Carcinogenesis vol.30 no.12 pp.2014–2022, 2009 doi:10.1093/carcin/bgp249 Advance Access publication October 20, 2009 A novel oncolytic adenovirus selectively silences the expression of tumor-associated STAT3 and exhibits potent antitumoral activity Zhiqiang Han y , Zhenya Hong y , Caihong Chen, Qinglei Gao, Danfeng Luo, Yong Fang, Yang Cao, TaoZhu, Xuefeng Jiang, Quanfu Ma, Wei Li, Lingfei Han, Daowen Wang, Gang Xu, Shixuan Wang, Li Meng, Jianfeng Zhou and Ding Ma Cancer Biology Research Center, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, Hubei 430030, China To whom correspondence should be addressed. Tel: þ86 27 83663351; Fax: þ86 27 83662681; Email: [email protected] Correspondence may also be addressed to Jianfeng Zhou. Tel: þ86 27 83662107; Fax: þ86 27 83662681; Email: [email protected] Tumor cells acquire the ability to proliferate uncontrollably, resist apoptosis, sustain angiogenesis and evade immune surveillance. Signal transducer and activator of transcription (STAT) 3 regu- lates all of these processes in a surprisingly large number of hu- man cancers. Consequently, the STAT3 protein is emerging as an ideal target for cancer therapy. This paper reports the generation of an oncolytic adenovirus (M4), which selectively blocks STAT3 signaling in tumor cells as a novel therapeutic strategy. M4 selec- tively replicated in tumor cells and expressed high levels of anti- sense STAT3 complementary DNA during the late phase of the viral infection in a replication-dependent manner. The viral prog- eny yield of M4 in tumor cells was much higher than that of the parent adenoviral mutants, Ad5/dE1A. M4 effectively silenced STAT3 and its target genes in tumor cells while sparing normal cells and exhibited potent antitumoral efficacy in vitro and in vivo. Systemic administration of M4 significantly inhibited tumor growth in an orthotopic gastric carcinoma mouse model, elimi- nated abdominal cavity metastases and prolonged survival time. In summary, M4 has low toxicity and great potential as a thera- peutic agent for different types of cancers. Introduction Signal transducer and activator of transcription (STAT) family proteins are transcription factors activated by many cytokines, growth factors and non-receptor tyrosine kinases (1,2). In normal cells, STAT proteins play central roles in cytokine signaling and modulate a variety of bi- ological functions by activating transcription of a diverse set of genes (3–5). Activation of STAT proteins is a reversible and tightly controlled process that typically lasts for a limited duration (6,7). However, con- stitutive activation of STAT proteins, especially STAT3, is detected at high frequency in a large number of human malignancies (6,8,9). Per- sistent activation of the STAT3 protein contributes to multiple malig- nant phenotypes such as aberrant proliferation, angiogenesis, invasion and immune evasion (10–12). Blocking STAT3 signaling has been shown to induce apoptosis in tumor cells, inhibit angiogenesis and enhance antitumor immune-mediated cytotoxicity (10,13,14). Thus, STAT3 has been identified as a potential high-yield target for drug development to treat many cancers (6,15,16). For the past several years, a number of approaches have been employed to block STAT3 signaling pathways. For example, small molecule compounds developed to block STAT3 signaling display potent antitumoral activity (17–19). While the safety and therapeutic efficacy of these promising agents is still await- ing evaluation in clinical trials, some fundamental concerns remain to be addressed. Given that STAT3 is a transcription factor ubiquitously expressed in both malignant and normal tissues, the clinical potential of these STAT3 inhibitors would rely on their tumor specificity and lack of toxicity to normal tissues in vivo. Therefore, it is important to develop alternative approaches for selective targeting of STAT3. Conditionally replicative oncolytic adenoviruses represent a prom- ising new approach in the treatment of cancer (20–22), and initial data obtained from clinical studies using oncolytic viruses have been en- couraging in terms of efficacy with minimal, if any, toxicity (23). A number of oncolytic adenoviruses have been engineered by deleting a small sequence in the E1A- or E1B-encoding regions to attenuate viral replication and cytolysis in normal tissues but not in tumor cells. Other attempts to enhance specific tumor cell lysis, including incorporating therapeutic transgenes into conditionally replicative oncolytic adenoviruses, have substantially augmented antitumoral ef- ficacy both in vitro and in vivo (24). We have previously reported a series of novel oncolytic adenoviral mutants in which we replaced the 6.7K/gp19K region with a fragment of antisense complementary DNA (cDNA) of a target gene in an E1A CR2 region-deleted adeno- viral mutant called Adv5/dE1A (25,26). The strategy of combining the oncolytic ability with the benefits of gene silencing has proven to be effective and has the potential to be generally applicable in a wide range of molecule-based anticancer therapeutics. This paper reports the generation of a novel adenoviral mutant called M4, which was constructed by replacing E3 adenovirus death protein (ADP) in Adv5/ dE1A with an antisense STAT3 cDNA following Hawkins’ gene de- livery system (27). Expression of antisense STAT3 cDNA in M4 mimics the expression kinetics of the native ADP gene and depends strongly on viral DNA replication. M4 showed potent antitumoral efficacy through selective cytolysis and silenced expression of STAT3 and its downstream target genes. Most strikingly, systemic adminis- tration of M4 alone significantly suppressed tumor growth in a human orthotopic gastric carcinoma mouse model and eliminated the dissem- ination in the abdominal cavity. Materials and methods Cell culture Breast cancer (MDA-MB-231), colorectal carcinoma (HCT-8) and lung carci- noma (A549) cell lines were purchased from the American Type Culture Col- lection (Rockville, MD). A prostate carcinoma (PC3M-1E8) cell line was obtained from the China Center for Type Culture Collection (Shanghai, People’s Republic of China). The human gastric cancer cell line (MKN-45) was a gift from Dr Li of the Shanghai Cancer Institute (Shanghai, People’s Republic of China). Human umbilical cord endothelial cells (HUVEC) were isolated as described previously (28). All the cell lines were grown in the recommended medium (GIBCO, Grand Island, NY) supplemented with 10% fetal bovine serum (HyClone, Logan, UT), 2 mmol/L glutamine, 100 U/ml penicillin and 100 lg/ml streptomycin and maintained in a 37°C atmosphere containing 5% CO 2 . Adenoviral mutants Ad5/dE1A with deletion of amino acids 121–129 in E1A CR2 was previously constructed (25). M4 was driven from Ad5/dE1A through replacement of the ADP open reading frame in the E3 region by a fragment of reverse STAT3 cDNA (bases 960–190). Ad5/dE1A/ADP was driven from Ad5/dE1A through deletion of the ADP open reading frame and used as a vector control for M4. The viral mutants were constructed by homologous recombination in the hu- man embryonic kidney 293 (HEK-293) cells. Abbreviations: ADP, adenovirus death protein; cDNA, complementary DNA; CPE, cytopathic effect; HEK-293, human embryonic kidney 293; HUVEC, human umbilical cord endothelial cells; i.t., intratumoral; MMP, matrix metal- loproteinase; MOI, multiplicity of infection; mRNA, messenger RNA; p.i., post-infection; RT-PCR, real-time PCR; siRNA, small interfering RNA; STAT, signal transducer and activator of transcription; TCID50, 50% tissue culture infective dose; wt-Ad5, wild-type adenovirus 5. y These authors contributed equally to this work. Ó The Author 2009. Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected] 2014 Downloaded from https://academic.oup.com/carcin/article/30/12/2014/2392052 by guest on 02 February 2022

