A short proregion of trialysin, a pore-forming protein of Triatoma infestans salivary glands, controls activity by folding the N-terminal lytic motif Rafael M. Martins 1 , Rogerio Amino 2 , Katia R. Daghastanli 3 , Iolanda M. Cuccovia 3 , Maria A. Juliano 4 and Sergio Schenkman 1 1 Departamento de Microbiologia, Imunologia e Parasitologia, Universidade Federal de Sa ˜o Paulo, Brazil 2 Departamento de Bioquı ´mica, Universidade Federal de Sa ˜o Paulo, Brazil 3 Departamento de Bioquı ´mica, Instituto de Quı ´mica, Universidade de Sa ˜o Paulo, Brazil 4 Departamento de Biofı ´sica, Universidade Federal de Sa ˜o Paulo, Brazil Hematophagous animals counteract physical and molecular barriers such as the epidermis and the inflammatory, hemostatic and immune systems of the hosts to fulfill their nutritional needs [1]. Therefore, their saliva has evolved for the specific task of circum- venting several biochemical cascades to facilitate blood acquisition. Triatomine insects are exclusive blood- feeders that transmit Chagas’ disease, acquiring Try- panosoma cruzi from the blood of infected mammalian hosts, and transmitting this parasite through their feces, instead of injecting protozoa during the bite, like anophelines, sand-flies, or tsetse flies [2]. In the salivary secretion of Triatoma infestans are found three differ- ent anticoagulant activities [3]: proteases [4], a sialidase [5], apyrases [6], an inhibitor of platelet aggregation [7], a Na + channel blocker [8], and a cytolytic protein named trialysin [9]. Trialysin is a pore-forming protein that permeabilizes several cell types, from bacteria to mammalian cells. Synthetic peptides based on the mature amphipathic N-terminus of trialysin (first 37 amino acids) induce bacterial, protozoal and mammalian cell membrane permeabilization, and their solution structures show characteristics of cationic amphipathic antimicrobial peptides [10]. The lytic activities presented by trialysin, as well as by other pore-forming proteins and peptides, must be well controlled in order to avoid destroying the membrane compartments during their synthesis and secretion. The cecropin and melittin antimicrobial peptides are synthesized as larger polypeptides that are Keywords membrane lysis; salivary gland; trialysin; Triatoma infestans; Trypanosoma cruzi Correspondence S. Schenkman, Departamento de Microbiologia, Imunologia e Parasitologia, Rua Botucatu 862 8 o andar 04023-062 Sa ˜o Paulo, SP, Brazil Fax: +55 11 5571 58 77 Tel: +55 11 5575 19 96 E-mail: [email protected] (Received 30 August 2007, revised 3 December 2007, accepted 2 January 2008) doi:10.1111/j.1742-4658.2008.06260.x Triatoma infestans (Hemiptera: Reduviidae) is a hematophagous insect that transmits the protozoan parasite Trypanosoma cruzi, the etiological agent of Chagas’ disease. Its saliva contains trialysin, a protein that forms pores in membranes. Peptides based on the N-terminus of trialysin lyse cells and fold into a-helical amphipathic segments resembling antimicrobial peptides. Using a specific antiserum against trialysin, we show here that trialysin is synthesized as a precursor that is less active than the protein released after saliva secretion. A synthetic peptide flanked by a fluorophore and a quencher including the acidic proregion and the lytic N-terminus of the protein is also less active against cells and liposomes, increasing activity upon proteolysis. Activation changes the peptide conformation as observed by fluorescence increase and CD spectroscopy. This mechanism of activa- tion could provide a way to impair the toxic effects of trialysin inside the salivary glands, thus restricting damaging lytic activity to the bite site. Abbreviations Abz, o-aminobenzoic acid; APMSF, (4-amidinophenyl)methanesulfonyl fluoride; GST, glutathione S-transferase; LUV, large unilamellar vesicle; NL, nonlytic; PGS, peptide between glycine and serine; Q-EDDnp, Gln-n-(2,4-dinitrophenyl)-ethylenediamine. 994 FEBS Journal 275 (2008) 994–1002 ª 2008 The Authors Journal compilation ª 2008 FEBS

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A short proregion of trialysin, a pore-forming proteinof Triatoma infestans salivary glands, controls activityby folding the N-terminal lytic motifRafael M. Martins1, Rogerio Amino2, Katia R. Daghastanli3, Iolanda M. Cuccovia3, Maria A. Juliano4
and Sergio Schenkman1
1 Departamento de Microbiologia, Imunologia e Parasitologia, Universidade Federal de Sao Paulo, Brazil
2 Departamento de Bioquımica, Universidade Federal de Sao Paulo, Brazil
3 Departamento de Bioquımica, Instituto de Quımica, Universidade de Sao Paulo, Brazil
4 Departamento de Biofısica, Universidade Federal de Sao Paulo, Brazil
Hematophagous animals counteract physical and
molecular barriers such as the epidermis and the
inflammatory, hemostatic and immune systems of the
hosts to fulfill their nutritional needs [1]. Therefore,
their saliva has evolved for the specific task of circum-
venting several biochemical cascades to facilitate blood
acquisition. Triatomine insects are exclusive blood-
feeders that transmit Chagas’ disease, acquiring Try-
panosoma cruzi from the blood of infected mammalian
hosts, and transmitting this parasite through their
feces, instead of injecting protozoa during the bite, like
anophelines, sand-flies, or tsetse flies [2]. In the salivary
secretion of Triatoma infestans are found three differ-
ent anticoagulant activities [3]: proteases [4], a sialidase
[5], apyrases [6], an inhibitor of platelet aggregation
[7], a Na+ channel blocker [8], and a cytolytic protein
named trialysin [9]. Trialysin is a pore-forming protein
that permeabilizes several cell types, from bacteria to
mammalian cells. Synthetic peptides based on the
mature amphipathic N-terminus of trialysin (first
37 amino acids) induce bacterial, protozoal and
mammalian cell membrane permeabilization, and their
solution structures show characteristics of cationic
amphipathic antimicrobial peptides [10].
