Herpetologica, 62(1), 2006, 71–89 Ó 2006 by The Herpetologists’ League, Inc. A REVISION OF SUB-SAHARAN CHALCIDES (SQUAMATA: SCINCIDAE), WITH REDESCRIPTIONS OF TWO EAST AFRICAN SPECIES ELI GREENBAUM 1,2,3 ,ANDREW C. CAMPBELL 1 , AND CHRISTOPHER J. RAXWORTHY 4 1 Division of Herpetology, Natural History Museum & Biodiversity Research Center, and Department of Ecology and Evolutionary Biology, 1345 Jayhawk Boulevard, The University of Kansas, Lawrence, KS 66045-7561, USA 4 Department of Herpetology, American Museum of Natural History, Central Park West at 79 th Street, New York, NY 10024-5192, USA ABSTRACT: There has been little recent work on the sub-Saharan species of the African and Eurasian genus Chalcides, possibly because the morphology of the genus seems to be highly conserved and specimens are generally rare. As a result, there is a lack of consensus concerning the taxonomic status and distributions of these species in the literature. Herein, we expand upon the brief original descriptions of C. bottegi and C. ragazzii, which we consider to be valid and allopatrically distributed species (separated by the Ethiopian highlands). Examination of populations previously assigned to C. ragazzii in the south-central Saharan highlands and the West African Niger River floodplain indicates that these populations are most similar to populations of C. ocellatus, and we assign them to this widespread, variable species. A disjunct population of C. ocellatus in Somalia is separated from its nearest conspecifics by the Ethiopian highlands and the Indian Ocean; however, we did not find morphological evidence to recognize it as a distinct species. All three species of Chalcides are readily distinguishable from the West African endemics C. armitagei, C. pulchellus, and C. thierryi, which are the only other sub-Saharan representatives for the genus. An identification key is provided for all six sub-Saharan Chalcides species. ABSTRAIT: Peu de travaille re ´cent a e ´te ´ effectue ´ sur des taxa subsahariens du genre Chalcides, possiblement a ` cause de la morphologie tre ´ s conserve ´e du genre et la rarete ´ generale des specimens de plusieurs des re ´ gions. En e ´ ffet, avis divers de la position taxonomique et de la distribution des taxa sont pre ´ sent a ` la litte ´rature scientifique. Nous e ´ largons les descriptions originaux bre ` ves de C. bottegi et C. ragazzii; nous les conside ´rons comme e ´ speces valides et distribue ´ es allopatriques (elles sont separe ´es par les re ´ gions montagneuses d’Ethiopie). Apre `s nos examens des populations des re ´ gions montagneuses de Sahara sud- central et du lit majeur du Niger en Afrique d’ouest attribue ´ auparavant de C. ragazzii, nous les avons transfe ´re ´es a ` C. ocellatus, une e ´spece re ´ pandue et variable. Une population disjointe de C. ocellatus en Somalie est separe ´e des populations de la me ˆ me e ´spece voisines par des re ´ gions montagneuses d’Ethiopie et l’ocean Indien, mais nous n’avons pas trouve ´ d’evidence morphologique pour la reconnaı ˆtre comme e ´ spece se ´ pare ´e. Ces trois e ´ speces Chalcides se distinguent facilement des e ´ speces ende ´ mique d’Afrique d’ouest: C. armitagei, C. pulchellus, et C. thierryi, qui sont les seules autres repre ´ sentants subsahariens. Un table d’identification est fourni de tous les six e ´speces de Chalcides subsahariennes. Key words: Africa; Chalcides; Distribution; Reptilia; Sahara; Scincidae; Taxonomy THE COMPLICATED taxonomic history of the African and Eurasian scincid Chalcides ocellatus began with the work of George A. Boulenger (G. Boulenger, 1887, 1890, 1891a,b, 1896, 1898), who recognized six varieties of this widespread species and de- scribed C. bottegi from Ethiopia. Edward Boulenger, George’s youngest son and cura- tor of reptiles at the Zoological Society of London (Adler, 1989), expanded upon his father’s work by arguing against the recogni- tion of C. bottegi, and summarized the morphology and color patterns of eight forms of C. ocellatus (E. Boulenger, 1920). Sub- sequently, North African and insular species of Chalcides received considerable taxonomic attention (e.g., Brown et al., 2000; Caputo, 1993; Caputo and Mellado, 1992; Caputo et al., 1995, 1999; Geniez et al., 2004; Klausewitz, 1954; Pasteur, 1981; Schleich et al., 1996), but relatively little has been published about the sub-Saharan species (e.g., Caputo et al., 1995; Greenbaum, 2005; Greenbaum and Rasmus- sen, 2004; Pasteur, 1981). Aside from the distinctive West African endemics C. armita- gei, C. pulchellus, and C. thierryi, sub-Saharan specimens of other Chalcides species have 2 PRESENT ADDRESS: Department of Biology, Villanova University, 800 Lancaster Avenue, Villanova, Pennsylvania 19085-1699 USA. 3 CORRESPONDENCE: e-mail, [email protected] 71

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Herpetologica, 62(1), 2006, 71–89� 2006 by The Herpetologists’ League, Inc.
A REVISION OF SUB-SAHARAN CHALCIDES(SQUAMATA: SCINCIDAE), WITH REDESCRIPTIONS
OF TWO EAST AFRICAN SPECIES
ELI GREENBAUM1,2,3, ANDREW C. CAMPBELL
1, AND CHRISTOPHER J. RAXWORTHY4
1Division of Herpetology, Natural History Museum & Biodiversity Research Center, andDepartment of Ecology and Evolutionary Biology, 1345 Jayhawk Boulevard,
The University of Kansas, Lawrence, KS 66045-7561, USA4Department of Herpetology, American Museum of Natural History, Central Park West at 79th Street,
New York, NY 10024-5192, USA
ABSTRACT: There has been little recent work on the sub-Saharan species of the African and Eurasian genusChalcides, possibly because the morphology of the genus seems to be highly conserved and specimens aregenerally rare. As a result, there is a lack of consensus concerning the taxonomic status and distributions ofthese species in the literature. Herein, we expand upon the brief original descriptions of C. bottegi and C.ragazzii, which we consider to be valid and allopatrically distributed species (separated by the Ethiopianhighlands). Examination of populations previously assigned to C. ragazzii in the south-central Saharanhighlands and the West African Niger River floodplain indicates that these populations are most similar topopulations of C. ocellatus, and we assign them to this widespread, variable species. A disjunct population ofC. ocellatus in Somalia is separated from its nearest conspecifics by the Ethiopian highlands and the IndianOcean; however, we did not find morphological evidence to recognize it as a distinct species. All three speciesof Chalcides are readily distinguishable from the West African endemics C. armitagei, C. pulchellus, and C.thierryi, which are the only other sub-Saharan representatives for the genus. An identification key is providedfor all six sub-Saharan Chalcides species.
ABSTRAIT: Peu de travaille recent a ete effectue sur des taxa subsahariens du genre Chalcides,possiblement a cause de la morphologie tres conservee du genre et la rarete generale des specimens deplusieurs des regions. En effet, avis divers de la position taxonomique et de la distribution des taxa sont presenta la litterature scientifique. Nous elargons les descriptions originaux breves de C. bottegi et C. ragazzii; nousles considerons comme especes valides et distribuees allopatriques (elles sont separees par les regionsmontagneuses d’Ethiopie). Apres nos examens des populations des regions montagneuses de Sahara sud-central et du lit majeur du Niger en Afrique d’ouest attribue auparavant de C. ragazzii, nous les avonstransferees a C. ocellatus, une espece repandue et variable. Une population disjointe de C. ocellatus enSomalie est separee des populations de la meme espece voisines par des regions montagneuses d’Ethiopie etl’ocean Indien, mais nous n’avons pas trouve d’evidence morphologique pour la reconnaıtre comme especeseparee. Ces trois especes Chalcides se distinguent facilement des especes endemique d’Afrique d’ouest:C. armitagei, C. pulchellus, et C. thierryi, qui sont les seules autres representants subsahariens. Un tabled’identification est fourni de tous les six especes de Chalcides subsahariennes.
Key words: Africa; Chalcides; Distribution; Reptilia; Sahara; Scincidae; Taxonomy
THE COMPLICATED taxonomic history of theAfrican and Eurasian scincid Chalcidesocellatus began with the work of GeorgeA. Boulenger (G. Boulenger, 1887, 1890,1891a,b, 1896, 1898), who recognized sixvarieties of this widespread species and de-scribed C. bottegi from Ethiopia. EdwardBoulenger, George’s youngest son and cura-tor of reptiles at the Zoological Society ofLondon (Adler, 1989), expanded upon hisfather’s work by arguing against the recogni-
tion of C. bottegi, and summarized themorphology and color patterns of eight formsof C. ocellatus (E. Boulenger, 1920). Sub-sequently, North African and insular speciesof Chalcides received considerable taxonomicattention (e.g., Brown et al., 2000; Caputo,1993; Caputo and Mellado, 1992; Caputo et al.,1995, 1999; Geniez et al., 2004; Klausewitz,1954; Pasteur, 1981; Schleich et al., 1996), butrelatively little has been published about thesub-Saharan species (e.g., Caputo et al., 1995;Greenbaum, 2005; Greenbaum and Rasmus-sen, 2004; Pasteur, 1981). Aside from thedistinctive West African endemics C. armita-gei, C. pulchellus, and C. thierryi, sub-Saharanspecimens of other Chalcides species have
2 PRESENT ADDRESS: Department of Biology, VillanovaUniversity, 800 Lancaster Avenue, Villanova, Pennsylvania19085-1699 USA.
3 CORRESPONDENCE: e-mail, [email protected]
71