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Carcinogenesis vol.30 no.12 pp.2014–2022, 2009doi:10.1093/carcin/bgp249Advance Access publication October 20, 2009
A novel oncolytic adenovirus selectively silences the expression of tumor-associatedSTAT3 and exhibits potent antitumoral activity
Zhiqiang Hany, Zhenya Hongy, Caihong Chen, QingleiGao, Danfeng Luo, Yong Fang, Yang Cao, Tao Zhu,Xuefeng Jiang, Quanfu Ma, Wei Li, Lingfei Han, DaowenWang, Gang Xu, Shixuan Wang, Li Meng, Jianfeng Zhou�
and Ding Ma�
Cancer Biology Research Center, Tongji Hospital, Tongji Medical College,Huazhong University of Science and Technology, Wuhan, Hubei 430030,China
�To whom correspondence should be addressed. Tel: þ86 27 83663351;Fax: þ86 27 83662681;Email: [email protected] may also be addressed to Jianfeng Zhou.Tel: þ86 27 83662107; Fax: þ86 27 83662681;Email: [email protected]
Tumor cells acquire the ability to proliferate uncontrollably, resistapoptosis, sustain angiogenesis and evade immune surveillance.Signal transducer and activator of transcription (STAT) 3 regu-lates all of these processes in a surprisingly large number of hu-man cancers. Consequently, the STAT3 protein is emerging as anideal target for cancer therapy. This paper reports the generationof an oncolytic adenovirus (M4), which selectively blocks STAT3signaling in tumor cells as a novel therapeutic strategy. M4 selec-tively replicated in tumor cells and expressed high levels of anti-sense STAT3 complementary DNA during the late phase of theviral infection in a replication-dependent manner. The viral prog-eny yield of M4 in tumor cells was much higher than that of theparent adenoviral mutants, Ad5/dE1A. M4 effectively silencedSTAT3 and its target genes in tumor cells while sparing normalcells and exhibited potent antitumoral efficacy in vitro and in vivo.Systemic administration of M4 significantly inhibited tumorgrowth in an orthotopic gastric carcinoma mouse model, elimi-nated abdominal cavity metastases and prolonged survival time.In summary, M4 has low toxicity and great potential as a thera-peutic agent for different types of cancers.
Introduction
Signal transducer and activator of transcription (STAT) family proteinsare transcription factors activated by many cytokines, growth factorsand non-receptor tyrosine kinases (1,2). In normal cells, STAT proteinsplay central roles in cytokine signaling and modulate a variety of bi-ological functions by activating transcription of a diverse set of genes(3–5). Activation of STAT proteins is a reversible and tightly controlledprocess that typically lasts for a limited duration (6,7). However, con-stitutive activation of STAT proteins, especially STAT3, is detected athigh frequency in a large number of human malignancies (6,8,9). Per-sistent activation of the STAT3 protein contributes to multiple malig-nant phenotypes such as aberrant proliferation, angiogenesis, invasionand immune evasion (10–12). Blocking STAT3 signaling has beenshown to induce apoptosis in tumor cells, inhibit angiogenesis and
enhance antitumor immune-mediated cytotoxicity (10,13,14). Thus,STAT3 has been identified as a potential high-yield target for drugdevelopment to treat many cancers (6,15,16). For the past several years,a number of approaches have been employed to block STAT3 signalingpathways. For example, small molecule compounds developed to blockSTAT3 signaling display potent antitumoral activity (17–19). While thesafety and therapeutic efficacy of these promising agents is still await-ing evaluation in clinical trials, some fundamental concerns remain tobe addressed. Given that STAT3 is a transcription factor ubiquitouslyexpressed in both malignant and normal tissues, the clinical potential ofthese STAT3 inhibitors would rely on their tumor specificity and lack oftoxicity to normal tissues in vivo. Therefore, it is important to developalternative approaches for selective targeting of STAT3.
Conditionally replicative oncolytic adenoviruses represent a prom-ising new approach in the treatment of cancer (20–22), and initial dataobtained from clinical studies using oncolytic viruses have been en-couraging in terms of efficacy with minimal, if any, toxicity (23). Anumber of oncolytic adenoviruses have been engineered by deletinga small sequence in the E1A- or E1B-encoding regions to attenuateviral replication and cytolysis in normal tissues but not in tumorcells. Other attempts to enhance specific tumor cell lysis, includingincorporating therapeutic transgenes into conditionally replicativeoncolytic adenoviruses, have substantially augmented antitumoral ef-ficacy both in vitro and in vivo (24). We have previously reporteda series of novel oncolytic adenoviral mutants in which we replacedthe 6.7K/gp19K region with a fragment of antisense complementaryDNA (cDNA) of a target gene in an E1A CR2 region-deleted adeno-viral mutant called Adv5/dE1A (25,26). The strategy of combiningthe oncolytic ability with the benefits of gene silencing has proven tobe effective and has the potential to be generally applicable in a widerange of molecule-based anticancer therapeutics. This paper reportsthe generation of a novel adenoviral mutant called M4, which wasconstructed by replacing E3 adenovirus death protein (ADP) in Adv5/dE1A with an antisense STAT3 cDNA following Hawkins’ gene de-livery system (27). Expression of antisense STAT3 cDNA in M4mimics the expression kinetics of the native ADP gene and dependsstrongly on viral DNA replication. M4 showed potent antitumoralefficacy through selective cytolysis and silenced expression of STAT3and its downstream target genes. Most strikingly, systemic adminis-tration of M4 alone significantly suppressed tumor growth in a humanorthotopic gastric carcinoma mouse model and eliminated the dissem-ination in the abdominal cavity.
Materials and methods
Cell culture
Breast cancer (MDA-MB-231), colorectal carcinoma (HCT-8) and lung carci-noma (A549) cell lines were purchased from the American Type Culture Col-lection (Rockville, MD). A prostate carcinoma (PC3M-1E8) cell line wasobtained from the China Center for Type Culture Collection (Shanghai, People’sRepublic of China). The human gastric cancer cell line (MKN-45) was a gift fromDr Li of the Shanghai Cancer Institute (Shanghai, People’s Republic of China).Human umbilical cord endothelial cells (HUVEC) were isolated as describedpreviously (28). All the cell lines were grown in the recommended medium(GIBCO, Grand Island, NY) supplemented with 10% fetal bovine serum(HyClone, Logan, UT), 2 mmol/L glutamine, 100 U/ml penicillin and 100lg/ml streptomycin and maintained in a 37�C atmosphere containing 5% CO2.