The lytic activities presented by trialysin, as well as
by other pore-forming proteins and peptides, must
be well controlled in order to avoid destroying the
membrane compartments during their synthesis and
secretion. The cecropin and melittin antimicrobial
peptides are synthesized as larger polypeptides that are
Keywords
membrane lysis; salivary gland; trialysin;
Triatoma infestans; Trypanosoma cruzi
Correspondence
S. Schenkman, Departamento de
Microbiologia, Imunologia e Parasitologia,
Rua Botucatu 862 8o andar 04023-062 Sao
Paulo, SP, Brazil
Fax: +55 11 5571 58 77
Tel: +55 11 5575 19 96
E-mail: [email protected]
(Received 30 August 2007, revised 3
December 2007, accepted 2 January 2008)
doi:10.1111/j.1742-4658.2008.06260.x
Triatoma infestans (Hemiptera: Reduviidae) is a hematophagous insect that
transmits the protozoan parasite Trypanosoma cruzi, the etiological agent
of Chagas’ disease. Its saliva contains trialysin, a protein that forms pores
in membranes. Peptides based on the N-terminus of trialysin lyse cells and
fold into a-helical amphipathic segments resembling antimicrobial peptides.
Using a specific antiserum against trialysin, we show here that trialysin is
synthesized as a precursor that is less active than the protein released after
saliva secretion. A synthetic peptide flanked by a fluorophore and a
quencher including the acidic proregion and the lytic N-terminus of the
protein is also less active against cells and liposomes, increasing activity
upon proteolysis. Activation changes the peptide conformation as observed
by fluorescence increase and CD spectroscopy. This mechanism of activa-
tion could provide a way to impair the toxic effects of trialysin inside the
salivary glands, thus restricting damaging lytic activity to the bite site.
Abbreviations
Abz, o-aminobenzoic acid; APMSF, (4-amidinophenyl)methanesulfonyl fluoride; GST, glutathione S-transferase; LUV, large unilamellar vesicle;
NL, nonlytic; PGS, peptide between glycine and serine; Q-EDDnp, Gln-n-(2,4-dinitrophenyl)-ethylenediamine.
994 FEBS Journal 275 (2008) 994–1002 ª 2008 The Authors Journal compilation ª 2008 FEBS

processed by proteolytic cleavage, resulting in one
active mature molecule [11,12]. On the other hand, one
prepromagainin polypeptide generates one magainin I
peptide between glycine and serine (PGS) and five mag-
ainin II (PGS-Gly10-Lys22) molecules [13]. The pro-
regions of magainin and melittin are composed of
negatively charged residues that may inhibit their action
[12,13]. Aureins [14], a-defensins (cryptidins) [15], derm-
aseptins [16] and latarcins are also examples of peptides
containing acidic proregions [17]. In contrast, apolar
residues constitute the proregion of cecropin [18]. A
large proregion of approximately 100 residues is present
in cathelicidins, which are mammalian proteins with
multiple functions, including antimicrobial activity [19].
There is structural evidence that the processing of cath-
elicidins is performed in dimeric, domain-swapped
structures that expose the cleavage site and, at the same
time, control their activities, impairing the antimicrobial
activity of the C-terminus [20].
Trialysin cDNA predicts a secretion signal followed
by an acidic domain composed of 33 amino acids,
which is not found in the mature protein. The activa-
tion of protrialysin into mature trialysin was suggested
to occur by limited proteolysis, as it is prevented by
(4-amidinophenyl)methanesulfonyl fluoride (APMSF),
which inhibits the major serine protease of T. infestans
saliva [4]. However, the precursor form was not previ-
ously identified, and it is unknown when it is activated
and whether it prevents lysis induced by trialysin
released with the saliva. To identify the precursor and
further understand how a potent lytic molecule is syn-
thesized and controlled, we raised antibodies that react
with trialysin [205 amino acids long, the nonlytic (NL)
fragment] and tested for the presence of different
forms of the protein and their activity in T. infestans.
We found that a precursor is stored in the salivary
glands and processed only after saliva is released. A
peptide containing the precursor region and the lytic
N-terminus of trialysin was synthesized, containing a
fluorophore and a quencher at the N-terminus and
C-terminus respectively. With use of this peptide,
evidence was obtained showing that the activation
mechanism of trialysin involves conformational
changes in this segment of the protein.
Results
Generation of specific anti-trialysin rabbit serum
Trialysin cDNA predicts a signal sequence followed by
an acidic domain, shown in bold and italicized letters
in Fig. 1A, upstream of the N-terminus sequence
detected in the protein isolated from T. infestans
saliva [9]. In order to detect precursor forms and under-
stand the mechanism of activation of T. infestans
protrialysin, we raised specific antibodies capable of
recognizing salivary trialysin. Attempts to obtain the
full-length precursor and mature forms of the recombi-
nant protein in heterologous systems to immunize
animals were unsuccessful. Therefore, a C-terminal NL
fragment of trialysin corresponding to the underlined
letters in Fig. 1A was subcloned into the pET-14b
plasmid and expressed in BL21 (DE3) pLysE Escheri-
chia coli cells. The recombinant protein was only
soluble in urea. It was purified by Ni-affinity and gel
exclusion chromatography, generating a single band of
21 kDa, as seen by Coomassie Blue staining after
SDS ⁄PAGE (Fig. 1B, lane C). It was used to immunize
rabbits, and the resulting immune serum specifically
recognized the recombinant protein in immunoblots
after SDS ⁄PAGE and a 24 kDa band in the T. infe-
stans saliva (Fig. 1B, lane IB). No signal was obtained
with preimmune or unrelated sera (data not shown).
A
B
Fig. 1. Sequence, expression and purification of the NL fragment
of trialysin. (A) The translated cDNA predicted for preprotrialysin.
The leader peptide sequence is shown in bold letters, the proregion
is shown in italics, and the NL fragment is underlined. The arrow
indicates the N-terminus of the proP7 peptide. Numbers show the
positions of amino acids according to the N-terminus of mature tri-
alysin (+1), the predicted signal sequence ()55, from M1 to A22),
the acidic proregion ()33, from A23 to R55), and the cleavage site
()1, R55). (B) SDS ⁄ PAGE of the NL sequence (NL2) of trialysin
expressed in E. coli BL21 (DE3) pLysE; purified by Ni-affinity chro-
matography after staining with Coomassie Blue (C) and reaction
with antibodies to NL2 trialysin (IB). On the right is shown the
T. infestans saliva stained with antibody to NL2. On the left are
shown the positions of 20 kDa and 25 kDa molecular mass mark-
ers (Benchmark, Invitrogen).
