www.elsevier.com/locate/brainres Available online at www.sciencedirect.com Research Report A novel analytical brain block tool to enable functional annotation of discriminatory transcript biomarkers among discrete regions of the fronto-limbic circuit in primate brain Clifton L. Dalgard a , David M. Jacobowitz a , Vijay K. Singh b , Kadharbatcha S. Saleem c , Robert J. Ursano d , Joshua M. Starr a , Harvey B. Pollard a,n a Department of Anatomy, Physiology and Genetics, and Center for Medical Proteomics, Uniformed Services University School of Medicine, Uniformed Services University of the Health Sciences (USUHS), 4301 Jones Bridge Road, Bethesda, MD 20914, United States b Department of Radiation Biology, Armed Forces Radiobiological Institute (AFRRI), Uniformed Services University of the Health Sciences (USUHS), 4301 Jones Bridge Road, Bethesda, MD 20914, United States c National Institute of Mental Health, National Institutes of Health, 9600 Rockville Pike, Bethesda, MD 20892, United States d Department of Psychiatry, and Center for the Study of Traumatic Stress, Uniformed Services University School of Medicine, Uniformed Services University of the Health Sciences (USUHS), 4301 Jones Bridge Road, Bethesda, MD 20914, United States article info Article history: Accepted 11 December 2014 Available online 18 December 2014 Keywords: Systems RNA-seq Area 25 Hippocampus Anterior cingulate cortex Amygdala abstract Fronto-limbic circuits in the primate brain are responsible for executive function, learning and memory, and emotions, including fear. Consequently, changes in gene expression in cortical and subcortical brain regions housing these circuits are associated with many important psychiatric and neurological disorders. While high quality gene expression profiles can be identified in brains from model organisms, primate brains have unique features such as Brodmann Area 25, which is absent in rodents, yet profoundly important in primates, including humans. The potential insights to be gained from studying the human brain are complicated by the fact that the post-mortem interval (PMI) is variable, and most repositories keep solid tissue in the deep frozen state. Consequently, sampling the important medial and internal regions of these brains is difficult. Here we describe a novel method for obtaining discrete regions from the fronto-limbic circuits of a 4 year old and a 5 year old, male, intact, frozen non-human primate (NHP) brain, for which the PMI is exactly known. The method also preserves high quality RNA, from which we use transcriptional profiling and a new algorithm to identify region-exclusive RNA signatures http://dx.doi.org/10.1016/j.brainres.2014.12.031 0006-8993/& 2015 Published by Elsevier B.V. n Correspondence to: Department of Anatomy, Physiology and Genetics, Uniformed Services University School of Medicine, Uniformed Services University of the Health Sciences, Building B, Room B2100, 4301 Jones Bridge Road, Bethesda, MD 20814, United States. Fax: þ1 301 295 2822. E-mail address: [email protected] (H.B. Pollard). brain research 1600 (2015) 42–58

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Available online at www.sciencedirect.com
www.elsevier.com/locate/brainres
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 4 2 – 5 8
http://dx.doi.org/10.0006-8993/& 2015 Pu
nCorrespondenceServices UniversityFax: þ1 301 295 2822
E-mail address:
Research Report
A novel analytical brain block tool to enablefunctional annotation of discriminatory transcriptbiomarkers among discrete regions of thefronto-limbic circuit in primate brain
Clifton L. Dalgarda, David M. Jacobowitza, Vijay K. Singhb,Kadharbatcha S. Saleemc, Robert J. Ursanod, Joshua M. Starra,Harvey B. Pollarda,n
aDepartment of Anatomy, Physiology and Genetics, and Center for Medical Proteomics, Uniformed Services UniversitySchool of Medicine, Uniformed Services University of the Health Sciences (USUHS), 4301 Jones Bridge Road, Bethesda,MD 20914, United StatesbDepartment of Radiation Biology, Armed Forces Radiobiological Institute (AFRRI), Uniformed Services University of theHealth Sciences (USUHS), 4301 Jones Bridge Road, Bethesda, MD 20914, United StatescNational Institute of Mental Health, National Institutes of Health, 9600 Rockville Pike, Bethesda,MD 20892, United StatesdDepartment of Psychiatry, and Center for the Study of Traumatic Stress, Uniformed Services University Schoolof Medicine, Uniformed Services University of the Health Sciences (USUHS), 4301 Jones Bridge Road, Bethesda,MD 20914, United States
a r t i c l e i n f o
Article history:
Accepted 11 December 2014
Fronto-limbic circuits in the primate brain are responsible for executive function, learning
and memory, and emotions, including fear. Consequently, changes in gene expression in
Available online 18 December 2014
Keywords:
Systems
RNA-seq
Area 25
Hippocampus
Anterior cingulate cortex
Amygdala
1016/j.brainres.2014.12.03blished by Elsevier B.V.
to: Department of Anatomof the Health [email protected]
a b s t r a c t
cortical and subcortical brain regions housing these circuits are associated with many
important psychiatric and neurological disorders. While high quality gene expression
profiles can be identified in brains from model organisms, primate brains have unique
features such as Brodmann Area 25, which is absent in rodents, yet profoundly important
in primates, including humans. The potential insights to be gained from studying the
human brain are complicated by the fact that the post-mortem interval (PMI) is variable,
and most repositories keep solid tissue in the deep frozen state. Consequently, sampling
the important medial and internal regions of these brains is difficult. Here we describe a
novel method for obtaining discrete regions from the fronto-limbic circuits of a 4 year old
and a 5 year old, male, intact, frozen non-human primate (NHP) brain, for which the PMI is
exactly known. The method also preserves high quality RNA, from which we use
transcriptional profiling and a new algorithm to identify region-exclusive RNA signatures
1
y, Physiology and Genetics, Uniformed Services University School of Medicine, Uniformed, Building B, Room B2100, 4301 Jones Bridge Road, Bethesda, MD 20814, United States.
u (H.B. Pollard).

b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 4 2 – 5 8 43
for Area 25 (NFκB and dopamine receptor signaling), the anterior cingulate cortex (LXR/RXR
signaling), the amygdala (semaphorin signaling), and the hippocampus (Caþþ and retinoic
acid signaling). The RNA signatures not only reflect function of the different regions, but
also include highly expressed RNAs for which function is either poorly understood, or
which generate proteins presently lacking annotated functions. We suggest that this new
approach will provide a useful strategy for identifying changes in fronto-limbic system
biology underlying normal development, aging and disease in the human brain.
& 2015 Published by Elsevier B.V.
1. Introduction
In human and non-human primate (NHP) brains, messengerRNA and microRNA expression patterns change profoundlyand coincidently during post-natal development, maturationand aging (Dannemann et al., 2012; Hu et al., 2011; Somelet al., 2010, 2011). Consequently, it has been widely antici-pated that analysis of changes in gene expression in corticaland subcortical brain regions of NHP, particularly thoseassociated with executive function and emotional control inthe fronto-limbic circuit, might yield mechanistic insightsinto many important human disorders. These disorders couldinclude entities as diverse as posttraumatic stress disorder(PTSD) (Admon et al., 2013; Choi et al., 2011; Myers et al., 2013;Taghva et al., 2013), major depressive disorder (Badawy et al.,2013; Engel et al., 2013; Godsil et al., 2013; Lozano et al., 2008;Seminowicz et al., 2004; Sibille et al., 2004), autism spectrumdisorders (Gotts et al., 2012), schizophrenia (Guillozet-Bongaarts et al., 2014; Lee, 2013; Penzes et al., 2013), anddementias associated with Alzheimer Disease (Grieve et al.,2005; Kensinger et al., 2002), Parkinson’s Disease (Bonelli andCummings, 2007; Ibarretxe-Bilbao et al., 2008), and cognitivedecline (Arnsten et al., 1995). However, very little is knownregarding the genomic and epigenomic signatures character-izing individual components of the healthy fronto-limbiccircuit, and how they might change as a function of age.We have therefore reasoned that there might be an opera-tional advantage to focusing experimental attention on theNHP brain. Importantly, the time between death and avail-ability for preservation can be minimized, and preciselycontrolled. By contrast, this is seldom true for the humanbrain, whether diseased or healthy, and circadian considera-tions also impact profoundly on the human brain transcrip-tion profile (Li et al., 2013). However, since human and NHPbrain genomes in the superior frontal gyrus precisely paralleleach other over the individual NHP and human lifespans(Somel et al., 2010), we have reasoned that there would be agreat advantage to developing methods to isolate discretefrontal and limbic NHP brain regions, while at the same timepreserving high quality RNA for genomic and epigenomicanalysis.
The transcription profile changes as a function of age in aregion-specific manner in both rodent cortex (Inukai et al.,2012; Lee et al., 2000; Loerch et al., 2008; Zahn et al., 2007), andhuman cortex (Erraji-Benchekroun et al., 2005; Hu et al., 2011;Lu et al., 2004). However, few parallels have been identifiedwhen directly comparing gene expression changes in brains
from human and other species. In the only study of this kindof which we are aware, Somel et al. (2010)) were able to mapparallel developmental and aging changes in microRNAs(miRs) and messenger RNAs (mRNAs) in gross samples ofthe surface-accessible superior frontal gyrus from frozenpostmortem brains of healthy Rhesus macaque, and in thesame region which had been dissected from frozen healthyhuman brains obtained from an NICHD/NIH-sponsored repo-sitory of frozen human brains. Relevantly, the major changesin gene expression in NHP and human superior frontal gyruscoordinately change with age when differences in lifespanare taken into account (Somel et al., 2010). However, it is notknown to what extent other parts of the brain parallel thesefindings, including components of the fronto-limbic circuit,Furthermore, inasmuch as most repositories of diseased andcontrol human brains are often available only in an intact,frozen state, it is a challenge to access and isolate centrallylocated brain regions. In addition, the time of death forhumans, especially “controls”, is seldom known with cer-tainty. For example, deaths from heart attack or stroke inotherwise psychologically healthy humans often occurs aftermidnight. By contrast, suicides usually occur during the day,when timing is more certain (Li et al., 2013). It is therefore thepurpose of this paper to describe a workflow for the frozenNHP brain that not only solves the microdissection problemfor deeper brain structures, but also yields high quality RNAthat can be used to identify quantitative, region-specific geneexpression from within the fronto-limbic circuit.
Our strategy has been to build on our personal experienceswith atlas-assisted isolation of discrete regions of freshrodent brains (Jacobowitz, 1974; Palkovits, 1973), and freshor fixed human brain (Jacobowitz et al., 1994). However, asdescribed above, substantial changes in approach needed tobe developed for microdissecting large frozen intact brains. Inthe case of brains from small rodents, we previously devel-oped the Jacobowitz Brain Block ™ (Jacobowitz, 1974). Here,we describe a new brain block, specifically designed for theNHP brain, that can be deployed for physically slicing ofeither fresh or frozen intact brains into a series of rostral-to-caudal 3 mm slices. Specific regions of frozen brain can thenbe further microdissected, and micropunched, to isolatespecific cortical and subcortical regions of the brain, whilealso preserving high quality RNA. Having verified that themethod generates intact, high quality RNA, we addressed thefollowing: (1) whether fronto-temporal regions from differentNHPs yield quantitatively similar levels of RNAs: we findr-values of 0.9 or better for two NHP brains of similar age; (2)

b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 4 2 – 5 844
whether RNA signatures can be identified that distinguishbetween Brodmann Area 25, anterior cingulate cortex (ACC),amygdala and hippocampus: we developed an algorithm toanswer this question using RNASeq data from next-generation transcription profiling; (3) whether region-specific RNA signatures have functional significance: the dataemphasize dopamine receptor and NFκB signaling in Area 25;LXR/RXR signaling in the ACC; semaphorin and glutamatereceptor signaling in amygdala; and long term potentiation(LTP), NMDA signaling, bipolar depression and temporal lobeepilepsy in hippocampus. It is as if the algorithm “automa-tically” picks out RNAs which characterize the most impor-tant individual functions of these regions relative to others inthe fronto-limbic circuit. We suggest that this new approachto developing region-specific RNA signatures provides a novelstrategy for identifying new mechanisms in fronto-limbicsystem biology which we anticipate underlie normal devel-opment, aging and disease in the human brain.
