This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
A multi-assay comparison of seed germination inhibition by Lonicera maackiiand co-occurring native shrubs
Ryan W. McEwan a,n,1, Lucia G. Arthur-Paratley b, Lynne K. Rieske c, Mary A. Arthur a
a Department of Forestry, University of Kentucky, 214 T.P. Cooper, Lexington, KY 40546, United Statesb Bryan Station High School, Lexington, KY 40505, United Statesc Department of Entomology, University of Kentucky, S 225 Ag. North, Lexington, KY 40546, United States
a r t i c l e i n f o
Article history:
Received 18 December 2008
Accepted 27 August 2009
Keywords:
Exotic species
Allelopathy
Phytotoxicity
Asimina triloba
Lindera benzoin
a b s t r a c t
Recent work suggests that germination inhibition via allelopathy may be an important component of
some species’ invasion ecology. We conducted four germination assays to test the inhibitory potential
of Amur honeysuckle (Lonicera maackii), a problematic invasive shrub. The subject species in the first
assay, Festuca arundinacea, exhibited a significant delay in germination when treated with extract from
ground honeysuckle foliage. In the second assay, two concentrations of L. maackii extract were created
by soaking foliage. F. arundinacea germination was not significantly influenced by the treatments while
germination in Impatiens wallerana was substantially decreased. The third assay compared the impact
of foliar extract from honeysuckle, with the native shrubs Lindera benzoin and Asimina triloba using F.
arundinaceae, I. wallerana, Coreopsis lanceolata and Poa pratensis as subject species. In this assay, I.
wallerana was strongly inhibited by L. maackii foliage; however, none of the other species were
significantly influenced by the treatments. In the fourth assay, fruit extracts from L. maackii and L.
benzoin were applied to the same four subject species. L. maackii fruit extract had an inhibitory
influence on seed germination of all subject species, and inhibition from L. benzoin was also noted.
Across all assays, there was a mixed reaction to extract from L. maackii and co-occurring native species
that was species-specific and dependent upon the extract source. Our findings provide support for the
idea that L. maackii has allelopathic activity, but further work is needed to (i) understand how broad the
impact may be across the wide variety of species that are found in its invasion range and (ii)
substantiate that the allelopathic effect has relevance in field environments.
& 2009 Elsevier GmbH. All rights reserved.
1. Introduction
Non-native invasive species often have negative consequencesfor native ecosystems and efforts to prevent their spread arecostly (Ehrenfeld, 2003; Evans et al., 2001; Lovett et al., 2006;Pimental et al., 2000). Invasive species move aggressively intonatural areas, reducing the richness and diversity of plantcommunities, and altering ecosystem function (Ehrenfeld, 2003;Hartman and McCarthy, 2007; Mack et al., 2000; Miller andGorchov, 2004). Although much is known about how invasivespecies impact native plant communities (Collier et al., 2002;Gorchov and Trisel, 2003), less is known about the mechanism(s)by which invasive species establish and maintain site dominance(Meekins and McCarthy, 2001). Some promising recent work hassuggested that belowground processes are important during plantinvasion, and may be key to understanding how invaders
proliferate in native ecosystems (Hierro and Callaway, 2003).Belowground effects of exotic species have included chemicalinterference (allelopathy), wherein native species’ growth andreproduction are inhibited by secondary chemicals exuded by theexotic species (Hierro and Callaway, 2003; Roberts and Anderson,2001; Skulman et al., 2004). Compelling evidence of allelopathiceffects of non-native invasive plants has come from a variety ofspecies and regions (Callaway and Ridenour, 2004; Hierro andCallaway, 2003; Prati and Bossdorf, 2004; Skulman et al., 2004).
The deciduous shrub Amur honeysuckle (Lonicera maackii) is anon-native, invasive plant that has proliferated rapidly in east-central North America in the past 30 years (Hutchinson andVankat, 1997; Luken and Thieret, 1996; Miller and Gorchov,2004). It was introduced to North America from its native Asia foruse as an ornamental, for wildlife cover, and to control soil erosion(Luken and Thieret, 1996). Like other plant invaders, L. maackii isencroaching into natural areas, usurping habitat space at theexpense of native species (Collier et al., 2002; Hutchinson andVankat, 1998; Miller and Gorchov, 2004). Once established,L. maackii reduces the growth and fecundity of native species(Gould and Gorchov, 2000; Gorchov and Trisel, 2003; Hartman
ARTICLE IN PRESS
Contents lists available at ScienceDirect
journal homepage: www.elsevier.de/flora
Flora
0367-2530/$ - see front matter & 2009 Elsevier GmbH. All rights reserved.
doi:10.1016/j.flora.2009.12.031
n Corresponding author. Tel.: +1 937 229 2558.
E-mail address: [email protected] (R.W. McEwan).1 Current address: Department of Biology, The University of Dayton, 300
College Park, Dayton, OH 45469, United States.
Flora 205 (2010) 475–483

Author's personal copyARTICLE IN PRESS
and McCarthy, 2004, 2007). Although the negative influence ofthis species on native plant communities is well established (e.g.,Collier et al., 2002; Gorchov and Trisel, 2003; Luken et al., 1997),the mechanism(s) of this influence are not well understood.
L. maackii negatively influences native plant communities in avariety of ways. It has a dense growth form, sprouts prolifically(Luken et al., 1995, 1997), and has a longer leaf phenology thanmost deciduous shrubs in the habitats it invades (McEwan et al.2009a). Therefore, light occlusion is almost certainly an im-portant factor limiting native species in areas where L. maackii
occurs. Preliminary evidence also suggests that L. maackii sitedominance may be facilitated by allelopathy. Skulman et al.(2004) provide compelling evidence of allelopathic activity inLonicera japonica Thunb, a congener of L. maackii that is also atroublesome invasive species in North America. Dorning andCipollini (2006) demonstrated that L. maackii inhibits seedgermination in both native and non-native forbs, withoutautotoxic effects. Cipollini et al. (2008b) also demonstrated thatL. maackii extract caused a reduction in Arabidopsis thaliana (L.)Heynh reproductive output across a range of nutrient conditions.McEwan et al. (2009b) found that L. maackii leaf material wastoxic to Lymantria dispar L., an insect herbivore with a broad hostrange. These studies strongly suggest that allelopathic activitycould be a contributor to the suite of characteristics that enableinvasion by L. maackii.