R. M. Martins et al. Trialysin precursor activation
FEBS Journal 275 (2008) 994–1002 ª 2008 The Authors Journal compilation ª 2008 FEBS 995
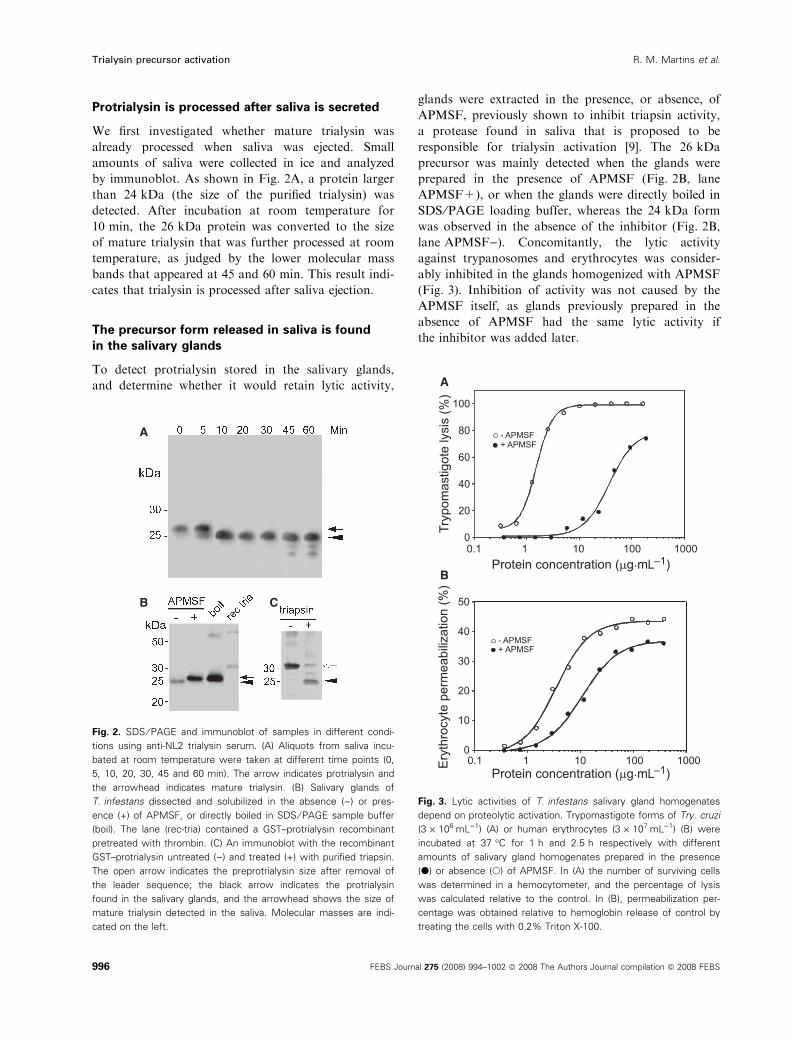
Protrialysin is processed after saliva is secreted
We first investigated whether mature trialysin was
already processed when saliva was ejected. Small
amounts of saliva were collected in ice and analyzed
by immunoblot. As shown in Fig. 2A, a protein larger
than 24 kDa (the size of the purified trialysin) was
detected. After incubation at room temperature for
10 min, the 26 kDa protein was converted to the size
of mature trialysin that was further processed at room
temperature, as judged by the lower molecular mass
bands that appeared at 45 and 60 min. This result indi-
cates that trialysin is processed after saliva ejection.
The precursor form released in saliva is found
in the salivary glands
To detect protrialysin stored in the salivary glands,
and determine whether it would retain lytic activity,
glands were extracted in the presence, or absence, of
APMSF, previously shown to inhibit triapsin activity,
a protease found in saliva that is proposed to be
responsible for trialysin activation [9]. The 26 kDa
precursor was mainly detected when the glands were
prepared in the presence of APMSF (Fig. 2B, lane
APMSF+), or when the glands were directly boiled in
SDS ⁄PAGE loading buffer, whereas the 24 kDa form
was observed in the absence of the inhibitor (Fig. 2B,
lane APMSF)). Concomitantly, the lytic activity
against trypanosomes and erythrocytes was consider-
ably inhibited in the glands homogenized with APMSF
(Fig. 3). Inhibition of activity was not caused by the
APMSF itself, as glands previously prepared in the
absence of APMSF had the same lytic activity if
the inhibitor was added later.
A
B C
Fig. 2. SDS ⁄ PAGE and immunoblot of samples in different condi-
tions using anti-NL2 trialysin serum. (A) Aliquots from saliva incu-
bated at room temperature were taken at different time points (0,
5, 10, 20, 30, 45 and 60 min). The arrow indicates protrialysin and
the arrowhead indicates mature trialysin. (B) Salivary glands of
T. infestans dissected and solubilized in the absence ()) or pres-
ence (+) of APMSF, or directly boiled in SDS ⁄ PAGE sample buffer
(boil). The lane (rec-tria) contained a GST–protrialysin recombinant
pretreated with thrombin. (C) An immunoblot with the recombinant
GST–protrialysin untreated ()) and treated (+) with purified triapsin.
The open arrow indicates the preprotrialysin size after removal of
the leader sequence; the black arrow indicates the protrialysin
found in the salivary glands, and the arrowhead shows the size of
mature trialysin detected in the saliva. Molecular masses are indi-
cated on the left.
A
B
Fig. 3. Lytic activities of T. infestans salivary gland homogenates
depend on proteolytic activation. Trypomastigote forms of Try. cruzi
(3 · 106ÆmL)1) (A) or human erythrocytes (3 · 107ÆmL)1) (B) were
incubated at 37 �C for 1 h and 2.5 h respectively with different
amounts of salivary gland homogenates prepared in the presence
(d) or absence (s) of APMSF. In (A) the number of surviving cells
was determined in a hemocytometer, and the percentage of lysis
was calculated relative to the control. In (B), permeabilization per-
centage was obtained relative to hemoglobin release of control by
treating the cells with 0.2% Triton X-100.
Trialysin precursor activation R. M. Martins et al.
996 FEBS Journal 275 (2008) 994–1002 ª 2008 The Authors Journal compilation ª 2008 FEBS

To identify which form of the protein would represent
the active trialysin, a glutathione S-transferase (GST)–
protrialysin fusion protein was expressed in E. coli. This
protein contained GST, a thrombin cleavage site, and
the protrialysin from amino acid )33 to the C-terminus
(see Fig. 1A). It was obtained from soluble E. coli
extracts and purified by chromatography in a glutathi-
one–Sepharose column. The fusion protein was largely
unstable, and it did not show lytic activity, precipitating
as the protein concentration increased in solution. It
could be processed by thrombin, generating a 30 kDa
protein band in SDS ⁄PAGE (Fig. 2C, lane triapsin)).The processed recombinant protein was also unable to
promote lysis. When partially purified triapsin was
added to the thrombin-cleaved 30 kDa protein, it was
processed to a 24 kDa band (Fig. 2C, lane triapsin+).
