The FASEB Journal express article 10.1096/fj.03-1154fje. Published online July 9, 2004. A new type of antimicrobial protein with multiple histidines from the hard tick, Amblyomma hebraeum Ren Lai,* Hajime Takeuchi,* Lee O. Lomas, † Jan Jonczy, ‡ Daniel J. Rigden,* Huw H. Rees,* and Philip C. Turner * *Cellular Regulation and Signaling Division, School of Biological Sciences, University of Liverpool, Liverpool, United Kingdom; † Ciphergen Biosystems Inc., Fremont, California; ‡ Novartis Animal Health Centre de Recherche Santé Animale, Switzerland Corresponding author: Huw H. Rees Cellular Regulation and Signaling Division, School of Biological Sciences, University of Liverpool, Biosciences Building, Crown Street, Liverpool, L69 7ZB, United Kingdom. E-mail: [email protected] ABSTRACT A novel 11 kDa antimicrobial protein, named as hebraein, and having a unique amino acid sequence, was purified from the hemolymph of fed female Amblyomma hebraeum ticks. A full-length cDNA clone encoding hebraein was isolated from a cDNA library made from tick synganglia. Hebraein consists of 102 amino acids, including 6 cysteine residues; has 9 histidines in its C-terminal domain that are mainly present as HX repeats; and has no significant similarity to any known protein. The secondary structure prediction is very clearly all α-helical (4–6 helices) except for a very short extension at the C terminus. Such high α-helical content is quite different from known antimicrobial proteins. Recombinant hebraein and a mutant lacking the histidine residues in the C-terminal domain were constructed and expressed. Assayed at the slightly acidic pH equivalent of fed female tick hemolymph, the wild-type and the histidine-rich recombinant hebraein had stronger antimicrobial activities than the histidine-deficient mutant. The pH-dependent properties of histidine-rich antimicrobial proteins may allow the design of agents that would function selectively in specific pH environments. The results from protein profiling of hemolymph, analyzed by surface-enhanced laser desorption/ionization time-of-flight (SELDI-TOF) mass spectrometry combined with ProteinChip technology and RT-PCR analysis suggested that this antimicrobial protein was up-regulated by blood feeding. Our findings describe a new type of antimicrobial protein with multiple cysteine and histidine residues, and with unique secondary structure. Key words: hebraein • innate immunity • pH-dependent • hemolymph • SELDI-TOF uring the past decade, endogenous antimicrobial peptides have become recognized as important, ubiquitous, and ancient contributors to the innate mechanisms that permit animals and plants to resist infection (1). A large number of antimicrobial peptides have been found in insects, amphibians, and mammals. Most of these antimicrobial peptides are amphipathic and possess either an α-helical or a cystine-stabilized β-sheet structure (1). Known antimicrobial peptides can be divided into several groups, which include linear peptides that form amphipathic and hydrophobic helices, cyclic peptides and small proteins that form β-sheet D Page 1 of 17 (page number not for citation purposes)

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The FASEB Journal express article 10.1096/fj.03-1154fje. Published online July 9, 2004.
A new type of antimicrobial protein with multiple histidines from the hard tick, Amblyomma hebraeum Ren Lai,* Hajime Takeuchi,* Lee O. Lomas,† Jan Jonczy,‡ Daniel J. Rigden,* Huw H. Rees,* and Philip C. Turner*
*Cellular Regulation and Signaling Division, School of Biological Sciences, University of Liverpool, Liverpool, United Kingdom; †Ciphergen Biosystems Inc., Fremont, California; ‡Novartis Animal Health Centre de Recherche Santé Animale, Switzerland
Corresponding author: Huw H. Rees Cellular Regulation and Signaling Division, School of Biological Sciences, University of Liverpool, Biosciences Building, Crown Street, Liverpool, L69 7ZB, United Kingdom. E-mail: [email protected]
ABSTRACT
A novel 11 kDa antimicrobial protein, named as hebraein, and having a unique amino acid sequence, was purified from the hemolymph of fed female Amblyomma hebraeum ticks. A full-length cDNA clone encoding hebraein was isolated from a cDNA library made from tick synganglia. Hebraein consists of 102 amino acids, including 6 cysteine residues; has 9 histidines in its C-terminal domain that are mainly present as HX repeats; and has no significant similarity to any known protein. The secondary structure prediction is very clearly all α-helical (4–6 helices) except for a very short extension at the C terminus. Such high α-helical content is quite different from known antimicrobial proteins. Recombinant hebraein and a mutant lacking the histidine residues in the C-terminal domain were constructed and expressed. Assayed at the slightly acidic pH equivalent of fed female tick hemolymph, the wild-type and the histidine-rich recombinant hebraein had stronger antimicrobial activities than the histidine-deficient mutant. The pH-dependent properties of histidine-rich antimicrobial proteins may allow the design of agents that would function selectively in specific pH environments. The results from protein profiling of hemolymph, analyzed by surface-enhanced laser desorption/ionization time-of-flight (SELDI-TOF) mass spectrometry combined with ProteinChip technology and RT-PCR analysis suggested that this antimicrobial protein was up-regulated by blood feeding. Our findings describe a new type of antimicrobial protein with multiple cysteine and histidine residues, and with unique secondary structure.
Key words: hebraein • innate immunity • pH-dependent • hemolymph • SELDI-TOF
uring the past decade, endogenous antimicrobial peptides have become recognized as important, ubiquitous, and ancient contributors to the innate mechanisms that permit animals and plants to resist infection (1). A large number of antimicrobial peptides have
been found in insects, amphibians, and mammals. Most of these antimicrobial peptides are amphipathic and possess either an α-helical or a cystine-stabilized β-sheet structure (1). Known antimicrobial peptides can be divided into several groups, which include linear peptides that form amphipathic and hydrophobic helices, cyclic peptides and small proteins that form β-sheet
D
Page 1 of 17(page number not for citation purposes)

structures, peptides with unique amino acid compositions, cyclic peptides with thio-ether groups in the ring, and lipopeptides terminating in an amino alcohol and macrocyclic knotted peptides (1). In spite of great primary structure diversity, most antimicrobial peptides documented are now characterized by a preponderance of cationic and hydrophobic amino acids, which are spatially organized in discrete sectors of the molecule (1). The only other known anionic antimicrobial peptide, apart from a proteolytic fragment of hemoglobin (2), is maximin H 5 from the amphibian, Bombina maxima (3).
On a global basis, ticks are second only to mosquitoes as vectors of disease-causing agents to humans, and they are the most important arthropod transmitting pathogens to other animal species (4). Ticks produce prostaglandins, anticoagulants, immunosuppressive anti-inflammatory protein molecules, and antimicrobial peptides that help overcome host defensive responses, such as keeping the blood sterile and preventing the blood from coagulation in the tick body (4). The innate immunity of ticks is difficult to understand, although several defensin-like antimicrobial peptides have been found in tick hemolymph and saliva (5, 6). During differential screening between fed and unfed female ticks, we found a cDNA encoding a putative protein (named as hebraein) possessing multiple histidine residues near its C terminus that we suspected to have antimicrobial activity, because of known antimicrobial peptides that are rich in certain specific amino acids, such as histidine or tryptophan (7–9). Here, we report the characterization of this novel antimicrobial protein that possesses a unique primary structure, from the fed female tick Amblyomma hebraeum.
