Botanical Journal ar the Linnean Society ; 2003, 142, 199-212. With 32 figures A new late Westphalian fossil marattialean fern from Nova Scotia JOSEF PŠENIČKA\ JIŘÍ BEK 2 , ERWIN L. ZODROW 3 , CHRISTOPHER J. CLEAU* FLS and ALAN R. HEMSLEy5 FLS I 'Deportment of Palaeontology, West-Bohemian Museum in Pilsen, Kopeckehosady 2, 301 36 Pilsen, Czech Republic 2Institute of Geology, Czech Academy of Sciences, Rozvojova 135, 16500 Prague 6, Czech Republic 3Department of Earth Sciences, University College ofCape Breton, Sydney, Nova Scotia, Canada B1P 6L2 "Department of Biodiversity and Systematic Biology, National Museums and Galleries of Wales, Cardiff, CF10 3Np, UK 'Laboratory for Experimental Palynology, Department of Earth Sciences, Cardiff University, . Cardiff CF10 3YE, UK Reeeived August 2002; aeeepted for publieation January 2003 Sydneia manleyi gen. et sp. nov. is based on part of a fertile frond from the upper Westphalian D of the Sydney Coalfield, Nova Scotia, Canada. It has small synangia composed of laterally fused sporangia that are elongate and with a circular cross-section. The sporangia yielded variably sized monolete and trilete spores with laevigate and microspinate ornamentation; intermediate forms were also observed. The spores can be correlated with the sporae dispersae species Latosporites minutus, Punetatosporites oeulus and Laevigatosporites minimus. Size distribution of the spores is variable and highly skewed, suggesting heterogeneity ofthe spore s within the sporangium. Spore ultra- structure indicates that the fossil is part of a fern, and the morphology of the spores and synangia indicate marat- tialean affinities. © 2003 The Linnean Society ofLondon, Botanieal Journal ofthe Linnean Soeiety, 2003,142,199- 212. ADDITIONAL KEYWORDS: Canada - Pennsylvanian - sporangia - spores. INTRODUCTION I Ferns with small, deeply incised pinnules were wide- spread in Late Carboniferous (Pennsylvanian) fioras of Europe and North America. Most belonged to the extinct order Urnatopteridales, and their fertile fronds are now referred to morphogenera such as Zeil- leria Kidst., 1884 and Urnatopteris Kldst., 1884. These eusporangiate ferns typically had pinnules with spo- rangia attached either singly or in loose clusters ('synangia'). Little is known about the habit of these plants but it is generally assumed that they were herbaceous. *Corresponding author. E-mail: [email protected] Another abundant group of eusporangiate ferns in these floras belong to the Marattiales. These were mostly tree-ferns and usually had linguaeform, pecopteroid pinnules, with the sporangia arranged in discrete synangia on their abaxial side. The synangia have an essentially radial symmetry in plan view, which contrasts with the oval to biseriate arrange- ment in most extant marattialeans (Hill & Camus, 1986). Much rarer were ferns with deeply incised pinnules, where the sporangia are arranged in an oval configuration more similar to that seen in synangia in extant marattialeans (Taylor, 1967). Zod- row et al. (2001) gave a preliminary report of such a fern found recently in the Westphalian D deposits of the Sydney Coalfield of Canada (Fig. 1) and the present paper gives a full description and systematic analysis. © 2003 The Linnean Society of London, Botanical Journal ar the Linnean. Society, 2003, 142, 199-212 199

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Botanical Journal ar the Linnean Society ; 2003, 142, 199-212. With 32 figures
A new late Westphalian fossil marattialean fern from NovaScotia
JOSEF PŠENIČKA\ JIŘÍ BEK2, ERWIN L. ZODROW3, CHRISTOPHER J. CLEAU* FLS andALAN R. HEMSLEy5 FLS
I 'Deportment of Palaeontology, West-Bohemian Museum in Pilsen, Kopeckehosady 2, 301 36 Pilsen,Czech Republic2Institute of Geology, Czech Academy of Sciences, Rozvojova 135, 16500 Prague 6, Czech Republic3Department of Earth Sciences, University College ofCape Breton, Sydney, Nova Scotia, Canada B1P 6L2"Department of Biodiversity and Systematic Biology, National Museums and Galleries of Wales,Cardiff, CF10 3Np, UK'Laboratory for Experimental Palynology, Department of Earth Sciences, Cardiff University,
. Cardiff CF10 3YE, UK
Reeeived August 2002; aeeepted for publieation January 2003
Sydneia manleyi gen. et sp. nov. is based on part of a fertile frond from the upper Westphalian D of the SydneyCoalfield, Nova Scotia, Canada. It has small synangia composed of laterally fused sporangia that are elongate andwith a circular cross-section. The sporangia yielded variably sized monolete and trilete spores with laevigate andmicrospinate ornamentation; intermediate forms were also observed. The spores can be correlated with the sporaedispersae species Latosporites minutus, Punetatosporites oeulus and Laevigatosporites minimus. Size distribution ofthe spores is variable and highly skewed, suggesting heterogeneity ofthe spore s within the sporangium. Spore ultra-structure indicates that the fossil is part of a fern, and the morphology of the spores and synangia indicate marat-tialean affinities. © 2003 The Linnean Society ofLondon, Botanieal Journal ofthe Linnean Soeiety, 2003,142,199-212.
ADDITIONAL KEYWORDS: Canada - Pennsylvanian - sporangia - spores.
INTRODUCTION
IFerns with small, deeply incised pinnules were wide-spread in Late Carboniferous (Pennsylvanian) fiorasof Europe and North America. Most belonged to theextinct order Urnatopteridales, and their fertilefronds are now referred to morphogenera such as Zeil-leria Kidst., 1884 and Urnatopteris Kldst., 1884. Theseeusporangiate ferns typically had pinnules with spo-rangia attached either singly or in loose clusters('synangia'). Little is known about the habit of theseplants but it is generally assumed that they wereherbaceous.
*Corresponding author. E-mail: [email protected]
Another abundant group of eusporangiate ferns inthese floras belong to the Marattiales. These weremostly tree-ferns and usually had linguaeform,pecopteroid pinnules, with the sporangia arranged indiscrete synangia on their abaxial side. The synangiahave an essentially radial symmetry in plan view,which contrasts with the oval to biseriate arrange-ment in most extant marattialeans (Hill & Camus,1986). Much rarer were ferns with deeply incisedpinnules, where the sporangia are arranged in anoval configuration more similar to that seen insynangia in extant marattialeans (Taylor, 1967). Zod-row et al. (2001) gave a preliminary report of such afern found recently in the Westphalian D deposits ofthe Sydney Coalfield of Canada (Fig. 1) and thepresent paper gives a full description and systematicanalysis.
© 2003 The Linnean Society of London, Botanical Journal ar the Linnean. Society, 2003, 142, 199-212 199

200 J. PŠENIČKA ET AL.
70'
Ir'
j/''-:; 7
-:
QUEBEC
Maritimes8asin
/'I:. lIos do laMadeleine
NOVA? t. SCOTIA
'I. f'.o •
c.ajajID
fraiOI
si50 TJ
r<l
"
I60"
""",10 Bay Bas;n
..'
