Eur. J. Biochem. 235, 103-113 (1996) 0 FEBS 1996 A novel dystrophidutrophin-associated protein is an enzymatically inactive member of the phosphoglucomutase superfamily Elena P. MOlSEEVA', Alexey M. BELKIN', Nigel K. SPURR?, Victor E. KOTELIANSKY4 and David R. CRITCHLEY' I Department of Biochemistry, University of Leicester, UK ' Imperial Cancer Research Fund, Clare Hall Laboratories, South Mimms, Potters Bar, UK Department of Cell Biology and Anatomy, University of North Carolina at Chapel Hill, North Carolina, USA Laboratoire de Physiopathologie du Developpement, CNRS et Ecole Normal Superieure, Paris, France (Received 4 September/3 Novcmber 1995) - EJB 95 1453/1 A 60-kDa protein localised in adherens-type cellular junctions, and previously called aciculin, has been found to interact with the cytoskeletal proteins dystrophin and utrophin IBelkin, A. M. & Burridge, K. (1995) .I. Biol. Chern. 270, 6328-63371. In this study, we report the complete sequence of this protein, and show that it is a novel member of the phosphoglucomutase (PGM) family of proteins. The PGM- related protein (PGM-RP), which contains 506 amino acids (55.6 kDa), is smaller than PGMl (566 amino acids, 61 kDa). The active site consensus sequences of prokaryotic and eukaryotic mutases are not con- served in PGM-RP, a finding consistent with the lack of enzymatic activity of PGM-RP in vitvo, and the absence of a phosphorylated intermediate in vivo. The organisation of the PGM-RP gene is essentially identical to that of PGMl. We propose that the PGM-RP gene, which we have mapped to human chromo- some 9qcen-ql3, evolved from the PGMl gene, and encodes a protein with a structural rather than an enzymatic role. PGM-RP is expressed predominantly in muscle with the highest levels in smooth muscle. The significance of the interaction between dystrophinhtrophin and an increasing number of cytoplasmic proteins including PGM-RP remains to be explored. Keywords: phosphoglucomutase ; adherens-type cellular junctions ; dystrophin ; utrophin. The gene mutated in the X-linked Duchenne and Becker- type muscular dystrophies encodes a 427-kDa protein (dystrophin) that is homologous to the a-actinin and speclrin family of cytoskeletal proteins 11, 21. Dystrophin is an elongated inolecule 13 I which contains an N-terminal actin-binding domain 141, a central rod-like domain containing 25 spectrin-like repeats, a cysteine-rich domain containing two incomplete EF-hand calcium-binding motifs, and a C-terminal region containing a possible leucine zipper [5]. It is localised on the cytoplasmic face of the sarcolemma where it may serve to protect the sarco- lemma from the mechanical stresses developed during muscle contraction 16, 71. Some information on the nature of the link between dystrophin and the membrane has recently been ob- tained, although the observation that the DMD gene gives rise to multiple dystrophin isoforins suggests that current models will require modification [S]. The C-terminal region of dystrophin is thought to contain binding sites for a complex of five trans- membrane proteins (adhalin, P-dystroglycan, 43-kDa DAG (dystrophin-associated glycoprotein), 35-kDa DAG and 25-kDa DAP (dystrophin-associated protein) four of which are in turn Correspondence to D. R. Critchley, Department of Biochemistry, University of Leicester, University Rd, Leicester LEI 7RH, UK Fax: +44 116 2523369. Ahhreviutions. PGM, phosphoglucomutase; PGM-RP, phosphoglu- coniulase-relatcd protein; DAG, dystropbin-associated glycoprotein; DAP, dystrophin-associated protein. Ennzynze. Phosphoglucomutase (EC 5.4.2.2). Note. The PGM-RP cDNA sequence data have been deposited with the GSDB/EMBL/NCBI sequence data banks and are available under acccssion number L40933. The sequences of exoiis 2, 3,S, 8 and 9 of the muse PGM-KP gene are available under accession numbers L42902- L42905 and L42907, respectively. linked to a large extracellular glycoprotein a-dystroglycan 12, 81. This latter protein binds to the laminin isoform merosin, provid- ing a link between the extracellular matrix and the muscle cy- toskeleton. At least three additional cytoplasmic proteins (al-, [jl-, and p2-syntrophins) have been identified that bind to the C- terminal region of dystrophin, although their function is un- known [2, 8, 91. al-Syntrophin and PI-syntrophin have been shown to bind to the alternatively spliced dystrophin exons 73 and 74, respectively 19, 101. The dystrophin-related protein, utrophin 121, has also been shown to interact with some of the same dystrophin-associated proteins [l I]. Utrophin is expressed in embryonic and regenerating muscle where it is localised to the sarcolemma, but it is replaced by dystrophin in adult skeletal muscle 121. However, it is expressed in many different cell types with high levels of expression in smooth muscle. Down-regula- tion of utrophin in cultured astroctyes using antisense oligonu- cleotides, leads to a marked reduction in cell-matrix contacts which implies that the utrophin cytoskeletal complex is also in- volved in cell adhesion [12]. In an attempt to identify additional muscle proteins localised at the membranekytoskeletal interface, we raised a panel of mo- noclonal antibodies to a human uterine smooth muscle cytoskel- eta1 extract. We identified a novel antigen (60-63 kDa) which was localised in a variety of adherens-type cellular junctions [I 3 1. This protein, initially called aciculin 11 41, has recently been shown to coimmunoprecipitate with dystrophin and utrophin [15]. In this study, we report the nucleotide and deduced amino acid sequence of this protein which turns out to be a novel men- ber of the phosphoglucomutase (PGM) family of proteins. Since the protein lacks phosphoglucomutase activity, we refer to the protein as the phosphoglucomutase-related protein or PGM-RP.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Eur. J. Biochem. 235, 103-113 (1996) 0 FEBS 1996
A novel dystrophidutrophin-associated protein is an enzymatically inactive member of the phosphoglucomutase superfamily Elena P. MOlSEEVA', Alexey M. BELKIN', Nigel K. SPURR?, Victor E. KOTELIANSKY4 and David R. CRITCHLEY'
I Department of Biochemistry, University of Leicester, UK
' Imperial Cancer Research Fund, Clare Hall Laboratories, South Mimms, Potters Bar, UK Department o f Cell Biology and Anatomy, University of North Carolina at Chapel Hill, North Carolina, USA
Laboratoire de Physiopathologie du Developpement, CNRS et Ecole Normal Superieure, Paris, France
(Received 4 September/3 Novcmber 1995) - EJB 95 1453/1
A 60-kDa protein localised in adherens-type cellular junctions, and previously called aciculin, has been found to interact with the cytoskeletal proteins dystrophin and utrophin IBelkin, A. M. & Burridge, K. (1995) .I. Biol. Chern. 270, 6328-63371. In this study, we report the complete sequence of this protein, and show that it is a novel member of the phosphoglucomutase (PGM) family of proteins. The PGM- related protein (PGM-RP), which contains 506 amino acids (55.6 kDa), is smaller than PGMl (566 amino acids, 61 kDa). The active site consensus sequences of prokaryotic and eukaryotic mutases are not con- served in PGM-RP, a finding consistent with the lack of enzymatic activity of PGM-RP in vitvo, and the absence of a phosphorylated intermediate in vivo. The organisation of the PGM-RP gene is essentially identical to that of PGMl. We propose that the PGM-RP gene, which we have mapped to human chromo- some 9qcen-ql3, evolved from the PGMl gene, and encodes a protein with a structural rather than an enzymatic role. PGM-RP is expressed predominantly in muscle with the highest levels in smooth muscle. The significance of the interaction between dystrophinhtrophin and an increasing number of cytoplasmic proteins including PGM-RP remains to be explored.
Keywords: phosphoglucomutase ; adherens-type cellular junctions ; dystrophin ; utrophin.
The gene mutated in the X-linked Duchenne and Becker- type muscular dystrophies encodes a 427-kDa protein (dystrophin) that is homologous to the a-actinin and speclrin family of cytoskeletal proteins 11, 21. Dystrophin is an elongated inolecule 13 I which contains an N-terminal actin-binding domain 141, a central rod-like domain containing 25 spectrin-like repeats, a cysteine-rich domain containing two incomplete EF-hand calcium-binding motifs, and a C-terminal region containing a possible leucine zipper [5] . It is localised on the cytoplasmic face of the sarcolemma where it may serve to protect the sarco- lemma from the mechanical stresses developed during muscle contraction 16, 71. Some information on the nature of the link between dystrophin and the membrane has recently been ob- tained, although the observation that the D M D gene gives rise to multiple dystrophin isoforins suggests that current models will require modification [S]. The C-terminal region of dystrophin is thought to contain binding sites for a complex of five trans- membrane proteins (adhalin, P-dystroglycan, 43-kDa DAG (dystrophin-associated glycoprotein), 35-kDa DAG and 25-kDa DAP (dystrophin-associated protein) four of which are in turn
Correspondence to D. R. Critchley, Department of Biochemistry, University of Leicester, University Rd, Leicester LEI 7RH, UK
Fax: +44 116 2523369. Ahhreviutions. PGM, phosphoglucomutase; PGM-RP, phosphoglu-
coniulase-relatcd protein; DAG, dystropbin-associated glycoprotein; DAP, dystrophin-associated protein.
Ennzynze. Phosphoglucomutase (EC 5.4.2.2). Note. The PGM-RP cDNA sequence data have been deposited with
the GSDB/EMBL/NCBI sequence data banks and are available under acccssion number L40933. The sequences of exoiis 2, 3,S, 8 and 9 of the m u s e PGM-KP gene are available under accession numbers L42902- L42905 and L42907, respectively.
linked to a large extracellular glycoprotein a-dystroglycan 12, 81. This latter protein binds to the laminin isoform merosin, provid- ing a link between the extracellular matrix and the muscle cy- toskeleton. At least three additional cytoplasmic proteins (a l - , [jl-, and p2-syntrophins) have been identified that bind to the C- terminal region of dystrophin, although their function is un- known [2, 8, 91. al-Syntrophin and PI-syntrophin have been shown to bind to the alternatively spliced dystrophin exons 73 and 74, respectively 19, 101. The dystrophin-related protein, utrophin 121, has also been shown to interact with some of the same dystrophin-associated proteins [l I]. Utrophin is expressed in embryonic and regenerating muscle where it is localised to the sarcolemma, but it is replaced by dystrophin in adult skeletal muscle 121. However, it is expressed in many different cell types with high levels of expression in smooth muscle. Down-regula- tion of utrophin in cultured astroctyes using antisense oligonu- cleotides, leads to a marked reduction in cell-matrix contacts which implies that the utrophin cytoskeletal complex is also in- volved in cell adhesion [12].
