A Scallop IGF Binding Protein Gene: Molecular Characterization and Association of Variants with Growth Traits Liying Feng, Xue Li, Qian Yu, Xianhui Ning, Jinzhuang Dou, Jiajun Zou, Lingling Zhang, Shi Wang, Xiaoli Hu*, Zhenmin Bao* Key Laboratory of Marine Genetics and Breeding (MGB), Ministry of Education, College of Marine Life Sciences, Ocean University of China, Qingdao, China Abstract Background: Scallops represent economically important aquaculture shellfish. The identification of genes and genetic variants related to scallop growth could benefit high-yielding scallop breeding. The insulin-like growth factor (IGF) system is essential for growth and development, with IGF binding proteins (IGFBPs) serving as the major regulators of IGF actions. Although an effect of IGF on growth was detected in bivalve, IGFBP has not been reported, and members of the IGF system have not been characterized in scallop. Results: We cloned and characterized an IGFBP (PyIGFBP) gene from the aquaculture bivalve species, Yesso scallop (Patinopecten yessoensis, Jay, 1857). Its full-length cDNA sequence was 1,445 bp, with an open reading frame of 378 bp, encoding 125 amino acids, and its genomic sequence was 10,193 bp, consisting of three exons and two introns. The amino acid sequence exhibited the characteristics of IGFBPs, including multiple cysteine residues and relatively conserved motifs in the N-terminal and C-terminal domains. Expression analysis indicated that PyIGFBP was expressed in all the tissues and developmental stages examined, with a significantly higher level in the mantle than in other tissues and a significantly higher level in gastrulae and trochophore larvae than in other stages. Furthermore, three single nucleotide polymorphisms (SNPs) were identified in this gene. SNP c.1054A.G was significantly associated with both shell and soft body traits in two populations, with the highest trait values in GG type scallops and lowest in AG type ones. Conclusion: We cloned and characterized an IGFBP gene in a bivalve, and this report also represents the first characterizing an IGF system gene in scallops. A SNP associated with scallop growth for both the shell and soft body was identified in this gene. In addition to providing a candidate marker for scallop breeding, our results also suggest the role of PyIGFBP in scallop growth. Citation: Feng L, Li X, Yu Q, Ning X, Dou J, et al. (2014) A Scallop IGF Binding Protein Gene: Molecular Characterization and Association of Variants with Growth Traits. PLoS ONE 9(2): e89039. doi:10.1371/journal.pone.0089039 Editor: Zhanjiang Liu, Auburn University, United States of America Received November 10, 2013; Accepted January 13, 2014; Published February 19, 2014 Copyright: ß 2014 Feng et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. Funding: This work was supported by National High-Tech R&D Program (863 Program, 2012AA10A402), National Natural Science Foundation of China (31272656), National Basic Research Program of China (973 Program, 2010CB126406), and Natural Science Foundation for Distinguished Young Scholars of Shandong Province (JQ201308). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. Competing Interests: The authors confirm that the co-author Shi Wang is a PLOS ONE Editorial Board member. This does not alter the authors’ adherence to PLOS ONE Editorial policies and criteria. The authors have also declared that no competing interests exist. * E-mail: [email protected] (XH); [email protected] (ZB) Introduction Scallops represent an economically important aquaculture species in Asian countries and are consumed worldwide. Among the varieties, the Yesso scallop (Patinopecten yessoensis, Jay, 1857) is the main scallop species cultured in Japan and has become one of the most important maricultural shellfish in northern China since it was introduced in 1982 [1]. Similar to other aquaculture species, genetic breeding aiming at improving the growth rate is one of the main focuses of the Yesso scallop farming industry. The identification of genes with a possible function in growth regulation and of genetic markers associated with growth could provide useful information for the genetic improvement of this species. The insulin-like growth factor (IGF) system which is the important component of the growth hormone axis, plays a pivotal role in cell growth and differentiation [2,3]. The system mainly includes two IGF ligands (IGF-I and IGF-II, members of a family of insulin related peptides), two IGF receptors (IGF-IR and IGF- IIR), and a family of IGF binding proteins (IGFBPs) [2]. The IGF ligands circulate in the plasma in complexes with IGFBPs with affinities that are equal to or higher than those of IGF-IR, which transports, stores and modulates the bioavailability of IGFs [3]. Therefore, IGFBPs are the major regulators of IGF activity [3]. In addition to modulating IGF bioactivity, the importance of IGFBPs for cell growth has also been indicated in IGF-independent mechanisms [4]. Although most of these studies were implemented in vertebrate species, the IGF system has also been increasingly studied in invertebrates, and its members have been characterized in bivalve species. For example, the insulin-related peptide was identified in Mytilus edulis [5], Crassostrea gigas [6] and Anodonta cygnea [7], and the PLOS ONE | www.plosone.org 1 February 2014 | Volume 9 | Issue 2 | e89039

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A Scallop IGF Binding Protein Gene: MolecularCharacterization and Association of Variants withGrowth TraitsLiying Feng, Xue Li, Qian Yu, Xianhui Ning, Jinzhuang Dou, Jiajun Zou, Lingling Zhang, Shi Wang,
Xiaoli Hu*, Zhenmin Bao*
Key Laboratory of Marine Genetics and Breeding (MGB), Ministry of Education, College of Marine Life Sciences, Ocean University of China, Qingdao, China
Abstract
Background: Scallops represent economically important aquaculture shellfish. The identification of genes and geneticvariants related to scallop growth could benefit high-yielding scallop breeding. The insulin-like growth factor (IGF) system isessential for growth and development, with IGF binding proteins (IGFBPs) serving as the major regulators of IGF actions.Although an effect of IGF on growth was detected in bivalve, IGFBP has not been reported, and members of the IGF systemhave not been characterized in scallop.
