ORIGINAL INVESTIGATION A reliable method to study cue-, priming-, and stress-induced reinstatement of cocaine self-administration in mice Guadalupe Soria & Maria Flavia Barbano & Rafael Maldonado & Olga Valverde Received: 10 March 2008 / Accepted: 21 April 2008 # Springer-Verlag 2008 Abstract Rationale Cocaine addiction is a relapsing psychiatric disorder with a high prevalence in developed countries. To date, the reinstatement model has been difficult to implement in mice. The design of an appropriate reinstate- ment model in mice is required in order to use genetically modified animals with the aim of clarifying the mechanisms involved in cocaine relapse. Objectives Our aim was to develop an appropriate model of reinstatement of cocaine-seeking behavior and to investi- gate the factors that can trigger this reinstatement by using an operant intravenous self-administration procedure in mice. Discrete cues, priming injection of cocaine, and exposure to stress were the stimuli used to reinstate cocaine-seeking behavior. Material and methods Mice were trained to acquire intravenous self-administration of cocaine (1 mg/kg per infusion) on a fixed ratio 1 (FR1) schedule of reinforce- ment. After achieving the acquisition criteria, animals were led to extinguish the operant behavior. Subsequently, under extinction conditions, mice were tested after the adminis- tration of a cocaine priming injection (10 mg/kg i.p.), the presentation of a light cue associated with cocaine administration, or the exposure to a stressful situation (0.21 mA electric footshock). Results Under our experimental conditions the three stimuli successfully reinstated an extinguished cocaine-seeking behavior. Reexposure to cocaine effects by a priming injection was revealed as the strongest stimulus, capable of reinstating cocaine-seeking behavior. Conclusions The effective reinstatement model that we have developed will become a useful tool for future understanding of the neurobiological basis of cocaine addiction and relapse, specifically, with the use of geneti- cally modified mice. Keywords Cocaine . Mice . Self-administration . Operant behavior . Extinction . Reinstatement . Drug seeking . Priming . Environmental cues . Stress Abbreviations i.p. intraperitoneally i.v. intravenous FR fixed ratio CPP conditioned place preference Psychopharmacology DOI 10.1007/s00213-008-1184-x G. Soria : M. F. Barbano : R. Maldonado Laboratorio de Neurofarmacología, Universitat Pompeu Fabra, Parc de Recerca Biomèdica de Barcelona. C/Dr. Aiguader 88, 08003 Barcelona, Spain O. Valverde (*) Grup de Recerca de Neurobiologia del Comportament (GRNC), Departament de Ciències Experimentals i de la Salut, Universitat Pompeu Fabra, Barcelona Biomedical Research Park, C/Doctor Aiguader 88, 08003 Barcelona, Spain e-mail: [email protected] Present address: G. Soria Department of Cerebral Ischemia and Neurodegeneration, Institut d’Investigacions Biomèdiques de Barcelona (CSIC), IDIBAPS, Rosselló, 161, 6th floor, 08036 Barcelona, Spain Present address: M. F. Barbano Instituto de Investigaciones Biotecnológicas, Universidad de General San Martín, INTI - Colectora Av.General Paz 5445, edificio 24, casilla de correos 30, CP 1650 San Martin, Buenos Aires, Argentina

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL INVESTIGATION
A reliable method to study cue-, priming-, and stress-inducedreinstatement of cocaine self-administration in mice
Guadalupe Soria & Maria Flavia Barbano &
Rafael Maldonado & Olga Valverde
Received: 10 March 2008 /Accepted: 21 April 2008# Springer-Verlag 2008
AbstractRationale Cocaine addiction is a relapsing psychiatricdisorder with a high prevalence in developed countries.To date, the reinstatement model has been difficult toimplement in mice. The design of an appropriate reinstate-ment model in mice is required in order to use geneticallymodified animals with the aim of clarifying the mechanismsinvolved in cocaine relapse.Objectives Our aim was to develop an appropriate model ofreinstatement of cocaine-seeking behavior and to investi-gate the factors that can trigger this reinstatement by using
an operant intravenous self-administration procedure inmice. Discrete cues, priming injection of cocaine, andexposure to stress were the stimuli used to reinstatecocaine-seeking behavior.Material and methods Mice were trained to acquireintravenous self-administration of cocaine (1 mg/kg perinfusion) on a fixed ratio 1 (FR1) schedule of reinforce-ment. After achieving the acquisition criteria, animals wereled to extinguish the operant behavior. Subsequently, underextinction conditions, mice were tested after the adminis-tration of a cocaine priming injection (10 mg/kg i.p.), thepresentation of a light cue associated with cocaineadministration, or the exposure to a stressful situation(0.21 mA electric footshock).Results Under our experimental conditions the three stimulisuccessfully reinstated an extinguished cocaine-seekingbehavior. Reexposure to cocaine effects by a priminginjection was revealed as the strongest stimulus, capableof reinstating cocaine-seeking behavior.Conclusions The effective reinstatement model that wehave developed will become a useful tool for futureunderstanding of the neurobiological basis of cocaineaddiction and relapse, specifically, with the use of geneti-cally modified mice.
Keywords Cocaine .Mice . Self-administration .
Operant behavior . Extinction . Reinstatement .
Drug seeking . Priming . Environmental cues . Stress
Abbreviationsi.p. intraperitoneallyi.v. intravenousFR fixed ratioCPP conditioned place preference
PsychopharmacologyDOI 10.1007/s00213-008-1184-x
G. Soria :M. F. Barbano :R. MaldonadoLaboratorio de Neurofarmacología, Universitat Pompeu Fabra,Parc de Recerca Biomèdica de Barcelona. C/Dr. Aiguader 88,08003 Barcelona, Spain
O. Valverde (*)Grup de Recerca de Neurobiologia del Comportament (GRNC),Departament de Ciències Experimentals i de la Salut,Universitat Pompeu Fabra,Barcelona Biomedical Research Park, C/Doctor Aiguader 88,08003 Barcelona, Spaine-mail: [email protected]
Present address:G. SoriaDepartment of Cerebral Ischemia and Neurodegeneration,Institut d’Investigacions Biomèdiques de Barcelona (CSIC),IDIBAPS,Rosselló, 161, 6th floor,08036 Barcelona, Spain
Present address:M. F. BarbanoInstituto de Investigaciones Biotecnológicas,Universidad de General San Martín,INTI - Colectora Av.General Paz 5445, edificio 24,casilla de correos 30,CP 1650 San Martin, Buenos Aires, Argentina

Introduction
Drug addiction is a chronic psychiatric disease character-ized by compulsive drug seeking and consumption fol-lowed by periods of abstinence and episodes of intensedrug craving which lead to high rates of relapse (DSM-IVTR 2004). The repeated cycles of cessation and relapseremain the major clinical problem in treating drug addiction(Miller and Gold 1994; Weiss et al. 2001). Cocaine is oneof the most widely used psychostimulant drugs of abuse.Thus, the prevalence of cocaine use in the European Unionis 12 million people, and it is estimated that 4.5 millionEuropeans currently consume cocaine (EMCDDA 2005;Watson 2007; Wiessing 2005). Cocaine produces euphoriceffects, increases motor activity (Woolverton and Johnson1992), and enhances dopaminergic activity by blockingdopamine reuptake by the binding of the dopaminetransporter (Rothman and Baumann 2003). In humans,relapse to cocaine and heroin abuse or the intense desire forthese drugs (craving) can be generated by reexposure to theconsumed drug, by stimuli usually associated with drugintake, and by stress (see Bossert et al. 2005 for a review).
