For permission to copy, contact [email protected] © 2007 Geological Society of America 3 ABSTRACT Microfossil assemblages and their shell geochemistry are widely used in paleocean- ography, but they can be significantly altered by subtle variations in preservation state. Clay-rich, hemipelagic sediments of the Paleogene Kilwa Group of coastal Tanzania host calcareous microfossils that are excep- tionally preserved, as evidenced by morpho- logical, taxonomic, and geochemical data. The planktonic foraminifera are preserved as glassy, translucent tests with original microgranular wall textures that resemble well-preserved modern specimens, and they arguably yield geochemical values that are relatively unaffected by recrystallization. The calcareous nannofossils are extraordi- narily diverse and represented by unique assemblage compositions that include dis- solution-susceptible taxa, especially holo- coccoliths and rhabdoliths, and fragile and very small (<3-μm) heterococcoliths, many of which are new taxa. Notably, the extant, deep–photic-zone taxon Gladiolithus is documented for the first time in the pre- Quaternary fossil record. The Kilwa Group calcareous nannofossil diversities are con- sistently higher than all coeval assemblages and provide a benchmark against which to compare other Paleogene biodiversity data. Highest diversities are preserved in hemipe- lagic, clay-rich lithologies and the greatest losses occur in lithified, carbonate-rich sedi- ments. Most of the lost diversity, however, is confined to distinct taxonomic groups (holococcoliths and Syracosphaerales), and in general the preservational potential of Paleogene coccolithophores was greater than in the modern oceans because a larger proportion of the biodiversity fell within the larger size fractions. For both foraminifera and coccolithophores, incorporation into impermeable clay-rich sediments that have never been deeply buried appears to have been critical in producing this Konservat- Lagerstätte preservation. Keywords: calcareous nannofossils, foraminifera, preservation, Lagerstätte, Paleogene, diversity. INTRODUCTION Konservat-Lagerstätten are extraordinary fos- sil occurrences characterized by unusual quality of preservation (Briggs, 2001). Most commonly, they preserve the soft parts of animals and pro- vide rare glimpses of the biology and biodi- versity of ancient ecosystems. Paleontologists studying fossils of unicellular protistan organ- isms have rarely thought of individual deposits as Lagerstätten because they routinely work with stratigraphically continuous data sets, compris- ing assemblages of many hundreds to thousands of specimens, which are considered to equate to relatively complete fossil records. Calcareous microfossils, in particular, are generally robust enough to provide stratigraphically useful data from a wide range of lithologies and depositional settings. There is increasing evidence, however, that microfossil assemblages and their geochem- ical signatures may be significantly altered by subtle or cryptic variations in preservation state (Pearson et al., 2001, 2007; Gibbs et al., 2004; Williams et al., 2005). One approach to assess- ing the potential magnitude of such change is to search for sequences with exceptional A Paleogene calcareous microfossil Konservat-Lagerstätte from the Kilwa Group of coastal Tanzania P.R. Bown † T. Dunkley Jones J.A. Lees R.D. Randell J.A. Mizzi Department of Earth Sciences, University College London, Gower Street, London WC1E 6BT, England, UK P.N. Pearson H.K. Coxall School of Earth, Ocean and Planetary Sciences, Cardiff University, Main Building, Park Place, Cardiff CF10 3YE, Wales, UK J.R. Young Department of Palaeontology, The Natural History Museum, London SW7 5BD, England, UK C.J. Nicholas Department of Geology, Trinity College, Dublin 2, Ireland A. Karega J. Singano Tanzania Petroleum Development Corporation, PO Box 2774, Dar-es-Salaam, Tanzania B.S. Wade Department of Geology and Geophysics, Texas A&M University, College Station, Texas 77843-3115, USA † E-mail: [email protected] GSA Bulletin; January/February 2008; v. 120; no. 1/2; p. 3–12; doi: 10.1130/B26261.1; 5 figures; 1 table.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

For permission to copy, contact [email protected]© 2007 Geological Society of America
3
ABSTRACT
Microfossil assemblages and their shell geochemistry are widely used in paleocean-ography, but they can be signifi cantly altered by subtle variations in preservation state. Clay-rich, hemipelagic sediments of the Paleogene Kilwa Group of coastal Tanzania host calcareous microfossils that are excep-tionally preserved, as evidenced by morpho-logical, taxonomic, and geochemical data. The planktonic foraminifera are preserved as glassy, translucent tests with original microgranular wall textures that resemble well-preserved modern specimens, and they arguably yield geochemical values that are relatively unaffected by recrystallization. The calcareous nannofossils are extraordi-narily diverse and represented by unique assemblage compositions that include dis-solution-susceptible taxa, especially holo-coccoliths and rhabdoliths, and fragile and very small (<3-µm) heterococcoliths, many of which are new taxa. Notably, the
extant, deep–photic-zone taxon Gladiolithus is documented for the fi rst time in the pre-Quaternary fossil record. The Kilwa Group calcareous nannofossil diversities are con-sistently higher than all coeval assemblages and provide a benchmark against which to compare other Paleogene biodiversity data. Highest diversities are preserved in hemipe-lagic, clay-rich lithologies and the greatest losses occur in lithifi ed, carbonate-rich sedi-ments. Most of the lost diversity, however, is confi ned to distinct taxonomic groups (holococcoliths and Syracosphaerales), and in general the preservational potential of Paleogene coccolithophores was greater than in the modern oceans because a larger proportion of the biodiversity fell within the larger size fractions. For both foraminifera and coccolithophores, incorporation into impermeable clay-rich sediments that have never been deeply buried appears to have been critical in producing this Konservat-Lagerstätte preservation.
Keywords: calcareous nannofossils, foraminifera, preservation, Lagerstätte, Paleogene, diversity.
INTRODUCTION
Konservat-Lagerstätten are extraordinary fos-sil occurrences characterized by unusual quality of preservation (Briggs, 2001). Most commonly, they preserve the soft parts of animals and pro-vide rare glimpses of the biology and biodi-versity of ancient ecosystems. Paleontologists studying fossils of unicellular protistan organ-isms have rarely thought of individual deposits as Lagerstätten because they routinely work with stratigraphically continuous data sets, compris-ing assemblages of many hundreds to thousands of specimens, which are considered to equate to relatively complete fossil records. Calcareous microfossils, in particular, are generally robust enough to provide stratigraphically useful data from a wide range of lithologies and depositional settings. There is increasing evidence, however, that microfossil assemblages and their geochem-ical signatures may be signifi cantly altered by subtle or cryptic variations in preservation state (Pearson et al., 2001, 2007; Gibbs et al., 2004; Williams et al., 2005). One approach to assess-ing the potential magnitude of such change is to search for sequences with exceptional
A Paleogene calcareous microfossil Konservat-Lagerstätte from the Kilwa Group of coastal Tanzania
P.R. Bown†
T. Dunkley JonesJ.A. LeesR.D. RandellJ.A. MizziDepartment of Earth Sciences, University College London, Gower Street, London WC1E 6BT, England, UK
P.N. PearsonH.K. CoxallSchool of Earth, Ocean and Planetary Sciences, Cardiff University, Main Building, Park Place, Cardiff CF10 3YE, Wales, UK
J.R. YoungDepartment of Palaeontology, The Natural History Museum, London SW7 5BD, England, UK
C.J. NicholasDepartment of Geology, Trinity College, Dublin 2, Ireland
A. KaregaJ. SinganoTanzania Petroleum Development Corporation, PO Box 2774, Dar-es-Salaam, Tanzania
B.S. WadeDepartment of Geology and Geophysics, Texas A&M University, College Station, Texas 77843-3115, USA
†E-mail: [email protected]
GSA Bulletin; January/February 2008; v. 120; no. 1/2; p. 3–12; doi: 10.1130/B26261.1; 5 fi gures; 1 table.

Bown et al.
4 Geological Society of America Bulletin, January/February 2008
preservation and use them as benchmarks against which to judge the extent of taphonomic and geochemical alteration in other sections.
Recent attempts to seek out exceptional fora-minifera for geochemical paleoclimate studies have targeted clay-rich, hemipelagic sediments (Wilson et al., 2002; Bice et al., 2003; Pearson et al., 2007). However, the paleobiological poten-tial of these predominantly shelf successions that host well-preserved microfossils remains largely unexploited. To a large extent, this is the result of the enormous amount of stratigraphic and paleoceanographic work that has accompa-nied the Deep Sea Drilling Project and Ocean Drilling Program since the late 1960s, and the rather uniform state of preservation that is typi-cally associated with such deep-sea chalks and oozes. Cenozoic nannofossil study in particular saw a slowing of taxonomic description after the switch from largely continental-shelf research to deep-sea studies, but the effect is less pro-nounced in Mesozoic research, which has con-tinued to rely on hemipelagic successions.
The aim of this paper is to exemplify excep-tional calcareous microfossil preservation through a description of the Paleogene Kilwa Group of Tanzania. These sediments represent a Konservat-Lagerstätte for calcareous micro-fossils and provide a benchmark against which to highlight the signifi cant effects that preser-vation can have on both microfossil diversity and geochemistry.
