A New In Vifro Culture Technique for Rat Embryos' MAURICE A. ROBKIN, THOMAS H. SHEPARD AND TAKASHI TANIMURA Department of Nuclear Engineering and Central Laboratory for Human Embryology, Department of Pediatrics, Uniuerszty of Washington, Seattle, Washington 98195 ABSTRACT A new technique for culturing embryos, which permits detailed observation and manipulation, is described. The method has been applied to rat embryos on day 10, 11, or 12 of gestation, Twenty-four-hour culture of day-11 rat embryos resulted in nearly normal increases in somite number, and embryo length and protein increment of about half that occurring in vivo. After 24 h of culture the embryos generally were translucent and showed no apparent areas of gross tissue necrosis. Sections from representative embryos are presented. The apparatus is extremely compact and requires only about 9 ml of medium which is sufficient to support three embryos. The small volume allows the mea- surement of metabolites consumed or liberated with reasonable accuracy. The method permits a detailed and continuous record to be made of the heart rate and the addition of drugs to and sampling of the culture medium. The apparatus, named the PLASMOM, was used to measure the effect of changes in the ambient temperature on the heart rate of day-11 rat embryos. The results showed a linear dependence of heart rate on temperature between 30 and 40" C. Teratologists and developmental biolo- gists have long recognized the advantages of being able directly to observe the devel- oping embryo. Nature permits this for oviparous species in a relatively straight- forward way. In the case of viviparous species nature is not so cooperative. From the early days of this century workers have attacked the problem of culturing growing mammalian embryos. Their conclusions (see e.g., Waddington and Waterman, '33; Nicholas and Rudnick, '34, '38; Nicholas, '38) tend to be fairly consistent. Although it is possible to grow presomite and early-somite embryos with tissue culture techniques in static me- dium, embryos at stages beyond about 30 somites require a circulating medium to survive for more than a very short period of time. Nicholas' ('38) techniques were not widely adopted and further advances in the culture of mammalian embryos were not made for 30 years. The work of New (New, '67; New and Daniel, '69) showed that, in fact, using a circulating culture medium would enable rat embryos to grow and develop in vitro past the 30-somite stage. This was true for embryos up to about 50 somites as measured by increases in somite number, crown-rump length, and protein content. This work put the in vitro culture of rat embryos on a solid foundation and pro- vided a dependable preparation for ex- periments in development, both normal and abnormal. New's ('67) apparatus, however, re- quires that the perfusing gas drive the circulation which results in fluid pulsa- tions that cause the embryos to be agitated on their attachments to their rafts. The gas bubbles cause frothing when human serum is used as the culture medium which in some cases requires the use of an antifoamant as an adulterant. In ad- dition the circulator must be maintained in a thermostatically controlled enclosure which inhibits access, observation, and manipulation. Tamarim and Jones ('69) have proposed a culture device, but we have had no experience with it. A desirable culture system should have the following attributes: (1) visual acces- sibility, (2) physical accessibility for in- strumentation and obtaining specimens of medium, (3) small internal volume, and 1 Supported by the NIH grants HD02392, HD00180, and HD00836, and by the Graduate School Research Fund, University of Washington. 2 Present Address: Department of Anatomy, Kyoto University, Kyoto, Japan. TERATOLOGY, 5: 367-376. 367

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A New In Vifro Culture Technique for Rat Embryos' MAURICE A. ROBKIN, THOMAS H. SHEPARD AND TAKASHI TANIMURA Department of Nuclear Engineering and Central Laboratory for Human Embryology, Department of Pediatrics, Uniuerszty of Washington, Seattle, Washington 98195
ABSTRACT A new technique for culturing embryos, which permits detailed observation and manipulation, is described. The method has been applied to rat embryos on day 10, 11, or 12 of gestation, Twenty-four-hour culture of day-11 rat embryos resulted in nearly normal increases in somite number, and embryo length and protein increment of about half that occurring in vivo. After 24 h of culture the embryos generally were translucent and showed no apparent areas of gross tissue necrosis. Sections from representative embryos are presented. The apparatus is extremely compact and requires only about 9 ml of medium which is sufficient to support three embryos. The small volume allows the mea- surement of metabolites consumed or liberated with reasonable accuracy. The method permits a detailed and continuous record to be made of the heart rate and the addition of drugs to and sampling of the culture medium. The apparatus, named the PLASMOM, was used to measure the effect of changes in the ambient temperature on the heart rate of day-11 rat embryos. The results showed a linear dependence of heart rate on temperature between 30 and 40" C.
Teratologists and developmental biolo- gists have long recognized the advantages of being able directly to observe the devel- oping embryo. Nature permits this for oviparous species in a relatively straight- forward way. In the case of viviparous species nature is not so cooperative.