Whereas acute drug reinforcing properties are essentialfor the initial phases of the addiction, other complexbehavioral processes are crucial for the consolidation ofthis chronic relapsing disorder (Koob and LeMoal 2001).Operant intravenous drug self-administration is the mostcomplete and reliable approach to investigate the addictivepotential of drugs in rodents. Several aspects of theaddiction process can be dissected by using this paradigm,including acquisition of an operant behavior to obtain thedrug, maintenance, extinction, and reinstatement of such abehavior. Most studies conducted in mice to evaluateextinction and reinstatement of drug-seeking behavior havebeen performed in the conditioned place preference (CPP)paradigm (Tzschentke 2007) or in operant models usingmonkeys and rats (Weiss 2005; Shaham et al. 2003; Le Folland Goldberg 2005). However, only three studies havemodeled cocaine reinstatement after extinction in mice(Highfield et al. 2002; Fuchs et al. 2003; Kruzich 2007).Furthermore, most of these studies investigated the potentialof cocaine priming administration and environmental cues toproduce reinstatement of an extinguished drug-seekingbehavior, whereas the exposure to stress was investigatedto a lesser extent. However, stressful conditions are the mostcommon situations precipitating reinstatement of an addic-tive behavior in humans (Bossert et al. 2005).
The lack of validated operant models of cocainereinstatement in mice is mainly due to the complexity ofthe experimental technique, namely, the surgical procedurein these animal species (Epstein et al. 2006). During the lastdecade, the use of genetically modified mice has led toimportant advances in the knowledge of the neurobiological
substrates underlying drug addiction. The existence of thesemutant animals represents a relevant tool to unveil theneurobiological mechanisms involved in relapse, so thedevelopment of a reliable drug reinstatement model in micebecomes imperative.
In the present study, we investigated the reinstatement ofa previously extinguished cocaine-seeking behavior byusing an operant self-administration procedure in outbredmice. The reinstatement of drug seeking was evaluatedunder three different experimental conditions: (1) presenta-tion of a visual cue associated with drug delivery, (2)priming injection of cocaine administered at the home cage,and (3) exposure to a stressful situation (an electricfootshock). Additionally, we describe and characterize herefor the first time the acquisition and extinction of cocaineself-administration in the case of outbred mice.
Materials and methods
Animals
Male CD1 outbred mice (Charles River, France) were used inthis study. The animals (weighing 20–25 g upon arrival at thelaboratory) were individually housed in clear plastic cages in ananimal vivarium maintained on a 12-h light-dark cycle (lightson at 0800 hours), at constant temperature (21±1°C) andhumidity (55±10%). Food andwater were provided ad libitum,except during experimental sessions. Mice were handled dailyin order to habituate them and minimize handling stress duringthe experiments. Animal care and experimental procedureswere in strict accordance with institutional and internationalstandards (the European Communities Council Directive 86/609/EEC, 24 November 1986) and were approved by the localEthics Committee (CEEA-PRBB).
Drugs
Cocaine hydrochloride was obtained from Ministerio deSanidad y Consumo (Spain) and prepared in sterile 0.9%physiological saline. We used a dose of 1 mg/kg perinfusion during the training phase of the self-administrationexperiments and 10 mg/kg intraperitoneally (i.p.) for thecocaine priming injection.
Operant cocaine self-administration
Apparatus Self-administration training and testing occurredin operant chambers (Model ENV-307A-CT, MED Asso-ciates, Inc., Georgia, VT, USA) equipped with tworetractable levers. One of them was selected as thereinforced lever for delivering the drug and the other asthe non-reinforced lever. Active pressing on the reinforced
Psychopharmacology

lever resulted in a cocaine infusion while pressing on thenon-reinforced lever had no consequences. Chambers weremade of aluminum and clear acrylic, had grid floorsconnected to an electrical shocker (ENV-414, MED Asso-ciates, Inc., St. Albans, VT, USA), and were housed insound- and light-attenuated boxes equipped with fans toprovide ventilation and ambient noise. A stimulus light,located above the reinforced lever, was paired contingentlywith the delivery of the drug. When mice responded on thereinforced lever, the chamber light went off, the stimuluslight went on, and a drug infusion was delivered. Cocainewas infused via a syringe that was mounted on a micro-infusion pump (PHM-100A, MED Associates, Inc., Georgia,VT, USA) and connected via Tygon tubing (0.96 mm o.d.,Portex Fine Bore Polythene Tubing, Portex Limited, Hythe,Kent, UK) to a single channel liquid swivel (375/25, InstechLaboratories, Plymouth Meeting, PA, USA) and to themouse intravenous (i.v.) catheters. The swivel was mountedon a counterbalanced arm above the operant chamber.
Surgery
Mice were anesthetized under isoflurane anesthesia (1.5–2.0%) and then implanted with indwelling intravenous silasticcatheters (Soria et al. 2005). Briefly, a 6-cm length of silastictubing (0.3 mm inner diameter, 0.6 mm outer diameter)(Silastic®, Dow Corning, Houdeng-Goegnies, Belgium) wasfitted to a 22-gauge steel cannula (Semat, Herts, UK) thatwas bent at a right angle and then embedded in a cement disk(Dentalon Plus, Heraeus Kulzer, Wehrheim, Germany) withan underlying nylon mesh. The catheter tubing was inserted1.3 cm into the right jugular vein and anchored with suture.The remaining tubing ran subcutaneously to the cannula,which exited at the midscapular region. All incisions weresutured and coated with antibiotic ointment (Bactroban,GlaxoSmithKline, Madrid, Spain). After surgery, animalswere allowed to recover for 3 days prior to initiation of self-administration sessions. The catheter was flushed daily with asaline solution. The patency of intravenous catheters wasevaluated periodically (approximately every 5–6 days) andwhenever drug self-administration behavior appeared todeviate dramatically from that observed previously byinfusion of 0.1 ml thiobarbital (5 mg/ml) through the catheter.If prominent signs of anesthesia were not apparent within 3 sof the infusion, the mouse was removed from the experiment.
Procedure
Cocaine self-administration training
Cocaine self-administration sessions were performed aspreviously described (Soria et al. 2005). Responding was
maintained by cocaine (1 mg/kg per injection) delivered in58.75 μl over 4 s. Daily self-administration started with apriming injection of the drug, lasted for 120 min, and wasconducted 7 days per week. The house light was on at thebeginning of the session for 3 s and off during theremaining time of the session. Each daily session startedwith the presentation of the reinforced and non-reinforcedlevers, a priming injection of the drug. and a 4-spresentation of the light cue (located above the reinforcedlever). Presses on the reinforced lever led to cocaineinfusions and the presentation of the light cue for 4 s. Micewere trained to lever press for cocaine under a fixed ratio 1(FR1) schedule of reinforcement. A 30-s time-out periodwas established after obtainment of each reinforcement.During this 30-s period, the cue light was off and no druginfusions were provided after pressing the reinforced lever.Non-reinforced lever presses and all the responses per-formed during the 30-s time-out period were also recorded.The session was terminated after 75 reinforcements weredelivered or after 2 h, whichever occurred first. The criteriafor the acquisition was achieved when mice maintained astable responding with less than 20% deviation from themean of the total number of cocaine infusions earned inthree consecutive sessions (80% of stability), with at least75% responding on the reinforced lever, and a minimum often reinforcements per session. Considering that relapsewas evaluated in a second step, only those animals showinga reliable drug self-administration behavior continued theexperiment. The percentage of animals which achieved theacquisition criteria was 78.13%. Once the acquisitioncriteria were achieved, the extinction period started.