>jni~fBay Sl George
Sub-8aslnQ
./ _,rJř./'
Co.J:..i.1
I"='] Onshore Mar~imes Basin
11111111 Discrete pre-Carboniferous uplift
oI
200J
Approximate offshore extent
/" Trnce of major fault
100I
KJlcrnetres
7
Figure 1. Location map of Sydney Coalfield (Sydney Sub-Basin) within the Carboniferous Maritimes Basin of Canada.
MATERIAL AND METHODS
The fossils investigated were collected from a 0.5 mthick mudstone above the Stubbart Seam, PrinceMine, Cape Breton, Nova Scotia, Canada. The mud-stone is rich in micro- and macrotioras (Zodrow, 1986)that are well preserved and include cuticles. Synangiaand cuticles were separated from the rock matrixusing 35% hydrotiuoric acid for 24 h. Cuticles weregently bleached in Schulze's solution (nitric acid withpotassium chlorate) for 3-30 min, and then drainedand washed in water. They were then percolated with10% potassium hydroxide solution to remove oxidationproducts, and finally washed again in water. To avoidcontamination, distilled water was used throughoutthe process. Cuticles, and some sporangia and theirspores, were mounted in glycerine jelly and examinedunder the light microscope using brightfield andNomarski illumination.
Some spores and whole sporangia were mounted onstubs and gold-coated for examination under a CAM-ECA SX100 scanning electron microscope (SEM).
Spores were also sectioned for study under TEM, usingtechniques described by Hemsley & Glasspool (1999).
All study specimens, including slide mounts of spo-rangia, spores, cuticles and SEM stubs, are perma-nently housed at the University College of CapeBreton, Sydney, Nova Scotia (Accession numbers989GF-271-272 and 989GF-274-277).
DESCRIPTION
The five pieces of mudstone that originally fittedtogether to form a slab with a surface area of450 x 300 mm preserve one contiguous frond segmentas a coalified compression. A thin covering of second-ary gypsum crystals obscures parts of the plant but itwas not removed because of the potential risk of dam-aging the delicate fructifications. The followingdescription assumes that the fossil is part of a frondand use s standard terminology for the description offern foliage (Tryon, 1960).
Two of the fragments (989GF-275, 989GF-276)together preserve a bipinnate fertile structure
Figradrac
© 2003 The Linnean Society of London, Batanical Journal ar the Linnean Society, 2003, 142, 199-212 © 2{

lanada.
, us ing(1999).
, of spa-perma-
)f Capeumbers
, fittedarea ofegmentsecand-t but itofdam-llawinga frondotion of
NF-276)I ucture
[99-212
NEW LATE WESTPHALIAN MARATTIALEAN FERN 201
C. 130 mm long and c. 50 mm wide (Fig. 2). Thisappears to have been attached to a lower order rachis,although their exact relationship cannot now be deter-mined due to unavoidable damage during collectingfrom the roof rocks. The penultimate rachis is hairyand tapers in width from 12 mm to 7 mm. Parts offiveor possibly six ultimate pinnae are preserved on oneside of this penultimate rachis, spaced 26 mm apart.These ultimate pinnae are 50-60 mm long and have arachis usually less than 2 mm wide. The pinnae are
attached at 30-40° to the main axis, but then archstrongly backwards, probably due to distortion duringfossilization (Fig. 3). Fragments of pinnae are alsopresent on the other side ofthe penultimate rachis butnone are attached.
The ultimate rachises are longitudinally fine1y stri-ate and bear alternately arranged pinnules, attachedat 60-80°. Adjacent pinnules are spaced 4-5 mmapart, and are 13-15 mm long and 5-7 mm wide. Theyhave a considerably reduced lamina, giving the pin-
Figures 2-4. Sydneia manleyi gen. et sp. nov. (holotype). Fig. 2. Whole specimen showing the characteristic wide mainrachis with pinnae attached on the dextral side. Scale bar = 20 mm. Fig. 3. Close-up of arched pinna attached to mainrachis. Scale bar = 5 mm. Fig. 4. Close-up of fertile pinnule. Scale bar = 2.5 mm.
© 2003 The Linnean Society of London, Botanical Journal ar the Linnean Society, 2003, 142, 199-212
202 J. PŠENIČKA ET AL.
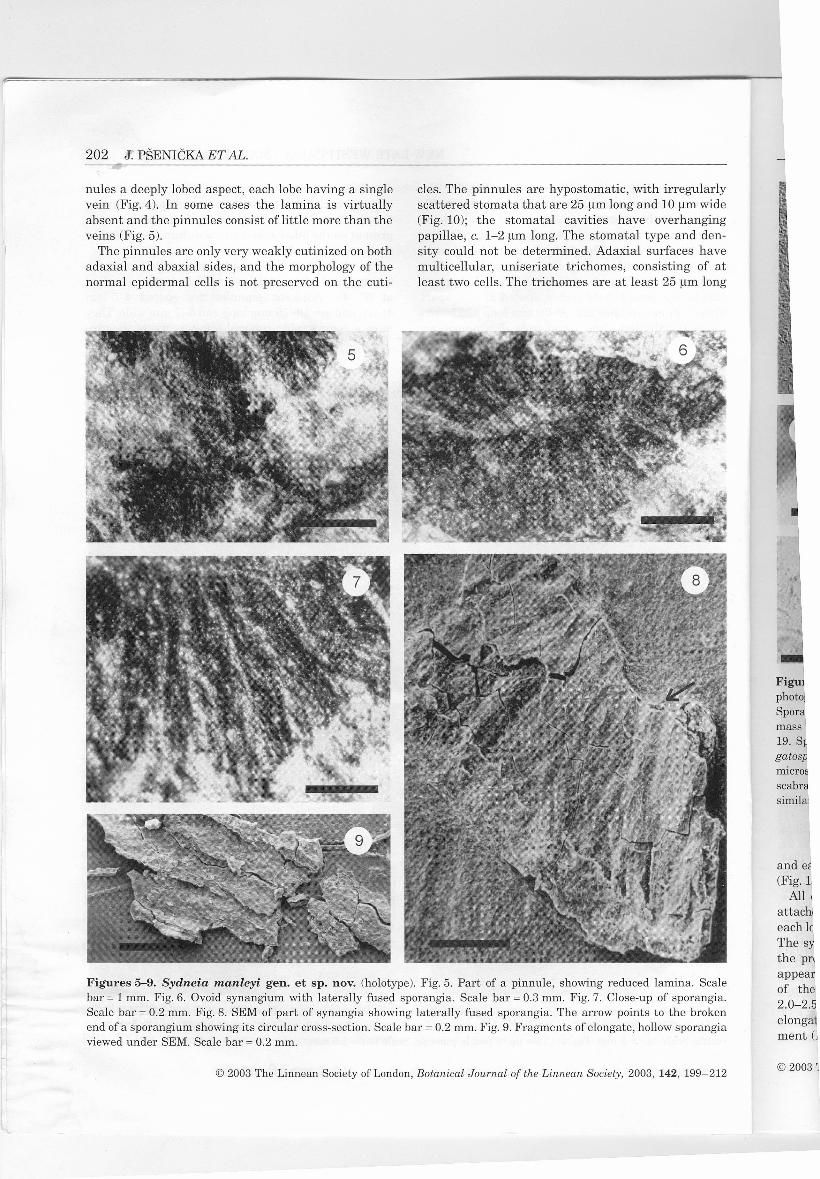
nule s a deeply lobed aspect, each lobe having a singlevein (Fig. 4). In some cases the lamina is virtuallyabsent and the pinnules consist oflittle more than theveins (Fig. 5).