MATERIALS AND METHODS
Animals
Adult ticks, Amblyomma hebraeum (Koch; Ixodidae), were reared in the laboratory according to the method of Kaufman and Phillips and Lomas et al. (10, 11). Engorged female ticks (1 g and larger) were allowed to detach spontaneously (generally 8–10 days after attachment). After detachment, ticks were maintained at 26°C and > 90% humidity.
SMART cDNA synthesis
Total RNA was extracted using TRIzol (Life Technologies, Ltd.) from the synganglia (n=50) dissected from fed or unfed female ticks. cDNA was synthesized by SMARTTM techniques by using a SMARTTM PCR cDNA synthesis kit (Clontech, Palo Alto, CA). The first strand was synthesized by using cDNA 3′ SMART CDS Primer II A, 5′-AAGCAGTGGTATCAACGCAGAGTACT (30) N-1N-3′ (N = A, C, G or T; N-1 = A, G or C), and SMART II A oligonucleotide, 5′-AAGCAGTGGTATCAACGCAGAGTACGCGGG-3′. The second strand was amplified using Advantage polymerase by 5′ PCR primer II A, 5′-AAGCAGTGGTATCAACGCAGAGT- 3′.
PCR-selectTM cDNA subtractive hybridization and cDNA library construction for differentially expressed genes
Clontech PCR – SelectTM cDNA Subtraction Kit (Clontech) was used for cDNA subtractive hybridization to compare two populations of synganglia mRNA from fed and unfed female ticks and obtain clones of genes that are differentially expressed in these two populations. Briefly, double-stranded cDNA was digested by Rsa I to produce tester cDNA that contains specific
Page 2 of 17(page number not for citation purposes)

(differentially expressed) transcripts and driver cDNAs that is a reference. After digestion, only tester cDNA was ligated with the adaptor provided with the kit. Adaptor-ligated tester cDNA was hybridized twice with driver cDNA according to the user manual from Clontech. After this step, differentially expressed cDNA was selectively amplified by a two-step PCR using Advantage cDNA polymerase mix according to the manufacturer’s instructions (Clontech). Finally, the PCR products were cloned into pGEM®-T Easy vector (Promega, Madison, WI). Both a forward subtracted library (cDNA from fed tick synganglia as the tester and cDNA from unfed tick synganglia as the driver) and a reverse subtracted library (using cDNA from unfed tick synganglia as the tester and cDNA from fed tick synganglia as the driver) were constructed, respectively.
Differential screening of the subtracted cDNA library
Individual white colonies from forward- and reverse-subtracted libraries were picked from X-gal/IPTG LB (Luria-Bertani)-ampicillin plates and used as templates for colony PCR. The colony PCR was performed as follows: 1 cycle of 94°C for 3 min and 30 cycles of 94°C for 30 s, 60°C for 30 s, 72°C for 3 min and 1 cycle of 72°C for 15 min using M13 forward and reverse primers (5′-GTAAAACGACGGCCAG-3′ and 5′-CAGGAAACAGCTATGAC-3′, respectively). Four identical blots were created by spotting NaOH-denatured PCR products onto Electran® nylon membranes (Merck). Membranes were denatured by soaking in a 0.6 N NaOH solution, neutralized in 0.5 M Tris-HCl, pH 7.5 buffer; washed with water; and cross-linked by UV. Membranes were hybridized separately with α-32P [dCTP]-labeled cDNA probes generated from mRNA samples extracted from either fed or unfed tick synganglia by using a Ready-To-GoTM DNA-labeling kit (-dCTP; Amersham Pharmacia Biotech, Frederick, MD), respectively. Prehybridzation and hybridization were performed using ULTRAhybTM (Ambion Inc., Austin, TX) under the conditions recommended by the manufacturer. The blots were washed at low stringency (2 × SSC and 0.1% SDS at room temperature, where 1 × SSC is 0.15 M NaCl, 0.015 M sodium citrate) and labeled spots were visualized by autoradiography. Clones that displayed differential hybridization were selected for further study, and the inserts were sequenced.
Hemolymph collection
Hemolymph was collected from engorged female ticks 4 days after detachment by cutting to make a small hole at the base of the forelegs, applying gentle pressure on the tick’s body, and drawing the clear liquid into a plastic micropipette (5–100 µl per tick). Hemolymph was diluted 1:1 in ice-cold 0.2 M acetic acid, and the sample was centrifuged at 10, 000 × g for 10 min, the supernatant retained and stored at –80°C until required.
Protein purification
Ni-CAMTM Histidine-tag affinity resin (0.5 ml; Sigma, St. Louis, MO) was transferred to a 20 ml chromatography column and washed with two volumes of deionized water and then three volumes of equilibration buffer (50 mM sodium phosphate, pH 8.0; 0.3 M sodium chloride; 10 mM imidazole). Acetic acid-treated hemolymph sample (0.5 ml) was then loaded onto the column at a flow rate of 20 ml/h and after all of the extract was loaded, the column was washed with buffer (the same as equilibration buffer) at a flow rate of 20 ml/h until no absorbance at 280 nm was observed. The proteins that were attached onto the Ni-CAMTM Histidine-tag affinity resin were eluted by elution buffer (50 mM sodium phosphate, pH 8.0; 0.3 M sodium chloride; 250 mM imidazole) at a flow rate of 20 ml/h. The eluate was filtered through a 50 kDa cutoff Centriprep filter (Millipore,
Page 3 of 17(page number not for citation purposes)

Bedford, CA), and the filtrate was concentrated by using a Centriprep (3 kDa cutoff). Concentrated filtrate was applied to a 5 × 250 mm Vydac C18 RP-HPLC (reversed-phase high-performance liquid chromatography) column (Sigma) equilibrated with 0.1% (v/v) trifluoroacetic acid/water. Elution was performed with an acetonitrile gradient (10–60% acetonitrile in 0.1% (v/v) trifluoroacetic acid/water over 50 min) at a flow rate of 0.7 ml/min. UV-absorbing peaks were collected, concentrated to a small volume by vacuum centrifugation, and assayed for antimicrobial activity. The fraction with antimicrobial activity was collected and purified further by C18 RP-HPLC under the same conditions and concentrated as before.
SDS-PAGE (PAGE)
About 2 µg of purified protein from HPLC was mixed with 10 µl of SDS sample buffer and boiled for 5 min before loading onto a 16% polyacrylamide gel. After electrophoresis, the gel was stained with Coomassie brilliant blue and destained to allow visualization of the protein.
Protein sequencing and matrix-assisted laser desorption ionization time-of-flight (MALDI-TOF) mass spectrometry
The purified peptide was subjected to sequence analysis by the automated Edman degradation method on an Applied Biosystems pulsed liquid-phase sequencer, model ABI 491. Mass measurement was performed in a MALDI-TOF instrument (Bruker Reflex), using α-cyano-4-hydroxycinnamic acid (HCCA) suspended in 100% acetonitrile as matrix.
