A novel parthenin analog exhibits anti-cancer activity: Activation of apoptotic signaling events through robust NO formation in human leukemia HL-60 cells Ajay Kumar a , Fayaz Malik a , Shashi Bhushan a , Bhahwal A. Shah b , Subhash C. Taneja b , Harish C. Pal a , Zahoor A. Wani a , Dilip M. Mondhe a , Jagdeep Kaur c , Jaswant Singh a,⇑ a Division of Pharmacology, Indian Institute of Integrative Medicine (Council of Scientific and Industrial Research), Canal Road, Jammu Tawi 180001, India b Division of Bio-organic Chemistry, Indian Institute of Integrative Medicine (Council of Scientific and Industrial Research), Canal Road, Jammu Tawi 180001, India c Department of Biotechnology, Panjab University, Chandigarh 160014, India article info Article history: Received 29 January 2011 Received in revised form 8 June 2011 Accepted 15 June 2011 Available online 29 June 2011 Keywords: Parthenin analog (P19) Apoptosis Nitric oxide (NO) NAC NF-jB abstract This study describes the anti-cancer activity of P19, an analog of parthenin. P19 induced apoptosis in HL- 60 cells and inhibited cell proliferation with 48 h IC50 of 3.5 lM. At 10 mg/kg dose, it doubled the median survival time of L1210 leukemic mice and at 25 mg/kg it inhibited Ehrlich ascites tumor growth by 60%. Investigation of the mechanism of P19 induced apoptosis in HL-60 cells revealed that N-acetyl-L-cysteine (NAC) and s-methylisothiourea (sMIT) could reverse several molecular events that lead to cell death by inhibiting nitric oxide (NO) formation. It selectively produced massive NO in cells while quenching the basal ROS levels with concurrent elevation of GSH. P19 disrupted mitochondrial integrity leading to cyto- chrome c release and caspase-9 activation. P19 also caused caspase-8 activation by selectively elevating the expression of DR4 and DR5. All these events lead to the activation of caspase-3 leading to PARP-1 cleavage and DNA fragmentation. However, knocking down of AIF by siRNA also suppressed the apoptosis substantially thus indicating caspase independent apoptosis, too. Further, contrary to enhanced iNOS expression, its transcription factor, NF-jB (p65) was cleaved with a simultaneous increase in cytosolic IjB-alpha. In addition, P19 potently inhibited pro-survival proteins pSTAT3 and survivin. The multi- modal pro-apoptotic activity of P19 raises its potential usefulness as a promising anti-cancer therapeutic. Ó 2011 Elsevier Ireland Ltd. All rights reserved. 1. Introduction Cancer cells accumulate several genetic and epigenetic muta- tions to cleverly evade self demise by deregulating apoptosis and setting a stage for initiation, promotion and progression of the malignant growth [1]. Agents that can trigger the process of apop- tosis in cancer cells are therefore, considered potentially important for the development of anti-cancer chemotherapeutics [2]. In this concern, we focused our attention on parthenin, a major sesquiter- pene lactone from the obnoxious weed Parthenium hysterophorus Linn. (Asteraecae) that grows wild in different regions of India. The plant causes contact dermatitis and allergic rhinitis in animals [3]. Despite the fact that Parthenium is considered a toxic plant its traditional uses are amply reported in literature [4]. One of its chemical constituent parthenin is reported to be responsible for cytotoxic effects of this plant [5,6]. Parthenin has also been re- ported to exhibit anti-tumor activity in mice [7]. In view of the re- ported toxicity of parthenin we attempted to prepare various analogs of parthenin modifying its functional groups to obtain some potent anti-cancer analog of lesser toxicity. In this endeavor we screened several semi-synthetic analogs of parthenin primarily for their apoptotic index and selected P19 among them as the pro- spective pro-apoptotic candidate for its detailed studies to develop it into a promising anticancer therapeutic lead. About 50% of the cancers are known to have p53 mutated. Therefore, we employed p53 null, human acute myeloid leukemia HL-60 cells [8]. Thus, the agents that can induce apoptosis via a p53-independent path- way should have a broader therapeutic potential not only in AML patients but also in other cancer diseases. Several earlier studies have demonstrated that nitric oxide (NO) is an important signaling messenger that plays an important role in many physiological and pathological conditions. Endogenous NO is generated from L-arginine by three major types of NO synthases (NOS), i.e. endothelial NOS, neural NOS, and inducible NOS (iNOS) while another form of NO has recently been reported from 0009-2797/$ - see front matter Ó 2011 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.cbi.2011.06.006 Abbreviations: AIF, apoptosis inducing factor; AML, acute myeloid leukemia; DR4, death receptor 4; DR5, death receptor 5; DRP-1, dynamin related protein-1; GSH, glutathione reduced; iNOS, inducible nitric oxide synthase; NAC, N-acetyl-L- cysteine; NF-jB, nuclear factor-kappa B; PARP-1, poly (ADP ribose) polymerase-1; pSTAT3, phosphorylated signal transducer and activator of transcription protein-3; ROS, reactive oxygen species; sMIT, s-methylisothiourea; TNFR1, tumor necrosis factor receptor 1. ⇑ Corresponding author. Tel.: +91 191 2569000x291; fax: +91 191 2569333. E-mail address: [email protected] (J. Singh). Chemico-Biological Interactions 193 (2011) 204–215 Contents lists available at ScienceDirect Chemico-Biological Interactions journal homepage: www.elsevier.com/locate/chembioint

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Chemico-Biological Interactions 193 (2011) 204–215
Contents lists available at ScienceDirect
Chemico-Biological Interactions
journal homepage: www.elsevier .com/locate /chembioint
A novel parthenin analog exhibits anti-cancer activity: Activation of apoptoticsignaling events through robust NO formation in human leukemia HL-60 cells
Ajay Kumar a, Fayaz Malik a, Shashi Bhushan a, Bhahwal A. Shah b, Subhash C. Taneja b, Harish C. Pal a,Zahoor A. Wani a, Dilip M. Mondhe a, Jagdeep Kaur c, Jaswant Singh a,⇑a Division of Pharmacology, Indian Institute of Integrative Medicine (Council of Scientific and Industrial Research), Canal Road, Jammu Tawi 180001, Indiab Division of Bio-organic Chemistry, Indian Institute of Integrative Medicine (Council of Scientific and Industrial Research), Canal Road, Jammu Tawi 180001, Indiac Department of Biotechnology, Panjab University, Chandigarh 160014, India
a r t i c l e i n f o
Article history:Received 29 January 2011Received in revised form 8 June 2011Accepted 15 June 2011Available online 29 June 2011
Keywords:Parthenin analog (P19)ApoptosisNitric oxide (NO)NACNF-jB
0009-2797/$ - see front matter � 2011 Elsevier Irelandoi:10.1016/j.cbi.2011.06.006
Abbreviations: AIF, apoptosis inducing factor; AMDR4, death receptor 4; DR5, death receptor 5; DRP-1GSH, glutathione reduced; iNOS, inducible nitric oxidcysteine; NF-jB, nuclear factor-kappa B; PARP-1, polypSTAT3, phosphorylated signal transducer and activatROS, reactive oxygen species; sMIT, s-methylisothiofactor receptor 1.⇑ Corresponding author. Tel.: +91 191 2569000x29
E-mail address: [email protected] (J. Singh).
a b s t r a c t
This study describes the anti-cancer activity of P19, an analog of parthenin. P19 induced apoptosis in HL-60 cells and inhibited cell proliferation with 48 h IC50 of 3.5 lM. At 10 mg/kg dose, it doubled the mediansurvival time of L1210 leukemic mice and at 25 mg/kg it inhibited Ehrlich ascites tumor growth by 60%.Investigation of the mechanism of P19 induced apoptosis in HL-60 cells revealed that N-acetyl-L-cysteine(NAC) and s-methylisothiourea (sMIT) could reverse several molecular events that lead to cell death byinhibiting nitric oxide (NO) formation. It selectively produced massive NO in cells while quenching thebasal ROS levels with concurrent elevation of GSH. P19 disrupted mitochondrial integrity leading to cyto-chrome c release and caspase-9 activation. P19 also caused caspase-8 activation by selectively elevatingthe expression of DR4 and DR5. All these events lead to the activation of caspase-3 leading to PARP-1cleavage and DNA fragmentation. However, knocking down of AIF by siRNA also suppressed the apoptosissubstantially thus indicating caspase independent apoptosis, too. Further, contrary to enhanced iNOSexpression, its transcription factor, NF-jB (p65) was cleaved with a simultaneous increase in cytosolicIjB-alpha. In addition, P19 potently inhibited pro-survival proteins pSTAT3 and survivin. The multi-modal pro-apoptotic activity of P19 raises its potential usefulness as a promising anti-cancer therapeutic.
� 2011 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
Cancer cells accumulate several genetic and epigenetic muta-tions to cleverly evade self demise by deregulating apoptosis andsetting a stage for initiation, promotion and progression of themalignant growth [1]. Agents that can trigger the process of apop-tosis in cancer cells are therefore, considered potentially importantfor the development of anti-cancer chemotherapeutics [2]. In thisconcern, we focused our attention on parthenin, a major sesquiter-pene lactone from the obnoxious weed Parthenium hysterophorusLinn. (Asteraecae) that grows wild in different regions of India.The plant causes contact dermatitis and allergic rhinitis in animals[3]. Despite the fact that Parthenium is considered a toxic plant its
d Ltd. All rights reserved.
L, acute myeloid leukemia;, dynamin related protein-1;e synthase; NAC, N-acetyl-L-(ADP ribose) polymerase-1;
or of transcription protein-3;urea; TNFR1, tumor necrosis
1; fax: +91 191 2569333.
traditional uses are amply reported in literature [4]. One of itschemical constituent parthenin is reported to be responsible forcytotoxic effects of this plant [5,6]. Parthenin has also been re-ported to exhibit anti-tumor activity in mice [7]. In view of the re-ported toxicity of parthenin we attempted to prepare variousanalogs of parthenin modifying its functional groups to obtainsome potent anti-cancer analog of lesser toxicity. In this endeavorwe screened several semi-synthetic analogs of parthenin primarilyfor their apoptotic index and selected P19 among them as the pro-spective pro-apoptotic candidate for its detailed studies to developit into a promising anticancer therapeutic lead. About 50% of thecancers are known to have p53 mutated. Therefore, we employedp53 null, human acute myeloid leukemia HL-60 cells [8]. Thus,the agents that can induce apoptosis via a p53-independent path-way should have a broader therapeutic potential not only in AMLpatients but also in other cancer diseases.
