A systematic approach to the recovery and identification of starches from carbonised deposits on ceramic vessels Hayley Saul a, * , Julie Wilson b , Carl P. Heron c , Aikaterini Glykou d , Sönke Hartz e , Oliver E. Craig a a BioArch, University of York, S-Block, Heslington, York, YO10 5DD, UK b Departments of Mathematics and Chemistry, University of York, Heslington, York, YO10 5DD, UK c Archaeological, Geographical and Environmental Sciences, University of Bradford, Bradford, BD7 1DP, UK d Graduate School ‘Human Development in Landscapes’, Institute of Prehistoric and Protohistoric Archaeology, University of Kiel, Germany e Stiftung Schleswig-Holsteinische Landesmuseen, Schlob Gottorf, D-24837 Schleswig, Germany article info Article history: Received 6 January 2011 Received in revised form 24 April 2012 Accepted 28 May 2012 Keywords: Starch Automated classification Transition to agriculture Wild plant foods Domestication Foodcrust Ceramic residues abstract Starch granules are being successfully recovered from an increasing range of artefacts. Here we present the recovery of starches from carbonised ceramic ‘foodcrusts’ from late Mesolithiceearly Neolithic residues at the site of Neustadt in northern Germany. A method for investigating background loading of residues with contaminant starches is proposed by comparing interior ‘foodcrusts’ versus exterior ‘sooting’, for the purposes of eliminating samples with insignificant quantities of grains from subsequent identification procedures. The classification of starches to plant taxon is traditionally achieved by manual observations and measurement of nominal and ratio morphological variables. Here, we present a method for the automated classification of granules, using software developed in-house. The results show that when multiple granules are considered, the species selected as modern reference examples can be classified to high levels of specificity. When applied to the archaeological samples we show that wild plant resources persist in importance across the transition to agriculture, with high proportions of granule forms consistent with acorn (Quercus sp.) occurring in all samples. Hazelnut (Corylus avellana) types are less well-represented suggesting it was not an important food in the context of pottery, and may have been over-represented in the repertoire of hunteregatherer resources. Cereals are not rep- resented in any of the samples, supporting the notion that their adoption may have been a slow process, occurring more gradually than for other domesticated foods, or that they were not initially processed in ceramic vessels. Ó 2012 Elsevier Ltd. All rights reserved. 1. Introduction Our understanding of the uses and values of plants in the past is limited in comparison to other foodstuffs, and this is especially true in prehistory. This is in large part because of the poor pres- ervation of macroscopic plant remains, the limited scope for bimolecular techniques to detect all but oily and waxy plant products (Charters et al., 1997; Evershed et al., 1991; Evershed, 2008), and also our inability to securely tie archaeobotanical remains in sediments with specific anthropogenic causes. Plant microfossils, such as pollen, starches and phytoliths, often preserve much better than other plant tissues and offer the best target for analysis. Recent methodological advances have led to the increased recovery and identification of plant microfossils from challenging substrates, such as dental calculus (Hardy et al., 2009; Henry and Piperno, 2008; Lalueza Fox and Péz-Pérez, 1994; Lalueza Fox et al., 1996), stone tool residues (Briuer, 1976; Kealhofer et al., 1999; Perry, 2004; Piperno and Holst, 1998; Piperno et al., 2000; 2004; Yang et al., 2009), coprolites (Horrocks, 2004, Horrocks et al., 2004, Reinhard and Danielson, 2005) and food residues associated with ceramic vessels (Boyd et al., 2008; Crowther, 2005; Staller and Thompson, 2002). Furthermore, there are now well established techniques for microfossil extraction from sediments that have been applied successfully (e.g. Balme and Beck, 2002; Horrocks, 2005, Horrocks and Nunn, 2007; Pearsall, 2002). The high temperatures to which carbonised deposits have been exposed can accelerate tissue degradation in plants making them more prone to subsequent organic degradation and enzymatic attack in soils (Haslam, 2004). However, the durability of many * Corresponding author. University of York, BioArch, S-Block, PO Box 373, Heslington, York YO10 5DD, UK. Tel.: þ44 1904 328806. E-mail addresses: [email protected], [email protected] (H. Saul). Contents lists available at SciVerse ScienceDirect Journal of Archaeological Science journal homepage: http://www.elsevier.com/locate/jas 0305-4403/$ e see front matter Ó 2012 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.jas.2012.05.033 Journal of Archaeological Science 39 (2012) 3483e3492

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

at SciVerse ScienceDirect
Journal of Archaeological Science 39 (2012) 3483e3492
Contents lists available
Journal of Archaeological Science
journal homepage: http : / /www.elsevier .com/locate/ jas
A systematic approach to the recovery and identification of starches fromcarbonised deposits on ceramic vessels
Hayley Saul a,*, Julie Wilson b, Carl P. Heron c, Aikaterini Glykou d, Sönke Hartz e, Oliver E. Craig a
aBioArch, University of York, S-Block, Heslington, York, YO10 5DD, UKbDepartments of Mathematics and Chemistry, University of York, Heslington, York, YO10 5DD, UKcArchaeological, Geographical and Environmental Sciences, University of Bradford, Bradford, BD7 1DP, UKdGraduate School ‘Human Development in Landscapes’, Institute of Prehistoric and Protohistoric Archaeology, University of Kiel, Germanye Stiftung Schleswig-Holsteinische Landesmuseen, Schlob Gottorf, D-24837 Schleswig, Germany
a r t i c l e i n f o
Article history:Received 6 January 2011Received in revised form24 April 2012Accepted 28 May 2012
Keywords:StarchAutomated classificationTransition to agricultureWild plant foodsDomesticationFoodcrustCeramic residues
* Corresponding author. University of York, BioAHeslington, York YO10 5DD, UK. Tel.: þ44 1904 3288
E-mail addresses: [email protected], hs140@hotm
0305-4403/$ e see front matter � 2012 Elsevier Ltd.http://dx.doi.org/10.1016/j.jas.2012.05.033
a b s t r a c t
Starch granules are being successfully recovered from an increasing range of artefacts. Here we presentthe recovery of starches from carbonised ceramic ‘foodcrusts’ from late Mesolithiceearly Neolithicresidues at the site of Neustadt in northern Germany. A method for investigating background loading ofresidues with contaminant starches is proposed by comparing interior ‘foodcrusts’ versus exterior‘sooting’, for the purposes of eliminating samples with insignificant quantities of grains from subsequentidentification procedures. The classification of starches to plant taxon is traditionally achieved by manualobservations and measurement of nominal and ratio morphological variables. Here, we present a methodfor the automated classification of granules, using software developed in-house. The results show thatwhen multiple granules are considered, the species selected as modern reference examples can beclassified to high levels of specificity. When applied to the archaeological samples we show that wildplant resources persist in importance across the transition to agriculture, with high proportions ofgranule forms consistent with acorn (Quercus sp.) occurring in all samples. Hazelnut (Corylus avellana)types are less well-represented suggesting it was not an important food in the context of pottery, andmay have been over-represented in the repertoire of hunteregatherer resources. Cereals are not rep-resented in any of the samples, supporting the notion that their adoption may have been a slow process,occurring more gradually than for other domesticated foods, or that they were not initially processed inceramic vessels.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Our understanding of the uses and values of plants in the pastis limited in comparison to other foodstuffs, and this is especiallytrue in prehistory. This is in large part because of the poor pres-ervation of macroscopic plant remains, the limited scope forbimolecular techniques to detect all but oily and waxy plantproducts (Charters et al., 1997; Evershed et al., 1991; Evershed,2008), and also our inability to securely tie archaeobotanicalremains in sediments with specific anthropogenic causes. Plantmicrofossils, such as pollen, starches and phytoliths, oftenpreserve much better than other plant tissues and offer the besttarget for analysis. Recent methodological advances have led to
rch, S-Block, PO Box 373,06.ail.co.uk (H. Saul).
All rights reserved.
the increased recovery and identification of plant microfossilsfrom challenging substrates, such as dental calculus (Hardy et al.,2009; Henry and Piperno, 2008; Lalueza Fox and Péz-Pérez, 1994;Lalueza Fox et al., 1996), stone tool residues (Briuer, 1976;Kealhofer et al., 1999; Perry, 2004; Piperno and Holst, 1998;Piperno et al., 2000; 2004; Yang et al., 2009), coprolites (Horrocks,2004, Horrocks et al., 2004, Reinhard and Danielson, 2005) andfood residues associated with ceramic vessels (Boyd et al., 2008;Crowther, 2005; Staller and Thompson, 2002). Furthermore, thereare now well established techniques for microfossil extractionfrom sediments that have been applied successfully (e.g. Balmeand Beck, 2002; Horrocks, 2005, Horrocks and Nunn, 2007;Pearsall, 2002).