In an attempt to identify additional muscle proteins localised at the membranekytoskeletal interface, we raised a panel of mo- noclonal antibodies to a human uterine smooth muscle cytoskel- eta1 extract. We identified a novel antigen (60-63 kDa) which was localised in a variety of adherens-type cellular junctions [I 3 1. This protein, initially called aciculin 11 41, has recently been shown to coimmunoprecipitate with dystrophin and utrophin [15]. In this study, we report the nucleotide and deduced amino acid sequence of this protein which turns out to be a novel men- ber of the phosphoglucomutase (PGM) family of proteins. Since the protein lacks phosphoglucomutase activity, we refer to the protein as the phosphoglucomutase-related protein or PGM-RP.

104 Moiseeva et al. (Eur: J . Hiochem. 235)
MATERIALS AND METHODS
Construction of a human uterus cDNA library and im- munoscreening for PGM-RP. Total RNA was purified from human uterus using the guanidinium thiocyanate phenokhloro- form extraction method [I 61. Poly(A)-rich RNA isolated using the Fast Track mRNA isolation kit (Tnvitrogen) was used to make a cDNA library in the LZAPII vector with the Uni-Zap XR/Gigapack 11 Gold Cloning kit (Stratagene). Approximately 1.9X10" independent clones were obtained and amplified (average insert size of 1.3 kb), and 5x10" clones were immu- noscreened with a rabbit polyclonal antibody raised against PGM-RP and anti-rabbit IgG conjugated to alkaline phosphatase (Promcga). Ten positive clones were obtained two of which (A111 and P87) reacted strongly with the antibody. Single plaques of these recombinants were converted to the Bluescript SK(-) plasmid by superinfection and excision with helper phage. Double-stranded plasmid templates were sequenced using the Circumvent Thermal Cycle Dideoxy DNA Sequencing kit (New England Biolabs). The strategies used to determine the complete sequence of the full-length cDNA (A1 11) encoding PGM-RP included subcloning and sequcncing rcstriction enzyme frag- ments, and the use of oligonucleotide sequencing primers de- signed according to the determined cDNA sequence. Sequences were analysed using the University of Wisconsin Genetics Com- puter Group software package, and proteins in data bases show- ing homology to PGM-RP were identified using the FASTA pro- gram.
Analysis of the PGM-RP mRNA. The approximate size of thc PGM-Ii'P mRNA was determined by Northern-blot analysis following standard procedures [ I 61. Total RNA isolated from human uterus smooth muscle and rabbit skelelal muscle was re- solved in formaldehyde gels and the RNA blotted to Hybond-N filters (Amersham). Hybridisation was performed using a PGM- RP probe spanning nucleotidcs 90-1609 labellcd with .'2P by nick-tr~inslation (Pharmacia). Filters were incubated in Ex- pressHyb hybridization solution (Clontech) at 68°C for 1 11 ac- cording to the manufacturer's protocol. The blot was stripped and hybridiscd with a human PCMI cDNA probe equivalent to nucleotides 303 - 1754 of the complete sequence [ 171.
Metabolic labelling of C2C12 myoblasts and immune precipitation of labelled proteins. C2C12 mouse myoblasts 1181 were grown in Dulbecco's modified Eagle's medium con- taining 10% newborn calf scrum. Cells (9-cm-diameter dishes) were pulse-labelled (4 h) with either 0.1 mCi/ml [ '*Plorthophos- phate i n phosphate-free medium or with 0.2 mCi/ml ["Slinethio- nine/cysteine in methionine-free and cysteine-frce medium. Cells were lysed in 1 in1 RTPA buffer (50 mM Tris, pH 7.5, 150 mM NaCI, 1 mM EDTA, 1 % Triton X-100, 0.5 % sodium deoxycholatc, 0.1 % SDS) containing the protease inhibitors 1 niM phenylrnethylsulfonyl fluoride, 1 mM benzamidine, 10 yg/ml leupeptin and 10 pg/ml aprotinin. Lysates were incu- bated with either 10 yg monoclonal antibody XIVFR (to specifi- cally precipitate PGM-RP) or 10 pg affinity-purified rabbit poly- clonal antibody to PGM-RP which also cross-reacts with PGMl [ 1.71. Iminune complexes were precipitated with protein-A-
Sepharose (Sigma) and labelled proteins were resolved by SDS/ PAGE (1 0% polyacrylamide gels) and detected by autoradiog- raphy.
Chromosome mapping. The gene encoding PGM-RP was mapped by PCR using DNA from a panel of human cell hybrids that have been described previously [19-221. PCR was per- formed with 0.1 yg DNA, 5 U Taq DNA polymerase (Boeh- ringer Mannheini) in 50 yI (if the buffer supplied.
The 5' primer A1 3 (5'-GCCCAAAGAGAGTGCTCA- GCGC-3') and the 3' primer A14 (5'-ACACCTATTGATATA- TTTTGTTGGAAGT-3') were designed according to the 3'-un- translated sequence, and were used in touchdown PCR as fol- lows. Eight cycles were performed under the following condi- tions: 1 min at 94"C, 45 s at the annealing temperature (the annealing temperature was decreased from 72°C to 66°C in steps of2"C every 2 cycles), 45 s at 72OC. 36 cycles were subse- quently carried out: 1 min at 94"C, 45 s at 66"C, 45 s at 72°C. Aliquots of the reaction were resolved i n 1 %J agarose gels and the product derived from the PGM-RP gene was detected by Southern blotting using a 5'-end-labelled oligonucleotide A4 (5'- GTTCTGATTAACAGCTGCCT-3') according to standard pro- cedures [16]. Hybridisation was performed at 51 "C using 1 X NaCI/Cit (0.15 M NaC1, 15 mM sodium citrate, pH 7). Alterna- tively, products of the first PCR reaction were diluted twenty times and reamplified in the second round of PCR with oligonu- cleotides A5 (5'-TTGCTACCTGTTTGTGCCTC) and A1 4. 30 cycles were carried out as follows: 1 min at 94"C, 1 min at 57"C, 45 s at 72°C.
Isolation and analysis of PGM-RP genomic clones. A hu- man PGM-RP cDNA probe (nucleotides 1-1609) was used to isolate mouse genornic clones from an ES cell genomic library in the vector L2001 (a generous gift from Dr Andrew J. H. Smith, MRC Laboratory for Molecular Biology, Cambridge). Restriction enzyme fragments containing coding sequence were identified by Southern-blot analysis and subcloned into the Bluescript plasmid. Exon boundaries were determincd by se- quencing using primers based on the human PGM-RP cDNA sequence. The position and orientation of exons on DNA frag- ments was verified by PCR. The distances betwecn exons on different DNA fragments contained within the same phage were determined by long distance PCR using phage DNA as templatc P31.
Detection and quantitation of PGM-RP in different tis- sues and cell lines. Expression of PGM-RP in different tissucs was detected by immunofluorescence on cryostat sections as de- scribed previously 1131. The amount of the PGM-RP in tissues or a variety of types of cultured cells (150 pg/sample) was deter- mined by a modified Western-blot protocol in which bound mo- noclonal antibody XIVFR was detected with an iodinated sec- ondary antibody (1 yg/ml containing 1 O7 cpm/ml). Radiolabelled bands detected by autoradiography were cut out, and the amount of radioactivity present was determined by scintillation count- ing. At least three independent samples were analysed for each tissue or cell type. The amount of radiolabelled antibody hound for each sample varied from the mean by < 10%.
- Fig. 1. Nucleotide sequence and deduced amino acid sequence of human PGM-RI? (A) Schematic diagram showing the relationship between thc Iwo PGM-RP cDNAs A1 I1 and ASS. The coding sequence (R) and tmtranxlatcd sequence (0) are indicated. The region of PGM-RP rccognised by monoclonal antibody XIVFS is mnrlted (t). The relative positions of several adclitional PGM-RP cDNAs isolated fmm the human smooth muscle library by screening with the A1 11 cDNA are also shown. These cDNAs are derived from unproccsscd R N A and contain both coding (R) and iiitron (diagonal line) sequences, and therefore provide information on the exoii boundaries of the human P C M - R P gene. The coordinates of thc nuclcotidc coding sequence are iis follows: A51, 169-334; AS2, 323-524; A46, 346-796; A99, 1028- 1387. (B) The complete nucleotidc and amino acid sequence of PGM-RP. The coordinates are indicated at the right. Amino acid residues (single lcttcr code) verified by peptide sequencing arc tmdcrlined. The nucleotide sequence wiis compiled from the two overlapping cDNAs A l l 1 and ASS.

