A Subset of Host B Lymphocytes Controls Melanoma Metastasis through a Melanoma Cell Adhesion Molecule/MUC18-Dependent Interaction: Evidence from Mice and Humans Fernanda I. Staquicini, 1 Anita Tandle, 2 Steven K. Libutti, 2 Jessica Sun, 1 Maya Zigler, 1 Menashe Bar-Eli, 1 Fabiana Aliperti, 3 Elizabeth C. Pe ´rez, 3 Jeffrey E. Gershenwald, 1 Mario Mariano, 3 Renata Pasqualini, 1 Wadih Arap, 1 and Jose ´ Daniel Lopes 3 1 The University of Texas M. D. Anderson Cancer Center, Houston, Texas; 2 Surgery Branch, National Cancer Institute, Bethesda, Maryland; and 3 Sa ˜o Paulo Federal University, Sa ˜o Paulo, Brazil Abstract Host immunity affects tumor metastasis but the corre- sponding cellular and molecular mechanisms are not entirely clear. Here, we show that a subset of B lymphocytes (termed B-1 population), but not other lymphocytes, has prometastatic effects on melanoma cells in vivo through a direct hetero- typic cell-cell interaction. In the classic B16 mouse mela- noma model, one mechanism underlying this phenomenon is a specific up-regulation and subsequent homophilic inter- action mediated by the cell surface glycoprotein MUC18 (also known as melanoma cell adhesion molecule). Presence of B-1 lymphocytes in a panel of tumor samples from melanoma patients directly correlates with MUC18 expression in mela- noma cells, indicating that the same protein interaction exists in humans. These results suggest a new but as yet unrecog- nized functional role for host B-1 lymphocytes in tumor metastasis and establish a biochemical basis for such observations. Our findings support the counterintuitive central hypothesis in which a primitive layer of the immune system actually contributes to tumor progression and metas- tasis in a mouse model and in melanoma patients. Given that monoclonal antibodies against MUC18 are in preclinical development but the reason for their antitumor activity is not well understood, these translational results are relevant in the setting of human melanoma and perhaps of other cancers. [Cancer Res 2008;68(20):8419–28] Introduction Studies addressing the role of the immune system in tumor growth and metastasis have yielded conflicting and often counterintuitive results. Over the 1970s, Prehn and colleagues proposed that the immune response mediated by lymphoid cells could paradoxically lead to tumor cell stimulation (1–3). To date, the interplay of immunity, inflammation, and cancer is still not entirely understood (4, 5). To add a further level of complexity— depending on the experimental model used—it is evident that host immunity can actually lead to enhancement, suppression, or even no effect at all on the metastatic potential of tumor cells, so that no global generalizations can be easily made (6). Specifically in the B16 mouse melanoma model, previous reports show that melanoma cells can be stimulated by lymphocytes (7) and that melanoma progression can indeed be delayed if tumor- bearing mice are rendered immunosuppressed (8). However, the basis for these intriguing experimental observations remains elusive. In particular, the relevance of cell subpopulations from the more primitive layers of the immune system such as B-1 lymphocytes (9–12) on tumor phenotype has not been fully elucidated, although clues for such a role have recently emerged (13, 14). Here, we have evaluated the cellular and molecular cross-talk by which B-1 lymphocytes affect melanoma growth and metastasis. First, we used the classic B16 mouse melanoma model to show that one mechanism accounting for this observation is the up- regulation and subsequent homophilic interaction of the cell surface glycoprotein MUC18 (also known as melanoma cell adhesion molecule). Next, we show that B-1 lymphocytes are also present in human tumors and directly correlate with MUC18 expression in melanoma cells, indicating that the same functional mechanism is conserved across species and likely active in human disease. Together, our results strongly suggest an important role for host B-1 lymphocytes in melanoma-derived metastasis and its corresponding biochemical basis in tumor-bearing mice and in patients. Materials and Methods Animals. Female mice were purchased and housed in the animal facilities of the University of Texas M. D. Anderson Cancer Center, Federal University of Sa ˜o Paulo, or University of Campinas. All animal procedures were approved by the respective Institutional Animal Care and Use Committee. Human specimens. Incidental human melanoma samples were obtained, through written informed consent, from patients treated at the Surgery Branch of the National Cancer Institute (NCI) or at The University of Texas M. D. Anderson Cancer Center. Reagents. Anti-MUC18 (mouse and human) antibodies were purchased from Santa Cruz Biotechnology and Zymed. Antibacteriophage (Sigma) and FITC-conjugated anti-human IgM, allophycocyanin-conjugated anti-human CD5, and phycoerythrin (PE)-conjugated anti-human MUC18 (BD Bio- sciences) were commercially obtained. MART-1 antibody was purchased from BioGenex and labeled with FITC by using EZ-Label FITC Protein Labeling kit (Pierce) and Zeba desalt spin columns (Pierce). Horsera- dish peroxidase (HRP)-conjugated anti-rabbit, PE-conjugated anti-mouse (Pharmingen), Cy3-conjugated anti-rabbit antibodies were purchased from Note: Supplementary data for this article are available at Cancer Research Online (http://cancerres.aacrjournals.org/). Requests for reprints: Wadih Arap and Renata Pasqualini, The University of Texas M. D. Anderson Cancer Center, 1515 Holcombe Boulevard, Houston, TX 77030. Phone: 713-792-3873; Fax: 713-745-2999; E-mail: [email protected] and [email protected] or Jose ´ Daniel Lopes, Federal University of Sa ˜o Paulo, Rua Botucatu 862, Sa ˜o Paulo 04023-062, Brazil. Phone: 55-011-5576-4532; Fax: 55-011- 5571-1095; E-mail: [email protected]. I2008 American Association for Cancer Research. doi:10.1158/0008-5472.CAN-08-1242 www.aacrjournals.org 8419 Cancer Res 2008; 68: (20). October 15, 2008 Research Article Research. on February 25, 2016. © 2008 American Association for Cancer cancerres.aacrjournals.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A Subset of Host B Lymphocytes Controls Melanoma Metastasis
through a Melanoma Cell Adhesion Molecule/MUC18-Dependent
Interaction: Evidence from Mice and Humans
Fernanda I. Staquicini,1Anita Tandle,
2Steven K. Libutti,
2Jessica Sun,
1Maya Zigler,
1
Menashe Bar-Eli,1Fabiana Aliperti,
3Elizabeth C. Perez,
3Jeffrey E. Gershenwald,
1
Mario Mariano,3Renata Pasqualini,
1Wadih Arap,
1and Jose Daniel Lopes
3
1The University of Texas M. D. Anderson Cancer Center, Houston, Texas; 2Surgery Branch, National Cancer Institute, Bethesda, Maryland;and 3Sao Paulo Federal University, Sao Paulo, Brazil
Abstract
Host immunity affects tumor metastasis but the corre-sponding cellular and molecular mechanisms are not entirelyclear. Here, we show that a subset of B lymphocytes (termedB-1 population), but not other lymphocytes, has prometastaticeffects on melanoma cells in vivo through a direct hetero-typic cell-cell interaction. In the classic B16 mouse mela-noma model, one mechanism underlying this phenomenonis a specific up-regulation and subsequent homophilic inter-action mediated by the cell surface glycoprotein MUC18 (alsoknown as melanoma cell adhesion molecule). Presence of B-1lymphocytes in a panel of tumor samples from melanomapatients directly correlates with MUC18 expression in mela-noma cells, indicating that the same protein interaction existsin humans. These results suggest a new but as yet unrecog-nized functional role for host B-1 lymphocytes in tumormetastasis and establish a biochemical basis for suchobservations. Our findings support the counterintuitivecentral hypothesis in which a primitive layer of the immunesystem actually contributes to tumor progression and metas-tasis in a mouse model and in melanoma patients. Given thatmonoclonal antibodies against MUC18 are in preclinicaldevelopment but the reason for their antitumor activity isnot well understood, these translational results are relevant inthe setting of human melanoma and perhaps of other cancers.[Cancer Res 2008;68(20):8419–28]
Introduction
Studies addressing the role of the immune system in tumorgrowth and metastasis have yielded conflicting and oftencounterintuitive results. Over the 1970s, Prehn and colleaguesproposed that the immune response mediated by lymphoid cellscould paradoxically lead to tumor cell stimulation (1–3). To date,the interplay of immunity, inflammation, and cancer is still notentirely understood (4, 5). To add a further level of complexity—
depending on the experimental model used—it is evident thathost immunity can actually lead to enhancement, suppression,or even no effect at all on the metastatic potential of tumor cells,so that no global generalizations can be easily made (6).Specifically in the B16 mouse melanoma model, previous reports
show that melanoma cells can be stimulated by lymphocytes (7)and that melanoma progression can indeed be delayed if tumor-bearing mice are rendered immunosuppressed (8). However, thebasis for these intriguing experimental observations remainselusive. In particular, the relevance of cell subpopulations fromthe more primitive layers of the immune system such as B-1lymphocytes (9–12) on tumor phenotype has not been fullyelucidated, although clues for such a role have recently emerged(13, 14).Here, we have evaluated the cellular and molecular cross-talk by
which B-1 lymphocytes affect melanoma growth and metastasis.First, we used the classic B16 mouse melanoma model to show thatone mechanism accounting for this observation is the up-regulation and subsequent homophilic interaction of the cellsurface glycoprotein MUC18 (also known as melanoma celladhesion molecule). Next, we show that B-1 lymphocytes are alsopresent in human tumors and directly correlate with MUC18expression in melanoma cells, indicating that the same functionalmechanism is conserved across species and likely active in humandisease. Together, our results strongly suggest an important rolefor host B-1 lymphocytes in melanoma-derived metastasis andits corresponding biochemical basis in tumor-bearing mice andin patients.