The high temperatures to which carbonised deposits have beenexposed can accelerate tissue degradation in plants making themmore prone to subsequent organic degradation and enzymaticattack in soils (Haslam, 2004). However, the durability of many

H. Saul et al. / Journal of Archaeological Science 39 (2012) 3483e34923484
plant microfossils is attested by their recovery from heavilycarbonised deposits that have also undergone mechanical pro-cessing in some instances. For example, pollen has been recoveredfrom Late Neolithic British vessels leading to the suggestion thatthey were used for the ritual preparation of henbane (Hyoscyamusniger) (Barclay and Russel-White, 1993) or as mead containers dueto the presence of meadowsweet peaks (Filipendula ulmaria)(Barclay, 1983). Criticisms of pollen as a useful food indicator havestressed the very small proportions being considered as evidence,and the high mobility of the microfossil (Long et al., 1999, 2000).Phytoliths, with their much greater thermal durability, are havingan increasingly consistent role in the analysis of charred plantdeposits (Lusteck and Thompson, 2007). For example, at KebaraCave in Israel concentrations of phytoliths from wood and barkfuel could be used to confirm the locations of prehistoric hearths(Albert et al., 2000). A current limitation of phytolith research is itsapplication outside arid and tropical regions, where the rate ofevapotranspiration is high (Madella et al., 2009). Thus, its useful-ness in a temperate European context is only recently beingsystematically explored (Saul et al., forthcoming), and the taxo-nomic resolution at which it is possible to operate using phytolithsis unknown.
In this paper we aim to test a range of carbonised ceramicsurface deposits formed experimentally and from prehistoricsubmerged contexts in Northern Germany (dating from 4600 calBC to 3700 cal BC) to see if starches preserve and can be extracted.Carbonised deposits on the inside of cooking pots (Fig. 1) are foundin a wide range of archaeological contexts (e.g. Boudin et al., 2009;Craig, 2004, Craig et al., 2007; Crombé et al., 2002; Andersen andMalmros, 1984) and may provide a good micro-environment forstarch preservation if degradation rates are retarded when they arein association with artefacts, as has been suggested by Haslam(2004). Crucially, plant microfossils identified within the matrixof carbonised surface residues are likely to derive from vessel use(Crowther, 2005).
In addition to testing whether starch granules survive incarbonised food residues on ceramics, here we trial automatedclassification of extracted granules from these contexts usingimage analysis and pattern recognition of both wild and domes-ticated plant starch morphologies (Wilson et al., 2010). The studyaims to answer the following three questions: i) Do starchespreserve in carbonised deposits from prehistoric temperatewaterlogged sites? ii) Are these starches representative of the use
Fig. 1. A carbonised ‘foodcrust’ deposit on the interior rim of a funnel beaker vessel.
of the vessel? iii) Can the starches be identified to a plant foodsource?
1.1. Ancient starch analysis
Starch granules are semi-crystalline, water insoluble grains(Miles et al., 1985) that can account for 16e24% of the total weightof a storage root, tuber crop or other underground storage organ(Hoover, 2001). Starches are carbohydrate polymers deployed byplants into energy storage organs. Within the field of plantmicrofossil analysis, starches have long been known for theirpotential as an indicator of diet, as they display morphologicalvariation that is taxonomically significant (Torrence and Barton,2006). Starches have a huge range of molecular and granularstructures that bear on the physico-chemical properties of granules,and their susceptibility to taphonomic processes (Leach, 1965;Sivak and Preiss, 1998). A temperature range of 50e85 �C is oftencited as the point at which gelatinisation of the grains begins(Hoover, 2001; Tang et al., 2001; Torrence and Barton, 2006), andwhilst this is true in general, it simplifies the potential range ofresponses to processes like cooking (Gaillard and Bowler, 1987;Henry et al., 2009). Grain size is related to starch hydration(Torrence and Barton, 2006), with water molecules inhabiting thecrystalline regions of the amylopectin molecules (Ratnayake andJackson, 2006). Small granules generally have less swellingpower, or water-binding capacity, meaning gelatinisation occursrelatively quicker than for larger granules. However, a higheramylose content also lowers swelling capacity, buffering thegranules from gelatinisation (Frederiksson et al., 1998; Tester andMorrison, 1990), so in theory gelatinisation in granules witha high amylose:amylopectin ratio could bemore effectively offset inlarger granules than smaller ones.
Starch degradation and morphological alteration is a complexissue, and current knowledge advocates as much potential for re-order as for catastrophic disorder of granule micro-structure asa consequence of cooking (Ratnayake and Jackson, 2006; Zarilloet al., 2008). This is especially true considering we know so littleabout these features in most wild northern European species.Annealing (low temperature heating with low water content), aswell as heatemoisture treatment (low water content with highheat) are known cooking methods that increase gelatinisationtemperatures, and in the former case also reduce leaching andgranular swelling (Tester and Debon, 2000). A survival potential isalso posed by resistant starch which is resilient against hydrolysis,because starch is tightly packed in granules in a radial pattern andis relatively dehydrated (Sajilata et al., 2006). Experiments withcooked tubers showed that in samples with gelatinised starcha proportion of the grains were resistant grain varieties, andresisted all thermal alterations to morphology (Gott et al., 2006).
Microscopy of archaeological starch granules in Egyptian breadwas being conducted as early as 1905 (Samuel, 1996). The use ofstarches as dietary indicators from carbonised pottery foodcrusts isincreasing in microfossil research although applications toarchaeological problems have been limited, with NewWorld maize(Zea mays) agriculture and Australasian taro (Colocasia esculenta)domestication being the main areas of research. Pottery residueanalyses of starch first showed that maize cultivationwas practicedwidely on the North American Great Plains well before Europeancontact (Boyd et al., 2006). A follow-up study of foodcrust starchextended the impact of maizewithmuch earlier evidence from LateWoodland sites on the eastern Canadian Plains around 700 AD(Boyd et al., 2008). In New Ireland Lapita pottery from Papua NewGuinea, the co-occurrence of raphides with starches consistentwith the Aroid family has suggested the processing of taro(C. esculenta) c. 3300 BP (Crowther, 2005). Additionally, ground and

H. Saul et al. / Journal of Archaeological Science 39 (2012) 3483e3492 3485
charred archaeological cereal starches were detected from prehis-toric Greek and Bulgarian sites in recognisable forms (Valamotiet al., 2008) and almost completely carbonised cell structures ofwheat (Triticum sp.) were recovered fromViking Age bread remainsfrom Våstergården, Denmark (Hansson and Isaksson, 1994).
Archaeological starches have contributed to debates about theimportance of plant foods from contexts as early as the PolishPalaeolithic at c. 11,380 � 95 BP. In this case starches werediscovered in preserved parenchyma tissue (Kubiak-Martens,1996). Remarkably, starches have been reported to endure sincethe Pleistocene c. 180,000 years ago at Sai Island in northernSudan, in association with pounding implements (Van Peer et al.,2003). Documentation of granules such as those in cookedEgyptian breads, in unaltered forms (Samuel, 1996) shows that thecomplex physico-chemical properties of such diverse grainmorphologies means gelatinisation does not necessarily followfrom cooking. The preservation of starch granules in carboniseddeposits from such diverse contexts and climates as the arid andsemi-arid climes of Australia (Atchinson and Fullager, 1998; Parrand Carter, 2003), tropical regions like Papua New Guinea(Barton and White, 1993; Therin et al., 1999), and temperatelocations like northern Europe (Hardy, 2004, 2007) is therefore ofgreat interest.
2. Materials and methods
2.1. Reference plant materials
Twelve modern reference species were selected mainly on thebasis of their occurrence as macrofossils on later prehistoric sites innorthern Europe, their availability, and their potential value asa starchy food in prehistory. Domesticated cereals are representedby einkorn (Triticum monococcum), whilst hazelnuts (Corylus avel-lana) and acorns (Quercus sp.) were chosen due to their impliedimportance as wild staples (Holst, 2010; Kubiak-Martens, 1999;Mason, 2000). Other species included are edible sedge and reedtypes (Cyperus longus, Typha latifolia, Sparganium erectum), lords-and-ladies tuber (Arum maculatum), bracken fiddlehead (Pteri-dium sp.), root of wild horseradish (Armoracia rusticana), mead-owsweet (F. ulmaria), and beechnut (Fagus sp.). Whilst clearly notexhaustive of the plants available during the period, these speciesare commonly represented on archaeological sites, and in manycases derive from plants that are high producers of edible compo-nents. Acorus calamus is not indigenous to northern Europe, andwas only naturalised in the sixteenth-century (Vojti�sková et al.,2004) and therefore should not be present in the prehistoricceramic vessels. However, including this species as a control allowsthe accuracy of the classification to be assessed, within theparameters of the dataset with which training was carried out. Thefact that granules were classified as A. calamus shows that they aremost morphologically similar to this species than to any other inthe training set and highlights the limitations of the referencecollection size.
2.2. Experimental foodcrusts
In order to study the effect of cooking on starch granules, wholeeinkorn (T. monococcum) and acorn (Quercus sp.) were boiledseparately in water for 3 h in replica ceramic vessels. Each cookingexperiment was repeated three times in the same vessel. Boilingwas chosen as it is a cooking process associated with the use ofceramic vessels, attested archaeologically by residues thatfrequently appear as a horizontal band on the rim of vessels. Duringthe experiment spot temperatures were recorded with a thermo-couple at regular intervals both in the liquid and on the surface of
the pot. Foodcrust rapidly accumulated on the inside of the vessels,to a thickness of up to 5 mm. The pots were air dried and sealed forfurther analysis.
