(This is a sample cover image for this issue. The actual cover is not yet available at this time.) This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

(This is a sample cover image for this issue. The actual cover is not yet available at this time.)
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
A multidisciplinary approach to reconstructing the chronology and environmentof southwestern European Neanderthals: the contribution of Teixoneres cave(Moià, Barcelona, Spain)
Juan Manuel López-García a,b,*, Hugues-Alexandre Blain a,b, Francesc Burjachs a,b,c, Anna Ballesteros a,b,Ethel Allué a,b, Gloria Ericka Cuevas-Ruiz d, Florent Rivals a,b, Ruth Blasco a,b, Juan Ignacio Morales a,b,Antonio Rodríguez Hidalgo a,b, Eudald Carbonell a,b,e, David Serrat f, Jordi Rosell a,b
a IPHES, Institut Català de Paleoecologia Humana i Evolució Social, C/Escorxador s/n, 43003 Tarragona, SpainbÀrea de Prehistòria, Universitat Rovira i Virgili (URV), Avinguda de Catalunya 35, 43002 Tarragona, Spainc ICREA (Institució Catalana de Recerca i Estudis Avançats), Barcelona, Catalonia, SpaindMuseo de Paleontología, Área Académica de Biología, Universidad Autónoma del Estado de Hidalgo, Ciudad Universitaria s/n, Carretera Pachuca-Tulancingo km 4.5,C.P. 42184 Pachuca, Hidalgo, Mexicoe Institute of Vertebrate Paleontology and Paleoanthropology of Beijing (IVPP), ChinafDepartament de Geodinàmica i Geofísica, Universitat de Barcelona, Facultat de Geologia, C/Martí i Franquès s/n, 08028 Barcelona, Spain
a r t i c l e i n f o
Article history:Received 17 January 2012Received in revised form5 April 2012Accepted 6 April 2012Available online xxx
Keywords:PollenCharcoalHerpetofaunaSmall and large mammalsBiochronologyPaleoenvironmentPaleoclimateNeanderthalsSouthwestern Europe
a b s t r a c t
According to pollen analysis and the 18O-isotope curve, the first part of the Late Pleistocene (ca 128e30ka) is mainly characterized by a dynamic that alternates cold phases (Heinrich Events) and temperatephases (interstadials). These rapid fluctuations provide the context for the Neanderthal occupations inthe northeastern part of the Iberian Peninsula. In this paper we present the chronological, environmentaland climatic data obtained by analyzing the pollen, the charcoal, the small vertebrates (amphibians,squamates and small mammals) and the large-mammal dental wear at the Neanderthal site of Teix-oneres cave, Northeastern Iberia. Levels II and III from this cavity have provided Mousterian industriesand other evidence of Neanderthal occupations, such as cut-marks in large-mammal bones. A multiproxystudy such as this constitutes a new approach to the chronological, environmental and climatic contextin which Neanderthal populations lived in southwestern Europe (Iberian Peninsula). The results allow usto establish a relative chronology for these two levels of between ca 30e90 ka and show that they areassociated with different environmental and climatic conditions: temperate and humid for Level III andcold and dry for Level II. This demonstrates that the Neanderthals were well adapted to the territory thatthey occupied, irrespective of the climatic conditions.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
The Late Pleistocene climatic history of the Iberian Peninsula hasbeen well known since 1995, when the Iberian marine margin wascored as part of the IMAGES program (International Marine GlobalChange Study). The samples obtained show the continuous anddetailed development of the north and south ecosystems of theIberian Peninsula and the climatic changes produced in thenortheast Atlantic over the last 140,000 years (Cacho et al., 1999;
Sánchez-Goñi and d’Errico, 2005). The Greenland 18O-isotopiccurve detects forty sudden changes in temperatures during the last123,000 years, called DansgaardeOeschger events (DeO events)(Johnsen et al., 1992; Dansgaard et al., 1993; Cacho et al., 1999).Among these oscillations certain cold phases marked by theconcentration of Iceberg Rafted Detritus (IRD), indicating an influxof icebergs to the Iberian shores (Cacho et al., 1999; Bard et al.,2000; Sánchez-Goñi et al., 2000; Naughton et al., 2009), havebeen identified; these are called Heinrich Events (Heinrich, 1988).Six of these phases have been clearly identified in the surveys of thesouthwestern Iberian margin. The climate of the Late Pleistocenebegan with Marine Isotope Stage 5 (MIS 5) (ca 128e74 ka BP),characterized by a first mild phase called the Eemian or MIS 5e (ca128e110 ka BP), which saw the maximum expansion of the
* Corresponding author. IPHES, Institut Català de Paleoecologia Humana i Evo-lució Social, C/Escorxador s/n, 43003 Tarragona, Spain.
E-mail address: [email protected] (J.M. López-García).
Contents lists available at SciVerse ScienceDirect
Quaternary Science Reviews
journal homepage: www.elsevier .com/locate/quascirev
0277-3791/$ e see front matter � 2012 Elsevier Ltd. All rights reserved.doi:10.1016/j.quascirev.2012.04.008
Quaternary Science Reviews 43 (2012) 33e44

Author's personal copy
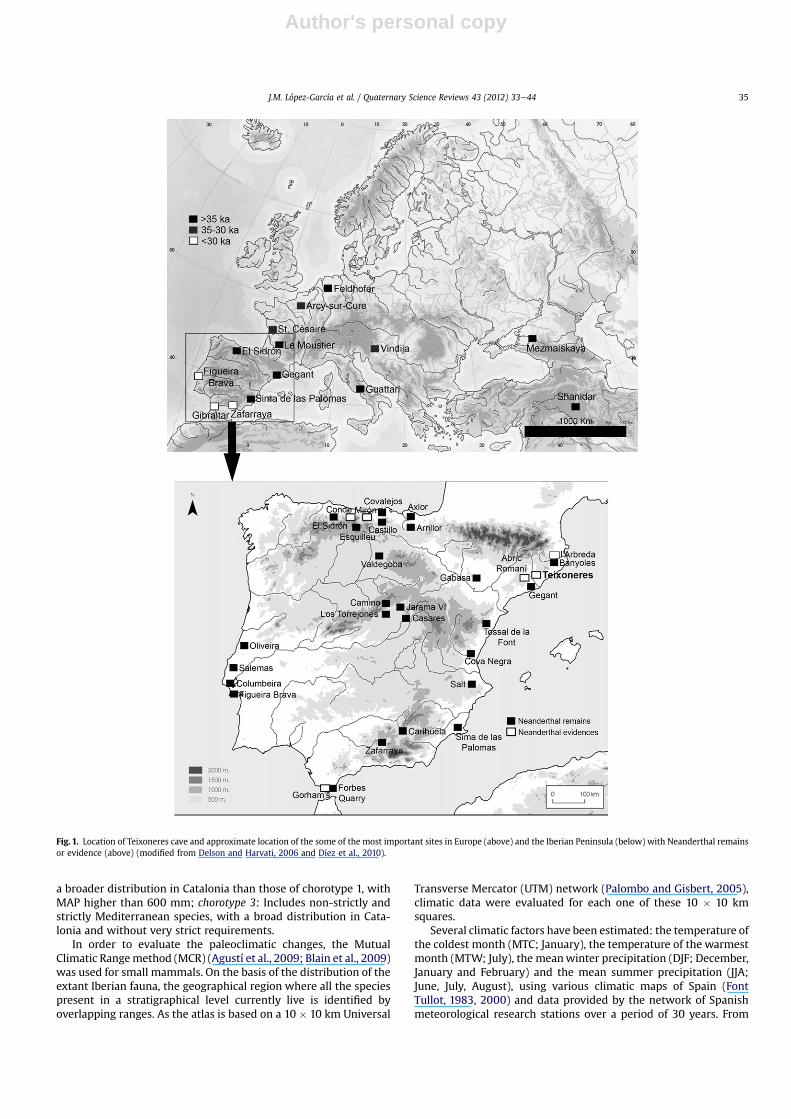
Mediterranean forest in southwestern Iberia. This phase is followedby four cold periods alternating with four mild periods (ca110e74 ka BP), characterized by the development of steppeformations in cold spells, together with the expansion of the openforest in warm spells (Shackleton et al., 2003; Sánchez-Goñi andd’Errico, 2005). The following period, occurring between 74 ande60 ka BP, is Marine Isotope Stage 4 (MIS 4), which is characterizedby summer isolation minima over the northern latitudes of thenorthern hemisphere, producing the maximum extension of icecaps, a sea-level hundreds of meters below the current level, andlow ocean temperatures (Sánchez-Goñi and d’Errico, 2005). After-wards, Marine Isotope Stage 3 (MIS 3) (ca 60e30 ka BP) is charac-terized by a dynamic that alternates between phases of forestdevelopment and expansion of semi-arid areas in accordance withthe warming and cooling, respectively, of the marine surfacetemperatures (Sánchez-Goñi and d’Errico, 2005; Fletcher andSánchez-Goñi, 2008). Such multiple fluctuations provide thecontext for the Neanderthal occupations in Europe. There are manyEuropean sites with Neanderthal remains (Delson and Harvati,2006), and in recent years there has been an increase in findingsof Neanderthal remains in the Iberian Peninsula (Díez et al., 2010)(Fig. 1). These have shown that the Neanderthals survived in Iberiaas late as ca 30e28 ka BP in the context of a climatic refugiumassociated with woodland formations (Finlayson et al., 2006;Finlayson and Carrión, 2007; Jennings et al., 2011). Against thisbackground of rapid climatic fluctuations and Neanderthal occu-pations in Iberia, we here present the chronological, environmentaland climatic data obtained by analyzing the small vertebrates(amphibians, squamates and small mammals), the dental wear inlarge mammals, and the pollen and charcoal records at the Nean-derthal occupation site of Teixoneres cave, putting our data intocontext with the climatic phases of the Late Pleistocene and withthe Neanderthal occupations in the rest of the Iberian Peninsula.