2. Results
2.1. Design of the NHP Brain block and knives for cuttingrostral to caudal slices
The primary objective of this workflow has been to obtainsuitable material from specific brain regions of post-mortem
rostral
caudal
Knife
Prefrontal cortex
Pituitary stalk
Cerebellum
OFC
IC
AMYG
Fig. 1 – Method for dissection and micropunching of frozen NHPJacobowitz Brain Block. Brain can be mounted rostral to caudal (mounted in the brain block. Ventral surface is “up”. Surface lantemporal lobe are shown. (c) Brain slices (6 mm) are laid out on adry ice is placed on top of the slices as they are deposited on thethe Saleem Rhesus macaque atlas, and cut out of the frozen slice(Brodmann Area 25). The area for mounting in a cryostat is marcingulate cortex. Details are as described for (d) and (f) amygdalentorhinal cortex. Details are as described for (d).
primate brain for transcription profiling. Our first step, there-fore, was to develop a NHP brain slicer. This slicer is a smallerversion of our human brain slicer (Jacobowitz et al., 1994),which consists of a series of 3 mm thick acrylic plates (seeFig. 1a). The plates have clearance for the movement ofrectangular knives (18�14 cm) which have been machinedfrom 0.019 in. thin stainless steel sheets, and sharpened onone edge. Two 0.5 cm diameter holes have been drilledthrough all the plates, below where the brain cavity has beenmachined, through which two 0.5 mm diameter stainlesssteel rods, threaded internally at both ends to accept screwsthat clamp the assembly together. As also seen in Fig. 1b, theassembled plates are engraved with consecutive numbers,suitable for anatomical reference.
2.2. Preparing the fresh and frozen, intact brain forcutting 6 mm rostral to caudal coronal slices
The optimum freezing protocol for NHP brain is to immerse itin powdered dry ice, and to transfer it to the �80 1C freezer.However, when brains are obtained from a distant source, itwill have been shipped in dry ice. So, this is where ourprotocol begins: the frozen brain is transferred to a �20 1Cfreezer for 3 days (e.g., Friday to Monday), to increase thetemperature. The next step is to transfer the brain to acryostat set at �6 to �10 1C, and leave it there for 5 h. Finally,the brain is placed in the NHP brain block (see Fig. 1b). The
Temporallobe
M.Prefrontal Cortex, Area 25
brain. (a) Brain block. Knife is inserted into one end of thesee arrow), or other orientations, as needed. (b) NHP brain isdmarks for prefrontal cortex, pituitary stalk, cerebellum andn aluminium sheet, resting on dry ice. Routinely, powderedplates. Brain region that has been identified by reference towith a Dremels dental drill. (d) Medial prefrontal cortexked in black ink with a fine Sharpies pencil. (e) Anteriora. Details are as described for(d) and (g) hippocampus and

b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 4 2 – 5 8 45
ventral surface is up, and the long axis is perpendicular to thecoronal plates in the block. The brain is cut in a rostral-caudaldirection. Parallel frontal slices of the brain are made atintervals of 6 mm (or any dimension of choice). In addition,the entire brain can be slightly shifted rostro-caudally alongthe cortical surface until the desired coronal angle of slicepreparation is reached. Importantly, fresh NHP brains can behandled exactly as described for the frozen brains, withoutthe preliminary �80 1C, �20 1C, �6 1C to �10 1C sequence ofsteps. However, immediately following the slicing procedure,fresh brain slices are laid out on thin metal plates (see Fig. 1c),and processed with powdered dry ice as described below.
2.3. Cutting the frozen, intact brain into 3 mm rostralto caudal coronal sections
A series of slicing blades (“knives”) are first equilibrated in anoven, maintained at a temperature of 75 1C. As shown inFig. 1a, the first knife serves as a “fence” to maintain theshape and position of that portion of the brain positioned inthe brain block. A slight pressure forward is placed at thecaudal end of the brain with the fingers of one hand, whichserves to provide rigidity to the brain. Simultaneously, withthe other reinforced gloved hand holding the knife, a down-ward sheering action of the knife produces the slice. Eachbrain slice, with its own knife, is retained in the block while aseries of slices are made. The knives are then seriallyremoved with the brain slice contained on the surface ofthe plate knife. By rotating the block 901, the knives are keptparallel to the surface of the table, thus facilitating theremoval of individual slices without slippage. As shown inFig. 1c, the slices can be arranged sequentially on aluminumplates. Each plate containing the slices is then quicklycovered with large amounts of powdered dry ice. After oneminute, the dry ice is removed with a brush, and the entiretray, holding the brain slices, is placed in a Ziploc™ bag andstored in the �80 1C freezer.
2.4. Using a stereotaxic atlas of the NHP brain to identifychosen brain regions
The sequence of steps to be followed depend on what regionsof the brain are of interest. In the present case we have beeninterested in regions associated with the fronto-limbic circuit,thought to be associated with executive function and emo-tional control. We have therefore focused specific attentionon the Medial prefrontal cortex (Area 25), anterior cingulatecortex (ACC), amygdala (AMG), and hippocampus (HIPPO). Asshown in Fig. 1d–g, we used the Saleem stereotactic atlas(Saleem et al., 2008; Saleem and Lonjothetis, 2012) to identifygross regions where these structures are found, and we haveoutlined likely regions with the fine tip of a twin tip Sharpie™marker. We then used a high speed Dremel drill to removethe segments of brain to sizes suitable for mounting andfurther sectioning in the cryostat.
2.5. Isolation of specific brain regions by micropunch
The macrodissected 6 mm slices were then mounted oncryostat chucks with OCT compound, and further sectioned
into 300 μm (0.3 mm) thick sections. These latter sections,starting approximately 1 mm away from the sliced surface,with relevant structures, were then placed on glass slides inthe cryostat. The slides were removed from the cryostat andslices thawed momentarily and quickly frozen on powdereddry ice. When sufficient sections had been cut and placed onslides, the specific regions of interest were punched out witha 1 mm diameter canula. Forty micropunches are collectedfrom each brain region, and blown directly into 350 μL ofQiazol Lysis Reagent (Qiagen, Valencia, CA). The remainder ofthe slides, and the tissue blocks, were then returned to the�80 1C freezer.
2.6. Pure high integrity RNA yield from micropunchedprimate brain regions
After Isolating total RNA from microdissected brain regions,samples were quantitatively analyzed by absorbance at230 nm, 260 nm and 280 nm to assess sample concentrationand nucleic acid purity. As summarized in Fig. 2a (Male brain#1) and Fig. 2b (Male brain #2), sufficient amounts of total RNA(4500 ng) with suitable yield (4 66 ng/μL and purity (42.0260/280 ratio) were observed from all brain regions sampled.Integrity of the isolated RNA samples from the post-mortemtissue was also observed to be suitable by automated electro-phoresis analysis (RNA integrity numbers Z8.0). Fig. 2cshows representative electropherograms for each region inMale Brain #1. A digital “gel” image of the distribution in each“lane” of the labchip is also shown (Fig. 2c-a). The twinprimary peaks in each “lane” represent 18S and 28S ribosomalRNA subunit presence and are utilized for RNA integrityassessment. In the representative example shown, totalamounts of total RNA in different brain regions were in therange of ca. 10 μg. Similar yields were observed for RNAisolated from 14 different NHP brains (Fig. 2d), of which 2are the frozen samples further analyzed here, and 12 ofwhich are from acutely obtained non-frozen NHP brains,which will be described elsewhere. We conclude that thisnew technique for RNA isolation yields high quality RNAfrom either frozen or acutely dissected NHP brains.
2.7. Transcriptome profiling of primate Area 25, anteriorcingulate cortex, amygdala and hippocampus
The informative extent of tissue material from primate brainregions by our workflow was assessed utilizing RNA-seq.Fig. 2d summarizes the sequencing metrics for analysis ofmRNA transcript abundance in different brain regions fromthe NHP fronto-limbic circuit. The numbers (#1 or #2),following the names of the brain regions, refer to sources inbrains from either NHP #1 or #2, described in Fig. 2a and b,respectively. The reads passing the quality filter of dataobtained from the Illumina GIIx is a function of multiplexingefficiency after pooling indexed samples, and sequencingdepth was aimed for approximately 20 million reads and atleast 12 million reads per sample. Alignment of reads to theRhesus macaque reference genome build resulted in 82–84% ofreads aligned for NHP #1 independent of region or sequencingdepth. Interestingly, we observed 72–78% aligned reads forNHP #2 independent of region or sequencing depth. Overall
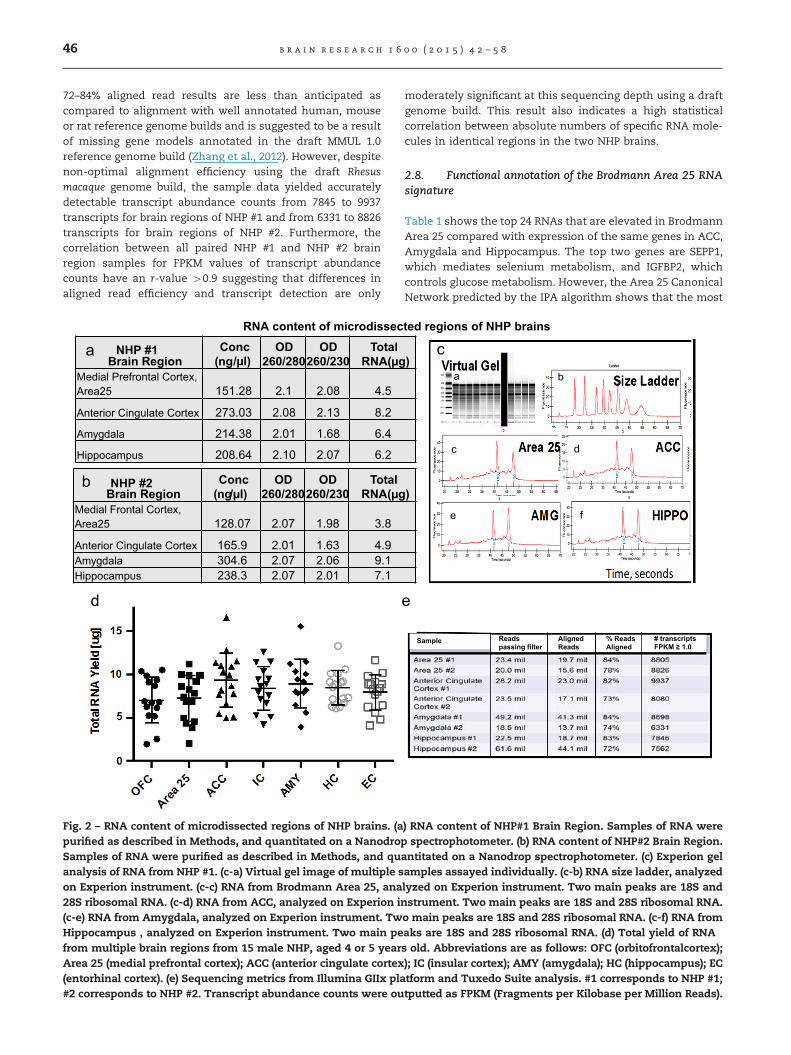
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 4 2 – 5 846
72–84% aligned read results are less than anticipated ascompared to alignment with well annotated human, mouseor rat reference genome builds and is suggested to be a resultof missing gene models annotated in the draft MMUL 1.0reference genome build (Zhang et al., 2012). However, despitenon-optimal alignment efficiency using the draft Rhesusmacaque genome build, the sample data yielded accuratelydetectable transcript abundance counts from 7845 to 9937transcripts for brain regions of NHP #1 and from 6331 to 8826transcripts for brain regions of NHP #2. Furthermore, thecorrelation between all paired NHP #1 and NHP #2 brainregion samples for FPKM values of transcript abundancecounts have an r-value 40.9 suggesting that differences inaligned read efficiency and transcript detection are only
Brain RegionConc
(ng/μl)OD
260/280OD
260/230Total
RNA(μgMedial Frontal Cortex, Area25 128.07 2.07 1.98 3.8
Anterior Cingulate Cortex 165.9 2.01 1.63 4.9Amygdala 304.6 2.07 2.06 9.1Hippocampus 238.3 2.07 2.01 7.1
RNA content of microdissec
NHP #2
Brain RegionConc
(ng/μl)OD
260/280OD
260/230Total
RNA(μgMedial Prefrontal Cortex, Area25 151.28 2.1 2.08 4.5
Anterior Cingulate Cortex 273.03 2.08 2.13 8.2
Amygdala 214.38 2.01 1.68 6.4
Hippocampus 208.64 2.10 2.07 6.2
NHP #1
Fig. 2 – RNA content of microdissected regions of NHP brains. (apurified as described in Methods, and quantitated on a NanodroSamples of RNA were purified as described in Methods, and quanalysis of RNA from NHP #1. (c-a) Virtual gel image of multiple son Experion instrument. (c-c) RNA from Brodmann Area 25, ana28S ribosomal RNA. (c-d) RNA from ACC, analyzed on Experion i(c-e) RNA from Amygdala, analyzed on Experion instrument. TwHippocampus , analyzed on Experion instrument. Two main pefrom multiple brain regions from 15 male NHP, aged 4 or 5 yearArea 25 (medial prefrontal cortex); ACC (anterior cingulate cortex(entorhinal cortex). (e) Sequencing metrics from Illumina GIIx pla#2 corresponds to NHP #2. Transcript abundance counts were ou
moderately significant at this sequencing depth using a draftgenome build. This result also indicates a high statisticalcorrelation between absolute numbers of specific RNA mole-cules in identical regions in the two NHP brains.