GEOLOGICAL SETTING
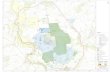
The Tanzania Drilling Project (TDP) is a paleoclimate research program that has targeted the recovery of Cretaceous-Paleogene sedi-ments with exceptionally preserved planktonic foraminifera through critical stratigraphic inter-vals (Pearson et al., 2004, 2006; Nicholas et al., 2006). Surface-exposure mapping and subsur-face coring have focused on the Kilwa Group (Santonian-Oligocene), which crops out along a coastal strip of southern Tanzania around the towns of Kilwa, Lindi, and Pande (Fig. 1). The group comprises a roughly kilometer-thick suc-cession of homogeneous, dark claystones, with variably developed secondary lithologies of siltstones, limestones, and sandstones. The clay-stones are typically uncemented and unlithifi ed, and have never been deeply buried. Based on paleontological and lithological considerations, the succession is thought to have been depos-ited in an outer-shelf to upper-slope environ-ment at water depths of 300–500 m (Nicholas et al., 2006). The passive margin continental shelf was located at around 19° S paleolatitude in the Eocene. The shelf was relatively narrow, and therefore the sediments also incorporate
shallower, inner-shelf components, such as intermittent carbonate beds with larger benthic foraminifera and terrestrial organic matter (van Dongen et al., 2006; Nicholas et al., 2006). The rich planktonic foraminifera and calcare-ous nannoplankton assemblages indicate fully marine, open-ocean conditions. Age determina-tion of the sediments was achieved by integrated micropaleontological analysis (nannofossils, palynology, and foraminifers), and initial bio-stratigraphic results are presented in Pearson et al. (2004, 2006) and Nicholas et al. (2006).
METHODS
Calcareous microfossils were studied using standard micropaleontological methods. Nan-nofossils were viewed as simple smear slides (Bown and Young, 1998), using transmitted-light microscopy in cross-polarized and phase-contrast light (507 samples), and on broken rock
surfaces using scanning electron microscopy (SEM). Assemblage data were collected from smear slides using semiquantitative and quanti-tative count methods (Bown and Young, 1998). Nannofossil species richness is a tally of all taxa occurring in one nannofossil zone. Size data were collected from digital light microscope images sourced from all the TDP sites (e.g., Bown, 2005a; Bown and Dunkley Jones, 2006). Foraminifera were prepared by gently disaggre-gating the sediment in tap water and washing over a 63-µm sieve. Residues were viewed using light microscopy and SEM.
KILWA GROUP MICROFOSSILS
The Paleogene Kilwa Group sediments con-tain planktonic foraminifera that in refl ected light have a translucent and refl ective, glassy appearance, comparable to well-preserved mod-ern specimens (Fig. 2A). High-resolution SEM
1 623
10
9
11
12
13
14a
14b
15a
15b
15c
16
17
1819
20
21
22
67
8
29
31
30
NP
Nf.
Zon
es
Early
Olig
ocen
e
TDP coresAge
Ma
O3
O2
O4
P5
E2E1
E3
E4
E5
E6
E7
E8
E9
E10
E11
E12
E13
E14
E15
E16
O1
P4c
P4b
55
54
53
56
52
49
51
47
48
50
46
44
45
43
42
41
10
87
3
2
6
39
38
40
37
Mid
dle
Eoce
neEa
rly E
ocen
eL.
Pal
eoce
ne
36
34
35
33
32
Late
Eoc
ene
PF
Zon
es
12 11
14
57
58
20
16
19
17
pars
4
13
18
24pars
N
KENYA
Pemba Is.
Zanzibar Is.
MafiaIs.
Dar esSalaam
Kilwa Masoko
Pande
Lindi
MOZAM-BIQUE
TANZANIA
MainN-Sroad
KilwaGroupoutcrop
IndianOcean
0 100km 0 20km
Mkazambo
A
B
Figure 1. (A) Stratigraphic extent of the TDP boreholes. Nannofossil zonation (NP Nf. Zones) from Martini (1971) and plank-tonic foraminifer zonation (PF Zones) from Berggren and Pearson (2005). Correlation of the two plankton zonation schemes and time scale are from the latter. (B) Location of the Kilwa Group in coastal Tanzania. Detailed location is given in Nicholas et al. (2006).
Tanzania microfossil Konservat-Lagerstätte
Geological Society of America Bulletin, January/February 2008 5
A
C
JI
E
H
B
D
F G
K
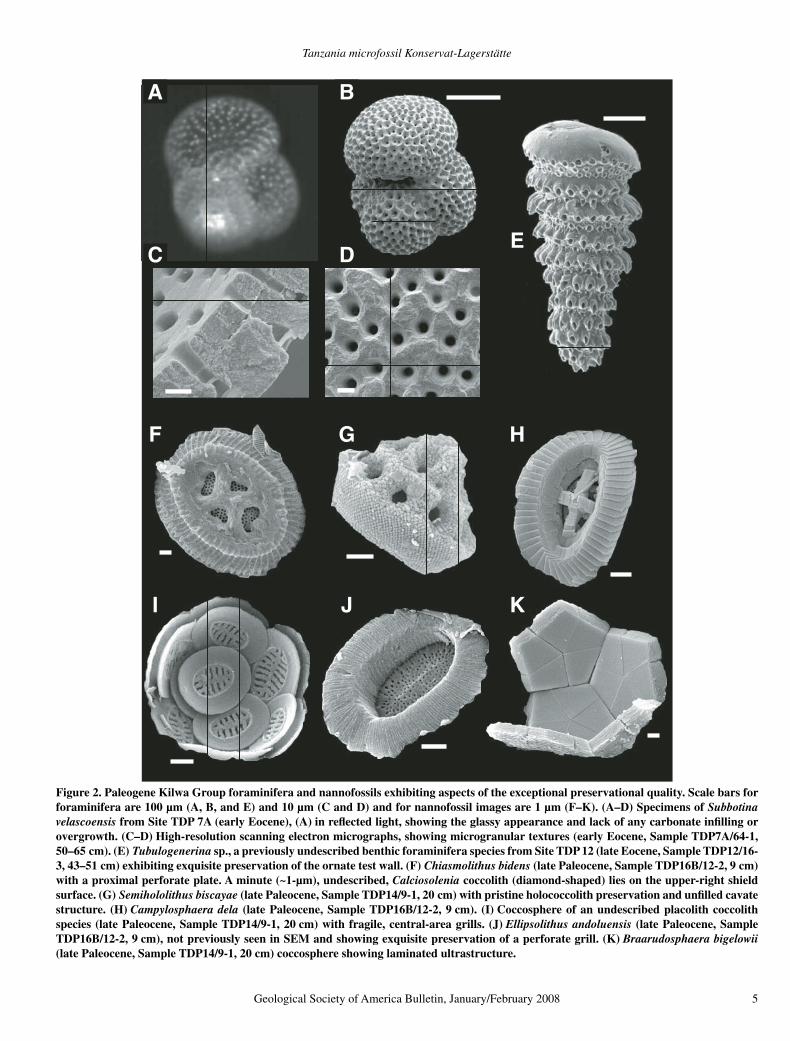
Figure 2. Paleogene Kilwa Group foraminifera and nannofossils exhibiting aspects of the exceptional preservational quality. Scale bars for foraminifera are 100 µm (A, B, and E) and 10 µm (C and D) and for nannofossil images are 1 µm (F–K). (A–D) Specimens of Subbotina velascoensis from Site TDP 7A (early Eocene), (A) in refl ected light, showing the glassy appearance and lack of any carbonate infi lling or overgrowth. (C–D) High-resolution scanning electron micrographs, showing microgranular textures (early Eocene, Sample TDP7A/64-1, 50–65 cm). (E) Tubulogenerina sp., a previously undescribed benthic foraminifera species from Site TDP 12 (late Eocene, Sample TDP12/16-3, 43–51 cm) exhibiting exquisite preservation of the ornate test wall. (F) Chiasmolithus bidens (late Paleocene, Sample TDP16B/12-2, 9 cm) with a proximal perforate plate. A minute (~1-µm), undescribed, Calciosolenia coccolith (diamond-shaped) lies on the upper-right shield surface. (G) Semihololithus biscayae (late Paleocene, Sample TDP14/9-1, 20 cm) with pristine holococcolith preservation and unfi lled cavate structure. (H) Campylosphaera dela (late Paleocene, Sample TDP16B/12-2, 9 cm). (I) Coccosphere of an undescribed placolith coccolith species (late Paleocene, Sample TDP14/9-1, 20 cm) with fragile, central-area grills. (J) Ellipsolithus andoluensis (late Paleocene, Sample TDP16B/12-2, 9 cm), not previously seen in SEM and showing exquisite preservation of a perforate grill. (K) Braarudosphaera bigelowii (late Paleocene, Sample TDP14/9-1, 20 cm) coccosphere showing laminated ultrastructure.

Bown et al.
6 Geological Society of America Bulletin, January/February 2008
reveals well-preserved microgranular wall tex-tures, hollow chambers, and large, well-defi ned pores (Figs. 2B–D), and specimens display morphological features that have not previously been observed (Fig. 2E). The exceptional pres-ervation of the tests is also indicated by their oxygen and carbon stable isotope values that are signifi cantly different than those from coeval sediments deposited at comparable latitudes, but which are less well preserved (Pearson et al., 2001, 2007; Stewart et al., 2004; Sexton et al., 2006). However, the taxonomy and diver-sity of these coeval foraminifera assemblages are broadly comparable, indicating that the robust shells conserve the overall composition of assemblages despite the differences in shell appearance, ultrastructure, and geochemistry.
