This article was downloaded by: [Laurent Marivaux] On: 03 May 2012, At: 09:04 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Journal of Vertebrate Paleontology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/ujvp20 A new basal phiomorph (Rodentia, Hystricognathi) from the late Oligocene of Lokone (Turkana Basin, Kenya) Laurent Marivaux a , Fabrice Lihoreau a , Fredrick Kyalo Manthi b & Stéphane Ducrocq c a Institut des Sciences de l’Évolution de Montpellier (ISE-M, UMR-CNRS 5554), C.c. 64, Université Montpellier 2, Place Eugène Bataillon, F-34095, Montpellier Cedex 05, France b Earth Sciences Department, National Museums of Kenya (NMK), Museum Hill, P.O. Box 40658, 00100, Nairobi, Kenya c Institut de Paléoprimatologie, Paléontologie Humaine: Évolution et Paléoenvironnements (IPHEP, UMR-CNRS 7262), Université de Poitiers UFR SFA, 40 avenue du Recteur Pineau, F-86022, Poitiers Cedex, France Available online: 03 May 2012 To cite this article: Laurent Marivaux, Fabrice Lihoreau, Fredrick Kyalo Manthi & Stéphane Ducrocq (2012): A new basal phiomorph (Rodentia, Hystricognathi) from the late Oligocene of Lokone (Turkana Basin, Kenya), Journal of Vertebrate Paleontology, 32:3, 646-657 To link to this article: http://dx.doi.org/10.1080/02724634.2012.657318 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article was downloaded by: [Laurent Marivaux]On: 03 May 2012, At: 09:04Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Journal of Vertebrate PaleontologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/ujvp20
A new basal phiomorph (Rodentia, Hystricognathi) fromthe late Oligocene of Lokone (Turkana Basin, Kenya)Laurent Marivaux a , Fabrice Lihoreau a , Fredrick Kyalo Manthi b & Stéphane Ducrocq ca Institut des Sciences de l’Évolution de Montpellier (ISE-M, UMR-CNRS 5554), C.c. 64,Université Montpellier 2, Place Eugène Bataillon, F-34095, Montpellier Cedex 05, Franceb Earth Sciences Department, National Museums of Kenya (NMK), Museum Hill, P.O. Box40658, 00100, Nairobi, Kenyac Institut de Paléoprimatologie, Paléontologie Humaine: Évolution et Paléoenvironnements(IPHEP, UMR-CNRS 7262), Université de Poitiers UFR SFA, 40 avenue du Recteur Pineau,F-86022, Poitiers Cedex, France
Available online: 03 May 2012
To cite this article: Laurent Marivaux, Fabrice Lihoreau, Fredrick Kyalo Manthi & Stéphane Ducrocq (2012): A new basalphiomorph (Rodentia, Hystricognathi) from the late Oligocene of Lokone (Turkana Basin, Kenya), Journal of VertebratePaleontology, 32:3, 646-657
To link to this article: http://dx.doi.org/10.1080/02724634.2012.657318
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses shouldbe independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims,proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly inconnection with or arising out of the use of this material.

Journal of Vertebrate Paleontology 32(3):646–657, May 2012© 2012 by the Society of Vertebrate Paleontology
ARTICLE
A NEW BASAL PHIOMORPH (RODENTIA, HYSTRICOGNATHI) FROM THE LATEOLIGOCENE OF LOKONE (TURKANA BASIN, KENYA)
LAURENT MARIVAUX,*,1 FABRICE LIHOREAU,1 FREDRICK KYALO MANTHI,2 and STEPHANE DUCROCQ3
1Institut des Sciences de l’Evolution de Montpellier (ISE-M, UMR-CNRS 5554), C.c. 64, Universite Montpellier 2, Place EugeneBataillon F-34095, Montpellier Cedex 05, France, [email protected], [email protected];2Earth Sciences Department, National Museums of Kenya (NMK), Museum Hill, P.O. Box 40658, 00100, Nairobi, Kenya,
[email protected];3Institut de Paleoprimatologie, Paleontologie Humaine: Evolution et Paleoenvironnements (IPHEP, UMR-CNRS 7262), Universite
de Poitiers UFR SFA, 40 avenue du Recteur Pineau, F-86022, Poitiers Cedex, France, [email protected]
ABSTRACT—Early hystricognathous rodents from Africa are primarily documented by two basal and extinct groups, theparaphyletic “Phiomyidae” and the Gaudeamuridae, which were particularly well diversified through the late Eocene andthe early Oligocene. However, in the absence of a comprehensive late Oligocene fossil record, the evolutionary history ofAfrican hystricognathous rodents during the end of the Paleogene is unclear. Continuing field efforts in the Lokichar Basinof Kenya (western Turkana Basin) have led to the discovery of dental remains of a ‘phiomyid’ from the Lokone site LOK 13.The dental pattern of this rodent is unusual among ‘phiomyids,’ which led us to propose here a new taxon: Turkanamys hex-alophus, gen. et sp. nov. Turkanamys shares a similar dental bauplan with early Afro-Asian hystricognaths (early ‘phiomyids’and ‘baluchimyines’), and as such appears evolutionarily conservative with respect to some Oligocene and all the Miocenehystricognaths. Conversely, Turkanamys is also highly autapomorphic in developing a hexalophodont pattern on upper mo-lars. The discovery of this peculiar rodent highlights the persistence of this ancestral ‘phiomorph’ group at the end of thePaleogene. Given the great familial diversity of hystricognaths in the early Miocene of East Africa, the virtual absence oftheir stem representatives in the late Oligocene record of the same province suggests either important taphonomic/samplingbiases or a real absence of these families in this province at that time. This would suggest that hystricognathous rodentsdiversified elsewhere and colonized East Africa by the late Oligocene–early Miocene transition.
INTRODUCTION
Despite a poor Paleogene fossil record, Africa has long ap-peared as being an early center of adaptive radiation of twophylogenetically distinct groups of Rodentia: Hystricognathi andAnomaluroidea. Early hystricognathous rodents from Africa(i.e., “Phiomorpha” sensu Lavocat, 1962; superfamily Thry-onomyoidea Wood, 1968) are primarily documented by the“Phiomyidae” (see Wood, 1955) and the Gaudeamuridae (seeSallam et al., 2011), two extinct families that were among themost common members of African mammal faunas during thelate Paleogene (especially through the late Eocene and the earlyOligocene). Early ‘phiomorphs’ are generally viewed as the stemgroup of the modern African cane rats (Thryonomyidea; Thryon-omys), dassie rats (Petromuridae; Petromus), mole rats (Bathy-ergidae; e.g., Bathyergus, Heterocephalus), modern Afro-Asianporcupines (Hystricidae; e.g., Hystrix, Atherurus), and SouthAmerican caviomorphs (e.g., Lavocat, 1976; Sallam et al., 2009;Antoine, Marivaux, et al., 2012). Historically, ‘phiomorphs’ werefirst reported and described early in the last century from theOligocene Jebel el-Qatrani Formation in Egypt (Fayum: Os-born, 1908; Wood, 1968), the early Miocene of Kenya-Uganda(Rusinga, Songhor: Lavocat, 1973), the late middle Eocene ofAlgeria (Bir el Ater: Jaeger et al., 1985), the early Oligocene ofLibya (Zallah: Fejfar, 1987), and the early Oligocene of Dhofar(Thaytiniti, Taqah: Thomas et al., 1989, 1992). Over the last re-cent years, several new Paleogene localities from Egypt (Fayum,BQ2, earliest late Eocene: Sallam et al., 2009; L41, latest Eocene:
*Corresponding author.
Holroyd, 1994; Sallam et al., 2011) and Libya (Dur At-Talah,late middle–middle late Eocene: Jaeger et al., 2010; Zallah, earlyOligocene: Coster et al., 2010), as well as from sub-SaharanAfrica such as Tanzania (Mbeya, late Oligocene: Stevens et al.,2006, 2009), Namibia (Sperrgebiet, ?middle Eocene: Pickfordet al., 2008), and Kenya (late Oligocene, Losodok: Rasmussenand Gutierrez, 2009; Lokone: Ducrocq et al., 2010), have sub-stantially increased the species diversity of Paleogene hystricog-nathous rodents in Africa. Phiomorph-like hystricognaths havesince been recorded in the Paleogene (late Eocene and earlyOligocene) of southern Asia and Asia Minor (“Baluchimyinae”;Flynn et al., 1986; Marivaux et al., 2000, 2002; Marivaux and Wel-comme, 2003; de Bruijn et al., 2003) and of South America (?LateEocene of Santa Rosa, Peru: Frailey and Campbell, 2004; EarlyOligocene of Gran Barranca, Argentina: Vucetich et al., 2010;late middle Eocene of Contamana, Peru: Antoine, Marivaux, etal., 2012), thereby highlighting the early evolutionary and paleo-biogeographic history of that widespread rodent group.