been treated as C. bottegi (G. Boulenger,1898); C. ocellatus (Le Berre, 1989; Pasteur,1981; Spawls et al., 2002; Welch, 1982), C.ocellatus bottegi (Drewes, 1972; Parker, 1932);C. ocellatus var. humilis (G. Boulenger, 1896);C. ocellatus ocellatus (Laurent and Gans, 1965;Loveridge, 1936); C. ocellatus tassiliensis(Angel, 1936; Angel and Lhote, 1938), C.ocellatus sacchii (Lanza, 1954; Lanza andCarfı, 1968); C. ragazzii (Pasteur, 1981), C. r.bottegi, and C. r. ragazzii (Caputo et al., 1995;Pasteur, 1981; Welch, 1982). Museums withholdings of sub-Saharan Chalcides have speci-mens catalogued under various combinationsof these taxa, thereby exacerbating the confu-sion. Moreover, many of these species (andother semifossorial scincids) are poorly repre-sented in museum collections because ofa dearth of fieldwork in the sub-Saharan Sahelof Africa, and the rapid escape behavior ofthese lizards (Greenbaum, 2005; Pasteur,1981; Spawls et al., 2002).
The most recent taxonomic analysis ofChalcides was performed by Pasteur (1981),who constructed a key for the genus andidentified five species groups, including thewidespread C. ocellatus group. Based on theinformation within his key, Pasteur (1981)noted African C. ocellatus have 26–34 scalerows at midbody, and are distributed fromMorocco to Zanzibar Island in East Africa. Inthe same key, Pasteur (1981) noted C. ragazziihas 20–26 scale rows at midbody, and isdistributed in northern Somalia and northernKenya to the Sahara in southern Algeria.Although Angel (1936) described C. ocellatustassiliensis from the highlands of southeasternAlgeria, Pasteur (1981) did not discuss hisreasons for considering this population to beC. ragazzii. In his discussion of the ‘‘ragazziicomplex,’’ Pasteur (1981:11) noted that C. r.ragazzii is limited to an undefined, relativelysmall part of the species range near the typelocality (Assab, southeastern Eritrea), whereasC. r. bottegi is distributed from the highlandsof Ethiopia to northern Kenya and northernSomalia. Pasteur (1981) noted a third, un-named subspecies from Sudan and Chad to Aır(eastern Hoggar Mountains), Niger, despiteciting Angel (1936) who named C. ocellatustassiliensis from this region. In a study ofmorphological variation of the genus Chal-cides, Caputo et al. (1995) seemed to follow
the taxonomy of Pasteur (1981) by treatingspecimens from Djibouti as C. r. ragazzii, andEthiopian specimens as C. r. bottegi.
In addition to the localities provided byPasteur (1981), a specimen from sub-SaharanWest Africa was collected by the senior andultimate authors from the Niger River flood-plain in Mali in 2001, and two additionalspecimens from Timbuktu, Mali, were discov-ered in the ZMUC collection. Herein, weexamine the taxonomic status of populations ofChalcides in East Africa, the Saharan and sub-Saharan highlands, and Niger River floodplain,and compare them to C. ocellatus from Sudan,North Africa, and Saudi Arabia.
MATERIALS AND METHODS
Operational taxonomic units for this studyincluded two putative East African species(C. bottegi and C. ragazzii), and allopatric andparapatric populations of C. ocellatus fromcoastal North Africa, Saudi Arabia, Sudan,Somalia, and West African Sahel and Saharawith previously noted differences in colorpattern, morphology, and scale counts. Mea-surements were made with dial calipers to thenearest 0.1 mm and include snout–vent length(SVL), head length (HL, measured from snoutto ear opening), head depth (HD, measured atcenter of frontal scale), maximum head width(HW, determined as maximum distance be-tween lateral margins of the head between thecenter of the frontal scale to jaw rictus), axilla–groin distance (AG, measured from posteriorinsertion of forelimb to anterior insertion ofhind limb), snout–forelimb length (SF, mea-sured from snout to anterior insertion offorelimb), forelimb length (FLL, measuredfrom proximal insertion of forelimb to end oflongest digit [excluding the claw]), hind limblength (HLL, measured from proximal in-sertion of hind limb to end of longest digit[excluding the claw]), frontal scale length(FSL, measured from anterior to posteriormargins), internasal width (IW, measuredfrom left to right margins), interparietal length(IL, measured from anterior to posteriormargins), ear-opening height (EOH, mea-sured from inferior to superior margin), ear-opening width (EOW, measured from left toright margin), eye height (EH, measuredacross center of eye from superior margin ofsupralabial to inferior margin of supraciliary),
72 HERPETOLOGICA [Vol. 62, No. 1

eye length (EL, measured along margin ofupper eyelid from anterior margin of post-ocular to posterior margin of preocular), lowereyelid window height (LEWH, measured atcenter of window from superior to inferiormargin), lower eyelid window length (LEWL,measured at center of window from left toright margin), internarial distance (IND,measured between medial margins of narialopenings), interocular distance (IOD, mea-sured between center of eyes where theycontact supraciliaries), eye–naris distance(END, measured from anterior border of eyeto posterior margin of narial opening), eye–eardistance (EED, measured from posteriorborder of eye to anterior margin of earopening), tail length (TL, measured from ventto tip of tail), tail width at base (TW, measuredfrom left to right margin of tail at vent), taildepth at base (TD, measured from superiorto inferior margin of tail at vent), length offorelimb Digits II–IV (measured from proxi-mal attachment to distal margin [excluding theclaws]), and length of hind limb Digits III–V(measured from proximal attachment to distalmargin [excluding the claws]). Meristic datawere recorded using the scale nomenclature ofKlausewitz (1954:193), except for the frenalthat we consider as the anterior loreal, andfrenoculars considered here as the 1st supra-ciliary and posterior loreal. Ventral scales werecounted from the mental (excluded) to theprecloacal scale (included). The concept of‘‘expanded vertebral’’ scales has not beenadequately defined in previous work onChalcides. Previous workers have tacitly sug-gested that expanded vertebrals are muchwider than long (e.g., Spawls et al., 2002),but all dorsal scales of the examined specimensare wider than long to varying degrees. Wedefine expanded vertebrals as those that arewider than paravertebrals, and provide ratiosto compare the width of vertebrals and para-vertebrals. Incisions to determine gender andmaturity were not permitted for most loanedspecimens; sexual dimorphism for all theseChalcides species is not evident based onexternal morphology. All specimens wereradiographed to determine the number ofpresacral vertebrae. Museum codes are fromLeviton et al. (1985) except for MakasutuWildlife Trust, Serrekunda, Gambia (MWTR);specimens examined are listed in Appendix I.
Because specimen samples from differentpopulations also included juveniles and sub-adults, we used shape and statistics describingmeristic characters that are not ontogeneticallyvariable. Exploratory covariance analyses re-gressed HL, HW, AG, SF, FLL, HLL, IND,IOD, END, EED, and lengths of Toes andFingers III and IV on SVL in 202 specimens(juveniles to adults) of C. ocellatus and sub-Saharan Chalcides of uncertain taxonomicstatus. Specimens with damaged or missingstructures were excluded from analyses. Taillength was not included in this analysis,because more than 80% of these specimenshad incomplete or regenerated tails. Signifi-cant interaction terms for species 3 SVLindicated significant differences in the slopesof regressions. Analyses of variance (ANOVA)and t-tests were used to test for differences inmeristic data among putative species. Toaccount for multiple tests and reduce theprobability of Type-I errors, the significancelevel was adjusted (P � 0.01).
RESULTS
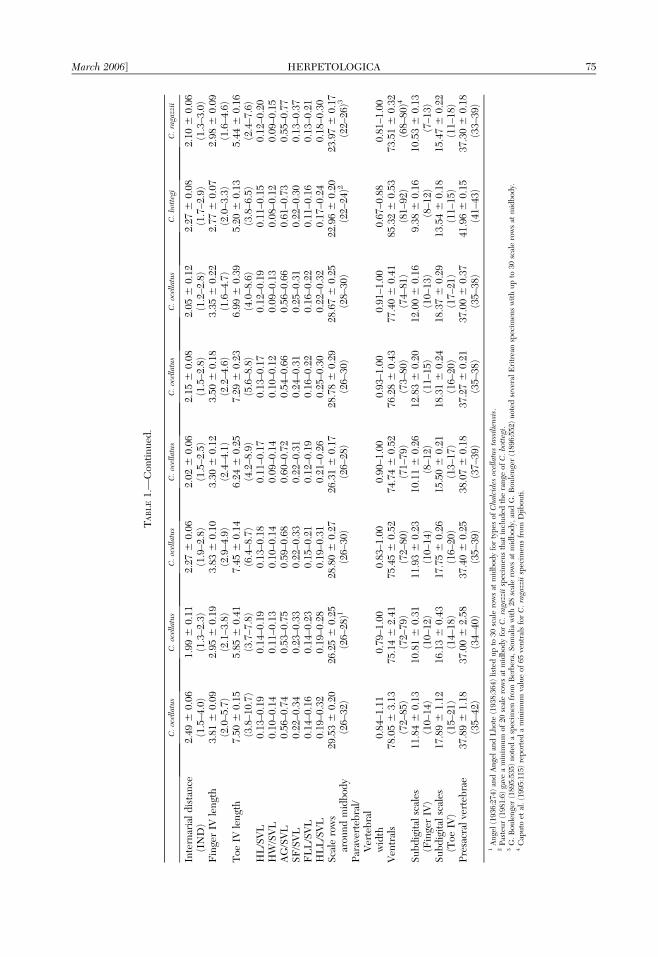
Morphological data, color pattern, distribu-tion, and proposed taxonomy of African popu-lations of Chalcides ocellatus, C. bottegi, andC. ragazzii are shown in Table 1. Populationsof C. ocellatus from the Saharan and sub-Saharan highlands and Sudan have signifi-cantly fewer scale rows at midbody (F5,107 525.46, P , 0.001), subdigital lamellae underFinger III (F5,106 5 13.66, P , 0.001), FingerIV (F5,107 5 11.42, P , 0.001), Toe III (F5,105 511.47, P , 0.001) and Toe IV (F5,106 5 19.65,P , 0.001) than conspecifics in coastal NorthAfrica, Nile River floodplain, Saudi Arabia,and Somalia. Populations of C. ocellatus fromthe North African coast and Somalia havesignificantly more ventrals than other popula-tions (F5,105 5 14.24, P , 0.001). Covarianceanalyses indicated significant slope differencesfor HL (P 5 0.002) between C. ocellatuspopulations from the North African coast andSudan, and for SF (P 5 0.007) betweenpopulations from Sudan and Somalia. Colorpattern (brownish vertebral stripe present orabsent), varies among and within populationsof C. ocellatus (Fig. 1). Although most of theabove differences involve the population fromSudan, we did not note any distinct, non-
March 2006] HERPETOLOGICA 73

