A preliminary investigation into calcareous dinoflagellate cysts and problematic microfossils from an expanded Cretaceous/Paleogene boundary section at Kulstirenden, Stevns Klint, Denmark Andrew D. Leighton * , Malcolm B. Hart, Christopher W. Smart School of Geography, Earth and Environmental Sciences, Drake Circus, University of Plymouth, Plymouth PL4 8AA, UK article info Article history: Received 26 May 2010 Accepted in revised form 27 May 2011 Available online 30 June 2011 Keywords: Calcareous dinoflagellate cysts K/Pg boundary Fish clay Stevns Klint Bolboforma abstract Located to the north of the Stevns Klint Peninsula (Denmark), Kulstirenden shows the transition from Cretaceous chalks to Danian carbonates across the Cretaceous e Paleogene (K/Pg) boundary. The K/Pg boundary at Kulstirenden is represented by the Fiskeler Member (Fish Clay), a streaked marl interval with a smectite rich, red oxidised layer at its base marking the boundary. The Fiskeler Member is important as it includes the iridium concentration linked to the bolide impact which may have caused the end-Cretaceous mass extinction and is at its maximum thickness at Kulstirenden (c.45 cm). Calcareous dinoflagellate cysts (Calciodinelloideae) have been investigated from within the fine fraction (45e125 mm) at Kulstirenden in order to understand the ecological implications of these enigmatic microfossils. Several species were found within the Fiskeler Member, including Orthopithonella collaris Wendler et al. (2001 , Rev. Palaeobot. Palynol. 115, 69e77). This species is of particular interest as it demonstrates reduced paratabulation and may indicate a transgressive period immediately after the K/ Pg boundary event. O. collaris has been described as a morphotype formed under post-K/Pg environ- mental conditions and is, therefore, described as a “disaster” taxon that marks the K/Pg boundary at Stevns Klint. The distribution of this taxon is more extensive than previously documented, disappearing as carbonate sedimentation returned in the Cerithium Limestone Member of the Danian. Several calcareous microfossil specimens that were found cannot be associated with the dinoflagellates and display Bolboforma-like features. While the origins of Bolboforma are enigmatic, the current finding expands their stratigraphic range and suggests an appearance of this group of organisms found at Stevns Klint may give an insight into the inception of this group into the earliest Danian. Ó 2011 Elsevier Ltd. All rights reserved. 1. Introduction Located south of Copenhagen, Stevns Klint is a cliff succession that exposes uppermost Maastrichtian and lowermost Danian sedi- ments, including the Cretaceous e Paleogene (K/Pg) boundary. At Stevns Klint, the K/Pg boundary is represented by a relatively thin, clay rich layer known as the Fiskeler Member (previously known as Fish Clay) which is preserved in the troughs between uppermost Maastrichtian chalk mounds and which is overlain by a gradation upwards into the overlying Danian limestones (Hart et al., 2005). The Fiskeler Member is widely regarded as containing the evidence of a bolide impact with a fall-out layer at its base (Alvarez et al., 1980; Smit, 1999). The changes in the biology and chemistry of the Fiskeler Member reflect the relatively short-term environmental changes associated with an extraterrestrial impact, probably associated with the impact crater at Chicxulub (Hildebrand et al., 1991; Wendler et al., 2001; Schulte et al., 2006, 2010; Keller, 2008). Other authors, however, have suggested that the Fiskeler Member was of local origin (Schmitz, 1988). What is not in question is that the Fiskeler Member records the transition from the Cretaceous to the Danian. This smectite rich, red layer (Bauluz et al., 2000) is present as infill within the troughs of the Cretaceous chalk mounds (see Schmitz et al., 1992, Fig. 1). The lateral extent of the Fiskeler Member is variable across the whole of the Stevns Peninsula (Surlyk et al., 2006). The geochemical and palaeontological aspects of the Fisk- eler Member have been previously studied at Stevns Klint, most notably by Schmitz et al. (1992), Frei and Frei (2002), Hart et al. (2004, 2005), Rasmussen et al. (2005) and Sepulveda et al. (2009). However, most studies (with the exception of Sepulveda et al. (2009)) have taken place at the most accessible (and famous) part * Corresponding author. Fax: þ44 (0) 1752 584776. E-mail address: [email protected] (A.D. Leighton). Contents lists available at ScienceDirect Cretaceous Research journal homepage: www.elsevier.com/locate/CretRes 0195-6671/$ e see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.cretres.2011.05.011 Cretaceous Research 32 (2011) 606e617

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

lable at ScienceDirect
Cretaceous Research 32 (2011) 606e617
Contents lists avai
Cretaceous Research
journal homepage: www.elsevier .com/locate/CretRes
A preliminary investigation into calcareous dinoflagellate cysts and problematicmicrofossils from an expanded Cretaceous/Paleogene boundary sectionat Kulstirenden, Stevns Klint, Denmark
Andrew D. Leighton*, Malcolm B. Hart, Christopher W. SmartSchool of Geography, Earth and Environmental Sciences, Drake Circus, University of Plymouth, Plymouth PL4 8AA, UK
a r t i c l e i n f o
Article history:Received 26 May 2010Accepted in revised form 27 May 2011Available online 30 June 2011
Keywords:Calcareous dinoflagellate cystsK/Pg boundaryFish clayStevns KlintBolboforma
* Corresponding author. Fax: þ44 (0) 1752 584776E-mail address: [email protected]
0195-6671/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.cretres.2011.05.011
a b s t r a c t
Located to the north of the Stevns Klint Peninsula (Denmark), Kulstirenden shows the transition fromCretaceous chalks to Danian carbonates across the Cretaceous e Paleogene (K/Pg) boundary. The K/Pgboundary at Kulstirenden is represented by the Fiskeler Member (Fish Clay), a streaked marl intervalwith a smectite rich, red oxidised layer at its base marking the boundary. The Fiskeler Member isimportant as it includes the iridium concentration linked to the bolide impact which may have causedthe end-Cretaceous mass extinction and is at its maximum thickness at Kulstirenden (c.45 cm).Calcareous dinoflagellate cysts (Calciodinelloideae) have been investigated from within the fine fraction(45e125 mm) at Kulstirenden in order to understand the ecological implications of these enigmaticmicrofossils. Several species were found within the Fiskeler Member, including Orthopithonella collarisWendler et al. (2001, Rev. Palaeobot. Palynol. 115, 69e77). This species is of particular interest as itdemonstrates reduced paratabulation and may indicate a transgressive period immediately after the K/Pg boundary event. O. collaris has been described as a morphotype formed under post-K/Pg environ-mental conditions and is, therefore, described as a “disaster” taxon that marks the K/Pg boundary atStevns Klint. The distribution of this taxon is more extensive than previously documented, disappearingas carbonate sedimentation returned in the Cerithium Limestone Member of the Danian. Severalcalcareous microfossil specimens that were found cannot be associated with the dinoflagellates anddisplay Bolboforma-like features. While the origins of Bolboforma are enigmatic, the current findingexpands their stratigraphic range and suggests an appearance of this group of organisms found at StevnsKlint may give an insight into the inception of this group into the earliest Danian.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Located south of Copenhagen, Stevns Klint is a cliff successionthat exposes uppermost Maastrichtian and lowermost Danian sedi-ments, including the Cretaceous e Paleogene (K/Pg) boundary. AtStevns Klint, the K/Pg boundary is represented by a relatively thin,clay rich layer known as the Fiskeler Member (previously known asFish Clay) which is preserved in the troughs between uppermostMaastrichtian chalk mounds and which is overlain by a gradationupwards into the overlying Danian limestones (Hart et al., 2005). TheFiskeler Member is widely regarded as containing the evidence ofa bolide impact with a fall-out layer at its base (Alvarez et al., 1980;Smit, 1999). The changes in the biology and chemistry of the Fiskeler
.(A.D. Leighton).