Recombinant hebraein expression vector
For expression of hebraein and its mutant, full-length cDNA was used as a template to amplify only the cDNA that begins at the initial methionine and ends at the first stop codon. A Kozak consensus sequence (ANNATGG) was added and the DNA amplified. For unaltered recombinant hebraein containing all 10 histidine residues in its C-terminal region, the forward primer was 5′-AAAATGGACGCTGTCTTCGCCTCCTGT-3′, and the reverse primer was Anti5-AS0 5′-CTAATGGTGGTCGTGGTCATGACC-3′. For the mutant without histidine residues in its C-terminal region, the same forward primer was used but the reverse primer was Anti5-AS1 5′- CTAGCGGGCGTCGGGATCGCAAGT-3′, which created a deletion lacking the C-terminal 18 residues (Fig. 1). PCR conditions were 95°C (3 min), and 30 cycles of 95°C (30 s), 68°C (30 s), and 72°C (3 min), followed by a 15-min extension period at 72°C. The PCR products were purified by the Wizard® SV Gel and PCR Clean-Up System (Promega) and were cloned into the vector pIB/V5-His TOPO (Invitrogen, Carlsbad, CA) following the manufacturer's instructions. Genes of interest that were inserted in the correct orientation were confirmed by sequencing the plasmid constructs using the sequencing primers provided by the manufacturer.
Expression of hebraein in an insect cell line (High Five)
High FiveTM cells (Invitrogen) cultured in High Five serum-free medium (Invitrogen) supplemented with Cellfectin® reagent in 24-well plates were used for transfection with hebraein, its mutant or antisense (control) construct plasmids, respectively. The three transfections for the recombinant hebraein, histidine-deficient mutant, and control were done in the same volume (2 ml) of High Five cells at the same concentration (2 × 106 cells/ml, >95% viability). The transfection procedure followed the manufacturer’s instructions. In each case, six transfections
Page 4 of 17(page number not for citation purposes)

were done in parallel; after 2 days of incubation, the supernatant was collected and stored at – 80°C for further analysis.
Antimicrobial assay
The standard bacterial and fungal strains used in antimicrobial assays were the gram-positive bacterium Staphylococcus aureus (Oxford), the gram-negative bacterium Escherichia coli (OP 50), and fungi strains Candida albicans SC 5314 and Candida glabrato ATCC 2001. Bacteria were first grown in LB broth to an OD600 nm of 0.8. A 10 µl aliquot of the bacteria was then taken and added to 8 ml of fresh LB broth with 0.7% agar and poured over a 90 mm Petri dish containing 25 ml of 1.5% agar in LB broth. After the top agar hardened, a 20 µl aliquot of the test sample filtered on a 0.22 µm Millipore filter was dropped onto the surface of the top agar and completely dried before incubation overnight at 37°C. If a sample contained antimicrobial activity, a clear zone formed on the surface of the top agar representing inhibition of bacterial growth. The minimal inhibitory concentration (MIC) was determined in liquid LB medium by incubating the bacteria in 96-well plates with LB broth and variable amounts of the sample tested. The MIC at which no visible growth occurred was recorded. In the assays of anti-fungal activity, fungi were cultured with variable amounts of the sample tested in yeast extract-peptone-dextrose broth (12). Three independent experiments were performed for each sample.
Because 9 histidine residues are clustered in the C-terminal region of hebraein (NH2……RHDHDHDHGHGHGHDHDPH-COOH) and a digestion site for trypsin (….RH…) precedes the first-histidine, trypsin was used to digest hebraein to obtain the C-terminal peptide fragment with the H-X repeats. Trypsin (10 µg; Promega) was incubated with 200 µg hebraein at 37°C for 24 h in 0.2 M NH4HCO3 solution (pH 8.0), and the digestion solution was loaded onto a Ni-CAMTM Histidine-tag affinity resin. The procedure was the same as for purification of hebraein from hemolymph (above). All the fractions (including the wash fraction and eluate) were subjected to filtration through a 10-kDa cutoff Centriprep filter, and the filtrates were concentrated by centrifugal vacuum evaporation and examined for their antimicrobial activities using the same method as described above.
SELDI profiling of hemolymph to study differential expression of hebraein
Lyophilized hemolymph samples were reconstituted in 5 µl of 50 mM Tris-HCl buffer containing 9 M urea and 1% CHAPS (pH 8). After vortexing, samples were diluted 1:10 in 50 mM Tris-HCl buffer (pH 8), then 10 µl was incubated on spots of a weak anionic CM10 ProteinChipTM Array (Ciphergen Biosystem Inc.) previously equilibrated in 50 mM ammonium acetate buffer (pH 4.5; 5 µl incubation for 5 min). After incubation for 30 min at ambient temperature and high humidity, the sample droplet was removed and the CM10 spot was subsequently washed twice with 50 mM Tris-HCl buffer (pH 8; 10 µl each wash). The spots were finally washed once with 5 µl of 5 mM HEPES buffer (pH 7.2), then allowed to air-dry. Two 0.5 µl applications of CHCA [α-cyano-7-hydroxycinnamic acid (Ciphergen Biosystems Inc.); 20% saturated in 50% acetonitrile, 0.1% trifluoroacetic acid (Sigma)] were added to each spot. Once dried, the ProteinChip array was inserted into the ProteinChip Reader (PBSIIc, Ciphergen Biosystems Inc.) and analyzed according to an automated data collection protocol. The ProteinChip Reader, a linear time-of-flight mass spectrometer equipped with a 337 nm nitrogen laser was operated in positive-ion mode. The total accelerating potential was +20 kV, and the extraction delay time was
Page 5 of 17(page number not for citation purposes)

set to 528 ns. A total of 65 transients were averaged over 66% of the target area in a linear sweep to generate each spectrum.
RT-PCR
PCR was performed to analyze gene expression of hebraein in A. hebraeum. Total RNA extraction from synganglia of fed and unfed female ticks and first-strand cDNA synthesis was the same as described above. One-tenth of each resulting cDNA was used in the subsequent PCR. The primers were forward primer, 5′-AAAATGGACGCTGTCTTCGCCTCCTGT-3′, and reverse primer, Anti5-AS0 5′-CTAATGGTGGTCGTGGTCATGACC-3′. PCR conditions were 95°C (3 min), and 30 cycles of 95°C (30 s), 68°C (30 s), and 72°C (3 min), followed by a 15-min extension period at 72°C. Control PCR was performed by using specific primers for tick actin (forward primer, actin-s 5′-GGAAACGTATGACGTGATAG-3′, and reverse primer, actin-as 5′-CGTGGCATACAGGCTTAGGACG-3′), using the same conditions as above, except for an annealing temperature of 57°C.