In Africa, the rare late Oligocene localities have yieldedonly a very limited record regarding hystricognathous rodentscompared with the late Eocene, early Oligocene, and earlyMiocene localities, where a wide array of ‘phiomorphs’ withwell-preserved specimens have been collected. For the lateOligocene, which ranges over 5 million years, only three genera(Metaphiomys, Kahawamys, and Diamantomys) are known froma few localities situated exclusively in sub-Saharan Africa. Inaddition, these rodents are rather poorly documented, knownonly by their lower dentition, including one tooth fragment(Stevens et al., 2006), two isolated teeth (Ducrocq et al., 2010),and two partial mandibles preserving worn teeth (Rasmussenand Gutierrez, 2009; Stevens et al., 2009). The preliminary report
646
Dow
nloa
ded
by [
Lau
rent
Mar
ivau
x] a
t 09:
04 0
3 M
ay 2
012

MARIVAUX ET AL.—LATE OLIGOCENE PHIOMYID RODENT FROM KENYA 647
FIGURE 1. Location maps and picture of the fossiliferous area. A, Lokone hill (black star) is situated in the Turkana Basin in Kenya; B, map showingthe position of the Lokone hill (black star) in the Lokichar Basin in the western Turkana Basin and the surrounding Cretaceous to Cenozoic rift basinsin northern-central Kenya (modified after Ducrocq et al., 2010); C, photograph of the fossiliferous sandstone of LOK 13 (foreground) from which therodent material described here was extracted after acid treatment (picture by F. Lihoreau).
of Ducrocq et al. (2010) on the late Oligocene Lokone faunal as-semblage from Kenya (Lokone Hill [LOK 1, 2, 12, 13], LokicharBasin, western Turkana Basin; Fig. 1) described one rodentlower molar (m2) from LOK 13, which was tentatively attributedto Metaphiomys cf. M. schaubi. A rodent lower incisor showing amultiserial enamel microstructure was also reported from LOK 1
(Ducrocq et al., 2010). The continuing field efforts on the Lokonesites, notably on LOK 13, associated with acid processing of thehardened sediments, have led to the recovery of additional fossilremains of that rodent. The new material now includes fragmentsof mandibles and isolated upper teeth, which allows revision ofour former taxonomic attribution of the Lokone ‘phiomorph’
Dow
nloa
ded
by [
Lau
rent
Mar
ivau
x] a
t 09:
04 0
3 M
ay 2
012
648 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 32, NO. 3, 2012
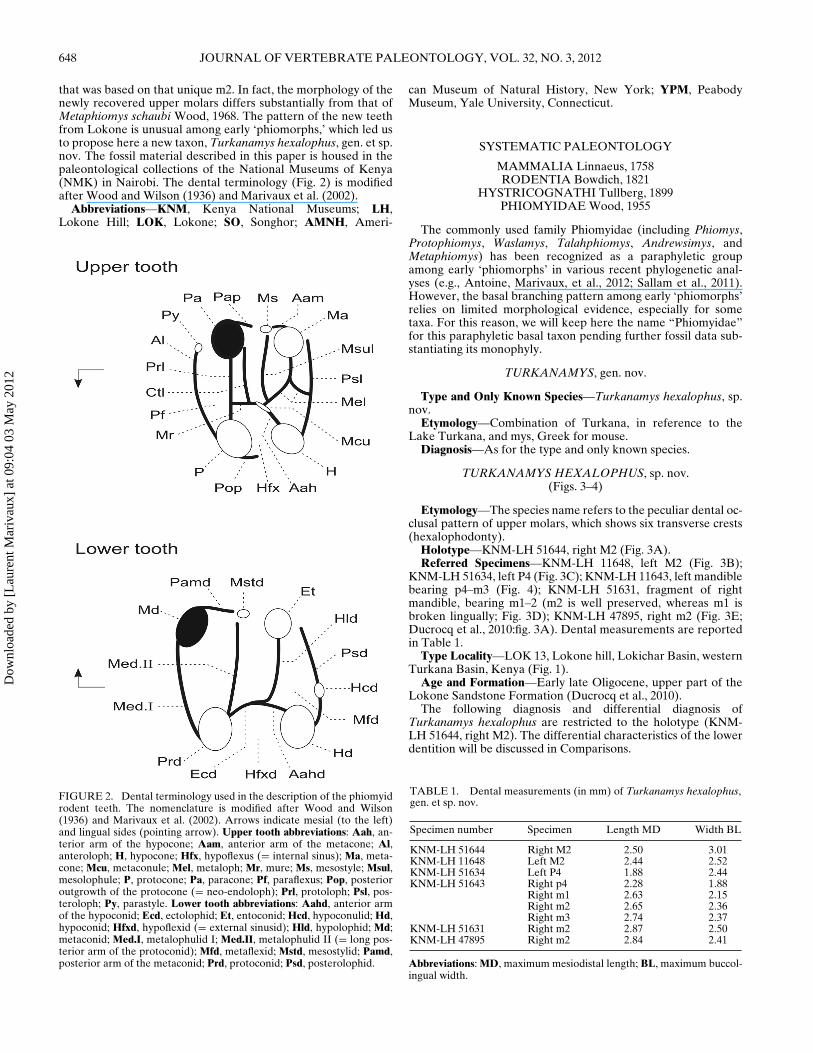
that was based on that unique m2. In fact, the morphology of thenewly recovered upper molars differs substantially from that ofMetaphiomys schaubi Wood, 1968. The pattern of the new teethfrom Lokone is unusual among early ‘phiomorphs,’ which led usto propose here a new taxon, Turkanamys hexalophus, gen. et sp.nov. The fossil material described in this paper is housed in thepaleontological collections of the National Museums of Kenya(NMK) in Nairobi. The dental terminology (Fig. 2) is modifiedafter Wood and Wilson (1936) and Marivaux et al. (2002).
Abbreviations—KNM, Kenya National Museums; LH,Lokone Hill; LOK, Lokone; SO, Songhor; AMNH, Ameri-
FIGURE 2. Dental terminology used in the description of the phiomyidrodent teeth. The nomenclature is modified after Wood and Wilson(1936) and Marivaux et al. (2002). Arrows indicate mesial (to the left)and lingual sides (pointing arrow). Upper tooth abbreviations: Aah, an-terior arm of the hypocone; Aam, anterior arm of the metacone; Al,anteroloph; H, hypocone; Hfx, hypoflexus (= internal sinus); Ma, meta-cone; Mcu, metaconule; Mel, metaloph; Mr, mure; Ms, mesostyle; Msul,mesolophule; P, protocone; Pa, paracone; Pf, paraflexus; Pop, posterioroutgrowth of the protocone (= neo-endoloph); Prl, protoloph; Psl, pos-teroloph; Py, parastyle. Lower tooth abbreviations: Aahd, anterior armof the hypoconid; Ecd, ectolophid; Et, entoconid; Hcd, hypoconulid; Hd,hypoconid; Hfxd, hypoflexid (= external sinusid); Hld, hypolophid; Md;metaconid; Med.I, metalophulid I; Med.II, metalophulid II (= long pos-terior arm of the protoconid); Mfd, metaflexid; Mstd, mesostylid; Pamd,posterior arm of the metaconid; Prd, protoconid; Psd, posterolophid.
can Museum of Natural History, New York; YPM, PeabodyMuseum, Yale University, Connecticut.
SYSTEMATIC PALEONTOLOGY
MAMMALIA Linnaeus, 1758RODENTIA Bowdich, 1821
HYSTRICOGNATHI Tullberg, 1899PHIOMYIDAE Wood, 1955
The commonly used family Phiomyidae (including Phiomys,Protophiomys, Waslamys, Talahphiomys, Andrewsimys, andMetaphiomys) has been recognized as a paraphyletic groupamong early ‘phiomorphs’ in various recent phylogenetic anal-yses (e.g., Antoine, Marivaux, et al., 2012; Sallam et al., 2011).However, the basal branching pattern among early ‘phiomorphs’relies on limited morphological evidence, especially for sometaxa. For this reason, we will keep here the name “Phiomyidae”for this paraphyletic basal taxon pending further fossil data sub-stantiating its monophyly.
TURKANAMYS, gen. nov.
Type and Only Known Species—Turkanamys hexalophus, sp.nov.
Etymology—Combination of Turkana, in reference to theLake Turkana, and mys, Greek for mouse.
Diagnosis—As for the type and only known species.
TURKANAMYS HEXALOPHUS, sp. nov.(Figs. 3–4)
Etymology—The species name refers to the peculiar dental oc-clusal pattern of upper molars, which shows six transverse crests(hexalophodonty).
Holotype—KNM-LH 51644, right M2 (Fig. 3A).Referred Specimens—KNM-LH 11648, left M2 (Fig. 3B);
KNM-LH 51634, left P4 (Fig. 3C); KNM-LH 11643, left mandiblebearing p4–m3 (Fig. 4); KNM-LH 51631, fragment of rightmandible, bearing m1–2 (m2 is well preserved, whereas m1 isbroken lingually; Fig. 3D); KNM-LH 47895, right m2 (Fig. 3E;Ducrocq et al., 2010:fig. 3A). Dental measurements are reportedin Table 1.
Type Locality—LOK 13, Lokone hill, Lokichar Basin, westernTurkana Basin, Kenya (Fig. 1).
Age and Formation—Early late Oligocene, upper part of theLokone Sandstone Formation (Ducrocq et al., 2010).
The following diagnosis and differential diagnosis ofTurkanamys hexalophus are restricted to the holotype (KNM-LH 51644, right M2). The differential characteristics of the lowerdentition will be discussed in Comparisons.
TABLE 1. Dental measurements (in mm) of Turkanamys hexalophus,gen. et sp. nov.