Moiseeva el al. (Eur J. Biochem. 235)
A
105
B
cggcctcttcgagggccagcgcaactacctgcccaactttatccagagcgtgctgtcgtccatcgacctgcgcgaccgtcagggctgcac
C A T G G T G G T G G G C A G C G A C G G C A G G T A C T T T A G C A G G A C G G A C T M V V G S D G R Y F S R T A I E I V V Q M A A A N G I G R T d
GATTATTGGACAGAATGGCATCTTGTCGACACCTGCGGTCTCCTGCATTATCAGGAAGATCAAGGCAGCTGGTGGAATCATTCTAACAGC I I G O N G I L S T P A V S C I I R K I K A A G G I I L T A
CAGCCACTGCCCTGGAGGACCAGGGGGAGAGTTTGGAGTG~GTTT~TGTTGCCAATGGAGGTCCTGCACCCGATGTTGTCTCAGAC~ S H C P G G P G G E F G V K F N V A N G G P A P D V V S D K
AATCTACCAAATCAGCAAAACGATTGAGGAATATGCTATATGTCCTGATCTCCGAATCGACCTATCTCGACTAGGAAGACAAGAATTTGA I Y Q I S K T I E E Y A I C P D L R I D L S R L G R Q E F D
CCTAGAAAACAAATTCAAACCATTCAGAGTGGAGATAGTGGACCCAGTGGATATCTATCTTAACCTCCTTCGGACCATCTTTGACTTTCA L E N K F K P F R V E I V D P V D I Y L N L L R T I F D F H
T G C C A T C A A G G G T T T G C T G A C T G G A C C C A G C C A A C T G A A G G T A I K G L L T G P S Q L K I R I D A M H G V M G P Y V R K V
TCTGTGTGATGAGCTGGGGGCCCCAGCCAATTCTGCAATAAACTGTGTTCCCCTGGAAGACTTTGGAGGGCAGCACCCTGACCCAAACCT L C D E L G A P A N S A T N C V P 1 , E D F G G Q H P D P N L
GACATATGCAACGACTCTTCTGGAAGCAATGAAAGGAGGAGAATATGGATTTGGAGCTGCATTTGATGCTGATGGGGACCGTTATATGAT T Y A T T L L E A M K G G E Y G F G A A F D A D G D R Y M I
CCTAGGCCAAAATGGCTTCTTTGTGAGCCCTTCTGACTCCCTGGCCATCATTGCTGCC~CCTCTCTTGCATTCCATATTTCCGTCAGAT L G Q N G F F V S P S D S L A I I A A N L S C I P Y F R Q M
GGGGGTCCGCGGGTTTGGGAGGAGTATGCCAACCAGCATGGCCCTGGACAGAGTGGCCAAATC~TGAAGGTCCCTGTATATGAGACCCC G V R G F G R S M P T S M A L D R V A K S M K V P V Y E T P
AGCTGGATGGAGATTCTTCTCAAATCTGATGGACTCAGGACGTTGCAATCTGTGTGGGG~GAGAGCTTTGGCACTGGCTCTGACCACCT A G W R F F S N L M D S G R C N L C G E E S F G T G S D H L
CCGAGAGAAGGATGGCCTGTGGGCTGTCTTGGTCTGGCTCTCCATTATTGCTGCCCGGAAGCAGAGTGTGGAGG~TTGTCCGAGATCA R E K D G L W A V L V W L S I I A A R K Q S V E E I V R D H
CTGGGCCAAATTTGGCCGCCACTACTATTGCAGGTTTGACTATGAGGGGTTGGATCCCAAGACGACATATTATATCATGAGGGACCTGGA W A K F G R H Y Y C R F D Y E G L D P K T T Y Y I M R D L E
GGCCCTGGTCACAGACAAATCCTTCATTGGCCAGCAGTTTGCTGTGGGGAGCCATGTCTACAGCGTGGCGAAGACGGATAGTTTTG~TA A L V T D K S F I G O O F A V G S H V Y S V A K T D S F E Y
CGTGGACCCTGTGGATGGCACTGTGACCAAGAAACAGGGGCCTAAGGATCATTTTCTCGGATGCATCACGGC~CATCTTCCGGCTCAGTTC V D P V D G T V T K K Q G L R I I F S D A S R L I F R L S S
CTCCAGTGGTGTGCGGGCCACCCTCAGACTGTACGCAGAGAGCTACGAGAGGGATCCCAGCGGCCATGACCAGGAGCCACAGGCAGTGCT S S G V R A T L R L Y A E S Y E R D P S G H D Q E P Q A V L
GAGCCCTCTCATAGCCATCGCACTGAAAATATCCCAGATTCATGAGAGAACTGGCCGGAGGGGACCCACTGTCATCACCtgaatagagga S P L I A I A L K I S Q I H E R T G R R G P T V I T
aagatcactcaccagggccaaagagagtgctcagcgggagatgcttcactgatgccttcttgctacctgtttgtgcctcttatgactttg
gaaaaacaaaaqatattttgcttttgqgggatagagggtgggtgggaaaagaaaaaaaatccatttggttttggttttgtcctattcctc
caaatgcagcagggcctttagttgtctgttaaagctgcactataatttggtatctacattttatcacacaaaggaacctccccttttgac
aacaactgggctaggcagctgttaatcacaacatttgtgcatcacttgtgccaagtgagaaaatgttctaaaatcacaagagagaacagt
gccagaatgaaactgaccctaagtcccaggtgcccctgggcaggcagaaggagacactcccagcatggaggagggtttatcttttcatcc
taggtcaggtctacaatqggggaaggttttattatagaactcccaacagcccacctcactcctgccacccacccgatggccctgcctccc
ccatcccatccccaacatccctgtaccaccttctctcacatcttctaaagctttgtacaaatcacaatggtgcacttccaacaaaatata
tcaataggtgttttcctctctcaaaaaaaaaaaaaaaaaaaa
90
1 8 0 30
270 60
360 90
450 1 2 0
540 1 5 0
630 1 8 0
72 0 210
810 240
900 2 7 0
990 300
1080 330
1170 360
1260 3 9 0
1350 420
1440 450
1530 480
1620 506
1710
1800
1890
1980
2070
2160
2250

106 Moiseeva ct al. (Eur: J. Biochenz. 235)
* I 2 B 1 2
- 7.5 - 4.4
- 2.4
- 1.35
Fig. 2. Northern-blot hyhridisation. Tola1 RNA isolated from rabbit skclctal muscle (lanes 1) and from human uterus (lanes 2) was separated in an agarose gel, blotted onto Hybond N and hybridised to a human PGM-RP probe (A). The blot was stripped and hybridised to a human P G M f probe (B). The positions of molecular s i x markers (kb) are indi- cated on the right.
RESULTS
Isolation and characterisation of cDNAs encoding a novel member of the phosphoglucomutase gene family. Since PGM- RP is most abundant in smooth muscle, we constructed a human uterine smooth muscle cDNA library in the expression vector AZAPII. The library was screened with a polyclonal antibody raised against PGM-RP which also cross-reacts with PGMl . Two clones containing the largest cDNA inserts were analysed in more detail. One was a partial cDNA (P87) encoding PGMl and was equivalent to nucleotides 372-2320 of the published human sequence [17]. The second cDNA ( A l l l ) encoded a fu- sion protein that was also recognised by a monoclonal antibody XIVF8 specific for PGM-RP. This clone was sequenced com- pletely on both strands. The cDNA (2286 bp) contained an open reading frame of 1609 nucleotides (1 - 1609) followed by 659 nucleotides of 3’-untranslated sequence preceding a poly(A) tail (Fig. 1). An additional cDNA clone ( A S ) encoding a fusion protein recognised by both the polyclonal and monoclonal anti- bodies was also isolated. The 5‘ end of the A85 cDNA was equivalent to nucleotide 1233 of the A l l l cDNA (codon 382) and the 3‘ end extended four nucleotides beyond that of the A1 11 cDNA (Fig. 1). The rest of the cDNA clones isolated rep- resented partially spliced mRNAs, with exodintron boundaries i n the same positions as they were found in the mouse PGM-RP gcne (Fig. 5 B).
The first ATG codon is at position 92 and is embedded in the sequence caccATGG which is close to the consensus eukaryotic translation initiation site 1241. The 5‘-untranslated region has a higher G + C content (65%) than the coding region (52%) and the 3’-untranslated region (46 %). Unfortunately, the N-terminus of the protein is blocked, and we have been unable to determine the N-terminal sequence directly. However, we have sequenced a human genomic DNA fragment encoding the first PGM-RP exon, and there is no additional 5’ ATG codon within 500 nucle- otides of the ATG identified in the cDNA sequence (data not shown). We have previously determined the amino acid se- quence of four peptides derived from the protein purified from human uterus smooth muscle, and covering a total of 69 amino acid residues [ 3 31. There was a near perfect match between the determined amino acid sequence and the deduced amino acid scquence of clone A1 11 confirming that this cDNA encodes a novel PGM-related protein, and not PGMI (Fig. 1). The only discrepancy is at amino acid 183 where there is an aspartic acid residue in the deduced sequence but a glutamatic acid residue in the determined sequence of PGM-RP. This discrepancy could be a result of mistakes in reverse transcription or DNA synthesis
during cDNA library construction, amplification or cloning, or it might represent a polymorphism. PGM-RP is predicted to con- tain 506 amino acids with a deduced molecular mass of 55.6 kDa, and is therefore smaller than PGMl (566 amino acids; 61 kDa). This is consistent with the relative mobilities of the two proteins in SDS/PAGE (Fig. 4).
The PGM-RP cDNA hybridised to a single 3.5-kb mRNA in Northern blots of human uterus smooth muscle RNA, but no signal was detected using rabbit skeletal muscle RNA (Fig. 2A). In contrast, the human PGMl cDNA hybridised to a single 2.5- kb mRNA in rabbit skeletal muscle (significantly smaller than that encoding PGM-RP), but 110 signal was detected in human smooth muscle (Fig. 2B). The PGM-RP mRNA in uterus is of low abundance as estimated by a ribonuclease protection assay (data not shown). Of the six PGM-RP cDNA clones we have isolated, four clones represent partly spliced mRNAs. This ob- servation is consistent with the proposal that the number of fully processed PGM-RP mRNA molecules in the uterus is small. The PGM-KP cDNA contains the sequence AACAAA (nucleotides 2240-2245) which is similar to a polyadenylation signal (AA- TAAA), and precedes the start of the poly(A) tail by 22-26 nucleotides. The 3’ untranslated sequence is therefore 658 - 662 nucleotides long. Assuming that the average length of a poly(A) tail is about 200-250 nucleotides, the total length of the coding sequence (1 51 8 nucleotides) plus the 3’hntranslated sequence including the poly(A) tail is about 2.4 kb. The observation that the PGM-RP cDNA hybridized to a 3.5-kb mRNA in smooth muscle suggests that the 5’-untranslated sequence must be of the order of 1 kb in length.
Comparison of the nucleotide and amino acid sequences of PGM-RP with other members of the PGM family. The nucle- otide sequence encoding human PGM-RP shows 70% identity with mammalian PGMI enzymes [17, 25, 261, 55% identity with yeast PGM [27], and 59% identity with bacterial PGM [28] (Fig. 3). The similarity between the PGM-RP and mammalian PGMl sequences breaks down 14 nucleotides after the start of the 3’-untranslated sequence, although the 3’-untranslated se- quences of mammalian PGMI enzymes are highly conserved. The similarity with the yeast PGMl nucleotide sequence is re- stricted to the first two thirds of the PGM-RP coding sequence (nucleotides 81 -1 132). Interestingly, the level of amino acid sequence identity of human PGM-RP to mammalian PGMl is about the same as that at the nucleotide level (Fig. 3). In con- trast, the level of amino acid sequence identity between the mammalian PGMI family (90-97 %) is substantially greater than that at the nucleotide sequence level (83 - 87 %). This im- plies that many of the nucleotide differences in the gene encod- ing the PGM-RP isoform lead to amino acid substitutions, whereas the nucleotide substitutions in the mammalian PGMI genes are generally in the third base position within a codon, and do not affect the amino acid sequence. Thus, PGM-RP is less conserved than other mammalian PGMl genes. The diver- gence in amino acid sequence is greatest in the so-called N- terminal domain 1 [29], which is 30% shorter in the human PGM-RP, and in the C-terminal domain 4, which contains twice as many amino acid differences as the rest of the sequence. PGM-RP shows a similar level of amino acid sequence identity to the Purumecium protein parafusin (a new member of the PGM family) 1301 as it does to yeast PGM (Fig. 3). Despite the low level of identity between the mammalian, yeast and bacterial PGM enzymes, peptide sequences that are in, or close to the active site of PGMl , according to the crystal structure of rabbit PGMl 1291, show a high degree of conservation. In contrast, the peptide sequences in PGM-RP equivalent to those thought to be in, or close to the active site, show a lower degree of conserva-

Moiseeva et al. ( E m J. Biochem. 235)
-7 i. 68 69 69 70 69 38 39 40 46
I93 97 90 96 46 47 45 51
1 9 1 96 91 45 46 44 50
! 93 97 46 47 45 51
92 45 46 45 51
I
: 46 47 45 51 ................
