ALCOHOL DEPENDENCY A molecular mechanism for choosing alcohol over an alternative reward Eric Augier 1 *, Estelle Barbier 1 , Russell S. Dulman 2 , Valentina Licheri 3 , Gaëlle Augier 1 , Esi Domi 1 , Riccardo Barchiesi 1 , Sean Farris 4 , Daniel Nätt 1 , R. Dayne Mayfield 4 , Louise Adermark 3 , Markus Heilig 1 Alcohol addiction leads to increased choice of alcohol over healthy rewards.We established an exclusive choice procedure in which ~15% of outbred rats chose alcohol over a high-value reward. These animals displayed addiction-like traits, including high motivation to obtain alcohol and pursuit of this drug despite adverse consequences. Expression of the g-aminobutyric acid (GABA) transporter GAT-3 was selectively decreased within the amygdala of alcohol-choosing rats, whereas a knockdown of this transcript reversed choice preference of rats that originally chose a sweet solution over alcohol.GAT-3 expression was selectively decreased in the central amygdala of alcohol-dependent people compared to those who died of unrelated causes. Impaired GABA clearance within the amygdala contributes to alcohol addiction, appears to translate between species, and may offer targets for new pharmacotherapies for treating this disorder. A lcohol use accounts for almost 5% of global disease burden (1), and the harm from al- cohol use has been estimated to exceed that due to heroin or cocaine (2). Basic neuro- science has identified brain circuits and molecular mechanisms that contribute to drug and alcohol reward, craving, and relapse (3, 4). These advances have, however, had limited im- pact on the treatment of addictive disorders, sug- gesting that research in model organisms needs to incorporate additional processes (3, 5). Once established, alcohol addiction—hereafter equated with alcoholism—is a chronic relapsing disorder in which alcohol use becomes compul- sive, i.e., continues despite negative consequences (6). Understanding the transition from controlled to compulsive alcohol use is a critical challenge for addiction research. In humans, only a subset of users transition to compulsive drug use (7, 8). In contrast, in commonly used animal models, nearly all rats learn to self-administer addict- ive drugs, including alcohol (9). This points to the possibility that focusing on self-administration may be insufficient to identify key mechanisms of addiction (5, 10). This realization has prompted important con- ceptual advances. When rats are allowed to self- administer cocaine over a prolonged period of time, only a minority of them develop compul- sive drug taking (11, 12). Furthermore, most rats will cease to self-administer cocaine when a high- value alternative becomes available, but a subset of animals continue self-administration despite the presence of an alternative (13, 14). Indeed, in epidemiological studies, only a minority of ex- posed people develop addiction (5, 10). Here, we set out to identify molecular mecha- nisms underlying the choice of alcohol over a nat- ural reward. Conceptually and methodologically, we built on the exclusive choice model pioneered with cocaine and heroin by Ahmed and colleagues ( 5, 13, 14). We first established an exclusive choice- based method to identify rats that continue to self-administer alcohol at the expense of a high- value alternative, a sweet solution, and assessed whether these animals show other characteristics of clinical alcoholism (15). We then used gene expression profiling to identify a molecular mech- anism that mediates compulsive alcohol drinking at the expense of other high-value options. Intense sweetness outcompetes alcohol reward in most but not all rats Sweetness is a fundamental reward that is highly conserved between humans and other animals (16). To tap into its value without the confound of caloric content, we used as alternative reward a solution of the noncaloric sweetener saccharin. In a first experiment (see fig. S1 for experimental timelines), rats (n = 32) were trained to self- administer 20% alcohol for about 10 weeks until reaching stable response rates on a fixed-ratio 3 (FR-3) reinforcement schedule (fig. S2, A and B). They were then offered daily sessions of mu- tually exclusive choice between alcohol and 0.04% saccharin (Fig. 1A). Overall, despite not having previously been trained to respond to saccharin, rats quickly started to choose more saccharin than alcohol (Fig. 1B). The percentage of choice for saccharin over alcohol became even higher when a more rewarding concentration of sac- charin, 0.2%, was introduced (Fig. 1C, significant effect of the saccharin concentration: F 1,31 = 21.56, p < 0.0001, h 2 = 0.41). Once operant responding had stabilized, the rats only chose alcohol over the alternative re- ward 26.2% of the time (Fig. 1C). However, al- though the vast majority of rats strongly preferred saccharin, a