Results: We cloned and characterized an IGFBP (PyIGFBP) gene from the aquaculture bivalve species, Yesso scallop(Patinopecten yessoensis, Jay, 1857). Its full-length cDNA sequence was 1,445 bp, with an open reading frame of 378 bp,encoding 125 amino acids, and its genomic sequence was 10,193 bp, consisting of three exons and two introns. The aminoacid sequence exhibited the characteristics of IGFBPs, including multiple cysteine residues and relatively conserved motifs inthe N-terminal and C-terminal domains. Expression analysis indicated that PyIGFBP was expressed in all the tissues anddevelopmental stages examined, with a significantly higher level in the mantle than in other tissues and a significantlyhigher level in gastrulae and trochophore larvae than in other stages. Furthermore, three single nucleotide polymorphisms(SNPs) were identified in this gene. SNP c.1054A.G was significantly associated with both shell and soft body traits in twopopulations, with the highest trait values in GG type scallops and lowest in AG type ones.
Conclusion: We cloned and characterized an IGFBP gene in a bivalve, and this report also represents the first characterizingan IGF system gene in scallops. A SNP associated with scallop growth for both the shell and soft body was identified in thisgene. In addition to providing a candidate marker for scallop breeding, our results also suggest the role of PyIGFBP in scallopgrowth.
Citation: Feng L, Li X, Yu Q, Ning X, Dou J, et al. (2014) A Scallop IGF Binding Protein Gene: Molecular Characterization and Association of Variants with GrowthTraits. PLoS ONE 9(2): e89039. doi:10.1371/journal.pone.0089039
Editor: Zhanjiang Liu, Auburn University, United States of America
Received November 10, 2013; Accepted January 13, 2014; Published February 19, 2014
Copyright: � 2014 Feng et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permitsunrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: This work was supported by National High-Tech R&D Program (863 Program, 2012AA10A402), National Natural Science Foundation of China(31272656), National Basic Research Program of China (973 Program, 2010CB126406), and Natural Science Foundation for Distinguished Young Scholars ofShandong Province (JQ201308). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing Interests: The authors confirm that the co-author Shi Wang is a PLOS ONE Editorial Board member. This does not alter the authors’ adherence toPLOS ONE Editorial policies and criteria. The authors have also declared that no competing interests exist.
* E-mail: [email protected] (XH); [email protected] (ZB)
Introduction
Scallops represent an economically important aquaculture
species in Asian countries and are consumed worldwide. Among
the varieties, the Yesso scallop (Patinopecten yessoensis, Jay, 1857) is
the main scallop species cultured in Japan and has become one of
the most important maricultural shellfish in northern China since
it was introduced in 1982 [1]. Similar to other aquaculture species,
genetic breeding aiming at improving the growth rate is one of the
main focuses of the Yesso scallop farming industry. The
identification of genes with a possible function in growth
regulation and of genetic markers associated with growth could
provide useful information for the genetic improvement of this
species.
The insulin-like growth factor (IGF) system which is the
important component of the growth hormone axis, plays a pivotal
role in cell growth and differentiation [2,3]. The system mainly
includes two IGF ligands (IGF-I and IGF-II, members of a family
of insulin related peptides), two IGF receptors (IGF-IR and IGF-
IIR), and a family of IGF binding proteins (IGFBPs) [2]. The IGF
ligands circulate in the plasma in complexes with IGFBPs with
affinities that are equal to or higher than those of IGF-IR, which
transports, stores and modulates the bioavailability of IGFs [3].
Therefore, IGFBPs are the major regulators of IGF activity [3]. In
addition to modulating IGF bioactivity, the importance of IGFBPs
for cell growth has also been indicated in IGF-independent
mechanisms [4].
Although most of these studies were implemented in vertebrate
species, the IGF system has also been increasingly studied in
invertebrates, and its members have been characterized in bivalve
species. For example, the insulin-related peptide was identified in
Mytilus edulis [5], Crassostrea gigas [6] and Anodonta cygnea [7], and the
PLOS ONE | www.plosone.org 1 February 2014 | Volume 9 | Issue 2 | e89039

insulin receptor-related receptor that showed an association with
insulin-like effects on growth was found in C. gigas [8]. Single
nucleotide polymorphisms (SNPs) associated with growth were
recently identified in the insulin-related peptide gene of C. gigas [9].
Meanwhile, growth-regulating effects of mammalian IGF on C.
gigas [8] and Pecten maximus [10] were also observed. These findings
suggest the possible role of the IGF system in the growth regulation
of bivalves. However, IGF system members have not been
characterized in scallops, and the important regulator of the
IGF system, IGFBP, has not been reported in any bivalve species.
In this study, using the transcriptome data of Yesso scallop
[11,12], a cDNA fragment encoding an IGFBP homolog was
found. Then, we cloned the full-length cDNA sequence and
obtained the genomic DNA sequence of the Yesso scallop IGFBP
gene (PyIGFBP). Its expression levels in adult tissues and
developmental stages were characterized. We also identified three
SNPs in the transcribed sequence of this gene and found that one
of them was significantly associated with the growth of both the
shell and soft body of the Yesso scallop. Our data suggest the
possible function of PyIGFBP in scallop growth regulation and
also provide a candidate locus for the selective breeding of Yesso
scallop.
Materials and Methods
Sample CollectionTwo Yesso scallop populations were collected from Zhangzidao
Fishery Group Co., Dalian, the leading producer of Yesso scallop
in China. Each population was established by the artificial
fertilization of more than 2,000 sexually mature scallops. A total
of 60 individuals (13 months old) were randomly sampled in
Population I, and 120 individuals (10 months old) were randomly
collected from Population II. For all scallops, the shell length (SL),
shell height (SH), body weight (BW), soft tissue weight (STW) and
adductor muscle weight (AMW) were measured. Tissues including
the mantle, gill, gonad, kidney, striated muscle and hepatopan-
creas were dissected, immediately frozen in liquid nitrogen and
stored at 280uC. Embryos and larvae, including newly fertilized
eggs, blastulae, gastrulae, trochophore larvae and D-shaped
larvae, were also collected and preserved at 280uC.
RNA Isolation and cDNA SynthesisTotal RNA was isolated from the tissues and embryos/larvae of
Yesso scallop using TRIzol reagent (Life Technologies, CA, USA)
following the manufacturer’s instructions. The first-strand cDNA
was synthesized according to the manufacturer’s instructions using
M-MLV Reverse Transcriptase (Promega, WI, USA) in a 25-mLvolume with 2 mg of total RNA as the template and 0.8 mM Oligo
(dT)18 (TaKaRa Biotechnology, Liaoning, China) as the primer.