TA
BL
E1.
—S
elec
ted
mea
sure
men
ts(i
nm
m),
mor
ph
olog
ical
rati
os,
mer
isti
cd
ata,
colo
rp
atte
rn,
hab
itat
s,an
dge
ogra
ph
icd
istr
ibu
tion
ofC
halc
ides
bott
egi,
C.
ocel
latu
s,an
dC
.rag
azzi
i.N
um
eric
dat
aar
em
ean
s(m
easu
rem
ents
inm
m)
6S
E,
follo
wed
by
ran
gein
par
enth
eses
.R
atio
sd
on
otin
clu
de
aver
age
orS
E.
C.o
cella
tus
C.o
cella
tus
C.o
cella
tus
C.
ocel
latu
sC
.oc
ella
tus
C.o
cella
tus
C.
bott
egi
C.
raga
zzii
N44
820
2018
1525
63
Dis
trib
uti
onN
orth
Afr
ican
coas
t(w
est
ofN
ileR
iver
)
Sah
aran
and
Sah
elia
nh
igh
lan
ds;
Nig
erR
iver
floo
dp
lain
Nile
Riv
er,
Egy
pt
Nile
Riv
er,
Su
dan
Sau
di
Ara
bia
Som
alia
Eas
tern
Su
dan
ton
orth
ern
Ken
ya
Eri
trea
ton
orth
wes
tern
Som
alia
Maj
orh
abit
atN
orth
Sah
aran
woo
dla
nd
s;M
edit
erra
nea
nfo
rest
s
Mon
tan
exe
ric
woo
dla
nd
;ri
ver
floo
dp
lain
Riv
erfl
ood
pla
inth
rou
ghS
ahar
aR
iver
floo
dp
lain
thro
ugh
Sah
ara
and
Sah
el
Sem
i-ar
idb
ush
lan
dB
ush
lan
dan
dth
icke
tS
avan
na
and
mon
tan
efo
rest
s
Xer
icgr
assl
and
and
mon
tan
efo
rest
Com
mon
colo
rp
atte
rnB
row
nve
rteb
ral
stri
pe,
dor
so-l
ater
alcr
eam
stri
pe,
late
ral
irre
gula
rd
ark
bro
wn
stri
pe,
nu
mer
ous
dar
kb
row
nsp
ots
wit
hw
hit
eoc
elli
Bro
wn
vert
ebra
lst
rip
e,d
orso
-lat
eral
pal
eb
row
nst
rip
e,la
tera
lir
regu
lar
dar
kb
row
nst
rip
e,n
um
erou
sd
ark
bro
wn
spot
sw
ith
wh
ite
ocel
li
Gro
un
dco
lor
bro
wn
orta
n;
nu
mer
ous
dar
kb
row
nsp
ots
wit
hw
hit
eoc
elli
Fai
nt
bro
wn
vert
ebra
lst
rip
e,gr
oun
dco
lor
bro
wn
orta
n;
nu
mer
ous
dar
kb
row
nsp
ots
wit
hw
hit
eoc
elli
Gro
un
dco
lor
bro
wn
orgr
ay;
nu
mer
ous
dar
kb
row
nsp
ots
wit
hw
hit
eoc
elli
Gro
un
dco
lor
crea
m;
nu
mer
ous
dar
kb
row
nsp
ots
wit
hw
hit
eoc
elli
Pal
eb
row
nve
rteb
ral
stri
pe,
dor
so-l
ater
alcr
eam
stri
pe,
late
ral
irre
gula
rd
ark
bro
wn
stri
pe,
nu
mer
ous
dar
kb
row
nsp
ots
wit
hw
hit
eoc
elli
Bro
wn
vert
ebra
lst
rip
e,d
orso
-lat
eral
crea
mst
rip
e,la
tera
lir
regu
lar
dar
kb
row
nst
rip
e,n
um
erou
sd
ark
bro
wn
spot
sw
ith
wh
ite
ocel
li
Lon
gitu
din
ald
orsa
lst
rip
esP
rese
nt
Pre
sen
tA
bse
nt
Pre
sen
tb
ut
fain
tA
bse
nt
Ab
sen
tP
rese
nt
Pre
sen
t
Sn
out–
ven
tle
ngt
h(S
VL
)
93.8
06
2.19
(54.
1–14
8.7)
79.1
66
6.37
(41.
0–97
.1)
88.1
16
2.21
(76.
6–10
7.5)
87.2
06
3.51
(61.
5–12
3.9)
83.9
46
4.67
(53.
0–12
5.6)
79.5
66
6.20
(36.
3–11
5.8)
97.0
26
3.74
(59.
8–13
7.2)
82.7
76
3.06
(36.
9–12
7.4)
Hea
dle
ngt
h(H
L)
13.9
76
0.32
(8.3
–24.
9)11
.99
60.
79(7
.8–1
4.0)
12.9
66
0.29
(11.
3–15
.9)
12.0
06
0.34
(9.2
–15.
2)12
.65
60.
55(8
.4–1
7.1)
11.7
56
0.66
(7.0
–14.
8)12
.54
60.
36(9
.1–1
5.6)
12.1
56
0.34
(7.4
–16.
8)M
axim
um
hea
dw
idth
(HW
)
10.7
66
0.29
(5.7
–19.
1)9.
296
0.78
(5.3
–11.
4)9.
816
0.32
(7.9
–13.
0)9.
776
0.33
(7.1
–12.
7)9.
086
0.58
(5.2
–13.
9)8.
396
0.56
(4.7
–11.
6)9.
486
0.31
(5.7
–12.
0)9.
056
0.31
(4.8
–14.
2)
Axi
lla–g
roin
len
gth
(AG
)
59.4
86
1.60
(33.
7–10
2.8)
48.3
86
4.46
(24.
0–64
.5)
55.9
46
1.45
(45.
2–68
.0)
57.1
66
2.41
(39.
2–82
.2)
51.9
96
3.30
(29.
9–77
.9)
48.8
96
4.16
(20.
6–76
.9)
64.6
76
2.67
(39.
3–94
.6)
53.0
56
2.14
(20.
1–87
.6)
Sn
out–
fore
limb
len
gth
(SF
)25
.47
60.
55(1
5.6–
40.6
)22
.39
61.
96(1
3.4–
27.3
)23
.11
60.
59(2
0.2–
29.1
)22
.88
60.
63(1
6.9–
26.9
)23
.05
61.
14(1
5.8–
32.9
)21
.17
61.
41(1
1.1–
28.8
)23
.72
60.
76(1
7.8–
32.9
)22
.33
60.
71(1
1.4–
33.6
)F
orel
imb
len
gth
(FL
L)
16.2
16
0.36
(9.4
–26.
9)13
.70
60.
85(9
.3–1
6.2)
15.4
56
0.41
(12.
6–18
.8)
13.7
26
0.48
(9.4
–18.
2)15
.77
60.
74(9
.7–2
1.5)
14.5
96
0.97
(6.9
–18.
4)13
.48
60.
36(9
.6–1
6.3)
13.3
46
0.42
(5.6
–20.
9)H
ind
limb
len
gth
(HL
L)
23.8
66
0.52
(12.
9–36
.2)
18.8
46
1.41
(11.
5–24
.7)
22.4
66
0.71
(15.
0–28
.4)
19.9
26
0.77
(13.
7–28
.8)
23.0
96
1.08
(14.
9–32
.5)
21.6
96
1.39
(11.
5–27
.0)
19.3
36
0.60
(12.
1–24
.1)
18.8
06
0.61
(10.
0–27
.0)
74 HERPETOLOGICA [Vol. 62, No. 1
TA
BL
E1.
—C
onti
nu
ed.
C.
ocel
latu
sC
.oc
ella
tus
C.o
cella
tus
C.o
cella
tus
C.
ocel
latu
sC
.oc
ella
tus
C.
bott
egi
C.r
agaz
zii
Inte
rnar
ial
dis
tan
ce(I
ND
)2.
496
0.06
(1.5
–4.0
)1.
996
0.11
(1.3
–2.3
)2.
276
0.06
(1.9
–2.8
)2.
026
0.06
(1.5
–2.5
)2.
156
0.08
(1.5
–2.8
)2.
056
0.12
(1.2
–2.8
)2.
276
0.08
(1.7
–2.9
)2.
106
0.06
(1.3
–3.0
)F
inge
rIV
len
gth
3.81
60.
09(2
.0–5
.7)
2.95
60.
19(2
.1–3
.8)
3.83
60.
10(2
.9–4
.9)
3.30
60.
12(2
.4–4
.1)
3.50
60.
18(2
.2–4
.6)
3.35
60.
22(1
.6–4
.7)
2.77
60.
07(2
.0–3
.3)
2.98
60.
09(1
.6–4
.6)
Toe
IVle
ngt
h7.
506
0.15
(3.8
–10.
7)5.
856
0.41
(3.7
–7.8
)7.
456
0.14
(6.4
–8.7
)6.
246
0.25
(4.2
–8.9
)7.
296
0.23
(5.6
–8.8
)6.
996
0.39
(4.0
–8.6
)5.
206
0.13
(3.8
–6.5
)5.
446
0.16
(2.4
–7.6
)H
L/S
VL
0.13
–0.1
90.
14–0
.19
0.13
–0.1
80.
11–0
.17
0.13
–0.1
70.
12–0
.19
0.11
–0.1
50.
12–0
.20
HW
/SV
L0.
10–0
.14
0.11
–0.1
30.
10–0
.14
0.09
–0.1
40.
10–0
.12
0.09
–0.1
30.
08–0
.12
0.09
–0.1
5A
G/S
VL
0.56
–0.7
40.
53–0
.75
0.59
–0.6
80.
60–0
.72
0.54
–0.6
60.
56–0
.66
0.61
–0.7
30.
55–0
.77
SF
/SV
L0.
22–0
.34
0.23
–0.3
30.
22–0
.33
0.22
–0.3
10.
24–0
.31
0.25
–0.3
10.
22–0
.30
0.13
–0.3
7F
LL
/SV
L0.
14–0
.16
0.14
–0.2
30.
15–0
.21
0.12
–0.1
90.
16–0
.22
0.16
–0.2
20.
11–0
.16
0.13
–0.2
1H
LL
/SV
L0.
19–0
.32
0.19
–0.2
80.
19–0
.31
0.21
–0.2
60.
25–0
.30
0.22
–0.3
20.
17–0
.24
0.18
–0.3
0S
cale
row
sar
oun
dm
idb
ody
29.5
36
0.20
(26–
32)
26.2
56
0.25
(26–
28)1
28.8
06
0.27
(26–
30)
26.3
16
0.17
(26–
28)
28.7
86
0.29
(26–
30)
28.6
76
0.25
(28–
30)
22.9
66
0.20
(22–
24)2
23.9
76
0.17
(22–
26)3
Par
aver
teb
ral/
Ver
teb
ral
wid
th0.
84–1
.11
0.79
–1.0
00.
83–1
.00
0.90
–1.0
00.
93–1
.00
0.91
–1.0
00.
67–0
.88
0.81
–1.0
0V
entr
als
78.0
56
3.13
(72–
85)
75.1
46
2.41
(72–
79)
75.4
56
0.52
(72–
80)
74.7
46
0.52
(71–
79)
76.2
86
0.43
(73–
80)
77.4
06
0.41
(74–
81)
85.3
26
0.53
(81–
92)
73.5
16
0.32
(68–
80)4
Su
bd
igit
alsc
ales
(Fin
ger
IV)
11.8
46
0.13
(10–
14)
10.8
16
0.31
(10–
12)
11.9
36
0.23
(10–
14)
10.1
16
0.26
(8–1
2)12
.83
60.
20(1
1–15
)12
.00
60.
16(1
0–13
)9.
386
0.16
(8–1
2)10
.53
60.
13(7
–13)
Su
bd
igit
alsc
ales
(Toe
IV)
17.8
96
1.12
(15–
21)
16.1
36
0.43
(14–
18)
17.7
56
0.26
(16–
20)
15.5
06
0.21
(13–
17)
18.3
16
0.24
(16–
20)
18.3
76
0.29
(17–
21)
13.5
46
0.18
(11–
15)
15.4
76
0.22
(11–
18)
Pre
sacr
alve
rteb
rae
37.8
96
1.18
(35–
42)
37.0
06
2.58
(34–
40)
37.4
06
0.25
(35–
39)
38.0
76
0.18
(37–
39)
37.2
76
0.21
(35–
38)
37.0
06
0.37
(35–
38)
41.9
66
0.15
(41–
43)
37.3
06
0.18
(33–
39)
1A
nge
l(1
936:
274)
and
An
gel
and
Lh
ote
(193
8:36
4)lis
ted
up
to30
scal
ero
ws
atm
idb
ody
for
typ
esof
Cha
lcid
esoc
ella
tus
tass
ilien
sis.
2P
aste
ur
(198
1:6)
gave
am
inim
um
of20
scal
ero
ws
atm
idb
ody
for
C.
raga
zzii
spec
imen
sth
atin
clu
ded
the
ran
geof
C.b
otte
gi.
3G
.B
oule
nge
r(1
895:
535)
not
eda
spec
imen
from
Ber
ber
a,S
omal
iaw
ith
28sc
ale
row
sat
mid
bod
y,an
dG
.B
oule
nge
r(1
896:
552)
not
edse
vera
lE
ritr
ean
spec
imen
sw
ith
up
to30
scal
ero
ws
atm
idb
ody.
4C
apu
toet
al.
(199
5:11
5)re
por
ted
am
inim
um
valu
eof
65ve
ntr
als
for
C.
raga
zzii
spec
imen
sfr
omD
jibou
ti.
March 2006] HERPETOLOGICA 75

overlapping characters that distinguish anypopulation from the others, and consider allof them them to represent one widespreadspecies (Fig. 2). For comparisons to otherspecies, data from all populations ofC. ocellatus were pooled.
Chalcides bottegi has significantly fewerscales around the midbody (t 5 20.77, df 565, P , 0.001), subdigital lamellae of FingerIII (t 5 14.44, df 5 84, P 5 0.000), Finger IV(t 5 10.99, df 5 51, P , 0.001), Toe III (t 5
12.39, df 5 49, P 5 0.000), and Toe IV (t 517.01, df 5 56, P , 0.001) than C. ocellatus.Chalcides bottegi has significantly more ven-trals (t 5 13.82, df 5 39, P , 0.001) andpresacral vertebrae (t 5 21.06, df 5 58, P ,0.001) than does C. ocellatus. Significant slopedifferences for HW (P , 0.001), Toe-IIIlength (P 5 0.008) and a marginally insignif-icant result for HLL (P 5 0.011) were notedbetween C. ocellatus and C. bottegi (Fig. 3).Chalcides bottegi also is distinguished from
FIG. 1.—Variation in dorsal color pattern in Chalcides ocellatus. (A) AMNH 148516 in life from Mali; (B) SMF 58326from Libya; (C) LACM 146091 from Egypt; (D) NMW 10437:5 from Sudan; (E) MZUF 21073 from Somalia; (F) CAS142002 from Saudi Arabia.
76 HERPETOLOGICA [Vol. 62, No. 1