In some experiments, lytic activity was detected,
although the resulting protein precipitated and became
inactive, as observed with the trialysin purified from the
saliva [9]. The same processed bands were obtained
when the GST fusion protein was directly incubated
with triapsin, confirming that the 24 kDa molecule
found in saliva is the active form of trialysin, and sug-
gesting that the presence of 10–15 amino acids of the
proregion of protrialysin found in the glands (instead of
the predicted 33-mer acidic propiece) is sufficient to inhi-
bit lytic activity.
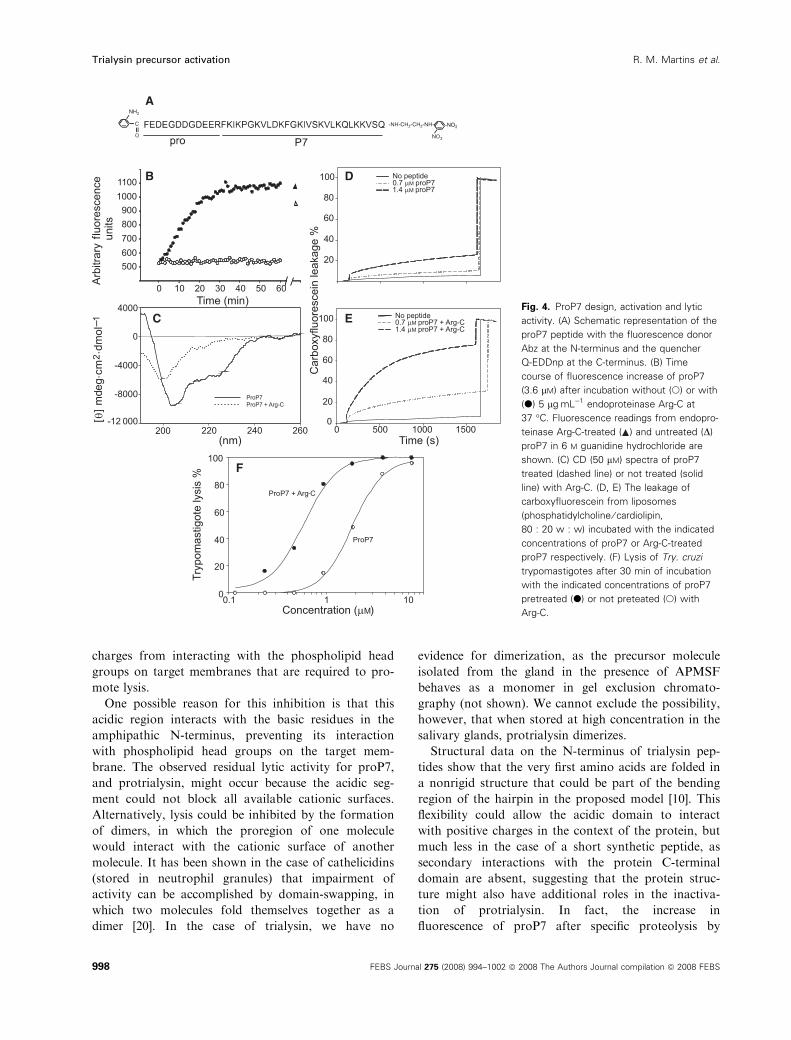
A synthetic peptide containing the proregion and
the N-terminus of trialysin is structured
The amino acid sequence of the N-terminus of the tri-
alysin precursor has not been determined so far either
by Edman degradation or by MS. To investigate how
a small additional negative prosequence could decrease
the lytic activity, a peptide containing 12 residues from
the proregion and 27 residues downstream of the
N-terminus of mature trialysin was synthesized. This
12 residue segment roughly corresponds to the differ-
ence observed between the mature and precursor
forms. The 27-mer segment was previously character-
ized as the lytic peptide P7 [10]. The new peptide
(proP7) is represented in Fig. 4A, and was synthesized
containing at the N-terminus the fluorescent probe
o-aminobenzoic acid (Abz), and at the C-terminus a
fluorescence quencher group Gln-N-(2,4-dinitrophe-
nyl)ethylenediamine (Q-EDDnp). When proP7 was
treated with endoproteinase Arg-C, which cleaves at
the unique Arg of proP7, it released the negative
prosequence from the P7 portion, and this was accom-
panied by an increase in fluorescence over time
(Fig. 4B). A similar increase in fluorescence was
observed when triapsin or saliva was added (not
shown). As quenching of fluorogenic peptides longer
than 40 amino acids is minimal, unless they are folded
[21], this increase in fluorescence suggests that proP7 is
folded before being processed. The fact that proP7
fluorescence in 6 m guanidine hydrochloride was higher
than the fluorescence in nondenaturing conditions con-
firms this hypothesis, although part of the increase in
fluorescence may be due to the peptide hydrolysis,
which abolishes intramolecular quenching. Evidence
that proP7 is structured and that it unfolds after
Arg-C treatment was also obtained by CD spectros-
copy (Fig. 4C). ProP7 contains 36% a-helix, decreas-
ing to 7% after Arg-C treatment.
Lysis increases after cleavage of proP7
Next, the lytic activities of proP7 and Arg-C-processed
peptide were compared by using artificial liposome
membranes (20 : 80 cardiolipin ⁄phosphatidylcholine)containing 6-carboxyfluorescein, which is a fluorophore
that autosuppresses its fluorescence at higher concen-
trations. Upon permeabilization, these liposomes
release the entrapped quenched 6-carboxyfluorescein,
diluting the fluorophore in the sample, and fluores-
cence increases. As expected, the Arg-C-processed
peptide was more effective at promoting liposome
permeabilization than proP7 (Fig. 4D,E). Similar
results were obtained when the lysis of trypanosomes
was assayed (Fig. 4F). These results indicate that a
small acidic portion of the proregion decreases the lysis
efficiency of trialysin in the model using a lytic
synthetic peptide based on the trialysin N-terminus.
Discussion
Here we describe trialysin processing from its accumu-
lation in the salivary glands of T. infestans until its
release during saliva ejection, when the full lytic capac-
ity of the protein is achieved. Judging by the cDNA
sequence, protrialysin should contain a 22 amino acid
prosequence migrating in SDS ⁄PAGE as a 32 kDa
protein. However, we only detected a shorter protein
(26 kDa) that accumulates in the glands and is released
during salivation. This 26 kDa precursor is less active
than mature trialysin, due to no more than 15 amino
acid residues in the proregion. This finding was con-
firmed using an engineered synthetic peptide contain-
ing 12 amino acids from the proregion followed by the
trialysin N-terminus. Upon proteolytic processing, the
precursor-like synthetic peptide unfolds, as observed in
the CD spectra, and lytic activity increases. We pro-
pose that the presence of a short acidic sequence
affects the trialysin N-terminus, preventing its positive
R. M. Martins et al. Trialysin precursor activation
FEBS Journal 275 (2008) 994–1002 ª 2008 The Authors Journal compilation ª 2008 FEBS 997
charges from interacting with the phospholipid head
groups on target membranes that are required to pro-
mote lysis.