All rights reserved.
Member reflect the relatively short-term environmental changesassociated with an extraterrestrial impact, probably associated withthe impact crater at Chicxulub (Hildebrand et al., 1991; Wendleret al., 2001; Schulte et al., 2006, 2010; Keller, 2008). Other authors,however, have suggested that the Fiskeler Member was of localorigin (Schmitz, 1988). What is not in question is that the FiskelerMember records the transition from the Cretaceous to the Danian.This smectite rich, red layer (Bauluz et al., 2000) is present as infillwithin the troughs of the Cretaceous chalk mounds (see Schmitzet al., 1992, Fig. 1). The lateral extent of the Fiskeler Member isvariable across the whole of the Stevns Peninsula (Surlyk et al.,2006). The geochemical and palaeontological aspects of the Fisk-eler Member have been previously studied at Stevns Klint, mostnotably by Schmitz et al. (1992), Frei and Frei (2002), Hart et al.(2004, 2005), Rasmussen et al. (2005) and Sepulveda et al. (2009).However, most studies (with the exception of Sepulveda et al.(2009)) have taken place at the most accessible (and famous) part

Fig. 1. Locality map of the Stevns Peninsula, Denmark (after Hart et al., 2004).
A.D. Leighton et al. / Cretaceous Research 32 (2011) 606e617 607
of the Stevns Klint succession which is located at Højerup and, toa certain extent, Rødvigwhere the Fiskeler Member is at a maximumthickness of c.7 cm (Fig. 1). As a result, resolution of data is oftenreduced across such an important boundary. This is in contrast withthe location a few km north at Kulstirenden where the FiskelerMember has a thickness of <45 cm.
The K/Pg boundary is marked by an important decrease incalcareous nannofossil absolute abundance (Bernaola and Monechi,2007). Although most authors state that a significant proportion ofthe Cretaceous species found in the Danian sediments (excludinga few persistent species) are reworked (see Tantawy, 2003 andreferences therein; Bown, 2005) a few authors suggest that severalCretaceous species survived the catastrophe though they disappearedafter a short time (Gardin and Monechi, 1998; Gardin, 2002). Calcar-eous dinoflagellate cysts show a high diversity throughout theCretaceous and are assumed to reflect conditions within the photiczone (Wendler andWillems, 2002). They also display a wide range ofcharacter traits and differing morphological ornamentation fromspecies to species such as cyst formation, cyst size, wall structure orcyst shape. Previous studies byWendler et al. (2001) andWendler andWillems (2002) were the first high-resolution analyses of the FiskelerMember and indicate the presence of a distinctive flora of calcareous
dinoflagellate cysts (Calciodinelloideae) within the thinner FiskelerMember at Højerup. Kienel (1994) also studied calcareous dinofla-gellate cysts across the K/Pg boundary within the Boreal realm,including Stevns Klint, but never investigated the Fiskeler Member.Calcareous dinoflagellate cysts are particularly well preservedthroughout the Fiskeler Member at Stevns Klint. This is in contrast tothe severe dissolution affecting some of the foraminifera (Hart et al.,2005). As a result of this the calcareous dinoflagellate cysts seem torecord a more complete record than the foraminifera.
The Subfamily Calciodinelloideae (Fensome et al., 1993) includesall dinoflagellates that form calcareous resting cysts as part of theirlife cycle (Wendler and Willems, 2002). These cysts are mostlyspherical in shape and do not show any signs of paratabulation in themajority of cases, although some species clearly reflect the peri-dinioid plate pattern of the corresponding motile dinoflagellate(Wendler et al., 2001). Wendler et al. (2001) describe a new species(Orthopithonella collaris Wendler et al., 2001) exclusively present atStevns Klint. O. collaris is very distinctive as it shows reduced para-tabulation and the authors conclude that it has a “pulse-like”occurrence immediately after the boundary event in the FiskelerMember. Species that display reduced paratabulation are of partic-ular interest as their occurrence in the stratigraphic record is char-acterised by short-term, often event-like abundance peaks inrelation to transgressive/regressive trends (Wendler et al., 2001) andMilankovitch cyclicity (Wendler et al., 2002). Studies of calcareousdinoflagellate cysts around the K/Pg boundary time interval arelimited, with the southern Atlantic Ocean and the Boreal realmpreferentially studied: see Willems (1994) and Hildebrand-Habelet al. (1999). In this study we focus on the distribution patterns ofspecific Calciodinelloideae (i.e., O. collaris, Pithonella spp.) across theK/Pg boundary along with some problematic findings that poten-tially can be grouped with Bolboformawithin the expanded FiskelerMember at Kulstirenden. This study aims to focus on the ecologicalinterpretations of these Calciodinelloideae.