C. ocellatus by a brownish vertebral stripe thatspans the vertebrals and medial half of theparavertebrals (brownish stripe spans verte-brals, paravertebrals, and dorsal scales lateralto paravertebrals in C. ocellatus; Fig. 4), andvertebrals that are always expanded (usuallyunexpanded or slightly expanded in C. ocella-tus, Table 1).
Chalcides ragazzii has significantly fewerscales around the midbody (t 5 18.80, df 5161, P , 0.001), ventrals (t 5 8.24, df 5 132,P , 0.001), subdigital lamellae of Finger III(t 5 6.11, df 5 148, P , 0.001), Finger IV (t 55.82, df 5 145, P , 0.001), Toe III (t 5 5.44,df 5 135, P 5 0.000), and Toe IV (t 5 7.73,df 5 110, P , 0.001) than does C. ocellatus. Amarginally nonsignificant difference in slopefor IND (P 5 0.013) was noted betweenC. ocellatus and C. ragazzii. Chalcides ragazziiis also distinguished from C. ocellatus by abrownish vertebral stripe that spans the verte-brals and medial half of the paravertebrals(brownish stripe spans vertebrals, paraverte-
brals, and dorsal scales lateral to paraverte-brals in C. ocellatus; Fig. 4).
Chalcides bottegi has significantly fewerdorsal scales (t 5 3.81, df 5 57, P , 0.001),subdigital lamellae of Finger III (t 5 7.47,df 5 76, P , 0.001), Finger IV (t 5 5.67, df 555, P , 0.001), Toe III (t 5 7.09, df 5 56, P ,0.001), and Toe IV (t 5 6.83, df 5 77, P ,0.001) than C. ragazzii. Chalcides bottegi hassignificantly more ventrals (t 5 19.19, df 5 42,P , 0.001) and presacral vertebrae (t 5 19.82,df 5 71, P , 0.001) than C. ragazzii.Significant slope differences for FLL (P 50.006), and IOD (P 5 0.005) were notedbetween C. bottegi and C. ragazzii (Fig. 5).Chalcides bottegi is also distinguished fromC. ragazzii by vertebrals that are always ex-panded (usually unexpanded or slightly ex-panded in C. ragazzii, Table 1). Based on thesedifferences and the separation of C. bottegiand C. ragazzii by extensive Ethiopian high-lands (Fig. 6), we recognize both taxa as validspecies and redescribe them below.
FIG. 2.—Northern Africa showing localities and literature records for Chalcides armitagei (open triangle), C. bottegi(open squares), C. pulchellus (closed triangles), C. ocellatus (closed circles), C. ragazzii (open circles), and C. thierryi(closed squares). Elevations above 1000 m are shown in dark gray; freshwater lakes are shaded in light gray. 1 5 literaturerecords of C. ocellatus in Morocco, Western Sahara, Mauritania, and Somalia from G. Boulenger (1898), Bons andGeniez (1996), Dekeyser and Villiers (1956), Lanza (1954), Laurent and Gans (1965), Spawls et al. (2002), and Geniezet al. (2004).
March 2006] HERPETOLOGICA 77

SPECIES REDESCRIPTIONS
Chalcides bottegi BoulengerFig. 7A,B
Chalcides bottegi G. Boulenger 1898:719–720;Mertens 1921:119–120.
Chalcides ragazzii Pasteur 1981:6–7; Caputoet al. 1995:101, 131; Schleich et al.1996:333–334.
Chalcides ragazzii bottegi Pasteur 1981:11–12;Welch 1982:71; Caputo et al. 1995:95, 98,116, 150; Schleich et al. 1996:348.
Chalcides ocellatus Spawls et al. 2002:120,156–157.
Chalcides ocellatus bottegi Drewes 1972:7, 10;Spawls et al. 2002:157.
Holotype.—MSNG 28569, adult (deter-mined by comparison to large sample ofadults) of unknown gender, collected betweenSancurar (48 N, 408 E) and Amarr (region
FIG. 4.—Photomicrographs of the dorsum of Chalcidesbottegi, North African C. ocellatus, and C. ragazzii justposterior to the forelimbs, showing variation in width of thebrownish vertebral stripe relative to the vertebrals, para-vertebrals, and dorsals just lateral to the paravertebrals. (A)MNHN 1990.285, C. ocellatus, showing a brown vertebralstripe with dark brown lateral borders spanning the vertebrals,paravertebrals, and dorsal scales lateral to the paravertebrals;(B) BMNH 1905.11.7.32, C. ragazzii, showing a brownvertebral stripe with dark brown lateral borders spanning thevertebrals and medial half of the paravertebrals; (C) NMK L-1645/2, C. bottegi, showing a pale brown vertebral stripe withbrown lateral borders spanning the vertebrals and medial halfof the paravertebrals. Scale bars 5 5 mm.
FIG. 3.—Bivariate plots for two characters that differsignificantly between Chalcides ocellatus (closed circles,solid line of best fit) and C. bottegi (open squares, dashedline of best fit). HW 5 head width.
78 HERPETOLOGICA [Vol. 62, No. 1

northwest of Lake Chew Bahir), Ethiopia (seemap in Vannutelli and Citerni, 1897) byVittorio Bottego in 1896.
Diagnosis.—Chalcides bottegi is a large,pentadactyl species with extensively expandedvertebral scales, a long tail, a vertebral palebrown stripe with lateral, thin brown bordersthat spans the vertebrals and medial half of theparavertebrals, a cream stripe on the lateralsides of the dorsum, and an irregular brownstripe on the flanks; the dorsum, flanks, and tailare extensively covered with brown spots andwhite ocelli. Chalcides bottegi is distinguishedfrom C. armitagei by the possession ofpentadactyl limbs (tridactyl in C. armitagei),six supralabials (5 in C. armitagei), six infrala-bials (4 in C. armitagei), one postnasal (absentin C. armitagei), 81–92 ventrals (94–96 inC. armitagei), and 41–43 presacral vertebrae(50–51 in C. armitagei) (Caputo et al., 1995).Chalcides bottegi is distinguished from C.pulchellus and C. thierryi by having a vertebralpale brown stripe with lateral, thin brownborders that spans the vertebrals and medialhalf of the paravertebrals, a cream stripe onthe lateral sides of the dorsum, and an irre-gular brown stripe on the flanks (no stripes inC. pulchellus and C. thierryi), and extensivebrown spots with white ocelli on the body (nowhite ocelli on the body in C. pulchellus andC. thierryi). Differences among populations ofC. bottegi, C. ocellatus and C. ragazzii areshown in Table 1. Chalcides bottegi is distin-guished from C. ocellatus by having a vertebralpale brown stripe with lateral, thin brownborders that spans the vertebrals and medialhalf of the paravertebrals, a cream stripe onthe lateral sides of the dorsum, and an irre-gular brown stripe on the flanks (no stripes orvertebral stripe spanning vertebrals, paraver-tebrals, and dorsals lateral to paravertebrals inC. ocellatus), always extensively expandedvertebral scales (usually unexpanded orslightly expanded in C. ocellatus), 22–24 scalerows around midbody (26–32 in C. ocellatus),and 41–43 presacral vertebrae (34–39 in all butthe North African population of C. ocellatus).Chalcides bottegi is distinguished fromC. ragazzii by having brown borders on eachside of the pale brown vertebral stripe (bolddark brown borders on a pale brown or brownvertebral stripe in C. ragazzii; Fig. 7), alwaysextensively expanded vertebral scales (usually
unexpanded or slightly expanded in C. ragaz-zii), 81–92 ventrals (68–80 in C. ragazzii),and 41–43 presacral vertebrae (33–39 inC. ragazzii).
Description of holotype.—An adult (basedon body size of other adult specimens) ofunknown gender, which has a ventral incisionand is missing all internal organs in the bodycavity; morphometric and meristic data areshown in Table 2. Head small, as wide as neck;snout short, sides converge to rounded point,slightly projects beyond labial margin; body aswide as neck, slightly depressed; limbs mod-erately short, forelimb laid forward reachesjust past ear opening, hind limb fails to reachaxilla by 60.2 mm; tail round in cross section,tapers gradually to fine point.
A pair of narrow, elongate supranasals forma suture at the median line, which contact therostral anteriorly, and the internasal posteri-orly; internasal contacts the anterior loreal, firstsupraocular, as well as supranasals and frontal;
FIG. 5.—Bivariate plots for two characters that differsignificantly between Chalcides ragazzii (open circles,solid line of best fit) and C. bottegi (open squares, dashedline of best fit). FLL 5 forelimb length, IOD 5interorbital distance.
March 2006] HERPETOLOGICA 79

frontal longer than wide, contacts first threesupraoculars laterally, and interparietal andboth parietals posteriorly; parietals large, meetbehind interparietal; interparietal shield-shaped, with parietal eye in central part ofscale; two unkeeled, subequal nuchals meet atangle behind interparietal; first supraocularlargest, slightly larger than second; third supra-ciliary longest on left side, fourth on right; nasalpierced by nostril at anterior edge, nostril aboveand anterior to suture between rostral and firstsupralabial; postnasal small, between nasal,supranasal, anterior loreal, and first two supra-labials; anterior loreal large, contacts posteriorloreal, first supraciliary, first supraocular, inter-nasal, supranasal, postnasal, and second supra-labial; posterior loreal about twice as high asfirst supraciliary; three small postsubocularsjust above sixth supralabial; lower eyelid witha rectangular, semi-transparent window; ear
opening much larger than nostril, ventrallytriangular or circular; mental large, wider thanrostral, bordered posteriorly by slightly largerpostmental; two anterior chin shields meetmedially behind postmental; vertebrals mod-erately expanded, about 2.7 times wider thanlong (1.3 times wider than paravertebrals);other dorsal, lateral, and ventral scales of thetrunk subequal, smooth, imbricate, hexagonal,about twice as wide as long; ventral scales ofmanus and pes juxtaposed, raised, and smooth.
Coloration of holotype in preservative.—Much of the original color pattern of thisspecimen has faded. Dorsum with a palebrown vertebral stripe (covers vertebrals andmedial half of paravertebrals) from parietals tobase of tail, flanked on each side by a creamstripe. Numerous dark brown spots containingwhite ocelli from dorsum of neck to tail. Anirregular, brown stripe from nares through eye
FIG. 6.—East Africa and the Arabian Peninsula illustrating the highlands of the Rift Valley, and collection localities forChalcides bottegi (open squares), C. ocellatus (closed circles), and C. ragazzii (open circles). 1 5 literature record of C.ocellatus from Lugh, Somalia (G. Boulenger, 1898; Lanza, 1954; Spawls et al., 2002). 2 5 literature record of C. ocellatusfrom Candala, Somalia (Laurent and Gans, 1965). 3 5 literature records of C. ocellatus from Ta’izz and Bajil, Yemen(Schmidt, 1953).
80 HERPETOLOGICA [Vol. 62, No. 1

