A Newly Isolated Reovirus Has the Simplest Genomic and Structural Organization of Any Reovirus Albert J. Auguste, a Jason T. Kaelber, b Eric B. Fokam, c Hilda Guzman, a Christine V. F. Carrington, d Jesse H. Erasmus, a Basile Kamgang, e Vsevolod L. Popov, a Joanita Jakana, b Xiangan Liu, b Thomas G. Wood, f Steven G. Widen, f Nikos Vasilakis, a Robert B. Tesh, a Wah Chiu, b Scott C. Weaver a,g Institute for Human Infections and Immunity, Center for Tropical Diseases, and Department of Pathology, University of Texas Medical Branch, Galveston, Texas, USA a ; National Center for Macromolecular Imaging, Verna and Marrs McLean Department of Biochemistry and Molecular Biology, and Department of Molecular Virology and Microbiology, Baylor College of Medicine, Houston, Texas, USA b ; Department of Zoology and Animal Physiology, University of Buea, Buea, Cameroon c ; Department of Preclinical Sciences, Faculty of Medical Sciences, University of the West Indies, St. Augustine, Trinidad and Tobago d ; Institut Pasteur in Bangui, Department of Virology, Bangui, Central African Republic e ; Department of Biochemistry and Molecular Biology, University of Texas Medical Branch, Galveston, Texas, USA f ; Sealy Center for Structural Biology and Molecular Biophysics, University of Texas Medical Branch, Galveston, Texas, USA g ABSTRACT A total of 2,691 mosquitoes representing 17 species was collected from eight locations in southwest Cameroon and screened for pathogenic viruses. Ten isolates of a novel reovirus (genus Dinovernavirus) were detected by culturing mosquito pools on Aedes albopictus (C6/36) cell cultures. A virus that caused overt cytopathic effects was isolated, but it did not infect vertebrate cells or produce detectable disease in infant mice after intracerebral inoculation. The virus, tentatively designated Fako virus (FAKV), represents the first 9-segment, double-stranded RNA (dsRNA) virus to be isolated in nature. FAKV appears to have a broad mos- quito host range, and its detection in male specimens suggests mosquito-to-mosquito transmission in nature. The structure of the T1 FAKV virion, determined to subnanometer resolution by cryoelectron microscopy (cryo-EM), showed only four pro- teins per icosahedral asymmetric unit: a dimer of the major capsid protein, one turret protein, and one clamp protein. While all other turreted reoviruses of known structures have at least two copies of the clamp protein per asymmetric unit, FAKV’s clamp protein bound at only one conformer of the major capsid protein. The FAKV capsid architecture and genome organization rep- resent the most simplified reovirus described to date, and phylogenetic analysis suggests that it arose from a more complex an- cestor by serial loss-of-function events. IMPORTANCE We describe the detection, genetic, phenotypic, and structural characteristics of a novel Dinovernavirus species isolated from mosquitoes collected in Cameroon. The virus, tentatively designated Fako virus (FAKV), is related to both single-shelled and partially double-shelled viruses. The only other described virus in this genus was isolated from cultured mosquito cells. It was previously unclear whether the phenotypic characteristics of that virus were reflective of this genus in nature or were altered during serial passaging in the chronically infected cell line. FAKV is a naturally occurring single-shelled reovirus with a unique virion architecture that lacks several key structural elements thought to stabilize a single-shelled reovirus virion, suggesting what may be the minimal number of proteins needed to form a viable reovirus particle. FAKV evolved from more complex an- cestors by losing a genome segment and several virion proteins. H uman and livestock diseases are commonly caused by viruses transmitted from enzootic hosts by arthropod vectors (1). Mosquito surveillance studies have been key to understanding the prevalence, distribution, and outbreak potential of the majority of these pathogens, as well as for the detection of novel pathogenic and nonpathogenic agents. The family Reoviridae consists of double-stranded RNA (dsRNA) viruses with 9 to 12 genome segments. Reoviruses have both a wide geographic distribution and wide host range, having been isolated from fungi, plants, insects, ticks, arachnids, fish, marine protists, crustaceans, mammals, and birds (2). They are divided into two subfamilies, Spinareovirinae and Sedoreovirinae, which are classified based on their core structures. Spinareovirinae viruses, such as the recently emerged human pathogen Melaka virus and the mammalian orthoreovirus type 3, are called “tur- reted” reoviruses, because they encode spikes (protrusions) on the surface of the virion. The subfamily Sedoreovirinae contains the “nonturreted” viruses with multiple smooth shells, which include human and other animal pathogens, such as rotaviruses and orbi- viruses (3, 4). In addition to this structural classification, amino acid identities of 30% in the polymerase sequence and the pres- Received 4 August 2014 Accepted 20 October 2014 Accepted manuscript posted online 29 October 2014 Citation Auguste AJ, Kaelber JT, Fokam EB, Guzman H, Carrington CVF, Erasmus JH, Kamgang B, Popov VL, Jakana J, Liu X, Wood TG, Widen SG, Vasilakis N, Tesh RB, Chiu W, Weaver SC. 2015. A newly isolated reovirus has the simplest genomic and structural organization of any reovirus. J Virol 89:676 – 687. doi:10.1128/JVI.02264-14. Editor: R. W. Doms Address correspondence to Scott C. Weaver, [email protected]. A.J.A., J.T.K., and E.B.F. contributed equally. Supplemental material for this article may be found at http://dx.doi.org/10.1128 /JVI.02264-14. Copyright © 2015, American Society for Microbiology. All Rights Reserved. doi:10.1128/JVI.02264-14 676 jvi.asm.org January 2015 Volume 89 Number 1 Journal of Virology on December 16, 2014 by MOODY MEDICAL LIBRARY http://jvi.asm.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A Newly Isolated Reovirus Has the Simplest Genomic and StructuralOrganization of Any Reovirus
Albert J. Auguste,a Jason T. Kaelber,b Eric B. Fokam,c Hilda Guzman,a Christine V. F. Carrington,d Jesse H. Erasmus,a
Basile Kamgang,e Vsevolod L. Popov,a Joanita Jakana,b Xiangan Liu,b Thomas G. Wood,f Steven G. Widen,f Nikos Vasilakis,a
Robert B. Tesh,a Wah Chiu,b Scott C. Weavera,g
Institute for Human Infections and Immunity, Center for Tropical Diseases, and Department of Pathology, University of Texas Medical Branch, Galveston, Texas, USAa;National Center for Macromolecular Imaging, Verna and Marrs McLean Department of Biochemistry and Molecular Biology, and Department of Molecular Virology andMicrobiology, Baylor College of Medicine, Houston, Texas, USAb; Department of Zoology and Animal Physiology, University of Buea, Buea, Cameroonc; Department ofPreclinical Sciences, Faculty of Medical Sciences, University of the West Indies, St. Augustine, Trinidad and Tobagod; Institut Pasteur in Bangui, Department of Virology,Bangui, Central African Republice; Department of Biochemistry and Molecular Biology, University of Texas Medical Branch, Galveston, Texas, USAf; Sealy Center forStructural Biology and Molecular Biophysics, University of Texas Medical Branch, Galveston, Texas, USAg
ABSTRACT
A total of 2,691 mosquitoes representing 17 species was collected from eight locations in southwest Cameroon and screened forpathogenic viruses. Ten isolates of a novel reovirus (genus Dinovernavirus) were detected by culturing mosquito pools on Aedesalbopictus (C6/36) cell cultures. A virus that caused overt cytopathic effects was isolated, but it did not infect vertebrate cells orproduce detectable disease in infant mice after intracerebral inoculation. The virus, tentatively designated Fako virus (FAKV),represents the first 9-segment, double-stranded RNA (dsRNA) virus to be isolated in nature. FAKV appears to have a broad mos-quito host range, and its detection in male specimens suggests mosquito-to-mosquito transmission in nature. The structure ofthe T�1 FAKV virion, determined to subnanometer resolution by cryoelectron microscopy (cryo-EM), showed only four pro-teins per icosahedral asymmetric unit: a dimer of the major capsid protein, one turret protein, and one clamp protein. While allother turreted reoviruses of known structures have at least two copies of the clamp protein per asymmetric unit, FAKV’s clampprotein bound at only one conformer of the major capsid protein. The FAKV capsid architecture and genome organization rep-resent the most simplified reovirus described to date, and phylogenetic analysis suggests that it arose from a more complex an-cestor by serial loss-of-function events.
IMPORTANCE
We describe the detection, genetic, phenotypic, and structural characteristics of a novel Dinovernavirus species isolated frommosquitoes collected in Cameroon. The virus, tentatively designated Fako virus (FAKV), is related to both single-shelled andpartially double-shelled viruses. The only other described virus in this genus was isolated from cultured mosquito cells. It waspreviously unclear whether the phenotypic characteristics of that virus were reflective of this genus in nature or were alteredduring serial passaging in the chronically infected cell line. FAKV is a naturally occurring single-shelled reovirus with a uniquevirion architecture that lacks several key structural elements thought to stabilize a single-shelled reovirus virion, suggestingwhat may be the minimal number of proteins needed to form a viable reovirus particle. FAKV evolved from more complex an-cestors by losing a genome segment and several virion proteins.
Human and livestock diseases are commonly caused by virusestransmitted from enzootic hosts by arthropod vectors (1).
Mosquito surveillance studies have been key to understanding theprevalence, distribution, and outbreak potential of the majority ofthese pathogens, as well as for the detection of novel pathogenicand nonpathogenic agents.
The family Reoviridae consists of double-stranded RNA(dsRNA) viruses with 9 to 12 genome segments. Reoviruses haveboth a wide geographic distribution and wide host range, havingbeen isolated from fungi, plants, insects, ticks, arachnids, fish,marine protists, crustaceans, mammals, and birds (2). They aredivided into two subfamilies, Spinareovirinae and Sedoreovirinae,which are classified based on their core structures. Spinareovirinaeviruses, such as the recently emerged human pathogen Melakavirus and the mammalian orthoreovirus type 3, are called “tur-reted” reoviruses, because they encode spikes (protrusions) on thesurface of the virion. The subfamily Sedoreovirinae contains the“nonturreted” viruses with multiple smooth shells, which includehuman and other animal pathogens, such as rotaviruses and orbi-
viruses (3, 4). In addition to this structural classification, aminoacid identities of �30% in the polymerase sequence and the pres-
Received 4 August 2014 Accepted 20 October 2014
Accepted manuscript posted online 29 October 2014
Citation Auguste AJ, Kaelber JT, Fokam EB, Guzman H, Carrington CVF, ErasmusJH, Kamgang B, Popov VL, Jakana J, Liu X, Wood TG, Widen SG, Vasilakis N,Tesh RB, Chiu W, Weaver SC. 2015. A newly isolated reovirus has the simplestgenomic and structural organization of any reovirus. J Virol 89:676 – 687.doi:10.1128/JVI.02264-14.
Editor: R. W. Doms
Address correspondence to Scott C. Weaver, [email protected].
A.J.A., J.T.K., and E.B.F. contributed equally.
Supplemental material for this article may be found at http://dx.doi.org/10.1128/JVI.02264-14.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.02264-14
676 jvi.asm.org January 2015 Volume 89 Number 1Journal of Virology
on Decem
ber 16, 2014 by MO
OD
Y M
ED
ICA
L LIBR
AR
Yhttp://jvi.asm
.org/D
ownloaded from