107
%
amlno
acid
iden-
tity
A
B PGMRP HPGMl HPGMla OPGMl OPGMla RP GM YPGMl YPGM2 PFUSP APGMP
PGMRP HPGMl OPGMl RP GM YPGMl YPGM2 PFUSP APGMP
PGMRP HPGMl OPGMl RP GM YPGMl YPGMZ PFUSP APGMP
PGMRP HPGMl OPGMl RPGM YPGMl YPGMZ PFUSP APGMP
PGMRP HPGMl OPGMl RPGM YPGMl YPGMZ PFUSP APGMP
PGMRP HPGMl OPGMl RPGM YPGMl YPGMZ PFUSP APGMP
PGMRP
HPGMl
HPGMl
OPGMl
OPGMl
RPGM
SPGMl
APGM
a
a
88 84
56 56 53
% nucleotide identity
MVVGSDGRYFSRTAIEIWQMAAANG MVKIVTVKTQAYQDQKPGTSGLRKRVKVFQSSANYAENFIQSIISTVEPAQRQEATLWGGDGRFYMKEAIQLIARIRAANG
MSDF--EEWISGTYRKMEEGPLPLLTFATAPYHDQKPGTSGLRKKTYYFEEKPCYLENFIQSIFFSIDLKDRQGSSLWGGDGRYFNKSAIETIVQMAAANG MVKIVTVKTKAYPDQKPGTSGLRKRVKVFQSSTNYAENFIQSIISTVEPAQRQEATLWGGDGRFYMKEAIQLIVRIAAANG
MSDS--DGWISGTYRKMEEGPLPLLTIRTAPYHDQKPGTSGLRKKTYYFEDKPCYLENFIQSIFFSIDLKDRQGSSLWGGDGRYFNKSAIETILQMAAANG MVKIVTVKTQAYPDQKPGTSGLRKRVKVFQGNANYAENFIQSIVSTVEPALRQEATLWGGDGRFYMTEAIQLIVRIAAANG
MSLLIDSVPTVAYKDQKPGTSGLRKKTKVFMDEPHYTENFIQATMQSI-PNGSEGTTLWGGDGRFYNDVIMNKIAAVGAANG MSFQIETVPTKPYEDQKPGTSGLRKKTKVFKDEPNYTENFIQSIMEAI-PEGSKGATLWGGDGRYYNDVILHKIAAIGAANG
WLFLLPLRLGHNLWRIEAPRVQV----TQPYAGQKPGTSGLRKKVSE-ATQPNYLENFVQSIFNTL-RKDELKNVLFVGGDGRYFNRQAIFSIIRLAYAND MIKTIKTTPYQDQKPGTSGLRKKVPVF-AQENYAENFIQSIFDALE--GFEGQTLVIGGDGRYYNREVIQKAI~G . .*.**** . . .. * .
IGRLIIGQNGILSTPAVSCIIRKIK------AAGGIILTASHCPGGPG-GEFGVKFNVANGGPAPDWSDKIYQISKTIEEYAICP-DL-R-IDLSRLGRQE I G R L V I G Q N G I L S T P A V S C I I R K I K - - - - - - A I G G I I L T ~ - G D F G I K F N I S N G G P A P E A I T D K I F Q I S K T I E E Y A V C P - D L - K - V D L G V L G K Q Q IGRLVIGQNGILSTPAVSCIIRKIK------AIGGIILTASHNPGGPN-GDFGIKFNISNGGPAPEAITDKIFQISKTIEEYAICP-DL-K-VDLGVLGKQQ IGRLVIGQNGILSTPAVSCIIRKIK------AIGGIILTASHNPGGPN-GDFGIKFNISNGGPAPEAITDKIFQISKTIEEYAICP-DL-K-VDLGVLGKQQ VRKLVIGQGGLLSTPAASHIIRTYEE---KCTGGGIILTASHNPGGPE-NDLGIKYNLPNGGPAPESVTNAIWEASKKLTHYKIIK-NFPK-LNLNKLGK-- IKKLVIGQHGLLSTPAASHIMRTYEE---KCTGG-IILTASHNPGGPE-NDMGIKYNLSNGGPAPESVTNAIWEISKKLTSYKIIK-DFPE-LDLGTIGK-- ISEVHVGQAGLMSTPASSHYIRKVNEEVGNC-IGGIILTASHNPGGKEHGDFGIKFMIRTGAPAPEDFTDQIYTHTTKIKEYLTVDYEFEKHINLDQIGVYK FGKVLVGQGGILSTPAASNVIRKYK------AFGGIVLSASHNPGGPT-EDFGIKYNIGNGGPAPEKITDAIYARSKVIDSYKISD-AA-D-IDLDKIG--S . .** *..**** .* .............................. ..* .*
FD---LENKFKPFRVEIVDPVDIYLNLLRTIFDF~IKGLLTGPSQ---LKIRID~HG~GPYVR~LCDELGAPANSAINC-VPLEDFGGQHPDPNLTYA FD---LENKFKPFTVEIVDSVEAYATMLRSIFDFSALKELLSGPNR---LKICID~HGWGPYVKKILCEELGAPANSAVNC-VPLEDFGGHtiPDPNLTYA FD---LENKFKPFTVEIVDSVEAYATMLRNIFDFNALKELLSGPNR---LKIRIDAMHGWGPYVKKILCEELGAPANSAVNC-VPLEDFGGHHPDPNLTYA FD---LENKFKPFTVEIVDSVEAYATMLRNIFDFNALKELLSGPNR---LKIRIDAMHGWGPYVKKILCEELGAPANSAVNC-VPLEDFGGHHPDPNLTYA ------ NQKYGPLLVDIIDPAKAYVQFLKEIFDFDLIKSFLAKQRKDKGWKLLFDSLNGITGPYGKAIFVDEFGLPAEEVLQ~HPLPDFGGLHPDPNLTYA ------ NKKYGPLLVDIIDITKDYVNFLKEIFDFDLIKKFIDNQRSTK~KLLFDS~GVTGPYGKAI~DEFGLPADEVLQ~HPSPDFGGMHPDPNLTYA FEGTRLEK--SHFEVKWDTVQDYTQLMQKLFDFDLLKGLFSN----KDFSFRFDGMHGVAGPYAKHIFGTLLGCSKESELLNCDPSEDFGGGHPDPNLTYA FK---VDE----LTVDVIDPVADYAALMEELFDFGAIRSLIAGG-----F~DSMSAVTGPYAVEILEKRLGAPKGSVRNA-TPLPDFGGHHPDPNLVHA . . . . . . . . . . . . . . . ............. **** ******
TTLLEAMKG------GE-YGFGAAFDADGDRYMILGQNGFFVSPSDSLAIIAANLSCIPYFRQMGVRGFGRSMPTSMALDRVAKSMKVPVYETPAGWRFFSN ADLVETMKS------GE-HDFGAqEPGPGPBNMILGKHGFFVNPSDSVAVIAANIFSIPYFQQTGVRGFARSMPTSGALDRVASATKIALYETPTGWKFFGN ADLVETMKS------GE-HDFGAAFDGDGDGDRNMILGKHGFFVNPSDSVAVIAANIFSIPYFQQTGVRGFARSMPTSGALDRVANATKIALYETPTGWKFFGN ADLVETMKS------GE-HDFGAAFDGDGDGD~MILGKHGFFVNPSDSVAVI~IFSIPYFQQTGVRGFARSMPTSGALDRVANATKIASYETPTGWKFFGN RTLVDRVDR-------EKIAFGAASDGDGDRNMIYGYGPAFVSPGDSVAIIAEYAPEIPYFAKQGIYGLARSFPTSSAIDRVAAKKGLRCYEVPTGWKFFCA SSLVKRVDR-------EFIEFGAASDGDGDRNMIYGYGPSFVSPGDSVAIIAEYAAEIPYFAKQGIYGLARSFPTSGAIDRVAKAHGLNCYEVPTFWKFFCA HDLVELLDIHKKKDVGTVPQFGAACDGDADRNMILGRQ-FFVTPSDSLAVIAANANLI--F-~GLLG~RSMPTSGALD~AA~GIKLFETPTGWKFFGN KELYDDVMS------PEGPDFGAASDGDGDRNMVVGK-GMFVTPSDSLAIIAANAKLAPGYA-AGISGIARSMPTSAAADRVAEKLGLGMYETPTGWKFFGN
* . * *** *.*.** *. * ** *.**.*.** . *. * .** * * * * *.** - .* *. *.**
LMDSGRCNLCGEESFGTGSDHLREKDGLWAVLWJLSIIAR-------RKQSVEEIVRDHWAKFGRHYYCRFDYEGLDPKTTYYIMRDLEALVTDKSFIGQQF LMDASKLSLCGEESEGTGSDHIREPlDGLWAVLAWLSILAT-------RKQSVEDILKDIiWQKHGRNFFTBXPXEEVEAEGANKMMKDLEALMFDRSFVGKQF LMDASKLSLCGEESFGTGSDHIREKDGLWAVLAWLSILAT-------RKQSVEDILKDHWHKFGRNFFTRYDYEEVEAEGATKMMKDLEA~FDRSFVGKQF LMDASKLSLCGEESFGTGSDHIREKDGLWAVLAWLSILAT-------RKQRVEDILKDHWQKFGRNFFTRYDYEEVEAEGANKMMKDLEALMLDRSFVGKQF LFDAKKLSICGEESFGTGSNHIREKDGLWAIIAWLNILAIYHRRNPEKEASIKTIQDEFWNEYGRTFFTRYDYEHIECEQAE~ALLSEFVSRPNVCGSHF LFDAKKLSICGEESFGTGSMHVREKDGVI~WLNILAIYNKHHPENEASIKTIQNEFWAKYGRTFFTRYDFEKVETEKANKIVDQL~~TKSGWNSAF LMDAGLINLCGEESFGTGSNHIREKDGIWAVLAWLTILA-HKNKNTDHFVTVEEIVTQYWQQFGRNYYSRYDYEQVDSAGAN~EHLKT---------- KF
