Accepted by M.R. de Carvalho: 20 Aug. 2013; published: 26 Sept. 2013 ZOOTAXA ISSN 1175-5326 (print edition) ISSN 1175-5334 (online edition) Copyright © 2013 Magnolia Press Zootaxa 3717 (1): 001–022 www.mapress.com/zootaxa/ Article 1 http://dx.doi.org/10.11646/zootaxa.3717.1.1 http://zoobank.org/urn:lsid:zoobank.org:pub:38CCFE74-24B5-4A51-9FB4-51838784D065 A new Brachyrhamdia (Siluriformes: Heptapteridae) from Rio Japurá basin, Brazil, with comments on its phylogenetic affinities, biogeography and mimicry in the genus VERONICA SLOBODIAN 1,3 & FLÁVIO ALICINO BOCKMANN 2,3 1 Laboratório de Ictiologia, Museu de Zoologia da Universidade de São Paulo, Caixa Postal 42494, 04218-970 São Paulo, SP, Brazil. E-mail: [email protected] 2 Laboratório de Ictiologia de Ribeirão Preto (LIRP), Departamento de Biologia, FFCLRP, Universidade de São Paulo, Av. dos Ban- deirantes 3900, 14040-901 Ribeirão Preto, SP, Brazil. E-mail: [email protected] 3 Programa de Pós-Graduação em Biologia Comparada, FFCLRP, Universidade de São Paulo, Av. dos Bandeirantes 3900, 14040- 901, Ribeirão Preto, SP, Brazil Abstract Brachyrhamdia thayeria is herein described as a new heptapterid species from Rio Japurá, a left margin tributary of the Rio Solimões, Amazonas basin, Brazil. The new species is diagnosed from all its congeners by having a putative autapo- morphy: dark oblique stripe across the caudal peduncle, originating below the anterior half of adipose fin and ending at base of rays of ventral caudal-fin lobe. Brachyrhamdia thayeria is considered closely related to B. imitator and B. ram- barrani with which it shares a low vertebral number and absence of lateral stripe along trunk. The new species shares ex- clusively with B. rambarrani the presence of a dark conspicuous bar along the dorsolateral region of trunk, a putative synapomorphy. Brachyrhamdia are distributed in lowland regions of northern South America where they inhabit small streams with running waters. The new species appears to have a mimetic association with the syntopic callichthyid Cory- doras arcuatus. Herein, we argue in favor of the hypothesis that the interaction between species of Brachyrhamdia and Pimelodella or Corydoras is Müllerian mimicry. Key words: taxonomy, biodiversity, morphology, Amazon basin Brachyrhamdia thayeria é descrita como uma nova espécie de heptapterídio do Rio Japurá, um afluente da margem es- querda do Rio Solimões, na porção média da bacia do Rio Amazonas, Brasil. A nova espécie é diagnosticada de suas con- gêneres com base em uma possível autapomorfia: uma faixa escura oblíqua no pedúnculo caudal, se originando logo abaixo da metade anterior da nadadeira adiposa e terminando na base dos raios do lobo ventral da nadadeira caudal. Brachyrhamdia thayeria é considerada estreitamente relacionada à B. imitator e B. rambarrani com base na posse de um número baixo de vértebras e da ausência de faixa lateral ao longo do tronco. A nova espécie compartilha exclusivamente com B. rambarrani a presença de uma barra escura na região dorsolateral do tronco, uma possível sinapomorfia. Brachy- rhamdia se distribui em terras baixas do norte da América do Sul, onde elas habitam pequenos riachos com água corrente. Esta nova espécie provavelmente apresenta uma associação mimética com Corydoras arcuatus, com a qual ocorre sin- topicamente. São fornecidos argumentos que favorecem a hipótese de que a interação entre espécies de Brachyrhamdia e de Pimelodella e Corydoras é um mimetismo Mülleriano. Palavras-chave: taxonomia, biodiversidade, morfologia, bacia Amazônica Introduction Brachyrhamdia Myers (1927) is a catfish genus belonging to the family Heptapteridae and distributed throughout the Río Orinoco, Guianas and Amazonas basins (including Rio Tocantins basin) (Bockmann & Guazzelli, 2003; Slobodian, 2013). Species of Brachyrhamdia resemble those belonging to Pimelodella Eigenmann & Eigenmann TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
ZOOTAXA
ISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2013 Magnolia Press
Zootaxa 3717 (1): 001–022
www.mapress.com/zootaxa/Article
http://dx.doi.org/10.11646/zootaxa.3717.1.1
http://zoobank.org/urn:lsid:zoobank.org:pub:38CCFE74-24B5-4A51-9FB4-51838784D065
A new Brachyrhamdia (Siluriformes: Heptapteridae) from Rio Japurá basin,
Brazil, with comments on its phylogenetic affinities,
biogeography and mimicry in the genus
VERONICA SLOBODIAN1,3 & FLÁVIO ALICINO BOCKMANN2,3
1Laboratório de Ictiologia, Museu de Zoologia da Universidade de São Paulo, Caixa Postal 42494, 04218-970 São Paulo, SP, Brazil.
E-mail: [email protected]ório de Ictiologia de Ribeirão Preto (LIRP), Departamento de Biologia, FFCLRP, Universidade de São Paulo, Av. dos Ban-
deirantes 3900, 14040-901 Ribeirão Preto, SP, Brazil. E-mail: [email protected] de Pós-Graduação em Biologia Comparada, FFCLRP, Universidade de São Paulo, Av. dos Bandeirantes 3900, 14040-
901, Ribeirão Preto, SP, Brazil
Abstract
Brachyrhamdia thayeria is herein described as a new heptapterid species from Rio Japurá, a left margin tributary of the
Rio Solimões, Amazonas basin, Brazil. The new species is diagnosed from all its congeners by having a putative autapo-
morphy: dark oblique stripe across the caudal peduncle, originating below the anterior half of adipose fin and ending at
base of rays of ventral caudal-fin lobe. Brachyrhamdia thayeria is considered closely related to B. imitator and B. ram-
barrani with which it shares a low vertebral number and absence of lateral stripe along trunk. The new species shares ex-
clusively with B. rambarrani the presence of a dark conspicuous bar along the dorsolateral region of trunk, a putative
synapomorphy. Brachyrhamdia are distributed in lowland regions of northern South America where they inhabit small
streams with running waters. The new species appears to have a mimetic association with the syntopic callichthyid Cory-
doras arcuatus. Herein, we argue in favor of the hypothesis that the interaction between species of Brachyrhamdia and
Pimelodella or Corydoras is Müllerian mimicry.
Key words: taxonomy, biodiversity, morphology, Amazon basin
Brachyrhamdia thayeria é descrita como uma nova espécie de heptapterídio do Rio Japurá, um afluente da margem es-
querda do Rio Solimões, na porção média da bacia do Rio Amazonas, Brasil. A nova espécie é diagnosticada de suas con-
gêneres com base em uma possível autapomorfia: uma faixa escura oblíqua no pedúnculo caudal, se originando logo
abaixo da metade anterior da nadadeira adiposa e terminando na base dos raios do lobo ventral da nadadeira caudal.
Brachyrhamdia thayeria é considerada estreitamente relacionada à B. imitator e B. rambarrani com base na posse de um
número baixo de vértebras e da ausência de faixa lateral ao longo do tronco. A nova espécie compartilha exclusivamente
com B. rambarrani a presença de uma barra escura na região dorsolateral do tronco, uma possível sinapomorfia. Brachy-
rhamdia se distribui em terras baixas do norte da América do Sul, onde elas habitam pequenos riachos com água corrente.
Esta nova espécie provavelmente apresenta uma associação mimética com Corydoras arcuatus, com a qual ocorre sin-
topicamente. São fornecidos argumentos que favorecem a hipótese de que a interação entre espécies de Brachyrhamdia e
de Pimelodella e Corydoras é um mimetismo Mülleriano.
Palavras-chave: taxonomia, biodiversidade, morfologia, bacia Amazônica
Introduction
Brachyrhamdia Myers (1927) is a catfish genus belonging to the family Heptapteridae and distributed throughout the Río Orinoco, Guianas and Amazonas basins (including Rio Tocantins basin) (Bockmann & Guazzelli, 2003; Slobodian, 2013). Species of Brachyrhamdia resemble those belonging to Pimelodella Eigenmann & Eigenmann
Accepted by M.R. de Carvalho: 20 Aug. 2013; published: 26 Sept. 2013 1

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
(1888) but differ in being smaller (up to 76 mm SL), having a stout body, and bearing remarkable coloration patterns (Sands & Black, 1985; Lundberg & McDade, 1986; Bockmann & Guazzelli, 2003).
Species of Brachyrhamdia are popular among fish hobbyists and usually imported along with specimens of callichthyids of the genus Corydoras La Cepède (1803) (Innes & Myers, 1950; Sands, 1984, 1985a; Sands & Black, 1985). Recurrent associations of Brachyrhamdia with similarly colored species of Corydoras, Pimelodella
and the hypoptopomatin Otocinclus Cope (1871) led to the suspicions of mimetic relationships among those catfishes (Innes & Myers, 1950; Sands, 1980; Sands, 1985b; Sands & Black, 1985; Lundberg & McDade, 1986; Axelrod & Burgess in Axelrod, 1987; Hercos et al., 2009).
The validity of Brachyrhamdia has been questioned at times, the genus being considered indistinguishable from Pimelodella (Schultz, 1944; Innes & Myers, 1950; Sands, 1985b; Lundberg & McDade, 1986; Axelrod & Burgess in Axelrod, 1987; Bockmann, 1998) and/or a possible junior synonym of Rhamdia Bleeker (1858) (Zarske, 2003). Given the uncertain phylogenetic placement of Brachyrhamdia among other basal genera of Heptapteridae (see Bockmann & Miquelarena, 2008) and the fact that its included species certainly compose a monophyletic group, we conservatively treat Brachyrhamdia as valid, following recent reviewers (Sands & Black, 1985; Lundberg & Mc Dade, 1986; Bockmann & Guazzelli, 2003; Ferraris, 2007; Bockmann & Miquelarena, 2008).
Presently, Brachyrhamdia contains five valid species (Bockmann & Guazzelli, 2003): B. imitator Myers (1927), the type species, occurring in Río Orinoco basin of Venezuela; B. heteropleura (Eigenmann, 1912), inhabiting Essequibo and Corantijn River basins of Guyana and Suriname, respectively, and Rio Branco basin of Brazil; B. marthae Sands & Black (1985), distributed throughout the Madeira-Mamoré-Madre de Díos system of Bolivia, Brazil, and Peru; B. meesi Sands & Black (1985), currently known only from aquarium exports; and B.
rambarrani (Axelrod & Burgess in Axelrod, 1987), distributed in Rio Negro and its affluent Rio Unini, in Brazil. However, at least two undescribed species of Brachyrhamdia occurring in Brazil are mentioned in the literature (e.g., Hercos et al., 2009; Queiroz & Hercos, 2009; Slobodian & Bockmann, 2011, 2013; Slobodian et al., 2011; Slobodian, 2013). One of them, reported from igarapés of Amanã Lake, an affluent of Rio Japurá, in Middle Amazon, Brazil (Queiroz & Hercos, 2009; Hercos et al., 2009; Slobodian et al., 2011), is herein described.
Material and methods
Measurements were taken as point-to-point distances with digital calipers under a dissecting scope, and are recorded to the nearest 0.1 mm, following Bockmann & de Pinna (2004) with modifications. All type specimens were measured, including that one which was cleared and stained (see below), whose measurements were taken prior to its preparation. Measurements of head parts are presented as proportions of head length, except for measurements of barbels that were converted to proportions of standard length; head length and measurements of body parts are given as proportions of SL. Methodology for taking data on meristics and fin positions followed Bockmann & Castro (2010). Anal-fin rays were counted individually, including the anterior splints and the two most posterior rays. Vertebral counts include the anteriormost elements modified into a complex vertebra (counted as five), all free vertebrae, and the compound caudal centrum (PU1+U1) counted as one (Lundberg & Baskin, 1969). For osteological analysis, specimens were cleared and stained according to the procedures of Taylor & Van Dyke (1985). Illustrations were prepared under a stereomicroscope with camera lucida and edited electronically, or prepared digitally with a stereomicroscope Leica MZ16 and digital camera Leica DFC295. Radiographs also were obtained with a Faxitron LX-60 digital system, Faxitron DX software. All digital radiographs were stored at Laboratório de Ictiologia de Ribeirão Preto, Universidade de São Paulo, Brazil, and copies were sent to each institution holding the radiographed specimen(s). Notations cited throughout the text include ‘c&s’ for cleared and stained specimens, ‘SL’ for standard length, ‘TL’ for total length, and ‘xr’ for x-rayed specimens. The map was made using Google Earth® (version 7.0.3.8542) and Quantum Gis (version 1.8.0) softwares.