2.3. Archaeological ceramic vessels
Samples were obtained from the site of Neustadt (LA 156,Kreis Ortholstein), northern Germany (54�0700500N/10�5601600E).The site is a substantial coastal dump of cultural debris thatspans the transition to agriculture, which occurred in thecenturies around 4100 cal BC in this region (Hartz et al., 2002).Based on the large numbers of terrestrial and marine mammalsfound, the site was likely to have been a hunting and fishingstation. The ceramics were dated based on their distinctivetypologies as belonging to the late Mesolithic Ertebølle (EBK)pointed-based style or Early Neolithic flat-based Funnel Beaker(TRB) form. Associated dates on faunal remains were in agree-ment with the typological dating (Glykou, 2011) and indicate thatthe site was used from ca. 4300 cal BC to 3800 cal BC. Severalauthors (e.g. Fischer, 2002) suggest that a shift from wild todomesticated plant foods, such as cereals, occurred during thisperiod, providing an additional incentive for selecting this site.All finds come from a peat context ranging from c. 0.2e0.6 m inthickness sealed under a stratified thick mud layer beneatha sand layer. The excavators consider the find layer to be com-pacted after deposition as a result of intensive productionactivities in this area, such as carcass processing, fishing and flintknapping. Although it was not possible to separate Ertebølle fromFunnel Beaker ceramics by visible stratigraphy, Ertebølle vesselswere predominant at the base of the vertical distribution, andFunnel Beakers in the upper portion (Glykou, 2011). The residuesare extremely well preserved (Fig. 1), in some cases to a thicknessof up to 3 mm. Twenty-six surface deposits were analysed from18 EBK and 8 TRB vessels. Four soot deposits were available fromthe vessels’ exteriors. These were sampled to provide negativecontrols as they are far less likely to derive from the vessels’contents than interior deposits. However, their position on thevessel, such as the rim or body, was recorded to allow thepossibility of over-spill to be assessed.
Soil samples were acquired from the stratum associated withthe ceramics, to provide further controls for contamination. Fiveseparate sub-samples of between 250 mg and 300 mg (averageweight 276 mg) were tested. Faster granule degradation rates havebeen proposed for starch granules in soil than for those associatedwith artefacts, due to factors such as increased enzyme accessibilityin the absence of the protective micro-environment of the ceramic(Haslam, 2004). Therefore interior versus exterior residue counts(mg�1) were used as the primary indicator of contamination, withthe soil samples serving as a supporting measure.
2.4. Starch extraction
Carbonised residues (w1 mg) from experimental and archaeo-logical vessels were removed with a scalpel, accurately weighedinto sterile plastic tubes and treatedwith hydrogen peroxide (H2O2;10%, 10 ml; 15e30 min), whilst gently disaggregating the carbon-ised matrix with a spatula. The tubes were centrifuged (2665 rcf;3 min) and the supernatant removed. After washing three timeswith UltraPure water, the remaining residue was made up to a 1 mlsuspension with UltraPure water. The supernatant, containingliberated starch granules, was added to glass microscope slides thatwere then left to dry at room temperature. This approach leads toquantities of fine carbonised detritus being included in the mounts,but eliminates the potential issue of differential settling rates ofplant microfossils, identified with phytoliths (Stromberg, 2007).
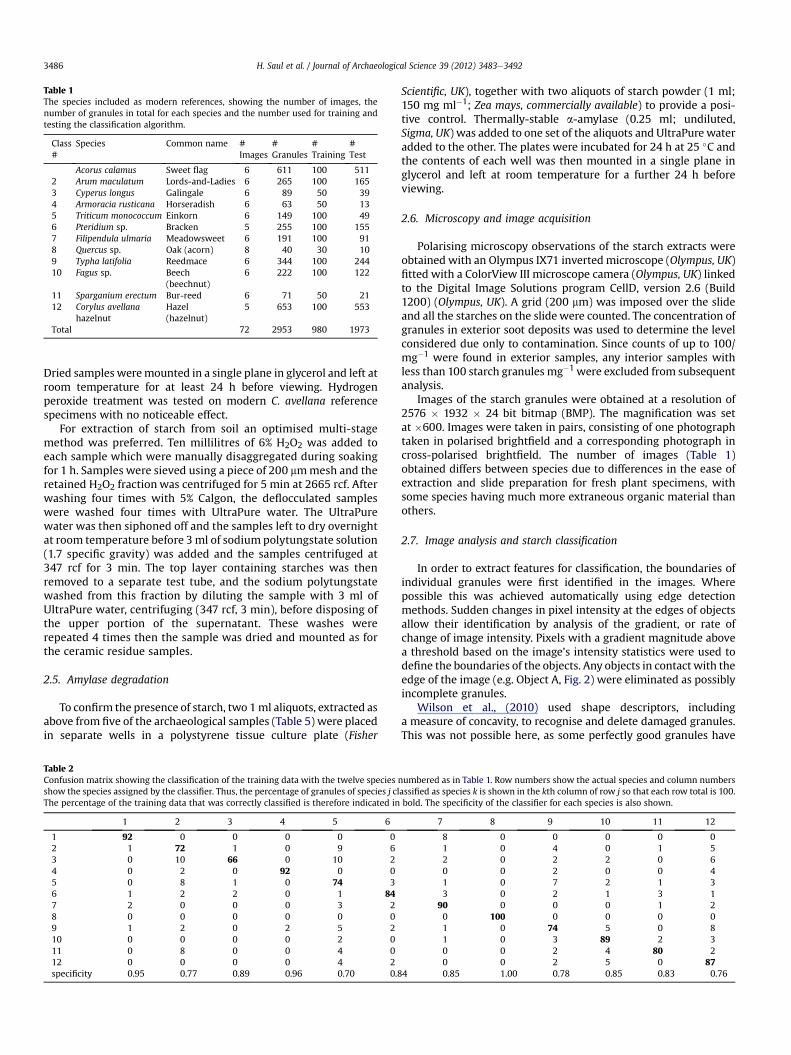
Table 1The species included as modern references, showing the number of images, thenumber of granules in total for each species and the number used for training andtesting the classification algorithm.
Class#
Species Common name #Images
#Granules
#Training
#Test
Acorus calamus Sweet flag 6 611 100 5112 Arum maculatum Lords-and-Ladies 6 265 100 1653 Cyperus longus Galingale 6 89 50 394 Armoracia rusticana Horseradish 6 63 50 135 Triticum monococcum Einkorn 6 149 100 496 Pteridium sp. Bracken 5 255 100 1557 Filipendula ulmaria Meadowsweet 6 191 100 918 Quercus sp. Oak (acorn) 8 40 30 109 Typha latifolia Reedmace 6 344 100 24410 Fagus sp. Beech
(beechnut)6 222 100 122
11 Sparganium erectum Bur-reed 6 71 50 2112 Corylus avellana
hazelnutHazel(hazelnut)
5 653 100 553
Total 72 2953 980 1973
H. Saul et al. / Journal of Archaeological Science 39 (2012) 3483e34923486
Dried samples weremounted in a single plane in glycerol and left atroom temperature for at least 24 h before viewing. Hydrogenperoxide treatment was tested on modern C. avellana referencespecimens with no noticeable effect.
For extraction of starch from soil an optimised multi-stagemethod was preferred. Ten millilitres of 6% H2O2 was added toeach sample which were manually disaggregated during soakingfor 1 h. Samples were sieved using a piece of 200 mmmesh and theretained H2O2 fraction was centrifuged for 5 min at 2665 rcf. Afterwashing four times with 5% Calgon, the deflocculated sampleswere washed four times with UltraPure water. The UltraPurewater was then siphoned off and the samples left to dry overnightat room temperature before 3ml of sodium polytungstate solution(1.7 specific gravity) was added and the samples centrifuged at347 rcf for 3 min. The top layer containing starches was thenremoved to a separate test tube, and the sodium polytungstatewashed from this fraction by diluting the sample with 3 ml ofUltraPure water, centrifuging (347 rcf, 3 min), before disposing ofthe upper portion of the supernatant. These washes wererepeated 4 times then the sample was dried and mounted as forthe ceramic residue samples.
2.5. Amylase degradation
To confirm the presence of starch, two 1ml aliquots, extracted asabove from five of the archaeological samples (Table 5) were placedin separate wells in a polystyrene tissue culture plate (Fisher
Table 2Confusion matrix showing the classification of the training data with the twelve speciesshow the species assigned by the classifier. Thus, the percentage of granules of species j clThe percentage of the training data that was correctly classified is therefore indicated in
1 2 3 4 5 6
1 92 0 0 0 0 02 1 72 1 0 9 63 0 10 66 0 10 24 0 2 0 92 0 05 0 8 1 0 74 36 1 2 2 0 1 847 2 0 0 0 3 28 0 0 0 0 0 09 1 2 0 2 5 210 0 0 0 0 2 011 0 8 0 0 4 012 0 0 0 0 4 2specificity 0.95 0.77 0.89 0.96 0.70 0.8
Scientific, UK), together with two aliquots of starch powder (1 ml;150 mg ml�1; Zea mays, commercially available) to provide a posi-tive control. Thermally-stable a-amylase (0.25 ml; undiluted,Sigma, UK) was added to one set of the aliquots and UltraPure wateradded to the other. The plates were incubated for 24 h at 25 �C andthe contents of each well was then mounted in a single plane inglycerol and left at room temperature for a further 24 h beforeviewing.