Materials and Methods
Animals. Female mice were purchased and housed in the animal
facilities of the University of Texas M. D. Anderson Cancer Center, Federal
University of Sao Paulo, or University of Campinas. All animal procedureswere approved by the respective Institutional Animal Care and Use
Committee.
Human specimens. Incidental human melanoma samples were
obtained, through written informed consent, from patients treated at theSurgery Branch of the National Cancer Institute (NCI) or at The University
of Texas M. D. Anderson Cancer Center.
Reagents. Anti-MUC18 (mouse and human) antibodies were purchasedfrom Santa Cruz Biotechnology and Zymed. Antibacteriophage (Sigma) and
FITC-conjugated anti-human IgM, allophycocyanin-conjugated anti-human
CD5, and phycoerythrin (PE)-conjugated anti-human MUC18 (BD Bio-
sciences) were commercially obtained. MART-1 antibody was purchasedfrom BioGenex and labeled with FITC by using EZ-Label FITC Protein
Labeling kit (Pierce) and Zeba desalt spin columns (Pierce). Horsera-
dish peroxidase (HRP)-conjugated anti-rabbit, PE-conjugated anti-mouse
(Pharmingen), Cy3-conjugated anti-rabbit antibodies were purchased from
Note: Supplementary data for this article are available at Cancer Research Online(http://cancerres.aacrjournals.org/).Requests for reprints:Wadih Arap and Renata Pasqualini, The University of Texas
M. D. Anderson Cancer Center, 1515 Holcombe Boulevard, Houston, TX 77030.Phone: 713-792-3873; Fax: 713-745-2999; E-mail: [email protected] [email protected] or Jose Daniel Lopes, Federal University of Sao Paulo,Rua Botucatu 862, Sao Paulo 04023-062, Brazil. Phone: 55-011-5576-4532; Fax: 55-011-5571-1095; E-mail: [email protected].
I2008 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-08-1242
www.aacrjournals.org 8419 Cancer Res 2008; 68: (20). October 15, 2008
Research Article
Research. on February 25, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

Jackson ImmunoResearch Laboratories. Keyhole limpet hemocyanin–conjugated peptides and synthetic peptides were synthesized and
conjugated to our specifications (AnaSpec).
Cell culture and coculture of B-1 lymphocytes and B16 melanomacells. B16-derived melanoma cells (The Jackson Laboratory) were culturedin RPMI-1640 (Sigma) containing 10% of fetal bovine serum (Cultilab),
antibiotics, and supplements. Purified B-1 lymphocytes were obtained as
described (15). Only samples showing >95% purity were used.
Tumor growth and experimental metastasis assays. We used astandard model (16, 17) to deplete B-1 lymphocytes in mice. Untreated,
radiated, or reconstituted cohorts of C57BL/6 mice received B16 cells i.v.
(105 per mouse). Mice were sacrificed and the number of colonies on the
surface of lungs was determined on day 15 after administration. Primarytumor growth into the mouse footpad was measured daily.
Phage display screening and binding assays.We used a random phage
library displaying the insert CX7C (C, cysteine; X, any residue) for selection
of peptides binding to melanoma cells after coculture with B-1 lympho-
cytes (18). As a preclearing step, 106 B16 cells without exposure to B-1
lymphocytes were detached, washed, and resuspended in RPMI-1640
containing 2% bovine serum albumin (BSA) plus 109 transducing units
(TU) of unselected phage library. Cells and phage were transferred to the
top of a nonmiscible organic lower phase [dibutyl phthalate/cyclohexane,
9:1 (v:v)] and centrifuged at 10,000 � g for 10 min. The unbound phage
population remaining in the aqueous upper phase (precleared library) was
collected into a fresh Eppendorf tube and incubated with 106 B16 cells
isolated after coculture with B-1 lymphocytes. Phage in the organic lower
phase was recovered from the cell pellet by bacterial host infection (19–24).
For phage binding assays to B16 melanoma, 106 cells before and after
coculture with B-1 lymphocytes were incubated with each specific phage
clone (109 TU) or negative controls. Melanoma cells and phage were
centrifuged through the organic phase and the cell-bound phage clones
were recovered by bacterial infection (18).
Immunocapture assays. Immunocapture experiments were with anti-MUC18 or IgG control antibodies, as described (19). ELISA with anti-IgG
confirmed equal molar concentration of IgG on each of the wells. Afterblocking with PBS containing 3% BSA, 30 Ag of protein from cell membrane
extracts were added onto the wells for overnight incubation. Following
washes, phage (2 � 109 TU) was added to each well. Bound phage were
recovered by bacterial infection.In vivo phage display. Homing of phage to s.c. tumors was performed as
described (25). Animals received 1 � 1010 TU of phage diluted in DMEM.
Tumors and control organs were collected after 6 h of circulation. Boundphage was recovered by bacterial infection (25).
Immunofluorescence and flow cytometry. B16 melanoma cells beforeand after coculture with B-1 lymphocytes were seeded in an eight-chamber
slide (Nalge Nunc International) and incubated with phage (109 TU). Cellswere washed, fixed, and incubated with an antibacteriophage antibody
followed by secondary antibody. For flow cytometry, melanoma cells or
purified B-1 lymphocytes were incubated with primary antibody anti-
MUC18 followed by PE-conjugated secondary antibody. To investigate thepresence of B-1 lymphocytes in human melanoma samples, cells were
isolated, washed, fixed, and permeabilized with BD Cytofix/Cytoperm (BD
Biosciences). Cells were stained either with isotype control or with specific
antibody. Cells were analyzed with a FACSCalibur machine (BD Bioscien-ces) equipped with CellQuest software.
Immunofluorescence and immunohistochemical staining forMUC18 detection in tissue specimens. Tissue specimens were sectioned,mounted, and air dried for 24 h. Antigen retrieval was performed with
0.1 mol/L citrate buffer (pH 6). Sections were stained with the UltraVision
Plus Detection System Anti-Polyvalent, HRP/AEC kit (LabVision Corp.) andcounterstained with Gill’s hematoxylin (Sigma). For immunofluorescence,
sections were washed, blocked, and incubated with specific antibodies and
Cy3-conjugated secondary antibodies.
Western blot and immunoprecipitation assays. Cells were lysed byusing PBS containing 250 mmol/L sucrose, 50 mmol/L octylglucoside, 1
mmol/L EDTA, and protease inhibitors, resolved in a 4% to 20% gradientSDS-PAGE gel, transferred to nitrocellulose membranes, and developed with
the enhanced chemiluminescence reagent (Amersham Pharmacia). Fordetection of phosphorylated extracellular signal-regulated kinase 1/2
(ERK1/2), total proteins were extracted as described (14).