One possible reason for this inhibition is that this
acidic region interacts with the basic residues in the
amphipathic N-terminus, preventing its interaction
with phospholipid head groups on the target mem-
brane. The observed residual lytic activity for proP7,
and protrialysin, might occur because the acidic seg-
ment could not block all available cationic surfaces.
Alternatively, lysis could be inhibited by the formation
of dimers, in which the proregion of one molecule
would interact with the cationic surface of another
molecule. It has been shown in the case of cathelicidins
(stored in neutrophil granules) that impairment of
activity can be accomplished by domain-swapping, in
which two molecules fold themselves together as a
dimer [20]. In the case of trialysin, we have no
evidence for dimerization, as the precursor molecule
isolated from the gland in the presence of APMSF
behaves as a monomer in gel exclusion chromato-
graphy (not shown). We cannot exclude the possibility,
however, that when stored at high concentration in the
salivary glands, protrialysin dimerizes.
Structural data on the N-terminus of trialysin pep-
tides show that the very first amino acids are folded in
a nonrigid structure that could be part of the bending
region of the hairpin in the proposed model [10]. This
flexibility could allow the acidic domain to interact
with positive charges in the context of the protein, but
much less in the case of a short synthetic peptide, as
secondary interactions with the protein C-terminal
domain are absent, suggesting that the protein struc-
ture might also have additional roles in the inactiva-
tion of protrialysin. In fact, the increase in
fluorescence of proP7 after specific proteolysis by
A
B D
C
F
E Fig. 4. ProP7 design, activation and lytic
activity. (A) Schematic representation of the
proP7 peptide with the fluorescence donor
Abz at the N-terminus and the quencher
Q-EDDnp at the C-terminus. (B) Time
course of fluorescence increase of proP7
(3.6 lM) after incubation without (s) or with
(d) 5 lgÆmL)1 endoproteinase Arg-C at
37 �C. Fluorescence readings from endopro-
teinase Arg-C-treated ( ) and untreated (D)
proP7 in 6 M guanidine hydrochloride are
shown. (C) CD (50 lM) spectra of proP7
treated (dashed line) or not treated (solid
line) with Arg-C. (D, E) The leakage of
carboxyfluorescein from liposomes
(phosphatidylcholine ⁄ cardiolipin,
80 : 20 w : w) incubated with the indicated
concentrations of proP7 or Arg-C-treated
proP7 respectively. (F) Lysis of Try. cruzi
trypomastigotes after 30 min of incubation
with the indicated concentrations of proP7
pretreated (d) or not preteated (s) with
Arg-C.
Trialysin precursor activation R. M. Martins et al.
998 FEBS Journal 275 (2008) 994–1002 ª 2008 The Authors Journal compilation ª 2008 FEBS

Arg-C indicates that it is folded, as fluorescence
quenching is not possible for a long, unfolded peptide
[21]. This is confirmed by fluorescence readings
obtained from both peptides in 6 m guanidine hydro-
chloride showing that intramolecular quenching is very
low when proP7 is unstructured. The CD data support
the notion that the proregion stabilizes a folded struc-
ture at the N-terminus of protrialysin, as Arg-C-trea-
ted proP7 and the N-terminus-spanning peptides are
poorly structured in water solution. The results
obtained using phospholipid liposomes also indicate
that unfolding and activation are directly correlated
with an increase in lysis, and that no other molecules
of the parasite are necessary for its activation.
We have previously observed that small variations
in the sequence of the trialysin N-terminus peptides
can modify its specificity for target cells [10]. The
mobility of the N-terminus seems to prevent lysis of
erythrocytes, as substitution of Gly and Pro residues in
this peptide end increases activity for these cells, but
not for trypanosomes. This could explain why the
acidic portion of trialysin, interacting with the basic
amino acids in the amphipathic helix, is less effective
in inhibiting lysis of erythrocytes as compared to try-
panosomes. Perhaps the different negative charges in
these membranes would compete differently for the
basic domain, and the protein or peptides would end
up inserting in the target with variable efficiency.
Other larger forms of trialysin precursors could not
be detected in the salivary glands, indicating that pro-
cessing must occur rapidly after protein synthesis, and
the stored precursor is nontoxic for the gland.
Attempts to express either the full-length predicted
protrialysin, or trialysin itself, in bacteria or in the
yeast Pichia pastoris have been unsuccessful so far.
Our results have indicated that both the predicted pro-
region and the whole N-terminus (the first two pre-
dicted amphipathic a-helices) need to be ablated or
fused to a bigger protein (NusA protein or GST; histi-
dine tags are insufficient) in order to be translated. On
the other hand, the obtained recombinant proteins
(GST–protrialysin or NL fragment) were unstable or
quickly precipitated during purification. These findings
suggest that protrialysin might require a proper envi-
ronment to first fold into an inactive protein, to be
further activated by proteolysis. The possibility cannot
be excluded that another salivary molecule would pre-
vent protrialysin activity, releasing trialysin after saliva
dilution. The lipids present in T. infestans salivary
glands could control trialysin activity. Cytochemical
analyses have shown a high lipid content in the D1 sal-
ivary glands [22], the gland where hemolytic activity
accumulates [23].
Acidic portions are also found in other cytolytic
protein toxins. For example, some pore-forming bacte-
rial toxins are synthesized as precursors with acidic
propieces: proaerolysin is synthesized and secreted by
Aeromonas hydrophila as a dimer that binds the glycan
core of glycosylphosphatidyl inositol-anchored proteins
on the cell surface, and it is processed by host pro-
teases, releasing a small C-terminal peptide, thus
enabling the toxin to oligomerize into the heptameric
channel [24]. The El Tor hemolysin of Vibrio cholerae
is also processed by many host proteases in different
sites at the acidic ⁄apolar propiece [25].
In this work, we have provided evidence that the
acidic portion of a pore-forming protein precursor
controls the lytic activity of the mature molecule. A
synthetic peptide that mimics lysis inhibition and is
suitable for proteolytic activation might be useful in
designing regulated antimicrobial compounds.
Experimental procedures
Insects and cells
T. infestans (males and females) were maintained at room
temperature and fed twice weekly on mice anesthetized with
0.2% (w ⁄ v) ketamine chlorhydrate and 0.12% (w ⁄ v) xyla-
zine chlorhydrate in NaCl ⁄Pi. Trypomastigote forms of the
Y strain of Try. cruzi human erythrocytes were obtained as
previously described [9].