Adenoviral mutants
Ad5/dE1A with deletion of amino acids 121–129 in E1A CR2 was previouslyconstructed (25). M4 was driven from Ad5/dE1A through replacement of theADP open reading frame in the E3 region by a fragment of reverse STAT3cDNA (bases 960–190). Ad5/dE1A/ADP was driven from Ad5/dE1A throughdeletion of the ADP open reading frame and used as a vector control for M4.The viral mutants were constructed by homologous recombination in the hu-man embryonic kidney 293 (HEK-293) cells.
Abbreviations: ADP, adenovirus death protein; cDNA, complementary DNA;CPE, cytopathic effect; HEK-293, human embryonic kidney 293; HUVEC,human umbilical cord endothelial cells; i.t., intratumoral; MMP, matrix metal-loproteinase; MOI, multiplicity of infection; mRNA, messenger RNA; p.i.,post-infection; RT-PCR, real-time PCR; siRNA, small interfering RNA; STAT,signal transducer and activator of transcription; TCID50, 50% tissue cultureinfective dose; wt-Ad5, wild-type adenovirus 5.
yThese authors contributed equally to this work.
� The Author 2009. Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected] 2014
Dow
nloaded from https://academ
ic.oup.com/carcin/article/30/12/2014/2392052 by guest on 02 February 2022

Detection of antisense STAT3 cDNA and gp19K/STAT3 cDNA fusionmessenger RNA
A549 cells were infected with virus at an multiplicity of infection (MOI) of 10,cultured for various times and collected for extraction of total RNA. A senseprimer (5#-GCTCACTCACGATGCTTCTC-3#) and an antisense primer (5#-ATTGACCAGCAGTATAGCCG-3#) were used for amplification of antisenseSTAT3 cDNA. For amplification of the fusion transcript containing sequencesof gp19K gene and antisense STAT3 cDNA, a 5#-end sense primer from thecoding region of gp19K (5#-GGTGGTTCTCCATAGCGCTT-3#) and a 3#-endantisense primer complimentary to the antisense STAT3 cDNA (5#-TACA-GACTGCAGCCACTGCG-3#) were used.
TCID50 to determine viral titers for burst assay
A549 cells were infected with different viruses at an MOI of 20. The viral yield wasdetermined by the limiting dilution method, known as the determination of 50%tissue culture infective dose (TCID50) described elsewhere (27). The data fromthree separate infection studies were expressed as plaque-forming unit per milliliterand normalized to virus production per cell based on the number of infected cells.
Real-time polymerase chain reaction
The cells were infected with various viral mutants at an MOI of 10. At varioustime points post-infection (p.i.), total RNA was isolated from the cells andtreated with DNase I for 30 min at 37�C. Real-time PCR (RT-PCR) was per-formed using the SYBR Premix Ex Taq Kit (TaKaRa Biotechnology Co., Ltd,Dalian, China). All fold differences in expression were normalized relative to18s RNA expression in the corresponding sample. Primers used for RT-PCR areas follows: gp19K—forward (F): 5#-TTTAAGGAGCCAGCCTGTAATGT-3#,reverse (R): 5#-GCAGCTTTTCATGTTCTGTGGT-3#; receptor internalizationand degradation (RID)-b—F: 5#-GCAATCATCTCTGTTATGGTGTTCT-3#,R: 5#-GGGGAAAGTTGGGTGGTTC-3#; 14.7K—F: 5#-AGGCCAAAGT-CACCTACGACA-3#, R: 5#-TCCCACCATGACCACCAA-3#; STAT3—F: 5#-CCAGTCAGTGACCAGGCAGAAG-3#, R: 5#-GCACGTACTCCATCGCT-GACA-3#; survivin—F: 5#-AGGTCATCTCGGCTGTTCCTG-3#, R: 5#-TCATCCTCACTGCGGCTGTC-3#; and STAT5a—F: 5#-AGGACTA-CAGTGGCCTGTCC-3#, R: 5#-GTGCTTCTTCAACACCTCCA-3#.
Western blot analysis
Standard western blot analysis was performed with antibodies for STAT3, phos-pho-STAT3 (Tyr705), vascular endothelial growth factor, matrix metalloproteinase(MMP)-9, c-myc, survivin, b-actin (Santa Cruz, CA) and Bcl-xL (Cell SignalingTechnology, Danvers, MA). The proteins were detected by using the Pierce(Pierce Biotechnology, Rockford, IL)-enhanced chemiluminescence system.
Apoptosis assay
Fluorescein isothiocyanate-annexin V and propidium iodide staining (BD Bio-sciences Pharmingen, San Diego, CA) were used to determine apoptosis ac-cording to our previously described methods (25).
Matrigel invasion assay
Twenty-four hours p.i., MDA-MB-231 or HCT-8 cells infected with the in-dicated viruses at an MOI of 10 were trypsinized and counted, and 2 � 105
cells were placed in matrigel-coated transwell inserts (8 lm pores). Cells wereallowed to invade through the matrigel for 22 h. Then, cells in the upperchamber were removed by using cotton swabs. Cells adhered to the outersurface that had invaded through the matrigel were fixed and stained withcrystal violet solution (Sigma Chemical Co., St Louis, MO). Cells werecounted at a magnification of �400 in 10 randomly selected fields, and themean number of cells per field was recorded.
In vitro angiogenesis evaluation
In vitro formation of tubular structures was studied on extracellular matrix asdescribed below. Briefly, 24-well plates were coated with cold matrigel (80 llper well of a solution of 9:1 EC Matrigel to 10� buffer). Then, 1 � 105
HUVEC cells suspended in the conditioned media collected from cell culturesof PC3M-1E8 or MKN-45 cells infected with 10 MOI of various viral mutantswere seeded in each well. After 16 h incubation, angiogenesis was assessedbased on formation of capillary-like structures. The number of capillary-liketubes was counted in five randomly chosen microscopic fields (Nikon Eclipse,TE2000-U), and the average was calculated.
Small interfering RNA preparation
Small interfering RNA (siRNA) against human STAT3 (5#-GGAGCAGCACCUU-CAGGAUTT-3#) was synthesized according to the published sequence (29), andnon-targeting control siRNA (5#-UACCGACUAAAGACAUCAAUU-3#) washigh-performance liquid chromatography purified. RNA oligonucleotides wereordered from Invitrogen (Shanghai, People’s Republic of China).
Mouse tumor models
Athymic BALB/c nude mice were purchased from the Animal ExperimentalCenter of Slaccas (Shanghai, People’s Republic of China). The 4- to 6-week-old mice were maintained in laminar flow cabinets under specific pathogen-free conditions. In the direct intratumoral (i.t.) injection studies, 3 � 106
PC3M-1E8 cells or 1 � 106 MKN-45 cells were suspended in 100 ll normalsaline, injected subcutaneously in the right flanks of mice and allowed to de-velop into a tumor of 4–6 mm in diameter before treatment. Then, 1 � 108 pfudoses of each virus were injected directly into tumors once daily for fiveconsecutive days (n 5 5 for each group). Tumors were measured in twodimensions with calipers twice weekly, and the volume was calculated aslength � width2 � 0.52. The mice were killed when the tumor volumewas .800 mm3 for PC3M-1E8 and 1800 mm3 for MKN-45. A parallel exper-iment was performed to determine STAT3 and its target protein levels (n 5 2for each group). For siRNA injection, 6 lg of siRNA with polyethyleniminecomplexes were injected into tumors once daily for five consecutive days (n53 for each group). Tumors were harvested from mice 3 or 15 days after in-jection and analyzed for STAT3 expression. The orthotopic gastric cancermouse model was established as described previously (30). Ten days afterorthotopic implantation, mice were randomly assigned into four groups (n 515 each group), and 2 � 108 pfu of virus was injected into the tail vein daily forfive consecutive days. Five mice were selected randomly from each group toexamine tumor growth and metastases and were then killed on day 42 afterimplantation. Ten mice were monitored for death and were euthanized whenthey appeared moribund. A parallel experiment was performed to determinein vivo viral replication or endogenous STAT3 protein levels (n 5 2 for eachgroup).