Osteological terminology follows Bockmann & Miquelarena (2008), with addition of Carvalho et al. (2013) for accessory element of ceratobranchials. Nomenclature for laterosensory canals follows Northcutt (1989), with details of supraorbital and infraorbital sensory canal systems and preoperculomandibular sensory canal system according to Arratia & Huaquín (1995) and Bockmann & Miquelarena (2008).
Institutional abbreviations are listed in Sabaj Pérez (2013). Specimens of the other species of Brachyrhamdia
SLOBODIAN & BOCKMANN2 · Zootaxa 3717 (1) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
examined and species of Pimelodella are listed in the Appendix; remaining heptapterids examined for comparative purposes are listed in Bockmann & Miquelarena (2008) and Bockmann & Castro (2010).
Results
Brachyrhamdia thayeria, n. sp.
Figures 1–5; Table 1
Brachyrhamdia n. sp. ‘Amanã’.—Slobodian & Bockmann, 2013 [phylogenetic relationships: 462].Brachyrhamdia sp. 1.—Queiroz & Hercos, 2009 [abundance in igarapés of Amanã Lake, sampled October, 2006: 63, unnumb.
table]; Hercos, Queiroz & Almeida, 2009 [fishes of the Amanã Reservation: 51, tab. 1; photograph of live specimen in aquarium: 156; brief description: 156; considered rare: 156; observed in shoals of Corydoras: 156; data on habitat and natural history: 156–157; distribution in Lago Amanã basin: 157, map].
[Nova espécie de] Brachyrhamdia.—Slobodian, Bockmann & Sousa [Rio Japurá basin: 192; similar to B. rambarrani: 192; possible syntopy with Corydoras arcuatus Elwin (1939) (Callichthyidae): 192].
Holotype. INPA 39534, 50.7 mm SL (xr), Brazil, Amazonas, Maraã, Lago Amanã, Rio Japurá basin, 02°32’06”S, 65°43’02”W, 44 m a.s.l., 14 Mar 2003, M. Catarino, J. Zuanon, L. M. Sousa et al.
Paratypes. ANSP 194110, 1, xy, 35.9 mm SL, Brazil, Amazonas, Amanã, Igarapé Baré, tributary to Rio Japurá, 02°17’15”S, 64°41’16”W, 44 m a.s.l., 6 Nov 2002, M. Catarino, L. M. Sousa et al.; INPA 25306, 2, xy, 37.9–41.0 mm SL, collected with the holotype; LIRP 10237, 1 xr, 44.5 mm SL, 1 c&s, 55.2 mm SL, collected with the holotype; INPA 25740, 1, xy, 31.3 mm SL, collected with ANSP 194110; INPA 25741, 2, xy, 36.8–37.1 mm SL, Brazil, Amazonas, Amanã, Igarapé Baré, tributary to Rio Japurá, 02°20’10”S, 64°42’43”W, 44 m a.s.l., 6 Nov 2002, coll. M. Catarino, L. M. Sousa et al.; MZUSP 114011, 1, xr, 34.1 mm SL, collected with INPA 25741.
Diagnosis. Brachyrhamdia thayeria is distinguished from all its congeners by a unique coloration pattern represented by a dark brown stripe at caudal region extending transversely from region immediately below middle of adipose-fin base to the base of the ventral caudal-fin lobe. It shares exclusively with B. rambarrani the presence of a dark brown band on dorsolateral region of body. In B. thayeria, however, the band originates just posterior to the eye and finishes on caudal peduncle just posterior to adipose-fin base, whereas in B. rambarrani the band begins just below the dorsal-fin origin. It further differs from B. rambarrani by the having the supraoccipital process roughly triangular (vs. roughly rectangular). Brachyrhamdia thayeria is distinguished from B.
heteropleura, B. marthae, and B. meesi by having a lower vertebral count, usually 35–36 vertebrae (vs. 37–39 vertebrae) and absence of dark lateral stripe along the trunk (vs. distinct stripe in B. marthae and B. meesi and faint stripe in B. heteropleura). It differs from B. heteropleura, B. imitator, and B. rambarrani in possessing the pharyngobranchial 1, which is otherwise missing in those species. Brachyrhamdia thayeria is promptly separated from B. marthae and B. meesi by having large, pointed, retrorse dentations along the basal portion of medial edge of pectoral spine (vs. dentations on medial edge of pectoral spine distinctly smaller, antrorse or perpendicularly oriented, and some with bifid apices).
Description. Morphometric data presented in Table 1. Body (Figure 1) stout, progressively more compressed caudally from cleithrum. Dorsal profile of body arched from snout tip to caudal peduncle. Ventral profile of body slightly convex from mandible to ventral-fin origin and approximately straight from this region to caudal peduncle. Greatest body depth at dorsal-fin origin. Pseudotympanum large, triangular, above posterior process of cleithrum (Figure 1). Posterior cleithral process slender, sharply pointed. Axillary pore as slit immediately above the posterior portion of pectoral-fin base, below posterior cleithral process. Urogenital papilla tubular, triangular, short. Anus and urogenital papilla adjacent. Anus between verticals through end of first third of pelvic fin and anterior portion of second third of pelvic fin; urogenital papilla between verticals through end of second third of pelvic fin and anterior portion of the last third of pelvic fin.
Head (Figure 1) long and deep; snout terminal. Eyes elliptical, large, with greatest length in horizontal axis, placed dorsolaterally. Limits of eye well defined by marked invagination, especially pronounced around anterior and dorsal rim. Anterior naris tubular; posterior naris completely surrounded by shallow skin flap except for narrow posterolateral region. Premaxilla without backward projecting angle. Premaxilla and dentary each with 5–6 rows of small viliform teeth. Anteriormost tooth row of each premaxilla with 29–31 teeth; anteriormost tooth row
Zootaxa 3717 (1) © 2013 Magnolia Press · 3A NEW SPECIES OF BRACHYRHAMDIA

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
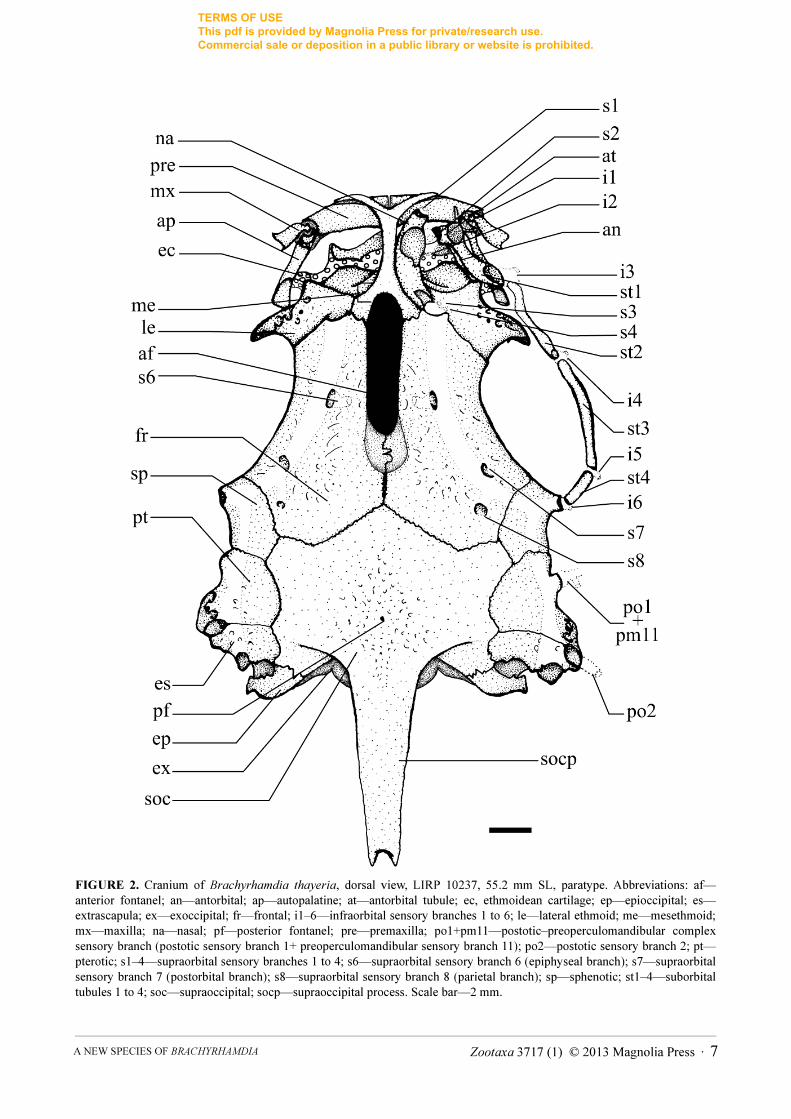
of each dentary with 37–42 teeth. Palate and vomer edentulous. Barbels relatively long, thin, and elliptical in cross-section (Figure 1). Tip of maxillary barbel extending to posterior margin of anal fin or sometimes beyond, reaching base of caudal-fin rays. Mental barbel, when parallel to main body axis, with distal tip finishing in region between anterior and mid-third of dorsal-fin base. Inner mental barbel, when lying parallel to main body axis, with distal tip finishing in region between middle and outer border of branchiostegal membrane. Outer mental barbel longer than inner barbel. Insertion of inner mental barbel anterior to origin of outer mental barbel. Supraoccipital process roughly triangular, with broad base and lateral edges converging posteriorly towards tip (Figure 2).
FIGURE 1. Brachyrhamdia thayeria, INPA 39534, 50.7 mm SL, holotype, Brazil, Amazonas, Maraã, Lago Amanã, Rio Japurá basin.
Branchiostegal membranes well developed, free, united to isthmus only at medial apex, and not connected to each other anteriorly. Branchiostegal rays 6 (9*)–7 (1). Branchial rakers short, 7 (4), 8 (5*), 9 (1) on first ceratobranchial (including one on angle formed with epibranchial), and 1 (1), 2 (6*), 3 (3) on first epibranchial. Pharyngobranchial 1 present (Figure 3).
Dorsal fin triangular, distally concave (Figure 1), reaching adipose fin when adpressed. Dorsal fin with i, 5 (1), i, 6 (8*), plus anteriormost spinelet. Cleared and stained specimen with abnormal extra posterior ray. Distance between dorsal and adipose fins short, about 60–80% of dorsal-fin base. Dorsal-fin rays supported by seven blade-like pterygiophores. Proximal tip of anteriormost pterygiophore of dorsal fin located between bifid neural spine of vertebra 4–5 (10*); proximal tip of posteriormost pterygiophore of dorsal fin located between space of neural (or pseudoneural) spine of vertebra 9 (1), 9–10 (7*), and 10 (2). Spinelet large, with wide base and rounded distal extremity. Supraneural triangular dorsally, with anterior point fitted into distal margin of supraoccipital process,
SLOBODIAN & BOCKMANN4 · Zootaxa 3717 (1) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
and with long ventral process abutting anterior edge of first pterygiophore. Distal extremity of first pterygiophore expanded, forming typical median, anteriormost nuchal plate 1. Distal extremity of second pterygiophore expanded laterally, forming typically paired nuchal plates 2. Dorsal-fin pterygiophores 1 and 2 proximate to each other, distally sutured.
TABLE 1. Morphometric data of Brachyrhamdia thayeria. n. sp., based on holotype (INPA 39534) and 9 paratypes
(ANSP 194110, INPA 25306, INPA 25740, INPA 25741, LIRP 10237, and MZUSP 114011). Abbreviation: H—
holotype; Max—maximum; Min—minimum; n—number of specimens; SD—standard deviation; x— average.