Reverse transcription-PCR. RNA was purified by using the Perfect
RNA Mini kit extraction method (Eppendorf). First-strand cDNA synthesiswas performed by using the SuperScript II Reverse Transcriptase kit
(Invitrogen). For the mouse MUC18 transcript amplification, we used the
primers 5¶-GGATCCTTGGCTTGCGCCCTCCGTCGG-3¶ and 5¶-CTAATGCC-TCAGATCGATGTATTTCTCTCC-3¶ under the same conditions for templatedenaturation and elongation but with the annealing temperature of 60jC.As a loading control, we used primers for the mouse glyceraldehyde-
3-phosphate dehydrogenase: 5¶-CGCCTGGTCACCAGGGCTGC-3¶ and 5¶-CA-CCACCCTGTTGCTGTAGCC-3¶.Design of small hairpin RNA and lentivirus production. Mouse
MUC18 small interfering RNA (siRNA) sequences 5¶-GGAGAGAAATAC-ATCGATC-3¶ and 5¶-GATCGATGTATTTCTCTCC-3¶ were obtained fromDharmacon (On-Target Plus, NM_023061). Nonspecific control siRNA
[nontargeting short hairpin RNA (shRNA)] sequences were 5¶-TAAGGC-TATGAAGAGATAC-3¶ and 5¶-GTATCTCTTCATAGCCTTA-3¶. shRNAsequences for both targeting and nontargeting were ligated into a lentiviralvector pLVTHM, which drives the expression of the green fluorescent
protein (GFP; a gift from Dr. Didier Trono, University of Geneva, Geneva,
Switzerland; ref. 26). The restriction enzymes Cla1 and Mlu1 were used.
The lentiviruses were produced by infecting human embryonic kidneycells (293FT) with the sequence-verified pLVTHM, the packing plasmid
(MD2G), and the envelope plasmid (PAX2), required for viral production.
GFP-positive cells were enriched to 100% by fluorescence-activated cellsorting (FACS).
Statistical analysis. Graphical analyses (balloon plots) were used to
depict protein expression levels based on flow cytometry results. Spear-
man’s rank correlation test was used to analyze the correlation betweennumber of B-1 lymphocytes and MUC18 expression profile on patients.
Statistical analysis of in vivo experiments was carried out by using Student’s
t tests as indicated.
Results
B-1 lymphocytes influence malignant melanoma metastasisin vivo . We first evaluated the role of B-1 lymphocytes inmelanoma growth and metastasis in vivo by selectively depletingthe predominantly B-1 lymphocyte population from peritoneal andpleural surfaces of mice (11, 12, 15, 27). We used external beamionizing radiation to deplete B-1 lymphocytes with no detectableeffect on other cell types (16, 17). We confirmed the depletion byflow cytometry analysis of cell surface markers: we observed asevere reduction in the B-1 lymphocyte population (typically over80% cell depletion) by using this procedure (Supplementary Figs. S1and S2). Next, we compared s.c. melanoma growth and experi-mental metastasis in radiated versus nonradiated (control) mice(Fig. 1A). In the radiated cohorts, we observed tumor growthsuppression (Fig. 1A, left) and marked reduction in melanomametastasis (Fig. 1A, right). In either case, reconstitution with totalperitoneal cells reverted tumor growth and metastasis to levelsundistinguishable from those observed in control nonradiatedmice. To evaluate which depleted cell population mediates thisphenomenon, we reconstituted radiated mice with either B-1lymphocytes (Fig. 1B) or all other resident peritoneal cells but B-1lymphocytes (Fig. 1C). We show that B-1 lymphocytes are necessaryand sufficient to revert the radiation-induced metastasis suppres-sion of melanoma. Finally, by using an unrelated genetic model ofimmunosuppression [X-linked immunodeficiency (Xid)], we alsoobserved melanoma metastasis inhibition when mutant mice(constitutively B-1 lymphocyte deficient; refs. 28, 29) werecompared with their otherwise isogenic wild-type counterparts
Cancer Research
Cancer Res 2008; 68: (20). October 15, 2008 8420 www.aacrjournals.org
Research. on February 25, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
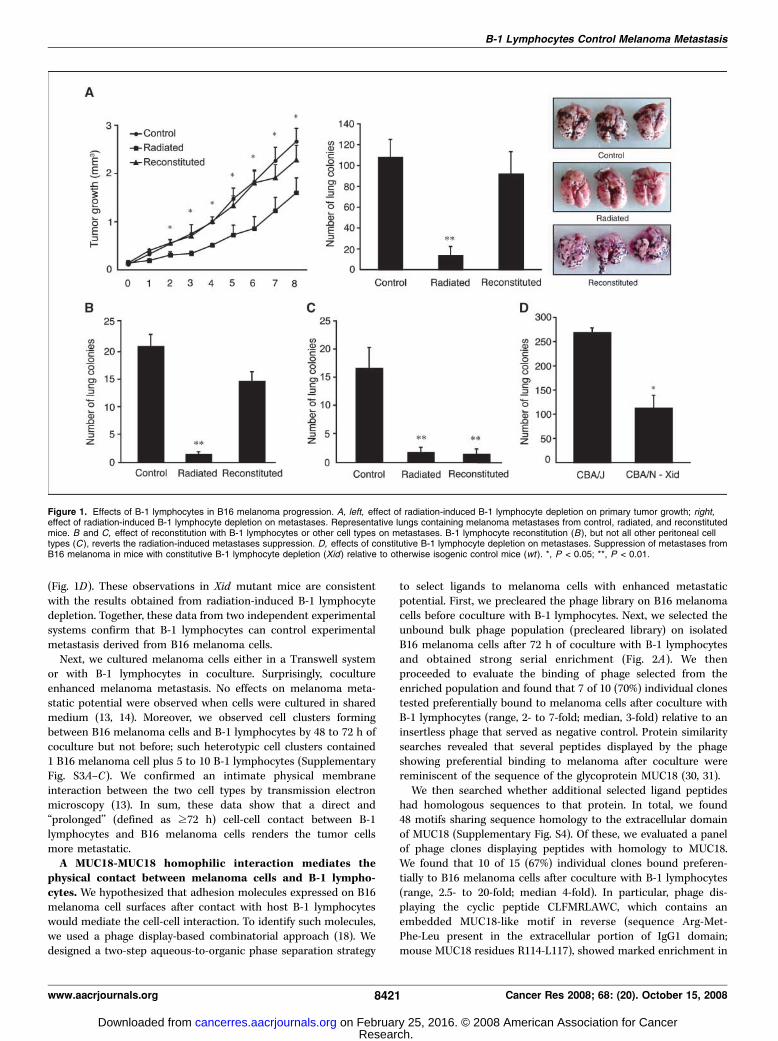
(Fig. 1D). These observations in Xid mutant mice are consistentwith the results obtained from radiation-induced B-1 lymphocytedepletion. Together, these data from two independent experimentalsystems confirm that B-1 lymphocytes can control experimentalmetastasis derived from B16 melanoma cells.Next, we cultured melanoma cells either in a Transwell system
or with B-1 lymphocytes in coculture. Surprisingly, cocultureenhanced melanoma metastasis. No effects on melanoma meta-static potential were observed when cells were cultured in sharedmedium (13, 14). Moreover, we observed cell clusters formingbetween B16 melanoma cells and B-1 lymphocytes by 48 to 72 h ofcoculture but not before; such heterotypic cell clusters contained1 B16 melanoma cell plus 5 to 10 B-1 lymphocytes (SupplementaryFig. S3A–C). We confirmed an intimate physical membraneinteraction between the two cell types by transmission electronmicroscopy (13). In sum, these data show that a direct and‘‘prolonged’’ (defined as z72 h) cell-cell contact between B-1lymphocytes and B16 melanoma cells renders the tumor cellsmore metastatic.A MUC18-MUC18 homophilic interaction mediates the
physical contact between melanoma cells and B-1 lympho-cytes. We hypothesized that adhesion molecules expressed on B16melanoma cell surfaces after contact with host B-1 lymphocyteswould mediate the cell-cell interaction. To identify such molecules,we used a phage display-based combinatorial approach (18). Wedesigned a two-step aqueous-to-organic phase separation strategy
to select ligands to melanoma cells with enhanced metastaticpotential. First, we precleared the phage library on B16 melanomacells before coculture with B-1 lymphocytes. Next, we selected theunbound bulk phage population (precleared library) on isolatedB16 melanoma cells after 72 h of coculture with B-1 lymphocytesand obtained strong serial enrichment (Fig. 2A). We thenproceeded to evaluate the binding of phage selected from theenriched population and found that 7 of 10 (70%) individual clonestested preferentially bound to melanoma cells after coculture withB-1 lymphocytes (range, 2- to 7-fold; median, 3-fold) relative to aninsertless phage that served as negative control. Protein similaritysearches revealed that several peptides displayed by the phageshowing preferential binding to melanoma after coculture werereminiscent of the sequence of the glycoprotein MUC18 (30, 31).We then searched whether additional selected ligand peptides
had homologous sequences to that protein. In total, we found48 motifs sharing sequence homology to the extracellular domainof MUC18 (Supplementary Fig. S4). Of these, we evaluated a panelof phage clones displaying peptides with homology to MUC18.We found that 10 of 15 (67%) individual clones bound preferen-tially to B16 melanoma cells after coculture with B-1 lymphocytes(range, 2.5- to 20-fold; median 4-fold). In particular, phage dis-playing the cyclic peptide CLFMRLAWC, which contains anembedded MUC18-like motif in reverse (sequence Arg-Met-Phe-Leu present in the extracellular portion of IgG1 domain;mouse MUC18 residues R114-L117), showed marked enrichment in
Figure 1. Effects of B-1 lymphocytes in B16 melanoma progression. A, left, effect of radiation-induced B-1 lymphocyte depletion on primary tumor growth; right,effect of radiation-induced B-1 lymphocyte depletion on metastases. Representative lungs containing melanoma metastases from control, radiated, and reconstitutedmice. B and C, effect of reconstitution with B-1 lymphocytes or other cell types on metastases. B-1 lymphocyte reconstitution (B), but not all other peritoneal celltypes (C ), reverts the radiation-induced metastases suppression. D, effects of constitutive B-1 lymphocyte depletion on metastases. Suppression of metastases fromB16 melanoma in mice with constitutive B-1 lymphocyte depletion (Xid ) relative to otherwise isogenic control mice (wt ). *, P < 0.05; **, P < 0.01.