minority, 4 rats out of 32 in the first experiment (12.5% of the population, Fig. 1D, left) continued to choose alcohol despite having access to a high-value alternative (alcohol-preferring, AP). Although this was a small number, it aligns well with human addiction rates (7, 8) and promp- ted us to expand the study of individual differences in choice behavior. Ultimately, this percentage was stable across a large number of rats from successive batches (see below; Fig. 1D, right). Alcohol preference was not influenced by hold- ing prior history of alcohol and saccharin self- administration identical (fig. S3). Extensive pre-exposure to saccharin does not affect subsequent alcohol choice People who go on to develop addictive disorders have typically had extensive exposure to sweet reward prior to initiating alcohol use. We there- fore tested whether this kind of preexposure would affect subsequent alcohol choice. A sep- arate group of rats was offered the opportunity to drink a 0.2% saccharin solution or water (ex- periment 2, n = 32 per group) in their home cage for 4 weeks, before operant training was initiated. Rats that had access to the sweet so- lution consumed extensive amounts of saccha- rin during the first week of exposure (101.6 ± 6.8 ml/day) and maintained this consumption throughout their 4 weeks of exposure (week 4: 89.8 ± 5.4 ml/day; fig. S4A). There was no significant difference in acquisition of alcohol self-administration between saccharin- and water- preexposed animals (fig. S4B, no main effect of group: saccharin versus water, F 1,61 = 0.13, p = 0.72; main effect of sessions, F 29,1769 = 5.28, p < 0.001; h 2 = 0.08 but no interaction between group and sessions: F 29,1769 = 0.98, p = 0.48). Extensive preexposure to saccharin also left acquisition of the choice procedure unaffected (fig. S4C, no main effect of group: saccharin ver- sus water, F 1,57 = 0.40, p = 0.53; main effect of sessions, F 18,1026 = 7.82, p <0.001; h 2 = 0.12 but no interaction between group and sessions: F 18,1026 = 1.00, p = 0.45). Once responding had stabilized, both groups strongly favored the sweet solution (22.3 ± 3.8% alcohol choice for the saccharin-exposed group versus 20.6 ± 4.0% for the water-exposed group; fig. S4D, no main effect of group: saccharin ver- sus water, F 1,61 = 0.32, p = 0.57). Again, a sub- population of animals chose alcohol over saccharin (fig. S4E, n = 3 in each group). Extensive pre- exposure to the sweet solution did not affect sampling (fig. S4F) or completed trials ( F 1,61 = 0.56, p = 0.46). Thus, the alcohol choice observed in experiment 1 was not sensitive to the individual’ s prior history of sweet-reward exposure. Finally, we investigated whether AP rats would persist in working for alcohol if the alternative reward was readily available. We used a modified choice paradigm in which rats had continuous access to both sources of reinforcement and found that AP rats maintained a strong preference for RESEARCH Augier et al., Science 360, 1321–1326 (2018) 22 June 2018 1 of 6 1 Center for Social and Affective Neuroscience, IKE, Linköping University, Linköping, 581 83, Sweden. 2 Center for Alcohol Research in Epigenetics, Department of Psychiatry, University of Illinois, Chicago, IL 60612, USA. 3 Department of Psychiatry and Neurochemistry, Sahlgrenska Academy, University of Göteborg, 413 90 Göteborg, Sweden. 4 The Waggoner Center for Alcohol and Addiction Research, University of Texas, Austin, TX 78712, USA. *Corresponding author. Email: [email protected] Corrected 9 July 2018. See full text. on January 25, 2020 http://science.sciencemag.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ALCOHOL DEPENDENCY
A molecular mechanism for choosingalcohol over an alternative rewardEric Augier1*, Estelle Barbier1, Russell S. Dulman2, Valentina Licheri3, Gaëlle Augier1,Esi Domi1, Riccardo Barchiesi1, Sean Farris4, Daniel Nätt1, R. Dayne Mayfield4,Louise Adermark3, Markus Heilig1
Alcohol addiction leads to increased choice of alcohol over healthy rewards.We establishedan exclusive choice procedure in which ~15% of outbred rats chose alcohol over ahigh-value reward. These animals displayed addiction-like traits, including high motivationto obtain alcohol and pursuit of this drug despite adverse consequences. Expression ofthe g-aminobutyric acid (GABA) transporter GAT-3 was selectively decreased within theamygdala of alcohol-choosing rats, whereas a knockdown of this transcript reversed choicepreference of rats that originally chose a sweet solution over alcohol. GAT-3 expressionwas selectively decreased in the central amygdala of alcohol-dependent people comparedto those who died of unrelated causes. Impaired GABA clearance within the amygdalacontributes to alcohol addiction, appears to translate between species, and may offertargets for new pharmacotherapies for treating this disorder.