Several earlier studies have demonstrated that nitric oxide (NO)is an important signaling messenger that plays an important role inmany physiological and pathological conditions. Endogenous NO isgenerated from L-arginine by three major types of NO synthases(NOS), i.e. endothelial NOS, neural NOS, and inducible NOS (iNOS)while another form of NO has recently been reported from

A. Kumar et al. / Chemico-Biological Interactions 193 (2011) 204–215 205
mitochondria [9,10]. Several studies have amply demonstratedthat NO can induce apoptosis in a variety of tumor cells [11,12],by mechanisms involving induction of stress proteins, mitochon-drial disruption, release of cytochrome c, and caspase activation[13]. In the recent past, we reported a lignan composition fromCedrus deodara that caused NO-mediated apoptotic cell death ofMolt-4 cells by activating both intrinsic and extrinsic pathways[14]. Nitric oxide in vivo may contribute to paracrine tumor-suppressive activities by triggering various signaling pathways. Aknown example is tamoxifen, which potently induced iNOS andNO formation in myoepithelial cells in culture in an estrogenreceptor-b-dependent manner [15]. This chemotherapeutic is usedfor high risk population of women with familial history of breastcancer. One potent transcription factor accounting for iNOS induc-tion is the nuclear factor-kappa-B (NF-jB), which is constitutivelyactivated in most of the cancer cell lines and in several cancertissues while its level is very low in resting cells [16]. Cells thatexpress constitutively activated NF-jB are resistant to various che-motherapeutic agents and radiation treatment [16]. Many previousstudies have implicated NF-jB in the development and progressionof cancer. NF-jB is also known to regulate the expression of severalanti-apoptotic genes such as IAPs, Bcl-2, Bcl-xL and survivin [17].In addition NF-jB regulates the expression of several genes in-volved in cancer cell proliferation, interestingly, some of thesegenes are also regulated by STAT3 (signal transducer and activatorof transcription protein-3). In fact STAT3 and NF-jB are closelylinked and both these transcription factors require each other fortheir persistent activation in the cancer cells [18]. STAT3 is foundto be crucial for growth of several cancer types but its expressionis dispensable for normal cells in postembryonic stage [19]. Thismakes NF-jB and STAT3 an ideal target for development of anti-cancer therapeutics. The present studies describe the multi-modalaction of P19 in killing cancer cells by targeting several such apop-totic signaling pathways where the selective generation of NO ap-pears indispensable initiating event in activating pro-apoptoticcascades while simultaneously inhibiting the activity of NF-jBand STAT3.
O
O
O
OO
Cl
Cl
Scheme 1. Structure of parthenin analog P19.
2. Materials and methods
2.1. Chemicals and antibodies
Dihydrorhoethidium (DHE), 2,7-dichlorodihydrofluoresceindiacetate (DCFH-DA), 4,5-diaminofluorescene-diacetate (DAF-2-DA), L-buthionine-S,R-sulfoximine (BSO), ethidium bromide,propidium iodide (PI), DNase-free RNase, proteinase K, 3-(4,5,-dimethylthiazole-2-yl)-2,5-diphenyltetrazolium bromide (MTT),N-acetyl-L-cysteine (NAC), s-methylisothiourea (sMIT), stauro-sporine, camptothecin, Fetal bovine serum were purchased fromM/s Sigma–Aldrich, USA; other reagents used were of analyticalgrade and available locally. Annexin V-FITC apoptosis detectionkit, Mitochondrial Membrane Sensor Kit and Apo Alert glutathionedetection kit were obtained from M/s BD Biosciences while Apoalertcaspases assay kits were from M/s B.D. Clontech; Anti-human anti-bodies to Bax (#SC-20067), Bid (#SC-6538), DRP-1 (#SC-21804),PARP-1 (#SC-8007), Bcl-2 (SC-7382), TNFR1 (#SC-1070), FAS (#SC-8009), DR4 (#SC-6824), DR5 (#SC-7192), NF-jB p65 (#SC-8008),NF-jB p50 (#SC-8414), pSTAT-3 (Ser727) (#SC-21876), CASPASE-3(#SC-7272), CASPASE-8 (#SC-56070), CASPASE-9 (#SC-56077), AIF(SC-5586), AIF siRNA (h) (SC-29193) survivin (#SC-8806), actin(#SC-8432), goat anti-rabbit IgG-HRP (#SC2030) and goat anti-mouse IgG-HRP (#SC2031), were from M/s Santa Cruz, USA; iNOS(#610432, clone 54) and cytochrome c (#556433, clone 7H8.2C12)were from M/s BD, Pharmingen, USA; actin (#CP01), iNOS(#482755) and rabbit anti-goat IgG-HRP (#401504) were from
Calbiochem, Germany; electrophoresis reagents and protein mark-ers were from M/s BIO-RAD, USA while Hyper film and ECL reagentsfrom M/s Amersham Biosciences, UK. The sources for other chemi-cals and reagents are same as used earlier [20,21].
2.2. Synthesis of parthenin analog P19
Parthenin, a sesquiterpene lactone was isolated from a plantweed P. hysterophorus. P19 [20-(4-chlorophenyl)-3-(4-chloropheny-lidene)-5,10-dimethyl-12-methylene-decahydro-1,2-(10,30-dioxo-cyclopenta[c])azuleno(4,5-b)furan-4,11-dione] (Scheme 1) wassynthesized from parthenin as reported earlier [22].
2.3. Cell culture, growth conditions and treatment
Human promyelocytic leukemia cell line HL-60 was obtainedfrom NCI, USA; MCF-7 and HeLa cells were obtained from NCCSPune, India; Human Gingival Fibroblast (hGF) cell line developedfrom healthy gingival [23] was provided kindly by Dr. Anil Bala-pure, Scientist, CDRI, Lucknow, India. The cells were grown inRPMI-1640 medium containing 10% FCS, 100 units pencillin/100 lg streptomycin per ml medium in CO2 incubator at 37 �Cwith 95% humidity and 5% CO2 gas environment. Cells were treatedwith P19 dissolved in DMSO while the untreated cultures receivedonly the vehicle (DMSO, <0.2%, v/v).
2.4. Housing and care of animals
Swiss albino mice (22–25 g), DBA/2 female mice and CDF1 malemice (18–23 g) were housed in standard size polycarbonate cagesfed with standard pellet diet (Gold Muhor, Lipton India Ltd.) andautoclaved water was given ad libitum. They were housed in con-trolled conditions of temperature (25 ± 2 �C), humidity (50–60%)and 12:12 h of light:dark cycle. The studies and number of animalsused were approved by the institutional ethics committee.
2.5. Cell proliferation assay
Cell proliferation was determined using 3-(4,5-dimethylthia-zole-2-yl)-2,5-diphenyltetrazolium bromide (MTT) as describedearlier [20]. HL-60 cells (2.0 � 104/200 ll) and adherent cultures(104/200 ll) of MCF-7, HeLa and hGF cells in 96 well culture plateswere treated with various concentrations of P19 for 48 h or varioustime periods. The MTT formazan crystals formed were dissolved in200 ll of DMSO; OD measured at 570 nm. The cytotoxicity of P19was expressed as the relative viability (% of untreated controlcells).

206 A. Kumar et al. / Chemico-Biological Interactions 193 (2011) 204–215
2.6. In vivo anti cancer studies
The studies for in vivo anti cancer activity in mouse modelswere conducted as per the protocols of National cancer Institute(NCI), USA [24].
2.7. Anti-cancer activity of P19 in L1210 lymphoid leukemia model inmice
L1210 lymphoid leukemia cells grown in the peritoneal cavityof DBA/2 female mice were collected from the animal harboring6–7 days old ascites. For testing, CDF1 males were used.5 � 105 cells were injected intraperitoneally in 18 CDF1 malesweighing 18–23 g on day 0. The next day, animals were random-ized and divided into three groups, containing seven animals each.Group I were treated with P19 at the dose of 10 mg/kg (i.p.) fornine consecutive days, Group II received 20 mg/kg (i.p.) of 5-fluoro-uracil as positive control. Group III served as control and it received0.2 ml normal saline (i.p.) for nine consecutive days. The mediansurvival time of animals in each group was calculated as describedearlier [24].
2.8. The effect of P19 on Ehrlich ascites tumor (solid) models in mice
Ehrlich Ascites Carcinoma (EAC) cells maintained in the perito-neal cavity of Swiss albino mice were collected from an animal hav-ing 8–10 days old ascitic tumor by aspirating ascetic fluid. Mice ofsingle sex weighing 18–23 g were selected for the experiment, andwere injected with 1 � 107 EAC cells intramuscularly in the rightthigh of each animal on day 0 and the treatment of P19 (15 and25 mg/kg, i.p.) started from day 1. The tumor weight was calculatedon day 13 in the animals treated for 9 days intraperitoneally (i.p.) ineach animal on day 0. On day 1, the animals were randomized anddivided into different groups. Treatment groups contained sevenanimals each and a control group contained 15 animals. P19 wasprepared in 1% Gum acacia in normal saline and was given intraper-itoneally from day 1 to 9. Similarly, Control group received 1% Gumacacia in normal saline, whilst 5-fluorouracil (5FU), 22 mg/kg i.p.,was used as a positive control. The average tumor weight for eachgroup was calculated, and the percent tumor growth inhibition intreated groups was calculated as explained earlier [24].
2.9. The effect of P19 on Sarcoma-180 (solid) model in mice
Sarcoma-180 cells maintained in the peritoneal cavity of balb/cmice were collected from an animal having 8–10 days old ascitictumor by aspirating ascetic fluid and 1 � 107 Sarcoma-180 cellswere injected intramuscularly into the right thigh of each animalon day 0 and the treatment of P19 (15 mg/kg, i.p.) started fromday 1. Treatment with 5-fluorouracil (5FU), 22 mg/kg i.p., wasgiven to a group of mice serving as positive control. Rest of theconditions used were same as that in EAT model.
2.10. Flow cytometric analysis of apoptosis and necrosis
HL-60 cells (1 � 106/ml) were treated with or without 10 lMP19 and 5 mM NAC for indicated time periods. Cells were collected,washed twice with PBS and suspended in 0.1 ml binding bufferprovided with apoptosis detection kit. Cells were stained with an-nexin V-FITC antibody and propidium iodide as per instructions ofthe manufacturer and scanned in FL-1 (FITC) vs. FL-2 (PI) channelson BD-LSR flow cytometer using quadrant analysis for evaluatingapoptotic and necrotic cell populations [14].
2.11. DNA cell cycle analysis
Cells were treated for various time periods with indicatedconcentrations of P19, processed, stained with PI and analyzedfor PI-DNA fluorescence by flow cytometry as described earlier[20]. The hypodiploid sub-G0/G1 DNA fraction predicts an apopto-tic cell fraction.
2.12. DNA agarose gel electrophoresis
DNA fragmentation was assessed by electrophoresis ofextracted genomic DNA and visualized in BIO-RAD Gel Documen-tation System as described earlier [20].
2.13. Measurement of intracellular peroxides/ROS, nitric oxide andmitochondrial membrane potential (Dwm) in HL 60 cells by flowcytometry
The levels of intracellular peroxides/ROS were analyzed byusing dihydroethidium (DHE) and 20,70-dichlorodihydrofluoresceindiacetate (DCFH-DA). Intracellular generation of nitric oxide wasmeasured by using cell permeable dye 4,5-diaminofluorescene-diacetate (DAF-2-DA). Mitochondrial membrane potential wasmeasured by using a Mitochondrial Membrane Sensor Kit contain-ing JC-1 dye as described earlier [20,21].