H Min Max x SD n
Total length (mm) 65.5 40.6 65.5 - - 10
Standard length (mm) 50.7 30.3 54.2 - - 10
As percentage of Standard Length
Body depth (dorsal) 25.3 22.6 28.4 24.7 1.77 10
Body width (dorsal) 17.1 15.8 20.4 18.1 1.39 10
Cleithral width 21.3 21.3 23.7 22.1 0.68 10
Head length 34.6 27.3 41.8 35.2 5.18 10
As percentage of Head Length
Head depth 63.6 54.6 79.0 65.1 9.43 10
Head width 60.9 53.3 78.3 63.7 9.71 10
Eye diameter (left) 22.7 18.6 28.2 22.9 3.16 10
Eye diameter (right) 22.9 19.9 28.7 23.5 3.11 10
Fleshy interorbital 29.6 28.0 38.3 32.4 3.72 10
Bony interorbital 24.5 21.5 30.0 25.6 2.90 10
Mouth gape 32.9 28.0 40.5 33.1 5.15 10
Snout length (left) 27.4 25.2 36.6 30.8 3.62 10
Snout length (right) 28.0 23.0 37.6 30.3 4.11 10
Distance between snout tip and posterior nare (left side) 20.1 13.8 24.4 18.8 3.10 10
Distance between snout tip and posterior nare (right side) 19.8 15.2 20.5 17.9 1.85 10
Anterior internarial width 13.3 11.8 17.3 14.0 1.93 10
Posterior internarial width 17.1 14.9 22.4 17.9 2.55 10
Intranarial length (left side) 12.0 8.5 16.0 12.0 2.72 10
Intranarial length (right side) 12.5 8.1 17.6 11.5 3.21 10
As percentage of Standard Length
Maxillary-barbel length (left side) 88.5 80.8 100.4 92.5 6.60 10
Maxillary-barbel length (right side) 84.3 82.3 103.2 95.4 6.85 10
Outer mental-barbel length (left side) 43.3 27.4 43.3 36.0 4.80 10
Outer mental-barbel length (right side) 39.2 31.7 40.5 37.0 2.86 10
Inner mental-barbel length (left side) 28.1 18.7 28.1 22.7 2.60 10
Inner mental-barbel length (right side) 27.2 20.5 27.2 23.0 2.48 7
Predorsal length 37.7 35.5 41.9 39.0 1.94 10
Distance between snout tip and end of dorsal-fin base 52.5 52.3 56.0 53.9 1.22 10
Distance between snout tip and dorsal-fin distal tip 64.1 59.3 67.9 64.5 2.79 10
Dorsal fin to adipose fin 9.4 8.3 12.0 9.7 1.06 10
......continued on the next page
Zootaxa 3717 (1) © 2013 Magnolia Press · 5A NEW SPECIES OF BRACHYRHAMDIA

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
TABLE 1. (Continued)
H Min Max x SD n
Dorsal-fin base 15.3 13.9 17.6 16.0 1.27 10
Length of first dorsal-fin ray (unbranched) 21.9 21.6 26.7 23.8 1.74 9
Length of rigid part of first dorsal-fin ray 18.8 16.2 24.5 20.4 2.26 10
Length of second dorsal-fin ray (first branched) 22.1 17.9 25.4 22.3 2.16 10
Length of third dorsal-fin ray (second branched) 19.6 15.9 22.4 20.2 1.84 10
Prepectoral length 25.1 25.1 29.9 27.6 1.55 10
Distance between snout tip and end of pectoral-fin base 28.7 28.7 33.4 31.1 1.56 10
Distance between snout tip and pectoral-fin distal tip 48.7 46.8 53.7 50.8 2.38 10
Length of first left pectoral-fin ray (unbranched) 24.2 24.2 27.8 26.3 1.19 9
Length of rigid part of first left dorsal-fin ray 21.6 21.6 26.6 23.8 1.62 10
Length of second left pectoral-fin ray (first branched) 22.0 17.7 24.4 21.6 2.28 9
Length of third left pectoral-fin ray (second branched) 17.7 12.6 22.5 18.6 3.40 8
Length of first right pectoral-fin ray (unbranched) - 22.9 27.2 25.0 1.32 8
Length of rigid part of first right dorsal-fin ray 19.6 19.6 25.1 22.8 1.79 9
Length of second right pectoral-fin ray (first branched) - 18.2 25.2 22.2 2.23 9
Length of third right pectoral-fin ray (second branched) - 14.6 22.4 18.2 2.29 8
Prepelvic length 54.9 50.6 58.9 53.9 2.40 10
Distance between snout tip and end pelvic-fin base 56.8 50.5 59.2 55.8 2.61 10
Distance between snout tip and pelvic-fin distal tip 69.1 64.4 71.2 68.0 1.90 10
Distance between pelvic fins 6.4 4.5 6.8 5.5 0.78 10
Length of first left pelvic-fin ray (unbranched) 13.4 9.7 15.4 13.4 1.65 10
Length of second left pelvic-fin ray (first branched) 14.2 10.0 16.3 14.3 1.69 10
Length of third left pelvic-fin ray (second branched) 13.2 11.0 17.6 15.1 1.94 10
Length of first right pelvic-fin ray (unbranched) 13.1 12.4 15.3 13.7 1.01 10
Length of second right pelvic-fin ray (first branched) 15.1 12.9 16.5 14.9 1.22 10
Length of third right pelvic-fin ray (second branched) 13.8 10.1 17.2 14.7 2.02 10
Anal-fin base 11.7 11.6 13.4 12.5 0.70 10
Preanal length 69.8 68.5 74.8 71.0 2.32 10
Distance between snout tip and end of anal-fin base 82.6 81.2 87.8 84.0 2.22 10
Distance between snout tip and anal-fin distal tip 90.7 89.1 93.6 91.3 1.64 10
Adipose-fin length 27.6 22.5 28.5 26.0 1.71 10
Preadipose length 61.6 59.1 69.9 63.3 2.79 10
Distance between snout tip and end of adipose-fin base 87.3 84.9 91.1 87.2 1.57 10
Adipose-fin depth 4.3 4.0 5.9 4.7 0.57 10
Caudal-peduncle length 12.4 12.0 14.2 13.0 0.80 10
Caudal-peduncle depth 10.3 10.3 11.6 10.9 0.39 10
Snout-anus distance 59.8 57.5 63.6 59.8 1.93 10
Snout-urogenital papilla distance 64.4 61.4 69.4 64.7 2.63 10
Anus-urogenital papilla distance 5.6 3.7 7.1 5.4 1.20 10
Length of dorsal caudal-fin lobe 30.0 20.5 30.0 27.7 3.03 9
Length of ventral caudal-fin lobe 26.5 22.1 30.5 27.2 2.26 10
SLOBODIAN & BOCKMANN6 · Zootaxa 3717 (1) © 2013 Magnolia Press
TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURE 2. Cranium of Brachyrhamdia thayeria, dorsal view, LIRP 10237, 55.2 mm SL, paratype. Abbreviations: af—anterior fontanel; an—antorbital; ap—autopalatine; at—antorbital tubule; ec, ethmoidean cartilage; ep—epioccipital; es—extrascapula; ex—exoccipital; fr—frontal; i1–6—infraorbital sensory branches 1 to 6; le—lateral ethmoid; me—mesethmoid; mx—maxilla; na—nasal; pf—posterior fontanel; pre—premaxilla; po1+pm11—postotic–preoperculomandibular complex sensory branch (postotic sensory branch 1+ preoperculomandibular sensory branch 11); po2—postotic sensory branch 2; pt—pterotic; s1–4—supraorbital sensory branches 1 to 4; s6—supraorbital sensory branch 6 (epiphyseal branch); s7—supraorbital sensory branch 7 (postorbital branch); s8—supraorbital sensory branch 8 (parietal branch); sp—sphenotic; st1–4—suborbital tubules 1 to 4; soc—supraoccipital; socp—supraoccipital process. Scale bar—2 mm.
Zootaxa 3717 (1) © 2013 Magnolia Press · 7A NEW SPECIES OF BRACHYRHAMDIA

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURE 3. Branchial arches of Brachyrhamdia thayeria, dorsal view, LIRP 10237, 55.2 mm SL, paratype. Gill rakers and dorsal elements of left gill arches not shown. Abbreviations: ac4—accessory element of ceratobranchial 4; cn—cartilaginous nodule; bb2–4—basibranchials 2 to 4; cb1–5—ceratobranchials 1 to 5; eb1–4—epibranchials 1 to 4; hb1–3—hypobranchials 1 to 3; pb1—pharyngobranchial 1; pb3–4—pharyngobranchials 3 and 4; tp—tooth plate; and up—uncinate process. Scale bar—1 mm.
FIGURE 4. Right pectoral spine of Brachyrhamdia thayeria, dorsal view, LIRP 10237, 55.2 mm SL, paratype. Scale bar—1 mm.
SLOBODIAN & BOCKMANN8 · Zootaxa 3717 (1) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURE 5. Caudal skeleton of Brachyrhamdia thayeria, LIRP 10237, 55.2 mm SL, paratype. Abbreviations: cf—caudal artery foramen; ep—epural; ha+has—complex hypurapophysis composed of hypurapophysis and secondary hypurapophysis; hu1+hu2—complex ventral plate formed by co-ossification of hypurals 1 and 2; hu3+hu4—complex dorsal plate formed by co-ossification of hypurals 3 and 4; hu5—hypural 5; ph—parhypural; pu1+u1—complex centrum composed of preural centrum 1 and ural centrum 1; pu2—preural centrum 2; and ur—uroneural. Scale bar—1 mm.
Pectoral fin triangular with concave distal border, I, 7 (10*). First pectoral-fin ray long, roughly straight, with proximal part rigid, forming a spine, and short distal tip flexible and distinctly segmented. Lateral margin of spiny portion of first pectoral-fin ray with 26 (3), 27 (1), 30 (3), 31 (2), 32 (1*) tiny, perpendicular to slightly antrorse dentations from basal portion to or slightly short of midpoint of spine, and with 4–5 smooth serrae along its distal third. Medial margin of spiny portion of first pectoral-fin ray with 7 (2), 8 (7), 10 (1*) retrorse dentations, from region just beyond base of spine to or slightly short of midpoint of spine, plus 1–2 unossified distalmost dentations (Figure 4).
Pelvic fin triangular with straight distal border, i,5 rays (10*). Anterior portion of pelvic-fin base at vertical through region just posterior of dorsal-fin base (Figure 1). Inner margins of pelvic-fin bases remote from each other. Tip of adpressed pelvic fin falling short of vertical through anal-fin origin. First ray unbranched, completely flexible, segmented, and distinctly shorter than second and third rays (first and second branched rays, respectively). Site of insertion of first pelvic-fin ray on basipterygium below vertebral centra 12 to 14, with variation as follows: between centra 12–13 (2), centrum 13 (3), and centra 13–14 (5*).
Anal fin with short base and convex distal border, supported by 12 (5), 13 (3*), and 14 (2) rays, including 7 (5) and 8 (5*) branched rays. Anal-fin rays with following branching pattern: v,7 (4), vi,7 (1), iv,8 (1), v,8 (2*), and vi,8 (2). Two or three anteriormost anal-fin rays vestigial, unsegmented, embedded in thick anterior fold. Origin of anal-fin base posterior to adipose-fin origin. End of anal-fin base at vertical through the anterior portion of last fourth of adipose-fin base. Tip of anteriormost pterygiophore of anal fin between hemal spines of vertebrae 19–20 (8*) and 20–21 (2). Tip of posteriormost pterygiophore of anal fin between hemal spines of vertebrae 24–25 (9*) and 25–26 (1).
Zootaxa 3717 (1) © 2013 Magnolia Press · 9A NEW SPECIES OF BRACHYRHAMDIA

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Adipose fin long, forming ascending elevated curve in lateral profile, with highest point approximately at middle third. Adipose fin merging gradually with back anteriorly, its origin difficult to pinpoint. Origin of adipose-fin base above vertebral centra 17 to 19, with variation as follows: centrum 17 (1), 18 (7*), and 19 (2). Posterior limit of adipose fin well defined, ending in rounded free lobe. End of adipose-fin base above vertebral centra 29 to 31, with variation as follows: between centra 29–30 (2), centrum 30 (4), between centra 30–31 (3*), and centrum
31 (1).Caudal fin (Figures 1 and 5) deeply forked, with dorsal lobe usually slightly longer than ventral lobe. Branched
caudal-fin rays divided two or three times. Central rays of caudal fin also branched, curved, and devoid of marginal expansions. Total caudal fin-rays 49 (2), 51 (5), 52 (2), and 53 (1*); with 24 (2), 25 (5), and 26 (3*) rays in dorsal lobe, and 24 (1), 25 (2), 26 (4), and 27 (3*) rays in ventral lobe. Dorsal lobe with 7 (10*) branched rays; ventral lobe with 8 (10*) branched rays. Parhypural not fused to hypural 1. Hypurals 1 and 2 completely co-ossified into single ventral caudal plate, without any vestige of suture. Hypurals 3 and 4 completely fused to each other. Hypural 5 completely free, not fused to hypural 4. Epural single, rod-like, autogenous. Hypurapophysis and secondary hypurapophysis fused, forming a continuous horizontal shelf (complex hypurapophysis) to base of hypural 2 (hypurapophysis “type C” of Lundberg & Baskin, 1969). Dorsal hypural plate with 8 (10*) rays. Dorsal caudal rays arranged as follows on dorsal hypural plate: 6 rays on hypurals 3+4 and 2 rays on hypural 5 (10*). Caudal rays on dorsal plate with following branching pattern: i,7 (10*). Ventral caudal plate (parhypural plus hypurals 1+2) with 9 (10*) rays. Ventral caudal rays arranged as follows on ventral caudal plate: 2 rays on parhypural and 7 rays on hypurals 1+2 (10*). Caudal rays on ventral plate with following branching pattern: i,8 (10*) rays. Base of central caudal-fin rays (lowermost ray of dorsal caudal-fin lobe and uppermost ray of ventral caudal-fin lobe) not articulating directly to caudal plates, floating near diastema.