2.6. Microscopy and image acquisition
Polarising microscopy observations of the starch extracts wereobtained with an Olympus IX71 invertedmicroscope (Olympus, UK)fitted with a ColorView III microscope camera (Olympus, UK) linkedto the Digital Image Solutions program CellD, version 2.6 (Build1200) (Olympus, UK). A grid (200 mm) was imposed over the slideand all the starches on the slide were counted. The concentration ofgranules in exterior soot deposits was used to determine the levelconsidered due only to contamination. Since counts of up to 100/mg�1 were found in exterior samples, any interior samples withless than 100 starch granules mg�1 were excluded from subsequentanalysis.
Images of the starch granules were obtained at a resolution of2576 � 1932 � 24 bit bitmap (BMP). The magnification was setat �600. Images were taken in pairs, consisting of one photographtaken in polarised brightfield and a corresponding photograph incross-polarised brightfield. The number of images (Table 1)obtained differs between species due to differences in the ease ofextraction and slide preparation for fresh plant specimens, withsome species having much more extraneous organic material thanothers.
2.7. Image analysis and starch classification
In order to extract features for classification, the boundaries ofindividual granules were first identified in the images. Wherepossible this was achieved automatically using edge detectionmethods. Sudden changes in pixel intensity at the edges of objectsallow their identification by analysis of the gradient, or rate ofchange of image intensity. Pixels with a gradient magnitude abovea threshold based on the image’s intensity statistics were used todefine the boundaries of the objects. Any objects in contact with theedge of the image (e.g. Object A, Fig. 2) were eliminated as possiblyincomplete granules.
Wilson et al., (2010) used shape descriptors, includinga measure of concavity, to recognise and delete damaged granules.This was not possible here, as some perfectly good granules have
numbered as in Table 1. Row numbers show the actual species and column numbersassified as species k is shown in the kth column of row j so that each row total is 100.bold. The specificity of the classifier for each species is also shown.
7 8 9 10 11 12
8 0 0 0 0 01 0 4 0 1 52 0 2 2 0 60 0 2 0 0 41 0 7 2 1 33 0 2 1 3 1
90 0 0 0 1 20 100 0 0 0 01 0 74 5 0 81 0 3 89 2 30 0 2 4 80 20 0 2 5 0 87
4 0.85 1.00 0.78 0.85 0.83 0.76
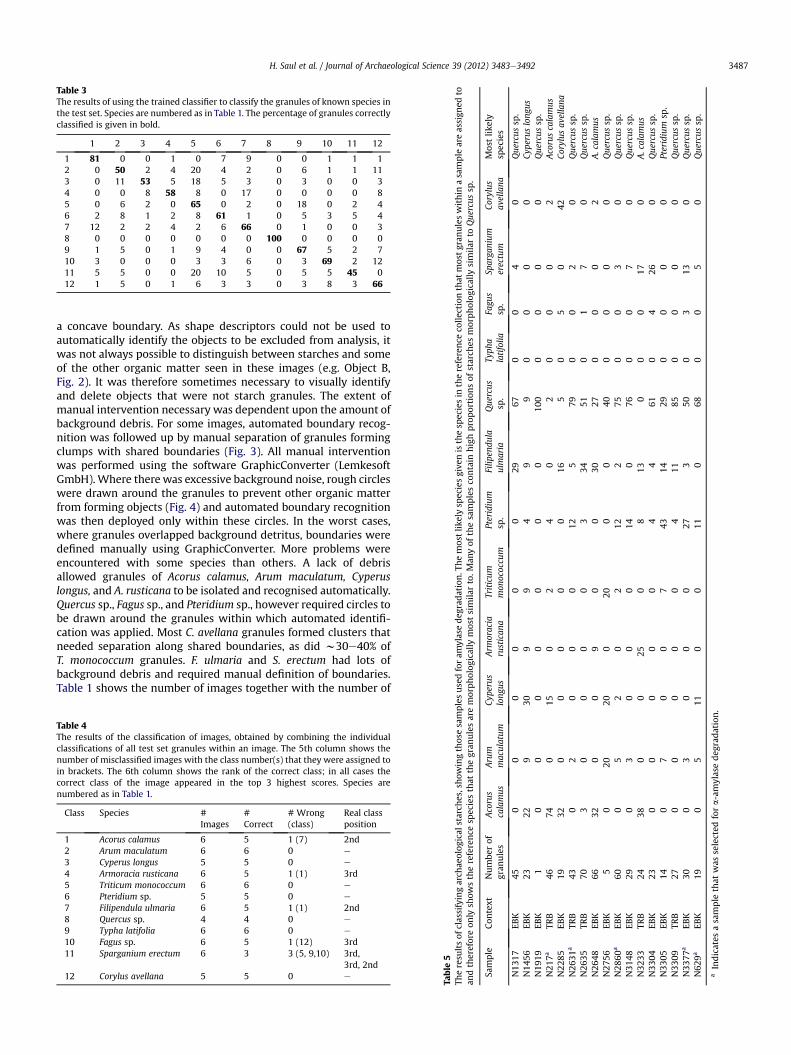
Table 3The results of using the trained classifier to classify the granules of known species inthe test set. Species are numbered as in Table 1. The percentage of granules correctlyclassified is given in bold.
1 2 3 4 5 6 7 8 9 10 11 12
1 81 0 0 1 0 7 9 0 0 1 1 12 0 50 2 4 20 4 2 0 6 1 1 113 0 11 53 5 18 5 3 0 3 0 0 34 0 0 8 58 8 0 17 0 0 0 0 85 0 6 2 0 65 0 2 0 18 0 2 46 2 8 1 2 8 61 1 0 5 3 5 47 12 2 2 4 2 6 66 0 1 0 0 38 0 0 0 0 0 0 0 100 0 0 0 09 1 5 0 1 9 4 0 0 67 5 2 710 3 0 0 0 3 3 6 0 3 69 2 1211 5 5 0 0 20 10 5 0 5 5 45 012 1 5 0 1 6 3 3 0 3 8 3 66
susedforam
ylasedeg
radation.T
hemostlik
elysp
eciesgive
nisthesp
eciesin
thereference
colle
ctionthat
mostgran
uleswithin
asample
areassign
edto
orpholog
ically
mostsimila
rto.M
anyof
thesamplesco
ntain
highproportion
sof
starch
esmorpholog
ically
simila
rto
Que
rcus
sp.
perus
ngus
Arm
oracia
rustican
aTriticum
mon
ococcu
mPteridium
sp.
Filip
endu
laulmaria
Que
rcus
sp.
Typh
alatifolia
Fagu
ssp
.Sp
arga
nium
erectum
Corylus
avellana
Mostlik
ely
species
00
029
670
04
0Que
rcus
sp.
99
49
90
00
0Cy
peruslong
us0
00
010
00
00
0Que
rcus
sp.
02
40
20
00
2Acoruscalamus
00
016
50
50
42Co
rylusav
ellana
00
125
790
02
0Que
rcus
sp.
00
334
510
17
0Que
rcus
sp.
90
030
270
00
2A.calam
us0
200
040
00
00
Que
rcus
sp.
02
122
750
03
0Que
rcus
sp.
00
140
760
07
0Que
rcus
sp.
250
813
00
017
0A.calam
us0
04
461
04
260
Que
rcus
sp.
07
4314
290
00
0Pteridium
sp.
00
411
850
00
0Que
rcus
sp.
00
273
500
313
0Que
rcus
sp.
00
110
680
05
0Que
rcus
sp.
H. Saul et al. / Journal of Archaeological Science 39 (2012) 3483e3492 3487
a concave boundary. As shape descriptors could not be used toautomatically identify the objects to be excluded from analysis, itwas not always possible to distinguish between starches and someof the other organic matter seen in these images (e.g. Object B,Fig. 2). It was therefore sometimes necessary to visually identifyand delete objects that were not starch granules. The extent ofmanual intervention necessary was dependent upon the amount ofbackground debris. For some images, automated boundary recog-nition was followed up by manual separation of granules formingclumps with shared boundaries (Fig. 3). All manual interventionwas performed using the software GraphicConverter (LemkesoftGmbH).Where there was excessive background noise, rough circleswere drawn around the granules to prevent other organic matterfrom forming objects (Fig. 4) and automated boundary recognitionwas then deployed only within these circles. In the worst cases,where granules overlapped background detritus, boundaries weredefined manually using GraphicConverter. More problems wereencountered with some species than others. A lack of debrisallowed granules of Acorus calamus, Arum maculatum, Cyperuslongus, and A. rusticana to be isolated and recognised automatically.Quercus sp., Fagus sp., and Pteridium sp., however required circles tobe drawn around the granules within which automated identifi-cation was applied. Most C. avellana granules formed clusters thatneeded separation along shared boundaries, as did w30e40% ofT. monococcum granules. F. ulmaria and S. erectum had lots ofbackground debris and required manual definition of boundaries.Table 1 shows the number of images together with the number of
Table 4The results of the classification of images, obtained by combining the individualclassifications of all test set granules within an image. The 5th column shows thenumber of misclassified images with the class number(s) that they were assigned toin brackets. The 6th column shows the rank of the correct class; in all cases thecorrect class of the image appeared in the top 3 highest scores. Species arenumbered as in Table 1.