to insertion of hind limb. Tail with transverserows of brown spots (all containing whiteocelli). Brown blotches on the posterior partsof the labials. Venter immaculate cream.
Variation.—Morphological variation of thisspecies is summarized in Table 1. One specimen(NMK L-1645/1) has a frontal scale fused to theinterparietal. Another specimen (CAS 131511)is unusual in having 2 postnasals and 2 posteriorloreals (right side only). The color pattern ofmost specimens resembles that of the holotype,except the vertebral stripe is pale brown witha slightly darker brown border between thevertebral stripe and cream lateral stripe, at leastto midbody. One specimen (MCZ 84185) differsfrom all others by having a rusty brown groundcolor, only slightly darker in the vertebral area,with no dark brown lines distinguishing thevertebral stripe from the paler lateral stripe. Thevalues of dorsal scales around midbody, ventrals,and presacral vertebrae differ slightly fromthose provided by Caputo et al. (1995:116) foreight specimens from ‘‘Abyssinia sensu lato,’’which likely include specimens of C. ragazzii as
defined herein. Unlike any other examinedspecies of Chalcides, the anterior loreal scaledoes not contact Supralabial 3 in most speci-mens of C. bottegi. However, there is somevariation of this character among specimens ofC. bottegi, and it should not be used asa diagnostic character for the species (Fig. 8).
Natural history.—Little is known about thenatural history of this species. At Lokomarinyang,specimens were found under stones in a rockyfield between 1645 and 1900 h at 34.88C. Thespecimen collected 1 km south of Lokomarinyangwas under a rock near low bushes at 1200–1300 hat 358C. At Kakuma, an individual was undera rock at 1230 h. Both specimens from Lokitaungwere under rocks, and one of them was buried inthe soil under a rock. One specimen fromEthiopia (BMNH 1969.1180) contained ants inits intestine. Spawls et al. (2002) suggested thatthis species is probably diurnal, although noobservations of foraging have been made.
Distribution.—Chalcides bottegi is knownfrom savanna and montane forests at low tomoderateelevationsofEthiopia,Kenya,andSudan.
FIG. 7.—Photographs of Chalcides bottegi and C. ragazzii showing unfaded dorsal color patterns and type specimens.(A) C. bottegi, NMK L-1678; (B) C. bottegi (holotype), MSNG 28569; (C) C. ragazzii, BMNH 1905.11.7.33; (D)C. ragazzii (holotype), BMNH 1946.8.21.31.
March 2006] HERPETOLOGICA 81

Etymology.—This species was named inhonor of Captain Vittorio Bottego, who seemsto have met his demise during the expeditionin which he collected the holotype (G.Boulenger, 1898).
Chalcides ragazzii BoulengerFig. 7C,D
Chalcides ocellatus var. Ragazzii G. Boulenger1890:444; G. Boulenger 1891b:12; G.Boulenger 1896:552.
Chalcides ocellatus var. humilis G. Boulenger1896:552.
Chalcides ocellatus bottegi Parker 1932:337, 361.Chalcides ocellatus humilis Mertens 1921:117,
119–120.Chalcides ocellatus ragazzii Mertens 1921:117,
119–120.
Chalcides ragazzii Pasteur 1981:2, 5–7, 14;Caputo et al. 1995:101, 131; Schleich et al.1996:333–334, 347; Spawls et al. 2002:157.
Chalcides ragazzii ragazzii Pasteur 1981:11–12, 14; Welch 1982:71; Caputo et al.1995:95, 98, 116, 150; Schleich et al.1996:348.
Chalcides ocellatus G. Boulenger 1895:535; G.Boulenger 1896:551–552; Le Berre1989:216–217; Spawls et al. 2002:156–157.
Holotype.—BMNH 1946.8.21.31 (formerlyBMNH 96.5.19.50), presumed subadult (de-termined by comparison to large sample ofadults) of unknown gender, collected by V.Ragazzi on February 1893 in Assab, Eritrea.
Diagnosis.—Chalcides ragazzii is a large,pentadactyl species with unexpanded or slight-ly expanded vertebral scales, a long tail, avertebral pale brown or brown stripe with
TABLE 2.—Selected measurements (in mm) and meristic data of the holotypes of Chalcides bottegi and C. ragazzii.
Measurement
C. bottegiMSNG28569
C. ragazziiBMNH
1946.8.21.31 Character
C. bottegiMSNG28569
C. ragazziiBMNH
1946.8.21.31
Snout–vent length 124.01 74.82 Supraoculars (L,R) 5,5 5,5Head length 15.2 10.7 Supraciliaries (L,R) 6,8 7,7Head depth 8.2 6.5 Supralabials (L,R) 6,6 6,6Maximum head width 10.7 7.5 Supralabials anterior to
1st subocular (L,R)4,4 4,4
Axilla–groin length 83.2 47.9 Infralabials (L,R) 6,6 6,6Snout–forelimb length 27.3 18.7 Scale rows around midbody 24 24Forelimb length 16.3 12.0 Ventrals from mental (excluded)
to precloacal (included)90 77
Hind limb length 23.0 15.3 Preoculars (L,R) 4,4 3,3Frontal scale length 5.9 4.2 Postnasals (L,R) 1,1 1,1Internasal width 4.1 2.8 Anterior and posterior loreals (L,R) 2,2 2,2Interparietal length 1.5 1.3 Postlabials (L,R) 2,2 2,2Ear-opening height 0.5 0.6 Postsuboculars (L,R) 3,3 2,2Ear-opening length 0.8 0.6 Subdigital scales Finger II (L,R) 8,8 8,8Eye height 1.4 1.2 Subdigital scales Finger III (L,R) 9,10 10,11Eye length 2.7 1.7 Subdigital scales Finger IV (L,R) 9,9 12,12Lower eyelid window length 1.5 1.1 Subdigital scales Finger V (L,R) 7,8 8,8Internarial distance 2.9 1.9 Subdigital scales Toe I (L,R) 7,6 6,6Interocular distance 6.2 4.9 Subdigital scales Toe II (L,R) 9,8 10,9Eye–naris distance 4.2 3.2 Subdigital scales Toe III (L,R) 12,11 14,14Eye–ear distance 6.6 4.5 Subdigital scales Toe IV (L,R) 14,13 17,—Tail length 97.5 91.4 Subdigital scales Toe V (L,R) 8,8 11,11Tail width 10.0 6.4 Presacral vertebrae 42 38Tail depth 8.9 6.2Finger II length 2.3 2.6Finger III length 3.5 3.3Finger IV length 2.8 3.0Toe III length 5.4 5.0Toe IV length 6.1 5.5Toe V length 2.8 3.0
1 G. Boulenger (1898) gave measurements for total length, head length, head width, ‘‘body,’’ forelimb, hind limb, and tail that are slightly larger (excepttail length) from the data provided here.
2 G. Boulenger (1890) noted SVL of this specimen was 83 mm.
82 HERPETOLOGICA [Vol. 62, No. 1

lateral, thin dark brown borders that spans thevertebrals and medial half of the paraverte-brals, a cream stripe on the lateral sides of thedorsum, and an irregular dark brown stripe onthe flanks; the dorsum, flanks, and tail areextensively to slightly covered with dark brownspots and white ocelli. Chalcides ragazzii isdistinguished from C. armitagei by the posses-sion of pentadactyl limbs (tridactyl in C.armitagei), six supralabials (5 in C. armitagei),six infralabials (4 in C. armitagei), one postnasal(absent in C. armitagei), 68–80 ventrals (94–96in C. armitagei), and 33–39 presacral vertebrae(50–51 in C. armitagei) (Caputo et al., 1995).Chalcides ragazzii is distinguished from C.pulchellus and C. thierryi by having a vertebralpale brown or brown stripe with lateral, thindark brown borders that spans the vertebralsand medial half of the paravertebrals, a creamstripe on the lateral sides of the dorsum, and anirregular dark brown stripe on the flanks (nostripes in C. pulchellus and C. thierryi),
numerous brown spots with white ocelli onthe body (no white ocelli in C. pulchellus and C.thierryi), and usually unexpanded or slightlyexpanded vertebrals (always extensively ex-panded vertebrals in C. pulchellus and C.thierryi). Differences among populations ofC. bottegi, C. ocellatus and C. ragazziii areshown in Table 1. Chalcides ragazzii isdistinguished from C. ocellatus by havinga vertebral pale brown or brown stripe withlateral, thin dark brown borders that spans thevertebrals and medial half of the paraverte-brals, a cream stripe on the lateral sides of thedorsum, and an irregular dark brown stripe onthe flanks (no stripes or vertebral stripespanning vertebrals, paravertebrals, and dor-sals lateral to paravertebrals in C. ocellatus),and 22–26 scale rows around midbody (26–32in C. ocellatus). Chalcides ragazzii is distin-guished from C. bottegi by having a vertebralpale brown or brown stripe bordered on eachside by a thin, dark brown stripe (pale brown
FIG. 8.—Photomicrographs of the right anterior heads of Chalcides bottegi, C. ocellatus and C. ragazzii showingvariation in position of the anterior loreal scale (AL) relative to Supralabials 1–3. (A) C. ocellatus, KU 69012, showingcontact of anterior loreal and Supralabials 2 and 3; (B) BMNH 1992.456, C. ragazzii, showing contact of anterior lorealand Supralabials 2 and 3; (C) NMK L-1643, C. bottegi, showing no contact between anterior loreal and Supralabial 3; (D)MCZ 84186, C. bottegi, showing contact between anterior loreal and Supralabials 2 and 3. Scale bars 5 1 mm.
March 2006] HERPETOLOGICA 83