We conducted a series of experiments to further our under-standing of the allelopathic potential of L. maackii. We hypothe-sized that (H1.1) L. maackii extract inhibits seed germination, andtested this hypothesis in a series of assays using four differentsubject species. One of the key aspects of allelopathy as a novelweapon associated with invasion success (Callaway and Ridenour,2004) is the absence of this capacity in native species. In order toassess whether allelopathy represents a novel advantage forL. maackii that is absent from native shrubs we included nativeshrubs in our assays and tested the hypothesis (H1.2) thatexudates from native shrubs found in the same habitats asL. maackii do not suppress seed germination.
2. Materials and Methods
2.1. Assay #1: influence of leaf extract from L. maackii and native
ericaceous shrubs on grass seed germination.
The influence of L. maackii foliage on germination ofF. arundinacea Schreb (Fayette Seed, Lexington, KY) was assessed.Festuca arundinacea is a grass species that is a common dominantof agricultural fields in areas where L. maackii is problematic, andtheir ecological interactions have important implications for old-field succession in the region. The influence of L. maackii onF. arundinacea germination was compared to that of two nativeericaceous shrubs, Rhododendron maximum L. and Kalmia latifolia
L. In this assay we also included the exotic congener, L. japonica asa treatment species. In November 2003, 20 g of leaf material wascollected from each species in forests of eastern Kentucky, USA. Ablender was used to macerate fresh leaf material in 200 ml ofdistilled water creating a 10:1 concentration (200 ml of solution;20 g of fresh leaves). This concentration is similar to that used inother studies of allelopathy (e.g., Dorning and Cipollini, 2006;Roberts and Anderson, 2001). The solution was filtered throughcheesecloth to remove leaf particles. Seeds of F. arundinacea
(n=25) were placed on two sheets of Whatman #1 filter paper, ina 100�20 mm Petri dish (n=3, per treatment), treated with 10 mlof either plant material solution or distilled water, and sealedwith Parafilm. Dishes were stored at 20 1C with a 12/12 h day–night light cycle. Germination was recorded every 24 h until
germination had ceased for at least two consecutive days. At theclose of the experiment root and shoot length of the seedlingswere measured.
2.2. Assay #2: influence of L. maackii foliar extract concentration on
grass and forb germination.
In the second assay we tested the germination response of agrass and a forb to two concentrations of L. maackii foliar extract. Inaddition to F. arundinacea, an Impatiens wallerana hybrid (‘‘dwarfwhite baby,’’ Ferry Morse Seeds, Fulton KY) was added to theexperiment. Adding I. wallerana was of ecological and experimentalinterest because (i) it added a forb for contrast with the response ofthe grass and (ii) Impatiens is a genus thought to be susceptible toL. maackii allelopathy (Dorning and Cipollini, 2006). In order toaddress the potential influence of distilled water, we used asimulated precipitation solution in the rest of the assays. Theprecipitation mimic was generated by adding chemical nutrients todistilled water until pH, conductivity, alkalinity, and concentrationsof NO3� , NH4+, Ca2+, Mg2+, K+, Na+, total organic carbon (TOC),Cl, and SO4� matched mean values from rain water collected in anunrelated, long-term precipitation monitoring project at RobinsonForest, in southeastern Kentucky. Leaves were collected in Novem-ber of 2006 and soaked in the precipitation mimic for 24 h, at a 10:1concentration (100 ml of solution, 10 g of fresh leaves). Seeds(n=25) of F. arundinacea and I. wallerana were placed on two sheetsof Whatman #1 filter paper in a 100�20 mm Petri dish (n=3).Dishes were treated with either full concentration (10:1), a 50%concentration (200 ml of solution, 10 g of fresh leaves), or thesimulated precipitation (i.e., control). Dishes were sealed and storedat 23 1C with a 15/9 h day/night light cycle. Germination wasrecorded every 24 h until germination had ceased for at least twoconsecutive days. At the close of the experiment root and shootlength of the seedlings were measured.
2.3. Assay #3: comparison of L. maackii and native shrub foliar
extracts on grass and forb germination.
The third assay assessed the allelopathic influence of L. maackii
foliar extracts compared to that of Lindera benzoin (L.) Blume andAsimina triloba (L.) Dunal, two native shrubs that have a similargrowth form, and are found in the same habitats, as L. maackii. Inaddition to F. arundinacea and I. wallerana, we added Poa pratensis L;(Fayette Seed, Lexington, KY) and Coreopsis lanceolata L. (LivingstonSeed Company, Columbus, OH). P. pratensis co-occurs with F.
arundinacea in old-fields where L. maackii is a problematic invaderand is also found in forest understories where it may be susceptibleto L. maackii influence. It also provided another grass species tocontrast with the response of F. arundinacea. C. lanceolata occurs inthe range of Lonicera invasion and provides contrast with I.
wallerana because Impatiens is a genus thought to be susceptibleto allelopathy, whereas the influence on Coreopsis is unknown.Treatment solutions were created by soaking fresh leaves (collectedin November of 2006) of the three test species in the precipitationmimic for 24 h, at a 10:1 ratio (100 ml of solution, 10 g of freshleaves). Seeds (n=20) of the subject species were placed on twosheets of Whatman #1 filter paper in a 100�20 mm Petri dish (n=3control; n=36 treatments), moistened with 8 ml of solution, sealed,and grown at 25/15 1C, 15/9 h day/night. Germination was recordedevery 24 h until germination had ceased for at least two consecutivedays. At the close of the experiment root and shoot length of theseedlings were measured.
R.W. McEwan et al. / Flora 205 (2010) 475–483476
Author's personal copyARTICLE IN PRESS
2.4. Assay #4: influence of L. maackii and L. benzoin fruit extracts on
grass and forb germination.