Specimen number Specimen Length MD Width BL
KNM-LH 51644 Right M2 2.50 3.01KNM-LH 11648 Left M2 2.44 2.52KNM-LH 51634 Left P4 1.88 2.44KNM-LH 51643 Right p4 2.28 1.88
Right m1 2.63 2.15Right m2 2.65 2.36Right m3 2.74 2.37
KNM-LH 51631 Right m2 2.87 2.50KNM-LH 47895 Right m2 2.84 2.41
Abbreviations: MD, maximum mesiodistal length; BL, maximum buccol-ingual width.
Dow
nloa
ded
by [
Lau
rent
Mar
ivau
x] a
t 09:
04 0
3 M
ay 2
012

MARIVAUX ET AL.—LATE OLIGOCENE PHIOMYID RODENT FROM KENYA 649
FIGURE 3. Isolated upper and lower dental remains of Turkanamys hexalophus, gen. et sp. nov. A, KNM-LH 51644, a right M2 (holotype) inocclusal (A1–2) and lingual (A3) views; B, KNM-LH 11648, a left M2 (buccally broken) in occlusal (B1–2) and lingual (B3) views; C, KNM-LH 51634,a left P4 (distally worn) in occlusal views (C1–2); D, KNM-LH 51631, a fragment of right mandible bearing m1–2 (m1 being badly broken lingually) inocclusal views (D1–2); E, KNM-LH 47895, a right m2 (figured in Ducrocq et al., 2010:fig. 3A [SEM micrograph]). Original drawings (A1, B1, C1, andD1) by Laurence Meslin, © CNRS-Meslin. The fossil specimens were photographed at the Kenya National Museum in Nairobi, with a digital cameracoupled with a stereomicroscope. Scale bar equals 1 mm.
Dow
nloa
ded
by [
Lau
rent
Mar
ivau
x] a
t 09:
04 0
3 M
ay 2
012

650 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 32, NO. 3, 2012
FIGURE 4. Lower toothrow of Turkanamys hexalophus, gen. et sp. nov. KNM-LH 11643, a left mandible bearing p4–m3 in buccal (A1) and occlusal(A2–3) views. Original drawing (A3) by Laurence Meslin, © CNRS-Meslin. Scale bar equals 1 mm.
Dow
nloa
ded
by [
Lau
rent
Mar
ivau
x] a
t 09:
04 0
3 M
ay 2
012

MARIVAUX ET AL.—LATE OLIGOCENE PHIOMYID RODENT FROM KENYA 651
Diagnosis—Differs from all other ‘phiomyids’ (e.g.,Metaphiomys, Phiomys, Protophiomys, Waslamys, Talah-phiomys, Andrewsimys) in having hexalophodont upper molars,which are characterized by well-developed anteroloph, pro-toloph, mesolophule, metaloph, and posteroloph and thedevelopment of an additional central transverse crest (i.e.,centroloph). Differs from Metaphiomys and Andrewsimys inpreserving a connection between the metaloph and the anteriorarm of hypocone. Differs from Phiomys in having a strongerdevelopment of the mesolophule, a narrower, deeper, andmore oblique hypoflexus, and in having slightly more bulbouscusps. Differs from Andrewsimys in having upper molars non-taeniodont (hypoflexus and paraflexus separated by the completelingual protoloph). Differs from Protophiomys, Waslamys, andTalahphiomys in bearing a complete mure, a strong developmentof the mesolophule, in having no trace of metaconule, and inhaving a metaloph-posteroloph connection (double connectionof the metaloph). Differs more specifically from Protophiomysand Waslamys in lacking the development of a neo-endoloph(posterior outgrowth of the protocone).
Differential Diagnosis—Differs from ‘baluchimyines’ (e.g.,Baluchimys, Lophibaluchia, Hodsahibia, Bugtimys, Lindsaya,Ottomania) in having a hexalophodont pattern on upper mo-lars, in showing a strong development of the mure, and in havinga metaloph-posteroloph connection (except for Lophibaluchia)and no trace of metaconule. Differs from Ottomania in lackingthe lingual anteroloph, and from Baluchimys in lacking the neo-endoloph. Differs from diamantomyids (e.g., Diamantomys) inhaving brachydont and bunolophodont teeth, thinner and trans-verse instead of strong and oblique crests, and in preserving aconnection between the metaloph and the anterior arm of thehypocone. Upper molars are more square-shaped and have theirhypoflexus (internal sinus) anteriorly oblique, forwardly ratherthan transversely oriented. Differs from gaudeamurids (i.e.,Gaudeamus) in having brachydont and hexalophodont teeth,with thinner transverse crests and well-distinct metaloph, whichis connected to the anterior arm of the hypocone, and in lackinga taeniodont pattern (lingual protoloph complete). Differs fromthryonomyids (e.g., Paraphiomys, Paraulacodus) in having hex-alophodont upper molars, a stronger (longer) development of themesolophule, less inflated cusps, in preserving a connection be-tween the metaloph and the anterior arm of the hypocone, andin having a deeper hypoflexus. Differs from Miocene thryono-myoids (from Kenya-Uganda [Lavocat, 1973]; e.g., Simonimys,Epiphiomys, Kenyamys, Myophiomys) in having hexalophodontupper molars and in preserving the mesial connection of the met-aloph. Differs more specifically from Epiphiomys in having thin-ner and lower transverse crests, and from Simonimys in hav-ing upper molars with more bulbous cusps and more square-shaped rather than rectangular outline. Differs from Kenyamysin having lower transverse crests, cusps more prominent, smallermesostyle, and in displaying an oblique rather than transversehypoflexus. Differs from Myophiomys in being less brachydont,more lophate (transverse crests more elevated), and in lacking alingual anteroloph.
DESCRIPTION
Upper Dentition
Upper Molars—KNM-LH 51644 (holotype; Fig. 3A) andKNM-LH 11648 (Fig. 3B) are identified as M2 rather than M1due to the position of the hypocone in both teeth, which oc-curs in a more buccal position than the protocone. Both uppermolars are quadrangular and tri-rooted (one large lingual rootand two small buccal roots). Despite the degree of wear, themain cusps (protocone, paracone, metacone, and hypocone) arebulbous and elevated, dominating the transverse crests. Thesecrests are well marked and sub-parallel. A strong, long longitu-
dinal mure is present, connecting the mesial end of the anteriorarm of the hypocone to the lingual part of the protoloph. As inmost ‘phiomyid’ species but also in some other families of earlyhystricognaths (i.e., ‘baluchimyines,’ diamantomyines), the an-teroloph, protoloph, metaloph, posteroloph, and the central crest(the mesolophule, the degree of development of which can bevariable) represent the main transverse crests. However, uppermolars of Turkanamys bear a distinct sixth transverse crest (cen-troloph), which develops between the protoloph and the mesolo-phule, thereby producing a hexalophodont pattern. This addi-tional crest is somewhat less elevated than the main crests. Thecentroloph joins the mure lingually and extends buccally, endingat the base of the distobuccal aspect of the paracone on KNM-LH11648 (Fig. 3B), or at the base of the distolingual aspect of theparacone on KNM-LH 51644 (Fig. 3A). The paracone and pro-tocone are buccolingually opposed and linked to each other bythe protoloph. The anteroloph is strong and connected linguallyto the protocone, but not to the paracone buccally (there is onlya narrow notch separating the anteroloph from the paracone).The anteroloph and the protoloph isolate a long, narrow, shallowparaflexus, which remains faintly open buccally. The hypocone issmaller than the protocone. The posteroloph links the hypoconeto the metacone. The hypocone bears a strong, long anterior arm,which extends obliquely mesiobuccally to connect to the mure.The lingual part of the mesolophule starts at the junction betweenthe mure and the anterior arm of the hypocone. The mesolophuleis sinuous in its lingual part, and continues its course buccally, be-ing slightly oblique, but parallel to the centroloph. On both up-per molars, the mesolophule ends in a minute mesostyle buccally.From the metacone, the metaloph extends lingually (straight inits buccal-most part), and turns mesially in its median part to con-nect the lingual-most part of the mesolophule. The metaloph isalso connected to the posteroloph by a thin, very short longitu-dinal enamel crestule. The posteroloph, associated with the ante-rior arm of the hypocone, the lingual-most part of the mesolo-phule, and the double-connection of the metaloph (metaloph-mesolophule and metaloph-posteroloph), delimits a small andcircular distal fovea. The hypocone and its anterior arm, associ-ated with the mure, the lingual-most part of the protoloph and theprotocone, circumscribe the hypoflexus (internal sinus), which isvery narrow, deep, oblique, and lingually open.
Upper Premolar—We identify as P4, an oval-shaped toothcharacterized by a buccolingual long axis (KNM-LH 51634; Fig.3C). The distal part of this premolar is poorly preserved, whichprevents any precise description. The paracone, protocone, andmetacone are distinct. The paracone appears as the largest cuspof the crown, but the metacone, which is broken, was certainly ofsimilar size. These two buccal cusps are twinned and separatedonly by a narrow notch. Just mesiolingual to the metacone, at thecenter of the tooth, there is a small cusp, that we interpret hereas an isolated metaconule. The protocone is crestiform (buccolin-gually compressed) and lingually opposed to the paracone. Bothcusps are linked by a strong protoloph and an anteroloph, whichis elevated (as high as the protoloph) and curved (convex). Theprotocone develops a strong posterior outgrowth, which extendsfar distally, thereby closing the hypoflexus lingually. On the dis-tolingual region of the tooth, the hypocone is crestiform, entirelymerged in the distal end of the posterior outgrowth of the proto-cone and displays an anterior arm. There is a strong longitudinalmure, which connects the lingual part of the protoloph and theanterior arm of the hypocone.