Class Species #Images
#Correct
# Wrong(class)
Real classposition
1 Acorus calamus 6 5 1 (7) 2nd2 Arum maculatum 6 6 0 e
3 Cyperus longus 5 5 0 e
4 Armoracia rusticana 6 5 1 (1) 3rd5 Triticum monococcum 6 6 0 e
6 Pteridium sp. 5 5 0 e
7 Filipendula ulmaria 6 5 1 (1) 2nd8 Quercus sp. 4 4 0 e
9 Typha latifolia 6 6 0 e
10 Fagus sp. 6 5 1 (12) 3rd11 Sparganium erectum 6 3 3 (5, 9,10) 3rd,
3rd, 2nd12 Corylus avellana 5 5 0 e
Table
5Th
eresu
ltsof
classifyingarch
aeolog
ical
starch
es,show
ingthosesample
andthereforeon
lysh
owsthereference
speciesthat
thegran
ulesarem
Sample
Con
text
Numbe
rof
gran
ules
Acorus
calamus
Arum
maculatum
Cy lo
N13
17EB
K45
00
0N14
56EB
K23
229
30N19
19EB
K1
00
0N21
7aTR
B46
740
15N22
85EB
K19
320
0N26
31a
TRB
430
20
N26
35TR
B70
30
0N26
48EB
K66
320
0N27
56EB
K5
020
20N28
60a
EBK
600
52
N31
48EB
K29
03
0N32
33TR
B24
380
0N33
04EB
K23
00
0N33
05EB
K14
07
0N33
09TR
B27
00
0N33
77a
EBK
300
30
N62
9aEB
K19
05
11
aIndicates
asample
that
was
selected
fora-amylasedeg
radation.

Fig. 2. A masked image of Arum maculatum granules with (A) an unusable starchgranule overlapping the edge of the image and (B) an unwanted piece of cellulose.Magnification �600.
Fig. 4. The manual intervention of rough black circles around the granules allows theedge detection to be restricted and only objects located within these areas considered.Magnification �600.
H. Saul et al. / Journal of Archaeological Science 39 (2012) 3483e34923488
granules extracted for each of the modern reference species. Formost species, well over 100 granules were extracted.
For each granule, a total of 26 feature variables were recordedfor classification. Variables were calculated based on size and shapeof the granule, its boundary curvature and concavity and the shapeof its polarisation cross. More detail on these variables can be foundin Wilson et al., (2010). In addition, statistical measures werecalculated from pixel intensities within the granule for both thepolarised and cross-polarised images as well as the gradientmagnitudes. Data for two-thirds of the granules (chosen randomly)from each modern species were used to train supervised learningalgorithms and the remaining data used as an independent test set.Four different classifiers were trained; a self-organisingmap (SOM),learning vector quantisation (LVQ) and support vector machines
Fig. 3. Without manual intervention, the clumps of hazelnut (Corylus avellana) gran-ules lead to composite objects as shown in (a). After boundaries between granules areadded manually, separate granules can be identified and treated individually insubsequent morphological analysis. Magnification �600.
(SVM), both (SVM_linear) and with a radial basis function kernel(SVM_RBF).
Classification of the training data was used to assess the internalconsistency (both sensitivity and specificity) of the individualclassifiers as well as combinations of classifiers. The best classifi-cation system was then applied to the test data from the modernreference starches not used during training, to test the extrapola-tion of the results to unseen data. The trained classifier was thenapplied to experimentally cooked examples of T. monococcum andacorn (Quercus sp.) to test classification after thermal alteration.Finally the classifier was applied to archaeological starches fromcarbonised surface deposits from Neustadt.
3. Results and discussion
3.1. Starch survival in experimentally carbonised deposits
Starch granules in various states of heat-alteration are preservedin carbonised deposits. This emphasises the still incompleteunderstanding of the processes and conditions of morphogenesis.The experimental acorn and einkorn cooking pots yielded608 granules mg�1 and 184 granules mg�1 respectively. These wereexposed to temperatures of at least 100 �C for more than 3 h andreached higher temperatures (>120 �C) as the water content of thepots decreased and the residues dehydrated on the ceramic surface.The granules survived repeated cooking in the vessels, confirmingthat thermal degradation does not occur uniformly at a thresholdtemperature (Torrence and Barton, 2006).
3.2. Starch survival in archaeological carbonised deposits
No granules were found in any of the archaeological samplesthat were subjected to a-amylase, whereas counts of between 252and 284 mg�1 were obtained for the five controls without amylase,suggesting conclusively that these are starch. The positive Z. mayscontrol showed no starch, whilst the negative Z. mays starches(>500 mg�1) remained unaltered. Therefore it would seem that, atleast in this burial environment, archaeological carbonised depositsof at least 6000 years old do contain starch. The minor discrepancybetween counts mg�1 of samples put forward for automated clas-sification and counts mg�1 of the same samples used for a-amylasedegradation may result in part from the non-uniform nature ofstarch content in the residues.
H. Saul et al. / Journal of Archaeological Science 39 (2012) 3483e3492 3489
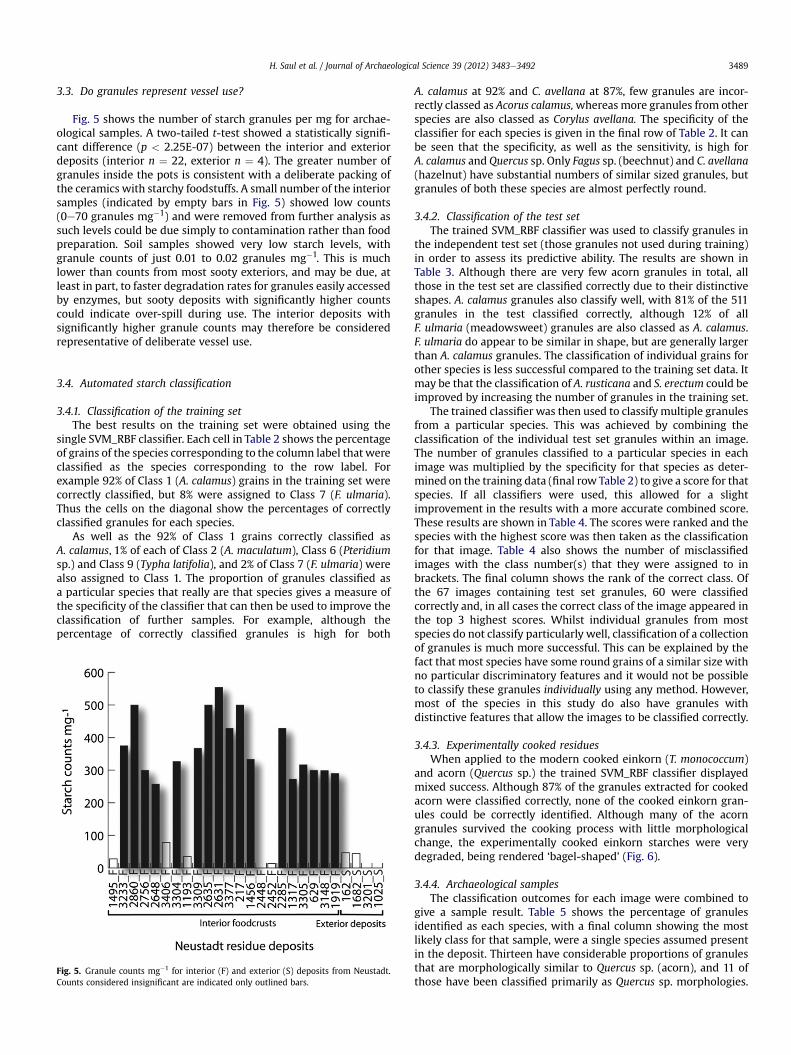
3.3. Do granules represent vessel use?
Fig. 5 shows the number of starch granules per mg for archae-ological samples. A two-tailed t-test showed a statistically signifi-cant difference (p < 2.25E-07) between the interior and exteriordeposits (interior n ¼ 22, exterior n ¼ 4). The greater number ofgranules inside the pots is consistent with a deliberate packing ofthe ceramics with starchy foodstuffs. A small number of the interiorsamples (indicated by empty bars in Fig. 5) showed low counts(0e70 granules mg�1) and were removed from further analysis assuch levels could be due simply to contamination rather than foodpreparation. Soil samples showed very low starch levels, withgranule counts of just 0.01 to 0.02 granules mg�1. This is muchlower than counts from most sooty exteriors, and may be due, atleast in part, to faster degradation rates for granules easily accessedby enzymes, but sooty deposits with significantly higher countscould indicate over-spill during use. The interior deposits withsignificantly higher granule counts may therefore be consideredrepresentative of deliberate vessel use.
3.4. Automated starch classification
3.4.1. Classification of the training setThe best results on the training set were obtained using the
single SVM_RBF classifier. Each cell in Table 2 shows the percentageof grains of the species corresponding to the column label that wereclassified as the species corresponding to the row label. Forexample 92% of Class 1 (A. calamus) grains in the training set werecorrectly classified, but 8% were assigned to Class 7 (F. ulmaria).Thus the cells on the diagonal show the percentages of correctlyclassified granules for each species.
As well as the 92% of Class 1 grains correctly classified asA. calamus, 1% of each of Class 2 (A. maculatum), Class 6 (Pteridiumsp.) and Class 9 (Typha latifolia), and 2% of Class 7 (F. ulmaria) werealso assigned to Class 1. The proportion of granules classified asa particular species that really are that species gives a measure ofthe specificity of the classifier that can then be used to improve theclassification of further samples. For example, although thepercentage of correctly classified granules is high for both
Fig. 5. Granule counts mg�1 for interior (F) and exterior (S) deposits from Neustadt.Counts considered insignificant are indicated only outlined bars.