Total vertebrae 35 (3)–36 (7*). Ribs 7 (6)–8 (4*) pairs. Distal extremities of ribs tapered. First complete (i.e., not bifid distally) hemal spine on vertebrae 14 (7)–15 (3*).
Laterosensory system (Figure 2). Head sensory canals with simple (unbranched) tubes ending in single pores. Supraorbital sensory canal continuous and connected to otic and infraorbital sensory canals posteriorly and, usually, to infraorbital anteriorly. Supraorbital sensory canal at least with five branches: s1, s2, s3, s6 (epiphyseal branch), and s8 (parietal branch). Left and right epiphyseal branches (s6) never fused to each other medially. Presence of s4 branch and pore variable: present on both head sides in 3 specimens, present on one head side only in 3 (*) specimens, and absent on both head sides in 1 specimen. Presence of s7 branch and pore (postorbital) variable: present on both head sides in 2 specimens, present on one head side only in 3 specimens, and absent on both head sides in 2 (*) specimens. S5 branch and pore absent. Supraorbital and infraorbital sensory canals anteriorly connected to each other through s2 and i2 branches (forming complex s2+i2 pore), except in one c&s specimen. Otic sensory canal short, without pores, and continuous with posterior limits of supra- and infraorbital sensory canals, anteriorly, and with anterior limit of postotic sensory canal, posteriorly. Postotic (or temporal) sensory canal extends from posterior limit of otic sensory canal to anterior limit of lateral line, with 3 branches (po1, po2, and po3). First postotic branch (po1) fused to posteriormost branch preoperculomandibular sensory (pm11), forming po1+pm11 complex pore. Infraorbital sensory canal with six branches, with second branch (i2) usually fused to second supraorbital branch (s2) (see above). Preoperculomandibular sensory canal with 11 branches and pores; anteriormost preoperculomandibular sensory branch (pm1) not fused to its antimeric branch. Lateral-line sensory canal continuous with postotic sensory canal anteriorly and extending to base of caudal-fin rays. First lateral-line pore below level of adjacent pores of lateral line.
Color in alcohol. Background body coloration (Figure 1) yellowish dorsally; dorsal and lateral regions of body with sparsely distributed dark melanophores, slightly more concentrated along limits between myomeres; a large area of trunk above the pectoral fin (covering the pseudotympanum) with dense concentration of melanophores. Each side of body with a broad, dark brown dorsolateral stripe extending from just behind the eye to region immediately posterior to adipose-fin base. Paired dorsolateral stripes separated anteriorly by supraoccipital process, converging anteriorly along midline of the dorsum from the dorsal-fin origin, although the region lateral to dorsal-fin base is sometimes unpigmented.
Top of head with heavy concentration of dark melanophores. Head with broad tranverse dark stripe from its posterodorsal part to region below eye (forming conspicuous subocular blotch). Ventral region of body lacking pigmentation except for region between gape and insertion of mental barbels; pectoral-fin base and region immediately posterior to pelvic fins with few melanophores. Maxillary barbel with brown pigment along its dorsal
SLOBODIAN & BOCKMANN10 · Zootaxa 3717 (1) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
surface; mental barbels weakly pigmented, with scattered brown pigment along their dorsal surfaces. Sparse dark melanophores along all barbels.
Dorsal-fin spine and distal portion of first branched dorsal-fin ray heavily pigmented; remaining rays with scattered melanophores; interradial membranes hyaline. First ray of pectoral fin darkly pigmented, remaining rays with few scattered melanophores along basal portion. Rays of pelvic fin with few scattered melanophores along basal portion. Adipose fin has brown pigmentation on its basal half. Anal fin almost hyaline, with sparse pigment on bases of rays. Caudal fin with brown pigment along rays.
Geographic distribution. Brachyrhamdia thayeria is known to occur in the Rio Japurá basin, a left margin tributary of the Rio Solimões, in Brazil, especially in the Lago Amanã system (Figure 6).
FIGURE 6. Map of northern South America, showing distribution of Brachyrhamdia thayeria.
Ecological notes. Brachyrhamdia thayeria inhabits the upper and middle courses of both white and black-water streams (“igarapés”) affluent to Lago Amanã. Specimens were captured in the water column associated with marginal vegetation or along sandy beaches in shallow streams (about 1 m depth) with 0.20 mg/l of dissolved
oxygen and conductivity 27.6–57.1 µS/cm3 (Hercos et al., 2009).Etymology. The specific epithet thayeria refers to Thayeria Eigenmann (1908), a genus of South American
characin that has a similar diagonal dark stripe on the caudal peduncle, and also in honor of the North American businessman and philanthropist Nathaniel Thayer, Jr. (1808–1883), who sponsored the expedition to Brazil led bySwiss zoologist Louis Agassiz from 1865 to 1866 (then known as Thayer Expedition), which was responsible for important discoveries of the Amazon fish fauna.
Zootaxa 3717 (1) © 2013 Magnolia Press · 11A NEW SPECIES OF BRACHYRHAMDIA

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Comparisons and phylogenetic relationships of Brachyrhamdia thayeria
A comprehensive phylogenetic analysis of Pimelodella and allied genera is necessary to demonstrate the validity of the genus Brachyrhamdia, an issue beyond the scope of this article and to be treated elsewhere (Slobodian & Bockmann, in prep.). For now it is enough to say that species currently assigned to Brachyrhamdia constitute a monophyletic unit. One of the derived characters shared by all species of Brachyrhamdia, and originally recognized by Myers (1927), is the almost completely closed posterior cranial fontanel. As in other species of Brachyrhamdia (Lundberg & McDade, 1986; Bockmann, 1998; Slobodian, 2013; pers. obs.), the posterior cranial fontanel in B. thayeria is almost fully obliterated, except for a small anterior opening, which corresponds to region immediately posterior to the epiphyseal bar, and by a second tiny rounded opening at the base of the supraoccipital process (Figure 2). Among heptapterids, a mostly closed posterior cranial fontanel is also present in Rhamdia
quelen (Quoy & Gaimard, 1824), Rhamdia enfurnada Bichuette & Trajano, 2005, and the monotypic Taunayia
Miranda Ribeiro, 1918 (Silfvergrip, 1996; Bockmann, 1998; Bichuette & Trajano, 2005). However, these three taxa share a different state of closure because the lateral limits of posterior fontanel are still noticeable as a suture line along the midline of the supraoccipital (vs. medial region of supraoccipital solid in Brachyrhamdia); also, the posterior remnant of posterior fontanel at the supraoccipital is an ovoid opening and distinctly larger than in Brachyrhamdia.
FIGURE 7. Right pectoral spines of Brachyrhamdia species, dorsal view: A—Brachyrhamdia heteropleura, USNM 226105, 32.5 mm SL; B—Brachyrhamdia imitator, MCP 15132, 34.7 mm SL; C—Brachyrhamdia marthae, USNM 305631, 26.5 mm SL; D—Brachyrhamdia rambarrani, INPA 9188, 47.3 mm SL. Scale bars—1 mm.
Myers (1927) also mentioned “…pectoral spines…with thorns along the basal of the posterior edge” as a diagnostic feature for Brachyrhamdia, as opposed to the condition exhibited by Goeldiella Eigenmann & Norris (1900) and Rhamdella Eigenmann & Eigenmann (1888), in which the dentations along posterior edge of the pectoral spine extend considerably beyond its basal half, often approaching its tip (Bockmann, 1998; Bockmann &
SLOBODIAN & BOCKMANN12 · Zootaxa 3717 (1) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Miquelarena, 2008, Slobodian, 2013). All species of Brachyrhamdia (Figure 7), including B. thayeria (Figure 4), have the condition as described by Myers (1927) for B. imitator. Although some species of Pimelodella may have a condition similar to that found in the pectoral spine of Brachyrhamdia (Lundberg & McDade, 1986), such as in P.
eutaenia Regan, 1913 and Pimelodella hartii (Steindachner, 1877) (cf. Eigenmann, 1917: pl. 35, figs. 25 and 35), the condition in the vast majority of Pimelodella species resembles that present in Goeldiella and Rhamdella (cf.
Eigenmann, 1917; Bockmann & Slobodian, in press). Because there is no additional evidence supporting a close relationship between the aforementioned species of Pimelodella (i.e., P. eutaenia and P. hartii) and Brachyrhamdia, there is no reason to reject this feature as a potential synapomorphy for Brachyrhamdia
(Bockmann, 1998; Slobodian, 2013).The lack of published data on the phylogenetic relationships among species of Brachyrhamdia prevents more
categorical statements about the affinities of B. thayeria. For now, we have identified two putatively derived states that suggest close affinities between B. thayeria, B. imitator, and B. rambarrani, which are discussed below.
1) Low number of vertebrae (usually 35–36). Most members of the Heptapteridae have 37–45 total vertebrae (Bockmann, 1998; Bockmann & Miquelarena, 2008), which is the presumed plesiomorphic state. The generic name Brachyrhamdia was given in allusion to the compact aspect of the body of its type species, B. imitator
(Myers, 1927). Such a habitus is reflected in a low vertebral count for this species: 34 (3), 35 (6*), and 36 (1) (Bockmann, 1998; pers. obs.). Lundberg & McDade (1986) reported similar counts for B. imitator, also ranging from 34 to 36, most commonly 35–36 vertebrae. This likely derived state is shared by B. thayeria, which has 35 (3)–36 (7) vertebrae, and B. rambarrani, which has 35 (1), 36 (4), and 37 (2) (pers. obs.). On the other hand, B.
heteropleura, B. marthae, and B. meesi have a more slender aspect (Sands & Black, 1985a; pers. obs.), which is expressed by their higher number of vertebrae: B. heteropleura has 38 (4), B. marthae has 37 (3) to 39 (1), and B.
meesi has 37 (2), 38 (2), and 39 (2) (Bockmann, 1998; Slobodian, 2013; pers. obs.).2) Midlateral stripe absent. The genus Brachyrhamdia is closely related to, or perhaps nested within
Pimelodella (cf. Bockmann, 1998; Bockmann & Miquelarena, 2008; Slobodian, 2013), a genus whose species typically have a dark midlateral stripe (Eigenmann, 1917; Bockmann & Slobodian, in press). Bockmann & Miquelarena (2008) tentatively considered the absence of a midlateral stripe as a plesiomorphy for the family Heptapteridae; therefore its absence in Brachyrhamdia would be considered a secondarily derived trait if the genus is nested within Pimelodella. A dark midlateral stripe is distinctly present in B. marthae and B. meesi (Sands & Black, 1985; pers. obs.). Although the stripe of Brachyrhamdia heteropleura is narrow (cf. Eigenmann, 1912: pl. 16, fig. 2; Ferreira et al., 2007: 150, unnumb. fig.; pers. obs.), it is undoubtedly present and discrete. The absence of a midlateral stripe along the body of B. thayeria (Figure 1), B. imitator (Myers, 1927; Lundberg & McDade, 1986; pers. obs.), and B. rambarrani (Axelrod & Burgess in Axelrod, 1987; Figure 8) is interpreted as a synapomorphy.
Brachyrhamdia thayeria (Figure 1) and B. rambarrani (Figure 8) resemble one another in having bilaterally paired dark bands along the dorsal half of the trunk (Axelrod & Burgess in Axelrod, 1987: 24–25, unnumb. figs.), an apomorphic condition suggestive of a sister-group relationship. However, these dark bands differ between species in their position, and they may be questionably homologous. In B. thayeria the band originates just behind the eye and terminates on the caudal peduncle just posterior to the adipose-fin base (Figure 1), whereas in B.
rambarrani, the origin and end of the band are significantly more posterior, extending from below the dorsal-fin origin to the insertion region of the dorsal procorrent caudal rays (Figure 8).