The mixture of total RNA and primer was heated at 95uC for
5 min and then chilled on ice. The first strand cDNA was then
synthesized at 42uC for 90 min after adding the reverse
transcriptase and its buffer. The cDNA was stored at 220uCand diluted to 1:40 for use as the template in PCR. To preclude
any DNA contamination, a control reaction without reverse
transcriptase was performed.
Full-length cDNA Cloning of PyIGFBPUsing the transcriptome data of the Yesso scallop [11,12], a
cDNA fragment of 1,277 bp that encoded an IGFBP homolog was
obtained. Then, the 39 and 59 rapid amplification of cDNA ends
(RACE) were performed to obtain the full-length cDNA from total
RNA of mantle tissue from one individual. The reactions were
completed according to the instructions of the SMARTTM RACE
cDNA Amplification Kit (Clontech, CA, USA). Specific primers
Igfbp5-f1 and Igfbp5-r1 (Table 1) were designed and used for the
39 and 59 RACE, respectively. The RACE products were purified
and ligated into the pMD18-T vector (TaKaRa Biotechnology),
and six recombinant plasmids from each RACE reaction were
sequenced by Sangon Biotech (Shanghai, China). The full-length
cDNA sequence was obtained by assembling the sequences of 39
and 59 RACE products with the 1,277 bp gene fragment.
Sequence AnalysisThe assembled cDNA sequence was evaluated by TBLASTX
against the National Center for Biotechnology Information
database (NCBI, http://www.ncbi.nlm.nih.gov/blast/) for anno-
tation. The genomic sequence of this gene was obtained by
searching the genomic data (unpublished) of the Yesso scallop
using the full-length cDNA sequence. Then, the genomic and
cDNA sequences were compared to obtain the genomic structure
of this gene. The deduced amino acid sequence was analyzed
using the simple modular architecture research tool (SMART)
(http://smart.embl-heidelberg.de/) to predict the conserved do-
mains. The SignalP 4.0 server (http://www.cbs.dtu.dk/services/
SignalP/) was used to determine the presence and location of the
signal peptide. The ClustalW2 multiple alignment program
(http://www.ebi.ac.uk/Tools/msa/clustalw2/) and the GeneDoc
multiple alignment editor (http://www.nrbsc.org/gfx/genedoc/
index.html) were used to achieve the multiple alignment of
PyIGFBP and IGFBP5s from other species.
Expression Analysis of PyIGFBP GeneThe expression levels of PyIGFBP in the adult tissues and
developmental stages of the Yesso scallop were analyzed using
real-time quantitative reverse transcription PCR (qRT-PCR). The
first-stand cDNA from adult tissues (mantle, gill, gonad, kidney,
striated muscle and hepatopancreas) of 12 Yesso scallops and from
embryos and larvae (fertilized egg, blastula, gastrula, trochophore
larva and D-shaped larva, n .500, three sets of samples for each
stage) was used as the template. For each PCR, three technical
repeats were performed. Specific primers Igfbp5-f5 and Igfbp5-r5
(Table 1), which corresponded to the sequences located in exon 2
and exon 3, respectively, were designed for the amplification of the
PyIGFBP cDNA fragment. Genes encoding DEAD-box RNA
helicase-like protein (HELI), ubiquitin (UBQ) and 60S ribosomal
protein L16 (RPL16) were selected as reference genes for the tissue
samples, and those encoding Cytochrome B (CB), Cytochrome C
(CC) and Histone H3.3 (His3.3) were used as reference genes for
the embryo/larva samples [13].
The reaction mix included 16 Real-time PCR Master Mix
containing SYBR Green dye (TOYOBO, Osaka, Japan), 0.4 mMeach primer and 2 mL of the cDNA template. The reaction was
performed as follows: initial denaturation at 95uC for 10 min,
followed by 40 cycles of 95uC for 15 s and 62.8uC for 1 min. At
the end of the PCR, a dissociation (from 95uC to 60uC) analysiswas performed to confirm that only one product was amplified.
The PCR products for PyIGFBP and the reference genes were
purified and sequenced by Sangon Biotech to verify the
specificity of the qRT-PCR products. All the reactions were
performed on a LightCycler 480 system (Roche Applied Science,
Penzberg, Germany), and the results were analyzed with Real-
time PCR Miner (http://www.miner.ewindup.info/) [14]. The
geometric means of the values generated with the three reference
genes were calculated for both tissue and embryo/larva samples
for normalization [15].
Scallop IGFBP Gene and SNP Associated with Growth
PLOS ONE | www.plosone.org 2 February 2014 | Volume 9 | Issue 2 | e89039
SNP Scanning and GenotypingGenomic DNA was extracted from the striated muscle of 180
scallops in the two populations, using the traditional phenol/
chloroform extraction method [16]. For SNP screening, the top
5 and bottom 5 scallops in Population I were selected based on
the SL values. Three primer pairs, Igfbp5-f2 and Igfbp5-r2,
Igfbp5-f3 and Igfbp5-r3 and Igfbp5-f4 and Igfbp5-r4 (Table 1)
were used to amplify the DNA fragments that spanned the
transcribed sequence of PyIGFBP. The 10-mL reaction mix
contained 16 Advantege2 PCR buffer, 0.2 mM each dNTP
(Life Technologies), 0.2 mM each primer, 50 ng of genomic
DNA and 0.2 mL of 506 Advantage2 Polymerase Mix
(Clontech). The cycling protocol was the following: 95uC for
5 min; 30 cycles of 95uC for 30 s, 63uC for 30 s and 72uC for
2 min; and a final extension at 72uC for 5 min. Products of the
target length were purified by gel extraction and sequenced by
Sangon Biotech. Sequences amplified from the same primer
pairs were compared among the 10 scallops using the
ClustalW2 program. Finally, three candidate SNPs (c.-117T.