B-1 Lymphocytes Control Melanoma Metastasis
www.aacrjournals.org 8421 Cancer Res 2008; 68: (20). October 15, 2008
Research. on February 25, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
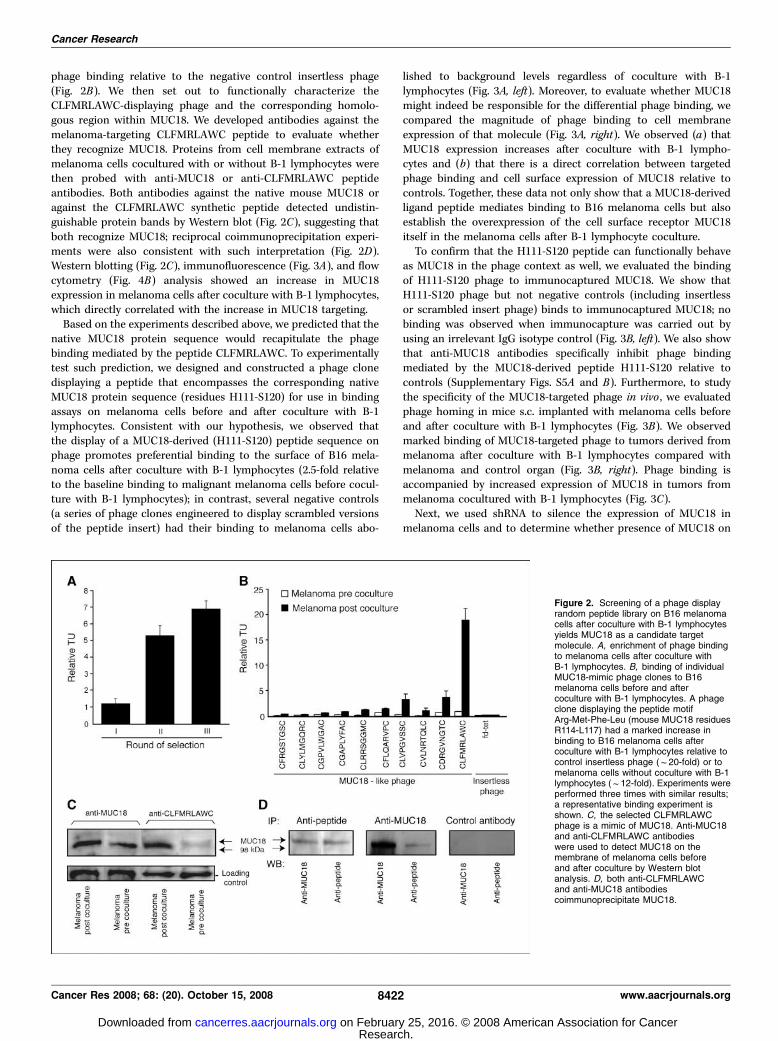
phage binding relative to the negative control insertless phage(Fig. 2B). We then set out to functionally characterize theCLFMRLAWC-displaying phage and the corresponding homolo-gous region within MUC18. We developed antibodies against themelanoma-targeting CLFMRLAWC peptide to evaluate whetherthey recognize MUC18. Proteins from cell membrane extracts ofmelanoma cells cocultured with or without B-1 lymphocytes werethen probed with anti-MUC18 or anti-CLFMRLAWC peptideantibodies. Both antibodies against the native mouse MUC18 oragainst the CLFMRLAWC synthetic peptide detected undistin-guishable protein bands by Western blot (Fig. 2C), suggesting thatboth recognize MUC18; reciprocal coimmunoprecipitation experi-ments were also consistent with such interpretation (Fig. 2D).Western blotting (Fig. 2C), immunofluorescence (Fig. 3A), and flowcytometry (Fig. 4B) analysis showed an increase in MUC18expression in melanoma cells after coculture with B-1 lymphocytes,which directly correlated with the increase in MUC18 targeting.Based on the experiments described above, we predicted that the
native MUC18 protein sequence would recapitulate the phagebinding mediated by the peptide CLFMRLAWC. To experimentallytest such prediction, we designed and constructed a phage clonedisplaying a peptide that encompasses the corresponding nativeMUC18 protein sequence (residues H111-S120) for use in bindingassays on melanoma cells before and after coculture with B-1lymphocytes. Consistent with our hypothesis, we observed thatthe display of a MUC18-derived (H111-S120) peptide sequence onphage promotes preferential binding to the surface of B16 mela-noma cells after coculture with B-1 lymphocytes (2.5-fold relativeto the baseline binding to malignant melanoma cells before cocul-ture with B-1 lymphocytes); in contrast, several negative controls(a series of phage clones engineered to display scrambled versionsof the peptide insert) had their binding to melanoma cells abo-
lished to background levels regardless of coculture with B-1lymphocytes (Fig. 3A, left). Moreover, to evaluate whether MUC18might indeed be responsible for the differential phage binding, wecompared the magnitude of phage binding to cell membraneexpression of that molecule (Fig. 3A, right). We observed (a) thatMUC18 expression increases after coculture with B-1 lympho-cytes and (b) that there is a direct correlation between targetedphage binding and cell surface expression of MUC18 relative tocontrols. Together, these data not only show that a MUC18-derivedligand peptide mediates binding to B16 melanoma cells but alsoestablish the overexpression of the cell surface receptor MUC18itself in the melanoma cells after B-1 lymphocyte coculture.To confirm that the H111-S120 peptide can functionally behave
as MUC18 in the phage context as well, we evaluated the bindingof H111-S120 phage to immunocaptured MUC18. We show thatH111-S120 phage but not negative controls (including insertlessor scrambled insert phage) binds to immunocaptured MUC18; nobinding was observed when immunocapture was carried out byusing an irrelevant IgG isotype control (Fig. 3B, left). We also showthat anti-MUC18 antibodies specifically inhibit phage bindingmediated by the MUC18-derived peptide H111-S120 relative tocontrols (Supplementary Figs. S5A and B). Furthermore, to studythe specificity of the MUC18-targeted phage in vivo , we evaluatedphage homing in mice s.c. implanted with melanoma cells beforeand after coculture with B-1 lymphocytes (Fig. 3B). We observedmarked binding of MUC18-targeted phage to tumors derived frommelanoma after coculture with B-1 lymphocytes compared withmelanoma and control organ (Fig. 3B, right). Phage binding isaccompanied by increased expression of MUC18 in tumors frommelanoma cocultured with B-1 lymphocytes (Fig. 3C).Next, we used shRNA to silence the expression of MUC18 in
melanoma cells and to determine whether presence of MUC18 on
Figure 2. Screening of a phage displayrandom peptide library on B16 melanomacells after coculture with B-1 lymphocytesyields MUC18 as a candidate targetmolecule. A, enrichment of phage bindingto melanoma cells after coculture withB-1 lymphocytes. B, binding of individualMUC18-mimic phage clones to B16melanoma cells before and aftercoculture with B-1 lymphocytes. A phageclone displaying the peptide motifArg-Met-Phe-Leu (mouse MUC18 residuesR114-L117) had a marked increase inbinding to B16 melanoma cells aftercoculture with B-1 lymphocytes relative tocontrol insertless phage (f20-fold) or tomelanoma cells without coculture with B-1lymphocytes (f12-fold). Experiments wereperformed three times with similar results;a representative binding experiment isshown. C, the selected CLFMRLAWCphage is a mimic of MUC18. Anti-MUC18and anti-CLFMRLAWC antibodieswere used to detect MUC18 on themembrane of melanoma cells beforeand after coculture by Western blotanalysis. D, both anti-CLFMRLAWCand anti-MUC18 antibodiescoimmunoprecipitate MUC18.