A revision of the upper Maastrichtian e Danian lithostratig-raphy of the succession exposed at Stevns Klint has recently beenprovided by Surlyk et al. (2006) who propose new names for theindividual members of the succession (Fig. 2). The beds examinedin this study belong to the Sigerslev Member (¼ White coccolithchalk), Fiskeler Member (¼ Fish Clay) and Cerithium LimestoneMember (¼ Cerithium Limestone). The Højerup Member (¼ “GreyChalk”) is absent in some sections at Kulstirenden, including thesection analysed in this study. This revised lithostratigraphicalterminology is used here.
1.1. Geological setting
Stevns Klint is the coastal cliff of the Stevns Peninsula in thesouth-eastern part of the Danish Basin (Anderskouv et al., 2007)which was part of an extensive NW European epeiric sea (Surlyk,1997). Bounded by the inverted Sorgenfrei e Tonquist Zone to thenorth east and the Ringkøbing e Fyn High to the south (Bjeragerand Surlyk, 2007), the Danish Basin was elongated in a NWeSEorientation (Håkansson and Thomas, 1999) and formed by rifting inthe earliest Permian through to the Jurassic, with subsequentthermal contraction in the Cretaceous (Anderskouv et al., 2007).The Stevns Klint area lies just to the northern edge of the Ring-købing e Fyn High (Fig. 3) and to the east is the major tectonicboundary of the Fennoscandian Border Zone (Hart et al., 2004). Asa result, the basement below the Cretaceous succession is exten-sively faulted (Hart et al., 2005), although the thrusting andbedding-plane slippage seen in some other, arguably more famous,K/Pg sections in Southern Spain (Agost and Caravaca de la Cruz) isnot observed in the Danish sections (Hart et al., 2004, 2005). Duringthe latest Maastrichtian, a narrow marine seaway crossed over the

Fig. 2. Lithostratigraphy for the MaastrichtianeDanian succession exposed at Stevns Klint (after Surlyk et al., 2006).
A.D. Leighton et al. / Cretaceous Research 32 (2011) 606e617608
Danish region and connected the initial North Atlantic to the northwest of the Danish region. The south west of the Danish Regionwasconnected through to the Tethyan area across Europe via the Cri-mean Basin (Håkansson and Thomas, 1999).
2. Materials and methods
Samples for micropalaeontological analysis were carefullyprepared in a clean environment at the University of Plymouth.Samples were processed using the White Spirit method of Brasier(1980). Where the samples were pure chalks or partially cementedlimestones, they were placed inside clean sealable bags and gentlycrushed down to a maximum size of large pebbles (according to theWentworth Scale) using an iron pestle and mortar before beingreturned to the same aluminium bowl and submerged in deionisedwater for a minimum of 12 h. These samples were returned toa specially designed iron pestle and mortar and gently crushed toafine sand/silt grademixture. Although these sampleswere subjectedto a more physical approach, there seems to be no indication that thiscaused excessive damage to the microfossils and this technique iswidely used in the preparation of “chalky” samples (see Heinburg,2005; Rasmussen et al., 2005). Samples that were processed in thisway are indicated in Fig. 4 with an asterisk (*). All samples were wetsieved through a 45 mmmesh and the residues collectedwerewashedinto filter paper and returned to their original aluminium bowlsbefore being placed in the oven at a temperature of 40 �C for a periodof at least 5 h. Once dried, the >45 mm residues were dry sieved on250 mm,150 mmand 63 mmsieves.Micropalaeontological analysiswasachieved using an eyelash sable brush under a binocular microscopeand mounted on a lightly gummed micropalaeontological slides. Allsampleswere processed and analysed in stratigraphical order in order
minimise contamination between samples. No chemicals were usedin the processing except White Spirit and, in line with processingtechnique of Hart et al. (2004, 2005). Checks of damage being causedto the microfossils and the bias that this might have caused weremade at all stages of the processing. It is, therefore, proposed thatthere is no evidence to suggest that the micropalaeontological resi-dues are not a good representation of the initial assemblages. Speci-mens that were selected for analysis under the scanning electronmicroscope (SEM) were mounted on a specimen stub with a double-sided sticker and coated twice with 12 Å gold (to avoid “charging”)using a sputter coater. SEM analysis and imaging was performed ona JEOL 5600 and 6100 SEM at an accelerating voltage of 15 kV.Calculations of the number of cysts/g were made using the equation;
#perg=1gram¼#counted*weight45�150=weightcounted*weightbulk
2.1. Site location
Kulstirenden is located w8 km north of Højerup towards thenorthern end of Stevns Klint (Hart et al., 2005) on the north-easternfacing sea cliffs. The succession at Kulstirenden has been over-looked by the majority of previous studies in favour of the moreaccessible sections at Højerup and Rødvig: see Schmitz et al. (1992)and references therein. The expanded Fiskeler Member at Kulstir-enden is located high in the cliff succession (Fig. 5), in contrast toHøjerup and Rødvig where the Fiskeler Member is accessibledirectly from the beach. Present along the Stevns Klint section isa prominent, regular line of massive black flints that can becorrelated throughout the succession at Højerup, Harvig and Kul-stirenden (Hart et al., 2005; Surlyk et al., 2006) and this providesa direct correlation between the various locations (Fig. 6). The

Fig. 3. Geological map of Denmark showing the position of Stevns Klint and Thisted (marking the location of other key K/Pg boundary sections such as Nye Kløv, Bulberg). (modifiedafter Hart et al., 2005).
A.D. Leighton et al. / Cretaceous Research 32 (2011) 606e617 609
Fiskeler Member rests directly on a hardground, with the whitechalks of the Sigerslev Member immediately below this hard-ground (Hart et al., 2005). The boundary between the FiskelerMember and the underlying white chalks of the Sigerslev Memberreflects an abrupt transition from carbonate to clay sedimentation,with the gradational return of carbonate sedimentation in theCerithium Limestone Member (Hart et al., 2005). The expandedFiskeler Member at Kulstirenden is shown in Fig. 7.