C, c.879C.T and c.1054A.G) were identified, with one in the
59 UTR and two in the 39 UTR (Figure 1).
The three SNPs were then genotyped in the 10 scallops used in
the SNP screening by the high-resolution melting (HRM) method
for locus verification and marker development [17]. For each
SNP, two primers and one probe were designed for the HRM
genotyping (Table 1). The 10-mL reaction mix contained 16PCR
buffer, 1.5 mM MgCl2, 0.5 U of Taq DNA polymerase (Ta-
KaRa), 0.2 mM each dNTP (Life Technologies), 0.1 mM forward
primer, 0.5 mM reverse primer, 16LCGreen Plus (Idaho Tech-
nology, UT, USA) and 20 ng of genomic DNA. The PCR reaction
was performed as follows: 95uC for 5 min; 60 cycles of 95uC for
40 s, 63uC for 40 s and 72uC for 40 s; and a final extension at
72uC for 5 min. Then, 3 ml of the probe (10 mM) was added to
each reaction mix, and the mixture was denatured at 95uC for
10 min and slowly cooled to 40uC. HRM genotyping was
immediately performed using a Light-Scanner instrument (Idaho
Technology) with continuous melting curve acquisition (10
acquisitions per uC) over a 0.1uC/s ramp from 40 to 95uC. The
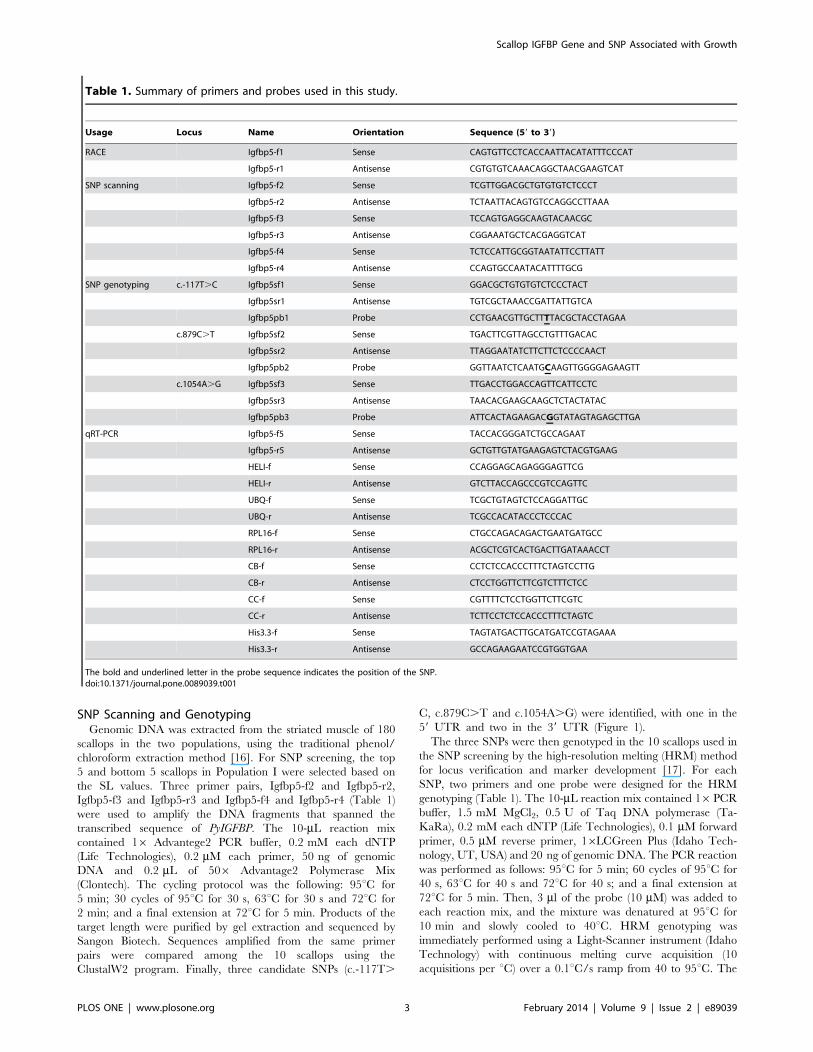
Table 1. Summary of primers and probes used in this study.
Usage Locus Name Orientation Sequence (59 to 39)
RACE Igfbp5-f1 Sense CAGTGTTCCTCACCAATTACATATTTCCCAT
Igfbp5-r1 Antisense CGTGTGTCAAACAGGCTAACGAAGTCAT
SNP scanning Igfbp5-f2 Sense TCGTTGGACGCTGTGTGTCTCCCT
Igfbp5-r2 Antisense TCTAATTACAGTGTCCAGGCCTTAAA
Igfbp5-f3 Sense TCCAGTGAGGCAAGTACAACGC
Igfbp5-r3 Antisense CGGAAATGCTCACGAGGTCAT
Igfbp5-f4 Sense TCTCCATTGCGGTAATATTCCTTATT
Igfbp5-r4 Antisense CCAGTGCCAATACATTTTGCG
SNP genotyping c.-117T.C Igfbp5sf1 Sense GGACGCTGTGTGTCTCCCTACT
Igfbp5sr1 Antisense TGTCGCTAAACCGATTATTGTCA
Igfbp5pb1 Probe CCTGAACGTTGCTTTTACGCTACCTAGAA
c.879C.T Igfbp5sf2 Sense TGACTTCGTTAGCCTGTTTGACAC
Igfbp5sr2 Antisense TTAGGAATATCTTCTTCTCCCCAACT
Igfbp5pb2 Probe GGTTAATCTCAATGCAAGTTGGGGAGAAGTT
c.1054A.G Igfbp5sf3 Sense TTGACCTGGACCAGTTCATTCCTC
Igfbp5sr3 Antisense TAACACGAAGCAAGCTCTACTATAC
Igfbp5pb3 Probe ATTCACTAGAAGACGGTATAGTAGAGCTTGA
qRT-PCR Igfbp5-f5 Sense TACCACGGGATCTGCCAGAAT
Igfbp5-r5 Antisense GCTGTTGTATGAAGAGTCTACGTGAAG
HELI-f Sense CCAGGAGCAGAGGGAGTTCG
HELI-r Antisense GTCTTACCAGCCCGTCCAGTTC
UBQ-f Sense TCGCTGTAGTCTCCAGGATTGC
UBQ-r Antisense TCGCCACATACCCTCCCAC
RPL16-f Sense CTGCCAGACAGACTGAATGATGCC
RPL16-r Antisense ACGCTCGTCACTGACTTGATAAACCT
CB-f Sense CCTCTCCACCCTTTCTAGTCCTTG
CB-r Antisense CTCCTGGTTCTTCGTCTTTCTCC
CC-f Sense CGTTTTCTCCTGGTTCTTCGTC
CC-r Antisense TCTTCCTCTCCACCCTTTCTAGTC
His3.3-f Sense TAGTATGACTTGCATGATCCGTAGAAA
His3.3-r Antisense GCCAGAAGAATCCGTGGTGAA
The bold and underlined letter in the probe sequence indicates the position of the SNP.doi:10.1371/journal.pone.0089039.t001
Scallop IGFBP Gene and SNP Associated with Growth
PLOS ONE | www.plosone.org 3 February 2014 | Volume 9 | Issue 2 | e89039

data were retrieved and analyzed using the Light Scanner software
(Idaho Technology), followed by manually curating the genotype
results. All three SNPs were successfully genotyped and further
genotyped in other individuals of population I. The SNP that
showed a significant association with growth traits in Population I
was further genotyped in Population II.