Despite their resemblance, B. thayeria markedly differs from B. rambarrani in having a dark brown band crossing the peduncle diagonally from the region just below the middle of adipose-fin base to the base of ventral caudal-fin lobe (Figure 1), while that stripe is definitively absent in B. rambarrani (cf. Axelrod & Burgess in
Axelrod, 1987; Figure 8). In addition, the dorsal fin is mostly hyaline, with dark pigmentation restricted to the spine and first branched ray in B. thayeria (Figure 1); in contrast, the dorsal fin of B. rambarrani has a large, roughly triangular dark mark on its anterobasal region (cf. Axelrod & Burgess in Axelrod, 1987; Figure 8). Furthermore, B. thayeria differs from B. rambarrani by having a roughly triangular supraoccipital process, its wide base narrowing progressively towards the tip (Figure 2). That feature is also seen in B. imitator (cf. fig. 6 in Lundberg & McDade, 1986), B. marthae, and B. meesi (Slobodian, 2013). Alternatively in B. rambarrani, the supraoccipital process has a narrow base and keeps its width for most of its length, exhibiting a roughly rectangular aspect shared by B. heteropleura (Slobodian, 2013).
According to Bockmann & Miquelarena (2008), coding Brachyrhamdia for the presence or absence of pharyngobranchial 1 is equivocal, since it is present in B. marthae and B. meesi and absent in B. imitator. This bone
Zootaxa 3717 (1) © 2013 Magnolia Press · 13A NEW SPECIES OF BRACHYRHAMDIA

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
is plesiomorphically present in basal lineages of the Heptapteridae, such as Pimelodella spp., Rhamdia laticauda
(Kner, 1857), and R. quelen (Silfvergrip, 1996; Bockmann & Miquelarena, 2008). The pharyngobranchial 1 was found on both sides of the branchial basket in the single c&s specimen of B. thayeria (Figure 3), while it is absent in B. heteropleura and B. rambarrani (Bockmann, 1998; Slobodian, 2013; pers. obs.). The phylogenetic significance of this character awaits a comprehensive cladistic analysis of Brachyrhamdia and allied genera, which is beyond the scope of the present work (Slobodian & Bockmann, in prep.). For now, it remains a characteristic further distinguishing B. thayeria from B. rambarrani.
FIGURE 8. Brachyrhamdia rambarrani, USNM 204694, 53.1 mm SL, paratype of Pimelodella rambarrani, Brazil, Amazonas, Rio Unini (Rio Negro basin). Photo by Vitor Masson.
The ornamentation pattern of the pectoral spine appears to be conserved within each species of Brachyrhamdia. In general, the pectoral spine of B. thayeria (Figure 4) is more akin to that of B. heteropleura
(Figure 7A), B. imitator (Lundberg & McDade, 1986: fig. 2B; Figure 7B), and B. rambarrani (Figure 7D), and bears 7–10 retrorse dentations on the basal portion of its medial edge; those elements in B. heteropleura and B.
imitator appear to be larger than those of B. thayeria (Figure 4) and B. rambarrani (Figure 7D). On the other hand, the hooks along the medial margin of the pectoral spine in B. marthae (Figure 7C) and B. meesi are distinctly
SLOBODIAN & BOCKMANN14 · Zootaxa 3717 (1) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
smaller than those of congeners, and are oriented forward (antrorse) and perpendicularly, with some having bifid apices (Figure 7C), most markedly in B. marthae. The more attenuated pattern of pectoral spine hooks in B.
marthae and B. meesi when compared to that of B. imitator was recognized by Sands & Black (1985). A more complete treatment of the characteristics associated with the pectoral spine will be presented in a forthcoming paper.
Galvis et al. (2006) illustrated and provided a brief description of an unknown species of Brachyrhamdia
found in Leticia and surroundings, in Colombia, in terra firme streams affluent to the Laguna Yahuarcaca system (side arm of the Río Amazonas) and possibly in the Río Yavari (J.I. Mojica, pers. comm.). The unknown species closely resembles B. thayeria, especially in color pattern, and is known as ‘Brachyrhamdia sp. (1)’ among hobbyists (http://www.scotcat.com/thedarkone/heptapteridae/tdo_heptapteridae.htm) (S. Grant, pers. obs.). Brachyrhamdia thayeria differs from the Leticia species by having 7 branched pectoral-fin rays, a widespread count among species of Brachyrhamdia (Sands & Black, 1985; Axelrod & Burgess in Axelrod, 1987; Bockmann, 1998; Slobodian, 2013; pers. obs.), and 6 pelvic-fin rays (i, 5), a plesiomorphic count among siluriforms (Grande & Lundberg, 1988) [vs. 9 branched pectoral-fin rays and 7 (i,6) pelvic-fin rays as reported by Galvis et al. (2006)]. We refrain from treating B. thayeria and the Leticia species as conspecific until specimens of the latter are closely examined.
Biogeography of Brachyrhamdia thayeria
The region where Brachyrhamdia thayeria is known to occur, situated about 40–50 meters above sea level, is typical Amazonian lowland formed by Tertiary sediments, and characterized by plan terrains drained by low-speed white-waters (Lima & Ribeiro, 2011). However, within this region, B. thayeria seems to prefer small streams not prone to flooding (“terra firme” streams), having comparatively lower sediment loads and consequently clearer (black) waters, and is associated with marginal vegetation and stream bottoms of sand or silt (Hercos et al., 2009; Queiroz & Hercos, 2009). It is interesting to note that the Leticia species of Brachyrhamdia occurs in the Laguna Yahuarcaca system, part of the lowlands of the Colombian Amazon and inhabits small black-water streams with densely forested surroundings (cf. Galvis et al., 2006), similar to habitats found in the Lago Amanã system.
Although most species of Brachyrhamdia occur in the northern lowlands of South America, B. heteropleura, B. rambarrani, and an undescribed species from the Rio Tocantins basin inhabit rivers draining ancient highland, shield areas (Lima & Ribeiro, 2011). These species occur in watersheds on ancient basaltic basements that possibly suffered subsidence and were overlain by sediments, thus acquiring characteristics of lowlands and supporting a mixed highland/lowland fish fauna. For example, Brachyrhamdia heteropleura occurs in the Branco (Amazonas basin) and Essequibo (Atlantic coastal basin) draining the Takutu graben, a tectonic block that sunk, allowing the formation of a continuous sedimentary basin between the two basins (Crawford et al., 1985; Milani & Thomaz-Filho, 2000; Lima & Ribeiro, 2011; Lujan & Armbruster, 2011). Brachyrhamdia rambarrani is reportedly endemic to middle and lower reaches of the Rio Negro basin, below São Gabriel da Cachoeira, which bear characteristics of Amazonian lowlands (Latrubesse & Franzinelli, 2005; Lima & Ribeiro, 2011). The undescribed species endemic to the Araguaia-Tocantins is in a system which settled on the tectonically-developed Araguaia and Tocantins depressions (Saadi et al., 2005; Lima & Ribeiro, 2011).
The genus Brachyrhamdia, as presently known, is distributed throughout the Amazon system (including the Rio Tocantins basin), part of the Orinoco River basin, and in the Essequibo and Corantijn river basins (Bockmann & Guazzelli, 2003; Slobodian, 2013). The distributional range of each species is well demarcated and non-overlapping (Slobodian, 2013). Taking into account the broad distribution of the genus, the disjunctive distribution patterns of its species and the fact that they are weakly vagile, the present-day distribution of Brachyrhamdia is attributed predominantly to extrinsic causes, indicating that lowlands of northern South America, although recent in age, are geomorphologically dynamic.
Commentaries on mimicry in Brachyrhamdia
Species of Brachyrhamdia draw attention because, unlike most species of Heptapteridae which are relatively uniformly colored, they exhibit remarkably different patterns of body pigmentation. Interestingly, at least some
Zootaxa 3717 (1) © 2013 Magnolia Press · 15A NEW SPECIES OF BRACHYRHAMDIA

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
species of Brachyrhamdia occur syntopically with other catfishes, especially with members of the callichthyid genus Corydoras, with which they share a striking resemblance in overall body shape and color pattern (Myers, 1927; Innes & Myers, 1950; Sands, 1984, 1985b; Sands & Black, 1985; Lundberg & McDade, 1986; Axelrod & Burgess in Axelrod, 1987). Brachyrhamdia thayeria has been collected together both with C. arcuatus and C. cf. nanus Nijssen & Isbrücker (1967) (A. P. Hercos, pers. comm.; L.M. Sousa, pers. comm.). Corydoras arcuatus, which occurs in the Upper Rio Amazonas basin, in Brazil, Colombia, Ecuador, and Peru (Reis, 2003), shares a striking resemblance with B. thayeria, including the dorsolateral dark band and the oblique stripe across the caudal peduncle (Elwin, 1939). The species of Brachyrhamdia from the region of Leticia, Colombia, also has been reported to occur in association with C. arcuatus (Galvis et al., 2006; J.I. Mojica, pers. comm.). Brachyrhamdia
imitator is usually caught together with Corydoras delphax Nijssen & Isbrücker, 1983 (Sands, 1984), a species endemic to Río Inírida and Upper Río Orinoco basins of Colombia (Reis, 2003) that is commonly misidentified as Corydoras melanistius Regan (1912) (Myers, 1927; Innes & Myers, 1950; Sands, 1980; Lundberg & McDade, 1986). Brachyrhamdia imitator and Corydoras delphax bear a striking resemblance, both having a stout body, finely mottled color pattern on trunk, dark mask across the eye, and dark region covering the anterodorsal portion of trunk and the anterior portion of dorsal fin (Myers, 1927; Sands, 1980; Nijssen & Isbrücker, 1983; Lundberg & McDade, 1986). Specimens of Brachyrhamdia marthae, possibly from Peru, are imported alongside Corydoras
pygmaeus Knaack, 1966 (Sands, 1985a; Sands & Black, 1985). In the Rio Madeira basin of Brazil, B. marthae is collected in association with shoals of Corydoras caudimaculatus Rössel (1961) and Corydoras trilineatus Cope (1872), sharing with the former a dark mark at base of caudal peduncle, and with the latter the presence of a dark lateral stripe (Bockmann & Slobodian, in press). Brachyrhamdia meesi is reportedly imported together with Pimelodella sp. (Sands, 1985b; Sands & Black, 1985), both species possessing a dark midlateral stripe. To our knowledge, there is no published information on the co-occurrence of B. rambarrani with species of Corydoras. There are, however, three species of Corydoras endemic to the Rio Negro that display the same color pattern as B.
rambarrani (Reis, 2003): C. adolfoi Burgess, 1982; C. duplicareus Sands, 1995; and C. serratus Sands, 1995. In the Rio Araguaia basin, Brazil, an undescribed species of Brachyrhamdia commonly occurs in close association with Corydoras araguaiaensis Sands (1990) (F.C.T. Lima, pers. comm.), a species endemic to that river basin (Reis, 2003). Both species are roughly similar in having narrow dark stripes along the trunk, formed by coalesced dots in the case of C. araguaiaensis (Sands, 1990). There are no records of Brachyrhamdia heteropleura being sighted or collected consistently with other catfishes.
Another morphological trait that allows species Brachyrhamdia to blend into shoals of Corydoras is their small and comparable size. Although adult individuals Brachyrhamdia are not as small as the heptapterids Gladioglanis
Ferraris & Mago-Leccia, 1989, Nemuroglanis Eigenmann & Eigenmann, 1988, and Pariolius Cope, 1872 (Bockmann & Guazzelli, 2003; Bockmann & Ferraris, 2005), they are significantly smaller than species of Pimelodella, a genus putatively closely related to Brachyrhamdia. Brachyrhamdia marthae and B. meesi are possibly the largest species of Brachyrhamdia, reaching 6–7 cm SL; other species of the genus are not longer than 5.5 cm SL (Bockmann & Guazzelli, 2003). On the other hand, adults of most species of Pimelodella are consistently longer than 9 cm SL, sometimes reaching more than 30 cm SL (Bockmann & Guazzelli, 2003). The maximum size of species of Corydoras ranges from 2.3 to 7.5 cm SL, and is usually 3.5–6 cm SL (Reis, 2003).
Because species of Brachyrhamdia display color patterns that are unusual for its family, but common among ecologically associated catfishes (especially Corydoras), it is assumed that these relationships involve mimicry, with species of Brachyrhamdia playing the mimics and the other catfishes being the models. Mimicry among South American catfishes is mostly based on anecdotal observations (Innes & Myers, 1950; Sands, 1980; Sands, 1985b; Sands & Black, 1985; Lundberg & McDade, 1986; Axelrod & Burgess in Axelrod, 1987), although a few studies have investigated mimicry more thoroughly (Axenrot & Kullander, 2003; Alexandrou et al., 2011).