Lower Dentition
The lower dentition is documented by three specimens: a leftmandible bearing p4 and m1–3 (KNM-LH 11643; Fig. 4); a frag-ment of right mandible bearing a well-preserved m2 and a broken
Dow
nloa
ded
by [
Lau
rent
Mar
ivau
x] a
t 09:
04 0
3 M
ay 2
012

652 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 32, NO. 3, 2012
m1 (KNM-LH 51631; Fig. 3D); an isolated right m2 (KNM-LH47895; Fig. 3E, figured in Ducrocq et al., 2010:fig. 3A).
Lower Molars—The molars (m1–3) are longer than wide andshare the same structural pattern. On KNM-LH 11643 (Fig. 4),which preserves a complete toothrow, the size of the molars in-creases from m1 to m3. The m3 is the largest tooth and differsfrom m1 and m2 in having a talonid slightly narrower than thetrigonid. On these three molars, the main cuspids (protoconid,metaconid, hypoconid, and entoconid) are bulbous and of similarsize. As on upper molars, the transverse cristids are well markedbut remain lower than the cuspids. The protoconid is slightly dis-tally displaced with respect to the metaconid. Mesiobucally andimmediately anterior to the protoconid, there is a short ante-rocingulid. Both mesial cuspids are linked by a strong and slightlycurved metalophulid I. The entoconid is slightly mesial to thehypoconid. The hypoconid develops a strong, long, oblique an-terior arm, which joins a straight hypolophid running from theentoconid. A strong, elevated ectolophid extends obliquely fromthe protoconid (its distal aspect) to the junction between the hy-polophid and the anterior arm of the hypoconid. There is notrace of mesoconid. The second transverse cristid is somewhatdifficult to describe inasmuch as it is highly variable. On KNM-LH 47895 (Fig. 3E), this cristid is complete (i.e., buccolinguallyuninterrupted), formed by a long posterior arm of the proto-conid (= metalophulid II), which extends lingually and ends ina minute (lingual) mesostylid. On KNM-LH 11643 (Fig. 4) andKNM-LH 51631 (Fig. 3D), it is made from two cristids that mergetogether; one being a short posterior arm of the protoconid (lin-gually directed), and the other one being a short cristid originat-ing from the mesostylid (buccomesially directed, i.e., oblique; lin-gual mesolophid-like structure?). The question as to whether thissecond transverse cristid represents a complete metalophulid IIor a composite cristid including a lingual mesolophid and a part ofthe posterior arm of the protoconid requires additional materialfor a better understanding of the pattern of variation. The meta-conid displays a long, moderately elevated posterior arm, whichprojects distally and ends just before the mesostylid (KNM-LH51631; Fig. 3D) or connects to it (KNM-LH 11643; Fig. 4). Theanterior arm of the entoconid is faintly developed and the trigo-nid remains open lingually as a result. Distally, there is a strongposterolophid, which bears a small hypoconulid. This latter isfaintly visible, appearing as a minute swelling, mesiodistally com-pressed, located on the middle of the posterolophid. There is noentoconid-posterolophid connection.
Lower Premolar—The p4 on KNM-LH 11643 (Fig. 4) is oval-shaped, with a trigonid slightly buccolingually narrower than thetalonid. The talonid is as high as the trigonid. The protoconid,metaconid, hypoconid, and entoconid are of similar size. The pro-toconid is slightly more distal in position than the metaconid. Theposterior arm of the protoconid is well developed and extendslingually to the metaconid distolingually, thereby forming a com-plete metalophulid II. Mesially, there is no anterocingulid, andthe metalophulid I connects the protoconid but not the meta-conid. The hypoconid is slightly mesiodistally compressed, andis buccolingually in line with the entoconid. The hypoconid andentoconid are linked by a strong and trenchant posterolophid.There is no hypoconulid on the posterolophid. The anterior armof the hypoconid is strong and long, and connects the ectolophid.The ectolophid extends mesiobucally and reaches the distal as-pect of the protoconid (complete ectolophid). This longitudinalcrest bears a small enamel swelling that might be a minute meso-conid. At the junction between the ectolophid and the anteriorarm of the hypoconid, there is a very low transverse cristid thatmight be a hypolophid (only the buccal part). This hypolophid-like structure (or mesolophid?) extends lingually but does notconnect the entoconid. This crest is rather directed toward a well-marked mesostylid. This mesostylid extends distally and mesially,and connects the entoconid and a strong, long posterior arm of
the metaconid, respectively. There is a longitudinal accessoryenamel crestulid, which is connected to the lingual extremity ofthe hypolophid-like structure and directed toward the middle ofthe metalophulid II.
Incisor Enamel Microstructure
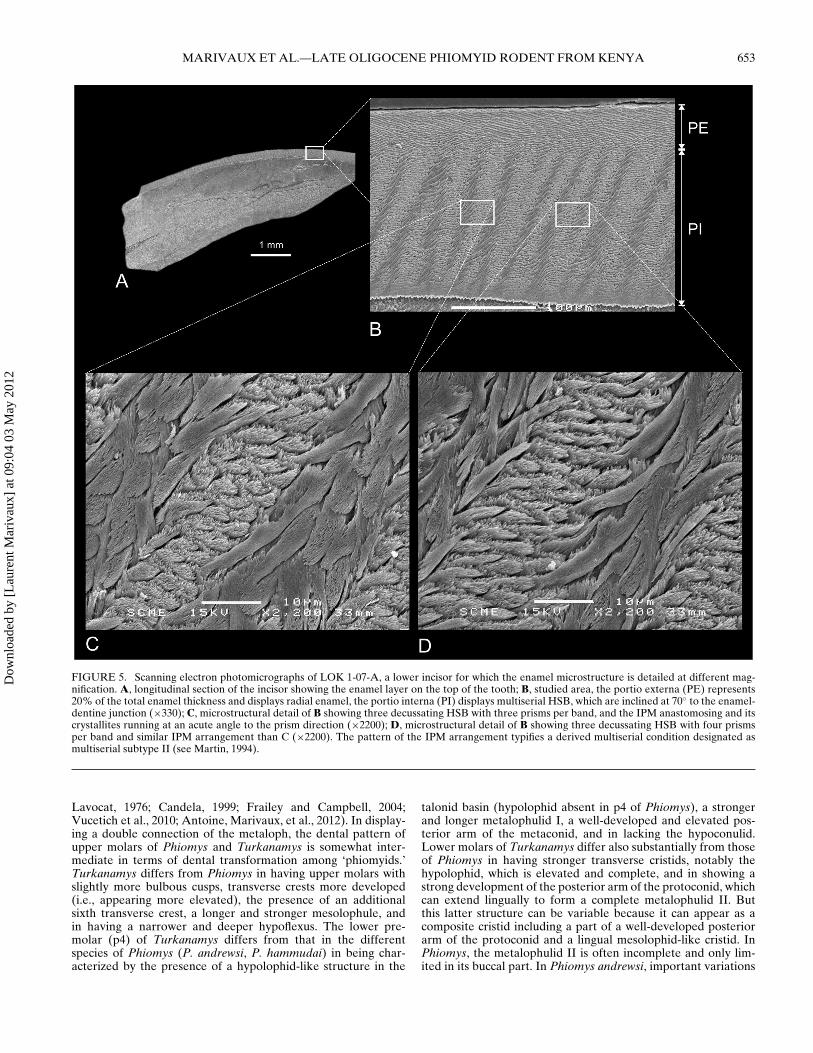
Among the fossil material collected from the residues of LOK1, one tooth is a lower rodent incisor (LOK 1-07-A). In overallproportion, the size of this incisor matches that of the molars ofTurkanamys hexalophus, which would therefore suggest that thisincisor could be tentatively referred to this taxon. A fragmentof this incisor was embedded in artificial resin, and subsequentlypolished longitudinally (Fig. 5A). The polished section was thenetched for 30 seconds with H3PO4 (37%) to make the microstruc-tural details visible (Fig. 5B–C). The enamel microstructure ofthis incisor was succinctly described in Ducrocq et al. (2010). Weprovide here a detailed description.
In longitudinal section, the total enamel thickness is about 0.23mm with two distinct layers, the portio interna (PI) and the por-tio externa (PE). The PE has radial enamel and constitutes about20% of the total enamel thickness. The PI, which is very thick,consists of decussating layers of prisms appearing as Hunter-Schreger bands (HSB), 3–4 prisms wide per band (oval in crosssection). The straight HSB are inclined at 70◦ to the enamel-dentine junction (EDJ). There is no transition zone between ad-jacent decussating HSB. The interprismatic matrix (IPM) in thePI is thin and never surrounds the prisms but it anastomosesand its crystallites run at an acute angle to the prism direction.Such a crystallite arrangement in PI determines typical multise-rial enamel (Korvenkontio, 1934). The pattern of the IPM ar-rangement further typifies a derived multiserial condition desig-nated as multiserial subtype II (see Martin, 1994).