RESULTS
Cloning of hebraein cDNA
In the course of our studies of genes that are differentially expressed in synganglia following feeding, we sequenced differentially expressed cDNAs from our forward subtracted library of female ticks. One of the clones contained an insert cDNA fragment of 0.7 kb encoding a small predicted protein with six cysteine residues and multiple histidine residues in its C-terminal region (Fig. 1, GenBank accession no AY437139). The mature protein predicted from the cDNA sequence was named as hebraein. Conceptual translation of the cDNA sequence suggested that hebraein (102 amino acids) is produced as a pro-protein consisting of 123 amino acid residues with 9 histidine residues in its C-terminal region. This latter feature suggested that this protein may have antimicrobial activity because of antimicrobial peptides that are rich in certain specific amino acids, such as histidine or tryptophan (7–9, 13–15).
The secondary structure of hebraein was predicted by using several software packages that included GOR3, GOR4, Hierarchical Neural Network, and NNPREDICT. The secondary structure prediction is very clearly all α-helical (4–6 helices) except for a very short extension at the C terminus, which is quite different from known antimicrobial peptides/proteins because most of these antimicrobial peptides are amphipathic and possess either a single α-helix or a cystine-stabilized β-sheet structure (1). All the predicted α-helices are evenly distributed throughout the whole protein.
Partial amino acid sequencing and molecular weight
Because the cDNA sequence for hebraein predicts a primary structure with a cluster of histidine residues in the C-terminal region, a histidine-tag affinity column was used to isolate hebraein from female tick hemolymph. The eluate from the histidine-tag affinity column was purified by reversed-phase high-performance liquid chromatography. A fraction containing antimicrobial activity was eluted at 30% acetonitrile in 0.1% trifluoracetic acid and purified further by RP-HPLC on the same system (Fig. 2). This fraction was collected and concentrated for protein sequencing. The N-terminal partial amino acid sequence obtained was
Page 6 of 17(page number not for citation purposes)

His-Leu-Glu-Leu-X-Lys-Lys-Gln-Lys-Asp-Gln-Val-Leu-Ala-Gly-Thr-Glu-Leu-Glu-X-X-Lys-X, which matched sufficiently well with the amino acid sequence at 22-38 deduced from the cDNA sequence, allowing for the problems of sequencing small amounts of material (Fig. 1). This shows that the protein is processed and secreted into the hemolymph from fed female ticks, although we cloned the cDNA from the synganglia mRNA of fed female ticks. The molecular weight of purified hebraein was approx. 11 kDa as determined by SDS-PAGE (Fig. 3), and this value is close to the predicted molecular weight for the cleaved polypeptide product predicted from the cDNA sequence (11434 Da). The MALDI-TOF analysis gave a mass of 11427.1, which matches well with the predicted molecular weight of the mature protein from the cDNA sequence and our SDS-PAGE determination and strongly suggests that the six cysteines form three disulfide bridges.
Recombinant hebraein was purified as described in Materials and Methods; however, it is not possible to purify the deletion mutant protein that has no histidine residues in its C-terminal region by this procedure. The supernatant containing mutant protein was successively passed through 30 kDa cutoff Centriprep and 10 kDa cutoff Centriprep filters, and then the material retained was purified by two-step RP-HPLC. The purified proteins were analyzed by SDS-PAGE and shown to be pure (Fig. 3).
Antimicrobial assays
Native hebraein purified from the hemolymph of fed female ticks and recombinant hebraeins expressed in insect cells were assayed for their antimicrobial activities under different pH conditions. The antimicrobial activities (minimal inhibitory concentration, MIC) of native hebraein, recombinant hebraein, as well as mutant protein, against S. aureus Oxford, E. coli OP 50, C. albicans SC 5314, and C. glabrato ATCC 2001 are shown in Table 1. The antimicrobial activities of the histidine-rich, wild-type hebraein samples are substantially greater at pH 6.2 than at pH 7.6. By contrast, the histidine-poor mutant exerted similar antimicrobial activities at both pH 6.2 and 7.6. At the physiological pH equivalent of fed female tick hemolymph (pH 6.2–6.4), the histidine-rich samples had stronger antimicrobial activities than the histidine-poor mutant, except against S. aureus. Although wild-type hebraein showed significant antimicrobial activity against C. glabrato, neither wild-type nor mutant showed antimicrobial activity against C. albicans.
The C-terminal peptide fragment (NH2-HDHDHDHGHGHGHDHDPH-COOH) of hebraein obtained by trypsin digestion did not display any detectable antimicrobial activity in our experiments with the four species of test organisms. It suggested that this fragment alone did not function as an antimicrobial factor.
SELDI profiling of hemolymph and RT-PCR
The hemolymph samples from female ticks (0 and 4 days after detachment) were assayed with CM10 ProteinChip arrays. The ProteinChip software provided by the manufacturer was used to identify clusters of peaks differentially present in these hemolymph samples. Among 35 clusters differentially present, one peak had a mass of 11425.3 ± 3 (Fig. 4) that matched well with the molecular weight of hebraein predicted from the gene sequence and MALDI-TOF mass spectrometry analysis. RT-PCR analysis revealed that the gene encoding hebraein in synganglia of female ticks was up-regulated by blood feeding (Fig. 5). The results from SELDI profiling and RT-PCR confirmed that the gene was differentially expressed during feeding, as was revealed
Page 7 of 17(page number not for citation purposes)

during cloning by differentially screening our library using 32P[dCTP]-labeled cDNA probes. Furthermore, SELDI profiling indicated that differential expression of hebraein occurred during the first four days post-engorgement.
DISCUSSION
Ticks have many opportunities to encounter microorganisms because of their special feeding behavior. Antimicrobial peptides/proteins are an innate immune system to prevent pathogenic microorganisms. In terms of co-evolution, perhaps ticks have developed multiple antimicrobial factors because they have to encounter a large diversity of pathogenic microbes that include bacteria, fungi, and viruses from their hosts during their life cycle. With the growing problem of pathogenic organisms that resist conventional antibiotics, ticks are becoming fruitful sources to find novel pharmaceutical substances to treat infections. Several defensin-like antimicrobial peptides, which are very common in insects, have been found in ticks (5, 6), and hebraein is now the second type of antimicrobial protein isolated from ticks.
Hebraein has a unique primary structure that consists of six evenly distributed cysteine residues and a histidine cluster in its C terminus that contains eight histidine-aspartic acid- or histidine-glycine-continuous repeats. The lack of similarity to any known proteins in existing databases suggested that hebraein is a novel protein. Secondary structure prediction suggests that hebraein is a protein with multiple α-helixes (4–6 α-helixes) that are distributed evenly. This multi-α-helical structure is quite different from that of known antimicrobial proteins/peptides many of which are peptides with a single α-helix (1).
Hebraein's novel primary structure, comprising six cysteine residues and a histidine-rich C terminus, suggests that it is a novel antimicrobial protein. However, when we were writing this paper, a cDNA sequence (GenBank accession number AY233212) cloned from another species of hard tick, Boophilus microplus, was published, and the predicted protein was named as microplusin (16). Microplusin has the same cysteine motif as hebraein, but it has only four histidine residues near its C terminus. Aligning hebraein with microplusin gives an identity of 62% and a similarity of 73% between these two sequences, and our secondary structure prediction suggested that microplusin has 4–6 α-helixes as does hebraein. The high identity and similar secondary structure between these two proteins from two different hard ticks suggests that they belong to the same protein family. Both members of this protein family have multiple histidine residues near their C terminus, suggesting that histidine residues play an important role, although more proof is needed to support this hypothesis. Although known antimicrobial peptides/proteins have encompassed a wide variety of structural motifs (17, 18), some new antimicrobial peptides/proteins with unique structural motifs are being found. The unique primary structure of hebraein and microplusin suggests that this family of proteins contains a novel structural motif.