The pinnules are only very weakly cutinized on bothadaxial and abaxial sides, and the morphology of thenormal epidermal cells is not preserved on the cuti-
cles. The pinnules are hypostomatic, with irregularlyscattered stomata that are 25 um long and 10 um wide(Fig. 10); the stomatal cavities have overhangingpapillae, c. 1-2 um long. The stomatal type and den-sity could not be determined. Adaxial surfaces havemulticellular, uniseriate trichomes, consisting of atleast two cells. The trichomes are at least 25 um long
Figures 5-9. Sydneia manleyi gen. et sp. nov. (holotype). Fig. 5. Part of a pinnule, showing reduced lamina. Scalebar = 1 mm. Fig.6. Ovoid synangium with laterally fused sporangia. Scale bar = 0.3 mm. Fig. 7. Close-up of sporangia.Scale bar = 0.2 mm. Fig. 8. SEM of part of synangia showing laterally fused sporangia. The arrow points to the brokenend of a sporangium showing its circular cross-section. Scale bar = 0.2 mm. Fig. 9. Fragments of elongate, hollow sporangiaviewed under SEM. Scale bar = 0.2 mm.
© 2003 The Linnean Society of London, Botanical Journal of the Linnean Society, 2003, 142, 199-212
mass19. S~gatosAmicrosscabrsimilal
and e~(Fig. 1
Allattaclieach 10The sythe prappearof the2.0-2.5elongament (l
© 20031

·rregularly10 um wideerhanging
e and den-lfaces have,ting of at25 um long
ina. Scalesporangia.he brokensporangia
2,199-212
NEW LATE WESTPHALIAN MARATTIALEAN FERN 203
Figures 10-19. Sydneia manleyigen. et sp. nov. (holotype). Fig. 10. Abaxial cuticle with three stomata (arrowed),photographed under Nomarski illumination. The storna on the left ofthe picture shows papillae. Scale bar = 50 um. Fig. 11.Sporangium after maceration, showing elongate epidermal cells in its distal part. Scale bar = 100 um, Fig. 12. Part of sporemass macerated from sporangium. Scale bar = 100 um. Fig. 13. Base of epidermal trichome. Scale bar = 50 um, Figs 14-19. Spores obtained from sporangia, viewed using transmitted light microscopy. Scale bars = 20 um. Figs 14,15 Laevi-gatosporites-type spores with laevigate exine. Fig. 16 Latosporites-type spore. Fig. 17 intermediate-type spore with finelymicrospinate to micogranulate exine, similar to Punctatosporites punctatus. Fig. 18 Intermediate-type spores with finelyscabrate exine, similar to Punctatosporites minutus. Fig. 19 Thick-walled, densely microspinate to microgranulate sporessimilar to Punctatosporites oculus.
and each has a thickened base 20-25 um in diameter(Fig. 13).
All of the pinnules are fertile, with a synangiumattached to the abaxial surface, near the distal end ofeach lobe (Fig. 4); there are c. 20 synangia per pinnule.The synangia are normally borne singly. However, inthe proximal parts of the larger pinnules they mayappear to be in pairs or even threes, due to forkingof the pinnule lobes (Fig. 4). The synangia are2.0-2.5 mm long and 1 mm wide, and consist of 20-30elongate sporangia in a simple oval-shaped arrange-ment (Fig. 6). Adjacent sporangia appear to be later-
ally fused along their entire length (Figs 7-9), givingthe synangium an open cup-shaped form, with a cen-tral cavity.
Each sporangium is 0.5-0.8 mm long and 0.3 mmwide (Fig. 11), with a circular cross-section (Fig.8),and an obtuse apex. The sporangia are exannulatewith a wall composed of longitudinal, polygonal cells150-200 um long and 20-30 um wide (Fig. 11).
The sporangia contain dense masses of ovoid tospherical spores (Figs 12, 14-25). Spore size rangesfrom 16 um to 39 um (Table 1), with a bimodal distri-bution (peaks at 25.0-26.9 um and 29.0-30.9 um).
© 2003 The Linnean Societyof London,Botanical Journal of the Linnean Society, 2003, 142, 199-212

~~---------------------------------------------------------------------------------------------- ..--~--~
Figures 20-25. Sydneia manleyi gen. et sp, nov. (holotype). Spores obtained from sporangia, viewed under SEM.Fig. 20. Circular spore with laevigate exine, similar to Latosporites minutus Bharadwaj, 1957. Scale bar = 5 um, Fig.21.Oval spore with laevigate exine, similar to Laeuigatosporites minimus (Wilson & Coe) Schopf, Wilson & Bentall, 1944.Scale bar = 5 um. Fig. 22. lrregularly scabrate exine of the Laeuigatosporites type. Scale bar = 10 um. Fig. 23. Spore withmicrospinate exine, similar to Punctatosporites occulus Smith & Butterworth, 1967. Scale bar = 5 um. Figs 24,25. Close-ups of exine with microspinate ornamentation. Scale bars = 5 um.
ab1'lltcZLm
204 J. PŠENIČKA ET AL.
Table 1. Morphometrics of the spores of Sydneia manleyi gen. et sp. nov
Palynomorph species Type Size um Minimum Maximum Average No. of spore s
Latosporites minutus laevigate 16.0 32.6 27.3 126Laeuigatosporites minimus laevigate 20.0 34.0Punctatosporites oculus microspinate 19.0 38.4 29.6 74
irtcww
tc(I
mh:(litaa
This bimodal distribution correlates with two morpho-logical types of spore.1. Monolete spores, with a <111mthick laevigate exinethat has several secondary folds (Figs 20,21). Thelaesurae extend for 1/2 to 3/4 of the spore radius. Thespores range in size from 16 11mto 33 11mwith an aver-age of 27.3 11m(Table 1) and often occur in clusters.
The circular forms correspond to the dispersed mor-phospecies Latosporites minutus Bharadwaj(Figs 16,20), and the oval forms to Laeuigatosporitesminimus (Wilson & Coe) Schopf, Wilson & Bentall(Figs 14,15,21).2. Trilete spores, with a 2-5 11mthick microspinateexine (Figs 23-25). Two rays of laesurae extend for
Fispba
frcaba
© 2003 The Linnean Society of London, Botanical Journal of the Linnean Society, 2003, 142, 199-212 © :

I. of spores
6
ider SEM.m. Fig.21..tall, 1944.Spore with,25.Close-
sed mor-haradwajosporites
(, Bentall
ospinatextend for
2,199-212
NEW LATE WESTPHALIAN MARATTIALEAN FERN 205
about 2/3 of the spore radius, sometimes accompaniedby a third short ray. The spores range in size from19 um to 39 um with an average of 29.6 um (Table 1).These spores never occur in clusters. They correspondto the dispersed morphospecies Punctatosporites ocu-lus Smith & Butterworth, which can include bothmonolete and trilete spores (Figs 19,23-25).
There are also intermediate spores 20 um to 34 umin size, that have an irregularly scabrate exine 1.5 umto 3 um thick (Fig. 22). Some of them are monoletewith laesurae extending 1/2 to 3/4 ofthe spore radius,whilst others are trilete with a short third ray, simil arto spores of the Punctatosporites oculus-type(Figs 17,18).