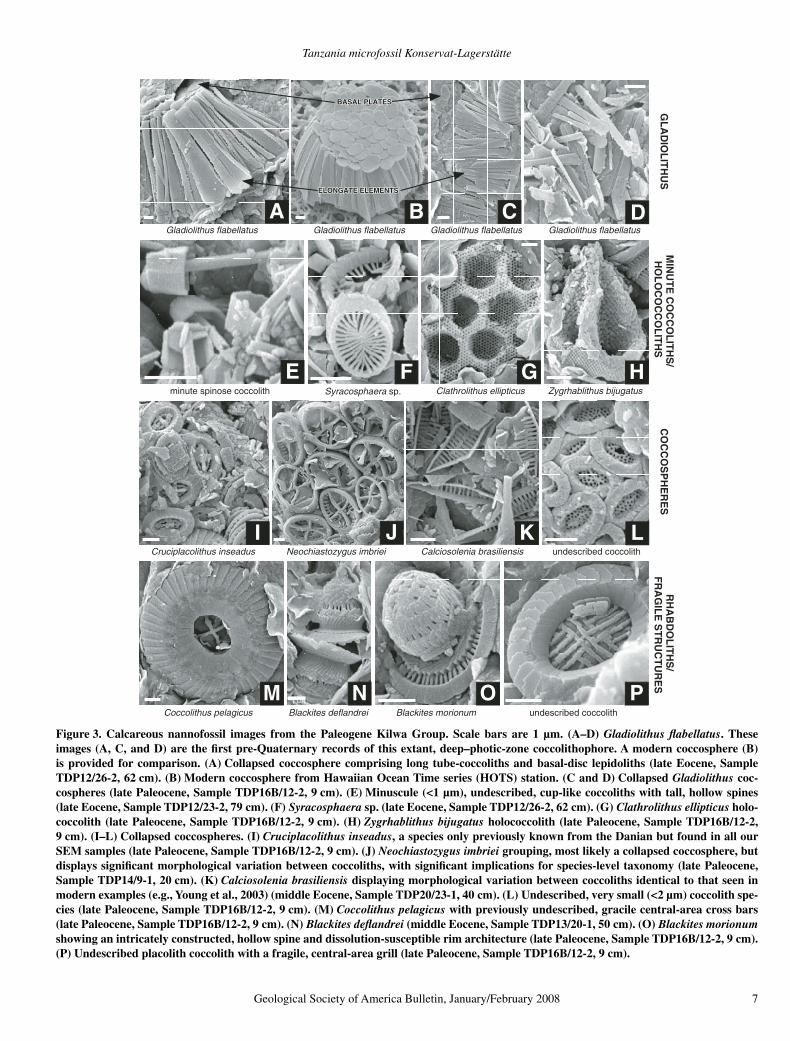
In contrast to the planktonic foraminifera, the Kilwa Group calcareous nannofossil assem-blages are extraordinarily diverse (around 360 species in total) and have distinct taxonomic compositions (Figs. 2 and 3). Comparative nan-nofossil species richness data are provided in Table 1 and Figure 4. More than a quarter of the Paleogene nannofossil diversity is due to the presence of holococcoliths (48 species, Figs. 2G and 3G–H) and several diverse coccolith groups, most notably representatives of the living fam-ily Rhabdosphaeraceae (informally known as rhabdoliths) (42 species, Figs. 3N–O). The assemblages also include small and/or fragile coccoliths (Figs. 2F and 3E–F), as well as larger forms with delicate central-area structures, which are usually only discernible using SEM (Figs. 2F, 2I–J, 3M, and 3P). The small, delicate forms include the extant genus Gladiolithus, documented in this paper for the fi rst time in the pre-Quaternary fossil record (Figs. 3A–D), and the extant genera Calciosolenia and Syra-cosphaera, which also have very poor fossil records (Figs. 2F, 3F, and 3K).
Gladiolithus was not identifi ed until SEM observation of rock surfaces revealed recurrent assemblages of abundant lath-like elements within coccolith concentrations. Almost com-plete, collapsed coccospheres were observed, along with entire tube coccoliths and the dis-tinctive lepidolith coccoliths (Figs. 3A–D). The Gladiolithus coccoliths and disaggregated ele-ments are extremely thin, and the calcite c-axis is oriented nearly perpendicular to the surface of the laths, so they show very low birefrin-gence and are thus virtually invisible in the light microscope. Gladiolithus and Calciosolenia are consistently present throughout the Paleo-gene succession and are often abundant. Syra-cosphaera is only documented in the middle Eocene to early Oligocene part of the Kilwa Group. Several samples have yielded concentra-tions of Calciosolenia that appear to represent
collapsed coccospheres (Fig. 3K) showing vari-morphism (changes in shape across the cocco-sphere), similar to that seen in modern species (Young et al., 2003). Coccospheres of placolith and non-placolith taxa are relatively common in smear slides and on rock surfaces (Figs. 2I, 2K, 3A, 3C, 3E, and 3I–L).
Other common, small coccoliths include minuscule (<1-µm) spinose forms that are, as yet, undescribed (Fig. 3E). They are not eas-ily classifi ed in existing fossil groups but are comparable to the extant Papposphaeraceae and “narrow-rimmed muroliths” (Young et al., 2003, p. 78), which have no previously docu-mented fossil record. Larger coccoliths with fragile central-area structures include new taxa that are diffi cult to place within existing fossil classifi cations (Figs. 2I and 3P). Well-known species with delicate structures that have not been previously observed (Figs. 2F, 2J, and 3M) are also preserved. Coccolithus pelagicus speci-mens, for example, are frequently seen with gracile, axial cross bars (Fig. 3M), demonstrat-ing a subtle morphological difference compared to modern populations, where single transverse bars are common (Young et al., 2003). Delicate central grills are occasionally reported in other coccolith groups, but are routinely observed in the Tanzania material, most notably in Cyclicar-golithus, Reticulofenestra, Chiasmolithus, and Cruciplacolithus (Figs. 2F and 3I).
DISCUSSION
Signifi cance of Kilwa Group Microfossil Assemblage Components
The geochemistry and visual appearance of the planktonic foraminifera shells indicate the exceptional quality of the larger Kilwa Group microfossils, but it is the extraordinary diversity and preservation of calcareous nannofossils that substantiates the unique Konservat-Lagerstätte status (Figs. 2–4). A component of the high nan-nofossil diversity can be explained by the pres-ence of broadly shelf-dwelling taxa (11%), such as Braarudosphaera, and some diversity is the result of updated taxonomic concepts based on the Kilwa Group research itself (Bown, 2005a; Bown and Dunkley Jones, 2006). However, most of the high diversity is unequivocally related to the quality of preservation and, in particular, the presence of holococcoliths, rhabdoliths, and small and fragile taxa.
Holococcoliths and RhabdolithsHolococcoliths and small rhabdoliths are
not routinely preserved in modern deep-sea sediments (Roth and Berger, 1975), and they are considered to be the most prone to dissolution
(Roth and Thierstein, 1972). Holococcoliths are constructed from minute, equidimensional cal-cite crystallites and formed during the haploid phase of the haplo-diplontic coccolithophore life cycle. They typically alternate with a dip-loid phase that produces the more commonly observed and robustly constructed heterococ-coliths (compare Figs. 2G and 2H). Over 90 holococcolith morphologies have been docu-mented in the modern ocean (32% of the total morphological diversity), but all of them are small (<3 µm) and none typically preserve in the sedimentary record (Young et al., 2003). Liv-ing holococcolith-bearing coccolithophores are widely distributed (Kleijne, 1991) and show no particular affi nity for shelf environments. Their absence from seafl oor sediments is simply the result of a preservational fi lter, which removes these highly dissolution-prone coccoliths. There are several larger, extinct holococcoliths (e.g., Zygrhablithus; Fig. 3H) that do consistently preserve in the fossil record, but even these are absent in sediments deposited in deeper waters (~3000 m Bown, 2005b). In general, fossil holococcolith preservation is patchy and largely restricted to hemipelagic, clay-rich lithologies. For the Paleogene and Late Cretaceous time intervals, the new taxa described from the Kilwa Group have effectively doubled the known holo-coccolith fossil diversity, with the addition of 25 Paleogene and 23 Late Cretaceous species (Bown, 2005a; Bown and Dunkley Jones, 2006; Lees, 2007).
Rhabdoliths are also widely distributed in the modern ocean (e.g., Boeckel et al., 2006) but have a patchy and inconsistent fossil record. They are typically spinose and can be large, but minimal dissolution leads to fragmenta-tion of the coccoliths and disarticulation of the spines. Again, their abundance and diver-sity in the Kilwa Group sediments is the result of exceptional preservation, as can be seen in the delicate rim and spine structures shown in Figures 3N and 3O.