* *. . * * * * * * * * * * *.*****.**.. * * *.* . . . . . . . . . . . . . . . . . AVGSHVYSVAKTDSFEYVDPVDGT~KKQGLRIIFSDASRLIFRLSSSSGV~TLRLYAESYERDPSGHDQEPQAVLSPLIAIALKISQIHERTGRRGPTVIT SANDKVYTVEKADNFEYSDPVDGSISRNQGLRLIFTDGSRIVFRLSGTGSAGATIRLYIDSYEKDVAKINQDPQ~~PLISIALKVSQLQERTGRTAPTVIT SANDKVYTVEKADNFEYHDPVDGSVS~QGLRLIFADGSRIIFRLSGTGSAGATIRLYIDSYEKDNAKINQDPQ~LAPLISIALKVSQLQERTGRTAPTVIT SANDKVYTVEKADNFEYSDPVDGSISKNOGLRLIFADGSRIIFRLSGTGSAGATIRLYIDSYEKDAAKINQDPQVM~PLISIALKVSQLQERTGRTAPTVIT PADESL-TVIDCGDFSYRD-LDGSISENQGLFVKFSNGTKFVLRLSGTGSSGATIRLYVEKYTDKKENYGQTADVFLKPVINSIVKFLRFKEILGTDEPTVRT PADESL-KVTDCGDFSYTD-LDGSVSDHQGLYVKLSNGARFVLRLSGTGSSGATIRLYIEKYCDDKSQYQKTAEEYLKPIINSVIKFLNFKQVLGTEEPTVRT QYFEQLKQGNKADIYDYVDPVDQSVSKNQGVRFVFGDGSRIIFRLSGTGSVGATIRIYFEQFEQQ--QIQHETATA~NIIKLGLEI8DIAQFTGRNEPTVIT
................................. t . * . . * * * *
LMDAGKVTICGEESFGTGSNHVREKDGLWAVLYWLNIVAA-------RKESVKDIVTKHWAEYGRNYYSRHDYEEVDSDARNTLVAILREKL--ATLPGTSY
GN----LKVAAADDFAYHDPVDQSVS~QGIRILFEGGSRIVLRLSGTGTAGATLRLYVERYEPDAARHGIETQSALADLISVADTIAGIKAHTADSEPTVIT
Fig. 3. Nucleotide and amino acid sequence identities between the human PGM-RP and other members of the PGM gene family. (A) Identity (%) bctween the following nucleotide and amino acid sequences: PGMRP, human PGM-RP; HPGM1, human PGMI 1161; HPGMla, human fast muscle PGMI isoform 1391 ; OPGMl , rabbit PGMI 1221 ; OPGMl a, rabbit muscle-specific PCMl isoform 1221 ; RPGM1, rat PGMI [23] ; SPGMl and SPGM2, Saccharornpces cerevisiae PGMI and PGM2 1241 ; PFUS, parafusin from Parumecium [27] ; APGM, Ajirohacteriurn tumeficiens [ 251. (B) A comparison of the complete amino acid sequences of the above proteins. The following are indicated: identical residue (.,); conserved residues (0). Amino acid residues at or close to the active sitc of rabbit PGMl 1261 are underlined. The amino acid sequence of OPGMla was translated i'roin the first methionine in the open reading frame.
108 Moisceva ct al. ( E u J. Rioclzenz. 235)
Table 1. Assignment of the human PGM-related protein gene to chromosome 9. +, chromosome present; -, chromosome absent; p, short arm present; 4, long arm present; *, chromosome fragment present; n d . , not determined. The following are indicated in the rcsults columns: +, PCR fragmcnt detected; -, PCR fragment not detected.
Hybrids Chromosome Hybrid- Second ization PCR ~ _ _ -
1 2 3 4 5 6 7 8 Y 10 I1 12 13 14 15 I6 17 18 19 20 21 22 X Y
GM072Y GM10826B GM102.53 HHW416 GM10114 MCP6BRA CLONE21 E C4a GMl0611 7628d JIU4 1 aA9602+ VE 289 GM 10479 HORLI PCTBAI.8 DLI 8TS GM I06 I2 GMI 0478 THYR1.3.3 PGME2Snu HORL9X 8.53 MOG2 DIS20 F4Sc13C112 CTP34B4 MOG34A4 ClOb2BU 640-63q12
A B C D
200-
116- 97-
68- - PGMl - PGMRP 45-
30-
a b c d a b c d a b c d a b c d
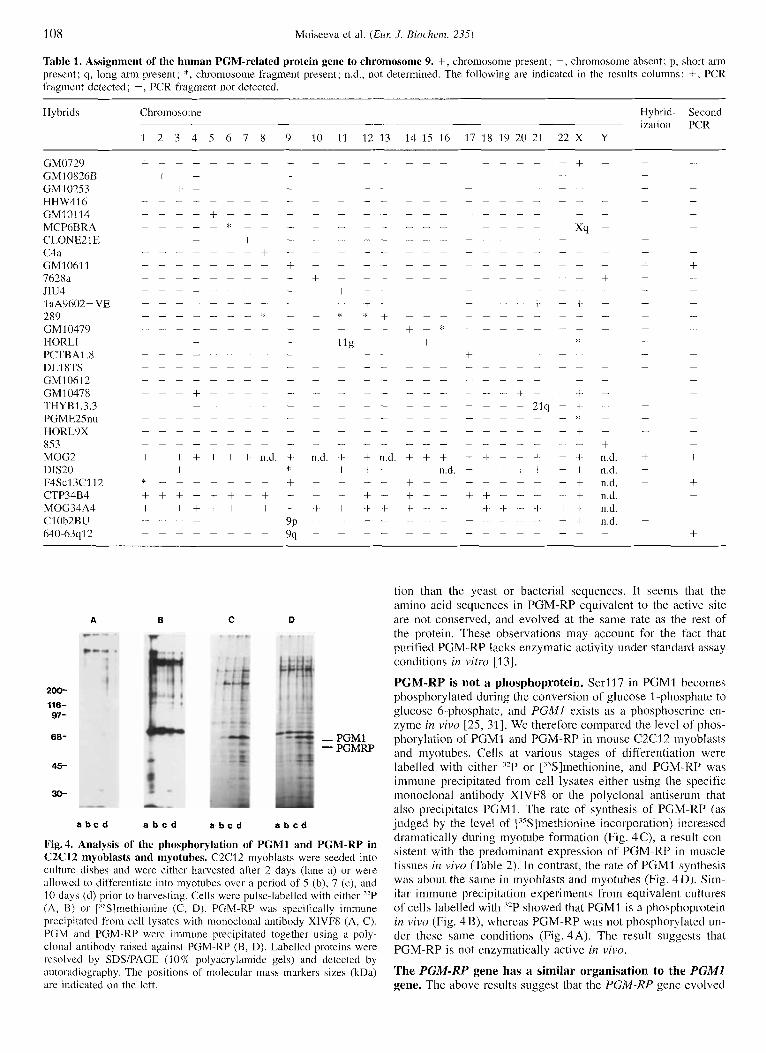
Fig.4. Analysis of the phosphorylation of PGMl and PGM-RP in C2C12 myoblasts and myotubes. C2C12 inyoblasts were seeded into culture dishes and were either harvcstcd after 2 days (lane a) or were allowed to differentiate into myotubcs over a period of .5 (b), 7 (c), and 10 days (d) prior to harvesting. Cells wcrc pulse-labelled with either "P (A, B) or j"S1methionine (C, D). PGM-RP was specifically immune prccipilated from cell lysates with monoclonal antibody XlVFX (A, C). PGM and PGM-KP were immune precipitated together using a poly- clonal antibody raised against PGM-RP (B, D). Labelled proteins were rcsolvcd by SDSlPAGE (1 0% polyacrylarnide gels) and detected by autoradiography. The positions of molecular inass markers sizes (kDa) are indicated on the left.
tion than the yeast or bacterial sequences. It seems that the amino acid sequences in PGM-RP equivalent to the active site are not conserved, and evolved at the same rate as the rest of the protein. These observations may account for the fact that purified PGM-RP lacks enzymatic activity under standard assay conditions in vitro 11 31.
PGM-RP is not a phosphoprotein. Serl17 i n PGMl becomes phosphorylated during the conversion of glucose 1 -phosphate to glucose 6-phosphate, and PGMI exists as a phosphoserine en- zyme in vivo [25, 311. We therefore compared the level of phos- phorylation of PGMI and PGM-RP in mouse C2C12 inyoblasts and myotubes. Cells at various stages of differentiation were labelled with either '*P or ["S]methionine, and PGM-RP was immune precipitated from cell lysates either using the specific monoclonal antibody XIVF8 or the polyclonal antiserum that also precipitates PGMI. The rate of synthesis of' PGM-RP (as judged by the level of 1"s lmethionine incorporation) increased dramatically during myotube formation (Fig. 4C), a result con- sistent with the predominant expression of PGM-RP in muscle tissues in vivo (Table 2). In contrast, the rate of PGMI synthesis was about the same in myoblasts and myotubes (Fig. 4D). Sim- ilar immune precipitation experiments from equivalent cultures of cells labelled with '*P showed that PGMI i s a phosphoprotein in vivo (Fig. 4B), whereas PGM-RP was not phosphorylated un- der these same conditions (Fig. 4A). The result suggests that PGM-RP is not enzymatically active in vivo.
