Hindawi Publishing Corporation BioMed Research International Volume 2013, Article ID 125492, 11 pages http://dx.doi.org/10.1155/2013/125492 Research Article A Novel Bone Morphogenetic Protein 2 Mutant Mouse, nBmp2NLS tm , Displays Impaired Intracellular Ca 2+ Handling in Skeletal Muscle Laura C. Bridgewater, 1 Jaime L. Mayo, 1 Bradley G. Evanson, 2 Megan E. Whitt, 1 Spencer A. Dean, 1 Joshua D. Yates, 1 Devin N. Holden, 1 Alina D. Schmidt, 1 Christopher L. Fox, 1 Saroj Dhunghel, 1 Kevin S. Steed, 1 Michael M. Adam, 1 Caitlin A. Nichols, 1 Sampath K. Loganathan, 1 Jeffery R. Barrow, 3 and Chad R. Hancock 2 1 Department of Microbiology and Molecular Biology, Brigham Young University, 775-A WIDB, Provo, UT 84602, USA 2 Department of Nutrition, Dietetics, and Food Science, Brigham Young University, Provo, UT 84602, USA 3 Department of Physiology and Developmental Biology, Brigham Young University, Provo, UT 84602, USA Correspondence should be addressed to Laura C. Bridgewater; laura [email protected] Received 18 June 2013; Revised 12 October 2013; Accepted 29 October 2013 Academic Editor: Kunikazu Tsuji Copyright © 2013 Laura C. Bridgewater et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. We recently reported a novel form of BMP2, designated nBMP2, which is translated from an alternative downstream start codon and is localized to the nucleus rather than secreted from the cell. To examine the function of nBMP2 in the nucleus, we engineered a gene-targeted mutant mouse model (nBmp2NLS tm ) in which nBMP2 cannot be translocated to the nucleus. Immunohistochemistry demonstrated the presence of nBMP2 staining in the myonuclei of wild type but not mutant skeletal muscle. e nBmp2NLS tm mouse exhibits altered function of skeletal muscle as demonstrated by a significant increase in the time required for relaxation following a stimulated twitch contraction. Force frequency analysis showed elevated force production in mutant muscles compared to controls from 10 to 60 Hz stimulation frequency, consistent with the mutant muscle’s reduced ability to relax between rapidly stimulated contractions. Muscle relaxation aſter contraction is mediated by the active transport of Ca 2+ from the cytoplasm to the sarcoplasmic reticulum by sarco/endoplasmic reticulum Ca 2+ ATPase (SERCA), and enzyme activity assays revealed that SERCA activity in skeletal muscle from nBmp2NLS tm mice was reduced to approximately 80% of wild type. ese results suggest that nBMP2 plays a role in the establishment or maintenance of intracellular Ca 2+ transport pathways in skeletal muscle. 1. Introduction Bone morphogenetic proteins (BMPs) are members of the transforming growth factor (TGF-) super family, and the first members of the family were identified by their ability to induce ectopic bone formation in animals [1, 2]. BMPs have since been shown to participate in multiple developmental pathways, including axis formation, limb patterning, heart development, neural crest cell migration, neurogenesis, apop- tosis, and others [3–12]. e BMP family of proteins is the largest subfamily in the TGF- superfamily, containing over twenty members including the growth and differentiation factors (GDFs) [5, 13]. BMP proteins are synthesized as preproproteins, which are directed by N-terminal signal peptides to the rough endo- plasmic reticulum (ER) for translation, processing, and sec- retion from the cell. While in the secretory pathway, BMP proproteins homodimerize by disulfide bonding and are cleaved by furin-type proprotein convertase enzymes to pro- duce the mature secreted BMP growth factors [14, 15]. Sec- reted BMPs bind to cell surface receptors and trigger cellular responses through the SMAD and the mitogen-activated protein kinase (MAPK) pathways [16–19]. Secreted BMP growth factors have been widely stud- ied since their discovery over two decades ago. Recently, however, we found that some BMP family members can be

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Hindawi Publishing CorporationBioMed Research InternationalVolume 2013 Article ID 125492 11 pageshttpdxdoiorg1011552013125492
Research ArticleA Novel Bone Morphogenetic Protein 2 Mutant MousenBmp2NLStm Displays Impaired Intracellular Ca2+ Handling inSkeletal Muscle
Laura C Bridgewater1 Jaime L Mayo1 Bradley G Evanson2 Megan E Whitt1
Spencer A Dean1 Joshua D Yates1 Devin N Holden1 Alina D Schmidt1
Christopher L Fox1 Saroj Dhunghel1 Kevin S Steed1 Michael M Adam1
Caitlin A Nichols1 Sampath K Loganathan1 Jeffery R Barrow3 and Chad R Hancock2
1 Department of Microbiology and Molecular Biology Brigham Young University 775-A WIDB Provo UT 84602 USA2Department of Nutrition Dietetics and Food Science Brigham Young University Provo UT 84602 USA3Department of Physiology and Developmental Biology Brigham Young University Provo UT 84602 USA
Correspondence should be addressed to Laura C Bridgewater laura bridgewaterbyuedu
Received 18 June 2013 Revised 12 October 2013 Accepted 29 October 2013
Academic Editor Kunikazu Tsuji
Copyright copy 2013 Laura C Bridgewater et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
We recently reported a novel form of BMP2 designated nBMP2 which is translated from an alternative downstream start codonand is localized to the nucleus rather than secreted from the cell To examine the function of nBMP2 in the nucleus we engineered agene-targetedmutantmousemodel (nBmp2NLStm) inwhichnBMP2 cannot be translocated to the nucleus Immunohistochemistrydemonstrated the presence of nBMP2 staining in the myonuclei of wild type but not mutant skeletal muscle The nBmp2NLStmmouse exhibits altered function of skeletal muscle as demonstrated by a significant increase in the time required for relaxationfollowing a stimulated twitch contraction Force frequency analysis showed elevated force production inmutant muscles comparedto controls from 10 to 60Hz stimulation frequency consistent with the mutant musclersquos reduced ability to relax between rapidlystimulated contractions Muscle relaxation after contraction is mediated by the active transport of Ca2+ from the cytoplasm to thesarcoplasmic reticulum by sarcoendoplasmic reticulum Ca2+ ATPase (SERCA) and enzyme activity assays revealed that SERCAactivity in skeletal muscle from nBmp2NLStm mice was reduced to approximately 80 of wild type These results suggest thatnBMP2 plays a role in the establishment or maintenance of intracellular Ca2+ transport pathways in skeletal muscle
1 Introduction
Bone morphogenetic proteins (BMPs) are members of thetransforming growth factor 120573 (TGF-120573) super family and thefirst members of the family were identified by their ability toinduce ectopic bone formation in animals [1 2] BMPs havesince been shown to participate in multiple developmentalpathways including axis formation limb patterning heartdevelopment neural crest cellmigration neurogenesis apop-tosis and others [3ndash12]
The BMP family of proteins is the largest subfamily inthe TGF-120573 superfamily containing over twenty membersincluding the growth and differentiation factors (GDFs) [5
13] BMP proteins are synthesized as preproproteins whichare directed byN-terminal signal peptides to the rough endo-plasmic reticulum (ER) for translation processing and sec-retion from the cell While in the secretory pathway BMPproproteins homodimerize by disulfide bonding and arecleaved by furin-type proprotein convertase enzymes to pro-duce the mature secreted BMP growth factors [14 15] Sec-reted BMPs bind to cell surface receptors and trigger cellularresponses through the SMAD and the mitogen-activatedprotein kinase (MAPK) pathways [16ndash19]
Secreted BMP growth factors have been widely stud-ied since their discovery over two decades ago Recentlyhowever we found that some BMP family members can be
2 BioMed Research International
translated in a novel alternative form that is translocated tothe nucleus rather than being secreted from the cell [20]BMP2 BMP4 and GDF5 all have nuclear variants which wenamed nBMP2 nBMP4 and nGDF5 respectively Nuclearlocalization is mediated in each case by a bipartite nuclearlocalization signal (NLS) that overlaps the furin proproteinconvertase cleavage site Cleavage at this site would destroythe NLS but because these proteins are translated from analternative downstream start codon and thus lack the N-terminal signal peptide they are translated in the cytoplasmrather than the ER and thus avoid contact with the proproteinconvertases in the Golgi apparatus The intact NLS directsnuclear translocation [20]
The conservation of nuclear variants among three distinctBMP family members suggested a physiologically importantrole for these novel nuclear proteins [20] To evaluate thein vivo requirement for nBMP2 we generated nBmp2NLStmtargeted mutant mice which produce nBMP2 that cannottranslocate to the nucleus The conventional BMP2 growthfactor however is still secreted and functions normally Herewe report that the nBmp2NLStm mouse exhibits impairedskeletal muscle relaxation rates suggesting a defect in theintracellular transport of Ca2+
2 Materials and Methods
21 Cell Culture and Transfections Rat chondrosarcoma(RCS) cells were maintained in Dulbeccorsquos Modified EaglersquosMedium (DMEM) supplemented with penicillin (50 umL)streptomycin (50120583gmL) L-glutamine (2mM) and 10 fetalbovine serum at 37∘C under 5 CO
2 Cells were passaged
every 3-4 daysTo determine the effect of theRKR toAAANLSmutation
on the nuclear localization of nBMP2 wtBmp2GFP orRKRmBmp2GFP fusion plasmids (both containing the GFPtag at the C-terminus of BMP2) were transfected into RCScells on Lab-Tek II Chamber Slides (ISC Bioexpress) usingthe TransIT-Jurkat Transfection Reagent (Mirus MadisonWI) according to themanufacturerrsquos instructions 48 hrs aftertransfection cells were fixed using 4 paraformaldehydenuclei were stained with a 1 1000 dilution of TOPRO-3iodide (Invitrogen Corporation Carlsbad CA) and slideswere mounted in Fluoromount-G (Southern Biotech Birm-inghamAL) and coverslippedCellswere imaged andnuclearlocalization was quantified using an Olympus IX81 laserconfocal microscope as previously described [20]
To assess BMP2 secretion RCS cells were seeded in25 cm2 culture flasks and transfected with HA-tagged expres-sion plasmids wtBmp2HA or RKRmBmp2HA (both con-taining the HA tag at the C-terminus of BMP2) as describedabove After 48 hrs culture medium was collected andHA-tagged proteins that had been secreted into the cul-ture medium were precipitated using EZview Red Anti-HA Affinity Gel (Sigma Saint Louis MO) according tothe manufacturerrsquos protocol to concentrate the proteinsPrecipitated proteins were separated by SDS-PAGE andanalyzed by immunoblot using an anti-HA primary anti-body
22 Research Animals Experimental procedures were ap-proved by the Institutional Animal Care and Use Com-mittee (IACUC) of Brigham Young University Mice werekept in a temperature-controlled (21-22∘C) room with a12 12 hr light-dark cycle and were fed standard rodent chowand water ad libitum
23 Construction of the nBmp2NLS119905119898 Targeting Vector Thetargeting vector was constructed using ldquorecombineeringrdquoas described in [21] Briefly a genomic clone containingthe BMP2 gene was retrieved from the BAC RP23-384M14(The BACPAC Resource Center Childrenrsquos Hospital OaklandResearch Institute Oakland CA) The BMP2 bipartite NLSwas mutated by first replacing the target sequence with thegalK gene and then replacing the galK gene with the desiredmutation to alter the protein sequence of theNLS (KREKRQ-AKHKQRKRLKwas changed toKREKRQAKHKQAAALK)as described in [22] Specifically the nucleotide sequence 51015840-CGGAAGCGC (coding forRKR) was replacedwith 51015840-GCGGCG GCC (coding for AAA)
The mutant nBMP2 gene was then retrieved from theBAC into pBluescript modified to contain a thymidinekinase gene (MC1TK) A male germline self-excising neocassette (tACN) was also inserted into the targeting constructbetween the BMP2 homology arms [23]
24 Generation of the nBmp2NLS119905119898 Mouse ThenBmp2NLStm targeting vector was linearized and electro-porated into 129Bl6 G4 ES cells (generously provided byAndras Nagy of the Samuel Lunenfeld Research Institute inToronto Canada) and targeted cell lines were subjected topositivenegative selection as described by Mansour et al[24 25] Cells were analyzed by Southern blot to determinethe presence of the targeted nBMP2-mut allele Two targetedcell lines were then microinjected into Bl6 host blasto-cysts and both gave rise to chimeras Chimeric males wereintercrossed with Bl6 females to generate nBmp2NLStm+minusmice Heterozygous mice were intercrossed to produce themale minusminus (mutant) and ++ (control) mice utilized in thisstudy Successful self-excision of the neo cassette (tACN) wasverified by PCR using primers that annealed to the BMP2gene regions bracketing the neo cassette (forward primerCCTGCAGCAAGAACAAAGCAGG reverse primerCCCCAACCTTGTCATCATTCACC) PCR product sizefrom wild type was 525 bp after insertion of the targetingvector and self-excision of the neo cassette it was 607 bp
25 Genotyping To distinguish the genotypes of progenyfromheterozygous intercrosses we performed two PCR reac-tions on DNA samples from each offspring One reactionemployed a primer set that detected the presence of the wildtype allele whereas the second identified the nBmp2NLStmallele Primers sets for both reactions used the same forwardprimer but the reverse primer for the wild type reactionbound only the wild type DNA sequence and the reverseprimer for the mutant reaction bound only the mutant DNAsequence DNA from wild type mice yielded the 201 bp pro-duct only in the wild type reaction DNA from homozygous
BioMed Research International 3
mutants yielded the same size product only in the mutantreaction and DNA from heterozygotes yielded the prod-uct in both reactions Sequences for the primers were asfollows forward primer GGCCCATTTAGAGGAGAA-CC wild type reverse primer TTGCAGCTGGACTTG-AGGCGCTTCCG mutant reverse primer TTGCAGCTG-GACTTGAGGGCCGCCGC The 9 bp sequence that wasaltered to convert RKR to AAA is underlined Reactionconditions are 94∘C 5min 94∘C 20 sec 615∘C 30 sec and68∘C 40 sec repeated 30x
26 Skeletal Morphometric Analysis Two wild type andthree homozygous mutant male mice 9-10 weeks old wereeuthanized by CO
2inhalation and eviscerated soft tissues
were removed in 1KOH and skeletons were stained in a0004Alizarin red in 1KOH solution and then cleared in agraded glycerin series (20 50 80 and 100)The lengthand the proximal distal and midshaft widths at both thewidest and narrowest diameters of the femur tibia humerusand radius both left and right limbs were measured usingdigital calipers Measurements were also taken of the scapulapelvis sternum rib cage skull and vertebral column
27 GripStrength Test Male mice five homozygous wildtypes and five homozygous mutants were analyzed usingthe gripstrength test at 206 plusmn 03 weeks (wild type) and207plusmn03weeks (mutant) of age for the firstmeasurement andmeasurements were repeated once a week for six weeks Eachmouse was placed on a wire mesh cage lid which was theninverted over the open cage filled with bedding and mousewas held suspended approximately 25 cm above the bedding[26] Holding time was recorded up to a maximum of 3min
28 Western Analysis of Muscle BMP2 To compare produc-tion of conventional BMP2 in wild type versus nBmp2NLStmmouse skeletal muscle cytoplasmic extracts were preparedfrom gastrocnemius and quadriceps muscle by homogeniz-ing tissue in ice-cold buffer (025M sucrose 10mMNaCl3mMMgCl
2 1 mMDTT 1mMPMSF and a protease inhi-
bitor cocktail) centrifuging for 5min at 500timesg and col-lecting supernatant Protein concentration was quantifiedby Bradford assay before western blot analysis using BMP2primary antibody N-14 from Santa Cruz (product sc-6895)Autoradiograms were scanned and bands quantified usingAlphaEase software
29 Immunohistochemistry Gastrocnemius muscle from 6-month-old male wild type and mutant mice was isolatedembedded in paraffin and cross-sectioned at 6 120583 thick-ness After deparaffinization and rehydration sections werestained with a biotin-tagged primary antibody NBP1-19751B(Novus Biologicals) Stainingwas visualized using the LSAB+System-HRPkit fromDakowith a hematoxylin counterstainSections were viewed and imaged using a Zeiss Imager A1microscope with an AxioCam HRC camera and AxioVision47 imaging software
210 In Situ Muscle Preparation Male mice were used in themuscle stimulation experiments at 509plusmn04 weeks (control)
and 439 plusmn 75 weeks (mutant) of age Muscle preparationwas similar to that described previously [27ndash29] Briefly micewere anesthetized with 70mgkg intraperitoneal injectionof sodium pentobarbital The hamstrings were cut awayfrom the gastrocnemius plantaris and soleus (GPS) musclecomplex and the femur was secured on both the medial andlateral sides of the knee by two 16-gauge pins to preventmovement The foot was also clamped to the platform toeliminate movement of the lower leg The Achilles tendonwas then secured to a Grass Force-Displacement TransducerFT03 level arm with a calibrated tension of 700 grams Thesciatic nerve was exposed tied off and cut An electrode wasplaced directly on the sciatic nerve to achieve stimulationAnimals were supported with 100 oxygen directly to thenose throughout the procedure
211 In Situ Isometric Contractions TheGPSmuscle complexwas stimulated via electrical stimulation (2-3V stimulation005ms square wave at a frequency of 150Hz with the useof a Grass S88X Stimulator) Both a force frequency analysisand a 2Hz twitch contraction protocol were used A forcefrequency curve was developed by stimulating the muscle forten pulses at varying frequencies (10 20 40 60 80 100 120and 140 pulsessec)Thepercent ofmaximal tetanic force pro-duction was determined over all frequencies tested In orderto examine the capacity for sustained muscle contractionstwitch contractions at the rate of 2 contractionssec for 10minutes were elicited
212 Contractile Function The GPS complex data was ana-lyzed using LabScribe2 by iWorx which captured data at1000Hz The muscle was stretched to achieve maximal forceas previously described [28] Several tetanic contractionswere elicited to stretch the muscle to the length that createdmaximal force Peak twitch force was evaluated as well asone-half relaxation times every 60 pulses for 10 minutesRelaxation was determined by measuring the time requiredfor the GPS muscle to relax to 50 of the peak force forindividual contractions
213 SERCA Enzyme Activity Assay
2131 SR Membrane Purification Sarcoplasmic reticulum(SR) membranes were prepared and purified as describedby Kosk-Kosicka with minor adaptations for small musclesamples [30] All steps were performed at 4∘C unless oth-erwise specified Skeletal muscle was removed from limbsand washed in 01mMEDTA pH 70 and then homogenizedin Solution 1 (10mMMOPS 10 sucrose 01mMEDTA pH70) and pH was adjusted to between 65 and 70 using 10NaOH Samples were centrifuged at 15000timesg for 20minSupernatant was collected and filtered through one layer ofgauze then centrifuged at 40000timesg for 90min The pelletwas resuspended in Solution 2 (10mMMOPS 06MKClpH 70) and allowed to incubate at room temperature for40min Again the preparation was centrifuged at 15000timesgfor 20min and the supernatant was collected and cen-trifuged at 40000timesg for 90min The resulting pellet wasresuspended in 1mL of Solution 3 (10mMMOPS 30
4 BioMed Research International
sucrose pH 70) Protein concentrations were determinedusing a standard Bradford protein assay and preparationswere quick-frozen in liquid nitrogen for short-term stor-age
2132 Measurement of SERCA Activity SERCA activitywas measured at both 15 and 30min reaction time pointsfor every sample and both Ca2+ dependent (assay buffer50mMTris-maleate pH 74 8mMMgCl
2 120mMKCl
1mMEGTA 10 120583Mionophore A23187 1008mMCaCl2to
yield 175 120583Mfree Ca2+) and Ca2+ independent (assay buffer50mMTris-maleate pH 74 8mMMgCl
2 120mMKCl
1mMEGTA 10 120583Mionophore A23187) reactions were per-formed as described by Kosk-Kosicka [30] Briefly 02 120583gof SR membrane preparation was added to each tube andenough water was added to give a final volume of 10 120583L85 120583L of the appropriate membrane assay buffer was thenadded to each tube Reactions were started 15 sec after thebuffer was added by adding 5 120583L 60mMATP and reactionswere capped vortexed and placed in a 37∘C water bathReactions were stopped precisely 15 or 30min after ATP wasadded by adding 300 120583L Lin Morales Reagent (see [30] forreagent composition) and vortexing Results were measuredas absorbance at a wavelength of 350 nm precisely 30 sec afterthe addition of Lin Morales Reagent The Ca2+ independentand dependent reactions were performed so that the baseATPase activity (Ca2+ independent) could be subtracted fromCa2+ dependent ATPase activity Results of mutant werenormalized to wild type SERCA activity
214 Data Analysis Analyses of data from gripstrengthtests morphometric measurements in situ isometric con-tractions contractile function and SERCA enzyme activ-ity experiments were performed using two-tailed unpairedStudentrsquos 119905-tests assuming unequal variance Significance wasset at 119875 lt 005
3 Results
31 nBmp2NLS119905119898Mouse Construction Mouse embryos lack-ing all BMP2 activity have been previously described [8]These embryos died at approximately 7 days of developmentdue to defects in the formation of the chorion and amnion [8]In order to examine the effects of nBMP2 inactivation sepa-rately from the complete BMP2 knockout it was necessary todevise a mutation scheme that would leave the conventionalsecreted form of BMP2 intact while preventing the functionof nBMP2 We previously demonstrated in tissue culturethat mutating the alternative start codon from which nBMP2translation initiates only results in a 50 reduction of nBMP2relative to controls suggesting that other alternative start sitescan be used to generate forms of BMP2 that localize to thenucleus if the primary alternative start site is mutated [20]Mutation of the alternative start site therefore was unlikelyto abolish nBMP2 in a mouse model
Instead we made specific alterations to the portion of thenBMP2 gene that encodes the bipartite NLS whose altera-tions were predicted to block nuclear translocation of nBMP2yet still allow synthesis and secretion of conventional BMP2
KREKRQAKHKQRKRLK
(a)
20kDa
wtBmp2HA RKRmBmp2HA
(b)
Figure 1 The RKR to AAA mutation in the bipartite NLS doesnot inhibit secretion of the conventional BMP2 growth factor (a)Detail of the nBMP2 NLS sequence Critical amino acids in thebipartite NLS are bold and underlined the furin recognition site isboxed and the site of cleavage to release the mature secreted growthfactor from the propeptide is marked by an arrow (b) Culturemedia from cells transfected with either wild type (wtBmp2HA) ormutant (RKRmBmp2HA) BMP2 expression plasmids was collectedand analyzed by western blot The HA-tagged growth factors weresecreted equally
A consensus bipartite NLS is characterized by the followingpattern two basic residues approximately 10 spacer residuesand another basic region consisting of 4 basic residues outof five The sequence of the nBMP2 NLS is shown in Figure1(a)The upstreamKR portion of this NLS sequence overlapstheR-X-(KR)-R furin recognition sequencewhere the BMP2proprotein is cleaved to release the mature growth factorand mutation of the KR would thus disrupt production ofthe secreted growth factor (Figure 1(a)) The downstreambasic RKRLK portion of the BMP2 NLS however does notaffect propeptide cleavage To determine whether mutationof this portion of the NLS was sufficient to prevent nuclearlocalization we constructed a mutant BMP2GFP expres-sion plasmid (called RKRmBmp2GFP) in which RKR wasreplaced with AAA and transfected it into cultured cellsThis mutation eliminated nuclear localization of the GFP-tagged BMP2 in cultured RCS cells just like the previouslydescribed KR RKR to AA AAAmutation did [20] To deter-mine whether BMP2 growth factor containing this mutationcould still be secreted anHA-tagged RKRmBmp2 expressionvector (RKRmBmp2HA) was transfected into cultured RCScells and culture medium was collected 48 hrs later HA-tagged proteins were immunoprecipitated from the mediumand visualized by immunoblotting The medium from cellstransfected with RKRmBmp2HA contained as much HA-tagged BMP2 growth factor as medium from cells transfectedwith the wild type wtBmp2HA plasmid indicating that theRKR to AAA did not disrupt synthesis or secretion of theconventional BMP2 growth factor (Figure 1(b))
Having demonstrated in cell culture that theRKR toAAAmutation blocked nuclear localization of nBMP2 while stillallowing secretion of normal quantities of the conventionallyprocessed and secreted BMP2 growth factor we constructeda targeting vector and generated nBmp2NLStm mutant micebearing theRKR toAAAmutation (Figure 2) Both heterozy-gous and homozygous mutant mice appeared morphologi-cally normal and were fertile
BioMed Research International 5
Targeting vector AAA
AAA
DrdI
E1 E2 E3
Wild-type allele
E1
E1
E2
E2
E3
E3
DrdI
DrdI
DrdI
DrdIDrdI
138 kbTargeted allele
61 kb
CreNeo
CreNeo
TK
(a)
138 kb wt allele
61 kb targeted allele
(b)
1000
750
500
++ minusminus
(bp)
(c)
0
02
04
06
08
1
12
14
Gastrocnemius Quadriceps
Wild typeMutant
(d)
Figure 2 Integration of the nBmp2NLStm targeting vector (a) Schematic of the nBmp2NLStm targeting vector recombining into genomicDNA The BMP2 exons are numbered and represented as black boxes while the neo cassette and thymidine kinase (TK) gene arerepresented as grey boxes Insertion of the neo cassette introduces a DrdI restriction site as indicated The mutation of the bipartite NLS(KREKRQAKHKQRKRLKS to KREKRQAKHKQAAALKS) in exon 3 is denoted byAAA (b)After aDrdI digest and Southern hybridizationusing a 31015840 flanking probe complementary to the region indicated with the striped box the wild type band was 138 kb and the targeted alleleband was 61 kb (c) To verify male germline self-excision of the neo cassette PCR was performed using primers that bound to BMP2 outsidetheneo cassette PCRproduced a 525 bp band fromwild typeDNAand a 607 bp if the targeting vectorwas inserted and theneo cassette excisedleaving a LoxP sequence (d) Expression levels of conventional BMP2weremeasured in skeletalmuscle bywestern blot of cytoplasmic extractsusing an anti-BMP2 antibody The bands representing BMP2 proprotein precursor of secreted BMP2 were quantified digitally and mutantswere normalized to wild types within each experiment before averaging between experiments 119899 = 4 for wild type gastrocnemius 119899 = 5 formutant gastrocnemius and 119899 = 3 for both wild type and mutant quadriceps
32 Verification of Normal Secreted BMP2 Function innBmp2NLS119905119898 Mice Work by others predicted that the RKRto AAA mutation would produce secreted BMP2 that wasstill able to bind its receptors activate downstream genesand induce ectopic bone formation but that it might haveincreased diffusion range through the extracellular matrix[31 32] Increased diffusion range could disrupt embryonicpatterning but the normal appearance of the mutant miceruled out major patterning abnormalities Any minor pat-terning abnormalities would likely be manifest in the skele-ton since secreted BMP2 plays an important role in skeletaldevelopment and limb patterning Skeletal preparations ofwild type and homozygous mutant mice were performedand digital caliper measurements were taken of limbs
pelvis scapula ribcage skull and vertebrae with multiplemeasurements at various positions on each bone No sig-nificant differences were detected between wild type andmutant mice in any of the bones measured supportingthe conclusion that the function of secreted BMP2 is notimpaired in nBmp2NLStm mice (see Supplemental Table 1 inSupplementtary Materials available online at httpdxdoi)org1011552013125492)
33 Upside-Down Hanging Ability Is Decreased innBmp2NLS119905119898 Mice Because nBmp2NLStm mice appearedphenotypically normal a series of preliminary tests wereperformed to detect subtle phenotypic changes In oneevaluation five wild type and five mutant mice were tested
6 BioMed Research International
0
50
100
150
200
1 2 3 4 5 6
Hol
d tim
e (s)
+minus
SE
Weeks
Wild typeMutant
lowast lowast
Figure 3 The nBmp2NLStm mice show impaired performance inthe gripstrength test The length of time mice could hold to theunderside of a wire mesh was measured once a week for six weeksin wild type and nBmp2NLStm mice Data are presented as mean plusmnSEM and lowastdenotes 119875 lt 005 119899 = 5 per group
once a week for six weeks to measure the length of time theycould cling to the underside of a wire mesh cage lid [26] Onaverage wild type mice held on 2-3 times as long as mutantsdid at every time point and the difference was significant(119875 lt 005) at weeks 3 and 4 (Figure 3) This trend suggestedthat we should consider the possibility of a neuromusculardefect in nBmp2NLStm mice [26]
34 nBMP2 Is Detectable in Myonuclei of Wild Type but NotnBmp2NLS119905119898 Mice To examine the expression of nBMP2and BMP2 in skeletal muscle gastrocnemius muscle fromwild type and mutant mice was paraffin embedded cross-sectioned and stained using a biotin-tagged primary anti-body against BMP2 (NBP1-19751G from Novus) and a Dakovisualization system The myotubes in skeletal muscle arelong multinucleated cells with the myonuclei positionedagainst the outer edges of the myotube Brown BMP2 stain-ing of myonuclei was detectable in wild type but notnBmp2NLStm muscle whereas nonnuclear BMP2 stainingwas detectable in both wild type and mutant This resultvalidated the effectiveness of the targeted NLS-inactivatingmutation in a whole animal and it also demonstrated thatnBMP2 is expressed in skeletal muscle of control animals(Figure 4)
The presence of extranuclear BMP2 staining in bothwild type and mutant skeletal muscle suggested that theexpression of conventional BMP2 was not inhibited bythe nBmp2NLStm mutation To further examine expressionlevels of the conventional protein cytoplasmic extracts wereprepared from gastrocnemius and quadriceps muscles andexamined by western blot The BMP2 proprotein precursorof secreted BMP2 is detectable as a 50 kDa band Levels of theproprotein were not different between wild type and mutantagain suggesting unaltered expression of conventional BMP2(Figure 2(d))
Table 1 Body weight age and muscle force characteristics
Control (119899 = 7) nBmp2NLStm (119899 = 7)
Age (weeks) 509 plusmn 04 439 plusmn 75
Body weight (g) 369 plusmn 11 353 plusmn 24
GPS muscle weight (mg) 1609 plusmn 96 1717 plusmn 43
Peak tetanic tension (g) 498 plusmn 30 567 plusmn 18
Peak tetanic tension(gmg muscle) 309 plusmn 005 331 plusmn 008lowast
Peak twitch tension (g) 1926 plusmn 97 2206 plusmn 51lowast
Peak twitch tension(gmg muscle) 120 plusmn 003 129 plusmn 004
Data are presented as means plusmn SEM Where indicated lowastindicates 119875 lt 005compared to control values
35 Skeletal Muscle Relaxation Times Are Prolonged innBmp2NLS119905119898 Mice To examine the possibility of a neuro-muscular defect we measured muscle performance in situThe peak tetanic tension of the gastrocnemius plantarisand soleus (GPS) muscle group was slightly but significantlyhigher in nBmp2NLStm mice compared to control micewhen normalized to muscle weight (Table 1) No significantdifferences were observed in the peak twitch tension afternormalization to muscle weight (Table 1) and no significantdifferences were observed in muscle fatigability in responseto a 10 minute 2Hz twitch protocol (Figure 5(a))
A significant difference did emerge however when relax-ation kinetics were examined Half-relaxation times wereprolonged by up to 42 in the nBmp2NLStm muscle com-pared to control (Figure 5(b))This prolonged relaxation timewas apparent in averaged twitch performance traces frominitial (Figure 6(a)) 2min (Figure 6(b)) and 6min (Figure6(c)) time points in the 2Hz twitch contraction protocol
36 Force Frequency Analysis Exhibits a Shift in nBmp2NLS119905119898Mouse Skeletal Muscle Whenmuscle twitches are stimulatedat increasing frequencies in situ the contractions eventuallybecome so frequent that the muscle is unable to completeone relaxation before the next contraction begins and twitchpeak traces begin to merge The force or tension generatedby the muscle increases as twitch peaks merge until forcereaches its maximum and thereafter remains level regardlessof additional increases in the stimulation frequency Thiscould be considered the in situ version of a muscle crampSlowed relaxation after contraction widens each twitch peakcausing peaks to merge at a lower stimulation frequencyshifting the force frequency curve leftward
Because the nBmp2NLStm mice showed slowed relax-ation after contraction we performed a force frequency ana-lysis The nBmp2NLStm mutant mice generated significantlyelevated relative forces at 10 20 40 and 60Hz (119875 lt001) The data collected fit an expected sigmoidal curveand showed a significant leftward shift as determined bythe average force achieved at each frequency compared tothe control (Figure 7) This shift in the force frequency pat-tern as well as the slowed relaxation rates after twitch
BioMed Research International 7
(a) (b)
(c) (d)
(e) (f)
(g) (h)
Figure 4 nBMP2 is detectable in myonuclei of wild type but not nBmp2NLStm mice Gastrocnemius muscles were sectioned and stained byimmunohistochemistry (a)ndash(d) are wild type (e)ndash(h) are mutant (a) and (e) show sections at 40x magnification and all other panels show100x The brown BMP2 antibody stain is evident in myonuclei of wild type muscle whereas myonuclei stain pale blue in the mutant muscleArrows indicate some (but not all) of the myonuclei
8 BioMed Research International
0
02
04
06
08
1
12
0 500 1000
Initi
al te
nsio
n (
)
Time (s)
(a)
0
5
10
15
20
25
30
0 500 1000
Tim
e (m
s)
Time (s)
Wild typeMutant
lowast
(b)
Figure 5 Muscle function is altered in nBmp2NLStm mice (a) Muscle fatigue in both control and nBmp2NLStm GPS muscle complex wassimilar in response to 2Hz twitch contractions for 10 minutes (b) Muscle relaxation was significantly delayed in nBmp2NLStm musclecompared to WT control muscle beginning at 60 seconds and persisting throughout the 10 minutes in which these measurements weremade Data are presented as mean plusmn SEM and lowast denotes 119875 lt 005 119899 = 7 per group Only the first significant difference is marked but allvalues from 60 seconds on were significantly different from control
contractions suggests impairment of Ca2+ sequestration bythe sarcoplasmic reticulum (SR) after muscle contraction
37 SERCA Activity Is Reduced in nBmp2NLS119905119898 SkeletalMuscle The relaxation phase of a skeletal muscle contractionoccurs when Ca2+ is pumped in an ATP-dependent mannerfrom the cytosol back into the SR by sarcoendoplasmicreticulum Ca2+ ATPases (SERCA) Measurement of SERCAactivity in skeletal muscle revealed that SERCA activityin nBmp2NLStm mice is reduced to 81 plusmn 1 of wild typeactivity (Figure 8) This observation is consistent with theslowed relaxation rate observed in nBmp2NLStm muscle
4 Discussion
Several members of the BMP protein family are expressedin variant forms that are translocated to the nucleus ratherthan being secreted from the cell [20] A major challengein characterizing the function of these nuclear variants ofBMPs is that the nuclear protein is translated from the samemRNA transcript as the conventional secreted BMP2 growthfactor as demonstrated by our prior observation that bothnBMP2 and BMP2 are produced from a transfected wild typeBMP2 cDNA expression vector in tissue culture [20] Thisprecludes usage of several of the most common methodsfor studying gene function including RNAi quantitative RT-PCR and gene knockout whichwould all affect andor detectthe conventional secreted BMP2 growth factor as well asnBMP2 Instead we knocked in anRKR toAAAmutation inthe bipartite NLS to prevent nuclear localization of nBMP2while allowing production of secreted BMP2