2. Teixoneres cave
Teixoneres cave is one of the caves belonging to the karsticsystem called the Toll caves. Located near the village of Moià(Barcelona, Spain) at 900 m a.s.l. (Fig. 1), it forms a course ofgalleries more than 2 km long that contains several archaeologicaldeposits fromdifferent chronologies. Some of them, such as the oneknown as the South Gallery of the Toll caves, have been partiallyexcavated since the 1950s and 1970s and report an importantHolocene sequence and Late Pleistocene paleontological record(Serra et al., 1957; Crusafont, 1960; Lumley, 1971; Guilaine et al.,1982). This complex was formed by the drainage system of Maltorrent, which modeled the Neogene limestone (CollsuspinaFormation) and configures the endokarstic landscape observednowadays. Teixoneres cave is thus a fossilized ancient outlet of theToll cave system. Teixoneres is a U-shaped cave 30 m long and withthree differentiated chambers called X, Y and Z. The cave has twoentrances, the main one providing access to chamber X and thesecond one leading to chamber Z. The latter is smaller and probablyopened more recently than the main one (Fig. 2B).
Archaeological work in the cave was started in the 1950s bya local speleological group. This group made three deep test dril-lings in themain chamber (chamber X) together with a longitudinaltrench from which they recovered some lithic remains and, aboveall, an important Pleistocene faunal assemblage. Later, in 1973another small excavation was carried out, focusing on the paleon-tological record; the cave was then closed until the current exca-vation project started in 2003.
Teixoneres is filled with a 6 m-thick sediment package con-taining at least 15 archaeo-paleontological levels. We worked onthe first part of the sequence (levels I to IV), belonging to the Late
Pleistocene. Levels I and IV probably correspond to warm and wetperiods, during which water and carbonate precipitation sealed thestratigraphy forming two continuous stalagmitic beds. Uranium-series dating has confidently situated the stalagmite of level IVwithin MIS 5c, with an average date of 100.3� 6.1 ka (Tissoux et al.,2006). The data are uncertain for the stalagmite of level I, whichprobably corresponds to MIS 2. Method correction to account forthe uranium contamination of the sample places level I in theLateglacial, between ca 14e16 ka BP (Tissoux et al., 2006) (Fig. 2A).
Extensive work carried out in the outer part of the cave, whichtoday is an open-air site due to the progressive collapse of thecornice, allowed the excavation of levels II, IIb, III and IIIb in thezone where the areas of most intense human activity wereexpected (Rosell et al., 2010).
3. Material and methods
3.1. Small vertebrates
The small-vertebrate fossil remains used for this study consist ofdisarticulated bone fragments collected by water-screening duringthe 2008e2010 excavation campaigns at Teixoneres cave. All thesediment was water-screened using superimposed 5 mm and0.5 mm-mesh screens, and bagged by level. In subsequent years,the fossils were processed, sorted and classified at the IPHES/URV(Tarragona, Spain). This assemblage includes a total of 1401 frag-ments that correspond to a minimum number of 636 small verte-brates, representing at least 36 taxa (Table 1). The fragments wereidentified following the general criteria of systematic paleontology(e.g. Van der Meulen, 1973; Pasquier, 1974; Damms, 1981; Reumer,1984; Sanchíz, 1984; Szyndlar, 1984; Sevilla, 1988; Bailon, 1991,1999; Blain, 2005, 2009; Sanchíz et al., 2002).
Specific identification of this material rests principally on thebest diagnostic elements: humerus, ilium, scapula and sacrum foranurans; skull elements for lizards and trunk vertebrae for snakes;mandible, maxilla and isolated teeth for shrews; mandible andisolated teeth for bats; first lower molars for the Arvicolinaesubfamily; and isolated teeth for Murinae and Glirinae subfamilies.
The fossils were grouped using the minimum number of indi-viduals (MNI) method, with the sample (i.e. from each level)determined by counting a diagnostic element, taking into account,whenever possible, laterality and sex (for anurans).
In order to reconstruct the environment at Teixoneres cave, themethod of habitat weightings (Evans et al., 1981; Andrews, 2006)was used, distributing each small-vertebrate taxon in the habitat(s)where it is possible to find it at present in the Iberian Peninsula.Habitats were divided into five types (according to Cuenca-Bescóset al., 2005, 2009; Blain et al., 2008): open land in which dry andwet meadows are distinguished, woodland and woodland marginareas, rocky areas and surrounding water areas. These types aredetailed as follows (Table 1): open dry: meadows with seasonalchange; open moist: evergreen meadows with dense pastures andsuitable topsoil; woodland: mature forest including woodlandmargins and forest patches, with moderate ground cover; water:along streams, lakes and ponds; and rocky: areas with suitablerocky or stony substratum. The “Ch1&Ch2” and “Ch3” curves (Fig. 4)were obtained from the percentage representation of the MNI byclassifying our taxa according to chorotypes established previouslyby Sans-Fuentes and Ventura (2000) and López-García et al. (2010).These chorotypes are detailed as follows (Table 1): chorotype 1:Includes species with mid-European requirements, with meansummer temperatures lower than 20 �C, mean annual tempera-tures (MAT) between 10 �C and 12 �C and mean annual precipita-tion (MAP) higher than 800 mm; chorotype 2: Includes mid-European species tolerant of Mediterranean conditions, with
J.M. López-García et al. / Quaternary Science Reviews 43 (2012) 33e4434
Author's personal copy
a broader distribution in Catalonia than those of chorotype 1, withMAP higher than 600 mm; chorotype 3: Includes non-strictly andstrictly Mediterranean species, with a broad distribution in Cata-lonia and without very strict requirements.
In order to evaluate the paleoclimatic changes, the MutualClimatic Rangemethod (MCR) (Agustí et al., 2009; Blain et al., 2009)was used for small mammals. On the basis of the distribution of theextant Iberian fauna, the geographical region where all the speciespresent in a stratigraphical level currently live is identified byoverlapping ranges. As the atlas is based on a 10 � 10 km Universal
Transverse Mercator (UTM) network (Palombo and Gisbert, 2005),climatic data were evaluated for each one of these 10 � 10 kmsquares.
Several climatic factors have been estimated: the temperature ofthe coldest month (MTC; January), the temperature of the warmestmonth (MTW; July), the meanwinter precipitation (DJF; December,January and February) and the mean summer precipitation (JJA;June, July, August), using various climatic maps of Spain (FontTullot, 1983, 2000) and data provided by the network of Spanishmeteorological research stations over a period of 30 years. From
Fig. 1. Location of Teixoneres cave and approximate location of the some of the most important sites in Europe (above) and the Iberian Peninsula (below) with Neanderthal remainsor evidence (above) (modified from Delson and Harvati, 2006 and Díez et al., 2010).
J.M. López-García et al. / Quaternary Science Reviews 43 (2012) 33e44 35

Author's personal copy
this variably sized region, the climatic parameters can be estimatedand compared with the weather station at Barcelona Airport(current data from Font-Tullot, 2000).
3.2. Paleobotany
Seventeen samples were recovered during the 2007 excavationcampaign from Chamber X (north section, squares N19/20) ofTeixoneres cave. The samples were recovered from a depth of155e380 cm, recovering a sample every 5 cm for level II and atirregular intervals of 20e40 cm for level III, according to the lith-ological features. The sediments were treated using the protocolproposed by Goeury and Beaulieu (1979), slightly modified by theelimination of acetolysis in accordance with Girard and Renault-Miskovsky (1969), and the protocol developed by Burjachs et al.(2003). The sediments were reduced by HCl; the humic acidswere washed away in NaOH; the sediments were floated in denseThoulet liquor, filtered with a glass fibre filter, and the filter wasdestroyed with 70% HF. All these steps were followed by washing inH2O distilled water and centrifuging. The final residue of eachsample was mounted in preparation for biological microscopy, anddiluted with glycerol.
The charcoal analyses are based on identification of the woodcell structure using a metallographic microscope with reflectedlight (Olympus BX41). Each piece of charcoal was fragmented byhand to observe the three anatomical sections of wood anatomy.
3.3. Dental wear
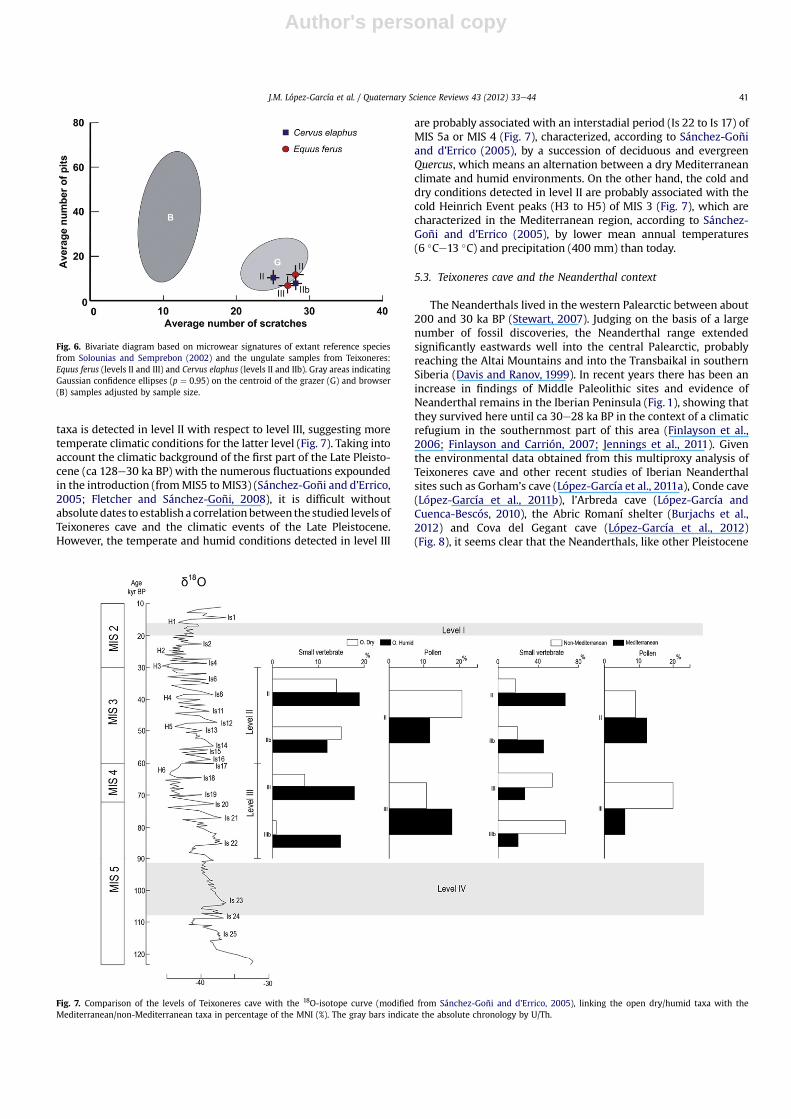
Toothmeso- andmicrowear analysis was used to study ungulateremains from levels II, IIb and III. Tooth wear provides insight intothe diet of the individuals, i.e. into the immediate behavior of thepopulation. The results of the mesowear analysis reflect the diet ofthe last few weeks or months (Fortelius and Solounias, 2000),whereas the microwear analysis yields information on the last fewdays or the last week (Mainland, 1998; Solounias and Semprebon,2002).