Extinction and reinstatement
The extinction procedure was modified from Highfield etal. (2002). In this phase, pressing the reinforced leverresulted in the activation of the pump to maintain the usualexperimental environment but animals did not receive druginfusions. Also, the stimulus light was off during thisperiod. Mice were given two 2-h daily extinction sessionsseparated by 1 h in which the levers were retracted (Fig. 1).Extinction sessions were conducted 6 days per week untilreaching the extinction criteria. These criteria wereachieved when, in the first 2 h of the extinction session,mice made a mean number of responses in two consecutiveextinction sessions of less than 30% of the responsesperformed during the last day of the cocaine training phase.A small percentage of animals never achieved the extinc-tion criteria (10%) and were excluded from the followingexperiments. The following day after achieving the extinc-tion criteria, three different experimental conditions wereevaluated to induce reinstatement of the cocaine self-administration behavior: the presentation of a conditioned
Psychopharmacology

discrete cue (light), a priming injection of cocaine (admin-istered i.p.), and exposure to a stressful situation (electricfootshock). We considered that an animal reliably relapsedwhen both of the following two conditions were met duringthe reinstatement test: (1) it doubled the number ofresponses on the reinforced lever with regard to extinctionand (2) it performed at least ten responses on the reinforcedlever.
Cue-induced reinstatement The conditions during the testday were the same as the training sessions except that nococaine was available. Indeed, the session started with a 5-spresentation of the light cue and subsequent presses on thereinforced lever led also to the presentation of the light cuefor 5 s.
Cocaine-induced reinstatement Animals received an injec-tion of cocaine (10 mg/kg i.p.) at their home cage andimmediately after were confined to the self-administrationboxes to start the reinstatement test. No cue light wasassociated with the reinforced lever pressing to avoid apossible context bias in the procedure. The remainingconditions during the test day were the same as during thetraining sessions except that no cocaine was available. Acontrol group of five animals received an i.p. injection ofsaline on the day before the cocaine-induced reinstatementtest and immediately after were confined to the self-administration box following the same procedure as forcocaine-injected animals. This control group makes itpossible to ensure that cocaine priming-induced relapsewas specific to cocaine administration and not triggered byother factors such as the injection procedure.
Stress-induced reinstatement Animals received a 2-s foot-shock (0.21 mA) every min during a 5-min period (totalfootshocks: 5) and immediately after levers were presentedin the self-administration boxes to start the reinstatementtest during a 2-h session. As in the case of cocaine-inducedreinstatement, no cue light or cocaine were presented afterpressing the reinforced lever.
Statistical analysis
The results were analyzed using a multivariate analysis ofvariance (MANOVA), with time (days) and lever (rein-forced vs non-reinforced) as the within-subjects factors,when analyzing acquisition and extinction of the self-administration behavior. In the case of reinstatementexperiments, phase of the experiment (last day of cocaine,first day of extinction, mean of extinction and relapse) aswell as lever (reinforced vs non-reinforced) were thewithin-subjects factors. In all of the cases, the dependentvariable was the number of lever presses accomplished in a2-h period. When significant overall interactions werefound, further analyses of partial interactions were carriedout. Post hoc analyses were performed with Fisher’s LSDtest when the initial p value was significant. All data wereanalyzed with Statistica software (StatSoft Inc., France). Aresult was considered significant if p<0.05. All the resultsare expressed as mean ± SEM.
Results
Acquisition and extinction of cocaine self-administrationbehavior in CD1 outbred mice
Three different groups of mice were trained to self-administer cocaine, one for each experimental conditionfor reinstatement. An analysis of variance among thesegroups was performed to evaluate any possible differenceduring the acquisition phase. The main effect of group wasnot significant (F(2,28)=0.46, p=0.636), and the results ofacquisition were then pooled for the three experimentalgroups (Fig. 2a). During the training period, mice readilylearned to discriminate between reinforced and non-reinforced levers, as revealed by the main effect of lever(F(1,30)=48.91, p<0.001). A significant interaction betweentime and lever (F(7,210)=3.76, p<0.001) indicated thatanimals increased the number of responses on thereinforced lever along the time. The mean acquisition timewas 8.31±0.52 days, and the mean drug intake during thelast 3 days of training was 24.87±1.43 mg/kg of cocaine.
During the extinction phase, mice were given two 2-h dailyextinction sessions separated by 1 h in which the levers were
Fig. 1 Different phases of the experimental design employed to testreinstatement of cocaine-seeking behavior. As described in the“Materials and methods” section, cocaine training sessions wereconducted during 2 h. After reaching the acquisition criteria, micestarted the extinction phase. The conditions during the extinctionphase were the same as in the training phase, with the exception thatcocaine was not available and no light cue was paired with thereinforced lever. Mice were given two 2-h daily extinction sessionsseparated by 1 h in which the levers were retracted. Once micereached the extinction criteria, the reinstatement of cocaine-seekingbehavior was tested (elicited by cue, cocaine priming, and electricfootshock). Comparisons of the number of responses were performedbetween the last day of cocaine self-administration, the mean of thefirst 2 h of the days in which animals reached the extinction criteria,and the test for reinstatement
Psychopharmacology
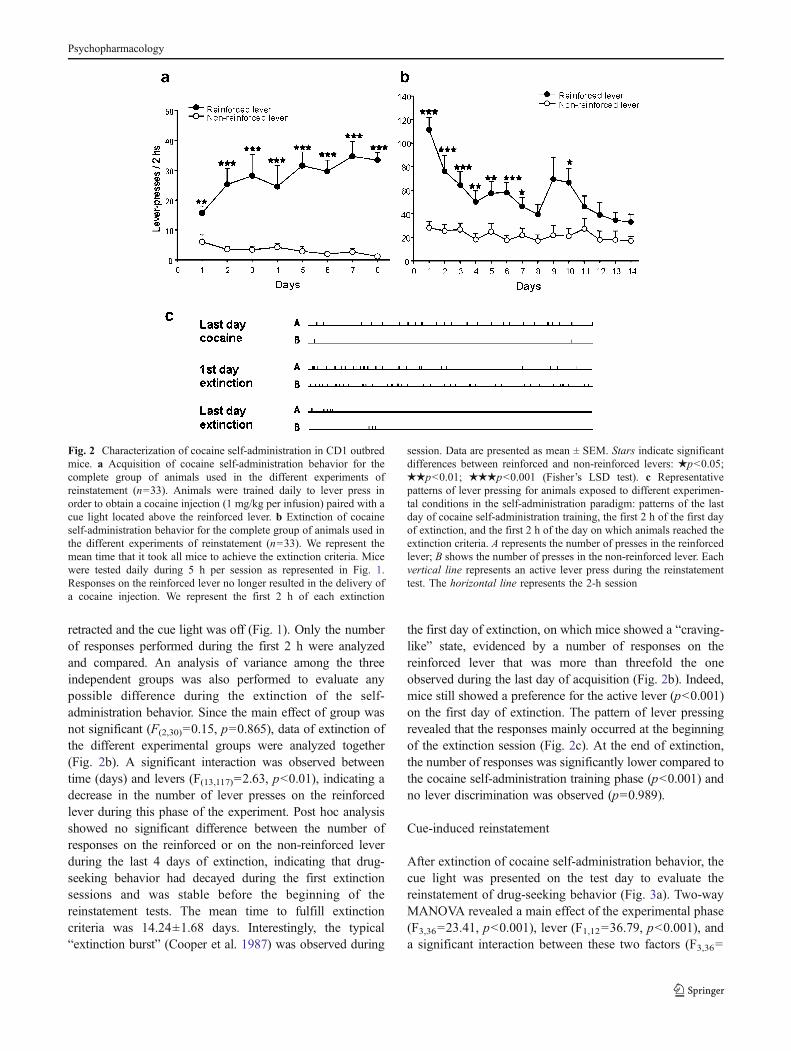
retracted and the cue light was off (Fig. 1). Only the numberof responses performed during the first 2 h were analyzedand compared. An analysis of variance among the threeindependent groups was also performed to evaluate anypossible difference during the extinction of the self-administration behavior. Since the main effect of group wasnot significant (F(2,30)=0.15, p=0.865), data of extinction ofthe different experimental groups were analyzed together(Fig. 2b). A significant interaction was observed betweentime (days) and levers (F(13,117)=2.63, p<0.01), indicating adecrease in the number of lever presses on the reinforcedlever during this phase of the experiment. Post hoc analysisshowed no significant difference between the number ofresponses on the reinforced or on the non-reinforced leverduring the last 4 days of extinction, indicating that drug-seeking behavior had decayed during the first extinctionsessions and was stable before the beginning of thereinstatement tests. The mean time to fulfill extinctioncriteria was 14.24±1.68 days. Interestingly, the typical“extinction burst” (Cooper et al. 1987) was observed during
the first day of extinction, on which mice showed a “craving-like” state, evidenced by a number of responses on thereinforced lever that was more than threefold the oneobserved during the last day of acquisition (Fig. 2b). Indeed,mice still showed a preference for the active lever (p<0.001)on the first day of extinction. The pattern of lever pressingrevealed that the responses mainly occurred at the beginningof the extinction session (Fig. 2c). At the end of extinction,the number of responses was significantly lower compared tothe cocaine self-administration training phase (p<0.001) andno lever discrimination was observed (p=0.989).