ence and sequence of conserved terminal nucleotide (nt) motifshave been used as criteria for the classification of genera withinthis family (2, 5–9).
The genus Dinovernavirus (subfamily Spinareovirinae) con-tains a single isolate designated Aedes pseudoscutellaris reovirus(APRV) that was isolated from the AP61 mosquito cell line (8),which was established in 1974 from A. pseudoscutellaris larvaefrom Fiji (10). Phylogenetic analysis of APRV sequences indicatesthat it represents a distinct genus within the family and that itsclosest genetic and structural relationships are with reoviruses inthe genera Cypovirus, Fijivirus, and Oryzavirus (8).
APRV is the only reovirus known to contain a 9-segmentdsRNA genome and, with the exception of cypoviruses (CPVs), isthe only other reovirus to have a single-shelled virion. The inner-most shell contains two interlocking peanut-shaped monomers ofthe major capsid protein, which are chemically but not structur-ally identical. Therefore, this T�1 layer has sometimes been de-scribed as “T�2” to emphasize the conformational nonequiva-lence of the two chemically identical monomers per asymmetricunit. Most reoviruses contain at least part of a second shell, com-prised mainly of trimers arranged in a T�13l lattice. In contrast tocypoviruses that form polyhedra/occlusion bodies, APRV is non-occluded (8). The structure of the cypovirus cytoplasmic polyhe-drosis virus is the highest-resolution single-particle cryoelectronmicroscopy (cryo-EM) structure solved to date (11). The single-shell capsid of cypoviruses is also thought to have an unusualstability (12, 13), which has been explained by the inclusion of thesmall protrusion (SP) domain in the major capsid protein, theN-terminal anchor of the major capsid protein, and the presenceof “clamp” (also called “cement”) proteins, which are homologs ofaquareovirus VP6 and orthoreovirus �2 (11, 14).
Here, we describe the first Dinovernavirus (family Reoviridae)to be detected in nature, which was isolated during a surveillancestudy in the Fako division of Cameroon in 2010 and tentativelydesignated Fako virus (FAKV). FAKV is related to both single-shelled and partially double-shelled viruses (3) and has minimalgenetic composition. These characteristics suggest that its virionstructure could shed light on the evolutionary history and struc-tural diversity of this family. A subnanometer three-dimensional(3D) structure of the FAKV virion determined by cryo-EM andsingle-particle reconstruction revealed a simplified virion archi-tecture with only four polypeptides per asymmetric unit. Wetherefore hypothesize that FAKV evolved from a more complexancestor by serial loss-of-function events within the subfamily.We also describe for the first time the genetic, ecological, andphenotypic characteristics of a naturally circulating dinovernavi-rus, as well as show evidence of mosquito-to-mosquito transmis-sion in nature.
MATERIALS AND METHODSMosquito collection and virus isolation. Between 15 July and 8 August2010, mosquitoes were collected at seven locations in Cameroon by usinga combination of landing collections and CDC light traps (John Hock,Gainesville, FL) baited with dry ice. After collection, mosquitoes weresorted by sex, date, and location of collection into conspecific pools rang-ing in size from one to 50 individuals. Pools were frozen at �80°C untilassayed for virus on cultures of Vero and C6/36 cells as described previ-ously (15, 16).
Virus purification and transmission electron microscopy. Aedes al-bopictus (C6/36) cells were grown to 80% confluence in eight 150-cm2
culture flasks and infected with FAKV at a multiplicity of infection (MOI)
of �0.1 PFU/cell. Three days postinfection, cell culture supernatants werecollected and virus was precipitated using 7% polyethylene glycol (PEG)8000 and 2.3% NaCl (wt/vol). Pellets containing virus were resuspendedin 4 ml of TEN (10 mM Tris-HCl, pH 7.5; 1 mM EDTA, pH 7.8; and 0.1 MNaCl) buffer, pH 7.8, and purified on a 20% to 70% continuous sucrosegradient by centrifugation at 210,000 � g for 1.5 h using an SW41Ti rotor(Beckman, Indianapolis, IN). The virus band was collected and applied toa 100-kDa Amicon filter (Millipore, Billerica, MA). Sucrose was then re-moved by 10 washes with TEN. Purified virus was harvested in 100 �l ofTEN with or without magnesium (10 mM) and stored at 4°C. Transmis-sion electron microscopy of thin sections from infected C6/36 cells andnegatively stained carbon-coated copper grids containing purified virionswas performed as previously described (16).
Virus propagation and RNA extraction. Virus was subsequently in-oculated onto an 80% confluent C6/36 cell monolayer in a 150-cm2 cul-ture flask. The culture medium was collected 6 days postinfection andcentrifuged at 1,860 � g to pellet cell debris. The supernatant was collectedand PEG was added to 7% (wt/vol), after which the mixture was incubatedat 4°C for 24 h prior to centrifugation at 3,100 � g for 40 min. Thesupernatant was removed and the virus pellet resuspended in 250 �l ofTEN buffer by repeated pipetting. Viral RNA was extracted from purifiedvirions using TRIzol LS (Life Technologies, Carlsbad, CA) according tothe manufacturer’s instructions.
Sequencing. The complete genome sequences for 4 isolates were de-termined using the Illumina HiSeq1000 platform (Illumina, Inc., San Di-ego, CA) as previously described (17). Intrahost genomic diversity wasassessed using VarScan v2 with a 3% threshold (18).
Sequence analysis. Nucleotide and open reading frame (ORF) se-quences were submitted to BLASTx and BLASTp to identify sequencesthat share homology with FAKV. Protein families were identified based onBLASTp analysis. Nucleotide and amino acid sequences for the ORFswithin all 9 segments were determined with EnzymeX using a 200-ntthreshold for each ORF (Aalsmeer, Netherlands).
Phylogenetic analysis. Previously published reovirus RNA-depen-dent RNA polymerase (RdRp) amino acid sequences (n � 29) were down-loaded from GenBank and aligned with the FAKV RdRp (ORF detectedwithin segment 2) translated sequence using the ClustalW program andthen manually adjusted in Se-Al (http://tree.bio.ed.ac.uk). Sequenceidentities were calculated using GENEIOUS v5.6 (Auckland, New Zea-land). A neighbor-joining (NJ) phylogenetic tree was constructed usingPAUP* version 4.0b (Sinauer Associates, Inc., Sunderland, MA, USA),with Mimoreovirus acting as an outgroup. Bootstrapping was used to as-sess the robustness of tree topologies using 1,000 replicate NJ trees.
Given the large sequence divergence within the subfamily Spinareo-virinae and to differentiate ancestral from derived clades, a Bayesian co-alescent analysis was performed using BEAST (19). BEAST utilizes aBayesian Markov chain Monte Carlo (MCMC) approach to jointly esti-mate trait diffusion patterns from sampled sequences, while concurrentlytaking into account phylogenetic uncertainty arising from both the se-quence data and trait diffusion processes. The statistical support for eachtrait under study is therefore obtained by implementing the analyses overall plausible trees, thereby removing the bias of using a parsimony ap-proach to assess trait evolution over a single phylogeny that may not betruly accurate. All GenBank reference RdRp sequences for the Spinareo-virinae were analyzed along with FAKV, assuming a Yule speciation pro-cess and an uncorrelated lognormal molecular clock (20). Tracer version1.5 (http://tree.bio.ed.ac.uk/software/tracer/) was used to monitor sta-tionarity and efficient topology mixing, as diagnosed using effective sam-ple size (ESS) statistics. TreeAnnotator version 1.8.0 (http://beast.bio.ed.ac.uk) was used to summarize the posterior tree distribution and FigTreeversion 1.3.1 (http://beast.bio.ed.ac.uk) to visualize the annotated maxi-mum clade credibility (MCC) tree.
Virus quantification. A plaque assay based on A. albopictus (C7/10)cells was used for quantification of FAKV as previously described (21),except that fixed cells were stained with 2% crystal violet.
Genetic and Structural Features of Dinovernaviruses
January 2015 Volume 89 Number 1 jvi.asm.org 677Journal of Virology
on Decem
ber 16, 2014 by MO
OD
Y M
ED
ICA
L LIBR
AR
Yhttp://jvi.asm
.org/D
ownloaded from

Cryoelectron microscopy. Purified virions were applied to 200-meshQuantifoil grids (2-�m hole size) and blotted inside a Vitrobot Mark IV(FEI Company) for 3.5 s at 6°C. From virions purified in TEN buffer only,1,385 particles at a pixel size of 2.78 Å/pixel were selected from data im-aged on a JEM3200FSC 300-kV cryoelectron microscope (JEOL Ltd., To-kyo, Japan) and a DE-20 camera (Direct Electron, LP, San Diego, CA) at16 frames per second. Motion correction and damage compensation wereperformed on a per-particle basis to combine information from the 32frames per specimen area (22, 23). From virions purified in TEN bufferwith supplemental magnesium, 2,464 full particles (intact capsid and ge-nome) were manually boxed with EMAN2 (24) from data imaged on aJEM2010F electron cryomicroscope (JEOL Ltd., Tokyo, Japan) and aUSA4000 charge-coupled device (CCD; Gatan, Pleasanton, CA) at 3.62Å/pixel.
Cryo-EM density map and interpretation. For 3D reconstruction,data derived from virus purified with and without Mg2 were divided intohalves, and each was refined independently. EMAN2.1 was used for auto-mated boxing, contrast transfer function correction, and generation ofdata-derived initial models. 2D alignment and 3D reconstruction wereperformed with EMAN2.1 (24) in the case of full particles and with mul-tipath simulated annealing (MPSA) (25) in the case of empty particles.The Fourier shell correlation (FSC) between the structures from indepen-dent halves was used to measure the resolution of the combined map (26).Visualization of the density map was performed in UCSF Chimera (27).All filtration operations were performed in EMAN2. To measure the vol-ume of regions of the asymmetric unit, semiautomatic segmentation ofthe capsid was performed using Segger (28).
To determine the location of secondary structural elements and clarifyprotein boundaries in the map, subvolumes were extracted, analyzed, andrepositioned in the original map. For each protein in the asymmetric unit,a subvolume extending 20 Å out from the location of its closest structur-ally characterized homolog from CPV was extracted. Gorgon was thenused to automatically skeletonize density and to manually identify sec-ondary structural elements in the extracted subvolume (29, 30).
Homology models for all three proteins were constructed using Mod-eler v9.8 (31, 32). Where secondary structural elements identified in Gor-gon corresponded to secondary structural elements in the homologymodel, the path between these elements was traced manually in Chimera.Regions of uncertainty were not traced or segmented; these include thetermini of the major capsid proteins. Finally, the partially traced pathswere used to define the borders of polypeptides in the original map.
Structural protein characterization. Proteins extracted from purifiedvirions were subjected to polyacrylamide gel electrophoresis under dena-turing and reducing conditions on a 4% to 20% gradient gel (Bio-Rad,Hercules, CA). Gels were stained with Coomassie blue or with the Silver-Quest silver staining kit (Life Technologies, Carlsbad, CA). Three bandswere excised from the silver-stained gel, and in-gel trypsin digestion wasperformed. Matrix-assisted laser desorption ionization–time of flightmass spectrometry (MALDI-TOF MS) was performed, and peptides wereidentified by searching the NCBI database or all predicted FAKV ORFswith Mascot (Matrix Science, Boston, MA).
In vitro culture of FAKV. Cell lines derived from African green mon-key kidney (Vero), baby hamster kidney (BHK-S), and mosquitoes (A.albopictus; C6/36 and C7/10) were inoculated with FAKV at a multiplicityof infection of 10 PFU/cell (titers based on C7/10 cell plaque assays), usingcell culture supernatant from infected C6/36 cells as the inoculum. Mos-quito cultures were incubated at 28°C and observed for cytopathic effects(CPE) for 7 days; mammalian cells were maintained at 37°C and observedfor 14 days. Dulbecco’s minimal essential medium (DMEM) containing5% fetal bovine serum (FBS), 1% penicillin-streptomycin, and 1% tryp-tose phosphate broth was used to maintain mosquito cells postinfection,and DMEM containing 2.5% FBS and 1% penicillin-streptomycin wasused to maintain mammalian cells. Culture fluid from each of the mam-malian cell lines was then serially blind-passaged three times, using undi-luted supernatant from the previous passage as the inoculum. Cells were
observed for CPE, and the third-passage supernatant was screened forFAKV RNA by reverse transcription (RT)-PCR using primers based onthe VP9 segment (available from authors on request). This method de-tected FAKV RNA at a titer of 0.01 PFU/ml. One-step RT-PCRs wereperformed with the Titan one-step RT-PCR kit (Roche Diagnostics, Indi-anapolis, IN).
Inoculation of mice. Culture medium from C6/36 cells infected withFAKV was used to inoculate intracerebrally 10 2-day-old CD1 mice (�105
PFU/ml). Mice were observed for signs of illness for 14 days. Animalexperiments were approved by the UTMB Institutional Animal Care andUse Committee (IACUC), protocol number 9505045.
Accession numbers. FAKV genomic sequences were submitted toGenBank under accession numbers KM978409 to KM978450. The Fakovirus full-virion structure was deposited in the Electron Microscopy DataBank under accession no. EMD-6002 and the empty-capsid structure un-der accession number EMD-6001.
RESULTSMosquito collection and FAKV isolation. A total of 138 poolscontaining 2,691 mosquitoes representing 17 species (see Table S1in the supplemental material) were collected and screened for ar-boviruses by CPE assays on Vero and C6/36 cells. Ten cytopathicviruses were isolated via screening on C6/36 cells (Table 1), but noCPE were observed on Vero cells. Locations and GPS coordinatesin decimal degrees where the 10 isolates were made include Ekona(4.12521, 9.19569), Malende (4.20305, 9.25555), and Muea(4.10224, 9.18398). Cytopathic effects were observed 4 dayspostinfection in both C6/36 and C7/10 cells (Fig. 1), which aredifferent clones of an A. albopictus cell line. These viruses couldnot be identified using standard immunofluorescence techniquesagainst commonly detected, medically important orthobunyavi-ruses, flaviviruses, alphaviruses, rhabdoviruses, and orbiviruses.To assess whether the observed CPE were a result of virus infec-tion, transmission electron microscopy of ultrathin sections ofinfected C6/36 cell monolayers was performed and revealed thepresence of a single-shelled, turreted virus that was similar in sizeand structure to turreted reoviruses such as cypoviruses, dinover-naviruses, and oryzaviruses. Virus particles clustered primarily inthe host cell cytoplasm and inside cytoplasmic vacuoles (see Fig.S1A and B in the supplemental material). Negative-stain imagingsuggested that FAKV virions were approximately 450 to 500 Å indiameter and were icosahedral in shape, with 6 of the 12 largeturrets visible in projection images (see Fig. S1C).
Analysis of FAKV nucleotide sequences. Illumina sequencingperformed on purified viral RNA revealed the presence of a nine-
TABLE 1 Viruses isolated in this study
Name ofvirus
Straindesignation Source mosquito species
Sourcegender
FAKVa CSW101 Eretmapodites dracaenae andEretmapodites inornatus
Male
FAKV CSW117 Aedes albopictus MaleFAKV CSW52 Aedes albopictus FemaleFAKV CSW69 Aedes simpsoni FemaleFAKV CSW72 Aedes albopictus FemaleFAKV CSW77 Aedes albopictus FemaleFAKV CSW79 Aedes albopictus FemaleFAKV CSW84 Aedes albopictus FemaleFAKV CSW87 Aedes africanus FemaleFAKV CSW90 Eretmapodites dracaenae Femalea FAKV, Fako virus.
Auguste et al.
678 jvi.asm.org January 2015 Volume 89 Number 1Journal of Virology
on Decem
ber 16, 2014 by MO
OD
Y M
ED
ICA
L LIBR
AR
Yhttp://jvi.asm
.org/D
ownloaded from