In the fourth assay we evaluated the allelopathic influence ofL. maackii and L. benzoin fruit on two grasses, F. arundinacea andP. pratensis, and two forbs, I. wallerana and C. lanceolata, using themethods and replication described in assay #3. Fruits were collectedin November of 2006 and treatment solutions were created bymacerating fruits in a blender containing a 10:1 concentration(100 ml of solution, 10 g of berries). Seeds (n=20) of the subjectspecies were placed upon two sheets of Whatman #1 filter paper ina 100�20 mm Petri dish (n=3 control; n=36 treatments), mois-tened with 8 ml of solution, sealed, and grown at 25/15 1C, 15/9 hday/night. Germination was recorded every 24 h until germinationhad ceased for at least two consecutive days. At the close of theexperiment root and shoot length of the seedlings were measured.
2.5. Statistical analysis
Data were screened for normality using the D’Agostino omnibustest (D’Agostino et al. 1990). The mean number of seeds germinatedthrough time was compared among treatments using repeatedmeasures analysis of variance (RMANOVA). For these analyses,germination was the response variable, leaf extract treatment wasthe between-subjects factor and time (day) was the within-subjectsfactor. Final germination was compared among treatments usinganalysis of variance (ANOVA) or the Kruskal–Wallis Rank ANOVA,depending on normality. Where significant model differences wereidentified by the ANOVA, post-hoc multiple comparisons were usedto determine differences among treatments; the Tukey–Kramermultiple comparison test was used for normally distributed data andthe Kruskal–Wallis multiple comparison Z test was used otherwise.All statistical procedures were conducted using NCSS (Hintze, 2001).
3. Results
3.1. Assay #1: influence of leaf extract from L. maackii and native
ericaceous shrubs on grass seed germination.
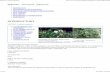
Foliar extracts from L. maackii significantly delayed germi-nation (%) of F. arundinacea seeds compared to native
ericaceous shrubs and the control (Fig. 1). The treatmentshad a highly significant (RMANOVA: F4,10 = 21.1, Po0.001)influence on seed germination. Germination of the control wasnearly at its maximum by day 5; in contrast, fescue seedstreated with L. maackii leaf extract began germination on dayfive and did not reach their maximum until day nine (Fig. 1).Germination of seeds treated with the native shrubs K. latifolia
and R. maximum was intermediate between invasivetreatments and the control for virtually the entire assay(Fig. 1). At the end of the assay, the treatments werestatistically indistinguishable (ANOVA: F4,10 = 1.19, P= 0.37),meaning that the (final) total germination was not reduced byany of the treatments (relative to the control). Extract from thetreatment species created statistical differences in final F.
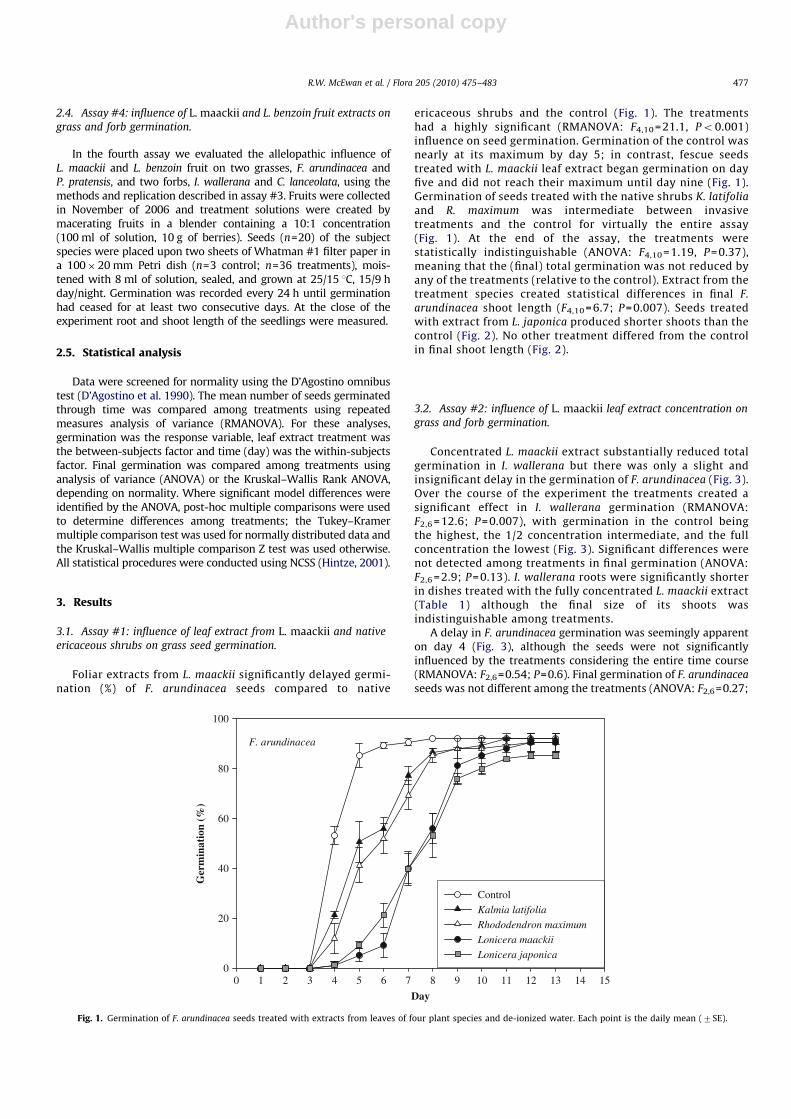
arundinacea shoot length (F4,10 = 6.7; P = 0.007). Seeds treatedwith extract from L. japonica produced shorter shoots than thecontrol (Fig. 2). No other treatment differed from the controlin final shoot length (Fig. 2).
3.2. Assay #2: influence of L. maackii leaf extract concentration on
grass and forb germination.
Concentrated L. maackii extract substantially reduced totalgermination in I. wallerana but there was only a slight andinsignificant delay in the germination of F. arundinacea (Fig. 3).Over the course of the experiment the treatments created asignificant effect in I. wallerana germination (RMANOVA:F2,6 = 12.6; P= 0.007), with germination in the control beingthe highest, the 1/2 concentration intermediate, and the fullconcentration the lowest (Fig. 3). Significant differences werenot detected among treatments in final germination (ANOVA:F2,6 = 2.9; P= 0.13). I. wallerana roots were significantly shorterin dishes treated with the fully concentrated L. maackii extract(Table 1) although the final size of its shoots wasindistinguishable among treatments.