From the early days of this century workers have attacked the problem of culturing growing mammalian embryos. Their conclusions (see e.g., Waddington and Waterman, '33; Nicholas and Rudnick, '34, '38; Nicholas, '38) tend to be fairly consistent. Although it is possible to grow presomite and early-somite embryos with tissue culture techniques in static me- dium, embryos at stages beyond about 30 somites require a circulating medium to survive for more than a very short period of time. Nicholas' ('38) techniques were not widely adopted and further advances in the culture of mammalian embryos were not made for 30 years.
The work of New (New, '67; New and Daniel, '69) showed that, in fact, using a circulating culture medium would enable rat embryos to grow and develop in vitro past the 30-somite stage. This was true for embryos up to about 50 somites as measured by increases in somite number,
crown-rump length, and protein content. This work put the in vitro culture of rat embryos on a solid foundation and pro- vided a dependable preparation for ex- periments in development, both normal and abnormal.
New's ('67) apparatus, however, re- quires that the perfusing gas drive the circulation which results in fluid pulsa- tions that cause the embryos to be agitated on their attachments to their rafts. The gas bubbles cause frothing when human serum is used as the culture medium which in some cases requires the use of an antifoamant as an adulterant. In ad- dition the circulator must be maintained in a thermostatically controlled enclosure which inhibits access, observation, and manipulation. Tamarim and Jones ('69) have proposed a culture device, but we have had no experience with it.
A desirable culture system should have the following attributes: (1) visual acces- sibility, (2) physical accessibility for in- strumentation and obtaining specimens of medium, (3) small internal volume, and
1 Supported by the NIH grants HD02392, HD00180, and HD00836, and by the Graduate School Research Fund, University of Washington.
2 Present Address: Department of Anatomy, Kyoto University, Kyoto, Japan.
TERATOLOGY, 5: 367-376. 367

368 M. A. ROBKIN, T. H. SHEPARD AND T. TANIMURA
Loser -
(4) simple construction and repair. Such a device has been constructed.
Twenty-four-hour culture experiments with day-11 rat embryos (24-26 somites) have given good growth results as deter- mined by the increases in somite number, crown-rump length, and protein content. The device has been named the PLASMOM and is described in detail below. In addi- tion an electronic method of measuring and recording the heart rates of these embryos is described and illustrative data from its use are given.
APPARATUS, MATERIALS, AND PROCEDURES
Photo
Tube Embryo Multiplying
A schematic of the PLASMOM system is shown in figure 1. Taking each part in order of circulation, the various compo- nents are described as follows:
Pump. The circulating medium is driven by a small-displacement roller pump with a torque control monitor which permits the turning rate to be accurately set (Cole-Parmer Ultra-Masterflex with
A
w
Laser Beam
Strip Chart Recorder
Laser Beam modulated by heart beat
Filter L,1 Water Bath
Fig. 1 Diagram of the components of the PLASMOM embryo culture device.
Model 7014 pumphead). Depending on the tubing used the pumping rate tends to change with time as the tubing fatigues. Silicone rubber tubing has the best fa- tigue vs. time characteristics, but is suf- ficiently permeable to oxygen to make i t impossible completely to saturate the nu- trient medium. Although Tygon tubing has an inferior fatigue rate it was pref- erable because it is relatively imperme- able to oxygen. Using this tubing the medium may be kept near saturation for the duration of the experiments so far at- tempted. Using a larger pumphead with larger diameter pump tubing permits higher flow rates with lower RPM and so prolongs tubing life. (This improvement must be balanced against an increase in the agitation of the embryos with the in- creased surging of the medium due to the increased volume displaced per pulse.)
Filter. The filter is a simple glass- wool-filled tube which serves to trap par- ticulates circulating with the medium. The glass wool may be autoclaved, and the system is assembled with the filter in the circuit.
The bubble trap and thermistor port is a cru- ciform glass construct which serves to provide a preferred diversion for any in- cidental bubbles flowing with the medium. The thermistor port is covered with a serum stopper, and needle-mounted therm- istors are inserted to monitor the temper- ature on both sides of the embryo cham- ber. One of the thermistors may be used to control the heating circuit, if desired.
Oxygen electrode port. The oxygen saturation level is monitored by a polaro- graphic-type Clark oxygen electrode (Yel- low Springs Instrument Co., Yellow Springs, Ohio) held in a glass bubble with small inlet and outlet lumens which give a rapid Aow of medium across the face o f the electrode membrane. The oxygen elec- trode is usually placed just upstream from the embryo chamber and monitors the relative oxygen saturation of the medium as it is delivered to the embryos. The oxy- gen level is calibrated at the start of each experiment at the appropriate tempera- ture by adjusting the readout value to 21% before the oxygen flow is started. Presumably the medium is in equilibrium with the atmosphere until the gas flow is
Bubble trap and thermistor port.