The PGM-RP gene has a similar organisation to the PGMI gene. The above results suggest that the PGM-KP gene evolved
Moiseeva et al. ( E m .I. Biochem. 235) 109
A 1
B
\ X1 , 5kb ,
PX e9 .. I 89
I I I bi3 B E E li
C
e x o n 2
e x o n 3
e x o n 5
e x o n 8
e x o n 9
cctggact actttcctgtctaaatgccttcaagtccatgaaccttggactgactatttcccgtattttttttttttagATTGGTCGACTG 27 I l e G l y A r q L e u
ATTATTGGGCAGAATGGCATCTTGTCGACCCCTGCAGTCTCCTGCATTATCAGG~GATC~GGCAGCTGGTGGGATCATTCTAACAGCC IleIleGlyGlnAsnGlyIleLeuSerThrProAlaValSerCysIleIleArgLysIleLysAlaAlaGlyGlyIleIleLeuThrAla
A G C C A C T G T C C T G G A G G A C C A G G G G G A G A G T T T G G A G T G A T G G A G g t a t g t g g t t c c a a a t a a g c c t t a a c t t a SerHisCysProGlyGlyProGlyGlyGluPheGlyValLysPheAsnValAlaAsnGlyG 81
cattttggatgtcaaattttagGTCCTGCACCAGATGTTGTCTCAGACAAAATCTACCAGATCAGC~CAATTGAGGAATATGCCATA 81 lyProAlaProAspValValSerAspLysIleTyrGlnIleSerLysThrIleGluGluTyrA~aIle
TGTCCAGATCTTCGAATTGACCTATCTCGACTGGGAAGACAAGAATTTGACCTGGAGAACAAGTTCAAACCATTCAGAGqtccagatatc CysProAspLeuArgIleAspLeuSerArgLeuGlyArgGlnGluPheAspLeuGluAsnLysPheLysProPheArgV 130
cccagggaagagtattcttcaggcatgtttatattttcttctatcttttttttttttaagTTATGGGACCCTATGTAG~GTTCTGT 172 alMetGlyProTyrValArgLysValLeuC
GTGATGAACTGGGAGCTCCAGCC~CTCCGCRATAAACTGTGTCCCACTAG~GACTTCGGAGGGCAGCACCCAGACCC~CCTGACCT ysAspGluLeuGlyAlaProAlaAsnSerAlaIleAsnCysValProLeuGluAspPheGlyGlyGlnHisProAspProAsnLeuThrT
ATGCAACCACCCTCCTGGAAGCCATGAAGGGGGGAGAATATGGATTTGGAGCTGCGTTTGATGCTGATGGGgtgagtgagaqgqtttcca yrAlaThrThrLeuLeuGluAlaMetLysGlyGlyGluTyrGlyPheGlyAlaAlaPheAspAlaAspGly 235
ttttgccttagGGTCTGATCACCTCCGAGAGAAAGATGGCCTCTGGGCTGTCTTGGTCTGGCTGTCCATTATTGCTGCCCGGAAGCAGAG 3 2 6 lySerAspH~sLeuArqGluLysAspGlyLeuTrpAlaValLeuValTrpLeuSerIleIleAlaAlaArgLysGlnSe
C G T G G A G G A A A T T G T T A G A G A C C A T T G G G C C A A A T A T G G C G G C C A C T A C T A C T G C A G g t g a g a a g g g t c a g q c t t t c t g c a t g t q t c t t g rValGluGluIleValArgAspHisTrpAlaLysTyrGlyGlyHisTyrTyrCysAr 3 7 1
ccaccctctgattgggttttgttttcctgtgtcctgctctcccccaatagGTTTGATTACGAAGGACTGGACTGGAGCCCAAGGC~CCTATTAC 3 7 1 gPheAspTyrGluGlyLeuGluProLysAlaThrTyrTyr
ATCATGAGGGATCTGGAGGCCCTAGTGACAGACAAGTCCTTCATTGGCCAGCAGTTTGCTGTAGGGAGCCACATTTACAGCATTGC~G IleMetArgAspLeuGluAlaLeuValThrAspLysSerPheIleGlyGlnGlnPheAlaValGlySerHisIleTyrSerIleAlaLys
ACTGACAGCTTTGAATATGTGGACCCCGTGGATGGCCCCGTGACCAAG~CAGgtatttgcagggcagtcgctgaaacagaggqatcag ThrAspSerPheGluTyrValAspProValAspGlyProValThrLysLysGln 432
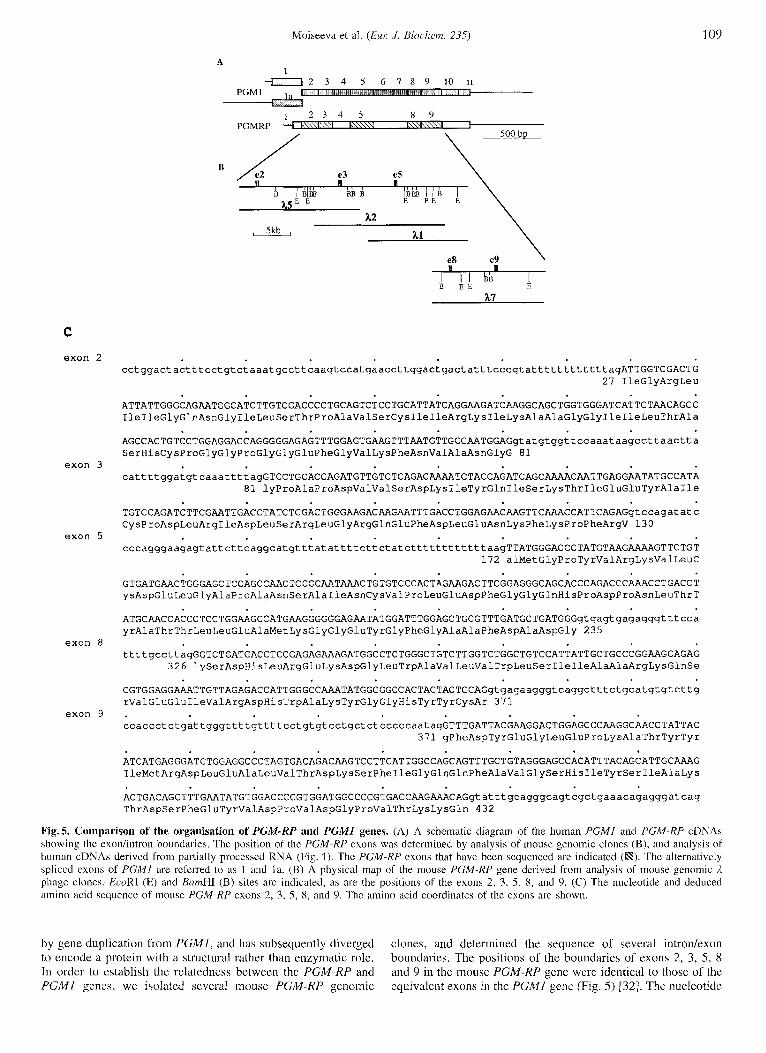
Fig.5. Comparison of the organisation of PGM-RP and PGMI genes. (A) A schcmatic diagram of the human P G M l and PGM-RP cDNAs showing the exonlintron boundarics. The position of thc PGM-RP exons was determined by analysis of mouse genomic clones (B), and analysis of human cDNAs derived from partially processed RNA (Fig. 1). The PGM-RP exons that have been sequcnced are indicated (R). The alternatively spliccd cxons of PGMl are rcfcrred to as 1 atid la . (B) A physical map of the mouse PGM-RP gene derived from analysis of mouse genomic 3, phage clones. EcoRI (E) and BamHI (B) sites arc indicated, as are the positions of the exons 2, 3, 5 , 8, and 9. (C) The nucleotide and deduced amino acid sequence of mouse PGM-RP exons 2, 3, 5 , 8, and 9. Thc amino acid coordinates of the exons are shown.
by gene duplication from PGMI, and has subsequently diverged to encode a protein with a structural rather than enzymatic role. In order to establish the relatedness between the PGM-RP and P G M l genes, we isolated several mouse PGM-RP genomic
clones, and determined the sequence of several introntexon boundaries. The positions of the boundaries of exons 2, 3, 5, 8 and 9 in the mouse PGM-KP gene were identical to those of the equivalent exons in the PGMl gene (Fig. 5) (321. The nucleotide

110 Moiseeva et al, (EM 1. Hiuchem. 235)
Tablc 2. Distribution of‘ PCM-KP in human tissues. +, positive iininuiiofluorescence staining in cryostat sections.
Source Tissue type PGM-RP
Skin
Gastrointestinal tract
Pancreas
Genitourinary tract
Livcr
Respiratory tract
Nervous systcin
Cardiovascular system
Connective tissue
Skcletal muscle
B rcast
Kidncy
epidermis hair follicle sebaceous gland erector pili
salivary gland epithelium visceral smooth muscle intestinal epithelium
ducts acini islets of Langerhans
smooth muscle epithelial tissue
hepatocytes bile ducts vascular smooth muscle capsula
smooth muscle epithelial tissue
neuroglia neurons Schwann cells
endothelium smooth muscle connective tissue cardiac muscle
fibroblasts adipoc ytes cartilage
in yotendinous junctions
ductal and alveolial epithelium vascular smooth muscle
glomcruli proximal and distal tubuli -
sequences of the mouse exons and the human cDNA sequence varied in the range 85-96% identity. At the amino acid level, the sequences had 90 % - 100 96 identity.
Chromosome assignment of the PGM-HP gene. Blots of hu- man geiiomic DNA cut with either EcoRI or BamHI were hy- bridised to a probe containing the 3’-untranslated region of the PGM-RP cDNA. This region shows no similarity with the PGMI gene. Single 3.7-kb EcoRI and 3.2-kb BarnHI fragments werc detected (data not shown) indicating that PGM-RP is en- coded by a single-copy gene. To determine the chromosomal location of the gene, DNA from a panel of human-rodent hy- brids was analysed by PCR using primers based on the sequence of the 3’-untranslated sequence of the gene (5’-primer nucleo- tides 1636- 1657 ; 3’-primer nucleotides 2261 -2234). PCR products werc resolved in agarose gels, and the product resulting from amplification of the PGM-related gene (626 bp) was de- tected by Southern blotting using a labelled oligonucleotide (nu- cleotides 1903- 1922) internal to the two primers. Alternatively, a nested 5‘ primer (iiucleotides 1680-1699) was used in a sec- ond PCR reaction to increase the specificity of the initial ampli- fication. In both cases, the specific PCR product was only de- tected when analyzed hybrids contained human chromosome 9 (Table 1). Furthermore, we also obtained the specific PCR prod-
Table 3. Quantitative analysis of PGM-RP expression in different human tissues. Thc relative levels of PGM-RP in human tissues were determined by Western blotting using a monoclonal antibody (XIVFII) specific for PGM-RP RS described in the Materials and Methods section. Bound monoclonal antibody XIVF8 was detected with a radiolabelled iodinated secondary antibody.
Tissues 10 - 4 X Radioactivity
Brain Skin Kidney Liver Skeletal muscle Cardiac muscle Aortic media Aortic intima Uterus smooth muscle
cpin
0.10 0.23 0.42 0.26 0.71 1.45 3.46 3.60 5.10
uct with DNA from hybrid 640-63 Yq, which contains the long arm of human chromosome 9. However, we did not obtain the PCR product using DNA from the ClOb2BU hybrid which con- tains only the short arm of chromosome 9, and the DIS20 hybrid which contains the 9q21-9qter fragment of human chromosome 9 translocated to chromosome 10. Therefore, the gene encoding PGM-RP is located on chromosome 9ycen-q13. The PGMl gene has been previously mapped to chromosome lp31 [ 171.