The exact benefits of the interactions among Brachyrhamdia and Corydoras and Brachyrhamdia and Pimelodella are yet to be established through field observations and under experimental conditions. It is speculated that it is a type of defensive mimicry complex inasmuch as all members share the defensive tripod formed by pungent spines in the dorsal and paired pectoral fins, that increase their effective size when erected, making it significantly more harmful to predators (Fine & Ladich, 2003). Sands (1980) noticed that, in aquarium conditions, individuals of Brachyrhamdia do not shoal with Corydoras except when feeding or stressed. Species of Corydoras
are further protected by bony body armor, composed of two longitudinal series of dermal plates (Reis, 1998). The distinctive color patterns of both Brachyrhamdia and Corydoras do not appear to give to them any cryptic
SLOBODIAN & BOCKMANN16 · Zootaxa 3717 (1) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
advantage. Instead, the color patterns seems to produce a disruptive effect against the uniformly plain backgrounds represented by bottoms of silt and sand in small tributaries with clear, shallow waters (Lundberg & McDade, 1986; Ferreira et al., 2007; Hercos et al., 2009; Queiroz & Hercos, 2009; F.C.T. Lima, pers. comm.).
The most common cases of defensive mimicry fit into categories of either Batesian or Müllerian mimicry. In Batesian mimicry, the mimic imitates the warning signals of the model but does not have the attributes that make it unprofitable (i.e., only the mimic benefits). In Müllerian mimicry, the mimic also imitates the warning signals of the model, but both have anti-predation attributes, setting a mutual benefit to the interaction (Remington, 1963; Wickler, 1968; Mallet & Joron, 1999). Although only Corydoras has body armor, both Brachyrhamdia and Corydoras are equipped with the defensive tripod. Therefore, the association between those taxa best fits the category of Müllerian mimicry.
On the other hand, individuals of Brachyrhamdia are usually fewer in number than its models (Sands, 1980; Galvis et al., 2006; J.I. Mojica, S. Grant; pers. comm.), which suggests Batesian mimicry. In this kind of interaction, mimics occur in lower proportions than the models since the presence of harmless mimics weakens the discouraging effect to occasional experimental attackers (Wickler, 1968; Huheey, 1988). However, the scarcity of individuals of Brachyrhamdia may be the product of a plesiomorphic behavior as heptapterids tend to be solitary or to shoal in small groups of up to 10 individuals (Bockmann & Guazzelli, 2003).
Populations involved in Batesian mimicry often produce a stable polymorphism, which is explained by frequency-dependent selection (Huheey, 1988; Mallet & Joron, 1999). Each species of Brachyrhamdia examined, including B. thayeria, is highly conserved in morphological terms, showing no evidence of polymorphism. This finding strengthens the hypothesis that the association between Brachyrhamdia and other catfishes, especially Corydoras, is Müllerian mimicry.
Alexandrou et al. (2011) reported shoaling of phylogenetically distant species of Corydoras displaying convergently evolved color patterns. Such Müllerian co-mimicry is maintained by distinct occupation of trophic niches, as is expressed by species-specific traits linked to resource acquisition (Alexandrou et al., 2011). Trophic niche partitioning may similarly occur between species of Brachyrhamdia and Corydoras in a mimicry complex; however, there is currently insufficient data on the food preferences of Brachyrhamdia to test this hypothesis. An alternative hypothesis is that by occurring naturally at low densities, species of Brachyrhamdia do not compete for food resources with their Corydoras models.
Acknowledgements
Special words of gratitude are addressed to L. Sousa, who drew our attention to this new species, and M. Sabaj Pérez, who did a particularly thorough review of the manuscript. We are grateful to the following people for providing assistance during visits and other facilities at their institutions, and for loaning material: R. Arrindell, M. Britto, H. Britski, P. Buckup, R. Castro, D. Catania, W. Costa, W. Crampton, C. Doria, J. de Figueiredo, C. Lucena, Z. Lucena, J. Lundberg, W. Ohara, R. Oliveira, O. Oyakawa, L. Page, L. Parenti, M. de Pinna, L. Rapp Py-Daniel, R. Reis, R. de Ruiter, M. Sabaj Pérez, S. Schaefer, and R. Vari. The following colleagues kindly shared with us their knowledge and helped us in getting literature: S. Grant, A. Hercos, J. Horne, S. Kullander, F. Lima, J. Mojica, W. Ohara, and L. Sousa. We are also indebted to Ricardo Castro who lent the photographic equipment and guided us in obtaining the photograph of the holotype. Marcelo Carvalho helped us with editorial issues for which we are very grateful. Research funding was provided by Conselho Nacional de Desenvolvimento Científico e Tecnológico (PROTAX/156515/2010-5 to VS, PROTAX Proc. No. 562268/2010-3 and Proc. No. 309752/2010-8 to FAB). Visit to ANSP by the first author was partially supported by the Böhlke Memorial Endowment Fund; additional support also provided by the All Catfish Species Inventory (NSF DEB-0315963). We also thank to Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) for providing funding for strengthening of conservation and research of scientific collections housed at Department of Biology of the Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto–USP (Proc. No. 2009/54931-0). This article is a result of the dissertation of VS, presented as part of requirements to obtain the Master’s degree for the Graduate Program in Comparative Biology, FFCLRP–USP, Brazil.
Zootaxa 3717 (1) © 2013 Magnolia Press · 17A NEW SPECIES OF BRACHYRHAMDIA

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
References
Alexandrou, M.A., Oliveira, C., Maillard, M., McGill, R.A.R., Newton, J., Creer, S. & Taylor, M.I. (2011) Competition and phylogeny determine community structure in Müllerian co-mimics. Nature, 469, 249–272. http://dx.doi.org/10.1038/nature09660
Arratia, G. & Huaquín, L. (1995) Morphology of the lateral line system and of the skin of diplomystid and certain primitive loricarioid catfishes and systematic and ecological considerations. Bonner Zoologische Monographien, 36, 1–110.
Axelrod, H.R. (1987) Two new species of catfishes (Siluriformes, Callichthyidae and Pimelodidae) from the Rio Unini, Amazonas, Brazil. Tropical Fish Hobbyist, 35, 22–25.
Axenrot, T.E. & Kullander, S.O. (2003) Corydoras diphyes (Siluriformes: Callichthyidae) and Otocinclus mimulus(Siluriformes: Loricariidae), two new species of catfishes from Paraguay, a case of mimetic association. Ichthyological Exploration of Freshwaters, 14, 249–272.
Bichuette, M.E. & Trajano, E. (2005) A new cave species of Rhamdia (Siluriformes: Heptapteridae) from Serra do Ramalho, northeastern Brazil, with notes on ecology and behavior. Neotropical Ichthyology, 3, 587–595. http://dx.doi.org/10.1590/s1679-62252005000400016
Bockmann, F.A. (1998) Análise Filogenética da Família Heptapteridae (Teleostei, Ostariophysi, Siluriformes) e Redefinição de seus Gêneros. Unpublished Ph.D. thesis, Universidade de São Paulo, São Paulo, Brazil, 599 pp.
Bockmann, F.A. & Castro, R.M.C. (2010) The blind catfish from the caves of Chapada Diamantina, State of Bahia, northeastern Brazil (Siluriformes: Heptapteridae): description, anatomy, phylogenetic relationships, biology, and biogeography. Neotropical Ichthyology, 8, 673–706. http://dx.doi.org/10.1590/S1679-62252010000400001
Bockmann, F.A. & Ferraris, C.J. Jr. (2005) Systematics of the Neotropical catfish genera Nemuroglanis Eigenmann and Eigenmann 1889, Imparales Schultz 1944, and Medemichthys Dahl 1961 (Siluriformes: Heptapteridae). Copeia, 2005, 124–137. http://dx.doi.org/10.1643/ci-04-019r1
Bockmann, F.A. & Guazzelli, G.M. (2003) Family Heptapteridae (Heptapterids). In: Reis, R.E., Kullander, S.O. & Ferraris, C.J. Jr. (Eds.), Check List of the Freshwater Fishes of South and Central America. Edipucrs, Porto Alegre, Brazil, pp. 406–431.
Bockmann, F.A. & Miquelarena, A.M. (2008) Anatomy and phylogenetic relationships of a new catfish species from northeastern Argentina with comments on the phylogenetic relationships of the genus Rhamdella Eigenmann and Eigenmann 1888 (Siluriformes, Heptapteridae). Zootaxa, 1780, 1–54.
Bockmann, F.A. & de Pinna, M.C.C. (2004) Heptapterus collettii Steindachner, 1881: a member of the Asian bagrid genus Olyra erroneously assigned to the Neotropical Fauna (Siluriformes: Ostariophysi). Copeia, 2004, 665–675. http://dx.doi.org/10.1643/ci-03-198r1
Bockmann, F.A. & Slobodian, V. (in press) Heptapteridae. In: Queiroz, L.J., Torrente-Vilara, G., Ohara, W.M., Silva, T.H.P., Zuanon, J. & Doria, C.R.C. (Eds.), Peixes do Rio Madeira. Vol. 3. Dialeto, São Paulo, Brazil, pp. 12–71.
Carvalho, M., Bockmann, F.A. & Carvalho, M.R. (2013) Homology of the fifth epibranchial and accessory elements of the ceratobranchials among gnathostomes: insights from the development of ostariophysans. PLoS ONE, 8, e62389. http://dx.doi.org/10.1371/journal.pone.0062389
Crawford, F.D., Szelewski, C.E. & Alvey, G.D. (1985) Geology and exploration in the Takutu graben of Guyana and Brazil. Journal of Petroleum Geology, 8, 5–36. http://dx.doi.org/10.1111/j.1747-5457.1985.tb00189.x
Eigenmann, C.H. (1912) The freshwater fishes of British Guiana, including a study of the ecological grouping of species, and the relation of the fauna of the plateau to that of the lowlands. Memoirs of the Carnegie Museum, 5, i–xxii + 1–578, pls. 1–103. http://dx.doi.org/10.5962/bhl.title.4686
Eigenmann, C.H. (1917) Pimelodella and Typhlobagrus. Memoirs of the Carnegie Museum, 7, 229–258, pls. 29–35.Elwin, M.G. (1939) Corydoras arcuatus, sp. n., an Amazonian catfish. Annals and Magazine of Natural History (Series 11), 3,
126–128, pl. 3. http://dx.doi.org/10.1080/03745481.1939.9723581
Ferraris, C.J. Jr. (2007) Checklist of catfishes, recent and fossil (Osteichthyes: Siluriformes), and catalogue of siluriform primary types. Zootaxa, 1418, 1–628.
Ferreira, E., Zuanon, J., Forsber, B., Goulding, M. & Briglia-Ferreira, S.R. (2007) Rio Branco. Peixes, Ecologia e Conservação de Roraima. Wust Ediciones, Lima, Peru, 201 pp.
Fine, M.L. & Ladich, F. (2003) Sound production, spine locking, and related adaptations. In: Arratia, G., Kapoor, B.G., Chardon, M. & Diogo, R. (Eds.), Catfishes. Science Publishers, Enfield, pp. 249–290.
Galvis, G., Mojica, J.I., Duque, S.R., Castellanos, C., Sánchez-Duarte, P., Arce, M., Gutiérrez, Á., Jiménez, L.F., Santos, M., Vejarano, S., Arbeláez, F., Prieto, E. & Leiva, M. (2006) Peces de Medio Amazonas. Región de Leticia. Conservación International, Editora Panamericana, Formas e Impresos, Bogotá, D.C., Colombia, 548 pp.
Grande, L. & Lundberg, J.G. (1988) Revision and redescription of the genus †Astephus (Siluriformes: Ictaluridae) with a discussion of its phylogenetic relationships. Journal of Vertebrate Paleontology, 8, 139–171. http://dx.doi.org/10.1080/02724634.1988.10011694
Hercos, A.P., Queiroz, H.L. & Almeida, H.L. (2009) Peixes Ornamentais do Amanã. Instituto de Desenvolvimento Sustentável Mamirauá, Tefé, Brazil, 241 pp.
SLOBODIAN & BOCKMANN18 · Zootaxa 3717 (1) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Huheey, J.E. (1988) Mathematical models of mimicry. American Naturalist, 131 (suppl.), 7–21. http://dx.doi.org/10.1086/284765
Innes, W.T. & Myers, G.S. (1950) The “Imitator catfish,” which mimics a Corydoras. The Aquarium, 19, 222–223.Latrubesse, E.M. & Franzinelli, E. (2005) The Late Quaternary evolution of the Negro River, Amazon, Brazil: implications for
island and floodplain formation in large anabranching tropical systems. Geomorphology, 70, 372–397. http://dx.doi.org/10.1016/j.geomorph.2005.02.014
Lima, F.C.T & Ribeiro, A.C. (2011) Continental-scale tectonic controls of biogeography and ecology. In: Albert, J.S. & Reis, R.E. (Eds.), Historical Biogeography of Neotropical Freshwater Fishes. University of California Press, Berkeley, U.S.A., pp. 145–164.