The RKR to AAA mutation has minimal effect on thefunction of secreted BMPs BMP2 growth factor that was
truncated N-terminally of either the R K or R still hadthe capability to induce ectopic bone formation to thesame extent as wild type BMP2 [31] Moreover deletion ormutation of the homologous sequence in BMP4 produceda protein that was functionally indistinguishable from wildtype BMP4 in receptor binding antagonist binding and tar-get gene responsiveness [32] Animal cap conjugation exper-iments with Xenopus embryos however suggested that thesemutations reduced the ability of Bmp4 to bind heparin sulfatein the extracellular matrix with the result of increased diffu-sion range [32] Together these experiments predicted thatBMP2 bearing the same mutation (RKR toAAA) would alsofunction normally in terms of receptor binding and targetgene activation but might have an increased diffusion range
Secreted BMP2 plays a critical role in dorsal-ventralpatterning of the early embryo in limb patterning and inskeletogenesis so an increased diffusion range of the secretedgrowth factor is likely to cause morphological abnormalitiesin mice [33ndash35] The nBmp2NLStm mouse however dis-played nomorphological or patterning abnormalities and nodifferences in the length width thickness or shape of any ofthe bones measured indicating that there is no physiologi-cally significant alteration in the diffusion range of BMP2 inthese mice We conclude that the RKR to AAA mutation inthe nBmp2NLStm mouse preserved the function of the sec-reted BMP2 growth factor The nBmp2NLStm mouse there-fore is a good model for studying the molecular and physio-logical functions of nBMP2 as distinct from secreted BMP2
The nBmp2NLStm mouse showed decreased ability tocling upside-down to awiremesh cage lid and immunohisto-chemistry showed that nBMP2 is detectable in skeletalmusclemyonuclei of wild type but not mutant animals Mutantmuscle showed no increase in fatigability in situ however
BioMed Research International 9
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(a)
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(b)
Wild typeMutant
0
25
50
75
100
0 50 100 150(ms)
Initi
al p
eak
tens
ion
()
(c)
Figure 6 Slowed muscle relaxation is evident in twitch contractions The average muscle twitch contraction for both wild typeand nBmp2NLStm mice is shown for the initial contractions (a) the contraction following 2 minutes of stimulation (b) and the contractionfollowing 6 minutes of stimulation (c) Each trace represents the average of 7 contractions
suggesting that the reduced hang time is not related to simplemuscle fatigue Instead the shifted force frequency curve andslowed relaxation rates in the mutant muscle suggest thatmutant mice may be more prone to muscle cramping if con-tractions merge during intense muscle activity as they doduring high-frequency in situ stimulations Slowed musclerelaxation does lead to cramping in the human syndromeBrodymyopathy where the length of time required for pump-ing Ca2+ back into the SR after each contraction is increaseddue to mutations in the gene for SERCA1 the pump thatperforms this function in skeletal muscle [36ndash40] Brodypatients have also reported problems with their hands ldquolock-ingrdquo after making a tight fist [40]This cramping can be pain-ful is most frequent after exercise and grows worse withage The slowed relaxation rates in both Brody patientsand nBmp2NLStm mice suggest the possibility that nBMP2may function in some aspect of the SERCA1 productionacti-vation pathway
Our demonstration of reduced SERCA1 activity inthe nBmp2NLStm mouse supports this hypothesis The phe-notype of the nBmp2NLStm mouse however could also
result from the dysfunction of several other proteins becausemuscle contractionrelaxation cycles are quite complex Con-tractions occur when an action potential leads to activationof Ca2+ release channels (ryanodine receptors) in the SRmembrane Ca2+ floods into the cytoplasm and binds tro-ponin C which modulates the function of tropomyosin sothatmyosin can bind strongly to actin and cause shortening ofthe sarcomere The contraction ceases when active pumping(by SERCA1) of Ca2+ back into the SR depletes cytoplasmicCa2+ levels causing release of Ca2+ from troponin C [4142] The nBmp2NLStm phenotype could be caused by leakyryanodine receptors (RyR) in the SRmembrane whichwouldlead to prolonged elevation of Ca2+ in the cytosol even ifthe SERCA pumps were working normally An increasedaffinity of troponinC for Ca2+ could also cause slowedmusclerelaxation by increasing the level of cytosolic Ca2+ depletionthat would have to occur before Ca2+ was released fromtroponin C Ongoing work in our lab on other physiologicalpathways that are controlled by intracellular Ca2+movementincluding neuronal signaling immune system activation and
10 BioMed Research International
0
25
50
75
100
0 50 100 150
Max
forc
e (
)
Frequency of stimulations (pulsess)
MutantWild type
lowastlowast
lowast
lowast
Figure 7 Force frequency analysis of control and nBmp2NLStmmuscle A significant shift in the force frequency curve was observedin nBmp2NLStm muscle compared to wild type supporting theevidence for slowed relaxation in mutants Data are presented asmean plusmn SEM and lowastdenotes 119875 lt 001 119899 = 6 per group
0
02
04
06
08
1
12
Wild type Mutant
Rela
tive S
ERCA
activ
ity
Figure 8 SERCA activity is reduced in nBmp2NLStm muscle com-pared to controls SR was isolated from control and mutant muscleand Ca2+ ATPase activity wasmeasured Data are presented asmeanplusmn SEM 119875 lt 005 119899 = 3 per group
cell cycle regulation indicates that these pathways are alsodisrupted in nBmp2NLStm mice We predict therefore thatnBMP2 affects aspects of intracellular Ca2+ transport that areshared among multiple pathways rather than those (such astroponin C) that are unique to skeletal muscle
5 Conclusions
In summary the muscle phenotype that we identified inthe nBmp2NLStm mouse suggests that a significant alter-ation in intracellular Ca2+ transport resulted from impedednuclear localization of nBMP2 The prolonged relaxationkinetics the shift in the force frequency curve and the
reduction in SERCA activity are most consistent with acondition of delayed Ca2+ uptake by the SR although otherpossibilities have not been ruled out We have recentlyobserved (unpublished data) that nBmp2NLStm mice alsohave cognitive deficits that are consistent with an intra-cellular Ca2+-handling defect in hippocampus It is likelythat Ca2+ handling is disrupted in other tissues as welland that nBMP2 will be found to function in multiplemolecular pathways that govern intracellular Ca2+ transportThe nBmp2NLStm mouse constitutes a valuable model forcharacterizing the functions of nBMP2 relative to Ca2+transport pathways and for distinguishing those functionsfrom the activities of secreted BMP2
Conflict of Interests
None of the authors have any conflict of interests or compet-ing interests
Acknowledgments
This work was supported by NIAMSNIH Grant AR48839to Laura C Bridgewater Jaime L Mayo was supported by agraduate fellowship from the BrighamYoungUniversity Can-cer ResearchCenterThanks are due toDrMario R Capecchiof the University of Utah for generously providing tech-nical assistance with the stem cell electroporation blastulamicroinjections and embryo transfer steps in production ofthe nBmp2NLStm mouse Recombineering plasmids pGalKpL452 and pL253 were obtained from Neal Copeland andNancy Jenkins of the Methodist Hospital Research Institutein Houston TX
References
[1] M R Urist ldquoBone formation by autoinductionrdquo Science vol150 no 698 pp 893ndash899 1965
[2] J M Wozney V Rosen A J Celeste et al ldquoNovel regulators ofbone formation molecular clones and activitiesrdquo Science vol242 no 4885 pp 1528ndash1534 1988
[3] D ChenM Zhao andG RMundy ldquoBonemorphogenetic pro-teinsrdquo Growth Factors vol 22 no 4 pp 233ndash241 2004
[4] A M Goldstein K C Brewer A M Doyle N Nagy and DJ Roberts ldquoBMP signaling is necessary for neural crest cellmigration and ganglion formation in the enteric nervous sys-temrdquo Mechanisms of Development vol 122 no 6 pp 821ndash8332005
[5] B L Hogan ldquoBone morphogenetic proteins multifunctionalregulators of vertebrate developmentrdquo Genes and Developmentvol 10 no 13 pp 1580ndash1594 1996
[6] YMishina ldquoFunction of bonemorphogenetic protein signalingduring mouse developmentrdquo Frontiers in Bioscience vol 8 ppd855ndashd869 2003
[7] T Schlange H-H Arnold and T Brand ldquoBMP2 is a positiveregulator of Nodal signaling during left-right axis formation inthe chicken embryordquo Development vol 129 no 14 pp 3421ndash3429 2002
[8] H Zhang and A Bradley ldquoMice deficient for BMP2 arenonviable and have defects in amnionchorion and cardiac
BioMed Research International 11
developmentrdquo Development vol 122 no 10 pp 2977ndash29861996
[9] C Kawamura M Kizaki and Y Ikeda ldquoBone morphogeneticprotein (BMP)-2 induces apoptosis in human myeloma cellsrdquoLeukemia and Lymphoma vol 43 no 3 pp 635ndash639 2002
[10] M Raida J H Clement K Ameri C Han R D Leek and A LHarris ldquoExpression of bone morphogenetic protein 2 in breastcancer cells inhibits hypoxic cell deathrdquo International Journal ofOncology vol 26 no 6 pp 1465ndash1470 2005
[11] Y Du and H Yip ldquoEffects of bone morphogenetic protein 2on Id expression and neuroblastoma cell differentiationrdquoDiffer-entiation vol 79 no 2 pp 84ndash92 2010
[12] J-C GuimondM Levesque P-LMichaud et al ldquoBMP-2 func-tions independently of SHH signaling and triggers cell con-densation and apoptosis in regenerating axolotl limbsrdquo BMCDevelopmental Biology vol 10 article 15 2010
[13] C C Rider and B Mulloy ldquoBone morphogenetic protein andgrowth differentiation factor cytokine families and their proteinantagonistsrdquo Biochemical Journal vol 429 no 1 pp 1ndash12 2010
[14] D B Constam and E J Robertson ldquoRegulation of bone mor-phogenetic protein activity by pro domains and proprotein con-vertasesrdquo Journal of Cell Biology vol 144 no 1 pp 139ndash149 1999
[15] J M Wozney ldquoBone morphogenetic proteinsrdquo Progress inGrowth Factor Research vol 1 no 4 pp 267ndash280 1989
[16] A Hiyama S S Gogate S Gajghate J Mochida I M Shapiroand M V Risbud ldquoBMP-2 and TGF-120573 stimulate expression of12057313-glucuronosyl transferase 1 (GlcAT-1) in nucleus pulposuscells through AP1 TonEBP and Sp1 role of MAPKsrdquo Journal ofBone and Mineral Research vol 25 no 5 pp 1179ndash1190 2010
[17] CAlarcon A-I ZaromytidouQXi et al ldquoNuclearCDKs driveSmad transcriptional activation and turnover in BMP andTGF-120573 pathwaysrdquo Cell vol 139 no 4 pp 757ndash769 2009
[18] J Lemonnier C Ghayor J Guicheux and J Caverzasio ldquoPro-tein kinase C-independent activation of protein kinase D isinvolved in BMP-2-induced activation of stress mitogen-acti-vated protein kinases JNK and p38 and osteoblastic cell differ-entiationrdquo Journal of Biological Chemistry vol 279 no 1 pp259ndash264 2004
[19] A Moustakas and C-H Heldin ldquoFrommono- to oligo-Smadsthe heart of thematter inTGF-120573 signal transductionrdquoGenes andDevelopment vol 16 no 15 pp 1867ndash1871 2002
[20] J E Felin J L Mayo T J Loos et al ldquoNuclear variants of bonemorphogenetic proteinsrdquo BMC Cell Biology vol 11 no 1 article20 2010
[21] N G Copeland N A Jenkins and D L Court ldquoRecombi-neering a powerful new tool for mouse functional genomicsrdquoNature Reviews Genetics vol 2 no 10 pp 769ndash779 2001
[22] S Warming N Costantino D L Court N A Jenkins and NG Copeland ldquoSimple and highly efficient BAC recombineeringusing galK selectionrdquo Nucleic Acids Research vol 33 no 4article e36 2005
[23] M Bunting K E Bernstein J M Greer M R Capecchi and KR Thomas ldquoTargeting genes for self-excision in the germ linerdquoGenes and Development vol 13 no 12 pp 1524ndash1528 1999
[24] S H George M Gertsenstein K Vintersten et al ldquoDevelop-mental and adult phenotyping directly frommutant embryonicstemcellsrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol 104 no 11 pp 4455ndash4460 2007
[25] S L Mansour K R Thomas and M R Capecchi ldquoDisruptionof the proto-oncogene int-2 in mouse embryo-derived stemcells a general strategy for targeting mutations to non-select-able genesrdquo Nature vol 336 no 6197 pp 348ndash352 1988
[26] V E Papaioannou and R R Behringer Mouse PhenotypesmdashAHandbook of Mutation Analysis Cold Spring Harbor Labora-tory Press Cold Spring Harbor NY USA 2005
[27] C R Hancock J J Brault RWWiseman R L Terjung and RA Meyer ldquo31P-NMR observation of free ADP during fatiguingrepetitive contractions of murine skeletal muscle lacking AK1rdquoThe American Journal of Physiology vol 288 no 6 pp C1298ndashC1304 2005
[28] C R Hancock E Janssen and R L Terjung ldquoSkeletal musclecontractile performance and ADP accumulation in adenylatekinase-deficient micerdquoThe American Journal of Physiology vol288 no 6 pp C1287ndashC1297 2005
[29] C R Hancock E Janssen and R L Terjung ldquoContraction-mediated phosphorylation of AMPK is lower in skeletal muscleof adenylate kinase-deficient micerdquo Journal of Applied Physiol-ogy vol 100 no 2 pp 406ndash413 2006
[30] D Kosk-Kosicka ldquoCalcium signaling protocolsrdquo in Methods inMolecular Biology D G Lambert Ed vol 114 pp 343ndash354Springer New York NY USA 1999
[31] F Hillger G Herr R Rudolph and E Schwarz ldquoBiophysicalcomparison of BMP-2 ProBMP-2 and the free pro-peptidereveals stabilization of the pro-peptide by the mature growthfactorrdquo Journal of Biological Chemistry vol 280 no 15 pp14974ndash14980 2005
[32] BOhkawara S Iemura P tenDijke andNUeno ldquoAction rangeof BMP is defined by its N-terminal basic amino acid corerdquoCurrent Biology vol 12 no 3 pp 205ndash209 2002
[33] E M de Robertis ldquoSpemannrsquos organizer and the self-regulationof embryonic fieldsrdquo Mechanisms of Development vol 126 no11-12 pp 925ndash941 2009
[34] D M Maatouk K-S Choi C M Bouldin and B D Harfe ldquoInthe limb AER Bmp2 and Bmp4 are required for dorsal-ventralpatterning and interdigital cell death but not limb outgrowthrdquoDevelopmental Biology vol 327 no 2 pp 516ndash523 2009
[35] B Robert ldquoBone morphogenetic protein signaling in limboutgrowth and patterningrdquo Development Growth and Differen-tiation vol 49 no 6 pp 455ndash468 2007
[36] A Odermatt K Barton V K Khanna et al ldquoThe mutation ofPro789 to Leu reduces the activity of the fast-twitch skeletalmuscle sarco(endo)plasmic reticulum Ca2+ ATPase (SERCA1)and is associated with Brody diseaserdquoHuman Genetics vol 106no 5 pp 482ndash491 2000
[37] A Odermatt P E Taschner V K Khanna et al ldquoMutationsin the gene-encoding SERCA1 the fast-twitch skeletal musclesarcoplasmic reticulumCa2+ ATPase are associated with Brodydiseaserdquo Nature Genetics vol 14 no 2 pp 191ndash194 1996
[38] A A Benders J H Veerkamp A Oosterhof et al ldquoCa2+ home-ostasis in Brodyrsquos disease A study in skeletal muscle and cul-tured muscle cells and the effects of dantrolene and verapamilrdquoJournal of Clinical Investigation vol 94 no 2 pp 741ndash748 1994
[39] I A Brody ldquoMuscle contracture induced by exercise Asyndrome attributable to decreased relaxing factorrdquo The NewEngland Journal of Medicine vol 281 no 4 pp 187ndash192 1969
[40] Y Zhang J Fujii M S Phillips et al ldquoCharacterization ofcDNA and genomic DNA encoding SERCA1 the Ca2+-ATPaseof human fast-twitch skeletal muscle sarcoplasmic reticulumand its elimination as a candidate gene for Brody diseaserdquoGenomics vol 30 no 3 pp 415ndash424 1995
[41] M Endo ldquoCalcium-induced calcium release in skeletal musclerdquoPhysiological Reviews vol 89 no 4 pp 1153ndash1176 2009
[42] M J Betzenhauser and A RMarks ldquoRyanodine receptor chan-nelopathiesrdquo Pflugers Archiv vol 460 no 2 pp 467ndash480 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013

2 BioMed Research International
translated in a novel alternative form that is translocated tothe nucleus rather than being secreted from the cell [20]BMP2 BMP4 and GDF5 all have nuclear variants which wenamed nBMP2 nBMP4 and nGDF5 respectively Nuclearlocalization is mediated in each case by a bipartite nuclearlocalization signal (NLS) that overlaps the furin proproteinconvertase cleavage site Cleavage at this site would destroythe NLS but because these proteins are translated from analternative downstream start codon and thus lack the N-terminal signal peptide they are translated in the cytoplasmrather than the ER and thus avoid contact with the proproteinconvertases in the Golgi apparatus The intact NLS directsnuclear translocation [20]
The conservation of nuclear variants among three distinctBMP family members suggested a physiologically importantrole for these novel nuclear proteins [20] To evaluate thein vivo requirement for nBMP2 we generated nBmp2NLStmtargeted mutant mice which produce nBMP2 that cannottranslocate to the nucleus The conventional BMP2 growthfactor however is still secreted and functions normally Herewe report that the nBmp2NLStm mouse exhibits impairedskeletal muscle relaxation rates suggesting a defect in theintracellular transport of Ca2+
2 Materials and Methods
21 Cell Culture and Transfections Rat chondrosarcoma(RCS) cells were maintained in Dulbeccorsquos Modified EaglersquosMedium (DMEM) supplemented with penicillin (50 umL)streptomycin (50120583gmL) L-glutamine (2mM) and 10 fetalbovine serum at 37∘C under 5 CO
2 Cells were passaged
every 3-4 daysTo determine the effect of theRKR toAAANLSmutation
on the nuclear localization of nBMP2 wtBmp2GFP orRKRmBmp2GFP fusion plasmids (both containing the GFPtag at the C-terminus of BMP2) were transfected into RCScells on Lab-Tek II Chamber Slides (ISC Bioexpress) usingthe TransIT-Jurkat Transfection Reagent (Mirus MadisonWI) according to themanufacturerrsquos instructions 48 hrs aftertransfection cells were fixed using 4 paraformaldehydenuclei were stained with a 1 1000 dilution of TOPRO-3iodide (Invitrogen Corporation Carlsbad CA) and slideswere mounted in Fluoromount-G (Southern Biotech Birm-inghamAL) and coverslippedCellswere imaged andnuclearlocalization was quantified using an Olympus IX81 laserconfocal microscope as previously described [20]
To assess BMP2 secretion RCS cells were seeded in25 cm2 culture flasks and transfected with HA-tagged expres-sion plasmids wtBmp2HA or RKRmBmp2HA (both con-taining the HA tag at the C-terminus of BMP2) as describedabove After 48 hrs culture medium was collected andHA-tagged proteins that had been secreted into the cul-ture medium were precipitated using EZview Red Anti-HA Affinity Gel (Sigma Saint Louis MO) according tothe manufacturerrsquos protocol to concentrate the proteinsPrecipitated proteins were separated by SDS-PAGE andanalyzed by immunoblot using an anti-HA primary anti-body
22 Research Animals Experimental procedures were ap-proved by the Institutional Animal Care and Use Com-mittee (IACUC) of Brigham Young University Mice werekept in a temperature-controlled (21-22∘C) room with a12 12 hr light-dark cycle and were fed standard rodent chowand water ad libitum
23 Construction of the nBmp2NLS119905119898 Targeting Vector Thetargeting vector was constructed using ldquorecombineeringrdquoas described in [21] Briefly a genomic clone containingthe BMP2 gene was retrieved from the BAC RP23-384M14(The BACPAC Resource Center Childrenrsquos Hospital OaklandResearch Institute Oakland CA) The BMP2 bipartite NLSwas mutated by first replacing the target sequence with thegalK gene and then replacing the galK gene with the desiredmutation to alter the protein sequence of theNLS (KREKRQ-AKHKQRKRLKwas changed toKREKRQAKHKQAAALK)as described in [22] Specifically the nucleotide sequence 51015840-CGGAAGCGC (coding forRKR) was replacedwith 51015840-GCGGCG GCC (coding for AAA)
The mutant nBMP2 gene was then retrieved from theBAC into pBluescript modified to contain a thymidinekinase gene (MC1TK) A male germline self-excising neocassette (tACN) was also inserted into the targeting constructbetween the BMP2 homology arms [23]
24 Generation of the nBmp2NLS119905119898 Mouse ThenBmp2NLStm targeting vector was linearized and electro-porated into 129Bl6 G4 ES cells (generously provided byAndras Nagy of the Samuel Lunenfeld Research Institute inToronto Canada) and targeted cell lines were subjected topositivenegative selection as described by Mansour et al[24 25] Cells were analyzed by Southern blot to determinethe presence of the targeted nBMP2-mut allele Two targetedcell lines were then microinjected into Bl6 host blasto-cysts and both gave rise to chimeras Chimeric males wereintercrossed with Bl6 females to generate nBmp2NLStm+minusmice Heterozygous mice were intercrossed to produce themale minusminus (mutant) and ++ (control) mice utilized in thisstudy Successful self-excision of the neo cassette (tACN) wasverified by PCR using primers that annealed to the BMP2gene regions bracketing the neo cassette (forward primerCCTGCAGCAAGAACAAAGCAGG reverse primerCCCCAACCTTGTCATCATTCACC) PCR product sizefrom wild type was 525 bp after insertion of the targetingvector and self-excision of the neo cassette it was 607 bp
25 Genotyping To distinguish the genotypes of progenyfromheterozygous intercrosses we performed two PCR reac-tions on DNA samples from each offspring One reactionemployed a primer set that detected the presence of the wildtype allele whereas the second identified the nBmp2NLStmallele Primers sets for both reactions used the same forwardprimer but the reverse primer for the wild type reactionbound only the wild type DNA sequence and the reverseprimer for the mutant reaction bound only the mutant DNAsequence DNA from wild type mice yielded the 201 bp pro-duct only in the wild type reaction DNA from homozygous
BioMed Research International 3
mutants yielded the same size product only in the mutantreaction and DNA from heterozygotes yielded the prod-uct in both reactions Sequences for the primers were asfollows forward primer GGCCCATTTAGAGGAGAA-CC wild type reverse primer TTGCAGCTGGACTTG-AGGCGCTTCCG mutant reverse primer TTGCAGCTG-GACTTGAGGGCCGCCGC The 9 bp sequence that wasaltered to convert RKR to AAA is underlined Reactionconditions are 94∘C 5min 94∘C 20 sec 615∘C 30 sec and68∘C 40 sec repeated 30x
26 Skeletal Morphometric Analysis Two wild type andthree homozygous mutant male mice 9-10 weeks old wereeuthanized by CO
2inhalation and eviscerated soft tissues
were removed in 1KOH and skeletons were stained in a0004Alizarin red in 1KOH solution and then cleared in agraded glycerin series (20 50 80 and 100)The lengthand the proximal distal and midshaft widths at both thewidest and narrowest diameters of the femur tibia humerusand radius both left and right limbs were measured usingdigital calipers Measurements were also taken of the scapulapelvis sternum rib cage skull and vertebral column
27 GripStrength Test Male mice five homozygous wildtypes and five homozygous mutants were analyzed usingthe gripstrength test at 206 plusmn 03 weeks (wild type) and207plusmn03weeks (mutant) of age for the firstmeasurement andmeasurements were repeated once a week for six weeks Eachmouse was placed on a wire mesh cage lid which was theninverted over the open cage filled with bedding and mousewas held suspended approximately 25 cm above the bedding[26] Holding time was recorded up to a maximum of 3min
28 Western Analysis of Muscle BMP2 To compare produc-tion of conventional BMP2 in wild type versus nBmp2NLStmmouse skeletal muscle cytoplasmic extracts were preparedfrom gastrocnemius and quadriceps muscle by homogeniz-ing tissue in ice-cold buffer (025M sucrose 10mMNaCl3mMMgCl
2 1 mMDTT 1mMPMSF and a protease inhi-
bitor cocktail) centrifuging for 5min at 500timesg and col-lecting supernatant Protein concentration was quantifiedby Bradford assay before western blot analysis using BMP2primary antibody N-14 from Santa Cruz (product sc-6895)Autoradiograms were scanned and bands quantified usingAlphaEase software
29 Immunohistochemistry Gastrocnemius muscle from 6-month-old male wild type and mutant mice was isolatedembedded in paraffin and cross-sectioned at 6 120583 thick-ness After deparaffinization and rehydration sections werestained with a biotin-tagged primary antibody NBP1-19751B(Novus Biologicals) Stainingwas visualized using the LSAB+System-HRPkit fromDakowith a hematoxylin counterstainSections were viewed and imaged using a Zeiss Imager A1microscope with an AxioCam HRC camera and AxioVision47 imaging software
210 In Situ Muscle Preparation Male mice were used in themuscle stimulation experiments at 509plusmn04 weeks (control)
and 439 plusmn 75 weeks (mutant) of age Muscle preparationwas similar to that described previously [27ndash29] Briefly micewere anesthetized with 70mgkg intraperitoneal injectionof sodium pentobarbital The hamstrings were cut awayfrom the gastrocnemius plantaris and soleus (GPS) musclecomplex and the femur was secured on both the medial andlateral sides of the knee by two 16-gauge pins to preventmovement The foot was also clamped to the platform toeliminate movement of the lower leg The Achilles tendonwas then secured to a Grass Force-Displacement TransducerFT03 level arm with a calibrated tension of 700 grams Thesciatic nerve was exposed tied off and cut An electrode wasplaced directly on the sciatic nerve to achieve stimulationAnimals were supported with 100 oxygen directly to thenose throughout the procedure
211 In Situ Isometric Contractions TheGPSmuscle complexwas stimulated via electrical stimulation (2-3V stimulation005ms square wave at a frequency of 150Hz with the useof a Grass S88X Stimulator) Both a force frequency analysisand a 2Hz twitch contraction protocol were used A forcefrequency curve was developed by stimulating the muscle forten pulses at varying frequencies (10 20 40 60 80 100 120and 140 pulsessec)Thepercent ofmaximal tetanic force pro-duction was determined over all frequencies tested In orderto examine the capacity for sustained muscle contractionstwitch contractions at the rate of 2 contractionssec for 10minutes were elicited
212 Contractile Function The GPS complex data was ana-lyzed using LabScribe2 by iWorx which captured data at1000Hz The muscle was stretched to achieve maximal forceas previously described [28] Several tetanic contractionswere elicited to stretch the muscle to the length that createdmaximal force Peak twitch force was evaluated as well asone-half relaxation times every 60 pulses for 10 minutesRelaxation was determined by measuring the time requiredfor the GPS muscle to relax to 50 of the peak force forindividual contractions
213 SERCA Enzyme Activity Assay
2131 SR Membrane Purification Sarcoplasmic reticulum(SR) membranes were prepared and purified as describedby Kosk-Kosicka with minor adaptations for small musclesamples [30] All steps were performed at 4∘C unless oth-erwise specified Skeletal muscle was removed from limbsand washed in 01mMEDTA pH 70 and then homogenizedin Solution 1 (10mMMOPS 10 sucrose 01mMEDTA pH70) and pH was adjusted to between 65 and 70 using 10NaOH Samples were centrifuged at 15000timesg for 20minSupernatant was collected and filtered through one layer ofgauze then centrifuged at 40000timesg for 90min The pelletwas resuspended in Solution 2 (10mMMOPS 06MKClpH 70) and allowed to incubate at room temperature for40min Again the preparation was centrifuged at 15000timesgfor 20min and the supernatant was collected and cen-trifuged at 40000timesg for 90min The resulting pellet wasresuspended in 1mL of Solution 3 (10mMMOPS 30
4 BioMed Research International
sucrose pH 70) Protein concentrations were determinedusing a standard Bradford protein assay and preparationswere quick-frozen in liquid nitrogen for short-term stor-age
2132 Measurement of SERCA Activity SERCA activitywas measured at both 15 and 30min reaction time pointsfor every sample and both Ca2+ dependent (assay buffer50mMTris-maleate pH 74 8mMMgCl
2 120mMKCl
1mMEGTA 10 120583Mionophore A23187 1008mMCaCl2to
yield 175 120583Mfree Ca2+) and Ca2+ independent (assay buffer50mMTris-maleate pH 74 8mMMgCl
2 120mMKCl
1mMEGTA 10 120583Mionophore A23187) reactions were per-formed as described by Kosk-Kosicka [30] Briefly 02 120583gof SR membrane preparation was added to each tube andenough water was added to give a final volume of 10 120583L85 120583L of the appropriate membrane assay buffer was thenadded to each tube Reactions were started 15 sec after thebuffer was added by adding 5 120583L 60mMATP and reactionswere capped vortexed and placed in a 37∘C water bathReactions were stopped precisely 15 or 30min after ATP wasadded by adding 300 120583L Lin Morales Reagent (see [30] forreagent composition) and vortexing Results were measuredas absorbance at a wavelength of 350 nm precisely 30 sec afterthe addition of Lin Morales Reagent The Ca2+ independentand dependent reactions were performed so that the baseATPase activity (Ca2+ independent) could be subtracted fromCa2+ dependent ATPase activity Results of mutant werenormalized to wild type SERCA activity
214 Data Analysis Analyses of data from gripstrengthtests morphometric measurements in situ isometric con-tractions contractile function and SERCA enzyme activ-ity experiments were performed using two-tailed unpairedStudentrsquos 119905-tests assuming unequal variance Significance wasset at 119875 lt 005
3 Results
31 nBmp2NLS119905119898Mouse Construction Mouse embryos lack-ing all BMP2 activity have been previously described [8]These embryos died at approximately 7 days of developmentdue to defects in the formation of the chorion and amnion [8]In order to examine the effects of nBMP2 inactivation sepa-rately from the complete BMP2 knockout it was necessary todevise a mutation scheme that would leave the conventionalsecreted form of BMP2 intact while preventing the functionof nBMP2 We previously demonstrated in tissue culturethat mutating the alternative start codon from which nBMP2translation initiates only results in a 50 reduction of nBMP2relative to controls suggesting that other alternative start sitescan be used to generate forms of BMP2 that localize to thenucleus if the primary alternative start site is mutated [20]Mutation of the alternative start site therefore was unlikelyto abolish nBMP2 in a mouse model
Instead we made specific alterations to the portion of thenBMP2 gene that encodes the bipartite NLS whose altera-tions were predicted to block nuclear translocation of nBMP2yet still allow synthesis and secretion of conventional BMP2
KREKRQAKHKQRKRLK
(a)
20kDa
wtBmp2HA RKRmBmp2HA
(b)
Figure 1 The RKR to AAA mutation in the bipartite NLS doesnot inhibit secretion of the conventional BMP2 growth factor (a)Detail of the nBMP2 NLS sequence Critical amino acids in thebipartite NLS are bold and underlined the furin recognition site isboxed and the site of cleavage to release the mature secreted growthfactor from the propeptide is marked by an arrow (b) Culturemedia from cells transfected with either wild type (wtBmp2HA) ormutant (RKRmBmp2HA) BMP2 expression plasmids was collectedand analyzed by western blot The HA-tagged growth factors weresecreted equally
A consensus bipartite NLS is characterized by the followingpattern two basic residues approximately 10 spacer residuesand another basic region consisting of 4 basic residues outof five The sequence of the nBMP2 NLS is shown in Figure1(a)The upstreamKR portion of this NLS sequence overlapstheR-X-(KR)-R furin recognition sequencewhere the BMP2proprotein is cleaved to release the mature growth factorand mutation of the KR would thus disrupt production ofthe secreted growth factor (Figure 1(a)) The downstreambasic RKRLK portion of the BMP2 NLS however does notaffect propeptide cleavage To determine whether mutationof this portion of the NLS was sufficient to prevent nuclearlocalization we constructed a mutant BMP2GFP expres-sion plasmid (called RKRmBmp2GFP) in which RKR wasreplaced with AAA and transfected it into cultured cellsThis mutation eliminated nuclear localization of the GFP-tagged BMP2 in cultured RCS cells just like the previouslydescribed KR RKR to AA AAAmutation did [20] To deter-mine whether BMP2 growth factor containing this mutationcould still be secreted anHA-tagged RKRmBmp2 expressionvector (RKRmBmp2HA) was transfected into cultured RCScells and culture medium was collected 48 hrs later HA-tagged proteins were immunoprecipitated from the mediumand visualized by immunoblotting The medium from cellstransfected with RKRmBmp2HA contained as much HA-tagged BMP2 growth factor as medium from cells transfectedwith the wild type wtBmp2HA plasmid indicating that theRKR to AAA did not disrupt synthesis or secretion of theconventional BMP2 growth factor (Figure 1(b))
Having demonstrated in cell culture that theRKR toAAAmutation blocked nuclear localization of nBMP2 while stillallowing secretion of normal quantities of the conventionallyprocessed and secreted BMP2 growth factor we constructeda targeting vector and generated nBmp2NLStm mutant micebearing theRKR toAAAmutation (Figure 2) Both heterozy-gous and homozygous mutant mice appeared morphologi-cally normal and were fertile
BioMed Research International 5
Targeting vector AAA
AAA
DrdI
E1 E2 E3
Wild-type allele
E1
E1
E2
E2
E3
E3
DrdI
DrdI
DrdI
DrdIDrdI
138 kbTargeted allele
61 kb
CreNeo
CreNeo
TK
(a)
138 kb wt allele
61 kb targeted allele
(b)
1000
750
500
++ minusminus
(bp)
(c)
0
02
04
06
08
1
12
14
Gastrocnemius Quadriceps
Wild typeMutant
(d)
Figure 2 Integration of the nBmp2NLStm targeting vector (a) Schematic of the nBmp2NLStm targeting vector recombining into genomicDNA The BMP2 exons are numbered and represented as black boxes while the neo cassette and thymidine kinase (TK) gene arerepresented as grey boxes Insertion of the neo cassette introduces a DrdI restriction site as indicated The mutation of the bipartite NLS(KREKRQAKHKQRKRLKS to KREKRQAKHKQAAALKS) in exon 3 is denoted byAAA (b)After aDrdI digest and Southern hybridizationusing a 31015840 flanking probe complementary to the region indicated with the striped box the wild type band was 138 kb and the targeted alleleband was 61 kb (c) To verify male germline self-excision of the neo cassette PCR was performed using primers that bound to BMP2 outsidetheneo cassette PCRproduced a 525 bp band fromwild typeDNAand a 607 bp if the targeting vectorwas inserted and theneo cassette excisedleaving a LoxP sequence (d) Expression levels of conventional BMP2weremeasured in skeletalmuscle bywestern blot of cytoplasmic extractsusing an anti-BMP2 antibody The bands representing BMP2 proprotein precursor of secreted BMP2 were quantified digitally and mutantswere normalized to wild types within each experiment before averaging between experiments 119899 = 4 for wild type gastrocnemius 119899 = 5 formutant gastrocnemius and 119899 = 3 for both wild type and mutant quadriceps
32 Verification of Normal Secreted BMP2 Function innBmp2NLS119905119898 Mice Work by others predicted that the RKRto AAA mutation would produce secreted BMP2 that wasstill able to bind its receptors activate downstream genesand induce ectopic bone formation but that it might haveincreased diffusion range through the extracellular matrix[31 32] Increased diffusion range could disrupt embryonicpatterning but the normal appearance of the mutant miceruled out major patterning abnormalities Any minor pat-terning abnormalities would likely be manifest in the skele-ton since secreted BMP2 plays an important role in skeletaldevelopment and limb patterning Skeletal preparations ofwild type and homozygous mutant mice were performedand digital caliper measurements were taken of limbs