I N VlTRO CULTURE OF RAT EMBRYOS 369
started. The flask is usually left open to the atmosphere so no hyperbaria develops, but by strengthening the glassware and closing the openings to the atmosphere the system could be operated hyperbari- cally with relatively little modification.
Embryo chamber . The embryo cham- ber is a high-quality glass tube of square cross section made from a cuvette for a spectrophotometer with round glass tub- ing fused on the ends to accommodate the silicone rubber connectors. The opti- cal properties are excellent as the oppo- site sides are parallel and carefully pol- ished. The internal dimension of the chamber used in our apparatus is about 7 mm. (Using a two-cell flashlight, we have obtained color motion pictures of the embryos at high enough magnifica- tion to see the flow ofindividual red blood cells in the yolk sac circulation.)
The lung of the PLASMOM is composed of a bundle of silicone rubber tubes immersed in a water-filled flask. The perfusing gas of the desired composi- tion is bubbled into the water and passes by diffusion through the tubes into the culture medium which is flowing inside the tubes. The silicone rubber is extremely permeable to oxygen, carbon dioxide, and water vapor, with transfer rates propor- tional to the difference in partial pressure across the tube walls. The diffusive trans- fer eliminates the bubbles in the medium. Since there is no direct contact there is no frothing of the serum and no danger of serum protein denaturation. Thus, there is no need to add an antifoaming agent to the medium. For a total volume of 10 ml flowing at about 10 ml/min the 02 sat- uration level increases from 21 to 80% in about 5 min when 95% 02 is used as the perfusing gas and the lung has a total of 50 linear feet of 0.020 X 0.037-inch tub- ing made up as a bundle of 25 %foot lengths.
W a t e r bath . The flask in which the tube bundle is immersed is supported in a heating mantle controlled by either one of the circuit thermistors or a third therm- istor in the water bath (if desired). The heated water warms the circulating me- dium at the same time that gas exchange occurs. Once the desired temperature is reached long-term thermostasis is excel-
Lung .
lent and there are practically no short- term fluctuations. The water bath does not interfere in any way with observability of the embryos since they are held well above the flask.
Assembly . The various parts of the circuit are connected by rubber tubing, and assembly and disassembly are very simple. The assembled system and the components described above are shown in figure 2.
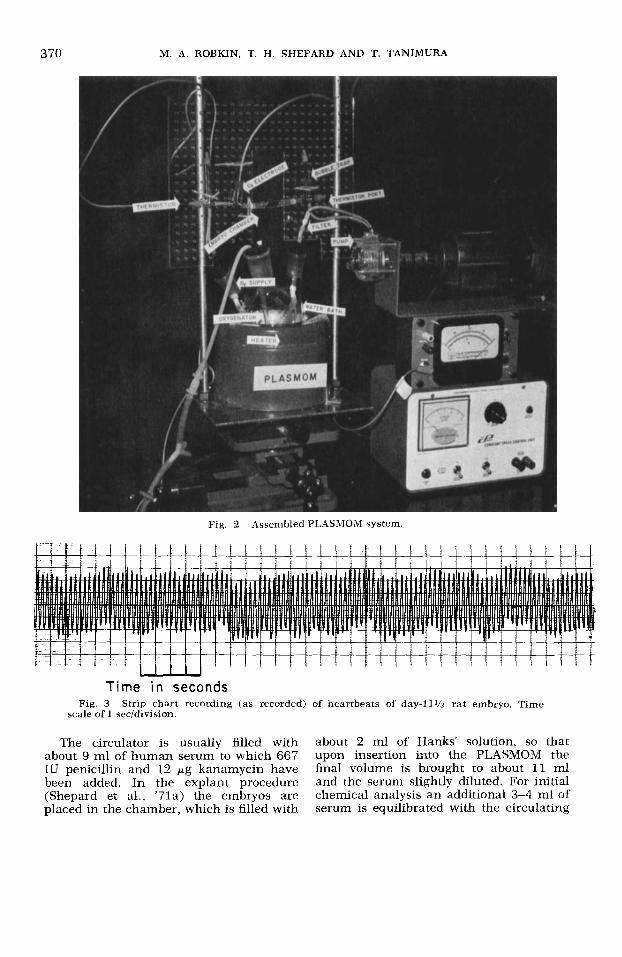
Heart-rate determinat ion. Automatic measurement of the embryonic heart rate is made by a method of transillumination using a laser and photomultiplying tube. The beam from a red (He-Ne) low-power laser (Coleman Instruments, Maywood, Ill.) is passed through the embryo. A lens focuses an image of the beating heart into the face of a photomultiplying tube, and the flicker produced by the beating heart is detected and recorded either by an os- cilloscope or strip chart recorder. The signal can also be passed into the input of a cardiotachometer, and the heart rate obtained directly. The rate also may be obtained from the oscilloscope or strip chart recorder, so that a simultaneous record of both the heartbeat pattern and rate is obtained. Embryos have been ex- posed to the laser for up to an hour with no apparent ill effect. Figure 3 shows a typical strip chart recording of individual heart beats of a day-1 1 rat embryo.