3. Results and analysis
Previous work at Højerup by Wendler et al. (2001) and Wendlerand Willems (2002) indicated the presence of calcareous dinofla-gellate cysts in the Cretaceous chalks and basal Danian marls. Thesestudies identified 12 distinct species across the K/Pg boundary.Wendler et al. (2001) and Wendler and Willems (2002) wet sievedthrough a 20 mmmesh, while this study wet sieved through a 45 mmmesh. Subsequently, some of the taxa described by Wendler et al.(2001) and Wendler and Willems (2002) are in the smaller sizerange (e.g., Cervisiella spp., Calcicarpinum spp., Lentodinella danica)and are not represented in this study. To fully investigate calcareousdinoflagellate cysts in their entirety at Stevns Klint, reprocessingusing a 20 mmmeshwill be undertaken in on-going research. Severalspecies of calcareous dinoflagellate cysts and other problematicmicrofossils were found in varying abundances over the stratigraphicinterval studied at Kulstirenden within the 45e63 mm and the63e150 mm size fractions used in this study. Identification, using
electronmicroscopy, indicates that calcareous dinoflagellate cysts areparticularly well preserved throughout the section, in comparisonwith the dissolution affecting some species of foraminifera. Wendleret al. (2001) also found this to be the case at Højerup, although thesampling interval they studied was small (>2 cm) and did not extendinto the Cerithium Limestone Member. “Pithonella” sp. recorded thelongest stratigraphical distribution, being present in every samplepicked within the Danian (Fig. 4). “Pithonella” sp. is distinct as it hasa surface crystal pattern that made it clearly distinguishable fromambiguous carbonate grains. “Pithonella” sp. was recorded as “Pith-onella” as this species displayed elongations along one axis and wassimilar (morphologically) to specimens described by Wendler andWillems (2002). O. collaris also extends over a long stratigraphicinterval, ranging from�16.5 cm toþ4.25 cm. The stratigraphic rangeof O. collaris is significant as it does not appear to have the “pulse-like” distribution just above the boundary described byWendler et al.(2001) despite the short-term geological duration of the Fish Claydeposition. The distribution of O. collaris is more extensive than theprevious lower-resolution study of Wendler et al. (2001) describedalthough it does appear to be a “disaster species”. New ecologicalinterpretations can be proposed from this occurrence and these arediscussed below. A quantative distribution for both O. collaris and“Pithonella” sp. is shown in Fig. 8, based on the numbers that werepicked from the 63e150 mm size fraction.
O. collaris cysts/g records a first occurrence at �16.5 cm witha peak in abundance at�13 cm of 14.38 cysts/g, coinciding with thetop of the black and grey streakedmarl.O. collaris is absent from the

Fig. 4. Sedimentary log of the Kulstirenden section. The scale is at zero within the Fiskeler Member as it is placed at a marked colour change that is present throughout the localsuccession.
Fig. 5. Sampling at Kulstirenden being undertaken by Prof. M. Hart, Prof. E. Håkansson and Dr C. Heinburg. The K/Pg boundary is indicated by the red dotted line. (For interpretationof the references to colour in this figure legend, the reader is referred to the web version of this article).
A.D. Leighton et al. / Cretaceous Research 32 (2011) 606e617610

Fig. 6. Lithological logs for the various sections investigated across Stevns Klint. Included is the new stratigraphical scheme of Surlyk et al. (2006) for the exposures at Stevns Klint.Modified after Hart et al. (2004).
A.D. Leighton et al. / Cretaceous Research 32 (2011) 606e617 611
next sample but is recorded in the sample above and then fluctu-ates around w3 cysts/g up section. O. collaris is absentfrom þ3.25 cm upwards in the 63e150 mm size fraction but is,however, recorded in the 45e63 mm size fraction at þ4.25 cm: it isnot recorded above this level. The upper stratigraphic range ofO. collaris coincides with the transition from the Fiskeler Member to
Fig. 7. Expanded Fish Clay section at Kulstirenden. 1) Højerup Member. 2) Fiskeler MemberMember and the Fiskeler Member. The exact boundary between the Fiskeler Member and thwith a dotted line.
the Cerithium Limestone Member. This is located in the sedimen-tary succession by a change from a marl to a limestone. “Pithonella”sp. cysts/g record a first appearance at the boundary layer sampleat �18 cm with a peak in abundance at �16.5 cm of 68.66 cysts/g.“Pithonella” sp. cysts/g fluctuate around w9 cysts/g to the limit ofsampling, the species being present in both the 45e63 mm and the
. 3) Cerithium Limestone Member. The K/Pg boundary is located between the Højerupe Cerithium Limestone Member is undefined and transitional, and as such is indicated

Orthopithonella collaris Wendler et al., 2001/g "Pithonella" sp./g
Fig. 8. Quantitative distribution of two calcareous dinocyst species (“Pithonella” sp. and Orthopithonella collaris) from within the Fiskeler Member at Kulstirenden.
Fig. 9. Stratigraphical distribution of calcareous dinoflagellate cysts at Kulstirenden, Stevns Klint. Species “A” e “Pithonella sphaerica”, sample KU29. Range; 18 cme17 cm. Scalebar ¼ 20 mm. Species “B” e “Pithonella” sp., sample KU28. Range; 18 cmeþ8.5 cm. Scale bar ¼ 10 mm. Species “C” e Orthopithonella collaris, sample KU25. Range; 16.5 cmeþ4.25 cm.Scale bar ¼ 20 mm. Species “D” e “Pirumella” sp., sample KU27. Range; 15 cme14 cm. Scale bar ¼ 10 mm. Species “E” e “micro-problematicum E”, sample KU29. Range; 18 cme17 cm.Scale bar ¼ 50 mm. Species “F” e “micro-problematicum F”, sample KU16. Range; þ4 cmeþ3 cm. Scale bar ¼ 50 mm. Species “G” e “micro-problematicum G”, sample KU39.Range; þ6.25 cmeþ8 cm. Scale bar ¼ 50 mm. Species “H” e “micro-problematicum H”, sample KU29. Range; 18 cme17 cm. Scale bar ¼ 20 mm.