2.14. Measurement of GSH contents in cells
Intracellular levels of GSH were estimated using the BD Apo-Alert™ glutathione detection kit employing monochlorobimane(MCB) reagent. Briefly, cells after various treatments were lysedand incubated with 2 mM MCB for 3 h at 37 �C. Reduced glutathi-one levels were assayed fluorometrically at 395/480 nm accordingto manufacturer’s protocol.
2.15. Caspase assays
For fluoremetric enzyme assay cells (2 � 106) were incubatedwith 10 lM P19 for indicated time periods. At the end of treatmentcells were washed in PBS and pellets lysed in the cell lysis buffer.Activities of caspase-3, -9/6 and -8 in the cell lysates were deter-mined fluorometrically, using BD ApoAlert caspase fluorescentassay kits. Caspase-3 and -8 employed fluorochrome conjugatedpeptides DEVD-AFC and IETD-AFC as substrates, respectively whilecaspase-9 employed LEHD-AMC. Release of AFC (7-amino-4-trifluoromethyl coumarin) and AMC (7-aminomethylcoumarin)was assayed according to the instructions provided in the manualby the supplier. Specific inhibitors were used as negative control todetermine whether fluorescence intensity changes were specificfor the activity of caspases. The peptide based inhibitors used wereDEVD-CHO for caspase-3, IETD-fmk for caspase-8 and LEHD-CHOfor caspase-9/6. Activation of caspases-3, -9 and -8 was also con-firmed by western blotting from the whole cell lysates.
2.16. Use of AIF-siRNA and caspase inhibitors to validate thecontribution of AIF and caspases to apoptosis
Expression of AIF was knocked-down in HL-60 cells by usingsiRNA according to manufacturer’s protocol (Santacruz Biotechnol-ogy Inc., USA). For inhibition of caspases cell permeable pan cas-pase inhibitor Z-VAD-fmk at 50 lV was added to cell cultures 1 hbefore 6 h treatment with 10 lV of P19 or 50 lV of b-boswellicacid butyrate (BA-145), used as positive control.
A. Kumar et al. / Chemico-Biological Interactions 193 (2011) 204–215 207
2.17. Preparation of cell lysates for western blots analysis
Treated and untreated HL-60 cells were centrifuged at 400g at4 �C, washed in PBS and cell pellets processed for preparation ofcytosolic, mitochondrial, nuclear and whole cell lysate fractionsas described earlier [21]. The conditions for the analysis of theexpression of various proteins by western blot analysis using spe-cific antibodies were same as reported earlier [21].
2.18. Statistical analysis
Data are presented as mean ± S.D. of the number of experimentsindicated. The comparisons were made between control and trea-ted groups and statistical significance was calculated by using Bon-ferroni test. The difference was considered to be statisticallysignificant if the p value was ⁄p < 0.05, ⁄⁄p < 0.01, ⁄⁄⁄p < 0.001.
3. Results
3.1. P19 inhibits cell proliferation
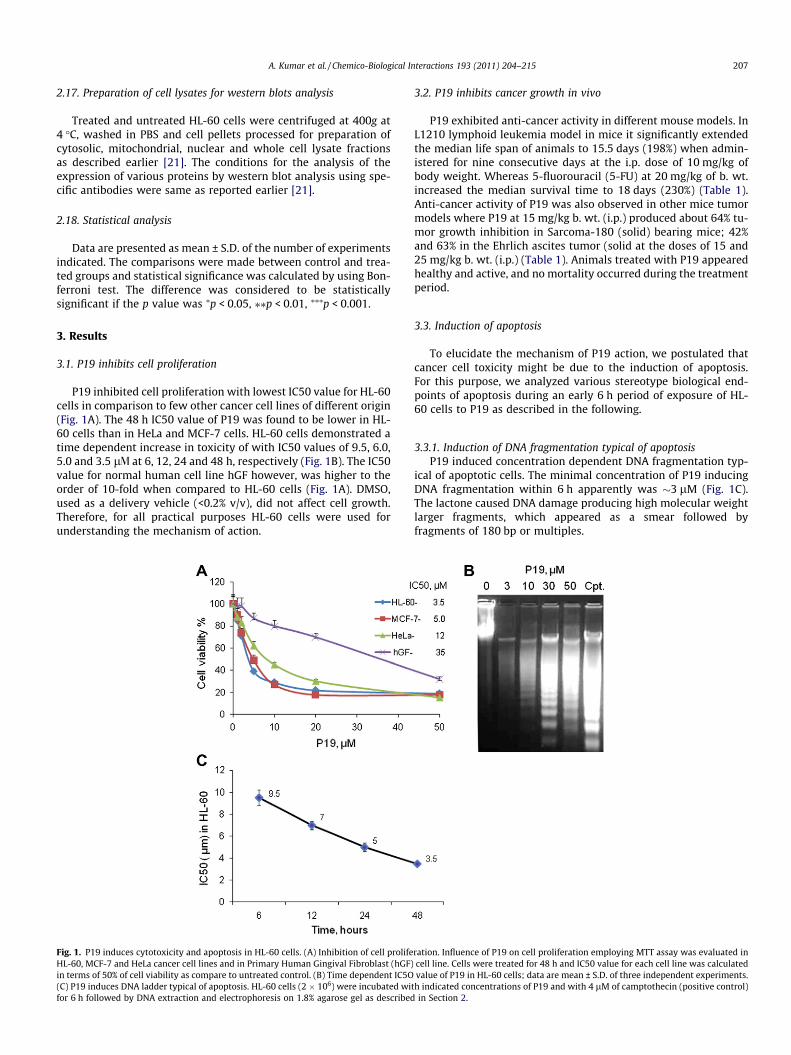
P19 inhibited cell proliferation with lowest IC50 value for HL-60cells in comparison to few other cancer cell lines of different origin(Fig. 1A). The 48 h IC50 value of P19 was found to be lower in HL-60 cells than in HeLa and MCF-7 cells. HL-60 cells demonstrated atime dependent increase in toxicity of with IC50 values of 9.5, 6.0,5.0 and 3.5 lV at 6, 12, 24 and 48 h, respectively (Fig. 1B). The IC50value for normal human cell line hGF however, was higher to theorder of 10-fold when compared to HL-60 cells (Fig. 1A). DMSO,used as a delivery vehicle (<0.2% v/v), did not affect cell growth.Therefore, for all practical purposes HL-60 cells were used forunderstanding the mechanism of action.
Fig. 1. P19 induces cytotoxicity and apoptosis in HL-60 cells. (A) Inhibition of cell prolifeHL-60, MCF-7 and HeLa cancer cell lines and in Primary Human Gingival Fibroblast (hGF)in terms of 50% of cell viability as compare to untreated control. (B) Time dependent IC5O(C) P19 induces DNA ladder typical of apoptosis. HL-60 cells (2 � 106) were incubated wifor 6 h followed by DNA extraction and electrophoresis on 1.8% agarose gel as described
3.2. P19 inhibits cancer growth in vivo
P19 exhibited anti-cancer activity in different mouse models. InL1210 lymphoid leukemia model in mice it significantly extendedthe median life span of animals to 15.5 days (198%) when admin-istered for nine consecutive days at the i.p. dose of 10 mg/kg ofbody weight. Whereas 5-fluorouracil (5-FU) at 20 mg/kg of b. wt.increased the median survival time to 18 days (230%) (Table 1).Anti-cancer activity of P19 was also observed in other mice tumormodels where P19 at 15 mg/kg b. wt. (i.p.) produced about 64% tu-mor growth inhibition in Sarcoma-180 (solid) bearing mice; 42%and 63% in the Ehrlich ascites tumor (solid at the doses of 15 and25 mg/kg b. wt. (i.p.) (Table 1). Animals treated with P19 appearedhealthy and active, and no mortality occurred during the treatmentperiod.
3.3. Induction of apoptosis
To elucidate the mechanism of P19 action, we postulated thatcancer cell toxicity might be due to the induction of apoptosis.For this purpose, we analyzed various stereotype biological end-points of apoptosis during an early 6 h period of exposure of HL-60 cells to P19 as described in the following.
3.3.1. Induction of DNA fragmentation typical of apoptosisP19 induced concentration dependent DNA fragmentation typ-
ical of apoptotic cells. The minimal concentration of P19 inducingDNA fragmentation within 6 h apparently was �3 lM (Fig. 1C).The lactone caused DNA damage producing high molecular weightlarger fragments, which appeared as a smear followed byfragments of 180 bp or multiples.
ration. Influence of P19 on cell proliferation employing MTT assay was evaluated incell line. Cells were treated for 48 h and IC50 value for each cell line was calculatedvalue of P19 in HL-60 cells; data are mean ± S.D. of three independent experiments.
th indicated concentrations of P19 and with 4 lM of camptothecin (positive control)in Section 2.
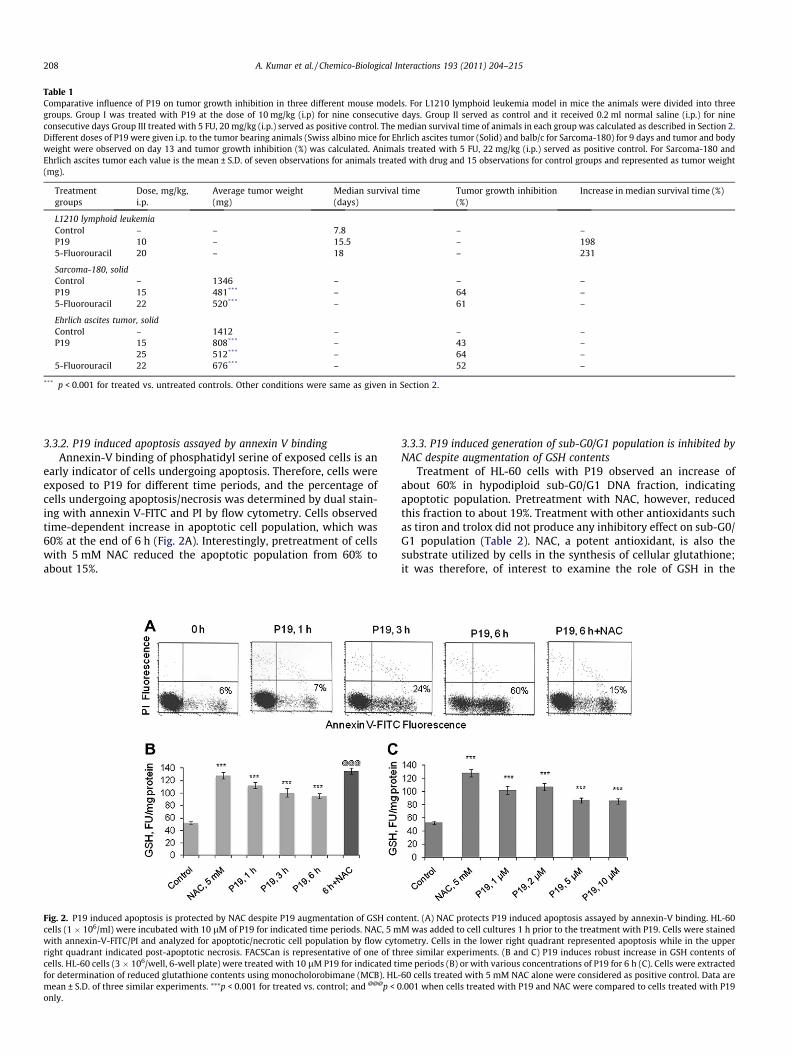
Table 1Comparative influence of P19 on tumor growth inhibition in three different mouse models. For L1210 lymphoid leukemia model in mice the animals were divided into threegroups. Group I was treated with P19 at the dose of 10 mg/kg (i.p) for nine consecutive days. Group II served as control and it received 0.2 ml normal saline (i.p.) for nineconsecutive days Group III treated with 5 FU, 20 mg/kg (i.p.) served as positive control. The median survival time of animals in each group was calculated as described in Section 2.Different doses of P19 were given i.p. to the tumor bearing animals (Swiss albino mice for Ehrlich ascites tumor (Solid) and balb/c for Sarcoma-180) for 9 days and tumor and bodyweight were observed on day 13 and tumor growth inhibition (%) was calculated. Animals treated with 5 FU, 22 mg/kg (i.p.) served as positive control. For Sarcoma-180 andEhrlich ascites tumor each value is the mean ± S.D. of seven observations for animals treated with drug and 15 observations for control groups and represented as tumor weight(mg).