A PLASMOM circuit with a single embryo chamber (as many chambers as desired may be used simultaneously) with room for three em- bryos has an internal volume of about 9 ml. This is a small enough volume so that the amounts of metabolites consumed or liberated may be measured with rea- sonable accuracy. Since the entire circuit of the PLASMOM is accessible small ali- quots of medium may be withdrawn from time to time for analysis.
Medium. Based on his experiments New ('67) concluded that the best results were obtained by culturing rat embryos in homologous serum. Tanimura and Shep- ard ('70) found no statistically significant differences between cultures using homol- ogous serum and human serum. The best results with human serum, however, were obtained with freshly drawn and prepared blood.
Sampling of the m e d i u m .
370 M. A. ROBKIN, T. H. SHEPARD AND T. TANIMURA
Fig. 2 Assembled PLASMOM system
Time i n seconds Fig. 3 Strip chart recording (as recorded) of heartbeats of day-11% rat embryo. Time
scale of 1 secldivision.
The circulator is usually filled with about 2 ml of Hanks' solution, so that about 9 ml of human serum to which 667 upon insertion into the PLASMOM the IU penicillin and 12 Fg kanamycin have final volume is brought to about 11 ml been added, In the explant procedure and the serum slightly diluted. For initial (Shepard et al., '71a) the embryos are chemical analysis an additional 3 - 4 ml of placed in the chamber, which is filled with serum is equilibrated with the circulating

IN VITRO CULTURE OF RAT EMBRYOS 371
medium using two hypodermic syringes as injection and withdrawal reservoirs. The extra serum is used for analysis and the net protein dilution owing to the Hanks' medium is 85% (2 ml out of 14- 15 ml total).
