešić 2008)
yes 90º or >90º straight no very long yes
R. auricularia Russian Far East,
excluding Khodutka
area (our data, list of
localities see. in
“Material examined”)
yes 90º or >90º straight no very long yes
R. auricularia Khodutka area,
Kamchatka (our data)
yes 90º or >90º straight no very long or
long
yes
Holotype of L. (R.)
hadutkae
Khodutka area,
Kamchatka (Kruglov &
Starobogatov 1989;
Kruglov 2005; our data)
yes 90º straight -- very long no data
Holotype of L. (R.)
thermokamtschatica
Khodutka area,
Kamchatka (Kruglov &
Starobogatov 1989;
Kruglov 2005; our data)
yes >90º straight -- long no data
BOLOTOV ET AL.588 · Zootaxa 3869 (5) © 2014 Magnolia Press

Kamchatka, Primorye, Sakhalin, and Kuriles (Fig. 7). Zhadin (1952) reported that this species is widespread on the
Pacific Basin, including Kamchatka, Amur Basin and Primorye, which is in agreement with our records. However,
Kruglov & Starobogatov (1993) noted that R. auricularia ranged in Europe, the western part of the Middle East
and Siberia, but not mentioned in the species records from the Far East. Thereafter, Starobogatov et al. (2004)
again reported that species ranged on the Far East, where it was recorded in rivers of the Pacific coast from
Kamchatka to Primorye. Meanwhile, Kruglov & Starobogatov (1989, 1993), Starobogatov et al. (2004) and
Kruglov (2005) listed many local endemic Radix species for different areas of the Russian Far East, which belong
to the sections Ussuriradix Kruglov & Starobogatov, 1989, Nipponiradix Kruglov & Starobogatov, 1989 and
Okhotiradix Kruglov & Starobogatov, 1989. Taxonomic position of these taxa is uncertain and they need in a
current revision based on mitochondrial and nuclear molecular markers.
Acknowledgements
This study was supported by grants from the President of Russia (no. MD-6465.2014.5) and the Ural Branch of
Russian Academy of Sciences (no. 12-M-45-2062 and 12-P-5-1014). We are grateful to Dr. M.V. Vinarski
(Museum of Siberian Aquatic Molluscs, Russia) for providing the high-resolution photos of type specimens of
Lymnaea (Radix) hadutkae and L. (R.) thermokamtschatica. We also are thankful to Dr. P.V. Kijashko and L.L.
Jarochnovich (ZIN, Russia) for their assistance in studies of lymnaeids from the ZIN collection. Special thanks go
to Dr. N. Friberg (National Environmental Research Institute, Oslo, Norway and University of Aarhus, Denmark),
Dr. B.O.L. Demars (The James Hutton Institute, UK), Mr. V.V. Komarov (Volcanoes of Kamchatka Nature Park,
Russia), Mr. O.N. Bespaliy (Arkhangelsk, Russia) and Mr. E.P. Dekin (Elisovo, Russia) for their great help during
our field works. We also thank K. Schniebs (Senckenberg Natural History Collections Dresden, Germany) and an
anonymous reviewer for improvements to the manuscript.
References
Albrecht, C., Wolff, C., Glöer, P. & Wilke, T. (2008) Concurrent evolution of ancient sister lakes and sister species: the
freshwater gastropod genus Radix in lakes Ohrid and Prespa. Hydrobiologia, 615, 157–167.
http://dx.doi.org/10.1007/s10750-008-9555-1
Folmer, O., Black, M., Hoeh, W., Lutz, R. & Vrijenhoek, R. (1994) DNA primers for amplification of mitochondrial
cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology, 3,
294–299.
Glöer, P. & Pešić, V. (2008) Radix skutaris n. sp., a new species from Montenegro (Gastropoda: Lymnaeidae). Mollusca, 26 (1),
83–88.
Kruglov, N.D. (2005) Molluscs of the family Lymnaeidae (Gastropoda: Pulmonata) in Europe and Northern Asia. SGPU
Publishing, Smolensk, 507 pp. [in Russian]
Kruglov, N.D. & Starobogatov, Ya.I. (1989) Morphology and taxonomy of the molluscs from the subgenus Radix of the genus
Lymnaea (Gastropoda, Pulmonata, Lymnaeidae) from Siberia and Far East of the USSR. Zoologicheskii Zhurnal, 68 (5),
17–30. [in Russian]
Kruglov, N.D. & Starobogatov, Ya.I. (1993) Guide to recent molluscs of northern Eurasia. 3. Annotated and illustrated
catalogue of species of the family Lymnaeidae (Gastropoda Pulmonata Lymnaeiformes) of Palaearctic and adjacent river
drainage areas. Part 1. Ruthenica, 3 (1), 65–92.