Under SEM, the small spores appear unorna-mented except for a coarse foveolate system on whathas been presumed to be the distal surface(Figs 20,21), which may be an effect caused bytaphonomy or maceration. They are surrounded byabundant tapetal material in the outer part of the
sporangial mass but are well separated within; afeature that could give rise to variation in thedegree of sculpturing shown by a population ofmature spores. In TEM section, the walls are 0.5 umto 1.0 um thick (Figs 27,28,31) and show no thicken-ing towards the presumed aperture. Folds visible inthe equatorial region are probably the result of com-pression of the spore (Fig. 29). Some sections showwhat may be a distinct, thin, 80-100 nm inner layer(Fig.30). The outer surface is finely irregular butdoes not reveal great depth to the minimal sculptur-ing observed. In some cases, the darker tapetalmaterial can be seen adherent to the outer wall sur-face (Fig. 26). Internally, the wall shows a more orless even texture (Figs 27-29,31) but there are someindications of a possible, compressed, coarse granu-larity with individual components reaching around0.1 11m (Fig.30). However, both the internal layerand the possible particulate nature could be arte-facts of preparation.
28
31
Figures 26-31. Sydneia manleyi gen. et sp. nov. (holotype).TEM sections through spore wall. Fig. 26. Amb and distalspore wall, demonstrating compressed lumen and wall homogeneity. Arrow indicates possible tapetal material. Scalebar = 750 nm. Fig. 27. Detail of isolated distal wall. Outer surface is uppermost. Scale bar = 400 nm. Fig. 28. Detail of ambwall showing scuff marks but no apparent internal structure. Scale bar = 400 nm. Fig. 29. Equatorial fold of spore resultingfrom compression. The markings present are artefacts of cutting. Scale bar = 400 nm. Fig. 30. High magnification detailacross a separated lumen showing possible inner layer. Scale bar = 200 nm. Fig.31. Detail of a distal wall. Scalebar = 315 nm.
© 2003The Linnean Societyof London,Botanical Jaurnal ar the Linnean Society, 2003, 142, 199-212

206 J. PŠENIČKA ET AL.
AFFINITIES OF IN SITU SPORE S
CORRELATIO WITH DISPERSEDSPORE TAXASma11 monolete dispersed spores with laevigate ormicrospinate ornamentation such as described hereare genera11y referred to as the Punctatosporites-Lae-vigatosporites-type. It is very rare for microspinate tomicrogranulate spore s of the Punctatosporites-typenot to be found in association with laevigate spores ofthe Laevigatosporites-Latosporites-type. The smallerlaevigate types of spores from the presently investi-gated specimens belong to the dispersed morphospe-cies Laevigatosporites perminutus Alpern andLatosporites minutus Bharadwaj (Bek & Opluštil,1998; Zodrow, Bek & Pšenička, 2000). These two mor-photaxa differ only in shape (circular to oval) which isinfl.uenced by diagenesis. The sma11er of the sculp-tured spores from the present specimen can becompared with Latosporites globosus Schemel, Punc-tatosporites minutus Ibrahim, or P granifer RPotonié& Kremp, depending on their size (Bek, 1998; Zodrowet al., 2000). Significantly, however, a11 similarmicrospinate spores have a thinner exine (usua11y<2 um), or are sma11er.
There is a continuous morphological gradationbetween the laevigate, thin-wa11ed spores, and themonolete and trilete microspinate thick-walled spores.The gradation is evident by the several intermediateforms, including sma11, relatively thin-wa11ed (c. 1 um)scabrate to finely microgranulate spores, and larger,more densely microgranulate to microspinate formswith a thicker (2-3 um) exine. These may be corre-lated with various dispersed spores of the Punc-tatosporites species, according to their size andsculpture. Sma11er scabrate to finely microspinateforms correspond with P minutus Ibrahim, whilstlarger finely microspinate spores resembleP granulatus Bharadwaj, P scabellus (Imgrund)R Potonié & Kremp, and P punctatus (Kosanke)R Potonié & Kremp.
We have included both monolete and trilete sporeswithin the same dispersed genus PunctatosporitesIbrahim. Dispersed species of this genus are variablein size, shape, sculpture and also in the type of tetradmark; most of species are monolete, but some of thempossess a trilete mark, always with short third ray.
SPORE ULTRASTRUCTUREThe range of Palaeozoic fern spores studied by TEM isdominated by botryopterids and scolecopterids (e.g.Millay & Taylor, 1982, 1984). These generally have rel-atively thin walls, which may be homogeneous ininternal structure or demonstrate some simple layer-ing. Millay & Taylor (1982) believed that this horno-geneity was a real feature and not one derived through
diagenesis. Observations on Mississipian spores byHemsley, Clayton & Galtier (1994) support this view.Also apparent in these early fern spores is the poor dif-ferentiation oflayers associated with the aperture andthe tendency for the retention of tapetal material asan outer (perisporal?) coating. Extant eusporangiatefern spore s have a generally more complex internalultrastucture (see Tryon & Lugardon, 1990).
In contrast, most pteridosperm pollen grains exam-ined by TEM demonstrate a distinct layering withinthe wall structure. A homogeneous internal layer ispresent as one quarter or less of the total wa11 thick-ness; this may reach 10 um or more, where sculptureis present. The outer part of the exine consists of whathas been termed an alveolate region, which is com-posed of a spongy material derived from an intercon-nected network or system of globules (Taylor, 1982).Much work has concentrated on po11en assumed to bemedullosalean (e.g. Taylor & Rothwe11, 1982; Kur-mann & Taylor, 1984) but subsequent studies on othergroups (e.g. Osborne & Taylor, 1994) seem to confirmthis general trend.
Although it is possible that such an outer layerstructure may be lost through diagenesis or prepara-tion, it seems improbable that a11indications of layer-ing would disappear. If compression was responsiblefor the loss of structure, we would expect to see athicker exine and, if the outer alveolar layers were tohave been removed, the preservation of the observedtapetal material would be inconsistent. Based onthese observations and the fundamental differencesbetween early fern spore ultrastructure and that ofpteridosperm pollens, it seems reasonable to concludethat the spores under study are indeed fern spores.
COMPARISONS
Sma11, laevigate, scabrate, microspinate or microgran-ulate spores with a monolete or trilete mark, similarto those extracted from the present specimens, areknown from various Carboniferous fertile fronds(Balme, 1995; Bek, 1998). It is impossible at this junc-ture to distinguish palynologically between thoseplants that produced the sma11er type of laevigatespores, but those that produced the larger ornamentedspores can sometimes be differentiated on that basis.Where possible, the following comparisons will there-fore ta ke into account palynological as we11as morpho-logical characters (Table 2).
RADSTOCKlA KrDST., 1923
This morphogenus was established by Kidston toaccommodate two species. The type had origina11ybeen referred to as Schizostachys sphenopteroidesKidst., 1888 but, as pointed out by Potonié (1892b),
© 2003 The Linnean Society of London, Botanical Journal af the Linnean Society, 2003, 142, 199-212
Tabll
Specir
SaaricSaaricSaaricRadstiSydnei
this sj
ston (schlaghe didvalid IschlagiH. bey:as eaclsporanhas sinsee als
Thephologiwhich Iston oattacherangia,ing strsubseqistructuiresentiI('line ar'1983).R. spherinterprein the Imakes i'type tosynangi:detailedCanadia
BrousRadstoctWestphamaterialthe spor:four, witlsynangitan anonreportedclearly SE
miche (I!