Saliva extraction and salivary glands extracts
Saliva was collected as previously described [9] from both
male and female insects 2 days after feeding. Salivary
glands were obtained by dissecting the insects by pulling
off the rostrum and exposing the thoracic viscera. The
glands were isolated from the esophagus and ducts, and
kept in ice-cold NaCl ⁄Pi. For SDS ⁄PAGE analysis,
glands were readily homogenized in SDS ⁄PAGE loading
buffer containing 1% 2-mercaptoethanol, and boiled for
5 min before electrophoresis. Otherwise, glands were
mechanically disrupted in ice-cold NaCl ⁄Pi containing or
not containing 200 lg of the serine protease inhibitor
APMSF (Roche Diagnostics, Indianapolis, IN, USA)
per mL, and centrifuged for 5 min at 14 000 g. The
collected supernatants were used for activity assays and
SDS ⁄PAGE analysis. The protein concentration was
determined by the Bradford technique, using BSA as
standard [26].
Activity assays
Lysis of trypomastigotes and permeabilization assays of
erythrocytes were performed as previously described [9,10],
R. M. Martins et al. Trialysin precursor activation
FEBS Journal 275 (2008) 994–1002 ª 2008 The Authors Journal compilation ª 2008 FEBS 999

using twofold dilutions of salivary glands extracts, or stock
solutions of the peptide proP7.
Recombinant protein expression and purification
An NL region of preprotrialysin (spanning the C-terminal
region between amino acids Met89 and Asp260, NL2)
was amplified by PCR using primers NDE15P15 89
(5¢-CCATATGAAGAAAGGAGCAGC-3¢) and Bam-LYS30
reverse (5¢-CGGGATCCTTAATCAATTTCAACTTC
ATC-3¢), and the protrialysin cDNA cloned in pGEM-T
Easy (Promega, Madison, WI, USA) as template [9] in
order to insert NdeI and BamHI restriction sites at the
5¢-terminus and 3¢-terminus. The amplified fragment was
inserted in the cloning vector pCR 2.1-TOPO (Invitrogen,
Carlsbad, CA, USA), and the reaction was used to trans-
form chemically competent E. coli DH5a. After sequence
confirmation, the obtained plasmid was digested with
restriction enzymes NdeI and BamHI (Fermentas Interna-
tional, Burlington, Canada), and the insert was purified
from agarose gel and ligated into pET-14b (Novagen,
EMD, Madison, WI, USA) previously digested with the
same restriction enzymes using a Rapid DNA Ligation Kit
(Promega). The ligation reaction was used to transform
E. coli DH5a, and the recovered plasmid (pET14b-NL2)
was used to transform BL21 (DE3) pLysE. The recombi-
nant protein expression was obtained in 300 mL of LB
medium cultures at 37 �C induced at A600 nm @ 0.6 with
0.6 mm isopropyl b-d-thioglucopyranoside (Sigma Chemical
Co., St Louis, MO, USA). Bacteria were collected after
overnight incubation by centrifugation at 3000 g for
10 min. The bacterial cell pellet was resuspended in 20 mm
Tris ⁄HCl (pH 8.0), 6 mm MgCl2, and 0.1% Triton X-100,
and lysis was obtained by three freeze–thawing cycles. The
lysate was centrifuged (15 000 g, 15 min, 4 �C), and the
insoluble pellet was extracted with 8.0 m urea. The insolu-
ble material was removed by centrifugation, and
urea-solubilized NL2 was purified by chromatography in
Ni–nitrilotriacetic acid agarose resin (Qiagen Inc., Chats-
worth, CA, USA) after elution with 100 mm sodium
phosphate, 10 mm Tris ⁄HCl, and 8 m urea (pH 4.3). NL2-
containing fractions were pooled, and the recombinant
protein was further purified by gel filtration in a Super-
dex HR200 column (GE Health Care do Brasil LTDA, Sao
Paulo, Brazil) equilibrated with 20 mm Tris ⁄HCl (pH 8.0),
300 mm NaCl and 8 m urea in an AktaPurifier system
(GE). The purified protein was visualized by SDS ⁄PAGE,
and selected samples were dialyzed twice against 1 L of
NaCl ⁄Pi at 4 �C to remove urea.
Protrialysin fused with GST was obtained from DH5acells transformed with the vector pGEX-2T containing pro-
trialysin cDNA. This construct was obtained after inserting
the restriction sites at the flanking regions of protrialysin
by PCR amplification. The template was the same as above,
and the reaction included oligonucleotides BAMH15P15
(5¢-CGGATCCGCTGAATATGAACTTG-3¢) and ECOR-
13LYS (5¢-CGAATTCTTAATCAATTTCAACTTC-3¢).Cells were grown at 37 �C in LB medium. At D600 nm = 1.5,
the culture was induced with 0.1 mm isopropyl b-d thioglu-
copyranoside, with subsequent growth overnight at 30 �C at
200 r.p.m. Afterwards, the culture was centrifuged at 3000 g
for 10 min, and the cell pellet was subjected to 10 pulses
(20 s each, at maximum power) of sonication in a Branson
Sonifier 450 (Branson Ultrasonics Corporation, Danbury,
CT, USA) in 20 mm Tris ⁄HCl (pH 8.0) and 5 mm EDTA
containing 0.1% Triton X-100 (v ⁄ v). Soluble proteins were
collected after centrifugation at 15 000 g for 20 min, and the
resulting supernatant was incubated with 1 mL of gluta-
thione–Sepharose 4B (GE) previously equilibrated in the
buffer used for cell lysis. The column was washed with
50 mL of lysis buffer, and bound proteins were eluted with
the same buffer containing 20 mm reduced glutathione after
an overnight incubation at 4 �C.
Antiserum production and immunoblotting
A suspension containing 100 lg of NL2 in 300 lL of
NaCl ⁄Pi was emulsified with the same volume of complete
Freund’s adjuvant (Sigma) and subcutaneously injected
throughout the dorsum of a female rabbit. Two consecu-
tive boosts in incomplete Freund’s adjuvant (Sigma) at
3 week intervals were administered, and blood was col-
lected from the ear marginal vein. For immunoblots,
SDS ⁄PAGE gels were wet-transferred to nitrocellulose
membranes (Hybond C-extra; GE), and total blotted
proteins were visualized by Ponceau S staining. The mem-
brane was incubated for 1 h in NaCl ⁄Pi containing 5%
nonfat dry milk and for 1 h with the antiserum diluted
1 : 5000 in the same solution. After three 10 min washes
in NaCl ⁄Pi, bound antibodies were detected after 1 h of
incubation with peroxidase-conjugated goat anti-(rabbit
IgG) (Santa Cruz Biotechnology, Santa Cruz, CA, USA)
and three 10 min NaCl ⁄Pi washes, and detected by
enhanced chemoluminescence (Pierce, Rockford, IL,
USA), using Hyperfilm-ECL (GE) for detection.