GladiolithusGladiolithus is the most surprising compo-
nent of the new diversity preserved in the Kilwa Group (Figs. 3A–D). It is one of a small number of living coccolithophores specifi cally adapted to life in the deep photic zone (100–200 m), and it produces highly modifi ed coccoliths and coccospheres that are suspected to be morpho-logical adaptations related to the low-light con-ditions (Young, 1994). Despite being abundant in the water column, the liths are rarely found at the seafl oor (Roth and Berger, 1975), and they have not been previously documented in sediments older than late Quaternary (Okada and Matsuoka, 1996). In fact, there has been
Tanzania microfossil Konservat-Lagerstätte
Geological Society of America Bulletin, January/February 2008 7
FR
AG
ILE
ST
RU
CT
UR
ES
CO
CC
OS
PH
ER
ES
HO
LO
CO
CC
OL
ITH
SG
LA
DIO
LIT
HU
SR
HA
BD
OL
ITH
S/
MIN
UT
E C
OC
CO
LIT
HS
/
Coccolithus pelagicus Blackites deflandrei Blackites morionum undescribed coccolith
Cruciplacolithus inseadus Neochiastozygus imbriei Calciosolenia brasiliensis undescribed coccolith
minute spinose coccolith Syracosphaera sp. Clathrolithus ellipticus Zygrhablithus bijugatus
Gladiolithus flabellatus Gladiolithus flabellatus Gladiolithus flabellatus Gladiolithus flabellatus
BASAL PLATESBASAL PLATES
ELONGATE ELEMENTSELONGATE ELEMENTS
BASAL PLATES
ELONGATE ELEMENTS
A DC
E
I
M P
HF
J L
O
B
K
N
G
Figure 3. Calcareous nannofossil images from the Paleogene Kilwa Group. Scale bars are 1 µm. (A–D) Gladiolithus fl abellatus. These images (A, C, and D) are the fi rst pre-Quaternary records of this extant, deep–photic-zone coccolithophore. A modern coccosphere (B) is provided for comparison. (A) Collapsed coccosphere comprising long tube-coccoliths and basal-disc lepidoliths (late Eocene, Sample TDP12/26-2, 62 cm). (B) Modern coccosphere from Hawaiian Ocean Time series (HOTS) station. (C and D) Collapsed Gladiolithus coc-cospheres (late Paleocene, Sample TDP16B/12-2, 9 cm). (E) Minuscule (<1 µm), undescribed, cup-like coccoliths with tall, hollow spines (late Eocene, Sample TDP12/23-2, 79 cm). (F) Syracosphaera sp. (late Eocene, Sample TDP12/26-2, 62 cm). (G) Clathrolithus ellipticus holo-coccolith (late Paleocene, Sample TDP16B/12-2, 9 cm). (H) Zygrhablithus bijugatus holococcolith (late Paleocene, Sample TDP16B/12-2, 9 cm). (I–L) Collapsed coccospheres. (I) Cruciplacolithus inseadus, a species only previously known from the Danian but found in all our SEM samples (late Paleocene, Sample TDP16B/12-2, 9 cm). (J) Neochiastozygus imbriei grouping, most likely a collapsed coccosphere, but displays signifi cant morphological variation between coccoliths, with signifi cant implications for species-level taxonomy (late Paleocene, Sample TDP14/9-1, 20 cm). (K) Calciosolenia brasiliensis displaying morphological variation between coccoliths identical to that seen in modern examples (e.g., Young et al., 2003) (middle Eocene, Sample TDP20/23-1, 40 cm). (L) Undescribed, very small (<2 µm) coccolith spe-cies (late Paleocene, Sample TDP16B/12-2, 9 cm). (M) Coccolithus pelagicus with previously undescribed, gracile central-area cross bars (late Paleocene, Sample TDP16B/12-2, 9 cm). (N) Blackites defl andrei (middle Eocene, Sample TDP13/20-1, 50 cm). (O) Blackites morionum showing an intricately constructed, hollow spine and dissolution-susceptible rim architecture (late Paleocene, Sample TDP16B/12-2, 9 cm). (P) Undescribed placolith coccolith with a fragile, central-area grill (late Paleocene, Sample TDP16B/12-2, 9 cm).
Bown et al.
8 Geological Society of America Bulletin, January/February 2008
no unequivocal documentation of deep-dwell-ing nannoplankton prior to the Neogene, and of the modern assemblage, only Florisphaera has a fossil record, stretching back to the late Mio-cene (Young, 1998). We have not observed Flo-risphaera in the Kilwa Group, suggesting that it evolved in the late Oligocene or Miocene.
The presence of abundant Gladiolithus is sig-nifi cant, not only because it confi rms the unique quality of preservation in the Kilwa Group but also because it indicates that a deep–photic-zone niche was exploited by the same group at least back to the late Paleocene (56 Ma). It also lends strong support to the interpretation of these depositional environments as being open ocean and deep water. Modern Gladiolithus are uncommon in shelf seas, and the deep–photic-zone community abundance is strongly corre-lated with water depth and excluded from mar-ginal basins (Okada, 1983).
Small Taxa, Delicate Structures, and Coccospheres
The occurrence of small coccoliths and frag-ile, central-area structures, such as those seen
in Calciosolenia and Syracosphaera (Figs. 3F and 3K), represents preservation that resembles well-preserved modern coccolithophore mate-rial. The presence of coccospheres provides additional paleobiological information that is lost when coccoliths are disaggregated. Gen-erally, only placolith coccoliths that mechani-cally interlock are found preserved as intact coccospheres in the fossil record (Figs. 2I, 3I, and 3L), while all other fossil taxa are virtu-ally unknown in this state. The Kilwa Group has yielded the only Cenozoic examples of undisturbed, collapsed coccospheres of non-placolith taxa, providing indications of origi-nal cell size, coccolith production per cell, and ranges of intraspecifi c morphological variabil-ity (Figs. 3A, 3C, 3E, 3J, and 3K).
Preservation of the Kilwa Group Microfossils
Preservation of the principal calcareous microfossil groups (planktonic foraminifera, benthic foraminifera, and calcareous nanno-plankton) can be affected by the initial degree of
shell calcifi cation and postmortem taphonomic and diagenetic processes, including bioturba-tion, erosion, dissolution, recrystallization, and overgrowth. The planktonic groups live high in the water column (0–200 m) and are exported to the seafl oor by simple sinking, in the case of foraminifera (Berger, 1971), or within marine snow aggregates and zooplanktonic fecal pel-lets, in the case of the smaller nannoplankton (Steinmetz, 1994). Much is known about the dissolution of planktonic foraminifera as they approach the lysocline and sink beneath the cal-cite compensation depth (Thunell and Honjo, 1981; Schmuker and Schiebel, 2002), but in shallower settings, the death assemblages of planktonic foraminifera are relatively faithful
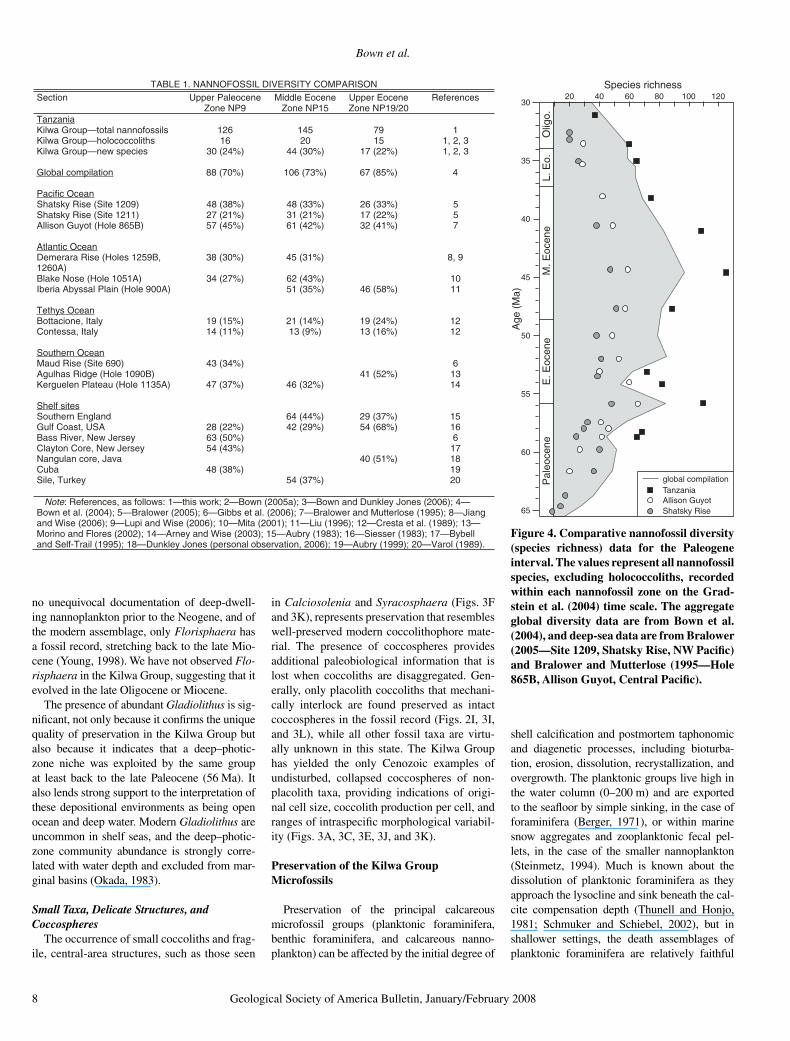
TABLE 1. NANNOFOSSIL DIVERSITY COMPARISON enecoelaP reppU noitceS
Zone NP9 Middle Eocene
Zone NP15 Upper EoceneZone NP19/20
References
Tanzania Kilwa Group—total nannofossils 126 145 79 1 Kilwa Group—holococcoliths 16 20 15 1, 2, 3 Kilwa Group—new species 30 (24%) 44 (30%) 17 (22%) 1, 2, 3
Global compilation 88 (70%) 106 (73%) 67 (85%) 4
Pacific Ocean Shatsky Rise (Site 1209) 48 (38%) 48 (33%) 26 (33%) 5 Shatsky Rise (Site 1211) 27 (21%) 31 (21%) 17 (22%) 5 Allison Guyot (Hole 865B) 57 (45%) 61 (42%) 32 (41%) 7
Atlantic Ocean Demerara Rise (Holes 1259B, 1260A)
38 (30%) 45 (31%) 8, 9
Blake Nose (Hole 1051A) 34 (27%) 62 (43%) 10 Iberia Abyssal Plain (Hole 900A) 51 (35%) 46 (58%) 11
Tethys Ocean
21 )%42( 91 )%41( 12 )%51( 91 ylatI ,enoicattoB 21 )%61( 31 )%9( 31 )%11( 41 ylatI ,assetnoC
Southern Ocean Maud Rise (Site 690) 43 (34%) 6 Agulhas Ridge (Hole 1090B) 41 (52%) 13
41 )%23( 64 )%73( 74 )A5311 eloH( uaetalP neleugreK
Shelf sites 51 )%73( 92 )%44( 46 dnalgnE nrehtuoS 61 )%86( 45 )%92( 24 )%22( 82 ASU ,tsaoC fluG
Bass River, New Jersey 63 (50%) 6 Clayton Core, New Jersey 54 (43%) 17
81 )%15( 04 avaJ ,eroc nalugnaN 91 )%83( 84 abuC 02 )%73( 45 yekruT ,eliS
Note: References, as follows: 1—this work; 2—Bown (2005a); 3—Bown and Dunkley Jones (2006); 4—Bown et al. (2004); 5—Bralower (2005); 6—Gibbs et al. (2006); 7—Bralower and Mutterlose (1995); 8—Jiang and Wise (2006); 9—Lupi and Wise (2006); 10—Mita (2001); 11—Liu (1996); 12—Cresta et al. (1989); 13—Morino and Flores (2002); 14—Arney and Wise (2003); 15—Aubry (1983); 16—Siesser (1983); 17—Bybell and Self-Trail (1995); 18—Dunkley Jones (personal observation, 2006); 19—Aubry (1999); 20—Varol (1989).