Immunohistochemistry and in situ hybridization
Immunohistochemical labeling of tissues and in situ hybridization stainingwere performed as described previously (30).
Statistical analysis
Statistical values are presented as mean ± SEM. The significance of differencesbetween groups was assessed by the two-tailed Student’s t-test. Statisticalsignificance was defined as P ,0.05. Survival was analyzed by the Kaplan–Meier method and differences were analyzed by a log-rank test.
Results
Construction and confirmation of M4
Two artificial features were combined into one wild-type adenovirus 5(wt-Ad5) genome to generate the M4 mutant adenovirus (Figure 1A).First, a 27 bp sequence from wt-Ad5 bases 920–946 was deleted togenerate Ad5/dE1A, which expresses a mutant E1A protein lackingthe CR2 domain necessary for pRb binding. Second, the E3 transcrip-tion unit of the Ad5/dE1A genome, corresponding to wt-Ad5 bases29 477–29 714 known to encode the ADP protein, was selectivelyexcised and substituted by a fragment of reverse STAT3 cDNA (bases960–190) to generate M4. In M4, antisense STAT3 cDNA was fused tothe 3# portion of the E3 gp19K gene to yield a single chimeric tran-script. Expression of the chimeric transcript was detected in A549cells infected with M4 (Figure 1B). The genomic structure of M4was further verified by restriction digest with ClaI, and the appearanceof a 0.77 kb-long band confirmed the insertion of STAT3 cDNA anddeletion in the E1A region (Figure 1C).
Antisense STAT3 cDNA expression from M4 mimicks theADP expression pattern
To test whether the expression pattern of reverse STAT3 cDNA in M4mimicked that of the native ADP gene, A549 cells were infected withM4 or wt-Ad5. The expression pattern of the antisense STAT3 trans-gene was determined and compared with that of the native ADP geneover the course of the viral infections. The transcriptional expressionof antisense STAT3 cDNA in M4 was nearly undetectable 12 h p.i. butwas at a high level 36 h after infection, similar to the expressionpattern of ADP (Figure 2A). In wt-Ad5, native ADP expression ishighly dependent on viral replication. To address whether reverseSTAT3 cDNA expression from M4 retained this feature, its expressionwas analyzed in the presence or absence of Ara-C, a viral DNAreplication inhibitor. To validate the experimental conditions, the na-tive adenoviral genes pVIII and gp19K were included as positive and
Oncolytic adenovirus silences the expression of STAT3
2015
Dow
nloaded from https://academ
ic.oup.com/carcin/article/30/12/2014/2392052 by guest on 02 February 2022

negative controls, respectively. pVIII is a structural gene from theadenoviral late transcription unit L4, and its expression is completelydependent on viral DNA replication. In contrast, gp19K is a viral earlyresponse gene whose transcription is much less dependent on DNAreplication. In the absence of Ara-C, transcripts of pVIII, gp19K or theantisense STAT3 were readily detected in A549 cells infected withM4, and these genes were expressed at levels similar to those obtainedwith wt-Ad5 infection (Figure 2B). In the presence of Ara-C, thetranscription of pVIII and ADP was almost completely abolished,whereas the expression of gp19K was not significantly changed.As expected, the expression of antisense STAT3 cDNA from M4mimicked the pattern of the native ADP gene and was almost com-pletely inhibited by the addition of Ara-C (Figure 2B). Next, wetested whether the transgene in M4 could be expressed in replica-tion-non-permissive cells. HUVECs were infected with wt-Ad5 orM4 and examined for transcript levels of ADP or antisense STAT3.While the transcripts of gp19K and ADP were easily detected in wt-Ad5-infected HUVECs, only gp19K transcripts were detected in M4-infected HUVECs, and antisense STAT3 cDNA transcripts werenearly undetectable (Figure 2C).
Expression of genes adjacent to the site of ADP transgene insertion ismaintained in M4
In wt-Ad5, the expression timing and levels of E3 genes are controlledby a number of cis signals. One of these signals is encoded within theADP gene (31). To address whether the artificial modification in M4altered the expression of E3 genes adjacent to the ADP gene, A549cells were infected with various viral mutants and the expression ofthe E3 genes gp19K, RIDb and 14.7 K, which surround the ADP gene,was examined by RT-PCR. Although there was a minor difference atdifferent times p.i., the expression timing and levels of the gp19K,RIDb and 14.7 K genes in Ad5/dE1A/ADP and M4 infection weresimilar to those seen in the wt-Ad5 infection (Figure 2D).
Fig. 1. Analyses of adenoviral vector M4. (A) A 27 nucleotide sequencefrom Ad5 bases 920 to 946, corresponding to the amino acid sequence of theE1A necessary for Rb protein binding, was deleted to generate Ad5/dE1A. Asingle region corresponding to E3 ADP from Ad5 bases 29477 to 29714 wasexcised from the Ad5/dE1A genome and substituted by a fragment of reverseSTAT3 cDNA (bases 960–190) with a ClaI restriction site introduced at eachend to generate M4. (B) Amplification of fusion mRNA containingsequences of the gp19K gene and antisense STAT3 cDNA in A549 cells 48 hafter infection with M4, wt-Ad5, Ad5/dE1A or Ad5/dE1A/ADP. (C) Thegenomic structure of M4 was confirmed by ClaI digestion. Because thenative ClaI restriction site (bp 917–922) was disrupted, the M4 genomecontained only two ClaI restriction sites located at the 5# end and 3# end ofthe reverse STAT3 cDNA. The appearance of a 0.77 kb band verified thepresence of the inserted STAT3 cDNA and deletion in the E1A region.
Fig. 2. Expression of antisense STAT3 cDNA and surrounding E3 genes in M4-infected cells. (A) A549 cells were infected with wt-Ad5 or M4 at an MOI of10 and harvested at the indicated time points p.i. Expression of ADP from wt-Ad5 or antisense STAT3 cDNA from M4 was detected by semiquantitative RT-PCR.(B) A549 cells infected with wt-Ad5 or M4 at an MOI of 10 in the presence or absence of Ara-C. Expression of gp19K, pVIII, ADP or antisense STAT3cDNA was determined by semiquantitative RT-PCR at the indicated time points p.i. (C) Semiquantitative RT-PCR examination of gp19K, pVIII, ADP andantisense STAT3 cDNA in HUVEC cells infected with wt-Ad5 or M4 at the indicated time points p.i. (D) RT-PCR examination of gp19K, receptor internalizationand degradation (RID)-b and 14.7 K in A549 cells infected with Ad5, Ad5/dE1A, Ad5/dE1A/ADP or M4 at the indicated time points p.i.