A. calamus at 92% and C. avellana at 87%, few granules are incor-rectly classed as Acorus calamus,whereas more granules from otherspecies are also classed as Corylus avellana. The specificity of theclassifier for each species is given in the final row of Table 2. It canbe seen that the specificity, as well as the sensitivity, is high forA. calamus and Quercus sp. Only Fagus sp. (beechnut) and C. avellana(hazelnut) have substantial numbers of similar sized granules, butgranules of both these species are almost perfectly round.
3.4.2. Classification of the test setThe trained SVM_RBF classifier was used to classify granules in
the independent test set (those granules not used during training)in order to assess its predictive ability. The results are shown inTable 3. Although there are very few acorn granules in total, allthose in the test set are classified correctly due to their distinctiveshapes. A. calamus granules also classify well, with 81% of the 511granules in the test classified correctly, although 12% of allF. ulmaria (meadowsweet) granules are also classed as A. calamus.F. ulmaria do appear to be similar in shape, but are generally largerthan A. calamus granules. The classification of individual grains forother species is less successful compared to the training set data. Itmay be that the classification of A. rusticana and S. erectum could beimproved by increasing the number of granules in the training set.
The trained classifier was then used to classify multiple granulesfrom a particular species. This was achieved by combining theclassification of the individual test set granules within an image.The number of granules classified to a particular species in eachimage was multiplied by the specificity for that species as deter-mined on the training data (final row Table 2) to give a score for thatspecies. If all classifiers were used, this allowed for a slightimprovement in the results with a more accurate combined score.These results are shown in Table 4. The scores were ranked and thespecies with the highest score was then taken as the classificationfor that image. Table 4 also shows the number of misclassifiedimages with the class number(s) that they were assigned to inbrackets. The final column shows the rank of the correct class. Ofthe 67 images containing test set granules, 60 were classifiedcorrectly and, in all cases the correct class of the image appeared inthe top 3 highest scores. Whilst individual granules from mostspecies do not classify particularly well, classification of a collectionof granules is much more successful. This can be explained by thefact that most species have some round grains of a similar size withno particular discriminatory features and it would not be possibleto classify these granules individually using any method. However,most of the species in this study do also have granules withdistinctive features that allow the images to be classified correctly.
3.4.3. Experimentally cooked residuesWhen applied to the modern cooked einkorn (T. monococcum)
and acorn (Quercus sp.) the trained SVM_RBF classifier displayedmixed success. Although 87% of the granules extracted for cookedacorn were classified correctly, none of the cooked einkorn gran-ules could be correctly identified. Although many of the acorngranules survived the cooking process with little morphologicalchange, the experimentally cooked einkorn starches were verydegraded, being rendered ‘bagel-shaped’ (Fig. 6).
3.4.4. Archaeological samplesThe classification outcomes for each image were combined to
give a sample result. Table 5 shows the percentage of granulesidentified as each species, with a final column showing the mostlikely class for that sample, were a single species assumed presentin the deposit. Thirteen have considerable proportions of granulesthat are morphologically similar to Quercus sp. (acorn), and 11 ofthose have been classified primarily as Quercus sp. morphologies.

Fig. 6. The degradation of experimentally cooked einkorn (Triticum monococcum)shows a distinctive ‘bagel’-shape, resulting in poor identification in automated anal-ysis. Magnification �600.
H. Saul et al. / Journal of Archaeological Science 39 (2012) 3483e34923490
These findings correspond strongly with manual observations ofthe granules, suggesting that a primary role for these vessels mayhave been the processing of plants with acorn-like starchmorphologies. These results are supported by the large quantitiesof acorn casings recovered from deposits at Neustadt (recorded as“N” in the macrofloral report). Other samples show equally highproportions of other species. For example, five samples suggestedible Pteridium sp., type starch in combination with C. avellana(n1), F. ulmaria, and S. erectum (n1) type starch. Little evidence forthe use of cereals was found. A moderate percentage (20%) ofgranules classified as T. monococcum (einkorn) typewas recorded inan Ertebølle vessel (N2756), albeit with a low granule count.Furthermore, none of the archaeological samples had starches thatresembled the degraded T. monococcum granules observed aftercooking. It could be that cereals, if present at all, were not processedin these pots or perhaps, as suggested by the cooking experiments,that cereal starches do not survive in this particular archaeologicalcontext. One reason for the poor preservation could be the lowproportion (25e27%) of amylose in cereal starch (Frederikssonet al., 1998), which may not be sufficient to offset gelatinisationduring formation of the foodcrusts.
In five samples, a high proportion of granules are classified asA. calamus although this species is not indigenous to Europe andwas not introduced until the 16th Century. This species thereforecould not have been cooked in Ertebølle or Early Neolithic vessels.The fact that granules were classified as A. calamus shows that theyare more similar morphologically to this species than to any otherspecies in the training set, and highlights the limitations of usingsuch a small reference set. For this reason, all the identificationsproposed above should be considered to be tentative and prelimi-nary, pending expansion of the reference dataset.
3.4.5. Wider significance of the findingsGranule forms similar to hazelnut are only represented in one
vessel as the most likely species, suggesting that macroscopicfindings of hazelnut (Kubiak-Martens,1999; Holst, 2010; Harild andKarg, 2009; Mason, 2004) may have overemphasised their impor-tance archaeologically, in relation to other important staples.Alternatively hazelnuts were not processed in ceramic vessels asfrequently as other plants. Conversely, starches with morphologiessimilar to acorn seem to have been intensively processed in thevessels at Neustadt, occurring in all of the samples analysed.Although this cannot be taken as definite proof of acorn processing,comparable practices are known ethnographically from Californian
Native Americans (Abrams and Nowacki, 2008; Keeley, 2002).Acorns are stripped of their outer casings, crushed roughly andheated repetitively in several changes of water to remove toxictannins.
In purely calorific terms, the intensive collection and process-ing of acorns for food produces similar suggested return rates tohazelnut (Rowley-Conwy, 1984, 303). However, unlike hazelnuts,Quercus sp., is a masting plant meaning it produces a substantialcrop bi-, tri-, or quadrennially, rather than annually (Abrams andNowacki, 2008). In order for acorns to make up an annualcontribution either a collection area spanning multiple staggeredmasting regions must be available, or storage of acorn must beemployed.
The results may be evidencing the second option. Acorns arearguably best stored as a processed and dried flour. Up to 31% oftheir content can be lipid (Bainbridge, 1985), compared to only1% for wheat (Wagnon, 1946), making them susceptible torancidity in their fresh form. In addition, rapid destruction ofa fresh store of acorns can occur with infestation by acornweevil (e.g. Curculio nucum and Cuculio glandinium), an insectcommonly found burrowed discretely into the achene. Consid-ering calories exclusively, a duel acorn and hazelnut plant stapleeasily matches the return rates for land mammals (Rowley-Conwy, 1984), and even outdoes them, especially if the sug-gested kill rate is too high as has been speculated (Rowley-Conwy, 1984). Considering the importance animal productshave played in interpretations of the transition to agriculture,these results would suggest plant resources have a higher profilerole than has previously been allowed for.
4. Conclusions
Here we have shown that starches can be extracted andidentified from carbonised residues adhering to ceramics.Comparisons of counts mg�1 from interior foodcrusts andexterior sooty deposits, as well as associated sediments showthat these starches are representative of plant processingactivities across the transition to agriculture at Neustadt,northern Germany. Based on these current findings, there is nosupport for the interpretation of a synchronous adoption ofdomesticated cereals with a change to Funnel Beakers, as istraditionally supposed (Fischer, 2002). However, further work isrequired to establish whether this absence of evidence is due tocompromised starch preservation following cooking ortaphonomy. Our results show that acorn (Quercus sp.) typestarch and grain forms similar to some species of edible sedgeand reed (Sparganium erectum, Cyperus longus) may have beenimportant plant resources in both the Mesolithic and EarlyNeolithic. Classification algorithms can only assign one of theclasses for which training data is available. The more repre-sentative the modern training data is, the more reliable theclassification will be. The most systematic survey of starchvariability including northern European species is to date rep-resented by Reichert (1913). Though not directly measuringvariability, the micrographs and accompanying descriptionssuggest considerable variation in granule histology, pointing tothe potential for increased identification accuracy with expan-sion of the reference collection. In addition to the inclusion offurther species, future improvements should include trainingwith experimentally cooked starch populations to account forheat alteration more accurately. Although the methods pre-sented here are aimed at classifying the plant starch part of theresidue which might only be a small fraction of the foods thatwere cooked, such systematic and extensive techniques mayfinally begin to rectify stunted debates about the importance of

H. Saul et al. / Journal of Archaeological Science 39 (2012) 3483e3492 3491
plants across the transition to agriculture in northern Europe(Zvelebil, 1994).
Acknowledgements
The authors would like to gratefully acknowledge the finan-cial support of the Arts and Humanities Research Council (AH/E008232/1). Some of the modern reference material was kindlygiven by The Royal Botanical Gardens at Kew, London. This workcould not have been achieved without the support of NickyMilner, and Richard Allen.