60 80 10020 40 120
55
50
65
60
35
30
45
40
Species richness
global compilationTanzania
Shatsky RiseAllison Guyot
Age
(M
a)
Pal
eoce
neE
. Eoc
ene
M. E
ocen
eL.
Eo.
Olig
o.Figure 4. Comparative nannofossil diversity (species richness) data for the Paleogene interval. The values represent all nannofossil species, excluding holococcoliths, recorded within each nannofossil zone on the Grad-stein et al. (2004) time scale. The aggregate global diversity data are from Bown et al. (2004), and deep-sea data are from Bralower (2005—Site 1209, Shatsky Rise, NW Pacifi c) and Bralower and Mutterlose (1995—Hole 865B, Allison Guyot, Central Pacifi c).

Tanzania microfossil Konservat-Lagerstätte
Geological Society of America Bulletin, January/February 2008 9
recorders of the overlying living plankton (Bé, 1977). However, in the modern ocean, calcar-eous nannoplankton are subject to far stronger taphonomic biases that signifi cantly reduce the exported and preserved diversity. This bias is highly correlated with coccolith size, and there appears to be a threshold in preservation poten-tial at 3 µm: 90% of the species with coccoliths >3 µm are found as fossils, compared with only 20% of those with coccoliths <3 µm (Young et al., 2005). This is not direct size selection, but rather the result of small coccoliths having higher surface-area-to-volume ratios, which increases their vulnerability to dissolution. Sed-iment trap and seafl oor samples show that the loss of small coccoliths takes place largely in the water column and is the result of grazing and/or dissolution while sinking, even well above the lysocline (Roth, 1994; Andruleit et al., 2004). Further selective dissolution and fragmenta-tion occurs within the sediment through inges-tion by sediment grazers and early diagenesis. With burial, diagenetic processes continue, and it is commonplace to observe deterioration of preservation with increasing depth in deep-sea cores. In carbonate-rich oozes, this involves increase in crystal size at the micron scale, with small crystals being selectively dissolved and larger ones overgrown (Wise, 1977). The net result of the various processes occurring in the water column, at the sediment surface, and dur-ing burial, is that even soft oozes are increas-ingly dominated by larger coccoliths. When the modern, global nannoplankton diversity is com-pared with the Holocene fossil record, the esti-mated preserved diversity is, at best, 54% but more typically around 30%. Preserved diversity is even less, if holococcolith morphologies are considered (20%–36%) (Young et al., 2003, 2005). These are signifi cantly high diversity losses that have serious implications for paleon-tological studies.
The exceptional preservation of calcareous nannofossils in the Paleogene Kilwa Group has resulted in assemblages that contain extraordi-narily high species richness, comprising new diversity in well-known families, alongside pres-ervation of small and delicate forms for which we have had no previous fossil record. The majority of this enhanced diversity is explained by preservation rather than paleoecology, and demonstrates the signifi cant effect of favorable taphonomic conditions. The same sediments host planktonic foraminifera assemblages that are not exceptionally diverse but which yield stable isotope values that are considered relatively unaffected by diagenesis. The glassy foraminifera tests, absence of infi lling, and pri-mary wall fabrics contrast with deep-sea ooze taphonomy, which is characterized by frosty or
white and chalky shells that are considered to result from recrystallization that includes both replacement and overgrowth/infi lling (Pearson et al., 2001; Sexton et al., 2006). Post-deposi-tional recrystallization arguably shifts the iso-topic values toward early diagenetic calcite and inferred seafl oor-environment values that are both colder, in terms of estimated paleotemper-atures, and more homogeneous (Pearson et al., 2001, 2007). The remarkable preservation of the calcareous nannofossils strongly corroborates this interpretation of minimal diagenetic modi-fi cation. The contrasting diversity records of the two microfossil groups, however, highlights the greater sensitivity to preservational modifi cation shown by the smaller-sized nannoplankton.
The quality of preservation is best explained by the clay-rich lithologies that have not been deeply buried. The clays isolate the calcite micro-fossils tests within an impermeable medium, preventing or inhibiting diagenetic recrystalliza-tion. This explanation is supported by excellent organic biomarker preservation that indicates thermal immaturity and low sediment perme-ability, which has inhibited organic matter bio-degradation (van Dongen et al., 2006). There is some variability in the preservation, from sample to sample and even across single SEM samples at the micron scale. Most likely, this refl ects hetero-geneous microenvironments within the sedimen-tary fabric, controlled by variations in grain size, porosity, permeability, and sediment chemistry.
Quantifying the Effects of Calcareous Nannofossil Preservation
Although the potential for preservational modifi cation of microfossil assemblages is universally acknowledged, the documenta-tion of preservation is inconsistent. Two main approaches have been used to record microfossil preservation—fi rst, qualitative, visual observa-tions, and second, indices based on indirect evi-dence, such as fragmentation, dissolution-sus-ceptibility rankings, and abundance comparisons (Berger, 1968; Roth and Thierstein, 1972; Roth and Krumbach, 1986; Le and Shackleton, 1992; Boeckel et al., 2006). More recently, geochemi-cal comparisons between different preservational classes of planktonic foraminifera have been attempted (Sexton et al., 2006). Visual assess-ment is largely subjective and greatly infl uenced by worker experience. Dissolution indices have been successful in foraminiferal studies but are not universally applied, and are rarely used in nannofossil studies (Matsuoka, 1990; Gibbs et al., 2004). Furthermore, both approaches may still fail to discriminate cryptic preservational effects that nevertheless signifi cantly alter both the taxonomic and geochemical composition of
a microfossil assemblage (Gibbs et al., 2004; Williams et al., 2005; Pearson et al., 2007).
To assess the preserved diversity of Paleogene nannofossils, we have used the Kilwa Group data as a benchmark against which to compare recorded diversities from coeval sections, repre-senting a range of preservation states, for three time slices (Table 1). It is striking that the global compilation returns lower diversities than the Kilwa Group (70%–85%), but the values are broadly comparable, given the uncertainties associated with composite literature surveys (Bown et al., 2004). The individual sections yield diversity values that are, in all cases, con-siderably less (9%–68%) than the Kilwa Group for each of the three time slices, but there are systematic discrepancies corresponding to sec-tion type. The Paleogene shelf sites, with report-edly good preservation (Clayton core—Bybell and Self-Trail, 1995; Bass River—Gibbs et al., 2006; Yazoo Clay, Gulf Coast—Siesser, 1983), host diversities ranging from 43%–68% and compare most favorably with the Kilwa Group values. Deep-sea sections, where carbonate-rich oozes and chalks dominate, yield values ranging from 21%–58%, while the lowest values (9%–24%) come from lithifi ed deepwater limestone sequences (Contessa, Italy). These sections are not all directly comparable, in particular those from higher latitudes (e.g., the Southern Ocean); however, the Eocene was a time of relatively low nannoplankton biogeographic differentiation, and its effects do not greatly bias the data (e.g., only nine species were absent from the Kilwa Group succession due to biogeography). Indeed, the Southern Ocean sites return relatively high diversity values, most likely refl ecting better preservation in more clay-rich lithologies.
The negative aspects of carbonate-rich lithol-ogies on the taphonomy of calcareous nannofos-sils are reasonably well known. However, there has been no serious attempt to quantify these effects, and the degree of taxonomic modifi -cation, highlighted in this paper, is probably greater than is generally perceived. This does not usually impact on the stratigraphic applica-tion of the group, which generally utilizes taxa selected for size and robustness, but it does have serious implications when assemblage abun-dance and diversity data are considered. That the least favorable diversity comparisons come from the carbonate-rich successions of tropical and subtropical latitudes is comparable to the morphological- and geochemical-based tapho-nomic observations from the study of planktonic foraminifera (Sexton et al., 2006).
The potential for large-magnitude nanno-fossil diversity loss is a signifi cant factor for those using fossil data in paleobiological or paleoceanographic interpretation. These losses
Bown et al.