Pre- and postexperimental maintenance. After assembly with new rubber tubing and glass components which have been soap and water washed the PLASMOM is wrapped in paper and sealed with auto- clave tape. After being steam sterilized, the assembly is left wrapped until used in the next experiment. New pumphead tubing is sterilized by circulating 70 % ethanol followed by distilled water rinses prior to connection into the PLASMOM circuit .
After each experiment all tubing is dis- carded and the glassware immediately rinsed with running tap water and then washed with soap and water. The tube bundle lung is copiously flushed with dis- tilled water to wash out as much medium as possible. After flushing the lung is boiled to denature any serum proteins ad- hering to the tubing and soaked overnight in a 2 mglml solution of pronase in 0.2 M Tris buffer at pH 8. The buffer solution is flushed out and the PLASMOM reas- sembled.
Explantation and assessment of the embryos. The methods of explantation and assessment of the embryos were de- scribed in Shepard ('71b).
RESULTS
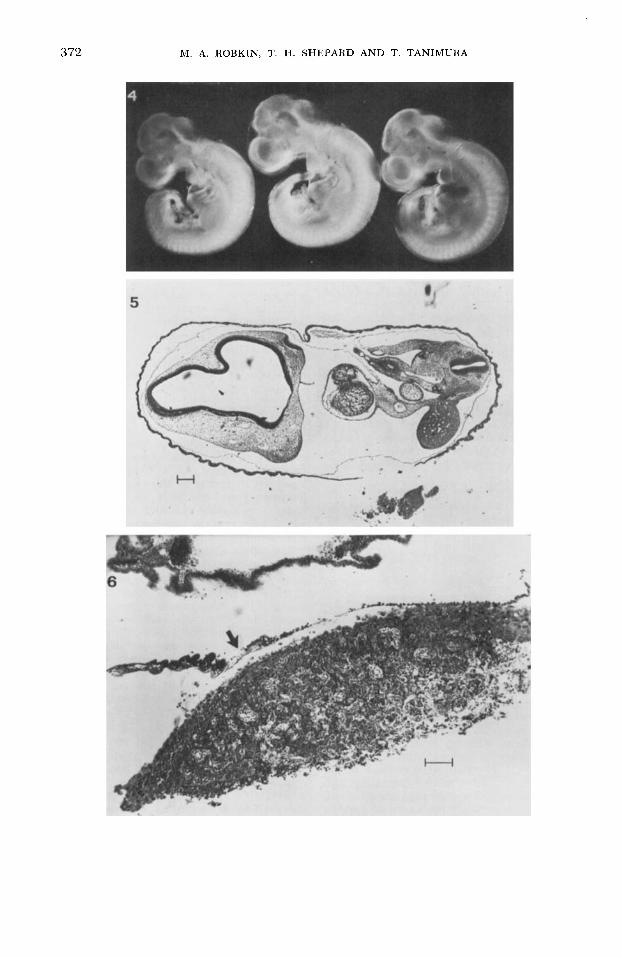
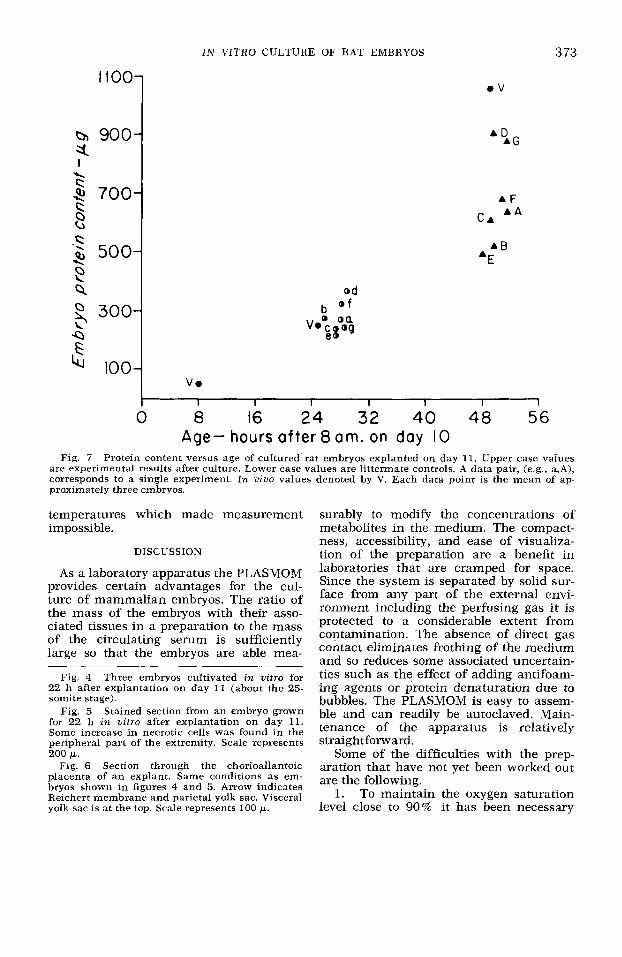
Embryonic growth. The PLASMOM circulator has been used in a series of 18- to 24-h growth experiments in which Sprague-Dawley rat embryos were ex- planted on day 11 of gestation. Figure 4 shows three embryos cultured for 20 h from explantation at day 11. These em- bryos were translucent and showed no apparent areas of gross tissue necrosis. Mitotic figures were present in histologic sections of the embryos. There were some necrotic areas in the center of the allan- toic placenta. Figures 5 and 6 show sec- tions of a day-11 embryo cultured €or 22 h. Figure 7 shows the protein content of the experimental embryos and littermate con- trols from each of seven experiments. Each data point represents the mean of approx-
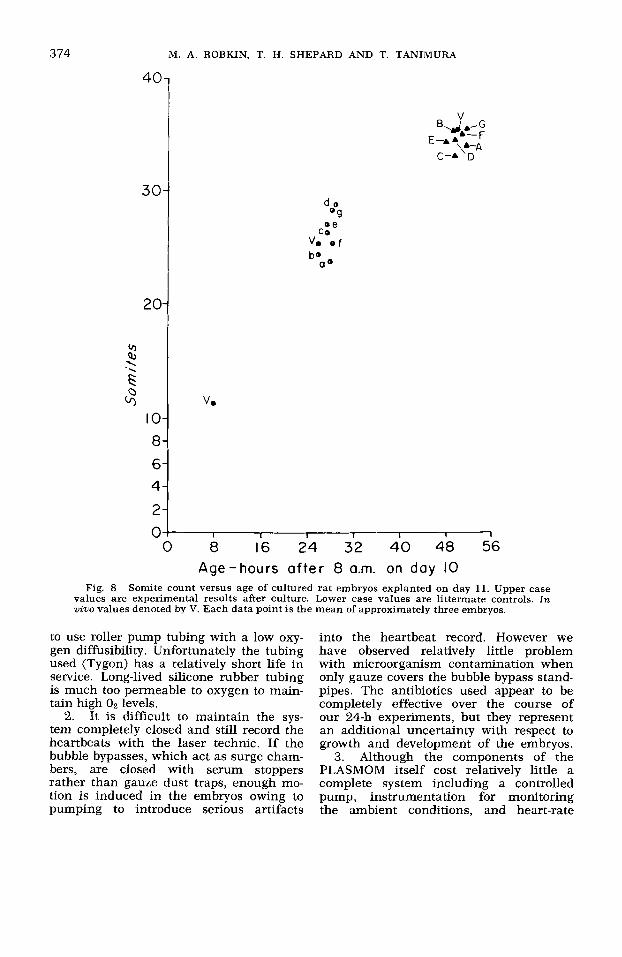
imately three embryos. Figure 8 shows the somite counts from the same experiments.