© 2003 Th

d
this species has nothing to do with Schizostachys. Kid-ston (1923) also suggested that Hymenotheca bey-schlagii Potonié (1892a) should be included, althoughhe did not actually use the combination (the earliestvalid publication of the combination Radstockia bey-schlagii appears to have been by Remy (1953)). In fact,H. beyschlagii differs fundamentally from Radstockia,as each reproductive body consists of a single largesporangium rather than a compound synangium, andhas since been transferred to Saaria by Remy (1953;see also comments below).
The present specimens show some superficial mor-phological similarities with the type of Radstockia,which has similar deeply incised fertile pinnules. Kid-ston originally interpreted the fertile structuresattached to the lobes of these pinnules as single spo-rangia, with transverse markings possibly represent-ing structures to aid dehiscence. However, mostsubsequent authors have instead interpreted thesestructures as synangia, the trans verse markings rep-resenting sporangia which are arranged in an oval('linear') configuration (Taylor, 1967; Brousmiche,1983). Having re-examined the holotype ofR. sphenopteroides, we tend to agree with this revisedinterpretation. However, the high rank of coalificationin the Radstock Coalfield (Kellaway & Welch, 1993)makes it impossible to macerate the Radstockia holo-type to obtain spores to determine the detail s of thesynangial structure. It is impossible therefore to give adetailed comparison between Radstockia and the newCanadian specimens.
Brousmiche (1983) assigned additional specimens toRadstockia sphenopteroides, this time from the upperWestphalian D of Saar-Lorraine. Based on this newmaterial, Brousmiche (1983; text-fig. 32) reconstructedthe sporangia ofRadstockia as occurring in clusters offour, with the clusters arranged in two rows to form asynangium. Although it is not clearly stated in the text,an anonymous referee of the present paper hasreported to us that the groups of four sporangia wereclearly seen by Brousmiche during maceration. Brous-miche (1983) also reported that her specimens yielded
NEW LATE WESTPHALIAN MARATTIALEAN FERN 207
y Table 2. Comparison of fructification morphologyv.
SynangiumShape and number of
Species Length (mm) Width (mm) sporangium/synangium
Saaria beyschlagii 5.5 2.0 ?Saaria weissii 3.0 1.0 falcate, elongate 112Saaria allosuroides 4.0 1.0Radstockia sphenopteroides 2.5 ? ?Sydneia manleyi 2.0-2.5 1.0 elongate 120-30
se11
l-
ns
Sporangium
Length (mm) Width (mm)
????0.5-0.8
????0.3
~-e.t1-
).
e
rli
r
eaodli
sIfe
1-
res
eed
oys),
exclusively trilete Punctatisporites-like spores with ascabrate or microgranulate-microverrucate ornamen-tation. However, as we cannot know the synangialstructure or spores of true Radstockia, it is impossibleto assign these Saar-Lorraine specimens to Radstockiawith any confidence. As pointed out by Brousmiche(1983; 130), her specimens have many features in com-mon with another Late Carboniferous fern, Crossoth-eca, in particular the synangia consisting of two rowsof elongate sporangia grouped in clusters of four.Although there are minor differences in the exine orna-mentation in the spores, it seems likely that the Brous-miche specimen in fact belongs to Crossotheca. Theyare clearly different from the Canadian specimens, inboth sporangial grouping within the synangia and themorphology of the spores that they contain.
A second species that has been referred to Radstockia(R. kidstonii Taylor, 1967) was based on a type from theMazon Creek tlora oflllinois, USA. The relatively shortoval synangia situated towards the margins of the fer-tile pinnules are of simil ar shape and configuration tothe presently studied synangia, but their relativelypoorer preservation makes it impossible to be certainthat the detailed configuration and morphology of thesporangia are the same. Unlike the Canadian speci-mens, Taylor (1967) interpreted the Mazon Creek spec-imen as having the synangia partly embedded withinthe pinnule limb, but it is difficult to see from his illus-trations on what evidence this was based. A more obvi-ous difference is that the fertile pinnules in the MazonCreek specimen have a much more developed limb oneither side of the veins. Also, Taylor described thespore s from here as being laevigate and larger(40-60 um), which is outside the range of the presentspore s (Table 1). He was uncertain as to whether theywere monolete or trilete, although the spore that he fig-ured on his plate 7 figure 5 is clearly trilete.
SAARIA REMY, 1953Remy established this morphogenus for two speciesincluded within Hymenotheca by Potonié (1892a):
2 © 2003The Linnean SocietyofLondon,Botanical Jaurnal ar the Linnean Saciety, 2003, 142, 199-212

208 J. PŠENIČKA ET AL.
H. beyschlagii Potonié and Hymenophyllum uieissiiSchimp., 1869. He also included Sphenopteris allo-suroides Gutbier, 1835, which has similarly shapedsynangia. Remy noted that these species ha ve repro-ductive structures with longitudinal furrows andtransverse marks, which are absent in the type spe-cies of Hymenotheca (H. dathei Potonié, 1892a). Kid-ston (1923) had made similar observations andsuggested that H. beyschlagii should be includedwithin his new morphogenus, Radstockia. However,Remy argued that Saaria differs from Radstockia inits frond architecture, and in having winged rachisesand stalked synangia. Furthermore, according toRemy, the Saaria synangia consist of elongate clustersof sporangia that are marginally attached and extendout from the ends of the pinnule lobes, in contrast tothe pendant synangia of Radstockia.
There is a superficial similarity between Saaria andthe present specimens, in that both consist of deeplydivided fertile pinnules, with synangia attached nearthe distal ends of the pinnule lobes. However, thesynangia in the present specimens are clearly pendanton the pinnule's abaxial surface, not marginallyattached as in Saaria. Furthermore, the Saariasynangia are significantly longer than wide and con-sist of about 12 sporangia, whereas the Canadiansynangia are less elongate and consist of 20-30 spo-rangia. It is difficult to compare the spores as Remy(1953) did not illustrate any detailed figures of themfrom Saaria. He described them as being laevigate,oval, 20 um long and 15 um wide, which would com-pare with the smaller types of spore found in theCanadian specimens. However, Remy was unable toobserve whether they were monolete or trilete, anddescribed only small thin-walled spores of the Laeui-gatosporites-Latosporites-types. No evidence wasfound for the larger types of ornamented spore, suchas found in our specimens.
ASTEROTHECOIDFERNSWe are referring here to the group of marattialeanferns with pecopteroid foliage that occurred abun-dantly in the late Pennsylvanian and earliest Permian.The difficult taxonomic and nomenclatural problemsthat surround these species have never been properlyresolved, especially as they relate to their genericassignment, and to the relationship between fossilspreserved as adpressions (coalification) and petrifac-tions (coal balls). Further discussion ofthese problemsis obviously beyond the scope of the present paper.
Synangia in the asterothecoid ferns compare withthe specimens described in the present paper in thatthey are pendant on the abaxial side of their fertilepinnules. Some also have comparable monolete sporeswith a fine surface ornamentation, such as the Minor
and Latifolia Groups of Scolecopteris (sensu Millay,1979), and Grandeuryella C.E.Weiss emend. Lesni-kowska & Galtier, 1992. The most obvious difference isin the gross morphology of the fertile pinnules, whichin all of these asterothecoid ferns is linguaeform withan unlobed margin ('pecopteroiď). Also, most tend tohave relatively small synangia of three to six (usuallyfour) sporangia in a more or less radial ('asterothecal')configuration. There are some cases where the synan-gia have more sporangia in an oval or bilateral ('lin-ear') configuration similar to that of the presentspecimens, such as Eoangiopteris Mamay, 1950 andGrandeuryella. However, the synangia in these mor-phogenera tend to have only a small central cavityseparating the component sporangia, whereas the spo-rangia in the present specimens appear to morewidely spreading and bell-like in configuration.