2.8. Functional annotation of the Brodmann Area 25 RNAsignature
Table 1 shows the top 24 RNAs that are elevated in BrodmannArea 25 compared with expression of the same genes in ACC,Amygdala and Hippocampus. The top two genes are SEPP1,which mediates selenium metabolism, and IGFBP2, whichcontrols glucose metabolism. However, the Area 25 CanonicalNetwork predicted by the IPA algorithm shows that the most
)
ted regions of NHP brains
)
Sample Reads passing filter
AlignedReads
% ReadsAligned
# transcriptsFPKM ≥ 1.0
) RNA content of NHP#1 Brain Region. Samples of RNA werep spectrophotometer. (b) RNA content of NHP#2 Brain Region.antitated on a Nanodrop spectrophotometer. (c) Experion gelamples assayed individually. (c-b) RNA size ladder, analyzedlyzed on Experion instrument. Two main peaks are 18S andnstrument. Two main peaks are 18S and 28S ribosomal RNA.o main peaks are 18S and 28S ribosomal RNA. (c-f) RNA fromaks are 18S and 28S ribosomal RNA. (d) Total yield of RNAs old. Abbreviations are as follows: OFC (orbitofrontalcortex);); IC (insular cortex); AMY (amygdala); HC (hippocampus); ECtform and Tuxedo Suite analysis. #1 corresponds to NHP #1;tputted as FPKM (Fragments per Kilobase per Million Reads).

Table 1 – Gene expression (FPKM) elevated in Brodmann Area 25, compared to ACC, amygdala and hippocampus in 4 and 5year old male NHP brains.
Gene symbol(MMU)
Gene symbol(HSA)
Gene Name Area25
ACC Amyg Hippo
MMU.2113 SEPP1 Selenoprotein P, plasma 1 65.8 11.7 27.8 15.4MMU.10509 IGFBP2 Insulin-like growth factor binding protein 2 56.5 19.1 12.0 18.4MMU.113 ERMLP Ermin, ERM-like protein 37.9 0.6 12.3 9.3MMU.4089 Protein EVI2A-like 26.9 0.6 8.7 7.1MMU.1583 CD9 CD9 antigen 26.1 2.2 12.9 7.9FOLH1 FOLH1 Folate hydrolase 23.6 4.8 10.7 7.0MMU.337 ENPP2 Ectonucleotidepyrophos-phatase/phosphodiesterase 2 20.7 1.3 8.4 6.4CDC42EP1 CDC4EP1 CDC42 effector protein (Rho GTPase binding)1 19.1 2.3 7.7 5.3MMU.15488 TS Thymidylate synthase 17.1 0.2 3.4 3.4RG59 RGS9 Regulator of G-protein signaling 9 15.7 1.0 3.2 1.2TSPAN15 TSPAN15 Tetraspanin 15 14.3 1.4 6.6 2.2MMU.3401 DRD2 Dopamine receptor D2 10.8 0.4 2.8 1.31n 1n Novel pseudogene 10.6 0.1 4.5 3.8COL9A3 COL9A3 Collagen type 9, alpha 3 10.6 1.3 3.9 2.62n DCLRE1B DNA crosslink repair 1B 10.0 2.6 3.4 3.3MMU.3402 DRD5 Dopamine receptor D5 8.2 3.8 3.2 0.6COCH COCH cochlin 7.4 1.7 3.2 0.8TMEM144 TMEM144 Transmembrane protein 144 7.4 0.3 2.8 1.8ASPA ASPA aspartoacylase 6.3 0.3 2.5 1.6SPI1 SPI1 Spleen focus forming virus(SFFV)proviral integration
oncogene6.0 1.1 2.9 2.6
MMU.3468 CYR61 Cystine rich-angiogenic inducer 61 5.0 1.7 2.1 1.7RELT RELT Tumor necrosis factor receptor 4.3 1.3 1.4 0.93n 4n Novel protein.coding 2.6 0.1 1.0 0.3HLX HLX H2.O-like homeobox 2.3 0.6 1.1 0.9
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 4 2 – 5 8 47
significant signaling pathway in this region is DopamineReceptor Signaling, followed by IGF-1 Signaling. Consistently,Dopamine Receptor D2 (DRD2, #12), a gene involved incognitive decline (Arnsten et al., 1995), and Dopamine Recep-tor D5 (DRD5, #16), a gene critical for object recognitionmemory (Saleem et al., 2008) are highly expressed. Othersignificant Dopamine Receptor categories in the IPA analysisare Dopamine-DARPP32 Feedback in cAMP signaling andG-Protein Coupled Receptor Signaling. Therefore, Area 25has a profound functional association with Dopamine Recep-tor signaling processes.
A further crucially important brain-specific connection toboth Dopamine Receptor function and IGFBP2 is aspartoacy-lase (ASPA, #19). This enzyme synthesizes N-Acetyl-asparticacid (NAA), the brain’s principal form of acetyl-coA (activatedacetate). The mRNA is expressed in neurons, but the meta-bolic product, NAA, is transferred to glia for use in eithermyelin lipid synthesis or ATP synthesis (Arun et al., 2010).Antipsychotic drugs such as haloperidol and clozapine,which bind to dopamine receptors, also increase levels ofNAA in human neurons (Arun et al., 2008). The RNA signaturefor Area 25 thus automatically identifies functional elementslinking dopamine receptors and glucose metabolism, alreadyknown from recent literature.
To further test for interactions that might functionallyinterconnect the significantly expressed RNAs in Area 25, weused the unbiased IPA algorithm to identify the Top Net-works. Fig. 3b shows Top Network (1), in which many of theRNAs in Area 25 are present, and are coded red, meaningelevated in expression. The hub-and-spoke analysis (Fig. 3c)
identifies at least 6 highly populated hubs. Of these, NFκBattracted further attention due to the fact that NFκB, p65directly drives expression of nearly 874 genes in the humangenome (Yang et al., 2013). Fig. 3d, shows that NFκB providesan integrating mechanism for at least 9 of the top 24 RNAs inArea 25. In the case of DRD2, the interaction of DRD2 with adopamine ligand activates NFκB signaling (Yang et al., 2003).In turn, NFκB binds to a κB sequence in the DRD2 promoterand drives expression of DRD2 (Bontempi et al., 2007).
In the case of other RNAs in Area 25, the #2 gene isinsulin-like growth factor binding protein 2 (IGFBP2), which islinked to an integrin/integrin-linked kinase/NFκB signalingpathway (Holmes et al., 2012). Other genes driving NFκB/RELA/p65 include CD9 (#5: tetraspanin Suzuki et al., 2009);CDC42 (#8: Hwang et al., 2012; Perona et al., 1997); TYMS (#9:TS, thymidine synthase; Wang et al., 2004); RELT (#21,TNFR;Sica et al., 2001). Indirectly, DRD2 also binds to RGS9 andtargets it to lipid rafts in brain (Celver et al., 2012). SEPPI, the#1 mRNA in Area 25, is a selenium transport protein, anisoform of which SEPPI suppresses NFκB activation by actionon the glucocorticoid receptor (Rock and Moos, 2009).As further shown in Fig. 3b, other hubs in the Area 25interactome connect to NFκB, suggesting an even closerrelationship between NFκB and Area 25 function.
2.9. Functional annotation of the anterior cingulate cortex(ACC) RNA signature
Table 2 shows the top 13 RNAs that are elevated in the anteriorcingulate cortex (ACC), compared with expression of the same
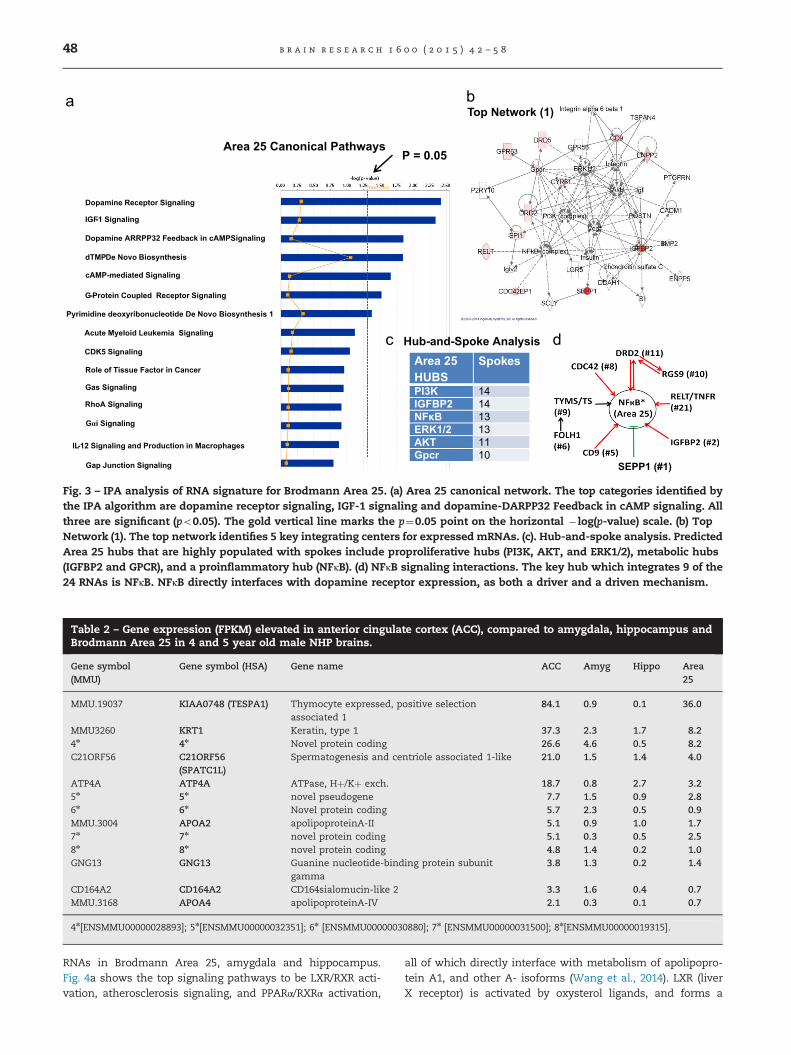
Table 2 – Gene expression (FPKM) elevated in anterior cingulate cortex (ACC), compared to amygdala, hippocampus andBrodmann Area 25 in 4 and 5 year old male NHP brains.