Ronquist, F., Teslenko, M., van der Mark, P., Ayres, D.L., Darling, A., Höhna, S., Larget, B., Liu, L., Suchard, M.A. &
Huelsenbeck, J.P. (2012) MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model
space. Systematic Biology, 61 (3), 539–542.
http://dx.doi.org/10.1093/sysbio/sys029
Schniebs, K., Glöer, P., Georgiev, D. & Hundsdoerfer, A.K. (2012) First record of Stagnicola montenegrinus Glöer & Pešić,
2009 (Mollusca: Gastropoda: Lymnaeidae) in Bulgaria and its taxonomic relationship to other European lymnaeids based
on molecular analysis. North-Western Journal of Zoology, 8 (1), 164–171.
Starobogatov, Ya.I., Prozorova, L.A., Bogatov, V.V. & Sayenko, E.M. (2004) Molluscs. In: S.J. Tsalolikhin (Ed.), Key to
freshwater invertebrates of Russia and adjacent lands (Vol. 6. Molluscs, Polychaetes, Nemerteans). Nauka, St-Petersburg,
pp. 9–492. [in Russian]
Stift, M., Michel, E., Sitnikova, T.Y., Mamonova, E.Y. & Sherbakov, D.Y. (2004) Palaearctic gastropod gains a foothold in the
dominion of endemics: range expansion and morphological change of Lymnaea (Radix) auricularia in Lake Baikal.
Hydrobiologia, 513 (1–3), 101–108.
http://dx.doi.org/10.1023/b:hydr.0000018175.37771.d6
Zootaxa 3869 (5) © 2014 Magnolia Press · 589RADIX SPP. FROM KHODUTKA GEOTHERMAL AREA
Tamura, K., Stecher, G., Peterson, D., Filipski, A. & Kumar, S. (2013) MEGA6: Molecular Evolutionary Genetics Analysis
version 6.0. Molecular Biology and Evolution, 30, 2725–2729.
http://dx.doi.org/10.1093/molbev/mst197
Vakin, E.A. (2003) Khodutkinskie thermal springs. Vestnik KRAUNZ. Series of the Earth Science, 2, 56–69. [in Russian]
Zhadin, V.I. (1952) Molluscs of fresh and brackish water of the USSR. Identification Guides on the Fauna of the USSR, 46,
1–374. [in Russian]
APPENDIX 1. List of studied Radix auricularia specimens from Russian Far East. Materials are deposited in the
Biological Museum of Institute of Ecological Problems of the North (INEP), the Ural Branch of Russian Academy
of Sciences, Arkhangelsk, Russian Federation.
COI NCBI’s
GenBank acc.
no.
Haplotype
Code
Specimen
Voucher
Locality Habitat type Latitude Longitude
KM067596 PAR1 INEP-378.1 Russia, Kamchatka, Paratunskie
hot springs
Geothermal 52°49'26" 158°09'48"
KM067598 PAR1 INEP-393.1 Russia, Kamchatka, Paratunskie
hot springs
Geothermal 52°49'26" 158°09'48"
KM067599 PAR1 INEP-386.2 Russia, Kamchatka, Paratunskie
hot springs
Geothermal 52°49'26" 158°09'48"
KM067600 PAR1 INEP-390.2 Russia, Kamchatka, Paratunskie
hot springs
Geothermal 52°49'26" 158°09'48"
KM067601 PAR1 INEP-398.2 Russia, Kamchatka, Paratunskie
hot springs
Non-
geothermal
52°49'26" 158°09'48"
KM067597 PAR1 INEP-379.1 Russia, Kamchatka, Paratunskie
hot springs
Geothermal 52°49'26" 158°09'48"
KM067602 KHO1 INEP-375.1 Russia, Kamchatka, Khodutka
geothermal area
Geothermal 52°06'27" 157°39'25"
KM067603 KHO1 INEP-385.2 Russia, Kamchatka, Khodutka
geothermal area
Geothermal 52°06'27" 157°39'25"
KM067604 KHO1 INEP-385.3 Russia, Kamchatka, Khodutka
geothermal area
Geothermal 52°06'27" 157°39'25"
KM067605 KHO1 INEP-491.1 Russia, Kamchatka, Khodutka
geothermal area
Geothermal 52°06'27" 157°39'25"
KM067606 KHO1 INEP-392.1 Russia, Kamchatka, Khodutka
geothermal area
Geothermal 52°06'27" 157°39'25"
KM111179 KHO2 INEP-491.4 Russia, Kamchatka, Khodutka
geothermal area
Geothermal 52°06'27" 157°39'25"
KM111180 KHO2 INEP-491.5 Russia, Kamchatka, Khodutka
geothermal area
Geothermal 52°06'27" 157°39'25"
KM067607 MAL1 INEP-117.1 Russia, Kamchatka, Malkinskie
hot springs
Geothermal 53°19'19" 157°32'14"
KM067608 MAL1 INEP-117.3 Russia, Kamchatka, Malkinskie
hot springs
Geothermal 53°19'19" 157°32'14"
KM067609 MAL1 INEP-119.3 Russia, Kamchatka, Malkinskie
hot springs
Geothermal 53°19'19" 157°32'14"
KM067610 MAL1 INEP-119.4 Russia, Kamchatka, Malkinskie
hot springs
Geothermal 53°19'19" 157°32'14"
KM067611 MAL1 INEP-119.5 Russia, Kamchatka, Malkinskie
hot springs
Geothermal 53°19'19" 157°32'14"
......continued on the next page
BOLOTOV ET AL.590 · Zootaxa 3869 (5) © 2014 Magnolia Press

APPENDIX 2. List of additional COI sequences of lymnaeid species obtained from NCBI’s GenBank including accession number,
taxon, specimen code, locality information and references.