Mesowear analysis, first introduced by Fortelius and Solounias(2000), is a method of categorizing the gross dental wear ofungulate molars by evaluating the relief and sharpness of cuspapices in ways that are correlated with the relative amounts ofattritive and abrasive dental wear. Low-abrasion diets, such asthose of browsers, generate an attritional (sharpening) relationshipbetween the upper and lower cusps. In contrast, abrasion-dominated (food-on-tooth) wear, associated with grazing diets,results in more rounded and blunted wear facets and less preciseocclusion. Only adults are found to have consistent signs of meso-wear (Rivals et al., 2007a); consequently, only adults were selectedfor study. Mesowear was scored on the paracone of the M2. Herewe employ the standardized method (mesowear “ruler”) intro-duced by Mihlbachler et al. (2011). The method is based on sevenmodern horse cusps (numbered 0e6), ranging in shape from highand sharp (stage 0) to completely blunt with no relief (stage 6). Theaverage value of the mesowear data from a single sample of fossildentitions is the “mesowear score” (Rivals et al., 2007b). Individualscores were then averaged for each sample.
Microwear analysis was performed using the method describedby Solounias and Semprebon (2002) and Semprebon et al. (2004).High-resolution silicone molds were made for all available upperand lower second molars, which were selected by excluding youngand old individuals. The casts were screened using a stereomicro-scope, and any specimens exhibiting signs of weathering wereexcluded. Microwear features were identified and quantified onhigh-resolution epoxy tooth casts at 35� magnification usinga stereomicroscope. Most microwear features can be categorized aspits and scratches of various sizes and textures. To approximate pitand scratch frequency, they are counted in a standard 0.4� 0.4 mmsquare area on the lingual (inner) band of enamel on the paraconeof the M2 or the protoconid of the m2.
4. Chronological and environmental data
4.1. Small vertebrates
4.1.1. TaphonomyAlthough a complete taphonomic study has not been performed
in Teixoneres cave, some preliminary remarks can be made. Thepattern of skeletal element frequencies indicates a good repre-sentation of small-vertebrate skeleton elements (Table 2). Theseshow little breakage (the 17.84% of the identified material isbroken), and the light-to-moderate digestion seen in microtineteeth, with rounded in the triangles, indicates that the bones wereprobably accumulated by an avian predator of category 1 (sensuAndrews, 1990), such as the barn owl (Tyto alba).
Although we are not able to specify the predator responsible forthe accumulation, there are no indications of alteration to suggestthat the Teixoneres cave assemblage is not representative of the
Fig. 2. A: Stratigraphic section of Teixoneres cave showing the absolute data for level Iand IV. B: Topographic map of the cavity.
J.M. López-García et al. / Quaternary Science Reviews 43 (2012) 33e4436
Author's personal copy
ecosystem in the immediate vicinity of the cave at the time whenthe remains were deposited.
From another standpoint, occasional predation on bats bynocturnal birds of prey might account for the isolated teeth atTeixoneres cave, though in-situ death might also account for these,since the species in question is known to frequent caves.
4.1.2. Small mammalsThe small-mammal remains from Teixoneres cave provided
a large sample of insectivores, bats and rodents from Levels II to IIIb,comprising 899 bone fragments that correspond to 443 individuals(Fig. 3; Table 1). The faunal list is composed of 6 insectivores (Eri-naceus europaeus, Talpa europaea, Crocidura russula, Neomys fodiens,Sorex minutus and Sorex coronatus), 5 chiropters (Rhinolophus fer-rumequinum, Rhinolophus gr. euryale-mehelyi, Myotis gr. myotis-blythii, Pispistrellus sp. and Miniopterus schreibersii) and 13 rodents(Arvicola sapidus, Microtus arvalis, Microtus agrestis, M. arvalis-agrestis, Microtus (Iberomys) cabrerae, Microtus (Terricola) duo-decimcostatus, Microtus (Terricola) gerbei, Pliomys lenki, Chionomysnivalis, Apodemus sylvaticus, Eliomys quercinus, Myoxus glis andHystrix sp.). To narrow the chronology of the site, we used threerodent species, two extinct (P. lenki and Hystrix sp.) and one extant(M. (Iberomys) cabrerae). The combination of these three species inthe levels of Teixoneres cave allows us to establish a chronology
older than 30 ka BP for Level IIb and between 60 and 90 ka BP forLevel III.
From a paleoenvironmental point of view, the small mammalassemblage from Teixoneres cave shows a predominance ofwoodland habitats throughout the sequence (more than 60% in alllevels), represented by the abundance of A. sylvaticus. Moreover,two clear environmental phases have been detected: the first one isfound in Level III, where there is an increase in humid meadows
Table 2Percentage of the skeletal representation (according to Andrews, 1990), taking intoaccount the diagnostic elements to identify the species.
Diagnostic elements % representation
Anurans Humerus 42.67IliumScapulaSacrum
Squamates Skull 52.38Trunk vertebrae
Insectivores Maxilla 52.27MandibuleIsolate teeth
Arvicolinae First lower molars 59.50Murinae Isolate teeth 43.72Gliridae Isolate teeth 49.05
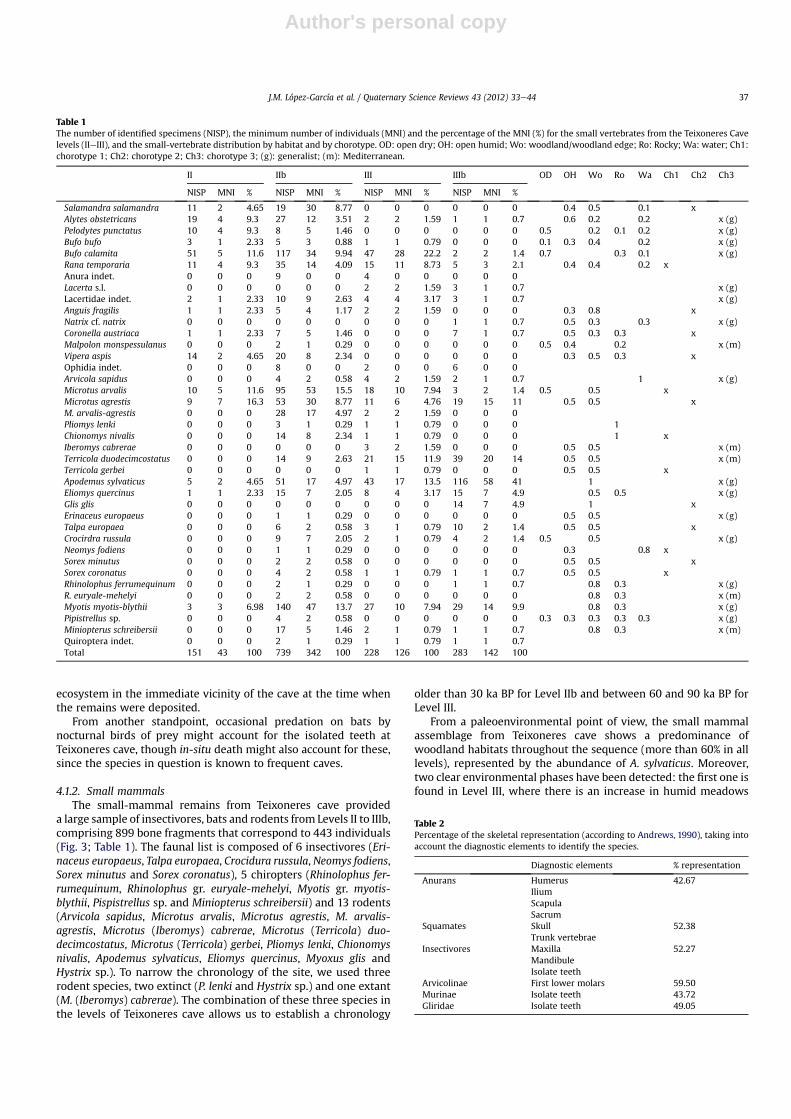
Table 1The number of identified specimens (NISP), the minimum number of individuals (MNI) and the percentage of the MNI (%) for the small vertebrates from the Teixoneres Cavelevels (IIeIII), and the small-vertebrate distribution by habitat and by chorotype. OD: open dry; OH: open humid; Wo: woodland/woodland edge; Ro: Rocky; Wa: water; Ch1:chorotype 1; Ch2: chorotype 2; Ch3: chorotype 3; (g): generalist; (m): Mediterranean.