Peptide synthesis and purification
The fluorescence resonance energy transfer peptide based
on the presumptive N-terminus of protrialysin found in the
salivary glands including the P7 region of mature trialysin
[10] was produced by solid-phase synthesis [27]. An auto-
mated benchtop simultaneous multiple solid-phase peptide
synthesizer (PSSM 8 system; Shimadzu, Japan) was used to
synthesize peptides, using the Fmoc procedure. All peptides
obtained were purified by semipreparative HPLC on an
Ecosil C-18 column using an Econosil C18 column (10 lm;
22.5 · 250 mm) and the following two-solvent system: sol-
vent A, 0.1% trifluoroacetic acid in water; and solvent B,
Trialysin precursor activation R. M. Martins et al.
1000 FEBS Journal 275 (2008) 994–1002 ª 2008 The Authors Journal compilation ª 2008 FEBS

0.1% trifluoroacetic acid in 90% acetonitrile and 10%
water. The molecular mass and purity of synthesized
peptides were checked by amino acid analysis and MALDI-
TOF MS, using a Tof-Spec-E from Micromass, Manches-
ter, UK. Further purification was performed using a
lRPC C2 ⁄C18 reverse-phase column in the Akta Purifier
system with 0.1% trifluoroacetic acid and a linear gradient
to 100% acetonitrile. Stock solutions of peptides were pre-
pared in dimethylsulfoxide ⁄water (20 : 80), and the peptide
concentrations were determined spectrophotometrically
using a molar extinction coefficient of 17.300 m)1Æcm)1 at
365 nm.
Fluorimetric measurements
Stock solutions of the peptide were diluted in the indicated
buffer solutions at 37 �C incubated with partially purified
triapsin (step 2 of [4]), or with 1 mgÆmL)1 trypsin (type VI,
bovine; Sigma) or 5 lgÆmL)1 Arg-C endoproteinase
(Calbiochem, EMD, San Diego, CA, USA). The proteo-
lytic cleavage of proP7 peptide was monitored by measuring
the fluorescence at kem = 420 nm after excitation at
kexc = 320 nm in a Synergy HT plate-reader spectrofluo-
rimeter (BioTek Instruments, Winooski, VT, USA).
CD
CD experiments were performed using a Jasco J-810 spec-
tropolarimeter (Jasco International Co. Ltd, Tokyo, Japan),
coupled to a peltier Jasco PFD-425S system for tempera-
ture control. ProP7 (50 lm) was digested with Arg-C in
5 mm Tris ⁄HCl (pH 7.4) at 37 �C for 16 h. After treatment,
NaCl was added to 10 mm and CD measurements were
carried out using a 0.1 mm cell in the spectral range 190–
260 nm, at 37 �C. Each spectrum is the average of four
consecutive scans. After baseline correction, the observed
ellipticity, h (mdeg) was converted to mean residue molar
ellipticity (h) (deg cm2Ædmol)1). The a-helix content was
calculated as previously described [28].
Liposome preparation and carboxyfluorescein
leakage assay
Large unilamellar vesicles (LUVs) were prepared from egg
phosphatidylcholine and bovine heart cardiolipin (80 : 20,
weight) dissolved in methanol and dried under an N2 flow.
The lipid film on the tube was hydrated in 10 mm Tris ⁄HCl
and 50 mm carboxyfluorescein, previously purified [29], and
adjusted to pH 8.0. This suspension was extruded through
11 rounds in a LiposoFast (Avestin Inc., Ottawa, Canada)
system containing two polycarbonate membranes (100 nm)
and applied to a Sephadex G-25 medium column equili-
brated in 10 mm Tris ⁄HCl (pH 8.0) and 0.3 m NaCl to
remove free carboxyfluorescein from LUVs. The phospho-
lipid content was determined according to Rouser [30].
LUVs were diluted in 1 mL of 10 mm Tris ⁄HCl (pH 8.0)
and 0.3 m NaCl, and fluorescence was measured in an
Hitachi F-2000 (Japan) spectrofluorimeter (kex = 490 nm
and kem = 512 nm) after addition of peptide solutions. At
the end of each experiment, total carboxyfluorescein fluores-
cence was recorded by the addition of 10% Triton X-100.
Acknowledgements
The authors would like to thank Claudio Rogerio de
Oliveira for assistance with cell cultures, Dr Izaura
Ioshico Hirata for performing amino acid analysis, and
Dr Luis Juliano Neto for helpful suggestions. This
work was supported by grants from Fundacao de
Amparo a Pesquisa do Estado de Sao Paulo (FAPESP)
and Conselho Nacional de Desenvolvimento Cientıfico
e Tecnologico (CNPq) from Brazil.
References
1 Ribeiro JM (1995) Blood-feeding arthropods: live syrin-
ges or invertebrate pharmacologists? Infect Agents Dis
4, 143–152.
2 Chagas C (1909) Nova tripanozomiase humana: estudos
sobre a morfolojia e o ciclo evolutivo do Schizotrypa-
num cruzi n. gen., n. sp., ajente etiolojico de nova entid-
ade morbida do homem. Mem Inst Oswaldo Cruz 1,
159–218.
3 Pereira MH, Souza ME, Vargas AP, Martins MS,
Penido CM & Diotaiuti L (1996) Anticoagulant activity
of Triatoma infestans and Panstrongylus megistus saliva
(Hemiptera ⁄Triatominae). Acta Trop 61, 255–261.
4 Amino R, Tanaka AS & Schenkman S (2001) Triapsin,
an unusual activatable serine protease from the saliva
of the hematophagous vector of Chagas’ disease Tria-
toma infestans (Hemiptera: Reduviidae). Insect Biochem
Mol Biol 31, 465–472.
5 Amino R, Porto RM, Chammas R, Egami MI &
Schenkman S (1998) Identification and characterization
of a sialidase released by the salivary gland of the
hematophagous insect Triatoma infestans. J Biol Chem
273, 24575–24582.
6 Faudry E, Lozzi SP, Santana JM, D’Souza-Ault M,
Kieffer S, Felix CR, Ricart CA, Sousa MV, Vernet T &
Teixeira AR (2004) Triatoma infestans apyrases belong
to the 5¢-nucleotidase family. J Biol Chem 279, 19607–
19613.