Z.Han et al.
2016
Dow
nloaded from https://academ
ic.oup.com/carcin/article/30/12/2014/2392052 by guest on 02 February 2022

M4 specifically depletes the STAT3 protein and its downstream targetgenes in cancerous but not in normal cells
To assess the efficacy of M4 in downregulating endogenous STAT3protein in cancer cells, PC3M-1E8 and MKN-45 cells were infectedwith M4 at an MOI of 10. Immunoblot analysis of whole-cell lysatesshowed significant reduction in expression of STAT3, p-STAT3 and itstarget proteins (Figure 3A and B) in a time-dependent manner. In con-trast, infection of normal cells (HUVEC) with M4 at an MOI of 100 didnot change the STAT3 protein levels (Figure 3C). These results dem-onstrate the selectivity of M4 to inhibit STAT3 expression in cancercells. In order to rule out the possibility that the reduction in theseproteins was the result of their degradation during apoptosis and hadnothing to do with specific downregulation of these genes by M4, weexamined the level of another endogenous protein, STAT5a. The resultsshow that the level of STAT5a protein was not affected in M4-infectedcells (Figure 3D). Moreover, c-myc and survivin, which are short-livedproteins, were not reduced by Ad5/dE1A or Ad5/dE1A/ADP even at96 h p.i. (Figure 3A and B). For M4 to reduce STAT3 target proteins,translation of STAT3 messenger RNA (mRNA) must be inhibitedthereby reducing the transcription of STAT3 target genes. We next de-termined the transcriptional level of STAT3, survivin and STAT5a inM4-infected cells by RT-PCR. The results show that significant down-
regulation of STAT3 mRNA levels was observed in the M4-infected cells and the decrease of survivin mRNA levels correspondedto the downregulation of STAT3, but the STAT5a mRNA levels were notsignificantly changed (Figure 3E). The data above show the specificityof M4 to selectively inhibit STAT3 and its target proteins in cancer cells.
Cytolytic potency of M4 and viral yield
In wt-Ad5, the ADP gene promotes the death of infected cells andparticipates in the release of the virus (32). To address how the sub-stitution of antisense STAT3 cDNA for the ADP gene altered cytolyticpotency, Ad5/dE1A, Ad5/dE1A/ADP and M4 were analyzed for theability to induce a cytopathic effect (CPE) in A549 cells. While nearlyall Ad5/dE1A-infected cells rounded up and detached by 72 h p.i.,many of the cells infected with Ad5/dE1A/ADP remained attachedand intact (Figure 4A). CPE was delayed in cells infected with M4compared with Ad5/dE1A but was earlier than those infected withAd5/dE1A/ADP (Figure 4A). In primary normal HUVECs, wt-Ad5caused complete CPE by 12 d p.i. at an MOI of 0.1. In contrast, M4,Ad5/dE1A and Ad5/dE1A/ADP exhibited no detectable CPE inHUVECs at an MOI of 100 (Figure 4B). To address whether theweakened CPE gave rise to a higher yield of progeny virions, A549cells were infected with Ad5/dE1A, Ad5/dE1A/ADP or M4, and then,
Fig. 3. Expression of STAT3 and downstream genes in cancer cells and normal primary cells infected with M4. PC-3M-1E8, MKN-45 or HUVEC cellswere infected with various viral mutants at an MOI of 10 and cultured for the designated time. Protein or mRNA was then extracted and subjected to western blotor real-time PCR analysis, respectively. Western blot analysis showed that STAT3 and its downstream targets were downregulated in both cancer cell lines infectedwith M4 but not those infected with Ad5/dE1A or Ad5/dE1A/ADP (A and B). In contrast, M4 had no effect on STAT3 expression in HUVECs (C). On theother hand, M4 did not affect the levels of other endogenous proteins, such as STAT5a and b-actin (D). RT-PCR analysis showed that a significant downregulationof STAT3 and survivin mRNA levels in the M4-infected cancer cells, but the mRNA levels of STAT5a were not changed significantly. The mRNA expressionof the genes in both kinds of cells at 0 h p.i. was set as 1 (E).
Oncolytic adenovirus silences the expression of STAT3
2017
Dow
nloaded from https://academ
ic.oup.com/carcin/article/30/12/2014/2392052 by guest on 02 February 2022
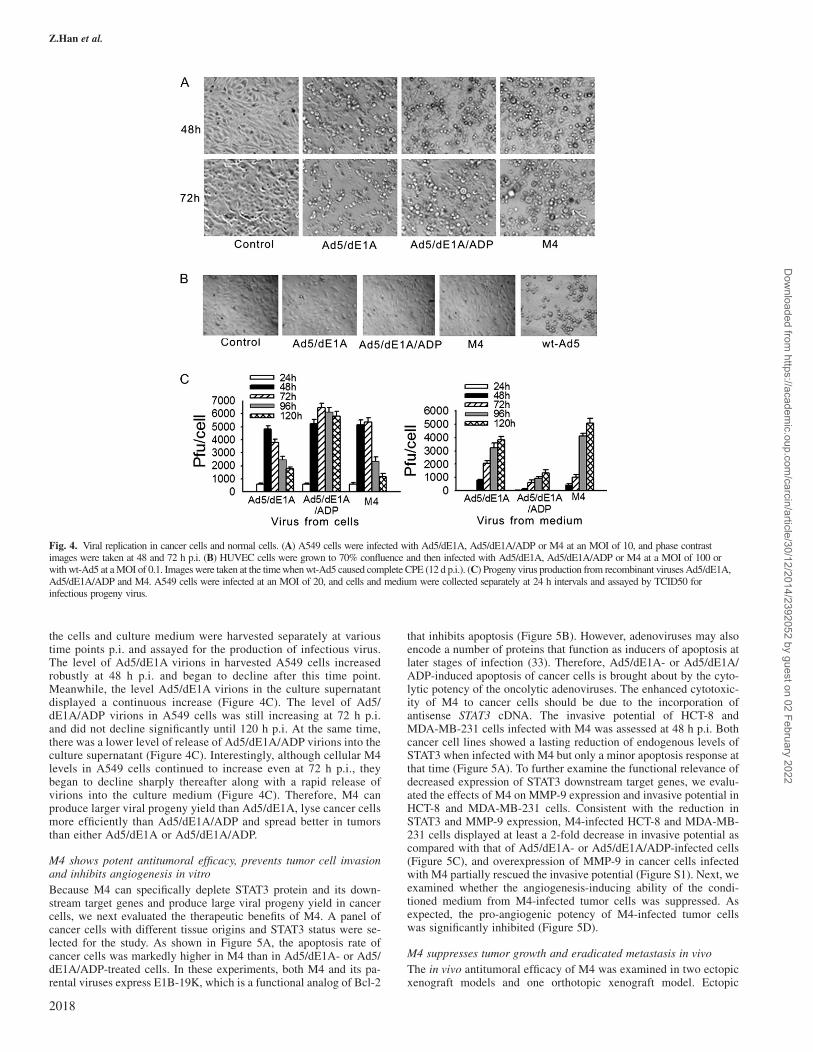
the cells and culture medium were harvested separately at varioustime points p.i. and assayed for the production of infectious virus.The level of Ad5/dE1A virions in harvested A549 cells increasedrobustly at 48 h p.i. and began to decline after this time point.Meanwhile, the level Ad5/dE1A virions in the culture supernatantdisplayed a continuous increase (Figure 4C). The level of Ad5/dE1A/ADP virions in A549 cells was still increasing at 72 h p.i.and did not decline significantly until 120 h p.i. At the same time,there was a lower level of release of Ad5/dE1A/ADP virions into theculture supernatant (Figure 4C). Interestingly, although cellular M4levels in A549 cells continued to increase even at 72 h p.i., theybegan to decline sharply thereafter along with a rapid release ofvirions into the culture medium (Figure 4C). Therefore, M4 canproduce larger viral progeny yield than Ad5/dE1A, lyse cancer cellsmore efficiently than Ad5/dE1A/ADP and spread better in tumorsthan either Ad5/dE1A or Ad5/dE1A/ADP.