Distribution of PGM-RP in human tissues. In order to estab- lish the tissue distribution of PGM-RP, human tissue sections were stained with monoclonal antibody XIVFX, which is spe- cific for PGM-RP. The major sites of immunoreactivity were found in visceral and vascular smooth muscle in various tissues, with some staining in heart muscle, and the myotendinous junc- tions of skeletal muscle (Table 2). To determine the relative lev- els of expression of the protein, tissue lysates were analysed by a SDS/PAGE and Western blotting. The highest levels of PGM- RP were found i n uterine and aortic smooth muscle with inter- mediate levels in cardiac muscle, and lesser amounts in skeletal muscle (Table 3). The trace amounts of the protein found in other tissues (kidney, liver, brain and skin) can be attributed to the presence of vascular smooth muscle cells within these tis- sues.
DISCUSSION
In the present study, we have determined the complete nucle- otide sequence of a 60/63-kDa protein which we previously identified as a novel component of adherens-type cell-cell and cell-matrix junctions [13]. Analysis of the deduced amino acid sequence shows that it is new member of the PGM family. PGM-RP contains 506 amino acids with a molecular mass of 55.6 kDa, and is significantly shorter than other members of the PGM family. This conclusion is entirely consistent with the mo- bility differences of PGMl and PGM-RP in SDS gels. We have also shown that PGM-RP expressed in Swiss 3T3 cells from the A1 1 1 cDNA has a mobility in SDS gels indistinguishable from that of the protein isolated from human uterus (data not shown). Although the A l l 1 cDNA (2286 bp) contains all of the coding sequence for PGM-RP, it hybridized to a 3.5-kb mRNA in hu- man uterus. Both the A1 l l and A85 cDNA clones contain the

Moiseeva et al. (Eur: J. Biochem. 235) 111
authentic 3’-untranslated region of the PGM-RP mRNA with AACAAA and GTGTTTT sequences, similar to the AATAAA polyadenylation signal and YGTGTTYY consensus sequence re- quired for efficient formation of the 3’ end of a mRNA 1331. Thus, the A l l l cDNA lacks much of the S’-untranslated se- quence which is about 1 kb in length. Interestingly, the mRNA for parafusin, another member of the PGM family identified in Paramecium, has a 0.9-kb 5‘-untranslated sequence [30 I. We have used the RACE protocol and primer extension in an at- tempt to synthesise the 5’ end of the cDNA, but the products stopped near the 5’ end of the A l l l cDNA. It appears that the 5’ end of the PGM-RP mRNA has a tightly folded tertiary struc- ture. It has been shown that the 5‘ ends of viral and rare cellular mRNAs with long Suntranslated regions have a complex sec- ondary structure that is involved in the formation of internal ribosome entry sites required for mRNA translation 1341.
Sequence alignments show that PGM-RP is less conserved than mammalian PGMl enzymes at both the nucleotide and amino acid sequence levels. Comparison of the mammalian 117, 25, 26, 311, yeast [27], and prokaryotic 1281 PGM enzymes re- veals consensus sequences in the active site around S e r l l 7 (T/ SASHNP), which acts as a phosphate acceptor in the phosphot- ransfer reaction involved in conversion of glucose 1 -phosphate to glucose 6-phosphate, and in the metal-binding site (F/ SDGDGDR). Several other mutases such as N-acetylglucosam- ine phosphomutase (Saccharornyces cerevisiae) [35] and phos- phomannomutase ( P . ~ e / ~ ~ l o m o n a s aeruginosa) 136 I contain very similar peptide sequences (TA/GSHNP and FDGDAEDR). Thus, these sequences are conserved in mutases with a wide range of substrate specificities among prokaryotes and eukary- otes. However, the analagous sequences i n PGM-RP (TASHCP and FDADGDR) deviate from these consensus sequences. Furthermore, the sequence QKPCTSGLRKKV near the N-ter- minus of all PCM enzymes is conserved, and forms part of the active site cleft 1311. PGM-RP is shorter than other PGM pro- teins and lacks the N-terminal region containing this sequence. The regions in PGM enzymes important in catalysis are there- fore mutated or deleted in PGM-RP. These observations are con- sistent with the observation that purified PGM-RP lacks enzy- matic activity in vitro, and it is not a phosphoprotein in vivo. Together these results provide compelling evidence that PGM- RP is not an enzyme.
To date, three human PGM genes ( P G M I - 3 ) have been identified and mapped to chromosomes 1, 4, and 6, respectively [I 7, 37 I. Evidence for a fourth gene (PGM4) encoding the PGM isoenzyme found in milk has been presented [38]. However, only PGMI has been sequenced. The novel amino acid sequence of PGM-RP, and the localisation of the gene to chromosome 9qcen-q13 confirms that it is distinct from P G M I , as well as the PGM2 and PGM3 genes. Since PGM-RP is predominantly expressed in smooth muscle, we also conclude that it is likely to be different from the PGM4 found in milk. The PGMI pro- tein, which is encoded by a highly polymorphic gene, is immu- nologically distinct from the PGM2 and PGM3 isoforms, but does show immunological similarities with the PGM-RP iden- tified here. Therefore, the PGMI and PGM-RP genes would be predicted to be the most closely related in evolutionary terms. Indeed, analysis of the PGM-RP gene shows that it has the same intron/exon structure as PGMl [32]. These results provide evi- dence that the PGM-RP gene arose by duplication of the an- cestral PGMl gene, and has evolved independently without con- servation of those residues at the active site of PGMI. We pro- pose that the PGM-RP gene has evolved to encode a protein with a structural rather than an enzymatic role in adherens junc- tions. The idea that PGM family members have evolved to fufill other roles is supported by the recent studies on parafusin 1301,
another PGM-related protein, which also lacks enzymatic activ- ity 1391, and is involved in exocytosis 1401 in Purumecium. A similar path of evolution has been described for the crystallin genes. The arginosuccinate lyase gene (62-crystallin gene) was duplicated during the evolution of birds. The second gene (61- crystallin gene), encodes a protein with a similar sequence, but which acts as a lens structural protein, with no enzyme activity in some species, or low activity in others 1411.
The function of PGM-RP remains to be established, but the fact that it coimmunoprecipitates with dystrophin and utrophin, and binds to dystrophin as assayed by gel filtration [ I S ] strongly suggests that it is part of the dystrophinhtrophin cytoskelal coni- plex. Adhalin, a-dystroglycan and &dystroglycan and the 3S- kDa dystrophin-associated protein are all resistant to alkaline extraction, whereas proteins such as dystrophin and syntrophin that are associated with the cytoplasmic face of this complex are extracted under these conditions [42]. PGM-RP is also extracted in alkaline buffers 1131, as would be expected of a cytoskeletal protein that is bound to dystrophinhtrophin on the cytoplasmic surface of the membrane. This conclusion is compatible with the tissue and subcelluar distribution of PGM-RP. In skeletal mus- cle, PGM-RP colocalises with dystrophin in the sarcolemma, and with dystrophin and utrophin in the myotendinous junction [ 13, IS, 43, 441. In cardiac muscle, PGM-RP co-localises with dystrophin at the sarcolemma, and with utrophin in intercalated discs [13, 43, 441. In cultured smooth muscle cells, PGM-RP colocalises with utrophin [ I 5 1. Analysis of the tissue distribution of PGM-RP shows that the highest levels of expression are in vascular and visceral smooth muscle, which more closely resem- bles the distribution of utrophin rather than dystrophin 1451. Lit- tle information is available on the dystrophirdutrophin-associ- ated protein complex in smooth muscle, but adhalin (SO kDa) is replaced by an immunologically related 70-kDa protein [ 461 and several new components have been identified that are not pre- sent in the complex isolated from skeletal and cardiac muscle [47]. PGM-RP is likely to be one of these new components.
The position of the PGM-RP-binding sites in dystrophin and utrophin has not been investigated. The C-terminal regions of both these proteins have been shown to bind the integral mem- brane protein p-dystroglycan, and the cytoplasmic syntrophins 12, 8, 481. It is likely that PGM-RP binds to dystrophinhtrophin at a site distinct from that recognised by these other proteins because antibodies to syntrophin precipitate a complex contain- ing PGM-RP, dystrophin, and syntrophin from C2C12 myotubes 11 51.
This work was supported by a grant from the Wellcome Trust to DRC, and a European Union Grant to VEK and DRC. AMB is a Muscu- lar Dystrophy Association postdoctoral fellow. We thank B. V. Shecho- nin (Institute of Experimental Cardiology, Moscow, Russian Federation) for help with the PGM-RP tissue distribution study, and Dr David White- house (University College London, England) and Dr Kathryn Lilley (University of Leicester, England) for helpful discussions.
REFERENCES 1. Koenig, M., Monaco, A. P. 8 Kunkel, L. M. (1988) The complete
sequence of dystrophin predicts a rod-shaped cytoskeletal protein, Cell 5.3, 219-228.
2. Tinsley, J. M., Blake, D. J., Zuellig, R. A. & Davies, K. E. (1994) Increasing complexity of the dystrophin-associated protein com- plex, Proc. Natl Acad. Sci. USA 91, 8307-8313.
3. Pons, F., Augier, N., Heilig, R., Leger, J., Mornet, D. & Eger, J. J. (1990) Isolated dystrophin molecules as seen by elcctron micro- scopy, Proc. Natl Acad. Sci. USA 87, 7851 -7855.
4. Hemmings, L., Kuhlman, P. A. & Critchley, D. R. (1992) Analysis of the actin-binding domain of cx-actinin by mutagenesis and tlem-

112 Moiseeva el al. ( E m J . Hioclzem. 235)
onstration that dystrophin contains a functionally homologous do- main, .I. Cell Hiol. 116, 1369-1380.
5. Blake, D. J., Tinslcy, J. M. & Davies, K. E. (1994) The emerging family of dystrophin-related proteins, Trends Cell Biol. 4 , 19-23.
6, Pctrof, B. J., Shrager, J. B., Stedman, H. H., Kelly, A. M. & Sweeney, H. L. (1993) Dystl-ophin protects the sarcolemma from stresses developed during muscle contraction, Proc. Null Acud.
7. Pastcrnak, C., Wong, S. & Elson, E. L. (1995) Mechanical function of dystrophin in muscle cells, J . Cell Biol. 128, 355-361.
8. Camphcll, K. P. (1995) Three muscular dystrophies: loss of cy- toskclcton-extracellular matrix linkage, Cell 80, 675 -679.