segment dsRNA genome totaling 23,170 nt. Approximately 41%of the reads in the prototype CSW77 sample mapped to the viralgenome, resulting in about 5.78 million read pairs mapped out of14.1 million total in the sample. The percentage of reads mappingto the viral genome in the other 3 strains ranged from 3.5% to20.5%. Host and viral sequences were also extracted from the re-sultant sequencing reads in an attempt to determine the presenceof any additional viral sequences, but none was detected in any ofthe 4 samples tested. For prototype strain CSW77, BLASTxsearches on translated amino acid sequences indicated the pres-ence of a Dinovernavirus (family Reoviridae) tentatively desig-nated Fako virus (FAKV), which belongs to a recently describedgenus containing a single, insect-specific virus known as Aedespseudoscutellaris reovirus (APRV) (8). Based on the RdRp, FAKVshared the highest amino acid sequence identity, at 89.7%, withAPRV, followed by the cypoviruses with �20% similarity (seeTable S2 in the supplemental material). Interestingly, submissionof FAKV sequences to UniProt Metagenomic and EnvironmentalSequences (UniMES) resulted in no matches, suggesting this virushas not been detected during previous metagenomic studies. All10 isolates of FAKV shared �99% sequence identity across 850 ntof the segment 9 sequenced, which suggests that they belong to asingle replicating lineage. This was further supported by completegenome sequencing of 4 additional isolates.
Table 2 shows the genome organization of the prototype FAKVstrain, CSW77. Segment names 1 through 9 were designated basedon similarity in sequence identity to APRV. The lengths of thevarious segments were very similar between FAKV and APRV,with the majority of the segments differing by only 1 to 3 nucleo-tides, except for segments 4 (28-nt difference) and 9 (7-nt differ-ence). The terminal sequences of FAKV were conserved, begin-ning with AGU at the 5= end and ending with AGU at the 3= end(Table 2). This pattern is conserved among the dinovernaviruses,cypoviruses, and fijiviruses.
We evaluated the intrahost genomic diversity of FAKV to de-termine if it increases its genetic diversity by incorporating largenumbers of single-nucleotide polymorphisms (SNPs), insertions,or deletions. We performed this analysis by realigning all Illuminasequencing reads against the viral genome. The results indicatedonly a few SNPs (see Table S4 in the supplemental material), sug-gesting limited intrahost sequence diversity.
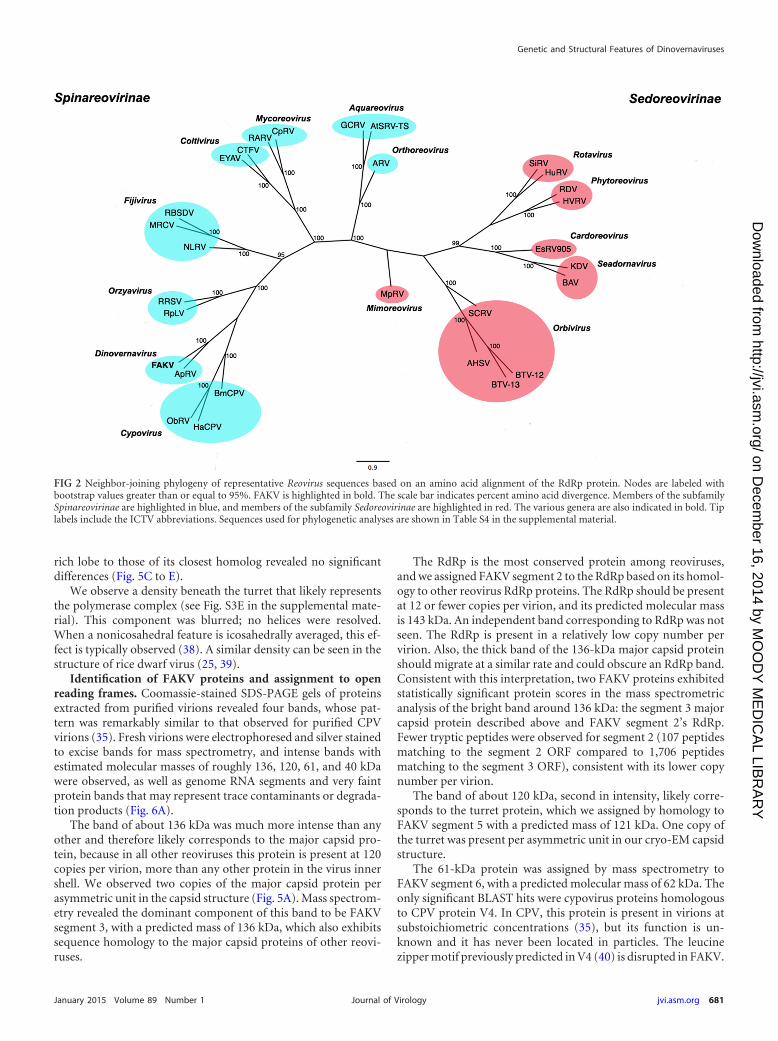
Phylogenetic analysis of FAKV. A neighbor-joining (NJ) phy-logeny based on RdRp amino acid sequences of 29 representativemembers of the family Reoviridae and prototype strain CSW77provided further support (i.e., 100% bootstrap) for FAKV’s as-
signment to the genus Dinovernavirus (Fig. 2). This phylogeneticposition is also supported by the inferred Bayesian MCC phylog-eny based on representative Spinareovirinae RdRp sequences (Fig.3). Table S3 in the supplemental material shows the species used inthe phylogenetic analyses. There was no evidence of reassortment,as all FAKV’s segments were more similar to the correspondingsegments of APRV (71 to 85% nucleotide identity) than they wereto segments of other reoviruses.
FAKV phenotypic characteristics. FAKV was detected in 3mosquito species within the genus Aedes and at least one specieswithin the genus Eretmapodites (Table 1); it was not detected inany of the other 4 mosquito genera tested, including known Culexand Anopheles arboviral vectors. FAKV was detected in male spec-imens of both A. albopictus and Eretmapodites dracaenae, suggest-ing vertical or venereal transmission in nature. Whether plants,vertebrates, or other invertebrates may play a role in FAKV’stransmission requires further testing.
Unlike APRV, the prototype virus of the Dinovernavirus genus,FAKV, caused cytopathic effects in mosquito cells; but like APRV,it appears to lack the ability to replicate in mammalian cells. Cy-topathic effects were apparent by day 5 postinfection in both A.albopictus cell lines tested. Figure 1 shows the typical CPE (delayedreplication and cell aggregation) and plaques created by FAKVinfection in C7/10 cells. Interestingly, FAKV lost the ability tocause CPE in C6/36 cells upon serial passaging (�5 passages) inthat line. Sequencing these multiply passaged strains might lead tothe identification of mutations associated with reduced cyto-pathogenicity. These results suggest that the failure of APRV tocause CPE reflects adaptation to replication in cell culture due toserial passaging in a cell line that was first established in 1974.Another difference observed between APRV and FAKV was thatAPRV lost infectivity after freezing at �20°C or �80°C, but FAKVremained infectious and showed no decline in titer following suchtreatment (stored for up to 2 years at �80°C). Treatment withvarious concentrations of NP-40 detergent did not abolish FAKVinfectivity (data not shown).
Lack of FAKV replication in vertebrate cells. FAKV did notproduce CPE and was not detected by RT-PCR after serial, blindpassages in either of the mammalian cell lines tested. Further-more, none of the 10 intracerebrally inoculated, 2-day-old miceshowed signs of illness or disease over the 14-day observationperiod. Our preliminary characterization of FAKV as an insect-specific reovirus supports previous observations with APRV andthe insect-specific nature of this genus.
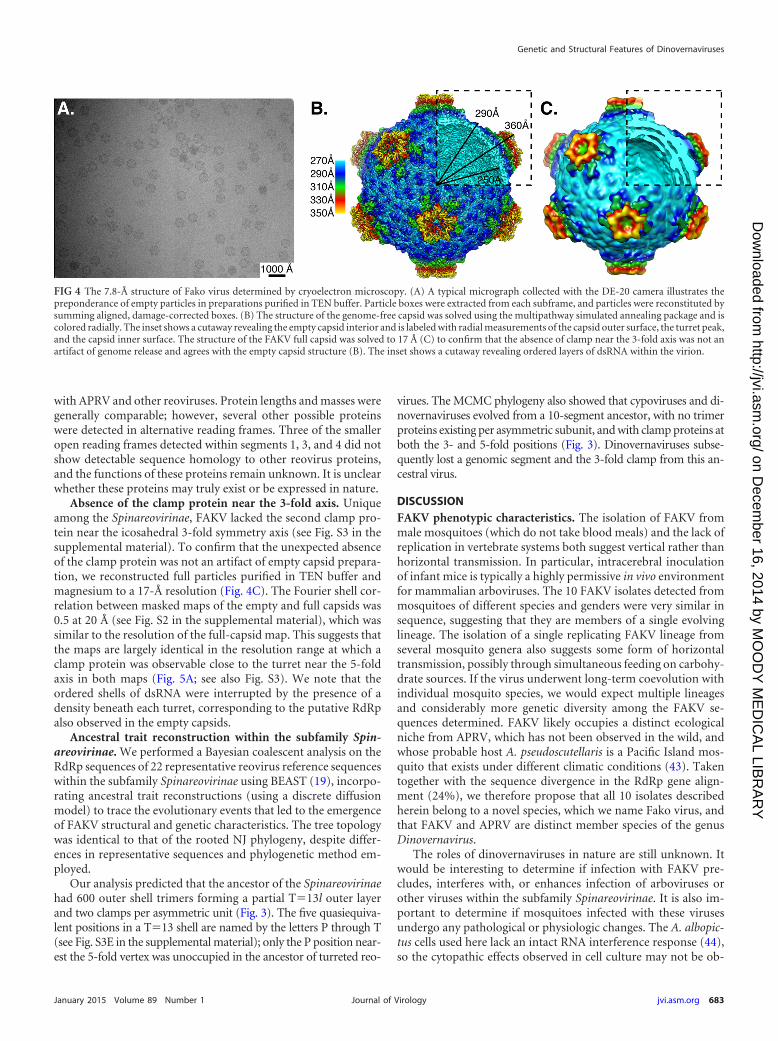
3D map of genome-free FAKV capsids. Cryo-EM micro-
FIG 1 Phase-contrast micrograph showing the cytopathic effects of CSW77 in representative cell lines: (A) negative control for C7/10; (B) C7/10 cells infectedwith CSW77. Note the significant syncytium formation and cell adhesion postinfection. (C) Typical plaques observed by CSW77 infection on C7/10 cells.
Genetic and Structural Features of Dinovernaviruses
January 2015 Volume 89 Number 1 jvi.asm.org 679Journal of Virology
on Decem
ber 16, 2014 by MO
OD
Y M
ED
ICA
L LIBR
AR
Yhttp://jvi.asm
.org/D
ownloaded from