M4 shows potent antitumoral efficacy, prevents tumor cell invasionand inhibits angiogenesis in vitro
Because M4 can specifically deplete STAT3 protein and its down-stream target genes and produce large viral progeny yield in cancercells, we next evaluated the therapeutic benefits of M4. A panel ofcancer cells with different tissue origins and STAT3 status were se-lected for the study. As shown in Figure 5A, the apoptosis rate ofcancer cells was markedly higher in M4 than in Ad5/dE1A- or Ad5/dE1A/ADP-treated cells. In these experiments, both M4 and its pa-rental viruses express E1B-19K, which is a functional analog of Bcl-2
that inhibits apoptosis (Figure 5B). However, adenoviruses may alsoencode a number of proteins that function as inducers of apoptosis atlater stages of infection (33). Therefore, Ad5/dE1A- or Ad5/dE1A/ADP-induced apoptosis of cancer cells is brought about by the cyto-lytic potency of the oncolytic adenoviruses. The enhanced cytotoxic-ity of M4 to cancer cells should be due to the incorporation ofantisense STAT3 cDNA. The invasive potential of HCT-8 andMDA-MB-231 cells infected with M4 was assessed at 48 h p.i. Bothcancer cell lines showed a lasting reduction of endogenous levels ofSTAT3 when infected with M4 but only a minor apoptosis response atthat time (Figure 5A). To further examine the functional relevance ofdecreased expression of STAT3 downstream target genes, we evalu-ated the effects of M4 on MMP-9 expression and invasive potential inHCT-8 and MDA-MB-231 cells. Consistent with the reduction inSTAT3 and MMP-9 expression, M4-infected HCT-8 and MDA-MB-231 cells displayed at least a 2-fold decrease in invasive potential ascompared with that of Ad5/dE1A- or Ad5/dE1A/ADP-infected cells(Figure 5C), and overexpression of MMP-9 in cancer cells infectedwith M4 partially rescued the invasive potential (Figure S1). Next, weexamined whether the angiogenesis-inducing ability of the condi-tioned medium from M4-infected tumor cells was suppressed. Asexpected, the pro-angiogenic potency of M4-infected tumor cellswas significantly inhibited (Figure 5D).
M4 suppresses tumor growth and eradicated metastasis in vivo
The in vivo antitumoral efficacy of M4 was examined in two ectopicxenograft models and one orthotopic xenograft model. Ectopic
Fig. 4. Viral replication in cancer cells and normal cells. (A) A549 cells were infected with Ad5/dE1A, Ad5/dE1A/ADP or M4 at an MOI of 10, and phase contrastimages were taken at 48 and 72 h p.i. (B) HUVEC cells were grown to 70% confluence and then infected with Ad5/dE1A, Ad5/dE1A/ADP or M4 at a MOI of 100 orwith wt-Ad5 at a MOI of 0.1. Images were taken at the time when wt-Ad5 caused complete CPE (12 d p.i.). (C) Progeny virus production from recombinant viruses Ad5/dE1A,Ad5/dE1A/ADP and M4. A549 cells were infected at an MOI of 20, and cells and medium were collected separately at 24 h intervals and assayed by TCID50 forinfectious progeny virus.
Z.Han et al.
2018
Dow
nloaded from https://academ
ic.oup.com/carcin/article/30/12/2014/2392052 by guest on 02 February 2022

xenograft models were first employed to test the therapeutic efficacyof i.t. injection of M4. Compared with control (Ad5/dE1A- or Ad5/dE1A/ADP-treated) tumors, those in mice that received M4 dis-played strong growth inhibition (Figure 6A). Control animals devel-oped symptoms like anorexia and weight loss before they were killed,whereas M4-treated animals remained symptom free. Confirming thein vitro findings, western blotting of tumor lysates also revealeda significant reduction in STAT3 protein levels and STAT3 down-stream target genes even 30 days after i.t. injection, and theeffect of downregulation was still obvious (Figure 6B and C). In
contrast, STAT3 siRNA produced a transitory downregulation ofSTAT3 protein by direct i.t. injection (Figure 6C). In the orthotopicxenograft model, primary tumor volume, local invasion and abdom-inal cavity metastasis were all significantly decreased in the animalstreated with M4 compared with Ad5/dE1A or Ad5/dE1A/ADP(Table 1), and M4 treatment significantly extended the survival time-of tumor-bearing mice (P , 0.01, Figure 6D). At 30 day after thelast systemic administration of M4, efficient viral replication wasfound in the viable tumor tissue and a significant number of necrotictumor cells was seen in adjacent tissue (Figure 6E). Depletion of
Fig. 5. Effects of M4 on tumor cell viability, invasion and angiogenesis in vitro. (A) Cancer cells (MDA-MB-231, HCT-8, PC3M-1E8 and MKN-45) wereinfected with the indicated viruses at an MOI of 10, and the rate of apoptosis was measured by fluorescein isothiocyanate–annexin Vand propidium iodide stainingat 48 h, 72 h and 96 h p.i. Western blotting was conducted on cell lysates from tumor cells with antibodies to STAT3 72 h p.i. (B) Semiquantitative RT-PCR analysisof E1B-19K in A549 cells infected with Ad5/dE1A, Ad5/dE1A/ADP or M4 at an MOI of 10 at the indicated times p.i. (C) Quantification of invasion asa percentage of the mock-transfected control. Bars represent the mean ± SEM. (D) The effects of conditioned medium from PC3M-1E8 or MKN-45 cells infectedwith the indicated viruses at an MOI of 10 on angiogenesis in vitro.
Oncolytic adenovirus silences the expression of STAT3
2019
Dow
nloaded from https://academ
ic.oup.com/carcin/article/30/12/2014/2392052 by guest on 02 February 2022

tumor-associated STAT3 protein was seen in mice treated by intra-venous injection of M4 but not Ad5/dE1A, Ad5/dE1A/ADP orphosphate-buffered saline (Figure 6F). To assess the adverse effectsof the treatment, the levels of alanine aminotransferase and aspartameaminotransferase in serum of mice were assayed. Compared with the
mice injected with phosphate-buffered saline, mice injected with M4also showed a slight increase in the levels of aspartame aminotrans-ferase and alanine aminotransferase; however, the increase was muchlower than those caused by wt-Ad5 (supplementary Figure S2 isavailable at Carcinogenesis Online).