7 Morita A, Isawa H, Orito Y, Iwanaga S, Chinzei Y &
Yuda M (2006) Identification and characterization of a
collagen-induced platelet aggregation inhibitor, triplatin,
from salivary glands of the assassin bug, Triatoma infe-
stans. FEBS J 273, 2955–2962.
R. M. Martins et al. Trialysin precursor activation
FEBS Journal 275 (2008) 994–1002 ª 2008 The Authors Journal compilation ª 2008 FEBS 1001

8 Dan A, Pereira MH, Pesquero JL, Diotaiuti L & Beirao
PS (1999) Action of the saliva of Triatoma infestans
(Heteroptera: Reduviidae) on sodium channels. J Med
Entomol 36, 875–879.
9 Amino R, Martins RM, Procopio J, Hirata IY, Juliano
MA & Schenkman S (2002) Trialysin, a novel pore-form-
ing protein from saliva of hematophagous insects acti-
vated by limited proteolysis. J Biol Chem 277, 6207–6213.
10 Martins RM, Sforca ML, Amino R, Juliano MA, Oy-
ama S Jr, Juliano L, Pertinhez TA, Spisni A & Schenk-
man S (2006) Lytic activity and structural differences of
amphipathic peptides derived from trialysin. Biochemis-
try 45, 1765–1774.
11 van Hofsten P, Faye I, Kockum K, Lee JY, Xanthopo-
ulos KG, Boman IA, Boman HG, Engstrom A, Andreu
D & Merrifield RB (1985) Molecular cloning, cDNA
sequencing, and chemical synthesis of cecropin B from
Hyalophora cecropia. Proc Natl Acad Sci USA 82,
2240–2243.
12 Kreil G (1973) Biosynthesis of melittin, a toxic peptide
from bee venom. Amino-acid sequence of the precursor.
Eur J Biochem 33, 558–566.
13 Terry AS, Poulter L, Williams DH, Nutkins JC, Gio-
vannini MG, Moore CH & Gibson BW (1988) The
cDNA sequence coding for prepro-PGS (prepro-magai-
nins) and aspects of the processing of this prepro-poly-
peptide. J Biol Chem 263, 5745–5751.
14 Chen T, Scott C, Tang L, Zhou M & Shaw C (2005)
The structural organization of aurein precursor cDNAs
from the skin secretion of the Australian green and
golden bell frog, Litoria aurea. Regul Pept 128, 75–83.
15 Wilson CL, Ouellette AJ, Satchell DP, Ayabe T, Lopez-
Boado YS, Stratman JL, Hultgren SJ, Matrisian LM &
Parks WC (1999) Regulation of intestinal alpha-defen-
sin activation by the metalloproteinase matrilysin in
innate host defense. Science 286, 113–117.
16 Amiche M, Seon AA, Pierre TN & Nicolas P (1999)
The dermaseptin precursors: a protein family with a
common preproregion and a variable C-terminal anti-
microbial domain. FEBS Lett 456, 352–356.
17 Kozlov SA, Vassilevski AA, Feofanov AV, Surovoy
AY, Karpunin DV & Grishin EV (2006) Latarcins,
antimicrobial and cytolytic peptides from the venom of
the spider Lachesana tarabaevi (Zodariidae) that exem-
plify biomolecular diversity. J Biol Chem 281, 20983–
20992.
18 Boman HG, Wade D, Boman IA, Wahlin B & Merri-
field RB (1989) Antibacterial and antimalarial proper-
ties of peptides that are cecropin–melittin hybrids.
FEBS Lett 259, 103–106.
19 Boman HG (2003) Antibacterial peptides: basic facts
and emerging concepts. J Intern Med 254, 197–215.
20 Sanchez JF, Hoh F, Strub MP, Aumelas A & Dumas C
(2002) Structure of the cathelicidin motif of protegrin-3
precursor: structural insights into the activation mecha-
nism of an antimicrobial protein. Structure 10, 1363–
1370.
21 Pimenta DC, Nantes IL, de Souza ES, Le Bonniec B,
Ito AS, Tersariol IL, Oliveira V, Juliano MA & Juliano
L (2002) Interaction of heparin with internally
quenched fluorogenic peptides derived from heparin-
binding consensus sequences, kallistatin and anti-throm-
bin III. Biochem J 366, 435–446.
22 Reis MM, Meirelles RM & Soares MJ (2003) Fine
structure of the salivary glands of Triatoma infestans
(Hemiptera: Reduviidae). Tissue Cell 35, 393–400.
23 Barth R (1954) Estudos anatomicos e histologicos sobre
a Subfmaılia Triatominae (Heteroptera, Reduviidae).
Mem Inst Oswaldo Cruz 52, 517–590.
24 Fivaz M, Abrami L, Tsitrin Y & van der Goot FG
(2001) Not as simple as just punching a hole. Toxicon
39, 1637–1645.
25 Nagamune K, Yamamoto K & Honda T (1997) Intra-
molecular chaperone activity of the pro-region of Vibrio
cholerae El Tor cytolysin. J Biol Chem 272, 1338–1343.
26 Bradford MM (1976) A rapid and sensitive method for
the quantitation of microgram quantities of protein
utilizing the principle of protein-dye binding. Anal
Biochem 72, 248–254.
27 Hirata IY, Cezari MHS, Nakaie CR, Boschov P,
Ito AS, Juliano MA & Juliano L (1994) Internally
quenched fluorogenic protease substrates: solid-phase
synthesis and fluorescence spectroscopy of peptides con-
taining ortho-aminobenzoyl ⁄dinitrophenyl groups asdonor–acceptor pairs. Lett Peptide Sci 1, 299–308.
28 Chen YH, Yang JT & Chau KH (1974) Determination
of the helix and beta form of proteins in aqueous solu-
tion by circular dichroism. Biochemistry 13, 3350–3359.
29 Weinstein JN, Yoshikami S, Henkart P, Blumenthal R
& Hagins WA (1977) Liposome–cell interaction: trans-
fer and intracellular release of a trapped fluorescent
marker. Science 195, 489–492.
30 Rouser G, Fkeischer S & Yamamoto A (1970) Two
dimensional thin layer chromatographic separation of
polar lipids and determination of phospholipids by
phosphorus analysis of spots. Lipids 5, 494–496.
Trialysin precursor activation R. M. Martins et al.
1002 FEBS Journal 275 (2008) 994–1002 ª 2008 The Authors Journal compilation ª 2008 FEBS
Related Documents