graphs of FAKV exhibited particles with a diameter of roughly 580Å, not including the protruding spikes (Fig. 4A). Both genome-associated and genome-free particles were observed, with ge-nome-free capsids predominating, especially in viral preparationspurified in the presence of EDTA. However, adding 50 mM EDTAto purified samples prepared without EDTA did not change theratio of genome-associated to genome-free particles after 1- or24-h incubation at 4°C (data not shown). This suggests that prep-aration conditions other than cation concentration contributed tothis ratio of empty to full particles. The structure of the FAKVgenome-free capsid was determined to a 7.8-Å resolution (Fig. 4B)by single-particle cryoelectron microscopy using icosahedral av-eraging and was deposited in the electron microscopy databank(EMD-6001). The resolution was estimated by independently re-constructing two halves of the data set (33), known as the currentgold standard (see Fig. S2 in the supplemental material). -Heliceswere well resolved in all polypeptides of the capsid, but the indi-vidual strands within �-sheets were not differentiated (Fig. 5).
FAKV was determined to be a single-shelled reovirus withgross morphology similar to that of Bombyx mori CPV or to riceragged stunt virus (RRSV) (see Fig. S3 in the supplemental mate-rial) (3, 34, 35). Each of the 60 asymmetric units in FAKV can besegmented into 4 density regions using Chimera and Gorgon,which are annotated in different colors (Fig. 5A). Similar resultswere obtained using Segger segmentation software (see Fig. S4).Two of these regions (cyan and yellow) formed an icosahedralshell with inner and outer radii of approximately 250 and 290 Å,respectively (Fig. 4B). When these densities were computationallyextracted and aligned to each other, they agreed to 7.9 Å at an FSCof 0.5, which is close to the resolution of the map. This structure isconsistent with the organization of all reoviruses, whereby eachasymmetric unit contains two copies of the major capsid proteinin similar conformations for a total of 120 monomers per capsid.Among the 26 -helices and 8 �-sheets identified with Gorgon, all-helices and 5 �-sheets of this major capsid protein were alsoseen in the major capsid protein of CPV, with which the predictedopen reading frame of FAKV segment 3 shares 24% amino acididentity (Fig. 5K).
The FAKV shell was decorated with 12 turrets protruding 360Å from the center of the particle, each with a radius of 75 Å. Aprotruding density occupying about 50,000 to 60,000 Å3 occurredonce per asymmetric unit adjacent to the turret (Fig. 4B and 5Aand F; see also Fig. S3 in the supplemental material). This regionwas occupied by the clamp (also termed “cement”) protein incypovirus (large protrusion protein [LPP]) (12) (Fig. 5B; see alsoFig. S3C), aquareovirus (VP6) (14), and orthoreovirus (�2) (4).When Gorgon was used to identify secondary structural elementsin this density (Fig. 5H), locations of 9 of 12 -helix and 1 of 2�-sheets matched to those in the CPV LPP clamp protein (Fig. 5G)with which it shared 23% amino acid identity. In contrast, fewerthan half of its helices and no �-sheets were shared with theaquareovirus VP6 clamp protein, with which it shared no se-quence homology detectable by BLASTp (36).
The organization of major capsid protein’s helix-rich lobe ishighly conserved among reoviruses (37), making it a useful bench-mark to validate the reconstruction and segmentation steps usedin our cryo-EM studies of the genome-free FAKV virion. In con-trast to the usually structurally divergent clamp protein, compar-ison of the segmented -helices in the major capsid protein helix-T
AB
LE2
Len
gth
sof
Fako
viru
sge
nom
ese
gmen
ts,p
redi
cted
prot
ein
s,an
dco
nse
rved
term
inal
sequ
ence
sat
the
5=an
d3=
ends
a
Segm
ent
orco
nse
nsu
sse
quen
ceL
engt
h(b
p)O
RF
len
gth
(s)
(OR
Fpo
siti
on;f
ram
e)P
rote
inle
ngt
h(s
)in
aa(m
ass
inkD
a)5=
-NC
Rle
ngt
h5=
-en
dse
quen
ce3=
-NC
Rle
ngt
h3=
-en
dse
quen
ce
13,
819
3,75
0(1
84–3
753;
F1)
and
498
(180
3–23
00;F
2)
1,18
9(1
31.1
)an
d16
5(1
8.3)
183
AG
UU
UA
AT
TC
CC
66U
UG
AU
CC
UA
AG
U2
3,75
23,
702
(28–
3729
;F
1)an
d22
2(3
–224
;F�
1)1,
233
(143
.3)
and
73(8
.0)
27A
GU
UA
AA
CC
GC
A23
UU
GU
UU
UU
AA
GU
33,
730
3,60
9(3
4–36
42;F
1)
and
279
(98–
376;
F2)
1,20
2(1
36.5
)an
d92
(10.
9)33
AG
UU
UA
AA
AC
CC
88U
UU
GA
UA
CU
AG
U4
3,40
33,
036
(269
–330
4;F
2)an
d31
2(2
5–33
6;F
1)1,
011
(117
.2)
and
103
(12.
2)26
8A
GU
UU
AA
AA
AC
C99
UU
AA
UC
CU
AA
GU
53,
227
3,17
1(1
7–31
87;F
2)
and
318
(928
–124
5;F�
3)1,
056
(121
.1)
and
105
(11.
6)16
AG
UU
AA
AA
CC
AC
40U
UU
AG
UA
AU
AG
U6
1,77
81,
623
(25–
1647
;F
1)54
0(6
2.2)
24A
GU
UU
AA
AA
CC
C13
1U
UU
GA
UA
CU
AG
U7
1,17
01,
047
(23–
1069
;F
2)34
8(3
9.6)
22A
GU
UA
AA
AA
CC
A10
1U
UU
AG
UA
AU
AG
U8
1,15
11,
038
(17–
1054
;F
2),3
66(5
25–8
90;F
3)
,an
d20
1(6
3–26
3;F�
1)34
5(3
9.6)
,121
(14.
4),a
nd
66(7
.9)
16A
GU
UU
AA
AU
CC
U97
UU
UG
AU
AA
UA
GU
91,
140
837
(89–
925;
F2)
278
(32.
1)88
AG
UU
AA
AA
CC
CA
215
UU
UA
GU
AA
UA
GU
Con
sen
sus
sequ
ence
AG
UU
----
----
----
----
---
----
----
----
----
-AG
Ua
Bol
dfac
ein
dica
tes
con
serv
edte
rmin
aln
ucl
eoti
des.
NC
R,n
onco
din
gre
gion
.
Auguste et al.
680 jvi.asm.org January 2015 Volume 89 Number 1Journal of Virology
on Decem
ber 16, 2014 by MO
OD
Y M
ED
ICA
L LIBR
AR
Yhttp://jvi.asm
.org/D
ownloaded from
rich lobe to those of its closest homolog revealed no significantdifferences (Fig. 5C to E).
We observe a density beneath the turret that likely representsthe polymerase complex (see Fig. S3E in the supplemental mate-rial). This component was blurred; no helices were resolved.When a nonicosahedral feature is icosahedrally averaged, this ef-fect is typically observed (38). A similar density can be seen in thestructure of rice dwarf virus (25, 39).
Identification of FAKV proteins and assignment to openreading frames. Coomassie-stained SDS-PAGE gels of proteinsextracted from purified virions revealed four bands, whose pat-tern was remarkably similar to that observed for purified CPVvirions (35). Fresh virions were electrophoresed and silver stainedto excise bands for mass spectrometry, and intense bands withestimated molecular masses of roughly 136, 120, 61, and 40 kDawere observed, as well as genome RNA segments and very faintprotein bands that may represent trace contaminants or degrada-tion products (Fig. 6A).
The band of about 136 kDa was much more intense than anyother and therefore likely corresponds to the major capsid pro-tein, because in all other reoviruses this protein is present at 120copies per virion, more than any other protein in the virus innershell. We observed two copies of the major capsid protein perasymmetric unit in the capsid structure (Fig. 5A). Mass spectrom-etry revealed the dominant component of this band to be FAKVsegment 3, with a predicted mass of 136 kDa, which also exhibitssequence homology to the major capsid proteins of other reovi-ruses.
The RdRp is the most conserved protein among reoviruses,and we assigned FAKV segment 2 to the RdRp based on its homol-ogy to other reovirus RdRp proteins. The RdRp should be presentat 12 or fewer copies per virion, and its predicted molecular massis 143 kDa. An independent band corresponding to RdRp was notseen. The RdRp is present in a relatively low copy number pervirion. Also, the thick band of the 136-kDa major capsid proteinshould migrate at a similar rate and could obscure an RdRp band.Consistent with this interpretation, two FAKV proteins exhibitedstatistically significant protein scores in the mass spectrometricanalysis of the bright band around 136 kDa: the segment 3 majorcapsid protein described above and FAKV segment 2’s RdRp.Fewer tryptic peptides were observed for segment 2 (107 peptidesmatching to the segment 2 ORF compared to 1,706 peptidesmatching to the segment 3 ORF), consistent with its lower copynumber per virion.
The band of about 120 kDa, second in intensity, likely corre-sponds to the turret protein, which we assigned by homology toFAKV segment 5 with a predicted mass of 121 kDa. One copy ofthe turret was present per asymmetric unit in our cryo-EM capsidstructure.
The 61-kDa protein was assigned by mass spectrometry toFAKV segment 6, with a predicted molecular mass of 62 kDa. Theonly significant BLAST hits were cypovirus proteins homologousto CPV protein V4. In CPV, this protein is present in virions atsubstoichiometric concentrations (35), but its function is un-known and it has never been located in particles. The leucinezipper motif previously predicted in V4 (40) is disrupted in FAKV.
FIG 2 Neighbor-joining phylogeny of representative Reovirus sequences based on an amino acid alignment of the RdRp protein. Nodes are labeled withbootstrap values greater than or equal to 95%. FAKV is highlighted in bold. The scale bar indicates percent amino acid divergence. Members of the subfamilySpinareovirinae are highlighted in blue, and members of the subfamily Sedoreovirinae are highlighted in red. The various genera are also indicated in bold. Tiplabels include the ICTV abbreviations. Sequences used for phylogenetic analyses are shown in Table S4 in the supplemental material.
Genetic and Structural Features of Dinovernaviruses
January 2015 Volume 89 Number 1 jvi.asm.org 681Journal of Virology
on Decem
ber 16, 2014 by MO
OD
Y M
ED
ICA
L LIBR
AR
Yhttp://jvi.asm
.org/D
ownloaded from