ZEILLERIA KrDST., 1884There is some similarity between the Canadian spec-imens and this morphogenus of Carboniferous ferns,which also has deeply incised pinnules bearing synan-gia that yield monolete spore s (Thomas & Crampton,1971). However, the Zeilleria synangia consist ofrather fewer sporangia (usually about five) and areattached to the tip s of the pinnules, rather than beingpendant (Kidston, 1923; Brousmiche, 1983). The tax-onomic position of Zeilleria has been a matter of somecontroversy, with Danzé (1956) placing it within hisextinct order Urnatopteridales, whereas Thomas &Crampton (1971) regarded it as a coenopteridaleanprogenitor of the Marattiales.
MEDULLOSALEANPOLLENORGANSThere is a superficial similarity in shape between thepresent synangia and certain of the complex pollen-bearing organs of the Medullosales, most notably thegenu s Whittleseya Newb. Whittleseya has a similarbell-shaped structure with a wide central cavity, andyields monolete palynomorphs. However, the Whittle-seya palynomorphs are monolete preopollen of theMonoletes-type (Taylor, 1973) with a layered ultra-structure (Millay & Taylor, 1976) that is quite differ-ent from that seen in the present specimens.Furthermore, there is no evidence that medullosaleanpollen-organs were ever attached to the pinnules, butwere instead attached directly to rachises of the frond(Stidd, 1981).
SYSTEMATICS
It is clear from the above comparisons that the Cana-dian specimens described in this paper cannot beaccommodated within any existing Pennsylvanian-
© 2003 The Linnean Society of London, Batanical Jaurnal af the Linneari Society, 2003, 142, 199-212

nsu Millay,end. Lesni-lifference isules, which.eform with.ost tend toiix (usuallyerothecal')the synan-ateral ('lin-e present1950 and
these mor-tral cavityas the spo-r to morebon.
dian spec-ous ferns,ing synan-rampton,
consist of,) and arehan being. The tax-er ofsomeithin his
'homas &;eridalean
tween the,x pollen-tably thea similarvity, and~Whittle-n of theed ultra-te differ-ecimens.lloealeanules, buthe frond
e Cana-nnot bevanian-
199-212
NEW LATE WESTPHALIAN MARATTIALEAN FERN 209
fructification morphogenus. Morphologically, theyshare some features with both pteridosperms andferns. In particular, the connate 'sporangia' arrangedin an oval synangiate configuration bear some resem-blance to the medullosalean Whittlesya-pollen organs.Our specimen was found exclusively associated withmedullosalean pinnules ofthe Linopteris obliqua type,although this species is normally regarded as havingborne Potoniea-pollen organs. The papillate stomata ofthe Canadian specimens also resemble certainmedullosaleans (e.g. Cleal & Shute, 1991, 1992; Zod-row & Cleal, 1998) but, as Palaeozoic fern cuticles arevery poorly known, the taxonomic significance of thischaracter is unclear. The monolete palynomorphs alsobear some resemblance to certain medullosalean pol-len (Millay & Taylor, 1976, 1979), although they arerather smaller than is typical.
However, the ultrastructure of the present palyno-morphs clearly indicates that they are pteridophyticspores rather than medullosalean pollen. The connatesporangia are rather unusual, but all other characterstend to suggest that these fossils are the remains of afern. The grouping of the exannulate sporangia intodistinct synangia and the monolete spores suggestthat it belongs to the Marattiales. The open bell-shaped synangia in which the sporangia are arrangedin an oval configuration are rather different fromthose normally associated with Pennsylvanian marat-tialeans (often referred to the family Asterothecaceaeor Psaroniaceae). In the latter, the synangia do nothave a significant central cavity and the sporangiatend to be in a circular ('asterothecal') configuration.
The nearest comparison with extant ferns is withMarattia Swartz, which has synangia with laterallyfused sporangia in an oval configuration, and whichproduces monolete spores (Hill & Camus, 1986;Camus, 1990). However, the Marattia synangia do nothave the same type of open cup-shaped configurationas the present specimens, and the pinnules have a farles s divided lamina. It is of course possible to derivethe Marattia-type of configuration from an ancestralform similar to that seen in our specimens (comparecomments by Taylor, 1967: 46), but we do not considerthat the evidence is conclusive. In our view, it is tax-onomically much safer to regard the Canadian speci-mens as being of an unknown marattialean family.The systematics of our new material may therefore besummarized as follows.
FlLICOPSIDA PlCHI -SERMOLLlMARATTlALES ENGLER & PRANTL
FAMlLY: UNKNOWNSYDNEIA PŠENlČKA ET AL. GEN. NOV.
Fertile frond segments with deeply incised ultimatepinnules. Each pinnule lobe has one pendant
synangium on abaxial surface. In the proximal part ofpinnae, synangia appear to be alternately in pairs, asa result of the bifurcation of the lateral veins of thelower pinnules. Synangia consist of connate sporangiain an oval configuration, with a wide central cavity .Sporangia elongate and exannulate, bearing bothmonolete and trilete, laevigate, scabrate tomicrospinate spores.
Type species: Sydneia manleyi sp. nov.
Etymology: Named after the city of Sydney, CapeBreton Island, Nova Scotia, Canada.
SYDNEIA MANLEYI PŠENlČKA ET AL. SP. NOV.(FlG.32)
Description: Synangia attached to club-shaped distalregion of pinnule lobes. Synangia 2.0-2.5 mm long,1 mm wide, and composed of20-30 sporangia. Sporan-gia 0.5-0.8 mm long, 0.3 mm wide, and have an obtuseapex. Monolete and trilete spores, ovoid to spherical,16-3911m in size, with a laevigate, scabrate ormicrospinate exine that is from 1 to 4 11mthick.
Holotype: 989GF-275 and 989GF-276 (Fig.2). Thehand specimen and all associated preparations from itare stored in the Palaeobotanical Collections,University College of Cape Breton, Sydney, NovaScotia, Canada.
Paratype: 989GF-274.
Locus typicus: Roof rocks of the Stubbart coal seam(upper Westphalian D), Prince Mine, Cape BretonIsland, Nova Scotia, Canada.
Etymology: Named after Dr Stephen Manley, formerDean ofthe School of Science and Technology, the Uni-versity College of Cape Breton, for his support of theexperimental geological sciences.
DI se us SlON
Sydneia manleyi is a distinctive type of marattialeanfern from the Pennsylvanian palaeotropical coal for-ests. It differs from most other marattialeans in ha v-ing deeply incised pinnules. There are some featuresthat encourage comparison with extant marattialeanssuch as Marattia, including the oval configuration ofthe synangia. However, on the whole S. manleyi dif-fers markedly from all other fossil and extant marat-tialeans, and is further evidence for the recognition ofthe diversity of Palaeozoic marattialeans.
Radstockia is widely quoted as the earliest evidenceof the Marattiaceae in the fossil record (e.g. Hill &
© 2003 The Linnean Society of London, Batanical Jaurnal ar the Linnean Saciety, 2003, 142, 199-212

210 J. PŠENIČKA ET AL.
3mm
Figure 32. Sydneia manleyi gen. et sp. nov. Reconstruction of part of the fertile frond showing a series of synangiaattached to pinnule. They are viewed from the top and slightly to the side. One of the synangia towards the end of thepinnule has been twisted around to show the lower surface (a).