10 Geological Society of America Bulletin, January/February 2008
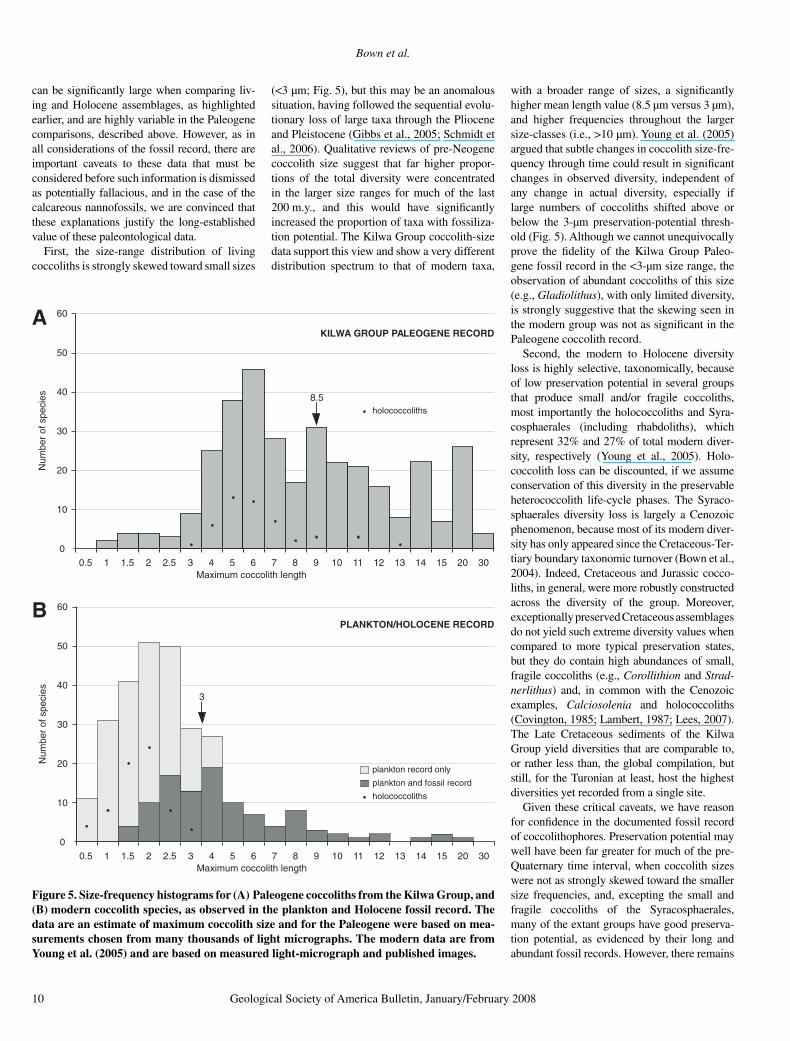
can be signifi cantly large when comparing liv-ing and Holocene assemblages, as highlighted earlier, and are highly variable in the Paleogene comparisons, described above. However, as in all considerations of the fossil record, there are important caveats to these data that must be considered before such information is dismissed as potentially fallacious, and in the case of the calcareous nannofossils, we are convinced that these explanations justify the long-established value of these paleontological data.
First, the size-range distribution of living coccoliths is strongly skewed toward small sizes
(<3 µm; Fig. 5), but this may be an anomalous situation, having followed the sequential evolu-tionary loss of large taxa through the Pliocene and Pleistocene (Gibbs et al., 2005; Schmidt et al., 2006). Qualitative reviews of pre-Neogene coccolith size suggest that far higher propor-tions of the total diversity were concentrated in the larger size ranges for much of the last 200 m.y., and this would have signifi cantly increased the proportion of taxa with fossiliza-tion potential. The Kilwa Group coccolith-size data support this view and show a very different distribution spectrum to that of modern taxa,
with a broader range of sizes, a signifi cantly higher mean length value (8.5 µm versus 3 µm), and higher frequencies throughout the larger size-classes (i.e., >10 µm). Young et al. (2005) argued that subtle changes in coccolith size-fre-quency through time could result in signifi cant changes in observed diversity, independent of any change in actual diversity, especially if large numbers of coccoliths shifted above or below the 3-µm preservation-potential thresh-old (Fig. 5). Although we cannot unequivocally prove the fi delity of the Kilwa Group Paleo-gene fossil record in the <3-µm size range, the observation of abundant coccoliths of this size (e.g., Gladiolithus), with only limited diversity, is strongly suggestive that the skewing seen in the modern group was not as signifi cant in the Paleogene coccolith record.
Second, the modern to Holocene diversity loss is highly selective, taxonomically, because of low preservation potential in several groups that produce small and/or fragile coccoliths, most importantly the holococcoliths and Syra-cosphaerales (including rhabdoliths), which represent 32% and 27% of total modern diver-sity, respectively (Young et al., 2005). Holo-coccolith loss can be discounted, if we assume conservation of this diversity in the preservable heterococcolith life-cycle phases. The Syraco-sphaerales diversity loss is largely a Cenozoic phenomenon, because most of its modern diver-sity has only appeared since the Cretaceous-Ter-tiary boundary taxonomic turnover (Bown et al., 2004). Indeed, Cretaceous and Jurassic cocco-liths, in general, were more robustly constructed across the diversity of the group. Moreover, exceptionally preserved Cretaceous assemblages do not yield such extreme diversity values when compared to more typical preservation states, but they do contain high abundances of small, fragile coccoliths (e.g., Corollithion and Strad-nerlithus) and, in common with the Cenozoic examples, Calciosolenia and holococcoliths (Covington, 1985; Lambert, 1987; Lees, 2007). The Late Cretaceous sediments of the Kilwa Group yield diversities that are comparable to, or rather less than, the global compilation, but still, for the Turonian at least, host the highest diversities yet recorded from a single site.
Given these critical caveats, we have reason for confi dence in the documented fossil record of coccolithophores. Preservation potential may well have been far greater for much of the pre-Quaternary time interval, when coccolith sizes were not as strongly skewed toward the smaller size frequencies, and, excepting the small and fragile coccoliths of the Syracosphaerales, many of the extant groups have good preserva-tion potential, as evidenced by their long and abundant fossil records. However, there remains
20 300.5 1 1.5 2 2.5 3 4 5 6 7 8 9 10 11 12 13 1514
0
10
20
30
40
50
60
Maximum coccolith length
Num
ber
of s
peci
es
20 300.5 1 1.5 2 2.5 3 4 5 6 7 8 9 10 11 12 13 1514
0
10
20
30
40
50
60
Maximum coccolith length
Num
ber
of s
peci
es 8.5
3
PLANKTON/HOLOCENE RECORD
KILWA GROUP PALEOGENE RECORD
plankton record only
plankton and fossil record
holococcoliths
holococcoliths
A
B
Figure 5. Size-frequency histograms for (A) Paleogene coccoliths from the Kilwa Group, and (B) modern coccolith species, as observed in the plankton and Holocene fossil record. The data are an estimate of maximum coccolith size and for the Paleogene were based on mea-surements chosen from many thousands of light micrographs. The modern data are from Young et al. (2005) and are based on measured light-micrograph and published images.

Tanzania microfossil Konservat-Lagerstätte
Geological Society of America Bulletin, January/February 2008 11
much to learn about the preservation of cocco-liths, and fi ne-fraction carbonate in general, and a need to develop protocols that allow for the adequate description, quantifi cation, and com-munication of this essential information. These issues are being addressed in the foraminifera and geochemistry communities, which, by and large, accept that qualitative, descriptive meth-ods of conveying preservation quality are no longer adequate (Pearson et al., 2001; Sexton et al., 2006). Instead, strict criteria that require high-resolution morphological analysis, or indi-rect geochemical methods, are being used to ensure effective documentation of preservation.
CONCLUSIONS
The Paleogene Kilwa Group sediments of coastal Tanzania host calcareous microfossils that are exceptionally preserved. The quality of preservation is demonstrated by the glassy appearance, wall ultrastructure, and stable isotope geochemistry of the planktonic fora-minifera, and high diversity of the calcareous nannofossils, which includes very small and fragile taxa. For both groups, this fossil material resembles well-preserved modern specimens. The glassy foraminifera shells provide stable isotope values that have been relatively unaf-fected by diagenesis and are providing valu-able new paleoclimate proxy records (Pearson et al., 2007). The taxonomic composition of these foraminiferal assemblages, however, is comparable to those from different taphonomic settings. By contrast, the calcareous nannofos-sils are remarkably diverse and have distinct assemblage compositions that are primarily the result of the exceptional preservation. They are characterized by the presence of dissolu-tion-susceptible and fragile taxa, in particular holococcoliths and rhabdoliths. The uniqueness of this Konservat-Lagerstätte is especially well demonstrated by the abundant occurrence of Gladiolithus, which is a delicate extant taxon that, until now, has had no documented fossil record prior to the Pleistocene.
For both foraminifera and coccolithophores, incorporation into impermeable, clay-rich sediments that have never been deeply buried appears to have been critical in producing the exceptional preservation. The enhanced diversity seen in the calcareous nannofossils highlights the different sensitivities of these two fossil groups to preservational modifi cation. The inte-grated taphonomic observations from both fossil groups, however, provide the maximum amount of information in support of the interpretation of both geochemical and paleontological proxies.