The 40-kDa band, which had higher intensity than the 61-kDaband, was assigned by mass spectrometry to FAKV segment 7,whose predicted molecular mass is 40 kDa. The only detectablehomologous sequence was APRV segment 7, which was previ-ously annotated as a putative nonstructural protein with noknown homologs (8). The cryo-EM structure of FAKV containedthree unique density regions per asymmetric unit, correspondingto the major capsid protein (segment 3), the turret protein (seg-ment 5), and the clamp protein. These three unique density re-gions should correspond to the three most abundant proteinsamong purified virions. This suggests that segment 7 is the FAKVclamp protein.
In support of the assignment of segment 7 as the FAKV clamp,we measured the volume of the clamp directly and by comparingthe clamp volume to the capsid volume. Assuming a specific vol-ume of 0.73 ml/g (41), the density of the putative clamp proteinregion would be occupied by roughly 40 to 50 kDa of protein.Local contrast variations were normalized in EMAN2.1, and theratio of clamp protein volume to total capsid volume was mea-
sured as 9.74%. Since an asymmetric unit consists of one turret of121 kDa and two major capsid proteins at 136 kDa each, the clampmass would be roughly 42 kDa. Therefore, of all the bands ob-served by electrophoresis of purified virions, the abundant bandcorresponding to segment 7 matches the measured clamp volumebetter than any other band.
In Oryzavirus, a 43-kDa clamp protein is produced from a67-kDa precursor (42), a process which also occurs in CPV (40).However, both the volume measurement by cryo-EM and densi-tometry of electrophoresed purified virions yielded estimates ofthe clamp mass that are consistent with the full mass of the seg-ment 7 open reading frame.
In summary, four polypeptides were identified and assigned toORFs in the FAKV asymmetric unit (Fig. 5A): the turret protein,two subunits of the major capsid protein dimer, and a clamp pro-tein adjacent to the turret structure. The RdRp and a protein ofunknown function were also present at less than one copy perasymmetric unit. Figure 6B indicates the putative functions of thevarious predicted proteins based on our study and comparisons
FIG 3 Bayesian MCC tree for the subfamily Spinareovirinae based on 1,232 amino acids of the RdRp protein. Terminal branches of the tree are colored accordingto the trimer positions of the taxon at the tip. Internal branches are colored according to the most probable (modal) trimer positions of their parental nodes. Allinternal nodes had posterior probabilities (clade credibilities) of �0.95. Clamp proteins, their positions, and their probabilities of occurrence at the relevantnodes are indicated in bold. The number of genomic segments each genus contains is highlighted in color. The scale bar represents nucleotide substitutions persite.
Auguste et al.
682 jvi.asm.org January 2015 Volume 89 Number 1Journal of Virology
on Decem
ber 16, 2014 by MO
OD
Y M
ED
ICA
L LIBR
AR
Yhttp://jvi.asm
.org/D
ownloaded from
with APRV and other reoviruses. Protein lengths and masses weregenerally comparable; however, several other possible proteinswere detected in alternative reading frames. Three of the smalleropen reading frames detected within segments 1, 3, and 4 did notshow detectable sequence homology to other reovirus proteins,and the functions of these proteins remain unknown. It is unclearwhether these proteins may truly exist or be expressed in nature.
Absence of the clamp protein near the 3-fold axis. Uniqueamong the Spinareovirinae, FAKV lacked the second clamp pro-tein near the icosahedral 3-fold symmetry axis (see Fig. S3 in thesupplemental material). To confirm that the unexpected absenceof the clamp protein was not an artifact of empty capsid prepara-tion, we reconstructed full particles purified in TEN buffer andmagnesium to a 17-Å resolution (Fig. 4C). The Fourier shell cor-relation between masked maps of the empty and full capsids was0.5 at 20 Å (see Fig. S2 in the supplemental material), which wassimilar to the resolution of the full-capsid map. This suggests thatthe maps are largely identical in the resolution range at which aclamp protein was observable close to the turret near the 5-foldaxis in both maps (Fig. 5A; see also Fig. S3). We note that theordered shells of dsRNA were interrupted by the presence of adensity beneath each turret, corresponding to the putative RdRpalso observed in the empty capsids.
Ancestral trait reconstruction within the subfamily Spin-areovirinae. We performed a Bayesian coalescent analysis on theRdRp sequences of 22 representative reovirus reference sequenceswithin the subfamily Spinareovirinae using BEAST (19), incorpo-rating ancestral trait reconstructions (using a discrete diffusionmodel) to trace the evolutionary events that led to the emergenceof FAKV structural and genetic characteristics. The tree topologywas identical to that of the rooted NJ phylogeny, despite differ-ences in representative sequences and phylogenetic method em-ployed.
Our analysis predicted that the ancestor of the Spinareovirinaehad 600 outer shell trimers forming a partial T�13l outer layerand two clamps per asymmetric unit (Fig. 3). The five quasiequiva-lent positions in a T�13 shell are named by the letters P through T(see Fig. S3E in the supplemental material); only the P position near-est the 5-fold vertex was unoccupied in the ancestor of turreted reo-
virues. The MCMC phylogeny also showed that cypoviruses and di-novernaviruses evolved from a 10-segment ancestor, with no trimerproteins existing per asymmetric subunit, and with clamp proteins atboth the 3- and 5-fold positions (Fig. 3). Dinovernaviruses subse-quently lost a genomic segment and the 3-fold clamp from this an-cestral virus.
DISCUSSIONFAKV phenotypic characteristics. The isolation of FAKV frommale mosquitoes (which do not take blood meals) and the lack ofreplication in vertebrate systems both suggest vertical rather thanhorizontal transmission. In particular, intracerebral inoculationof infant mice is typically a highly permissive in vivo environmentfor mammalian arboviruses. The 10 FAKV isolates detected frommosquitoes of different species and genders were very similar insequence, suggesting that they are members of a single evolvinglineage. The isolation of a single replicating FAKV lineage fromseveral mosquito genera also suggests some form of horizontaltransmission, possibly through simultaneous feeding on carbohy-drate sources. If the virus underwent long-term coevolution withindividual mosquito species, we would expect multiple lineagesand considerably more genetic diversity among the FAKV se-quences determined. FAKV likely occupies a distinct ecologicalniche from APRV, which has not been observed in the wild, andwhose probable host A. pseudoscutellaris is a Pacific Island mos-quito that exists under different climatic conditions (43). Takentogether with the sequence divergence in the RdRp gene align-ment (24%), we therefore propose that all 10 isolates describedherein belong to a novel species, which we name Fako virus, andthat FAKV and APRV are distinct member species of the genusDinovernavirus.
The roles of dinovernaviruses in nature are still unknown. Itwould be interesting to determine if infection with FAKV pre-cludes, interferes with, or enhances infection of arboviruses orother viruses within the subfamily Spinareovirinae. It is also im-portant to determine if mosquitoes infected with these virusesundergo any pathological or physiologic changes. The A. albopic-tus cells used here lack an intact RNA interference response (44),so the cytopathic effects observed in cell culture may not be ob-
FIG 4 The 7.8-Å structure of Fako virus determined by cryoelectron microscopy. (A) A typical micrograph collected with the DE-20 camera illustrates thepreponderance of empty particles in preparations purified in TEN buffer. Particle boxes were extracted from each subframe, and particles were reconstituted bysumming aligned, damage-corrected boxes. (B) The structure of the genome-free capsid was solved using the multipathway simulated annealing package and iscolored radially. The inset shows a cutaway revealing the empty capsid interior and is labeled with radial measurements of the capsid outer surface, the turret peak,and the capsid inner surface. The structure of the FAKV full capsid was solved to 17 Å (C) to confirm that the absence of clamp near the 3-fold axis was not anartifact of genome release and agrees with the empty capsid structure (B). The inset shows a cutaway revealing ordered layers of dsRNA within the virion.
Genetic and Structural Features of Dinovernaviruses
January 2015 Volume 89 Number 1 jvi.asm.org 683Journal of Virology
on Decem
ber 16, 2014 by MO
OD
Y M
ED
ICA
L LIBR
AR
Yhttp://jvi.asm
.org/D
ownloaded from

served in vivo or in the natural vectors identified in this study.Intrathoracic inoculation of Aedes aegypti mosquitoes with FAKVhad no visible effect on these mosquitoes or their survivability.
Both the in vitro and in vivo systems used in this study representhighly permissive environments for the replication and detectionof mammalian arboviruses. The inability to detect FAKV or FAKVgenomic RNA from these systems provides strong support for theinsect-specific nature of this genus. However, further studies us-ing a more diverse array of vertebrate cell cultures, in vivo models,or perhaps more sensitive techniques, such as real-time PCRs, arenecessary to truly preclude the possibility of FAKV replication in
vertebrate cells. Also of importance is the question of whether ornot FAKV has the ability to infect plants. This would add anotherlevel of complexity within the FAKV transmission cycle. Mosqui-toes do feed on the nectar of plants as a primary sugar source;hence, the opportunity exists for transmission between thesehosts, as does the possibility for horizontal transmission betweenmosquitoes.
Stability of the T�1 capsid. FAKV, like CPV, is a true single-shelled virus; it lacks trimers at any of the T�13l positions. Themechanism of capsid stability in CPV has been previously inves-tigated, because this has hitherto been the only reovirus for which
FIG 5 Protein composition of FAKV virions. Three segmented asymmetric units of FAKV (A) are colored by chain with different tints of the same color,representing symmetry-related copies of a protein. There are four proteins per asymmetric unit: a turret (purple) at the 5-fold axis, a dimer of the major capsidprotein (blue and yellow), and a clamp protein (red) that binds to the major capsid protein surface. In contrast, the previously published structure of CPV (B)(11) and all other turreted reoviruses contain one clamp for each major capsid protein (red and green), and some have an additional clamp spanning the 2-foldaxis. The turret protein (C to E), clamp protein (F to H), and major capsid protein (I to K) are contrasted between CPV and FAKV. The FAKV density map ofeach protein (C, F, I; colored as described above) was segmented from the 7.8-Å empty capsid reconstruction. A CPV map was obtained from the EM Data Bank(EMDB5256) (11) and filtered to similar resolution (D, G, J; gray). The FAKV-helices (light green) and �-sheets (dark green) were compared to the -helices(blue) and �-strands (cyan) of the CPV atomic structure (PDB 3IZX). While most secondary structural elements in the turret (E) and major capsid protein (K)are conserved, the clamp protein varies greatly around a conserved helix-barrel core (H).
Auguste et al.
684 jvi.asm.org January 2015 Volume 89 Number 1Journal of Virology
on Decem
ber 16, 2014 by MO
OD
Y M
ED
ICA
L LIBR
AR
Yhttp://jvi.asm
.org/D
ownloaded from

an inner-shell-only structure is stable (13, 45). Among the pro-posed factors conferring this stability include (i) the insertion ofthe SP domain (12), (ii) the N-terminal anchor, which extends theinterface between the two major capsid chains (11), and (iii) thetwo clamp proteins (4). While visualization of the N-terminalanchor is not possible at this resolution, FAKV virions are func-tional and stable without an SP domain insertion (Fig. 4C) andhalf the usual complement of clamp proteins. Interestingly, ourresults show that several of the structural features previouslythought to account for the stability and maintenance of a single-shelled reovirus are in fact dispensable. Fewer proteins than pre-viously proposed are necessary for its stability. Further work isnecessary to investigate the specific biophysical mechanisms thatconfer the stability and determine the degree of stability of theFAKV capsid.
Evolution of the family Reoviridae. FAKV clusters togetherwith APRV (8) in the genus Dinovernavirus that contains 9-seg-ment turreted reoviruses. Because serial passages not only causemutations but may also result in major genetic changes, such asgenome segmentation, it was previously unclear if APRV origi-nated as a 10-segment virus and lost a genomic segment duringcell culture passaging (46). Previous work has also shown thatsome segmented viruses may have an accessory segment that is notnecessary for further replication of the virus (47). However, ourdata now confirm the existence of an extant, fully functional9-segment reovirus, the first nine-segment dsRNA genome virusto be observed in nature. The T�1 inner layer consists of twointerlocking peanut-shaped monomers. Each monomer has a he-lix-rich lobe and a mixed /� lobe. This fold and organization areconserved within the T�1 Totiviridae and Cystoviridae as well (48,49). Thus, Totiviridae, Cystoviridae, and Reoviridae are 3 familiessometimes nicknamed “T�2,” because they have 2 copies of themajor capsid protein per asymmetric unit. Currently, it is unclearwhether Reoviridae and/or Cystoviridae is the product of a hori-zontal gene transfer event combining a T�1 Totiviridae inner cap-
sid shell with a T�13l Birnaviridae outer shell (50) or whether theTotiviridae evolved by deletion of the T�13l layer from a morecomplex reovirus or cystovirus ancestor (48). Within the reovirusfamily, results of the Bayesian analyses suggest that FAKV evolvedfrom a double-shelled ancestor by loss of the outer T�13l coat oftrimers. Its sister genus Cypovirus is also single-shelled, but it car-ries polyhedrin, a trimeric protein whose central �-sandwich issuspiciously similar to the fold observed in the second-shell trim-ers of more basal reoviruses (51). Polyhedrin forms intracellularnanocrystals, which protect virions from harsh conditions in oc-clusions. FAKV is nonoccluded and lacks any homolog to thesetrimeric proteins. The most parsimonious explanation is that theT�13l capsid trimer lost its structural function in an ancestor ofsingle-shelled reoviruses, and then the gene was neofunctional-ized (into polyhedrin) in cypoviruses but deleted in dinovernavi-ruses.
Unique among all turreted reoviruses, the FAKV clamp pro-tein (corresponding to aquareovirus VP6 and orthoreovirus �2)binds only one site per asymmetric unit. In all structurally char-acterized turreted reoviruses, the clamp protein binds at least twodifferent sites, each interacting predominantly with one of the twomajor capsid proteins in an asymmetric unit: a 5-fold proximaland 3-fold proximal clamp. Orthoreoviruses contain a thirdclamp binding site near the 2-fold symmetry axis. These interac-tions are similar but not identical, as the clamp can also interactwith the turret or with dissimilarly oriented domains (3, 4, 11, 14).No protein density was observed that would sterically hinder theclamp from binding at its usual site. Because this clamp is presentnear the 5-fold axis at one copy per asymmetric unit, it must beproperly expressed and folded. We therefore conclude that subtlechanges to the binding interface, the clamp, or both are probablyresponsible for the evolution of this synapomorphy in FAKV. De-termining the structure of the FAKV capsid at atomic resolutionwill permit valuable comparisons to the structure of its closestrelative, allowing for a better understanding of how the clamp
FIG 6 Electrophoresis of FAKV proteins. (A) Purified FAKV virions were analyzed by SDS-PAGE and silver staining. Decreasing 10-fold dilutions in water ofthe same FAKV preparation are indicated by a triangle beneath lanes 1 to 3. (B) Putative open reading frames identified in the nine-segment genome of FAKVare colored to match their location in the virion. (C) Protein chains of one asymmetric unit of FAKV were segmented using Chimera and Gorgon. The folds andpositions of these chains were similar to the turret, major capsid, or clamp of other Spinareovirinae. Open reading frames and protein chains are coloredgoldenrod and teal for two copies of the major capsid protein, purple for the turret protein, and red for the clamp protein. Lines indicate assignment of anSDS-PAGE band to a structural ORF.
Genetic and Structural Features of Dinovernaviruses
January 2015 Volume 89 Number 1 jvi.asm.org 685Journal of Virology
on Decem
ber 16, 2014 by MO
OD
Y M
ED
ICA
L LIBR
AR
Yhttp://jvi.asm
.org/D
ownloaded from