Alcohol use accounts for almost 5% of globaldisease burden (1), and the harm from al-cohol use has been estimated to exceed thatdue to heroin or cocaine (2). Basic neuro-science has identified brain circuits and
molecular mechanisms that contribute to drugand alcohol reward, craving, and relapse (3, 4).These advances have, however, had limited im-pact on the treatment of addictive disorders, sug-gesting that research in model organisms needsto incorporate additional processes (3, 5).Once established, alcohol addiction—hereafter
equated with alcoholism—is a chronic relapsingdisorder in which alcohol use becomes compul-sive, i.e., continues despite negative consequences(6). Understanding the transition from controlledto compulsive alcohol use is a critical challenge foraddiction research. In humans, only a subset ofusers transition to compulsive drug use (7, 8).In contrast, in commonly used animal models,nearly all rats learn to self-administer addict-ive drugs, including alcohol (9). This points tothe possibility that focusing on self-administrationmay be insufficient to identify key mechanismsof addiction (5, 10).This realization has prompted important con-
ceptual advances. When rats are allowed to self-administer cocaine over a prolonged period oftime, only a minority of them develop compul-sive drug taking (11, 12). Furthermore, most ratswill cease to self-administer cocaine when a high-value alternative becomes available, but a subsetof animals continue self-administration despite
the presence of an alternative (13, 14). Indeed,in epidemiological studies, only a minority of ex-posed people develop addiction (5, 10).Here, we set out to identify molecular mecha-
nisms underlying the choice of alcohol over a nat-ural reward. Conceptually and methodologically,we built on the exclusive choice model pioneeredwith cocaine and heroin by Ahmed and colleagues(5, 13, 14). We first established an exclusive choice-based method to identify rats that continue toself-administer alcohol at the expense of a high-value alternative, a sweet solution, and assessedwhether these animals show other characteristicsof clinical alcoholism (15). We then used geneexpression profiling to identify a molecular mech-anism that mediates compulsive alcohol drinkingat the expense of other high-value options.
Intense sweetness outcompetes alcoholreward in most but not all rats
Sweetness is a fundamental reward that is highlyconserved between humans and other animals(16). To tap into its valuewithout the confound ofcaloric content, we used as alternative reward asolution of the noncaloric sweetener saccharin.In a first experiment (see fig. S1 for experimentaltimelines), rats (n = 32) were trained to self-administer 20% alcohol for about 10 weeks untilreaching stable response rates on a fixed-ratio 3(FR-3) reinforcement schedule (fig. S2, A and B).They were then offered daily sessions of mu-tually exclusive choice between alcohol and 0.04%saccharin (Fig. 1A). Overall, despite not havingpreviously been trained to respond to saccharin,rats quickly started to choose more saccharinthan alcohol (Fig. 1B). The percentage of choicefor saccharin over alcohol became even higherwhen a more rewarding concentration of sac-charin, 0.2%, was introduced (Fig. 1C, significanteffect of the saccharin concentration: F1,31 =21.56, p < 0.0001, h2 = 0.41).Once operant responding had stabilized, the
rats only chose alcohol over the alternative re-
ward 26.2% of the time (Fig. 1C). However, al-though the vast majority of rats strongly preferredsaccharin, a minority, 4 rats out of 32 in the firstexperiment (12.5% of the population, Fig. 1D, left)continued to choose alcohol despite having accessto a high-value alternative (alcohol-preferring,AP). Although this was a small number, it alignswell with human addiction rates (7, 8) and promp-ted us to expand the study of individual differencesin choice behavior. Ultimately, this percentagewas stable across a large number of rats fromsuccessive batches (see below; Fig. 1D, right).Alcohol preference was not influenced by hold-ing prior history of alcohol and saccharin self-administration identical (fig. S3).