Native hebraein and its recombinant forms displayed their antimicrobial activities against the Gram positive bacterium S. aureus, the Gram negative bacterium E. coli, and the fungus C. glabrato and showed without doubt that hebraein is an antimicrobial protein with wide specificity. Compared with native and recombinant hebraein, the histidine-poor mutant protein displayed very weak antifungal activity in our experiments. This finding suggests that histidine residues in the primary structure play an important role in the antifungal activity of hebraein, although the C-terminal peptide fragment (-106HDHDHDHGHGHGHDHDPH123) of hebraein did not display any detectable antimicrobial activity in our experiments. However, it is necessary to do further
Page 8 of 17(page number not for citation purposes)

work to identify the actual role of the C-terminal histidine residues. It was noted that the antimicrobial activities of native and recombinant hebraein are substantially greater at pH 6.0 (physiological pH of engorged tick) than at pH 7.6, while the histidine-poor mutant exerted similar antimicrobial activities at both pH 6.0 and 7.6. It seems that hebraein exerts its activity by a pH-dependent behavior as has been found in other histidine-rich antimicrobial peptides, such as histatin-like peptides (19). Histidine residues have a pKa of ~6.5 that makes histidine-rich peptides have high net-positive charges at pH 6.0 yet are relatively uncharged at pH 7.6. The positive charge of antimicrobial peptides undoubtedly facilitates their interactions with anionic microbial components, such as lipopolysaccharide, and increases their antimicrobial potency (19). When a tick blood-feeds, the physiological environment becomes acidic, which increases the positive charges on hebraein and possibly increases its antimicrobial potency. In terms of evolution, a histidine-rich antimicrobial protein like hebraein is very useful to selectively exert antimicrobial function during blood-feeding. Interestingly, the histidine-poor mutant still has high antimicrobial activity against S. aureus. These data may suggest that the activity against S. aureus is independent from the histidine cluster. Furthermore, hebraein showed antimicrobial activity against C. galbrato but not against C. albicans. This may mean that hebraein can work in different ways in different species. Furthermore, the apparent difference in antimicrobial spectrum between hebraein and microplusin could be due to the 27% difference between these peptides, the details of the assays used, or the choice of microorganisms tested (16).
The up-regulated expression of genes for other antimicrobial peptides (defensins), in response to blood-feeding has been observed in the blood-sucking insect, Stomoxys calcitrans (20) and in the soft tick, Ornithodoros moubata (6). The clone-encoding hebraein was isolated from the forward differentially expressed cDNA library; its differential expression was confirmed further by RT-PCR and by SELDI-TOF mass spectrometry. When ticks attach onto their hosts, their living temperature and pH environment are changed, and ticks imbibe a large amount of blood from their hosts. Possibly, one of these changes provides the trigger leading to elevated hebraein expression. However, further work is necessary to identify the mechanism of regulation of hebraein expression.
More diversity of antimicrobial protein types makes understanding the mechanisms of antimicrobial action more complex. At the same time, however, this diversity provides more opportunities for designing novel and effective antimicrobial agents. Taken together: 1) this discovery of tick hebraein with its all helical structure composed of 4–6 α-helixes adds a new member to the increasing family of antimicrobial proteins/peptides; 2) histidine-rich antimicrobial peptides with pH-dependent antimicrobial activity that can render them active at low pH and relatively inactive at neutral pH are advantageous during a tick’s blood-feeding when it encounters an acidic environment; 3) the pH-dependent properties of histidine-rich antimicrobial proteins may allow the design of agents that would function selectively in specific pH environments, for example, hebraein may be a useful template for developing antimicrobial peptides intended to function in acidic environments such as the gastric lumen or vagina of mammals (19); 4) the existence of hebraein in A. hebraeum and microplusin, its homologue in another ixodid tick, B. microplus, suggests another antimicrobial protein/peptide family that is specific for the tick’s innate immunity, apart from the defensin-like peptides, although only two members of this new family have been found as yet. More work is needed to elucidate the 3D-structure of hebraein, identify its complete antimicrobial spectrum, its antimicrobial potency, and its innate immunity mechanism. Indeed, during revision of this manuscript, another histidine rich antimicrobial peptide was reported in the leech, Theromyzon tessulatum (21).
Page 9 of 17(page number not for citation purposes)

ACKNOWLEDGMENTS
We thank the Wellcome Trust for financial support. We are grateful to S. Corrigan for tick maintenance, Mohammad Tabish, Mark Wilkinson, and Mark Laddick for providing microbial strains, and Meng Qingxiong for protein sequencing and mass measurement.