Camus, 1986; Cleal, 1993; Collinson, 1996), basedmainly on the evidence presented by Taylor (1967)from the species that he called Radstockia kidstonii.ln fact, Taylor was correctly circumspect aboutwhether or not Radstockia was a primitive member ofthe Marattiaceae, as neither his specimen nor theholotype of Radstockia are sufficiently well preservedfor details ofthe synangial structure to be determined.Sydneia shares some features with extant marattiale-ans, in particular the oval configuration of the synan-gia, but as pointed out earlier there are also a numberof significant differences. It is more likely in fact thatSydneia is a Late Palaeozoic marattialean that is dif-ferent from both the Asterothecaceae and Maratti-aceae. Far more likely candidates for primitiveMarattiaceae plants are Qasimia from the Late Per-mian of Saudi Arabia (Hill, Wagner & EI-Khayal,1985) or even Danaeites saraepontanus Stur from theWestphalian of Saar-Lorraine (e.g. Laveine, 1969),both ofwhich have fronds and synangia that are moresimilar to the living plants.
This is not the first report of morphologically differ-ent types of spores occurring in the same parent plant.Spores of two or three different morphological types,belonging to dispersed spore species of two or threegenera, are known from several marattialeans(Guennel & Neavel, 1961; Horst, 1967; Laveine, 1969;Coquel & Brousmiche-Delcambre, 1996; Lesnikowska& Willard, 1997). All these authors described sporan-
gia with an outer zone containing thick-walled laevi-gate to microspinate spores (with an apical crassitude,i.e. Torispora-type), and an inner zone with laevigate(Laevigatosporites-Latosporites-type) to microspinate,microgranulate (Punctatosporites-type) spores. Thethinner-walled spore s are always more numerous thanthe thicker-walled spores. It is possible that the outerzone of thicker-walled spores represents a protectivelayer enveloping the thinner-walled spores. The samearrangement was clearly present in Sydneia manleyiand is an additional argument for the species havingmarattialean affinities.
Thomas (1987) has argued that the study of in situspores can play an important role in refining the tax-onomy of dispersed spores. This will be crucial if weare to be able to use dispersed palynology accuratelyfor vegetational studies. There are a number of earlierrecords of spores found in Late Carboniferous aster-othecoid marattialeans (Barthel, 1967; Laveine, 1969;Lesnikowska & Willard, 1997), but the present paperpresents the first evidence from a non-asterothecoidmarattialean of this age.
ACKNOWLEDGEMENTS
ELZ appreciates the financial support received fromthe Natural Science and Engineering Research Coun-cil of Canada, the American Chemical Society, Petro-leum Research Fund # 37539-AC8, and from the Nova
© 2003 The Linnean SocietyofLondon,Botanical Journal ar the Linnean Society, 2003, 142, 199-212
E
B(
SBrl
bti
CalecpA
Cle~rec
Cleafro:Zeti;46:
ClealboniPaly
CollinwithfamilPteri.349-,
Coquelspore:MaralConsiipalync
Danzé ,fougěr.
la Fraiterrain
GuennelSporeo
Gutbierauer Se.
Hemsleyon a laLochHu
© 2003 Th<

ynangiaid of the
~dlaevi-ssitude,ievigatespinate,'es. TheJUS thanne outer-otectivehe samemanleyi; having
If in situthe tax-ial if wecurately)fearlieris aster-ne, 1969;nt paper.othecoid
ved fromch Coun-;y,Petro-the Nova
, 199-212
NEW LATE WESTPHALIAN MARATTIALEAN FERN 211
Scotia Museum. We acknowledge Mrs AnnetteTownsend, National Museums and Galleries ofWales,Cardiff, for the hand drawing of a part of the fertilefrond of Sydneia manleyi.
REFERENCESBalme BE. 1995. Fossil in situ spores and pollen grains: An
annotated catalogue. Review of Palaeobotany and Palynology87: 81-323.
Barthel M. 1967. Sporen in situ aus dem Erzgebirgischen undDoehlener Becken. Monatsberichte der Deutschenakademieder Wissenschaften zu Berliri 9: 345-352.
Bek J. 1998. Spore populations of some plants of Lycophyta,Sphenophyta, Pteridophyta and Progymnospermophytafrom Carboniferous limnic basins of the Czech Republic.Unpublished PhD Thesis, Prague: Institute of Geology,Czech Academy of Sciences (in Czech).
Bek J, Opluštil S. 1998. Some lycopsid, sphenopsid andpteropsid fructifications and their miospores from the UpperCarboniferous basins of the Bohemian Massif. Palaeonto-graphica B 248: 127-161.
Brousmiche C. 1983. Les [ougěres sphénoptéridiennes dubassin houiller Sarro-Lorrain, Lil1e:Société Géolologique duNord, Publication no. 10.
Camus JM. 1990. Marattiaceae. In: Kramer KU, Green PS,eds. The families and genera of vascular plants. I. Pterido-phytes and gymnosperms. Berlin: Springer, 174-180.
Cleal CJ. 1993. Pteridophyta. In: Benton MJ, ed. The fossilrecord 2. London: Chapman & Hall, 779-794.
Cleal CJ, Shute CH. 1991. The Carboniferous pteridospermfrond Neuropteris heterophylLa (Brongniart) Sternberg. Bul-letin ofthe British Museum (Natural History), Geology Series46: 153-174.
Cleal CJ, Shute CH. 1992. Epidermal features of some Car-boniferous neuropteroid fronds. Review of Palaeobotany andPalynology 71: 191-206.
Collinson ME. 1996. 'What use are fossil ferns?' - 20 years on:with a review of the fossil history of extant pteridiophytefamilies and genera. In: Camus JM, Gibby M, Johns RJ, eds.Pteridology in perspective. Kew: Royal Botanic Gardens,349-394.
Coquel R, Brousmiche-Delcamhre C. 1996. Comparaisonsspores in situ-spores dispersées chez quelques Equisetales,Marattiales et Lépidodendrales du Carbonifere supérieur.Considérations sur l'utilisation de morphoespeces en paléo-palynologia. Review of Paléobiologie 151: 121-154.
Danzé J. 1956. Contribution a l'étude des Sphenopteris. Lesfougěres Sphénoptéridiennes du bassin houil1er du Nord dela France. Études Géologiques pour l'Atlas Topographie Sou-terraine 1 (2): 1-568.
Guennel GK, Neavel RC. 1961. Torispora securis (Balme).Spore or sporangial wall cell? Micropaleontology 7: 207-212.
Gutbier A. 1835. Abdrilcke und Versteinerungen des Zwick-auer Schwarzkohlengebirges. Zwickau: G. Richter.
Hemsley AR, Clayton G, Galtier J. 1994. Further studieson a late Tournaisian (Lower Carboniferous) flora fromLoch Humphrey Burn, Scotland: spore taxonomy and ultra-
structure. Review of Palaeobotany and Palynology 81: 213-231.
Hemsley AR, Glasspool IJ. 1999. Megaspore ultrastructure.In: Jones TP, Rowe NP, eds. Fossil plants and spores: moderntechniques. London: Geological Society, 121-125.
Hill CR, Camus JM. 1986. Evolutionary cladistics of marat-tialean ferns. Bulletin of the British Museum (Natural His-tory), Botany Series 14: 219-300.