Treatmentgroups
Dose, mg/kg,i.p.
Average tumor weight(mg)
Median survival time(days)
Tumor growth inhibition(%)
Increase in median survival time (%)
L1210 lymphoid leukemiaControl – – 7.8 – –P19 10 – 15.5 – 1985-Fluorouracil 20 – 18 – 231
Sarcoma-180, solidControl – 1346 – – –P19 15 481*** – 64 –5-Fluorouracil 22 520*** – 61 –
Ehrlich ascites tumor, solidControl – 1412 – – –P19 15 808*** – 43 –
25 512*** – 64 –5-Fluorouracil 22 676*** – 52 –
*** p < 0.001 for treated vs. untreated controls. Other conditions were same as given in Section 2.
208 A. Kumar et al. / Chemico-Biological Interactions 193 (2011) 204–215
3.3.2. P19 induced apoptosis assayed by annexin V bindingAnnexin-V binding of phosphatidyl serine of exposed cells is an
early indicator of cells undergoing apoptosis. Therefore, cells wereexposed to P19 for different time periods, and the percentage ofcells undergoing apoptosis/necrosis was determined by dual stain-ing with annexin V-FITC and PI by flow cytometry. Cells observedtime-dependent increase in apoptotic cell population, which was60% at the end of 6 h (Fig. 2A). Interestingly, pretreatment of cellswith 5 mM NAC reduced the apoptotic population from 60% toabout 15%.
Fig. 2. P19 induced apoptosis is protected by NAC despite P19 augmentation of GSH concells (1 � 106/ml) were incubated with 10 lM of P19 for indicated time periods. NAC, 5 mwith annexin-V-FITC/PI and analyzed for apoptotic/necrotic cell population by flow cytoright quadrant indicated post-apoptotic necrosis. FACSCan is representative of one of thcells. HL-60 cells (3 � 106/well, 6-well plate) were treated with 10 lM P19 for indicated tifor determination of reduced glutathione contents using monocholorobimane (MCB). HL-mean ± S.D. of three similar experiments. ⁄⁄⁄p < 0.001 for treated vs. control; and @@@p < 0only.
3.3.3. P19 induced generation of sub-G0/G1 population is inhibited byNAC despite augmentation of GSH contents
Treatment of HL-60 cells with P19 observed an increase ofabout 60% in hypodiploid sub-G0/G1 DNA fraction, indicatingapoptotic population. Pretreatment with NAC, however, reducedthis fraction to about 19%. Treatment with other antioxidants suchas tiron and trolox did not produce any inhibitory effect on sub-G0/G1 population (Table 2). NAC, a potent antioxidant, is also thesubstrate utilized by cells in the synthesis of cellular glutathione;it was therefore, of interest to examine the role of GSH in the
tent. (A) NAC protects P19 induced apoptosis assayed by annexin-V binding. HL-60M was added to cell cultures 1 h prior to the treatment with P19. Cells were stainedmetry. Cells in the lower right quadrant represented apoptosis while in the upperree similar experiments. (B and C) P19 induces robust increase in GSH contents ofme periods (B) or with various concentrations of P19 for 6 h (C). Cells were extracted60 cells treated with 5 mM NAC alone were considered as positive control. Data are.001 when cells treated with P19 and NAC were compared to cells treated with P19
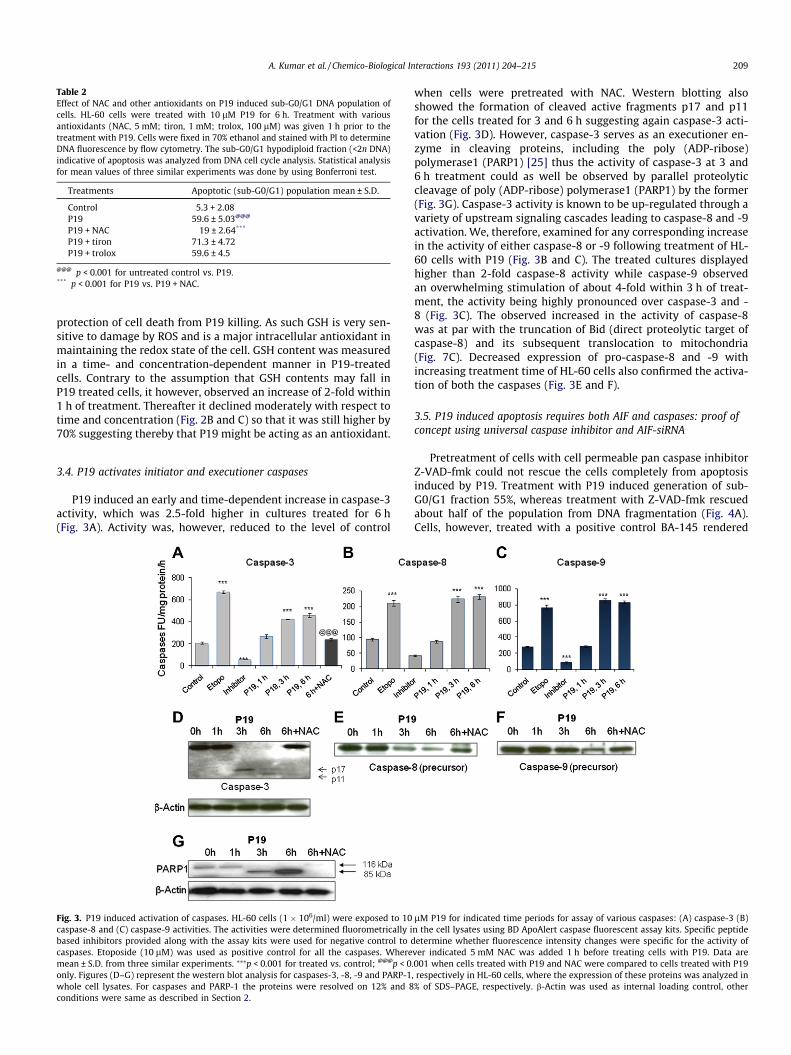
Table 2Effect of NAC and other antioxidants on P19 induced sub-G0/G1 DNA population ofcells. HL-60 cells were treated with 10 lM P19 for 6 h. Treatment with variousantioxidants (NAC, 5 mM; tiron, 1 mM; trolox, 100 lM) was given 1 h prior to thetreatment with P19. Cells were fixed in 70% ethanol and stained with PI to determineDNA fluorescence by flow cytometry. The sub-G0/G1 hypodiploid fraction (<2n DNA)indicative of apoptosis was analyzed from DNA cell cycle analysis. Statistical analysisfor mean values of three similar experiments was done by using Bonferroni test.
Treatments Apoptotic (sub-G0/G1) population mean ± S.D.
Control 5.3 + 2.08P19 59.6 ± 5.03@@@
P19 + NAC 19 ± 2.64***
P19 + tiron 71.3 ± 4.72P19 + trolox 59.6 ± 4.5
@@@ p < 0.001 for untreated control vs. P19.*** p < 0.001 for P19 vs. P19 + NAC.
A. Kumar et al. / Chemico-Biological Interactions 193 (2011) 204–215 209
protection of cell death from P19 killing. As such GSH is very sen-sitive to damage by ROS and is a major intracellular antioxidant inmaintaining the redox state of the cell. GSH content was measuredin a time- and concentration-dependent manner in P19-treatedcells. Contrary to the assumption that GSH contents may fall inP19 treated cells, it however, observed an increase of 2-fold within1 h of treatment. Thereafter it declined moderately with respect totime and concentration (Fig. 2B and C) so that it was still higher by70% suggesting thereby that P19 might be acting as an antioxidant.
3.4. P19 activates initiator and executioner caspases
P19 induced an early and time-dependent increase in caspase-3activity, which was 2.5-fold higher in cultures treated for 6 h(Fig. 3A). Activity was, however, reduced to the level of control
Fig. 3. P19 induced activation of caspases. HL-60 cells (1 � 106/ml) were exposed to 10caspase-8 and (C) caspase-9 activities. The activities were determined fluorometrically ibased inhibitors provided along with the assay kits were used for negative control to dcaspases. Etoposide (10 lM) was used as positive control for all the caspases. Wheremean ± S.D. from three similar experiments. ⁄⁄⁄p < 0.001 for treated vs. control; @@@p < 0only. Figures (D–G) represent the western blot analysis for caspases-3, -8, -9 and PARP-1whole cell lysates. For caspases and PARP-1 the proteins were resolved on 12% and 8conditions were same as described in Section 2.
when cells were pretreated with NAC. Western blotting alsoshowed the formation of cleaved active fragments p17 and p11for the cells treated for 3 and 6 h suggesting again caspase-3 acti-vation (Fig. 3D). However, caspase-3 serves as an executioner en-zyme in cleaving proteins, including the poly (ADP-ribose)polymerase1 (PARP1) [25] thus the activity of caspase-3 at 3 and6 h treatment could as well be observed by parallel proteolyticcleavage of poly (ADP-ribose) polymerase1 (PARP1) by the former(Fig. 3G). Caspase-3 activity is known to be up-regulated through avariety of upstream signaling cascades leading to caspase-8 and -9activation. We, therefore, examined for any corresponding increasein the activity of either caspase-8 or -9 following treatment of HL-60 cells with P19 (Fig. 3B and C). The treated cultures displayedhigher than 2-fold caspase-8 activity while caspase-9 observedan overwhelming stimulation of about 4-fold within 3 h of treat-ment, the activity being highly pronounced over caspase-3 and -8 (Fig. 3C). The observed increased in the activity of caspase-8was at par with the truncation of Bid (direct proteolytic target ofcaspase-8) and its subsequent translocation to mitochondria(Fig. 7C). Decreased expression of pro-caspase-8 and -9 withincreasing treatment time of HL-60 cells also confirmed the activa-tion of both the caspases (Fig. 3E and F).