Fig. 6. Relative antitumor efficacy after treatment with Ad5/dE1A, Ad5/dE1A/ADP or M4 in athymic BALB/c nude mice. (A) In vivo efficacy of M4 onsubcutaneous established PC3M-1E8 and MKN-45 tumors. (B) Western blot analysis of STAT3, vascular endothelial growth factor (VEGF) and MMP-9 in bothxenograft tumors treated with phosphate-buffered saline (PBS), Ad5/dE1A, Ad5/dE1A/ADP or M4 30 d after i.t. injection. (C) Western blot analysis of STAT3 andVEGF in both xenograft tumors treated with PBS, non-targeting control siRNA, STAT3 siRNA or M4 3 or 15 d after i.t. injection. (D) Kaplan–Meier survivalcurves following administration of PBS, Ad5/dE1A, Ad5/dE1A/ADP or M4. Each group included 10 animals. (E) Viral replication was assayed byimmunohistochemistry and in situ hybridization. Immunohistochemistry displays viral particles (short arrows) stained yellow in cancer cells (upper panel), andin situ hybridization shows cells containing viral particles (short arrows) stained dark blue (lower panel) 30 d after the last systemic administration of M4.(F) Western blot analysis of STAT3 in orthotopic tumors at day 30 after the last intravenous injection of viruses or PBS.
Table I. Effects of M4 on tumor growth and metastases in an orthotopic model of human gastric carcinoma 6 weeks after treatment
Parameter PBS Ad5/dE1A Ad5/dE1A/ADP M4
Primary tumor volume (cm3) 1.860 ± 0.247 1.608 ± 0.203 1.855 ± 0.164 0.544 ± 0.180a
Local invasive growth in the liver 5/5b 5/5 5/5 2/5Peritoneal dissemination 5/5 4/5 5/5 1/5
Six weeks after treatment, the volumes of the gastric tumors and the extent of metastatic involvement were determined. Data are reported as mean ± SEM.aP ,0.01 compared with phosphate-buffered saline (PBS), Ad5/dE1A or Ad5/dE1A/ADP.bData are shown as the number of mice in which local invasive growth or peritoneal metastases were observed/number of mice evaluable.
Z.Han et al.
2020
Dow
nloaded from https://academ
ic.oup.com/carcin/article/30/12/2014/2392052 by guest on 02 February 2022

Discussion
Although numerous lines of evidence have validated STAT3 asa promising target for cancer therapy, the development of agents thatinhibit STAT3 with sufficient potency and good tumor selectivity hasproven to be a formidable task. For example, one of most promisingapproaches to targeting STAT3 is to inhibit phosphorylation of STAT3using small molecule compounds. However, some studies have dem-onstrated that unphosphorylated STAT3 is also important for onco-genesis (34,35). The main contribution of the current study is thedevelopment of M4 for selective depletion of STAT3 protein in tumorcells but not in normal cells. By combining the oncolytic properties ofAd5 with the benefits of targeting the STAT3 protein by antisenseRNA, M4 demonstrated potent antitumoral efficacy and even moreattractively great potential to control metastases.
Three bioengineered features are responsible for the potency andselectivity of M4. First, M4 contains a 27 bp deletion in the E1A CR2necessary for pRb binding. This modification enables M4 to replicateefficiently in and lyse cells with a defective G1–S checkpoint, which isseen in almost all tumor cells as a result of a loss of function of in theRb pathway. Second, the current design employed the ADP region forinsertion of the therapeutic transgene, which lies directly downstreamof the gp19K gene in the E3 transcription unit and has the samepolyadenylation site as the gp19K gene. Unlike gp19K, which beginsto be greatly amplified in early stage of viral infection, expression ofADP is driven mainly by the adenovirus major late promoter and onlyoperates at a high level during the late phase of the viral infection ina replication-dependent manner (27). Our data demonstrate that M4generated by this strategy allowed for a latent cytolysis, favoringhigher viral production without undermining the final oncolytic po-tency. Third, placing reverse STAT3 cDNA into the deleted ADP locusin M4 enabled the highly selective expression of transcripts of anti-sense STAT3 in tumor but not normal cells and subsequently depletionof tumor cell STAT3 levels.
It is true that the oncolytic adenovirus-infected tumor cell shoulddie from oncolysis anyway. However, complete oncolysis only occursin in vitro cell culture models. On the other hand, many obstaclesin vivo exist to limit the abilities of oncolytic adenoviruses, whichis clearly exemplified by clinical trials with oncolytic adenovirus (36).Therefore, many investigators (including our group) have successfullydeveloped a variety of oncolytic adenoviral mutants by incorporatingdifferent transgenes, such as antisense plk1 or chk1 cDNA (25,26).Because STAT3 is a direct transcriptional activator of a large numberof downstream genes, targeting STAT3 appears to have more theoret-ically advantageous and is unique. In addition to the high frequency ofconstitutive activation of STAT3 in tumors, STAT3 activation hasadditive effects for maintaining multiple malignant behaviors suchas proliferation, angiogenesis and immune evasion (6). Oncolyticadenoviral mutants armed with antisense plk1 or chk1 cDNA are un-likely to produce bystander effects, which could limit the potency oftumor eradication. On the other hand, silencing of STAT3 has beenreported to elicit potent bystander effects, which might be caused bymechanisms such as reactivation of antitumor immunity, inhibition ofangiogenesis and reduction of the release of soluble cytokines (6).Most recently, compelling evidence has shown that inhibiting STAT3signaling elicits multicomponent antitumor immunity and reversesimmune tolerance (10). Our study showed that when armed withantisense STAT3 cDNA, M4 significantly enhanced its ability to in-duce apoptosis and inhibit invasive potential and angiogenesis ofcancer cells compared with Ad5/dE1A or Ad5/dE1A/ADP. One ofthe most interesting findings was that the human metastatic gastrictumor mouse model responded to systemic administration of M4 witha substantially reduced incidence of metastases. The unexpected elim-ination of metastatic progression by M4 is of great significance and isan advance from the failure of M1 as a single agent to reduce thefrequency of metastatic progression in our previous publication (26).The anti-metastatic potential of M4 could in part be due to the in-hibition of angiogenesis and loss of invasiveness, which are controlledby STAT3.
Concerns exist about the clinical potential of oncolytic adenoviralmutants, including M4. Up to now, the major route of administrationof adenovirus therapy in clinical use has been direct i.t. injection. Forthis reason, adenovirus gene therapy is now generally regarded as anadjuvant approach for local tumor control, which severely limits thepotential of oncolytic adenoviruses in vivo. However, we recentlyshowed that systemic administration of oncolytic adenovirus M1 al-lows it to effectively spread and replicate in local and distant metas-tases (30). The current study provides further evidence that systemicadministration of oncolytic adenovirus M4 could prevent the meta-static progression of primary tumors by selectively silencing tumor-associated STAT3. Whether the present findings can be translated intoclinical benefits remains to be determined in future clinical trials.Because the STAT3 protein is difficult to target, the current approachis an attractive potential treatment strategy. The findings presentedhere are encouraging enough to warrant further laboratory and clinicalevaluation.