COMPARISONS
In developing a hexalophodont rather than pentalophodontpattern on the upper molars, Turkanamys hexalophus is uniqueamong early ‘phiomyid’ rodents. However, except for the pres-ence of an additional transverse crest on the upper molars (i.e.,centroloph), the lower and upper dentitions of Turkanamysare typically ‘phiomyid’-like. The cusp and crest arrangementthat characterizes upper molars of T. hexalophus is somewhatreminiscent of that of Phiomys, notably P. hammudai (latemiddle–middle late Eocene; Jaeger et al., 2010), P. andrewsi, andP. paraphiomyoides (early Oligocene; Wood, 1968). Upper mo-lars of Phiomys and Turkanamys exhibit a double connectionof the metaloph, one with the mesolophule, the other one withthe posteroloph. This double connection of the metaloph associ-ated with the anterior arm of the hypocone and the posterolophdelimits a small, rounded fovea. The metaloph-mesolophule (ormetaloph-anterior arm of the hypocone connection) character-izes the basal-most ‘phiomyids’ (late middle–early late Eocene;especially Protophiomys and Waslamys; Jaeger et al., 1985, 2010;Sallam et al., 2009), but also other basal hystricognaths, notably‘baluchimyines’ (late Eocene, early Oligocene; e.g., Baluchimys,Hodsahibia, Bugtimys, Ottomania, Confiniummys; Flynn et al.,1986; Marivaux et al., 2000, 2002; de Bruijn et al., 2003; Mari-vaux and Welcomme, 2003) and one stem caviomorph (late mid-dle Eocene; Canaanimys; Antoine, Marivaux, et al., 2012). Ametaloph-posteroloph connection is characteristic of more ad-vanced ‘phiomyids’ (Oligocene; e.g., Metaphiomys; Wood, 1968;Fejfar, 1987), advanced ‘baluchimyines’ (early Oligocene; e.g.,Lophibaluchia; Flynn et al., 1986), but also diamantomyids (earlyMiocene; e.g., Diamantomys; Lavocat, 1973), basal thryonomy-oids (early Miocene; e.g., Simonimys, Epiphiomys, Kenyamys,Paraphiomys, Paraulacodus; Lavocat, 1973; de Bruijn and Hus-sain, 1985), and most caviomorphs (Eocene-Oligocene; e.g.,Cachiyacuy, Eobranisamys, Branisamys, Eoincamys, Incamys;
Dow
nloa
ded
by [
Lau
rent
Mar
ivau
x] a
t 09:
04 0
3 M
ay 2
012
MARIVAUX ET AL.—LATE OLIGOCENE PHIOMYID RODENT FROM KENYA 653
FIGURE 5. Scanning electron photomicrographs of LOK 1-07-A, a lower incisor for which the enamel microstructure is detailed at different mag-nification. A, longitudinal section of the incisor showing the enamel layer on the top of the tooth; B, studied area, the portio externa (PE) represents20% of the total enamel thickness and displays radial enamel, the portio interna (PI) displays multiserial HSB, which are inclined at 70◦ to the enamel-dentine junction (×330); C, microstructural detail of B showing three decussating HSB with three prisms per band, and the IPM anastomosing and itscrystallites running at an acute angle to the prism direction (×2200); D, microstructural detail of B showing three decussating HSB with four prismsper band and similar IPM arrangement than C (×2200). The pattern of the IPM arrangement typifies a derived multiserial condition designated asmultiserial subtype II (see Martin, 1994).
Lavocat, 1976; Candela, 1999; Frailey and Campbell, 2004;Vucetich et al., 2010; Antoine, Marivaux, et al., 2012). In display-ing a double connection of the metaloph, the dental pattern ofupper molars of Phiomys and Turkanamys is somewhat inter-mediate in terms of dental transformation among ‘phiomyids.’Turkanamys differs from Phiomys in having upper molars withslightly more bulbous cusps, transverse crests more developed(i.e., appearing more elevated), the presence of an additionalsixth transverse crest, a longer and stronger mesolophule, andin having a narrower and deeper hypoflexus. The lower pre-molar (p4) of Turkanamys differs from that in the differentspecies of Phiomys (P. andrewsi, P. hammudai) in being char-acterized by the presence of a hypolophid-like structure in the
talonid basin (hypolophid absent in p4 of Phiomys), a strongerand longer metalophulid I, a well-developed and elevated pos-terior arm of the metaconid, and in lacking the hypoconulid.Lower molars of Turkanamys differ also substantially from thoseof Phiomys in having stronger transverse cristids, notably thehypolophid, which is elevated and complete, and in showing astrong development of the posterior arm of the protoconid, whichcan extend lingually to form a complete metalophulid II. Butthis latter structure can be variable because it can appear as acomposite cristid including a part of a well-developed posteriorarm of the protoconid and a lingual mesolophid-like cristid. InPhiomys, the metalophulid II is often incomplete and only lim-ited in its buccal part. In Phiomys andrewsi, important variations
Dow
nloa
ded
by [
Lau
rent
Mar
ivau
x] a
t 09:
04 0
3 M
ay 2
012

654 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 32, NO. 3, 2012
involving the posterior arm of the protoconid and the mesolophidare observed. The metalophulid II is sometime incomplete andonly limited in its buccal part. In some specimens, the mesolophidis attached to the lingual tip of the posterior arm of the proto-conid (e.g., in m2 of AMNH 13271, Wood, 1968:fig. 1G; in m2 ofYPM 18021, Wood, 1968:fig. 2A). In some teeth, the mesolophidis short and isolated, extending from the mesostylid toward theposterior arm of the protoconid but does not connect this lat-ter (e.g., in m1 of YPM 18064, Wood, 1968:fig. 1F). Turkanamysdiffers from P. andrewsi in having no trace of mesoconid. InPhiomys hammudai, the posterior arm of the protoconid doesnot form a complete metalophulid II and the area involving themesolophid is quite variable. In P. hammudai and Turkanamys,the posterior arm of the metaconid projects distally toward themesostylid. Lower molars of Turkanamys are also characterizedby a slight mesial displacement of the entoconid, which is lin-gually opposed to the hypoconid in lower molars of Phiomys.Considering this mesial displacement of the entoconid withrespect to the hypoconid, as well as a slight distal displacementof the protoconid with respect to the metaconid, and the de-gree of development of the transverse cristids (i.e., marked ele-vation), the occlusal pattern of lower molars of Turkanamys bet-ter matches the condition found in Metaphiomys (Wood, 1968;Fejfar, 1987). Our original description of KNM-LH 47895 (m2;Fig. 3E; also figured in Ducrocq et al., 2010:fig. 3A) had led usto refer this isolated lower tooth to Metaphiomys cf. M. schaubi.The new dental material now attributed to Turkanamys allowsus to better assess the dental differences with Metaphiomys. Thedistinction between lower molars of both taxa is, however, ten-uous. In particular, Turkanamys tends to have a lingual wallbetter marked, whereas Metaphiomys does not. Indeed, lowermolars of Turkanamys display a stronger and longer posteriorarm of the metaconid, which connects a small mesostylid (absentin Metaphiomys), the posterolophid is higher and longer and itconnects the distal aspect of the entoconid, thereby closing themetaflexid lingually (open in M. schaubi, more or less closed inM. beadnelli). Some lower molars of Turkanamys are also char-acterized by a complete metalophulid II, whereas in species ofMetaphiomys, this character is highly variable (Metalophulid IIvery often incomplete). Lower molar of Turkanamys display notrace of protospur extending backward from the posterior armof the protoconid as it does in Metaphiomys. Lower molar ofMetaphiomys display also no trace of lingual mesolophid as itdoes in some teeth of Turkanamys and Phiomys. In contrast,the dental pattern of upper molars of Turkanamys differs radi-cally from that of Metaphiomys. Upper molars of Metaphiomyshave no metaloph-mesolophule connection (only the metaloph-posteroloph connection), they lack the centroloph, have nomesostyle, and show a lesser development of the mesolophule.Finally, in contrast to Metaphiomys, Turkanamys replaced up-per and lower deciduous premolars with permanent premolars.Turkanamys differs also substantially from Kahawamys (Stevenset al., 2009) in its larger size, and mainly in having longer teeth,which bear a stronger development of the posterior arm of theprotoconid (which is absent in Kahawamys) and a longer hy-polophid. Lower molars of Kahawamys are also characterizedby a stronger development of the lingual wall, with no trace ofmesostylid.