Extensive pre-exposure to saccharindoes not affect subsequentalcohol choice
People who go on to develop addictive disordershave typically had extensive exposure to sweetreward prior to initiating alcohol use. We there-fore tested whether this kind of preexposurewould affect subsequent alcohol choice. A sep-arate group of rats was offered the opportunityto drink a 0.2% saccharin solution or water (ex-periment 2, n = 32 per group) in their homecage for 4 weeks, before operant training wasinitiated. Rats that had access to the sweet so-lution consumed extensive amounts of saccha-rin during the first week of exposure (101.6 ±6.8 ml/day) and maintained this consumptionthroughout their 4 weeks of exposure (week 4:89.8 ± 5.4 ml/day; fig. S4A). There was nosignificant difference in acquisition of alcoholself-administration between saccharin- and water-preexposed animals (fig. S4B, no main effect ofgroup: saccharin versus water, F1,61 = 0.13, p =0.72; main effect of sessions, F29,1769 = 5.28, p <0.001; h2 = 0.08 but no interaction betweengroup and sessions: F29,1769 = 0.98, p = 0.48).Extensive preexposure to saccharin also leftacquisition of the choice procedure unaffected(fig. S4C, no main effect of group: saccharin ver-sus water, F1,57 = 0.40, p = 0.53; main effect ofsessions, F18,1026 = 7.82, p <0.001; h2 = 0.12 butno interaction between group and sessions:F18,1026 = 1.00, p = 0.45).Once responding had stabilized, both groups
strongly favored the sweet solution (22.3 ± 3.8%alcohol choice for the saccharin-exposed groupversus 20.6 ± 4.0% for the water-exposed group;fig. S4D, no main effect of group: saccharin ver-sus water, F1,61 = 0.32, p = 0.57). Again, a sub-population of animals chose alcohol over saccharin(fig. S4E, n = 3 in each group). Extensive pre-exposure to the sweet solution did not affectsampling (fig. S4F) or completed trials (F1,61 = 0.56,p = 0.46). Thus, the alcohol choice observed inexperiment 1 was not sensitive to the individual’sprior history of sweet-reward exposure.Finally, we investigated whether AP rats would
persist in working for alcohol if the alternativerewardwas readily available.We used amodifiedchoice paradigm in which rats had continuousaccess to both sources of reinforcement and foundthat AP rats maintained a strong preference for
RESEARCH
Augier et al., Science 360, 1321–1326 (2018) 22 June 2018 1 of 6
1Center for Social and Affective Neuroscience, IKE,Linköping University, Linköping, 581 83, Sweden. 2Centerfor Alcohol Research in Epigenetics, Department ofPsychiatry, University of Illinois, Chicago, IL 60612, USA.3Department of Psychiatry and Neurochemistry,Sahlgrenska Academy, University of Göteborg, 413 90Göteborg, Sweden. 4The Waggoner Center for Alcoholand Addiction Research, University of Texas, Austin,TX 78712, USA.*Corresponding author. Email: [email protected]
Corrected 9 July 2018. See full text. on January 25, 2020
http://science.sciencemag.org/
Dow
nloaded from

alcohol despite the concurrent availability of sac-charin throughout five consecutive sessions (fig. S5).
Alcohol-choosing rats showaddiction-like behaviorsOwing the low frequency of the AP phenotype,we repeated the first experiment on severalbatches of rats and screened them for preferenceduring the choice procedure before carrying outfurther behavioral testing (Fig. 1D, right). Wethen characterized the subpopulation of animalsthat chose alcohol over saccharin with regard toaddiction-like behaviors (experiments 3 and 4)and compared them to the much larger sub-population of rats that stopped working for al-cohol when an alternative high-value option was
available (saccharin-preferring, SP). Only 95 ratsout of 620 tested on the choice procedure con-tinued to choose alcohol despite having access toa high-value alternative (15.3% of the population).We first assessed the motivation of AP and
SP rats to obtain and consume alcohol or sac-charin using a progressive ratio reinforcementschedule (17). AP rats showed a higher motiva-tion to obtain alcohol than SP rats (Fig. 1E, leftbars), whereas the motivation to obtain sac-charin did not statistically differ between thetwo groups (Fig. 1E, right bars; interaction be-tween group (AP versus SP) × reinforcer (alcoholversus saccharin; F1,90 = 31.13, p < 0.0001; h2 = 0.26;post hoc comparison AP versus SP rats for alcoholp < 0.0001, AP versus SP rats for saccharin p = 0.33).