The Kilwa Group calcareous nannofossil diversities are consistently higher than all coeval
assemblages, and even slightly higher than com-posite global estimates. These comparisons dem-onstrate the degree of taxonomic modifi cation that can result from varying preservation states. The highest diversities are preserved in hemipe-lagic, clay-rich lithologies and the greatest losses occur in lithifi ed, carbonate-rich sediments. The majority of the lost diversity, however, is con-fi ned to distinct taxonomic groups, and espe-cially the holococcoliths and rhabdoliths (Syra-cosphaerales). The preservational potential of Paleogene coccolithophores may well have been signifi cantly greater than in the modern oceans because a larger proportion of the biodiversity fell within the larger size fractions.
Study of the Kilwa Group hemipelagic sedi-ments has highlighted the signifi cant effects that preservation can have on both the diversity and geochemistry of calcareous microfossils. These exceptionally preserved fossils are providing high-quality paleontological and geochemical paleoclimate proxy information, and, for the calcareous nannoplankton, this includes paleo-biological and biodiversity data that are cur-rently unique for this fossil group.
ACKNOWLEDGMENTS
We thank the Tanzania Commission for Science and Technology for permission to conduct this study, the Tanzania Petroleum Development Corporation for fi eldwork support, and the Natural Environment Research Council (Grant NE/C510508/1) and Uni-versity College London Graduate School for funding the research.
REFERENCES CITED
Andruleit, H., Rogalla, U., and Stäger, S., 2004, From liv-ing communities to fossil assemblages: Origin and fate of coccolithophorids in the northern Arabian Sea: Micropaleontology, v. 50, supplement 1, p. 5–21, doi: 10.2113/50.Suppl_1.5.
Arney, J.E., and Wise, S.W., 2003, Paleocene-Eocene nan-nofossil biostratigraphy of ODP Leg 183, Kerguelen Plateau: Proceedings of the Ocean Drilling Program, Scientifi c Results, v. 183, p. 1–59.
Aubry, M.-P., 1983, Biostratigraphie du Paléogène épiconti-nental de l’Europe du Nord-Ouest. Étude fondée sur les nannofossiles calcaires: Documents des Laboratoires de Géologie Lyon, no. 89, p. 1–317.
Aubry, M.-P., 1999, Late Paleocene–Early Eocene sedimen-tary history in western Cuba: Implications for the LPTM and for regional tectonic history: Micropaleontology, v. 45, supplement 2, p. 5–18, doi: 10.2307/1486101.
Bé, A.W.H., 1977, Recent planktonic foraminifera, in Ram-say, A.T.S., ed., Oceanic micropalaeontology, v. 1: London, Academic Press, p. 1–100.
Berger, W.H., 1968, Planktonic foraminifera: Selective solution and paleoclimatic interpretation: Deep-Sea Research, v. 15, p. 31–43.
Berger, W.H., 1971, Sedimentation of planktonic formin-ifera: Marine Geology, v. 11, p. 325–358, doi: 10.1016/0025-3227(71)90035-1.
Berggren, W.A., and Pearson, P.N., 2005, A revised tropi-cal to subtropical Paleogene planktonic foraminiferal zonation: Journal of Foraminiferal Research, v. 35, p. 279–298, doi: 10.2113/35.4.279.
Bice, K.L., Huber, B.T., and Norris, R.D., 2003, Extreme polar warmth during the Cretaceous greenhouse?: Paradox of the late Turonian δ18O record at Deep Sea
Drilling Project site 511: Paleoceanography, v. 18, p. 1031, doi: 10.1029/2002PA000848.
Boeckel, B., Baumann, K.-H., Henrich, R., and Kinkel, H., 2006, Coccolith distribution patterns in South Atlantic and Southern Ocean surface sediments in relation to environmental gradients: Deep-sea Research: Part I, Oceanographic Research Papers, v. 53, p. 1073–1099, doi: 10.1016/j.dsr.2005.11.006.
Bown, P.R., 2005a, Palaeogene calcareous nannofossils from the Kilwa and Lindi areas of coastal Tanzania (Tanza-nia Drilling Project 2003-4): Journal of Nannoplankton Research, v. 27, p. 21–95.
Bown, P.R., 2005b, Calcareous nannofossil biostratigraphy of the Cenozoic of Leg 198 Site 1208, Shatsky Rise, northwest Pacifi c Ocean, in Bralower, T.J., et al., eds., Proceedings of the Ocean Drilling Program, scientifi c results: v. 198 [Online] http://www-odp.tamu.edu/ publications/198_SR/104/104.htm.
Bown, P.R., and Dunkley Jones, T., 2006, New Paleogene calcareous nannofossil taxa from coastal Tanzania: Tanzania Drilling Project Sites 11 to 14: Journal of Nannoplankton Research, v. 28, p. 17–34.
Bown, P.R., and Young, J.R., 1998, Techniques, in Bown, P.R., ed., Calcareous nannofossil biostratigraphy: Lon-don, Kluwer Academic, p. 16–28.
Bown, P.R., Lees, J.A., and Young, J.R., 2004, Calcareous nannoplankton evolution and diversity through time, in Thierstein, H., and Young, J.R., eds., Coccolitho-phores—From molecular processes to global impact: Berlin, Springer-Verlag, p. 481–508.
Bralower, T.J., 2005, Paleocene–Early Oligocene calcare-ous nannofossil biostratigraphy, ODP Leg 198 Sites 1209, 1210, and 1211 (Shatsky Rise, Pacifi c Ocean), in Bralower, T.J., et al., eds., Proceedings of the Ocean Drilling Program, scientifi c results: v. 198 [Online] http://www-odp.tamu.edu/publications/198_SR/115/115.htm.
Bralower, T.J., and Mutterlose, J., 1995, Calcareous nan-nofossil biostratigraphy of Site 865, Allison Guyot, Central Pacifi c Ocean: A tropical Paleogene reference section: Proceedings of the Ocean Drilling Program, Scientifi c Results, v. 143, p. 31–74.
Briggs, D.E.G., 2001, Exceptionally preserved fossils, in Briggs, D.E.G., and Crowther P.R., eds., Palaeobiology 2: Oxford, Blackwell Publishing, p. 328–332.
Bybell, L.M., and Self-Trail, J.M., 1995, Evolutionary, biostratigraphic, and taxonomic study of calcareous nannofossils from the continuous Paleocene-Eocene boundary section in New Jersey: U.S. Geological Sur-vey Professional Paper, v. 1554, p. 1–36.
Covington, M., 1985, New morphologic information on Cretaceous nannofossils from the Niobrara Forma-tion (Upper Cretaceous) of Kansas: Geology, v. 13, p. 683–686, doi: 10.1130/0091-7613(1985)13<683:NMIOCN>2.0.CO;2.
Cresta, S., Monechi, S., and Parisi, G., 1989, Mesozoic-Ceno-zoic stratigraphy in the Umbria-Marche area: Memorie descrittive della carta geologica d’Italia, v. 39, p. 1–185.
Gibbs, S., Shackleton, N.J., and Young, J.R., 2004, Identi-fi cation of dissolution patterns in nannofossil assem-blages: A high-resolution comparison of synchronous records from Ceara Rise, ODP Leg 154: Paleoceanog-raphy, v. 19, p. PA1029, doi: 10.1029/2003PA000958.
Gibbs, S., Young, J.R., Bralower, T.J., and Shackleton, N.J., 2005, Nannofossil evolutionary events in the mid Plio-cene: An assessment of the degree of synchrony in the extinction of Reticulofenestra pseudoumbilicus and Sphenolithus abies: Palaeogeography, Palaeoclimatol-ogy, Palaeoecology, v. 217, p. 155–172, doi: 10.1016/j.palaeo.2004.11.005.
Gibbs, S.J., Bown, P.R., Sessa, J.A., Bralower, T.J., and Wilson, P.A., 2006, Nannoplankton origination and extinction across the Paleocene-Eocene Ther-mal Maximum: Science, v. 314, p. 1770–1773, doi: 10.1126/ science.1133902.
Gradstein, F.M., Ogg, J.G., and Smith, A.G., 2004, A geo-logical timescale 2004: Cambridge, Cambridge Uni-versity Press, 610 p.
Jiang, S., and Wise, S.W., 2006, Surface-water chemistry and fertility variations in the tropical Atlantic across the Paleocene/Eocene Thermal Maximum as evi-denced by calcareous nannoplankton from ODP Leg 207, Hole 1259B: Revue de Micropaléontology, v. 49, p. 227–244, doi: 10.1016/j.revmic.2006.10.002.

Bown et al.
12 Geological Society of America Bulletin, January/February 2008
Kleijne, A., 1991, Holococcolithophorids from the Indian Ocean, Red Sea, Mediterranean and North Atlantic Ocean: Marine Micropaleontology, v. 17, p. 1–76, doi: 10.1016/0377-8398(91)90023-Y.
Lambert, B., 1987, Nannofossiles calcaires de l’Albien supérieur et du Vraconnien du Cameroun méridional: Cahiers de Micropaléontologie, v. 2, p. 33–96.
Le, J., and Shackleton, N.J., 1992, Carbonate dissolution fl uctuations in the western equatorial Pacifi c during the late Quaternary: Paleoceanography, v. 7, p. 21–42.
Lees, J.A., 2007, New and rarely reported calcareous nannofos-sils from the Late Cretaceous of coastal Tanzania: Out-crop samples and Tanzania Drilling Project Sites 5, 9 and 15: Journal of Nannoplankton Research, v. 29, p. 39–65.
Liu, L., 1996, Eocene calcareous nannofossils from the Ibe-ria Abyssal Plain: Proceedings of the Ocean Drilling Program, Scientifi c Results, v. 149, p. 61–78.