pelvis scapula ribcage skull and vertebrae with multiplemeasurements at various positions on each bone No sig-nificant differences were detected between wild type andmutant mice in any of the bones measured supportingthe conclusion that the function of secreted BMP2 is notimpaired in nBmp2NLStm mice (see Supplemental Table 1 inSupplementtary Materials available online at httpdxdoi)org1011552013125492)
33 Upside-Down Hanging Ability Is Decreased innBmp2NLS119905119898 Mice Because nBmp2NLStm mice appearedphenotypically normal a series of preliminary tests wereperformed to detect subtle phenotypic changes In oneevaluation five wild type and five mutant mice were tested
6 BioMed Research International
0
50
100
150
200
1 2 3 4 5 6
Hol
d tim
e (s)
+minus
SE
Weeks
Wild typeMutant
lowast lowast
Figure 3 The nBmp2NLStm mice show impaired performance inthe gripstrength test The length of time mice could hold to theunderside of a wire mesh was measured once a week for six weeksin wild type and nBmp2NLStm mice Data are presented as mean plusmnSEM and lowastdenotes 119875 lt 005 119899 = 5 per group
once a week for six weeks to measure the length of time theycould cling to the underside of a wire mesh cage lid [26] Onaverage wild type mice held on 2-3 times as long as mutantsdid at every time point and the difference was significant(119875 lt 005) at weeks 3 and 4 (Figure 3) This trend suggestedthat we should consider the possibility of a neuromusculardefect in nBmp2NLStm mice [26]
34 nBMP2 Is Detectable in Myonuclei of Wild Type but NotnBmp2NLS119905119898 Mice To examine the expression of nBMP2and BMP2 in skeletal muscle gastrocnemius muscle fromwild type and mutant mice was paraffin embedded cross-sectioned and stained using a biotin-tagged primary anti-body against BMP2 (NBP1-19751G from Novus) and a Dakovisualization system The myotubes in skeletal muscle arelong multinucleated cells with the myonuclei positionedagainst the outer edges of the myotube Brown BMP2 stain-ing of myonuclei was detectable in wild type but notnBmp2NLStm muscle whereas nonnuclear BMP2 stainingwas detectable in both wild type and mutant This resultvalidated the effectiveness of the targeted NLS-inactivatingmutation in a whole animal and it also demonstrated thatnBMP2 is expressed in skeletal muscle of control animals(Figure 4)
The presence of extranuclear BMP2 staining in bothwild type and mutant skeletal muscle suggested that theexpression of conventional BMP2 was not inhibited bythe nBmp2NLStm mutation To further examine expressionlevels of the conventional protein cytoplasmic extracts wereprepared from gastrocnemius and quadriceps muscles andexamined by western blot The BMP2 proprotein precursorof secreted BMP2 is detectable as a 50 kDa band Levels of theproprotein were not different between wild type and mutantagain suggesting unaltered expression of conventional BMP2(Figure 2(d))
Table 1 Body weight age and muscle force characteristics
Control (119899 = 7) nBmp2NLStm (119899 = 7)
Age (weeks) 509 plusmn 04 439 plusmn 75
Body weight (g) 369 plusmn 11 353 plusmn 24
GPS muscle weight (mg) 1609 plusmn 96 1717 plusmn 43
Peak tetanic tension (g) 498 plusmn 30 567 plusmn 18
Peak tetanic tension(gmg muscle) 309 plusmn 005 331 plusmn 008lowast
Peak twitch tension (g) 1926 plusmn 97 2206 plusmn 51lowast
Peak twitch tension(gmg muscle) 120 plusmn 003 129 plusmn 004
Data are presented as means plusmn SEM Where indicated lowastindicates 119875 lt 005compared to control values
35 Skeletal Muscle Relaxation Times Are Prolonged innBmp2NLS119905119898 Mice To examine the possibility of a neuro-muscular defect we measured muscle performance in situThe peak tetanic tension of the gastrocnemius plantarisand soleus (GPS) muscle group was slightly but significantlyhigher in nBmp2NLStm mice compared to control micewhen normalized to muscle weight (Table 1) No significantdifferences were observed in the peak twitch tension afternormalization to muscle weight (Table 1) and no significantdifferences were observed in muscle fatigability in responseto a 10 minute 2Hz twitch protocol (Figure 5(a))
A significant difference did emerge however when relax-ation kinetics were examined Half-relaxation times wereprolonged by up to 42 in the nBmp2NLStm muscle com-pared to control (Figure 5(b))This prolonged relaxation timewas apparent in averaged twitch performance traces frominitial (Figure 6(a)) 2min (Figure 6(b)) and 6min (Figure6(c)) time points in the 2Hz twitch contraction protocol
36 Force Frequency Analysis Exhibits a Shift in nBmp2NLS119905119898Mouse Skeletal Muscle Whenmuscle twitches are stimulatedat increasing frequencies in situ the contractions eventuallybecome so frequent that the muscle is unable to completeone relaxation before the next contraction begins and twitchpeak traces begin to merge The force or tension generatedby the muscle increases as twitch peaks merge until forcereaches its maximum and thereafter remains level regardlessof additional increases in the stimulation frequency Thiscould be considered the in situ version of a muscle crampSlowed relaxation after contraction widens each twitch peakcausing peaks to merge at a lower stimulation frequencyshifting the force frequency curve leftward
Because the nBmp2NLStm mice showed slowed relax-ation after contraction we performed a force frequency ana-lysis The nBmp2NLStm mutant mice generated significantlyelevated relative forces at 10 20 40 and 60Hz (119875 lt001) The data collected fit an expected sigmoidal curveand showed a significant leftward shift as determined bythe average force achieved at each frequency compared tothe control (Figure 7) This shift in the force frequency pat-tern as well as the slowed relaxation rates after twitch
BioMed Research International 7
(a) (b)
(c) (d)
(e) (f)
(g) (h)
Figure 4 nBMP2 is detectable in myonuclei of wild type but not nBmp2NLStm mice Gastrocnemius muscles were sectioned and stained byimmunohistochemistry (a)ndash(d) are wild type (e)ndash(h) are mutant (a) and (e) show sections at 40x magnification and all other panels show100x The brown BMP2 antibody stain is evident in myonuclei of wild type muscle whereas myonuclei stain pale blue in the mutant muscleArrows indicate some (but not all) of the myonuclei
8 BioMed Research International
0
02
04
06
08
1
12
0 500 1000
Initi
al te
nsio
n (
)
Time (s)
(a)
0
5
10
15
20
25
30
0 500 1000
Tim
e (m
s)
Time (s)
Wild typeMutant
lowast
(b)
Figure 5 Muscle function is altered in nBmp2NLStm mice (a) Muscle fatigue in both control and nBmp2NLStm GPS muscle complex wassimilar in response to 2Hz twitch contractions for 10 minutes (b) Muscle relaxation was significantly delayed in nBmp2NLStm musclecompared to WT control muscle beginning at 60 seconds and persisting throughout the 10 minutes in which these measurements weremade Data are presented as mean plusmn SEM and lowast denotes 119875 lt 005 119899 = 7 per group Only the first significant difference is marked but allvalues from 60 seconds on were significantly different from control
contractions suggests impairment of Ca2+ sequestration bythe sarcoplasmic reticulum (SR) after muscle contraction
37 SERCA Activity Is Reduced in nBmp2NLS119905119898 SkeletalMuscle The relaxation phase of a skeletal muscle contractionoccurs when Ca2+ is pumped in an ATP-dependent mannerfrom the cytosol back into the SR by sarcoendoplasmicreticulum Ca2+ ATPases (SERCA) Measurement of SERCAactivity in skeletal muscle revealed that SERCA activityin nBmp2NLStm mice is reduced to 81 plusmn 1 of wild typeactivity (Figure 8) This observation is consistent with theslowed relaxation rate observed in nBmp2NLStm muscle
4 Discussion
Several members of the BMP protein family are expressedin variant forms that are translocated to the nucleus ratherthan being secreted from the cell [20] A major challengein characterizing the function of these nuclear variants ofBMPs is that the nuclear protein is translated from the samemRNA transcript as the conventional secreted BMP2 growthfactor as demonstrated by our prior observation that bothnBMP2 and BMP2 are produced from a transfected wild typeBMP2 cDNA expression vector in tissue culture [20] Thisprecludes usage of several of the most common methodsfor studying gene function including RNAi quantitative RT-PCR and gene knockout whichwould all affect andor detectthe conventional secreted BMP2 growth factor as well asnBMP2 Instead we knocked in anRKR toAAAmutation inthe bipartite NLS to prevent nuclear localization of nBMP2while allowing production of secreted BMP2
The RKR to AAA mutation has minimal effect on thefunction of secreted BMPs BMP2 growth factor that was
truncated N-terminally of either the R K or R still hadthe capability to induce ectopic bone formation to thesame extent as wild type BMP2 [31] Moreover deletion ormutation of the homologous sequence in BMP4 produceda protein that was functionally indistinguishable from wildtype BMP4 in receptor binding antagonist binding and tar-get gene responsiveness [32] Animal cap conjugation exper-iments with Xenopus embryos however suggested that thesemutations reduced the ability of Bmp4 to bind heparin sulfatein the extracellular matrix with the result of increased diffu-sion range [32] Together these experiments predicted thatBMP2 bearing the same mutation (RKR toAAA) would alsofunction normally in terms of receptor binding and targetgene activation but might have an increased diffusion range
Secreted BMP2 plays a critical role in dorsal-ventralpatterning of the early embryo in limb patterning and inskeletogenesis so an increased diffusion range of the secretedgrowth factor is likely to cause morphological abnormalitiesin mice [33ndash35] The nBmp2NLStm mouse however dis-played nomorphological or patterning abnormalities and nodifferences in the length width thickness or shape of any ofthe bones measured indicating that there is no physiologi-cally significant alteration in the diffusion range of BMP2 inthese mice We conclude that the RKR to AAA mutation inthe nBmp2NLStm mouse preserved the function of the sec-reted BMP2 growth factor The nBmp2NLStm mouse there-fore is a good model for studying the molecular and physio-logical functions of nBMP2 as distinct from secreted BMP2
The nBmp2NLStm mouse showed decreased ability tocling upside-down to awiremesh cage lid and immunohisto-chemistry showed that nBMP2 is detectable in skeletalmusclemyonuclei of wild type but not mutant animals Mutantmuscle showed no increase in fatigability in situ however
BioMed Research International 9
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(a)
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(b)
Wild typeMutant
0
25
50
75
100
0 50 100 150(ms)
Initi
al p
eak
tens
ion
()
(c)
Figure 6 Slowed muscle relaxation is evident in twitch contractions The average muscle twitch contraction for both wild typeand nBmp2NLStm mice is shown for the initial contractions (a) the contraction following 2 minutes of stimulation (b) and the contractionfollowing 6 minutes of stimulation (c) Each trace represents the average of 7 contractions
suggesting that the reduced hang time is not related to simplemuscle fatigue Instead the shifted force frequency curve andslowed relaxation rates in the mutant muscle suggest thatmutant mice may be more prone to muscle cramping if con-tractions merge during intense muscle activity as they doduring high-frequency in situ stimulations Slowed musclerelaxation does lead to cramping in the human syndromeBrodymyopathy where the length of time required for pump-ing Ca2+ back into the SR after each contraction is increaseddue to mutations in the gene for SERCA1 the pump thatperforms this function in skeletal muscle [36ndash40] Brodypatients have also reported problems with their hands ldquolock-ingrdquo after making a tight fist [40]This cramping can be pain-ful is most frequent after exercise and grows worse withage The slowed relaxation rates in both Brody patientsand nBmp2NLStm mice suggest the possibility that nBMP2may function in some aspect of the SERCA1 productionacti-vation pathway
Our demonstration of reduced SERCA1 activity inthe nBmp2NLStm mouse supports this hypothesis The phe-notype of the nBmp2NLStm mouse however could also
result from the dysfunction of several other proteins becausemuscle contractionrelaxation cycles are quite complex Con-tractions occur when an action potential leads to activationof Ca2+ release channels (ryanodine receptors) in the SRmembrane Ca2+ floods into the cytoplasm and binds tro-ponin C which modulates the function of tropomyosin sothatmyosin can bind strongly to actin and cause shortening ofthe sarcomere The contraction ceases when active pumping(by SERCA1) of Ca2+ back into the SR depletes cytoplasmicCa2+ levels causing release of Ca2+ from troponin C [4142] The nBmp2NLStm phenotype could be caused by leakyryanodine receptors (RyR) in the SRmembrane whichwouldlead to prolonged elevation of Ca2+ in the cytosol even ifthe SERCA pumps were working normally An increasedaffinity of troponinC for Ca2+ could also cause slowedmusclerelaxation by increasing the level of cytosolic Ca2+ depletionthat would have to occur before Ca2+ was released fromtroponin C Ongoing work in our lab on other physiologicalpathways that are controlled by intracellular Ca2+movementincluding neuronal signaling immune system activation and
10 BioMed Research International
0
25
50
75
100
0 50 100 150
Max
forc
e (
)
Frequency of stimulations (pulsess)
MutantWild type
lowastlowast
lowast
lowast
Figure 7 Force frequency analysis of control and nBmp2NLStmmuscle A significant shift in the force frequency curve was observedin nBmp2NLStm muscle compared to wild type supporting theevidence for slowed relaxation in mutants Data are presented asmean plusmn SEM and lowastdenotes 119875 lt 001 119899 = 6 per group
0
02
04
06
08
1
12
Wild type Mutant
Rela
tive S
ERCA
activ
ity
Figure 8 SERCA activity is reduced in nBmp2NLStm muscle com-pared to controls SR was isolated from control and mutant muscleand Ca2+ ATPase activity wasmeasured Data are presented asmeanplusmn SEM 119875 lt 005 119899 = 3 per group
cell cycle regulation indicates that these pathways are alsodisrupted in nBmp2NLStm mice We predict therefore thatnBMP2 affects aspects of intracellular Ca2+ transport that areshared among multiple pathways rather than those (such astroponin C) that are unique to skeletal muscle
5 Conclusions
In summary the muscle phenotype that we identified inthe nBmp2NLStm mouse suggests that a significant alter-ation in intracellular Ca2+ transport resulted from impedednuclear localization of nBMP2 The prolonged relaxationkinetics the shift in the force frequency curve and the
reduction in SERCA activity are most consistent with acondition of delayed Ca2+ uptake by the SR although otherpossibilities have not been ruled out We have recentlyobserved (unpublished data) that nBmp2NLStm mice alsohave cognitive deficits that are consistent with an intra-cellular Ca2+-handling defect in hippocampus It is likelythat Ca2+ handling is disrupted in other tissues as welland that nBMP2 will be found to function in multiplemolecular pathways that govern intracellular Ca2+ transportThe nBmp2NLStm mouse constitutes a valuable model forcharacterizing the functions of nBMP2 relative to Ca2+transport pathways and for distinguishing those functionsfrom the activities of secreted BMP2
Conflict of Interests
None of the authors have any conflict of interests or compet-ing interests
Acknowledgments
This work was supported by NIAMSNIH Grant AR48839to Laura C Bridgewater Jaime L Mayo was supported by agraduate fellowship from the BrighamYoungUniversity Can-cer ResearchCenterThanks are due toDrMario R Capecchiof the University of Utah for generously providing tech-nical assistance with the stem cell electroporation blastulamicroinjections and embryo transfer steps in production ofthe nBmp2NLStm mouse Recombineering plasmids pGalKpL452 and pL253 were obtained from Neal Copeland andNancy Jenkins of the Methodist Hospital Research Institutein Houston TX
References
[1] M R Urist ldquoBone formation by autoinductionrdquo Science vol150 no 698 pp 893ndash899 1965
[2] J M Wozney V Rosen A J Celeste et al ldquoNovel regulators ofbone formation molecular clones and activitiesrdquo Science vol242 no 4885 pp 1528ndash1534 1988
[3] D ChenM Zhao andG RMundy ldquoBonemorphogenetic pro-teinsrdquo Growth Factors vol 22 no 4 pp 233ndash241 2004
[4] A M Goldstein K C Brewer A M Doyle N Nagy and DJ Roberts ldquoBMP signaling is necessary for neural crest cellmigration and ganglion formation in the enteric nervous sys-temrdquo Mechanisms of Development vol 122 no 6 pp 821ndash8332005
[5] B L Hogan ldquoBone morphogenetic proteins multifunctionalregulators of vertebrate developmentrdquo Genes and Developmentvol 10 no 13 pp 1580ndash1594 1996
[6] YMishina ldquoFunction of bonemorphogenetic protein signalingduring mouse developmentrdquo Frontiers in Bioscience vol 8 ppd855ndashd869 2003
[7] T Schlange H-H Arnold and T Brand ldquoBMP2 is a positiveregulator of Nodal signaling during left-right axis formation inthe chicken embryordquo Development vol 129 no 14 pp 3421ndash3429 2002
[8] H Zhang and A Bradley ldquoMice deficient for BMP2 arenonviable and have defects in amnionchorion and cardiac
BioMed Research International 11
developmentrdquo Development vol 122 no 10 pp 2977ndash29861996
[9] C Kawamura M Kizaki and Y Ikeda ldquoBone morphogeneticprotein (BMP)-2 induces apoptosis in human myeloma cellsrdquoLeukemia and Lymphoma vol 43 no 3 pp 635ndash639 2002
[10] M Raida J H Clement K Ameri C Han R D Leek and A LHarris ldquoExpression of bone morphogenetic protein 2 in breastcancer cells inhibits hypoxic cell deathrdquo International Journal ofOncology vol 26 no 6 pp 1465ndash1470 2005
[11] Y Du and H Yip ldquoEffects of bone morphogenetic protein 2on Id expression and neuroblastoma cell differentiationrdquoDiffer-entiation vol 79 no 2 pp 84ndash92 2010
[12] J-C GuimondM Levesque P-LMichaud et al ldquoBMP-2 func-tions independently of SHH signaling and triggers cell con-densation and apoptosis in regenerating axolotl limbsrdquo BMCDevelopmental Biology vol 10 article 15 2010
[13] C C Rider and B Mulloy ldquoBone morphogenetic protein andgrowth differentiation factor cytokine families and their proteinantagonistsrdquo Biochemical Journal vol 429 no 1 pp 1ndash12 2010
[14] D B Constam and E J Robertson ldquoRegulation of bone mor-phogenetic protein activity by pro domains and proprotein con-vertasesrdquo Journal of Cell Biology vol 144 no 1 pp 139ndash149 1999
[15] J M Wozney ldquoBone morphogenetic proteinsrdquo Progress inGrowth Factor Research vol 1 no 4 pp 267ndash280 1989
[16] A Hiyama S S Gogate S Gajghate J Mochida I M Shapiroand M V Risbud ldquoBMP-2 and TGF-120573 stimulate expression of12057313-glucuronosyl transferase 1 (GlcAT-1) in nucleus pulposuscells through AP1 TonEBP and Sp1 role of MAPKsrdquo Journal ofBone and Mineral Research vol 25 no 5 pp 1179ndash1190 2010
[17] CAlarcon A-I ZaromytidouQXi et al ldquoNuclearCDKs driveSmad transcriptional activation and turnover in BMP andTGF-120573 pathwaysrdquo Cell vol 139 no 4 pp 757ndash769 2009
[18] J Lemonnier C Ghayor J Guicheux and J Caverzasio ldquoPro-tein kinase C-independent activation of protein kinase D isinvolved in BMP-2-induced activation of stress mitogen-acti-vated protein kinases JNK and p38 and osteoblastic cell differ-entiationrdquo Journal of Biological Chemistry vol 279 no 1 pp259ndash264 2004
[19] A Moustakas and C-H Heldin ldquoFrommono- to oligo-Smadsthe heart of thematter inTGF-120573 signal transductionrdquoGenes andDevelopment vol 16 no 15 pp 1867ndash1871 2002
[20] J E Felin J L Mayo T J Loos et al ldquoNuclear variants of bonemorphogenetic proteinsrdquo BMC Cell Biology vol 11 no 1 article20 2010
[21] N G Copeland N A Jenkins and D L Court ldquoRecombi-neering a powerful new tool for mouse functional genomicsrdquoNature Reviews Genetics vol 2 no 10 pp 769ndash779 2001
[22] S Warming N Costantino D L Court N A Jenkins and NG Copeland ldquoSimple and highly efficient BAC recombineeringusing galK selectionrdquo Nucleic Acids Research vol 33 no 4article e36 2005
[23] M Bunting K E Bernstein J M Greer M R Capecchi and KR Thomas ldquoTargeting genes for self-excision in the germ linerdquoGenes and Development vol 13 no 12 pp 1524ndash1528 1999
[24] S H George M Gertsenstein K Vintersten et al ldquoDevelop-mental and adult phenotyping directly frommutant embryonicstemcellsrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol 104 no 11 pp 4455ndash4460 2007
[25] S L Mansour K R Thomas and M R Capecchi ldquoDisruptionof the proto-oncogene int-2 in mouse embryo-derived stemcells a general strategy for targeting mutations to non-select-able genesrdquo Nature vol 336 no 6197 pp 348ndash352 1988
[26] V E Papaioannou and R R Behringer Mouse PhenotypesmdashAHandbook of Mutation Analysis Cold Spring Harbor Labora-tory Press Cold Spring Harbor NY USA 2005
[27] C R Hancock J J Brault RWWiseman R L Terjung and RA Meyer ldquo31P-NMR observation of free ADP during fatiguingrepetitive contractions of murine skeletal muscle lacking AK1rdquoThe American Journal of Physiology vol 288 no 6 pp C1298ndashC1304 2005
[28] C R Hancock E Janssen and R L Terjung ldquoSkeletal musclecontractile performance and ADP accumulation in adenylatekinase-deficient micerdquoThe American Journal of Physiology vol288 no 6 pp C1287ndashC1297 2005
[29] C R Hancock E Janssen and R L Terjung ldquoContraction-mediated phosphorylation of AMPK is lower in skeletal muscleof adenylate kinase-deficient micerdquo Journal of Applied Physiol-ogy vol 100 no 2 pp 406ndash413 2006
[30] D Kosk-Kosicka ldquoCalcium signaling protocolsrdquo in Methods inMolecular Biology D G Lambert Ed vol 114 pp 343ndash354Springer New York NY USA 1999
[31] F Hillger G Herr R Rudolph and E Schwarz ldquoBiophysicalcomparison of BMP-2 ProBMP-2 and the free pro-peptidereveals stabilization of the pro-peptide by the mature growthfactorrdquo Journal of Biological Chemistry vol 280 no 15 pp14974ndash14980 2005
[32] BOhkawara S Iemura P tenDijke andNUeno ldquoAction rangeof BMP is defined by its N-terminal basic amino acid corerdquoCurrent Biology vol 12 no 3 pp 205ndash209 2002
[33] E M de Robertis ldquoSpemannrsquos organizer and the self-regulationof embryonic fieldsrdquo Mechanisms of Development vol 126 no11-12 pp 925ndash941 2009
[34] D M Maatouk K-S Choi C M Bouldin and B D Harfe ldquoInthe limb AER Bmp2 and Bmp4 are required for dorsal-ventralpatterning and interdigital cell death but not limb outgrowthrdquoDevelopmental Biology vol 327 no 2 pp 516ndash523 2009
[35] B Robert ldquoBone morphogenetic protein signaling in limboutgrowth and patterningrdquo Development Growth and Differen-tiation vol 49 no 6 pp 455ndash468 2007
[36] A Odermatt K Barton V K Khanna et al ldquoThe mutation ofPro789 to Leu reduces the activity of the fast-twitch skeletalmuscle sarco(endo)plasmic reticulum Ca2+ ATPase (SERCA1)and is associated with Brody diseaserdquoHuman Genetics vol 106no 5 pp 482ndash491 2000
[37] A Odermatt P E Taschner V K Khanna et al ldquoMutationsin the gene-encoding SERCA1 the fast-twitch skeletal musclesarcoplasmic reticulumCa2+ ATPase are associated with Brodydiseaserdquo Nature Genetics vol 14 no 2 pp 191ndash194 1996
[38] A A Benders J H Veerkamp A Oosterhof et al ldquoCa2+ home-ostasis in Brodyrsquos disease A study in skeletal muscle and cul-tured muscle cells and the effects of dantrolene and verapamilrdquoJournal of Clinical Investigation vol 94 no 2 pp 741ndash748 1994
[39] I A Brody ldquoMuscle contracture induced by exercise Asyndrome attributable to decreased relaxing factorrdquo The NewEngland Journal of Medicine vol 281 no 4 pp 187ndash192 1969
[40] Y Zhang J Fujii M S Phillips et al ldquoCharacterization ofcDNA and genomic DNA encoding SERCA1 the Ca2+-ATPaseof human fast-twitch skeletal muscle sarcoplasmic reticulumand its elimination as a candidate gene for Brody diseaserdquoGenomics vol 30 no 3 pp 415ndash424 1995
[41] M Endo ldquoCalcium-induced calcium release in skeletal musclerdquoPhysiological Reviews vol 89 no 4 pp 1153ndash1176 2009
[42] M J Betzenhauser and A RMarks ldquoRyanodine receptor chan-nelopathiesrdquo Pflugers Archiv vol 460 no 2 pp 467ndash480 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013

BioMed Research International 3
mutants yielded the same size product only in the mutantreaction and DNA from heterozygotes yielded the prod-uct in both reactions Sequences for the primers were asfollows forward primer GGCCCATTTAGAGGAGAA-CC wild type reverse primer TTGCAGCTGGACTTG-AGGCGCTTCCG mutant reverse primer TTGCAGCTG-GACTTGAGGGCCGCCGC The 9 bp sequence that wasaltered to convert RKR to AAA is underlined Reactionconditions are 94∘C 5min 94∘C 20 sec 615∘C 30 sec and68∘C 40 sec repeated 30x
26 Skeletal Morphometric Analysis Two wild type andthree homozygous mutant male mice 9-10 weeks old wereeuthanized by CO
2inhalation and eviscerated soft tissues
were removed in 1KOH and skeletons were stained in a0004Alizarin red in 1KOH solution and then cleared in agraded glycerin series (20 50 80 and 100)The lengthand the proximal distal and midshaft widths at both thewidest and narrowest diameters of the femur tibia humerusand radius both left and right limbs were measured usingdigital calipers Measurements were also taken of the scapulapelvis sternum rib cage skull and vertebral column
27 GripStrength Test Male mice five homozygous wildtypes and five homozygous mutants were analyzed usingthe gripstrength test at 206 plusmn 03 weeks (wild type) and207plusmn03weeks (mutant) of age for the firstmeasurement andmeasurements were repeated once a week for six weeks Eachmouse was placed on a wire mesh cage lid which was theninverted over the open cage filled with bedding and mousewas held suspended approximately 25 cm above the bedding[26] Holding time was recorded up to a maximum of 3min
28 Western Analysis of Muscle BMP2 To compare produc-tion of conventional BMP2 in wild type versus nBmp2NLStmmouse skeletal muscle cytoplasmic extracts were preparedfrom gastrocnemius and quadriceps muscle by homogeniz-ing tissue in ice-cold buffer (025M sucrose 10mMNaCl3mMMgCl
2 1 mMDTT 1mMPMSF and a protease inhi-
bitor cocktail) centrifuging for 5min at 500timesg and col-lecting supernatant Protein concentration was quantifiedby Bradford assay before western blot analysis using BMP2primary antibody N-14 from Santa Cruz (product sc-6895)Autoradiograms were scanned and bands quantified usingAlphaEase software
29 Immunohistochemistry Gastrocnemius muscle from 6-month-old male wild type and mutant mice was isolatedembedded in paraffin and cross-sectioned at 6 120583 thick-ness After deparaffinization and rehydration sections werestained with a biotin-tagged primary antibody NBP1-19751B(Novus Biologicals) Stainingwas visualized using the LSAB+System-HRPkit fromDakowith a hematoxylin counterstainSections were viewed and imaged using a Zeiss Imager A1microscope with an AxioCam HRC camera and AxioVision47 imaging software
210 In Situ Muscle Preparation Male mice were used in themuscle stimulation experiments at 509plusmn04 weeks (control)
and 439 plusmn 75 weeks (mutant) of age Muscle preparationwas similar to that described previously [27ndash29] Briefly micewere anesthetized with 70mgkg intraperitoneal injectionof sodium pentobarbital The hamstrings were cut awayfrom the gastrocnemius plantaris and soleus (GPS) musclecomplex and the femur was secured on both the medial andlateral sides of the knee by two 16-gauge pins to preventmovement The foot was also clamped to the platform toeliminate movement of the lower leg The Achilles tendonwas then secured to a Grass Force-Displacement TransducerFT03 level arm with a calibrated tension of 700 grams Thesciatic nerve was exposed tied off and cut An electrode wasplaced directly on the sciatic nerve to achieve stimulationAnimals were supported with 100 oxygen directly to thenose throughout the procedure
211 In Situ Isometric Contractions TheGPSmuscle complexwas stimulated via electrical stimulation (2-3V stimulation005ms square wave at a frequency of 150Hz with the useof a Grass S88X Stimulator) Both a force frequency analysisand a 2Hz twitch contraction protocol were used A forcefrequency curve was developed by stimulating the muscle forten pulses at varying frequencies (10 20 40 60 80 100 120and 140 pulsessec)Thepercent ofmaximal tetanic force pro-duction was determined over all frequencies tested In orderto examine the capacity for sustained muscle contractionstwitch contractions at the rate of 2 contractionssec for 10minutes were elicited
212 Contractile Function The GPS complex data was ana-lyzed using LabScribe2 by iWorx which captured data at1000Hz The muscle was stretched to achieve maximal forceas previously described [28] Several tetanic contractionswere elicited to stretch the muscle to the length that createdmaximal force Peak twitch force was evaluated as well asone-half relaxation times every 60 pulses for 10 minutesRelaxation was determined by measuring the time requiredfor the GPS muscle to relax to 50 of the peak force forindividual contractions
213 SERCA Enzyme Activity Assay
2131 SR Membrane Purification Sarcoplasmic reticulum(SR) membranes were prepared and purified as describedby Kosk-Kosicka with minor adaptations for small musclesamples [30] All steps were performed at 4∘C unless oth-erwise specified Skeletal muscle was removed from limbsand washed in 01mMEDTA pH 70 and then homogenizedin Solution 1 (10mMMOPS 10 sucrose 01mMEDTA pH70) and pH was adjusted to between 65 and 70 using 10NaOH Samples were centrifuged at 15000timesg for 20minSupernatant was collected and filtered through one layer ofgauze then centrifuged at 40000timesg for 90min The pelletwas resuspended in Solution 2 (10mMMOPS 06MKClpH 70) and allowed to incubate at room temperature for40min Again the preparation was centrifuged at 15000timesgfor 20min and the supernatant was collected and cen-trifuged at 40000timesg for 90min The resulting pellet wasresuspended in 1mL of Solution 3 (10mMMOPS 30
4 BioMed Research International
sucrose pH 70) Protein concentrations were determinedusing a standard Bradford protein assay and preparationswere quick-frozen in liquid nitrogen for short-term stor-age
2132 Measurement of SERCA Activity SERCA activitywas measured at both 15 and 30min reaction time pointsfor every sample and both Ca2+ dependent (assay buffer50mMTris-maleate pH 74 8mMMgCl
2 120mMKCl
1mMEGTA 10 120583Mionophore A23187 1008mMCaCl2to
yield 175 120583Mfree Ca2+) and Ca2+ independent (assay buffer50mMTris-maleate pH 74 8mMMgCl
2 120mMKCl
1mMEGTA 10 120583Mionophore A23187) reactions were per-formed as described by Kosk-Kosicka [30] Briefly 02 120583gof SR membrane preparation was added to each tube andenough water was added to give a final volume of 10 120583L85 120583L of the appropriate membrane assay buffer was thenadded to each tube Reactions were started 15 sec after thebuffer was added by adding 5 120583L 60mMATP and reactionswere capped vortexed and placed in a 37∘C water bathReactions were stopped precisely 15 or 30min after ATP wasadded by adding 300 120583L Lin Morales Reagent (see [30] forreagent composition) and vortexing Results were measuredas absorbance at a wavelength of 350 nm precisely 30 sec afterthe addition of Lin Morales Reagent The Ca2+ independentand dependent reactions were performed so that the baseATPase activity (Ca2+ independent) could be subtracted fromCa2+ dependent ATPase activity Results of mutant werenormalized to wild type SERCA activity
214 Data Analysis Analyses of data from gripstrengthtests morphometric measurements in situ isometric con-tractions contractile function and SERCA enzyme activ-ity experiments were performed using two-tailed unpairedStudentrsquos 119905-tests assuming unequal variance Significance wasset at 119875 lt 005
3 Results
31 nBmp2NLS119905119898Mouse Construction Mouse embryos lack-ing all BMP2 activity have been previously described [8]These embryos died at approximately 7 days of developmentdue to defects in the formation of the chorion and amnion [8]In order to examine the effects of nBMP2 inactivation sepa-rately from the complete BMP2 knockout it was necessary todevise a mutation scheme that would leave the conventionalsecreted form of BMP2 intact while preventing the functionof nBMP2 We previously demonstrated in tissue culturethat mutating the alternative start codon from which nBMP2translation initiates only results in a 50 reduction of nBMP2relative to controls suggesting that other alternative start sitescan be used to generate forms of BMP2 that localize to thenucleus if the primary alternative start site is mutated [20]Mutation of the alternative start site therefore was unlikelyto abolish nBMP2 in a mouse model
Instead we made specific alterations to the portion of thenBMP2 gene that encodes the bipartite NLS whose altera-tions were predicted to block nuclear translocation of nBMP2yet still allow synthesis and secretion of conventional BMP2
KREKRQAKHKQRKRLK
(a)
20kDa
wtBmp2HA RKRmBmp2HA
(b)
Figure 1 The RKR to AAA mutation in the bipartite NLS doesnot inhibit secretion of the conventional BMP2 growth factor (a)Detail of the nBMP2 NLS sequence Critical amino acids in thebipartite NLS are bold and underlined the furin recognition site isboxed and the site of cleavage to release the mature secreted growthfactor from the propeptide is marked by an arrow (b) Culturemedia from cells transfected with either wild type (wtBmp2HA) ormutant (RKRmBmp2HA) BMP2 expression plasmids was collectedand analyzed by western blot The HA-tagged growth factors weresecreted equally
A consensus bipartite NLS is characterized by the followingpattern two basic residues approximately 10 spacer residuesand another basic region consisting of 4 basic residues outof five The sequence of the nBMP2 NLS is shown in Figure1(a)The upstreamKR portion of this NLS sequence overlapstheR-X-(KR)-R furin recognition sequencewhere the BMP2proprotein is cleaved to release the mature growth factorand mutation of the KR would thus disrupt production ofthe secreted growth factor (Figure 1(a)) The downstreambasic RKRLK portion of the BMP2 NLS however does notaffect propeptide cleavage To determine whether mutationof this portion of the NLS was sufficient to prevent nuclearlocalization we constructed a mutant BMP2GFP expres-sion plasmid (called RKRmBmp2GFP) in which RKR wasreplaced with AAA and transfected it into cultured cellsThis mutation eliminated nuclear localization of the GFP-tagged BMP2 in cultured RCS cells just like the previouslydescribed KR RKR to AA AAAmutation did [20] To deter-mine whether BMP2 growth factor containing this mutationcould still be secreted anHA-tagged RKRmBmp2 expressionvector (RKRmBmp2HA) was transfected into cultured RCScells and culture medium was collected 48 hrs later HA-tagged proteins were immunoprecipitated from the mediumand visualized by immunoblotting The medium from cellstransfected with RKRmBmp2HA contained as much HA-tagged BMP2 growth factor as medium from cells transfectedwith the wild type wtBmp2HA plasmid indicating that theRKR to AAA did not disrupt synthesis or secretion of theconventional BMP2 growth factor (Figure 1(b))
Having demonstrated in cell culture that theRKR toAAAmutation blocked nuclear localization of nBMP2 while stillallowing secretion of normal quantities of the conventionallyprocessed and secreted BMP2 growth factor we constructeda targeting vector and generated nBmp2NLStm mutant micebearing theRKR toAAAmutation (Figure 2) Both heterozy-gous and homozygous mutant mice appeared morphologi-cally normal and were fertile