Compared with more basal ‘phiomyids’ (i.e., late Eoceneforms: Protophiomys, Waslamys, Talahphiomys), which arebunodont to bunolophodont (crests and cristids present but lowand often short), the dental pattern of Turkanamys is muchmore lophodont (crests and cristids elevated and long), al-though the main cusps and cuspids still remain well pronounced(bunolophodont). In addition, upper molars of Turkanamys haveno metaconule; they lack the neo-endoloph, which generallycloses the hypoflexus lingually (notably in Waslamys and insome species of Protophiomys), but they develop a strong
longitudinal mure (connection between the protoloph andthe mesial extremity of the anterior arm of the hypocone),thereby closing the hypoflexus mediobuccally. Lower molars ofTurkanamys also differ in exhibiting a strong development ofthe metalophulid II (very short posterior arm of the protoconidin Protophiomys, Waslamys, and Talahphiomys) and a stronglyreduced (virtually indistinct) hypoconulid on the posterolophid.In comparing Turkanamys with early Miocene thryonomyoidrodents from Kenya-Uganda (e.g., Paraphiomys, Diamantomys,Simonimys, Kenyamys, Epiphiomys, Lavocatomys, Myophiomys,Andrewsimys, Ugandamys; Lavocat, 1973; Winkler et al., 2005;Holroyd and Stevens, 2009), Turkanamys appears morpholog-ically quite conservative. Except for the hexalophodonty of itsupper molars, Turkanamys preserves the mesial connection ofthe metaloph, a condition as mentioned above that is otherwisefound only in basal ‘phiomyids’ and more generally in early hys-tricognaths. Almost all thryonomyoids from the early Miocene ofKenya-Uganda, except one form referred to Phiomys andrewsi(KNM-SO 884; Lavocat, 1973; but see Holroyd and Stevens,2009), are morphologically advanced over Turkanamys in havinglost the mesial connection of the metaloph, and in developingonly the metaloph-posteroloph connection. In addition, all theseearly Miocene taxa are also very specialized and somewhat diver-gent in different ways from Turkanamys. For instance, Simon-imys has molars that are longer than wide, with well-developed(elevated) but slender transverse crests and cristids, and the maincusps and cuspids are nearly indistinct. In contrast, Epiphiomysand Paraphiomys exhibit very inflated cusps and cuspids, and thetransverse and longitudinal crests and cristids are very thick (mas-sive). The minute taxon Kenyamys displays lower molars with adental pattern that is close to that of lower molars in Turkanamys(highly crested, complete metalophulid II, strong, elevated poste-rior arm of the metaconid, entoconid-posterolophid connection).The same is true for lower molars of Lavocatomys (Holroydand Stevens, 2009; Phiomys andrewsi sensu Lavocat, 1973)that are characterized, as in Turkanamys, by a bunolophodontpattern but with a lesser development of the posterior arm of theprotoconid (incomplete metalophulid II). However, the cusp andcrest arrangement of upper molars of Kenyamys (high transversecrests, cusps nearly indistinct, strong mesostyle and transversehypoflexus) differs radically from that observed in Turkanamys.The small ‘phiomyid,’ Andrewsimys, differs also substantiallyfrom Turkanamys in having taeniodont upper molars (absenceof the lingual protoloph, and hypoflexus confluent with theparaflexus). The dental pattern of Myophiomys is also highlydivergent from that of Turkanamys, but also from that of allother early Miocene thryonomyoids. Upper and lower teeth ofMyophiomys are particularly low crowned (brachydont) and lowcrested (crests and cristids very low and thin), and the main cuspsand cuspids are moderately bulbous and salient (bunodonty).Finally, among the thryonomyoid rodents recorded in EastAfrica near the Paleogene/Neogene transition, Diamantomys(D. timius and D. luederitzi; Stromer, 1926; Lavocat, 1973;Rasmussen and Gutierrez, 2009) may appear as one of the mostdivergent taxa. Diamantomys is characterized by high-crowned(hypsodont) and rectangular teeth, with very thick, oblique crestsand cristids, and very sharp cusps and cuspids. The dental patternof Diamantomys is highly specialized, and as such it is divergentfrom that of Turkanamys and of ‘phiomyids’ in general.
DISCUSSION AND CONCLUSIONS
The evolutionary history of rodents in Africa during theend of the Paleogene, especially during the late Oligocene, israther misunderstood because of a very limited fossil record(e.g., Winkler et al., 2010). For this time period, which rangesover 5 million years, only three genera of hystricognathous ro-dents (Metaphiomys, Kahawamys, and Diamantomys) have been
Dow
nloa
ded
by [
Lau
rent
Mar
ivau
x] a
t 09:
04 0
3 M
ay 2
012

MARIVAUX ET AL.—LATE OLIGOCENE PHIOMYID RODENT FROM KENYA 655
so far reported from a couple of localities situated exclusivelyin sub-Saharan East Africa (Tanzania and Kenya; Stevens etal., 2006, 2009; Rasmussen and Gutierrez, 2009; Ducrocq etal., 2010). In the absence of a comprehensive late Oligocenefossil record, it remains therefore quite difficult to link thediverse late Eocene–early Oligocene rodent faunas of Saha-ran Africa and Oman with the highly diversified and well-sampled early Miocene African faunas. Paleogene rodent faunasincluded primarily ‘phiomorphs’ (“Phiomyidae,” Gaudeamuri-dae, and Myophiomyidae) and anomaluroids (“Zegdoumyidae”and Anomaluridae), whereas early Miocene faunas included awide array of ‘phiomorphs’ (Thryonomyoidea [Thryonomyidea,“Phiomyidae,” Myophiomyidae, Kenyamyidae, Diamantomyi-dae], Hystricoidea [Hystricidae], and Bathyergoidea [Bathyergi-dae]) and Anomaluridae, but also Pedetidae, Ctenodactylidae,Nesomyidae, Pseudocricetodontidae, Spalacidae, and Sciuridae(Lavocat, 1973; Pickford et al., 2003; Winkler et al., 2010). Itis generally admitted that some of these rodent groups (no-tably ctenodactylids, muroids, and sciuroids) entered Africa fromEurasia by the earliest Miocene, as part of the faunal inter-changes (e.g., Winkler et al., 2010; Orliac et al., 2010; Antoine,Metais, et al., 2012) linked to the collision between the Afro-Arabian and Eurasian plates (e.g., Jolivet and Faccenna, 2000). Incontrast, hystricognaths (‘phiomorphs’), as well as anomaluroidsand possibly pedetids, evolved within Africa from the Eoceneto the early Miocene and beyond (Lavocat, 1973; Sallam, Seif-fert, and Simons, 2010; Sallam, Seiffert, et al., 2010; Marivaux etal., 2011). Stem ‘phiomorphs’ seem to have invaded Africa fromAsia at least by the late middle Eocene and achieved their initialadaptive radiation on that continent (Jaeger et al., 1985; Mari-vaux et al., 2000, 2002; Sallam et al., 2009; Marivaux and Jaegerin Jaeger et al., 2010), whereas anomaluroids show a greater an-tiquity in Africa, which can be traced back to the early Eocene (asdocumented by the “Zegdoumyidae”; Vianey-Liaud et al., 1994;Marivaux et al., 2011).
The discovery of the ‘phiomyid’ Turkanamys hexalophus high-lights the persistence of this ancestral ‘phiomorph’ group atthe end of the Paleogene. In fact, Turkanamys shares a simi-lar dental bauplan with early Afro-Asian hystricognaths (early‘phiomyids’ and ‘baluchimyines’), and as such it appears some-what evolutionarily conservative (primitive) with respect to someOligocene (e.g., Metaphiomys, Gaudeamus, Diamantomys) andall the Miocene forms. Conversely, Turkanamys is also highlyautapomorphic in developing a hexalophodont pattern on theupper molars, which might indicate some dietary specializa-tion. Interestingly, the subtlety of the morphological dental dif-ferences between Phiomys and Turkanamys may indicate thatthey are closely related taxa. However, in some other morpho-logical details, Turkanamys is more advanced (or specialized)over Phiomys, which suggests that Turkanamys could repre-sent a late-diverging form of the Phiomys lineage, but withoutMiocene representative. In sum, Turkanamys appears as a spe-cialized ‘phiomyid,’ which seems relatively remote phylogenet-ically from the taxa documenting the numerous early Miocenehystricognath families from Kenya-Uganda (Lavocat, 1973; Win-kler et al., 2005). In other words, Turkanamys can be consid-ered as a relic ‘phiomyid’ species, a conclusion that has alsobeen proposed by Ducrocq et al. (2011) regarding the para-pithecoid species recorded in Lokone (LOK 13; Lokonepithe-cus). The late Eocene through early Miocene record in Africashows that hystricognathous rodents were particularly abundantand diversified. However, the important diversity of this grouprecorded as early as the early Miocene, including seven familiescontra three for the Paleogene, would suggest a second majoradaptive radiation in Africa that probably occurred during thelate Oligocene. This time window is so far only documented inEast Africa, and the hystricognath record is still limited because
it consists only of two members of the ancestral ‘phiomorph’group (‘phiomyid’: Metaphiomys cf. beadnelli and Turkanamyshexalophus), one representative of the Miocene family Diaman-tomyidae (Diamantomys timius), and Kahawamys mbeyaensis ofuncertain familial affinities. Miocene thryonomyids (e.g., Para-phiomys, Paraulacodus) and diamantomyids are clearly nestedwithin the early ‘phiomorph’ radiation (Wood, 1968; Lavocat,1973; Marivaux et al., 2002, 2004; Lopez-Antonanzas et al., 2004;Lopez-Antonanzas and Sen, 2005; Sallam et al., 2011), whereashystricids seem to be more closely related to gaudeamurids (Sal-lam et al., 2011). Kenyamyidae and Myophiomyidae are alsoprobably late diverging ‘phiomyid’-like rodents, but their phy-logenetic positions have never been clearly established. In ad-dition, the phylogenetic origin of the Bathyergidae among earlyhystricognaths remains obscure, inasmuch as there is no firmlyestablished Paleogene record for this group. However, bathyer-goids seem to have diverged from the thryonomyoid lineages dur-ing the early Oligocene (Sallam et al., 2009). According to thesephylogenetic assumptions, most Miocene families of hystricog-naths are supposed to be present during the Oligocene. The lateOligocene diversity of hystricognathous rodents in Africa is thusclearly undersampled. Given the great familial diversity of hys-tricognaths in the early Miocene of East Africa, the virtual ab-sence of their stem representatives in the late Oligocene record ofthe same province implies either important taphonomic/samplingbiases or a real absence of these families in this province at thattime. This would indicate that hystricognathous rodents diversi-fied elsewhere in Africa and colonized East Africa by the earlyMiocene. The early evolutionary and biogeographic history ofhystricognathous rodents on the vast African landmass is there-fore far from being well understood given the scarce and veryscattered fossil record.