A delay in F. arundinacea germination was seemingly apparenton day 4 (Fig. 3), although the seeds were not significantlyinfluenced by the treatments considering the entire time course(RMANOVA: F2,6=0.54; P=0.6). Final germination of F. arundinacea
seeds was not different among the treatments (ANOVA: F2,6=0.27;
F. arundinacea
Day
0
Ger
min
atio
n (%
)
0
20
40
60
80
100
ControlKalmia latifolia
Rhododendron maximum
Lonicera maackii
Lonicera japonica
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Fig. 1. Germination of F. arundinacea seeds treated with extracts from leaves of four plant species and de-ionized water. Each point is the daily mean (7SE).
R.W. McEwan et al. / Flora 205 (2010) 475–483 477
Author's personal copyARTICLE IN PRESS
F. arundinacea
Day0
Ger
min
atio
n (%
)
0
20
40
60
80
100
I. wallerana
Ger
min
atio
n (%
)
0
20
40
60
80
100
Control
1/2 Concentration
Full
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
Fig. 3. Germination of I. wallerana and F. arundinacea seeds treated with two concentrations of L. maackii extract and a precipitation mimic control solution. Each point is
the daily mean (7SE).
Treatment species
Control
Fin
a F
. aru
ndin
acea
sho
ot le
ngth
(cm
)
0
1
2
3
4
5
6A
AB
B
A
AB
L. maackii L. japonica R. maximum K. latifolia
Fig. 2. Final shoot length of F. arundinacea seedlings treated with extracts from four plant species and a de-ionized water control. Each histogram bar represents the mean
(7SE), and statistical differences (Po0.05) are indicated by letters above the bars.
R.W. McEwan et al. / Flora 205 (2010) 475–483478
Author's personal copyARTICLE IN PRESS
P=0.77; Fig. 3). F. arundinacea seeds did exhibit differences ingrowth due to L. maackii extract; those treated with the fullconcentration produced longer roots and shoots than either the1/2 concentration or the control (Table 1). Confusingly,F. arundinacea seedlings treated with 1/2 concentration hadshoots that were shorter than either the control or fullconcentration treatment (Table 1).
3.3. Assay #3: comparison of L. maackii and native shrub leaf
extracts on grass and forb germination.
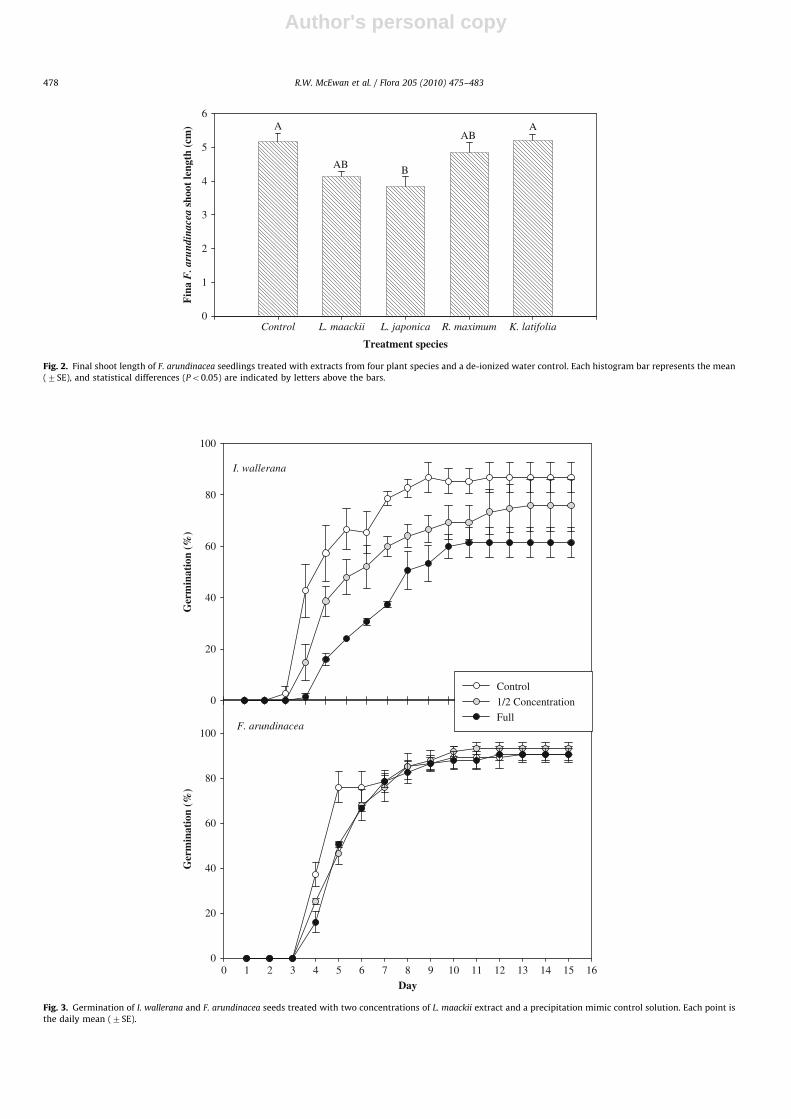
Leaf extracts from L. maackii and native shrubs similarlyinfluenced seed germination (Fig. 4). Leaf extract from L. maackii
created an apparent delay in the germination of F. arundinacea onday 4 that was also evident for extracts of A. triloba and L. benzoin
Table 1Mean (7SE) final seedling root and shoot length (mm) of F. arundinacea and I. wallerana seeding treated with leaf extract from L. maackii at two concentrations and a rain
water mimic control.
Treatments I. wallerana F. arundinacea
Root Shoot Root Shoot
Full concentration 05.170.2a 5.770.3 47.370.3a 58.070.3a½ concentration 11.571.8b 7.171.5 26.771.3b 49.371.0cControl 21.977.8b 6.271.5 31.972.3b 54.170.5b
H2=6.49 F2,6=0.33 F2,6=47.0 F2,6=42.5
P=0.039 P=0.72 Po0.001 Po0.001
Statistical differences (Po0.05) among treatments (within columns) indicated by bold letters, and statistical values are posted in the bottom of columns. H values are
reported for Kruskal–Wallis rank ANOVA, F values for parametric ANOVA.