BioMed Research International 5
Targeting vector AAA
AAA
DrdI
E1 E2 E3
Wild-type allele
E1
E1
E2
E2
E3
E3
DrdI
DrdI
DrdI
DrdIDrdI
138 kbTargeted allele
61 kb
CreNeo
CreNeo
TK
(a)
138 kb wt allele
61 kb targeted allele
(b)
1000
750
500
++ minusminus
(bp)
(c)
0
02
04
06
08
1
12
14
Gastrocnemius Quadriceps
Wild typeMutant
(d)
Figure 2 Integration of the nBmp2NLStm targeting vector (a) Schematic of the nBmp2NLStm targeting vector recombining into genomicDNA The BMP2 exons are numbered and represented as black boxes while the neo cassette and thymidine kinase (TK) gene arerepresented as grey boxes Insertion of the neo cassette introduces a DrdI restriction site as indicated The mutation of the bipartite NLS(KREKRQAKHKQRKRLKS to KREKRQAKHKQAAALKS) in exon 3 is denoted byAAA (b)After aDrdI digest and Southern hybridizationusing a 31015840 flanking probe complementary to the region indicated with the striped box the wild type band was 138 kb and the targeted alleleband was 61 kb (c) To verify male germline self-excision of the neo cassette PCR was performed using primers that bound to BMP2 outsidetheneo cassette PCRproduced a 525 bp band fromwild typeDNAand a 607 bp if the targeting vectorwas inserted and theneo cassette excisedleaving a LoxP sequence (d) Expression levels of conventional BMP2weremeasured in skeletalmuscle bywestern blot of cytoplasmic extractsusing an anti-BMP2 antibody The bands representing BMP2 proprotein precursor of secreted BMP2 were quantified digitally and mutantswere normalized to wild types within each experiment before averaging between experiments 119899 = 4 for wild type gastrocnemius 119899 = 5 formutant gastrocnemius and 119899 = 3 for both wild type and mutant quadriceps
32 Verification of Normal Secreted BMP2 Function innBmp2NLS119905119898 Mice Work by others predicted that the RKRto AAA mutation would produce secreted BMP2 that wasstill able to bind its receptors activate downstream genesand induce ectopic bone formation but that it might haveincreased diffusion range through the extracellular matrix[31 32] Increased diffusion range could disrupt embryonicpatterning but the normal appearance of the mutant miceruled out major patterning abnormalities Any minor pat-terning abnormalities would likely be manifest in the skele-ton since secreted BMP2 plays an important role in skeletaldevelopment and limb patterning Skeletal preparations ofwild type and homozygous mutant mice were performedand digital caliper measurements were taken of limbs
pelvis scapula ribcage skull and vertebrae with multiplemeasurements at various positions on each bone No sig-nificant differences were detected between wild type andmutant mice in any of the bones measured supportingthe conclusion that the function of secreted BMP2 is notimpaired in nBmp2NLStm mice (see Supplemental Table 1 inSupplementtary Materials available online at httpdxdoi)org1011552013125492)
33 Upside-Down Hanging Ability Is Decreased innBmp2NLS119905119898 Mice Because nBmp2NLStm mice appearedphenotypically normal a series of preliminary tests wereperformed to detect subtle phenotypic changes In oneevaluation five wild type and five mutant mice were tested
6 BioMed Research International
0
50
100
150
200
1 2 3 4 5 6
Hol
d tim
e (s)
+minus
SE
Weeks
Wild typeMutant
lowast lowast
Figure 3 The nBmp2NLStm mice show impaired performance inthe gripstrength test The length of time mice could hold to theunderside of a wire mesh was measured once a week for six weeksin wild type and nBmp2NLStm mice Data are presented as mean plusmnSEM and lowastdenotes 119875 lt 005 119899 = 5 per group
once a week for six weeks to measure the length of time theycould cling to the underside of a wire mesh cage lid [26] Onaverage wild type mice held on 2-3 times as long as mutantsdid at every time point and the difference was significant(119875 lt 005) at weeks 3 and 4 (Figure 3) This trend suggestedthat we should consider the possibility of a neuromusculardefect in nBmp2NLStm mice [26]
34 nBMP2 Is Detectable in Myonuclei of Wild Type but NotnBmp2NLS119905119898 Mice To examine the expression of nBMP2and BMP2 in skeletal muscle gastrocnemius muscle fromwild type and mutant mice was paraffin embedded cross-sectioned and stained using a biotin-tagged primary anti-body against BMP2 (NBP1-19751G from Novus) and a Dakovisualization system The myotubes in skeletal muscle arelong multinucleated cells with the myonuclei positionedagainst the outer edges of the myotube Brown BMP2 stain-ing of myonuclei was detectable in wild type but notnBmp2NLStm muscle whereas nonnuclear BMP2 stainingwas detectable in both wild type and mutant This resultvalidated the effectiveness of the targeted NLS-inactivatingmutation in a whole animal and it also demonstrated thatnBMP2 is expressed in skeletal muscle of control animals(Figure 4)
The presence of extranuclear BMP2 staining in bothwild type and mutant skeletal muscle suggested that theexpression of conventional BMP2 was not inhibited bythe nBmp2NLStm mutation To further examine expressionlevels of the conventional protein cytoplasmic extracts wereprepared from gastrocnemius and quadriceps muscles andexamined by western blot The BMP2 proprotein precursorof secreted BMP2 is detectable as a 50 kDa band Levels of theproprotein were not different between wild type and mutantagain suggesting unaltered expression of conventional BMP2(Figure 2(d))
Table 1 Body weight age and muscle force characteristics
Control (119899 = 7) nBmp2NLStm (119899 = 7)
Age (weeks) 509 plusmn 04 439 plusmn 75
Body weight (g) 369 plusmn 11 353 plusmn 24
GPS muscle weight (mg) 1609 plusmn 96 1717 plusmn 43
Peak tetanic tension (g) 498 plusmn 30 567 plusmn 18
Peak tetanic tension(gmg muscle) 309 plusmn 005 331 plusmn 008lowast
Peak twitch tension (g) 1926 plusmn 97 2206 plusmn 51lowast
Peak twitch tension(gmg muscle) 120 plusmn 003 129 plusmn 004
Data are presented as means plusmn SEM Where indicated lowastindicates 119875 lt 005compared to control values
35 Skeletal Muscle Relaxation Times Are Prolonged innBmp2NLS119905119898 Mice To examine the possibility of a neuro-muscular defect we measured muscle performance in situThe peak tetanic tension of the gastrocnemius plantarisand soleus (GPS) muscle group was slightly but significantlyhigher in nBmp2NLStm mice compared to control micewhen normalized to muscle weight (Table 1) No significantdifferences were observed in the peak twitch tension afternormalization to muscle weight (Table 1) and no significantdifferences were observed in muscle fatigability in responseto a 10 minute 2Hz twitch protocol (Figure 5(a))
A significant difference did emerge however when relax-ation kinetics were examined Half-relaxation times wereprolonged by up to 42 in the nBmp2NLStm muscle com-pared to control (Figure 5(b))This prolonged relaxation timewas apparent in averaged twitch performance traces frominitial (Figure 6(a)) 2min (Figure 6(b)) and 6min (Figure6(c)) time points in the 2Hz twitch contraction protocol
36 Force Frequency Analysis Exhibits a Shift in nBmp2NLS119905119898Mouse Skeletal Muscle Whenmuscle twitches are stimulatedat increasing frequencies in situ the contractions eventuallybecome so frequent that the muscle is unable to completeone relaxation before the next contraction begins and twitchpeak traces begin to merge The force or tension generatedby the muscle increases as twitch peaks merge until forcereaches its maximum and thereafter remains level regardlessof additional increases in the stimulation frequency Thiscould be considered the in situ version of a muscle crampSlowed relaxation after contraction widens each twitch peakcausing peaks to merge at a lower stimulation frequencyshifting the force frequency curve leftward
Because the nBmp2NLStm mice showed slowed relax-ation after contraction we performed a force frequency ana-lysis The nBmp2NLStm mutant mice generated significantlyelevated relative forces at 10 20 40 and 60Hz (119875 lt001) The data collected fit an expected sigmoidal curveand showed a significant leftward shift as determined bythe average force achieved at each frequency compared tothe control (Figure 7) This shift in the force frequency pat-tern as well as the slowed relaxation rates after twitch
BioMed Research International 7
(a) (b)
(c) (d)
(e) (f)
(g) (h)
Figure 4 nBMP2 is detectable in myonuclei of wild type but not nBmp2NLStm mice Gastrocnemius muscles were sectioned and stained byimmunohistochemistry (a)ndash(d) are wild type (e)ndash(h) are mutant (a) and (e) show sections at 40x magnification and all other panels show100x The brown BMP2 antibody stain is evident in myonuclei of wild type muscle whereas myonuclei stain pale blue in the mutant muscleArrows indicate some (but not all) of the myonuclei
8 BioMed Research International
0
02
04
06
08
1
12
0 500 1000
Initi
al te
nsio
n (
)
Time (s)
(a)
0
5
10
15
20
25
30
0 500 1000
Tim
e (m
s)
Time (s)
Wild typeMutant
lowast
(b)
Figure 5 Muscle function is altered in nBmp2NLStm mice (a) Muscle fatigue in both control and nBmp2NLStm GPS muscle complex wassimilar in response to 2Hz twitch contractions for 10 minutes (b) Muscle relaxation was significantly delayed in nBmp2NLStm musclecompared to WT control muscle beginning at 60 seconds and persisting throughout the 10 minutes in which these measurements weremade Data are presented as mean plusmn SEM and lowast denotes 119875 lt 005 119899 = 7 per group Only the first significant difference is marked but allvalues from 60 seconds on were significantly different from control
contractions suggests impairment of Ca2+ sequestration bythe sarcoplasmic reticulum (SR) after muscle contraction
37 SERCA Activity Is Reduced in nBmp2NLS119905119898 SkeletalMuscle The relaxation phase of a skeletal muscle contractionoccurs when Ca2+ is pumped in an ATP-dependent mannerfrom the cytosol back into the SR by sarcoendoplasmicreticulum Ca2+ ATPases (SERCA) Measurement of SERCAactivity in skeletal muscle revealed that SERCA activityin nBmp2NLStm mice is reduced to 81 plusmn 1 of wild typeactivity (Figure 8) This observation is consistent with theslowed relaxation rate observed in nBmp2NLStm muscle
4 Discussion
Several members of the BMP protein family are expressedin variant forms that are translocated to the nucleus ratherthan being secreted from the cell [20] A major challengein characterizing the function of these nuclear variants ofBMPs is that the nuclear protein is translated from the samemRNA transcript as the conventional secreted BMP2 growthfactor as demonstrated by our prior observation that bothnBMP2 and BMP2 are produced from a transfected wild typeBMP2 cDNA expression vector in tissue culture [20] Thisprecludes usage of several of the most common methodsfor studying gene function including RNAi quantitative RT-PCR and gene knockout whichwould all affect andor detectthe conventional secreted BMP2 growth factor as well asnBMP2 Instead we knocked in anRKR toAAAmutation inthe bipartite NLS to prevent nuclear localization of nBMP2while allowing production of secreted BMP2
The RKR to AAA mutation has minimal effect on thefunction of secreted BMPs BMP2 growth factor that was
truncated N-terminally of either the R K or R still hadthe capability to induce ectopic bone formation to thesame extent as wild type BMP2 [31] Moreover deletion ormutation of the homologous sequence in BMP4 produceda protein that was functionally indistinguishable from wildtype BMP4 in receptor binding antagonist binding and tar-get gene responsiveness [32] Animal cap conjugation exper-iments with Xenopus embryos however suggested that thesemutations reduced the ability of Bmp4 to bind heparin sulfatein the extracellular matrix with the result of increased diffu-sion range [32] Together these experiments predicted thatBMP2 bearing the same mutation (RKR toAAA) would alsofunction normally in terms of receptor binding and targetgene activation but might have an increased diffusion range
Secreted BMP2 plays a critical role in dorsal-ventralpatterning of the early embryo in limb patterning and inskeletogenesis so an increased diffusion range of the secretedgrowth factor is likely to cause morphological abnormalitiesin mice [33ndash35] The nBmp2NLStm mouse however dis-played nomorphological or patterning abnormalities and nodifferences in the length width thickness or shape of any ofthe bones measured indicating that there is no physiologi-cally significant alteration in the diffusion range of BMP2 inthese mice We conclude that the RKR to AAA mutation inthe nBmp2NLStm mouse preserved the function of the sec-reted BMP2 growth factor The nBmp2NLStm mouse there-fore is a good model for studying the molecular and physio-logical functions of nBMP2 as distinct from secreted BMP2
The nBmp2NLStm mouse showed decreased ability tocling upside-down to awiremesh cage lid and immunohisto-chemistry showed that nBMP2 is detectable in skeletalmusclemyonuclei of wild type but not mutant animals Mutantmuscle showed no increase in fatigability in situ however
BioMed Research International 9
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(a)
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(b)
Wild typeMutant
0
25
50
75
100
0 50 100 150(ms)
Initi
al p
eak
tens
ion
()
(c)
Figure 6 Slowed muscle relaxation is evident in twitch contractions The average muscle twitch contraction for both wild typeand nBmp2NLStm mice is shown for the initial contractions (a) the contraction following 2 minutes of stimulation (b) and the contractionfollowing 6 minutes of stimulation (c) Each trace represents the average of 7 contractions
suggesting that the reduced hang time is not related to simplemuscle fatigue Instead the shifted force frequency curve andslowed relaxation rates in the mutant muscle suggest thatmutant mice may be more prone to muscle cramping if con-tractions merge during intense muscle activity as they doduring high-frequency in situ stimulations Slowed musclerelaxation does lead to cramping in the human syndromeBrodymyopathy where the length of time required for pump-ing Ca2+ back into the SR after each contraction is increaseddue to mutations in the gene for SERCA1 the pump thatperforms this function in skeletal muscle [36ndash40] Brodypatients have also reported problems with their hands ldquolock-ingrdquo after making a tight fist [40]This cramping can be pain-ful is most frequent after exercise and grows worse withage The slowed relaxation rates in both Brody patientsand nBmp2NLStm mice suggest the possibility that nBMP2may function in some aspect of the SERCA1 productionacti-vation pathway
Our demonstration of reduced SERCA1 activity inthe nBmp2NLStm mouse supports this hypothesis The phe-notype of the nBmp2NLStm mouse however could also
result from the dysfunction of several other proteins becausemuscle contractionrelaxation cycles are quite complex Con-tractions occur when an action potential leads to activationof Ca2+ release channels (ryanodine receptors) in the SRmembrane Ca2+ floods into the cytoplasm and binds tro-ponin C which modulates the function of tropomyosin sothatmyosin can bind strongly to actin and cause shortening ofthe sarcomere The contraction ceases when active pumping(by SERCA1) of Ca2+ back into the SR depletes cytoplasmicCa2+ levels causing release of Ca2+ from troponin C [4142] The nBmp2NLStm phenotype could be caused by leakyryanodine receptors (RyR) in the SRmembrane whichwouldlead to prolonged elevation of Ca2+ in the cytosol even ifthe SERCA pumps were working normally An increasedaffinity of troponinC for Ca2+ could also cause slowedmusclerelaxation by increasing the level of cytosolic Ca2+ depletionthat would have to occur before Ca2+ was released fromtroponin C Ongoing work in our lab on other physiologicalpathways that are controlled by intracellular Ca2+movementincluding neuronal signaling immune system activation and
10 BioMed Research International
0
25
50
75
100
0 50 100 150
Max
forc
e (
)
Frequency of stimulations (pulsess)
MutantWild type
lowastlowast
lowast
lowast
Figure 7 Force frequency analysis of control and nBmp2NLStmmuscle A significant shift in the force frequency curve was observedin nBmp2NLStm muscle compared to wild type supporting theevidence for slowed relaxation in mutants Data are presented asmean plusmn SEM and lowastdenotes 119875 lt 001 119899 = 6 per group
0
02
04
06
08
1
12
Wild type Mutant
Rela
tive S
ERCA
activ
ity
Figure 8 SERCA activity is reduced in nBmp2NLStm muscle com-pared to controls SR was isolated from control and mutant muscleand Ca2+ ATPase activity wasmeasured Data are presented asmeanplusmn SEM 119875 lt 005 119899 = 3 per group
cell cycle regulation indicates that these pathways are alsodisrupted in nBmp2NLStm mice We predict therefore thatnBMP2 affects aspects of intracellular Ca2+ transport that areshared among multiple pathways rather than those (such astroponin C) that are unique to skeletal muscle
5 Conclusions
In summary the muscle phenotype that we identified inthe nBmp2NLStm mouse suggests that a significant alter-ation in intracellular Ca2+ transport resulted from impedednuclear localization of nBMP2 The prolonged relaxationkinetics the shift in the force frequency curve and the
reduction in SERCA activity are most consistent with acondition of delayed Ca2+ uptake by the SR although otherpossibilities have not been ruled out We have recentlyobserved (unpublished data) that nBmp2NLStm mice alsohave cognitive deficits that are consistent with an intra-cellular Ca2+-handling defect in hippocampus It is likelythat Ca2+ handling is disrupted in other tissues as welland that nBMP2 will be found to function in multiplemolecular pathways that govern intracellular Ca2+ transportThe nBmp2NLStm mouse constitutes a valuable model forcharacterizing the functions of nBMP2 relative to Ca2+transport pathways and for distinguishing those functionsfrom the activities of secreted BMP2
Conflict of Interests
None of the authors have any conflict of interests or compet-ing interests
Acknowledgments
This work was supported by NIAMSNIH Grant AR48839to Laura C Bridgewater Jaime L Mayo was supported by agraduate fellowship from the BrighamYoungUniversity Can-cer ResearchCenterThanks are due toDrMario R Capecchiof the University of Utah for generously providing tech-nical assistance with the stem cell electroporation blastulamicroinjections and embryo transfer steps in production ofthe nBmp2NLStm mouse Recombineering plasmids pGalKpL452 and pL253 were obtained from Neal Copeland andNancy Jenkins of the Methodist Hospital Research Institutein Houston TX
References
[1] M R Urist ldquoBone formation by autoinductionrdquo Science vol150 no 698 pp 893ndash899 1965
[2] J M Wozney V Rosen A J Celeste et al ldquoNovel regulators ofbone formation molecular clones and activitiesrdquo Science vol242 no 4885 pp 1528ndash1534 1988
[3] D ChenM Zhao andG RMundy ldquoBonemorphogenetic pro-teinsrdquo Growth Factors vol 22 no 4 pp 233ndash241 2004
[4] A M Goldstein K C Brewer A M Doyle N Nagy and DJ Roberts ldquoBMP signaling is necessary for neural crest cellmigration and ganglion formation in the enteric nervous sys-temrdquo Mechanisms of Development vol 122 no 6 pp 821ndash8332005
[5] B L Hogan ldquoBone morphogenetic proteins multifunctionalregulators of vertebrate developmentrdquo Genes and Developmentvol 10 no 13 pp 1580ndash1594 1996
[6] YMishina ldquoFunction of bonemorphogenetic protein signalingduring mouse developmentrdquo Frontiers in Bioscience vol 8 ppd855ndashd869 2003
[7] T Schlange H-H Arnold and T Brand ldquoBMP2 is a positiveregulator of Nodal signaling during left-right axis formation inthe chicken embryordquo Development vol 129 no 14 pp 3421ndash3429 2002
[8] H Zhang and A Bradley ldquoMice deficient for BMP2 arenonviable and have defects in amnionchorion and cardiac
BioMed Research International 11
developmentrdquo Development vol 122 no 10 pp 2977ndash29861996
[9] C Kawamura M Kizaki and Y Ikeda ldquoBone morphogeneticprotein (BMP)-2 induces apoptosis in human myeloma cellsrdquoLeukemia and Lymphoma vol 43 no 3 pp 635ndash639 2002
[10] M Raida J H Clement K Ameri C Han R D Leek and A LHarris ldquoExpression of bone morphogenetic protein 2 in breastcancer cells inhibits hypoxic cell deathrdquo International Journal ofOncology vol 26 no 6 pp 1465ndash1470 2005
[11] Y Du and H Yip ldquoEffects of bone morphogenetic protein 2on Id expression and neuroblastoma cell differentiationrdquoDiffer-entiation vol 79 no 2 pp 84ndash92 2010
[12] J-C GuimondM Levesque P-LMichaud et al ldquoBMP-2 func-tions independently of SHH signaling and triggers cell con-densation and apoptosis in regenerating axolotl limbsrdquo BMCDevelopmental Biology vol 10 article 15 2010
[13] C C Rider and B Mulloy ldquoBone morphogenetic protein andgrowth differentiation factor cytokine families and their proteinantagonistsrdquo Biochemical Journal vol 429 no 1 pp 1ndash12 2010
[14] D B Constam and E J Robertson ldquoRegulation of bone mor-phogenetic protein activity by pro domains and proprotein con-vertasesrdquo Journal of Cell Biology vol 144 no 1 pp 139ndash149 1999
[15] J M Wozney ldquoBone morphogenetic proteinsrdquo Progress inGrowth Factor Research vol 1 no 4 pp 267ndash280 1989
[16] A Hiyama S S Gogate S Gajghate J Mochida I M Shapiroand M V Risbud ldquoBMP-2 and TGF-120573 stimulate expression of12057313-glucuronosyl transferase 1 (GlcAT-1) in nucleus pulposuscells through AP1 TonEBP and Sp1 role of MAPKsrdquo Journal ofBone and Mineral Research vol 25 no 5 pp 1179ndash1190 2010
[17] CAlarcon A-I ZaromytidouQXi et al ldquoNuclearCDKs driveSmad transcriptional activation and turnover in BMP andTGF-120573 pathwaysrdquo Cell vol 139 no 4 pp 757ndash769 2009
[18] J Lemonnier C Ghayor J Guicheux and J Caverzasio ldquoPro-tein kinase C-independent activation of protein kinase D isinvolved in BMP-2-induced activation of stress mitogen-acti-vated protein kinases JNK and p38 and osteoblastic cell differ-entiationrdquo Journal of Biological Chemistry vol 279 no 1 pp259ndash264 2004
[19] A Moustakas and C-H Heldin ldquoFrommono- to oligo-Smadsthe heart of thematter inTGF-120573 signal transductionrdquoGenes andDevelopment vol 16 no 15 pp 1867ndash1871 2002
[20] J E Felin J L Mayo T J Loos et al ldquoNuclear variants of bonemorphogenetic proteinsrdquo BMC Cell Biology vol 11 no 1 article20 2010
[21] N G Copeland N A Jenkins and D L Court ldquoRecombi-neering a powerful new tool for mouse functional genomicsrdquoNature Reviews Genetics vol 2 no 10 pp 769ndash779 2001
[22] S Warming N Costantino D L Court N A Jenkins and NG Copeland ldquoSimple and highly efficient BAC recombineeringusing galK selectionrdquo Nucleic Acids Research vol 33 no 4article e36 2005
[23] M Bunting K E Bernstein J M Greer M R Capecchi and KR Thomas ldquoTargeting genes for self-excision in the germ linerdquoGenes and Development vol 13 no 12 pp 1524ndash1528 1999
[24] S H George M Gertsenstein K Vintersten et al ldquoDevelop-mental and adult phenotyping directly frommutant embryonicstemcellsrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol 104 no 11 pp 4455ndash4460 2007
[25] S L Mansour K R Thomas and M R Capecchi ldquoDisruptionof the proto-oncogene int-2 in mouse embryo-derived stemcells a general strategy for targeting mutations to non-select-able genesrdquo Nature vol 336 no 6197 pp 348ndash352 1988
[26] V E Papaioannou and R R Behringer Mouse PhenotypesmdashAHandbook of Mutation Analysis Cold Spring Harbor Labora-tory Press Cold Spring Harbor NY USA 2005
[27] C R Hancock J J Brault RWWiseman R L Terjung and RA Meyer ldquo31P-NMR observation of free ADP during fatiguingrepetitive contractions of murine skeletal muscle lacking AK1rdquoThe American Journal of Physiology vol 288 no 6 pp C1298ndashC1304 2005
[28] C R Hancock E Janssen and R L Terjung ldquoSkeletal musclecontractile performance and ADP accumulation in adenylatekinase-deficient micerdquoThe American Journal of Physiology vol288 no 6 pp C1287ndashC1297 2005
[29] C R Hancock E Janssen and R L Terjung ldquoContraction-mediated phosphorylation of AMPK is lower in skeletal muscleof adenylate kinase-deficient micerdquo Journal of Applied Physiol-ogy vol 100 no 2 pp 406ndash413 2006
[30] D Kosk-Kosicka ldquoCalcium signaling protocolsrdquo in Methods inMolecular Biology D G Lambert Ed vol 114 pp 343ndash354Springer New York NY USA 1999
[31] F Hillger G Herr R Rudolph and E Schwarz ldquoBiophysicalcomparison of BMP-2 ProBMP-2 and the free pro-peptidereveals stabilization of the pro-peptide by the mature growthfactorrdquo Journal of Biological Chemistry vol 280 no 15 pp14974ndash14980 2005
[32] BOhkawara S Iemura P tenDijke andNUeno ldquoAction rangeof BMP is defined by its N-terminal basic amino acid corerdquoCurrent Biology vol 12 no 3 pp 205ndash209 2002
[33] E M de Robertis ldquoSpemannrsquos organizer and the self-regulationof embryonic fieldsrdquo Mechanisms of Development vol 126 no11-12 pp 925ndash941 2009
[34] D M Maatouk K-S Choi C M Bouldin and B D Harfe ldquoInthe limb AER Bmp2 and Bmp4 are required for dorsal-ventralpatterning and interdigital cell death but not limb outgrowthrdquoDevelopmental Biology vol 327 no 2 pp 516ndash523 2009
[35] B Robert ldquoBone morphogenetic protein signaling in limboutgrowth and patterningrdquo Development Growth and Differen-tiation vol 49 no 6 pp 455ndash468 2007
[36] A Odermatt K Barton V K Khanna et al ldquoThe mutation ofPro789 to Leu reduces the activity of the fast-twitch skeletalmuscle sarco(endo)plasmic reticulum Ca2+ ATPase (SERCA1)and is associated with Brody diseaserdquoHuman Genetics vol 106no 5 pp 482ndash491 2000
[37] A Odermatt P E Taschner V K Khanna et al ldquoMutationsin the gene-encoding SERCA1 the fast-twitch skeletal musclesarcoplasmic reticulumCa2+ ATPase are associated with Brodydiseaserdquo Nature Genetics vol 14 no 2 pp 191ndash194 1996
[38] A A Benders J H Veerkamp A Oosterhof et al ldquoCa2+ home-ostasis in Brodyrsquos disease A study in skeletal muscle and cul-tured muscle cells and the effects of dantrolene and verapamilrdquoJournal of Clinical Investigation vol 94 no 2 pp 741ndash748 1994
[39] I A Brody ldquoMuscle contracture induced by exercise Asyndrome attributable to decreased relaxing factorrdquo The NewEngland Journal of Medicine vol 281 no 4 pp 187ndash192 1969
[40] Y Zhang J Fujii M S Phillips et al ldquoCharacterization ofcDNA and genomic DNA encoding SERCA1 the Ca2+-ATPaseof human fast-twitch skeletal muscle sarcoplasmic reticulumand its elimination as a candidate gene for Brody diseaserdquoGenomics vol 30 no 3 pp 415ndash424 1995
[41] M Endo ldquoCalcium-induced calcium release in skeletal musclerdquoPhysiological Reviews vol 89 no 4 pp 1153ndash1176 2009
[42] M J Betzenhauser and A RMarks ldquoRyanodine receptor chan-nelopathiesrdquo Pflugers Archiv vol 460 no 2 pp 467ndash480 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013

4 BioMed Research International
sucrose pH 70) Protein concentrations were determinedusing a standard Bradford protein assay and preparationswere quick-frozen in liquid nitrogen for short-term stor-age
2132 Measurement of SERCA Activity SERCA activitywas measured at both 15 and 30min reaction time pointsfor every sample and both Ca2+ dependent (assay buffer50mMTris-maleate pH 74 8mMMgCl
2 120mMKCl
1mMEGTA 10 120583Mionophore A23187 1008mMCaCl2to
yield 175 120583Mfree Ca2+) and Ca2+ independent (assay buffer50mMTris-maleate pH 74 8mMMgCl
2 120mMKCl
1mMEGTA 10 120583Mionophore A23187) reactions were per-formed as described by Kosk-Kosicka [30] Briefly 02 120583gof SR membrane preparation was added to each tube andenough water was added to give a final volume of 10 120583L85 120583L of the appropriate membrane assay buffer was thenadded to each tube Reactions were started 15 sec after thebuffer was added by adding 5 120583L 60mMATP and reactionswere capped vortexed and placed in a 37∘C water bathReactions were stopped precisely 15 or 30min after ATP wasadded by adding 300 120583L Lin Morales Reagent (see [30] forreagent composition) and vortexing Results were measuredas absorbance at a wavelength of 350 nm precisely 30 sec afterthe addition of Lin Morales Reagent The Ca2+ independentand dependent reactions were performed so that the baseATPase activity (Ca2+ independent) could be subtracted fromCa2+ dependent ATPase activity Results of mutant werenormalized to wild type SERCA activity
214 Data Analysis Analyses of data from gripstrengthtests morphometric measurements in situ isometric con-tractions contractile function and SERCA enzyme activ-ity experiments were performed using two-tailed unpairedStudentrsquos 119905-tests assuming unequal variance Significance wasset at 119875 lt 005
3 Results
31 nBmp2NLS119905119898Mouse Construction Mouse embryos lack-ing all BMP2 activity have been previously described [8]These embryos died at approximately 7 days of developmentdue to defects in the formation of the chorion and amnion [8]In order to examine the effects of nBMP2 inactivation sepa-rately from the complete BMP2 knockout it was necessary todevise a mutation scheme that would leave the conventionalsecreted form of BMP2 intact while preventing the functionof nBMP2 We previously demonstrated in tissue culturethat mutating the alternative start codon from which nBMP2translation initiates only results in a 50 reduction of nBMP2relative to controls suggesting that other alternative start sitescan be used to generate forms of BMP2 that localize to thenucleus if the primary alternative start site is mutated [20]Mutation of the alternative start site therefore was unlikelyto abolish nBMP2 in a mouse model
Instead we made specific alterations to the portion of thenBMP2 gene that encodes the bipartite NLS whose altera-tions were predicted to block nuclear translocation of nBMP2yet still allow synthesis and secretion of conventional BMP2
KREKRQAKHKQRKRLK
(a)
20kDa
wtBmp2HA RKRmBmp2HA
(b)
Figure 1 The RKR to AAA mutation in the bipartite NLS doesnot inhibit secretion of the conventional BMP2 growth factor (a)Detail of the nBMP2 NLS sequence Critical amino acids in thebipartite NLS are bold and underlined the furin recognition site isboxed and the site of cleavage to release the mature secreted growthfactor from the propeptide is marked by an arrow (b) Culturemedia from cells transfected with either wild type (wtBmp2HA) ormutant (RKRmBmp2HA) BMP2 expression plasmids was collectedand analyzed by western blot The HA-tagged growth factors weresecreted equally
A consensus bipartite NLS is characterized by the followingpattern two basic residues approximately 10 spacer residuesand another basic region consisting of 4 basic residues outof five The sequence of the nBMP2 NLS is shown in Figure1(a)The upstreamKR portion of this NLS sequence overlapstheR-X-(KR)-R furin recognition sequencewhere the BMP2proprotein is cleaved to release the mature growth factorand mutation of the KR would thus disrupt production ofthe secreted growth factor (Figure 1(a)) The downstreambasic RKRLK portion of the BMP2 NLS however does notaffect propeptide cleavage To determine whether mutationof this portion of the NLS was sufficient to prevent nuclearlocalization we constructed a mutant BMP2GFP expres-sion plasmid (called RKRmBmp2GFP) in which RKR wasreplaced with AAA and transfected it into cultured cellsThis mutation eliminated nuclear localization of the GFP-tagged BMP2 in cultured RCS cells just like the previouslydescribed KR RKR to AA AAAmutation did [20] To deter-mine whether BMP2 growth factor containing this mutationcould still be secreted anHA-tagged RKRmBmp2 expressionvector (RKRmBmp2HA) was transfected into cultured RCScells and culture medium was collected 48 hrs later HA-tagged proteins were immunoprecipitated from the mediumand visualized by immunoblotting The medium from cellstransfected with RKRmBmp2HA contained as much HA-tagged BMP2 growth factor as medium from cells transfectedwith the wild type wtBmp2HA plasmid indicating that theRKR to AAA did not disrupt synthesis or secretion of theconventional BMP2 growth factor (Figure 1(b))
Having demonstrated in cell culture that theRKR toAAAmutation blocked nuclear localization of nBMP2 while stillallowing secretion of normal quantities of the conventionallyprocessed and secreted BMP2 growth factor we constructeda targeting vector and generated nBmp2NLStm mutant micebearing theRKR toAAAmutation (Figure 2) Both heterozy-gous and homozygous mutant mice appeared morphologi-cally normal and were fertile
BioMed Research International 5
Targeting vector AAA
AAA
DrdI
E1 E2 E3
Wild-type allele
E1
E1
E2
E2
E3
E3
DrdI
DrdI
DrdI
DrdIDrdI
138 kbTargeted allele
61 kb
CreNeo
CreNeo
TK
(a)
138 kb wt allele
61 kb targeted allele
(b)
1000
750
500
++ minusminus
(bp)
(c)
0
02
04
06
08
1
12
14
Gastrocnemius Quadriceps
Wild typeMutant
(d)
Figure 2 Integration of the nBmp2NLStm targeting vector (a) Schematic of the nBmp2NLStm targeting vector recombining into genomicDNA The BMP2 exons are numbered and represented as black boxes while the neo cassette and thymidine kinase (TK) gene arerepresented as grey boxes Insertion of the neo cassette introduces a DrdI restriction site as indicated The mutation of the bipartite NLS(KREKRQAKHKQRKRLKS to KREKRQAKHKQAAALKS) in exon 3 is denoted byAAA (b)After aDrdI digest and Southern hybridizationusing a 31015840 flanking probe complementary to the region indicated with the striped box the wild type band was 138 kb and the targeted alleleband was 61 kb (c) To verify male germline self-excision of the neo cassette PCR was performed using primers that bound to BMP2 outsidetheneo cassette PCRproduced a 525 bp band fromwild typeDNAand a 607 bp if the targeting vectorwas inserted and theneo cassette excisedleaving a LoxP sequence (d) Expression levels of conventional BMP2weremeasured in skeletalmuscle bywestern blot of cytoplasmic extractsusing an anti-BMP2 antibody The bands representing BMP2 proprotein precursor of secreted BMP2 were quantified digitally and mutantswere normalized to wild types within each experiment before averaging between experiments 119899 = 4 for wild type gastrocnemius 119899 = 5 formutant gastrocnemius and 119899 = 3 for both wild type and mutant quadriceps
32 Verification of Normal Secreted BMP2 Function innBmp2NLS119905119898 Mice Work by others predicted that the RKRto AAA mutation would produce secreted BMP2 that wasstill able to bind its receptors activate downstream genesand induce ectopic bone formation but that it might haveincreased diffusion range through the extracellular matrix[31 32] Increased diffusion range could disrupt embryonicpatterning but the normal appearance of the mutant miceruled out major patterning abnormalities Any minor pat-terning abnormalities would likely be manifest in the skele-ton since secreted BMP2 plays an important role in skeletaldevelopment and limb patterning Skeletal preparations ofwild type and homozygous mutant mice were performedand digital caliper measurements were taken of limbs
pelvis scapula ribcage skull and vertebrae with multiplemeasurements at various positions on each bone No sig-nificant differences were detected between wild type andmutant mice in any of the bones measured supportingthe conclusion that the function of secreted BMP2 is notimpaired in nBmp2NLStm mice (see Supplemental Table 1 inSupplementtary Materials available online at httpdxdoi)org1011552013125492)
33 Upside-Down Hanging Ability Is Decreased innBmp2NLS119905119898 Mice Because nBmp2NLStm mice appearedphenotypically normal a series of preliminary tests wereperformed to detect subtle phenotypic changes In oneevaluation five wild type and five mutant mice were tested
6 BioMed Research International
0
50
100
150
200
1 2 3 4 5 6