A.D. Leighton et al. / Cretaceous Research 32 (2011) 606e617612
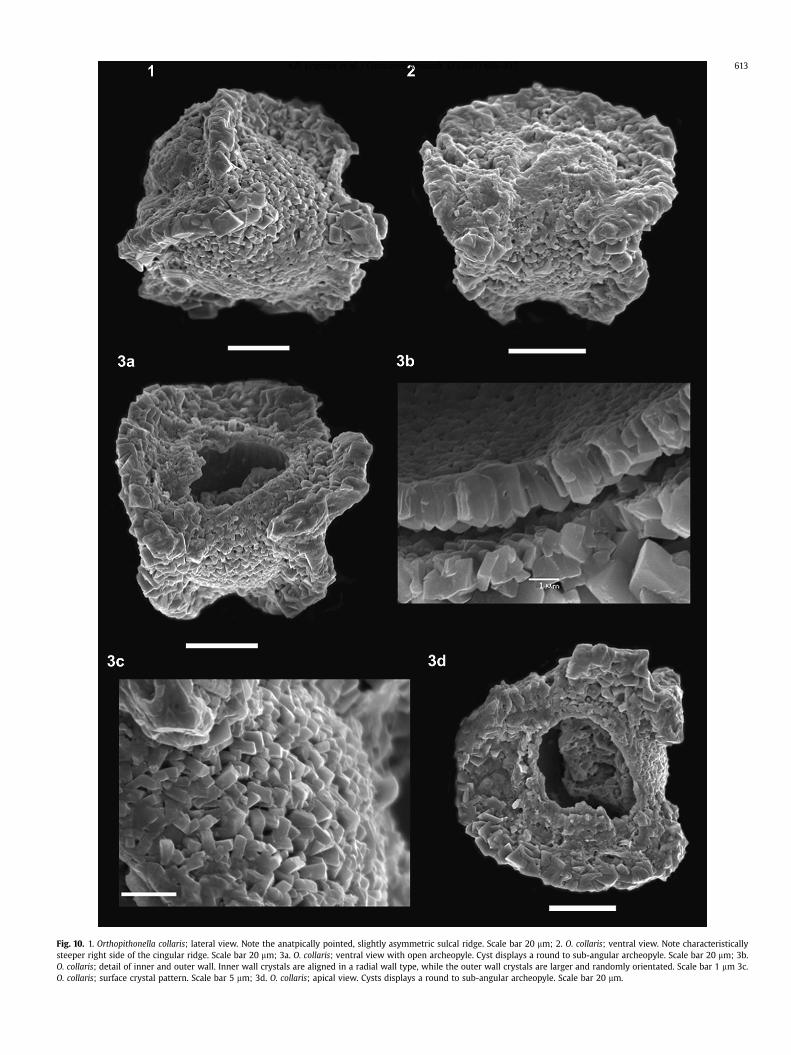
Fig. 10. 1. Orthopithonella collaris; lateral view. Note the anatpically pointed, slightly asymmetric sulcal ridge. Scale bar 20 mm; 2. O. collaris; ventral view. Note characteristicallysteeper right side of the cingular ridge. Scale bar 20 mm; 3a. O. collaris; ventral view with open archeopyle. Cyst displays a round to sub-angular archeopyle. Scale bar 20 mm; 3b.O. collaris; detail of inner and outer wall. Inner wall crystals are aligned in a radial wall type, while the outer wall crystals are larger and randomly orientated. Scale bar 1 mm 3c.O. collaris; surface crystal pattern. Scale bar 5 mm; 3d. O. collaris; apical view. Cysts displays a round to sub-angular archeopyle. Scale bar 20 mm.
A.D. Leighton et al. / Cretaceous Research 32 (2011) 606e617 613

Fig. 11. 1. Species “E” e “micro-problematicum E”, sample KU29. Range; 18 cme17 cm. Scale bar 50 mm; 2. Species “H” e “micro-problematicum H”, sample KU29. Range;18 cme17 cm. Scale bar 20 mm; 3. Species “D” e “Pirumella” sp., sample KU27. Range; 15 cme14 cm. Scale bar 10 mm; 4. Species “G” e “micro-problematicum G”, sample KU39.Range; þ6.25 cmeþ8 cm. Scale bar 50 mm; 5. Species “F” e “micro-problematicum F”, sample KU16. Range; þ4 cmeþ3 cm. Scale bar 50 mm; 6. Species “A” e “Pithonella sphaerica”,sample KU29. Range; 18 cme17 cm. Scale bar 20 mm; 7. Species “B” e “Pithonella” sp., sample KU28. Range; 18 cmeþ8.5 cm. Scale bar 10 mm; 8a. Species “B” e “Pirumella kra-sheninnikovii(?)”, sample KU28. Range; 18 cmeþ8.5 cm. Scale bar 10 mm; 8b. Species “B” e “Pirumella krasheninnikovii(?)”. Surface crystal pattern. Scale bar 5 mm.

A.D. Leighton et al. / Cretaceous Research 32 (2011) 606e617 615
63e150 mm size fractions in all samples analysed in the Danian.With our present sampling at Kulstirenden the upper stratigraphiclimit cannot be confined.
4. Discussion and conclusions
Studies of calcareous dinoflagellate cysts at Stevns Klintcomprise the work of Kienel (1994), Wendler et al. (2001) andWendler and Willems (2002). The distribution of the calcareousdinoflagellate cysts and other problematic microfossils foundwithin the Fiskeler Member in this study are presented in Fig. 9.Scanning electron microscope images of O. collaris are shown inFig. 10, while all other calcareous dinoflagellate cysts are illustratedin Fig. 11. Our principle findings are presented below.
4.1. Orthopithonella collaris Wendler et al., 2001
The first record of O. collaris (Fig. 10) is only found in the FiskelerMember at Højerup (Wendler et al., 2001). The species was, there-fore, described as being a possible biostratigraphic and ecologicalmarker for post-K/Pg environmental conditions in continental shelfareas by Wendler et al. (2001). O. collaris displays reduced para-tabulation and is, therefore, of particular interest as occurrences ofcalcareous dinoflagellate cysts with reduced paratabulation in thestratigraphic record are characteristic of short-term, often “eventlike” abundance peaks in relation to transgressive/regressive trendsand Milankovitch cyclicity (Wendler et al., 2001, 2002; Wendler andWillems, 2002). The reduced paratabulation pattern is also unique toO. collaris which clearly distinguishes it from other calcareousdinoflagellate cysts. At Kulstirenden (this study), O. collaris firstappears in the basal Danian and is absent at the top of the FiskelerMember, not being present in any size fraction of the samples fromthe Cerithium Limestone Member. The peak abundance of O. collarisis at the top of the black and grey streaked marl (sample KU25). Ourwork confirms the findings of Wendler et al. (2001) that this speciesis “Orthopithonella” based on the radial wall type of the orientation ofthe c-axiswall crystals (Fig.10 3b) as defined by Kohring et al. (2005).Its occurrence, solely within the Fiskeler Member at both Højerupand Kulstirenden, suggests that this species is a “disaster species”formed during a period of environmental stress. Wendler andWillems (2002) propose that O. collaris is stress-controlled andstate that this is themost “straightforward explanation” of the short-term appearance of O. collaris. This view is supported by Kohringet al. (2005) who state that character traits in calcareous dinofla-gellate cysts are triggered by palaeoceanographic parameters such assea-level changes, water temperature, nutrient conditions, etc.. Thelack of information about O. collarismakes the interpretation of it asa world-wide or even regional marker for the K/Pg boundaryimpossible. However, O. collaris can be used as a marker for the K/Pgboundary at Stevns Klint as two separate areas (Højerup and Kul-stirenden) record its occurrence within the Fiskeler Member. It isalso suggested that the reduced paratabulation is ecologically trig-gered and not a genetically fixed character trait. Assumptions canonly be made as to why this occurs. Perhaps the paratabulation maybe related to buoyancy or changes in temperature, nutrient supply orsalinity caused by the K/Pg event.