C. lanceolata
Day
P. pratensis
I. wallerana
Day
0
Ger
min
atio
n (%
)
0
20
40
60
80
100
F. arundinacea
Ger
min
atio
n (%
)
0
20
40
60
80
100
Lonicera
Asimina
Lindera
Control
2 4 6 8 10 12 14 16 18 20 22 0 2 4 6 8 10 12 14 16 18 20 22
Fig. 4. Germination of four subject species when treated with leaf extract from three treatment species and a precipitation mimic control solution. Each point is the daily
mean (7SE).
R.W. McEwan et al. / Flora 205 (2010) 475–483 479

Author's personal copyARTICLE IN PRESS
(Fig. 4A); however, across the time course there were nosignificant differences among treatments (RMANOVA: F3,17=0.71;P=0.56). Likewise, P. pratensis germination was not significantlyinfluenced by the treatments (RMANOVA: F3,17=0.32; P=0.81;Fig. 4B). Neither F. arundinacea nor P. pratensis exhibited significantdifferences in final germination (Fig. 4A,B).
Germination of I. wallerana seeds was strongly influenced byour treatments (Fig. 4C). Across the time course, there was asignificant effect of treatment (RMANOVA: F3,17=6.41; P=0.004),and there was an apparent relationship wherein germination of I.
wallerana seeds treated with A. triloba and L. benzoin wasintermediate between the control solution and L. maackii solution(Fig. 4C). Final germination was different among treatments(ANOVA: F3,17=5.87; P=0.006), with germination of I. wallerana
seeds treated with L. maackii extract significantly lower than allother treatments (Fig. 4C; Tukey–Kramer multiple comparisontest, Po0.05). In contrast, C. lanceolata seed germination was notdistinguishable among treatments (Fig. 4D).
Final seedling size was generally not influenced by leafextracts. The only significant effect was for P. pratensis roots(Table 2). Unexpectedly, both the L. maackii and L. benzoin extracttreatments resulted in significantly longer P. pratensis roots thanthe A. triloba extract and the control (Table 2). Shoot length didnot differ among treatments for any species.
3.4. Assay #4: influence of L. maackii and L. benzoin fruit extracts on
grass and forb germination.
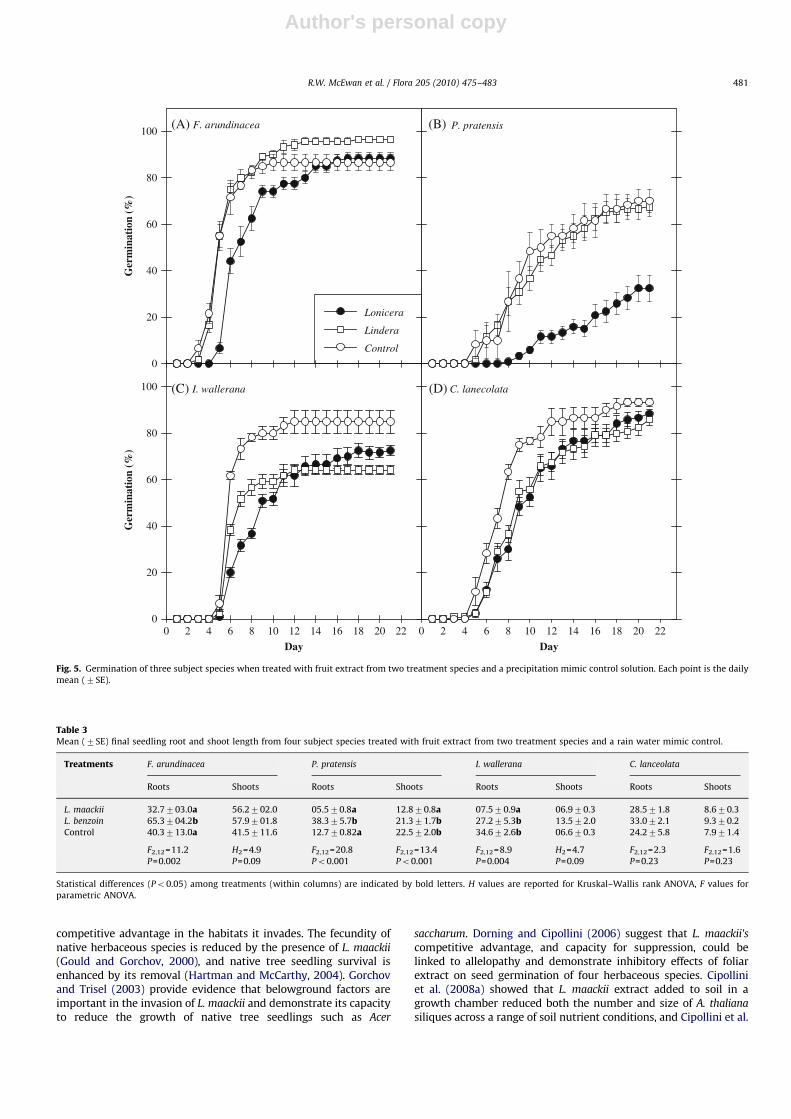
Extract from the fruit of L. maackii influenced germination ofboth grasses and forbs, while L. benzoin fruit extract affected onlyforbs (Fig. 5). In F. arundinacea, there was a highly significanteffect of the treatments on germination (RMANOVA: F2,12=30.9;Po0.001). Through most of the time course, germination of seedstreated with L. maackii was clearly separated from the other twotreatments (Fig. 5A). There was a significant effect of thetreatments on final germination (ANOVA: F2,12=6.14; P=0.01)where the seeds treated with L. maackii were indistinguishablefrom the control, and germination for both were statisticallylower than seeds treated with L. benzoin fruit pulp (Tukey–Kramermultiple comparison test, Po0.05). P. pratensis seeds weremarkedly influenced by the treatments (RMANOVA: F2,12=25.2;Po0.001) and exhibited substantially lower germination whentreated with L. maackii fruit extract than when treated with L.
benzoin or the control (Fig. 5B). Final germination reflected thisgeneral pattern—there was a significant effect of the treatments(ANOVA: F2,12=17.66; Po0.001), and the L. maackii treatmentwas significantly separated from the other two treatments(Tukey–Kramer multiple comparison test, Po0.05).