Heart rate and ambient temperature. To illustrate the use of the PLASMOM system further, the relation between the embryonic heart rate and the ambient temperature was determined. A length of rubber tubing was submerged in the water bath and connected to a separate pump drawing from an ice-water bath. This cool- ing coil provided for rapid cooling of the water bath and hence the circulating serum. The embryos were explanted into the PLASMOM.
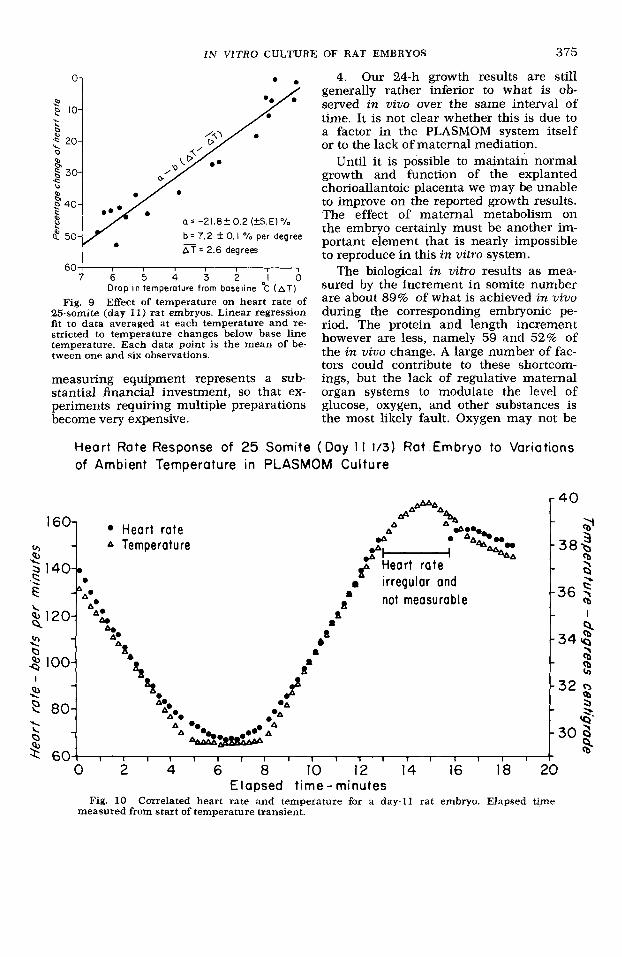
The temperature of the medium at the inlet and outlet of the embryo chamber was monitored by needle-mounted therm- istors and the heart rate monitored by the laser-transillumination technique de- scribed above. The results of the experi- ment are presented in two graphs (figs. 9, 10). In the first, pooled data for the heart rate of several embryos are plotted against ambient temperature between 30 and 40" C.
The experiments were done by first stabilizing the heart rate at a nominal base line temperature of 37-39' C. How- ever, since the initial base line values varied somewhat each experiment was analyzed in terms of the change in tem- perature from the base line. Since the heart rates tended to become erratic above 39" C only experiments done below 39" C were analyzed.
A null hypothesis of a linear response was made and the data were regression- fitted to a straight lineof the formRate = of a - b (AT - AT), where AT is the mean of the temperature changes over the pooled data. The results showed a temper- ature coefficient of 7.2 & 0.1 % (value * SE) change in heart rate per degree cen- tigrade change in temperature. Student's t value for the fit is t = 77 with 16 de- grees of freedom which yields a probabil- ity of rejection of the null hypothesis of linearity of P < 0.000001 (Abromowitz and Stegun, '64).
In the second graph (fig. 10) the tem- perature and heart rate are plotted against time for a single embryo to illustrate the close correlation. The gaps in the figure near 40" C are due to the irregularity observed in the heartbeat at these higher
372 M. A. ROBKIN, T. H. SHEPARD AND T. TANIMURA
IN VITRO CULTURE OF RAT EMBRYOS 3 73
IIOO.
b 900. a. I \
\ 700. s Ll
300- \ -D G l' 100-
.V
A F A A
C A
od
b O f
V.