Cue-induced reinstatement
After extinction of cocaine self-administration behavior, thecue light was presented on the test day to evaluate thereinstatement of drug-seeking behavior (Fig. 3a). Two-wayMANOVA revealed a main effect of the experimental phase(F3,36=23.41, p<0.001), lever (F1,12=36.79, p<0.001), anda significant interaction between these two factors (F3,36=
Fig. 2 Characterization of cocaine self-administration in CD1 outbredmice. a Acquisition of cocaine self-administration behavior for thecomplete group of animals used in the different experiments ofreinstatement (n=33). Animals were trained daily to lever press inorder to obtain a cocaine injection (1 mg/kg per infusion) paired with acue light located above the reinforced lever. b Extinction of cocaineself-administration behavior for the complete group of animals used inthe different experiments of reinstatement (n=33). We represent themean time that it took all mice to achieve the extinction criteria. Micewere tested daily during 5 h per session as represented in Fig. 1.Responses on the reinforced lever no longer resulted in the delivery ofa cocaine injection. We represent the first 2 h of each extinction
session. Data are presented as mean ± SEM. Stars indicate significantdifferences between reinforced and non-reinforced levers: ★p<0.05;★★p<0.01; ★★★p<0.001 (Fisher’s LSD test). c Representativepatterns of lever pressing for animals exposed to different experimen-tal conditions in the self-administration paradigm: patterns of the lastday of cocaine self-administration training, the first 2 h of the first dayof extinction, and the first 2 h of the day on which animals reached theextinction criteria. A represents the number of presses in the reinforcedlever; B shows the number of presses in the non-reinforced lever. Eachvertical line represents an active lever press during the reinstatementtest. The horizontal line represents the 2-h session
Psychopharmacology
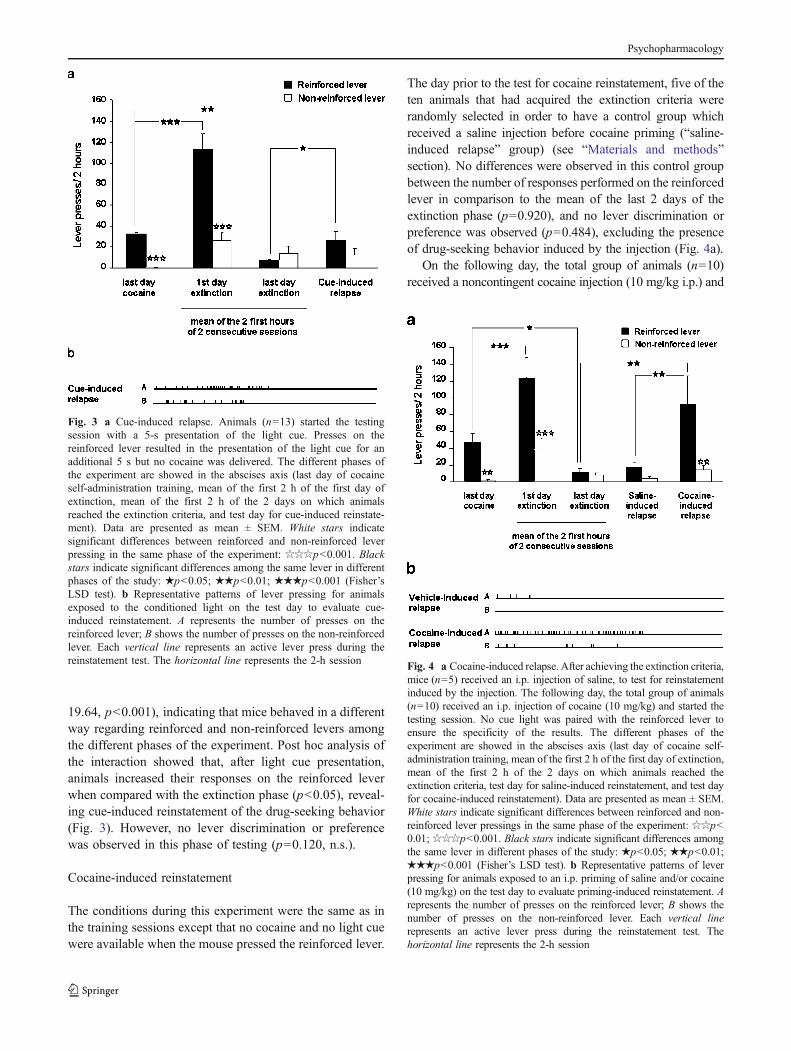
19.64, p<0.001), indicating that mice behaved in a differentway regarding reinforced and non-reinforced levers amongthe different phases of the experiment. Post hoc analysis ofthe interaction showed that, after light cue presentation,animals increased their responses on the reinforced leverwhen compared with the extinction phase (p<0.05), reveal-ing cue-induced reinstatement of the drug-seeking behavior(Fig. 3). However, no lever discrimination or preferencewas observed in this phase of testing (p=0.120, n.s.).
Cocaine-induced reinstatement
The conditions during this experiment were the same as inthe training sessions except that no cocaine and no light cuewere available when the mouse pressed the reinforced lever.
The day prior to the test for cocaine reinstatement, five of theten animals that had acquired the extinction criteria wererandomly selected in order to have a control group whichreceived a saline injection before cocaine priming (“saline-induced relapse” group) (see “Materials and methods”section). No differences were observed in this control groupbetween the number of responses performed on the reinforcedlever in comparison to the mean of the last 2 days of theextinction phase (p=0.920), and no lever discrimination orpreference was observed (p=0.484), excluding the presenceof drug-seeking behavior induced by the injection (Fig. 4a).