vertebral stripe with brown borders in C.ragazzii; Fig. 7), usually unexpanded or slightlyexpanded vertebrals (always extensively ex-panded vertebrals in C. bottegi), 68–80 ventrals(81–92 in C. bottegi), and 33–39 presacralvertebrae (41–43 in C. bottegi).
Description of holotype.—The holotype(Fig. 7D) is likely a subadult (many other adultspecimens from Eritrea are larger) and ofunknown sex; morphometric and meristic dataare shown in Table 2. Head small, as wide asneck; snout short, sides converging to roundedpoint, slightly projecting beyond labial margin;body slightly wider than neck, slightly de-pressed; limbs moderately short, forelimb laidforward reaches just past ear opening, hindlimb fails to reach axilla by 32.6 mm; tail roundin cross section, tapering gradually to fine point.
A pair of narrow, elongate supranasals formsa suture at the median line, which contact therostral anteriorly, and the internasal posteri-orly; internasal contacts the loreal, first supra-ocular, as well as supranasals and frontal;frontal longer than wide, contacts first threesupraoculars laterally, and interparietal andboth parietals posteriorly; parietals large, meet-ing behind interparietal; interparietal shield-shaped, with parietal eye in central part of scale;two unkeeled nuchals (unequal in width) meetat angle behind interparietal; first supraocularlargest, slightly larger than second; secondsupraciliary longest; nasal pierced by nostril atanterior edge, nostril above suture betweenrostral and first supralabial; postnasal small,between nasal, supranasal, loreal, and first twosupralabials; anterior loreal large, contactsposterior loreal and first supraciliary, firstsupraocular, internasal, supranasal, postnasal,and second and third supralabials; posteriorloreal about twice as high as first supraciliary;two small postsuboculars just above sixthsupralabial; lower eyelid with a rectangular,semi-transparent window; ear opening muchlarger than nostril, ventrally triangular tocircular; mental large, wider than rostral,bordered posteriorly by one equally largepostmental; two anterior chin shields meetmedially behind postmental; vertebral scalesslightly expanded, about two times wider thanlong (only 0.1 mm wider than paravertebrals);other dorsal, lateral, and ventral scales of thetrunk subequal, smooth, imbricate, hexagonal,
about twice as wide as long; ventral scales ofmanus and pes juxtaposed, raised, and smooth.
Coloration of holotype in preservative.—Much of the original color pattern of thisspecimen has faded. Dorsal color tan, gradinginto immaculate cream on the venter. Faintevidence of a dark brown stripe from frontalscale across center of dorsum, including bothvertebral scales and medial half of adjacentparavertebrals. White ocelli in brown spotsbecome more numerous posteriorly on theflanks and dorsum. Ocelli arranged in transverserows on tail, but not reaching to venter. A brown,irregular stripe present from point just anteriorto narial opening through eye and ear opening toabout 1 cm past insertion of forelimb, about 3.5scales wide. Posterior borders of labials andsome head plates edged in brown.
Variation.—Morphological variation in thisspecies is shown in Table 1. Most specimenshave a color pattern similar to the holotype,but the pale brown or brown vertebral stripe,dark brown lateral borders of the stripe, creamstripes on the lateral sides of the dorsum, anddark brown, irregular lateral stripes of theflanks are much more contrasting than theholotype. The number of dark brown spots onthe dorsum is variable; one specimen fromSomalia (BMNH 1905.10.30.103) lacks dorsalspots except in the region just proximal to thebase of the tail. Specimens from Harar, Ethiopia(BMNH 1911.12.13.11–14, 1970.1462–67),have more of an olive-brown background color(possibly an artifact of specimen preservation)with less contrast between the vertebral stripeand paler lateral stripe of the dorsum; someof these specimens have an additional darkbrown stripe in the center of the brownvertebral stripe, similar (but not identical) tothe color pattern of C. ocellatus from Libya(Fig. 1B). Specimens from Djibouti havea relatively darker brown ground color thanother specimens, with less contrast betweenthe darker vertebral stripe and paler lateralstripe. Most specimens have an immaculatecream venter, but some have pale brownpigmentation under the chin. One specimen(BMNH 1937.12.5.758) has seven supralabialson the right side and six on the left; anotherspecimen from a nearby locality has only fivesupralabials (both sides of body). Caputo et al.(1995:115) reported a minimum value of 65 ven-trals for seven specimens from Obock, Djibouti.
84 HERPETOLOGICA [Vol. 62, No. 1

Natural history.—Little is known about thenatural history of this species. One specimenfrom Somalia (BMNH 1937.12.5.753) hadorthopteran body parts in its punctured stomach.
Distribution.—Chalcides ragazzii is knownfrom xeric grassland and montane forest atsea level to moderate elevations (1515 m) ofDjibouti, Eritrea, Ethiopia, and northwesternSomalia.
Etymology.—This species was named inhonor of the collector of the holotype, Dr.V. Ragazzi.
KEY TO THE SUB-SAHARAN SPECIES
OF CHALCIDES
1. Limbs tridactyl, less than six supralabials, lessthan six infralabials, postnasal absent, more than93 ventrals, more than 45 presacral vertebrae(Gambia) - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - C. armitageiLimbs pentadactyl, six (or more) supralabials, sixinfralabials, postnasal present, fewer than 93ventrals, fewer than 45 presacral vertebrae - - - - - - - - - - 2
2. Vertebral scales extensively expanded (para-vertebral width/vertebral width , 0.9) - - - - - - - - - - - - - - - - 3Vertebral scales not expanded, or only weaklyexpanded (paravertebral width/vertebral width. 0.9) - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - 5
3. Vertebral body stripe present, and brown spotswith white ocelli markings on body (easternSudan to northern Kenya) - - - - - - - - - - - - - - - - - - - - - C. bottegiNo vertebral body stripe, and no brown spotswith white ocelli markings on body - - - - - - - - - - - - - - - - - - - - 4
4. Five phalanges in Toe IV, 22–24 scales atmidbody, 79–86 ventrals, 13–17 subdigitallamellae under Toe IV, 37–42 presacral verte-brae, brown spots of tail without white borders(western Guinea to southeastern Senegal andsoutheastern Mali) - - - - - - - - - - - - - - - - - - - - - - - - - - C. pulchellusFour phalanges in Toe IV, 20 (rarely 22) scales atmidbody, 85–91 ventrals, 9–13 subdigital lamel-lae under Toe IV, 41–44 presacral vertebrae,brown spots of tail usually with white borders(northern Ghana to eastern Nigeria) - - - - - - - C. thierryi
5. 22–26 scale rows at midbody, brown vertebralstripe present and spanning 4 mid-dorsal scalerows (Eritrea to northwestern Somalia) - - - C. ragazzii26–32 scale rows at midbody; brown vertebralstripe present and spanning 6 mid-dorsal scalerows (Sahara and sub-Sahara) or absent (So-malia) - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - C. ocellatus
DISCUSSION
Our results support the current taxonomicstatus of C. ocellatus throughout Africa and theArabian Peninsula, and our recognition of C.bottegi and C. ragazzii should provide taxo-nomic stability to a group that has been ina state of flux for more than a century. However,
given the variation of color pattern amongparapatric populations of C. ocellatus examinedin this study (Fig. 1), we are skeptical oftaxonomic recognitions based solely on minordifferences in color-pattern characters (e.g.,some Moroccan species and subspecies of theC. ocellatus group, Schleich et al., 1996:333).Conversely, many morphological features andnumbers of scales are similar among popula-tions of C. ocellatus (Table 1), which could bea result of conservative evolution, recent (5000years bp) speciation events resulting fromaridification in the Sahara (Burgess et al.,2004), or character convergence. For example,we did not detect morphological differencesbetween the potentially isolated C. ocellatusspecimens from Somalia and those in otherparts of Africa and the Arabian Peninsula. Wefind this surprising considering that theseregions have been separated by the Ethiopianhighlands and the Gulf of Aden (Fig. 6) for atleast 5–7 million years (Arnold, 1987; Coleman,1993; Joger, 1987; Smith, 1989). Moreover,among the drier ecoregions of mainland Africa,the Horn of Africa contains the highest levels ofendemism for birds, mammals, and reptiles(Burgess et al., 2004; Kingdon, 1989). In theoriginal description of C. ocellatus sacchii fromLugh, Somalia, Lanza (1954) diagnosed his newsubspecies from the Mediterranean form C.ocellatus linosae based on minor differences incolor pattern and number of scale rows aroundthe midbody. But when compared to a morecomprehensive data set, none of Lanza’s (1954)characters or any others examined by usdistinguished the Somalian population of C.ocellatus from conspecifics elsewhere in Africaor Saudi Arabia (Table 1).
Explaining the disjunct distribution of C.ocellatus in Somalia is problematic, and thesimilarity in color pattern and morphologybetween this population and conspecifics inSaudi Arabia (Table 1) is intriguing. Thissimilarity suggests possible dispersal betweenthe Horn of Africa and the Arabian Peninsula,perhaps across the shallow (137 m) B�ab-al-Mandeb seafloor during several periods of lowsea levels (at least 120 m lower than present) inthe Pleistocene (Coleman, 1993; Joger, 1987;Rohling et al., 1998). Wildman et al. (2004)noted that at least 62 species of mammals occuron both sides of the Red Sea and concluded thatthis ‘‘point[s] to a Pleistocene connection across
March 2006] HERPETOLOGICA 85

the straits.’’ However, consulting mammaliandistribution sources for the Arabian Peninsula(Harrison and Bates, 1991) and northeast Africa(Kingdon, 1997) reveals that half of thesespecies are bats capable of flying across theseaway, and most of the remaining species havea continuous distribution from the ArabianPeninsula across the Sinai Peninsula to EastAfrica. Moreover, the Red Sea seems to havebeen an effective biogeographic barrier todispersal for most herpetological species—noamphibians and up to 27 reptile species occuron both sides of the Red Sea in Eritrea andSomalia (Arnold, 1987; Frost, 2004). Like thesituation with mammals, several of these reptileshave a continuous or nearly continuous distri-bution from Somalia (and other areas of EastAfrica) across the Sinai Peninsula to the ArabianPeninsula (Joger, 1987; Leviton et al., 1992;Spawls et al., 2002), and thus dispersal acrossthe Red Sea does not necessarily account fortheir current distribution. Given this evidenceand the absence of C. ocellatus from the Africanside of the B�ab-al-Mandeb putative landbridge,the Somali population of C. ocellatus mightrepresent a geographically isolated relict pop-ulation in the Horn of Africa, and it is possiblethat molecular studies will support the recogni-tion of this allopatric population as C. sacchii.
The occurrence of isolated populations ofC. ocellatus in the Sahelian Niger River flood-plain and montane xeric vegetation of thedisjunct highlands around the periphery of theSahara is consistent with the distribution ofrelict populations of dozens of other terrestrialamphibians, reptiles, and mammals (Kingdon,1997; Le Berre, 1989; Schleich et al., 1996). It islikely that C. ocellatus, like many other terres-trial tetrapods, was once widespread in northernAfrica in the Palearctic and Afrotropical realms,but became trapped in pockets of relativelymesic habitats as the cooling and aridification ofthe Sahara progressed over the course of theMiocene to the present (Borkin, 1999; Burgesset al., 2004; Douady et al., 2003; Zachos et al.,2001). In West Africa, C. ocellatus is replaced byC. armitagei, C. pulchellus, and C. thierryi insavanna mosaic habitats south of the Sahel,whereas C. bottegi replaces C. ocellatus in thesavanna and bushland habitats southeast of theSahel in East Africa (Greenbaum, 2005).
The regional endemism shown by C. bottegifor the western and southern flanks of the
Ethiopian Highlands, and C. ragazzii for theEthiopian xeric grasslands and shrublands, isalso noteworthy. The historical factors respon-sible for producing these patterns of regionalendemism in the Horn of Africa are not yet wellunderstood for Chalcides, but may possibly belinked with glacial refugia associated with theEthiopian Highlands (Fjeldsa and Lovett,1997) or localized climatic conditions. Theabsence of sympatric C. ocellatus in these areasis especially intriguing considering the other-wise very broad distribution of this species.
A recent mtDNA analysis of skinks suggestedthat Chalcides is paraphyletic with respect toSphenops (Brandley et al., 2005), and therefore,Chalcides may include more species thancurrently recognized. A molecular study willbe useful to further explore phylogeneticrelationships within the genus Chalcides, andtest the hypotheses and distribution patternsdiscussed above. Our morphological evidencesuggests a strong phenetic similarity among C.bottegi, C. pulchellus, and C. thierryi. All ofthese species, relative to C. ocellatus and C.ragazzii, have extensively expanded vertebralscales, an elongate body, reduced limbs, andhigh numbers of presacral vertebrae (Table 1;Greenbaum, 2005). These morphological fea-tures have been associated with adaptations toenter narrow spaces in search of food or shelter(Caputo et al., 1995; Gans, 1975), which isconsistent with the known natural history ofthese species. Based on these shared charac-ters, which we suspect are apomorphic, wehypothesize that these species (all distributedsouth of the Sahel) are closely related. Of keyinterest to future molecular studies are poten-tial cryptic species, hybridization, trends inbody elongation and limb reduction (Caputo etal., 1995), and the role of the Sahara andEthiopian highlands in the evolution of Chal-cides and other terrestrial vertebrates (Douadyet al., 2003; Fjeldsa and Lovett, 1997).
Despite reasonably comprehensive collectingin Mali (e.g., Bohme, 1985; Bohme et al., 1996;Joger, 1981; Joger and Lambert, 1996, 1997;Papenfuss, 1969), C. ocellatus has not beenreported until the present study, underscoringthe importance of continued collecting in theSahel from Mauritania to Sudan. Moreover,given the recent discovery of Chalcides inwestern Guinea, additional efforts are neededin sub-Saharan grasslands and dry forests
86 HERPETOLOGICA [Vol. 62, No. 1