Statistical AnalysisThe scallops from the two populations were grouped according
to their genotypes. The chi-squared test was used to examine
Hardy-Weinberg equilibrium (HWE) in the populations. For each
growth trait, the mean value and standard deviation were
calculated for each genotype group. A comparison of the means
for each trait among the different genotypes was performed using
one way ANOVA with a post-hoc test. The comparison of the
expression levels of PyIGFBP among adult tissues and among
developmental stages was performed using one way ANOVA with
a post-hoc test. P values of less than 0.05 were considered
statistically significant.
Results and Discussion
Characterization of PyIGFBP Gene SequenceUsing the transcriptome data of the Yesso scallop [11,12], a
1,277-bp cDNA fragment of the PyIGFBP gene was obtained.
After 59 and 39 RACE, the full-length sequence of PyIGFBP
(1,468 bp) was obtained. The gene contained an open reading
frame (ORF) of 378 bp (encoding 125 amino acids), a 59 UTR of
206 bp and a 39 UTR of 884 bp. A putative polyadenylation
signal (AATAAA) was identified at nucleotide positions 1,211 to
1,216. A comparison of the cDNA and genomic DNA sequence of
this gene showed that PyIGFBP consisted of three exons (405, 97
and 944 bp) and two introns (6465 and 2282 bp), whereas all the
identified IGFBPs except IGFBP3 (five exons) in vertebrate species
are encoded by four exons [2]. As the genomic information of
mollusca IGFBPs is currently unavailable in the public database
(NCBI), whether the three-exon structure is PyIGFBP-specific or
common in mollusca species is unknown. All the intron-exon
boundaries of PyIGFBP conformed to the GT-AG rule [18]. The
structure of the PyIGFBP gene is shown in Figure 1. The genomic
and cDNA sequences of PyIGFBP have been deposited in the
GenBank database under accession numbers KF801669 and
KF801670, respectively.
TBLASTX analysis showed that the deduced amino acid
sequence was most similar to IGFBPs, especially IGFBP5, from
other species. To analyze the sequence characteristics of
PyIGFBP, the amino acid sequences for PyIGFBP and IGFBP5s
in other species were aligned (Figure 2). IGFBPs have been
reported to contain conserved cysteine-rich N-terminal and C-
terminal domains and a highly variable central linker domain [2].
The alignment showed that there was a long sequence deletion in
PyIGFBP in the central region. However, the N-terminal and C-
terminal regions were relatively more conserved, which is
consistent with the characteristics of IGFBPs. IGFBPs have also
been characterized by a conserved GCGCCXXC motif in the N-
terminal domain and a CWCV motif in the C-terminal domain,
although the significance of these motifs is not yet known [2,19].
The PyIGFBP exhibited two amino acid differences in the
corresponding regions of the GCGCxxC motif (-CRCCXXC)
and the CWCV motif (CQNV) (Figure 2). Meanwhile, the number
of cysteine residues in the N-terminal and C-terminal regions of
vertebrate IGFBP5s is 12 and 6, respectively, and 11 and 1,
respectively, in PyIGFBP, mainly due to the deletion of amino
acids in the PyIGFBP midregion compared with those from
vertebrates. The 20 amino acids in the N-terminal region of
PyIGFBP were predicted to be a cleavable signal peptide
(Figure 2).