II IIb III IIIb OD OH Wo Ro Wa Ch1 Ch2 Ch3
NISP MNI % NISP MNI % NISP MNI % NISP MNI %
Salamandra salamandra 11 2 4.65 19 30 8.77 0 0 0 0 0 0 0.4 0.5 0.1 xAlytes obstetricans 19 4 9.3 27 12 3.51 2 2 1.59 1 1 0.7 0.6 0.2 0.2 x (g)Pelodytes punctatus 10 4 9.3 8 5 1.46 0 0 0 0 0 0 0.5 0.2 0.1 0.2 x (g)Bufo bufo 3 1 2.33 5 3 0.88 1 1 0.79 0 0 0 0.1 0.3 0.4 0.2 x (g)Bufo calamita 51 5 11.6 117 34 9.94 47 28 22.2 2 2 1.4 0.7 0.3 0.1 x (g)Rana temporaria 11 4 9.3 35 14 4.09 15 11 8.73 5 3 2.1 0.4 0.4 0.2 xAnura indet. 0 0 0 9 0 0 4 0 0 0 0 0Lacerta s.l. 0 0 0 0 0 0 2 2 1.59 3 1 0.7 x (g)Lacertidae indet. 2 1 2.33 10 9 2.63 4 4 3.17 3 1 0.7 x (g)Anguis fragilis 1 1 2.33 5 4 1.17 2 2 1.59 0 0 0 0.3 0.8 xNatrix cf. natrix 0 0 0 0 0 0 0 0 0 1 1 0.7 0.5 0.3 0.3 x (g)Coronella austriaca 1 1 2.33 7 5 1.46 0 0 0 7 1 0.7 0.5 0.3 0.3 xMalpolon monspessulanus 0 0 0 2 1 0.29 0 0 0 0 0 0 0.5 0.4 0.2 x (m)Vipera aspis 14 2 4.65 20 8 2.34 0 0 0 0 0 0 0.3 0.5 0.3 xOphidia indet. 0 0 0 8 0 0 2 0 0 6 0 0Arvicola sapidus 0 0 0 4 2 0.58 4 2 1.59 2 1 0.7 1 x (g)Microtus arvalis 10 5 11.6 95 53 15.5 18 10 7.94 3 2 1.4 0.5 0.5 xMicrotus agrestis 9 7 16.3 53 30 8.77 11 6 4.76 19 15 11 0.5 0.5 xM. arvalis-agrestis 0 0 0 28 17 4.97 2 2 1.59 0 0 0Pliomys lenki 0 0 0 3 1 0.29 1 1 0.79 0 0 0 1Chionomys nivalis 0 0 0 14 8 2.34 1 1 0.79 0 0 0 1 xIberomys cabrerae 0 0 0 0 0 0 3 2 1.59 0 0 0 0.5 0.5 x (m)Terricola duodecimcostatus 0 0 0 14 9 2.63 21 15 11.9 39 20 14 0.5 0.5 x (m)Terricola gerbei 0 0 0 0 0 0 1 1 0.79 0 0 0 0.5 0.5 xApodemus sylvaticus 5 2 4.65 51 17 4.97 43 17 13.5 116 58 41 1 x (g)Eliomys quercinus 1 1 2.33 15 7 2.05 8 4 3.17 15 7 4.9 0.5 0.5 x (g)Glis glis 0 0 0 0 0 0 0 0 0 14 7 4.9 1 xErinaceus europaeus 0 0 0 1 1 0.29 0 0 0 0 0 0 0.5 0.5 x (g)Talpa europaea 0 0 0 6 2 0.58 3 1 0.79 10 2 1.4 0.5 0.5 xCrocirdra russula 0 0 0 9 7 2.05 2 1 0.79 4 2 1.4 0.5 0.5 x (g)Neomys fodiens 0 0 0 1 1 0.29 0 0 0 0 0 0 0.3 0.8 xSorex minutus 0 0 0 2 2 0.58 0 0 0 0 0 0 0.5 0.5 xSorex coronatus 0 0 0 4 2 0.58 1 1 0.79 1 1 0.7 0.5 0.5 xRhinolophus ferrumequinum 0 0 0 2 1 0.29 0 0 0 1 1 0.7 0.8 0.3 x (g)R. euryale-mehelyi 0 0 0 2 2 0.58 0 0 0 0 0 0 0.8 0.3 x (m)Myotis myotis-blythii 3 3 6.98 140 47 13.7 27 10 7.94 29 14 9.9 0.8 0.3 x (g)Pipistrellus sp. 0 0 0 4 2 0.58 0 0 0 0 0 0 0.3 0.3 0.3 0.3 0.3 x (g)Miniopterus schreibersii 0 0 0 17 5 1.46 2 1 0.79 1 1 0.7 0.8 0.3 x (m)Quiroptera indet. 0 0 0 2 1 0.29 1 1 0.79 1 1 0.7Total 151 43 100 739 342 100 228 126 100 283 142 100
J.M. López-García et al. / Quaternary Science Reviews 43 (2012) 33e44 37

Author's personal copy
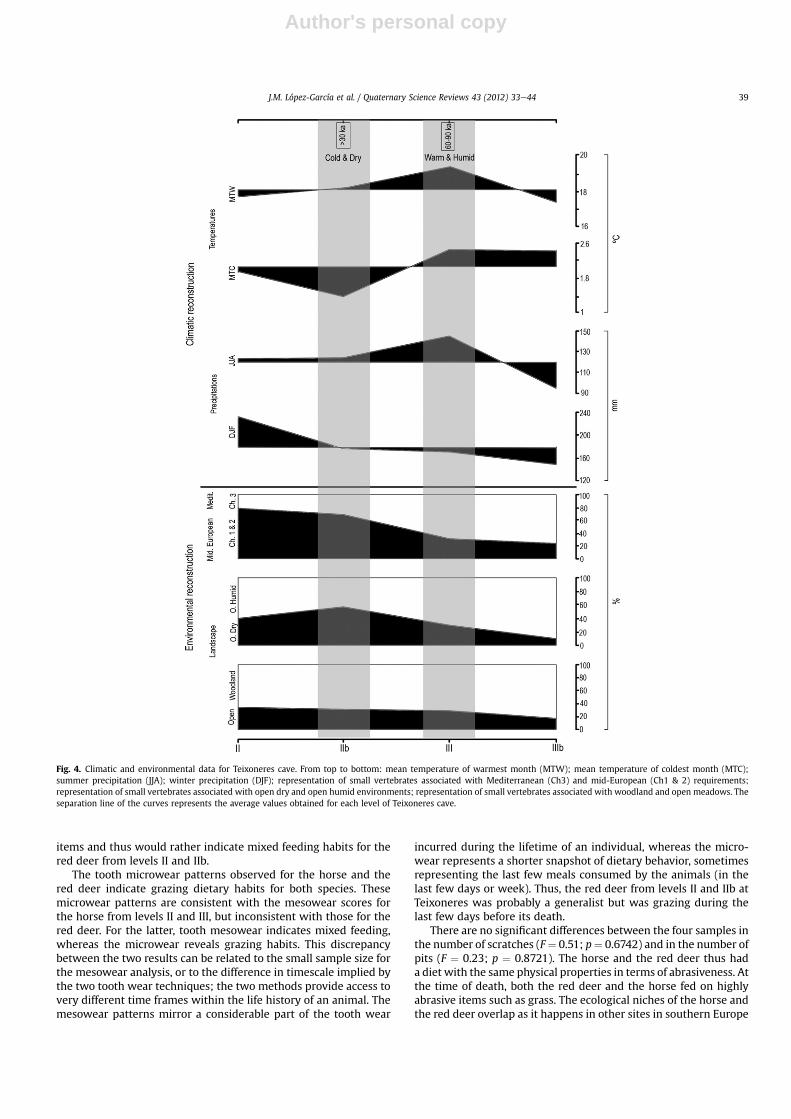
and water streams, represented by the species Microtus agrestris,Microtus (Terricola) duodecimcostatus and A. sapidus; the secondone is found in Level IIb, where there is an increase in dry meadowsand rocky/stony areas, as well as a progressive decrease in waterstreams, represented by an abundance of M. arvalis and C. nivalisand the presence of P. lenki. The data obtained by the MCR methodindicate, in general, that temperatures were lower than today(between �8.3 �C and �5.3 �C) and the precipitation was higher(between þ181 mm and þ322 mm) (current data from Font-Tullot,2000). Furthermore, we observed in Level III high values fortemperatures (coldest and warmest month) probably in responseto summer precipitation (JJA). In Level IIb there is also a largedecrease in winter temperatures together with low values forwinter and summer precipitation (Fig. 4).
4.1.3. Amphibians and squamate reptilesTogether with small mammals, the excavations at Teixoneres
cave also delivered some 502 bones of amphibians and squamatereptiles, which correspond to 193 individuals and 13 taxa (Table 1):one salamander (Salamandra salamandra), five anurans (Alytesobstetricans, Pelodytes punctatus, Bufo bufo, Bufo calamita and Ranatemporaria), three lizards (Lacerta s.l., Lacertidae indet. and Anguisfragilis) and four snakes (Natrix cf. natrix, Coronella austriaca, Mal-polon monspessulanus and Vipera aspis).
Taxa typical of temperate humid areas and/or woodland edgesare predominant (S. salamandra, A. obstetricans, P. punctatus, B. bufo,R. temporaria, A. fragilis, N. natrix, C. austriaca and V. aspis), sug-gesting large extensions of humidmeadows andwoodland patches.In particular, S. salamandra is not present today in areas with pro-longed periods of aridity (Buckley and Alcobendas, 2002). Wood-lands are well developed throughout the sequence with thepresence of S. salamandra, B. bufo, R. temporaria, A. fragilis andV. aspis. The presence of water in the vicinity of the cave is wellattested by the occurrence of anurans and the grass snake N. natrix.Although the remains are less abundant in levels III and IIIb, theamphibians and squamate reptiles suggest temperate-to-coldhumid conditions for all levels. The representation of open-dryenvironments shows a small increase from the base (IIIb and III)to the top (IIb and II) of the sequence, concordant with the resultsfor the small mammals. Noteworthy is that the presence of thethermophilous snake M. monspessulanus in level IIb suggests thatMATs are higher than 10 �C (Blásquez and Pleguezuelos, 2002), andthe presence of S. salamandra suggests that the climate is not verydry in levels II and IIb, especially during summer.