9. Suzuki, A,, Yoshida, M. & Ozawa, E. (1995) Mammalian (11- and /&syntrophin bind to the alternative splice-prone region of the dystrophin COOH terminus, J . Cell Biol. 128, 373-381.
10. Ahn, A. H. &. Kunkel, L. M. (1995) Syntrophin binds to an alterna- tivcly spliced exon of dystrophin, J . Cell B id . 128, 363-371.
11, Matsumura, K., Tomc, F. M. S., Collin, H., Azibi, K., Chaouch, M., Kaplan, J.-C., Fardeau, M. & Campbell, K. P. (1992) Deficiency of the 50 K dystrophin-associated glycoprotein in severe child- hood autosomal recessive muscular dystrophy, Nufure 359, 320- 322.
12. Khurana, T. S., Kunkel, L. M., Fredcrickson, A. D., Carhonetto, S. & Watkins, S. C. (1 995) Interaction of chromosoine-6-encode~~ dystrophin related protein with the extracellular matrix, J. Cell Sci. 108, 173 - 185.
13. Belkin, A . M., Klimanskaya, 1. V., Lukashcv, M. T., Lilley, K., Critchley, D. R. & Koteliansky, V. E. (1994) A novel phosphoglu- coniutasc-related protein is concentrated in adherens junctions of muscle and nonmuscle cells, J. Cell Sci. 107, 159-173.
14. Belkin, A. M. & Burridge, K. (1994) Expression and localization of the phosphoglucoinutase-related cytoskeletal protein, aciculin, in skeletal muscle, .I. Cell Sci. 107, 1993-2003.
IS. Belkin, A. M. &. Burridge, K. (1995) Association of aciculin with dystrophin and utrophin, J. Biol. Chem. 270, 6327-6337.
16. Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989) Molecular clon- ing: a luhorutory manual, 2nd edn, Cold Spring Harbor Labora- tory, Cold Spring Harbor NY.
17. Whitehouse, D. B., Putt, W., Lovegrove, J. U., Morrison, K., Hol- lyoake, M., Fox, M. F., Hopkinson, D. A. & Edwards, Y. H. (1992) Phosphoglucomutase 1 : complete human and rabbit mRNA sequences and direct mapping of this highly polymorphic marker on human chromosome 1, Proc. Nafl Accrd. Sci. USA 89,
18. Blau, H. M., Chiu, C.-P. & Wcbster, C. (1983) Cytoplasmic activa- tion of human nuclear genes in stable heterocaryons, Cell 32,
19. Spurr, N. K., Cough, A,, Goodfellow, P. J., Goodfellow, P. N., Lee, M. G. & Nurse, P. (1988) Evolutionary conservation of the human homologue ol' the yeast cell cycle control gene ccic2 and assign- ment of Cd2 to chromosome 10, Hum. Genet. 78, 333-337.
20. Kelsell, D. P., Rooke, L., Warne, D., Bouzyk, M., Cullin, L., Cox, S., West, L., Povey, S. & Spurr, N. K. (1995) Development of a panel of monochromosotnal somatic cell hybrids for rapid gene mapping, Ann. Hum. Genet. 59, 233-241.
21. Shephard, E. A,, Povey, S., Spurr, N. K. & Phillips, I. R. (1991) Chromosomal localization of a cytochromc h, gene to human chromosome 18 and a cytochrome h, pseudogene to the X chro- mosome, Genonzic.7 / I , 302-308.
22. Heisterkamp, N., Groffen, J., Stcphenson, J. R., Spurr, N. K., Good- fellow, P. N., Solomon, E., Carritt, R. & Bodmer, W. F. (1982) Chromosomal localization of human cellular homologues of two viral oncogenes, Nature 299, 747-749.
23. Barncs, W. M. (1994) PCR amplification of up to 35-kb DNA with high fidelity and high yield from /z bacteriophage templates, Proc.
24. Kozak, M. (1 987) An analysis of 5'-noncoding sequences from 699 vcrtehrate messenger RNAs, Nucleic Acid.s Res. IS, 8126-81
25. Lee, Y. S., Marks, A. R, Gureckas, N., Lacro, R., Nadal-Ginard, B. & Kim, D. H. (1992) Purification, characterization and molec- ular cloning of a 60-kDa phosphoprotcin in rabbit skeletal sarco- plasmic reticulnm which is an isoform of phosphoglucomutase,
Sci. USA 90, 3710-3714.
41 1-415.
'I 1 71 - 1 1 80.
N d A C U ~ Sci. USA 91, 2216-2220.
J. Hiol. Chem. 267, 21 080-21 088.
26. Rivera, A. A., Elton, T. S., Dey, N. B., Bounelis, P. & Marchase, K. B. (1993) Isolation and expression of a rat liver cDNA cncoding phosphoglucomutase, Gene (Amst.) 133, 261 -266.
27. Boles, E., Liebetrau, W., Hofmann, M. & Zimmermann, F. K. (1994) A family of hcxosephosphate mutases in Succharomyr,e.s cerevi- .sine, Eur: J. Biochem. 220, 83-96.
28. Uttaro, A. D. & Udalge, R. A. (1994) A chromosomal cluster of genes encoding ADP-glucose synthetase, glycogen synthase and phosphoglucomutase in Agrobacferium runieficiens, Gene (Amsf.)
29. Dai, J.-B., Liu, Y., Ray, W. J. & Konno, M. (1992) Th,c crystal structure of muscle phosphoglucomutase refined at 2.7-Angstrom resolution, J . Biol Chem. 267, 6322-6337.
30. Subramanian, S. V., Wyroba, E., Andersen, A. P. & Satir, B. H. (1 994) Cloning and sequencing of parafusin, a calcium-dependent exocytosis-related phosphoglycoprotein, Proc. Nut1 Acud. Sci.
31. Ray, W. J . Jr, Hermodson, M. A,, Puvathingal, J . M. & Mahoney, W. C. (1 983) The complete amino acid sequence of rabbit muscle phosphoglucomutase, J . Biol. Chem. 258, 91 66-9174.
32. Putt, W., Ives, J. H., Hollyoake, M., Hopkinson, D. A,, Whilehouse, D. B. & Edwards, Y. H. (1993) Phosphoglucomutasc 1 : a gene with two promoters and a duplicated first exon, Biochem. J. 296,
33. McLauchlan, J., Gaffney, D., Whitton, J. L. & Clements, J . B. (1 985) The consensus sequence YGTGTTYY located down- stream from the AATAAA signal is required for efficient forma- tion of mRNA 3' termini, Nucleic Acids Res, I.?, 1347-1368.
34. Altmann, M. & Trachsel, H. (1993) Regulation of translation initia- tion and modulation of cellular physiology, Trends Biochem. Sci. 18, 429-432.
35. Hofmann, M., Boles, E. & Zimmermann, F. K. (1994) Characteriza- tion of the essential yeast gene encoding N-acetylglucosainine- phosphate mutase, Eur: J. Hiochem. 221, 741 -747.
36. Coyne, M. J., Kristin, J. R., Russell, S., Coyle, C. L. & Goldberg, J. B. (1994) The Pseudnmonus aeruginosu algC gene encodes phosphoglucomutase, required for the synthesis of a complete lipopolysaccharide core, .I. Bacteriol. 176, 3500- 3507.
37. McAlpine, P. J., Stranc, L. C., Boucheix, C. & Shows, T. B. (1990) The 1990 catalog of mapped genes and report of the nomenclature committee, Cytogenet. Celf Genet. 55, 5 -76.
38. Cantu, 1. M. & Ibarra, B. (1982) Phosphoglucoinutase: evidence for a new locus expressed in human milk, Science 216, 639- 640.
39. Anderscn, A. P., Wyroba, E., Reichman, M., Zhao, H. & Satir, H. (1994) Thc activity of parafusin is distinct from that of phospho- glucomutase in the unicellular eukaryote Purcirnecium, Rioclwm. Biophys. Res. Commuii 200, 1353-1358.
40. Gilligan, D. M. & Satir, B. H. (1982) Protcin phosphorylation/ dephosphorylation and stimulus-secretion coupling in wild type and mutant Parumecium, J. B id . Chem. 257, 13 903 - 13 906.
41, Wistow, G. (1993) Lens crystallins: gene recruitment and evolution- ary dynamism, Trend.s Biochem. Sci. 18, 301 -306.
42. Ervasti, J. M. & Campbell, K. P. (1991) Membrane organization of the dystrophin-glycoprotein complex, Cell 66, 1 121 - 11 31.
43. Byers, T. J., Kunkel, L. M. & Watkins, S. C. (1991) Thc subcellular distribution of dystrophin in mouse skeletal, cardiac, and smooth muscle, J. Cell Bid . 115, 41 1-421.
44. Pons, F., Robert, A,, Fabbrizio, E., Hugon, G., Califano, J.-C., Feh- rentz, J. A., Martinez, J. & Mornet, D. (1994) Utrophin localisa- tion in normal and dystrophin-deficient heart, Circulation 90, 369-374.
45. Man, N., Ellis, J. M., Love, D. R., Davies, K. E., Gatter, K. C., Dickson, G. & Morris, G. E. (1991) Localization of the DMDL gene-encoded dystrophin-related protein using a panel of nineteen monoclonal antibodies: presence at neuromuscular junctions, in the sarcolemma of dystrophic skeletal muscle, in vascular and other smooth muscles, and in proliferating brain cell lines, J. Cell
46. Mizuno, Y., Yoshida, M., Yamamoto, H., Hirai, S. & Ozawa, E. (1 993) Distribution of dystrophin isoforins and dystrophin-associ- ated proteins 43DAG (A3a) and SODAG (A2) in various monkey tissues, J . Hiochem. 114, 936-941.
150, 117-122.
USA 91, 9832-9836.
41 7 -422.
Biol. 115, 1695-1700.

Moiseeva ct al. (EZN J . Biochem. 235) 113
47. Kfiiinarcy, N. R., Vidal, A., Frochner, S. C. & Sealock, R. (1994) 48. Suzuki, A,, Yoshida, M., Hayashi, K., Mizuno, Y., Hagiwara, Y. & Association of utrophin and inultiplc dystrophin short forins with Ozawa, E. (I 994) Molecular organisation at the glycoprolein- the mammalian M , 58 000 dystrophin-associatcd protein complex-binding site of dystrophin : threc dystrophin-associated (synlrophin), ./. Biol. Chem. 269, 2870-2876. proteins bind directly t o the carboxy-terminal portion of
dystrophin, Eur: J. Biochem. 220, 283-292.
Related Documents