References
Abrams, M.D., Nowacki, G., 2008. Native Americans as active and passive promotersof mast and fruit trees in eastern USA. Holocene 18 (7), 1123e1137.
Albert, R.M., Weiner, S., Bar-Yosef, O., Meignen, L., 2000. Phytoliths in the MiddlePalaeolithic deposits of Kebara Cave, Mt. Carmel, Israel: study of plant materialsused for fuel and other purposes. J. Archaeol. Sci. 27 (10), 931e947.
Andersen, S., Malmros, C., 1984. “Madskorpe” på Ertebøllekar fra Tybrind Vig.Aarbøger for Nordisk Oldkyndighed of Historie, 78e95.
Atchinson, J.L., Fullager, R., 1998. Starch residues on pounding implements fromJinmium rock-shelter. In: Fullager, R. (Ed.), A Closer Look: Recent AustralianStudies of Stone Tools. University of Sydney, Sydney, pp. 109e125.
Bainbridge, D.A., 1985. Acorns as Food: Oak Bibliography #1. Sierra Nature Prints,Twain Harte, California.
Balme, J., Beck, W.E., 2002. Starch and charcoal: useful measures of activity areas inarchaeological rockshelters. J. Archaeol. Sci. 29, 157e166.
Barclay, G.J., 1983. Sites of the third millennium BC to the first millennium AD atNorth mains, Strathallan, Perthshire. Proc. Soc. Antiq. Scot. 113, 122e281.
Barclay, G.J., Russel-White, C.J., 1993. Excavations in the ceremonial complex of thefourth to second millenium BC at Balfarg/Balbirinie, Glenrothes, Fife. Proc. Soc.Antiq. Scot. 123, 43e210.
Barton, H., White, J.P., 1993. Use of stone and shell artefacts at Balof 2, new Ireland,Papua new Guinea. Asian Perspect. 32, 169e181.
Boudin, M., Van Strydonck, M., Crombé, P., 2009. Radiocarbon dating of pottery foodcrusts: reservoir effect or not? The case of the Swifterbant pottery from Doel‘Deurganckdok’ (Belgium). In: Crombé, P., Van Strydonck, M., Sargant, J.,Boudin, M., Bats, M. (Eds.), Chronology and Evolution within the Mesolithic ofNorth-west Europe. Proceedings of an International Meeting, Brussels, May30theJune 1st 2007. Cambridge Scholars Publishing, Newcastle, pp. 727e746.
Boyd, M., Surette, C., Nicholson, B.A., 2006. Archaeobotanical evidence of prehistoricmaize (Zea mays) consumption at the northern edge of the Great Plains.J. Archaeol. Sci. 33 (8), 1129e1140.
Boyd, M., Varney, T., Surette, C., Surrette, J., 2008. Reassessing the northern limit ofmaize consumption in North America: stable isotope, plant microfossil, andtrace element content of carbonised food residue. J. Archaeol. Sci. 35,2545e2556.
Briuer, F.L., 1976. New clues to stone tool function: plant and animal residues. Am.Antiq. 41, 478e484.
Charters, S., Evershed, R.P., Quye, A., Blinkhorn, P., Reeves, V., 1997. Simulationexperiments for determining the use of ancient pottery vessels: the behaviours ofepicuticular leaf wax during boiling of a leafy vegetable. J. Archaeol. Sci. 24, 1e7.
Craig, O.E., 2004. Organic analysis of <<food crusts>> from sites in the ScheldeValley, Belgium: a preliminary evaluation. Notae Praehistoricae 24, 209e217.
Craig, O.E., Forster, M., Andersen, S.H., Koch, E., Crombé, P., Milner, N., Stern, B.,Bailey, G., Heron, C., 2007. Molecular demonstration of marine and fresh-water products in prehistoric pottery from northern Europe. Archaeom 49,135e142.
Crombé, P., Perdaen, Y., Sargant, J., Van Royen, J.-P., Van Strydonck, M., 2002. TheMesolithiceNeolithic transition in the sandy lowlands of Belgium: newevidence. Antiq 76, 699e706.
Crowther, A., 2005. Starch residues on Undecorated Lapita pottery from Anir, newIreland. Oceania 40, 62e66.
Evershed, R.P., 2008. Experimental approaches to the identification of absorbedorganic residues in archaeological ceramics. World Archaeol. 40 (1), 26e47.
Evershed, R.P., Heron, C., Goad, L.J., 1991. Epicuticular leaf wax componentspreserved in potsherds as chemical indicators of leafy vegetables in ancientdiet. Antiq 65, 540e544.
Fischer, A., 2002. Food for feasting? In: Fischer, A., Kristiansen, K. (Eds.), The Neo-lithisation of Denmark e 150 years of Debate Sheffield Archaeological Mono-graphs, vol. 12. J. R. Collis, Sheffield, pp. 343e393.
Frederiksson, H., Silverio, J., Andersson, R., Eliasson, A.-C., Ǻman, P., 1998. Theinfluence of amylose and amylopectin characteristics on gelatinisation andretrogradation properties of different starches. Carbohydr. Polym. 35, 119e134.
Gaillard, T., Bowler, P., 1987. Morphology and composition of starch. In: Gaillard, T.(Ed.), Starch: Properties and Potential. Critical Reposits on Applied Chemistry,vol. 13. John Wiley and Sons, Brisbane, pp. 55e79.
Glykou, A., 2011. Neustadt LA 156- ein submariner Fundplatz des späten Meso-lithikums und frühesten Neolithikums in Schleswig-Holstein. Untersuchungen
zur Subsistenzstrategie der letzten Jäger, Sammler und Fischer an der nord-deutschen ostseeküste. PhD thesis, University of Kiel, Germany.
Gott, B., Barton, H., Samuel, D., Torrence, R., 2006. Biology of starch. In: Torrence, R.,Barton, H. (Eds.), Ancient Starch Research. Left Coast Press, California,pp. 35e46.
Hansson, A.-M., Isaksson, S., 1994. Analyses of charred organic remains. LaborativArkeologi 7, 21e29.
Hardy, B.L., 2004. Neanderthal behaviour and stone tool function at the MiddlePalaeolithic site of La Quina, France. Antiq 78, 547e565.
Hardy, K., 2007. Food for thought: starch in Mesolithic diet. Meso. Misc. 18 (2), 2e11.Hardy, K., Blakeney, T., Copeland, L., Kirkham, J., Wrangham, R., Collins, M., 2009.
Starch granules, dental calculus and new perspectives on ancient diet.J. Archaeol. Sci. 36 (2), 248e255.
Harild, J.A., Karg, S., 2009. Analysis of plant macrofossil remains in gyttja samplesfrom Ronæs Skov. In: Andersen, S. (Ed.), Ronæs Skov: marinarkæologiskeundersøgelser af kystboplads fra Ertebølletid, vol. 64. Jysk Arkæologisk SelskabsSkrifter, pp. 233e236.
Hartz, S., Heinrich, D., Lübke, H., 2002. Coastal Farmers e the Neolithisation ofnorthern-most Germany. In: Fischer, A., Kristiansen, K. (Eds.), The Neo-lithisation of Denmark e 150 Years of Debate. J. R. Collis Publications, Sheffield,pp. 321e340.
Haslam, M., 2004. The decomposition of starch grains in soils: implications forarchaeological residue analysis. J. Archaeol. Sci. 31 (12), 1715e1734.
Henry, A.G., Piperno, D.R., 2008. Using plant microfossils from dental calculus torecover human diet: a case study from Tell al-Raq�a’I, Syria. J. Archaeol. Sci. 35,1943e1950.
Henry, A.G., Hudson, H.F., Piperno, D.R., 2009. Changes in starch grain morphologiesfrom cooking. J. Archaeol. Sci. 36 (3), 915e922.
Holst, D., 2010. Hazelnut economy of early Holocene hunteregatherers: a casestudy from Mesolithic Duvensee, northern Germany. J. Archaeol. Sci. 37 (11),2871e2880.
Hoover, R., 2001. Composition, molecular structure, and physicochemical propertiesof the tuber and root starches: a review. Carbohydr. Polym. 45, 253e267.
Horrocks, M., 2004. Polynesian plant subsistence in prehistoric New Zealand:a summary of the microfossil evidence. New Zeal. J. Bot. 42, 321e334.
Horrocks, M., 2005. A combined procedure for recovering phytoliths and starchresidues from soils, sedimentary deposits and similar materials. J. Archaeol. Sci.32, 1169e1175.
Horrocks, M., Nunn, P.D., 2007. Evidence for introduced taro (Colocasia esculenta)and lesser yam (Dioscorea esculenta) in Lapita-era (c. 3050e2500 cal. Yr B.P)deposits from Bourewa, southwest Viti Levu Island. Fiji. J. Arch. Sci. 34,739e748.
Horrocks, M., Irwin, G., Jones, M., Sutton, D., 2004. Starch grains and xylem cells ofsweet potato (Ipomoea batatas) and bracken (Pteridium esculentum) in archae-ological deposits from northern New Zealand. J. Archaeol. Sci. 31 (3), 251e258.
Kealhofer, L., Torrence, R., Fullager, R., 1999. Integrating phytoliths within use-wear/residue studies of stone tools. J. Arch. Sci. 26, 527e546.