4.2. Paleobotany
In general, the sequence of Teixoneres cave shows a differentenvironments but with woodland and open areas in both levels (IIand III). The woodland areas were principally composed of pinesand oaks (Quercus spp.). The arboreal pollen (AP) values are above50% throughout the sequence.
Among the herbaceous taxa are Poaceae, Chenopodiacea andAsteraceae, which are present throughout this period. On the otherhand, the steppe taxon Artemisia is only present in level II. Inaddition, a semi-open landscape is well attested by the majorpresence of heliophilous taxa such as Cupressaceae, Corylus, Cis-taceae, Erica, Ephedra and herbs in general (Fig. 5).
Judging the results by levels, level III shows indications ofrelative moisture, with a high representation of Corylus and Pter-idophyta spores, whereas level II shows dry conditions, given thehigh representation values for Artemisia.
Charcoal remains at Teixoneres cave are not abundant (35fragments), probably due to questions of preservation; however,evidence of burnt bones, thermal alterations present on lithics, andcharcoal in archaeological levels suggest that the origin of theanthracological material is anthropic, i.e. the charcoal fragmentscome from hearths. The results from Levels II, IIb and III show up to4 taxa, including Buxus sempervirens, Pinus pinea/pinaster type,Pinus sylvestris type and Quercus sp. deciduous (Table 3). Thepresence of P. sylvestris is in accordance with other Middle Paleo-lithic sequences in the NE of the Iberian Peninsula (Allué, 2002).
P. sylvestris type groves were the most widespread forestformation, being well distributed from the coast to the Pyrenees.The occurrence of other taxa such as Buxus or Quercus in charcoalassemblages occurred during periods with milder climates,permitting mesothermophilous taxa to spread out (Burjachs et al.,2012).
4.3. Dental wear
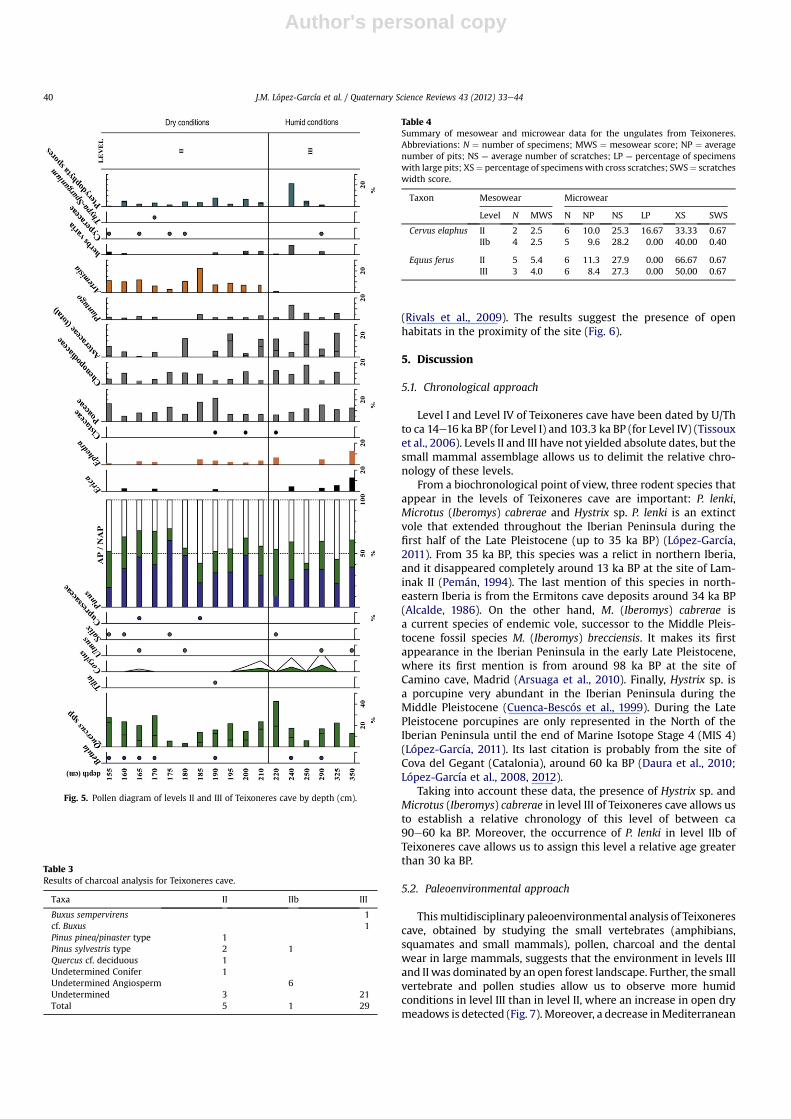
When compared to the extant reference species from Forteliusand Solounias, the tooth mesowear scores for the horse (Equusferus) indicate the presence of highly abrasive items in the diet(Table 4). This suggests grazing dietary habits for this species inlevels II and III. For the red deer (Cervus elaphus), the mesowearscores are lower than those observed for the horse but the samplesize is rather small to allow definite dietary conclusions. Themesowear scores are consistent with a diet based on low-abrasion
Fig. 3. Some small mammal remains recovered from levels II and III of Teixoneres cave.A: first lower (m1) left molar Pliomys lenki (occlusal and labial views; B: m1 rightMicrotus (Terricola) duodecimcostatus (occlusal view); C: m1 left Arvicola sapidus(occlusal view); D: m1 left Microtus (Iberomys) cabrerae (occlusal view); E: m1 rightChionomys nivalis (occlusal view); F: left mandible Neomys fodiens (posterior andlingual views). Scale 1 mm.
J.M. López-García et al. / Quaternary Science Reviews 43 (2012) 33e4438
Author's personal copy
items and thus would rather indicate mixed feeding habits for thered deer from levels II and IIb.
The tooth microwear patterns observed for the horse and thered deer indicate grazing dietary habits for both species. Thesemicrowear patterns are consistent with the mesowear scores forthe horse from levels II and III, but inconsistent with those for thered deer. For the latter, tooth mesowear indicates mixed feeding,whereas the microwear reveals grazing habits. This discrepancybetween the two results can be related to the small sample size forthe mesowear analysis, or to the difference in timescale implied bythe two tooth wear techniques; the two methods provide access tovery different time frames within the life history of an animal. Themesowear patterns mirror a considerable part of the tooth wear
incurred during the lifetime of an individual, whereas the micro-wear represents a shorter snapshot of dietary behavior, sometimesrepresenting the last few meals consumed by the animals (in thelast few days or week). Thus, the red deer from levels II and IIb atTeixoneres was probably a generalist but was grazing during thelast few days before its death.
There are no significant differences between the four samples inthe number of scratches (F¼ 0.51; p¼ 0.6742) and in the number ofpits (F ¼ 0.23; p ¼ 0.8721). The horse and the red deer thus hada diet with the same physical properties in terms of abrasiveness. Atthe time of death, both the red deer and the horse fed on highlyabrasive items such as grass. The ecological niches of the horse andthe red deer overlap as it happens in other sites in southern Europe
Fig. 4. Climatic and environmental data for Teixoneres cave. From top to bottom: mean temperature of warmest month (MTW); mean temperature of coldest month (MTC);summer precipitation (JJA); winter precipitation (DJF); representation of small vertebrates associated with Mediterranean (Ch3) and mid-European (Ch1 & 2) requirements;representation of small vertebrates associated with open dry and open humid environments; representation of small vertebrates associated with woodland and open meadows. Theseparation line of the curves represents the average values obtained for each level of Teixoneres cave.
J.M. López-García et al. / Quaternary Science Reviews 43 (2012) 33e44 39
Author's personal copy
(Rivals et al., 2009). The results suggest the presence of openhabitats in the proximity of the site (Fig. 6).
5. Discussion
5.1. Chronological approach
Level I and Level IV of Teixoneres cave have been dated by U/Thto ca 14e16 ka BP (for Level I) and 103.3 ka BP (for Level IV) (Tissouxet al., 2006). Levels II and III have not yielded absolute dates, but thesmall mammal assemblage allows us to delimit the relative chro-nology of these levels.
From a biochronological point of view, three rodent species thatappear in the levels of Teixoneres cave are important: P. lenki,Microtus (Iberomys) cabrerae and Hystrix sp. P. lenki is an extinctvole that extended throughout the Iberian Peninsula during thefirst half of the Late Pleistocene (up to 35 ka BP) (López-García,2011). From 35 ka BP, this species was a relict in northern Iberia,and it disappeared completely around 13 ka BP at the site of Lam-inak II (Pemán, 1994). The last mention of this species in north-eastern Iberia is from the Ermitons cave deposits around 34 ka BP(Alcalde, 1986). On the other hand, M. (Iberomys) cabrerae isa current species of endemic vole, successor to the Middle Pleis-tocene fossil species M. (Iberomys) brecciensis. It makes its firstappearance in the Iberian Peninsula in the early Late Pleistocene,where its first mention is from around 98 ka BP at the site ofCamino cave, Madrid (Arsuaga et al., 2010). Finally, Hystrix sp. isa porcupine very abundant in the Iberian Peninsula during theMiddle Pleistocene (Cuenca-Bescós et al., 1999). During the LatePleistocene porcupines are only represented in the North of theIberian Peninsula until the end of Marine Isotope Stage 4 (MIS 4)(López-García, 2011). Its last citation is probably from the site ofCova del Gegant (Catalonia), around 60 ka BP (Daura et al., 2010;López-García et al., 2008, 2012).