Hol
d tim
e (s)
+minus
SE
Weeks
Wild typeMutant
lowast lowast
Figure 3 The nBmp2NLStm mice show impaired performance inthe gripstrength test The length of time mice could hold to theunderside of a wire mesh was measured once a week for six weeksin wild type and nBmp2NLStm mice Data are presented as mean plusmnSEM and lowastdenotes 119875 lt 005 119899 = 5 per group
once a week for six weeks to measure the length of time theycould cling to the underside of a wire mesh cage lid [26] Onaverage wild type mice held on 2-3 times as long as mutantsdid at every time point and the difference was significant(119875 lt 005) at weeks 3 and 4 (Figure 3) This trend suggestedthat we should consider the possibility of a neuromusculardefect in nBmp2NLStm mice [26]
34 nBMP2 Is Detectable in Myonuclei of Wild Type but NotnBmp2NLS119905119898 Mice To examine the expression of nBMP2and BMP2 in skeletal muscle gastrocnemius muscle fromwild type and mutant mice was paraffin embedded cross-sectioned and stained using a biotin-tagged primary anti-body against BMP2 (NBP1-19751G from Novus) and a Dakovisualization system The myotubes in skeletal muscle arelong multinucleated cells with the myonuclei positionedagainst the outer edges of the myotube Brown BMP2 stain-ing of myonuclei was detectable in wild type but notnBmp2NLStm muscle whereas nonnuclear BMP2 stainingwas detectable in both wild type and mutant This resultvalidated the effectiveness of the targeted NLS-inactivatingmutation in a whole animal and it also demonstrated thatnBMP2 is expressed in skeletal muscle of control animals(Figure 4)
The presence of extranuclear BMP2 staining in bothwild type and mutant skeletal muscle suggested that theexpression of conventional BMP2 was not inhibited bythe nBmp2NLStm mutation To further examine expressionlevels of the conventional protein cytoplasmic extracts wereprepared from gastrocnemius and quadriceps muscles andexamined by western blot The BMP2 proprotein precursorof secreted BMP2 is detectable as a 50 kDa band Levels of theproprotein were not different between wild type and mutantagain suggesting unaltered expression of conventional BMP2(Figure 2(d))
Table 1 Body weight age and muscle force characteristics
Control (119899 = 7) nBmp2NLStm (119899 = 7)
Age (weeks) 509 plusmn 04 439 plusmn 75
Body weight (g) 369 plusmn 11 353 plusmn 24
GPS muscle weight (mg) 1609 plusmn 96 1717 plusmn 43
Peak tetanic tension (g) 498 plusmn 30 567 plusmn 18
Peak tetanic tension(gmg muscle) 309 plusmn 005 331 plusmn 008lowast
Peak twitch tension (g) 1926 plusmn 97 2206 plusmn 51lowast
Peak twitch tension(gmg muscle) 120 plusmn 003 129 plusmn 004
Data are presented as means plusmn SEM Where indicated lowastindicates 119875 lt 005compared to control values
35 Skeletal Muscle Relaxation Times Are Prolonged innBmp2NLS119905119898 Mice To examine the possibility of a neuro-muscular defect we measured muscle performance in situThe peak tetanic tension of the gastrocnemius plantarisand soleus (GPS) muscle group was slightly but significantlyhigher in nBmp2NLStm mice compared to control micewhen normalized to muscle weight (Table 1) No significantdifferences were observed in the peak twitch tension afternormalization to muscle weight (Table 1) and no significantdifferences were observed in muscle fatigability in responseto a 10 minute 2Hz twitch protocol (Figure 5(a))
A significant difference did emerge however when relax-ation kinetics were examined Half-relaxation times wereprolonged by up to 42 in the nBmp2NLStm muscle com-pared to control (Figure 5(b))This prolonged relaxation timewas apparent in averaged twitch performance traces frominitial (Figure 6(a)) 2min (Figure 6(b)) and 6min (Figure6(c)) time points in the 2Hz twitch contraction protocol
36 Force Frequency Analysis Exhibits a Shift in nBmp2NLS119905119898Mouse Skeletal Muscle Whenmuscle twitches are stimulatedat increasing frequencies in situ the contractions eventuallybecome so frequent that the muscle is unable to completeone relaxation before the next contraction begins and twitchpeak traces begin to merge The force or tension generatedby the muscle increases as twitch peaks merge until forcereaches its maximum and thereafter remains level regardlessof additional increases in the stimulation frequency Thiscould be considered the in situ version of a muscle crampSlowed relaxation after contraction widens each twitch peakcausing peaks to merge at a lower stimulation frequencyshifting the force frequency curve leftward
Because the nBmp2NLStm mice showed slowed relax-ation after contraction we performed a force frequency ana-lysis The nBmp2NLStm mutant mice generated significantlyelevated relative forces at 10 20 40 and 60Hz (119875 lt001) The data collected fit an expected sigmoidal curveand showed a significant leftward shift as determined bythe average force achieved at each frequency compared tothe control (Figure 7) This shift in the force frequency pat-tern as well as the slowed relaxation rates after twitch
BioMed Research International 7
(a) (b)
(c) (d)
(e) (f)
(g) (h)
Figure 4 nBMP2 is detectable in myonuclei of wild type but not nBmp2NLStm mice Gastrocnemius muscles were sectioned and stained byimmunohistochemistry (a)ndash(d) are wild type (e)ndash(h) are mutant (a) and (e) show sections at 40x magnification and all other panels show100x The brown BMP2 antibody stain is evident in myonuclei of wild type muscle whereas myonuclei stain pale blue in the mutant muscleArrows indicate some (but not all) of the myonuclei
8 BioMed Research International
0
02
04
06
08
1
12
0 500 1000
Initi
al te
nsio
n (
)
Time (s)
(a)
0
5
10
15
20
25
30
0 500 1000
Tim
e (m
s)
Time (s)
Wild typeMutant
lowast
(b)
Figure 5 Muscle function is altered in nBmp2NLStm mice (a) Muscle fatigue in both control and nBmp2NLStm GPS muscle complex wassimilar in response to 2Hz twitch contractions for 10 minutes (b) Muscle relaxation was significantly delayed in nBmp2NLStm musclecompared to WT control muscle beginning at 60 seconds and persisting throughout the 10 minutes in which these measurements weremade Data are presented as mean plusmn SEM and lowast denotes 119875 lt 005 119899 = 7 per group Only the first significant difference is marked but allvalues from 60 seconds on were significantly different from control
contractions suggests impairment of Ca2+ sequestration bythe sarcoplasmic reticulum (SR) after muscle contraction
37 SERCA Activity Is Reduced in nBmp2NLS119905119898 SkeletalMuscle The relaxation phase of a skeletal muscle contractionoccurs when Ca2+ is pumped in an ATP-dependent mannerfrom the cytosol back into the SR by sarcoendoplasmicreticulum Ca2+ ATPases (SERCA) Measurement of SERCAactivity in skeletal muscle revealed that SERCA activityin nBmp2NLStm mice is reduced to 81 plusmn 1 of wild typeactivity (Figure 8) This observation is consistent with theslowed relaxation rate observed in nBmp2NLStm muscle
4 Discussion
Several members of the BMP protein family are expressedin variant forms that are translocated to the nucleus ratherthan being secreted from the cell [20] A major challengein characterizing the function of these nuclear variants ofBMPs is that the nuclear protein is translated from the samemRNA transcript as the conventional secreted BMP2 growthfactor as demonstrated by our prior observation that bothnBMP2 and BMP2 are produced from a transfected wild typeBMP2 cDNA expression vector in tissue culture [20] Thisprecludes usage of several of the most common methodsfor studying gene function including RNAi quantitative RT-PCR and gene knockout whichwould all affect andor detectthe conventional secreted BMP2 growth factor as well asnBMP2 Instead we knocked in anRKR toAAAmutation inthe bipartite NLS to prevent nuclear localization of nBMP2while allowing production of secreted BMP2
The RKR to AAA mutation has minimal effect on thefunction of secreted BMPs BMP2 growth factor that was
truncated N-terminally of either the R K or R still hadthe capability to induce ectopic bone formation to thesame extent as wild type BMP2 [31] Moreover deletion ormutation of the homologous sequence in BMP4 produceda protein that was functionally indistinguishable from wildtype BMP4 in receptor binding antagonist binding and tar-get gene responsiveness [32] Animal cap conjugation exper-iments with Xenopus embryos however suggested that thesemutations reduced the ability of Bmp4 to bind heparin sulfatein the extracellular matrix with the result of increased diffu-sion range [32] Together these experiments predicted thatBMP2 bearing the same mutation (RKR toAAA) would alsofunction normally in terms of receptor binding and targetgene activation but might have an increased diffusion range
Secreted BMP2 plays a critical role in dorsal-ventralpatterning of the early embryo in limb patterning and inskeletogenesis so an increased diffusion range of the secretedgrowth factor is likely to cause morphological abnormalitiesin mice [33ndash35] The nBmp2NLStm mouse however dis-played nomorphological or patterning abnormalities and nodifferences in the length width thickness or shape of any ofthe bones measured indicating that there is no physiologi-cally significant alteration in the diffusion range of BMP2 inthese mice We conclude that the RKR to AAA mutation inthe nBmp2NLStm mouse preserved the function of the sec-reted BMP2 growth factor The nBmp2NLStm mouse there-fore is a good model for studying the molecular and physio-logical functions of nBMP2 as distinct from secreted BMP2
The nBmp2NLStm mouse showed decreased ability tocling upside-down to awiremesh cage lid and immunohisto-chemistry showed that nBMP2 is detectable in skeletalmusclemyonuclei of wild type but not mutant animals Mutantmuscle showed no increase in fatigability in situ however
BioMed Research International 9
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(a)
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(b)
Wild typeMutant
0
25
50
75
100
0 50 100 150(ms)
Initi
al p
eak
tens
ion
()
(c)
Figure 6 Slowed muscle relaxation is evident in twitch contractions The average muscle twitch contraction for both wild typeand nBmp2NLStm mice is shown for the initial contractions (a) the contraction following 2 minutes of stimulation (b) and the contractionfollowing 6 minutes of stimulation (c) Each trace represents the average of 7 contractions
suggesting that the reduced hang time is not related to simplemuscle fatigue Instead the shifted force frequency curve andslowed relaxation rates in the mutant muscle suggest thatmutant mice may be more prone to muscle cramping if con-tractions merge during intense muscle activity as they doduring high-frequency in situ stimulations Slowed musclerelaxation does lead to cramping in the human syndromeBrodymyopathy where the length of time required for pump-ing Ca2+ back into the SR after each contraction is increaseddue to mutations in the gene for SERCA1 the pump thatperforms this function in skeletal muscle [36ndash40] Brodypatients have also reported problems with their hands ldquolock-ingrdquo after making a tight fist [40]This cramping can be pain-ful is most frequent after exercise and grows worse withage The slowed relaxation rates in both Brody patientsand nBmp2NLStm mice suggest the possibility that nBMP2may function in some aspect of the SERCA1 productionacti-vation pathway
Our demonstration of reduced SERCA1 activity inthe nBmp2NLStm mouse supports this hypothesis The phe-notype of the nBmp2NLStm mouse however could also
result from the dysfunction of several other proteins becausemuscle contractionrelaxation cycles are quite complex Con-tractions occur when an action potential leads to activationof Ca2+ release channels (ryanodine receptors) in the SRmembrane Ca2+ floods into the cytoplasm and binds tro-ponin C which modulates the function of tropomyosin sothatmyosin can bind strongly to actin and cause shortening ofthe sarcomere The contraction ceases when active pumping(by SERCA1) of Ca2+ back into the SR depletes cytoplasmicCa2+ levels causing release of Ca2+ from troponin C [4142] The nBmp2NLStm phenotype could be caused by leakyryanodine receptors (RyR) in the SRmembrane whichwouldlead to prolonged elevation of Ca2+ in the cytosol even ifthe SERCA pumps were working normally An increasedaffinity of troponinC for Ca2+ could also cause slowedmusclerelaxation by increasing the level of cytosolic Ca2+ depletionthat would have to occur before Ca2+ was released fromtroponin C Ongoing work in our lab on other physiologicalpathways that are controlled by intracellular Ca2+movementincluding neuronal signaling immune system activation and
10 BioMed Research International
0
25
50
75
100
0 50 100 150
Max
forc
e (
)
Frequency of stimulations (pulsess)
MutantWild type
lowastlowast
lowast
lowast
Figure 7 Force frequency analysis of control and nBmp2NLStmmuscle A significant shift in the force frequency curve was observedin nBmp2NLStm muscle compared to wild type supporting theevidence for slowed relaxation in mutants Data are presented asmean plusmn SEM and lowastdenotes 119875 lt 001 119899 = 6 per group
0
02
04
06
08
1
12
Wild type Mutant
Rela
tive S
ERCA
activ
ity
Figure 8 SERCA activity is reduced in nBmp2NLStm muscle com-pared to controls SR was isolated from control and mutant muscleand Ca2+ ATPase activity wasmeasured Data are presented asmeanplusmn SEM 119875 lt 005 119899 = 3 per group
cell cycle regulation indicates that these pathways are alsodisrupted in nBmp2NLStm mice We predict therefore thatnBMP2 affects aspects of intracellular Ca2+ transport that areshared among multiple pathways rather than those (such astroponin C) that are unique to skeletal muscle
5 Conclusions
In summary the muscle phenotype that we identified inthe nBmp2NLStm mouse suggests that a significant alter-ation in intracellular Ca2+ transport resulted from impedednuclear localization of nBMP2 The prolonged relaxationkinetics the shift in the force frequency curve and the
reduction in SERCA activity are most consistent with acondition of delayed Ca2+ uptake by the SR although otherpossibilities have not been ruled out We have recentlyobserved (unpublished data) that nBmp2NLStm mice alsohave cognitive deficits that are consistent with an intra-cellular Ca2+-handling defect in hippocampus It is likelythat Ca2+ handling is disrupted in other tissues as welland that nBMP2 will be found to function in multiplemolecular pathways that govern intracellular Ca2+ transportThe nBmp2NLStm mouse constitutes a valuable model forcharacterizing the functions of nBMP2 relative to Ca2+transport pathways and for distinguishing those functionsfrom the activities of secreted BMP2
Conflict of Interests
None of the authors have any conflict of interests or compet-ing interests
Acknowledgments
This work was supported by NIAMSNIH Grant AR48839to Laura C Bridgewater Jaime L Mayo was supported by agraduate fellowship from the BrighamYoungUniversity Can-cer ResearchCenterThanks are due toDrMario R Capecchiof the University of Utah for generously providing tech-nical assistance with the stem cell electroporation blastulamicroinjections and embryo transfer steps in production ofthe nBmp2NLStm mouse Recombineering plasmids pGalKpL452 and pL253 were obtained from Neal Copeland andNancy Jenkins of the Methodist Hospital Research Institutein Houston TX
References
[1] M R Urist ldquoBone formation by autoinductionrdquo Science vol150 no 698 pp 893ndash899 1965
[2] J M Wozney V Rosen A J Celeste et al ldquoNovel regulators ofbone formation molecular clones and activitiesrdquo Science vol242 no 4885 pp 1528ndash1534 1988
[3] D ChenM Zhao andG RMundy ldquoBonemorphogenetic pro-teinsrdquo Growth Factors vol 22 no 4 pp 233ndash241 2004
[4] A M Goldstein K C Brewer A M Doyle N Nagy and DJ Roberts ldquoBMP signaling is necessary for neural crest cellmigration and ganglion formation in the enteric nervous sys-temrdquo Mechanisms of Development vol 122 no 6 pp 821ndash8332005
[5] B L Hogan ldquoBone morphogenetic proteins multifunctionalregulators of vertebrate developmentrdquo Genes and Developmentvol 10 no 13 pp 1580ndash1594 1996
[6] YMishina ldquoFunction of bonemorphogenetic protein signalingduring mouse developmentrdquo Frontiers in Bioscience vol 8 ppd855ndashd869 2003
[7] T Schlange H-H Arnold and T Brand ldquoBMP2 is a positiveregulator of Nodal signaling during left-right axis formation inthe chicken embryordquo Development vol 129 no 14 pp 3421ndash3429 2002
[8] H Zhang and A Bradley ldquoMice deficient for BMP2 arenonviable and have defects in amnionchorion and cardiac
BioMed Research International 11
developmentrdquo Development vol 122 no 10 pp 2977ndash29861996
[9] C Kawamura M Kizaki and Y Ikeda ldquoBone morphogeneticprotein (BMP)-2 induces apoptosis in human myeloma cellsrdquoLeukemia and Lymphoma vol 43 no 3 pp 635ndash639 2002
[10] M Raida J H Clement K Ameri C Han R D Leek and A LHarris ldquoExpression of bone morphogenetic protein 2 in breastcancer cells inhibits hypoxic cell deathrdquo International Journal ofOncology vol 26 no 6 pp 1465ndash1470 2005
[11] Y Du and H Yip ldquoEffects of bone morphogenetic protein 2on Id expression and neuroblastoma cell differentiationrdquoDiffer-entiation vol 79 no 2 pp 84ndash92 2010
[12] J-C GuimondM Levesque P-LMichaud et al ldquoBMP-2 func-tions independently of SHH signaling and triggers cell con-densation and apoptosis in regenerating axolotl limbsrdquo BMCDevelopmental Biology vol 10 article 15 2010
[13] C C Rider and B Mulloy ldquoBone morphogenetic protein andgrowth differentiation factor cytokine families and their proteinantagonistsrdquo Biochemical Journal vol 429 no 1 pp 1ndash12 2010
[14] D B Constam and E J Robertson ldquoRegulation of bone mor-phogenetic protein activity by pro domains and proprotein con-vertasesrdquo Journal of Cell Biology vol 144 no 1 pp 139ndash149 1999
[15] J M Wozney ldquoBone morphogenetic proteinsrdquo Progress inGrowth Factor Research vol 1 no 4 pp 267ndash280 1989
[16] A Hiyama S S Gogate S Gajghate J Mochida I M Shapiroand M V Risbud ldquoBMP-2 and TGF-120573 stimulate expression of12057313-glucuronosyl transferase 1 (GlcAT-1) in nucleus pulposuscells through AP1 TonEBP and Sp1 role of MAPKsrdquo Journal ofBone and Mineral Research vol 25 no 5 pp 1179ndash1190 2010
[17] CAlarcon A-I ZaromytidouQXi et al ldquoNuclearCDKs driveSmad transcriptional activation and turnover in BMP andTGF-120573 pathwaysrdquo Cell vol 139 no 4 pp 757ndash769 2009
[18] J Lemonnier C Ghayor J Guicheux and J Caverzasio ldquoPro-tein kinase C-independent activation of protein kinase D isinvolved in BMP-2-induced activation of stress mitogen-acti-vated protein kinases JNK and p38 and osteoblastic cell differ-entiationrdquo Journal of Biological Chemistry vol 279 no 1 pp259ndash264 2004
[19] A Moustakas and C-H Heldin ldquoFrommono- to oligo-Smadsthe heart of thematter inTGF-120573 signal transductionrdquoGenes andDevelopment vol 16 no 15 pp 1867ndash1871 2002
[20] J E Felin J L Mayo T J Loos et al ldquoNuclear variants of bonemorphogenetic proteinsrdquo BMC Cell Biology vol 11 no 1 article20 2010
[21] N G Copeland N A Jenkins and D L Court ldquoRecombi-neering a powerful new tool for mouse functional genomicsrdquoNature Reviews Genetics vol 2 no 10 pp 769ndash779 2001
[22] S Warming N Costantino D L Court N A Jenkins and NG Copeland ldquoSimple and highly efficient BAC recombineeringusing galK selectionrdquo Nucleic Acids Research vol 33 no 4article e36 2005
[23] M Bunting K E Bernstein J M Greer M R Capecchi and KR Thomas ldquoTargeting genes for self-excision in the germ linerdquoGenes and Development vol 13 no 12 pp 1524ndash1528 1999
[24] S H George M Gertsenstein K Vintersten et al ldquoDevelop-mental and adult phenotyping directly frommutant embryonicstemcellsrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol 104 no 11 pp 4455ndash4460 2007
[25] S L Mansour K R Thomas and M R Capecchi ldquoDisruptionof the proto-oncogene int-2 in mouse embryo-derived stemcells a general strategy for targeting mutations to non-select-able genesrdquo Nature vol 336 no 6197 pp 348ndash352 1988
[26] V E Papaioannou and R R Behringer Mouse PhenotypesmdashAHandbook of Mutation Analysis Cold Spring Harbor Labora-tory Press Cold Spring Harbor NY USA 2005
[27] C R Hancock J J Brault RWWiseman R L Terjung and RA Meyer ldquo31P-NMR observation of free ADP during fatiguingrepetitive contractions of murine skeletal muscle lacking AK1rdquoThe American Journal of Physiology vol 288 no 6 pp C1298ndashC1304 2005
[28] C R Hancock E Janssen and R L Terjung ldquoSkeletal musclecontractile performance and ADP accumulation in adenylatekinase-deficient micerdquoThe American Journal of Physiology vol288 no 6 pp C1287ndashC1297 2005
[29] C R Hancock E Janssen and R L Terjung ldquoContraction-mediated phosphorylation of AMPK is lower in skeletal muscleof adenylate kinase-deficient micerdquo Journal of Applied Physiol-ogy vol 100 no 2 pp 406ndash413 2006
[30] D Kosk-Kosicka ldquoCalcium signaling protocolsrdquo in Methods inMolecular Biology D G Lambert Ed vol 114 pp 343ndash354Springer New York NY USA 1999
[31] F Hillger G Herr R Rudolph and E Schwarz ldquoBiophysicalcomparison of BMP-2 ProBMP-2 and the free pro-peptidereveals stabilization of the pro-peptide by the mature growthfactorrdquo Journal of Biological Chemistry vol 280 no 15 pp14974ndash14980 2005
[32] BOhkawara S Iemura P tenDijke andNUeno ldquoAction rangeof BMP is defined by its N-terminal basic amino acid corerdquoCurrent Biology vol 12 no 3 pp 205ndash209 2002
[33] E M de Robertis ldquoSpemannrsquos organizer and the self-regulationof embryonic fieldsrdquo Mechanisms of Development vol 126 no11-12 pp 925ndash941 2009
[34] D M Maatouk K-S Choi C M Bouldin and B D Harfe ldquoInthe limb AER Bmp2 and Bmp4 are required for dorsal-ventralpatterning and interdigital cell death but not limb outgrowthrdquoDevelopmental Biology vol 327 no 2 pp 516ndash523 2009
[35] B Robert ldquoBone morphogenetic protein signaling in limboutgrowth and patterningrdquo Development Growth and Differen-tiation vol 49 no 6 pp 455ndash468 2007
[36] A Odermatt K Barton V K Khanna et al ldquoThe mutation ofPro789 to Leu reduces the activity of the fast-twitch skeletalmuscle sarco(endo)plasmic reticulum Ca2+ ATPase (SERCA1)and is associated with Brody diseaserdquoHuman Genetics vol 106no 5 pp 482ndash491 2000
[37] A Odermatt P E Taschner V K Khanna et al ldquoMutationsin the gene-encoding SERCA1 the fast-twitch skeletal musclesarcoplasmic reticulumCa2+ ATPase are associated with Brodydiseaserdquo Nature Genetics vol 14 no 2 pp 191ndash194 1996
[38] A A Benders J H Veerkamp A Oosterhof et al ldquoCa2+ home-ostasis in Brodyrsquos disease A study in skeletal muscle and cul-tured muscle cells and the effects of dantrolene and verapamilrdquoJournal of Clinical Investigation vol 94 no 2 pp 741ndash748 1994
[39] I A Brody ldquoMuscle contracture induced by exercise Asyndrome attributable to decreased relaxing factorrdquo The NewEngland Journal of Medicine vol 281 no 4 pp 187ndash192 1969
[40] Y Zhang J Fujii M S Phillips et al ldquoCharacterization ofcDNA and genomic DNA encoding SERCA1 the Ca2+-ATPaseof human fast-twitch skeletal muscle sarcoplasmic reticulumand its elimination as a candidate gene for Brody diseaserdquoGenomics vol 30 no 3 pp 415ndash424 1995
[41] M Endo ldquoCalcium-induced calcium release in skeletal musclerdquoPhysiological Reviews vol 89 no 4 pp 1153ndash1176 2009
[42] M J Betzenhauser and A RMarks ldquoRyanodine receptor chan-nelopathiesrdquo Pflugers Archiv vol 460 no 2 pp 467ndash480 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
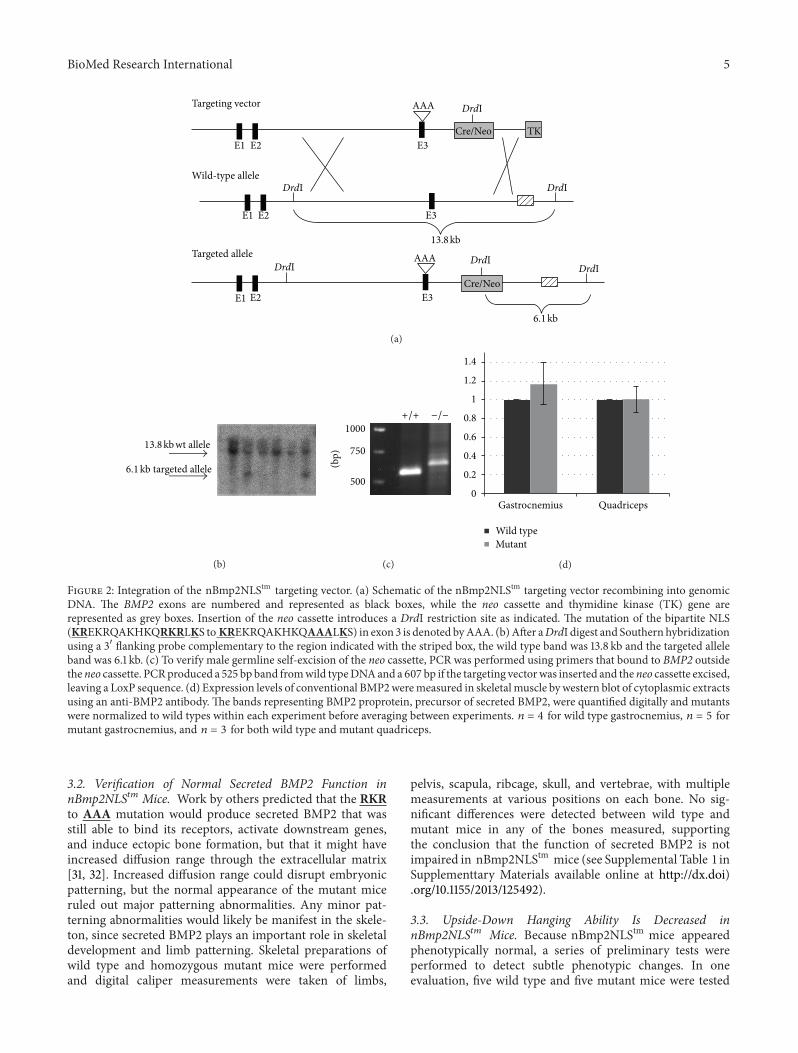
BioMed Research International 5
Targeting vector AAA
AAA
DrdI
E1 E2 E3
Wild-type allele
E1
E1
E2
E2
E3
E3
DrdI
DrdI
DrdI
DrdIDrdI
138 kbTargeted allele
61 kb
CreNeo
CreNeo
TK
(a)
138 kb wt allele
61 kb targeted allele
(b)
1000
750
500
++ minusminus
(bp)
(c)
0
02
04
06
08
1
12
14
Gastrocnemius Quadriceps
Wild typeMutant
(d)
Figure 2 Integration of the nBmp2NLStm targeting vector (a) Schematic of the nBmp2NLStm targeting vector recombining into genomicDNA The BMP2 exons are numbered and represented as black boxes while the neo cassette and thymidine kinase (TK) gene arerepresented as grey boxes Insertion of the neo cassette introduces a DrdI restriction site as indicated The mutation of the bipartite NLS(KREKRQAKHKQRKRLKS to KREKRQAKHKQAAALKS) in exon 3 is denoted byAAA (b)After aDrdI digest and Southern hybridizationusing a 31015840 flanking probe complementary to the region indicated with the striped box the wild type band was 138 kb and the targeted alleleband was 61 kb (c) To verify male germline self-excision of the neo cassette PCR was performed using primers that bound to BMP2 outsidetheneo cassette PCRproduced a 525 bp band fromwild typeDNAand a 607 bp if the targeting vectorwas inserted and theneo cassette excisedleaving a LoxP sequence (d) Expression levels of conventional BMP2weremeasured in skeletalmuscle bywestern blot of cytoplasmic extractsusing an anti-BMP2 antibody The bands representing BMP2 proprotein precursor of secreted BMP2 were quantified digitally and mutantswere normalized to wild types within each experiment before averaging between experiments 119899 = 4 for wild type gastrocnemius 119899 = 5 formutant gastrocnemius and 119899 = 3 for both wild type and mutant quadriceps
32 Verification of Normal Secreted BMP2 Function innBmp2NLS119905119898 Mice Work by others predicted that the RKRto AAA mutation would produce secreted BMP2 that wasstill able to bind its receptors activate downstream genesand induce ectopic bone formation but that it might haveincreased diffusion range through the extracellular matrix[31 32] Increased diffusion range could disrupt embryonicpatterning but the normal appearance of the mutant miceruled out major patterning abnormalities Any minor pat-terning abnormalities would likely be manifest in the skele-ton since secreted BMP2 plays an important role in skeletaldevelopment and limb patterning Skeletal preparations ofwild type and homozygous mutant mice were performedand digital caliper measurements were taken of limbs
pelvis scapula ribcage skull and vertebrae with multiplemeasurements at various positions on each bone No sig-nificant differences were detected between wild type andmutant mice in any of the bones measured supportingthe conclusion that the function of secreted BMP2 is notimpaired in nBmp2NLStm mice (see Supplemental Table 1 inSupplementtary Materials available online at httpdxdoi)org1011552013125492)
33 Upside-Down Hanging Ability Is Decreased innBmp2NLS119905119898 Mice Because nBmp2NLStm mice appearedphenotypically normal a series of preliminary tests wereperformed to detect subtle phenotypic changes In oneevaluation five wild type and five mutant mice were tested
6 BioMed Research International
0
50
100
150
200
1 2 3 4 5 6
Hol
d tim
e (s)
+minus
SE
Weeks
Wild typeMutant
lowast lowast
Figure 3 The nBmp2NLStm mice show impaired performance inthe gripstrength test The length of time mice could hold to theunderside of a wire mesh was measured once a week for six weeksin wild type and nBmp2NLStm mice Data are presented as mean plusmnSEM and lowastdenotes 119875 lt 005 119899 = 5 per group
once a week for six weeks to measure the length of time theycould cling to the underside of a wire mesh cage lid [26] Onaverage wild type mice held on 2-3 times as long as mutantsdid at every time point and the difference was significant(119875 lt 005) at weeks 3 and 4 (Figure 3) This trend suggestedthat we should consider the possibility of a neuromusculardefect in nBmp2NLStm mice [26]
34 nBMP2 Is Detectable in Myonuclei of Wild Type but NotnBmp2NLS119905119898 Mice To examine the expression of nBMP2and BMP2 in skeletal muscle gastrocnemius muscle fromwild type and mutant mice was paraffin embedded cross-sectioned and stained using a biotin-tagged primary anti-body against BMP2 (NBP1-19751G from Novus) and a Dakovisualization system The myotubes in skeletal muscle arelong multinucleated cells with the myonuclei positionedagainst the outer edges of the myotube Brown BMP2 stain-ing of myonuclei was detectable in wild type but notnBmp2NLStm muscle whereas nonnuclear BMP2 stainingwas detectable in both wild type and mutant This resultvalidated the effectiveness of the targeted NLS-inactivatingmutation in a whole animal and it also demonstrated thatnBMP2 is expressed in skeletal muscle of control animals(Figure 4)
The presence of extranuclear BMP2 staining in bothwild type and mutant skeletal muscle suggested that theexpression of conventional BMP2 was not inhibited bythe nBmp2NLStm mutation To further examine expressionlevels of the conventional protein cytoplasmic extracts wereprepared from gastrocnemius and quadriceps muscles andexamined by western blot The BMP2 proprotein precursorof secreted BMP2 is detectable as a 50 kDa band Levels of theproprotein were not different between wild type and mutantagain suggesting unaltered expression of conventional BMP2(Figure 2(d))
Table 1 Body weight age and muscle force characteristics
Control (119899 = 7) nBmp2NLStm (119899 = 7)
Age (weeks) 509 plusmn 04 439 plusmn 75
Body weight (g) 369 plusmn 11 353 plusmn 24
GPS muscle weight (mg) 1609 plusmn 96 1717 plusmn 43
Peak tetanic tension (g) 498 plusmn 30 567 plusmn 18
Peak tetanic tension(gmg muscle) 309 plusmn 005 331 plusmn 008lowast
Peak twitch tension (g) 1926 plusmn 97 2206 plusmn 51lowast
Peak twitch tension(gmg muscle) 120 plusmn 003 129 plusmn 004
Data are presented as means plusmn SEM Where indicated lowastindicates 119875 lt 005compared to control values
35 Skeletal Muscle Relaxation Times Are Prolonged innBmp2NLS119905119898 Mice To examine the possibility of a neuro-muscular defect we measured muscle performance in situThe peak tetanic tension of the gastrocnemius plantarisand soleus (GPS) muscle group was slightly but significantlyhigher in nBmp2NLStm mice compared to control micewhen normalized to muscle weight (Table 1) No significantdifferences were observed in the peak twitch tension afternormalization to muscle weight (Table 1) and no significantdifferences were observed in muscle fatigability in responseto a 10 minute 2Hz twitch protocol (Figure 5(a))
A significant difference did emerge however when relax-ation kinetics were examined Half-relaxation times wereprolonged by up to 42 in the nBmp2NLStm muscle com-pared to control (Figure 5(b))This prolonged relaxation timewas apparent in averaged twitch performance traces frominitial (Figure 6(a)) 2min (Figure 6(b)) and 6min (Figure6(c)) time points in the 2Hz twitch contraction protocol
36 Force Frequency Analysis Exhibits a Shift in nBmp2NLS119905119898Mouse Skeletal Muscle Whenmuscle twitches are stimulatedat increasing frequencies in situ the contractions eventuallybecome so frequent that the muscle is unable to completeone relaxation before the next contraction begins and twitchpeak traces begin to merge The force or tension generatedby the muscle increases as twitch peaks merge until forcereaches its maximum and thereafter remains level regardlessof additional increases in the stimulation frequency Thiscould be considered the in situ version of a muscle crampSlowed relaxation after contraction widens each twitch peakcausing peaks to merge at a lower stimulation frequencyshifting the force frequency curve leftward
Because the nBmp2NLStm mice showed slowed relax-ation after contraction we performed a force frequency ana-lysis The nBmp2NLStm mutant mice generated significantlyelevated relative forces at 10 20 40 and 60Hz (119875 lt001) The data collected fit an expected sigmoidal curveand showed a significant leftward shift as determined bythe average force achieved at each frequency compared tothe control (Figure 7) This shift in the force frequency pat-tern as well as the slowed relaxation rates after twitch
BioMed Research International 7
(a) (b)
(c) (d)
(e) (f)
(g) (h)
Figure 4 nBMP2 is detectable in myonuclei of wild type but not nBmp2NLStm mice Gastrocnemius muscles were sectioned and stained byimmunohistochemistry (a)ndash(d) are wild type (e)ndash(h) are mutant (a) and (e) show sections at 40x magnification and all other panels show100x The brown BMP2 antibody stain is evident in myonuclei of wild type muscle whereas myonuclei stain pale blue in the mutant muscleArrows indicate some (but not all) of the myonuclei
8 BioMed Research International
0
02
04
06
08
1
12
0 500 1000
Initi
al te
nsio
n (
)
Time (s)
(a)
0
5
10
15
20
25
30
0 500 1000
Tim
e (m
s)
Time (s)
Wild typeMutant
lowast
(b)
Figure 5 Muscle function is altered in nBmp2NLStm mice (a) Muscle fatigue in both control and nBmp2NLStm GPS muscle complex wassimilar in response to 2Hz twitch contractions for 10 minutes (b) Muscle relaxation was significantly delayed in nBmp2NLStm musclecompared to WT control muscle beginning at 60 seconds and persisting throughout the 10 minutes in which these measurements weremade Data are presented as mean plusmn SEM and lowast denotes 119875 lt 005 119899 = 7 per group Only the first significant difference is marked but allvalues from 60 seconds on were significantly different from control
contractions suggests impairment of Ca2+ sequestration bythe sarcoplasmic reticulum (SR) after muscle contraction
37 SERCA Activity Is Reduced in nBmp2NLS119905119898 SkeletalMuscle The relaxation phase of a skeletal muscle contractionoccurs when Ca2+ is pumped in an ATP-dependent mannerfrom the cytosol back into the SR by sarcoendoplasmicreticulum Ca2+ ATPases (SERCA) Measurement of SERCAactivity in skeletal muscle revealed that SERCA activityin nBmp2NLStm mice is reduced to 81 plusmn 1 of wild typeactivity (Figure 8) This observation is consistent with theslowed relaxation rate observed in nBmp2NLStm muscle
4 Discussion
Several members of the BMP protein family are expressedin variant forms that are translocated to the nucleus ratherthan being secreted from the cell [20] A major challengein characterizing the function of these nuclear variants ofBMPs is that the nuclear protein is translated from the samemRNA transcript as the conventional secreted BMP2 growthfactor as demonstrated by our prior observation that bothnBMP2 and BMP2 are produced from a transfected wild typeBMP2 cDNA expression vector in tissue culture [20] Thisprecludes usage of several of the most common methodsfor studying gene function including RNAi quantitative RT-PCR and gene knockout whichwould all affect andor detectthe conventional secreted BMP2 growth factor as well asnBMP2 Instead we knocked in anRKR toAAAmutation inthe bipartite NLS to prevent nuclear localization of nBMP2while allowing production of secreted BMP2
The RKR to AAA mutation has minimal effect on thefunction of secreted BMPs BMP2 growth factor that was
truncated N-terminally of either the R K or R still hadthe capability to induce ectopic bone formation to thesame extent as wild type BMP2 [31] Moreover deletion ormutation of the homologous sequence in BMP4 produceda protein that was functionally indistinguishable from wildtype BMP4 in receptor binding antagonist binding and tar-get gene responsiveness [32] Animal cap conjugation exper-iments with Xenopus embryos however suggested that thesemutations reduced the ability of Bmp4 to bind heparin sulfatein the extracellular matrix with the result of increased diffu-sion range [32] Together these experiments predicted thatBMP2 bearing the same mutation (RKR toAAA) would alsofunction normally in terms of receptor binding and targetgene activation but might have an increased diffusion range