Wendler andWillems (2002) alternatively suggest that O. collariscould have been a latitudinal invader in comparison with organicdinoflagellates which apparently show a pulse-like temperature-related latitudinal migration event in the aftermath of the K/Pgimpact (Brinkhuis et al., 1998). Wendler et al. (2001) suggests thatO. collariswas an invader from the Boreal realm based on thework ofBrinkhuis et al. (1998) who proposed a pulse-like northwardmigration of a Tethyan, warm water, organic-walled dinoflagellatecyst species during the deposition of the Fiskeler Member. Hart et al.
(2005) suggest that the FiskelerMembermarks amaximum floodingsurface, suggesting that the appearance of O. collaris may be indic-ative of a transgressive interval within the Fiskeler Member. Theappearance and increase in abundance of the benthic foraminiferidTappanina selmensiswithin the Fiskeler Member (Hart et al., 2011) isfurther evidence of this sea-level rise as T. selmensis is thought to bea deeper water taxon, normally indicative of an outer shelf setting(Schmitz et al.,1992). A rise in sea-level across the boundary and intothe earliest Danian is indicated by Schmitz et al. (1992, Fig. 2) andthis may explain the calcareous dinoflagellate cyst record.
4.2. “Pithonella” sp.
“Pithonella” sp. is the most abundant calcareous dinoflagellatecyst within the section, being present in the 45e63 mm and63e150 mm size fractions. “Pithonella” sp. has a micro-crystallinewall structure comprising of columnar crystals and is consideredin this study tobe a Cretaceous form that has been reworked into theDanian samples. Upper Cretaceous chalks characteristically containa high abundance of spherical calcareousmicrofossils determined tobe pithonelloid by their wall type (Wendler andWillems, 2004). Itspeak abundance occurs in sample KU28. This record does not reflectthe peak abundance of the “in situ” O. collaris in sample KU25. Thelowermost Danian samples also show evidence of reworking ofuppermost Cretaceous foraminifera (e.g., Bolivinoides spp). This isfurther evidence of the reworking of the uppermost Cretaceouschalks during the deposition of the basal Danain. Distribution ofupwards-decreasing numbers is suggestive of reworking and has noecological significance other than availability of a “source”.
4.3. The “Bolboforma” mystery?
For some of the specimens present in the Fiskeler Member anassociationwith calcareous dinoflagellates is questionable. SpecimensE-H in Fig. 9 display no evidence of dinoflagellate characteristics (e.g.,archaeopyle type or theca tabulation) and are larger than mostcalcareous dinoflagellate cysts which are 30e90 mm in diameter.These specimens are more representative of Bolboforma, which arenormally characterised by a calcitic monocrystalline sphaeroidal testwith, or without, inner cysts (Spiegler et al., 2005) and are charac-teristic of predominantly temperate to cool regions in both theNorthern and Southern Hemispheres (Spiegler, 1999). Spiegler et al.(2005) describe Bolboforma tests as being mostly sphaeroidal andsingle-chambered with a conical or tubular neck ending in a simpleaperture. Such forms have affinities with protophytic algae(Spezzaferri and Rögl, 2004). Sepulveda et al. (2009) examined indetail the geochemistry of the Fiskeler Member and stated thatphotosynthesis, although diminished, persisted across the K/Pgboundary. Sepulveda et al. (2009) also concluded that primary algalproduction in a neritic high-latitude ecosystem rapidly recoveredafter the K/Pg event. These findings are significant in the context ofthe proposed affinity of Bolboforma to protophytic algae. Test sizesrange from 60e240 mm, at an average of less than 150 mm in diameter.The surface morphology is characterised by reticulation, spines,knobs, ridges or flanges (Spiegler et al., 2005). These characters areseen in specimens E-H in Fig. 9. A tubular neck appears to be presentin specimen H (Fig. 11.2). The determination of these specimens asbeing Bolboforma implies an expansion of the stratigraphic range ofthis “genus” by 12Ma to the K/Pg boundary. The previously statedrange of Bolboforma is late Early Eocene to Late Pliocene (Spiegleret al., 2005). Identification of these microfossils as Bolboformawould require the first occurrence of the taxon to be lowered byw12 Ma. Spiegler (1999) indicated that bolboformids are marine,calcareous microfossils of “uncertain origin” with Spiegler et al.(2005) stating that their origin remains enigmatic. Our specimens

A.D. Leighton et al. / Cretaceous Research 32 (2011) 606e617616
do not appear to be reworked as they are very well preserved. Theydisplay almost pristine preservation in samples which record severecarbonate dissolution in other microfossils (e.g., foraminifera). Bol-boformids are recorded as showing strong resistance to dissolution(Spezzaferri and Rögl, 2004).