usually tolerates differences in the binding site arising from thenonequivalent positions occupied by the capsid proteins on whichit sits.
In summary, FAKV arose from a more complex ancestor byserial loss-of-function events, including the following: the loss ofthe second shell, possibly through an Oryzavirus-like intermediatehaving only three of the usual 10 copies of the outer shell protein(3); the loss of one copy per asymmetric unit of the clamp protein;and the loss of one genomic segment from a 10-segment ancestor(Fig. 3). It is an interesting finding that the single-shelled reovi-ruses are those that are restricted to insect hosts.
Conclusions. We report the first detection and isolation of a9-segment reovirus from nature and present the first structuralstudies of a member of this genus. The complete genome se-quences presented here should be helpful in the development ofdetection methods to study the distribution of this genus, as wellas aid in further understanding the evolutionary history of thefamily Reoviridae. With only four polypeptides per asymmetricunit (one turret, two major capsid proteins, and one clamp) and 9segments, FAKV illustrates a minimal but functional reovirus anddemonstrates the dispensability of several key structural features.The detection and characterization of FAKV increases our knowl-edge of the known geographic distribution, genomic characteris-tics, genetic diversity, and host range of another group of insect-specific viruses as well as the family Reoviridae.
ACKNOWLEDGMENTS
This work was supported by NIH grant P41GM103832 to W.C., NIHcontract HHSN272201000040I/HHSN27200004/D04 to R.B.T. and N.V.,Robert Welch Foundation grant Q1242 to W.C., a fellowship from theJohn S. Dunn Foundation to E.B.F., and the Robert E. Shope InternationalFellowship in Infectious Diseases from the American Society of TropicalMedicine and Hygiene to A.J.A. A.J.A. is supported by the James W.McLaughlin endowment fund. J.T.K. is supported by the NanobiologyInterdisciplinary Graduate Training Program (NIBIB T32EB009379)through the Gulf Coast Consortia.
We thank Matthew Baker and Andrew Debevec for helpful sugges-tions, Jill Thompson at the UTMB Recombinant DNA core for assistingwith the next-generation sequencing, Xuemei Luo for performing massspectrometry experiments, and Naoyuki Miyazaki for furnishing the mapof rice ragged stunt virus.
Funding agencies had no role in study design, data collection andanalysis, the decision to publish, or preparation of the manuscript.
REFERENCES1. Weaver SC, Reisen WK. 2010. Present and future arboviral threats. An-
tiviral Res 85:328 –345. http://dx.doi.org/10.1016/j.antiviral.2009.10.008.2. Attoui H, Mertens PPC, Becnel J, Belaganahalli S, Bergoin M, Brussard
CP, Chappell Ciarlet JDM, del Vas M, Dermody TS, Dormitzer PR,Duncan R, Fcang Q, Graham R, Guglielmi KM, Harding RM, HillmanB, Makkay A, Marzachi C, Matthijnssens J, Milne RG, Mohd Jaafar F,Mori H, Noordeloos AA, Omura T, Patton JT, Rao S, Maan M, StoltzD, Suzuki N, Upadhyaya NM, Wei C, Zhou H. 2012. Family Reoviridae,p 541– 637. In King AMQ, Adams MJ, Carstens EB, Lefkowitz EJ (ed),Virus taxonomy: ninth report of the International Committee on Taxon-omy of Viruses. Academic Press, London, United Kingdom.
3. Miyazaki N, Uehara-Ichiki T, Xing L, Bergman L, Higashiura A, Na-kagawa A, Omura T, Cheng RH. 2008. Structural evolution of Reoviridaerevealed by Oryzavirus in acquiring the second capsid shell. J Virol 82:11344 –11353. http://dx.doi.org/10.1128/JVI.02375-07.
4. Reinisch KM, Nibert ML, Harrison SC. 2000. Structure of the reoviruscore at 3.6 Å resolution. Nature 404:960 –967. http://dx.doi.org/10.1038/35010041.
5. Anzola JV, Xu ZK, Asamizu T, Nuss DL. 1987. Segment-specific in-verted repeats found adjacent to conserved terminal sequences in wound
tumor virus genome and defective interfering RNAs. Proc Natl Acad Sci US A 84:8301– 8305. http://dx.doi.org/10.1073/pnas.84.23.8301.
6. Attoui H, Fang Q, Mohd Jaafar F, Cantaloube JF, Biagini P, de MiccoP, de Lamballerie X. 2002. Common evolutionary origin of aquareovi-ruses and orthoreoviruses revealed by genome characterization of goldenshiner reovirus, grass carp reovirus, striped bass reovirus, and golden idereovirus (genus Aquareovirus, family Reoviridae). J Gen Virol 83:1941–1951.
7. Attoui H, Mohd Jaafar F, Biagini P, Cantaloube JF, de Micco P, Murphy FA,de Lamballerie X. 2002. Genus Coltivirus (family Reoviridae): genomic andmorphologic characterization of Old World and New World viruses. ArchVirol 147:533–561. http://dx.doi.org/10.1007/s007050200005.
8. Attoui H, Mohd Jaafar F, Belhouchet M, Biagini P, Cantaloube JF, deMicco P, de Lamballerie X. 2005. Expansion of family Reoviridae toinclude nine-segmented dsRNA viruses: isolation and characterization ofa new virus designated Aedes pseudoscutellaris reovirus assigned to a pro-posed genus (Dinovernavirus). Virology 343:212–223. http://dx.doi.org/10.1016/j.virol.2005.08.028.
9. Attoui H, Jaafar FM, Belhouchet M, de Micco P, de Lamballerie X,Brussaard CP. 2006. Micromonas pusilla reovirus: a new member of thefamily Reoviridae assigned to a novel proposed genus (Mimoreovirus). JGen Virol 87:1375–1383. http://dx.doi.org/10.1099/vir.0.81584-0.
10. Varma MG, Pudney M, Leake CJ. 1974. Cell lines from larvae of Aedes(Stegomyia) malayensis colless and Aedes (S) pseudoscutellaris (Theobald)and their infection with some arboviruses. Trans R Soc Trop Med Hyg68:374 –382. http://dx.doi.org/10.1016/0035-9203(74)90152-7.
11. Yu X, Ge P, Jiang J, Atanasov I, Zhou ZH. 2011. Atomic model of CPVreveals the mechanism used by this single-shelled virus to economicallycarry out functions conserved in multishelled reoviruses. Structure 19:652– 661. http://dx.doi.org/10.1016/j.str.2011.03.003.
12. Zhou Z, Zhang H, Jakana J, Lu X, Zhang J. 2003. Cytoplasmic polyhe-drosis virus structure at 8 Å by electron cryomicroscopy structural basis ofcapsid stability and mRNA processing regulation. Structure 11:651– 663.http://dx.doi.org/10.1016/S0969-2126(03)00091-1.
13. Zhang H, Yu X-K, Lu X-Y, Zhang J-Q, Zhou ZH. 2002. Molecularinteractions and viral stability revealed by structural analyses of chemicallytreated cypovirus capsids. Virology 298:45–52. http://dx.doi.org/10.1006/viro.2002.1473.
14. Cheng L, Fang Q, Shah S, Atanasov IC, Zhou ZH. 2008. Subnanometer-resolution structures of the grass carp reovirus core and virion. J Mol Biol382:213–222. http://dx.doi.org/10.1016/j.jmb.2008.06.075.
15. Auguste AJ, Volk SM, Arrigo NC, Martinez R, Ramkissoon V, AdamsAP, Thompson NN, Adesiyun AA, Chadee DD, Foster JE, Travassos DaRosa AP, Tesh RB, Weaver SC, Carrington CV. 2009. Isolation andphylogenetic analysis of Mucambo virus (Venezuelan equine encephalitiscomplex subtype IIIA) in Trinidad. Virology 392:123–130. http://dx.doi.org/10.1016/j.virol.2009.06.038.
16. Vasilakis N, Forrester NL, Palacios G, Nasar F, Savji N, Rossi SL,Guzman H, Wood TG, Popov V, Gorchakov R, Gonzalez AV, HaddowAD, Watts DM, da Rosa AP, Weaver SC, Lipkin WI, Tesh RB. 2013.Negevirus: a proposed new taxon of insect-specific viruses with wide geo-graphic distribution. J Virol 87:2475–2488. http://dx.doi.org/10.1128/JVI.00776-12.
17. Auguste AJ, Carrington CV, Forrester NL, Popov VL, Guzman H,Widen SG, Wood TG, Weaver SC, Tesh RB. 2014. Characterization of anovel Negevirus and a novel Bunyavirus isolated from Culex (Culex) de-clarator mosquitoes in Trinidad. J Gen Virol 95:481– 485. http://dx.doi.org/10.1099/vir.0.058412-0.
18. Koboldt DC, Zhang Q, Larson DE, Shen D, McLellan MD, Lin L, MillerCA, Mardis ER, Ding L, Wilson RK. 2012. VarScan 2: somatic mutationand copy number alteration discovery in cancer by exome sequencing.Genome Res 22:568 –576. http://dx.doi.org/10.1101/gr.129684.111.
19. Drummond AJ, Suchard MA, Xie D, Rambaut A. 2012. Bayesian phy-logenetics with BEAUti and the BEAST 1.7. Mol Biol Evol 29:1969 –1973.http://dx.doi.org/10.1093/molbev/mss075.
20. Drummond AJ, Ho SY, Phillips MJ, Rambaut A. 2006. Relaxed phylo-genetics and dating with confidence. PLoS Biol 4:e88. http://dx.doi.org/10.1371/journal.pbio.0040088.
21. Nasar F, Palacios G, Gorchakov RV, Guzman H, Da Rosa APT, Savji N,Popov VL, Sherman MB, Lipkin WI, Tesh RB. 2012. Eilat virus, a uniquealphavirus with host range restricted to insects by RNA replication. ProcNatl Acad Sci U S A 109:14622–14627. http://dx.doi.org/10.1073/pnas.1204787109.
Auguste et al.
686 jvi.asm.org January 2015 Volume 89 Number 1Journal of Virology
on Decem
ber 16, 2014 by MO
OD
Y M
ED
ICA
L LIBR
AR
Yhttp://jvi.asm
.org/D
ownloaded from