REFERENCES
1. Richard, M. E., and Hans, J. V. (1999) Diversity of antimicrobial peptides and their mechanisms of action. Biochim. Biophys. Acta 1462, 11–28
2. Brogden, K. A., de Lucca, A. J., Bland, J., and Elliott, S. (1996) Isolation of an ovine pulmonary surfactant-associated anionic peptide bactericidal for Pasteurella haemolytica. Proc. Natl. Acad. Sci. USA 93, 412–416
3. Lai, R., Liu, H., Hui, L. W., and Zhang, Y. (2002) An anionic antimicrobial peptide from toad Bombina maxima. Biochem. Biophys. Res. Commun. 295, 796–799
4. Yuan, J., Bowman, A. S., Aljamali, M., Payne, M. R., Tucker, J. S., Dillwith, J. W., Essenberg, R. C., and Sauer, J. R. (2000) Prostaglandin E(2)-stimulated secretion of protein in the salivary glands of the lone star tick via a phosphoinositide signaling pathway. Insect Biochem. Mol. Biol. 30, 1099–1106
5. Johns, R., Sonenshine, D. E., and Hynes, W. L. (2001) Identification of a defensin from the hemolymph of the American dog tick, Dermacentor variabilis. Insect Biochem. Mol. Biol. 31, 957–865
6. Nakajima, Y., van der Goes van Naters-Yasui, A., Taylor, D., and Yamakawa, M. (2002) Antibacterial peptide defensin is involved in midgut immunity of the soft tick, Ornithodoros moubata. Insect Molec. Biol. 11, 611–618
7. Lee, I. H., Zhao, C., Nguyen, T., Menzel, L., Waring, A. J., Sherman, M. A., and Lehrer, R. I. (2001) Clavaspirin, an antibacterial and haemolytic peptide from Styela clava. J. Pept. Res. 58, 445–456
8. Gusman, H., Travis, J., Helmerhorst, E. J., Potempa, J., Troxler, R. F., and Oppenheim, F. G. (2001) Salivary histatin 5 is an inhibitor of both host and bacterial enzymes implicated in periodontal disease. Infect. Immun. 69, 1402–1408
9. Schibli, D. J., Epand, R. F., Vogel, H. J., and Epand, R. M. (2002) Tryptophan-rich antimicrobial peptides: comparative properties and membrane interactions. Biochem. Cell Biol. 80, 667–677
10. Kaufman, W. R., and Phillips, J. E. (1973) Ion and water-balance in Ixodid tick Dermacentor-andersoni. 1. Routes of ion and water excretion. J. Exp. Biol. 58, 523–536
11. Lomas, L. O., Turner, P. C., and Rees, H. H. (1997) A novel neuropeptide-endocrine interaction controlling ecdysteroid production in Ixodid ticks. Proc. R. Soc. Lond. B. Biol. Sci. 264, 589–596
Page 10 of 17(page number not for citation purposes)

12. Lai, R., Zheng, Y. T., Shen, J. H., Liu, G. J., Liu, H., Lee, W. H., Tang, S. Z., and Zhang, Y. (2002) Antimicrobial peptides from skin secretions of Chinese red belly toad Bombina maxima. Peptides 23, 427–435
13. Edgerton, M., Koshlukova, S. E., Araujo, M. W., Patel, R. C., Dong, J., and Bruenn, J. A. (2000) Salivary histatin 5 and human neutrophil defensin 1 kill Candida albicans via shared pathways. Antimicrob. Agents Chemother. 44, 3310–3316
14. Edgerton, M., Koshlukova, S. E., Lo, T. E., Chrzan, B. G., Straubinger, R. M., and Raj, P. A. (1998) Candidacidal activity of salivary histatins - Identification of a histatin 5-binding protein on Candida albicans. J. Biol. Chem. 273, 20438–20447
15. Selsted, M. E., Novotny, M. J., Morris, W. L., Tang, Y. Q., Smith, W., and Cullor, J. S. (1992) Indolicidin, a novel bactericidal tridecapeptide amide from neutrophils. J. Biol. Chem. 267, 4292–4295
16. Fogaca, A. C., Lorenzini, D. M., Kaku, L. M., Esteves, E., Bulet, P., and Daffre, S. (2004) Cysteine-rich antimicrobial peptides of the cattle tick Boophilus microplus: isolation, structural characterization and tissue expression profile. Dev. Comp. Immunol. 28, 191–200
17. Kobayashi, S., Hirakura, Y., and Matsuzaki, K. (2001) Bacteria-selective synergism between the antimicrobial peptides alpha-helical magainin 2 and cyclic beta-sheet tachyplesin I: Toward cocktail therapy. Biochemistry 40, 14330–14335
18. Yan, H., and Hancock, R. E. (2001) Synergistic interactions between mammalian antimicrobial defense peptides. Antimicrob. Agents Chemother. 45, 1558–1560
19. Lee, I. H., Cho, Y., and Lehrer, R. I. (1997) Effects of pH and salinity on the antimicrobial properties of clavanins. Infect. Immun. 65, 2898–2903
20. Lehane, M. J., Wu, D., and Lehane, S. M. (1997) Midgut-specific immune molecules are produced by the blood-sucking insect Stomoxys calcitrans. Proc. Natl. Acad. Sci. USA 94, 11502–11507
21. Tasiemski, A., Vandenbulcke, F., Mitta, G., Lemoine, J., Lefebvre, C., Sautiere, P.-E., and Salzet, M. (2004) Molecular characterization of two novel antimicrobial peptides inducible upon bacterial challenge in an annelid: the leech, Theromyzon tessulatum J. Biol. Chem., In press
Received January 16, 2004; accepted June 8, 2004.
Page 11 of 17(page number not for citation purposes)
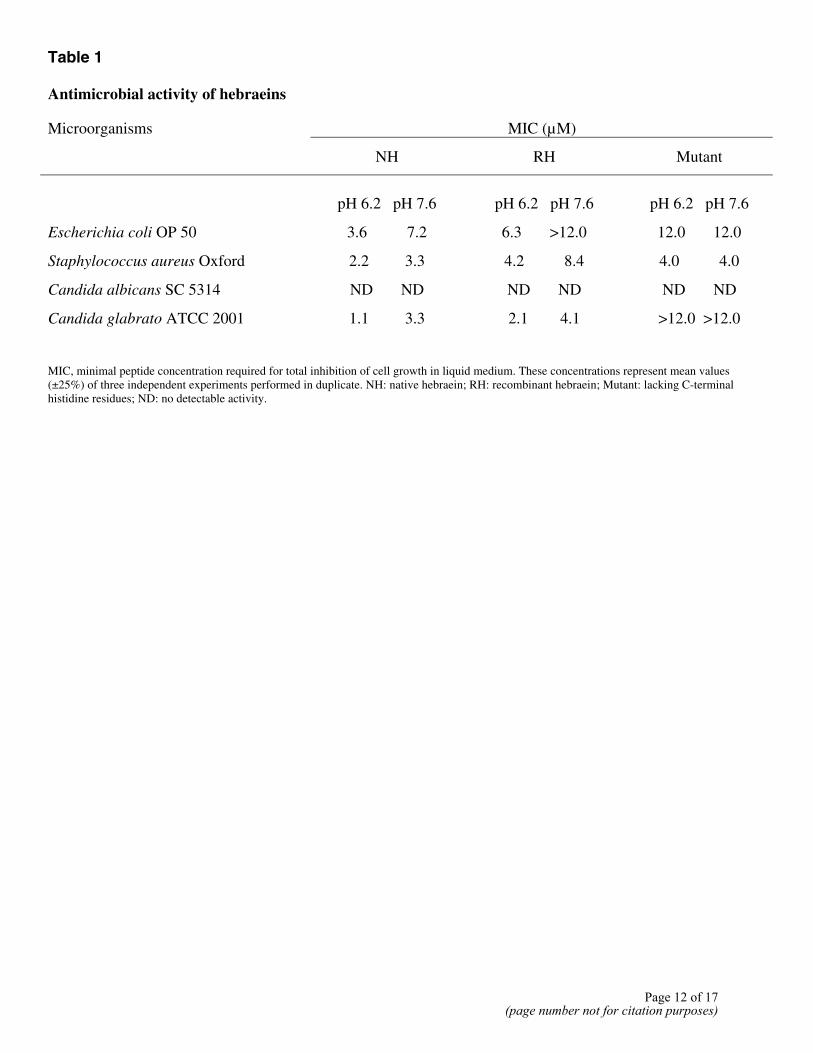
Table 1 Antimicrobial activity of hebraeins
Microorganisms MIC (µM)
NH RH Mutant
pH 6.2 pH 7.6 pH 6.2 pH 7.6 pH 6.2 pH 7.6
Escherichia coli OP 50 3.6 7.2 6.3 >12.0 12.0 12.0
Staphylococcus aureus Oxford 2.2 3.3 4.2 8.4 4.0 4.0
Candida albicans SC 5314 ND ND ND ND ND ND
Candida glabrato ATCC 2001 1.1 3.3 2.1 4.1 >12.0 >12.0
MIC, minimal peptide concentration required for total inhibition of cell growth in liquid medium. These concentrations represent mean values (±25%) of three independent experiments performed in duplicate. NH: native hebraein; RH: recombinant hebraein; Mutant: lacking C-terminal histidine residues; ND: no detectable activity.