On the following day, the total group of animals (n=10)received a noncontingent cocaine injection (10 mg/kg i.p.) and
Fig. 3 a Cue-induced relapse. Animals (n=13) started the testingsession with a 5-s presentation of the light cue. Presses on thereinforced lever resulted in the presentation of the light cue for anadditional 5 s but no cocaine was delivered. The different phases ofthe experiment are showed in the abscises axis (last day of cocaineself-administration training, mean of the first 2 h of the first day ofextinction, mean of the first 2 h of the 2 days on which animalsreached the extinction criteria, and test day for cue-induced reinstate-ment). Data are presented as mean ± SEM. White stars indicatesignificant differences between reinforced and non-reinforced leverpressing in the same phase of the experiment: ☆☆☆p<0.001. Blackstars indicate significant differences among the same lever in differentphases of the study: ★p<0.05; ★★p<0.01; ★★★p<0.001 (Fisher’sLSD test). b Representative patterns of lever pressing for animalsexposed to the conditioned light on the test day to evaluate cue-induced reinstatement. A represents the number of presses on thereinforced lever; B shows the number of presses on the non-reinforcedlever. Each vertical line represents an active lever press during thereinstatement test. The horizontal line represents the 2-h session Fig. 4 a Cocaine-induced relapse. After achieving the extinction criteria,
mice (n=5) received an i.p. injection of saline, to test for reinstatementinduced by the injection. The following day, the total group of animals(n=10) received an i.p. injection of cocaine (10 mg/kg) and started thetesting session. No cue light was paired with the reinforced lever toensure the specificity of the results. The different phases of theexperiment are showed in the abscises axis (last day of cocaine self-administration training, mean of the first 2 h of the first day of extinction,mean of the first 2 h of the 2 days on which animals reached theextinction criteria, test day for saline-induced reinstatement, and test dayfor cocaine-induced reinstatement). Data are presented as mean ± SEM.White stars indicate significant differences between reinforced and non-reinforced lever pressings in the same phase of the experiment: ☆☆p<0.01; ☆☆☆p<0.001. Black stars indicate significant differences amongthe same lever in different phases of the study: ★p<0.05; ★★p<0.01;★★★p<0.001 (Fisher’s LSD test). b Representative patterns of leverpressing for animals exposed to an i.p. priming of saline and/or cocaine(10 mg/kg) on the test day to evaluate priming-induced reinstatement. Arepresents the number of presses on the reinforced lever; B shows thenumber of presses on the non-reinforced lever. Each vertical linerepresents an active lever press during the reinstatement test. Thehorizontal line represents the 2-h session
Psychopharmacology
immediately after were confined to the self-administrationboxes for the reinstatement test, under the same conditionsas previously described for the “saline-induced relapse”group (Fig. 4a). As in the case of cue-induced reinstate-ment, two-way MANOVA revealed a main effect of thephase of the experiment (F3,24=8.06, p<0.001) and lever(F1,8=14.77, p<0.01) as well as a significant interactionbetween these two factors (F3,24=4.15, p<0.05), indicatingthat mice behaved differentially with regard to the reinforcedand the non-reinforced levers among the different phases ofthe experiment. Post hoc analysis showed that cocaineadministration induced an increase in the responses on thereinforced lever when compared with responses duringthe last day of extinction. Besides, a preference for thereinforced lever was observed (p<0.01) during cocainetesting, revealing altogether that cocaine priming induced areliable reinstatement of drug-seeking behavior.
Stress-induced reinstatement
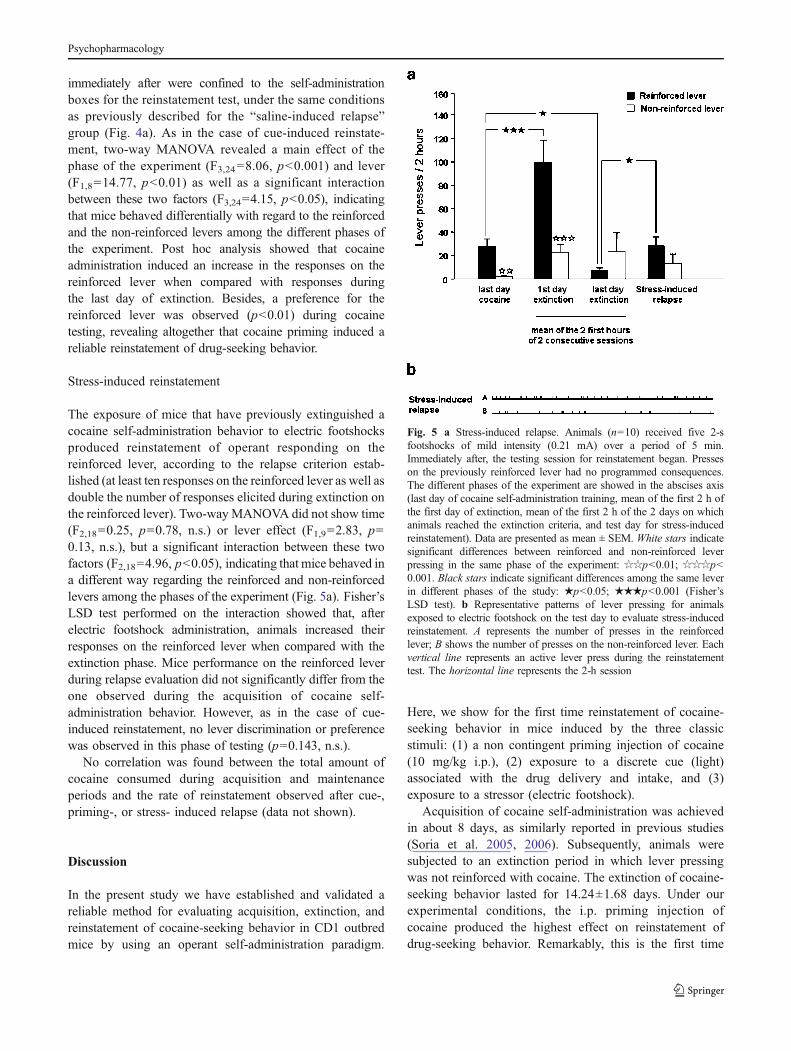
The exposure of mice that have previously extinguished acocaine self-administration behavior to electric footshocksproduced reinstatement of operant responding on thereinforced lever, according to the relapse criterion estab-lished (at least ten responses on the reinforced lever as well asdouble the number of responses elicited during extinction onthe reinforced lever). Two-wayMANOVA did not show time(F2,18=0.25, p=0.78, n.s.) or lever effect (F1,9=2.83, p=0.13, n.s.), but a significant interaction between these twofactors (F2,18=4.96, p<0.05), indicating that mice behaved ina different way regarding the reinforced and non-reinforcedlevers among the phases of the experiment (Fig. 5a). Fisher’sLSD test performed on the interaction showed that, afterelectric footshock administration, animals increased theirresponses on the reinforced lever when compared with theextinction phase. Mice performance on the reinforced leverduring relapse evaluation did not significantly differ from theone observed during the acquisition of cocaine self-administration behavior. However, as in the case of cue-induced reinstatement, no lever discrimination or preferencewas observed in this phase of testing (p=0.143, n.s.).
No correlation was found between the total amount ofcocaine consumed during acquisition and maintenanceperiods and the rate of reinstatement observed after cue-,priming-, or stress- induced relapse (data not shown).
Discussion
In the present study we have established and validated areliable method for evaluating acquisition, extinction, andreinstatement of cocaine-seeking behavior in CD1 outbredmice by using an operant self-administration paradigm.