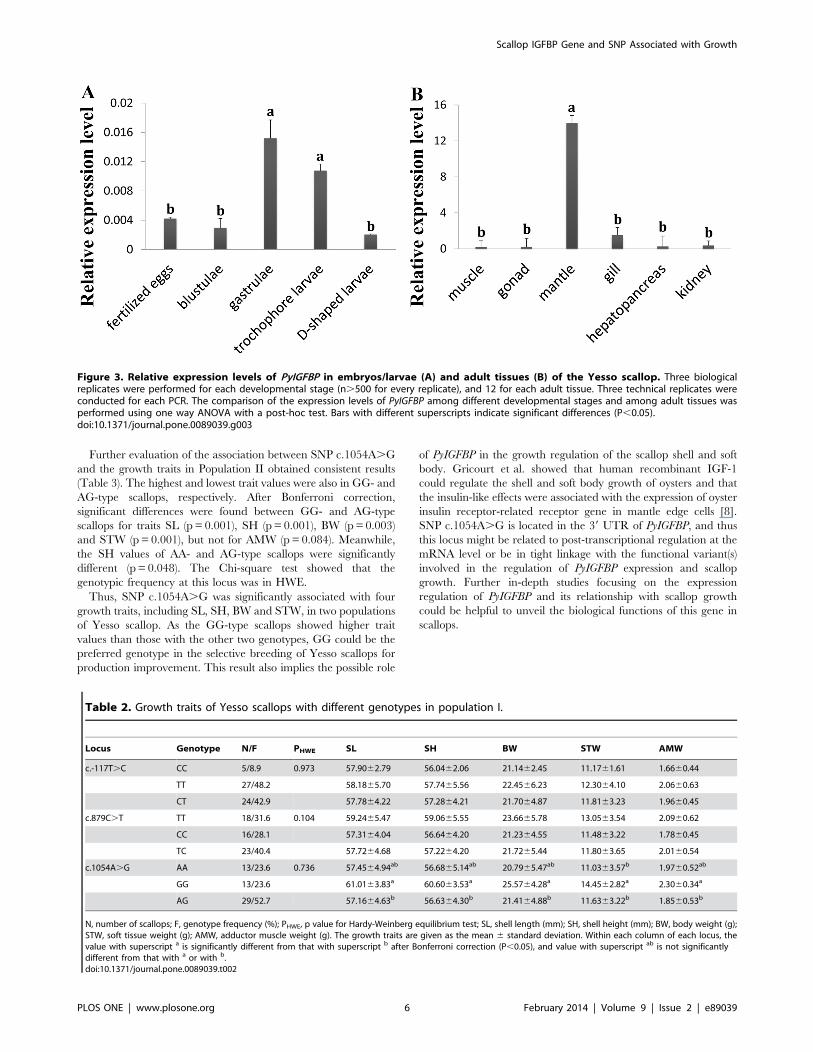
Spatiotemporal Expression of PyIGFBPThe expression of PyIGFBP in the embryos/larvae and adult
tissues of Yesso scallop was analyzed using qRT-PCR. The
amplification efficiencies for PyIGFBP and the reference genes
were all greater than 0.96 based on the analysis by Real-time PCR
Miner. PyIGFBP expression was detected in all the developmental
stages sampled, including fertilized egg, blastula, gastrula, trocho-
phore larva and D-shaped larva (Figure 3A). PyIGFBP mRNA
Figure 1. The structure of the PyIGFBP gene and protein. The gene contains three exons. The 59 and 39 UTR (light blue) and exons (blue) areshown relative to their lengths. The location of the three SNPs (c.-117T.C, c.879C.T and c.1054A.G) is indicated with a star. The position of theGCGCCXXC and CWCV motifs is indicated with an arrow, respectively.doi:10.1371/journal.pone.0089039.g001
Scallop IGFBP Gene and SNP Associated with Growth
PLOS ONE | www.plosone.org 4 February 2014 | Volume 9 | Issue 2 | e89039

detected in newly fertilized eggs implies that the transcripts might
be maternally derived and involved in the early development of
scallop embryos. At the gastrula stage, the expression level of
PyIGFBP increased sharply with a significant difference from that
in the fertilized eggs and blastulae. Then, the expression was
slightly downregulated in trochophore larvae. In D-shaped larvae,
the transcription level of PyIGFBP was the lowest among the stages
tested. The significantly higher level of PyIGFBP mRNA detected
in gastrulae and trochophore larvae suggests that the gene might
participate in the major organo-morphogenetic events.
PyIGFBP expression was also detected in all the adult tissues
analyzed (Figure 3B), with a significantly higher level of PyIGFBP
expression in the mantle, approximately 47, 68, 8, 55 and 30-fold
higher than that in striated muscle, gonad, gill, hepatopancreas
and kidney, respectively. No significant difference in the PyIGFBP
expression level was detected among the other five tissues. The
mantle is the main tissue in charge of the formation and growth of
the shell in bivalves [20,21]. The calcite and aragonite in the shell
are transformed from amorphous calcium carbonate through the
regulation by proteins secreted from epithelial cells in the outer
mantle tissues [20–23]. Many genes expressed in the mantle are
responsible for the biomineralization in this process [24–27]. The
effect of IGF system members on mantle growth and shell
formation has been demonstrated in oysters [8]. In mammals,
bone is one of the major target tissues of IGF-1 [28–30]. IGFBP5
was considered to be the major IGFBP in bone, thereby playing an
important role in the modulation of biomineralization [19,31].
PyIGFBP is more similar to IGFBP5 than to other IGFBP
members, as revealed by BLAST analysis. The much higher
expression level of PyIGFBP in mantle tissue implies its possible
role in scallop shell growth.
Association between PyIGFBP SNPs and Growth TraitsWe further investigated the sequence variants in PyIGFBP. A
total of three SNPs, named c.-117T.C, c.879C.T and
c.1054A.G, were found in 10 individuals of Population I
(Figure 1). SNP c.-117T.C was in the 59 UTR, and the latter
two were in the 39 UTR. These SNPs were then genotyped in the
other scallops of Population I using HRM assays. The genotypes of
all the three SNPs were in HWE (Table 2). Potential associations
between the genotypes of each locus and each of the five growth
traits (SL, SH, BW, STW and AMW) were analyzed via one-way
ANOVA with a post-hoc test. Significant associations were
detected only for one of the three SNPs, c.1054A.G (Table 2).
For all of the growth traits measured, the highest and lowest trait
values were in scallops with the GG and AG genotypes,
respectively, and AA type individuals showed intermediate values.
After Bonferroni correction, significant differences in the trait
values were detected between GG- and AG-type scallops
(p = 0.048 for SL, p = 0.029 for SH, p= 0.048 for BW, p= 0.045
for STW and p=0.029 for AMW) and between AA- and GG-type
ones for STW (p= 0.033).
Figure 2. Alignment of the amino acid sequences of PyIGFBP with IGFBP5s from other species. Identical and similar residues are shaded.The putative SP (signal peptide) is indicated with horizontal arrows above the alignment. The characteristic motif of IGFBPs, GCGCCXXC and CWCV, isunderlined and marked with a black box, respectively. The 12 conserved cysteine residues are denoted with stars. The black arrowheads mark theintron-exon boundaries in the PyIGFBP gene. GenBank accession numbers for IGFBP5s are the following: D. rerio (NP_001119935.1); S. salar(ABO36535.1); X. tropicalis (NP_001016042.1); M. musculus (AAH54812.1); H. sapiens (CAG33090.1).doi:10.1371/journal.pone.0089039.g002
Scallop IGFBP Gene and SNP Associated with Growth
PLOS ONE | www.plosone.org 5 February 2014 | Volume 9 | Issue 2 | e89039
Further evaluation of the association between SNP c.1054A.G
and the growth traits in Population II obtained consistent results
(Table 3). The highest and lowest trait values were also in GG- and
AG-type scallops, respectively. After Bonferroni correction,
significant differences were found between GG- and AG-type
scallops for traits SL (p = 0.001), SH (p= 0.001), BW (p= 0.003)
and STW (p= 0.001), but not for AMW (p= 0.084). Meanwhile,
the SH values of AA- and AG-type scallops were significantly
different (p = 0.048). The Chi-square test showed that the
genotypic frequency at this locus was in HWE.