Lupi, C., and Wise, S.W., 2006, Calcareous nannofossil bio-stratigraphic framework for middle Eocene sediments from ODP Hole 1260A, Demerara Rise: Revue de Micropaléontology, v. 49, p. 245–253, doi: 10.1016/j.revmic.2006.10.001.
Martini, E., 1971, Standard Tertiary and Quaternary calcare-ous nannoplankton zonation, in Farinacci, A., ed., Pro-ceedings of the Second Planktonic Conference, Roma 1970, v. 2: Rome, Edizioni Tecnoscienza, p. 739–785.
Matsuoka, H., 1990, A new method to evaluate dissolution of calcium carbonate in deep sea sediments: Palaeon-tological Society of Japan: Transactions and Proceed-ings, v. 157, p. 430–434.
Mita, I., 2001, Data report: Early to Late Eocene calcareous nannofossil assemblages of sites 1051 and 1052, Blake Nose, northwestern Atlantic Ocean: Proceedings of the Ocean Drilling Program, Scientifi c Results, v. 171B, p. 1–28.
Morino, M., and Flores, J.A., 2002, Data report: Calcareous nannofossil data from the Eocene to Oligocene, Leg 177, Site 1090B: Proceedings of the Ocean Drilling Program, Scientifi c Results, v. 177, p. 1–9.
Nicholas, C.J., Pearson, P.N., Bown, P.R., Dunkley Jones, T., Huber, B.T., Karega, A., Lees, J.A., McMillan, I.K., O’Halloran, A., Singano, J.M., and Wade, B.S., 2006, Stratigraphy and sedimentology of the Upper Cre-taceous to Paleogene Kilwa Group, southern coastal Tanzania: Journal of African Earth Sciences, v. 45, p. 431–466, doi: 10.1016/j.jafrearsci.2006.04.003.
Okada, H., 1983, Modern nannofossil assemblages in sedi-ments of coastal and marginal seas along the western Pacifi c Ocean: Utrecht Micropaleontological Bulletins, v. 30, p. 171–187.
Okada, H., and Matsuoka, M., 1996, Lower-photic nanno-fl ora as an indicator of the late Quaternary monsoonal paleo-record in the tropical Indian Ocean, in Mogui-levsky, A., and Whatley, R., eds., Microfossils and oce-anic environments: Aberystwyth, University of Wales, Aberystwyth Press, p. 231–245.
Pearson, P.N., Ditchfi eld, P.W., Singano, J.M., Harcourt-Brown, K.G., Nicholas, C.J., Olsson, R.K., Shackleton,
N.J., and Hall, M.A., 2001, Warm tropical sea surface temperatures in the Late Cretaceous and Eocene epochs: Nature, v. 413, p. 481–487, doi: 10.1038/35097000.
Pearson, P.N., Nicholas, C.J., Singano, J.M., Bown, P.R., Coxall, H.K., van Dongen, B.E., Huber, B.T., Karega, A., Lees, J.A., Msaky, E., Pancost, R.D., Pearson, M., and Roberts, A.P., 2004, Paleogene and Cretaceous sediment cores from the Kilwa and Lindi areas of coastal Tanzania: Tanzania Drilling Project Sites 1 to 5: Journal of African Earth Sciences, v. 39, p. 25–62, doi: 10.1016/j.jafrearsci.2004.05.001.
Pearson, P.N., Nicholas, C.J., Singano, J.M., Bown, P.R., Coxall, H.K., van Dongen, B.E., Huber, B.T., Karega, A., Lees, J.A., MacLeod, K., McMillan, I.K., Pancost, R.D., Pearson, M., and Msaky, E., 2006, Tanzania Drill-ing Project Sites 6 to 10: Further Paleogene and Cre-taceous sediment cores from the Kilwa area of coastal Tanzania: Journal of African Earth Sciences, v. 45, p. 279–317, doi: 10.1016/j.jafrearsci.2006.02.005.
Pearson, P.N., van Dongen, B.E., Nicholas, C.J., Pancost, R.D., Schouten, S., Singano, J.M., and Wade, B.S., 2007, Stable warm tropical climate through the Eocene epoch: Geology, v. 35, p. 211–214, doi: 10.1130/G23175A.1.
Roth, P.H., 1994, Distribution of coccoliths in oceanic sedi-ments, in Winter, A., and Siesser, W.G., eds., Cocco-lithophores: Cambridge, Cambridge University Press, p. 199–218.
Roth, P.H., and Berger, W.H., 1975, Distribution and dis-solution of coccoliths in the south and central Pacifi c: Cushman Foundation for Foraminiferal Research: Spe-cial Publication, v. 13, p. 87–113.
Roth, P.H., and Krumbach, K.R., 1986, Middle Cretaceous calcareous nannofossil biogeography and preservation in the Atlantic and Indian oceans: Implications for palaeoceanography: Marine Micropaleontology, v. 10, p. 235–266, doi: 10.1016/0377-8398(86)90031-9.
Roth, P.H., and Thierstein, H., 1972, Calcareous nanno-plankton: Leg 14 of the Deep Sea Drilling Project: Initial Reports of the Deep Sea Drilling Project, v. 14, p. 421–485.
Schmidt, D., Lazarus, D., Young, J.R., and Kucera, M., 2006, Biogeography and evolution of body size in marine plankton: Earth-Science Reviews, v. 78, p. 239–266, doi: 10.1016/j.earscirev.2006.05.004.
Schmuker, B., and Schiebel, R., 2002, Planktic foraminifers and hydrography of the eastern and northern Caribbean Sea: Marine Micropaleontology, v. 46, p. 387–403, doi: 10.1016/S0377-8398(02)00082-8.
Sexton, P.F., Wilson, P.A., and Pearson, P.N., 2006, Micro-structural and geochemical perspectives on planktic foraminiferal preservation: “Glassy” versus “frosty”: Geochemistry, Geophysics, Geosystems, v. 7, Q12P19, doi: 10.1029/2006GC001291.
Siesser, W.G., 1983, Paleogene calcareous nannoplankton biostratigraphy: Mississippi, Alabama and Tennes-see: Mississippi Bureau of Geology: Bulletin, v. 125, p. 1–61.
Steinmetz, J.C., 1994, Sedimentation of coccolithophores, in Winter, A., and Siesser, W.G., eds., Coccolithophores: Cambridge, Cambridge University Press, p. 179–197.
Stewart, D.R.M., Pearson, P.N., Ditchfi eld, P.W., and Sin-gano, J.M., 2004, Miocene tropical Indian Ocean temperatures: Evidence from three exceptionally pre-served foraminiferal assemblages from Tanzania: Jour-nal of African Earth Sciences, v. 40, p. 173–190, doi: 10.1016/j.jafrearsci.2004.09.001.
Thunell, R.C., and Honjo, S., 1981, Planktonic foraminiferal fl ux to the deep ocean: Sediment trap results from the tropical Atlantic and the Central Pacifi c: Marine Geol-ogy, v. 40, p. 237, doi: 10.1016/0025-3227(81)90142-0.
van Dongen, B.E., Talbot, H.M., Schouten, S., Pearson, P.N., and Pancost, R.D., 2006, Well-preserved Palaeogene and Cretaceous biomarkers from the Kilwa area, Tan-zania: Organic Geochemistry, v. 37, p. 539–557, doi: 10.1016/j.orggeochem.2006.01.003.
Varol, O., 1989, Eocene calcareous nannofossils from Sile (northwest Turkey): Revista Espanola de Micropaleon-tología, v. 21, p. 273–320.
Williams, M., Haywood, A.M., Claus-Dieter, H., and Wilkin-son, I.P., 2005, Effi cacy of δ18O data from Pliocene planktonic foraminifer calcite for spatial sea surface temperature reconstruction: Comparison with a fully coupled ocean-atmosphere GCM and fossil assemblage data for the mid-Pliocene: Geological Magazine, v. 142, p. 399–417, doi: 10.1017/S0016756805000828.
Wilson, P.A., Norris, R.D., and Cooper, M.J., 2002, Test-ing the Cretaceous greenhouse hypothesis using glassy foraminiferal calcite from the core of the Turonian tropics on Demerara Rise: Geology, v. 30, p. 607–610, doi: 10.1130/0091-7613(2002)030<0607:TTCGHU>2.0.CO;2.
Wise, S.W., 1977, Chalk formation: Early diagenesis, in Anderson, N.R., and Malahoff, A., eds., The fate of fos-sil fuel CO
2 in the oceans: New York, Plenum Publish-
ing, p. 717–739.Young, J.R., 1994, Functions of coccoliths, in Winter, A.,
and Siesser, W.G., eds., Coccolithophores: Cambridge, Cambridge University Press, p. 63–82.
Young, J.R., 1998, Neogene, in Bown, P.R., ed., Calcare-ous nannofossil biostratigraphy: London, Kluwer Aca-demic, p. 225–265.
Young, J.R., Geisen, M., Cros, L., Kleijne, A., Sprengel, C., Probert, I., and Ostergaard, J., 2003, A guide to extant coccolithophore taxonomy: Journal of Nannoplankton Research, v. 1, Special Issue, p. 1–125.
Young, J.R., Geisen, M., and Probert, I., 2005, A review of selected aspects of coccolithophore biology with impli-cations for paleodiversity estimates: Micropaleontology, v. 51, p. 267–288, doi: 10.2113/ gsmicropal.51.4.267.
MANUSCRIPT RECEIVED 11 MAY 2007REVISED MANUSCRIPT RECEIVED 13 AUGUST 2007MANUSCRIPT ACCEPTED 24 AUGUST 2007
Printed in the USA
Related Documents