*—These species were used as outgroups for Bayesian analysis.
**—Data source: (1) Bargues, M.D., Artigas, P., Khoubbane, M., Ortiz, P., Naquira, C. and Mas-Coma, S. (2011) Human and
animal fascioliasis vectors in Cajamarca, Peru: DNA assessment of Galba truncatula, Galba neotropica and Lymnaea
schirazensis and analysis of vector overlap implications (unpubl.); (2) International Barcode of Life (iBOL), Biodiversity
Institute of Ontario, University of Guelph, Canada (unpubl.); (3) Oheimb, P.V.v., Albrecht, C., Riedel, F., Du, L., Yang, J. et al.
(2011) Freshwater biogeography and limnological evolution of the Tibetan Plateau—insights from a plateau-wide distributed
gastropod taxon (Radix spp.). PLoS ONE, 6(10), e26307. doi:10.1371/journal.pone.0026307; (4) Vinarski, M. V., Schniebs, K.,
Glöer, P., & Hundsdoerfer, A. K. (2011) The taxonomic status and phylogenetic relationships of the genus Aenigmomphiscola
Kruglov and Starobogatov, 1981 (Gastropoda: Pulmonata: Lymnaeidae). Journal of Natural History, 45(33–34), 2049–2068.
doi: 10.1080/00222933.2011.574800; (5) Quintela, M., Johansson, M.P., Kristjansson, B.K., Barreiro, R., Laurila, A. (2014)
AFLPs and mitochondrial haplotypes reveal local adaptation to extreme thermal environments in a freshwater gastropod. PLoS
ONE, 9(7), e101821. doi:10.1371/journal.pone.0101821; (6) Albrecht, C., Wolff, C., Glöer, P., & Wilke, T. (2008) Concurrent
evolution of ancient sister lakes and sister species: the freshwater gastropod genus Radix in lakes Ohrid and Prespa.
Hydrobiologia, 615, 157–167. doi: 10.1007/978-1-4020-9582-5_11
APPENDIX 1. (continued)
COI NCBI’s
GenBank acc.
no.
Haplotype
Code
Specimen
Voucher
Locality Habitat type Latitude Longitude
KM067612 PLO1 INEP-231.1 Russia, Kamchatka, Plotnikova
River (Bolshaya Basin)
Non-
geothermal
53°06'14" 157°45'16"
KM067613 PLO1 INEP-231.3 Russia, Kamchatka, Plotnikova
River (Bolshaya Basin)
Non-
geothermal
53°06'14" 157°45'16"
KM067614 ALI1 INEP-241 Russia, Kunashir Island, Aliger
Lake
Non-
geothermal
44°02'50" 145°44'38"
KM067615 TYM1 INEP-498 Russia, Sakhalin Island, Tym
River
Non-
geothermal
50°54'45" 142°39'48"
KM067618 RAZ1 INEP-500 Russia, Primorye, small pond near
Razdolnaya River
Non-
geothermal
43°46'47" 131°57'38"
KM067616 USS1 INEP-499.1 Russia, Primorye, Ussuri River
(Amur Basin)
Non-
geothermal
44°04'52" 133°50'43"
KM067617 USS2 INEP-499.3 Russia, Primorye, Ussuri River
(Amur Basin)
Non-
geothermal
44°04'52" 133°50'43"
Haplotype
code
COI NCBI’s
GenBank acc. no.
Species Specimen
Voucher
Locality Data
source**
TRU1 HE610435 Galba truncatula* No data Peru, Cajamarca (1)
PAL1 GU680909 Lymnaea stagnalis* 09PROBE-
02278
Canada, Manitoba (2)
TAJ1 JN794513 Radix auricularia UGSB 7532 Tajikistan, Gorno-Badakhshan (3)
TIB1 JN794432 R. cf. auricularia (clade 1) UGSB 7400 China, Tibet (3)
GER1 FR797876 R. auricularia Moll S1332 Germany (4)
GER2 FR797877 R. auricularia Moll S1333 Germany (4)
GER3 FR797878 R. auricularia Moll S1313 Germany (4)
GER4 FR797879 R. auricularia Moll 53070 Germany (4)
BAL1 JN624792 R. balthica No data Iceland, Lake Mývatn (5)
REL1 EU818822 R. relicta No data Albania, Pole, Lake Ohrid (6)
AMP1 EU818837 R. ampla No data Austria, Salzburger Land, Lake
Wallersee
(6)
LAB1 EU818832 R. labiata No data Albania, Dibre, Cidhne, gorge of
Pr. i Setes
(6)
Zootaxa 3869 (5) © 2014 Magnolia Press · 591RADIX SPP. FROM KHODUTKA GEOTHERMAL AREA
Related Documents