Page 12 of 17(page number not for citation purposes)

Fig. 1
Figure 1. Nucleotide and deduced amino acid sequence of the precursor hebraein. Nucleotide numbers are indicated on the right, and amino acid numbers are indicated on the left. The open reading frame is shown in uppercase letters, and the 5′- and 3′-non-coding regions are shown in lowercase letters. The putative polyadenylation signal is double-underlined. The predicted signal peptide is in bold italics, and the N-terminal amino acid sequence experimentally obtained from the purified protein is shown in the box. The stop codon is indicated by asterisk. The position where the Anti5-AS1 primer anneals is indicated by single underlining.
Page 13 of 17(page number not for citation purposes)
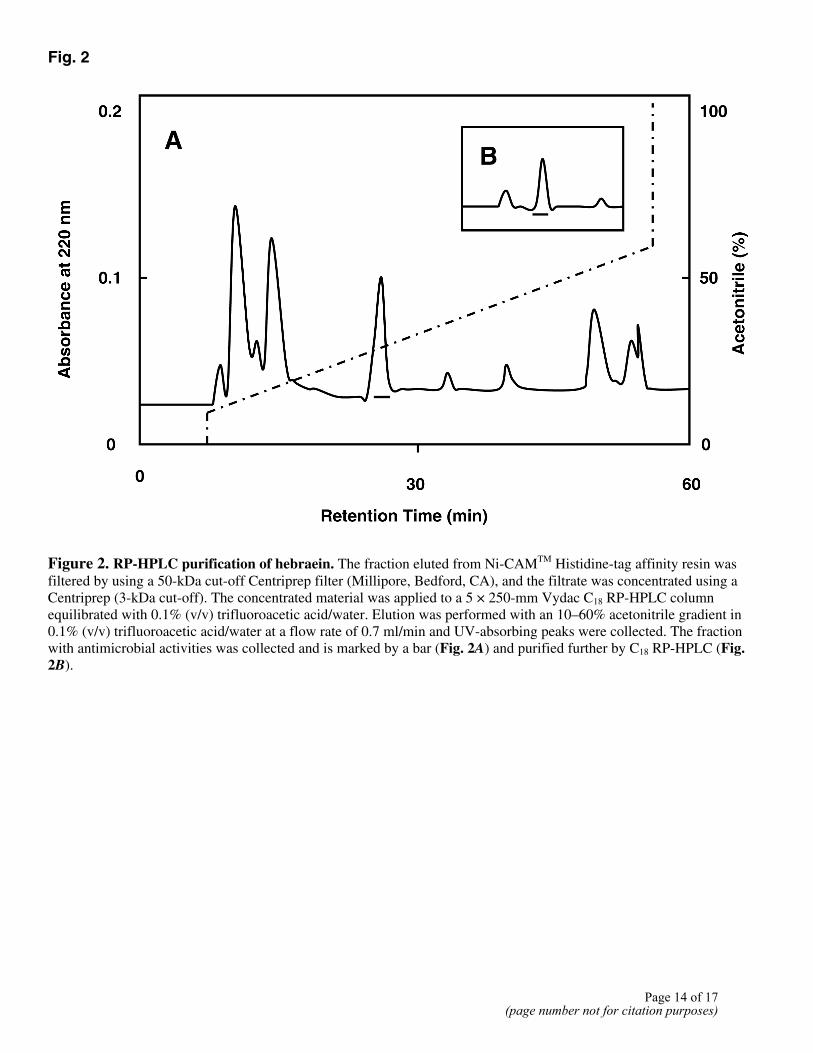
Fig. 2
Figure 2. RP-HPLC purification of hebraein. The fraction eluted from Ni-CAMTM Histidine-tag affinity resin was filtered by using a 50-kDa cut-off Centriprep filter (Millipore, Bedford, CA), and the filtrate was concentrated using a Centriprep (3-kDa cut-off). The concentrated material was applied to a 5 × 250-mm Vydac C18 RP-HPLC column equilibrated with 0.1% (v/v) trifluoroacetic acid/water. Elution was performed with an 10–60% acetonitrile gradient in 0.1% (v/v) trifluoroacetic acid/water at a flow rate of 0.7 ml/min and UV-absorbing peaks were collected. The fraction with antimicrobial activities was collected and is marked by a bar (Fig. 2A) and purified further by C18 RP-HPLC (Fig. 2B).
Page 14 of 17(page number not for citation purposes)
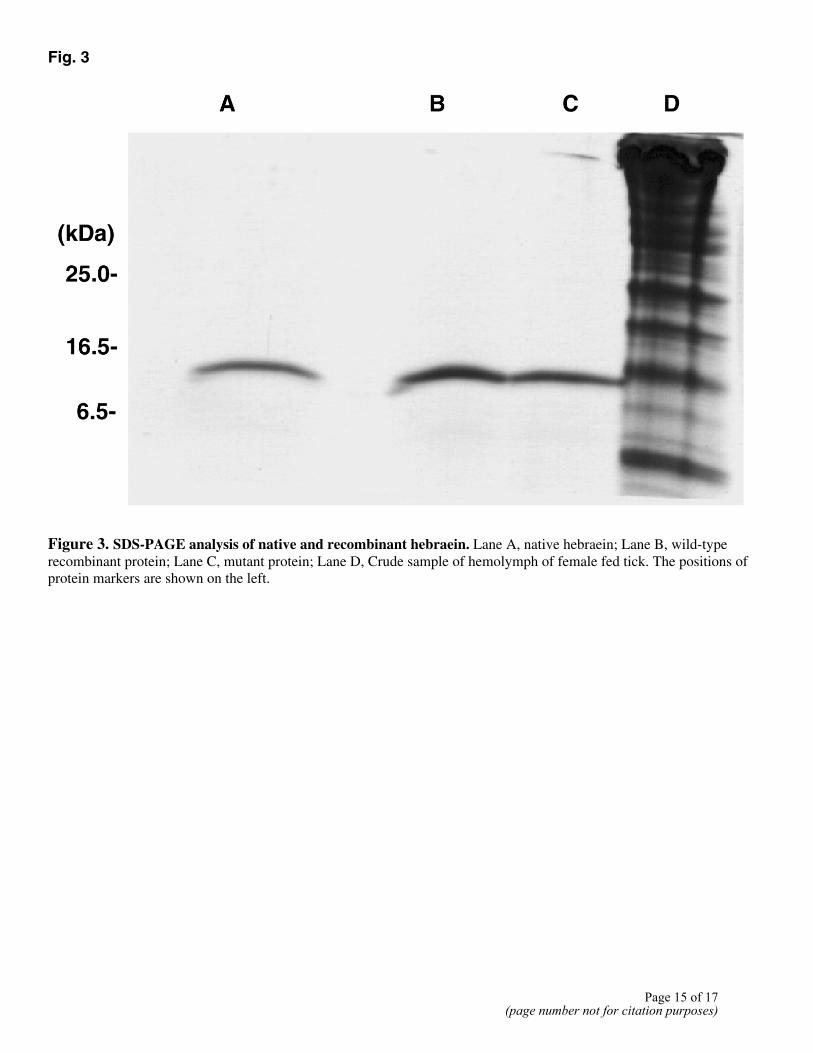
Fig. 3
Figure 3. SDS-PAGE analysis of native and recombinant hebraein. Lane A, native hebraein; Lane B, wild-type recombinant protein; Lane C, mutant protein; Lane D, Crude sample of hemolymph of female fed tick. The positions of protein markers are shown on the left.