Germination of I. wallerana seeds was significantly influencedby the fruit extracts (RMANOVA: F2,12=16.09; Po0.001). Through-out the experiment, the control exhibited the (apparently) highestgermination (Fig. 5C, D). At the end of the experiment, finalgermination was significantly influenced by the treatments(ANOVA: F2,12=12.86; P=0.001) with the control statisticallyseparated from both treatments (Tukey–Kramer multiple compar-ison test, Po0.05), which were indistinguishable from oneanother C. lanceolata seed germination was significantly influ-enced by the treatments over the time course of the experiment(RMANOVA: F2,12=5.16; P=0.02), and seeds treated with fruitsolution from L. maackii and L. benzoin exhibited a similar delay ingermination (Fig. 5D). By the end of the experiment, though, therewere no treatment differences (ANOVA: F2,12=12.86; P=0.16).
Seedling response to fruit extracts differed (Table 3). The rootlength of F. arundinacea treated with L. benzoin fruit extract wassignificantly greater than either the L. maackii berry treatment orthe control, which were indistinguishable (Table 3). Shoot lengthof F. arundinacea did not differ among treatments. P. pratensis rootlength and shoot length were significantly shorter for seedlingstreated with L. maackii berry extract (Table 3). Final root length ofI. wallerana seedlings treated with L. maackii berry extract weresignificantly shorter either the L. benzoin treatment or the control.Shoot length of I. wallerana, and root and shoot length of C.
lanceolata, did not differ among treatments (Table 3).
4. Discussion
One puzzling aspect of plant invasion is that the species inquestion are often infrequent and innocuous in native habitats,but proliferate aggressively and form near-monoculture stands intheir invaded habitats. Numerous explanations for plant inva-siveness have been proposed, but one key component of thedisparity in ecological activity may be that in their invadedhabitats these plants contain secondary compounds that facilitateacquisition of habitat space (Callaway and Ridenour, 2004; Hierroand Callaway, 2003; Prati and Bossdorf, 2004). An allelopathiccomponent in the invasion ecology of diffuse knapweed has beendemonstrated (Centaurea diffusa Lam.; Callaway and Aschehoug,2000), and preliminary work suggests that allelopathy may play arole in garlic mustard (Prati and Bossdorf, 2004) and Japanesehoneysuckle (Skulman et al., 2004) invasions. Overall, a growingbody of work supports the idea that allelopathy is a contributingfactor in some plant invasions.
Mounting evidence suggests that allelopathy plays a role in L.
maacki invasion ecology. L. maackii is a relatively minor componentof plant communities in its native habitat, but forms dense shrublayers in deciduous forests of eastern North America (Luken et al.,1995; Luken and Thieret, 1996), suggesting it has a strong
Table 2Mean (7SE) final seedling root and shoot length (mm) from four subject species treated with leaf extract from three treatment species and a rain water mimic control.
Treatments F. arundinacea P. pratensis I. wallerana C. lanceolata
Root Shoot Root Shoot Root Shoot Root Shoot
L. maackii 81.579.4 57.175.2 35.673.9a 23.670.6 19.172.2 8.170.6 42.771.3 10.170.40
L. benzoin 76.973.4 57.672.6 40.572.8a 23.672.2 19.171.1 7.070.6 41.374.8 9.770.8
A. triloba 59.473.7 51.371.6 11.971.4b 19.171.8 20.772.7 11.773.70 37.471.0 9.370.6
Control 53.472.1 48.174.3 18.072.9b 20.671.6 41.9711.8 9.276.4 39.670.5 8.370.3
H3=15.13 H3=6.28 H3=15.13 H3=7.31 H3=3.16 H3=5.03 H3=6.00 H3=4.76
P=0.06 P=0.10 P=0.002 P=0.063 P=0.34 P=0.16 P=0.11 P=0.19
Statistical differences (Po0.05) among treatments (within columns) indicated by bold letters, and statistical values are posted in the bottom of columns. All tests reported
here were from Kruskal–Wallis rank ANOVAs, followed by multiple comparison Z tests.
R.W. McEwan et al. / Flora 205 (2010) 475–483480
Author's personal copyARTICLE IN PRESS
competitive advantage in the habitats it invades. The fecundity ofnative herbaceous species is reduced by the presence of L. maackii
(Gould and Gorchov, 2000), and native tree seedling survival isenhanced by its removal (Hartman and McCarthy, 2004). Gorchovand Trisel (2003) provide evidence that belowground factors areimportant in the invasion of L. maackii and demonstrate its capacityto reduce the growth of native tree seedlings such as Acer
saccharum. Dorning and Cipollini (2006) suggest that L. maackii’s
competitive advantage, and capacity for suppression, could belinked to allelopathy and demonstrate inhibitory effects of foliarextract on seed germination of four herbaceous species. Cipolliniet al. (2008a) showed that L. maackii extract added to soil in agrowth chamber reduced both the number and size of A. thaliana
siliques across a range of soil nutrient conditions, and Cipollini et al.
Table 3Mean (7SE) final seedling root and shoot length from four subject species treated with fruit extract from two treatment species and a rain water mimic control.
Treatments F. arundinacea P. pratensis I. wallerana C. lanceolata
Roots Shoots Roots Shoots Roots Shoots Roots Shoots
L. maackii 32.7703.0a 56.2702.0 05.570.8a 12.870.8a 07.570.9a 06.970.3 28.571.8 8.670.3
L. benzoin 65.3704.2b 57.9701.8 38.375.7b 21.371.7b 27.275.3b 13.572.0 33.072.1 9.370.2
Control 40.3713.0a 41.5711.6 12.770.82a 22.572.0b 34.672.6b 06.670.3 24.275.8 7.971.4
F2,12=11.2 H2=4.9 F2,12=20.8 F2,12=13.4 F2,12=8.9 H2=4.7 F2,12=2.3 F2,12=1.6
P=0.002 P=0.09 Po0.001 Po0.001 P=0.004 P=0.09 P=0.23 P=0.23
Statistical differences (Po0.05) among treatments (within columns) are indicated by bold letters. H values are reported for Kruskal–Wallis rank ANOVA, F values for
parametric ANOVA.