Secreted BMP2 plays a critical role in dorsal-ventralpatterning of the early embryo in limb patterning and inskeletogenesis so an increased diffusion range of the secretedgrowth factor is likely to cause morphological abnormalitiesin mice [33ndash35] The nBmp2NLStm mouse however dis-played nomorphological or patterning abnormalities and nodifferences in the length width thickness or shape of any ofthe bones measured indicating that there is no physiologi-cally significant alteration in the diffusion range of BMP2 inthese mice We conclude that the RKR to AAA mutation inthe nBmp2NLStm mouse preserved the function of the sec-reted BMP2 growth factor The nBmp2NLStm mouse there-fore is a good model for studying the molecular and physio-logical functions of nBMP2 as distinct from secreted BMP2
The nBmp2NLStm mouse showed decreased ability tocling upside-down to awiremesh cage lid and immunohisto-chemistry showed that nBMP2 is detectable in skeletalmusclemyonuclei of wild type but not mutant animals Mutantmuscle showed no increase in fatigability in situ however
BioMed Research International 9
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(a)
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(b)
Wild typeMutant
0
25
50
75
100
0 50 100 150(ms)
Initi
al p
eak
tens
ion
()
(c)
Figure 6 Slowed muscle relaxation is evident in twitch contractions The average muscle twitch contraction for both wild typeand nBmp2NLStm mice is shown for the initial contractions (a) the contraction following 2 minutes of stimulation (b) and the contractionfollowing 6 minutes of stimulation (c) Each trace represents the average of 7 contractions
suggesting that the reduced hang time is not related to simplemuscle fatigue Instead the shifted force frequency curve andslowed relaxation rates in the mutant muscle suggest thatmutant mice may be more prone to muscle cramping if con-tractions merge during intense muscle activity as they doduring high-frequency in situ stimulations Slowed musclerelaxation does lead to cramping in the human syndromeBrodymyopathy where the length of time required for pump-ing Ca2+ back into the SR after each contraction is increaseddue to mutations in the gene for SERCA1 the pump thatperforms this function in skeletal muscle [36ndash40] Brodypatients have also reported problems with their hands ldquolock-ingrdquo after making a tight fist [40]This cramping can be pain-ful is most frequent after exercise and grows worse withage The slowed relaxation rates in both Brody patientsand nBmp2NLStm mice suggest the possibility that nBMP2may function in some aspect of the SERCA1 productionacti-vation pathway
Our demonstration of reduced SERCA1 activity inthe nBmp2NLStm mouse supports this hypothesis The phe-notype of the nBmp2NLStm mouse however could also
result from the dysfunction of several other proteins becausemuscle contractionrelaxation cycles are quite complex Con-tractions occur when an action potential leads to activationof Ca2+ release channels (ryanodine receptors) in the SRmembrane Ca2+ floods into the cytoplasm and binds tro-ponin C which modulates the function of tropomyosin sothatmyosin can bind strongly to actin and cause shortening ofthe sarcomere The contraction ceases when active pumping(by SERCA1) of Ca2+ back into the SR depletes cytoplasmicCa2+ levels causing release of Ca2+ from troponin C [4142] The nBmp2NLStm phenotype could be caused by leakyryanodine receptors (RyR) in the SRmembrane whichwouldlead to prolonged elevation of Ca2+ in the cytosol even ifthe SERCA pumps were working normally An increasedaffinity of troponinC for Ca2+ could also cause slowedmusclerelaxation by increasing the level of cytosolic Ca2+ depletionthat would have to occur before Ca2+ was released fromtroponin C Ongoing work in our lab on other physiologicalpathways that are controlled by intracellular Ca2+movementincluding neuronal signaling immune system activation and
10 BioMed Research International
0
25
50
75
100
0 50 100 150
Max
forc
e (
)
Frequency of stimulations (pulsess)
MutantWild type
lowastlowast
lowast
lowast
Figure 7 Force frequency analysis of control and nBmp2NLStmmuscle A significant shift in the force frequency curve was observedin nBmp2NLStm muscle compared to wild type supporting theevidence for slowed relaxation in mutants Data are presented asmean plusmn SEM and lowastdenotes 119875 lt 001 119899 = 6 per group
0
02
04
06
08
1
12
Wild type Mutant
Rela
tive S
ERCA
activ
ity
Figure 8 SERCA activity is reduced in nBmp2NLStm muscle com-pared to controls SR was isolated from control and mutant muscleand Ca2+ ATPase activity wasmeasured Data are presented asmeanplusmn SEM 119875 lt 005 119899 = 3 per group
cell cycle regulation indicates that these pathways are alsodisrupted in nBmp2NLStm mice We predict therefore thatnBMP2 affects aspects of intracellular Ca2+ transport that areshared among multiple pathways rather than those (such astroponin C) that are unique to skeletal muscle
5 Conclusions
In summary the muscle phenotype that we identified inthe nBmp2NLStm mouse suggests that a significant alter-ation in intracellular Ca2+ transport resulted from impedednuclear localization of nBMP2 The prolonged relaxationkinetics the shift in the force frequency curve and the
reduction in SERCA activity are most consistent with acondition of delayed Ca2+ uptake by the SR although otherpossibilities have not been ruled out We have recentlyobserved (unpublished data) that nBmp2NLStm mice alsohave cognitive deficits that are consistent with an intra-cellular Ca2+-handling defect in hippocampus It is likelythat Ca2+ handling is disrupted in other tissues as welland that nBMP2 will be found to function in multiplemolecular pathways that govern intracellular Ca2+ transportThe nBmp2NLStm mouse constitutes a valuable model forcharacterizing the functions of nBMP2 relative to Ca2+transport pathways and for distinguishing those functionsfrom the activities of secreted BMP2
Conflict of Interests
None of the authors have any conflict of interests or compet-ing interests
Acknowledgments
This work was supported by NIAMSNIH Grant AR48839to Laura C Bridgewater Jaime L Mayo was supported by agraduate fellowship from the BrighamYoungUniversity Can-cer ResearchCenterThanks are due toDrMario R Capecchiof the University of Utah for generously providing tech-nical assistance with the stem cell electroporation blastulamicroinjections and embryo transfer steps in production ofthe nBmp2NLStm mouse Recombineering plasmids pGalKpL452 and pL253 were obtained from Neal Copeland andNancy Jenkins of the Methodist Hospital Research Institutein Houston TX
References
[1] M R Urist ldquoBone formation by autoinductionrdquo Science vol150 no 698 pp 893ndash899 1965
[2] J M Wozney V Rosen A J Celeste et al ldquoNovel regulators ofbone formation molecular clones and activitiesrdquo Science vol242 no 4885 pp 1528ndash1534 1988
[3] D ChenM Zhao andG RMundy ldquoBonemorphogenetic pro-teinsrdquo Growth Factors vol 22 no 4 pp 233ndash241 2004
[4] A M Goldstein K C Brewer A M Doyle N Nagy and DJ Roberts ldquoBMP signaling is necessary for neural crest cellmigration and ganglion formation in the enteric nervous sys-temrdquo Mechanisms of Development vol 122 no 6 pp 821ndash8332005
[5] B L Hogan ldquoBone morphogenetic proteins multifunctionalregulators of vertebrate developmentrdquo Genes and Developmentvol 10 no 13 pp 1580ndash1594 1996
[6] YMishina ldquoFunction of bonemorphogenetic protein signalingduring mouse developmentrdquo Frontiers in Bioscience vol 8 ppd855ndashd869 2003
[7] T Schlange H-H Arnold and T Brand ldquoBMP2 is a positiveregulator of Nodal signaling during left-right axis formation inthe chicken embryordquo Development vol 129 no 14 pp 3421ndash3429 2002
[8] H Zhang and A Bradley ldquoMice deficient for BMP2 arenonviable and have defects in amnionchorion and cardiac
BioMed Research International 11
developmentrdquo Development vol 122 no 10 pp 2977ndash29861996
[9] C Kawamura M Kizaki and Y Ikeda ldquoBone morphogeneticprotein (BMP)-2 induces apoptosis in human myeloma cellsrdquoLeukemia and Lymphoma vol 43 no 3 pp 635ndash639 2002
[10] M Raida J H Clement K Ameri C Han R D Leek and A LHarris ldquoExpression of bone morphogenetic protein 2 in breastcancer cells inhibits hypoxic cell deathrdquo International Journal ofOncology vol 26 no 6 pp 1465ndash1470 2005
[11] Y Du and H Yip ldquoEffects of bone morphogenetic protein 2on Id expression and neuroblastoma cell differentiationrdquoDiffer-entiation vol 79 no 2 pp 84ndash92 2010
[12] J-C GuimondM Levesque P-LMichaud et al ldquoBMP-2 func-tions independently of SHH signaling and triggers cell con-densation and apoptosis in regenerating axolotl limbsrdquo BMCDevelopmental Biology vol 10 article 15 2010
[13] C C Rider and B Mulloy ldquoBone morphogenetic protein andgrowth differentiation factor cytokine families and their proteinantagonistsrdquo Biochemical Journal vol 429 no 1 pp 1ndash12 2010
[14] D B Constam and E J Robertson ldquoRegulation of bone mor-phogenetic protein activity by pro domains and proprotein con-vertasesrdquo Journal of Cell Biology vol 144 no 1 pp 139ndash149 1999
[15] J M Wozney ldquoBone morphogenetic proteinsrdquo Progress inGrowth Factor Research vol 1 no 4 pp 267ndash280 1989
[16] A Hiyama S S Gogate S Gajghate J Mochida I M Shapiroand M V Risbud ldquoBMP-2 and TGF-120573 stimulate expression of12057313-glucuronosyl transferase 1 (GlcAT-1) in nucleus pulposuscells through AP1 TonEBP and Sp1 role of MAPKsrdquo Journal ofBone and Mineral Research vol 25 no 5 pp 1179ndash1190 2010
[17] CAlarcon A-I ZaromytidouQXi et al ldquoNuclearCDKs driveSmad transcriptional activation and turnover in BMP andTGF-120573 pathwaysrdquo Cell vol 139 no 4 pp 757ndash769 2009
[18] J Lemonnier C Ghayor J Guicheux and J Caverzasio ldquoPro-tein kinase C-independent activation of protein kinase D isinvolved in BMP-2-induced activation of stress mitogen-acti-vated protein kinases JNK and p38 and osteoblastic cell differ-entiationrdquo Journal of Biological Chemistry vol 279 no 1 pp259ndash264 2004
[19] A Moustakas and C-H Heldin ldquoFrommono- to oligo-Smadsthe heart of thematter inTGF-120573 signal transductionrdquoGenes andDevelopment vol 16 no 15 pp 1867ndash1871 2002
[20] J E Felin J L Mayo T J Loos et al ldquoNuclear variants of bonemorphogenetic proteinsrdquo BMC Cell Biology vol 11 no 1 article20 2010
[21] N G Copeland N A Jenkins and D L Court ldquoRecombi-neering a powerful new tool for mouse functional genomicsrdquoNature Reviews Genetics vol 2 no 10 pp 769ndash779 2001
[22] S Warming N Costantino D L Court N A Jenkins and NG Copeland ldquoSimple and highly efficient BAC recombineeringusing galK selectionrdquo Nucleic Acids Research vol 33 no 4article e36 2005
[23] M Bunting K E Bernstein J M Greer M R Capecchi and KR Thomas ldquoTargeting genes for self-excision in the germ linerdquoGenes and Development vol 13 no 12 pp 1524ndash1528 1999
[24] S H George M Gertsenstein K Vintersten et al ldquoDevelop-mental and adult phenotyping directly frommutant embryonicstemcellsrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol 104 no 11 pp 4455ndash4460 2007
[25] S L Mansour K R Thomas and M R Capecchi ldquoDisruptionof the proto-oncogene int-2 in mouse embryo-derived stemcells a general strategy for targeting mutations to non-select-able genesrdquo Nature vol 336 no 6197 pp 348ndash352 1988
[26] V E Papaioannou and R R Behringer Mouse PhenotypesmdashAHandbook of Mutation Analysis Cold Spring Harbor Labora-tory Press Cold Spring Harbor NY USA 2005
[27] C R Hancock J J Brault RWWiseman R L Terjung and RA Meyer ldquo31P-NMR observation of free ADP during fatiguingrepetitive contractions of murine skeletal muscle lacking AK1rdquoThe American Journal of Physiology vol 288 no 6 pp C1298ndashC1304 2005
[28] C R Hancock E Janssen and R L Terjung ldquoSkeletal musclecontractile performance and ADP accumulation in adenylatekinase-deficient micerdquoThe American Journal of Physiology vol288 no 6 pp C1287ndashC1297 2005
[29] C R Hancock E Janssen and R L Terjung ldquoContraction-mediated phosphorylation of AMPK is lower in skeletal muscleof adenylate kinase-deficient micerdquo Journal of Applied Physiol-ogy vol 100 no 2 pp 406ndash413 2006
[30] D Kosk-Kosicka ldquoCalcium signaling protocolsrdquo in Methods inMolecular Biology D G Lambert Ed vol 114 pp 343ndash354Springer New York NY USA 1999
[31] F Hillger G Herr R Rudolph and E Schwarz ldquoBiophysicalcomparison of BMP-2 ProBMP-2 and the free pro-peptidereveals stabilization of the pro-peptide by the mature growthfactorrdquo Journal of Biological Chemistry vol 280 no 15 pp14974ndash14980 2005
[32] BOhkawara S Iemura P tenDijke andNUeno ldquoAction rangeof BMP is defined by its N-terminal basic amino acid corerdquoCurrent Biology vol 12 no 3 pp 205ndash209 2002
[33] E M de Robertis ldquoSpemannrsquos organizer and the self-regulationof embryonic fieldsrdquo Mechanisms of Development vol 126 no11-12 pp 925ndash941 2009
[34] D M Maatouk K-S Choi C M Bouldin and B D Harfe ldquoInthe limb AER Bmp2 and Bmp4 are required for dorsal-ventralpatterning and interdigital cell death but not limb outgrowthrdquoDevelopmental Biology vol 327 no 2 pp 516ndash523 2009
[35] B Robert ldquoBone morphogenetic protein signaling in limboutgrowth and patterningrdquo Development Growth and Differen-tiation vol 49 no 6 pp 455ndash468 2007
[36] A Odermatt K Barton V K Khanna et al ldquoThe mutation ofPro789 to Leu reduces the activity of the fast-twitch skeletalmuscle sarco(endo)plasmic reticulum Ca2+ ATPase (SERCA1)and is associated with Brody diseaserdquoHuman Genetics vol 106no 5 pp 482ndash491 2000
[37] A Odermatt P E Taschner V K Khanna et al ldquoMutationsin the gene-encoding SERCA1 the fast-twitch skeletal musclesarcoplasmic reticulumCa2+ ATPase are associated with Brodydiseaserdquo Nature Genetics vol 14 no 2 pp 191ndash194 1996
[38] A A Benders J H Veerkamp A Oosterhof et al ldquoCa2+ home-ostasis in Brodyrsquos disease A study in skeletal muscle and cul-tured muscle cells and the effects of dantrolene and verapamilrdquoJournal of Clinical Investigation vol 94 no 2 pp 741ndash748 1994
[39] I A Brody ldquoMuscle contracture induced by exercise Asyndrome attributable to decreased relaxing factorrdquo The NewEngland Journal of Medicine vol 281 no 4 pp 187ndash192 1969
[40] Y Zhang J Fujii M S Phillips et al ldquoCharacterization ofcDNA and genomic DNA encoding SERCA1 the Ca2+-ATPaseof human fast-twitch skeletal muscle sarcoplasmic reticulumand its elimination as a candidate gene for Brody diseaserdquoGenomics vol 30 no 3 pp 415ndash424 1995
[41] M Endo ldquoCalcium-induced calcium release in skeletal musclerdquoPhysiological Reviews vol 89 no 4 pp 1153ndash1176 2009
[42] M J Betzenhauser and A RMarks ldquoRyanodine receptor chan-nelopathiesrdquo Pflugers Archiv vol 460 no 2 pp 467ndash480 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
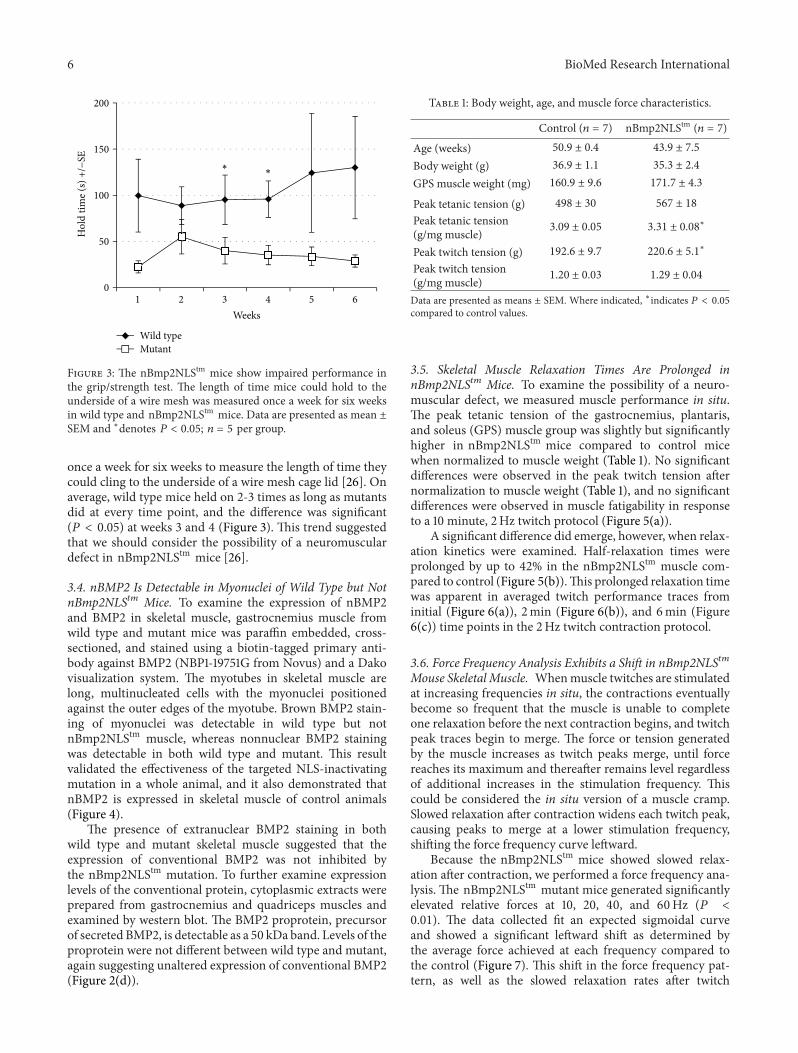
6 BioMed Research International
0
50
100
150
200
1 2 3 4 5 6
Hol
d tim
e (s)
+minus
SE
Weeks
Wild typeMutant
lowast lowast
Figure 3 The nBmp2NLStm mice show impaired performance inthe gripstrength test The length of time mice could hold to theunderside of a wire mesh was measured once a week for six weeksin wild type and nBmp2NLStm mice Data are presented as mean plusmnSEM and lowastdenotes 119875 lt 005 119899 = 5 per group
once a week for six weeks to measure the length of time theycould cling to the underside of a wire mesh cage lid [26] Onaverage wild type mice held on 2-3 times as long as mutantsdid at every time point and the difference was significant(119875 lt 005) at weeks 3 and 4 (Figure 3) This trend suggestedthat we should consider the possibility of a neuromusculardefect in nBmp2NLStm mice [26]
34 nBMP2 Is Detectable in Myonuclei of Wild Type but NotnBmp2NLS119905119898 Mice To examine the expression of nBMP2and BMP2 in skeletal muscle gastrocnemius muscle fromwild type and mutant mice was paraffin embedded cross-sectioned and stained using a biotin-tagged primary anti-body against BMP2 (NBP1-19751G from Novus) and a Dakovisualization system The myotubes in skeletal muscle arelong multinucleated cells with the myonuclei positionedagainst the outer edges of the myotube Brown BMP2 stain-ing of myonuclei was detectable in wild type but notnBmp2NLStm muscle whereas nonnuclear BMP2 stainingwas detectable in both wild type and mutant This resultvalidated the effectiveness of the targeted NLS-inactivatingmutation in a whole animal and it also demonstrated thatnBMP2 is expressed in skeletal muscle of control animals(Figure 4)
The presence of extranuclear BMP2 staining in bothwild type and mutant skeletal muscle suggested that theexpression of conventional BMP2 was not inhibited bythe nBmp2NLStm mutation To further examine expressionlevels of the conventional protein cytoplasmic extracts wereprepared from gastrocnemius and quadriceps muscles andexamined by western blot The BMP2 proprotein precursorof secreted BMP2 is detectable as a 50 kDa band Levels of theproprotein were not different between wild type and mutantagain suggesting unaltered expression of conventional BMP2(Figure 2(d))
Table 1 Body weight age and muscle force characteristics
Control (119899 = 7) nBmp2NLStm (119899 = 7)
Age (weeks) 509 plusmn 04 439 plusmn 75
Body weight (g) 369 plusmn 11 353 plusmn 24
GPS muscle weight (mg) 1609 plusmn 96 1717 plusmn 43
Peak tetanic tension (g) 498 plusmn 30 567 plusmn 18
Peak tetanic tension(gmg muscle) 309 plusmn 005 331 plusmn 008lowast
Peak twitch tension (g) 1926 plusmn 97 2206 plusmn 51lowast
Peak twitch tension(gmg muscle) 120 plusmn 003 129 plusmn 004
Data are presented as means plusmn SEM Where indicated lowastindicates 119875 lt 005compared to control values
35 Skeletal Muscle Relaxation Times Are Prolonged innBmp2NLS119905119898 Mice To examine the possibility of a neuro-muscular defect we measured muscle performance in situThe peak tetanic tension of the gastrocnemius plantarisand soleus (GPS) muscle group was slightly but significantlyhigher in nBmp2NLStm mice compared to control micewhen normalized to muscle weight (Table 1) No significantdifferences were observed in the peak twitch tension afternormalization to muscle weight (Table 1) and no significantdifferences were observed in muscle fatigability in responseto a 10 minute 2Hz twitch protocol (Figure 5(a))
A significant difference did emerge however when relax-ation kinetics were examined Half-relaxation times wereprolonged by up to 42 in the nBmp2NLStm muscle com-pared to control (Figure 5(b))This prolonged relaxation timewas apparent in averaged twitch performance traces frominitial (Figure 6(a)) 2min (Figure 6(b)) and 6min (Figure6(c)) time points in the 2Hz twitch contraction protocol
36 Force Frequency Analysis Exhibits a Shift in nBmp2NLS119905119898Mouse Skeletal Muscle Whenmuscle twitches are stimulatedat increasing frequencies in situ the contractions eventuallybecome so frequent that the muscle is unable to completeone relaxation before the next contraction begins and twitchpeak traces begin to merge The force or tension generatedby the muscle increases as twitch peaks merge until forcereaches its maximum and thereafter remains level regardlessof additional increases in the stimulation frequency Thiscould be considered the in situ version of a muscle crampSlowed relaxation after contraction widens each twitch peakcausing peaks to merge at a lower stimulation frequencyshifting the force frequency curve leftward
Because the nBmp2NLStm mice showed slowed relax-ation after contraction we performed a force frequency ana-lysis The nBmp2NLStm mutant mice generated significantlyelevated relative forces at 10 20 40 and 60Hz (119875 lt001) The data collected fit an expected sigmoidal curveand showed a significant leftward shift as determined bythe average force achieved at each frequency compared tothe control (Figure 7) This shift in the force frequency pat-tern as well as the slowed relaxation rates after twitch
BioMed Research International 7
(a) (b)
(c) (d)
(e) (f)
(g) (h)
Figure 4 nBMP2 is detectable in myonuclei of wild type but not nBmp2NLStm mice Gastrocnemius muscles were sectioned and stained byimmunohistochemistry (a)ndash(d) are wild type (e)ndash(h) are mutant (a) and (e) show sections at 40x magnification and all other panels show100x The brown BMP2 antibody stain is evident in myonuclei of wild type muscle whereas myonuclei stain pale blue in the mutant muscleArrows indicate some (but not all) of the myonuclei
8 BioMed Research International
0
02
04
06
08
1
12
0 500 1000
Initi
al te
nsio
n (
)
Time (s)
(a)
0
5
10
15
20
25
30
0 500 1000
Tim
e (m
s)
Time (s)
Wild typeMutant
lowast
(b)
Figure 5 Muscle function is altered in nBmp2NLStm mice (a) Muscle fatigue in both control and nBmp2NLStm GPS muscle complex wassimilar in response to 2Hz twitch contractions for 10 minutes (b) Muscle relaxation was significantly delayed in nBmp2NLStm musclecompared to WT control muscle beginning at 60 seconds and persisting throughout the 10 minutes in which these measurements weremade Data are presented as mean plusmn SEM and lowast denotes 119875 lt 005 119899 = 7 per group Only the first significant difference is marked but allvalues from 60 seconds on were significantly different from control
contractions suggests impairment of Ca2+ sequestration bythe sarcoplasmic reticulum (SR) after muscle contraction
37 SERCA Activity Is Reduced in nBmp2NLS119905119898 SkeletalMuscle The relaxation phase of a skeletal muscle contractionoccurs when Ca2+ is pumped in an ATP-dependent mannerfrom the cytosol back into the SR by sarcoendoplasmicreticulum Ca2+ ATPases (SERCA) Measurement of SERCAactivity in skeletal muscle revealed that SERCA activityin nBmp2NLStm mice is reduced to 81 plusmn 1 of wild typeactivity (Figure 8) This observation is consistent with theslowed relaxation rate observed in nBmp2NLStm muscle
4 Discussion
Several members of the BMP protein family are expressedin variant forms that are translocated to the nucleus ratherthan being secreted from the cell [20] A major challengein characterizing the function of these nuclear variants ofBMPs is that the nuclear protein is translated from the samemRNA transcript as the conventional secreted BMP2 growthfactor as demonstrated by our prior observation that bothnBMP2 and BMP2 are produced from a transfected wild typeBMP2 cDNA expression vector in tissue culture [20] Thisprecludes usage of several of the most common methodsfor studying gene function including RNAi quantitative RT-PCR and gene knockout whichwould all affect andor detectthe conventional secreted BMP2 growth factor as well asnBMP2 Instead we knocked in anRKR toAAAmutation inthe bipartite NLS to prevent nuclear localization of nBMP2while allowing production of secreted BMP2
The RKR to AAA mutation has minimal effect on thefunction of secreted BMPs BMP2 growth factor that was
truncated N-terminally of either the R K or R still hadthe capability to induce ectopic bone formation to thesame extent as wild type BMP2 [31] Moreover deletion ormutation of the homologous sequence in BMP4 produceda protein that was functionally indistinguishable from wildtype BMP4 in receptor binding antagonist binding and tar-get gene responsiveness [32] Animal cap conjugation exper-iments with Xenopus embryos however suggested that thesemutations reduced the ability of Bmp4 to bind heparin sulfatein the extracellular matrix with the result of increased diffu-sion range [32] Together these experiments predicted thatBMP2 bearing the same mutation (RKR toAAA) would alsofunction normally in terms of receptor binding and targetgene activation but might have an increased diffusion range
Secreted BMP2 plays a critical role in dorsal-ventralpatterning of the early embryo in limb patterning and inskeletogenesis so an increased diffusion range of the secretedgrowth factor is likely to cause morphological abnormalitiesin mice [33ndash35] The nBmp2NLStm mouse however dis-played nomorphological or patterning abnormalities and nodifferences in the length width thickness or shape of any ofthe bones measured indicating that there is no physiologi-cally significant alteration in the diffusion range of BMP2 inthese mice We conclude that the RKR to AAA mutation inthe nBmp2NLStm mouse preserved the function of the sec-reted BMP2 growth factor The nBmp2NLStm mouse there-fore is a good model for studying the molecular and physio-logical functions of nBMP2 as distinct from secreted BMP2
The nBmp2NLStm mouse showed decreased ability tocling upside-down to awiremesh cage lid and immunohisto-chemistry showed that nBMP2 is detectable in skeletalmusclemyonuclei of wild type but not mutant animals Mutantmuscle showed no increase in fatigability in situ however
BioMed Research International 9
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(a)
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(b)
Wild typeMutant
0
25
50
75
100
0 50 100 150(ms)
Initi
al p
eak
tens
ion
()
(c)
Figure 6 Slowed muscle relaxation is evident in twitch contractions The average muscle twitch contraction for both wild typeand nBmp2NLStm mice is shown for the initial contractions (a) the contraction following 2 minutes of stimulation (b) and the contractionfollowing 6 minutes of stimulation (c) Each trace represents the average of 7 contractions
suggesting that the reduced hang time is not related to simplemuscle fatigue Instead the shifted force frequency curve andslowed relaxation rates in the mutant muscle suggest thatmutant mice may be more prone to muscle cramping if con-tractions merge during intense muscle activity as they doduring high-frequency in situ stimulations Slowed musclerelaxation does lead to cramping in the human syndromeBrodymyopathy where the length of time required for pump-ing Ca2+ back into the SR after each contraction is increaseddue to mutations in the gene for SERCA1 the pump thatperforms this function in skeletal muscle [36ndash40] Brodypatients have also reported problems with their hands ldquolock-ingrdquo after making a tight fist [40]This cramping can be pain-ful is most frequent after exercise and grows worse withage The slowed relaxation rates in both Brody patientsand nBmp2NLStm mice suggest the possibility that nBMP2may function in some aspect of the SERCA1 productionacti-vation pathway
Our demonstration of reduced SERCA1 activity inthe nBmp2NLStm mouse supports this hypothesis The phe-notype of the nBmp2NLStm mouse however could also
result from the dysfunction of several other proteins becausemuscle contractionrelaxation cycles are quite complex Con-tractions occur when an action potential leads to activationof Ca2+ release channels (ryanodine receptors) in the SRmembrane Ca2+ floods into the cytoplasm and binds tro-ponin C which modulates the function of tropomyosin sothatmyosin can bind strongly to actin and cause shortening ofthe sarcomere The contraction ceases when active pumping(by SERCA1) of Ca2+ back into the SR depletes cytoplasmicCa2+ levels causing release of Ca2+ from troponin C [4142] The nBmp2NLStm phenotype could be caused by leakyryanodine receptors (RyR) in the SRmembrane whichwouldlead to prolonged elevation of Ca2+ in the cytosol even ifthe SERCA pumps were working normally An increasedaffinity of troponinC for Ca2+ could also cause slowedmusclerelaxation by increasing the level of cytosolic Ca2+ depletionthat would have to occur before Ca2+ was released fromtroponin C Ongoing work in our lab on other physiologicalpathways that are controlled by intracellular Ca2+movementincluding neuronal signaling immune system activation and
10 BioMed Research International
0
25
50
75
100
0 50 100 150
Max
forc
e (
)
Frequency of stimulations (pulsess)
MutantWild type
lowastlowast
lowast
lowast
Figure 7 Force frequency analysis of control and nBmp2NLStmmuscle A significant shift in the force frequency curve was observedin nBmp2NLStm muscle compared to wild type supporting theevidence for slowed relaxation in mutants Data are presented asmean plusmn SEM and lowastdenotes 119875 lt 001 119899 = 6 per group
0
02
04
06
08
1
12
Wild type Mutant
Rela
tive S
ERCA
activ
ity
Figure 8 SERCA activity is reduced in nBmp2NLStm muscle com-pared to controls SR was isolated from control and mutant muscleand Ca2+ ATPase activity wasmeasured Data are presented asmeanplusmn SEM 119875 lt 005 119899 = 3 per group
cell cycle regulation indicates that these pathways are alsodisrupted in nBmp2NLStm mice We predict therefore thatnBMP2 affects aspects of intracellular Ca2+ transport that areshared among multiple pathways rather than those (such astroponin C) that are unique to skeletal muscle
5 Conclusions
In summary the muscle phenotype that we identified inthe nBmp2NLStm mouse suggests that a significant alter-ation in intracellular Ca2+ transport resulted from impedednuclear localization of nBMP2 The prolonged relaxationkinetics the shift in the force frequency curve and the
reduction in SERCA activity are most consistent with acondition of delayed Ca2+ uptake by the SR although otherpossibilities have not been ruled out We have recentlyobserved (unpublished data) that nBmp2NLStm mice alsohave cognitive deficits that are consistent with an intra-cellular Ca2+-handling defect in hippocampus It is likelythat Ca2+ handling is disrupted in other tissues as welland that nBMP2 will be found to function in multiplemolecular pathways that govern intracellular Ca2+ transportThe nBmp2NLStm mouse constitutes a valuable model forcharacterizing the functions of nBMP2 relative to Ca2+transport pathways and for distinguishing those functionsfrom the activities of secreted BMP2
Conflict of Interests
None of the authors have any conflict of interests or compet-ing interests
Acknowledgments
This work was supported by NIAMSNIH Grant AR48839to Laura C Bridgewater Jaime L Mayo was supported by agraduate fellowship from the BrighamYoungUniversity Can-cer ResearchCenterThanks are due toDrMario R Capecchiof the University of Utah for generously providing tech-nical assistance with the stem cell electroporation blastulamicroinjections and embryo transfer steps in production ofthe nBmp2NLStm mouse Recombineering plasmids pGalKpL452 and pL253 were obtained from Neal Copeland andNancy Jenkins of the Methodist Hospital Research Institutein Houston TX
References
[1] M R Urist ldquoBone formation by autoinductionrdquo Science vol150 no 698 pp 893ndash899 1965
[2] J M Wozney V Rosen A J Celeste et al ldquoNovel regulators ofbone formation molecular clones and activitiesrdquo Science vol242 no 4885 pp 1528ndash1534 1988
[3] D ChenM Zhao andG RMundy ldquoBonemorphogenetic pro-teinsrdquo Growth Factors vol 22 no 4 pp 233ndash241 2004
[4] A M Goldstein K C Brewer A M Doyle N Nagy and DJ Roberts ldquoBMP signaling is necessary for neural crest cellmigration and ganglion formation in the enteric nervous sys-temrdquo Mechanisms of Development vol 122 no 6 pp 821ndash8332005
[5] B L Hogan ldquoBone morphogenetic proteins multifunctionalregulators of vertebrate developmentrdquo Genes and Developmentvol 10 no 13 pp 1580ndash1594 1996
[6] YMishina ldquoFunction of bonemorphogenetic protein signalingduring mouse developmentrdquo Frontiers in Bioscience vol 8 ppd855ndashd869 2003
[7] T Schlange H-H Arnold and T Brand ldquoBMP2 is a positiveregulator of Nodal signaling during left-right axis formation inthe chicken embryordquo Development vol 129 no 14 pp 3421ndash3429 2002
[8] H Zhang and A Bradley ldquoMice deficient for BMP2 arenonviable and have defects in amnionchorion and cardiac
BioMed Research International 11
developmentrdquo Development vol 122 no 10 pp 2977ndash29861996
[9] C Kawamura M Kizaki and Y Ikeda ldquoBone morphogeneticprotein (BMP)-2 induces apoptosis in human myeloma cellsrdquoLeukemia and Lymphoma vol 43 no 3 pp 635ndash639 2002
[10] M Raida J H Clement K Ameri C Han R D Leek and A LHarris ldquoExpression of bone morphogenetic protein 2 in breastcancer cells inhibits hypoxic cell deathrdquo International Journal ofOncology vol 26 no 6 pp 1465ndash1470 2005
[11] Y Du and H Yip ldquoEffects of bone morphogenetic protein 2on Id expression and neuroblastoma cell differentiationrdquoDiffer-entiation vol 79 no 2 pp 84ndash92 2010
[12] J-C GuimondM Levesque P-LMichaud et al ldquoBMP-2 func-tions independently of SHH signaling and triggers cell con-densation and apoptosis in regenerating axolotl limbsrdquo BMCDevelopmental Biology vol 10 article 15 2010
[13] C C Rider and B Mulloy ldquoBone morphogenetic protein andgrowth differentiation factor cytokine families and their proteinantagonistsrdquo Biochemical Journal vol 429 no 1 pp 1ndash12 2010
[14] D B Constam and E J Robertson ldquoRegulation of bone mor-phogenetic protein activity by pro domains and proprotein con-vertasesrdquo Journal of Cell Biology vol 144 no 1 pp 139ndash149 1999
[15] J M Wozney ldquoBone morphogenetic proteinsrdquo Progress inGrowth Factor Research vol 1 no 4 pp 267ndash280 1989
[16] A Hiyama S S Gogate S Gajghate J Mochida I M Shapiroand M V Risbud ldquoBMP-2 and TGF-120573 stimulate expression of12057313-glucuronosyl transferase 1 (GlcAT-1) in nucleus pulposuscells through AP1 TonEBP and Sp1 role of MAPKsrdquo Journal ofBone and Mineral Research vol 25 no 5 pp 1179ndash1190 2010
[17] CAlarcon A-I ZaromytidouQXi et al ldquoNuclearCDKs driveSmad transcriptional activation and turnover in BMP andTGF-120573 pathwaysrdquo Cell vol 139 no 4 pp 757ndash769 2009
[18] J Lemonnier C Ghayor J Guicheux and J Caverzasio ldquoPro-tein kinase C-independent activation of protein kinase D isinvolved in BMP-2-induced activation of stress mitogen-acti-vated protein kinases JNK and p38 and osteoblastic cell differ-entiationrdquo Journal of Biological Chemistry vol 279 no 1 pp259ndash264 2004
[19] A Moustakas and C-H Heldin ldquoFrommono- to oligo-Smadsthe heart of thematter inTGF-120573 signal transductionrdquoGenes andDevelopment vol 16 no 15 pp 1867ndash1871 2002
[20] J E Felin J L Mayo T J Loos et al ldquoNuclear variants of bonemorphogenetic proteinsrdquo BMC Cell Biology vol 11 no 1 article20 2010
[21] N G Copeland N A Jenkins and D L Court ldquoRecombi-neering a powerful new tool for mouse functional genomicsrdquoNature Reviews Genetics vol 2 no 10 pp 769ndash779 2001
[22] S Warming N Costantino D L Court N A Jenkins and NG Copeland ldquoSimple and highly efficient BAC recombineeringusing galK selectionrdquo Nucleic Acids Research vol 33 no 4article e36 2005
[23] M Bunting K E Bernstein J M Greer M R Capecchi and KR Thomas ldquoTargeting genes for self-excision in the germ linerdquoGenes and Development vol 13 no 12 pp 1524ndash1528 1999
[24] S H George M Gertsenstein K Vintersten et al ldquoDevelop-mental and adult phenotyping directly frommutant embryonicstemcellsrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol 104 no 11 pp 4455ndash4460 2007
[25] S L Mansour K R Thomas and M R Capecchi ldquoDisruptionof the proto-oncogene int-2 in mouse embryo-derived stemcells a general strategy for targeting mutations to non-select-able genesrdquo Nature vol 336 no 6197 pp 348ndash352 1988
[26] V E Papaioannou and R R Behringer Mouse PhenotypesmdashAHandbook of Mutation Analysis Cold Spring Harbor Labora-tory Press Cold Spring Harbor NY USA 2005
[27] C R Hancock J J Brault RWWiseman R L Terjung and RA Meyer ldquo31P-NMR observation of free ADP during fatiguingrepetitive contractions of murine skeletal muscle lacking AK1rdquoThe American Journal of Physiology vol 288 no 6 pp C1298ndashC1304 2005
[28] C R Hancock E Janssen and R L Terjung ldquoSkeletal musclecontractile performance and ADP accumulation in adenylatekinase-deficient micerdquoThe American Journal of Physiology vol288 no 6 pp C1287ndashC1297 2005
[29] C R Hancock E Janssen and R L Terjung ldquoContraction-mediated phosphorylation of AMPK is lower in skeletal muscleof adenylate kinase-deficient micerdquo Journal of Applied Physiol-ogy vol 100 no 2 pp 406ndash413 2006