Gene symbol(MMU)
Gene symbol (HSA) Gene name ACC Amyg Hippo Area25
MMU.19037 KIAA0748 (TESPA1) Thymocyte expressed, positive selectionassociated 1
84.1 0.9 0.1 36.0
MMU3260 KRT1 Keratin, type 1 37.3 2.3 1.7 8.24n 4n Novel protein coding 26.6 4.6 0.5 8.2C21ORF56 C21ORF56
(SPATC1L)Spermatogenesis and centriole associated 1-like 21.0 1.5 1.4 4.0
ATP4A ATP4A ATPase, Hþ/Kþ exch. 18.7 0.8 2.7 3.25n 5n novel pseudogene 7.7 1.5 0.9 2.86n 6n Novel protein coding 5.7 2.3 0.5 0.9MMU.3004 APOA2 apolipoproteinA-II 5.1 0.9 1.0 1.77n 7n novel protein coding 5.1 0.3 0.5 2.58n 8n novel protein coding 4.8 1.4 0.2 1.0GNG13 GNG13 Guanine nucleotide-binding protein subunit
gamma3.8 1.3 0.2 1.4
CD164A2 CD164A2 CD164sialomucin-like 2 3.3 1.6 0.4 0.7MMU.3168 APOA4 apolipoproteinA-IV 2.1 0.3 0.1 0.7
4n[ENSMMU00000028893]; 5n[ENSMMU00000032351]; 6n [ENSMMU00000030880]; 7n [ENSMMU00000031500]; 8n[ENSMMU00000019315].
Top Network (1)
Area 25 Canonical Pathways
Hub-and-Spoke AnalysisArea 25 HUBS
Spokes
PI3K 14IGFBP2 14NFκB 13ERK1/2 13AKT 11Gpcr 10
SEPP1 (#1)
P = 0.05
Dopamine Receptor Signaling
IGF-1 Signaling
Dopamine ARRPP32 Feedback in cAMPSignaling
dTMPDe Novo Biosynthesis
cAMP-mediated Signaling
G-Protein Coupled Receptor Signaling
Pyrimidine deoxyribonucleotide De Novo Biosynthesis 1
Acute Myeloid Leukemia Signaling
CDK5 Signaling
Role of Tissue Factor in Cancer
Gas Signaling
RhoA Signaling
Gαi Signaling
IL-12 Signaling and Production in Macrophages
Gap Junction Signaling
Fig. 3 – IPA analysis of RNA signature for Brodmann Area 25. (a) Area 25 canonical network. The top categories identified bythe IPA algorithm are dopamine receptor signaling, IGF-1 signaling and dopamine-DARPP32 Feedback in cAMP signaling. Allthree are significant (po0.05). The gold vertical line marks the p¼0.05 point on the horizontal � log(p-value) scale. (b) TopNetwork (1). The top network identifies 5 key integrating centers for expressed mRNAs. (c). Hub-and-spoke analysis. PredictedArea 25 hubs that are highly populated with spokes include proproliferative hubs (PI3K, AKT, and ERK1/2), metabolic hubs(IGFBP2 and GPCR), and a proinflammatory hub (NFκB). (d) NFκB signaling interactions. The key hub which integrates 9 of the24 RNAs is NFκB. NFκB directly interfaces with dopamine receptor expression, as both a driver and a driven mechanism.
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 4 2 – 5 848
RNAs in Brodmann Area 25, amygdala and hippocampus.Fig. 4a shows the top signaling pathways to be LXR/RXR acti-vation, atherosclerosis signaling, and PPARα/RXRα activation,
all of which directly interface with metabolism of apolipopro-tein A1, and other A- isoforms (Wang et al., 2014). LXR (liverX receptor) is activated by oxysterol ligands, and forms a

ACC: Canonical Pathways
Hub-and-Spoke AnalysisACC HUBS Spokes
UBC 24ATP4A 18
Top Network (1)P= 0.05
Novel or UncharacterizedProtein Coding
Genes
Rank
ENSMMUG_28893 #3ENSMMUG_30880 #7ENSMMUG_31500 #9ENSMMUG_19315 #10
Unknown Proteins
LXR/RXRSignaling
TESLA1(#1)
ATP4A(#5)
APOA4(#13)
APOA2(#8)
LXR/RXR Activation
IL-12 Signaling and Production in Macrophages
Clathrin-mediated Endocytosis Signaling
PPARα/RXRα Activation
Atherosclerosis Signaling
Acute Phase Response Signaling
Production of Nitric Oxide and ROS in Macrophages
G Protein Signaling Mediated by TUBBY
Antiproliferative Role of Somatostatin Receptor 2
Fig. 4 – IPA analysis of RNA signature for anterior cingulate cortex. (a) Anterior cingulate cortex canonical network. There areat least 9 categories identified by the IPA program as being significant to the set of 13 RNAs identified in the ACC. Top amongthese with relevance to the brain are LXR/RXR activation. (b) Top Network (1). The predicted ACC hubs identify 2 principalintegrating centers, UBC and ATP4A (c) Hub-and-spoke analysis. There are two well populated hubs: UBC (ubiquitin) andATP4A (ATPase, Hþ/Kþ exchanger). (d) Unknown proteins. Four of the 13 ACC RNAs code for novel or uncharacterizedproteins. Thus ca. 25% of the RNA signature in the ACC is unknown. (e) LXR/RXR Signaling. Of the 13 ACC-specific RNAs, 4 ofthem (ca. 25%) are associated with LXR/RXR signaling. These are nuclear receptors which are metabolic products of vitamin A,for which mRNA levels are significantly reduced in the postmortem dorsolateral prefrontal cortex/anterior cingulate cortex(ACC) from elderly depressed patients who did not die from suicide (Qi et al., 2013).
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 4 2 – 5 8 49
heterodimer with RXR (retinoic X acid receptor). The mRNAs forthese receptors are significantly reduced in the postmortemdorsolateral prefrontal cortex/anterior cingulate cortex (ACC)from elderly depressed patients who did not die from suicide(Qi et al., 2013). Thus they may play an important role in ACCfunction. The Top Network (1) in Fig. 4b emphasizes predictedmultiple interactions with apolipoprotein A isoforms. Cerebralhypoperfusion in rats has been reported to directly impact onlevels of LXR/RXR signaling and apolipoprotein A1 levels (Wanget al., 2014). It has also been suggested that elevated apo-A1 inthe forebrain may have neuroprotective properties by decreas-ing amyloid beta secretion (Koldamova et al., 2003). However,Fig. 4c emphasizes the fact that 5 of the 13 amygdala-specificRNAs code for novel or uncharacterized protein coding genes,and that another codes for a novel pseudogene. Thus almosthalf of the RNAs making up the specific ACC RNA signature areproteins with no known function. Nonetheless, a major hub isfound for ATP4A (Fig. 4d). This hub points to LXR/RXR activa-tion, and Fig. 4e shows that 4 of the 7 identified mRNAs areassociated with this signaling pathway. This result suggests
that if this strategy for automatically identifying functionallyimportant RNAs has importance for the dependent functions,but also in identifying the potential functions of theseunknown proteins.
2.10. Functional annotation of the amygdala RNAsignature
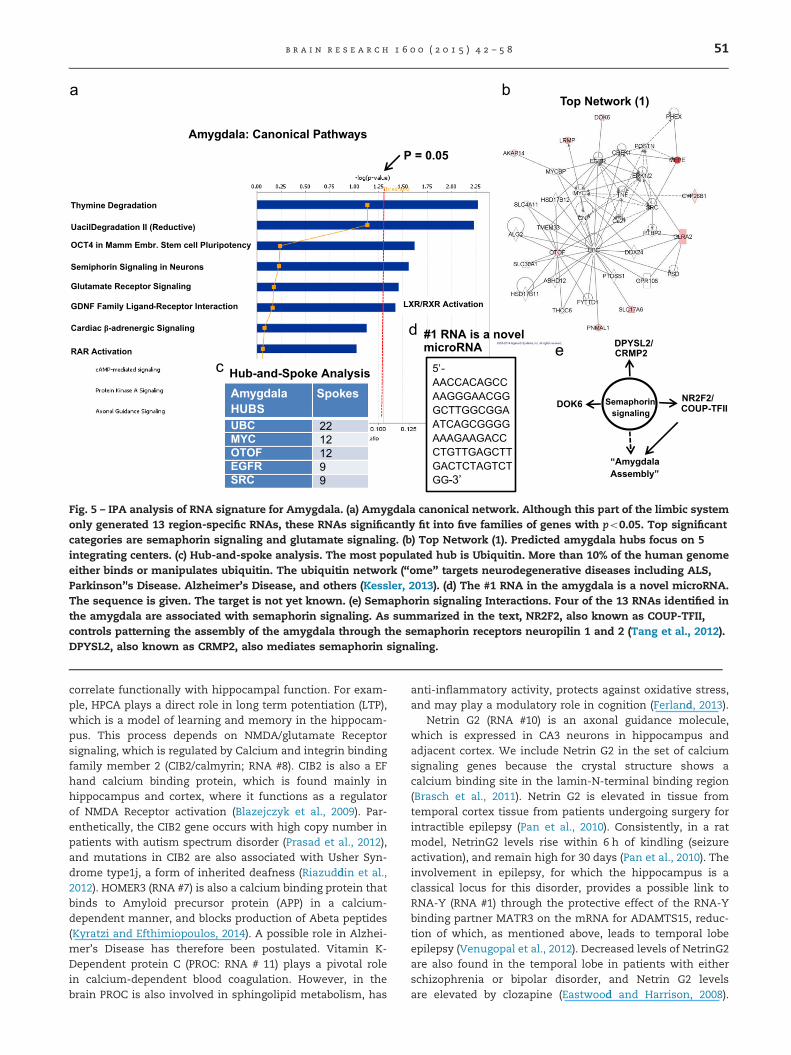
Table 3 shows the top 13 RNAs that are elevated in theamygdala, compared with expression of the same genes inBrodmann Area 25, ACC, and hippocampus. The top RNA inthis brain region is a novel microRNA. The sequence, which isshown at the foot of Table 3, and in Fig. 5d, presently lacks anidentified target mRNA. The amygdala canonical pathwayanalysis in Fig. 5a identifies two different significant signalingpathways: “semaphorin signaling in neurons” and “glutamatereceptor signaling”. As summarized in Fig. 5e, the #2 mRNA,NR2F2, also known as COUP-TFII, controls patterning theassembly of the amygdala through the semaphorin receptorsneuropilin 1 and 2 (Tang et al., 2012). DPYSL2, also known as
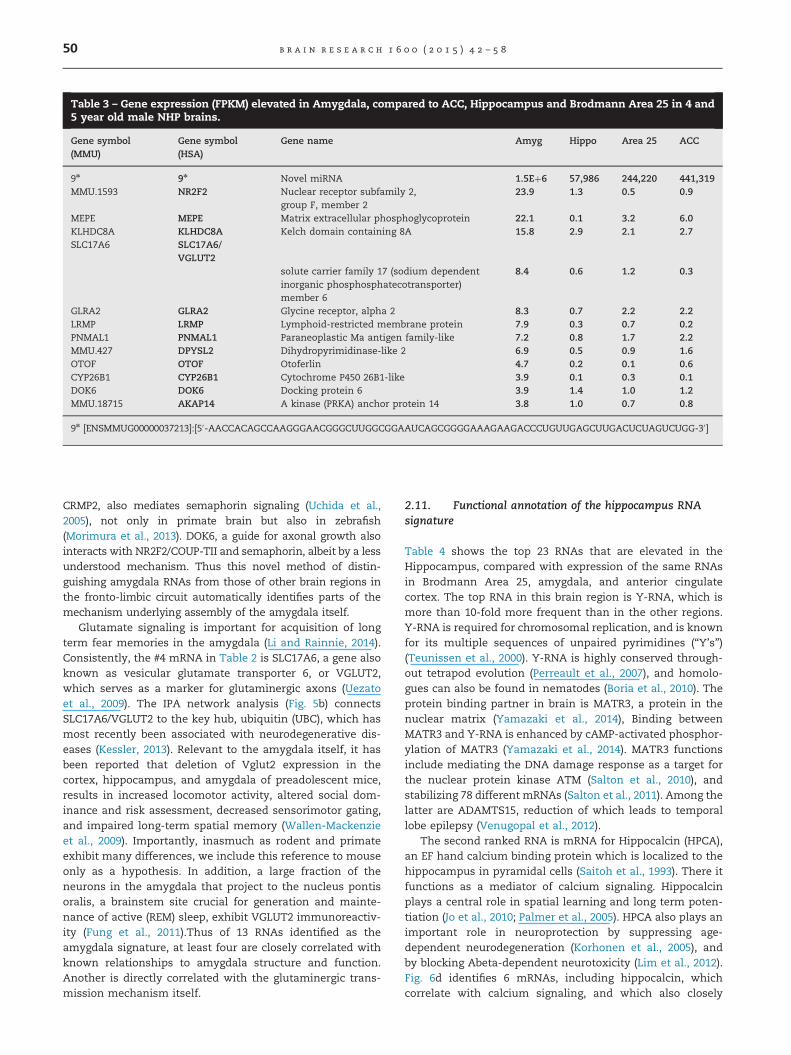
Table 3 – Gene expression (FPKM) elevated in Amygdala, compared to ACC, Hippocampus and Brodmann Area 25 in 4 and5 year old male NHP brains.