(Greenbaum, 2005; Greenbaum and Carr,2005). Although the natural habitats of theChalcides discussed in this paper face numer-ous threats (Burgess et al., 2004; Stuart andAdams, 1990), not one of the species (except C.armitagei) has a distribution sufficiently small towarrant threatened status (IUCN, 2001). Theproximity of several of the collection localities tohuman habitations and major cities suggeststhat these semi-fossorial lizards do not reactnegatively to human disturbance.
Acknowledgments.—We thank the curators and staffof the numerous museums that allowed us to examinespecimens—especially the African and European institu-tions that were understanding and patient with us as weadapted to new laws that regulate shipment of specimenspreserved in ethanol. Our (EG and CJR) fieldwork in Malicorresponded with the 2001 US–West Africa Workshop:Niger River Aquatic and Riparian Vertebrate Conservation,which was funded by an NSF grant (INT-0080699) to F.Pezold of the University of Louisiana at Monroe. We thankthe Mali authorities for research permits and our fieldcompanion Omar Attum for assistance in Mali. G. Pasteurkindly discussed his research with Chalcides. R. Houserand S. McEathron assisted with GIS data. The manuscriptwas improved by comments from W. E. Duellman, O.Torres-Carvajal, L. Trueb, and two anonymous reviewers.
LITERATURE CITED
ADLER, K. (Ed.). 1989. Contributions to the History ofHerpetology. Contributions to Herpetology No. 5.Society for the Study of Amphibians and Reptiles,Oxford, Ohio, U.S.A.
ANGEL, M. F. 1936. Sur quelques formes nouvelles dereptiles et de batraciens du Sahara central. Bulletin de laSociete zoologique de France 61:273–277.
ANGEL, F., AND H. LHOTE. 1938. Reptiles et amphibiens duSahara central et du Soudan. Bulletin du Comited’Etudes historiques et scientifiques de l’Afriqueoccidentale Francaise 21:347–381.
ARNOLD, E. N. 1987. Zoogeography of the reptiles andamphibians of Arabia. Pp. 245–256. In F. Krupp, W.Schneider, and R. Kinzelbach (Eds.), Proceedings of theSymposium on the Fauna and Zoogeography of theMiddle East. Tubinger Atlas des Vorderen Orients,Beiheft A 28, Wiesbaden, Germany.
BOHME, W. 1985. Zoogeographical patterns of the lizardfauna of the African sub-Saharan savanna belt, withpreliminary description of a new chameleon. Pp. 471–478. In K. Schuchmann (Ed.), Proceedings of theInternational Symposium on African Vertebrates.Bonn, Germany.
BOHME, W., H. MEINIG, AND M.-O. RODEL. 1996. Newrecords of amphibians and reptiles from Burkina Fasoand Mali. British Herpetological Society Bulletin (56):7–26.
BONS, J., AND P. GENIEZ. 1996. Amphibiens et Reptiles duMaroc (Sahara Occidental compris) Atlas Biogeogra-phique. Asociacion Herpetologica Espanola, Barcelona,Spain.
BORKIN, L. J. 1999. Distribution of amphibians in NorthAfrica, Europe, Western Asia, and the former SovietUnion. Pp. 329–420. In W. E. Duellman (Ed.), Patternsof Distribution of Amphibians: a Global Perspective.The Johns Hopkins University Press, Baltimore, Mary-land, U.S.A.
BOULENGER, E. G. 1920. On some lizards of the genusChalcides. Proceedings of the Zoological Society ofLondon 1920:77–83.
BOULENGER, G. A. 1887. Catalogue of the Lizards in theBritish Museum (Natural History), 2nd ed., Vol. 3.Lacertidae, Gerrhosauridae, Scincidae, Anelytropidae,Dibamidae, Chamaeleontidae. Taylor and Francis,London, U.K.
———. 1890. On the varieties of Chalcides ocellatus,Forsk. Annals and Magazine of Natural History 5:444–445.
———. 1891a. Catalogue of the reptiles and batrachiansof Barbary (Morocco, Algeria, Tunisia). Transactions ofthe Zoological Society of London 13:93–164.
———. 1891b. On some reptiles collected by Sig. L.Bricchetti Robecchi in Somaliland. Annali del MuseoCivicio di Storia Naturale di Genova 12:3–15.
———. 1895. An account of the reptiles and batrachianscollected by Dr. A. Donaldson Smith in western Somali-land and the Galla country. Proceedings of theZoological Society of London 1895:530–540.
———. 1896. A list of the reptiles and batrachianscollected by Dr. Ragazzi in Shoa and Eritrea. Annalidel Museo Civicio di Storia Naturale di Genova 16:546–554.
———. 1898. Concluding report of the late Capt.Bottego’s collection of reptiles and batrachians fromSomaliland and British East Africa. Annali delMuseo Civicio di Storia Naturale di Genova 18:715–723.
BRANDLEY, M. C., A. SCHMITZ, AND T. W. REEDER. 2005.Partitioned Bayesian analyses, partition choice, and thephylogenetic relationships of scincid lizards. SystematicBiology 54:373–390.
BROWN, R. P., R. CAMPOS-DELGADO, AND J. PESTANO. 2000.Mitochondrial DNA evolution and population history ofthe Tenerife skink Chalcides viridanus. MolecularEcology 9:1061–1067.
BURGESS, N., J. D’AMICO HALES, E. UNDERWOOD, E.DINERSTEIN, D. OLSON, I. ITOUA, J. SCHIPPER, T.RICKETTS, AND K. NEWMAN. 2004. Terrestrial Ecoregionsof Africa and Madagascar: a Conservation Assessment.Island Press, Washington, D.C., U.S.A.
CAPUTO, V. 1993. Taxonomy and evolution of the Chalcideschalcides complex (Reptilia, Scincidae) with descriptionof two new species. Museo Regionale di ScienzeNaturali Bolletino, Torino 11:47–120.
CAPUTO, V., B. LANZA, AND R. PALMIERI. 1995. Bodyelongation and limb reduction in the genus ChalcidesLaurenti 1768 (Squamata Scincidae): a comparativestudy. Tropical Zoology 8:95–152.
CAPUTO, V., AND J. MELLADO. 1992. A new species ofChalcides (Reptilia: Scincidae) from northeastern Mo-rocco. Bolletino di Zoologia 59:335–342.
CAPUTO, V., M. SORICE, AND L. CRESCIMBENI. 1999. Amolecular taxonomy of some Mediterranean scincidlizards, genus Chalcides Laurenti 1768 (Reptilia,Scincidae). Russian Journal of Herpetology 6:23–32.
March 2006] HERPETOLOGICA 87

COLEMAN, R. G. 1993. Geologic Evolution of the Red Sea.Oxford Monographs on Geology and Geophysics No. 24.Oxford University Press, New York, New York, U.S.A.
DEKEYSER, P. L., AND A. VILLIERS. 1956. Contributiona l’etude du peuplement de la Mauritanie: Notationsecologiques et biogeographiques sur la faune de l’Adrar.Memoires de l’Institut Francais d’Afrique Noire 44:1–222.
DOUADY, C. J., F. CATZEFLIS, J. RAMAN, M. S. SPRINGER,AND M. J. STANHOPE. 2003. The Sahara as a vicariantagent, and the role of Miocene climatic events, in thediversification of the mammalian order Macroscelidea(elephant shrews). Proceedings of the National Academyof Sciences, USA 100:8325–8330.
DREWES, R. C. 1972. Report on a collection of reptiles andamphibians from the Ilemi Triangle, southwesternSudan. Occasional Papers of the California Academyof Sciences (100):1–14.
FJELDSA, J., AND J. C. LOVETT. 1997. Geographical patternsof old and young species in African forest biota: thesignificance of specific montane areas as evolutionarycentres. Biodiversity and Conservation 6:325–346.
FROST, D. R. 2004. Amphibian Species of the World: anOnline Reference. Version 3.0 (22 August, 2004). Elec-tronic Database accessible at http://research.amnh.org/herpetology/amphibia/index.html. American Museumof Natural History, New York, New York, U.S.A.
GANS, C. 1975. Tetrapod limblessness: evolution andfunctional corollaries. American Zoologist 15:455–467.
GENIEZ, P., J. A. MATEO, M. GENIEZ, AND J. PETHER. 2004.The Amphibians and Reptiles of the Western Sahara.Edition Chimaira, Frankfurt am Main, Germany.
GREENBAUM, E. 2005. Systematics of West African skinksin the Chalcides thierryi group: composition, distribu-tion, and redescription of types. African Journal ofHerpetology 54:17–29.
GREENBAUM, E., AND J. L. CARR. 2005. The herpetofauna ofUpper Niger National Park, Guinea, West Africa.Scientific Papers, Natural History Museum, The Uni-versity of Kansas (37):1–21.
GREENBAUM, E., AND J. B. RASMUSSEN. 2004. Chalcidesthierryi pulchellus (Cylindrical skink). Attempted pre-dation. Herpetological Review 35:166–167.
HARRISON, D. L., AND P. J. J. BATES. 1991. The Mammals ofArabia. Harrison Zoological Museum, Kent, U.K.
IUCN. 2001. IUCN Red List Categories, Version 3.1.Prepared by the IUCN Species Survival Commission.IUCN, Gland, Switzerland.
JOGER, U. 1981. Zur Herpetofaunistik Westafrikas. BonnerZoologische Beitrage 32:297–340.
JOGER, U. 1987. An interpretation of reptile zoogeographyin Arabia, with special reference to Arabian herpeto-fauna relations with Africa. Pp. 257–271. In F. Krupp,W. Schneider and R. Kinzelbach (Eds.), Proceedingsof the Symposium on the Fauna and Zoogeography ofthe Middle East. Tubinger Atlas des Vorderen Orients,Beiheft A 28, Wiesbaden, Germany.
JOGER, U., AND M. R. K. LAMBERT. 1996. Analysis of theherpetofauna of the Republic of Mali, I. Annotatedinventory, with description of a new Uromastyx (Sauria:Agamidae). Journal of African Zoology 110:21–51.
JOGER, U., AND M. R. K. LAMBERT. 1997. Analysis of theherpetofauna of the Republic of Mali, II. Speciesdiversity and biogeographic assemblages. Pp. 189–202.
In H. Ulrich (Ed.), Tropical Biodiversity and System-atics. Proceedings of the International Symposium onBiodiversity and Systematics in Tropical Ecosystems,Bonn 1974, Bonn, Germany.
KINGDON, J. 1989. Island Africa: The Evolution of Africa’sRare Animals and Plants. Princeton University Press,Princeton, New Jersey, U.S.A.
KINGDON, J. 1997. The Kingdon Field Guide to AfricanMammals. Academic Press, San Diego, California,U.S.A.
KLAUSEWITZ, W. 1954. Eidonomische, taxonomische undtiergeographische Untersuchungen uber den Rassenk-reis der Scinciden Chalcides chalcides und Ch. striatus.Senckenbergiana 34:187–203.
LANZA, B. 1954. Su due nuove razze geografiche delChalcides ocellatus (Forskal) (Reptilia, Scincidae).Monitore Zoologico Italiano 62:161–173.
LANZA, B., AND S. CARFI. 1968. Gli scincidi della Somalia(Reptilia, Squamata). Monitore Zoologico Italiano,Supplemento 2:207–260.
LAURENT, R. F., AND C. GANS. 1965. Lizards. Pp. 25–45. InC. Gans, R. F. Laurent, and H. Pandit (Eds.), Notes ona Herpetological Collection from the Somali Republic.Annales Musee Royal de l’Afrique Centrale, Serie 8,Sciences Zoologiques (134):1–93þ 10 plates.
LE BERRE, M. 1989. Faune du Sahara 1: Poissons—Amphibiens Reptiles. Lechevalier—R. Chabaud, Paris,France.
LEVITON, A. E., S. C. ANDERSON, K. ADLER, AND S. A.MINTON. 1992. Handbook to Middle East Amphibiansand Reptiles. Society for the Study of Amphibians andReptiles, Oxford, Ohio, U.S.A.
LEVITON, A. E., R. H. GIBBS, JR., E. HEAL, AND C. E.DAWSON. 1985. Standards in herpetology and ichthyol-ogy: Part I. Standard symbolic codes for institutionalresource collections in herpetology and ichthyology.Copeia 1985:802–832.
LOVERIDGE, A. 1936. African reptiles and amphibians inField Museum of Natural History. Publications, Field Mu-seum of Natural History (Zoological Series) 22:1–111.
MERTENS, R. 1921. Zur Kenntis der geographischenFormen von Chalcides ocellatus Forskal, (Rept. Lac.).Senckenbergiana 3:116–120.
PAPENFUSS, T. J. 1969. Preliminary analysis of the reptilesof arid central West Africa. The Wasmann Journal ofBiology 27:249–325.
PARKER, H. W. 1932. Two collections of reptiles andamphibians from British Somaliland. Proceedings of theZoological Society of London 335–367.
PASTEUR, G. 1981. A survey of the species groups of theOld World scincid genus Chalcides. Journal of Herpe-tology 15:1–16.
ROHLING, E. J., M. FENTON, F. J. JORISSEN, P. BERTRAND,G. GANSSEN, AND J. P. CAULET. 1998. Magnitude of sea-level lowstands of the past 500,000 years. Nature394:162–165.
SCHLEICH, H. H., W. KASTLE, AND K. KABISCH. 1996.Amphibians and Reptiles of North Africa: Biology,Systematics, Field Guide. Koeltz Scientific Books,Koenigstein, Germany.
SCHMIDT, K. P. 1953. Amphibians and reptiles of Yemen.Fieldiana Zoology 34:253–261.
88 HERPETOLOGICA [Vol. 62, No. 1