Lujan, N.K. & Armbruster, J.W. (2011) The Guyana shield. In: Albert, J.S. & Reis, R.E. (Eds.), Historical Biogeography of Neotropical Freshwater Fishes. University of California Press, Berkeley, U.S.A., pp. 211–224.
Lundberg, J.G. & Baskin, J.N. (1969) The caudal skeleton of the catfishes, order Siluriformes. American Museum Novitates, no. 2398, 1–49.
Lundberg, J.G. & McDade, L.A. (1986) On the South American catfish Brachyrhamdia imitator Myers (Siluriformes, Pimelodidae), with a phylogenetic evidence for a large intrafamilial lineage. Notulae Naturae (Philadelphia), no. 463, 1–24.
Mallet, J. & Joron, M. (1999) Evolution of diversity in warning color and mimicry: polymorphisms, shifting balance, and speciation. Annual Review of Ecology and Systematics, 30, 201–233. http://dx.doi.org/10.1146/annurev.ecolsys.30.1.201
Milani, E.J. & Thomaz-Filho, A. (2000) Sedimentary basins of South America. In: Cordani, U.G., Milani, E.J. & Campos, D.A. (Eds.), Tectonic Evolution of South America. Academia Brasileira de Ciências/Departamento Nacional de Produção Mineral, Rio de Janeiro, Brazil, pp. 389–449.
Myers, G.S. (1927) Descriptions of new South American freshwater-fishes collected by Dr. Carl Ternetz. Bulletin of the Museum of Comparative Zoology, 68, 107–135.
Nijssen, H. & Isbrücker, I.J.H. (1983) Review of the genus Corydoras from Colombia, with descriptions of two new species (Pisces, Siluriformes, Callichthyidae). Beaufortia, 33, 53–71.
Northcutt, G. (1989) The phylogenetic distribution and innervation of craniate mechanoreceptive lateral line: neurobiology and evolution. In: Coombs, S.P., Görner, P. & Münz, H. (Eds.), The Mechanosensory Lateral Line. Springer Verlag, New York, pp. 17–78.
Queiroz, H.L. & Hercos, A.P. (Orgs.) (2009) Plano de Manejo das Áreas de Coleta de Peixes Ornamentais da Reserva Amaná (PORA). Instituto de Desenvolvimento Sustentável Mamirauá/Ministério da Ciência e Tecnologia; Zoological Society of London, Tefé, Brazil, 89 pp.
Reis, R.E. (1998) Anatomy and phylogenetic analysis of the neotropical callichthyid catfishes (Ostariophysi, Siluriformes). Zoological Journal of the Linnean Society, 124, 105–168. http://dx.doi.org/10.1111/j.1096-3642.1998.tb00571.x
Reis, R.E. (2003) Family Callichthyidae (Armored catfishes). In: Reis, R.E., Kullander, S.O. & Ferraris, C.J. Jr. (Eds.), Check List of the Freshwater Fishes of South and Central America. Edipucrs, Porto Alegre, Brazil, pp. 291–309.
Remington, C.L. (1963) Historical backgrounds of mimicry. Proceedings of the International Congress of Zoology, 16, 145–149.
Saadi, A., Bezerra, F.H.R., Costa, R.D., Igreja, H.L.S. & Franzinelli, E. (2005) Neotectônica da Plataforma Brasileira. In:Souza, C.R.G., Suguio, K., Oliveira, A.M.S. & Oliveira, P.E. (Eds.), Quaternário do Brasil. Holos Editora, Ribeirão Preto, Brazil, pp. 211–234.
Sabaj Pérez, M.H. (Ed.) (2013) Standard Symbolic Codes For Institutional Resource Collections in Herpetology and Ichthyology: An Online Reference. Version 4.0 (28 June 2013). Society of Ichthyologists and Herpetologists, Washington, D.C., 56 pp. Available from: http://www.asih.org/sites/default/files/documents/resources/symbolic_codes_for_ collections_v4.0_sabajperez_2013.pdf (Accessed 28 July 2013)
Sands, D.D. (1980) Corydoras, or are they? Notes on colour pattern sharing. Aquarist and Pondkeeper, 45, 40–42.Sands, D.D. (1984) Catfishes of the World. Vol. 3. Dunure Enterprises, Dunure, 145 pp.Sands, D.D. (1985a) Brachyrhamdia meesi, Sands and Black 1985. In: Sands, D. (Ed.), Catfishes of the World. Vol. 3, Suppl.
(first set). Self published, pp. 58b.Sands, D.D. (1985b) Brachyrhamdia, cryptic or mimetic catfishes from South America. Zoomimesis, camouflage or mimicry.
In: Sands, D. (Ed.), Catfishes of the World. Vol. 3, Suppl. (first set). Self published, pp. 58(9)–58(11).Sands, D.D. (1990) A new species of callichthyid catfish, Corydoras araguaiaensis (Siluriformes, Callichthyidae) from the Rio
Araguaia, Brazil. In: Sands, D. (Ed.), Catfishes of the World. Suppl. Self published, pp. 1–2.Sands, D.D. & Black, B.K. (1985) Two new species of Brachyrhamdia, Myers, 1927, from Brazil and Peru, together with a
redefinition of the genus. In: Sands, D. (Ed.), Catfishes of the World. Vol. 3, Suppl. (first set). Self published, pp. 58(1)–58(8).
Schultz, L.P. (1944) The catfishes of Venezuela, with descriptions of thirty-eight new forms. Proceedings of the United States National Museum, 94, 173–338, pls. 1–14. http://dx.doi.org/10.5479/si.00963801.94-3172.173
Silfvergrip, A.M.C. (1996) A Systematic Revision of the Neotropical Catfish Genus Rhamdia (Teleostei, Pimelodidae). Stockholm University, Stockholm, 156 pp., 8 pls.
Zootaxa 3717 (1) © 2013 Magnolia Press · 19A NEW SPECIES OF BRACHYRHAMDIA

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Slobodian, V. (2013) Taxonomia, Sistemática e Biogeografia de Brachyrhamdia Myers, 1927 (Siluriformes: Heptapteridae), com uma investigação sobre seu mimetismo com outros Siluriformes. Unpublished Master diss., Universidade de São Paulo, Ribeirão Preto, Brazil.
Slobodian, V. & Bockmann, F.A. (2011) The catfish genus Brachyrhamdia Myers, 1927 (Pisces: Siluriformes: Heptapteridae): a study case of the shield distribution pattern in northern South America. In: Abstracts from XXX Annual Meeting of the Willi Hennig Society Meeting. São José do Rio Preto, Brazil, pp. 192.
Slobodian, V. & Bockmann, F.A. (2013) Taxonomy, systematics and biogeography of Brachyrhamdia Myers, 1927 (Siluriformes: Heptapteridae) and its mimetism with other siluriforms. In: 2013 Joint Meeting of Ichthyologists and Herpetologists, Albuquerque, New Mexico, 10-15 July, 2013, Abstract. University of New Mexico, Albuquerque, pp. 462.
Slobodian, V., Bockmann, F.A. & Sousa, L.M. (2011) Uma nova espécie do gênero Brachyrhamdia Myers, 1927 (Siluriformes: Heptapteridae) do Rio Japurá, Estado do Amazonas. In: Ferreira, E.J.G., Amadio, S.A., Silva, C.P.D. & Porto, J.I.R. (Eds.), Livro de Programação e Resumos, XIX Encontro Brasileiro de Ictiologia. INPA, AIA, SBI, Manaus, Brazil, pp. 970.
Taylor, W.R. & Van Dyke, G.C. (1985) Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium, 9, 107–119.
Wickler, W. (1968) Mimicry in Plant and Animals. World University Press, Weidenfeld & Nicolson, London, 255 pp.Zarske, A. (2003) Wiederbeschreibung von Rhamdia marthae (Sands & Black, 1985) (Teleostei, Siluriformes, Pimelodidae).
Zoologische Abhandlungen, Staatliches Museum für Tierkunde in Dresden, 53, 47–55.
APPENDIX. Comparative material is listed below. Specimens are preserved in ethyl alcohol, except when noticed
otherwise.
Brachyrhamdia heteropleura: AMNH 52410, 35, 2 c&s, 30.8–43.3 mm SL, Suriname, Nickerie district, Mawatai Creek, Small inlets on sand bar opposite entrance; AMNH 54970, 1, 33.1 mm SL, Suriname, Nickerie district, small stream south of Tiger Falls on Corantijn River, km 405; ANSP 179737, 20, 25.0–39.6 mm SL, Guyana, Essequibo District, Rupununi River, sand beach and inlet at Karanambo; INPA 7906, 2, Brazil, Roraima, Bonfim, Igarapé do Arraia, km 114 on Boa Vista-Bonfim road; LIRP 7415, 3, 33.6–37.6 mm SL, Brazil, Roraima, Mucajaí, Rio Mucajaí [Rio Branco basin]; LIRP 7419, 6, 30.5–35.3, Brazil, Roraima, Boa Vista, Rio Cauamé, near waterfall of Igarapé Au-Au [Rio Branco basin]; USNM 226105, 13, 2 c&s, 29.3–36 mm SL, Suriname, Nickerie District, unnamed tributary to Sisa Creek, north side, approximately 700m downstream of crossing of Amotopo to Camp Geologie Road.
Brachyrhamdia imitator: CAS 54359, 1, xy, 69.4 mm SL, holotype, Venezuela, Bolivar, Río Orinoco system, Caño de Quiribana, near Caicara; AMNH 58322, 3, 36.1–76.7 mm SL, Ecuador, aquarium trade; ANSP 135585, 337, 26–54 mm SL, Venezuela, Bolívar, Caño Puerto Cabello at Puerto Cabello; ANSP 135601, 20, 30–38.5 mm SL, Venezuela, Bolívar, Caño Chuapo ca. 20 min downstream from Jabillal [opposite bank] on Río Caura; ANSP 135635, 1, 30 mm SL, Venezuela, Bolívar, Caño Barranca ca. 1 1/4 hours downstream from Jabillal [opposite bank] on Río Caura; ANSP 135740, 1, 31.9 mm SL, Venezuela, Bolívar, Río Urbana [Urbani] on Maripa-Las Trincheras road; ANSP 135867, 1, 39.2 mm SL, Venezuela, Bolívar, Río Caura at Jabillal; ANSP 139617, 2, 30.6–30.8 mm SL, Venezuela, Bolívar, sandbar along Río Mato; ANSP 139850, 100, 2 c&s, 28–47 mm SL, Venezuela, Bolívar, small [caño] tributary of Río Mato [left bank]; ANSP 160248, 100, 28.4–42.1 mm SL, Venezuela, Bolívar, Río Caura at puerto Las Majadas; ANSP 160457, 1, 31.4 mm SL, Venezuela, Bolívar, Caño Puerto Cabello, at crossing of road to Las Trincheras, 34km S of Ciudad Bolívar-Caicara highway; ANSP 160525, 185, 27.7–41.8 mm SL, Venezuela, Bolívar, Río Caura at puerto Las Majadas [fishing village]; MCP 15132, 8, 1 c&s, 34.7–39.9 mm SL, Venezuela, Bolívar, Río Caura at puerto Las Majadas; MZUSP 84669, 4, 33.5–36.2 mm SL, Venezuela, Bolívar, Río Caura at puerto Las Majadas; SU 16179, 2, 40.7 mm SL, Venezuela, Bolívar, Caño de Quiribana into Río Orinoco; SU 48666, 1, Venezuela, Bolívar, Caño de Quiribana.