Gene symbol(MMU)
Gene symbol(HSA)
Gene name Amyg Hippo Area 25 ACC
9n 9n Novel miRNA 1.5Eþ6 57,986 244,220 441,319MMU.1593 NR2F2 Nuclear receptor subfamily 2,
group F, member 223.9 1.3 0.5 0.9
MEPE MEPE Matrix extracellular phosphoglycoprotein 22.1 0.1 3.2 6.0KLHDC8A KLHDC8A Kelch domain containing 8A 15.8 2.9 2.1 2.7SLC17A6 SLC17A6/
VGLUT2solute carrier family 17 (sodium dependentinorganic phosphosphatecotransporter)member 6
8.4 0.6 1.2 0.3
GLRA2 GLRA2 Glycine receptor, alpha 2 8.3 0.7 2.2 2.2LRMP LRMP Lymphoid-restricted membrane protein 7.9 0.3 0.7 0.2PNMAL1 PNMAL1 Paraneoplastic Ma antigen family-like 7.2 0.8 1.7 2.2MMU.427 DPYSL2 Dihydropyrimidinase-like 2 6.9 0.5 0.9 1.6OTOF OTOF Otoferlin 4.7 0.2 0.1 0.6CYP26B1 CYP26B1 Cytochrome P450 26B1-like 3.9 0.1 0.3 0.1DOK6 DOK6 Docking protein 6 3.9 1.4 1.0 1.2MMU.18715 AKAP14 A kinase (PRKA) anchor protein 14 3.8 1.0 0.7 0.8
9n [ENSMMUG00000037213]:[50-AACCACAGCCAAGGGAACGGGCUUGGCGGAAUCAGCGGGGAAAGAAGACCCUGUUGAGCUUGACUCUAGUCUGG-30]
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 4 2 – 5 850
CRMP2, also mediates semaphorin signaling (Uchida et al.,2005), not only in primate brain but also in zebrafish(Morimura et al., 2013). DOK6, a guide for axonal growth alsointeracts with NR2F2/COUP-TII and semaphorin, albeit by a lessunderstood mechanism. Thus this novel method of distin-guishing amygdala RNAs from those of other brain regions inthe fronto-limbic circuit automatically identifies parts of themechanism underlying assembly of the amygdala itself.
Glutamate signaling is important for acquisition of longterm fear memories in the amygdala (Li and Rainnie, 2014).Consistently, the #4 mRNA in Table 2 is SLC17A6, a gene alsoknown as vesicular glutamate transporter 6, or VGLUT2,which serves as a marker for glutaminergic axons (Uezatoet al., 2009). The IPA network analysis (Fig. 5b) connectsSLC17A6/VGLUT2 to the key hub, ubiquitin (UBC), which hasmost recently been associated with neurodegenerative dis-eases (Kessler, 2013). Relevant to the amygdala itself, it hasbeen reported that deletion of Vglut2 expression in thecortex, hippocampus, and amygdala of preadolescent mice,results in increased locomotor activity, altered social dom-inance and risk assessment, decreased sensorimotor gating,and impaired long-term spatial memory (Wallen-Mackenzieet al., 2009). Importantly, inasmuch as rodent and primateexhibit many differences, we include this reference to mouseonly as a hypothesis. In addition, a large fraction of theneurons in the amygdala that project to the nucleus pontisoralis, a brainstem site crucial for generation and mainte-nance of active (REM) sleep, exhibit VGLUT2 immunoreactiv-ity (Fung et al., 2011).Thus of 13 RNAs identified as theamygdala signature, at least four are closely correlated withknown relationships to amygdala structure and function.Another is directly correlated with the glutaminergic trans-mission mechanism itself.
2.11. Functional annotation of the hippocampus RNAsignature
Table 4 shows the top 23 RNAs that are elevated in theHippocampus, compared with expression of the same RNAsin Brodmann Area 25, amygdala, and anterior cingulatecortex. The top RNA in this brain region is Y-RNA, which ismore than 10-fold more frequent than in the other regions.Y-RNA is required for chromosomal replication, and is knownfor its multiple sequences of unpaired pyrimidines (“Y’s”)(Teunissen et al., 2000). Y-RNA is highly conserved through-out tetrapod evolution (Perreault et al., 2007), and homolo-gues can also be found in nematodes (Boria et al., 2010). Theprotein binding partner in brain is MATR3, a protein in thenuclear matrix (Yamazaki et al., 2014), Binding betweenMATR3 and Y-RNA is enhanced by cAMP-activated phosphor-ylation of MATR3 (Yamazaki et al., 2014). MATR3 functionsinclude mediating the DNA damage response as a target forthe nuclear protein kinase ATM (Salton et al., 2010), andstabilizing 78 different mRNAs (Salton et al., 2011). Among thelatter are ADAMTS15, reduction of which leads to temporallobe epilepsy (Venugopal et al., 2012).
The second ranked RNA is mRNA for Hippocalcin (HPCA),an EF hand calcium binding protein which is localized to thehippocampus in pyramidal cells (Saitoh et al., 1993). There itfunctions as a mediator of calcium signaling. Hippocalcinplays a central role in spatial learning and long term poten-tiation (Jo et al., 2010; Palmer et al., 2005). HPCA also plays animportant role in neuroprotection by suppressing age-dependent neurodegeneration (Korhonen et al., 2005), andby blocking Abeta-dependent neurotoxicity (Lim et al., 2012).Fig. 6d identifies 6 mRNAs, including hippocalcin, whichcorrelate with calcium signaling, and which also closely
Amygdala: Canonical Pathways
Hub-and-Spoke Analysis
Top Network (1)
P = 0.05
Semaphorinsignaling
DPYSL2/CRMP2
DOK6 NR2F2/COUP-TFII
“Amygdala Assembly”
5’-AACCACAGCCAAGGGAACGGGCTTGGCGGAATCAGCGGGGAAAGAAGACCCTGTTGAGCTTGACTCTAGTCTGG-3’
#1 RNA is a novel microRNA
LXR/RXR Activation
Thymine Degradation
OCT4 in Mamm Embr. Stem cell Pluripotency.
UacilDegradation II (Reductive)
Semiphorin Signaling in Neurons
Glutamate Receptor Signaling
GDNF Family Ligand Receptor Interaction-
Cardiac β-adrenergic Signaling
RAR Activation
Amygdala HUBS
Spokes
UBCMYCOTOFEGFRSRC
22121299
Fig. 5 – IPA analysis of RNA signature for Amygdala. (a) Amygdala canonical network. Although this part of the limbic systemonly generated 13 region-specific RNAs, these RNAs significantly fit into five families of genes with po0.05. Top significantcategories are semaphorin signaling and glutamate signaling. (b) Top Network (1). Predicted amygdala hubs focus on 5integrating centers. (c) Hub-and-spoke analysis. The most populated hub is Ubiquitin. More than 10% of the human genomeeither binds or manipulates ubiquitin. The ubiquitin network (“ome” targets neurodegenerative diseases including ALS,Parkinson’'s Disease. Alzheimer’s Disease, and others (Kessler, 2013). (d) The #1 RNA in the amygdala is a novel microRNA.The sequence is given. The target is not yet known. (e) Semaphorin signaling Interactions. Four of the 13 RNAs identified inthe amygdala are associated with semaphorin signaling. As summarized in the text, NR2F2, also known as COUP-TFII,controls patterning the assembly of the amygdala through the semaphorin receptors neuropilin 1 and 2 (Tang et al., 2012).DPYSL2, also known as CRMP2, also mediates semaphorin signaling.
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 4 2 – 5 8 51
correlate functionally with hippocampal function. For exam-ple, HPCA plays a direct role in long term potentiation (LTP),which is a model of learning and memory in the hippocam-pus. This process depends on NMDA/glutamate Receptorsignaling, which is regulated by Calcium and integrin bindingfamily member 2 (CIB2/calmyrin; RNA #8). CIB2 is also a EFhand calcium binding protein, which is found mainly inhippocampus and cortex, where it functions as a regulatorof NMDA Receptor activation (Blazejczyk et al., 2009). Par-enthetically, the CIB2 gene occurs with high copy number inpatients with autism spectrum disorder (Prasad et al., 2012),and mutations in CIB2 are also associated with Usher Syn-drome type1j, a form of inherited deafness (Riazuddin et al.,2012). HOMER3 (RNA #7) is also a calcium binding protein thatbinds to Amyloid precursor protein (APP) in a calcium-dependent manner, and blocks production of Abeta peptides(Kyratzi and Efthimiopoulos, 2014). A possible role in Alzhei-mer’s Disease has therefore been postulated. Vitamin K-Dependent protein C (PROC: RNA # 11) plays a pivotal rolein calcium-dependent blood coagulation. However, in thebrain PROC is also involved in sphingolipid metabolism, has
anti-inflammatory activity, protects against oxidative stress,and may play a modulatory role in cognition (Ferland, 2013).
Netrin G2 (RNA #10) is an axonal guidance molecule,which is expressed in CA3 neurons in hippocampus andadjacent cortex. We include Netrin G2 in the set of calciumsignaling genes because the crystal structure shows acalcium binding site in the lamin-N-terminal binding region(Brasch et al., 2011). Netrin G2 is elevated in tissue fromtemporal cortex tissue from patients undergoing surgery forintractible epilepsy (Pan et al., 2010). Consistently, in a ratmodel, NetrinG2 levels rise within 6 h of kindling (seizureactivation), and remain high for 30 days (Pan et al., 2010). Theinvolvement in epilepsy, for which the hippocampus is aclassical locus for this disorder, provides a possible link toRNA-Y (RNA #1) through the protective effect of the RNA-Ybinding partner MATR3 on the mRNA for ADAMTS15, reduc-tion of which, as mentioned above, leads to temporal lobeepilepsy (Venugopal et al., 2012). Decreased levels of NetrinG2are also found in the temporal lobe in patients with eitherschizophrenia or bipolar disorder, and Netrin G2 levelsare elevated by clozapine (Eastwood and Harrison, 2008).

Table 4 – Gene expression (FPKM) elevated in hippocampus, compared to ACC, amygdala and Brodmann Area 25 in 4 and 5year old male NHP brains.
Gene symbol(MMU)
Gene symbol(HSA)
Gene name Hippo Area25
ACC Amyg
Y RNA Y RNA Y RNA 2246.9 185.2 56.4 180.0HPCA HPCA Hippocalcin, neuron-specific calcium binding protein 1298.8 300.3 418.7 340.0TTR TTR Transthyretin 1080.6 1.6 2.1 125.4RAB26 RAB26 Ras-related protein Rab-26-like 242.0 116.7 103.7 118.1CPLX2 CPLX2 Complexin 2 161.3 41.9 67.1 66.1LMO1 LMO1 LIM domain only 1 (rhombotin 1) 153.2 5.6 15.0 55.8HOMER3 HOMER3 HOMER3 102.4 21.3 16.0 12.8CIB2 CIB2 Calcium and integrin-binding family member 2 99.7 21.6 28.4 23.2AQP3 AQP3 Aquaporin 3 39.5 1.6 1.7 7.7NTNG2 NTNG2 Netrin G2 34.9 12.7 8.6 15.1PROC PROC Vitamin K-dependent protein C 17.2 1.9 2.5 4.7CST1 CST1 Cystatin SA 13.3 1.9 3.4 4.9WNT4 WNT4 Wingless-type MMTV integration site family, member 4 12.9 0.9 1.7 5.3KCNS3 KCNS3 Potassium voltage gated channel 8.6 1.2 2.9 2.4WFIKKN2 WFIKKN2 WAP-follistatin/kazal, immunoglobulin, kunitz and netrin
domain containing 27.8 0.7 0.4 0.4
GFRA3 GFRA3 GDNF family receptor alpha 3 6.8 1.7 0.7 1.8MMU.19523 HSP27 Heat shock protein 27 6.5 0.7 2.3 1.0MSC MSC musculin 5.2 1.5 1.4 1.9FOLR1 FOLR1 Folate receptor alpha 4.4 0.3 0.4 1.3MFAP5 MFAP5 Microfibrillar associated protein 5 4.2 0 0 0WNK4 WNK4 With No Lysines 4 2.6 0.3 0.3 0.7SCL13A4 SLC13A4 Solute carrier family 13 (sodium/sulfate symporter) member
42.4 0.5 0.2 0.6
CCNO CCNO Cyclin O 2.2 1.0 1.1 0.4
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 4 2 – 5 852
Clozapine is an atypical antipsychotic medication used totreat treatment-resistant schizophrenia (Cheine et al., 1999).