Hill CR, Wagner RH, EI-Khayal AA. 1985. Qasimia gen.nov., an early Marattia-like fern from the Permian of SaudiArabia. Scripta Geologica 79: 1-50.
Horst U. 1967. The genus Bicoloria and its phylogenetic rela-tionships. Review of Palaeobotany and Palynology 5: 101-1l0.
Kellaway GA, Welch FBA. 1993. Geology of the Bristol dis-trict. Memoirs of the Geological Survey of Great Britain.E250: 1-199.
Kidston R. 1884. On the fructification of Zeilleria (Sphenopt-eris) delicatula, Sternberg, sp., with remarks on Urnatopt-eris (Sphenopteris) tenella Brongt. & HymenophylLites(Sphenopteris) quadridactylites, Gutbier, sp. Quarterly Jour-nal of the Geological Society, London 40: 590-598, pl. 25.
Kidston R. 1888. On the fossil flora of the Radstock Series ofthe Somerset and Bristol Coal Field (Upper Coal Measures),Transactions of the Royal Society of Edinburgh 33: 335-417,plates 18-28.
Kidston R. 1923. Fossil plants of the Carboniferous rocks ofGreat Britain. Parts 1-4. Memoirs ofthe Geological Survey ofGreat Britain, Palaeontology 2: 1-376.
Kurmann MH, Taylor TN. 1984. The ultrastructure of Bou-laya fertilis (Medullosales) pollen. Pollen et Spores 26: 109-116.
Laveine J-P. 1969. Quelques Pécoptéridinées houillěres a lalumiere de la palynologie. PolLen et Spores 11: 619-668,plates 1-17.
Lesnikowska AD, Galtier J. 1992. Permineralized Maratti-ales from the Stephanian and Autunian of central France: areinvestigation of Grandeuryella renaultii (Stur) Weissemend. Review of Palaeobotany and Palynology 72: 299-315.
Lesnikowska AD, Willard DA. 1997. Two new species ofScolecopteris (Marattiales), sources of Torisopra securisBalme and Thymospora thiessenii (Kosanke) Wilson et Ven-katachala. Review of Palaeobotany and Palynology 95: 211-225.
Mamay S. 1950. Some American Carboniferous fern fructifi-cations. Annals of the Missouri Botanical Garden 37: 409-477.
Millay MA. 1979. Studies on Paleozoic marattialeans: a mono-graph of the American species of Scolecopteris. Palaeonto-graphica B 169: 1-69.
Millay MA, Taylor TN. 1976. Evolutionary trends in fossilgymnosperm pollen. Review of Palaeobotany and Palynology21: 65-91.
Millay MA, Taylor TN. 1979. Paleozoic seed fern pollenorgans. Botanical Reviews 45: 301-375.
Millay MA, Taylor TN. 1982. The ultrastructure of Paleozoicfern spores. 1.Botryopteris. American Journal of Botany 69:1148-1155.
© 2003 The Linnean Society of London, Botanical Journal of the Linnean Society, 2003, 142, 199-212

.--r---------------
Millay MA, Taylor TN. 1984. The ultrastructure of Paleozoicfern spores. II. Scolecopteris (Marattiales). Palaeontogroph-ica B 194: 1-13.
Osborne JM, Taylor TN. 1994. Comparative ultrastructureof fossil gymnosperm pollen and its phylogenetic implica-tions. In: Kurmann MH, Doyle JA, eds. Ultrastructure offos-sil spores and pollen. Kew: Royal Botanic Gardens, 99-12I.
Potonié H. 1892a. Ueber einige Carbonfarne. Jahrbuch derKoniglicli Preussischen Geologischen Landesanstalt und Ber-gakademie, Berlin 10: 21-27, plates 2-5.
Potonié H. 1892b. Ueber einige Carbonfarne. II. Theil. Jahr-buch der Koniglicli Preussischen Geologischen Landesanstaltund Bergakademie, Berlin 11: 11-39, plates 7-9.
Remy W. 1953. Untersuchungen uber einige Fruktifikationenvon Farnen und Pteridospermen aus dem mitteleuropais-chen Karbon und Perm. Abhandlungen der Deutschen Akad-emie der Wissenschaften zu Berlin, Klasse [iir Mathematischeund Allgemeine Naturwissenschaft 1952 (2): 5-38, 7 plates.
Schimper W. 1869. Traité de paléontologie oégétale, 1. Paris:J. B. Balliere et Fils.
Stidd BM. 1981. The current status ofmedullosan seed ferns.Reuiew of Palaeobotany and Palynology 32: 63-10I.
Taylor TN. 1967. On the structure and phylogenetic relation-ships of the fern Radstockia Kidston. Palaeontology 10: 43-46.
Taylor TN. 1973. Combined transmission and scanning elec-tron microscopy of in situ Palaeozoic spores. Palaeontology16: 765-776.
Taylor TN. 1982. Ultrastructural studies of Paleozoic seed
fern pollen: sporoderm development. Reuiew of Palaeobotanyand Palynology 37: 29-53.
Taylor TN, Rothwell GW. 1982. Studies of seed fern pollen:Development ofthe exine in Monoletes (Medullosales). Amer-ican Journal of Botany 69: 570-578.
Thomas BA. 1987. The use of in situ spores for defining spe-cies of dispersed spores. Reuiew of Palaeobotany and Palynol-ogy 51: 227-233.
Thomas BA, Crampton DM. 1971. A fertile Zeilleriaauoldensis from the British Upper Carboniferous. Reuiew ofPalaeobotany and Palynology 11: 283-295.
Tryon R. 1960. A glossary of some terms relating to the fernleaf. Taxon 9: 104-109.
Tryon AF, Lugardon B. 1990. Spores of the Pteridophyta.New York: Springer.
Zodrow EL. 1986. Succession of paleobotanical events: evi-dence for mid-Westphalian D floral changes, Morien Group(Late Pennsylvanian, Nova Scotia). Reuiew of Palaeobotanyand Palynology 47: 293-326.
Zodrow EL, Bek J, Pšenička J. 2000. In situ pecopteridmicrospores from near the Westphalian D - Cantabrianboundary in Sydney Coalfield, Nova Scotia, Canada. News-letter on Carboniferous Stratigraphy 18: 33-35.
Zodrow EL, Cleal CJ. 1998. Revision of the pteridospermfoliage Alethopteris and Lonchopteridium (Upper Carbonif-erous), Sydney Coalfield, Nova Scotia, Canada. Palaeon-tographca B 247: 65-122, 14 plates.
Zodrow EL, Pšenička J, Bek J, Hemsley AR, Cleal CJ.2001. Microspores or pollen? Newsletter on CarboniferousStratigraphy 19: 49-50.
© 2003 The Linnean Society of London, Botanical Journal of the Linnean Society, 2003, 142, 199-212
i~
Related Documents






![Cross-Fertilization of Westphalian Approaches to ...law.emory.edu/eilr/_documents/volumes/34/4/lone.pdfLONE_5.26.20 5/26/2020 3:14 PM 2020] CROSS-FERTILIZATION OF WESTPHALIAN APPROACHES](https://static.cupdf.com/doc/110x72/5f6000b2a4652457536b6f02/cross-fertilization-of-westphalian-approaches-to-lawemoryedueilrdocumentsvolumes344lonepdf.jpg)