Cancer Research
Cancer Res 2008; 68: (20). October 15, 2008 8422 www.aacrjournals.org
Research. on February 25, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
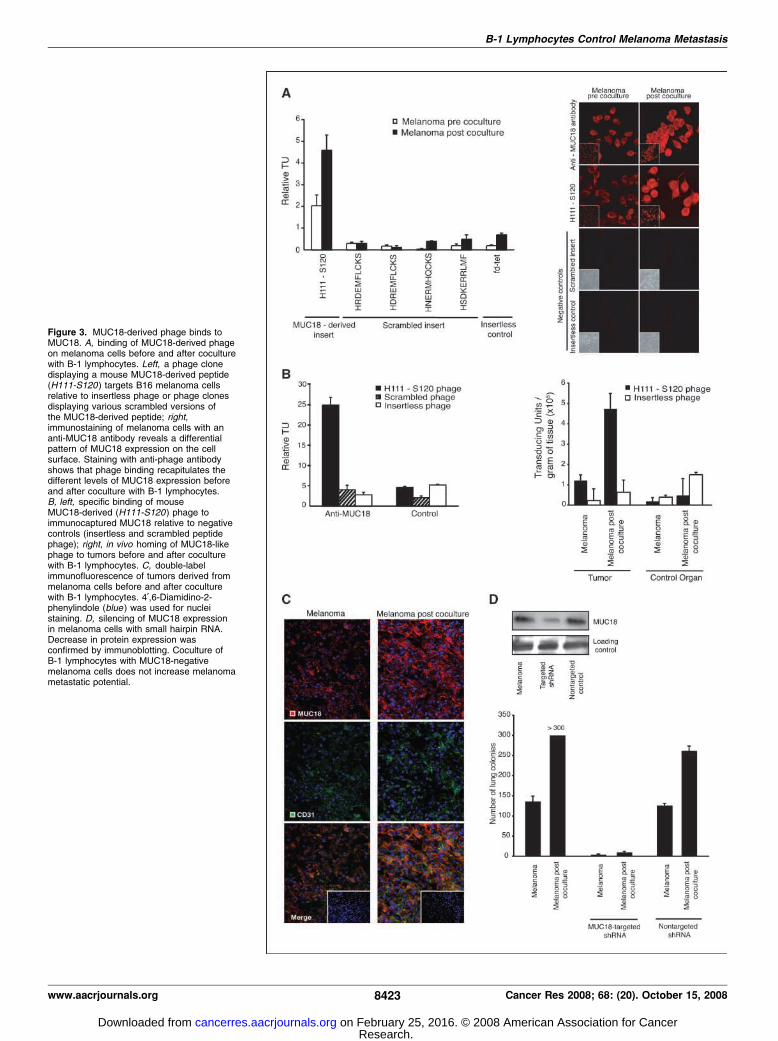
Figure 3. MUC18-derived phage binds toMUC18. A, binding of MUC18-derived phageon melanoma cells before and after coculturewith B-1 lymphocytes. Left, a phage clonedisplaying a mouse MUC18-derived peptide(H111-S120) targets B16 melanoma cellsrelative to insertless phage or phage clonesdisplaying various scrambled versions ofthe MUC18-derived peptide; right,immunostaining of melanoma cells with ananti-MUC18 antibody reveals a differentialpattern of MUC18 expression on the cellsurface. Staining with anti-phage antibodyshows that phage binding recapitulates thedifferent levels of MUC18 expression beforeand after coculture with B-1 lymphocytes.B, left, specific binding of mouseMUC18-derived (H111-S120) phage toimmunocaptured MUC18 relative to negativecontrols (insertless and scrambled peptidephage); right, in vivo homing of MUC18-likephage to tumors before and after coculturewith B-1 lymphocytes. C, double-labelimmunofluorescence of tumors derived frommelanoma cells before and after coculturewith B-1 lymphocytes. 4¶,6-Diamidino-2-phenylindole (blue ) was used for nucleistaining. D, silencing of MUC18 expressionin melanoma cells with small hairpin RNA.Decrease in protein expression wasconfirmed by immunoblotting. Coculture ofB-1 lymphocytes with MUC18-negativemelanoma cells does not increase melanomametastatic potential.
B-1 Lymphocytes Control Melanoma Metastasis
www.aacrjournals.org 8423 Cancer Res 2008; 68: (20). October 15, 2008
Research. on February 25, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
the cell surface is required for the biological phenomenon to occur.Decrease in expression of MUC18 was confirmed by immunoblot-ting (Fig. 3D). MUC18-depleted cells were cocultured with B-1lymphocytes for 72 h and injected i.v. into mice. We used B16and B16 transduced with nontargeting shRNA as controls. As pre-viously observed, coculture of MUC18-expressing melanoma cells(parental B16 or B16 transduced with nontargeting shRNA) withB-1 lymphocytes increases melanoma metastasis. However, suchprometastatic effect is abrogated when MUC18-negative cells areused, a result consistent with our hypothesis that a MUC18-MUC18-mediated cell interaction renders melanoma cells moremetastatic. Furthermore, a marked decrease in the number oflung colonies was observed in animals inoculated with MUC18-negative cells, again supporting the importance of this moleculein metastasis.Taken together, these results confirm the specificity of the
interaction and support the concept that a MUC18-MUC18homophilic interaction mediates the physical contact betweenB16 cells and B-1 lymphocytes. To gain insight into the molecularbasis of such interaction, we generated a panel of phage tocombine alanine scanning site-directed mutagenesis and bindingassays. Compared with wild-type H111-S120 phage, we identifiedfour key residues (Arg114, Cys118, Lys119, and Ser120) in MUC18whose mutation abolished phage binding to melanoma cellsregardless of coculture with B-1 lymphocytes (SupplementaryFig. S5C). Results of these mutational studies again indicatebinding specificity.B-1 lymphocytes express MUC18. Given that phage selected to
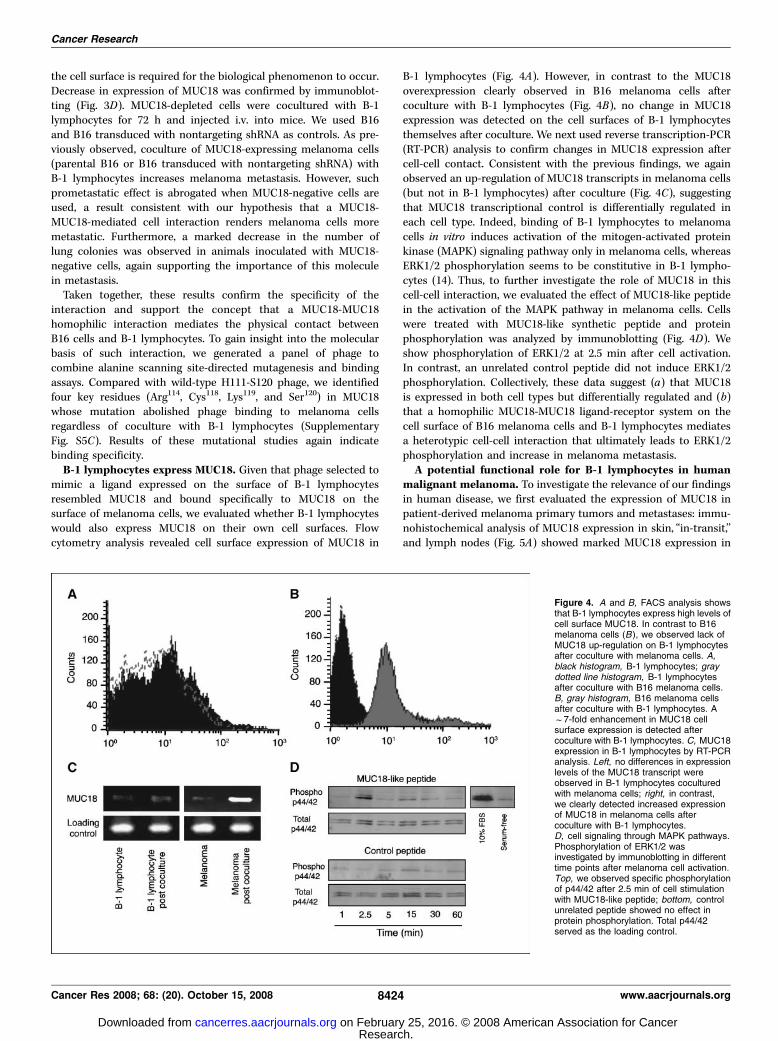
mimic a ligand expressed on the surface of B-1 lymphocytesresembled MUC18 and bound specifically to MUC18 on thesurface of melanoma cells, we evaluated whether B-1 lymphocyteswould also express MUC18 on their own cell surfaces. Flowcytometry analysis revealed cell surface expression of MUC18 in
B-1 lymphocytes (Fig. 4A). However, in contrast to the MUC18overexpression clearly observed in B16 melanoma cells aftercoculture with B-1 lymphocytes (Fig. 4B), no change in MUC18expression was detected on the cell surfaces of B-1 lymphocytesthemselves after coculture. We next used reverse transcription-PCR(RT-PCR) analysis to confirm changes in MUC18 expression aftercell-cell contact. Consistent with the previous findings, we againobserved an up-regulation of MUC18 transcripts in melanoma cells(but not in B-1 lymphocytes) after coculture (Fig. 4C), suggestingthat MUC18 transcriptional control is differentially regulated ineach cell type. Indeed, binding of B-1 lymphocytes to melanomacells in vitro induces activation of the mitogen-activated proteinkinase (MAPK) signaling pathway only