3.5. P19 induced apoptosis requires both AIF and caspases: proof ofconcept using universal caspase inhibitor and AIF-siRNA
Pretreatment of cells with cell permeable pan caspase inhibitorZ-VAD-fmk could not rescue the cells completely from apoptosisinduced by P19. Treatment with P19 induced generation of sub-G0/G1 fraction 55%, whereas treatment with Z-VAD-fmk rescuedabout half of the population from DNA fragmentation (Fig. 4A).Cells, however, treated with a positive control BA-145 rendered
lM P19 for indicated time periods for assay of various caspases: (A) caspase-3 (B)n the cell lysates using BD ApoAlert caspase fluorescent assay kits. Specific peptideetermine whether fluorescence intensity changes were specific for the activity of
ver indicated 5 mM NAC was added 1 h before treating cells with P19. Data are.001 when cells treated with P19 and NAC were compared to cells treated with P19, respectively in HL-60 cells, where the expression of these proteins was analyzed in% of SDS–PAGE, respectively. b-Actin was used as internal loading control, other
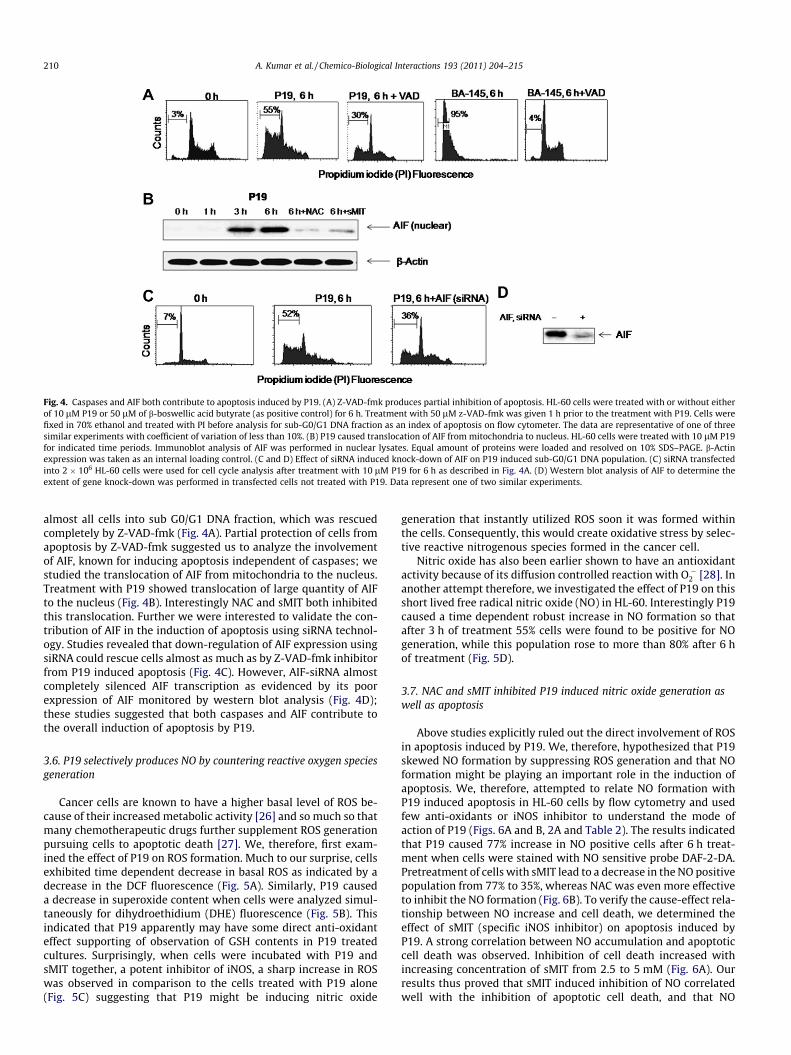
Fig. 4. Caspases and AIF both contribute to apoptosis induced by P19. (A) Z-VAD-fmk produces partial inhibition of apoptosis. HL-60 cells were treated with or without eitherof 10 lM P19 or 50 lM of b-boswellic acid butyrate (as positive control) for 6 h. Treatment with 50 lM z-VAD-fmk was given 1 h prior to the treatment with P19. Cells werefixed in 70% ethanol and treated with PI before analysis for sub-G0/G1 DNA fraction as an index of apoptosis on flow cytometer. The data are representative of one of threesimilar experiments with coefficient of variation of less than 10%. (B) P19 caused translocation of AIF from mitochondria to nucleus. HL-60 cells were treated with 10 lM P19for indicated time periods. Immunoblot analysis of AIF was performed in nuclear lysates. Equal amount of proteins were loaded and resolved on 10% SDS–PAGE. b-Actinexpression was taken as an internal loading control. (C and D) Effect of siRNA induced knock-down of AIF on P19 induced sub-G0/G1 DNA population. (C) siRNA transfectedinto 2 � 106 HL-60 cells were used for cell cycle analysis after treatment with 10 lM P19 for 6 h as described in Fig. 4A. (D) Western blot analysis of AIF to determine theextent of gene knock-down was performed in transfected cells not treated with P19. Data represent one of two similar experiments.
210 A. Kumar et al. / Chemico-Biological Interactions 193 (2011) 204–215
almost all cells into sub G0/G1 DNA fraction, which was rescuedcompletely by Z-VAD-fmk (Fig. 4A). Partial protection of cells fromapoptosis by Z-VAD-fmk suggested us to analyze the involvementof AIF, known for inducing apoptosis independent of caspases; westudied the translocation of AIF from mitochondria to the nucleus.Treatment with P19 showed translocation of large quantity of AIFto the nucleus (Fig. 4B). Interestingly NAC and sMIT both inhibitedthis translocation. Further we were interested to validate the con-tribution of AIF in the induction of apoptosis using siRNA technol-ogy. Studies revealed that down-regulation of AIF expression usingsiRNA could rescue cells almost as much as by Z-VAD-fmk inhibitorfrom P19 induced apoptosis (Fig. 4C). However, AIF-siRNA almostcompletely silenced AIF transcription as evidenced by its poorexpression of AIF monitored by western blot analysis (Fig. 4D);these studies suggested that both caspases and AIF contribute tothe overall induction of apoptosis by P19.
3.6. P19 selectively produces NO by countering reactive oxygen speciesgeneration
Cancer cells are known to have a higher basal level of ROS be-cause of their increased metabolic activity [26] and so much so thatmany chemotherapeutic drugs further supplement ROS generationpursuing cells to apoptotic death [27]. We, therefore, first exam-ined the effect of P19 on ROS formation. Much to our surprise, cellsexhibited time dependent decrease in basal ROS as indicated by adecrease in the DCF fluorescence (Fig. 5A). Similarly, P19 causeda decrease in superoxide content when cells were analyzed simul-taneously for dihydroethidium (DHE) fluorescence (Fig. 5B). Thisindicated that P19 apparently may have some direct anti-oxidanteffect supporting of observation of GSH contents in P19 treatedcultures. Surprisingly, when cells were incubated with P19 andsMIT together, a potent inhibitor of iNOS, a sharp increase in ROSwas observed in comparison to the cells treated with P19 alone(Fig. 5C) suggesting that P19 might be inducing nitric oxide
generation that instantly utilized ROS soon it was formed withinthe cells. Consequently, this would create oxidative stress by selec-tive reactive nitrogenous species formed in the cancer cell.
Nitric oxide has also been earlier shown to have an antioxidantactivity because of its diffusion controlled reaction with O��2 [28]. Inanother attempt therefore, we investigated the effect of P19 on thisshort lived free radical nitric oxide (NO) in HL-60. Interestingly P19caused a time dependent robust increase in NO formation so thatafter 3 h of treatment 55% cells were found to be positive for NOgeneration, while this population rose to more than 80% after 6 hof treatment (Fig. 5D).
3.7. NAC and sMIT inhibited P19 induced nitric oxide generation aswell as apoptosis
Above studies explicitly ruled out the direct involvement of ROSin apoptosis induced by P19. We, therefore, hypothesized that P19skewed NO formation by suppressing ROS generation and that NOformation might be playing an important role in the induction ofapoptosis. We, therefore, attempted to relate NO formation withP19 induced apoptosis in HL-60 cells by flow cytometry and usedfew anti-oxidants or iNOS inhibitor to understand the mode ofaction of P19 (Figs. 6A and B, 2A and Table 2). The results indicatedthat P19 caused 77% increase in NO positive cells after 6 h treat-ment when cells were stained with NO sensitive probe DAF-2-DA.Pretreatment of cells with sMIT lead to a decrease in the NO positivepopulation from 77% to 35%, whereas NAC was even more effectiveto inhibit the NO formation (Fig. 6B). To verify the cause-effect rela-tionship between NO increase and cell death, we determined theeffect of sMIT (specific iNOS inhibitor) on apoptosis induced byP19. A strong correlation between NO accumulation and apoptoticcell death was observed. Inhibition of cell death increased withincreasing concentration of sMIT from 2.5 to 5 mM (Fig. 6A). Ourresults thus proved that sMIT induced inhibition of NO correlatedwell with the inhibition of apoptotic cell death, and that NO
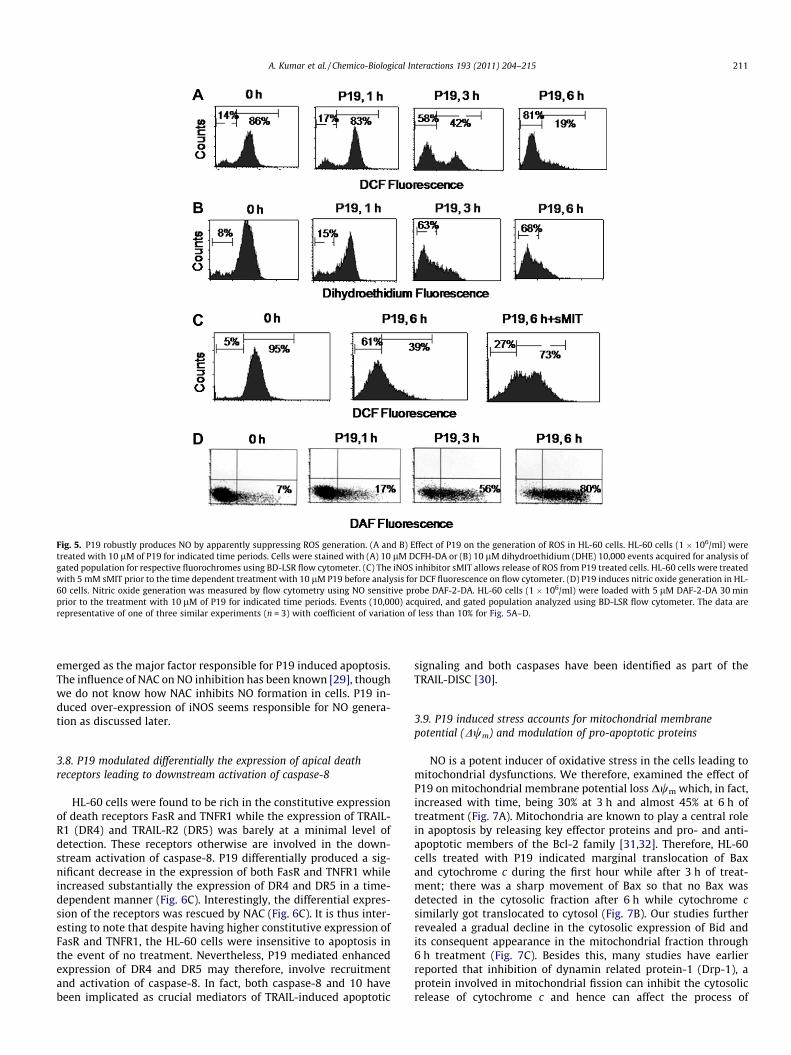
Fig. 5. P19 robustly produces NO by apparently suppressing ROS generation. (A and B) Effect of P19 on the generation of ROS in HL-60 cells. HL-60 cells (1 � 106/ml) weretreated with 10 lM of P19 for indicated time periods. Cells were stained with (A) 10 lM DCFH-DA or (B) 10 lM dihydroethidium (DHE) 10,000 events acquired for analysis ofgated population for respective fluorochromes using BD-LSR flow cytometer. (C) The iNOS inhibitor sMIT allows release of ROS from P19 treated cells. HL-60 cells were treatedwith 5 mM sMIT prior to the time dependent treatment with 10 lM P19 before analysis for DCF fluorescence on flow cytometer. (D) P19 induces nitric oxide generation in HL-60 cells. Nitric oxide generation was measured by flow cytometry using NO sensitive probe DAF-2-DA. HL-60 cells (1 � 106/ml) were loaded with 5 lM DAF-2-DA 30 minprior to the treatment with 10 lM of P19 for indicated time periods. Events (10,000) acquired, and gated population analyzed using BD-LSR flow cytometer. The data arerepresentative of one of three similar experiments (n = 3) with coefficient of variation of less than 10% for Fig. 5A–D.