5. Conclusions
Calcareous dinoflagellate cysts across the K/Pg boundary at Kul-stirendenareabundant, ornateandwellpreserved.This isparticularlythe case for samples from within the Fiskeler Member. O. collaris isfoundexclusively in the FiskelerMemberatKulstirendenandappearsto be a genuine “disaster” species immediately following the K/Pgevent. O. collaris has a greater stratigraphic occurrence than previ-ously described, largely due to the higher sampling resolution of theFiskeler Member at Kulstirenden. To fully investigate the calcareousdinoflagellate cysts and the other problematic microfossils at StevnsKlint, processing using a 20 mmmesh would be required, in linewithWendler et al. (2001) and Wendler and Willems (2002). O. collarisdisplays reduced paratabulation, which is thought to indicatea transgressiveperiod immediatelyabove theK/Pgboundary. This risein sea-level is supported by the foraminiferal and sedimentologicalrecord. Cysts that display a distinctive paratabulation and is thoughtto reflect a narrow palaeoecological niche (Wendler and Willems,2004) and often occur in environments of highly fluctuating condi-tions. The paleobiogeographical occurrence ofO. collaris is impossibleto determine as this species has only been described from StevnsKlint. Wendler et al. (2001) interpreted O. collaris as being eithera morphotype formed under environmental stress or an invader intothe Boreal realm from higher or lower latitudes. Since no otheroccurrence of O. collaris has been documented it is, therefore,suggestive of a specieswhichwas formed under conditions related tothe post-K/Pg boundary event. If O. collaris is recorded in other K/Pgboundary sections world-wide then it would become a potentialmarker species for the K/Pg event. IfO. collaris is not found elsewherethen it must be an ecophenotype. Some other calcareous dinoflagel-late cysts that appear in the Fiskeler Member, or indeed the wholeKulstirenden section, are enigmatic as some display characteristictraits of bolboformid taxa. The potential relationship of Bolboformawith protophytic algae is also significant with the biomarker data ofSepulveda et al. (2009) and may point towards the earliest recoveryplayers after the K/Pg boundary. Should these species be interpretedas Bolboforma then this would lower their first stratigraphic occur-rence by w12 Ma and suggest that the origins of Bolboforma arerelated to the K/Pg boundary event.
Acknowledgements
We thank Professors Finn Surlyk and Ekart Håkansson (bothKøbenhavns Universitet) for advice on the sections at Stevns Klint.We also extend our thanks to Dr Jens Wendler for helpful discus-sions on the section.
References
Alvarez, L., Alvarez, W., Asaro, F., Michel, H., 1980. Extraterrestrial cause for theCretaceouseTertiary extinction. Science 208, 1095e1108.
Anderskouv, K., Damholt, T., Surlyk, F., 2007. Late Maastrichtian chalk mounds,Stevns Klint, Denmark e combined physical and biogenic structures. Sedi-mentary Geology 200, 57e72.
Bauluz, B., Peacor, D.R., Elliott, W.C., 2000. Coexisting altered glass and FeeNi oxidesat the CretaceouseTertiary boundary, Stevns Klint (Denmark): direct evidenceof meteorite impact. Earth and Planetary Science Letters 182, 127e136.
Bernaola, G., Monechi, S., 2007. Calcareous nannofossil extinction and survivorshipacross the CretaceousePaleogene boundary at Walvis ridge (ODP Hole 1262C,south Atlantic Ocean). Palaeogeography, Palaeoclimatology, Palaeoecology 255,132e156.
Bjerager, M., Surlyk, F., 2007. Benthic palaeoecology of Danian deep-shelf bryozoansmounds in the Danish Basin. Palaeogeography, Palaeoclimatology, Palae-oecology 250, 184e215.
Bown, P.R., 2005. Selective calcareous nannoplankton survivorship at the Creta-ceouseTertiary boundary. Geology 33, 653e656.
Brasier, M.D., 1980. Microfossils. George Allen & Unwin, London. 162e168.Brinkhuis, H., Bujak, J.P., Smit, J., Versteegh, G.J.M., Visscher, H., 1998. Dinofla-
gellate-based sea surface temperature reconstructions across the Cretaceous-Tertiary boundary. Palaeogeography, Palaeoclimatology, Palaeoecology 141,67e83.
Fensome, R.A., Taylor, F.J.R., Norris, G., Sarjeant, W.A.S., Wharton, D.I., Williams, G.L.,1993. A Classification of Living and Fossil Dinoflagellates, 7 (Special Publica-tions), 351 p.
Frei, R., Frei, K.M., 2002. A multi-isotopic and trace element investigation of theCretaceous-Tertiary boundary layer at Stevns Klint, Denmark e inferences forthe origin and nature of siderophile and lithophile element geochemicalanomalies. Earth and Planetary Science Letters 203, 691e708.
Gardin, S., 2002. Late Maastrichtian to early Danian calcareous nannofossils at Elles(Northwest Tunisia). A tale of one million years across the KeT boundary.Palaeogeography, Palaeoclimatology, Palaeoecology 178, 211e231.
Gardin, S., Monechi, S., 1998. Palaeoecological change in middle to low latitudecalcareous nannoplankton at the Cretaceous/Tertiary boundary. Bulletin de laSociété Géologique de France 169, 709e723.
Håkansson, E., Thomas, E., 1999. Benthic extinction and recovery patterns at the K/Tboundary in shallow water carbonates, Denmark. Palaeogeography, Palae-oclimatology, Palaeoecology 154, 67e85.
Hart, M.B., Feist, S.E., Price, G.D., Leng, M.J., 2004. Re-appraisal of the K/T boundarysuccession at Stevns Klint, Denmark. Journal of the Geological Society, London161, 885e892.
Hart, M.B., Feist, S., Håkansson, E., Heinburg, C., Price, G.D., Leng, M.J.,Watkinson, M.P., 2005. The Cretaceous-Palaeogene boundary succession atStevns Klint, Denmark: foraminifers and stable isotope stratigraphy. Palae-ogeography, Palaeoclimatology, Palaeoecology 224, 6e26.
Hart, M.B., Searle, S.R., Feist, S.E., Leighton, A.D., Price, G.D., Smart, C.W., Twitchett,R.J., 2011. The distribution of benthic foraminifera across the Cretaceous-Palaeogene boundary in Texas (Brazos River) and Denmark (Stevns Klint). In:Keller, G., Adatte,T. (Eds.), The End-Cretaceous Mass Extinction and the Chic-xulub Impact in Texas, SEPM Special Publication No. 100.
Heinburg, C., 2005. Morphotype biostratigraphy, diachronism, and bivalve recoveryin the earliest Danian of Denmark. Bulletin of the Geological Society ofDenmark 52, 81e95.
Hildebrand, A.R., Penfield, G.T., Kring, D.A., Pilkington, M., Camargo, Z.A., 1991.Chicxulub crater: a possible Cretaceous/Tertiary boundary impact crater in theYucatan Peninsula. Geology 19, 867e871.
Hildebrand-Habel, T., Willems, H., Versteegh, G.J.M., 1999. Variations in calcareousdinoflagellate associations from the maastrichtian to middle eocene of thewestern south Atlantic Ocean (Sao Paulo Plateau, DSDP Leg 39, Site 356).Review of Palaeobotany and Palynology 106, 57e87.