in melanoma cells, whereasERK1/2 phosphorylation seems to be constitutive in B-1 lympho-cytes (14). Thus, to further investigate the role of MUC18 in thiscell-cell interaction, we evaluated the effect of MUC18-like peptidein the activation of the MAPK pathway in melanoma cells. Cellswere treated with MUC18-like synthetic peptide and proteinphosphorylation was analyzed by immunoblotting (Fig. 4D). Weshow phosphorylation of ERK1/2 at 2.5 min after cell activation.In contrast, an unrelated control peptide did not induce ERK1/2phosphorylation. Collectively, these data suggest (a) that MUC18is expressed in both cell types but differentially regulated and (b)that a homophilic MUC18-MUC18 ligand-receptor system on thecell surface of B16 melanoma cells and B-1 lymphocytes mediatesa heterotypic cell-cell interaction that ultimately leads to ERK1/2phosphorylation and increase in melanoma metastasis.A potential functional role for B-1 lymphocytes in human
malignant melanoma. To investigate the relevance of our findingsin human disease, we first evaluated the expression of MUC18 inpatient-derived melanoma primary tumors and metastases: immu-nohistochemical analysis of MUC18 expression in skin, ‘‘in-transit,’’and lymph nodes (Fig. 5A) showed marked MUC18 expression in
Figure 4. A and B, FACS analysis showsthat B-1 lymphocytes express high levels ofcell surface MUC18. In contrast to B16melanoma cells (B ), we observed lack ofMUC18 up-regulation on B-1 lymphocytesafter coculture with melanoma cells. A,black histogram, B-1 lymphocytes; graydotted line histogram, B-1 lymphocytesafter coculture with B16 melanoma cells.B, gray histogram, B16 melanoma cellsafter coculture with B-1 lymphocytes. Af7-fold enhancement in MUC18 cellsurface expression is detected aftercoculture with B-1 lymphocytes. C, MUC18expression in B-1 lymphocytes by RT-PCRanalysis. Left, no differences in expressionlevels of the MUC18 transcript wereobserved in B-1 lymphocytes coculturedwith melanoma cells; right, in contrast,we clearly detected increased expressionof MUC18 in melanoma cells aftercoculture with B-1 lymphocytes.D, cell signaling through MAPK pathways.Phosphorylation of ERK1/2 wasinvestigated by immunoblotting in differenttime points after melanoma cell activation.Top, we observed specific phosphorylationof p44/42 after 2.5 min of cell stimulationwith MUC18-like peptide; bottom, controlunrelated peptide showed no effect inprotein phosphorylation. Total p44/42served as the loading control.
Cancer Research
Cancer Res 2008; 68: (20). October 15, 2008 8424 www.aacrjournals.org
Research. on February 25, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
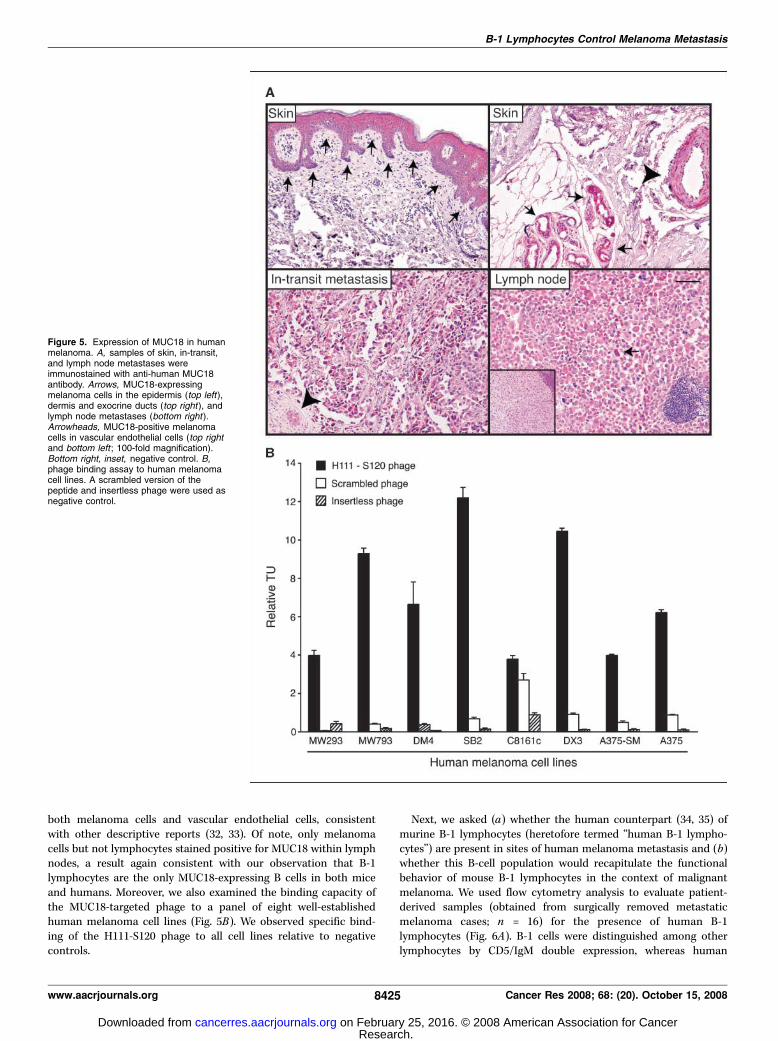
both melanoma cells and vascular endothelial cells, consistentwith other descriptive reports (32, 33). Of note, only melanomacells but not lymphocytes stained positive for MUC18 within lymphnodes, a result again consistent with our observation that B-1lymphocytes are the only MUC18-expressing B cells in both miceand humans. Moreover, we also examined the binding capacity ofthe MUC18-targeted phage to a panel of eight well-establishedhuman melanoma cell lines (Fig. 5B). We observed specific bind-ing of the H111-S120 phage to all cell lines relative to negativecontrols.
Next, we asked (a) whether the human counterpart (34, 35) ofmurine B-1 lymphocytes (heretofore termed ‘‘human B-1 lympho-cytes’’) are present in sites of human melanoma metastasis and (b)whether this B-cell population would recapitulate the functionalbehavior of mouse B-1 lymphocytes in the context of malignantmelanoma. We used flow cytometry analysis to evaluate patient-derived samples (obtained from surgically removed metastaticmelanoma cases; n = 16) for the presence of human B-1lymphocytes (Fig. 6A). B-1 cells were distinguished among otherlymphocytes by CD5/IgM double expression, whereas human
Figure 5. Expression of MUC18 in humanmelanoma. A, samples of skin, in-transit,and lymph node metastases wereimmunostained with anti-human MUC18antibody. Arrows, MUC18-expressingmelanoma cells in the epidermis (top left),dermis and exocrine ducts (top right ), andlymph node metastases (bottom right ).Arrowheads, MUC18-positive melanomacells in vascular endothelial cells (top rightand bottom left ; 100-fold magnification).Bottom right, inset, negative control. B,phage binding assay to human melanomacell lines. A scrambled version of thepeptide and insertless phage were used asnegative control.