0 8 16 24 32 40 48 56 Age- hours after 8 am. on day 10
Fig. 7 Protein content versus age of cultured rat embryos explanted on day 11. Upper case values are experimental results after culture. Lower case values are littermate controls. A data pair, (e.g.. a,A), corresponds to a single experiment. In vivo values denoted by V. Each data point is the mean of ap- proximately three embryos.
temperatures which made measurement impossible.
DISCUSSION
As a laboratory apparatus the PLASMOM provides certain advantages for the cul- ture of mammalian embryos. The ratio of the mass of the embryos with their asso- ciated tissues in a preparation to the mass of the circulating serum is sufficiently large so that the embryos are able mea-
Fig. 4 Three embryos cultivated in vitro for 22 h after explantation on day 11 (about the 25- somite stage).
Fig. 5 Stained section from an embryo grown for 22 h in nitro after explantatibn on day 11. Some increase in necrotic cells was found in the peripheral part of the extremity. Scale represents
Fig. 6 Section through the chorioallantoic placenta of an explant. Same conditions as em- bryos shown in figures 4 and 5. Arrow indicates Reichert membrane and parietal yolk sac. Visceral yolk sac is at the top. Scale represents 100 p.
200 p.
surably to modify the concentrations of metabolites in the medium. The compact- ness, accessibility, and ease of visualiza- tion of the preparation are a benefit in laboratories that are cramped for space. Since the system is separated by solid sur- face from any part of the external envi- ronment including the perfusing gas it is protected to a considerable extent from contamination. The absence of direct gas contact eliminates frothing of the medium and so reduces some associated uncertain- ties such as the effect of adding antifoam- ing agents or protein denaturation due to bubbles. The PLASMOM is easy to assem- ble and can readily be autoclaved. Main- tenance of the apparatus is relatively straightforward.
Some of the difficulties with the prep- aration that have not yet been worked out are the following.
To maintain the oxygen saturation level close to 90% it has been necessary
1.
3 74
40
3 0
20
cr, Q, '4 .s G
10 8 6 4
2 0
M. A. ROBKIN, T. H. SHEPARD AND T. TANIMURA
be a o
v.
I I I I I I I
0 8 16 24 32 40 48 56 Age-hours a f te r 8 a.m. on day 10
Fig. 8 Somite count versus age of cultured rat embryos explanted on day 11. Upper case values are experimental results after culture. Lower case values are littermate controls. In wino values denoted by V. Each data point is the mean of approximately three embryos.
to use roller pump tubing with a low oxy- gen diffusibility. Unfortunately the tubing used (Tygon) has a relatively short life in service. Long-lived silicone rubber tubing is much too permeable to oxygen to main- tain high 0 2 levels.
It is difficult to maintain the sys- tem completely closed and still record the heartbeats with the laser technic. If the bubble bypasses, which act as surge cham- bers, are closed with serum stoppers rather than gauze dust traps, enough mo- tion is induced in the embryos owing to pumping to introduce serious artifacts
2.
into the heartbeat record. However we have observed relatively little problem with microorganism contamination when only gauze covers the bubble bypass stand- pipes. The antibiotics used appear to be completely effective over the course of our 24-h experiments, but they represent an additional uncertainty with respect to growth and development of the embryos.
3. Although the components of the PLASMOM itself cost relatively little a complete system including a controlled pump, instrumentation for monitoring the ambient conditions, and heart-rate
I N VITRO CULTURE OF RAT EMBRYOS 375
A A
cr, - A Temper at u re .n I6O- Heart rate
9 Heart rate 5 140-0 .s E . irregular and
-"A* not measurable A. a
A. L \
8120- & a
2 2. A 2 b t a 9 P, 100-
x P, 4.
& A&-
-
- da I
A. *A .*A
P 80-
$ 6 0 I I I I I I I I I I I I I I
A. A%
- AA .&A A '4
. D
a = -21.8f0.2(fS.E)% b = 7.2 f 0. I O h per degree 3 = 2.6 degrees
- 40
2
9,
- 3 8 3 cb \
-36 5 Q I
-34 cb
2 - 3 2 3 - 3. - 3 0 3
5
a -
60 / I I 1
7 6 5 4 3 2 1 0 Drop in temperature from baseline "C ( A T )
Fig. 9 Effect of temperature on heart rate of 25-somite (day 11) rat embryos. Linear regression fit to data averaged at each temperature and re- stricted to temperature changes below base line temperature. Each data point is the mean of be- tween one and six observations.
measuring equipment represents a sub- stantial financial investment, so that ex- periments requiring multiple preparations become very expensive.