Keller, G., 2008. Cretaceous climate, volcanism, impacts, and biotic effects. Creta-ceous Research 29, 754e771.
Kienel, U., 1994. Die Entwicklung der kalkigen Nannofossilien und der kalkigenDinoflagellaten-Zysten an der Kreide/Tertiär-Grenze in Westbrandenburg imVergleich mit Profilen in Nordjütland und Seeland (Dänemark). Berliner Geo-wissenschaftliche Abhandlungen 12, 87.
Kohring, R., Gottschling, M., Keupp, H., 2005. Examples for character traits andpalaeoecological significance of calcareous dinoflagellates. PaläontologischeZeitschrift 79, 79e91.
Rasmussen, J.A., Heinburg, C., Håkansson, E., 2005. Planktonic foraminifers,biostratigraphy and the diachronous nature of the lowermost Danian Cerithiumlimestone at Stevns Klint, Denmark. Bulletin of the Geological Society ofDenmark 52, 113e131.
Schmitz, B., Keller, G., Stenvall, O., 1992. Stable isotope and foraminiferal changesacross the Cretaceous-Tertiary boundary at Stevns Klint, Denmark: argumentsfor long term oceanic instability before and after bolide-impact event. Palae-ogeography, Palaeoclimatology, Palaeoecology 96, 233e260.
Schmitz, B., 1988. Origin of microlayering in worldwide distributed Ir-rich marineCretaceous/Tertiary boundary clays. Geology 16, 1068e1072.
Schulte, P., Speijer, R., Mai, H., Kontny, A., 2006. The Cretaceous-Paleogene (K-P)boundary at Brazos, Texas: sequence stratigraphy, depositional events and theChicxulub impact. Sedimentary Geology 184, 77e109.
Schulte, P., Alegret, L., Arenillas, I., Arz, J.A., Barton, P.J., Brown, P.R., Bralower, T.J.,Christeson, G.L., Claeys, P., Cockell, C.S., Collins, G.S., Deutsch, A., Goldin, T.J.,Goto, K., Grajales-Nishimura, J.M., Grieve, R.A.F., Gulick, S.P.S., Johnson, K.R.,Kiessling, W., Koeberl, C., Kring, D.A., Macleod, K.G., Matsui, T., Melosh, J.,Montanari, A., Morgan, J.V., Neal, C.R., Nichols, D.J., Norris, R.D., Pierazo, E.,Ravizza, G., Rebolledo-Vieyra, M., Reimond, W.U., Robin, E., Salge, T., Speijer, R.P.,Sweet, A.R., Urrutia-Fucugauchi, J., Vajda, V., Whalen, M.T., Willumsen, P.S.,2010. The chicxulub asteroid impact and mass extinction at the Cretaceous-Paleogene boundary. Science 327, 1214e1218.
Sepulveda, J., Wendler, J.E., Summons, R.E., Hinrichs, K., 2009. Rapid Resurgence ofmarine productivity after the Cretaceous-Paleogene mass extinction. Science326, 129e132.
Smit, J., 1999. The global stratigraphy of the Cretaceous-Tertiary boundary impactejecta. Annual Review of Earth and Planetary Sciences 27, 75e113.

A.D. Leighton et al. / Cretaceous Research 32 (2011) 606e617 617
Spezzaferri, S., Rögl, F., 2004. Bolboforma (Phytoplankton Incertae Sedis), Bach-mayerella and other Calciodinelloidea (Phytoplankton) from the middleMiocene of the AlpineeCarpathian Foredeep (Central Paratethys). Journal ofMicropalaeontology 23, 139e152.
Spiegler, D., 1999. Bolboforma biostratigraphy from the Hatton-Rockall Basin (NorthAtlantic). In: Raymo, M.E., Jansen, E., Blum, P., Herbert, T.D. (Eds.), Proceedingsof the Ocean Drilling Program, Scientific Results, vol. 162, pp. 35e49. Texas.
Spiegler, D., Spezzaferri, K., Spezzaferri, S., 2005. Bolboforma e an overview. Pal-äontologische Zeitschrift 79, 167e181.
Surlyk, F., 1997. A cool-water carbonate ramp with bryozoan mounds: late Creta-ceouseDanian of the Danish Basin. In: Jamrd, N.P., Clarke, J.D.A. (Eds.), CoolWater Carbonates. Society for Sedimentary Geology, vol. 56, pp. 293e307(Special Publications).
Surlyk, F., Damholt, T., Bjerager, M., 2006. Stevns Klint, Denmark: uppermostMaastrichtian chalk, Cretaceous-Tertiary boundary, and lower Danian bryo-zoans mound complex. Bulletin of the Geological Society of Denmark 54, 1e48.
Tantawy, A.A., 2003. Calcareous nannofossil biostratigraphy and paleoecology of theCretaceouseTertiary transition in the central eastern desert of Egypt. MarineMicropaleontology 47, 323e356.
Wendler, J., Wendler, I., Willems, H., 2001. Orthopithonella collaris sp. nov., a newcalcareous dinoflagellate cyst from the K/T boundary (Fish Clay, Stevns Klint/Denmark). Review of Palaeobotany and Palynology 115, 69e77.
Wendler, J., Willems, H., 2002. Distribution pattern of calcareous dinoflagellatecysts across the Cretaceous-Tertiary boundary (Fish Clay, Stevns Klint,Denmark): implications for our understanding of species-selective extinction.In: Koeberl, C., MacLeod, K.G. (Eds.), Catastrophic Events and Mass Extinctions:Impacts and Beyond, pp. 265e276. Geological Society of America Special Paper356, Colorado.
Wendler, J., Willems, H., 2004. Pithonelloid wall-type of the Late Cretaceouscalcareous dinoflagellate cyst genus tetratropis. Review of Palaeobotany andPalynology 129, 133e140.
Wendler, J., Gräfe, K., Willems, H., 2002. Reconstruction of mid-Cenomanianorbitally forced palaeoenvironmental changes based on calcareous dino-fagellate cysts. Palaeogeography, Palaeoclimatology, Palaeoecology 179,19e41.
Willems, H., 1994. New calcareous dinoflagellates from the upper Cretaceouswhite chalk of northern Germany. Review of Palaeobotany and Palynology84, 57e72.
Related Documents