Keeley, J., 2002. Native American impacts on fire regimes of the California coastalranges. J. Biogeogr. 29, 303e320.
Kubiak-Martens, L., 1996. Evidence for possible use of plant foods in Palaeolithic andMesolithic diet from the site of Calowanie in the central part of the Polish Plain.Veg. Hist. Archaeobot. 5, 33e38.
Kubiak-Martens, L., 1999. The plant food component of the diet at the late Meso-lithic (Ertebølle) settlement at Tybrind Vig, Denmark. Veg. Hist. Archaeobot. 8,117e127.
Lalueza Fox, C., Péz-Pérez, A., 1994. Dietary information through the examination ofplant phytoliths on the enamel surface of human dentition. J. Archaeol. Sci. 21,29e34.
Lalueza Fox, C., Juan, J., Albert, R.M., 1996. Phytolith analysis on dental calculus,enamel surface, and burial soil: information about diet and palaeoenvironment.Am. J. Phys. Anth. 101, 101e113.
Leach, H.W., 1965. Gelatinization of starch. In: Whistler, R.L., Paschall, E.F. (Eds.),Starch: Chemistry and Technology. Academic Press, New York, pp. 289e307.
Long, D.J., Milburn, P., Bunting, M.J., Topping, R., Holden, T.G., 1999. Black henbane(Hyoscyamus niger L.) in the Scottish Neolithic: a re-evaluation of palynologicalfindings from grooved ware pottery at Balfarg Riding School and Henge, Fife.J. Archaeol. Sci. 26, 45e52.
Long, D.J., Tipping, R., Holder, T.G., Bunting, M.J., Milburn, P., 2000. The uses ofhenbane (Hyoscyamus niger L.) as a hallucinogenic at Neolithic ‘ritual’ sites:a re-evaluation. Antiq 74, 49e53.
Lusteck, R.K., Thompson, R.G., 2007. Residues of maize in North American pottery:what phytoliths can add to the story of maize. In: Barnard, H., Eerkens, J.W.(Eds.), Theory and Practice of Archaeological Residue Analysis. British Archae-ological Reports, vol. 1650. Archaeopress, Oxford, pp. 8e16.
Madella, M., Jones, K., Echlin, P., Powers-Jones, A., Moore, M., 2009. Plant wateravailability and analytical microscopy of phytoliths: implications for ancientirrigation in arid zones. Quat. Int. 193 (1-2), 32e40.
Mason, S.L.R., 2000. Fire and Mesolithic subsistence e managing oaks for acorns innorthwest Europe? Palaeogeogr. Palaeoclimatol. Palaeoecol. 164 (1e4),139e150.
Mason, S., 2004. Archaeobotanical analysis e Møllegabet II. In: Skaarup, J., Grøn, O.(Eds.), Møllegabet II: A Submerged Mesolithic Settlement in Southern Denmark.British Archaeological Reports International Series, vol. 1328, pp. 122e143.
Miles, M., Morris, V.J., Orford, P.D., Ring, S.G., 1985. The roles of amylose and amylo-pectin in theGelation and retrogradation of starch. Carbohydr. Res.135, 271e281.

H. Saul et al. / Journal of Archaeological Science 39 (2012) 3483e34923492
Parr, J.F., Carter, M., 2003. Phytolith and starch analysis of sediment samples fromtwo archaeological sites on Dauar Island, Torres Strait, northeastern Australia.Veg. Hist. Archaeobot. 12, 131e141.
Pearsall, D.M., 2002. Maize is still ancient in prehistoric Ecuador: the view from RealAlto, with comments on Staller and Thompson. J. Archaeol. Sci. 29, 51e55.
Perry, L., 2004. Starch analyses reveal the relationship between tool type andfunction: an example from the Orinoco valley of Venezuela. J. Archaeol. Sci. 31,1069e1081.
Piperno, D.R., Holst, I., 1998. The presence of starch grains on prehistoric stone toolsfrom the humid Neotropics: indications of early tuber use and agriculture inPanama. J. Archaeol. Sci. 25, 765e776.
Piperno, D.R., Ranere, A.J., Holst, I., Hansell, P., 2000. Starch grains reveal early rootcrop horticulture in the Panamanian tropical forest. Nature 407, 894e897.
Piperno, D.R., Weiss, E., Holst, I., Nadel, D., 2004. Processing of wild cereal grains inthe Upper Palaeolithic revealed by starch grain analysis. Nature 430, 670e673.
Ratnayake,W.S., Jackson,D.S., 2006.Gelatinization andsolubilityof corn starchduringheating in excess water: new insights. J. Agric. Food Chem. 54, 3712e3716.
Reichert, E.T., 1913. The Differentiation and Specificity of Starches in Relation toGenera, Species, Etc. No. 173. Carnegie Institute Publication, Washington.
Reinhard, K.J., Danielson, D.R., 2005. Pervasiveness of phytoliths in prehistoricsouthwestern diet and implications for regional and temporal trends for dentalmicrowear. J. Archaeol. Sci. 32, 981e988.
Rowley-Conwy, P., 1984. The laziness of the short-distance hunter: the origins ofagriculture in Western Denmark. J. Anthropol. Archaeol. 3, 300e324.
Sajilata, M.G., Singhal, R.S., Kulkarni, P.R., 2006. Resistant starch e a review. Comp.Rev. Food Sci. Food Saf. 5, 1e17.
Samuel, D., 1996. Investigations of ancient Egyptian baking and brewing methodsby correlative microscopy. Science 273, 488e490.
Saul, H., Fischer, A., Glykou, A., Hartz, S., Madella, M., Craig, O., forthcoming. Phy-toliths in pottery reveal prehistoric hunter-gatherers’ taste for spice.
Sivak, M.N.S., Preiss, J., 1998. Occurrence of starch. In: Sivak, M.N.S., Preiss, J. (Eds.),Starch: Basic Science to Biotechnology. Academic Press, San Diego, pp. 1e12.
Staller, J.E., Thompson, R.G., 2002. A multidisciplinary approach to understandingthe initial introduction of maize into coastal Ecuador. J. Archaeol. Sci. 29, 33e50.
Stromberg, C.A.E., 2007. Can slide preparation methods cause size biases in phy-tolith assemblages? Results from a preliminary study. In: Madella, M., Zurro, D.,Jones, M.K. (Eds.), Places, People and Plants: Using Phytoliths in Archaeologyand Paleoecology. Oxbow Books, Oxford, pp. 1e12.
Tang, H., Ando, H., Watanabe, K., Takeda, Y., Mitsunaga, T., 2001. Physiochemicalproperties and structure of large, medium and small starches in fractions ofnormal barley endosperm. Carbohyd. Res. 330, 241e248.
Tester, R.F., Debon, S.J.J., 2000. Annealing of starch- a review. Int. J. Biol. Macromol.27 (1), 1e12.
Tester, R.F., Morrison, W.R., 1990. Swelling and gelatinisation of cerealstarches I. Effects of amylopectin, amylose and lipids. Cereal Chem. 67,551e559.
Therin, M.R., Fullager, R., Torrence, R., 1999. Starch in sediment: a new approachto the study of subsistence and land use in Papua New Guinea. In:Gosden, C., Hather, J. (Eds.), Prehistory of Food. Routledge, London,pp. 438e462.
Torrence, R., Barton, H. (Eds.), 2006. Ancient Starch Research. Left Coast Press,California.
Valamoti, S.-M., Samuel, D., Bayram, M., Marinova, E., 2008. Prehistoric cereal foodsfrom Greece and Bulgaria: investigation of starch microstructure in experi-mental and archaeological charred remains. Veg. Hist. Archaeobot 17 (1),265e276.
Van Peer, P., Fullager, R., Stokes, S., Bailey, R.M., Moeyersons, J., Steenhoudt, F.,Geerts, A., Vanderbeken, T., De Dapper, M., Geus, F., 2003. The early to MiddleStone Age transition and the emergence of modern human behaviour at site 8-B-11, Sai Island, Sudan. J. Hum. Evol. 45, 187e193.
Vojti�sková, L., Munzarová, E., Votrubová, O., Rihova, A., Ju�ricová, B., 2004. Growthand biomass allocation of sweet flag (Acorus calamus L.) under differentnutrient conditions. Hydrobiologia 518 (1e3), 9e22.
Wagnon, K.A., 1946. Acorns as feed for range cattle. West. Livestock Journal 25 (6),92e94.
Wilson, J., Hardy, K., Allen, R., Copeland, L., Wrangham, R., Collins, M., 2010. Auto-mated classification of starch granules using supervised pattern recognition ofmorphological properties. J. Archaeol. Sci. 37 (3), 594e604.
Yang, X., Yu, J., Lü, H., Cui, T., Guo, J., Ge, Q., 2009. Starch grain analysis revealsfunction of grinding stone tools at Shanghai site, Beijing. Sci. China Ser. D EarthSci. 52 (8), 1164e1171.
Zarillo, S., Pearsall, D.M., Raymond, J.S., Tisdale, M.A., Quon, D.J., 2008. Directlydated starch residues document early formative maize (Zea mays L.) in tropicalEcuador. Proc. Nat. Acad. Sci. 105 (13), 5006e5011.
Zvelebil, M., 1994. Plant use in the Mesolithic and its role in the transition tofarming. Proc. Prehist. Soc. 60, 35e74.
Related Documents