Taking into account these data, the presence of Hystrix sp. andMicrotus (Iberomys) cabrerae in level III of Teixoneres cave allows usto establish a relative chronology of this level of between ca90e60 ka BP. Moreover, the occurrence of P. lenki in level IIb ofTeixoneres cave allows us to assign this level a relative age greaterthan 30 ka BP.
5.2. Paleoenvironmental approach
Thismultidisciplinary paleoenvironmental analysis of Teixonerescave, obtained by studying the small vertebrates (amphibians,squamates and small mammals), pollen, charcoal and the dentalwear in large mammals, suggests that the environment in levels IIIand II was dominated by an open forest landscape. Further, the smallvertebrate and pollen studies allow us to observe more humidconditions in level III than in level II, where an increase in open drymeadows is detected (Fig. 7). Moreover, a decrease inMediterranean
Fig. 5. Pollen diagram of levels II and III of Teixoneres cave by depth (cm).
Table 3Results of charcoal analysis for Teixoneres cave.
Taxa II IIb III
Buxus sempervirens 1cf. Buxus 1Pinus pinea/pinaster type 1Pinus sylvestris type 2 1Quercus cf. deciduous 1Undetermined Conifer 1Undetermined Angiosperm 6Undetermined 3 21Total 5 1 29
Table 4Summary of mesowear and microwear data for the ungulates from Teixoneres.Abbreviations: N ¼ number of specimens; MWS ¼ mesowear score; NP ¼ averagenumber of pits; NS ¼ average number of scratches; LP ¼ percentage of specimenswith large pits; XS¼ percentage of specimens with cross scratches; SWS¼ scratcheswidth score.
Taxon Mesowear Microwear
Level N MWS N NP NS LP XS SWS
Cervus elaphus II 2 2.5 6 10.0 25.3 16.67 33.33 0.67IIb 4 2.5 5 9.6 28.2 0.00 40.00 0.40
Equus ferus II 5 5.4 6 11.3 27.9 0.00 66.67 0.67III 3 4.0 6 8.4 27.3 0.00 50.00 0.67
J.M. López-García et al. / Quaternary Science Reviews 43 (2012) 33e4440
Author's personal copy
taxa is detected in level II with respect to level III, suggesting moretemperate climatic conditions for the latter level (Fig. 7). Taking intoaccount the climatic background of the first part of the Late Pleisto-cene (ca 128e30 ka BP) with the numerous fluctuations expoundedin the introduction (fromMIS5 toMIS3) (Sánchez-Goñi and d’Errico,2005; Fletcher and Sánchez-Goñi, 2008), it is difficult withoutabsolute dates to establish a correlationbetween the studied levels ofTeixoneres cave and the climatic events of the Late Pleistocene.However, the temperate and humid conditions detected in level III
are probably associated with an interstadial period (Is 22 to Is 17) ofMIS 5a or MIS 4 (Fig. 7), characterized, according to Sánchez-Goñiand d’Errico (2005), by a succession of deciduous and evergreenQuercus, which means an alternation between a dry Mediterraneanclimate and humid environments. On the other hand, the cold anddry conditions detected in level II are probably associated with thecold Heinrich Event peaks (H3 to H5) of MIS 3 (Fig. 7), which arecharacterized in the Mediterranean region, according to Sánchez-Goñi and d’Errico (2005), by lower mean annual temperatures(6 �Ce13 �C) and precipitation (400 mm) than today.
5.3. Teixoneres cave and the Neanderthal context
The Neanderthals lived in the western Palearctic between about200 and 30 ka BP (Stewart, 2007). Judging on the basis of a largenumber of fossil discoveries, the Neanderthal range extendedsignificantly eastwards well into the central Palearctic, probablyreaching the Altai Mountains and into the Transbaikal in southernSiberia (Davis and Ranov, 1999). In recent years there has been anincrease in findings of Middle Paleolithic sites and evidence ofNeanderthal remains in the Iberian Peninsula (Fig. 1), showing thatthey survived here until ca 30e28 ka BP in the context of a climaticrefugium in the southernmost part of this area (Finlayson et al.,2006; Finlayson and Carrión, 2007; Jennings et al., 2011). Giventhe environmental data obtained from this multiproxy analysis ofTeixoneres cave and other recent studies of Iberian Neanderthalsites such as Gorham’s cave (López-García et al., 2011a), Conde cave(López-García et al., 2011b), l’Arbreda cave (López-García andCuenca-Bescós, 2010), the Abric Romaní shelter (Burjachs et al.,2012) and Cova del Gegant cave (López-García et al., 2012)(Fig. 8), it seems clear that the Neanderthals, like other Pleistocene
Fig. 6. Bivariate diagram based on microwear signatures of extant reference speciesfrom Solounias and Semprebon (2002) and the ungulate samples from Teixoneres:Equus ferus (levels II and III) and Cervus elaphus (levels II and IIb). Gray areas indicatingGaussian confidence ellipses (p ¼ 0.95) on the centroid of the grazer (G) and browser(B) samples adjusted by sample size.
Fig. 7. Comparison of the levels of Teixoneres cave with the 18O-isotope curve (modified from Sánchez-Goñi and d’Errico, 2005), linking the open dry/humid taxa with theMediterranean/non-Mediterranean taxa in percentage of the MNI (%). The gray bars indicate the absolute chronology by U/Th.
J.M. López-García et al. / Quaternary Science Reviews 43 (2012) 33e44 41

Author's personal copy
hominids in Europe (Van Andel and Tzedakis, 1996; Stewart, 2007;Kahlke et al., 2011; Messager et al., 2011, among others), were welladapted to the different climatic and environmental conditions ofthe Late Pleistocene (MIS5 to MIS3) of Iberia. Moreover, as occurredduring the Early toMiddle Pleistocenewith other representatives ofthe genus Homo in southwestern Europe (Kahlke et al., 2011; Leroyet al., 2011), the data obtained for Teixoneres cave and the othersites mentioned above (Fig. 8) imply that the dispersal and survivalof the Neanderthals in the Iberian Peninsula seem to be closelyassociated with the existence of woodland formations, as previ-ously suggested by Finlayson and Carrión (2007). Although thereare differences in percentage of forest formations from the south tothe north of Iberia (Fig. 8), being the southern Iberia (Gorham’scave) more much forested (85%) than the northern parts of Iberia(El Conde, A. Romaní, l’Arbreda or Gegant) where the woodlandranges between 55 and 70%. These data are in accordance with theattribution of the southernmost Iberian Peninsula as a climaticrefugium, where Neanderthals survived until ca 28 ka BP (Finlaysonet al., 2006; Finlayson and Carrión, 2007; Jennings et al., 2011,among others). Furthermore, the southwestern fringes of thePalearctic were probably richer in forested habitats which requiredsome rainfall, and the Neanderthals probably preferred this kind ofhabitat to open landscape (Stewart, 2007).
6. Conclusions
This multiproxy analysis of Teixoneres cave based on the smallvertebrates (amphibians, squamates and small mammals), pollen,charcoal and the dental wear in large mammals allows us toestablish the following conclusions:
1) On the basis of the rodent assemblages, the presence of P. lenki,Microtus (Iberomys) cabrerae and Hystrix sp. in Teixoneres caveallows us to reduce the chronology for Levels II and III tobetween ca 30 and 90 ka BP.
2) The environmental and climatic results show that the twolevels of Teixoneres cave (III and II) are associated with an openforest landscape, with temperate and humid conditions forLevel III and cold and dry conditions for Level II. This suggeststhat the Neanderthals who lived around Teixoneres cave werewell adapted to the territory they inhabited.
3) Finally, the study of Teixoneres cave shows that, as in othersites in the Iberian Peninsula, the Middle Paleolithic occupa-tions are closely related to the presence of woodland forma-tions, irrespective of the climatic conditions.
Acknowledgments
Authors are grateful to Prof. José Carrión and two anonymousreviewers for valuable comments on the manuscript. This paper ispart of the Spanish Ministry of Science and Innovation projectsCGL2009-07896, HAR2008-01984 and HAR2010-19957 and theGeneralitat de Catalunya grants SGR2009-324 and SGR2009-188.J.M.L-G. has been supported by a postdoctoral grant from theJuan de la Cierva Subprogram (JCI-2009-04026), with the financialsponsorship of the Spanish Ministry of Science and Innovation. A.B.and R.B. are the beneficiaries of a predoctoral research fellowship(FI) from the Generalitat de Catalunya, financed by the EuropeanSocial Fund, and A.R.H. is the beneficiary of a predoctoral researchfellowship (FPI) from the Ministry of Science and Innovation of theSpanish Government (CGL2009-12703-C03-02).
References
Agustí, J., Blain, H.-A., Cuenca-Bescós, G., Bailon, S., 2009. Climate forcing of firsthominid dispersal in Western Europe. Journal of Human Evolution 57, 815e821.
Alcalde, G., 1986. frLes faunes de rongeurs du Pléistocène supérieur et de l’Holocènede Catalogne (Espagne) et leurs significations paléoécologiques et paléoclima-tiques. Diplôme E.P.H.E, Paris.
Allué, E., 2002. Dinámica de la vegetación y explotación del combustible leñosodurante el Pleistoceno Superior y el Holoceno del Noreste de la PenínsulaIbérica a partir del análisis antracológico. PhD Thesis, Universitat Rovira i Virgili,Tarragona.
Andrews, P., 1990. Owls, Caves and Fossils. Predation, Preservation and Accumula-tion of Small Mammal Bones in Caves, With An Analysis of the Pleistocene CaveFaunas from Westbury-sub-Mendip. The University of Chicago Press, Somerset,UK, London.
Andrews, P., 2006. Taphonomic effects of faunal impoverishment and faunal mix-ing. Palaeogeography Palaeoclimatology Palaeoecology 241, 572e589.
Arsuaga, J.L., Baquedano, E., Pérez-González, A., Sala, M.T.N., García, N., Álvarez-Lao, D., Laplana, C., Huguet, R., Sevilla, P., Blain, H.-A., Quam, R., Ruiz-Zapata, M.B., Sala, P., García, M.J.G., Uzquiano, P., Pantoja, A., 2010. El yaciminetoarqueo-paleontológico del Pleistoceno Superior de la Cueva del Camino en elCalvero de la Higuera (Pinilla del Valle, Madrid). Zona Arqueológica 13,422e442.