22. Bammes BE, Chen DH, Jin L, Bilhorn RB. 2013. Visualizing and correctingdynamic specimen processes in TEM using a direct detection device. MicroscMicroanal 19:1320–1321.
23. Wang Z, Hyrc C, Bammes B, Afoine P, Jakana J, Chen DH, Liu X, BakerM, Kao C, Ludtke S, Schmid M, Adams P, Chiu W. 2014. An atomicmodel of brome mosaic virus using direct electron detection and real-space optimization. Nat Commun 5:4808. http://dx.doi.org/10.1038/ncomms5808.
24. Tang G, Peng L, Baldwin PR, Mann DS, Jiang W, Rees I, Ludtke SJ. 2007.EMAN2: an extensible image processing suite for electron microscopy. JStruct Biol 157:38–46. http://dx.doi.org/10.1016/j.jsb.2006.05.009.
25. Liu X, Jiang W, Jakana J, Chiu W. 2007. Averaging tens to hundreds oficosahedral particle images to resolve protein secondary structure ele-ments using a multi-path simulated annealing optimization algorithm. JStruct Biol 160:11–27. http://dx.doi.org/10.1016/j.jsb.2007.06.009.
26. Henderson R, Sali A, Baker ML, Carragher B, Devkota B, Downing KH,Egelman EH, Feng Z, Frank J, Grigorieff N, Jiang W, Ludtke SJ,Medalia O, Penczek PA Rosenthal PB, Rossmann MG, Schmid MF,Schroder GF, Steven AC, Stokes DL, Westbrook JD, Wriggers W, YangH, Young J, Berman HM, Chiu W, Kleywegt GJ, Lawson CL. 2012.Outcome of the first electron microscopy validation task force meeting.Structure 20:205–214. http://dx.doi.org/10.1016/j.str.2011.12.014.
27. Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM,Meng EC, Ferrin TE. 2004. UCSF Chimera—a visualization system forexploratory research and analysis. J Comput Chem 25:1605–1612. http://dx.doi.org/10.1002/jcc.20084.
28. Pintilie GD, Zhang J, Goddard TD, Chiu W, Gossard DC. 2010.Quantitative analysis of cryo-EM density map segmentation by watershedand scale-space filtering, and fitting of structures by alignment to regions.J Struct Biol 170:427– 438. http://dx.doi.org/10.1016/j.jsb.2010.03.007.
29. Baker ML, Abeysinghe SS, Schuh S, Coleman RA, Abrams A, MarshMP, Hryc CF, Ruths T, Chiu W, Ju T. 2011. Modeling protein structureat near atomic resolutions with Gorgon. J Struct Biol 174:360 –373. http://dx.doi.org/10.1016/j.jsb.2011.01.015.
30. Baker ML, Ju T, Chiu W. 2007. Identification of secondary structureelements in intermediate-resolution density maps. Structure 15:7–19.http://dx.doi.org/10.1016/j.str.2006.11.008.
31. Sali A, Blundell TL. 1993. Comparative protein modeling by satisfactionof spatial restraints. J Mol Biol 234:779 – 815. http://dx.doi.org/10.1006/jmbi.1993.1626.
32. Pintilie G, Chiu W. 2012. Comparison of Segger and other methods forsegmentation and rigid-body docking of molecular components incryo-EM density maps. Biopolymers 97:742–760. http://dx.doi.org/10.1002/bip.22074.
33. Rosenthal PB, Henderson R. 2003. Optimal determination of particleorientation, absolute hand, and contrast loss in single-particle electroncryomicroscopy. J Mol Biol 333:721–745. http://dx.doi.org/10.1016/j.jmb.2003.07.013.
34. Hill CL, Booth TF, Prasad BVV. 1999. The structure of a cypovirus andthe functional organization of dsRNA viruses. Nat Struct Biol 6:565–568.http://dx.doi.org/10.1038/9347.
35. Zhang H, Zhang J, Yu X, Lu X, Zhang Q, Jakana J, Zhang X, Zhou ZH,Chen DH. 1999. Visualization of protein-RNA interactions in cytoplas-mic polyhedrosis virus visualization of protein-RNA interactions in cyto-plasmic polyhedrosis virus. J Virol 73:1624 –1629.
36. Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W,Lipman DJ. 1997. Gapped BLAST and PSI-BLAST: a new generation of
protein database search programs. Nucleic Acids Res 25:3389 –3402. http://dx.doi.org/10.1093/nar/25.17.3389.
37. Bamford DH, Grimes JM, Stuart DI. 2005. What does structure tell usabout virus evolution? Curr Opin Struct Biol 15:655– 663. http://dx.doi.org/10.1016/j.sbi.2005.10.012.
38. Huiskonen JT, de Haas F, Bubeck D, Bamford DH, Fuller SD, ButcherSJ. 2006. Structure of the bacteriophage phi6 nucleocapsid suggests amechanism for sequential RNA packaging. Structure 14:1039 –1048. http://dx.doi.org/10.1016/j.str.2006.03.018.
39. Zhou ZH, Baker ML, Jiang W, Dougherty M, Jakana J, Dong G, Lu G,Chiu W. 2001. Electron cryomicroscopy and bioinformatics suggest pro-tein fold models for rice dwarf virus. Nat Struct Biol 8:868 – 873. http://dx.doi.org/10.1038/nsb1001-868.
40. Hagiwara K, Matsumoto T. 2000. Nucleotide sequences of genome seg-ments 6 and 7 of Bombyx mori cypovirus 1, encoding the viral structuralproteins V4 and V5, respectively. J Gen Virol 81:1143–1147.
41. Squire PG, Himmel ME. 1979. Hydrodynamics and protein hydration.Arch Biochem Biophys 196:165–177. http://dx.doi.org/10.1016/0003-9861(79)90563-0.
42. Upadhyaya NM, Zinkowsky E, Kositratana W, Waterhouse PM. 1996.The M(r) 43K major capsid protein of rice ragged stunt oryzavirus is apost-translationally processed product of a M(r) 67,348 polypeptide en-coded by genome segment 8. Arch Virol 141:1689 –1701. http://dx.doi.org/10.1007/BF01718292.
43. Marks EN. 1954. A review of the Aedes scutellaris subgroup with a study ofvariation in Aedes Pseudoscutellaris (Theobald): Diptera–Culicidae. BullBrit Mus (Nat Hist) London Ent 3:347– 414.
44. Brackney DE, Scott JC, Sagawa F, Woodward JE, Miller NA, SchilkeyFD, Mudge J, Wilusz J, Olson KE, Blair CD, Ebel GD. 2010. C6/36Aedes albopictus cells have a dysfunctional antiviral RNA interference re-sponse. PLoS Negl Trop Dis 4:e856. http://dx.doi.org/10.1371/journal.pntd.0000856.
45. Hagiwara K, Naitow H. 2003. Assembly into single-shelled virus-likeparticles by major capsid protein VP1 encoded by genome segment S1 ofBombyx mori cypovirus 1. J Gen Virol 84:2439 –2441. http://dx.doi.org/10.1099/vir.0.19216-0.
46. Garcia-Arriaza J, Manrubia SC, Toja M, Domingo E, Escarmis C. 2004.Evolutionary transition toward defective RNAs that are infectious by com-plementation. J Virol 78:11678 –11685. http://dx.doi.org/10.1128/JVI.78.21.11678-11685.2004.
47. Schirmer A, Link D, Cognat V, Moury B, Beuve M, Meunier A, BragardC, Gilmer D, Lemaire O. 2005. Phylogenetic analysis of isolates of beetnecrotic yellow vein virus collected worldwide. J Gen Virol 86:2897–2911.http://dx.doi.org/10.1099/vir.0.81167-0.
48. Bamford DH, Burnett RM, Stuart DI. 2002. Evolution of viral structure.Theor Popul Biol 61:461– 470. http://dx.doi.org/10.1006/tpbi.2002.1591.
49. Butcher SJ, Dokland T, Ojala PM, Bamford DH, Fuller SD. 1997. Inter-mediates in the assembly pathway of the double-stranded RNA virus phi6.EMBO J 16:4477–4487. http://dx.doi.org/10.1093/emboj/16.14.4477.
50. Coulibaly F, Chevalier C, Gutsche I, Pous J, Navaza J, Bressanelli S,Delmas B, Rey FA. 2005. The birnavirus crystal structure reveals struc-tural relationships among icosahedral viruses. Cell 120:761–772. http://dx.doi.org/10.1016/j.cell.2005.01.009.
51. Coulibaly F, Chiu E, Ikeda K, Gutmann S, Haebel PW, Schulze-BrieseC, Mori H, Metcalf P. 2007. The molecular organization of cypoviruspolyhedra. Nature 446:97–101. http://dx.doi.org/10.1038/nature05628.
Genetic and Structural Features of Dinovernaviruses
January 2015 Volume 89 Number 1 jvi.asm.org 687Journal of Virology
on Decem
ber 16, 2014 by MO
OD
Y M
ED
ICA
L LIBR
AR
Yhttp://jvi.asm
.org/D
ownloaded from
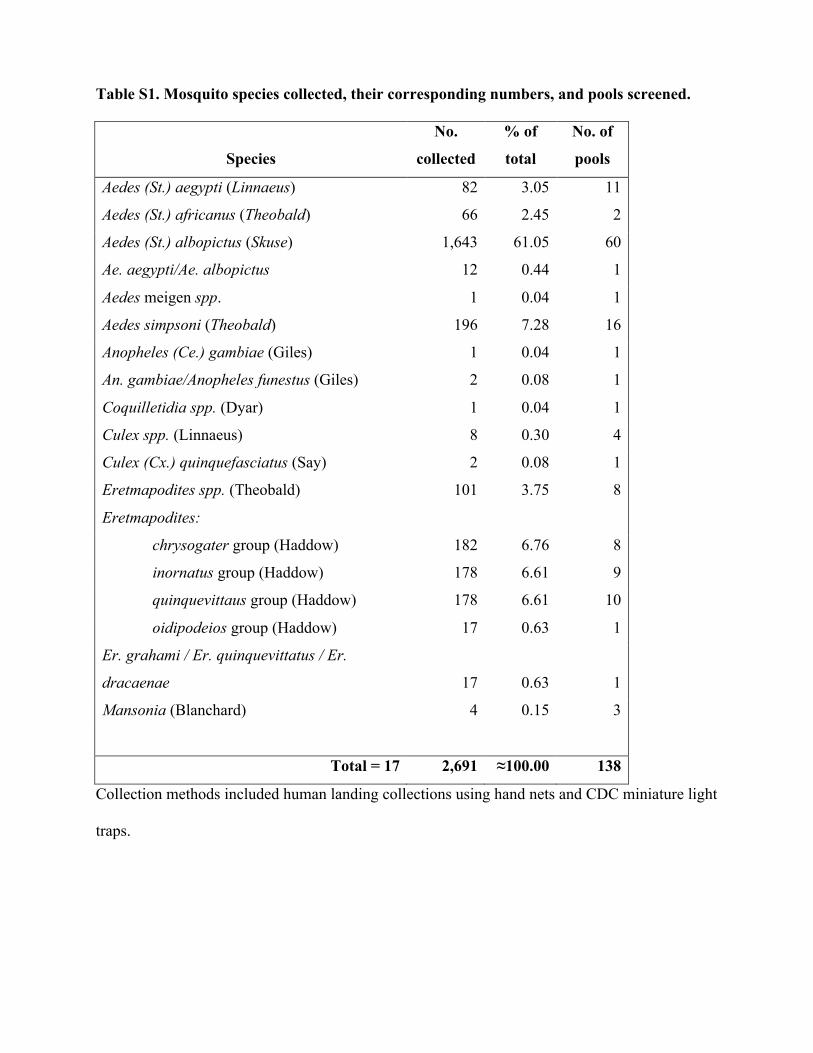
Table S1. Mosquito species collected, their corresponding numbers, and pools screened.
Species
No.
collected
% of
total
No. of
pools
Aedes (St.) aegypti (Linnaeus) 82 3.05 11
Aedes (St.) africanus (Theobald) 66 2.45 2
Aedes (St.) albopictus (Skuse) 1,643 61.05 60
Ae. aegypti/Ae. albopictus 12 0.44 1
Aedes meigen spp. 1 0.04 1
Aedes simpsoni (Theobald) 196 7.28 16
Anopheles (Ce.) gambiae (Giles) 1 0.04 1
An. gambiae/Anopheles funestus (Giles) 2 0.08 1
Coquilletidia spp. (Dyar) 1 0.04 1
Culex spp. (Linnaeus) 8 0.30 4
Culex (Cx.) quinquefasciatus (Say) 2 0.08 1
Eretmapodites spp. (Theobald) 101 3.75 8
182 6.76 8
178 6.61 9
178 6.61 10
Eretmapodites:
chrysogater group (Haddow)
inornatus group (Haddow)
quinquevittaus group (Haddow)
oidipodeios group (Haddow) 17 0.63 1
Er. grahami / Er. quinquevittatus / Er.
dracaenae 17 0.63 1
Mansonia (Blanchard) 4 0.15 3
Total = 17 2,691 ≈100.00 138
Collection methods included human landing collections using hand nets and CDC miniature light
traps.
5

Table S2. Percentage amino acid sequence identities for the RdRp within the subfamily Spinareovirinae.
Virus name*
FAKV ARV AtSRV-TS GCRV EYAV CTFV RARV CpRV NLRV MRCV RBSDV APRV ObRV BmCPV HaCPV RRSV RpLV
FAKV 100 6.9 7.2 7.4 10.9 11.3 11.6 12.3 11.7 12.7 12.5 89.7 22.4 21.5 25.2 18.6 20 ARV 100 40.8 41.4 8.1 8 8.1 7.5 6.3 7.7 6.9 7 7.3 8.1 6.6 8.5 6.7 AtSRV-TS
100 59 7.3 7.5 8.6 7.4 7.1 8.5 8 7.1 7.5 7.8 7.2 7.8 7
GCRV 100 7.9 8 7.4 6.5 6.6 7.7 8.2 7.2 7.5 8.6 6.5 8 6.7 EYAV 100 85.8 26.4 25.1 11.1 10.8 10.4 11 11.1 9.8 11.4 11.5 10.6 CTFV 100 26 24.8 11.2 10.7 10.5 11.5 11.5 9.9 11.6 11.8 10.8 RARV 100 37.5 10.3 10.5 10.5 11.2 11.5 10.3 11.4 11 12.3 CpRV 100 11.6 11.1 10.7 12.6 11.9 11.3 12.7 12.2 13.2 NLRV 100 25.9 26.2 11.5 11.1 10.8 10.9 11.5 11.6 MRCV 100 77.9 12.7 13.5 12.1 13.3 13.7 13.6 RBSDV 100 12.7 13.2 11.7 13 13.9 13.3 APRV 100 23.3 21.7 24.9 18 20 ObRV 100 23.5 37.3 20.1 20.2 BmCPV 100 28.5 21.3 20.7 HaCPV 100 22.4 24.3 RRSV 100 35.4 RpLV 100
*Virus abbreviations are described in Table S3.