Thus, SNP c.1054A.G was significantly associated with four
growth traits, including SL, SH, BW and STW, in two populations
of Yesso scallop. As the GG-type scallops showed higher trait
values than those with the other two genotypes, GG could be the
preferred genotype in the selective breeding of Yesso scallops for
production improvement. This result also implies the possible role
of PyIGFBP in the growth regulation of the scallop shell and soft
body. Gricourt et al. showed that human recombinant IGF-1
could regulate the shell and soft body growth of oysters and that
the insulin-like effects were associated with the expression of oyster
insulin receptor-related receptor gene in mantle edge cells [8].
SNP c.1054A.G is located in the 39 UTR of PyIGFBP, and thus
this locus might be related to post-transcriptional regulation at the
mRNA level or be in tight linkage with the functional variant(s)
involved in the regulation of PyIGFBP expression and scallop
growth. Further in-depth studies focusing on the expression
regulation of PyIGFBP and its relationship with scallop growth
could be helpful to unveil the biological functions of this gene in
scallops.
Figure 3. Relative expression levels of PyIGFBP in embryos/larvae (A) and adult tissues (B) of the Yesso scallop. Three biologicalreplicates were performed for each developmental stage (n.500 for every replicate), and 12 for each adult tissue. Three technical replicates wereconducted for each PCR. The comparison of the expression levels of PyIGFBP among different developmental stages and among adult tissues wasperformed using one way ANOVA with a post-hoc test. Bars with different superscripts indicate significant differences (P,0.05).doi:10.1371/journal.pone.0089039.g003
Table 2. Growth traits of Yesso scallops with different genotypes in population I.
Locus Genotype N/F PHWE SL SH BW STW AMW
c.-117T.C CC 5/8.9 0.973 57.9062.79 56.0462.06 21.1462.45 11.1761.61 1.6660.44
TT 27/48.2 58.1865.70 57.7465.56 22.4566.23 12.3064.10 2.0660.63
CT 24/42.9 57.7864.22 57.2864.21 21.7064.87 11.8163.23 1.9660.45
c.879C.T TT 18/31.6 0.104 59.2465.47 59.0665.55 23.6665.78 13.0563.54 2.0960.62
CC 16/28.1 57.3164.04 56.6464.20 21.2364.55 11.4863.22 1.7860.45
TC 23/40.4 57.7264.68 57.2264.20 21.7265.44 11.8063.65 2.0160.54
c.1054A.G AA 13/23.6 0.736 57.4564.94ab 56.6865.14ab 20.7965.47ab 11.0363.57b 1.9760.52ab
GG 13/23.6 61.0163.83a 60.6063.53a 25.5764.28a 14.4562.82a 2.3060.34a
AG 29/52.7 57.1664.63b 56.6364.30b 21.4164.88b 11.6363.22b 1.8560.53b
N, number of scallops; F, genotype frequency (%); PHWE, p value for Hardy-Weinberg equilibrium test; SL, shell length (mm); SH, shell height (mm); BW, body weight (g);STW, soft tissue weight (g); AMW, adductor muscle weight (g). The growth traits are given as the mean 6 standard deviation. Within each column of each locus, thevalue with superscript a is significantly different from that with superscript b after Bonferroni correction (P,0.05), and value with superscript ab is not significantlydifferent from that with a or with b.doi:10.1371/journal.pone.0089039.t002
Scallop IGFBP Gene and SNP Associated with Growth
PLOS ONE | www.plosone.org 6 February 2014 | Volume 9 | Issue 2 | e89039
Acknowledgments
We thank the Zhangzidao Fishery Group (Dalian, China) for assisting in
collection of the scallop samples.
Author Contributions
Conceived and designed the experiments: ZB XH. Performed the
experiments: LF XL QY XN JZ. Analyzed the data: LF JD LZ SW.
Contributed reagents/materials/analysis tools: ZB. Wrote the paper: XH
LF.
References
1. Wang Q (1984) Introduction of Japanese scallop and prospect of culture it innorthern China. Fisheries Science 3: 24–27.
2. Hwa V, Oh Y, Rosenfeld RG (1999) The insulin-like growth factor-bindingprotein (IGFBP) superfamily. Endocr Rev 20: 761–787.
3. Jones JI, Clemmons DR (1995) Insulin-like growth factors and their binding
proteins: biological actions. Endocr Rev 16: 3–34.4. Oh Y, Yamanaka Y, Wilson E, Kim H-S, Vorwerk P, et al. (1998) IGF-
independent actions of IGFBPs. In: Takano K, Hizuka N, Takahashi S-I (eds)Molecular Mechanisms to Regulate the Activities of Insulin-Like Growth
Factors. Elsevier, Amsterdam, 123–133.5. Kellner-Cousin K, Mialhe E, Mathieu M (1994) Identification of insulin-like
peptides in cerebral ganglia neurosecretory cells of the mussel Mytilus edulis.
Tissue Cell 26: 891–899.6. Hamano K, Awaji M (2002) cDNA cloning and expression analysis of insulin-
related peptide gene and prohormone convertase gene in the Pacific oyster,Crassostrea gigas. Fisheries Science 68: 769–721.
7. Shipilov V, Shpakov A, Rusakov YI (2005) Pleiotropic action of insulin-like
peptides of mollusk, Anodonta cygnea. Ann N Y Acad Sci 1040: 464–465.8. Gricourt L, Bonnec G, Boujard D, Mathieu M, Kellner K (2003) Insulin-like
system and growth regulation in the Pacific oyster Crassostrea gigas: hrIGF-1 effecton protein synthesis of mantle edge cells and expression of an homologous
insulin receptor-related receptor. Gen Comp Endocrinol 134: 44–56.9. Cong R, Li Q, Kong L (2013) Polymorphism in the insulin-related peptide gene
and its association with growth traits in the Pacific oyster Crassostrea gigas.