[30] D Kosk-Kosicka ldquoCalcium signaling protocolsrdquo in Methods inMolecular Biology D G Lambert Ed vol 114 pp 343ndash354Springer New York NY USA 1999
[31] F Hillger G Herr R Rudolph and E Schwarz ldquoBiophysicalcomparison of BMP-2 ProBMP-2 and the free pro-peptidereveals stabilization of the pro-peptide by the mature growthfactorrdquo Journal of Biological Chemistry vol 280 no 15 pp14974ndash14980 2005
[32] BOhkawara S Iemura P tenDijke andNUeno ldquoAction rangeof BMP is defined by its N-terminal basic amino acid corerdquoCurrent Biology vol 12 no 3 pp 205ndash209 2002
[33] E M de Robertis ldquoSpemannrsquos organizer and the self-regulationof embryonic fieldsrdquo Mechanisms of Development vol 126 no11-12 pp 925ndash941 2009
[34] D M Maatouk K-S Choi C M Bouldin and B D Harfe ldquoInthe limb AER Bmp2 and Bmp4 are required for dorsal-ventralpatterning and interdigital cell death but not limb outgrowthrdquoDevelopmental Biology vol 327 no 2 pp 516ndash523 2009
[35] B Robert ldquoBone morphogenetic protein signaling in limboutgrowth and patterningrdquo Development Growth and Differen-tiation vol 49 no 6 pp 455ndash468 2007
[36] A Odermatt K Barton V K Khanna et al ldquoThe mutation ofPro789 to Leu reduces the activity of the fast-twitch skeletalmuscle sarco(endo)plasmic reticulum Ca2+ ATPase (SERCA1)and is associated with Brody diseaserdquoHuman Genetics vol 106no 5 pp 482ndash491 2000
[37] A Odermatt P E Taschner V K Khanna et al ldquoMutationsin the gene-encoding SERCA1 the fast-twitch skeletal musclesarcoplasmic reticulumCa2+ ATPase are associated with Brodydiseaserdquo Nature Genetics vol 14 no 2 pp 191ndash194 1996
[38] A A Benders J H Veerkamp A Oosterhof et al ldquoCa2+ home-ostasis in Brodyrsquos disease A study in skeletal muscle and cul-tured muscle cells and the effects of dantrolene and verapamilrdquoJournal of Clinical Investigation vol 94 no 2 pp 741ndash748 1994
[39] I A Brody ldquoMuscle contracture induced by exercise Asyndrome attributable to decreased relaxing factorrdquo The NewEngland Journal of Medicine vol 281 no 4 pp 187ndash192 1969
[40] Y Zhang J Fujii M S Phillips et al ldquoCharacterization ofcDNA and genomic DNA encoding SERCA1 the Ca2+-ATPaseof human fast-twitch skeletal muscle sarcoplasmic reticulumand its elimination as a candidate gene for Brody diseaserdquoGenomics vol 30 no 3 pp 415ndash424 1995
[41] M Endo ldquoCalcium-induced calcium release in skeletal musclerdquoPhysiological Reviews vol 89 no 4 pp 1153ndash1176 2009
[42] M J Betzenhauser and A RMarks ldquoRyanodine receptor chan-nelopathiesrdquo Pflugers Archiv vol 460 no 2 pp 467ndash480 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
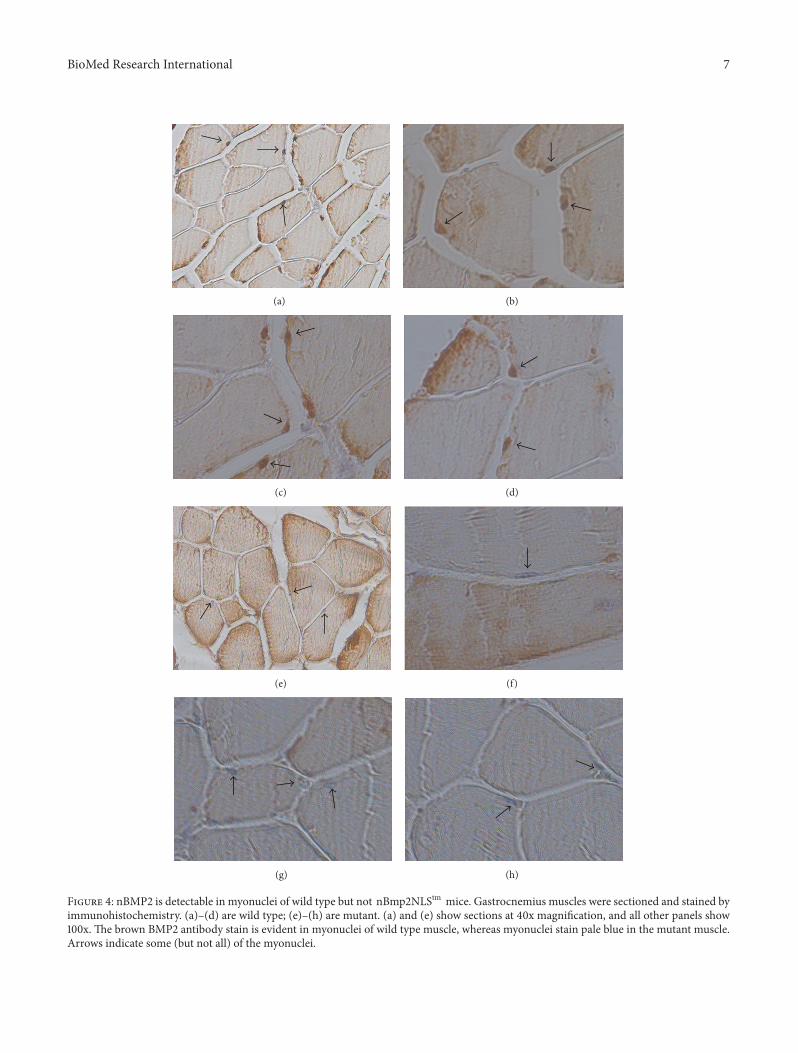
BioMed Research International 7
(a) (b)
(c) (d)
(e) (f)
(g) (h)
Figure 4 nBMP2 is detectable in myonuclei of wild type but not nBmp2NLStm mice Gastrocnemius muscles were sectioned and stained byimmunohistochemistry (a)ndash(d) are wild type (e)ndash(h) are mutant (a) and (e) show sections at 40x magnification and all other panels show100x The brown BMP2 antibody stain is evident in myonuclei of wild type muscle whereas myonuclei stain pale blue in the mutant muscleArrows indicate some (but not all) of the myonuclei
8 BioMed Research International
0
02
04
06
08
1
12
0 500 1000
Initi
al te
nsio
n (
)
Time (s)
(a)
0
5
10
15
20
25
30
0 500 1000
Tim
e (m
s)
Time (s)
Wild typeMutant
lowast
(b)
Figure 5 Muscle function is altered in nBmp2NLStm mice (a) Muscle fatigue in both control and nBmp2NLStm GPS muscle complex wassimilar in response to 2Hz twitch contractions for 10 minutes (b) Muscle relaxation was significantly delayed in nBmp2NLStm musclecompared to WT control muscle beginning at 60 seconds and persisting throughout the 10 minutes in which these measurements weremade Data are presented as mean plusmn SEM and lowast denotes 119875 lt 005 119899 = 7 per group Only the first significant difference is marked but allvalues from 60 seconds on were significantly different from control
contractions suggests impairment of Ca2+ sequestration bythe sarcoplasmic reticulum (SR) after muscle contraction
37 SERCA Activity Is Reduced in nBmp2NLS119905119898 SkeletalMuscle The relaxation phase of a skeletal muscle contractionoccurs when Ca2+ is pumped in an ATP-dependent mannerfrom the cytosol back into the SR by sarcoendoplasmicreticulum Ca2+ ATPases (SERCA) Measurement of SERCAactivity in skeletal muscle revealed that SERCA activityin nBmp2NLStm mice is reduced to 81 plusmn 1 of wild typeactivity (Figure 8) This observation is consistent with theslowed relaxation rate observed in nBmp2NLStm muscle
4 Discussion
Several members of the BMP protein family are expressedin variant forms that are translocated to the nucleus ratherthan being secreted from the cell [20] A major challengein characterizing the function of these nuclear variants ofBMPs is that the nuclear protein is translated from the samemRNA transcript as the conventional secreted BMP2 growthfactor as demonstrated by our prior observation that bothnBMP2 and BMP2 are produced from a transfected wild typeBMP2 cDNA expression vector in tissue culture [20] Thisprecludes usage of several of the most common methodsfor studying gene function including RNAi quantitative RT-PCR and gene knockout whichwould all affect andor detectthe conventional secreted BMP2 growth factor as well asnBMP2 Instead we knocked in anRKR toAAAmutation inthe bipartite NLS to prevent nuclear localization of nBMP2while allowing production of secreted BMP2
The RKR to AAA mutation has minimal effect on thefunction of secreted BMPs BMP2 growth factor that was
truncated N-terminally of either the R K or R still hadthe capability to induce ectopic bone formation to thesame extent as wild type BMP2 [31] Moreover deletion ormutation of the homologous sequence in BMP4 produceda protein that was functionally indistinguishable from wildtype BMP4 in receptor binding antagonist binding and tar-get gene responsiveness [32] Animal cap conjugation exper-iments with Xenopus embryos however suggested that thesemutations reduced the ability of Bmp4 to bind heparin sulfatein the extracellular matrix with the result of increased diffu-sion range [32] Together these experiments predicted thatBMP2 bearing the same mutation (RKR toAAA) would alsofunction normally in terms of receptor binding and targetgene activation but might have an increased diffusion range
Secreted BMP2 plays a critical role in dorsal-ventralpatterning of the early embryo in limb patterning and inskeletogenesis so an increased diffusion range of the secretedgrowth factor is likely to cause morphological abnormalitiesin mice [33ndash35] The nBmp2NLStm mouse however dis-played nomorphological or patterning abnormalities and nodifferences in the length width thickness or shape of any ofthe bones measured indicating that there is no physiologi-cally significant alteration in the diffusion range of BMP2 inthese mice We conclude that the RKR to AAA mutation inthe nBmp2NLStm mouse preserved the function of the sec-reted BMP2 growth factor The nBmp2NLStm mouse there-fore is a good model for studying the molecular and physio-logical functions of nBMP2 as distinct from secreted BMP2
The nBmp2NLStm mouse showed decreased ability tocling upside-down to awiremesh cage lid and immunohisto-chemistry showed that nBMP2 is detectable in skeletalmusclemyonuclei of wild type but not mutant animals Mutantmuscle showed no increase in fatigability in situ however
BioMed Research International 9
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(a)
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(b)
Wild typeMutant
0
25
50
75
100
0 50 100 150(ms)
Initi
al p
eak
tens
ion
()
(c)
Figure 6 Slowed muscle relaxation is evident in twitch contractions The average muscle twitch contraction for both wild typeand nBmp2NLStm mice is shown for the initial contractions (a) the contraction following 2 minutes of stimulation (b) and the contractionfollowing 6 minutes of stimulation (c) Each trace represents the average of 7 contractions
suggesting that the reduced hang time is not related to simplemuscle fatigue Instead the shifted force frequency curve andslowed relaxation rates in the mutant muscle suggest thatmutant mice may be more prone to muscle cramping if con-tractions merge during intense muscle activity as they doduring high-frequency in situ stimulations Slowed musclerelaxation does lead to cramping in the human syndromeBrodymyopathy where the length of time required for pump-ing Ca2+ back into the SR after each contraction is increaseddue to mutations in the gene for SERCA1 the pump thatperforms this function in skeletal muscle [36ndash40] Brodypatients have also reported problems with their hands ldquolock-ingrdquo after making a tight fist [40]This cramping can be pain-ful is most frequent after exercise and grows worse withage The slowed relaxation rates in both Brody patientsand nBmp2NLStm mice suggest the possibility that nBMP2may function in some aspect of the SERCA1 productionacti-vation pathway
Our demonstration of reduced SERCA1 activity inthe nBmp2NLStm mouse supports this hypothesis The phe-notype of the nBmp2NLStm mouse however could also
result from the dysfunction of several other proteins becausemuscle contractionrelaxation cycles are quite complex Con-tractions occur when an action potential leads to activationof Ca2+ release channels (ryanodine receptors) in the SRmembrane Ca2+ floods into the cytoplasm and binds tro-ponin C which modulates the function of tropomyosin sothatmyosin can bind strongly to actin and cause shortening ofthe sarcomere The contraction ceases when active pumping(by SERCA1) of Ca2+ back into the SR depletes cytoplasmicCa2+ levels causing release of Ca2+ from troponin C [4142] The nBmp2NLStm phenotype could be caused by leakyryanodine receptors (RyR) in the SRmembrane whichwouldlead to prolonged elevation of Ca2+ in the cytosol even ifthe SERCA pumps were working normally An increasedaffinity of troponinC for Ca2+ could also cause slowedmusclerelaxation by increasing the level of cytosolic Ca2+ depletionthat would have to occur before Ca2+ was released fromtroponin C Ongoing work in our lab on other physiologicalpathways that are controlled by intracellular Ca2+movementincluding neuronal signaling immune system activation and
10 BioMed Research International
0
25
50
75
100
0 50 100 150
Max
forc
e (
)
Frequency of stimulations (pulsess)
MutantWild type
lowastlowast
lowast
lowast
Figure 7 Force frequency analysis of control and nBmp2NLStmmuscle A significant shift in the force frequency curve was observedin nBmp2NLStm muscle compared to wild type supporting theevidence for slowed relaxation in mutants Data are presented asmean plusmn SEM and lowastdenotes 119875 lt 001 119899 = 6 per group
0
02
04
06
08
1
12
Wild type Mutant
Rela
tive S
ERCA
activ
ity
Figure 8 SERCA activity is reduced in nBmp2NLStm muscle com-pared to controls SR was isolated from control and mutant muscleand Ca2+ ATPase activity wasmeasured Data are presented asmeanplusmn SEM 119875 lt 005 119899 = 3 per group
cell cycle regulation indicates that these pathways are alsodisrupted in nBmp2NLStm mice We predict therefore thatnBMP2 affects aspects of intracellular Ca2+ transport that areshared among multiple pathways rather than those (such astroponin C) that are unique to skeletal muscle
5 Conclusions
In summary the muscle phenotype that we identified inthe nBmp2NLStm mouse suggests that a significant alter-ation in intracellular Ca2+ transport resulted from impedednuclear localization of nBMP2 The prolonged relaxationkinetics the shift in the force frequency curve and the
reduction in SERCA activity are most consistent with acondition of delayed Ca2+ uptake by the SR although otherpossibilities have not been ruled out We have recentlyobserved (unpublished data) that nBmp2NLStm mice alsohave cognitive deficits that are consistent with an intra-cellular Ca2+-handling defect in hippocampus It is likelythat Ca2+ handling is disrupted in other tissues as welland that nBMP2 will be found to function in multiplemolecular pathways that govern intracellular Ca2+ transportThe nBmp2NLStm mouse constitutes a valuable model forcharacterizing the functions of nBMP2 relative to Ca2+transport pathways and for distinguishing those functionsfrom the activities of secreted BMP2
Conflict of Interests
None of the authors have any conflict of interests or compet-ing interests
Acknowledgments
This work was supported by NIAMSNIH Grant AR48839to Laura C Bridgewater Jaime L Mayo was supported by agraduate fellowship from the BrighamYoungUniversity Can-cer ResearchCenterThanks are due toDrMario R Capecchiof the University of Utah for generously providing tech-nical assistance with the stem cell electroporation blastulamicroinjections and embryo transfer steps in production ofthe nBmp2NLStm mouse Recombineering plasmids pGalKpL452 and pL253 were obtained from Neal Copeland andNancy Jenkins of the Methodist Hospital Research Institutein Houston TX
References
[1] M R Urist ldquoBone formation by autoinductionrdquo Science vol150 no 698 pp 893ndash899 1965
[2] J M Wozney V Rosen A J Celeste et al ldquoNovel regulators ofbone formation molecular clones and activitiesrdquo Science vol242 no 4885 pp 1528ndash1534 1988
[3] D ChenM Zhao andG RMundy ldquoBonemorphogenetic pro-teinsrdquo Growth Factors vol 22 no 4 pp 233ndash241 2004
[4] A M Goldstein K C Brewer A M Doyle N Nagy and DJ Roberts ldquoBMP signaling is necessary for neural crest cellmigration and ganglion formation in the enteric nervous sys-temrdquo Mechanisms of Development vol 122 no 6 pp 821ndash8332005
[5] B L Hogan ldquoBone morphogenetic proteins multifunctionalregulators of vertebrate developmentrdquo Genes and Developmentvol 10 no 13 pp 1580ndash1594 1996
[6] YMishina ldquoFunction of bonemorphogenetic protein signalingduring mouse developmentrdquo Frontiers in Bioscience vol 8 ppd855ndashd869 2003
[7] T Schlange H-H Arnold and T Brand ldquoBMP2 is a positiveregulator of Nodal signaling during left-right axis formation inthe chicken embryordquo Development vol 129 no 14 pp 3421ndash3429 2002
[8] H Zhang and A Bradley ldquoMice deficient for BMP2 arenonviable and have defects in amnionchorion and cardiac
BioMed Research International 11
developmentrdquo Development vol 122 no 10 pp 2977ndash29861996
[9] C Kawamura M Kizaki and Y Ikeda ldquoBone morphogeneticprotein (BMP)-2 induces apoptosis in human myeloma cellsrdquoLeukemia and Lymphoma vol 43 no 3 pp 635ndash639 2002
[10] M Raida J H Clement K Ameri C Han R D Leek and A LHarris ldquoExpression of bone morphogenetic protein 2 in breastcancer cells inhibits hypoxic cell deathrdquo International Journal ofOncology vol 26 no 6 pp 1465ndash1470 2005
[11] Y Du and H Yip ldquoEffects of bone morphogenetic protein 2on Id expression and neuroblastoma cell differentiationrdquoDiffer-entiation vol 79 no 2 pp 84ndash92 2010
[12] J-C GuimondM Levesque P-LMichaud et al ldquoBMP-2 func-tions independently of SHH signaling and triggers cell con-densation and apoptosis in regenerating axolotl limbsrdquo BMCDevelopmental Biology vol 10 article 15 2010
[13] C C Rider and B Mulloy ldquoBone morphogenetic protein andgrowth differentiation factor cytokine families and their proteinantagonistsrdquo Biochemical Journal vol 429 no 1 pp 1ndash12 2010
[14] D B Constam and E J Robertson ldquoRegulation of bone mor-phogenetic protein activity by pro domains and proprotein con-vertasesrdquo Journal of Cell Biology vol 144 no 1 pp 139ndash149 1999
[15] J M Wozney ldquoBone morphogenetic proteinsrdquo Progress inGrowth Factor Research vol 1 no 4 pp 267ndash280 1989
[16] A Hiyama S S Gogate S Gajghate J Mochida I M Shapiroand M V Risbud ldquoBMP-2 and TGF-120573 stimulate expression of12057313-glucuronosyl transferase 1 (GlcAT-1) in nucleus pulposuscells through AP1 TonEBP and Sp1 role of MAPKsrdquo Journal ofBone and Mineral Research vol 25 no 5 pp 1179ndash1190 2010
[17] CAlarcon A-I ZaromytidouQXi et al ldquoNuclearCDKs driveSmad transcriptional activation and turnover in BMP andTGF-120573 pathwaysrdquo Cell vol 139 no 4 pp 757ndash769 2009
[18] J Lemonnier C Ghayor J Guicheux and J Caverzasio ldquoPro-tein kinase C-independent activation of protein kinase D isinvolved in BMP-2-induced activation of stress mitogen-acti-vated protein kinases JNK and p38 and osteoblastic cell differ-entiationrdquo Journal of Biological Chemistry vol 279 no 1 pp259ndash264 2004
[19] A Moustakas and C-H Heldin ldquoFrommono- to oligo-Smadsthe heart of thematter inTGF-120573 signal transductionrdquoGenes andDevelopment vol 16 no 15 pp 1867ndash1871 2002
[20] J E Felin J L Mayo T J Loos et al ldquoNuclear variants of bonemorphogenetic proteinsrdquo BMC Cell Biology vol 11 no 1 article20 2010
[21] N G Copeland N A Jenkins and D L Court ldquoRecombi-neering a powerful new tool for mouse functional genomicsrdquoNature Reviews Genetics vol 2 no 10 pp 769ndash779 2001
[22] S Warming N Costantino D L Court N A Jenkins and NG Copeland ldquoSimple and highly efficient BAC recombineeringusing galK selectionrdquo Nucleic Acids Research vol 33 no 4article e36 2005
[23] M Bunting K E Bernstein J M Greer M R Capecchi and KR Thomas ldquoTargeting genes for self-excision in the germ linerdquoGenes and Development vol 13 no 12 pp 1524ndash1528 1999
[24] S H George M Gertsenstein K Vintersten et al ldquoDevelop-mental and adult phenotyping directly frommutant embryonicstemcellsrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol 104 no 11 pp 4455ndash4460 2007
[25] S L Mansour K R Thomas and M R Capecchi ldquoDisruptionof the proto-oncogene int-2 in mouse embryo-derived stemcells a general strategy for targeting mutations to non-select-able genesrdquo Nature vol 336 no 6197 pp 348ndash352 1988
[26] V E Papaioannou and R R Behringer Mouse PhenotypesmdashAHandbook of Mutation Analysis Cold Spring Harbor Labora-tory Press Cold Spring Harbor NY USA 2005
[27] C R Hancock J J Brault RWWiseman R L Terjung and RA Meyer ldquo31P-NMR observation of free ADP during fatiguingrepetitive contractions of murine skeletal muscle lacking AK1rdquoThe American Journal of Physiology vol 288 no 6 pp C1298ndashC1304 2005
[28] C R Hancock E Janssen and R L Terjung ldquoSkeletal musclecontractile performance and ADP accumulation in adenylatekinase-deficient micerdquoThe American Journal of Physiology vol288 no 6 pp C1287ndashC1297 2005
[29] C R Hancock E Janssen and R L Terjung ldquoContraction-mediated phosphorylation of AMPK is lower in skeletal muscleof adenylate kinase-deficient micerdquo Journal of Applied Physiol-ogy vol 100 no 2 pp 406ndash413 2006
[30] D Kosk-Kosicka ldquoCalcium signaling protocolsrdquo in Methods inMolecular Biology D G Lambert Ed vol 114 pp 343ndash354Springer New York NY USA 1999
[31] F Hillger G Herr R Rudolph and E Schwarz ldquoBiophysicalcomparison of BMP-2 ProBMP-2 and the free pro-peptidereveals stabilization of the pro-peptide by the mature growthfactorrdquo Journal of Biological Chemistry vol 280 no 15 pp14974ndash14980 2005
[32] BOhkawara S Iemura P tenDijke andNUeno ldquoAction rangeof BMP is defined by its N-terminal basic amino acid corerdquoCurrent Biology vol 12 no 3 pp 205ndash209 2002
[33] E M de Robertis ldquoSpemannrsquos organizer and the self-regulationof embryonic fieldsrdquo Mechanisms of Development vol 126 no11-12 pp 925ndash941 2009
[34] D M Maatouk K-S Choi C M Bouldin and B D Harfe ldquoInthe limb AER Bmp2 and Bmp4 are required for dorsal-ventralpatterning and interdigital cell death but not limb outgrowthrdquoDevelopmental Biology vol 327 no 2 pp 516ndash523 2009
[35] B Robert ldquoBone morphogenetic protein signaling in limboutgrowth and patterningrdquo Development Growth and Differen-tiation vol 49 no 6 pp 455ndash468 2007
[36] A Odermatt K Barton V K Khanna et al ldquoThe mutation ofPro789 to Leu reduces the activity of the fast-twitch skeletalmuscle sarco(endo)plasmic reticulum Ca2+ ATPase (SERCA1)and is associated with Brody diseaserdquoHuman Genetics vol 106no 5 pp 482ndash491 2000
[37] A Odermatt P E Taschner V K Khanna et al ldquoMutationsin the gene-encoding SERCA1 the fast-twitch skeletal musclesarcoplasmic reticulumCa2+ ATPase are associated with Brodydiseaserdquo Nature Genetics vol 14 no 2 pp 191ndash194 1996
[38] A A Benders J H Veerkamp A Oosterhof et al ldquoCa2+ home-ostasis in Brodyrsquos disease A study in skeletal muscle and cul-tured muscle cells and the effects of dantrolene and verapamilrdquoJournal of Clinical Investigation vol 94 no 2 pp 741ndash748 1994
[39] I A Brody ldquoMuscle contracture induced by exercise Asyndrome attributable to decreased relaxing factorrdquo The NewEngland Journal of Medicine vol 281 no 4 pp 187ndash192 1969
[40] Y Zhang J Fujii M S Phillips et al ldquoCharacterization ofcDNA and genomic DNA encoding SERCA1 the Ca2+-ATPaseof human fast-twitch skeletal muscle sarcoplasmic reticulumand its elimination as a candidate gene for Brody diseaserdquoGenomics vol 30 no 3 pp 415ndash424 1995
[41] M Endo ldquoCalcium-induced calcium release in skeletal musclerdquoPhysiological Reviews vol 89 no 4 pp 1153ndash1176 2009
[42] M J Betzenhauser and A RMarks ldquoRyanodine receptor chan-nelopathiesrdquo Pflugers Archiv vol 460 no 2 pp 467ndash480 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
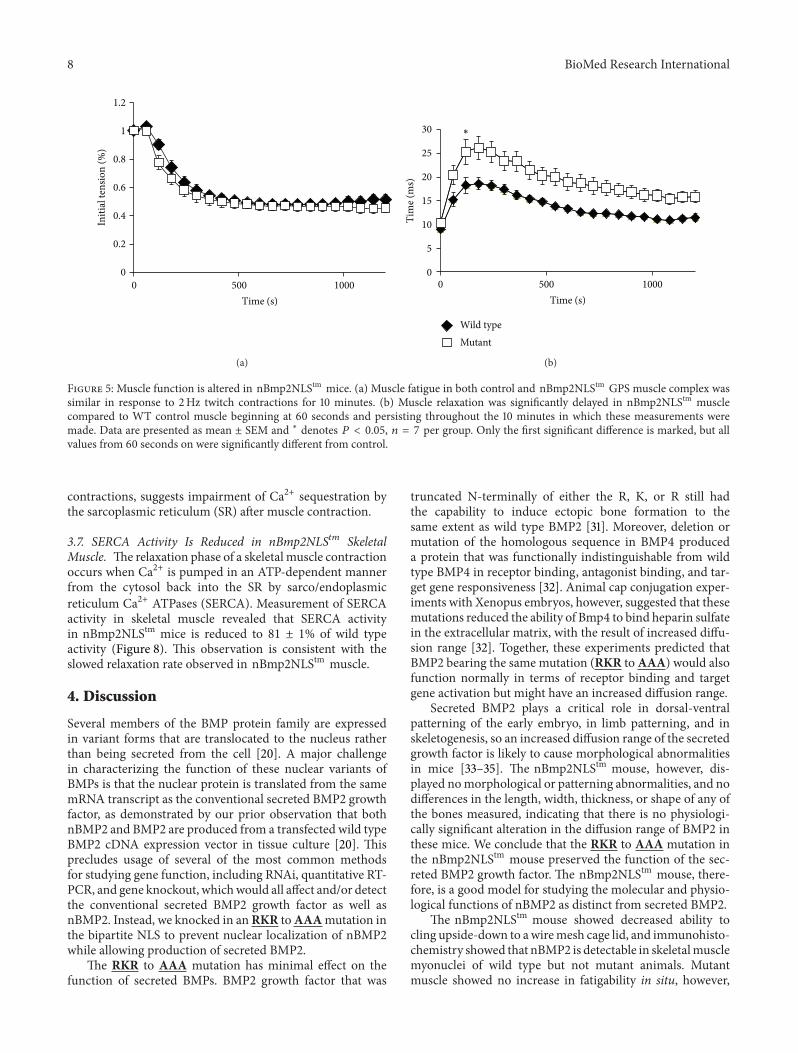
8 BioMed Research International
0
02
04
06
08
1
12
0 500 1000
Initi
al te
nsio
n (
)
Time (s)
(a)
0
5
10
15
20
25
30
0 500 1000
Tim
e (m
s)
Time (s)
Wild typeMutant
lowast
(b)
Figure 5 Muscle function is altered in nBmp2NLStm mice (a) Muscle fatigue in both control and nBmp2NLStm GPS muscle complex wassimilar in response to 2Hz twitch contractions for 10 minutes (b) Muscle relaxation was significantly delayed in nBmp2NLStm musclecompared to WT control muscle beginning at 60 seconds and persisting throughout the 10 minutes in which these measurements weremade Data are presented as mean plusmn SEM and lowast denotes 119875 lt 005 119899 = 7 per group Only the first significant difference is marked but allvalues from 60 seconds on were significantly different from control
contractions suggests impairment of Ca2+ sequestration bythe sarcoplasmic reticulum (SR) after muscle contraction
37 SERCA Activity Is Reduced in nBmp2NLS119905119898 SkeletalMuscle The relaxation phase of a skeletal muscle contractionoccurs when Ca2+ is pumped in an ATP-dependent mannerfrom the cytosol back into the SR by sarcoendoplasmicreticulum Ca2+ ATPases (SERCA) Measurement of SERCAactivity in skeletal muscle revealed that SERCA activityin nBmp2NLStm mice is reduced to 81 plusmn 1 of wild typeactivity (Figure 8) This observation is consistent with theslowed relaxation rate observed in nBmp2NLStm muscle
4 Discussion
Several members of the BMP protein family are expressedin variant forms that are translocated to the nucleus ratherthan being secreted from the cell [20] A major challengein characterizing the function of these nuclear variants ofBMPs is that the nuclear protein is translated from the samemRNA transcript as the conventional secreted BMP2 growthfactor as demonstrated by our prior observation that bothnBMP2 and BMP2 are produced from a transfected wild typeBMP2 cDNA expression vector in tissue culture [20] Thisprecludes usage of several of the most common methodsfor studying gene function including RNAi quantitative RT-PCR and gene knockout whichwould all affect andor detectthe conventional secreted BMP2 growth factor as well asnBMP2 Instead we knocked in anRKR toAAAmutation inthe bipartite NLS to prevent nuclear localization of nBMP2while allowing production of secreted BMP2
The RKR to AAA mutation has minimal effect on thefunction of secreted BMPs BMP2 growth factor that was
truncated N-terminally of either the R K or R still hadthe capability to induce ectopic bone formation to thesame extent as wild type BMP2 [31] Moreover deletion ormutation of the homologous sequence in BMP4 produceda protein that was functionally indistinguishable from wildtype BMP4 in receptor binding antagonist binding and tar-get gene responsiveness [32] Animal cap conjugation exper-iments with Xenopus embryos however suggested that thesemutations reduced the ability of Bmp4 to bind heparin sulfatein the extracellular matrix with the result of increased diffu-sion range [32] Together these experiments predicted thatBMP2 bearing the same mutation (RKR toAAA) would alsofunction normally in terms of receptor binding and targetgene activation but might have an increased diffusion range
Secreted BMP2 plays a critical role in dorsal-ventralpatterning of the early embryo in limb patterning and inskeletogenesis so an increased diffusion range of the secretedgrowth factor is likely to cause morphological abnormalitiesin mice [33ndash35] The nBmp2NLStm mouse however dis-played nomorphological or patterning abnormalities and nodifferences in the length width thickness or shape of any ofthe bones measured indicating that there is no physiologi-cally significant alteration in the diffusion range of BMP2 inthese mice We conclude that the RKR to AAA mutation inthe nBmp2NLStm mouse preserved the function of the sec-reted BMP2 growth factor The nBmp2NLStm mouse there-fore is a good model for studying the molecular and physio-logical functions of nBMP2 as distinct from secreted BMP2
The nBmp2NLStm mouse showed decreased ability tocling upside-down to awiremesh cage lid and immunohisto-chemistry showed that nBMP2 is detectable in skeletalmusclemyonuclei of wild type but not mutant animals Mutantmuscle showed no increase in fatigability in situ however
BioMed Research International 9
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(a)
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(b)
Wild typeMutant
0
25
50
75
100
0 50 100 150(ms)
Initi
al p
eak
tens
ion
()
(c)
Figure 6 Slowed muscle relaxation is evident in twitch contractions The average muscle twitch contraction for both wild typeand nBmp2NLStm mice is shown for the initial contractions (a) the contraction following 2 minutes of stimulation (b) and the contractionfollowing 6 minutes of stimulation (c) Each trace represents the average of 7 contractions
suggesting that the reduced hang time is not related to simplemuscle fatigue Instead the shifted force frequency curve andslowed relaxation rates in the mutant muscle suggest thatmutant mice may be more prone to muscle cramping if con-tractions merge during intense muscle activity as they doduring high-frequency in situ stimulations Slowed musclerelaxation does lead to cramping in the human syndromeBrodymyopathy where the length of time required for pump-ing Ca2+ back into the SR after each contraction is increaseddue to mutations in the gene for SERCA1 the pump thatperforms this function in skeletal muscle [36ndash40] Brodypatients have also reported problems with their hands ldquolock-ingrdquo after making a tight fist [40]This cramping can be pain-ful is most frequent after exercise and grows worse withage The slowed relaxation rates in both Brody patientsand nBmp2NLStm mice suggest the possibility that nBMP2may function in some aspect of the SERCA1 productionacti-vation pathway
Our demonstration of reduced SERCA1 activity inthe nBmp2NLStm mouse supports this hypothesis The phe-notype of the nBmp2NLStm mouse however could also
result from the dysfunction of several other proteins becausemuscle contractionrelaxation cycles are quite complex Con-tractions occur when an action potential leads to activationof Ca2+ release channels (ryanodine receptors) in the SRmembrane Ca2+ floods into the cytoplasm and binds tro-ponin C which modulates the function of tropomyosin sothatmyosin can bind strongly to actin and cause shortening ofthe sarcomere The contraction ceases when active pumping(by SERCA1) of Ca2+ back into the SR depletes cytoplasmicCa2+ levels causing release of Ca2+ from troponin C [4142] The nBmp2NLStm phenotype could be caused by leakyryanodine receptors (RyR) in the SRmembrane whichwouldlead to prolonged elevation of Ca2+ in the cytosol even ifthe SERCA pumps were working normally An increasedaffinity of troponinC for Ca2+ could also cause slowedmusclerelaxation by increasing the level of cytosolic Ca2+ depletionthat would have to occur before Ca2+ was released fromtroponin C Ongoing work in our lab on other physiologicalpathways that are controlled by intracellular Ca2+movementincluding neuronal signaling immune system activation and
10 BioMed Research International
0
25
50
75
100
0 50 100 150
Max
forc
e (
)
Frequency of stimulations (pulsess)
MutantWild type
lowastlowast
lowast
lowast
Figure 7 Force frequency analysis of control and nBmp2NLStmmuscle A significant shift in the force frequency curve was observedin nBmp2NLStm muscle compared to wild type supporting theevidence for slowed relaxation in mutants Data are presented asmean plusmn SEM and lowastdenotes 119875 lt 001 119899 = 6 per group
0
02
04
06
08
1
12
Wild type Mutant
Rela
tive S
ERCA
activ
ity
Figure 8 SERCA activity is reduced in nBmp2NLStm muscle com-pared to controls SR was isolated from control and mutant muscleand Ca2+ ATPase activity wasmeasured Data are presented asmeanplusmn SEM 119875 lt 005 119899 = 3 per group
cell cycle regulation indicates that these pathways are alsodisrupted in nBmp2NLStm mice We predict therefore thatnBMP2 affects aspects of intracellular Ca2+ transport that areshared among multiple pathways rather than those (such astroponin C) that are unique to skeletal muscle
5 Conclusions
In summary the muscle phenotype that we identified inthe nBmp2NLStm mouse suggests that a significant alter-ation in intracellular Ca2+ transport resulted from impedednuclear localization of nBMP2 The prolonged relaxationkinetics the shift in the force frequency curve and the
reduction in SERCA activity are most consistent with acondition of delayed Ca2+ uptake by the SR although otherpossibilities have not been ruled out We have recentlyobserved (unpublished data) that nBmp2NLStm mice alsohave cognitive deficits that are consistent with an intra-cellular Ca2+-handling defect in hippocampus It is likelythat Ca2+ handling is disrupted in other tissues as welland that nBMP2 will be found to function in multiplemolecular pathways that govern intracellular Ca2+ transportThe nBmp2NLStm mouse constitutes a valuable model forcharacterizing the functions of nBMP2 relative to Ca2+transport pathways and for distinguishing those functionsfrom the activities of secreted BMP2
Conflict of Interests
None of the authors have any conflict of interests or compet-ing interests
Acknowledgments
This work was supported by NIAMSNIH Grant AR48839to Laura C Bridgewater Jaime L Mayo was supported by agraduate fellowship from the BrighamYoungUniversity Can-cer ResearchCenterThanks are due toDrMario R Capecchiof the University of Utah for generously providing tech-nical assistance with the stem cell electroporation blastulamicroinjections and embryo transfer steps in production ofthe nBmp2NLStm mouse Recombineering plasmids pGalKpL452 and pL253 were obtained from Neal Copeland andNancy Jenkins of the Methodist Hospital Research Institutein Houston TX
References
[1] M R Urist ldquoBone formation by autoinductionrdquo Science vol150 no 698 pp 893ndash899 1965
[2] J M Wozney V Rosen A J Celeste et al ldquoNovel regulators ofbone formation molecular clones and activitiesrdquo Science vol242 no 4885 pp 1528ndash1534 1988
[3] D ChenM Zhao andG RMundy ldquoBonemorphogenetic pro-teinsrdquo Growth Factors vol 22 no 4 pp 233ndash241 2004
[4] A M Goldstein K C Brewer A M Doyle N Nagy and DJ Roberts ldquoBMP signaling is necessary for neural crest cellmigration and ganglion formation in the enteric nervous sys-temrdquo Mechanisms of Development vol 122 no 6 pp 821ndash8332005
[5] B L Hogan ldquoBone morphogenetic proteins multifunctionalregulators of vertebrate developmentrdquo Genes and Developmentvol 10 no 13 pp 1580ndash1594 1996
[6] YMishina ldquoFunction of bonemorphogenetic protein signalingduring mouse developmentrdquo Frontiers in Bioscience vol 8 ppd855ndashd869 2003
[7] T Schlange H-H Arnold and T Brand ldquoBMP2 is a positiveregulator of Nodal signaling during left-right axis formation inthe chicken embryordquo Development vol 129 no 14 pp 3421ndash3429 2002
[8] H Zhang and A Bradley ldquoMice deficient for BMP2 arenonviable and have defects in amnionchorion and cardiac
BioMed Research International 11
developmentrdquo Development vol 122 no 10 pp 2977ndash29861996
[9] C Kawamura M Kizaki and Y Ikeda ldquoBone morphogeneticprotein (BMP)-2 induces apoptosis in human myeloma cellsrdquoLeukemia and Lymphoma vol 43 no 3 pp 635ndash639 2002
[10] M Raida J H Clement K Ameri C Han R D Leek and A LHarris ldquoExpression of bone morphogenetic protein 2 in breastcancer cells inhibits hypoxic cell deathrdquo International Journal ofOncology vol 26 no 6 pp 1465ndash1470 2005
[11] Y Du and H Yip ldquoEffects of bone morphogenetic protein 2on Id expression and neuroblastoma cell differentiationrdquoDiffer-entiation vol 79 no 2 pp 84ndash92 2010
[12] J-C GuimondM Levesque P-LMichaud et al ldquoBMP-2 func-tions independently of SHH signaling and triggers cell con-densation and apoptosis in regenerating axolotl limbsrdquo BMCDevelopmental Biology vol 10 article 15 2010
[13] C C Rider and B Mulloy ldquoBone morphogenetic protein andgrowth differentiation factor cytokine families and their proteinantagonistsrdquo Biochemical Journal vol 429 no 1 pp 1ndash12 2010
[14] D B Constam and E J Robertson ldquoRegulation of bone mor-phogenetic protein activity by pro domains and proprotein con-vertasesrdquo Journal of Cell Biology vol 144 no 1 pp 139ndash149 1999
[15] J M Wozney ldquoBone morphogenetic proteinsrdquo Progress inGrowth Factor Research vol 1 no 4 pp 267ndash280 1989
[16] A Hiyama S S Gogate S Gajghate J Mochida I M Shapiroand M V Risbud ldquoBMP-2 and TGF-120573 stimulate expression of12057313-glucuronosyl transferase 1 (GlcAT-1) in nucleus pulposuscells through AP1 TonEBP and Sp1 role of MAPKsrdquo Journal ofBone and Mineral Research vol 25 no 5 pp 1179ndash1190 2010
[17] CAlarcon A-I ZaromytidouQXi et al ldquoNuclearCDKs driveSmad transcriptional activation and turnover in BMP andTGF-120573 pathwaysrdquo Cell vol 139 no 4 pp 757ndash769 2009
[18] J Lemonnier C Ghayor J Guicheux and J Caverzasio ldquoPro-tein kinase C-independent activation of protein kinase D isinvolved in BMP-2-induced activation of stress mitogen-acti-vated protein kinases JNK and p38 and osteoblastic cell differ-entiationrdquo Journal of Biological Chemistry vol 279 no 1 pp259ndash264 2004
[19] A Moustakas and C-H Heldin ldquoFrommono- to oligo-Smadsthe heart of thematter inTGF-120573 signal transductionrdquoGenes andDevelopment vol 16 no 15 pp 1867ndash1871 2002
[20] J E Felin J L Mayo T J Loos et al ldquoNuclear variants of bonemorphogenetic proteinsrdquo BMC Cell Biology vol 11 no 1 article20 2010
[21] N G Copeland N A Jenkins and D L Court ldquoRecombi-neering a powerful new tool for mouse functional genomicsrdquoNature Reviews Genetics vol 2 no 10 pp 769ndash779 2001
[22] S Warming N Costantino D L Court N A Jenkins and NG Copeland ldquoSimple and highly efficient BAC recombineeringusing galK selectionrdquo Nucleic Acids Research vol 33 no 4article e36 2005
[23] M Bunting K E Bernstein J M Greer M R Capecchi and KR Thomas ldquoTargeting genes for self-excision in the germ linerdquoGenes and Development vol 13 no 12 pp 1524ndash1528 1999
[24] S H George M Gertsenstein K Vintersten et al ldquoDevelop-mental and adult phenotyping directly frommutant embryonicstemcellsrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol 104 no 11 pp 4455ndash4460 2007