I. wallerana
Day0
Ger
min
atio
n (%
)
0
20
40
60
80
100
(A) (B)
(C) (D)
F. arundinaceaG
erm
inat
ion
(%)
0
20
40
60
80
100
Lonicera
Lindera
Control
C. lanecolata
Day
P. pratensis
2 4 6 8 10 12 14 16 18 20 22 0 2 4 6 8 10 12 14 16 18 20 22
Fig. 5. Germination of three subject species when treated with fruit extract from two treatment species and a precipitation mimic control solution. Each point is the daily
mean (7SE).
R.W. McEwan et al. / Flora 205 (2010) 475–483 481

Author's personal copyARTICLE IN PRESS
(2008b) identified the phenolic compounds apigenin and chloro-genic acid as potentially responsible for this allelopathic activity.
Here we provide further evidence that allelopathy may be afactor in L. maackii invasion. We document clear instances of seedgermination delay and overall inhibition by extracts from L.
maackii. For example, L. maackii consistently influenced I.
wallerana germination and increasing the concentration of L.
maackii leaf extract increased its inhibitory effect. Our strongestevidence of allelopathic activity came from assays that used fruitextracts. L. maackii fruits prolifically, and though these fruit areeaten and dispersed by frugivorous birds (Bartuszevige andGorchov, 2006; Ingold and Craycraft, 1983), there is often anabundance of berries remaining which fall to the ground resultingin a potentially substantial input to the soil. Berry extract from L.
maackii significantly reduced total germination in three of the fourspecies we tested. The fruits of many frugivore-dispersed speciescontain secondary chemicals that defend the fruit against pests,pathogens, inappropriate dispersers, etc. (Herrera, 1982), and itmay be that one or more of these compounds in L. maackii fruitsuppresses seed germination in some species. In any case, our datasuggest that L. maackii has germination-inhibiting constituents infruit pulp and in leaves, and further supports the idea thatallelopathy may play a role in the invasion ecology of this species.
Although our data provide evidence of allelopathic activity in L.
maackii, they also emphasize the difficulties inherent in establishingthe ecological relevance of this activity. For instance, the putativeallelopathic effect of macerated leaves on F. arundinacea germinationwas not present in assays where seeds were treated with extractobtained from soaking leaves. There was a relatively strong apparentallelopathic effect of L. maackii leaf extract (obtained by soaking) on I.
wallerana, but this effect was not apparent for the other species. Theinfluence of L. maackii berry extract was consistent and obvious;however, a similar reduction in forb seed germination was producedby L. benzoin extract and the ecological meaningfulness of inhibitoryproperties in fruit pulp is uncertain. Seedling size data muddled thepattern further as final root length of some subject species wasgreater when exposed to L. maackii extract, while other subjectspecies exhibited shorter roots and shoots. Overall, the responses to L.
maackii extracts were complex, species-specific, tissue-specific, andsometimes indistinguishable from effects created by co-occurringnative species.
Our study adds to a growing body of research that suggestsinvasive species, and specifically L. maackii, may have allelopathicactivity (e.g., Dorning and Cipollini, 2006; Prati and Bossdorf, 2004;Skulman et al., 2004), and also emphasizes the complexities inherentin understanding this phenomenon. Further work is needed to morefully understand the role of allelopathy as a mechanism of speciesinvasion, and to clarify its ecological relevance. First, the complexand species-specific nature of the responses in our study suggestthat additional multi-species screenings are needed to identifysusceptible species groups. Second, the relevance of the potentialallelopathic activity must be explored under more realistic condi-tions. Cipollini et al. (2008) used activated carbon in a field setting inan attempt to separate the influence of allelopathy from that ofcompetition, and found that carbon (and the associated removal ofallelochemicals) had no positive effect, perhaps because the carbonalso neutralized beneficial compounds in the soil. Gorchov andGould (2000) found that the fecundity of three annual herbs wasactually higher in plots where L. maackii had been removed than inareas where it had never occurred, leading to their contention thatthe major mechanism for the negative effects of L. maackii is not dueto the accumulation of allelopathic compounds. More work of thissort is needed. If allelopathy is to be considered a significantcomponent of the invasion ecology of L. maackii, it must bedemonstrated under field conditions. Future work should alsorecognize that allelopathic activity is most likely only one feature
in a suite of attributes that enable invasive species to dominatehabitat space in communities they invade. Identifying theseattributes, and clarifying the relative importance of each, willfurther our ecological understanding of these problematic speciesand facilitate the management of plant invasions.
Acknowledgements
We thank Juan-Carlos Aguilar, Laura Baird, Adrienne Kinney,Ryan Readnower, and Neil Wilson for assistance with datacollection. Thanks also to Millie Hamilton for assistance increating the precipitation mimic solution and general lab support.This work was partially funded by a University of KentuckyFaculty Research Support Grant, and with support from theUniversity of Kentucky College of Agriculture and Department ofForestry. This is a publication of the Kentucky AgriculturalExperiment Station (09-09-091).
References
Bartuszevige, A.M., Gorchov, D.L., 2006. Avian seed dispersal of an invasive shrub.Biological Invasions 8, 1013–1022.
Callaway, R.M., Aschehoug, E.T., 2000. Invasive plants versus their new and oldneighbors: a mechanism for exotic invasions. Science 290, 521–523.
Callaway, R.M., Ridenour, W.M., 2004. Novel weapons: invasive success and theevolution of increased competitive ability. Frontiers in Ecology and theEnvironment 2, 436–443.
Cipollini, D., Stevenson, R., Cipollini, K., 2008a. Contrasting effects of allelochem-icals from two invasive plants on the performance of a nonmycorrhizal plant.International Journal of Plant Sciences 169, 371–375.
Cipollini, D., Stevenson, R., Enright, S., Eyles, A., Bonello, P., 2008b. Phen-olic metabolites in leaves of the invasive shrub, Lonicera maackii, and theirpotential phytotoxic and anti-herbivore effects. Journal of Chemical Ecology 34,144–152.