B-1 Lymphocytes Control Melanoma Metastasis
www.aacrjournals.org 8425 Cancer Res 2008; 68: (20). October 15, 2008
Research. on February 25, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

MUC18-expressing tumor cells were identified within MART-1+
melanoma cells. Expression of surface markers was graphicallyrepresented as a ‘‘balloon plot’’ (Fig. 6A). We observed a directcorrelation between increasing number of human B-1 lymphocyteswithin the tumors and increasing expression of MUC18 onmelanoma cells (r = 0.6; P < 0.05). Histologic analysis of repre-sentative melanomas with mild (patients 1 and 3), moderate(patients 8 and 9), and marked (patient 13) levels of MUC18expression on melanoma cells illustrates differential expressionof the protein (Fig. 6B). Together, these results establish thathuman B-1 lymphocytes are present in melanoma metastasesand that such presence accounts for increased expression ofMUC18 in human melanoma cells. As such, the MUC18 homophilicmechanism of heterotypic cell interaction seems clearly pre-served across species and it is likely functional in humanmelanoma as well.
Discussion
It is often assumed that host immunity assembles its com-ponents to curb disease; however, the functional relationshipbetween cellular immune system and tumor cells is far from clear.In mice, B-1 lymphocytes arise very early during the ontogeny ofthe immune system compared with the more evolved layers. Such
cell lineage share several features with primitive B lymphocytessuch as neonatal development, limited antibody repertoire, self-renewal capacity, and regulatory feedback in adults. B-1 lympho-cytes can be readily identified by their cell surface phenotype(IgMhigh, IgDlow, CD23�, B220low, and CD11b+) and can be furthersubdivided in B-1a (CD5+) and B-1b (CD5�) subsets (9–12).We evaluated whether B-1 lymphocytes influence tumor
progression in the well-established B16 melanoma mouse model.We found that B-1 lymphocyte depletion markedly decreases lungmetastasis from melanoma cells. B-1 lymphocyte reconstitutionspecifically inhibits this phenomenon and restores metastasis tobaseline levels. These results show that B-1 lymphocytes have a keyrole in melanoma growth and metastasis.Given that B-1 lymphocytes are known to produce and release
high amounts of cytokines, we originally designed experiments toevaluate whether a soluble factor such as, for instance, interleukin-10 (36) might account for the observed metastasis enhancement.Unexpectedly, we found instead that a direct physical interactionbetween B-1 lymphocytes and melanoma cells is needed formetastasis. These findings led to the hypothesis that cell surfacemolecules are likely to mediate the phenomenon.To gain insight into the molecular mechanism of this cell-cell
interaction and the resulting biological effects, we attempted toidentify the functionally active molecules expressed at the surface
Figure 6. Correlation between numberof B-1 lymphocytes and MUC18 expressionin human melanoma. A, flow cytometryanalysis of melanoma samples isgraphically represented as a balloon plot.A positive correlation is observed betweenincreasing number of B-1 lymphocyteswithin the tumors and increasingexpression of MUC18 on melanoma cells.B, histologic analysis of representativesamples stained for MUC18 (20-foldmagnification).
Cancer Research
Cancer Res 2008; 68: (20). October 15, 2008 8426 www.aacrjournals.org
Research. on February 25, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

of each cell type. To that end, we selected a combinatorial peptidelibrary on melanoma cells after coculture with B-1 lymphocytes. Bycomparing sequences of selected peptides with those available inprotein databases, we found that ligand motifs were similar to thecell surface glycoprotein MUC18. Because MUC18 is an adhe-sion molecule that correlates with tumor growth and metastasis(32, 37, 38), we next evaluated whether a MUC18-dependentcell interaction would influence melanoma metastasis. First, weestablished that MUC18 serves as the partner on the surface of B16melanoma cells (‘‘receptor’’). Because MUC18-like peptides alsobind to MUC18 itself, we reasoned that MUC18 could also serveas the partner on the surface of B-1 lymphocytes (‘‘ligand’’). If so,one would predict that the ligand-receptor system affecting B16melanoma cells through B-1 lymphocytes is actually a MUC18-MUC18 interaction. Our data show that this is indeed the casebecause (a) MUC18-like peptides enhance phage binding tomelanoma cells after coculture, (b) antibodies against MUC18specifically inhibit phage targeting, and (c) MUC18 is also abundanton the surface of B-1 lymphocytes. Together, our findings indicatethat an unrecognized biochemical interaction between MUC18expressed on the surface of B16 melanoma cells and B-1 lympho-cytes regulates metastatic potential. Although the identification ofa MUC18-like motif ‘‘in reverse’’ may originally have suggested anantiparallel MUC18-MUC18 interaction, in vitro and in vivoexperiments performed with a phage clone designed to displaythe corresponding native MUC18 sequence indicate that thisparticular molecular mimicry is not affected by peptide orienta-tion. However, the same may not necessarily be true for the nativeMUC18 protein itself (due to potential steric hindrance). Therefore,whether this protein-protein interaction is influenced by orienta-tion of the native protein expressed on the surface of melanomaand B-1 cells remains uncertain; a full understanding of thestructural requirements for protein-protein or protein-peptide willlikely have to wait for the elucidation of the X-ray crystal structuresof MUC18-MUC18 and of MUC18-CLFMRLAWC complexes.To evaluate whether these observations are relevant in human
disease, we investigated the distribution of B-1 lymphocytes inpatients with melanoma. Although the origin and characteristics ofthe ‘‘human B-1 lymphocyte’’ counterpart are still poorly defined, aconsiderable proportion of IgM+ B cells in the human peritonealcavity are CD5+, a phenotypic hallmark of mouse B-1 lymphocytes.Mice and human B-1 lymphocytes are both largely responsible forthe production of autoreactive IgM antibodies in patients withcertain autoimmune diseases (34, 35).We detected IgM+/CD5+ B cells, presumably human B-1
lymphocytes, in 100% of the analyzed samples from a cohort ofpatients with metastatic melanoma (n = 16), suggesting that B-1
lymphocytes likely play a functional role in human cancer.Importantly, the presence of B-1 lymphocytes within tumorsdirectly correlates with increased expression of MUC18 inmelanoma cells (38), again supporting the relevance of a functionaland active MUC18-dependent cross-talk between human B-1lymphocytes and melanoma.Notably, disruption of MUC18-dependent cell interactions may
be of therapeutic value. In fact, (a) anti-MUC18 antibodies haveshown promise in preclinical models (37, 39) and (b) overexpres-sion of MUC18 occurs in human melanoma among other tumortypes (40, 41). One might speculate that if a MUC18-dependentinteraction accounts for malignant melanoma homing to lung vas-culature for instance, then anti-MUC18 antibodies and/or MUC18-based peptidomimetics might simultaneously block the functionalprotein-protein interaction from both partners. Considering thatreattachment of circulating tumor cells is a rate-limiting step inmetastasis, our observations might help explain the difference ofmagnitude of effect we observed between the (mild to moderate)B16 tumor growth suppression relative to lung (marked) metastasisinhibition in B-1 lymphocyte-deficient mice.In summary, we show that human and mouse melanoma cells
can subvert a putatively defensive function of the immune systemthrough a heterotypic cell-cell interaction with primitive B cellsand that this molecular cross-talk can influence tumor progressionwith adverse net effects to the host. New mechanism-basedstrategies, such as physical elimination or functional modulation ofhuman B-1 lymphocytes (through radiation or specific antibodies),may be considered as an experimental therapy against humanmelanomas; also, blocking MUC18 with antibodies or targetingMUC18 for ligand-directed delivery of agents in patients withmelanoma may have translational potential.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
Received 4/2/2008; revised 7/31/2008; accepted 8/11/2008.Grant support: Fundacao de Amparo a Pesquisa do Estado de Sao Paulo (J.D.
Lopes), Department of Defense (F.I. Staquicini, R. Pasqualini, and W. Arap), NCI (R.Pasqualini and W. Arap), Gillson-Longenbaugh Foundation, and AngelWorks. F.I.Staquicini received a predoctoral fellowship from the Fundacao de Amparo a Pesquisado Estado de Sao Paulo.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.We thank Dr. Richmond Prehn, Dr. Isaiah J. Fidler, Dr. Leonore A. Herzenberg, and
Dr. Olivera J. Finn for critical reading of the manuscript and helpful advice, Dr. RenatoMortara for confocal microscopy assistance, Dr. Alexander Lazar for immunohisto-chemistry, and Dr. John Wunderlich from the TIL Laboratory (NCI Surgery Branch) forhuman melanoma samples.