A. Kumar et al. / Chemico-Biological Interactions 193 (2011) 204–215 211
emerged as the major factor responsible for P19 induced apoptosis.The influence of NAC on NO inhibition has been known [29], thoughwe do not know how NAC inhibits NO formation in cells. P19 in-duced over-expression of iNOS seems responsible for NO genera-tion as discussed later.
3.8. P19 modulated differentially the expression of apical deathreceptors leading to downstream activation of caspase-8
HL-60 cells were found to be rich in the constitutive expressionof death receptors FasR and TNFR1 while the expression of TRAIL-R1 (DR4) and TRAIL-R2 (DR5) was barely at a minimal level ofdetection. These receptors otherwise are involved in the down-stream activation of caspase-8. P19 differentially produced a sig-nificant decrease in the expression of both FasR and TNFR1 whileincreased substantially the expression of DR4 and DR5 in a time-dependent manner (Fig. 6C). Interestingly, the differential expres-sion of the receptors was rescued by NAC (Fig. 6C). It is thus inter-esting to note that despite having higher constitutive expression ofFasR and TNFR1, the HL-60 cells were insensitive to apoptosis inthe event of no treatment. Nevertheless, P19 mediated enhancedexpression of DR4 and DR5 may therefore, involve recruitmentand activation of caspase-8. In fact, both caspase-8 and 10 havebeen implicated as crucial mediators of TRAIL-induced apoptotic
signaling and both caspases have been identified as part of theTRAIL-DISC [30].
3.9. P19 induced stress accounts for mitochondrial membranepotential (Dwm) and modulation of pro-apoptotic proteins
NO is a potent inducer of oxidative stress in the cells leading tomitochondrial dysfunctions. We therefore, examined the effect ofP19 on mitochondrial membrane potential loss Dwm which, in fact,increased with time, being 30% at 3 h and almost 45% at 6 h oftreatment (Fig. 7A). Mitochondria are known to play a central rolein apoptosis by releasing key effector proteins and pro- and anti-apoptotic members of the Bcl-2 family [31,32]. Therefore, HL-60cells treated with P19 indicated marginal translocation of Baxand cytochrome c during the first hour while after 3 h of treat-ment; there was a sharp movement of Bax so that no Bax wasdetected in the cytosolic fraction after 6 h while cytochrome csimilarly got translocated to cytosol (Fig. 7B). Our studies furtherrevealed a gradual decline in the cytosolic expression of Bid andits consequent appearance in the mitochondrial fraction through6 h treatment (Fig. 7C). Besides this, many studies have earlierreported that inhibition of dynamin related protein-1 (Drp-1), aprotein involved in mitochondrial fission can inhibit the cytosolicrelease of cytochrome c and hence can affect the process of
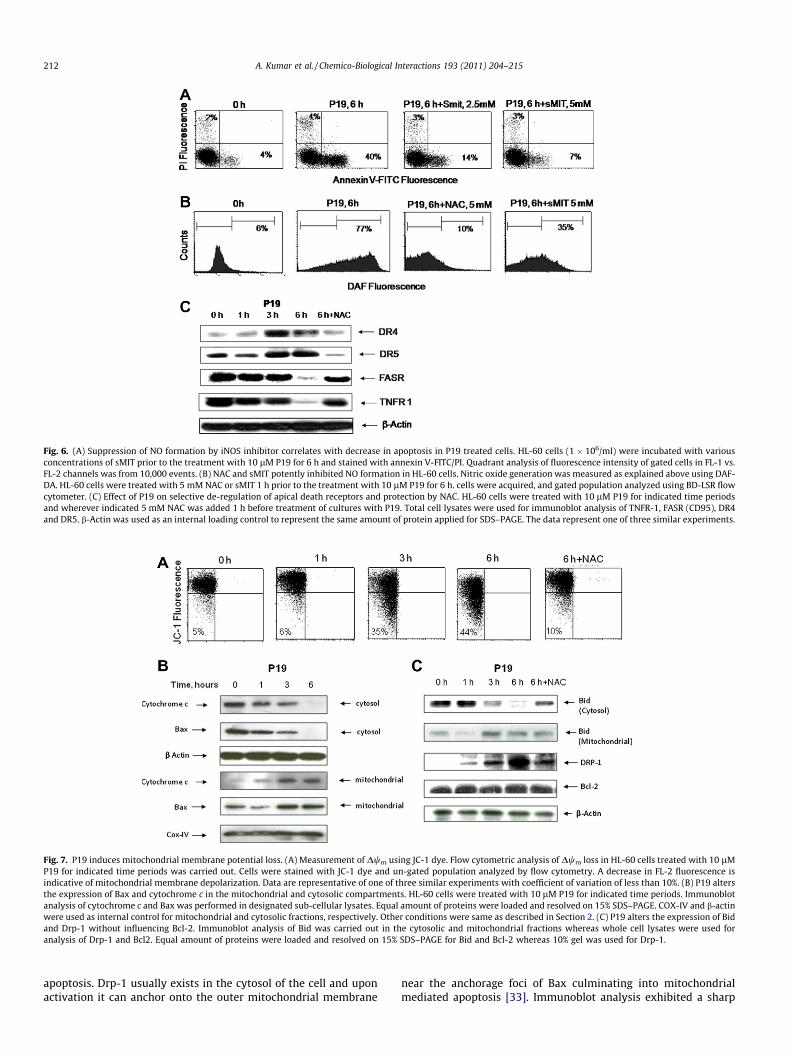
Fig. 6. (A) Suppression of NO formation by iNOS inhibitor correlates with decrease in apoptosis in P19 treated cells. HL-60 cells (1 � 106/ml) were incubated with variousconcentrations of sMIT prior to the treatment with 10 lM P19 for 6 h and stained with annexin V-FITC/PI. Quadrant analysis of fluorescence intensity of gated cells in FL-1 vs.FL-2 channels was from 10,000 events. (B) NAC and sMIT potently inhibited NO formation in HL-60 cells. Nitric oxide generation was measured as explained above using DAF-DA. HL-60 cells were treated with 5 mM NAC or sMIT 1 h prior to the treatment with 10 lM P19 for 6 h. cells were acquired, and gated population analyzed using BD-LSR flowcytometer. (C) Effect of P19 on selective de-regulation of apical death receptors and protection by NAC. HL-60 cells were treated with 10 lM P19 for indicated time periodsand wherever indicated 5 mM NAC was added 1 h before treatment of cultures with P19. Total cell lysates were used for immunoblot analysis of TNFR-1, FASR (CD95), DR4and DR5. b-Actin was used as an internal loading control to represent the same amount of protein applied for SDS–PAGE. The data represent one of three similar experiments.
Fig. 7. P19 induces mitochondrial membrane potential loss. (A) Measurement of Dwm using JC-1 dye. Flow cytometric analysis of Dwm loss in HL-60 cells treated with 10 lMP19 for indicated time periods was carried out. Cells were stained with JC-1 dye and un-gated population analyzed by flow cytometry. A decrease in FL-2 fluorescence isindicative of mitochondrial membrane depolarization. Data are representative of one of three similar experiments with coefficient of variation of less than 10%. (B) P19 altersthe expression of Bax and cytochrome c in the mitochondrial and cytosolic compartments. HL-60 cells were treated with 10 lM P19 for indicated time periods. Immunoblotanalysis of cytochrome c and Bax was performed in designated sub-cellular lysates. Equal amount of proteins were loaded and resolved on 15% SDS–PAGE. COX-IV and b-actinwere used as internal control for mitochondrial and cytosolic fractions, respectively. Other conditions were same as described in Section 2. (C) P19 alters the expression of Bidand Drp-1 without influencing Bcl-2. Immunoblot analysis of Bid was carried out in the cytosolic and mitochondrial fractions whereas whole cell lysates were used foranalysis of Drp-1 and Bcl2. Equal amount of proteins were loaded and resolved on 15% SDS–PAGE for Bid and Bcl-2 whereas 10% gel was used for Drp-1.
212 A. Kumar et al. / Chemico-Biological Interactions 193 (2011) 204–215
apoptosis. Drp-1 usually exists in the cytosol of the cell and uponactivation it can anchor onto the outer mitochondrial membrane
near the anchorage foci of Bax culminating into mitochondrialmediated apoptosis [33]. Immunoblot analysis exhibited a sharp

A. Kumar et al. / Chemico-Biological Interactions 193 (2011) 204–215 213
rise in the expression of Drp-1 in P19-treated cells, whereas itsexpression was below the level of detection in control (Fig. 7C).Further over expression of Drp-1 occurred concurrent to Bax trans-location to mitochondria reinforcing cells to tread the path ofapoptosis. It was interesting to note that P19 did not cause any sig-nificant change in the expression of anti-apoptotic protein Bcl-2(Fig. 7C), which otherwise is a predominant regulator of cyto-chrome c from the mitochondria.