Recently, Pickford et al. (2008) have reported the discovery ofa diverse rodent assemblage from the middle Eocene (Lutetian)of the Sperrgebiet, Namibia. This is the southernmost Paleo-gene mammal sites of sub-Saharan Africa. Surprisingly, thisassemblage includes a mixture of primitive forms including a‘zegdoumyid’ (Glibia namibiensis [Zegdoumys namibiensis sensuMarivaux et al., 2011]) and a basal ‘phiomyid’ (Protophiomyscf. algeriensis), together with taxa that show closer affinities toMiocene rodents: a diamantomyid (Prepomonomys bogenfelsi),a thryonomyid (Apodecter cf. stromeri), a myophiomyid (Sili-camys cingulatus), and a bathyergid (cf. Bathyergoides sp.). Thisfind would thus extend back to the middle Eocene the first oc-currence of most of the Oligo-Miocene hystricognathous rodentfamilies. A middle Eocene age for the deposits that have yieldedthe “Zegdoumyidae” and the basal “Phiomyidae” is reasonablyconceivable. However, the presence of typical Miocene generaor even species in this assemblage would imply a temporal exten-sion of about 25 million years for these taxa, whereas the meanand/or median mammalian species duration is generally between2.1 and 2.6 millions years (Alroy, 2000; Vrba and DeGusta,2004; Holroyd and Stevens, 2009). Given this mixed assemblageincluding primitive rodents and an evolutionarily advanced setof hystricognaths, it may be expected that these fossils do notbelong to a single faunal unit, and that the Sperrgebiet depositsrepresent a multiphased carbonated depression filling. Regard-less the Miocene taxa, the presence of a stem anomaluroids(“Zegdoumyidae”) and a stem ‘phiomorphs’ in the Sperrgebietdeposits indicates that both groups had a widespread distributionin Africa during some part of the Eocene, which ranged fromthe northernmost to southernmost latitudes. The vast Africanlandmass, which probably displayed various ecosystems duringthe Paleogene, has certainly housed several epicenters of rodentradiation. Only paleontological field efforts will provide informa-tion for highlighting the evolutionary and biogeographic historyof these African mammal groups during the early Tertiary.
Dow
nloa
ded
by [
Lau
rent
Mar
ivau
x] a
t 09:
04 0
3 M
ay 2
012

656 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 32, NO. 3, 2012
ACKNOWLEDGMENTS
We thank the Ministry of Science and Technology of Kenyafor permission to conduct our research in the Turkana Basin.We acknowledge I. O. Farah (Director General of NMK), E.Mbua, M. Muungu, and the staff of the NMK for their assis-tance with collections and administrative authorizations. We arethankful to M. G. Leakey and R. E. Leakey, without whom theLokone project would not have been initiated. We warmly thankJ.-J. Tiercelin for his valuable help on the field. Logistical andfield assistance were provided by M. Macharwas, F. Emekui, J.Makemkou, A. Londung, J. Manai, and people of Loperot andLomeloku. Many thanks to M. Vianey-Liaud, A. Ramdarshan,J.-R. Boisserie, and P.-O. Antoine for advices and fruitful discus-sions on rodent evolution. Special thanks are extended to A.-L.Charruault and M. Orliac for technical assistances, and L. Mes-lin for scientific drawings. Finally, we thank the three anony-mous reviewers, who have provided constructive remarks on themanuscript. The Agence Nationale de la Recherche (ANR) pro-grams PALASIAFRICA ANR-08-JCJC-0017 (ANR-ERC) andANR-09-BLAN-0238 supported this research. ISE-M publica-tion no. 2012-001.
LITERATURE CITED
Alroy, J. 2000. New methods for quantifying macroevolutionary patternsand processes. Paleobiology 26:707–733.
Antoine, P.-O., G. Metais, M. J. Orliac, J.-Y. Crochet, L. J. Flynn, L.Marivaux, A. R. Rajpar, G. Roohi, and J.-L. Welcomme. 2012.Mammalian Neogene biostratigraphy of the Sulaiman Province,Pakistan; in X. Wang, L. J. Flynn, and M. Fortelius (eds.), NeogeneTerrestrial Mammalian Biostratigraphy and Chronology of Asia.Columbia University Press, New York.
Antoine, P.-O., L. Marivaux, D. A. Croft, G. Billet, M. Ganerød, C.Jaramillo, T. Martin, M. J. Orliac, J. Tejada, A. J. Altamirano, F.Duranthon, G. Fanjat, S. Rousse, and R. S. Gismondi. 2012. Mid-dle Eocene rodents from Peruvian Amazonia reveal the pattern andtiming of caviomorph origins and biogeography. Proceedings of theRoyal Society of London B 279:1319–1326.
Bowdich, T. E. 1821. An analysis of the natural classifications of Mam-malia for the use of students and travellers; in J. Smith (ed.), Volume115, Paris.
Candela, A. M. 1999. The evolution of the molar pattern of the Erethizon-tidae (Rodentia, Hystricognathi) and the validity of ParasteiromysAmeghino, 1904. Palaeovertebrata 28:53–73.
Coster, P., M. Benammi, V. Lazzari, G. Billet, T. Martin, M. Salem,A. A. Bilal, Y. Chaimanee, M. Schuster, X. Valentin, M. Brunet,and J.-J. Jaeger. 2010. Gaudeamus lavocati sp. nov. (Rodentia, Hys-tricognathi) from the early Oligocene of Zallah, Libya: first Africancaviomorph? Naturwissenschaften 97:697–706.
de Bruijn, H., and S. T. Hussain. 1985. Thryonomyidae from the lowerMarchar Formation of Sindh, Pakistan. Palaeontology 88:155–166.
de Bruijn, H., E. Unay, G. Sarac, and A. Yılmaz. 2003. A rodent as-semblage from the Eo/Oligocene boundary interval near Sungulu,Lesser Caucasus, Turkey; pp. 47–76 in N. Lopez-Martınez, P.Pelaez-Campomanes, and M. Henandez Fernandez (eds.), Colo-quios de Paleontologia. Volumen Extraordinario 1 En honor alDr. Remmert Daams. Surrounding Fossil Mammals: Dating, Evolu-tion and Paleoenvirontment, Servicio de Publicaciones UniversidadComplutense de Madrid.
Ducrocq, S., K. F. Manthi, and F. Lihoreau. 2011. First record of a para-pithecid primate from the Oligocene of Kenya. Journal of HumanEvolution 61:327–331.
Ducrocq, S., J.-R. Boisserie, J.-J. Tiercelin, C. Delmer, G. Garcia, K. F.Manthi, L. Marivaux, O. Otero, S. Peigne, P. Tassy, and F. Lihoreau.2010. New Oligocene vertebrate localities from Northern Kenya(Turkana Basin). Journal of Vertebrate Paleontology 30:293–299.
Fejfar, O. 1987. Oligocene rodents from Zallah Oasis, Libya. MunchnerGeowiss Abhandlungen A 10:265–268.
Flynn, L. J., L. L. Jacobs, and I. U. Cheema. 1986. Baluchimyinae, a newctenodactyloid rodent subfamily from the Miocene of Baluchistan.American Museum Novitates 2841:1–58.
Frailey, C. D., and K. E. Campbell. 2004. Paleogene rodents from Amazo-nian Peru: the Santa Rosa local fauna; pp. 71–130 in K. E. Campbell(ed.), The Paleogene Mammalian Fauna of Santa Rosa, AmazonianPeru. Natural History Museum of Los Angeles County, Los Ange-les, California.
Holroyd, P. 1994. An examination of dispersal origins for Fayum mam-mals. Ph.D. dissertation, Duke University, Durham, North Carolina,328 pp.
Holroyd, P. A., and N. J. Stevens. 2009. Differentiation of Phiomysandrewsi from Lavocatomys aequatorialis (n. gen., n. sp.) (Ro-dentia: Thryonomyoidea) in the Oligo-Miocene interval on con-tinental Africa. Journal of Vertebrate Paleontology 29:1331–1334.
Jaeger, J.-J., C. Denys, and B. Coiffait. 1985. New Phiomorpha andAnomaluridae from the late Eocene of North-West Africa: phylo-genetic implications; pp. 567–588 in W. P. Luckett and J.-L. Harten-berger (eds.), Evolutionary Relationships among Rodents, A Multi-disciplinary Analysis. Nato, New York.
Jaeger, J.-J., L. Marivaux, M. Salem, A. A. Bilal, Y. Chaimanee, B.Marandat, X. Valentin, P. Duringer, M. Schuster, M. Benammi, E.Metais, and M. Brunet. 2010. New rodent assemblages from theEocene Dur at-Talhah escarpment (Sahara of Central Libya): sys-tematic, biochronologic and paleobiogeographic implications. Zoo-logical Journal of the Linnean Society 160:195–213.
Jolivet, L., and C. Faccenna. 2000. Mediterranean extension and theAfrica-Eurasia collision. Tectonics 19:1095–1106.
Korvenkontio, V. A. 1934. Mikroskopische Untersuchungen an Nager-incisiven unter Hinweis auf die Schmelzstruktur der Backenzahne.Annales Zoologici Societatis Zoologici—Botanicae FennicaeVanamo 2:1–274.