B-1 Lymphocytes Control Melanoma Metastasis
www.aacrjournals.org 8427 Cancer Res 2008; 68: (20). October 15, 2008
References
1. Prehn RT. Perspectives on oncogenesis: does immu-nity stimulate or inhibit neoplasia? J ReticuloendothelSoc 1971;10:1–16.2. Prehn RT. The immune reaction as a stimulator oftumor growth. Science 1972;176:170–1.3. Wexler H, Sindelar WF, Ketcham AS. The role ofimmune factors in the survival of circulating tumorcells. Cancer 1976;37:1701–6.4. Coussens LM, Werb Z. Inflammation and cancer.Nature 2002;420:860–7.5. de Visser KE, Korets LV, Coussens LM. De novocarcinogenesis promoted by chronic inflammation is Blymphocyte dependent. Cancer Cell 2005;7:411–23.
6. Fidler IJ. Molecular biology of cancer: invasion andmetastasis In: Devita VT, Hellman S, Rosenberg SA,editors. Cancer: principles and practice of oncology. Vol.1. New York: Lippincott-Raven; 1997. p. 135–52.7. Bartholomaeus WN, Bray AE, Papadimitriou JM, KeastD. Immune response to a transplantable malignantmelanoma in mice. J Natl Cancer Inst 1974;53:1065–72.8. Fidler IJ, Gersten DM. Effect of syngeneic lymphocyteson the vascularity, growth and induced metastasis of theB16 melanoma. In: Crispen RG, editor, Neoplasmimmunity: experimental and clinical. Vol. 3. Amsterdam:Elsevier North Holland; 1980. p. 3–15.9. Berland R, Wortis HH. Origins and functions of B-1cells with notes on the role of CD5. Annu Rev Immunol2002;20:253–300.
10. Hardy RR, Hayakawa K. Development and physiologyof Ly-1 B and its human homolog, Leu-1 B. Immunol Rev1986;93:53–79.11. Kantor AB, Herzenberg LA. Origin of murine B celllineages. Annu Rev Immunol 1993;11:501–38.12. Fagarasan S, Watanabe N, Honjo T. Generation,expansion, migration and activation of mouse B1 cells.Immunol Rev 2000;176:205–15.13. Staquicini FI. B-1 lymphocytes modulate the meta-static potential of murine melanoma through a specificinteraction mediated by MUC18 [dissertation]. SaoPaulo (Brazil): Federal University of Sao Paulo; 2004.14. Perez EC, Machado J, Jr., Aliperti F, Freymuller E,Mariano M, Lopes JD. B-1 lymphocytes increasemetastatic behavior of melanoma cells through the
Research. on February 25, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

Cancer Research
Cancer Res 2008; 68: (20). October 15, 2008 8428 www.aacrjournals.org
extracellular signal-regulated kinase pathway. CancerSci 2008;99:920–8.15. Almeida SR, Aroeira LS, Freymuller E, et al. Mouse B-1 cell-derived mononuclear phagocyte, a novel cellularcomponent of acute non-specific inflammatory exudate.Int Immunol 2001;13:1193–201.16. e Brito RR, De Lorenzo BH, Xander P, et al. Role ofdistinct immune components in the radiation-inducedabrogation of systemic lupus erythematosus develop-ment in mice. Lupus 2007;16:947–54.17. Popi AF, Lopes JD, Mariano M. Interleukin-10 secretedby B-1 cells modulates the phagocytic activity of murinemacrophages in vitro . Immunology 2004;113:348–54.18. Giordano RJ, Cardo-Vila M, Lahdenranta J, PasqualiniR, Arap W. Biopanning and rapid analysis of selectiveinteractive ligands. Nat Med 2001;7:1249–53.19. Pasqualini R, Koivunen E, Kain R, et al. AminopeptidaseN is a receptor for tumor-homing peptides and a target forinhibiting angiogenesis. Cancer Res 2000;60:722–7.20. Cardo-Vila M, Arap W, Pasqualini R. avh5 integrin-dependent programmed cell death triggered by apeptide mimic of annexin V. Mol Cell 2003;11:1151–62.21. Marchio S, Lahdenranta J, Schlingemann RO, et al.Aminopeptidase A is a functional target in angiogenicblood vessels. Cancer Cell 2004;5:151–62.22. Kolonin MG, Saha PK, Chan L, Pasqualini R, Arap W.Reversal of obesity by targeted ablation of adiposetissue. Nat Med 2004;10:625–32.23. Arap MA, Lahdenranta J, Mintz PJ, et al. Cell surfaceexpression of the stress response chaperone GRP78enables tumor targeting by circulating ligands. CancerCell 2004;6:275–84.
24. Hajitou A, Trepel M, Lilley CE, et al. A hybrid vectorfor ligand-directed tumor targeting and molecularimaging. Cell 2006;125:385–98.25. Pasqualini R, Ruoslahti E. Organ targeting in vivousing phage display peptide libraries. Nature 1996;380:364–6.26. Wiznerowicz M, Trono D. Conditional suppression ofcellular genes: lentivirus vector-mediated drug-inducibleRNA interference. J Virol 2003;77:8957–61.27. Kantor AB. The development and repertoire of B-1cells (CD5 B cells). Immunol Today 1991;12:389–91.28. Khan WN, Alt FW, Gerstein RM, et al. Defective B celldevelopment and function in Btk-deficient mice.Immunity 1995;3:283–99.29. Santos-Lima EC, Vasconcellos R, Reina-San-Martin B,et al. Significant association between the skewed naturalantibody repertoire of Xid mice and resistance toTrypanosoma cruzi infection. Eur J Immunol 2001;31:634–45.30. Shih IM. The role of CD146 (Mel-CAM) in biology andpathology. J Pathol 1999;189:4–11.31. Yang H, Wang S, Liu Z, et al. Isolation andcharacterization of mouse MUC18 cDNA gene, andcorrelation of MUC18 expression in mouse mela-noma cell lines with metastatic ability. Gene 2001;265:133–45.32. Jean D, Gershenwald JE, Huang S, et al. Lossof AP-2 results in up-regulation of MCAM/MUC18 andan increase in tumor growth and metastasis of humanmelanoma cells. J Biol Chem 1998;273:16501–8.33. Luca M, Hunt B, Bucana CD, Johnson JP, Fidler IJ,Bar-Eli M. Direct correlation between MUC18 expres-
sion and metastatic potential of human melanoma cells.Melanoma Res 1993;3:35–41.34. Casali P, Burastero SE, Nakamura M, Inghirami G,Notkins AL. Human lymphocytes making rheumatoidfactor and antibody to ssDNA belong to Leu-1+ B-cellsubset. Science 1987;236:77–81.35. Hardy RR, Hayakawa K, Shimizu M, Yamasaki K,Kishimoto T. Rheumatoid factor secretion from humanLeu-1+ B cells. Science 1987;236:81–3.36. Gieni RS, Umetsu DT, DeKruyff RH. Ly1� (CD5�) Bcells produce interleukin (IL)-10. Cell Immunol 1997;175:164–70.37. McGary EC, Heimberger A, Mills L, et al. A fullyhuman antimelanoma cellular adhesion molecule/MUC18 antibody inhibits spontaneous pulmonarymetastasis of osteosarcoma cells in vivo . Clin CancerRes 2003;9:6560–6.38. McGary EC, Lev DC, Bar-Eli M. Cellular adhesionpathways and metastatic potential of human melanoma.Cancer Biol Ther 2002;1:459–65.39. Mills L, Tellez C, Huang S, et al. Fully humanantibodies to MCAM/MUC18 inhibit tumor growth andmetastasis of human melanoma. Cancer Res 2002;62:5106–14.40. Filshie RJ, Zannettino AC, Makrynikola V, et al.MUC18, a member of the immunoglobulin superfamily,is expressed on bone marrow fibroblasts and a subset ofhematological malignancies. Leukemia 1998;12:414–21.41. Wu GJ, Fu P, Chiang CF, et al. Increased expression ofMUC18 correlates with the metastatic progression ofmouse prostate adenocarcinoma in the TRAMP model.J Urol 2005;173:1778–83.
Research. on February 25, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

2008;68:8419-8428. Cancer Res Fernanda I. Staquicini, Anita Tandle, Steven K. Libutti, et al. and HumansMolecule/MUC18-Dependent Interaction: Evidence from MiceMetastasis through a Melanoma Cell Adhesion A Subset of Host B Lymphocytes Controls Melanoma
Updated version
http://cancerres.aacrjournals.org/content/68/20/8419
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2008/10/13/68.20.8419.DC1.html
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/68/20/8419.full.html#ref-list-1
This article cites 38 articles, 10 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/68/20/8419.full.html#related-urls
This article has been cited by 5 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on February 25, 2016. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
Related Documents