3.10. P19 induced activation of iNOS is unlikely influenced by itstranscription factor NF-jB and transactivator PARP1
NF-jB is a transcription factor for vast array of genes, includingiNOS. It up-regulates the expression of iNOS by binding to its pro-moter whereas PARP-1 helps NF-jB in its task by acting as transac-tivator [34] leading ultimately to enhanced production of nitricoxide (NO). Interestingly, P19 caused a sharp increase in theexpression of iNOS with significantly high level being attained at6 h (Fig. 8A). The increase in NO formation (Fig. 4D) appeared tobe correlated with iNOS activation. The expression of NF-jB p65(Rel-A) on the contrary, was suppressed due to cleavage in the nu-clear and cytosolic fractions at 3 and 6 h treatments (Fig. 8A).Strong inhibitory effect of P19 on the expression of NF-jB p50(the most common functional partner of Rel-A) further suggestssuppressive action of P19 on NF-jB signaling. Coinciding withthe inhibitory action of P19 on NF-jB expression, the activity ofits regulatory protein Ijb also observed a moderate increase inthe cytosolic fraction (Fig. 8A). Ijb generally exists in a complexform with NF-jB and thereby regulates its translocation to the nu-cleus [16]. P19 increasingly caused localization of NF-jB p65 intothe cytosol in a time dependent manner whereas its expressionin the nucleus decreased correspondingly. P19 thus seemed to ex-ert a dual impact on the inhibition of NF-jB activity by reducingthe overall expression through cleavage and by stabilizing theNF-jB–Ijb complex in the cytosol and preventing its translocationto the nucleus (Fig. 8A). Analysis for the expression of PARP-1 alsorevealed that it becomes totally cleaved after 3 h treatment. Thecleaved 85kD product, however, exhibited a highly enhancedexpression indicating almost complete cleavage of PARP-1. All
Fig. 8. (A) P19 induced expression of iNOS is not dependent on the repression of NF-jB a10 lM P19. Samples were separated on 8% gel for iNOS and PARP-1 analysis whereas 1loading control. Other conditions were same as described in Section 2. (B) Effect of P19 oon 8% gel whereas for survivin 15% gel was used. Density of each band for STAT3 and s
these alterations in the expression of these genes were rescuedby NAC, a potent inhibitor of NO formation in this study (Figs. 8Aand 3G). This unusual observation made us to believe about theinvolvement of NO in the inhibition of NF-jB and cleavage ofPARP-1. Our results thus suggest that expression of NF-jB (p65/p50) is certainly governed by NO level and that P19 induced iNOSactivation is regulated by some transcription factor/s other thanNF-jB, because the activity of the later is inhibited by P19. Thissuggests that P19 might be activating other factors involved inthe enhanced expression of iNOS other than NF-jB and PARP-1in HL-60 cells.
3.11. P19 inhibited the expression of cancer cell survival proteinspSTAT3 and survivin
The family of signal transducer and activator of transcription(STAT) proteins, particularly STAT3 play a crucial role in selectivelyinducing and maintaining pro-carcinogenic inflammatory micro-environment both at the initiation and during cancer progression[18]. The expression of pSTAT3 (ser727) in P19 treated HL-60 cellsexhibited a predominant decline of about 90% through 6 h (Fig. 8B).Another important anti-apoptotic protein that plays a pivotal rolein cancer is survivin, a member of IAP family. The protein is ex-pressed in embryonic stage and in most tumor cells, but is notdetectable in normal adult cells [35]. HL-60 cells under standardgrowth conditions showed high expression of survivin which de-clined substantially in the P19 treated cells. NAC again suppressedall these changes (Fig. 8B).
4. Discussion
The results of the present study describe that a novel semi-syn-thetic analog of parthenin (P19), a sesquiterpene lactone, activatedapoptosis in acute myelogenous leukemia HL-60 cells and inhib-ited tumor growth in mice. Intervention of apoptosis is the hall-mark in the development of anticancer therapeutics because allcancers evade apoptosis [36]. P19 is a unique molecule which in-duced apoptosis by interfering with many apoptosis signal trans-duction pathways and important cell survival molecular targets
nd PARP-1. Cells were incubated with or without 5 mM NAC before treatment with0% gel was used for analysis of NF-jB and IjB while b-actin was used as internal
n the expression of STAT3 and surviving. For STAT3 whole cell lysates were resolvedurvivin was calculated using Quantity One software of Bio-RAD.

214 A. Kumar et al. / Chemico-Biological Interactions 193 (2011) 204–215
investigated in HL-60 cells. Its anti-cancer activity was evidentfrom doubling of the survival time of L1210 leukemic mice andinhibition of Sarcoma-180 (solid) and Ehrlich ascites tumor (EAT)models in mice.
The key factor ascribed to cell cytotoxicity is the nitric oxide(NO) produced by P19 with parallel induction of iNOS, and we de-scribe that P19 is a selective inducer of NO. Production of NO andconcomitant iNOS activation are selectively inhibited by both sMIT,an iNOS inhibitor and NAC, a potent anti-oxidant. P19 as such didnot appear to produce any ROS but in the presence of iNOS inhib-itor, a substantial amount of superoxide was formed, which per-haps is instantly utilized by an overwhelming amount of NOleading to the production of potent oxidizing and nitrating species.Both good and ugly aspects of NO are well known [37]. In this casehowever, NO primarily appears responsible for the toxic and detri-mental effects in cancer cells. Unlike other sesquiterpene lactonesare known to produce ROS in cancer cells [38]. P19 exhibited mildanti-oxidant activity because it quenched the basal ROS level andsimultaneously raised the GSH content in cells. Again as compareto parthenin toxicity, which is ascribed to adduct formation withglutathione and cysteine [6], P19, on the contrary, did not depleteGSH.
The lactone activated both initiator caspase-8 and -9 and theexecutioner caspase-3 demonstrating thereby the involvement ofboth intrinsic and extrinsic signaling cascades in cell death. Mito-chondria appeared to be a major target of P19 toxicity as it causedmitochondrial membrane potential loss and altered the ratio ofanti-and pro-apoptotic proteins expression. For instance, Bax gotcompletely translocated to mitochondria after P19 treatment andcytochrome c correspondingly was released to cytosol. The releaseof cytochrome c seems to be consistent with the activation of cas-pase-9 and therefore, intrinsic pathway of apoptosis. Completetranslocation of cytochrome c from mitochondria is suggestive ofstrong mitochondrial stress while Bax alone however, may not beable to release all cytochrome c. We therefore, examined the roleof another important protein known for its role in the fission ofmitochondria during stress, which might aid and abet the completetranslocation of cytochrome c. Our results clearly demonstrate atime-dependent increase in the cellular expression of Drp-1, withmaximum expression being at 6 h of treatment; the protein virtu-ally was below the level of detection in the untreated cells.
Further activation of caspase-8 suggests the involvement ofextrinsic pathways of apoptosis induced by P19, which seems tobe regulated by upstream over expression of DR4 and DR5 amongother death receptors studied. Caspase-8 activation reportedlytruncates Bid to tBid responsible for cross talk between extrinsicand intrinsic signaling pathways of apoptosis and this was evi-denced from more than 80% Bid disappeared from cytosol presum-ably to tBid which got translocated to mitochondria where inassociation with Bax it forms mitochondrial membrane pores forallowing cytochrome c release to cytosol [39]. When NO produc-tion was inhibited by NAC, the cytosolic level of Bid was restoredto normal. This clearly suggested that P19 strongly abrogates mito-chondrial functions because of NO mediated stress. Further, mito-chondrial stress produced by P19 also leads to AIF translocationfrom mitochondria to the nucleus. In this concern, our data sug-gested that AIF contributes significantly towards the induction ofapoptosis by P19, and this was confirmed by the fact that P19 in-duced apoptosis could not be blocked completely by pre-treatingcells with pan caspase inhibitor. On the other hand, siRNA usedto knock-down the expression of AIF could also substantially blockapoptosis. Therefore, our data clearly suggested that P19 utilizesAIF and caspases together to induce apoptosis in HL-60 cells. Weare unable to explain the low cytotoxic effect of P19 in p53 wildtype HeLa cells, which warrant further separate studies, whenp53 is also known to play a role in the translocation of AIF to the
nucleus during the induction of apoptosis. Contrary to these, somerecent studies however, suggested also a p53- independent trans-location of AIF to the nucleus [40]. Our present studies neverthe-less, clearly predicted that P19 activated both caspases- and AIF-dependent apoptosis in HL-60 cells.
There are several transcriptional factors involved in the activa-tion of iNOS, while some earlier studies have amply demonstratedthe role of tumor necrosis factor-related apoptosis-inducing ligand(TRAIL or DR) as a key component in the activation of iNOS andstimulation of caspase-3 in mediating apoptosis in human hemato-logic malignancies derived cell lines [41]. Our studies also supportthis fact that among the four apical death receptors examined, P19selectively stimulated TRAIL-R1 (DR4) and TRAIL-R2 (DR5), theinhibition of which by NAC reversed iNOS activation and henceNO formation. Thus P19 mediated DR4 and DR5 activation is onefactor, which promotes the cytotoxicity of leukemia cell line bystimulating NO and executioner caspase-3. Unlike constitutivelyhigher level of TNFR1 and Fas receptor in HL-60 cells, DR-4 andDR-5 are poorly expressed in these cells. Therefore, activation ofthe later selectively by P19 may be responsible for activatingextrinsic/caspase-8 dependent apoptosis.
Again to explore the cause and effect of increased activity ofiNOS we analyzed its transcription factor NF-jB (p65/p50) andits transactivator PARP1 [42], that could affect the expression ofiNOS. We observed a sharp decline in the expression of p65 sub-unit and its cleavage irrespective of its location in the cell com-partment. P19 caused a time-dependent inhibition of NF-jB/p65translocation into the nucleus with its corresponding parallel risein the cytosol presumably because of its stabilization due to NF-jB–Ijb complex formation. Further, P19 induced PARP1 cleavageoccurred simultaneous to NF-jB inhibition when these events in-versely correlated with the expression of iNOS and generation ofNO. This clearly indicated that neither NF-jB nor PARP1 may beresponsible for the enhanced expression of iNOS however; weare still trying to find out the factors responsible for iNOS activa-tion. TNFR1 is also known to regulate the activation of NF-jBthrough activating IKK, which facilitates the degradation of Ijbthrough phosphorylation [43]. The inhibitory effect of P19 onTNFR1 expression, therefore, corroborates with the decreasedactivity of NF-jB. Again, NF-jB has been shown to up-regulatethe expression of FasR (CD95) [44] by binding to its promoter re-gion. Decreased expression of CD95 could also be related to NF-jBinhibition. Restoration of NF-jB, TNFR1 and CD95 after treatmentwith NAC suggested that expression of all the three proteins is ef-fected by NO.
In addition, P19 also targets proteins like STAT3 and survivin,which regulate cancer cell survival by a different mechanism andare being exploited independently for development of anticancertherapeutics [19]. STATs play an important role at all levels oftumorigenesis and are responsible for generating pro-proliferativesignals like survivin and cyclin D1 and up-regulate anti-apoptoticproteins, e.g. Bcl-XL and Bcl-2. P19 mediated inhibition of pSTAT3corroborated with strong inhibition of pro-survival proteinsurvivin.
In conclusion, P19 is a novel semi-synthetic analogue whichpossesses strong anti-cancer activity mediated by NO inducedapoptosis in HL-60 cells. Among all the targets of P19, NF-jB andSTAT3 seem to be most important due to their central role in tumorprogression. Our studies suggest that P19 is a multi-target anti-cancer therapeutic lead which may be found useful in treating can-cer diseases.
Conflict of interest statement
The authors declare that they have no conflict of interest.