SMITH, A. 1989. The Great Rift: Africa’s Changing Valley.Sterling Publishing Company, Inc., New York, NewYork, U.S.A.
SPAWLS, S., K. HOWELL, R. DREWES, AND J. ASHE. 2002.A Field Guide to the Reptiles of East Africa. AcademicPress, San Diego, California, U.S.A.
STUART, S. N., AND R. J. ADAMS. 1990. Biodiversity in sub-Saharan Africa and its Islands. Occasional Papers ofthe IUCN Species Survival Commission, No. 6. OxfordPress, Oxford, U.K.
VANNUTELLI, L., AND C. CITERNI. 1897. Relazione prelim-inare sui risultati geografici della seconda spedizionecondotta dal capitano Vittorio Bottego nell’ Africaorientale. Bolletino della Societa Geographica Italiana10:320–330þmap.
WELCH, K. R. G. 1982. Herpetology of Africa: a Checklistand Bibliography of the Orders Amphisbaenia, Sauriaand Serpentes. Robert E. Krieger Publishing Company,Malabar, Florida, U.S.A.
WILDMAN, D. E., T. J. BERGMAN, A. AL-AGHBARI, K. N.STERNER, T. K. NEWMAN, J. E. PHILLIPS-CONROY, C. J.JOLLY, AND T. R. DISOTELL. 2004. Mitochondrialevidence for the origin of hamadryas baboons. Molec-ular Phylogenetics and Evolution 32:287–296.
ZACHOS, J., M. PAGANI, L. SLOAN, E. THOMAS, AND K.BILLUPS. 2001. Trends, rhythms, and aberrations inglobal climate 65 Ma to present. Science 292:686–693.
Accepted: 16 August 2005Associate Editor: Joseph Mendelson
APPENDIX I
Specimens Examined
Some localities (marked with one asterisk) could not begeoreferenced accurately, and are not mapped. Twolocalities (marked with two asterisks) are questionable,and data from these specimens were not included in thestudy. Data from Chalcides pulchellus and C. thierryi weretaken from Greenbaum (2005).
Chalcides armitagei. GAMBIA: north of Kartong village(MWTR 003).
Chalcides bottegi. ETHIOPIA: between Sancurar andAmarr (*)(MSNG 28569 [holotype]); Bikalal Hill, 13 km NGhimbi (BMNH 1969.1180); Zegi, Lake Tsana (BMNH1902.12.13.59–65, FMNH 1870); km 400 along rd. fromGondar to Asmara (*) (MCZ 84185); 40 km S Inda Selassie(MCZ 84186). KENYA: Turkana: Kakuma, 587 m (NMKL-1643); Lokitaung, 749 m (NMK L-1645/1–3). Sam-buru: Samburu Hills (*)(NMK L-1678). SUDAN: Bados(MCZ 8804); Ilemi triangle, Lokomarinyang, 576 m (CAS131511–12, 131515, NMK L-1642/1–3); ca. 1 km SLokomarinyang, 576 m (CAS 131535).
Chalcides ocellatus. ALGERIA: Djanet, Tassili desAzdjers (syntypes of C. o. tassiliensis) (MNHN 1936.090–91); Alger ‘‘n8 255’’ (MNHN 1990.252, 1990.256–58,1990.285–89); Massif de l’Aures (MNHN 1997.5091–97);Aın Sefra (MNHN A84, 1974.306–07, 1974.309–11,1974.426, NMW 10440:9); El Mereier (NMW 10440:2,10440:4–5); Touggourt (NMW 10440:7–8); Tomassime (*)(NMW 10440:3); Ourlana (NMW 10440:1); west AlgerianSahara (*) (NMW 10440:6); 13 km S Oran (LACM121610–11); Oran Arzew (KU 23478–79). CHAD: Ennedi
(MNHN 1967.530–31). EGYPT: Cairo (NHMB 5330,6512–14); Andura (*)(NHMB 5397); Fayum, Al-Fayyum(KU 69011–13); Giza (LACM 146062, 146064–65,146069–70); Giza, El Mansura (LACM 146091, 146098);Kafr Hakim (LACM 146060); Qalyubiya, Kafr Farouk(LACM 146086, 146094); no specific locality (*) (NHMB4788–89). LIBYA: western El Agheila, 30 km S Abiseia-Oase (SMF 58326); western Barce, Wadi Tuega (*)(SMF58325); Bengasi (SMF 34464, 35552); Derna (SMF34473); Uadi Sharshara ca. Tarhuna (NHMB 16694).NIGER: Aır, oued Goufat (MNHN 1936.89). MALI:Timbuktu (ZMUC R-47.6–7); between Kona and Sevare,148 449 060 N, 038 569 100 W (AMNH 148516). SAUDIARABIA: Al nar (*)(NMW 10437:6); Khasawiyah, 168 569
N, 428 379 E, 20 m (CAS 134148); Jiddah, 218 299 N, 398
109 E (CAS 139515); Jiddah, 218 299 N, 398 119 E, 20 m(CAS 102396, 113713); Jiddah, 218 309 N, 348 129 E, ca. sealevel (CAS 142002, 144204); Jiddah, ca. 20 m (CAS113712, 113722, 139745–47); Jizan (CAS 135565, 139739);Jizan, 168 539 N, 428 299 E (CAS 136555); Ras-Al-Bourge,Jizan, 168 539 180 N, 428 329 000 E (CAS 134152–53); RoyalSaudi Navy Reservation south of Jiddah [in area 21 07–2120 N, from coast to 6 km. inland], ca. sea level (CAS136512). SOMALIA: Bari, Bender Cassim (5 Bosasso)(MZUF 10538–40); Bosasso, Hotel Kamal, 118 179 7.80 N,498 109 43.80 E, 15 m (CAS 227565, 227567); Bari, Gard(5 Gardo) (MZUF 10537); Lower Shebelli, Afgoi, Scebeli(MZUF 21072–74); Mogadishu (MZUF 21068–69); Balad,30 km N Mogadishu (MZUF 5056, 9393); no specificlocality (*) (NMW 10591:1–2); Berbera (**) (NHMB5566). SUDAN: Halfa, 26 km S Wadi Halfa, Abka (FMNH58579–85); Magatha, Atbara (NMW 10437:1); Khartoum(MCZ 22992–93, NMW 10437:2–4, 10437:7); UmmRarnad, Kordofan (NMW 10437:5); Kordofan Region(NMW 10438:1–5). TUNISIA: beach of Kelibia (MNHN1990.229–30).
Chalcides ragazzii. DJIBOUTI: Foret du Day, autourdu village Day, 1450 m (MNHN 1999.9271–74); Bank-ouale, bords de l’oued qui va de la foret du Day versBankouale, 118 48.2499 N, 428 37.8249 E, 1250 m (MNHN1999.9275); Dittilou, Berges de l’Oued du campement deDittilou, 118 46.8649 N, 428 41.6109 E, 800 m (MNHN1999.9270); region d’Ali-Sabieh, oued Obodley (MNHN1996.239). ERITREA: Assab (BMNH 1946.8.21.31 [ho-lotype]); ‘‘Adi-Cai,h ed Adi-Ugri’’ (MZUF 9041–44);‘‘Adi-CaiS^h ed Adi-Ugri’’ (MZUF 163); Adi Ugri (MZUF9392). ETHIOPIA: ca. Harrar (BMNH 1911.12.13.11–14); Harar (BMNH 1970.1462–67); Zegi, Lake Tana (**)(MCZ 112212). SOMALIA: Berbera (NHMB 5563–65);near Berbera (BMNH 1905.10.30.102–09, 1905.10.30.111–12, 116); Wagga, Goolis Mts. ca. Berbera, 909–1212 m (BMNH 1905.11.7.32–37); Dagah Shabell,10.11668 N, 45.1338 E (BMNH 1931.8.1.169); Hargeisa(BMNH 1992.455–56); 4 km SSW Hargeisa (BMNH1996.443); Borama District, 108 209 N, 458 E, 1515 m(BMNH 1937.12.5.748); Borama District, 108 309 N, 428
409 E, 1061 m (BMNH 1937.12.5.749); Borama District,108 109 N, 428 509 E, 1212–1515 m (BMNH1937.12.5.750–53); Borama District, 108 209 N, 508 E,1061 m (BMNH 1937.12.5.754); Borama District, 108 059N, 438 E, 1515 m (BMNH 1937.12.5.755–59); island offZeilah (BMNH 1937.12.5.776); no specific locality (*)(NMW 10592:1–3).
March 2006] HERPETOLOGICA 89
Related Documents