Lavocat, R. 1962. Reflexions sur l’origine et la structure du groupedes Rongeurs. Colloques internationaux Problemes actuels dePaleontologie (Evolution des Vertebres). CNRS, Paris.
Lavocat, R. 1973. Les Rongeurs du Miocene d’Afrique Orientale.Memoires et Travaux de l’Institut de Montpellier de l’Ecole Pra-tique des Hautes Etudes 1:1–284.
Lavocat, R. 1976. Rongeurs Caviomorphes de l’Oligocene de Bolivie; IIRongeurs du bassin Deseadien de Salla-Luribay. Palaeovertebrata7:15–90.
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundumclasses, ordines, genera, species cum characteribus, differentris, syn-omymis, locis, Editis decima, reformata, Volume 1. Laurentii Salvii,Stockholm, 824 pp.
Lopez-Antonanzas, R., and S. Sen. 2005. New species of Paraphiomys(Rodentia Thryonomyidae) from the Lower Miocene of As-Sarrar,saudi Arabia. Palaeontology 48:223–233.
Lopez-Antonanzas, R., S. Sen, and P. Mein. 2004. Systematics and phy-logeny of the cane rats (Rodentia: Thryonomyidae). ZoologicalJournal of the Linnean Society 142:423–444.
Marivaux, L., and J.-L. Welcomme. 2003. Diatomyid and baluchimyinerodents from the Oligocene of Pakistan (Bugti Hills, Balochistan):systematic and paleobiogeographic implications. Journal of Verte-brate Paleontology 23:420–434.
Marivaux, L., M. Vianey-Liaud, and J.-J. Jaeger. 2004. High-level phy-logeny of early Tertiary rodents: dental evidence. Zoological Jour-nal of the Linnean Society 142:105–132.
Marivaux, L., J.-L. Welcomme, M. Vianey-Liaud, and J.-J. Jaeger. 2002.The role of Asia in the origin and diversification of hystricognathousrodents. Zoologica Scripta 31:225–239.
Marivaux, L., M. Benammi, S. Ducrocq, J.-J. Jaeger, and Y. Chaima-nee. 2000. A new baluchimyine rodent from the Late Eocene ofthe Krabi Basin (Thailand): paleobiogeographic and biochronologicimplications. Comptes rendus de l’Academie des Sciences, Paris331:427–433.
Marivaux, L., M. Adaci, M. Bensalah, H. Gomes Rodrigues, L. Hautier,M. Mahboubi, F. Mebrouk, R. Tabuce, and M. Vianey-Liaud. 2011.Zegdoumyidae (Rodentia, Mammalia), stem anomaluroid rodentsfrom the early to middle Eocene of Algeria (Gour Lazib, WesternSahara): new dental evidence. Journal of Systematic Palaeontology9:563–588.
Martin, T. 1994. African origin of caviomorph rodents is indicated by in-cisor enamel microstructure. Paleobiology 20:5–13.
Orliac, M. J., J.-R. Boisserie, L. MacLatchy, and F. Lihoreau. 2010.Early Miocene hippopotamids (Cetartiodactyla) constrain the phy-logenetic and spatiotemporal settings of hippopotamid origin.
Dow
nloa
ded
by [
Lau
rent
Mar
ivau
x] a
t 09:
04 0
3 M
ay 2
012

MARIVAUX ET AL.—LATE OLIGOCENE PHIOMYID RODENT FROM KENYA 657
Proceedings of the National Academy of Sciences of the UnitedStates of America 107:11871–11876.
Osborn, H. F. 1908. New fossil mammals from the Fayum Oligocene,Egypt. American Museum of Natural History Bulletin 26:415–424.
Pickford, M., B. Senut, D. Gommery, and E. Musiime. 2003. New catar-rhine fossils from Moroto II, early Middle Miocene (ca 17.5 Ma)Uganda. Comptes Rendus Palevol 2:649–662.
Pickford, M., B. Senut, J. Morales, P. Mein, and I. M. Sanchez. 2008.Mammalia from the Lutetian of Namibia. Memoirs of the Geologi-cal Survey of Namibia 20:465–514.
Rasmussen, D. T., and M. Gutierrez. 2009. A mammalian fauna fromthe Late Oligocene of Northwestern Kenya. Palaeontographica288:1–52.
Sallam, H. M., E. R. Seiffert, and E. L. Simons. 2010. A highly derivedanomalurid rodent from the earliest late Eocene of Egypt. Palaeon-tology 53:803–813.
Sallam, H. M., E. R. Seiffert, and E. L. Simons. 2011. Craniodental mor-phology and systematics of a new family of hystricognathous rodents(Gaudeamuridae) from the Late Eocene and Early Oligocene ofEgypt. Plos One 6:1–29.
Sallam, H. M., E. R. Seiffert, E. L. Simons, and C. Brindley. 2010. A large-bodied anomaluroid rodent from the earliest late Eocene of Egypt:phylogenetic and biogeographic implications. Journal of VertebratePaleontology 30:1579–1593.
Sallam, H. M., E. R. Seiffert, M. E. Steiper, and E. L. Simons. 2009. Fos-sil and molecular evidence constrain scenarios for the early evolu-tionary and biogeographic history of hystricognathous rodents. Pro-ceedings of the National Academy of Sciences of the United Statesof America 106:16722–16727.
Stevens, N. J., P. A. Holroyd, E. M. Roberts, P. M. O’Connor, and M.D. Gottfried. 2009. Kahawamys mbeyaensis (n. gen., n. sp.) (Roden-tia: Thryonomyoidea) from the Late Oligocene Rukwa Rift Basin,Tanzania. Journal of Vertebrate Paleontology 29:631–634.
Stevens, N. J., P. M. O’Connor, M. D. Gottfried, E. M. Roberts,S. Ngasala, and M. R. Dawson. 2006. Metaphiomys (Rodentia:Phiomyidae) from the Paleogene of Southwestern Tanzania. Jour-nal of Vertebrate Paleontology 80:407–410.
Stromer, E. 1926. Reste land und Susswasser Bewohnender Wirbeltiereaus den Diamantfeldern Deutsch Sudwestafrikas; pp. 107–153 in E.Kaiser (ed.), Die Diamantenwuste Sudwestafrika. Reimer Dietrich,Berlin.
Thomas, H., J. Roger, S. Sen, and Z. Al-Sulaimani. 1992. Early Oligocenevertebrates from Dhofar (Sultanate of Oman); pp. 283–293 in A.
Sadek (ed.), Geology of the Arab World. Cairo University, Cairo,Egypt.
Thomas, H., S. Roger, S. Sen, C. Bourdillon-de-Grissac, and Z.Al-Sulaimani. 1989. Decouverte de vertebres fossiles dansl’Oligocene inferieur du Dhofar (Sultanat d’Oman). Geobios 22:101–120.
Tullberg, T. 1899. Ueber das system der nagethiere: eine phylogenetis-che studie. Nova Acta Regiae Societatis Scientiarum Upsaliens18:1–514.
Vianey-Liaud, M., J.-J. Jaeger, J.-L. Hartenberger, and M. Mahboubi.1994. Les rongeurs de l’Eocene d’Afrique Nord-Occidentale (GlibZegdou (Algerie) et Chambi (Tunisie)) et l’origine des Anomaluri-dae. Palaeovertebrata 23:93–118.
Vrba, E. S., and D. DeGusta. 2004. Do species populations really startsmall? New perspectives from the Late Neogene fossil record ofAfrican mammals. Philosophical Transactions of the Royal Society,London 359:285–293.
Vucetich, M. G., E. C. Vieytes, M. E. Perez, and A. A. Carlini. 2010. Therodents from La Cantera and the early evolution of caviomorphsin South America; pp. 193–205 in R. H. Madden, A. A. Carlini,M. G. Vucetich, and R. F. Kay (eds.), The Paleontology of GranBarranca. Evolution and Environmental Change through the Mid-dle Cenozoic of Patagonia. Cambridge University Press, Cambridge,U.K.
Winkler, A. J., C. Denys, and D. M. Avery. 2010. Fossil rodents of Africa;pp. 263–304 in W. J. Sanders and L. Werdelin (eds.), Cenozoic Mam-mals of Africa. University of California Press, Berkeley, California.
Winkler, A. J., L. MacLatchy, and M. Mafabi. 2005. Small rodents and alagomorph from the early Miocene Bukwa locality, eastern Uganda.Palaeontologia Electronica 8(1): Article 24A, 12 pp.
Wood, A. E. 1955. A revised classification of the rodents. Journal ofMammalogy 36:165–187.
Wood, A. E. 1968. Part II: The African Oligocene Rodentia; pp. 23–105 inJ. E. Remington (ed.), Early Cenozoic Mammalian Faunas FayumProvince, Egypt. Peabody Museum of Natural History Yale Univer-sity, New Haven.
Wood, A. E., and R. W. Wilson. 1936. A suggested nomenclature forthe cusps of the cheek teeth of Rodents. Journal of Paleontology10:388–391.
Submitted September 19, 2011; revisions received December 19, 2011;accepted January 9, 2012.Handling editor: Guillermo Rougier.
Dow
nloa
ded
by [
Lau
rent
Mar
ivau
x] a
t 09:
04 0
3 M
ay 2
012
Related Documents