Brachyrhamdia marthae: RMNH 29424, 1, xy, 76 mm SL, holotype, Peru, aquarium import; AMNH 225416, 1, 27.2 mm SL, Bolivia, Santa Cruz, Río Ichilo, ca. 54km south of Boca Chapare; AMNH 39747, 1, Bolivia, Beni, Río Itenez, 2km SE of Costa Marques, in Brazil; AMNH 77576, 4, 22.6–26.8 mm SL, Bolivia, Beni, Río Mamoré, ca. 5km Southeast of Limoquije; UF 128932, 1, 24 mm SL, Peru, Loreto, Caño Yarina, affluent of Río Pacaya, in Pacaya-Samiria National Reserve; UF 128977, 5, 23.3–33 mm SL, Peru, Loreto, Caño Yarina, affluent of Río Pacaya, in Pacaya-Samiria National Reserve; UF 129036, 2, 29–32.5 mm SL, Peru, Loreto, Caño Yarina, affluent of Río Pacaya, in Pacaya-Samiria National Reserve; MCP 15555, 3, 26.5–28.0 mm SL, Brazil, Mato Grosso, Pontes e Lacerda, Rio Guaporé and flooded area in Pontes e Lacerda [Rio Madeira basin]; MZUSP 30771, 3, 32.8–45.1 mm SL, Brazil, Acre, Tarauacá, Rio Tarauacá [Rio Juruá basin]; LIRP 10040, 30, 23.6–32.9 mm SL, Brazil, Rondônia, Corumbiara, unnamed igarapé at Estrada dos Bois, affluent of Rio Corumbiara; UFRO-I 3277, 1, 25.6 mm SL, Brazil, Rondônia, Guarajá-mirim, Rio Pacaás Novos, near to confluence with Rio Mamoré; USNM 305631, 15, 2 c&s, 26.5–33.2 mm SL, Bolivia, Departamento del Beni, Provincia General José Ballivia Segurola, borrow pit by road at 1.5km W, Río Matos crossing 45 air km E San Borja; USNM 305864, 40, 2 c&s, 34.1–42.2 mm SL, Bolivia, Departamento del Beni, E. B. B. Trapiche La Pascana camp, ancient path of Río Maniqui; USNM 305891, 3, Bolivia, Departamento del Beni, E.B.B., Trapiche La Pascana camp, ancient path of Río Maniqui.
Brachyrhamdia meesi: RMNH 29425, 1, xy, 56.5 mm SL, holotype, Brazil, near Belém; INPA 8047, 1, xy, 1 c&s, 43.37 mm SL, Brazil, Amazonas, Manaus, unnamed igarapé near to the road to pisciculture, km 21 of AM-010 highway; MZUSP
SLOBODIAN & BOCKMANN20 · Zootaxa 3717 (1) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
87209, 1, xy, 57.7 mm SL, aquarium trade; UFRJ 0397, 2 c&s, 42.8–45.2 mm SL, aquarium trade; LIRP 9079, 1 c&s, aquarium trade.
Brachyrhamdia rambarrani: MZUSP 37693, 1, xy, 55 mm SL, holotype, Brazil, Amazonas, Rio Unini, tributary of Rio Negro; USNM 288460, 1, xy, 53 mm SL, paratype, Brazil, Amazonas, Rio Unini, tributary of Rio Negro; INPA 9188, 5, xr, 1 c&s, 47.3–59.4 mm SL, Brazil, Amazonas, São Gabriel da Cachoeira, Igarapé Iazinho, Morro Seis Lagos, below bridge crossing São Gabriel-Cucuí road.
Brachyrhamdia sp. (Araguaia-Tocantins): INPA 7949, 4, 1 c&s, 46.2–59 mm SL, Brazil, Pará, Tucuruí, Rio Tocantins, about 50 km upriver from dam of hydroelectric plant of Tucuruí [UHE Tucuruí]; INPA 21492, 2, 24.3–25.0 mm SL, Brazil, Tocantins, Rio Araguaia, at beach near to hotel, in State Park of Cantão [Parque Estadual de Cantão]; MZUSP 4941, 4, 1 c&s, 35.4–49.9 mm SL, Brazil, Goiás, Gurupi, unnamed stream on Belém-Brasília highway, 30 km N of Gurupi; MZUSP 49462, 2, 30.9–32.6 mm SL, Brazil, Tocantins, Barreira do Piqui, unnamed stream affluent of Rio Água Fria, tributary of Rio Javaés (Rio Araguaia basin), on Araguaçu-Barreira do Piqui road, ca. 28 km N of Araguaçu; MZUSP 52248, 25, 31.9–44.1 mm SL, Brazil, Tocantins, Sandolândia, Rio Água Fria, in marginal lagoons, on road between Sandolândia and Barra do Rio Verde, at Lagoão farm; MZUSP 73368, 3, 27.5–35.7 mm SL, Brazil, Tocantins, Araguaçu, Rio Água Fria, at Praia Alta 2 farm, 27 km N of Araguaçu, on Araguaçu/Barreira do Piqui road; MZUSP 100151, 1, 38.2 mm SL, Corixo da Saudade (Corixinho), 25 km NW of Cocalinho on MT-326 highway, affluent of Rio Araguaia; UFRJ 2294, 3, 26.3–32.6 mm SL, Brazil, Tocantins, Araguaçu, Rio Javaés [Rio Araguaia basin], 40km N of Araguaçu; UFRJ 2295, 1 c&s, 30.4 mm SL, Brazil, Tocantins, Rio Javaés [Rio Araguaia basin], 40km N of Araguaçu.
Pimelodella australis Eigenmann (1917): LIRP 424, 15, 2 c&s, 69.7–95.4 mm SL, Brazil, Rio Grande do Sul, Osório, Arroio das Pedras [Rio Maquiné basin]; MCP 23926, 5, 74.4–77.8 mm SL, Brazil, Rio Grande do Sul, Triunfo, Arroio Bom Jardim [Rio Caí basin], III Pólo Petroquímico, near SITEL; MCP 34905, 3, 59.1–67.3 mm SL, Brazil, Rio Grande do Sul, Roque Gonzales, Arroio Cinamomo, affluent of Rio Ijuí [Rio Uruguai basin]; MZUSP 25030, 5 c&s, Brazil, Rio Grande do Sul, Porto Alegre, Belém-Novo, unnamed stream near Porto Alegre [Lago Guaíba system].
Pimelodella avanhandavae Eigenmann (1917): LIRP 6538, 1, Brazil, Minas Gerais, Araguari, Rio Paranaíba [Upper Rio Paraná basin], just downstream of dam of the hydroelectric plant of Emborcação [UHE Emborcação]; LIRP 7792, 1, 134.5 mm SL, 1 c&s, 143.7 mm SL, Brazil, Goiás, São Simão, Rio Paranaíba [Upper Rio Paraná basin], beside bridge of BR-365 highway, downstream from hydroelectric plant of São Simão [UHE São Simão].
Pimelodella boliviana Eigenmann (1917): LIRP 10020, 2, 63.5–77.2 mm SL Brazil, Rondônia, Porto Velho, Rio Madeira, near mouth of Rio São Lourenço; LIRP 10030, 1, 77.4 mm SL, Brazil, Rondônia, Porto Velho, mouth of Igarapé Arara, affluent of Rio Madeira.
Pimelodella boschmai Van der Stigchel (1964): LIRP 8148, 5, 2 c&s, 72.6–93.9 mm SL Brazil, São Paulo, Pirassununga, main channel of Rio Mogi-Guaçu, just downstream of Cachoeira das Emas waterfall.
Pimelodella cristata (Müller & Troschell, 1848): ANSP 175787, 1, Guyana, Burro Burro River, Perai Rocks Landing [plus nearby sites], Siparuni VIII-2; ANSP 175788, 2, 179.3 mm SL, Guyana, unnamed creek tributary to Essequibo River, approx. 18min upstream from Maipuri campsite, Siparuni VIII-2; ANSP 177224, 21, Guyana, Essequibo, Burro Burro River, at Water Dog Falls; ANSP 177225, 14, Guyana, Essequibo, Essequibo River, unnamed black water creek 2km downstream from Paddle Rock campside.
Pimelodella gracilis (Valenciennes, 1835): LIRP 101, 4, 60.3–64.6 mm SL, Brazil, São Paulo, Cajuru, unnamed creek affluent of Rio Pardo [Upper Rio Paraná basin], at Santa Carlota farm; LIRP 156, 3, 38.9–48.9 mm SL, Brazil, São Paulo, Cajuru, Córrego Barro Branco, near to mouth of Rio Pardo [Upper Rio Paraná basin], at Santa Carlota farm; LIRP 206, 32, 82.0–93.3 mm SL, Brazil, São Paulo, Santa Rosa de Viterbo, Rio Pardo [Upper Rio Paraná basin]; LIRP 304, 9, 76.4–110.4 mm SL, Brazil, São Paulo, Ribeirão Preto, Rio Pardo [Upper Rio Paraná basin]; LIRP 668, 4, Brazil, Rio Pardo [Upper Rio Paraná basin]; LIRP 1625, 1, 78.6 mm SL, Brazil, São Paulo, Teodoro Sampaio, Córrego São Carlos, affluent of Rio Paranapanema [Upper Rio Paraná basin], at Morro do Diabo State Park [Parque Estadual do Morro do Diabo]; LIRP 1697, 1, 90.0 mm SL, Brazil, Paraná, Paranavaí, Córrego Santa Clara, affluent of Rio Paranapanema [Upper Rio Paraná basin], at Rancho Zinco farm e Cristo Rei; LIRP 2924, 16, 43.7–90.3 mm SL, Brazil, São Paulo, Cerqueira César, Córrego Virado, affluent of Upper Rio Paranapanema [Upper Rio Paraná basin], at Santa Cecília farm; LIRP 2925, 1, 102.1 mm SL, Brazil, São Paulo, Angatuba, Córrego Poço das Antas, affluent of Upper Rio Paranapanema [Upper Rio Paraná basin]; LIRP 2926, 7, 2 c&s, 65.8–84.0 mm SL, Brazil, São Paulo, Olímpia, Córrego das Laranjeiras, Três Corações farm, affluent of Rio Turvo, affluent of Rio Grande [Upper Rio Paraná basin].
Pimelodella howesi Fowler (1940): ANSP 69036, 1, 79.3 mm SL, holotype, Bolivia, Río Chimore, Boca Chapare; ANSP 69037, 69056, 25, 33–90 mm TL, paratypes, Bolivia, Río Chimore, Boca; ANSP 84281, 29, Bolivia, Río Chapare, Todos Santos; ANSP 88497, 40, Bolivia, Cochabamba, Río Chimore, Boca Chapare.
Pimelodella lateristriga (Lichtenstein, 1823): LIRP 273, 1, 90.2 mm SL, Brazil, Minas Gerais, Joanésia, Rio Santo Antonio [Rio Doce basin]; LIRP 6839, 3, 108.3–116.9 mm SL, Brazil, Espírito Santo, Mimoso do Sul, Rio Itabapoana; LIRP 7847, 1, 1 c&s, 87.2 mm SL, Brazil, Espírito Santo, São José do Calçado, Rio Itabapoana, ca. 300m downstream from powerhouse of hydroelectric plant of Rosal [UHE Rosal].
Pimelodella meeki Eigenmann (1910): CAS 06600, 2, 113.1 mm SL, Brazil, Goiás, Corrégo do Sape; CAS 06698, 1, Brazil, Goiás, Vae-vem brook [Córrego Vai-e-vem].
Pimelodella megalops Eigenmann (1912): CAS 63717, 5, paratypes, Guyana, Potaro River at Tumatumari, Essequibo River drainage.
Pimelodella mucosa Eigenmann & Ward (1907): CAS 63720, 1, 97.4 mm SL, holotype, Paraguay, Río Paraguay, Bahia Negra on West bank.
Zootaxa 3717 (1) © 2013 Magnolia Press · 21A NEW SPECIES OF BRACHYRHAMDIA

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Pimelodella serrata Eigenmann (1917): MZUSP 55671, 1, Brazil, Amazonas, Rio Madeira, 1.9 km downstream from Paraná do Urucurituba; MZUSP 56871, 1, Brazil, Amazonas, Rio Madeira; USNM 305561, 16, Bolivia, Departamento del Beni, Provincia General José Ballivia Segurola, Río Matos below road crossing, 48 km E San Borja.
Pimelodella spelaea Trajano, Reis & Bichuette (2004): LIRP 8160, 5, 68.0–79.0 mm SL, Brazil, Goiás, São Domingos de Goiás, São Bernardo Cave, subterranean stream tributary to Rio São Bernardo.
Pimelodella steindachneri Eigenmann (1917): MZUSP 74535, 1, Brazil, Rondônia, Arquimedes, unnamed igarapé at Boa Vida farm; MZUSP 57333, 1, Brazil, Amazonas, Rio Madeira, near mouth.
Pimelodella taenioptera Miranda Ribeiro (1914): LIRP 710, 1, Brazil, Mato Grosso, Barão de Melgaço, Rio Cuiabá, near to Hotel Tuiuiu.
SLOBODIAN & BOCKMANN22 · Zootaxa 3717 (1) © 2013 Magnolia Press
Related Documents