Here, we show for the first time reinstatement of cocaine-seeking behavior in mice induced by the three classicstimuli: (1) a non contingent priming injection of cocaine(10 mg/kg i.p.), (2) exposure to a discrete cue (light)associated with the drug delivery and intake, and (3)exposure to a stressor (electric footshock).
Acquisition of cocaine self-administration was achievedin about 8 days, as similarly reported in previous studies(Soria et al. 2005, 2006). Subsequently, animals weresubjected to an extinction period in which lever pressingwas not reinforced with cocaine. The extinction of cocaine-seeking behavior lasted for 14.24±1.68 days. Under ourexperimental conditions, the i.p. priming injection ofcocaine produced the highest effect on reinstatement ofdrug-seeking behavior. Remarkably, this is the first time
Fig. 5 a Stress-induced relapse. Animals (n=10) received five 2-sfootshocks of mild intensity (0.21 mA) over a period of 5 min.Immediately after, the testing session for reinstatement began. Presseson the previously reinforced lever had no programmed consequences.The different phases of the experiment are showed in the abscises axis(last day of cocaine self-administration training, mean of the first 2 h ofthe first day of extinction, mean of the first 2 h of the 2 days on whichanimals reached the extinction criteria, and test day for stress-inducedreinstatement). Data are presented as mean ± SEM. White stars indicatesignificant differences between reinforced and non-reinforced leverpressing in the same phase of the experiment: ☆☆p<0.01; ☆☆☆p<0.001. Black stars indicate significant differences among the same leverin different phases of the study: ★p<0.05; ★★★p<0.001 (Fisher’sLSD test). b Representative patterns of lever pressing for animalsexposed to electric footshock on the test day to evaluate stress-inducedreinstatement. A represents the number of presses in the reinforcedlever; B shows the number of presses on the non-reinforced lever. Eachvertical line represents an active lever press during the reinstatementtest. The horizontal line represents the 2-h session
Psychopharmacology

that a cocaine i.p. priming injection triggers the reinstate-ment of an operant responding for cocaine in mice.Moreover, an important discriminative effect between thereinforced and the non-reinforced lever was observed forcocaine-induced reinstatement. The dose of cocaine prim-ing (10 mg/kg i.p.) was chosen based on previous studiesevaluating cocaine rewarding effects (Martin et al. 2000). Inaddition, this dose is known to increase the extracellularlevels of dopamine in the mice nucleus accumbens (Soria etal. 2005). These data support the hypothesis that ourobservations involving cocaine priming were due to thepsychotropic effects of the drug. A possible effect of stressinduced by the injection procedure was ruled out by the factthat animals receiving a saline injection did not reinstatecocaine-seeking behavior. A limitation of our experimentalprocedure is that mice also were primed with an infusion ofcocaine (1 mg/kg) during training, in order to facilitate theacquisition of the self-administration behavior. Recently, ithas been shown that cocaine may act both as a discrimi-native stimulus as well as a reinforcer in the reinstatementparadigm currently used (Shalev et al. 2002; Keiflin et al.2008). Under our experimental conditions, we were notable to dissociate the portion of reinstatement induced bythe discriminative properties of cocaine from the oneinduced by its incentive motivational properties. Neverthe-less, now that we have validated this model of reinstate-ment in mice, further investigation aimed at dissociating thecontribution of the discriminative and motivational proper-ties of cocaine in reinstatement is warranted.
Another important finding was that both the light cueand, for the first time, a footshock stressor reinstatedcocaine-seeking behavior in CD1 outbred mice. In bothcases the number of lever pressings during the relapsetesting day was significantly increased specifically on thereinforced lever. However, no discrimination was foundbetween the reinforced and non-reinforced levers whenstress or discrete cues were tested to induce reinstatement.
Intravenous self-administration is probably the mostreliable approach to investigate the abuse potential of drugsin animals (Collins et al. 1984). The technical complexityof this method is the reason why the majority of thesestudies have been performed in rats (Shaham et al. 2000;Weiss et al. 2000; De Vries et al. 2001). As a result, micestudies testing reinstatement of drug seeking have currentlyemployed the CPP paradigm, which is based on classicconditioning (Daza-Losada et al. 2007; Tzschentke 2007).Several authors have demonstrated cocaine relapse after awithdrawal period using the CPP paradigm (Itzhak andMartin 2002; Graham et al. 2007; Ribeiro do Couto et al.2005). Nonetheless, only three studies have tried tocharacterize the reinstatement of cocaine-seeking behaviorin mice after a period of extinction by using the i.v. self-administration paradigm. First, Highfield et al. (2002)
showed reinstatement of cocaine seeking in 129X1/SvJmice. Several discrepancies exist between our study andthat conducted by Highfield et al. (2002); whereas 129/SvJmice exhibited a modest effect of cocaine priming onreinstatement of cocaine-seeking behavior, in our studycocaine priming was the most significant condition toreinstate drug seeking. Under their experimental conditions,the cocaine dose and the route of administration weredifferent in comparison to our experiment (6 mg/kg i.v.versus 10 mg/kg i.p.). It should be noted that the 129/SvJstrain is known to be less sensitive to cocaine-inducedbehavioral responses than CD1 outbred or C57BL/6Jstrains (Schlussman et al. 1998; Miner 1997). All theabove-mentioned factors could account for the modesteffect of cocaine-induced relapse observed in 129/SvJanimals. It is important to note that the 129/SvJ mouse isnot the most suitable strain to investigate a complexphenomenon such as drug addiction since several cognitivedeficits have been described in this particular strain(Phillips et al. 1999; Crawley 2003; Bailey et al. 2006).
Fuchs et al. (2003) also investigated cocaine reinstate-ment in mice after extinction and showed that conditionedstimuli associated with cocaine administration reinstateddrug-seeking behavior in C57BL/6J mice. The meannumber of responses during training and testing was muchhigher than the one observed under our experimentalconditions. However, Fuchs and colleagues (2003) trainedtheir animals during the dark period of the circadian cycle,the active period for most rodents. Also, animals weretrained to self-administer sucrose during an overnightsession of 16 h before the intravenous surgery. Therefore,both the circadian moment and the previous training historymay account for the differences observed in the perfor-mance of mice in both studies. On the other hand, theprevious work failed to demonstrate cocaine-inducedreinstatement of responding when cocaine (doses rangingfrom 1 to 40 mg/kg) was delivered i.p. This disparity withour results could be explained by the fact that Fuchs et al.(2003) exposed the animals to changes in the food regimeduring the different phases of the study. In our case, thisfactor is not present since animals were trained directly toself-administer cocaine in a satiated state. Moreover, themice strain used in the above-cited study was C57BL/6J,whereas here, for the first time, we have used a CD1outbred background mouse to study cocaine reinstatementbehavior. CD1 mice are a common outbred albino stock,and the outbred mating strategy makes it likely that thesemice will have more variable genes (alleles), phenotype,and, possibly, more variable behavior than the inbredstrains (Festing 1999). Thus, the use of CD1 mice may beadvantageous, since outbred mice are considered to reflectmore natural heterogeneous populations (Adams et al.2002). In addition, considering the present results, CD1
Psychopharmacology

seems to be the most suitable strain for relapse studies inmice, since all three experimental conditions evaluatedproduced a reliable reinstatement of the operant behavior.
Recently, Kruzich (2007) reported an operant method forcocaine craving and relapse in mice. The author performedconditioned- and cocaine-primed reinstatement in the sameC57BL/6J mice subjected to a food restriction regime. Thisauthor used a complex experimental design where micereceived cocaine in a contingent way inside the operantchamber, whereas we administered a non contingent i.p.injection of cocaine outside the operant chamber. Underthe experimental conditions reported by Kruzich (2007), itwas not possible to ensure that the observed reinstatementof cocaine-seeking behavior was only due to the effects ofcocaine per se or both to its effects and the presence ofenvironmental visual cues associated with drug delivery.