Cipollini, K.A., McClain, G.Y., Cipollini, D., 2008. Separating effects of allelo-pathy and shading by Alliaria petiolata and Lonicera maackii on growth,reproduction and survival of Impatiens capensis. American Midland Naturalist160, 117–128.
Collier, M.H., Vankat, J.L., Hughes, M.R., 2002. Diminished plant richness andabundance below Lonicera maackii, an invasive shrub. American MidlandNaturalist 147, 60–71.
D’Agostino, R.B., Belanger, A., D’Agostino Jr, R.B., 1990. A suggestion for usingpowerful and informative tests of normality. The American Statistician 44,316–321.
Dorning, M., Cipollini, D., 2006. Leaf and root extracts of the invasive shrub,Lonicera maackii, inhibit seed germination of three herbs with no autotoxiceffects. Plant Ecology 184, 287–296.
Ehrenfeld, J.G., 2003. Effects of exotic plant invasions on soil nutrient cyclingprocesses. Ecosystems 6, 503–523.
Evans, R.D., Rimer, R., Sperry, L., Belnap, J., 2001. Exotic plant invasion alters nitrogendynamics in an arid grassland. Ecological Applications 11, 1301–1310.
Gorchov, D.L., Trisel, D.E., 2003. Competitve effects of the invasive shrub, Loniceramaackii (Rupr.) Herder (Caprifoliaceae), on the growth and survival of nativetree seedlings. Plant Ecology 166, 13–24.
Gould, A.M.A., Gorchov, D.L., 2000. Effects of the exotic invasive shrub Loniceramaackii on the survival and fecundity of three species of native annuals.American Midland Naturalist 144, 36–50.
Hartman, K.M., McCarthy, B.C., 2004. Resoration of a forest understory after theremoval of an invasive shrub, Amur honeysuckle (Lonicera maackii). Restora-tion Ecology 12, 154–165.
Hartman, K.M., McCarthy, B.C., 2007. A dendro-ecological study of forestoverstorey productivity following the invasion of the non-indigenous shrubLonicera maackii. Journal of Applied Vegetation Science 10, 3–14.
Herrera, C.M., 1982. Defense of ripe fruit from pests: its significance in relation toplant–disperser interactions. The American Naturalist 120, 218–241.
Hierro, J.L., Callaway, R.M., 2003. Allelopathy and exotic plant invasions. Plant andSoil 256, 29–39.
Hintze, J. 2001. NCSS and PASS. Number Cruncher Statistical Systems. Kaysville, UT.Hutchinson, T.F., Vankat, J.L., 1997. Invasibility and effects of Amur honeysuckle in
southwestern Ohio forests. Conservation Biology 11, 1117–1124.Hutchinson, T.F., Vankat, J.L., 1998. Landscape structure and spread of the exotic
shrub Lonicera maackii (Amur honeysuckle) in southwestern Ohio forests.American Midland Naturalist 139, 383–390.
Ingold, J.L., Craycraft, M.J., 1983. Avian frugivory on honeysuckle (Lonicera) insouthwestern Ohio in Fall. Journal of the Ohio Academy of Science 83, 256–258.
Lovett, G.C., Canham, C.D., Arthur, M.A., Weathers, K.C., Fitzhugh, R.D., 2006. Forestecosystem response to exotic pests and pathogens in eastern North America.BioScience 56, 395–405.
R.W. McEwan et al. / Flora 205 (2010) 475–483482

Author's personal copyARTICLE IN PRESS
Luken, J.O., Thieret, J.W., 1996. Amur honeysuckle, Its fall from grace. BioScience46, 18–24.
Luken, J.O., Tholemeier, T.C., Kuddes, L.M., Kunkel, 1995. Performance, plasticity,and acclimation of the nonindigenous shrub Lonicera maackii (Caprifoliaceae)in contrasting light environments. Canadian Journal of Botany 73, 1953–1961.
Luken, J.O., Kuddes, L.M., Tholemeier, T.C., Haller, D.M., 1997. Comparativeresponses of Lonicera maackii (Amur honeysuckle) and Lindera benzoin(spicebush) to increased light. American Midland Naturalist 138, 331–343.
Mack, R.N., Simberloff, D., Lonsdale, L.M., Evans, H., Clout, M., Bazzaz, F.A., 2000.Biotic invasions: causes, epidemiology, global consequences, and control.Ecological Applications 10, 689–710.
McEwan, R.W., Birchfield, M.K., Schoergendorfer, A., Arthur, M.A., 2009a. Leafphenology and freeze tolerance of the invasive shrub Amur honeysuckle andpotential native competitors. Journal of the Torrey Botanical Society 136, 212–220.
McEwan, R.W., Rieske, L.K., Arthur, M.A., 2009b. Potential interactions betweeninvasive woody shrubs and the gypsy moth (Lymantria dispar), an invasiveinsect herbivore. Biological Invasions 11, 1053–1058.
Meekins, J.F., McCarthy, B.C., 2001. Effect of environmental variation on theinvasive success of a non-indigenous forest herb. Ecological Applications 11,1336–1348.
Miller, K.E., Gorchov, D.L., 2004. The invasive shrub, Lonicera maackii, reducesgrowth and fecundity of perennial forest herbs. Oecologia 139, 359–375.
Pimental, D., Lach, L., Zuniga, R., Morrison, D., 2000. Environmental and eco-nomic costs of nonindengenous species in the United States. BioScience 50,53–65.
Prati, D., Bossdorf, O., 2004. Allelopathic inhibition of germination by Alliariapetiolata (Brassicaceae). American Journal of Botany 91, 285–288.
Roberts, K.J., Anderson, R.C., 2001. Effect of garlic mustard [Alliaria petiolata (Beib.Cavara and Grande)] extracts on plants and arbuscular mycorrhizal (AM) fungi.American Midland Naturalist 146, 146–152.
Skulman, B.W., Mattice, J.D., Cain, M.D., Gbur, E.E., 2004. Evidence for allelopathicinterference of Japanese honeysuckle (Lonicera japonica) to loblolly andshortleaf pine regeneration. Weed Science 52, 433–439.
R.W. McEwan et al. / Flora 205 (2010) 475–483 483
Related Documents