Supplementary material
Supplementary data and Figures S1 and S2 can be found at http://carcin.oxfordjournals.org/
Funding
National High Technology Research and Development Program ofChina 2007AA021001; Major State Basic Research DevelopmentProgram of China 2009CB521800; National Natural Science Foun-dation of China 30672227, 30700895 and 30770914.
Acknowledgements
Conflict of Interest Statement: None declared.
References
1.Darnell,J.E.Jr (1997) STATs and gene regulation. Science, 277, 1630–1635.2.Bromberg,J. et al. (2000) The role of STATs in transcriptional control and
their impact on cellular function. Oncogene, 19, 2468–2473.3.Bromberg,J. (2000) Signal transducers and activators of transcription as
regulators of growth, apoptosis and breast development. Breast CancerRes., 2, 86–90.
4.Smithgall,T.E. et al. (2000) Control of myeloid differentiation and survivalby Stats. Oncogene, 19, 2612–2618.
5.Rawlings,J.S. et al. (2004) The JAK/STAT signaling pathway. J. Cell Sci.,117, 1281–1283.
6.Yu,H. et al. (2004) The STATs of cancer—new molecular targets come ofage. Nat. Rev. Cancer, 4, 97–105.
7.Groner,B. et al. (2008) The function of Stat3 in tumor cells and theirmicroenvironment. Semin. Cell Dev. Biol., 19, 341–350.
8.Turkson,J. et al. (2000) STAT proteins: novel molecular targets for cancerdrug discovery. Oncogene, 19, 6613–6626.
9.Buettner,R. et al. (2002) Activated STAT signaling in human tumors pro-vides novel molecular targets for therapeutic intervention. Clin. CancerRes., 8, 945–954.
10.Wang,T. et al. (2004) Regulation of the innate and adaptive immune re-sponses by Stat-3 signaling in tumor cells. Nat. Med., 10, 48–54.
11.Al Zaid Siddiquee,K. et al. (2008) STAT3 as a target for inducing apoptosisin solid and hematological tumors. Cell Res., 18, 254–267.
12.Chen,Z. et al. (2008) STAT3: a critical transcription activator in angiogen-esis. Med. Res. Rev., 28, 185–200.
13.Grandis,J.R. et al. (2000) Constitutive activation of Stat3 signaling abro-gates apoptosis in squamous cell carcinogenesis in vivo. Proc. Natl Acad.Sci. USA, 97, 4227–4232.
14.Niu,G. et al. (2002) Constitutive Stat3 activity up-regulates VEGF expres-sion and tumor angiogenesis. Oncogene, 21, 2000–2008.
15.Darnell,J. (2005) Validating Stat3 in cancer therapy. Nat. Med., 11, 595–596.16.Costantino,L. et al. (2008) STAT3 as a target for cancer drug discovery.
Curr. Med. Chem., 15, 834–843.17.Schust,J. et al. (2006) Stattic: a small-molecule inhibitor of STAT3 activa-
tion and dimerization. Chem. Biol., 13, 1235–1242.18. Iwamaru,A. et al. (2007) A novel inhibitor of the STAT3 pathway induces
apoptosis in malignant glioma cells both in vitro and in vivo. Oncogene, 26,2435–2444.
Oncolytic adenovirus silences the expression of STAT3
2021
Dow
nloaded from https://academ
ic.oup.com/carcin/article/30/12/2014/2392052 by guest on 02 February 2022

19.Xu,X. et al. (2009) Chemical probes that competitively and selectivelyinhibit STAT3 activation. PLoS One, 4, e4783.
20.Hermiston,T. (2000) Gene delivery from replication-selective viruses: arm-ing guided missiles in the war against cancer. J. Clin. Invest., 105,1169–1172.
21.Mathis,J.M. et al. (2005) Oncolytic adenoviruses—selective retargeting totumor cells. Oncogene, 24, 7775–7791.
22.Rein,D.T. et al. (2006) Current developments in adenovirus-based cancergene therapy. Future. Oncol., 2, 137–143.
23.Kanerva,A. et al. (2005) Adenoviruses for treatment of cancer. Ann. Med.,37, 33–43.
24.Kay,M.A. et al. (2001) Viral vectors for gene therapy: the art of turninginfectious agents into vehicles of therapeutics. Nat. Med., 7, 33–40.
25.Zhou,J. et al. (2005) Novel oncolytic adenovirus selectively targets tumor-associated polo-like kinase 1 and tumor cell viability. Clin. Cancer Res., 11,8431–8440.
26.Gao,Q. et al. (2006) Selective targeting of checkpoint kinase 1 intumor cells with a novel potent oncolytic adenovirus. Mol. Ther., 13,928–937.
27.Hawkins,L.K. et al. (2001) Gene delivery from the E3 region of replicatinghuman adenovirus: evaluation of the ADP region. Gene Ther., 8, 1132–1141.
28.McGettrick,H.M. et al. (2006) Chemokine- and adhesion-dependentsurvival of neutrophils after transmigration through cytokine-stimulatedendothelium. J. Leukoc. Biol., 79, 779–788.
29.Zhou,W. et al. (2006) STAT3 is required but not sufficient for EGFreceptor-mediated migration and invasion of human prostate carcinomacell lines. Br. J. Cancer, 95, 164–171.
30.Huang,X. et al. (2008) Biodistribution and kinetics of the novel selectiveoncolytic adenovirus M1 after systemic administration. Mol. Cancer Ther.,7, 1624–1632.
31.Wold,W.S. et al. (1995) E3 transcription unit of adenovirus. Curr. Top.Microbiol. Immunol., 199, 237–274.
32.Horwitz,M. (2004) Function of adenovirus E3 proteins and their interac-tions with immunoregulatory cell proteins. J. Gene Med., 6, 172–183.
33.Teodoro,J. et al. (1997) Regulation of apoptosis by viral gene products.J. Virol., 71, 1739–1746.
34.Yang,J. et al. (2005) Novel roles of unphosphorylated STAT3 in oncogen-esis and transcriptional regulation. Cancer Res., 65, 939–947.
35.Lassmann,S. et al. (2007) STAT3 mRNA and protein expression in colo-rectal cancer: effects on STAT3-inducible targets linked to cell survival andproliferation. J. Clin. Pathol., 60, 173–179.
36.Khuri,F.R. et al. (2000) A controlled trial of intratumoral ONYX-015,a selectively-replicating adenovirus, in combination with cisplatin and 5-fluorouracil in patients with recurrent head and neck cancer. Nat. Med., 6,879–885.
Received June 10, 2009; revised September 24, 2009;accepted October 10, 2009
Z.Han et al.
2022
Dow
nloaded from https://academ
ic.oup.com/carcin/article/30/12/2014/2392052 by guest on 02 February 2022
Related Documents