In contrast to our data, the three studies discussed above(Highfield et al. 2002; Fuchs et al. 2003; Kruzich 2007)demonstrated a significant discrimination between activeand inactive lever responses on the cue-induced reinstate-ment test. The discrepancy could be due to the fact that theyused compound conditioned stimuli consisting in a lightplaced above the reinforced lever plus a tone generator for2 s, whereas we only used the cue light situated above thereinforced lever. Their conditions would result in a strongerincentive value attributed to the conditioned stimuli and, asa consequence, to a stronger effect on the reinstatement ofcocaine seeking when presenting the compound stimulus.
In the case of reinstatement induced by electric stimulation,the use of a cocaine-paired cue during extinction andreinstatement phases is known to be critical for shock-inducedreinstatement in rats (Shelton and Beardsley 2005). Therefore,the absence of such a cue during extinction and reinstatementphases could account for the relatively weak effect of shockobserved in stress-induced reinstatement in our study.
Human and animal studies have suggested that stress isone of the most important factors to trigger relapse ofaddictive behaviors after a period of abstinence (Kita et al.1999; Todd 2004). Indeed, stress was recently revealed as acrucial factor in producing relapse and is strongly depen-dent on the individual genetically determined emotionality(Bilkei-Gorzo et al. 2008; LeDoux 2000). Footshock stressis a general method to induce reinstatement of drug-seekingbehavior in experimental animals (Weiss 2005). Moreover,stress facilitates the acquisition of psychostimulant self-administration in operant paradigms (Goeders and Guerin1994; Piazza et al. 1990). In a very recent study, nicotine-seeking behavior was reinstated after a brief exposure tofootshock in mice presenting high responsivity to stress, butnot in those presenting low responsivity (Bilkei-Gorzo et al.2008). Several studies have also used food deprivation as astressor in mice (Highfield et al. 2002) and rats (Shalev etal. 2001; De Vry et al. 1989) to reinstate cocaine- and
heroin-seeking behavior. However, inconsistent results havebeen found when evaluating the reinstatement of operantresponses using food deprivation, probably due to theparticipation of additional metabolic factors. As alreadystated, under our experimental conditions the lack ofmetabolic state manipulations of the organism allows usto attribute the results specifically to stress.
A body of evidence shows that the neuronal mechanismsunderlying stress-induced drug seeking, cues, or primingare not identical. Cocaine-induced reinstatement resultsfrom activation of the ventral tegmental area-dorsal pre-frontal cortex dopamine pathway, which is responsible forthe activation of the dorsal prefrontal cortex-nucleusaccumbens core glutamatergic pathway, with a subsequentactivation of the accumbens core-ventral pallidum pathway,presumably via a γ-aminobutyric acid (GABA) pathway(Kalivas and McFarland 2003; Maier and Watkins 2005).Several studies have proposed the role of both basolateraland central amygdala in discrete cue-induced reinstatementwhere the reexposure to the cue is earned contingently byresponding on the drug-associated lever (Di Ciano andEveritt 2004; Bossert et al. 2005; Davis et al. 2006).Concerning stress, recent neuroanatomical studies indicatethat stress-induced activation of the mesocorticolimbicdopamine and glutamatergic system is critically involvedin the reinstatement of drug-seeking responses (Piazza andLe Moal 1996; McFarland et al. 2004).
In conclusion, we have characterized and validated for thefirst time a model in CD1 outbred mice to evaluateacquisition, extinction, and reinstatement of cocaine-seekingbehavior by using the self-administration paradigm. Takinginto account the elevated relapse rates observed in cocaineaddiction, this method is of high interest to investigate theneurobiological basis involved in this phenomenon, by usinggenetically modified mice. A better understanding of theneural and molecular events underlying relapse will becrucial to design more effective therapeutic actions and toestablish convenient preventive policies.
Acknowledgments The authors wish to thank S.A. Bura and Dr. B.Rubi for critical reading of the manuscript. This study was supportedby grants from Spanish MCYT (SAF2004/0568; SAF2007/60249)and MSC (Plan Nacional sobre Drogodependencias, conv, 2006) to O.V., and (SAF2007-64062) to R.M., and European Communities(Genaddict LSHMCT-2004-005166; Phecomp LSHMCT-2006-037669) to R.M. G.S. was supported by a fellowship from SpanishMCYT associated with the SAF2004/0568 grant. M.F.B. wassupported by a postdoctoral fellowship from the Fyssen Foundation.
References
Adams B, Fitch T, Chaney S, Gerlai R (2002) Altered performancecharacteristics in cognitive tasks: comparison of the albino ICRand CD1 mouse strains. Behav Brain Res 133:351–361
Psychopharmacology

Bailey KR, Rustay NR, Crawley JN (2006) Behavioral phenotypingof transgenic and knockout mice: practical concerns and potentialpitfalls. ILAR J 47:124–131
Bilkei-Gorzo A, Rácz I, Michel K, Darvas M, Maldonado R, ZimmerA (2008) A common genetic predisposition to stress sensitivityand stress-induced nicotine craving. Biol Psychiatry 63:164–171
Bossert JM, Ghitza UE, Lu L, Epstein DH, Shaham Y (2005)Neurobiology of relapse to heroin and cocaine seeking: an updateand clinical implications. Eur J Pharmacol 526:36–50
Collins RJ, Weeks JR, Cooper MM, Good PI, Russell RR (1984)Prediction of abuse liability of drugs using IV self-administrationby rats. Psychopharmacology 82:6–13
Cooper JO, Heron TE, Heward WL (1987) Applied behavior analysis.Macmillan, New York
Crawley JN (2003) Behavioral phenotyping of rodents. Comp Med53:140–146
Davis M, Myers KM, Chhatwal J, Ressler KJ (2006) Pharmacologicaltreatments that facilitate extinction of fear: relevance to psycho-therapy. NeuroRx 3:82–96
Daza-Losada M, Ribeiro Do Couto B, Manzanedo C, Aguilar MA,Rodríguez-Arias M, Miñarro J (2007) Rewarding effects andreinstatement of MDMA-induced CPP in adolescent mice.Neuropsychopharmacology 32:1750–1759
De Vries TJ, Shaham Y, Homberg JR, Crombag H, Schuurman K,Dieben J, Vanderschuren LJ, Schoffelmeer AN (2001) Acannabinoid mechanism in relapse to cocaine seeking. Nat Med7:1151–1154
De Vry J, Donselaar I, Van Ree JM (1989) Food deprivation andacquisition of intravenous cocaine self-administration in rats:effect of naltrexone and haloperidol. J Pharmacol Exp Ther251:735–740
Diagnostic and statistical manual of mental disorders, 4th edn. Textrevision (DSM-IVTR) (2004) American Psychiatric Association,Washington, DC
Di Ciano P, Everitt BJ (2004) Direct interactions between thebasolateral amygdala and nucleus accumbens core underliecocaine-seeking behavior by rats. J Neurosci 24:7167–7173