A. Kumar et al. / Chemico-Biological Interactions 193 (2011) 204–215 215
Acknowledgments
We are thankful to Dr. Sarang Bani and Prashant Singh for theirhelp in the use of flow cytometer. Thanks are due to the Council ofScientific and Industrial Research, India, for financial support forthis work and for senior research fellowship to Ajay Kumar.
References
[1] G. Klein, Cancer, apoptosis, and non-immune surveillance, Cell Death Differ. 11(2004) 13–17.
[2] K.H. Lee, Anticancer drug design based on plant-derived natural products, J.Biomed. Sci. 6 (1999) 236–250.
[3] G.H.N. Towers, J.C. Mitchel, E. Rodriguez, D.F. Bannet, R.P.V. Subba, Biology andchemistry of Parthenium hysterophorus L., a problem weed in India, J. Sci. Ind.Res. India 36 (1977) 672–684.
[4] G.L. Sharma, K.K. Bhutani, Plant based antiamoebic drugs. Part II. Amoebicidalactivity of parthenin isolated from Parthenium hysterophorus, Planta Med. 54(1988) 20–22.
[5] A. Ramos, R. Rivero, A. Visozo, J. Piloto, A. Garcia, Parthenin, a sesquiterpenelactone of Parthenium hysterophorus L. is a high toxicity clastogen, Mutat. Res.514 (2002) 19–27.
[6] A.K. Pickman, G.H.N. Towers, P.V.S. Rao, Coronopilin-another majorsesquiterpene lactone in Parthenium hysterophorus, Phytochemistry 19(1980) 2206–2207.
[7] D. Mew, F. Balza, G.H.N. Towers, J.G. Levy, Anti-tumor effects of thesequiterpene lactone parthenin, Planta Med. 49 (1982) 23–27.
[8] G.S. Wu, W.S. El-Deiry, Apoptotic death of tumor cells correlates withchemosensitivity, independent of p53 or bcl-2, Clin. Cancer Res. 2 (1996)623–633.
[9] L.J. Ignarro, G.M. Buga, K.S. Wood, R.E. Byrns, G. Chaudhuri, Endothelium-derived relaxing factor produced and released from artery and vein is nitricoxide, Proc. Natl. Acad. Sci. 84 (1987) 9265–9269.
[10] C. Nathan, Nitric oxide as secretory product of mammalian cells, FASEB J. 6(1992) 3051–3064.
[11] S. Cui, J.S. Reichner, R.B. Mateo, J.E. Albina, Activated murine macrophagesinduce apoptosis in tumor cells through nitric oxide-dependent or -independent mechanisms, Cancer Res. 54 (1994) 2462–2467.
[12] D.S. Tendler, C. Bao, T. Wang, E.L. Huang, E.A. Ratovitski, D.A. Pardoll, C.J.Lowenstein, Intersection of interferon and hypoxia signal transductionpathways in nitric oxide-induced tumor apoptosis, Cancer Res. 61 (2001)3682–3688.
[13] J.I. Chao, P.C. Kuo, T.S. Hsu, Down-regulation of survivin in nitric oxide-inducedcell growth inhibition and apoptosis of the human lung carcinoma cells, J. Biol.Chem. 279 (2004) 20267–20276.
[14] S. Bhushan, J. Singh, J.M. Rao, A.K. Saxena, G.N. Qazi, A novel lignancomposition from Cedrus deodara induces apoptosis and early nitric oxidegeneration in human leukemia Molt-4 and HL-60 cells, Nitric Oxide-Biol.Chem. 14 (2006) 72–88.
[15] Z.M. Shao, W.J. Radziszewski, S.H. Barsky, Tamoxifen enhances myoepithelialcell suppression of human breast carcinoma progression in vitro by twodifferent effector mechanisms, Cancer Lett. 157 (2000) 133–144.
[16] G. Sethi, B. Sung, B.B. Aggarwal, Nuclear factor-jB activation: from bench tobedside, Exp. Biol. Med. 233 (2008) 21–31.
[17] K.S. Ahn, B.B. Aggarwal, Transcription factor NF-kappaB: a sensor for smokeand stress signals, Ann. NY Acad. Sci. 1056 (2005) 218–233.
[18] H. Yu, D. Pardoll, R. Jove, STATs in cancer inflammation and immunity: aleading role for STAT3, Nat. Rev. Can. 9 (2009) 798–809.
[19] K. Schlessinger, D.E. Levy, Malignant transformation but not normal cellgrowth depend on STAT3, Cancer Res. 65 (2005) 5828–5834.
[20] A. Kumar, F. Malik, S. Bhushan, V.K. Sethi, A.K. Shahi, J. Kaur, S.C. Taneja, G.N.Qazi, J. Singh, An essential oil and its major constituent isointermedeol induceapoptosis by increased expression of mitochondrial cytochrome c and apicaldeath receptors in human leukaemia HL-60 cells, Chem.-Biol. Interact. 171(2008) 332–347.
[21] F. Malik, A. Kumar, S. Bhushan, S. Khan, A. Bhatia, K.A. Suri, G.N. Qazi, J. Singh,Reactive oxygen species generation and mitochondrial dysfunction in theapoptotic cell death of human myeloid leukemia HL-60 cells by a dietarycompound with aferin A with concomitant protection by N-acetyl cysteine,Apoptosis 12 (2007) 2115–2133.
[22] B.A. Shah, S.C. Taneja, V.K. Sethi, P. Gupta, S.S. Andotra, S.S. Chimni,G.N. Qazi, The formation of novel 1,3-dioxolanes: atypical: Baylis–Hillman reaction of a sesquiterpene lactone parthenin, Tetrahedron Lett.48 (2007) 955–960.
[23] M. Nigam, V. Ranjan, S. Srivastava, R. Sharma, A. Balapure, Centchromaninduces G0/G1 arrest and caspase-dependent apoptosis involvingmitochondrial membrane depolarization in MCF-7 and MDA MB-231 humanbreast cancer cells, Life Sci. 82 (2008) 577–590.
[24] R.I. Geran, N.H. Greenberg, M.M. Macdonald, A.M. Schumacher, B.J. Abbott,Protocols for screening chemical agents and natural products against animaltumours and other biological systems (third ed.), Cancer Chemother Rep. 3(1972) 1–103.
[25] S. Nagata, Apoptosis by death factor, Cell 88 (1997) 355–365.[26] T.P. Szatrowski, C.F. Nathan, Production of large amounts of hydrogen peroxide
by human tumor cells, Cancer Res. 51 (1991) 794–798.[27] M. Adachi, Y. Zhang, X. Zhao, T. Minami, R. Kawamura, Y. Hinoda, K. Imai,
Synergistic effect of histone deacetylase inhibitors FK228 and m-carboxycinnamic acid bis-hydroxamide with proteasome inhibitors PSI andPS-341 against gastrointestinal adenocarcinoma cells, Clin. Cancer Res. 10(2004) 3853–3862.
[28] D.A. Wink, K.M. Miranda, M.G. Espey, R.M. Pluta, S.J. Hewett, C. Colton, M.Vitek, M. Feelisch, M.B. Grisham, Mechanisms of the antioxidant effects ofnitric oxide, Antioxid. Redox Signling 3 (2001) 203–213.
[29] S. Bergamini, C. Rota, R. Canali, M. Staffieri, F. Daneri, A. Bini, F. Giovannini, A.Tomasi, A. Iannone, N-acetylcysteine inhibits in vivo nitric oxide production byinducible nitric oxide synthase, Nitric Oxide 5 (2001) 349–360.
[30] J.L. Bodmer, N. Holler, S. Reynard, P. Vinciguerra, P. Schneider, P. Juo, J. Blenis, J.Tschopp, TRAIL receptor-2 signals apoptosis through FADD and caspase-8, Nat.Cell Biol. 4 (2000) 241–243.
[31] N.N. Danial, S.J. Korsmeyer, Cell death: critical control points, Cell 116 (2004)205–219.
[32] X. Wang, The expanding role of mitochondria in apoptosis, Genes Dev. 15(2001) 2922–2933.
[33] V. Parra, V. Eisner, M. Chiong, A. Criollo, F. Moraga, A. Garcia, S. Härtel, E.Jaimovich, A. Zorzano, C. Hidalgo, S. Lavandero, Changes in mitochondrialdynamics during ceramide-induced cardiomyocyte early apoptosis,Cardiovasc. Res. 77 (2008) 387–397.
[34] H. Nakajima, H. Nagaso, N. Kakui, M. Ishikawa, T. Hiranuma, S. Hoshiko, Criticalrole of the automodification of poly (ADP-ribose) polymerase-1 in nuclearfactor-kappaB-dependent gene expression in primary cultured mouse glialcells, J. Biol. Chem. 279 (2004) 42774–42786.
[35] F. Li, G. Ambrosini, E.Y. Chu, J. Plescia, S. Tognin, P.C. Marchisio, D.C. Altieri,Control of apoptosis and mitotic spindle checkpoint by survivin, Nature 396(1998) 580–584.
[36] D. Hanahan, R.A. Weinberg, The hallmarks of cancer, Cell 100 (2000) 57–70.[37] J.S. Beckman, W.H. Koppenol, Nitric oxide, superoxide and peroxinitrite: the
good, the bad and ugly, Am. J. Physiol. 271 (1996) C1424–C1437.[38] W. Wang, M. Adachi, R. Kawamura, H. Sakamoto, T. Hayashi, T. Ishida, K. Imai,
Y. Shinomura, Parthenolide-induced apoptosis in multiple myeloma cellsinvolves reactive oxygen species generation and cell sensitivity depends oncatalase activity, Apoptosis 11 (2006) 2225–2235.
[39] R. Eskes, S. Desagher, B. Antonsson, J.C. Martinou, Bid induces theoligomerization and insertion of bax into the outer mitochondrialmembrane, Mol. Cell Biol. 20 (2000) 929–935.
[40] B. Ory, F. Blanchard, S. Battaglia, F. Gouin, F. Redini, D. Heymann, Zoledronicacid activates the DNA S-phase checkpoint and induces osteosarcoma celldeath characterized by apoptosis-inducing factor and endonuclease-Gtranslocation independently of p53 and retinoblastoma status, Mol.Pharmacol. 71 (2007) 333–343.
[41] P. Secchiero, A. Gonelli, C. Celeghini, P. Mirandola, L. Guidotti, G. Visani, S.Capitani, G. Zauli, Activation of the nitric oxide synthase pathway represents akey component of tumor necrosis factor-related apoptosis-inducing ligand-mediated cytotoxicity on hematologic malignancies, Blood 98 (2001) 2220–2228.
[42] S. Beneke, A. Bürkle, Poly (ADP-ribosyl)ation in mammalian ageing, NucleicAcids Res. 35 (2007) 7456–7465.
[43] G. Chen, D.V. Goeddel, TNF-R1 signaling: a beautiful pathway, Science 296(2002) 1634–1635.
[44] H. Chan, D.P. Bartos, L.B. Owen-Schaub, Activation-dependent transcriptionalregulation of the human fas promoter requires NF-jB p50-p65 recruitment,Mol. Cell Biol. 19 (1999) 2098–2108.
Related Documents