[25] S L Mansour K R Thomas and M R Capecchi ldquoDisruptionof the proto-oncogene int-2 in mouse embryo-derived stemcells a general strategy for targeting mutations to non-select-able genesrdquo Nature vol 336 no 6197 pp 348ndash352 1988
[26] V E Papaioannou and R R Behringer Mouse PhenotypesmdashAHandbook of Mutation Analysis Cold Spring Harbor Labora-tory Press Cold Spring Harbor NY USA 2005
[27] C R Hancock J J Brault RWWiseman R L Terjung and RA Meyer ldquo31P-NMR observation of free ADP during fatiguingrepetitive contractions of murine skeletal muscle lacking AK1rdquoThe American Journal of Physiology vol 288 no 6 pp C1298ndashC1304 2005
[28] C R Hancock E Janssen and R L Terjung ldquoSkeletal musclecontractile performance and ADP accumulation in adenylatekinase-deficient micerdquoThe American Journal of Physiology vol288 no 6 pp C1287ndashC1297 2005
[29] C R Hancock E Janssen and R L Terjung ldquoContraction-mediated phosphorylation of AMPK is lower in skeletal muscleof adenylate kinase-deficient micerdquo Journal of Applied Physiol-ogy vol 100 no 2 pp 406ndash413 2006
[30] D Kosk-Kosicka ldquoCalcium signaling protocolsrdquo in Methods inMolecular Biology D G Lambert Ed vol 114 pp 343ndash354Springer New York NY USA 1999
[31] F Hillger G Herr R Rudolph and E Schwarz ldquoBiophysicalcomparison of BMP-2 ProBMP-2 and the free pro-peptidereveals stabilization of the pro-peptide by the mature growthfactorrdquo Journal of Biological Chemistry vol 280 no 15 pp14974ndash14980 2005
[32] BOhkawara S Iemura P tenDijke andNUeno ldquoAction rangeof BMP is defined by its N-terminal basic amino acid corerdquoCurrent Biology vol 12 no 3 pp 205ndash209 2002
[33] E M de Robertis ldquoSpemannrsquos organizer and the self-regulationof embryonic fieldsrdquo Mechanisms of Development vol 126 no11-12 pp 925ndash941 2009
[34] D M Maatouk K-S Choi C M Bouldin and B D Harfe ldquoInthe limb AER Bmp2 and Bmp4 are required for dorsal-ventralpatterning and interdigital cell death but not limb outgrowthrdquoDevelopmental Biology vol 327 no 2 pp 516ndash523 2009
[35] B Robert ldquoBone morphogenetic protein signaling in limboutgrowth and patterningrdquo Development Growth and Differen-tiation vol 49 no 6 pp 455ndash468 2007
[36] A Odermatt K Barton V K Khanna et al ldquoThe mutation ofPro789 to Leu reduces the activity of the fast-twitch skeletalmuscle sarco(endo)plasmic reticulum Ca2+ ATPase (SERCA1)and is associated with Brody diseaserdquoHuman Genetics vol 106no 5 pp 482ndash491 2000
[37] A Odermatt P E Taschner V K Khanna et al ldquoMutationsin the gene-encoding SERCA1 the fast-twitch skeletal musclesarcoplasmic reticulumCa2+ ATPase are associated with Brodydiseaserdquo Nature Genetics vol 14 no 2 pp 191ndash194 1996
[38] A A Benders J H Veerkamp A Oosterhof et al ldquoCa2+ home-ostasis in Brodyrsquos disease A study in skeletal muscle and cul-tured muscle cells and the effects of dantrolene and verapamilrdquoJournal of Clinical Investigation vol 94 no 2 pp 741ndash748 1994
[39] I A Brody ldquoMuscle contracture induced by exercise Asyndrome attributable to decreased relaxing factorrdquo The NewEngland Journal of Medicine vol 281 no 4 pp 187ndash192 1969
[40] Y Zhang J Fujii M S Phillips et al ldquoCharacterization ofcDNA and genomic DNA encoding SERCA1 the Ca2+-ATPaseof human fast-twitch skeletal muscle sarcoplasmic reticulumand its elimination as a candidate gene for Brody diseaserdquoGenomics vol 30 no 3 pp 415ndash424 1995
[41] M Endo ldquoCalcium-induced calcium release in skeletal musclerdquoPhysiological Reviews vol 89 no 4 pp 1153ndash1176 2009
[42] M J Betzenhauser and A RMarks ldquoRyanodine receptor chan-nelopathiesrdquo Pflugers Archiv vol 460 no 2 pp 467ndash480 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
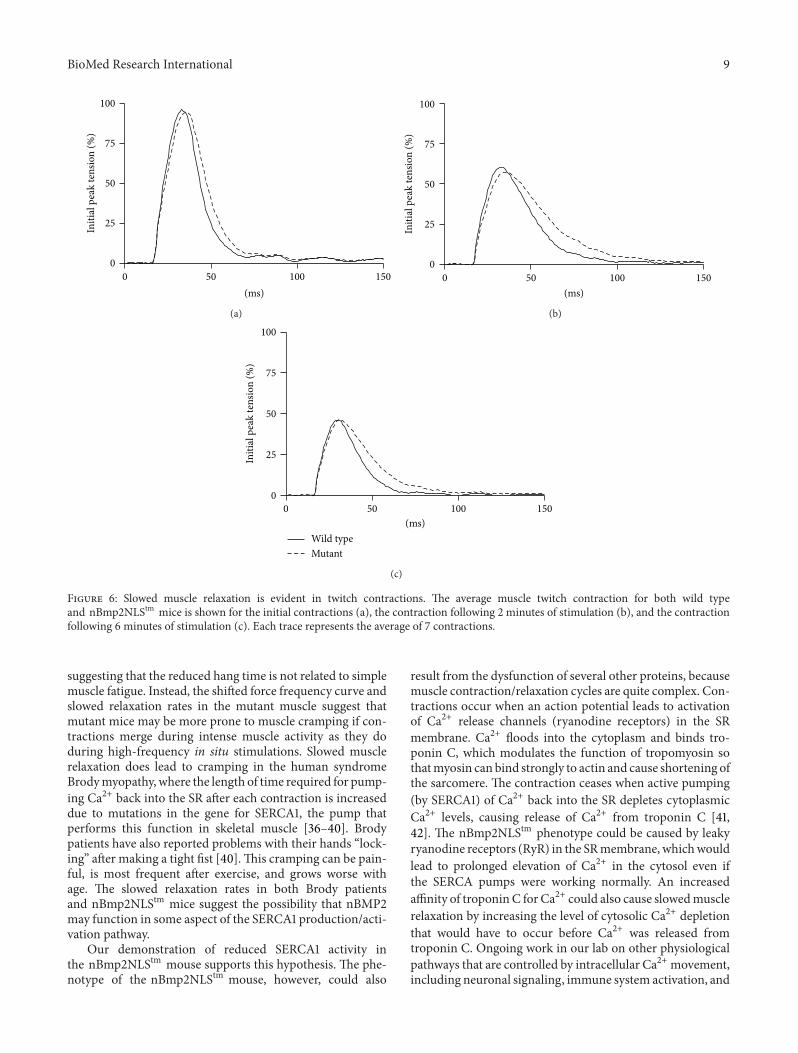
BioMed Research International 9
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(a)
0
25
50
75
100
0 50 100 150
Initi
al p
eak
tens
ion
()
(ms)
(b)
Wild typeMutant
0
25
50
75
100
0 50 100 150(ms)
Initi
al p
eak
tens
ion
()
(c)
Figure 6 Slowed muscle relaxation is evident in twitch contractions The average muscle twitch contraction for both wild typeand nBmp2NLStm mice is shown for the initial contractions (a) the contraction following 2 minutes of stimulation (b) and the contractionfollowing 6 minutes of stimulation (c) Each trace represents the average of 7 contractions
suggesting that the reduced hang time is not related to simplemuscle fatigue Instead the shifted force frequency curve andslowed relaxation rates in the mutant muscle suggest thatmutant mice may be more prone to muscle cramping if con-tractions merge during intense muscle activity as they doduring high-frequency in situ stimulations Slowed musclerelaxation does lead to cramping in the human syndromeBrodymyopathy where the length of time required for pump-ing Ca2+ back into the SR after each contraction is increaseddue to mutations in the gene for SERCA1 the pump thatperforms this function in skeletal muscle [36ndash40] Brodypatients have also reported problems with their hands ldquolock-ingrdquo after making a tight fist [40]This cramping can be pain-ful is most frequent after exercise and grows worse withage The slowed relaxation rates in both Brody patientsand nBmp2NLStm mice suggest the possibility that nBMP2may function in some aspect of the SERCA1 productionacti-vation pathway
Our demonstration of reduced SERCA1 activity inthe nBmp2NLStm mouse supports this hypothesis The phe-notype of the nBmp2NLStm mouse however could also
result from the dysfunction of several other proteins becausemuscle contractionrelaxation cycles are quite complex Con-tractions occur when an action potential leads to activationof Ca2+ release channels (ryanodine receptors) in the SRmembrane Ca2+ floods into the cytoplasm and binds tro-ponin C which modulates the function of tropomyosin sothatmyosin can bind strongly to actin and cause shortening ofthe sarcomere The contraction ceases when active pumping(by SERCA1) of Ca2+ back into the SR depletes cytoplasmicCa2+ levels causing release of Ca2+ from troponin C [4142] The nBmp2NLStm phenotype could be caused by leakyryanodine receptors (RyR) in the SRmembrane whichwouldlead to prolonged elevation of Ca2+ in the cytosol even ifthe SERCA pumps were working normally An increasedaffinity of troponinC for Ca2+ could also cause slowedmusclerelaxation by increasing the level of cytosolic Ca2+ depletionthat would have to occur before Ca2+ was released fromtroponin C Ongoing work in our lab on other physiologicalpathways that are controlled by intracellular Ca2+movementincluding neuronal signaling immune system activation and
10 BioMed Research International
0
25
50
75
100
0 50 100 150
Max
forc
e (
)
Frequency of stimulations (pulsess)
MutantWild type
lowastlowast
lowast
lowast
Figure 7 Force frequency analysis of control and nBmp2NLStmmuscle A significant shift in the force frequency curve was observedin nBmp2NLStm muscle compared to wild type supporting theevidence for slowed relaxation in mutants Data are presented asmean plusmn SEM and lowastdenotes 119875 lt 001 119899 = 6 per group
0
02
04
06
08
1
12
Wild type Mutant
Rela
tive S
ERCA
activ
ity
Figure 8 SERCA activity is reduced in nBmp2NLStm muscle com-pared to controls SR was isolated from control and mutant muscleand Ca2+ ATPase activity wasmeasured Data are presented asmeanplusmn SEM 119875 lt 005 119899 = 3 per group
cell cycle regulation indicates that these pathways are alsodisrupted in nBmp2NLStm mice We predict therefore thatnBMP2 affects aspects of intracellular Ca2+ transport that areshared among multiple pathways rather than those (such astroponin C) that are unique to skeletal muscle
5 Conclusions
In summary the muscle phenotype that we identified inthe nBmp2NLStm mouse suggests that a significant alter-ation in intracellular Ca2+ transport resulted from impedednuclear localization of nBMP2 The prolonged relaxationkinetics the shift in the force frequency curve and the
reduction in SERCA activity are most consistent with acondition of delayed Ca2+ uptake by the SR although otherpossibilities have not been ruled out We have recentlyobserved (unpublished data) that nBmp2NLStm mice alsohave cognitive deficits that are consistent with an intra-cellular Ca2+-handling defect in hippocampus It is likelythat Ca2+ handling is disrupted in other tissues as welland that nBMP2 will be found to function in multiplemolecular pathways that govern intracellular Ca2+ transportThe nBmp2NLStm mouse constitutes a valuable model forcharacterizing the functions of nBMP2 relative to Ca2+transport pathways and for distinguishing those functionsfrom the activities of secreted BMP2
Conflict of Interests
None of the authors have any conflict of interests or compet-ing interests
Acknowledgments
This work was supported by NIAMSNIH Grant AR48839to Laura C Bridgewater Jaime L Mayo was supported by agraduate fellowship from the BrighamYoungUniversity Can-cer ResearchCenterThanks are due toDrMario R Capecchiof the University of Utah for generously providing tech-nical assistance with the stem cell electroporation blastulamicroinjections and embryo transfer steps in production ofthe nBmp2NLStm mouse Recombineering plasmids pGalKpL452 and pL253 were obtained from Neal Copeland andNancy Jenkins of the Methodist Hospital Research Institutein Houston TX
References
[1] M R Urist ldquoBone formation by autoinductionrdquo Science vol150 no 698 pp 893ndash899 1965
[2] J M Wozney V Rosen A J Celeste et al ldquoNovel regulators ofbone formation molecular clones and activitiesrdquo Science vol242 no 4885 pp 1528ndash1534 1988
[3] D ChenM Zhao andG RMundy ldquoBonemorphogenetic pro-teinsrdquo Growth Factors vol 22 no 4 pp 233ndash241 2004
[4] A M Goldstein K C Brewer A M Doyle N Nagy and DJ Roberts ldquoBMP signaling is necessary for neural crest cellmigration and ganglion formation in the enteric nervous sys-temrdquo Mechanisms of Development vol 122 no 6 pp 821ndash8332005
[5] B L Hogan ldquoBone morphogenetic proteins multifunctionalregulators of vertebrate developmentrdquo Genes and Developmentvol 10 no 13 pp 1580ndash1594 1996
[6] YMishina ldquoFunction of bonemorphogenetic protein signalingduring mouse developmentrdquo Frontiers in Bioscience vol 8 ppd855ndashd869 2003
[7] T Schlange H-H Arnold and T Brand ldquoBMP2 is a positiveregulator of Nodal signaling during left-right axis formation inthe chicken embryordquo Development vol 129 no 14 pp 3421ndash3429 2002
[8] H Zhang and A Bradley ldquoMice deficient for BMP2 arenonviable and have defects in amnionchorion and cardiac
BioMed Research International 11
developmentrdquo Development vol 122 no 10 pp 2977ndash29861996
[9] C Kawamura M Kizaki and Y Ikeda ldquoBone morphogeneticprotein (BMP)-2 induces apoptosis in human myeloma cellsrdquoLeukemia and Lymphoma vol 43 no 3 pp 635ndash639 2002
[10] M Raida J H Clement K Ameri C Han R D Leek and A LHarris ldquoExpression of bone morphogenetic protein 2 in breastcancer cells inhibits hypoxic cell deathrdquo International Journal ofOncology vol 26 no 6 pp 1465ndash1470 2005
[11] Y Du and H Yip ldquoEffects of bone morphogenetic protein 2on Id expression and neuroblastoma cell differentiationrdquoDiffer-entiation vol 79 no 2 pp 84ndash92 2010
[12] J-C GuimondM Levesque P-LMichaud et al ldquoBMP-2 func-tions independently of SHH signaling and triggers cell con-densation and apoptosis in regenerating axolotl limbsrdquo BMCDevelopmental Biology vol 10 article 15 2010
[13] C C Rider and B Mulloy ldquoBone morphogenetic protein andgrowth differentiation factor cytokine families and their proteinantagonistsrdquo Biochemical Journal vol 429 no 1 pp 1ndash12 2010
[14] D B Constam and E J Robertson ldquoRegulation of bone mor-phogenetic protein activity by pro domains and proprotein con-vertasesrdquo Journal of Cell Biology vol 144 no 1 pp 139ndash149 1999
[15] J M Wozney ldquoBone morphogenetic proteinsrdquo Progress inGrowth Factor Research vol 1 no 4 pp 267ndash280 1989
[16] A Hiyama S S Gogate S Gajghate J Mochida I M Shapiroand M V Risbud ldquoBMP-2 and TGF-120573 stimulate expression of12057313-glucuronosyl transferase 1 (GlcAT-1) in nucleus pulposuscells through AP1 TonEBP and Sp1 role of MAPKsrdquo Journal ofBone and Mineral Research vol 25 no 5 pp 1179ndash1190 2010
[17] CAlarcon A-I ZaromytidouQXi et al ldquoNuclearCDKs driveSmad transcriptional activation and turnover in BMP andTGF-120573 pathwaysrdquo Cell vol 139 no 4 pp 757ndash769 2009
[18] J Lemonnier C Ghayor J Guicheux and J Caverzasio ldquoPro-tein kinase C-independent activation of protein kinase D isinvolved in BMP-2-induced activation of stress mitogen-acti-vated protein kinases JNK and p38 and osteoblastic cell differ-entiationrdquo Journal of Biological Chemistry vol 279 no 1 pp259ndash264 2004
[19] A Moustakas and C-H Heldin ldquoFrommono- to oligo-Smadsthe heart of thematter inTGF-120573 signal transductionrdquoGenes andDevelopment vol 16 no 15 pp 1867ndash1871 2002
[20] J E Felin J L Mayo T J Loos et al ldquoNuclear variants of bonemorphogenetic proteinsrdquo BMC Cell Biology vol 11 no 1 article20 2010
[21] N G Copeland N A Jenkins and D L Court ldquoRecombi-neering a powerful new tool for mouse functional genomicsrdquoNature Reviews Genetics vol 2 no 10 pp 769ndash779 2001
[22] S Warming N Costantino D L Court N A Jenkins and NG Copeland ldquoSimple and highly efficient BAC recombineeringusing galK selectionrdquo Nucleic Acids Research vol 33 no 4article e36 2005
[23] M Bunting K E Bernstein J M Greer M R Capecchi and KR Thomas ldquoTargeting genes for self-excision in the germ linerdquoGenes and Development vol 13 no 12 pp 1524ndash1528 1999
[24] S H George M Gertsenstein K Vintersten et al ldquoDevelop-mental and adult phenotyping directly frommutant embryonicstemcellsrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol 104 no 11 pp 4455ndash4460 2007
[25] S L Mansour K R Thomas and M R Capecchi ldquoDisruptionof the proto-oncogene int-2 in mouse embryo-derived stemcells a general strategy for targeting mutations to non-select-able genesrdquo Nature vol 336 no 6197 pp 348ndash352 1988
[26] V E Papaioannou and R R Behringer Mouse PhenotypesmdashAHandbook of Mutation Analysis Cold Spring Harbor Labora-tory Press Cold Spring Harbor NY USA 2005
[27] C R Hancock J J Brault RWWiseman R L Terjung and RA Meyer ldquo31P-NMR observation of free ADP during fatiguingrepetitive contractions of murine skeletal muscle lacking AK1rdquoThe American Journal of Physiology vol 288 no 6 pp C1298ndashC1304 2005
[28] C R Hancock E Janssen and R L Terjung ldquoSkeletal musclecontractile performance and ADP accumulation in adenylatekinase-deficient micerdquoThe American Journal of Physiology vol288 no 6 pp C1287ndashC1297 2005
[29] C R Hancock E Janssen and R L Terjung ldquoContraction-mediated phosphorylation of AMPK is lower in skeletal muscleof adenylate kinase-deficient micerdquo Journal of Applied Physiol-ogy vol 100 no 2 pp 406ndash413 2006
[30] D Kosk-Kosicka ldquoCalcium signaling protocolsrdquo in Methods inMolecular Biology D G Lambert Ed vol 114 pp 343ndash354Springer New York NY USA 1999
[31] F Hillger G Herr R Rudolph and E Schwarz ldquoBiophysicalcomparison of BMP-2 ProBMP-2 and the free pro-peptidereveals stabilization of the pro-peptide by the mature growthfactorrdquo Journal of Biological Chemistry vol 280 no 15 pp14974ndash14980 2005
[32] BOhkawara S Iemura P tenDijke andNUeno ldquoAction rangeof BMP is defined by its N-terminal basic amino acid corerdquoCurrent Biology vol 12 no 3 pp 205ndash209 2002
[33] E M de Robertis ldquoSpemannrsquos organizer and the self-regulationof embryonic fieldsrdquo Mechanisms of Development vol 126 no11-12 pp 925ndash941 2009
[34] D M Maatouk K-S Choi C M Bouldin and B D Harfe ldquoInthe limb AER Bmp2 and Bmp4 are required for dorsal-ventralpatterning and interdigital cell death but not limb outgrowthrdquoDevelopmental Biology vol 327 no 2 pp 516ndash523 2009
[35] B Robert ldquoBone morphogenetic protein signaling in limboutgrowth and patterningrdquo Development Growth and Differen-tiation vol 49 no 6 pp 455ndash468 2007
[36] A Odermatt K Barton V K Khanna et al ldquoThe mutation ofPro789 to Leu reduces the activity of the fast-twitch skeletalmuscle sarco(endo)plasmic reticulum Ca2+ ATPase (SERCA1)and is associated with Brody diseaserdquoHuman Genetics vol 106no 5 pp 482ndash491 2000
[37] A Odermatt P E Taschner V K Khanna et al ldquoMutationsin the gene-encoding SERCA1 the fast-twitch skeletal musclesarcoplasmic reticulumCa2+ ATPase are associated with Brodydiseaserdquo Nature Genetics vol 14 no 2 pp 191ndash194 1996
[38] A A Benders J H Veerkamp A Oosterhof et al ldquoCa2+ home-ostasis in Brodyrsquos disease A study in skeletal muscle and cul-tured muscle cells and the effects of dantrolene and verapamilrdquoJournal of Clinical Investigation vol 94 no 2 pp 741ndash748 1994
[39] I A Brody ldquoMuscle contracture induced by exercise Asyndrome attributable to decreased relaxing factorrdquo The NewEngland Journal of Medicine vol 281 no 4 pp 187ndash192 1969
[40] Y Zhang J Fujii M S Phillips et al ldquoCharacterization ofcDNA and genomic DNA encoding SERCA1 the Ca2+-ATPaseof human fast-twitch skeletal muscle sarcoplasmic reticulumand its elimination as a candidate gene for Brody diseaserdquoGenomics vol 30 no 3 pp 415ndash424 1995
[41] M Endo ldquoCalcium-induced calcium release in skeletal musclerdquoPhysiological Reviews vol 89 no 4 pp 1153ndash1176 2009
[42] M J Betzenhauser and A RMarks ldquoRyanodine receptor chan-nelopathiesrdquo Pflugers Archiv vol 460 no 2 pp 467ndash480 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
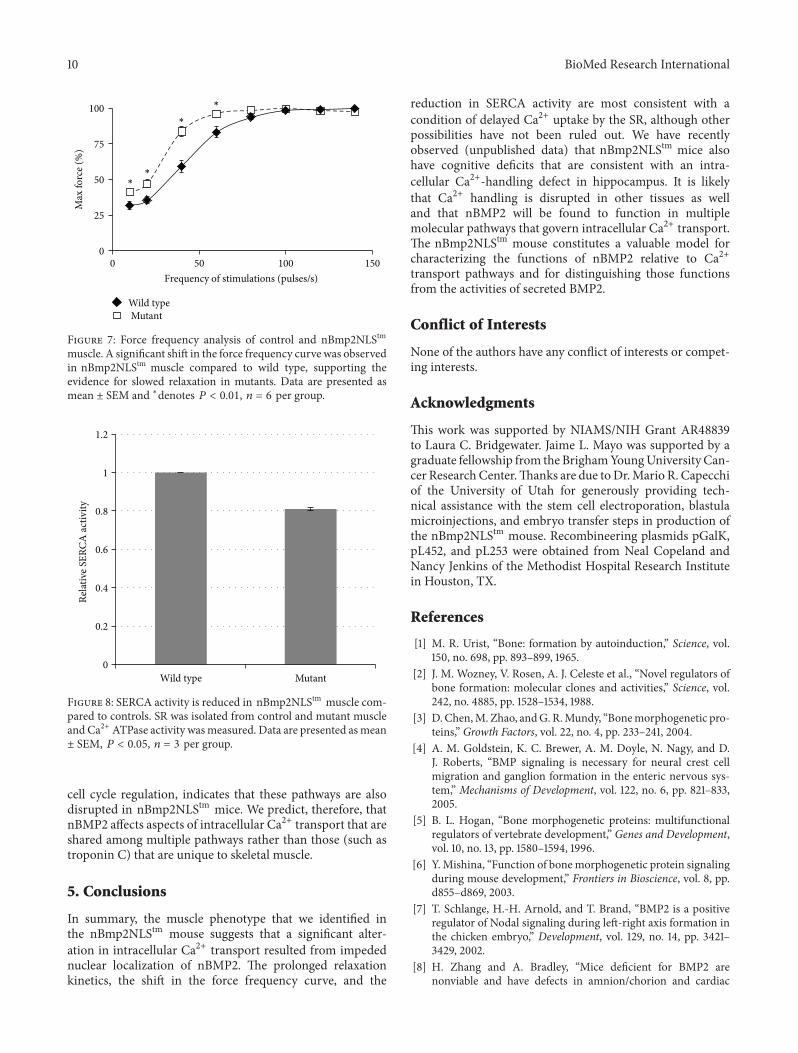
10 BioMed Research International
0
25
50
75
100
0 50 100 150
Max
forc
e (
)
Frequency of stimulations (pulsess)
MutantWild type
lowastlowast
lowast
lowast
Figure 7 Force frequency analysis of control and nBmp2NLStmmuscle A significant shift in the force frequency curve was observedin nBmp2NLStm muscle compared to wild type supporting theevidence for slowed relaxation in mutants Data are presented asmean plusmn SEM and lowastdenotes 119875 lt 001 119899 = 6 per group
0
02
04
06
08
1
12
Wild type Mutant
Rela
tive S
ERCA
activ
ity
Figure 8 SERCA activity is reduced in nBmp2NLStm muscle com-pared to controls SR was isolated from control and mutant muscleand Ca2+ ATPase activity wasmeasured Data are presented asmeanplusmn SEM 119875 lt 005 119899 = 3 per group
cell cycle regulation indicates that these pathways are alsodisrupted in nBmp2NLStm mice We predict therefore thatnBMP2 affects aspects of intracellular Ca2+ transport that areshared among multiple pathways rather than those (such astroponin C) that are unique to skeletal muscle
5 Conclusions
In summary the muscle phenotype that we identified inthe nBmp2NLStm mouse suggests that a significant alter-ation in intracellular Ca2+ transport resulted from impedednuclear localization of nBMP2 The prolonged relaxationkinetics the shift in the force frequency curve and the
reduction in SERCA activity are most consistent with acondition of delayed Ca2+ uptake by the SR although otherpossibilities have not been ruled out We have recentlyobserved (unpublished data) that nBmp2NLStm mice alsohave cognitive deficits that are consistent with an intra-cellular Ca2+-handling defect in hippocampus It is likelythat Ca2+ handling is disrupted in other tissues as welland that nBMP2 will be found to function in multiplemolecular pathways that govern intracellular Ca2+ transportThe nBmp2NLStm mouse constitutes a valuable model forcharacterizing the functions of nBMP2 relative to Ca2+transport pathways and for distinguishing those functionsfrom the activities of secreted BMP2
Conflict of Interests
None of the authors have any conflict of interests or compet-ing interests
Acknowledgments
This work was supported by NIAMSNIH Grant AR48839to Laura C Bridgewater Jaime L Mayo was supported by agraduate fellowship from the BrighamYoungUniversity Can-cer ResearchCenterThanks are due toDrMario R Capecchiof the University of Utah for generously providing tech-nical assistance with the stem cell electroporation blastulamicroinjections and embryo transfer steps in production ofthe nBmp2NLStm mouse Recombineering plasmids pGalKpL452 and pL253 were obtained from Neal Copeland andNancy Jenkins of the Methodist Hospital Research Institutein Houston TX
References
[1] M R Urist ldquoBone formation by autoinductionrdquo Science vol150 no 698 pp 893ndash899 1965
[2] J M Wozney V Rosen A J Celeste et al ldquoNovel regulators ofbone formation molecular clones and activitiesrdquo Science vol242 no 4885 pp 1528ndash1534 1988
[3] D ChenM Zhao andG RMundy ldquoBonemorphogenetic pro-teinsrdquo Growth Factors vol 22 no 4 pp 233ndash241 2004
[4] A M Goldstein K C Brewer A M Doyle N Nagy and DJ Roberts ldquoBMP signaling is necessary for neural crest cellmigration and ganglion formation in the enteric nervous sys-temrdquo Mechanisms of Development vol 122 no 6 pp 821ndash8332005
[5] B L Hogan ldquoBone morphogenetic proteins multifunctionalregulators of vertebrate developmentrdquo Genes and Developmentvol 10 no 13 pp 1580ndash1594 1996
[6] YMishina ldquoFunction of bonemorphogenetic protein signalingduring mouse developmentrdquo Frontiers in Bioscience vol 8 ppd855ndashd869 2003
[7] T Schlange H-H Arnold and T Brand ldquoBMP2 is a positiveregulator of Nodal signaling during left-right axis formation inthe chicken embryordquo Development vol 129 no 14 pp 3421ndash3429 2002
[8] H Zhang and A Bradley ldquoMice deficient for BMP2 arenonviable and have defects in amnionchorion and cardiac
BioMed Research International 11
developmentrdquo Development vol 122 no 10 pp 2977ndash29861996
[9] C Kawamura M Kizaki and Y Ikeda ldquoBone morphogeneticprotein (BMP)-2 induces apoptosis in human myeloma cellsrdquoLeukemia and Lymphoma vol 43 no 3 pp 635ndash639 2002
[10] M Raida J H Clement K Ameri C Han R D Leek and A LHarris ldquoExpression of bone morphogenetic protein 2 in breastcancer cells inhibits hypoxic cell deathrdquo International Journal ofOncology vol 26 no 6 pp 1465ndash1470 2005
[11] Y Du and H Yip ldquoEffects of bone morphogenetic protein 2on Id expression and neuroblastoma cell differentiationrdquoDiffer-entiation vol 79 no 2 pp 84ndash92 2010
[12] J-C GuimondM Levesque P-LMichaud et al ldquoBMP-2 func-tions independently of SHH signaling and triggers cell con-densation and apoptosis in regenerating axolotl limbsrdquo BMCDevelopmental Biology vol 10 article 15 2010
[13] C C Rider and B Mulloy ldquoBone morphogenetic protein andgrowth differentiation factor cytokine families and their proteinantagonistsrdquo Biochemical Journal vol 429 no 1 pp 1ndash12 2010
[14] D B Constam and E J Robertson ldquoRegulation of bone mor-phogenetic protein activity by pro domains and proprotein con-vertasesrdquo Journal of Cell Biology vol 144 no 1 pp 139ndash149 1999
[15] J M Wozney ldquoBone morphogenetic proteinsrdquo Progress inGrowth Factor Research vol 1 no 4 pp 267ndash280 1989
[16] A Hiyama S S Gogate S Gajghate J Mochida I M Shapiroand M V Risbud ldquoBMP-2 and TGF-120573 stimulate expression of12057313-glucuronosyl transferase 1 (GlcAT-1) in nucleus pulposuscells through AP1 TonEBP and Sp1 role of MAPKsrdquo Journal ofBone and Mineral Research vol 25 no 5 pp 1179ndash1190 2010
[17] CAlarcon A-I ZaromytidouQXi et al ldquoNuclearCDKs driveSmad transcriptional activation and turnover in BMP andTGF-120573 pathwaysrdquo Cell vol 139 no 4 pp 757ndash769 2009
[18] J Lemonnier C Ghayor J Guicheux and J Caverzasio ldquoPro-tein kinase C-independent activation of protein kinase D isinvolved in BMP-2-induced activation of stress mitogen-acti-vated protein kinases JNK and p38 and osteoblastic cell differ-entiationrdquo Journal of Biological Chemistry vol 279 no 1 pp259ndash264 2004
[19] A Moustakas and C-H Heldin ldquoFrommono- to oligo-Smadsthe heart of thematter inTGF-120573 signal transductionrdquoGenes andDevelopment vol 16 no 15 pp 1867ndash1871 2002
[20] J E Felin J L Mayo T J Loos et al ldquoNuclear variants of bonemorphogenetic proteinsrdquo BMC Cell Biology vol 11 no 1 article20 2010
[21] N G Copeland N A Jenkins and D L Court ldquoRecombi-neering a powerful new tool for mouse functional genomicsrdquoNature Reviews Genetics vol 2 no 10 pp 769ndash779 2001
[22] S Warming N Costantino D L Court N A Jenkins and NG Copeland ldquoSimple and highly efficient BAC recombineeringusing galK selectionrdquo Nucleic Acids Research vol 33 no 4article e36 2005
[23] M Bunting K E Bernstein J M Greer M R Capecchi and KR Thomas ldquoTargeting genes for self-excision in the germ linerdquoGenes and Development vol 13 no 12 pp 1524ndash1528 1999
[24] S H George M Gertsenstein K Vintersten et al ldquoDevelop-mental and adult phenotyping directly frommutant embryonicstemcellsrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol 104 no 11 pp 4455ndash4460 2007
[25] S L Mansour K R Thomas and M R Capecchi ldquoDisruptionof the proto-oncogene int-2 in mouse embryo-derived stemcells a general strategy for targeting mutations to non-select-able genesrdquo Nature vol 336 no 6197 pp 348ndash352 1988
[26] V E Papaioannou and R R Behringer Mouse PhenotypesmdashAHandbook of Mutation Analysis Cold Spring Harbor Labora-tory Press Cold Spring Harbor NY USA 2005
[27] C R Hancock J J Brault RWWiseman R L Terjung and RA Meyer ldquo31P-NMR observation of free ADP during fatiguingrepetitive contractions of murine skeletal muscle lacking AK1rdquoThe American Journal of Physiology vol 288 no 6 pp C1298ndashC1304 2005
[28] C R Hancock E Janssen and R L Terjung ldquoSkeletal musclecontractile performance and ADP accumulation in adenylatekinase-deficient micerdquoThe American Journal of Physiology vol288 no 6 pp C1287ndashC1297 2005
[29] C R Hancock E Janssen and R L Terjung ldquoContraction-mediated phosphorylation of AMPK is lower in skeletal muscleof adenylate kinase-deficient micerdquo Journal of Applied Physiol-ogy vol 100 no 2 pp 406ndash413 2006
[30] D Kosk-Kosicka ldquoCalcium signaling protocolsrdquo in Methods inMolecular Biology D G Lambert Ed vol 114 pp 343ndash354Springer New York NY USA 1999
[31] F Hillger G Herr R Rudolph and E Schwarz ldquoBiophysicalcomparison of BMP-2 ProBMP-2 and the free pro-peptidereveals stabilization of the pro-peptide by the mature growthfactorrdquo Journal of Biological Chemistry vol 280 no 15 pp14974ndash14980 2005
[32] BOhkawara S Iemura P tenDijke andNUeno ldquoAction rangeof BMP is defined by its N-terminal basic amino acid corerdquoCurrent Biology vol 12 no 3 pp 205ndash209 2002
[33] E M de Robertis ldquoSpemannrsquos organizer and the self-regulationof embryonic fieldsrdquo Mechanisms of Development vol 126 no11-12 pp 925ndash941 2009
[34] D M Maatouk K-S Choi C M Bouldin and B D Harfe ldquoInthe limb AER Bmp2 and Bmp4 are required for dorsal-ventralpatterning and interdigital cell death but not limb outgrowthrdquoDevelopmental Biology vol 327 no 2 pp 516ndash523 2009
[35] B Robert ldquoBone morphogenetic protein signaling in limboutgrowth and patterningrdquo Development Growth and Differen-tiation vol 49 no 6 pp 455ndash468 2007
[36] A Odermatt K Barton V K Khanna et al ldquoThe mutation ofPro789 to Leu reduces the activity of the fast-twitch skeletalmuscle sarco(endo)plasmic reticulum Ca2+ ATPase (SERCA1)and is associated with Brody diseaserdquoHuman Genetics vol 106no 5 pp 482ndash491 2000
[37] A Odermatt P E Taschner V K Khanna et al ldquoMutationsin the gene-encoding SERCA1 the fast-twitch skeletal musclesarcoplasmic reticulumCa2+ ATPase are associated with Brodydiseaserdquo Nature Genetics vol 14 no 2 pp 191ndash194 1996
[38] A A Benders J H Veerkamp A Oosterhof et al ldquoCa2+ home-ostasis in Brodyrsquos disease A study in skeletal muscle and cul-tured muscle cells and the effects of dantrolene and verapamilrdquoJournal of Clinical Investigation vol 94 no 2 pp 741ndash748 1994
[39] I A Brody ldquoMuscle contracture induced by exercise Asyndrome attributable to decreased relaxing factorrdquo The NewEngland Journal of Medicine vol 281 no 4 pp 187ndash192 1969
[40] Y Zhang J Fujii M S Phillips et al ldquoCharacterization ofcDNA and genomic DNA encoding SERCA1 the Ca2+-ATPaseof human fast-twitch skeletal muscle sarcoplasmic reticulumand its elimination as a candidate gene for Brody diseaserdquoGenomics vol 30 no 3 pp 415ndash424 1995
[41] M Endo ldquoCalcium-induced calcium release in skeletal musclerdquoPhysiological Reviews vol 89 no 4 pp 1153ndash1176 2009
[42] M J Betzenhauser and A RMarks ldquoRyanodine receptor chan-nelopathiesrdquo Pflugers Archiv vol 460 no 2 pp 467ndash480 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013

BioMed Research International 11
developmentrdquo Development vol 122 no 10 pp 2977ndash29861996
[9] C Kawamura M Kizaki and Y Ikeda ldquoBone morphogeneticprotein (BMP)-2 induces apoptosis in human myeloma cellsrdquoLeukemia and Lymphoma vol 43 no 3 pp 635ndash639 2002
[10] M Raida J H Clement K Ameri C Han R D Leek and A LHarris ldquoExpression of bone morphogenetic protein 2 in breastcancer cells inhibits hypoxic cell deathrdquo International Journal ofOncology vol 26 no 6 pp 1465ndash1470 2005
[11] Y Du and H Yip ldquoEffects of bone morphogenetic protein 2on Id expression and neuroblastoma cell differentiationrdquoDiffer-entiation vol 79 no 2 pp 84ndash92 2010
[12] J-C GuimondM Levesque P-LMichaud et al ldquoBMP-2 func-tions independently of SHH signaling and triggers cell con-densation and apoptosis in regenerating axolotl limbsrdquo BMCDevelopmental Biology vol 10 article 15 2010
[13] C C Rider and B Mulloy ldquoBone morphogenetic protein andgrowth differentiation factor cytokine families and their proteinantagonistsrdquo Biochemical Journal vol 429 no 1 pp 1ndash12 2010
[14] D B Constam and E J Robertson ldquoRegulation of bone mor-phogenetic protein activity by pro domains and proprotein con-vertasesrdquo Journal of Cell Biology vol 144 no 1 pp 139ndash149 1999
[15] J M Wozney ldquoBone morphogenetic proteinsrdquo Progress inGrowth Factor Research vol 1 no 4 pp 267ndash280 1989
[16] A Hiyama S S Gogate S Gajghate J Mochida I M Shapiroand M V Risbud ldquoBMP-2 and TGF-120573 stimulate expression of12057313-glucuronosyl transferase 1 (GlcAT-1) in nucleus pulposuscells through AP1 TonEBP and Sp1 role of MAPKsrdquo Journal ofBone and Mineral Research vol 25 no 5 pp 1179ndash1190 2010
[17] CAlarcon A-I ZaromytidouQXi et al ldquoNuclearCDKs driveSmad transcriptional activation and turnover in BMP andTGF-120573 pathwaysrdquo Cell vol 139 no 4 pp 757ndash769 2009
[18] J Lemonnier C Ghayor J Guicheux and J Caverzasio ldquoPro-tein kinase C-independent activation of protein kinase D isinvolved in BMP-2-induced activation of stress mitogen-acti-vated protein kinases JNK and p38 and osteoblastic cell differ-entiationrdquo Journal of Biological Chemistry vol 279 no 1 pp259ndash264 2004
[19] A Moustakas and C-H Heldin ldquoFrommono- to oligo-Smadsthe heart of thematter inTGF-120573 signal transductionrdquoGenes andDevelopment vol 16 no 15 pp 1867ndash1871 2002
[20] J E Felin J L Mayo T J Loos et al ldquoNuclear variants of bonemorphogenetic proteinsrdquo BMC Cell Biology vol 11 no 1 article20 2010
[21] N G Copeland N A Jenkins and D L Court ldquoRecombi-neering a powerful new tool for mouse functional genomicsrdquoNature Reviews Genetics vol 2 no 10 pp 769ndash779 2001
[22] S Warming N Costantino D L Court N A Jenkins and NG Copeland ldquoSimple and highly efficient BAC recombineeringusing galK selectionrdquo Nucleic Acids Research vol 33 no 4article e36 2005
[23] M Bunting K E Bernstein J M Greer M R Capecchi and KR Thomas ldquoTargeting genes for self-excision in the germ linerdquoGenes and Development vol 13 no 12 pp 1524ndash1528 1999
[24] S H George M Gertsenstein K Vintersten et al ldquoDevelop-mental and adult phenotyping directly frommutant embryonicstemcellsrdquoProceedings of theNational Academy of Sciences of theUnited States of America vol 104 no 11 pp 4455ndash4460 2007
[25] S L Mansour K R Thomas and M R Capecchi ldquoDisruptionof the proto-oncogene int-2 in mouse embryo-derived stemcells a general strategy for targeting mutations to non-select-able genesrdquo Nature vol 336 no 6197 pp 348ndash352 1988
[26] V E Papaioannou and R R Behringer Mouse PhenotypesmdashAHandbook of Mutation Analysis Cold Spring Harbor Labora-tory Press Cold Spring Harbor NY USA 2005
[27] C R Hancock J J Brault RWWiseman R L Terjung and RA Meyer ldquo31P-NMR observation of free ADP during fatiguingrepetitive contractions of murine skeletal muscle lacking AK1rdquoThe American Journal of Physiology vol 288 no 6 pp C1298ndashC1304 2005
[28] C R Hancock E Janssen and R L Terjung ldquoSkeletal musclecontractile performance and ADP accumulation in adenylatekinase-deficient micerdquoThe American Journal of Physiology vol288 no 6 pp C1287ndashC1297 2005
[29] C R Hancock E Janssen and R L Terjung ldquoContraction-mediated phosphorylation of AMPK is lower in skeletal muscleof adenylate kinase-deficient micerdquo Journal of Applied Physiol-ogy vol 100 no 2 pp 406ndash413 2006
[30] D Kosk-Kosicka ldquoCalcium signaling protocolsrdquo in Methods inMolecular Biology D G Lambert Ed vol 114 pp 343ndash354Springer New York NY USA 1999
[31] F Hillger G Herr R Rudolph and E Schwarz ldquoBiophysicalcomparison of BMP-2 ProBMP-2 and the free pro-peptidereveals stabilization of the pro-peptide by the mature growthfactorrdquo Journal of Biological Chemistry vol 280 no 15 pp14974ndash14980 2005
[32] BOhkawara S Iemura P tenDijke andNUeno ldquoAction rangeof BMP is defined by its N-terminal basic amino acid corerdquoCurrent Biology vol 12 no 3 pp 205ndash209 2002
[33] E M de Robertis ldquoSpemannrsquos organizer and the self-regulationof embryonic fieldsrdquo Mechanisms of Development vol 126 no11-12 pp 925ndash941 2009
[34] D M Maatouk K-S Choi C M Bouldin and B D Harfe ldquoInthe limb AER Bmp2 and Bmp4 are required for dorsal-ventralpatterning and interdigital cell death but not limb outgrowthrdquoDevelopmental Biology vol 327 no 2 pp 516ndash523 2009
[35] B Robert ldquoBone morphogenetic protein signaling in limboutgrowth and patterningrdquo Development Growth and Differen-tiation vol 49 no 6 pp 455ndash468 2007
[36] A Odermatt K Barton V K Khanna et al ldquoThe mutation ofPro789 to Leu reduces the activity of the fast-twitch skeletalmuscle sarco(endo)plasmic reticulum Ca2+ ATPase (SERCA1)and is associated with Brody diseaserdquoHuman Genetics vol 106no 5 pp 482ndash491 2000
[37] A Odermatt P E Taschner V K Khanna et al ldquoMutationsin the gene-encoding SERCA1 the fast-twitch skeletal musclesarcoplasmic reticulumCa2+ ATPase are associated with Brodydiseaserdquo Nature Genetics vol 14 no 2 pp 191ndash194 1996
[38] A A Benders J H Veerkamp A Oosterhof et al ldquoCa2+ home-ostasis in Brodyrsquos disease A study in skeletal muscle and cul-tured muscle cells and the effects of dantrolene and verapamilrdquoJournal of Clinical Investigation vol 94 no 2 pp 741ndash748 1994
[39] I A Brody ldquoMuscle contracture induced by exercise Asyndrome attributable to decreased relaxing factorrdquo The NewEngland Journal of Medicine vol 281 no 4 pp 187ndash192 1969
[40] Y Zhang J Fujii M S Phillips et al ldquoCharacterization ofcDNA and genomic DNA encoding SERCA1 the Ca2+-ATPaseof human fast-twitch skeletal muscle sarcoplasmic reticulumand its elimination as a candidate gene for Brody diseaserdquoGenomics vol 30 no 3 pp 415ndash424 1995
[41] M Endo ldquoCalcium-induced calcium release in skeletal musclerdquoPhysiological Reviews vol 89 no 4 pp 1153ndash1176 2009
[42] M J Betzenhauser and A RMarks ldquoRyanodine receptor chan-nelopathiesrdquo Pflugers Archiv vol 460 no 2 pp 467ndash480 2010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Related Documents