Bailon, S., 1991. Amphibiens et reptiles du Pliocène et du Quaternaire de France etd’Espagne: mise en place et évolution des faunes. Ph.D. Thesis, University ofParis VII, 499 pp., 89 pls.
Bailon, S.,1999.DifférenciationostéologiquedesAnoures (Amphibia,Anura)deFrance.In:Desse, J., Desse-Berset, N. (Eds.), Fichesd’ostéologie animale pour l’archéologie,Série C: varia. Centre de Recherches Archéologiques-CNRS, Valbonne.
Bard, E., Rostek, F., Turon, J.L., Gendreau, S., 2000. Hydrological impact of Heinrichevents in the subtropical Northeast Atlantic. Science 289, 1321e1324.
Blain, H.-A., Bailon, S., Cuenca-Bescós, G., 2008. The EarlyeMiddle Pleistocenepalaeoenvironmental change based on the squamate reptile and amphibianproxy at the Gran Dolina site, Atapuerca, Spain. Palaeogeography Palae-oclimatology Palaeoecology 261, 177e192.
Blain, H.-A., Bailon, S., Cuenca-Bescós, G., Arsuaga, J.L., Bermúdez de Castro, J.M.,Carbonell, E., 2009. Long-term climate record inferred from early-middlePleistocene amphibian and squamate reptile assemblages at the Gran Dolinacave, Atapuerca, Spain. Journal of Human Evolution 56, 55e65.
Blain, H.-A., 2005. Contribution de la paléoherpétofaune (Amphibia & Squamata) àla connaissance de l’évolution du climat et du paysage du Pliocène supérieur auPléistocène moyen d’Espagne. Muséum national d’Histoire naturelle, Institut dePaléontologie humaine, Paris, France, 402 pp.
Blain, H.-A., 2009. Contribution de la paléoherpétofaune (Amphibia & Squamata) àla connaissance de l’évolution du climat et du paysage du Pliocène supérieur auPléistocène moyen d’Espagne. Treballs del Museu de Geologia de Barcelona 16,39e170 pp.
Blásquez, M.C., Pleguezuelos, J.M., 2002. Malpolon monspessulanus. In:Pleguezuelos, J.M., Márquez, M., Lizana, M. (Eds.), Atlas y libro rojo de losAnfibios y Reptiles de España, Dirección General de Conservación de la Natu-raleza-Asociación Herpetologica Española, Madrid, pp. 284e286.
Buckley, D., Alcobendas, M., 2002. Salamandra salamandra. In: Pleguezuelos, J.M.,Márquez, M., Lizana, M. (Eds.), Atlas y libro rojo de los Anfibios y Reptiles deEspaña, Dirección General de Conservación de la Naturaleza-Asociación Her-petologica Española, Madrid, pp. 55e57.
Burjachs, F., López-Sáez, J.A., Iriarte, M.J., 2003. Metodología arqueopalinológica. In:Buxó, R., Piqué, R. (Eds.), La recogida de muestras en arqueobotánica: objetivosy propuestas metodológicas. Museu d’Arqueologia de Catalunya, Barcelona,pp. 11e18.
Burjachs, F., López-García, J.M., Allué, E., Blain, H.-A., Rivals, F., Bennàsar, M.,Expósito, I., 2012. Palaeoecology of Neanderthals during DansgaardeOeschgercycles in northeastern Iberia (Abric Romaní): from regional to global scale.Quaternary International 247, 26e37.
Fig. 8. Environmental comparison between Teixoneres cave, El Conde (data fromLópez-García et al., 2011b), l’Arbreda (data from López-García and Cuenca-Bescós,2010), Romaní (data from Burjachs et al., 2012), Gorham’s (data from López-Garcíaet al., 2011a) and Gegant (data from López-García et al., 2012).
J.M. López-García et al. / Quaternary Science Reviews 43 (2012) 33e4442

Author's personal copy
Cacho, I., Grimalt, O.J., Pelejero, C., Canals, M., Sierro, F.J., Flores, J.A., Shackleton, N.,1999. Dansgaard-Oeschger and Heinrich event imprints in Alboran Sea paleo-temperatures. Paleocenography 14, 698e705.
Crusafont, M., 1960. Le Quaternaire espagnol et sa faune de Mammifères. Essai desynthèse. Anthropos 1, 55e64.
Cuenca-Bescós, G., Canudo, J.I., Laplana, C., 1999. Análisis bioestratigráfico del Pleisto-ceno medio del yacimiento de Galería (Sierra de Atapuerca, Burgos). In:Carbonell, E., Rosas, A., Díez, J.C. (Eds.), Atapuerca: Ocupaciones humanas y paleo-ecología del yacimiento de Galería. Junta de Castilla y León, Burgos, pp. 189e210.
Cuenca-Bescós, G., Rofes, J., García Pimienta, J.C., 2005. Environmental changeacross the early-middle Pleistocene transition: small mammalian evidencefrom the Trinchera Dolina cave, Atapuerca, Spain. In: Head, M.J., Gibbard, P.L.(Eds.), Early-Middle Pleistocene Transitions: The LandeOcean Evidence.Geological Society Special Issue, London, pp. 277e388.
Damms, R., 1981. The dental pattern of the dormice Dryomys, Myomimus, Micro-dyromys and Peridryomys. Micropaleontological Bulletins 3, 1e115.
Dansgaard, W., Johnsen, S.J., Clausen, H.B., Dahl-Jensen, D., Gundestrup, N.S.,Hammer, C.U., Hvidberg, C.S., Steffensen, J.P., Sveinbjörnsdottir, A.E., Jouzel, J.,Bond, G., 1993. Evidence for general instability of past climate from a 250-kyrice-core record. Nature 364, 218e220.
Daura, J., Sanz, M., Pike, A.W.G., Subirà, M.E., Fornós, J.J., Fullola, J.M., Julià, R.,Zilhao, J., 2010. Stratigraphic context and direct dating of the Neandertalmandible from Cova del Gegant (Sitges, Barcelona). Journal of Human Evolution59, 109e122.
Davis, R.S., Ranov, V.A., 1999. Recent work on the Paleolithic of central Asia.Evolutionary Anthropology 8, 186e193.
Delson, E., Harvati, K., 2006. Return of the last Neanderthal. Nature 443, 762e763.Díez, J.C., Pérez, S., Martínez, J., 2010. Restos de neandertales y hiénidos en la
Península Ibérica. Zona Arqueológica 13, 230e243.Evans, E.M.N., Van Couvering, J.A.H., Andrews, P., 1981. Palaeoecology of Miocene
sites in western Kenya. Journal of Human Evolution 10, 99e116.Finlayson, C., Carrión, J.S., 2007. Rapid ecological turnover and its impact on
Neanderthal and other human populations. Trends in Ecology & Evolution 22(4), 213e222.
Finlayson, C., Giles-Pacheco, F., Rodriguez-Vidal, J., Fa, D.A., Gutiérrez-López, J.M.,Pérez, A.S., Finlayson, G., Allué, E., Baena-Preysler, J., Cáceres, I., Carrión, J.S.,Fernández-Jalvo, Y., Gleed-Owen, C.P., Jiménez-Espejo, F.J., López, P., López-Sáez, J.A., Riquelme-Cantal, J.A., Sánchez-Marco, A., Giles-Guzmán, F., Brown, K.,Fuentes, N., Valarino, C.A., Villalpando, A., Stringer, C.B., Martínez-Ruiz, F.,Sakamoto, T., 2006. Late survival of Neanderthals at the southernmost extremeof Europe. Nature 443, 850e853.
Fletcher, W., Sánchez-Goñi, M.A., 2008. Orbital and sub-orbital scale climateimpacts on vegetation of the western Mediterranean basin over the last 48,000years. Quaternary Research 70, 451e464.
Font Tullot, I., 1983. Atlas Climático de España. Ministerio de Transportes, Turismo yComunicaciones, Instituto Nacional de Meteorología, Madrid.
Font-Tullot, I., 2000. Climatología de España y Portugal. Ediciones Universidad deSalamanca, Salamanca, 422 pp.
Fortelius, M., Solounias, N., 2000. Functional characterization of ungulate molarsusing the abrasion-attrition wear gradient: a new method for reconstructingpaleodiets. American Museum Novitates 3301, 1e36.
Girard, M., Renault-Miskovsky, J., 1969. Nouvelles techniques de préparation enPalynologie appliqués à trois sédiments du Quaternaire final de l’Abri Cornille(Istres-Bouches du Rhône). Bulletin de l’Association Française pour l’Étude duQuaternaire 4, 275e284.
Goeury, Cl, Beaulieu, J.-L. de, 1979. À propos de la concentration du pollen à l’aide dela liqueur de Thoulet dans les sédiments minéraux. Pollen et Spores XXI (1e2),239e251.
Guilaine, J., Barbaza, M., Geddes, D., Vernet, J.-L., 1982. Prehistoric human adapta-tions in Catalonia (Spain). Journal of Field Archaeology 9, 407e416.
Heinrich, H., 1988. Origin and consequences of cyclic ice rafting in the northeastAtlantic ocean during the past 130.000 years. Quaternary Research 29,142e152.
Jennings, R., Finlayson, C., Fa, D., Finlayson, G., 2011. Southern Iberia as a refuge forthe last Neanderthal populations. Journal of Biogeography 38, 1873e1885.
Johnsen, S.J., Clausen, H.B., Dansgaard, W., Fuhrer, K., Gundestrup, N., Hammer, C.U.,Iversen, P., Jouzel, J., Stauffer, B., Steffensen, J.P., 1992. Irregular glacial inter-stadials recorded in a new Greenland ice core. Nature 359, 311e313.