4. Our 24-h growth results are still generally rather inferior to what is ob- served in viuo over the same interval of time. It is not clear whether this is due to a factor in the PLASMOM system itself or to the lack of maternal mediation.
Until it is possible to maintain normal growth and function of the explanted chorioallantoic placenta we may be unable to improve on the reported growth results. The effect of maternal metabolism on the embryo certainly must be another im- portant element that is nearly impossible to reproduce in this in vitro system.
The biological in vitro results as mea- sured by the increment in somite number are about 89% of what is achieved in uiuo during the corresponding embryonic pe- riod. The protein and length increment however are less, namely 59 and 52% of the in viuo change. A large number of fac- tors could contribute to these shortcom- ings, but the lack of regulative maternal organ systems to modulate the level of glucose, oxygen, and other substances is the most likely fault. Oxygen may not be
Heart Rate Response of 25 Somite (Day 1 1 113) Rat Embryo to Variations of Ambient Temperature in PLASMOM Culture
Elapsed time - minutes Fig. 10 Correlated heart rate and temperature for a day-I1 rat embryo. Elapsed time
measured from start of temperature transient.

3 76 M. A. ROBKIN, T. H. SHEPARD AND T. TANIMURA
a limiting factor since New and Coppola (‘70) have grown 29-somite rat embryos in 2 atm of oxygen for 40-45 h and found no increase in protein and somites over those grown in 1 atm. Tanimura and Shepard (‘70) reported high utilization of glucose by rat embryos in vitro, and we have evidence from heart-rate increases that addition of extra glucose to the sys- tem after 8-10 h is a benefit. We have also observed that the lactate in the 10 ml of medium doubles during a 20-h culture period .
An exact comparison with the results obtained by New (’67) is not possible be- cause he did not give in vivo values after day 11 and most of his measurements were made after 40-h culture periods. His best 40-h results after explanting 22- to 28- somite embryos were means of 43 somites and 1152 mg of protein. Our explanted embryos, starting at 24 somites and 236 mg of protein, attained the 33.5-somite stage and 632 mg of protein in a 22-h in vitro period. A discrepant increase in somite number
over protein during in vitro culture was also reported by Berry (‘68) in rats, and Klein et al. (‘64) in chicks. Our studies, in view of the lack of any marked cell death, suggest that increased embryonic protein and morphogenesis may not be completely integrated.
LITERATURE CITED
Abromowitz, M., and I. A. Stegun 1964 Hand- book of Mathematical Functions with Formulas,
Graphs and Mathematical Tables. National Bureau of Standards, Washington, D. C.
Berry, C. 1968 Comparison of in vivo and in vitro growth of the rat foetus. Nature, 219:
Klein, N. W., E. McConnell and D. J. Riquier 1964 Enhanced growth and survival of ex- planted chick embryos cultured under high levels of oxygen. Dev. Biol., 10: 17-44.
New. D. A. T. 1967 Develoument of exulanted
92-93.
embryos in circulating medium. J. Embryol. Exp. Morph., 17: 513-525.
New, D. A. T., and P. T. Coppola 1970 Devel- opment of explanted rat fetuses in hyperbaric oxygen. Teratology, 3: 153-159.
New, D. A. T., and J. C. Daniel 1969 Cultiva- tion of rat embryos explanted at 7.5 and 8.5 days of gestation. Nature, 223: 515516.
Nicholas, J. S. 1938 The development of rat embryos in a circulatory medium. Anat. Rec., 70: 199-210.
Nicholas, J. S., and D. Rudnick 1934 Develop- ment of rat embryos in tissue culture. Proc. Nat. Acad. Sci., 20: 656-658.
1938 Development of rat embryos of egg-cylinder to head-fold stages in plasma cul- ture. J. Exp. Zool., 78: 205-232.
Shepard, T. H., T. Tanimura and M. A. Robkin 1971a In vitro studies for analysis of terato- genic events. In: Malformations Conghitales des Mammifkes. H. Tuchmann-Duplessis, ed. Masson, Paris, pp. 51-66.
1971b Energy metabolism in early mammalian embryos. Dev. Biol. Suppl, 4: 42- 58.
Tamann, A., and K. W. Jones 1968 A circulat- ing medium system permitting manipulation during culture of postimplantation embryos. Acta Embryol. Morph. Exp., 10: 288-301.
Tanimura, T., and T. H. Shepard 1970 Glucose metabolism by rat embryos in vitro. Proc. SOC. Exp. Biol. Med., 135: 51-54.
Waddington, C. H., and A. J. Waterman 1933 The development in vitro of young rabbit em- bryos. J. Anat., 67: 355-370.
Related Documents