Biochem Syst Ecol 46: 36–43.10. Giard W, Lebel JM, Boucaud-Camou E, Favrel P (1998) Effects of vertebrate
growth factors on digestive gland cells from the mollusc Pecten maximus L.: anin vitro study. J Comp Physiol B 168: 81–86.
11. Hou R, Bao Z, Wang S, Su H, Li Y, et al. (2011) Transcriptome sequencing and
de novo analysis for Yesso scallop (Patinopecten yessoensis) using 454 GS FLX. PloSOne 6: e21560.
12. Wang S, Hou R, Bao Z, Du H, He Y, et al. (2013) Transcriptome sequencing ofZhikong scallop (Chlamys farreri) and comparative transcriptomic analysis with
Yesso scallop (Patinopecten yessoensis). PLoS One 8: e63927.13. Feng L, Yu Q, Li X, Ning X, Wang J, et al. (2013) Identification of reference
genes for qRT-PCR analysis in Yesso Scallop Patinopecten yessoensis. PloS One 8:
e75609.14. Zhao S, Fernald RD (2005) Comprehensive algorithm for quantitative real-time
polymerase chain reaction. J comput biol 12: 1047–1064.15. Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, et al. (2002)
Accurate normalization of real-time quantitative RT-PCR data by geometric
averaging of multiple internal control genes. Genome Biol 3: research0034.
16. Sambrook J (2001) Molecular Cloning: A Laboratory Mannual, Third Edition.
New York: Cold Spring Harbor Laboratory Press.
17. Guo H, Bao Z, Li J, Lian S, Wang S, et al. (2012) Molecular characterization of
TGF-b type I receptor gene (Tgfbr1) in Chlamys farreri, and the association of
allelic variants with growth traits. PloS One 7: e51005.
18. Sharp PA (1981) Speculations on RNA splicing. Cell 23: 643–646.
19. Rosenzweig SA (2004) What’s new in the IGF-binding proteins? Growth Horm
IGF Res 14: 329–336.
20. Marin F, Luquet G, Marie B, Medakovic D (2008) Molluscan shell proteins:
primary structure, origin, and evolution. Cur Top Dev Biol 80: 209–276.
21. Kinoshita S, Wang N, Inoue H, Maeyama K, Okamoto K, et al. (2011) Deep
sequencing of ESTs from nacreous and prismatic layer producing tissues and a
screen for novel shell formation-related genes in the pearl oyster. PLoS One 6:
e21238.
22. Addadi L, Raz S, Weiner S (2003) Taking advantage of disorder: amorphous
calcium carbonate and its roles in biomineralization. Adv Mater 15: 959–970.
23. Sarashina I, Endo K (2006) Skeletal matrix proteins of invertebrate animals:
Comparative analysis of their amino acid sequences. Paleontol Res 10: 311–336.
24. Fang D, Xu G, Hu Y, Pan C, Xie L (2011) Identification of genes directly
involved in shell formation and their functions in pearl oyster, Pinctada fucata.
PLoS One 6: e21860.
25. Shi Y, Yu C, Gu Z, Zhan X, Wang Y, et al. (2013) Characterization of the pearl
oyster (Pinctada martensii) mantle transcriptome unravels biomineralization genes.
Mar Biotechnol 15: 175–187.
26. ClarkMS ThorneMAS, Vieira FA, Cardoso JCR, PowerDM, etal. (2010)
Insights into shell deposition in the Antarctic bivalve Laternula elliptica: gene
discovery in the mantle transcriptome using 454 pyrosequencing. BMC
Genomics 11: 362.
27. Joubert C, Piquemal D, Marie B, Manchon L, Pierrat F, et al. (2010)
Transcriptome and proteome analysis of Pinctada margaritifera calcifying mantle
and shell: focus on biomineralization. BMC Genomics 11: 613.
28. McCarthy TL, Centrella M, Canalis E (1989) Insulin-like growth factor (IGF)
and bone. Connect Tissue Res 20: 277–282.
29. Mohan S, Baylink DJ (1991) Bone growth factors. Clin Orthop Relat Res 263:
30–48.
30. Niemann I, Hannemann A, Nauck M, Spielhagen C, Volzke H, et al. (2013)
The association between insulin-like growth factor I and bone turnover markers
in the general adult population. Bone 56: 184–190.
31. Luther GA, Lamplot J, Chen X, Rames R, Wagner ER, et al. (2013) IGFBP5
domains exert distinct inhibitory effects on the tumorigenicity and metastasis of
human osteosarcoma. Cancer Lett 336: 222–230.
Table 3. Growth traits of Yesso scallops with different genotypes at SNP c.1054A.G in population II.
Genotype N/F PHWE SL SH BW STW AMW
AA 30/25.6 0.890 38.7965.74ab 40.3765.86a 7.4563.21ab 4.2562.10ab 0.8060.78
GG 29/24.8 42.1266.29a 42.3866.49a 8.5863.44a 4.9561.89a 0.8060.34
AG 58/49.6 35.9165.96b 37.0465.77b 5.9063.07b 3.2961.77 b 0.5560.31
N, number of scallops; F, genotype frequency (%); PHWE, p value for Hardy-Weinberg equilibrium test; SL, shell length (mm); SH, shell height (mm); BW, body weight (g);STW, soft tissue weight (g); AMW, adductor muscle weight (g). The growth traits are given as the mean 6 standard deviation. Within each column, the value withsuperscript a is significantly different from that with superscript b after Bonferroni correction (P,0.05), and value with superscript ab is not significantly different fromthat with a or with b.doi:10.1371/journal.pone.0089039.t003
Scallop IGFBP Gene and SNP Associated with Growth
PLOS ONE | www.plosone.org 7 February 2014 | Volume 9 | Issue 2 | e89039
Related Documents