Kahlke, R.-D., García, N., Kostopoulos, D.S., Lacombat, F., Lister, A.M., Mazza, P.P.A.,Spassov, N., Titov, V.V., 2011. Western Palaearctic palaeoenvironmental condi-tions during the Early and early Middle Pleistocene inferred from largemammal communities, and implications for hominin dispersal in Europe.Quaternary Science Reviews 30, 1368e1395.
Leroy, S.A.G., Arpe, K., Mikolajewicz, U., 2011. Vegetation context and climatic limitsof the Early Pleistocene hominin dispersal in Europe. Quaternary ScienceReviews 30, 1448e1463.
López-García, J.M., Cuenca-Bescós, G., 2010. Évolution climatique durant le Pléis-tocène supérieur en Catalogne (Nord-Est de l’Espagne) d’après l’étude desmicromammifères. Quaternaire 21, 249e257.
López-García, J.M., Blain, H.-A., Cuenca-Bescós, G., Arsuaga, J.L., 2008. Chrono-logical, environmental and climatic precisions on the Neanderthal site of theCova del Gegant (Sitges, Barcelona, Spain). Journal of Human Evolution 55,1151e1155.
López-García, J.M., Blain, H.-A., Allué, E., Bañuls, S., Bargalló, A., Martín, P.,Morales, J.I., Pedro, M., Rodriguez, A., Solé, A., Oms, F.X., 2010. First fossil
evidence of an “interglacial refugium” in the Pyrenean region. Natur-wissenschaften 97, 753e761.
López-García, J.M., Cuenca-Bescós, G., Finlayson, C., Brown, C., Giles-Pacheco, F.,2011a. Palaeoenvironmental and palaeoclimatic proxies of the Gorham’s cavesmall mammal sequence, Gibraltar, southern Iberia. Quaternary International243, 137e142.
López-García, J.M., Cuenca-Bescós, G., Blain, H.-A., Álvarez-Lao, D., Uzquiano, P.,Adán, G., Arbizu, M., Arsuaga, J.L., 2011b. Palaeoenvironment and palaeoclimateof the Mousterian-Aurignacian transition in northern Iberia: The small-vertebrate assemblage from Cueva del Conde (Santo Adriano, Asturias). Jour-nal of Human Evolution 61, 108e116.
López-García, J.M., Blain, H.-A., Sanz, M., Daura, J., 2012. A coastal reservoir ofterrestrial resources for Neanderthal populations in north-eastern Iberia:palaeoenvironmental data inferred from the small-vertebrate assemblage ofCova del Gegant, Sitges, Barcelona. Journal of Quaternary Science 27,105e113.
López-García, 2011. Los micromamíferos del Pleistoceno superior de la PenínsulaIbérica. Evolución de la diversidad taxonómica y cambios paleoambientales ypaleoclimáticos. Editorial Académica Española, Saarbrücken, 368 pp.
Lumley, H. de, 1971. Le Paléolithique inférieur et moyen du Midi méditerranéendans son cadre géologique, II, Bas-Languedoc, Roussillon, Catalogne. V supplé-ment à Gallia Préhistoire. Centre national de la recherche scientifique 2 tomes,443e445.
Mainland, I.L., 1998. The lamb’s last supper: the role of dental microwear analysis inreconstructing livestock diet in the past. In: Charles, M., Halstead, P., Jones, G.(Eds.), Fodder: Archaeological, Historical and Ethnographic Studies. Environ-mental Archaeology, vol. 1, pp. 55e62.
Messager, E., Lebreton, V., Marquet, L., Russo-Ermoli, E., Orain, R., Renault-Miskovsky, J., Lordkipanidze, D., Despriée, J., Peretto, C., Arzarello, M., 2011.Palaeonvironments of early hominins in temperate and Mediterranean Eurasia:new paleobotanical data from Palaeolithic key-sites and synchronous naturalsequences. Quaternary Science Reviews 30, 1439e1447.
Mihlbachler, M.C., Rivals, F., Solounias, N., Semprebon, G.M., 2011. Dietary changeand evolution of horses in North America. Science 331, 1178e1181.
Naughton, F., Sánchez-Goñi, M.F., Kageyama, M., Bard, E., Duprat, J., Cortijo, E.,Desprat, S., Malaizé, B., Joly, C., Rostek, F., Turon, J.-L., 2009. Wet to dry climatictrend in north-western Iberia within Heinrich events. Earth and PlanetaryScience Letters 284, 329e342.
Palombo, J.L., Gisbert, J., 2005. Atlas de los Mamíferos Terrestres de España.Dirección General para la Biodiversidad, Madrid, 564 pp.
Pasquier, L., 1974. Dynamique évolutive d’un sous-genre de Muridae, Apodemus(Sylvaemus). Étude biométrique des caractères dentaires de populations fos-siles et actuelles d’Europe occidentale. Ph.D. Thesis. Université de Montpellier,France.
Pemán, E., 1994. Los Micromamíferos de Laminak II (Berriatua, Bizkaia). Koibe 21,225e233.
Reumer, J.W.F., 1984. Ruscinian and early Pleistocene Soricidae (Insectivora, Mam-malia) from Tegelen (The Netherlands) and Hungary. Scripta Geologica 73,1e173.
Rivals, F., Mihlbachler, M.C., Solounias, N., 2007a. Effect of ontogenetic-age distri-bution in fossil samples on the interpretation of ungulate paleodiets using themesowear method. Journal of Vertebrate Paleontology 27, 763e767.
Rivals, F., Solounias, N., Mihlbachler, M.C., 2007b. Evidence for geographic vari-ation in the diets of late Pleistocene and early Holocene Bison in NorthAmerica, and differences from the diets of recent Bison. Quaternary Research68, 338e346.
Rivals, F., Schulz, E., Kaiser, T.M., 2009. Late andmiddle Pleistocene ungulates dietarydiversity in Western Europe indicate variations of Neanderthal paleoenviron-ments through time and space. Quaternary Science Reviews 28, 3388e3400.
Rosell, J., Blasco, R., Rivals, F., Chacón, M.G., Menéndez, L., Morales, J.I., Rodríguez-Hidalgo, A., Cebrià, A., Carbonell, E., Serrat, D., 2010. A stop along the way: therole of Neanderthal groups at level III of Teixoneres cave (Moià, Barcelona,Spain). Quaternaire 21 (2), 139e154.
Sánchez-Goñi, M.F., Turon, J.-L., Eynaud, F., Gendreau, S., 2000. European climaticresponse to millennial-scale changes in the atmosphere-ocean system duringthe Last Glacial Period. Quaternary Research 54, 394e403.
Sánchez-Goñi, M.F., d’Errico, F., 2005. La historia de la vegetación y el clima delúltimo ciclo climático (OIS5-OIS1, 140.000e10.000 años BP) en la PenínsulaIbérica y su posible impacto sobre los grupos paleolíticos. Museo y Centro deInvestigación de Altamira. Monografías 20, 115e129.
Sanchíz, B., Tejedo, M., Sánchez-Herráiz, M.J., 2002. Osteological differentiationamong Iberian Pelodytes (Anura, Pelodytidae). Graellsia 58, 35e68.
Sanchíz, F.B., 1984. Análisis filogenético de la tribu Alytini (Anura, Discoglossidae)mediante el estudio de su morfoestructura ósea. In: Hemmer, H., Alcover, J.A.(Eds.), Historia Biológica del Ferreret (Baleaphrine muletensis). Ed. Moll, Palmade Mallorca, pp. 61e108.
Sans-Fuentes, M.A., Ventura, J., 2000. Distribution patterns of the small mammals(Insectivora, Rodentia) in a transitional zone between the Eurosiberian and theMediterranean regions. Journal of Biogeography 27, 755e764.
Semprebon, G.M., Godfrey, L.R., Solounias, N., Sutherland, M.R., Jungers, W.L., 2004.Can low-magnification stereomicroscopy reveal diet? Journal of HumanEvolution 47, 115e144.
Serra, J.D.C., Villalta, J.F., Thomas, J., Fuste, M., 1957. Livret Guide des excursions B2-B3. Alentours de Barcelona et Moià. V Congrès International del INQUA, Madrid-Barcelona 3, 5e25 pp.
J.M. López-García et al. / Quaternary Science Reviews 43 (2012) 33e44 43

Author's personal copy
Sevilla, P., 1988. Estudio paleontológico de los Quirópteros del Cuaternario Español.Paleontología y Evolución 22, 113e233.
Shackleton, N.J., Sánchez-Goñi, M.F., Pailler, D., Lancelot, Y., 2003. Marine IsotopeSubstage 5e and the Eemian Interglacial. Global and Planetary Change 36,151e155.
Solounias, N., Semprebon, G., 2002. Advances in the reconstruction of ungulateecomorphology with application to early fossil equids. American MuseumNovitates 3366, 1e49.
Stewart, J.R., 2007. Neanderthal extinctions as part of the faunal change in Europeduring Oxygen Isotope Stage 3. Acta Zoologica Cracoviensia 50A (1e2), 93e124.
Szyndlar, Z., 1984. Fossil snakes from Poland. Acta Zoologica Cracoviensia 28, 1e156.Tissoux,H., Falguéres, C., Bahain, J.-J., Rosell, J., Cebrià, A., Carbonell, R., Serrat, D., 2006.
Datation par les séries de l’Uranium des occupations moustériennes de la grottede Teixoneres (Moià, Province de Barcelone, Espagne). Quaternaire 17 (1), 27e33.
Van Andel, T.H., Tzedakis, P.C., 1996. Paleolithic landscapes of Europe and environs,150,000e25,000 years ago: an overview. Quaternary Science Reviews 15,481e500.
Van der Meulen, A.J., 1973. Middle Pleistocene small mammals from the MontePeglia (Orvieto, Italy) with special reference to the phylogeny of Microtus(Arvicolidae, Rodentia). Quaternaria 16, 1e144.
J.M. López-García et al. / Quaternary Science Reviews 43 (2012) 33e4444
Related Documents