Wingless-type MMTV integration site family member 4(WNT4; RNA #13) is included in the set of calcium signalinggenes because it drives calcium/NFAT signaling (Burn et al.,2011). WNT4 is also responsible for anterior–posterior position-ing in the developing vertebrate brain (Lyuksyutova et al., 2003).WNT4 has been shown to play a role in adult brain in survival,function, and plasticity of neurons (Inestrosa and Arenas, 2010),including adult hippocampal neurogenesis (Lie et al., 2005).WNT4, via the upstream wingless (Wnt)-frizzled (Fz) signalingpathways, has also been found to impact mood disorders andmay therefore be important for symptoms of depressionoriginating from the hippocampus (Duman and Voleti, 2012).GDNF family receptor alpha 3 (GFRA3: RNA # 16) marks neuronsin the pyramidal layer of Ammons’s Horn, and in the hilus andgranular layer of the fascia dentata (Serra et al., 2005). Expres-sion varies as a function of age, and functions to promote bothneuronal survival and regulate differentiation (Sariola andSaarma, 2003). Thus both WNT4, and GFRA3 play vital roles inthe differentiation and function of the hippocampus.
The third ranked RNA in the hippocampus is the mRNAfor transthyretin (TTR), a retinol binding protein recentlyshown to be significantly reduced in CSF and plasma fromschizophrenia in patients (Wan et al., 2006). The entire net-work of retinoic acid signaling genes is expressed in differentregions of the rat brain, including hypothalamus (Helfer et al.,2012), where retinoic acid metabolites and Wnt/β-catenin areboth regulated by the photoperiod. TTR mediates uptake andtransport of retinol (vitamin A), for downstream conversion
into the retinaldehyde and retinoic acid, and subsequentsignaling. These signaling ligands regulate expression of theAlzheimer Disease related amyloid precursor protein (APP),analogous with HOMER3 (#7). Retinoic acid signaling inhippocampus is itself driven by calcium signaling (Kimet al., 2014), and regulates 4 genes: LMO1 (#6) (Rice et al.,2004); TTR (#3) (Brouillette and Quirion, 2008); AQP3 (#9) (Songet al., 2008); and shares regulation of Netrin G2 (#10) (Koreckaet al., 2013) with the calcium signaling mechanism.
3. Discussion
The fronto-limbic circuit, which manages executive function,learning and memory, emotions, and fear behavior, has been ofclinical and academic concern for more than 100 years (Newmanand Harris, 2009; Papez, 1995; Shah et al., 2012; Triarhou, 2008).These are functions that most distinguish humans, and to alarge extent non-human primates, including Rhesus macaque,from lower animals. Yet how the components of the circuitfunction, both as individual units in the circuit, and as anintegrated circuit, remains poorly understood. Here we haveidentified RNA transcript markers which significantly discrimi-nate among four regions of the fronto-limbic circuit: BrodmannArea 25, amygdala, anterior cingulate cortex (ACC), and Hippo-campus. By using an algorithm to distinguish the most region-specific RNAs from among 9320 RNAs that are accuratelydetectable in specific brain regions, we have identified smallsubsets, of ca. 25 RNAs each, that have critical developmental

Hippocampus: Canonical Pathways
P = 0.05
Ca++
Signaling
HPCA(#2)
CIB2(#8)
(NMDA, LTP)
PROC(#11)
WNT4(#13)
NetrinG2(#10)(epilepsy)
(clozapine)(bipolar disorder, schizophrenia)
Y-RNA(#1)
(hippocampalneurogenesis)
HOMER3(#7)
(cognition)
(APP)
RetinoicAcid
TTR(#3) AQP3
(#9)
LMO1(#6)
Hub-and-Spoke AnalysisHippocampus HUBS
Spokes
ESR1 14TTR 14IL-6 14IFNG 12HNF10 10XBP1 9ERK1/2 9APP 7CXCL8 7IKBKB 6
Top Network (1)
GFRA3(#16)
Glutamate Receptor Signaling
GDNF Family-Ligand Interactions
Basal Cell Carcinoma SignalingWnt/GSK-3b Signaling in Path. of Influenza
Virus entry via Endocytic PathwayDeath Receptor Signaling
Fig. 6 – IPA analysis of RNA signature for Hippocampus. (a) Hippocampus canonical network. The Canonical Pathwaysanalysis is less informative than for other pathways. The reason may rest in many functions being focused on just a relativelyfew RNAs (however, see Fig. 4d). (b). Top Network (1). The predicted hippocampal hubs emphasize four integrating centers.(c) Hub-and-spoke analysis. There are four well populated hubs, falling into two categories: inflammation (TNF [tumornecrosis factor alpha], IFNG [interferon gamma], MMP2 [matrix metalloprotease 2] and proliferation (ERK1/2). (d) Calciumsignaling Interactions. The key to understanding the integrating principal in the hippocampal RNA signature is the commoncalcium-activation properties shared by 6 of the RNAs, and the interactions through common hippocampal functions.These include LTP (long term potentiation), a learning and memory model, epilepsy, cognition, bipolar depression, andschizophrenia. One of the genes (NetrinG2, #10) is a target for clozapine, an anti-schizophrenia drug.
b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 4 2 – 5 8 53
and clinical importance for each of the regions. These findingsinclude dopamine receptor and NFκB signaling in Area 25;semaphorin and glutamate receptor signaling in amygdala, longterm potentiation (LTP), NMDA signaling, bipolar depression andtemporal lobe epilepsy in hippocampus; and vitamin A metabo-lism in the ACC. This initial analysis enriched for RNA transcriptmarkers which characterize the most important individualfunctions of these regions in the fronto-limbic circuit. Only theACC remains an enigma in this analysis because almost half ofthe ACC-specific RNAs code for proteins with no currentlyknown function. In addition, the #1 gene is abundant relativeto the other three limbic areas, but is a non-coding RNA ofunknown function. Presumably identifying the function of theseunknown proteins, and non-coding transcripts may significantlyshed functional and developmental light on the ACC. These RNAsignatures are thus valuable additions to the neurobiology toolkitby providing molecular mechanisms responsible for region-specific development and differentiation, and for identifyingpotential therapeutic targets of psychological and psychiatric
disease that are considered to characterize each region in thecircuit.
3.1. Functional importance of RNA signatures andregion-specific organizing signaling networks in thefronto-limbic circuit
Circuitry in the brain is often used as an integrating paradigm toexplain complex behavior. Simplistically, connections within thefronto-limbic circuit integrate executive function and cognitionwith anxiety and fear memories, which have evolutionarilyimportant functions for survival and propagation of the species.This integrative function reaches an apogee in primate brain.However, to understand how the different regions of the circuitwork, a certain level of reductionistic analysis has always beenneeded. We find that all regions in this circuit share approxi-mately 9320 genes. Thus identifying which ones are importantfor regional function is challenging if the approach is one-gene-at-a-time. In fact, this one gene-at-a time approach, taking over a

b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 4 2 – 5 854
century to perform, has yielded the information we needed tovalidate the algorithm-dependent “automatic” identification offunctionally important RNAs in just four regions of the fronto-limbic circuit. The algorithm thus has the advantage of filteringout contributions from common cellular elements such as glia orblood vessels. This results in limiting identifications to mostlyneuron-specific RNAs. The integrating principals, represented bymanually curated “hubs” in each of the Figures, may only beimportant as helpful management tools. Alternatively, they maytruly represent functional differences for neurons in each of thefour regions. The fact that the RNA signatures for each of theregions are so different suggests that these management toolsmay have some mechanistic significance.
3.2. Novel method for accessing internal brain regions foranalysis from intact, frozen brains
The region-specific RNA signatures are possible because of animportant methodological advance, described here for the firsttime, that allows for micropunching interior brain regions fromfrozen intact brains, and obtaining quality RNA from thosemicropunches. Most available sources of brain for molecularanalysis are in brain repositories where the samples are keptfrozen at �80 1C. The consequence is that regions from thesurface of the frozen brain can be easily chiseled off for analysis(Somel et al., 2010). However, internal brain structures, includingmost of the component regions of the fronto-limbic circuit, arenot immediately available. Here we have solved this problem inthe following way: (i) by developing a brain block suitable forholding the Rhesus macaque brain in a reproducible manner forcoronal slicing; (ii) by lowering the temperature of the solid brainto ca. �10 1C for a limited time; (iii) by heating the individualknife blades to 75 1C just prior to use; and (iv) by immersing eachslice immediately in powdered dry ice to return the sample tothe �80 1C condition. This method thus exposes the sub-zerosolid brain for only a limited time to higher than �80 1Cconditions. Second, while the hot knife cuts through the frozenbrain, it may also heat-inactivate susceptible enzymes that maybe exposed on the cut surfaces as the hot knife passes throughthe tissue. However, cryostat sections for micropunching camefrom deeper within the 6mm initial slice, and were therefore areca 1mm away from the transiently heated, cut surfaces (seeSection 2.5). The new method, therefore, reproducibly generateshigh quality RNA from which functional insights into brainfunction can be dependably inferred. We have also used thistechnique on frozen segments of human brain, and on freshNHP brain, with equivalently high quality yields of RNA. Wetherefore believe that this new method of accessing deepstructures in frozen, intact brain will permit more efficient useof existing human brain bank samples.
3.3. Studies of non-human primate brain provide valuablefor insights into human brain function
Increasing attention is presently being given to the NHPbrain, as a more accurate model for human brain structureand function (Hara et al., 2012; Lu et al., 2012; Marvanovaet al., 2003). For example, it has recently been reported that
the genomic responses of the mouse to injuries as diverse asinfection, burns, and trauma do not significantly phenocopythe temporal relationships of gene expression in response toparallel injuries in humans (Seok et al., 2013). Furthermore,many centrally mediated drug side effects that are readilyobserved in humans, can also be observed in NHPs, but arenot easily accessible in pre-clinical rodent models (Porsolt,2013). A telling example for specific behavioral parallelsbetween NHP and humans is a depression-susceptibilityorthologue in the Rhesus macaque genome for a commonhuman polymorphism of the serotonin transporter gene(rh5-HTTLPR(s)). NHPs bearing this polymorphism have simi-lar behavior and brain morphology to humans with the samepolymorphism (Jedema et al., 2010). This polymorphism alsoimpacts on the human cingulate-amygdala interactions asso-ciated with susceptibility to depression (Pezawas et al., 2005),and also alters the functional integration of emotional infor-mation between amygdala and medial prefrontal cortex(Brodman Area 25). Deep brain stimulation of Area 25 hasbeen successfully deployed for treatment of Major DepressiveDisorder (MDD) (Kennedy et al., 2011; Mayberg et al., 2005). Bycontrast, an operational definition of susceptibility to depres-sion in rodent models is a significant challenge. However,since rodent brain and behavior will remain an importantexperimental system for the foreseeable future, we suggestthat the close genomic relationship between NHP and humanchanges, measured over a lifetime (Somel et al., 2010), can beused with confidence to verify the human-relevance of easilystudied behavioral and genomic responses to aging, drugs orinjury in the rodent brain. Our focus on NHP genomics, brainand behavior is therefore of particular importance as 21stcentury neuroscience inexorably turns towards identifyingbiological pathways in human brain that contribute to com-plex cognitive and emotional behavior (Hariri et al., 2006).