Page 15 of 17(page number not for citation purposes)
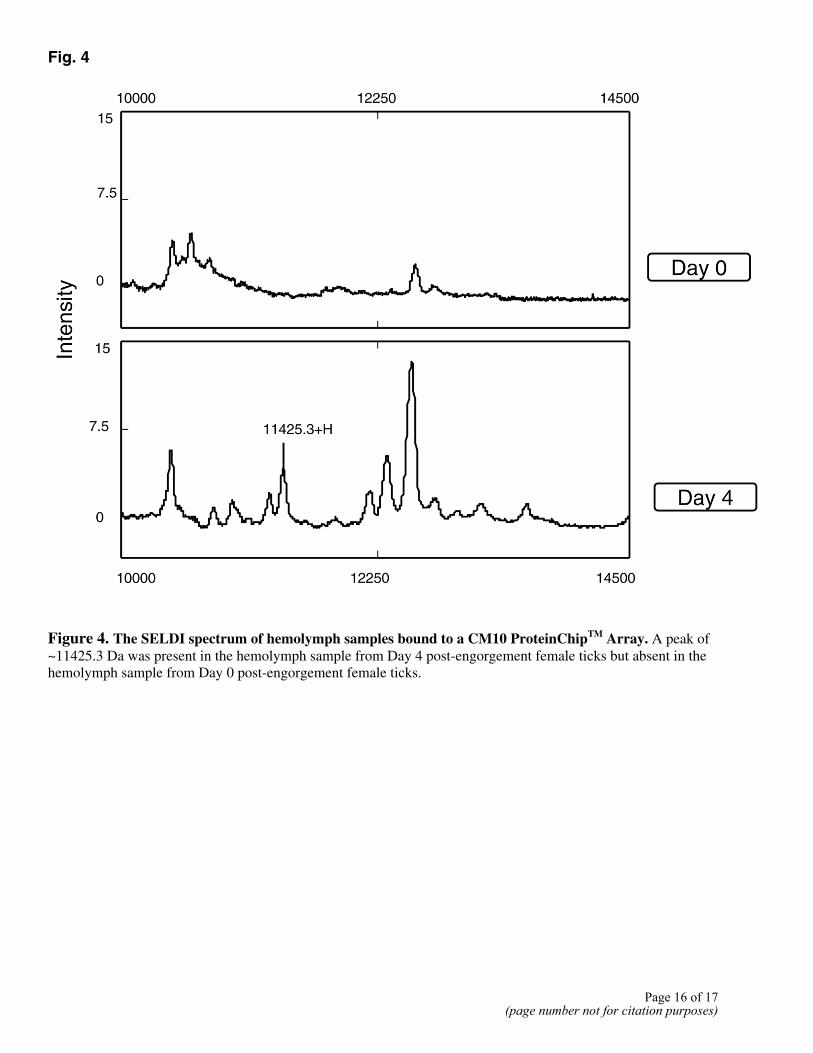
Fig. 4
Figure 4. The SELDI spectrum of hemolymph samples bound to a CM10 ProteinChipTM Array. A peak of ~11425.3 Da was present in the hemolymph sample from Day 4 post-engorgement female ticks but absent in the hemolymph sample from Day 0 post-engorgement female ticks.
Page 16 of 17(page number not for citation purposes)
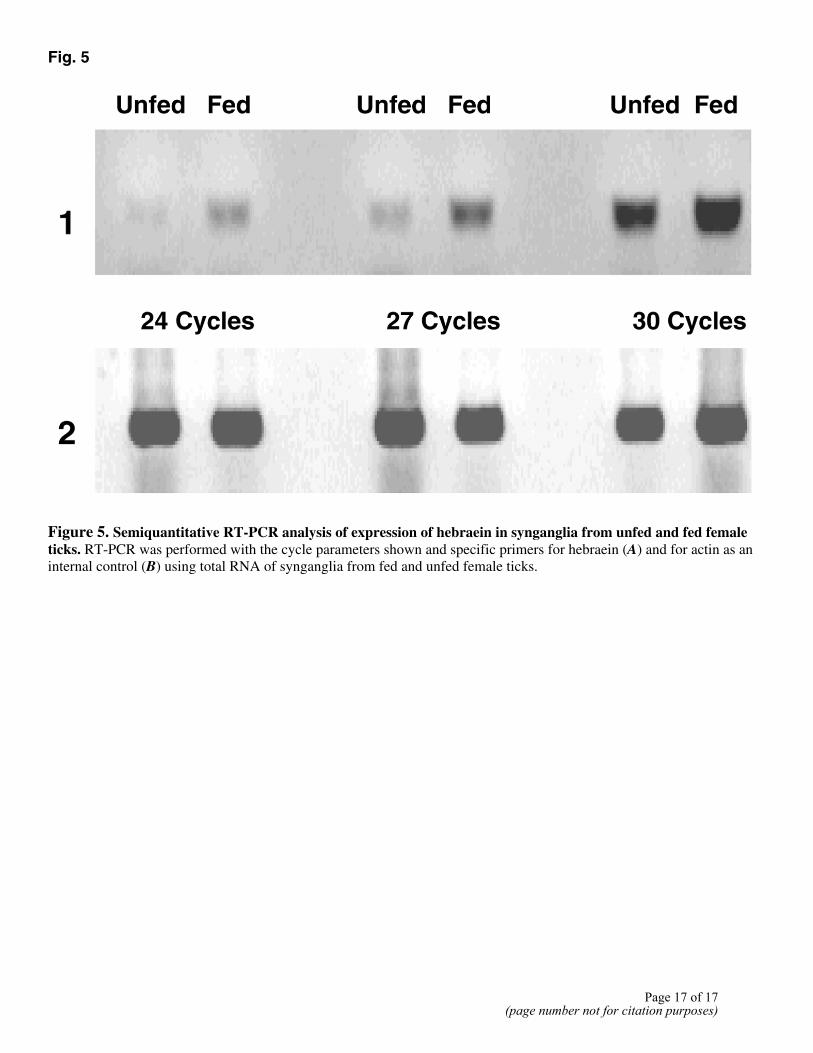
Fig. 5
Figure 5. Semiquantitative RT-PCR analysis of expression of hebraein in synganglia from unfed and fed female ticks. RT-PCR was performed with the cycle parameters shown and specific primers for hebraein (A) and for actin as an internal control (B) using total RNA of synganglia from fed and unfed female ticks.
Page 17 of 17(page number not for citation purposes)
Related Documents