EMCDDA (European Monitoring Center for Drugs and DrugAddiction) (2005) Annual report on the state of the drugsproblem in Europe. Luxembourg: Office for official publicationsof the European Communities (http://annualreport.emcdda.eu.int)
Epstein DH, Preston KL, Stewart J, Shaham Y (2006) Toward a modelof drug relapse: an assessment of the validity of the reinstatementprocedure. Psychopharmacology 189:1–16
Festing MFW (1999) Warning: the use of heterogeneous mice mayseriously damage your research. Neurobiol Aging 20:237–244
Fuchs RA, See RE, Middaugh LD (2003) Conditioned stimulus-induced reinstatement of extinguished cocaine seeking inC57BL/6 mice: a mouse model of drug relapse. Brain Res973:99–106
Goeders NE, Guerin GF (1994) Non-contingent electric footshockfacilitates the acquisition of intravenous cocaine self-administra-tion in rats. Psychopharmacology 114:63–70
Graham DL, Edwards S, Bachtell RK, DiLeone RJ, Rios M, Self DW(2007) Dynamic BDNF activity in nucleus accumbens withcocaine use increases self-administration and relapse. Nat Neuro-sci 10:1029–1037
Highfield DA, Mead AN, Grimm JW, Rocha BA, Shaham Y (2002)Reinstatement of cocaine seeking in 129X1/SvJ mice: effects ofcocaine priming, cocaine cues and food deprivation. Psycho-pharmacology 161:417–424
Itzhak Y, Martin JL (2002) Cocaine-induced conditioned placepreference in mice: induction, extinction and reinstatement byrelated psychostimulants. Neuropsychopharmacology 26:130–134
Kalivas PW, McFarland K (2003) Brain circuitry and the reinstate-ment of cocaine-seeking behavior. Psychopharmacology 168:44–56
Keiflin R, Isingrini E, Cador M (2008) Cocaine-induced reinstatementin rats: evidence for a critical role of cocaine stimulus properties.Psychopharmacology 197:649–660
Kita T, Okamoto M, Kubo K, Tanaka T, Nakashima T (1999)Enhancement of sensitization to nicotine-induced ambulatorystimulation by psychological stress in rats. Prog Neuropsycho-pharmacol Biol Psychiatry 23:893–903
Koob GF, LeMoal M (2001) Drug addiction, dysregulation of reward,and allostasis. Neuropsychopharmacology 24:97–129
Kruzich PJ (2007) Does response-contingent access to cocainereinstate previously extinguished cocaine-seeking behavior inC57BL/6J mice? Brain Res 1149:165–171
LeDoux JE (2000) Emotion circuits in the brain. Annu Rev Neurosci23:155–184
Le Foll B, Goldberg SR (2005) Control of the reinforcing effects ofnicotine by associated environmental stimuli in animals andhumans. Trends Pharmacol Sci 26:287–293
Maier SF, Watkins LR (2005) Stressor controllability and learnedhelplessness: the roles of the dorsal raphe nucleus, serotonin, andcorticotropin-releasing factor. Neurosci Biobehav Rev 29:829–841
Martin M, Ledent C, Parmentier M, Maldonado R, Valverde O (2000)Cocaine, but not morphine, induces conditioned place preferenceand sensitization to locomotor responses in CB1 knockout mice.Eur J Neurosci 12:4038–4046
McFarland K, Davidge SB, Lapish CC, Kalivas PW (2004) Limbicand motor circuitry underlying foot-shock-induced reinstatementof cocaine-seeking behavior. J Neurosci 24:1551–1560
Miller NS, Gold MS (1994) Dissociation of “conscious desire”(craving) from and relapse in alcohol and cocaine dependence.Ann Clin Psychiatry 6:99–106
Miner LL (1997) Cocaine reward and locomotor activity in C57BL/6Jand 129/SvJ inbred mice and their F1 cross. Pharmacol BiochemBehav 58:25–30
Phillips TJ, Hen R, Crabbe JC (1999) Complications associated withgenetic background effects in research using knockout mice.Psychopharmacology 147:5–7
Piazza PV, Le Moal ML (1996) Pathophysiological basis ofvulnerability to drug abuse: role of an interaction between stress,glucocorticoids, and dopaminergic neurons. Annu Rev PharmacolToxicol 36:359–378
Piazza PV, Deminiere JM, Le Moal M, Simon H (1990) Stress- andpharmacologically-induced behavioral sensitization increasesvulnerability to acquisition of amphetamine self-administration.Brain Res 514:22–26
Ribeiro do Couto B, Aguilar MA, Rodríguez-Arias M, Miñarro J(2005) Cross-reinstatement by cocaine and amphetamine ofmorphine-induced place preference in mice. Behav Pharmacol16:253–259
Rothman RB, Baumann MH (2003) Monoamine transporters andpsychostimulant drugs. Eur J Pharmacol 479:23–40
Schlussman SD, Ho A, Zhou Y, Curtis AE, Kreek MJ (1998) Effectsof “binge” pattern cocaine on stereotypy and locomotor activityin C57BL/6J and 129/J mice. Pharmacol Biochem Behav60:593–599
Shaham Y, Erb S, Stewart J (2000) Stress-induced relapse to heroinand cocaine seeking in rats: a review. Brain Res Brain Res Rev33:13–33
Shaham Y, Shalev U, Lu L, De Wit H, Stewart J (2003) Thereinstatement model of drug relapse: history, methodology andmajor findings. Psychopharmacology 168:3–20
Psychopharmacology

Shalev U, Yap J, Shaham Y (2001) Leptin attenuates acute fooddeprivation-induced relapse to heroin seeking. J Neurosci 21:RC129
Shalev U, Grimm JW, Shaham Y (2002) Neurobiology of relapse toheroin and cocaine seeking: a review. Pharmacol Rev 54:1–42
Shelton KL, Beardsley PM (2005) Interaction of extinguishedcocaine-conditioned stimuli and footshock on reinstatement inrats. Int J Comp Psychol 18:154–166
Soria G, Mendizábal V, Touriño C, Robledo P, Ledent C, ParmentierM, Maldonado R, Valverde O (2005) Lack of CB1 cannabinoidreceptor impairs cocaine self-administration. Neuropsychophar-macology 30:1670–1680
Soria G, Castañé A, Ledent C, Parmentier M, Maldonado R, Valverde O(2006) The lack ofA2Aadenosine receptors diminishes the reinforcingefficacy of cocaine. Neuropsychopharmacology 31:978–987
Todd M (2004) Daily processes in stress and smoking: effects ofnegative events, nicotine dependence, and gender. PsycholAddict Behav 18:31–39
Tzschentke TM (2007) Measuring reward with the conditioned placepreference (CPP) paradigm: update of the last decade. AddictBiol 12:227–462
Watson R (2007) Cocaine use rises in Europe while overall drug uselevels out. BMJ 335:1117
Weiss F (2005) Neurobiology of craving, conditioned reward andrelapse. Curr Opin Pharmacol 5:9–19
Weiss F, Maldonado-Vlaar CS, Parsons LH, Kerr TM, Smith DL,Ben-Shahar O (2000) Control of cocaine-seeking behavior bydrug-associated stimuli in rats: effects on recovery of extin-guished operant-responding and extracellular dopamine levels inamygdala and nucleus accumbens. Proc Natl Acad Sci USA97:4321–4326
Weiss F, Ciccocioppo R, Parsons LH, Katner S, Liu X, Zorrilla EP,Valdez GR, Ben-Shahar O, Angeletti S, Richter RR (2001)Compulsive drug-seeking behavior and relapse. Neuroadapta-tion, stress, and conditioning factors. Ann N Y Acad Sci 937:1–26
Wiessing L (2005) European drugs agency highlights trends in druguse and problems affecting drug users. Euro Surveill 10:E051215.3
Woolverton WL, Johnson KM (1992) Neurobiology of cocaine abuse.Trends Pharmacol Sci 13:193–200
Psychopharmacology
Related Documents