4. Experimental procedures
4.1. Animals
All animal studies were approved by the Uniformed ServicesUniversity University of the Health Sciences (USUHS) Institu-tional Animal Care and Use Committee (IACUC) and wereconducted in a facility accredited by the Association forAssessment and Accreditation of Laboratory Animal CareInternational (AAALAC). Frozen NHP brains were alsoobtained from Tulane Primate Center.
4.2. RNA isolation from micropunched brain regions
Tissue micropunches in QIAzol reagent were homogenizedusing a Bioruptor Standard UCD-200 (Diagenode, Denville, NJ)for 2 cycles of 30 s on and 30 s off at high power. Afterincubation at room temperature for 5 min, total RNA wasisolated using the miRNeasy Mini Kit (Qiagen). A final elutionvolume of 30 μL of RNAse free water is added to the spincolumn and centrifuged at 412,000 rpm for 1 min. Isolatedtotal RNA concentration is quantified on a NanoDrop ND-1000 Spectrophotometer (Thermo Scientific, Wilmington, DE)

b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 4 2 – 5 8 55
and integrity is determined on an Experion AutomatedElectrophoresis System (Bio-Rad) before storage at �80 1C.
4.3. RNA expression analysis of micro-punched braintissue by sequencing (mRNA-seq)
RNA samples with RNA integrity number values 47.0 wereused as input for sequencing library preparation using theTruSeq RNA Sample Preparation Kit v2 (Illumina, San Diego,CA). Sequencing libraries were validated for size and integrityusing an Experion Automated Electrophoresis System(BioRad), and library concentration was determined usingthe KAPA Library Quantification Kits for Illumina (KAPABiosystems). 12–15 pmol of Library were used as input forclustering by a c-Bot (Illumina) using the TruSeq PE ClusterKit—GAIIx (Illumina) before sequencing on a Genome Analy-zer IIx (Illumina). Sequencing was performed as a multiplexwith paired-end reads of 68 bp length with an indexing readof 7 bp in between sequencing reads. FASTQ data weregenerated using CASAVA, and raw reads were analyzed andgroomed using FASTQC and FASTQ Groomer, respectively.FPKM values for transcriptome expression levels were gener-ated using Bowtie 2, TopHat 2 and Cufflinks 2 and the Rhesusmacaque reference genome (MMUL 1.0 latest genebuild May2010). FPKM values were used for Comparative Marker Selec-tion and differential gene analysis by Cuffdiff (see Fig. 2e).
4.4. Analysis algorithm to identify region-enriched or-specific RNA transcripts in primate brain
One task of our analysis was to identify transcripts which areenriched in one of the four brain regions relative to the otherthree, and do so for each of the four regions. We identified thecandidate differently enriched transcripts in each specificbrain region from a subset of accurately detected transcriptswith FPKM values 41.0. This subset of 9320 transcripts wasanalyzed using Comparative Marker Selection in GenePattern2.0 (Broad Institute, MIT) to identify marker transcripts withp-value o0.05. Candidate transcripts were then tested for aFPKM fold-ratio 42.0 as compared to all other brain regionsindividually. Furthermore, candidate differentially enrichedtranscripts were only selected if these conditions weresatisfied in both of the subjects profiled. FPKM values ofregion-specific differences in expression were tabulated.
4.5. Bioinformatics of region-specific RNAs
The set of region-specific RNAs were used as input forpathway analysis in Ingenuity Pathway Analysis (IPA) soft-ware platform to test for significant connectivity in transcriptfunction, pathway, regulatory activity and/or mechanismrelationships. The network analysis identifies “hubs” towhich multiple targets may connect as “spokes”. High occu-pancy hubs are considered to identify important functionalintegrators. Manual curation of the knowledge base from thisanalysis identified contextual hubs that were not apparentfrom simple relationships.
Disclaimer
The opinions expressed herein belong solely to the authors.They do not, nor should they be, interpreted as representativeor endorsed by the Uniformed Services University of theHealth Sciences, the Department of Defense, or any otheragency of the federal government.
Acknowledgments
This work was supported by in part by AFRRI grant no. CBM.RAD.01.10. AR.005(G3B2EO) to VKS; and by CDMRP-PTSD(PTO74415) to HBP. Additionally, the authors would like tothank Ms. Xiuying Zhang for her expert help in preparing themanuscript, and the excellent technical assistance from Ms.Tinghua Chen. Gene expression data are deposited in theGene Expression Omnibus (GEO) database: http://www.ncbi.nlm.nih.gov/geo (accession number GSE64797).
r e f e r e n c e s
Admon, R., Milad, M.R., Hendler, T., 2013. A causal model of post-traumatic stress disorder: disentangling predisposed fromacquired neural abnormalities. Trends Cogn. Sci. 17, 337–347.
Arnsten, A.F., et al., 1995. Dopamine D2 receptor mechanismscontribute to age-related cognitive decline: the effects ofquinpirole on memory and motor performance in monkeys.J. Neurosci. 15, 3429–3439.
Arun, P., et al., 2008. Antipsychotic drugs increase N-acetylaspartate and N-acetylaspartylglutamate in SH-SY5Yhuman neuroblastoma cells. J. Neurochem. 106, 1669–1680.
Arun, P., et al., 2010. Metabolic acetate therapy for the treatmentof traumatic brain injury. J. Neurotrauma 27, 293–298.
Badawy, R.A., et al., 2013. Capturing the epileptic trait: corticalexcitability measures in patients and their unaffectedsiblings. Brain 136, 1177–1191.
Blazejczyk, M., et al., 2009. Biochemical characterization andexpression analysis of a novel EF-hand Ca2þ binding proteincalmyrin2 (Cib2) in brain indicates its function in NMDAreceptor mediated Ca2þ signaling. Arch. Biochem. Biophys.487, 66–78.
Bonelli, R.M., Cummings, J.L., 2007. Frontal-subcortical circuitryand behavior. Dialogues Clin. Neurosci. 9, 141–151.
Bontempi, S., et al., 2007. Identification and characterization oftwo nuclear factor-kappaB sites in the regulatory region of thedopamine D2 receptor. Endocrinology 148, 2563–2570.
Boria, I., et al., 2010. Nematode sbRNAs: homologs of vertebrate YRNAs. J. Mol. Evol. 70, 346–358.
Brasch, J., et al., 2011. Crystal structure of the ligand bindingdomain of netrin G2. J. Mol. Biol. 414, 723–734.
Brouillette, J., Quirion, R., 2008. Transthyretin: a key gene involvedin the maintenance of memory capacities during aging.Neurobiol. Aging 29, 1721–1732.
Burn, S.F., et al., 2011. Calcium/NFAT signalling promotes earlynephrogenesis. Dev. Biol. 352, 288–298.
Celver, J., Sharma, M., Kovoor, A., 2012. D(2)-Dopamine receptorstarget regulator of G protein signaling 9-2 to detergent-resistant membrane fractions. J. Neurochem. 120, 56–69.
Cheine, M.V., Wahlbeck, K., Rimon, M., 1999. Pharmacologicaltreatment of schizophrenia resistant to first-line treatment: acritical systematic review and meta-analysis. Int. J. PsychiatryClin. Pract. 3, 159–169.

b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 4 2 – 5 856
Choi, K., et al., 2011. Analysis of kinase gene expression in thefrontal cortex of suicide victims: implications of fear andstress. Front. Behav. Neurosci. 5, 46.
Dannemann, M., et al., 2012. Transcription factors are targeted bydifferentially expressed miRNAs in primates. Genome Biol.Evol. 4, 552–564.
Duman, R.S., Voleti, B., 2012. Signaling pathways underlying thepathophysiology and treatment of depression: novelmechanisms for rapid-acting agents. Trends Neurosci. 35,47–56.
Eastwood, S.L., Harrison, P.J., 2008. Decreased mRNA expressionof netrin-G1 and netrin-G2 in the temporal lobe inschizophrenia and bipolar disorder.Neuropsychopharmacology 33, 933–945.
Engel Jr., J., et al., 2013. Connectomics and epilepsy. Curr. Opin.Neurol. 26, 186–194.
Erraji-Benchekroun, L., et al., 2005. Molecular aging in humanprefrontal cortex is selective and continuous throughout adultlife. Biol. Psychiatry 57, 549–558.
Ferland, G., 2013. Vitamin K and brain function. Semin. Thromb.Hemost. 39, 849–855.
Fung, S.J., et al., 2011. Projection neurons from the centralnucleus of the amygdala to the nucleus pontis oralis.J. Neurosci. Res. 89, 429–436.
Godsil, B.P., et al., 2013. The hippocampal-prefrontal pathway: theweak link in psychiatric disorders? Eur.Neuropsychopharmacol. 23, 1165–1181.
Gotts, S.J., et al., 2012. Fractionation of social brain circuits inautism spectrum disorders. Brain 135, 2711–2725.
Grieve, S.M., et al., 2005. Preservation of limbic and paralimbicstructures in aging. Hum. Brain Mapp. 25, 391–401.
Guillozet-Bongaarts, A.L., et al., 2014. Altered gene expression inthe dorsolateral prefrontal cortex of individuals withschizophrenia. Mol. Psychiatry 19, 478–485.
Hara, Y., Rapp, P.R., Morrison, J.H., 2012. Neuronal andmorphological bases of cognitive decline in aged rhesusmonkeys. Age (Dordr) 34, 1051–1073.
Hariri, A.R., Drabant, E.M., Weinberger, D.R., 2006. Imaginggenetics: perspectives from studies of genetically drivenvariation in serotonin function and corticolimbic affectiveprocessing. Biol. Psychiatry 59, 888–897.
Helfer, G., et al., 2012. Photoperiod regulates vitamin A and Wnt/beta-catenin signaling in F344 rats. Endocrinology 153,815–824.
Holmes, K.M., et al., 2012. Insulin-like growth factor-bindingprotein 2-driven glioma progression is prevented by blocking aclinically significant integrin, integrin-linked kinase, and NF-kappaB network. Proc. Natl. Acad. Sci. U.S.A. 109, 3475–3480.
Hu, H.Y., et al., 2011. MicroRNA expression and regulation inhuman, chimpanzee, and macaque brains. PLoS Genet. 7,e1002327.
Hwang, J.R., et al., 2012. Upregulation of CD9 in ovarian cancer isrelated to the induction of TNF-alpha gene expression andconstitutive NF-kappaB activation. Carcinogenesis 33, 77–83.
Ibarretxe-Bilbao, N., et al., 2008. Hippocampal head atrophypredominance in Parkinson’s disease with hallucinations andwith dementia. J. Neurol. 255, 1324–1331.
Inestrosa, N.C., Arenas, E., 2010. Emerging roles of Wnts in theadult nervous system. Nat. Rev. Neurosci. 11, 77–86.
Inukai, S., et al., 2012. Novel microRNAs differentially expressedduring aging in the mouse brain. PLoS One 7, e40028.
Jacobowitz, D.M., 1974. Removal of discrete fresh regions of therat brain. Brain Res. 80, 111–115.
Jacobowitz, D.M., Sullivan, J.V., Fitze, P.E., 1994. Human brainslicer: a method for cutting coronal slices of fresh and fixedhuman brains. Brain Res. Bull. 33, 461–463.
Jedema, H.P., et al., 2010. Cognitive impact of genetic variation ofthe serotonin transporter in primates is associated with
differences in brain morphology rather than serotonin
neurotransmission. Mol. Psychiatr. 15 (512-22), 446.Jo, J., et al., 2010. Muscarinic receptors induce LTD of NMDAR
EPSCs via a mechanism involving hippocalcin, AP2 and PSD-
95. Nat. Neurosci. 13, 1216–1224.Kennedy, S.H., et al., 2011. Deep brain stimulation for treatment-
resistant depression: follow-up after 3 to 6 years. Am. J.