Table S3. Sequences used in phylogenetic analyses and their associated details 10
Species Isolate
(strain) name
Location of
isolation
Abbreviation Accession
number*
Genus Dinovernavirus
Aedes pseudoscutellaris reovirus
Fako virus
APRV
CSW77
France
Cameroon
ApRV
FAKV
YP_443936
KM978416
Genus Oryzavirus
Raspberry latent virus
Rice ragged stunt virus
RpLV9A
Thai
USA, Canada
Thailand
RpLV
RRSV
ADO27688
NP_620541
Genus Cypovirus
Heliothis armigera cypovirus 5
Bombyx mori cypovirus 1
Operophtera brumata cypovirus
China
China
ObRV-19
China
China
Orkney Isles
HaCPV
BmCPV
ObRV
YP_001883321
AAR88092
ABB17221
Genus Fijivirus
Rice black streaked dwarf virus
Mal del rio cuarto virus
Nilaparvata lugens reovirus
RBSDV
MRCV
Izumo
China
Argentina
Japan
RBSDV
MRCV
NLRV
NP_620452
AAO73182
BAA08542
Genus Mycoreovirus
Cryphonectria parasitica mycoreovirus
Rosellinia anti-rot virus
9B21
W370
USA
Japan
CpRV
RARV
AAP45577
BAC98431
Genus Coltivirus
Colorado tick fever virus
Eyach virus
Florio; N-7180
FR578
USA
Europe
CTFV
EYAV
NP_690891
AAM18342
Genus Aquareovirus
Grass carp reovirus
Atlantic salmon reovirus
NA
AtSRV-TS
China
Tasmania
GCRV
AtSRV-TS
AAG10436
ABO32573
Genus Orthoreovirus
Avian orthoreovirus
S1133
USA
ARV
ABF82230
Genus Phytoreovirus
Rice dwarf virus
Homalodisca vitripennis reovirus
Chinese strain
Fillmore
China
USA
RDV
HvRV
NP_620544
YP_002790884
Genus Rotavirus
Human rotavirus A
Simian rotavirus A
GER1H-09
SA11
Germany
South Africa
HuRV
SiRV
ACV73799
P22678
Genus Orbivirus
Blue tongue virus
BTV12
Taiwan
BTV-12
ADD62087

Blue tongue virus
African horse sickness virus
St. Croix river virus
BTV13
AHSV-9
SCRV
USA
Africa
USA
BTV-13
AHSV
SCRV
AAA87363
AAC40586
AAG34363
Genus Mimoreovirus
Micromonas pusilla reovirus
MpRV
Netherlands
MpRV
YP_654545
Genus Seadornavirus
Banna virus
Kadipiro virus
JKT-6423
JKT-7075
Indonesia
Indonesia
BAV
KDV
NP_694469
NP_694468
Genus Cardoreovirus
Eriocheir sinensis reovirus
905
China
EsRV905
AAT11887
*RdRp amino acid sequences used.

Table S4. SNP diversity within the 9 segments of the CSW77 genome at a minimum
frequency of 3%.
Segment Position Reference Variant Cons Coverage Frequency Segment 1 71 G T K 117062 12.06% Segment 1 83 C A M 96675 8.29% Segment 1 159 A C M 87830 12.94% Segment 1 196 C A M 117575 11.50% Segment 1 280 C A M 80190 10.18% Segment 1 458 G A R 70172 10.85% Segment 1 552 G A R 42296 11.16% Segment 1 600 G A R 68527 10.17% Segment 1 640 A C M 60210 10.27% Segment 1 675 T C Y 58575 9.87% Segment 1 731 C T Y 46843 6.53% Segment 1 753 G A R 60760 9.97% Segment 1 857 A G R 80118 9.47% Segment 1 988 A G R 45003 3.11% Segment 1 1020 T C Y 51983 11.35% Segment 1 1854 G A R 21960 9.59% Segment 1 2484 T C Y 5033 12.12% Segment 1 2876 G A R 9616 11.78% Segment 1 2928 A G R 14970 7.37% Segment 1 3161 T C Y 6246 11.66% Segment 1 3285 G A R 5929 12.38% Segment 1 3317 G A R 7024 11.66% Segment 1 3513 A G R 10658 9.14% Segment 1 3560 C T Y 10119 6.89% Segment 2 903 T C Y 21671 4.93% Segment 2 2661 A G R 10189 4.83% Segment 3 541 T A W 36480 9.01% Segment 3 828 A G R 47573 6.56% Segment 3 1398 T C Y 75974 4.79% Segment 3 2100 T C Y 15943 41.34% Segment 3 2803 A G R 16716 13.90% Segment 4 646 A C M 45626 12.72% Segment 4 690 T G K 54915 4.97% Segment 4 1408 T C Y 74435 13.96% Segment 4 2620 G A R 33055 9.87% Segment 5 1384 G A R 27447 5.10% Segment 6 1763 G A R 777 4.25% Segment 6 1766 T G K 371 8.89% Segment 6 1768 T G K 254 12.99% Segment 8 11 C A M 1723 4.53% Segment 8 323 C A M 69787 36.46% Segment 8 728 C T Y 29468 22.67% Segment 8 971 A G R 14294 40.30% Segment 8 1138 G A A 24 95.83% Segment 9 383 A G R 43051 5.38% Segment 9 983 C T Y 26446 18.49%

Figure S1. Transmission electron microscopy (TEM) photographs of ultrathin sections of
infected C6/36 cells showing the ultrastructure of CSW77 virus. Virus particles are
approximately 450 Å in diameter and cluster in the host cell cytoplasm (A, B; indicated by red
arrows) and inside cytoplasmic vacuoles (A; indicated by blue arrows). Scale bar = 100 nm.
Ultrastructure of CSW77 virus as observed under negative staining with uranyl acetate (C).
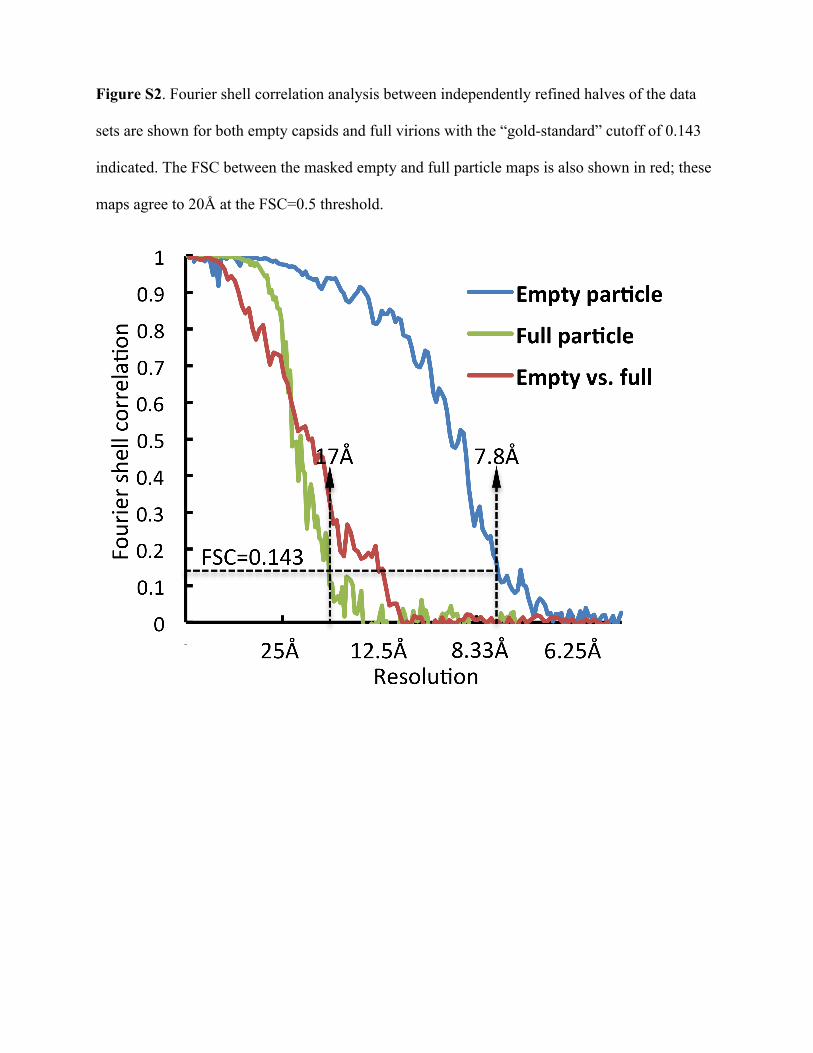
Figure S2. Fourier shell correlation analysis between independently refined halves of the data
sets are shown for both empty capsids and full virions with the “gold-standard” cutoff of 0.143
indicated. The FSC between the masked empty and full particle maps is also shown in red; these
maps agree to 20Å at the FSC=0.5 threshold.

Figure S3. Comparative morphology of FAKV, CPV, RRSV, and GCRV. The 7.8Å structure of
empty capsid (A,E), 17Å structure of FAKV full virion (C), the 3.1Å structure of CPV (B)
(EMD-5256) (1), and the 3.3Å structure of GCRV (F) (EMD-5160) (2) were filtered to 20Å to
compare to the 21Å structure of RRSV (D) (3). A 5-fold and a 3-fold symmetry axis are labeled
on the FAKV capsid (figure A, a pentagon and triangle, respectively). All four viruses contain a
clamp near the turret (labeled in B), but FAKV lacks any protrusion near the 3-fold symmetry
axis. Only CPV contains SP domains, which are seen as small knobs on either side of the 5-fold-
proximal turret. Five quasi-equivalent positions exist in a T=13l lattice, named P, Q, R, S, and T.
The trimeric outer-shell proteins of RRSV (D) and GCRV (F) are labeled with this
nomenclature, and GCRV's empty P position is also indicated. None of these positions are
occupied in FAKV nor in CPV. A cutaway of the FAKV capsid (E) shows a density under the
penton that likely corresponds to the location of the RdRp.


Figure S4. Semi-automatic segmentation of the Fako virus capsid. An asymmetric unit of FAKV
is segmented into four regions roughly corresponding to the turret (purple), clamp (red), and
major capsid protein dimer (cyan and yellow). The boundaries were similar to those obtained
when segmenting with Gorgon and Chimera (Fig. 6B).

References
1. Yu X, Ge P, Jiang J, Atanasov I, Zhou ZH. 2011. Atomic model of CPV reveals the
mechanism used by this single-shelled virus to economically carry out functions
conserved in multishelled reoviruses. Structure 19:652-661.
2. Zhang X, Jin L, Fang Q, Hui WH, Zhou ZH. 2010. 3.3 Å cryo-EM structure of a
nonenveloped virus reveals a priming mechanism for cell entry. Cell 141:472-482.
3. Miyazaki N, Uehara-Ichiki T, Xing L, Bergman L, Higashiura A, Nakagawa A,
Omura T, Cheng RH. 2008. Structural evolution of Reoviridae revealed by Oryzavirus
in acquiring the second capsid shell. J. Virol. 82:11344-11353.
Related Documents