Psychiatry 168, 502–510.Kensinger, E.A., et al., 2002. Effects of normal aging and
Alzheimer’s disease on emotional memory. Emotion 2,
118–134.Kessler, B.M., 2013. Ubiquitin—omics reveals novel networks and
associations with human disease. Curr. Opin. Chem. Biol. 17,
59–65.Kim, K.D., et al., 2014. Calcium signaling via Orai1 is essential for
induction of the nuclear orphan receptor pathway to drive
Th17 differentiation. J. Immunol. 192, 110–122.Koldamova, R.P., et al., 2003. 22R-hydroxycholesterol and 9-cis-
retinoic acid induce ATP-binding cassette transporter A1
expression and cholesterol efflux in brain cells and decrease
amyloid beta secretion. J. Biol. Chem. 278, 13244–13256.Korecka, J.A., et al., 2013. Phenotypic characterization of retinoic
acid differentiated SH-SY5Y cells by transcriptional profiling.
PLoS One 8, e63862.Korhonen, L., et al., 2005. Hippocalcin protects against caspase-
12-induced and age-dependent neuronal degeneration. Mol.
Cell. Neurosci. 28, 85–95.Kyratzi, E., Efthimiopoulos, S., 2014. Calcium regulates the
interaction of amyloid precursor protein with Homer3 protein.
Neurobiol. Aging 35, 2053–2063.Lee, C.K., Weindruch, R., Prolla, T.A., 2000. Gene-expression
profile of the ageing brain in mice. Nat. Genet. 25, 294–297.Lee, D., 2013. Decision making: from neuroscience to psychiatry.
Neuron 78, 233–248.Li, C., Rainnie, D.G., 2014. Bidirectional regulation of synaptic
plasticity in the basolateral amygdala induced by D1-like
family of dopamine receptors and group II metabotropic
glutamate receptors. J. Physiol..Li, J.Z., et al., 2013. Circadian patterns of gene expression in the
human brain and disruption in major depressive disorder.
Proc. Natl. Acad. Sci. U.S.A. 110, 9950–9955.Lie, D.C., et al., 2005. Wnt signalling regulates adult hippocampal
neurogenesis. Nature 437, 1370–1375.Lim, Y.A., et al., 2012. Role of hippocalcin in mediating Abeta
toxicity. Biochim. Biophys. Acta 1822, 1247–1257.Loerch, P.M., et al., 2008. Evolution of the aging brain
transcriptome and synaptic regulation. PLoS One 3, e3329.Lozano, A.M., et al., 2008. Subcallosal cingulate gyrus deep brain
stimulation for treatment-resistant depression. Biol.
Psychiatry 64, 461–467.Lu, J., et al., 2012. Effect of blast exposure on the brain structure
and cognition in Macaca fascicularis. J. Neurotrauma 29,
1434–1454.Lu, T., et al., 2004. Gene regulation and DNA damage in the ageing
human brain. Nature 429, 883–891.Lyuksyutova, A.I., et al., 2003. Anterior–posterior guidance of
commissural axons by Wnt-frizzled signaling. Science 302,
1984–1988.Marvanova, M., et al., 2003. Microarray analysis of nonhuman
primates: validation of experimental models in neurological
disorders. FASEB J. 17, 929–931.Mayberg, H.S., et al., 2005. Deep brain stimulation for treatment-
resistant depression. Neuron 45, 651–660.Morimura, R., et al., 2013. Phosphorylation of Dpsyl2 (CRMP2) and
Dpsyl3 (CRMP4) is required for positioning of caudal primary
motor neurons in the zebrafish spinal cord. Dev. Neurobiol. 73,
911–920.

b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 4 2 – 5 8 57
Myers, B., et al., 2013. Central stress-integrative circuits: forebrainglutamatergic and GABAergic projections to the dorsomedialhypothalamus, medial preoptic area, and bed nucleus of thestria terminalis. Brain Struct. Funct..
Newman, J.D., Harris, J.C., 2009. The scientific contributions ofPaul D. MacLean (1913–2007). J. Nerv. Ment. Dis. 197, 3–5.
Palkovits, M., 1973. Isolated removal of hypothalamic or otherbrain nuclei of the rat. Brain Res. 59, 449–450.
Palmer, C.L., et al., 2005. Hippocalcin functions as a calciumsensor in hippocampal LTD. Neuron 47, 487–494.
Pan, Y., et al., 2010. Abnormal expression of netrin-G2 in temporallobe epilepsy neurons in humans and a rat model. Exp.Neurol. 224, 340–346.
Papez, J.W., 1995. A proposed mechanism of emotion. 1937.J. Neuropsychiatry Clin. Neurosci. 7, 103–112.
Penzes, P., et al., 2013. Developmental vulnerability of synapsesand circuits associated with neuropsychiatric disorders.J. Neurochem. 126, 165–182.
Perona, R., et al., 1997. Activation of the nuclear factor-kappaB byRho, CDC42, and Rac-1 proteins. Genes Dev. 11, 463–475.
Perreault, J., Perreault, J.P., Boire, G., 2007. Ro-associated Y RNAsin metazoans: evolution and diversification. Mol. Biol. Evol. 24,1678–1689.
Pezawas, L., et al., 2005. 5-HTTLPR polymorphism impacts humancingulate-amygdala interactions: a genetic susceptibilitymechanism for depression. Nat. Neurosci. 8, 828–834.
Porsolt, R.D., 2013. The usefulness of non-human primates incentral nervous system safety pharmacology. J. Pharmacol.Toxicol. Methods 68, 23–29.
Prasad, A., et al., 2012. A discovery resource of rare copy numbervariations in individuals with autism spectrum disorder. G3(Bethesda) 2, 1665–1685.
Qi, X.R., et al., 2013. Abnormal retinoid and TrkB signaling in theprefrontal cortex in mood disorders. Cereb. Cortex.
Riazuddin, S., et al., 2012. Alterations of the CIB2 calcium- andintegrin-binding protein cause Usher syndrome type 1J andnonsyndromic deafness DFNB48. Nat. Genet. 44, 1265–1271.
Rice, A.M., et al., 2004. Analysis of the relationship between Scltranscription factor complex protein expression patterns andthe effects of LiCl on ATRA-induced differentiation in blastcells from patients with acute myeloid leukemia. Leuk. Res.28, 1227–1237.
Rock, C., Moos, P.J., 2009. Selenoprotein P regulation by theglucocorticoid receptor. Biometals 22, 995–1009.
Saitoh, S., et al., 1993. Distribution of hippocalcin mRNA andimmunoreactivity in rat brain. Neurosci. Lett. 157, 107–110.
Saleem, K.S., Kondo, H., Price, J.L., 2008. Complementary circuitsconnecting the orbital and medial prefrontal networks withthe temporal, insular, and opercular cortex in the macaquemonkey. J. Comp. Neurol. 506, 659–693.
Saleem, K.S., Lonjothetis, N.K., 2012. Combined MRI andHistology Atlas Of The Rhesus Monkey Brain In SterotaxicCoordinates, Vol. Elsevier/Academic Press, San Diego.
Salton, M., et al., 2010. Involvement of Matrin 3 and SFPQ/NONOin the DNA damage response. Cell Cycle 9, 1568–1576.
Salton, M., et al., 2011. Matrin 3 binds and stabilizes mRNA. PLoSOne 6, e23882.
Sariola, H., Saarma, M., 2003. Novel functions and signallingpathways for GDNF. J. Cell Sci. 116, 3855–3862.
Seminowicz, D.A., et al., 2004. Limbic-frontal circuitry in majordepression: a path modeling metanalysis. Neuroimage 22,409–418.
Seok, J., et al., 2013. Genomic responses in mouse models poorlymimic human inflammatory diseases. Proc. Natl. Acad. Sci. U.S.A. 110, 3507–3512.
Serra, M.P., et al., 2005. Ret, GFRalpha-1, GFRalpha-2 andGFRalpha-3 receptors in the human hippocampus and fasciadentata. Int. J. Dev. Neurosci. 23, 425–438.
Shah, A., Jhawar, S.S., Goel, A., 2012. Analysis of the anatomy of
the Papez circuit and adjoining limbic system by fiber
dissection techniques. J. Clin. Neurosci. 19, 289–298.Sibille, E., et al., 2004. Gene expression profiling of depression and
suicide in human prefrontal cortex. Neuropsychopharmacology
29, 351–361.Sica, G.L., et al., 2001. RELT, a new member of the tumor necrosis
factor receptor superfamily, is selectively expressed in
hematopoietic tissues and activates transcription factor
NF-kappaB. Blood 97, 2702–2707.Somel, M., et al., 2010. MicroRNA, mRNA, and protein expression
link development and aging in human and macaque brain.
Genome Res. 20, 1207–1218.Somel, M., et al., 2011. MicroRNA-driven developmental
remodeling in the brain distinguishes humans from other
primates. PLoS Biol. 9, e1001214.Song, X., et al., 2008. Nicotinamide attenuates aquaporin 3
overexpression induced by retinoic acid through inhibition of
EGFR/ERK in cultured human skin keratinocytes. Int. J. Mol.
Med. 22, 229–236.Suzuki, M., et al., 2009. Tetraspanin CD9 negatively regulates
lipopolysaccharide-induced macrophage activation and lung
inflammation. J. Immunol. 182, 6485–6493.Taghva, A., et al., 2013. Posttraumatic stress disorder:
neurocircuitry and implications for potential deep brain
stimulation. Stereotact. Funct. Neurosurg. 91, 207–219.Tang, K., et al., 2012. COUP-TFII controls amygdala patterning by
regulating neuropilin expression. Development 139,
1630–1639.Teunissen, S.W., et al., 2000. Conserved features of Y RNAs: a
comparison of experimentally derived secondary structures.
Nucleic Acids Res. 28, 610–619.Triarhou, L.C., 2008. Centenary of Christfried Jakob’s discovery of
the visceral brain: an unheeded precedence in affective
neuroscience. Neurosci. Biobehav. Rev. 32, 984–1000.Uchida, Y., et al., 2005. Semaphorin3A signalling is mediated via
sequential Cdk5 and GSK3beta phosphorylation of CRMP2:
implication of common phosphorylating mechanism
underlying axon guidance and Alzheimer’s disease. Genes
Cells 10, 165–179.Uezato, A., Meador-Woodruff, J.H., McCullumsmith, R.E., 2009.
Vesicular glutamate transporter mRNA expression in the
medial temporal lobe in major depressive disorder, bipolar
disorder, and schizophrenia. Bipolar Disord. 11, 711–725.Venugopal, A.K., et al., 2012. Transcriptomic profiling of medial
temporal lobe epilepsy. J. Proteomics Bioinf., 5.Wallen-Mackenzie, A., et al., 2009. Restricted cortical and
amygdaloid removal of vesicular glutamate transporter 2 in
preadolescent mice impacts dopaminergic activity and
neuronal circuitry of higher brain function. J. Neurosci. 29,
2238–2251.Wan, C., et al., 2006. Dysregulation of retinoid transporters
expression in body fluids of schizophrenia patients.
J. Proteome Res. 5, 3213–3216.Wang, L., et al., 2014. Dynamic changes of Apo A1 mediated by
LXR/RXR/ABCA1 pathway in brains of the aging rats with
cerebral hypoperfusion. Brain Res. Bull. 100, 84–92.Wang, W., et al., 2004. Mechanistic and predictive profiling of
5-Fluorouracil resistance in human cancer cells. Cancer Res.
64, 8167–8176.Yamazaki, F., et al., 2014. pY RNA1-s2: a highly retina-enriched
small RNA that selectively binds to Matrin 3 (Matr3). PLoS One
9, e88217.Yang, J., et al., 2013. A probabilistic approach to learn chromatin
architecture and accurate inference of the NF-kappaB/RelA
regulatory network using ChIP-Seq. Nucleic Acids Res. 41,
7240–7259.

b r a i n r e s e a r c h 1 6 0 0 ( 2 0 1 5 ) 4 2 – 5 858
Yang, M., et al., 2003. Requirement of Gbetagamma and c-Src inD2 dopamine receptor-mediated nuclear factor-kappaBactivation. Mol. Pharmacol. 64, 447–455.
Zahn, J.M., et al., 2007. AGEMAP: a gene expression database foraging in mice. PLoS Genet. 3, e201.
Zhang, X., Goodsell, J., Norgren Jr., R.B., 2012. Limitations of the
rhesus macaque draft genome assembly and annotation. BMC
Genomics 13, 206.
Related Documents