We then investigated whether AP rats wouldmaintain their alcohol drinking despite negativeconsequences: quinine adulteration or footshockpunishment. AP rats showed a robust resistanceto quinine adulteration (Fig. 1F, main effect ofgroup, F1,40 = 4.48, p < 0.05; h2 = 0.10). Theirchoice behavior remained unaffected even whenalcohol was adulterated with a highly aversiveconcentration of quinine, 200 mg/liter. Therewas no statistical difference in the preferencescore for quinine alone when separately testedagainst water (F1,40 = 0.09, p = 0.77; fig. S6A),indicating that the resistance to quinine adul-teration in AP rats was not due to an altered sen-sitivity to the quinine taste. Additionally, therewas no statistical difference in preference score for
Augier et al., Science 360, 1321–1326 (2018) 22 June 2018 2 of 6
Fig. 1. Addiction-like behaviors in rats that choose alcohol over ahigh-value alternative reward. (A) Schematic representation of thediscrete choice procedure. (B) Percentage alcohol choice (±SEM) duringacquisition (n = 32). (C) Stabilized percentage alcohol choice (±SEM;n = 32) (D) Individual distribution. (E) Selectively elevated motivation toobtain alcohol but not saccharin in alcohol-choosing rats [mean breakpoint± SEM during separate sessions of progressive ratio responding for
20% EtOH (left bars) or 0.2% saccharin (right bars; n = 53 for SP rats,39 for AP rats)]. (F) Compulsive alcohol drinking in alcohol-choosing rats,shown by resistance to quinine adulteration of the alcohol solution(percentage change ± SEM in quinine-adulterated alcohol drinking; n = 28for SP rats, 14 for AP rats). (G) Compulsive alcohol drinking, shown byresistance to footshock (percentage change ± SEM in footshock-punished alcohol self-administration; n = 66 for SP rats, 41 for AP rats).
RESEARCH | RESEARCH ARTICLE
Corrected 9 July 2018. See full text. on January 25, 2020
http://science.sciencemag.org/
Dow
nloaded from
a 0.2% solution of saccharin (F1,40 = 0.51, p = 0.48)or 20% ethanol (F1,24 = 0.48, p = 0.49), indicatingthat alcohol preference during choice could not beexplained by preexisting individual differences inpreference for the taste of saccharin or alcohol.Consistent with the quinine adulteration ex-
periment, AP, but not SP, rats also maintainedtheir responding for alcohol when drug deliverywas paired with a contingent footshock punish-ment, (18) {Fig. 1G; Kruskal-Wallis nonparametricanalysis of variance, significant group effect:[H(3,218) = 73.7, p < 0.001]; significant dif-ference between AP and SP rats at 0.2 mA (p <0.001), but not at 0.1 mA (p = 1)}. This was notcaused by any difference in pain sensitivity be-tween AP and SP rats (fig. S6B). Shock-resistantalcohol consumption in this procedure per secould potentially be explained by complex pro-cesses such as counterconditioning or latentinhibition (19). However, because of the strongcorrelation of shock resistance with alcohol choiceand also with resistance to quinine adulteration,we believe that such a mechanism is less likely.Instead, the most parsimonious explanation ap-pears to be that increased appetitive motivationof AP rats for alcohol allows them to overcomethe aversion to footshock punishment.Together, these results show that AP rats dis-
play a constellation of behavioral traits that re-semble those considered diagnostic for addiction(15): (i) The user gives up valuable social andrecreational activities because of substanceuse, here modeled by an increase in choice ofalcohol over the high-value alternative reward,saccharin; (ii) the user shows an increased mo-tivation to obtain and take the drug, spendingexcessive time and effort on its procurement
and consumption, heremodeled using an elevatedbreakpoint on a progressive-ratio schedule (11, 17);and (iii) the user continues drug use despite itsharmful and negative consequences, heremodeledusing continued alcohol self-administration de-spite quinine adulteration or delivery of a shockpunishment contingent with drug delivery (11, 12).Notably, the difference in total alcohol exposureduring self-administration training and choicewas minimal (fig. S7), making it unlikely thataddiction-like behavior in AP rats resulted fromalcohol exposure per se.
Locomotor reactivity to novelty does notpredict alcohol choice
Because locomotor reactivity to a novel envi-ronment predicts aspects of cocaine seeking andtaking (20, 21), we investigated whether this be-havioral marker would also be associated withalcohol choice and other alcohol-related behav-iors (experiment 5). We found that neither al-cohol self-administration nor alcohol choice wasassociated with reactivity to novelty (figs. S8 andS9); nor was reactivity to novelty associated withanxiety-like behaviors (fig. S10).
The GABA transporter GAT-3 isdown-regulated in the amygdalaof AP rats
To identify molecular substrates of alcohol choice,we carried out a gene expression screen in severalbrain regions thought to be involved in alcohol-taking behaviors in rodent models (22) (experi-ment 6). We used a custom Nanostring nCounterarray that contains probes targeting 310 tran-scripts previously hypothesized to be involvedin drug addiction (23, 24). Using this highly
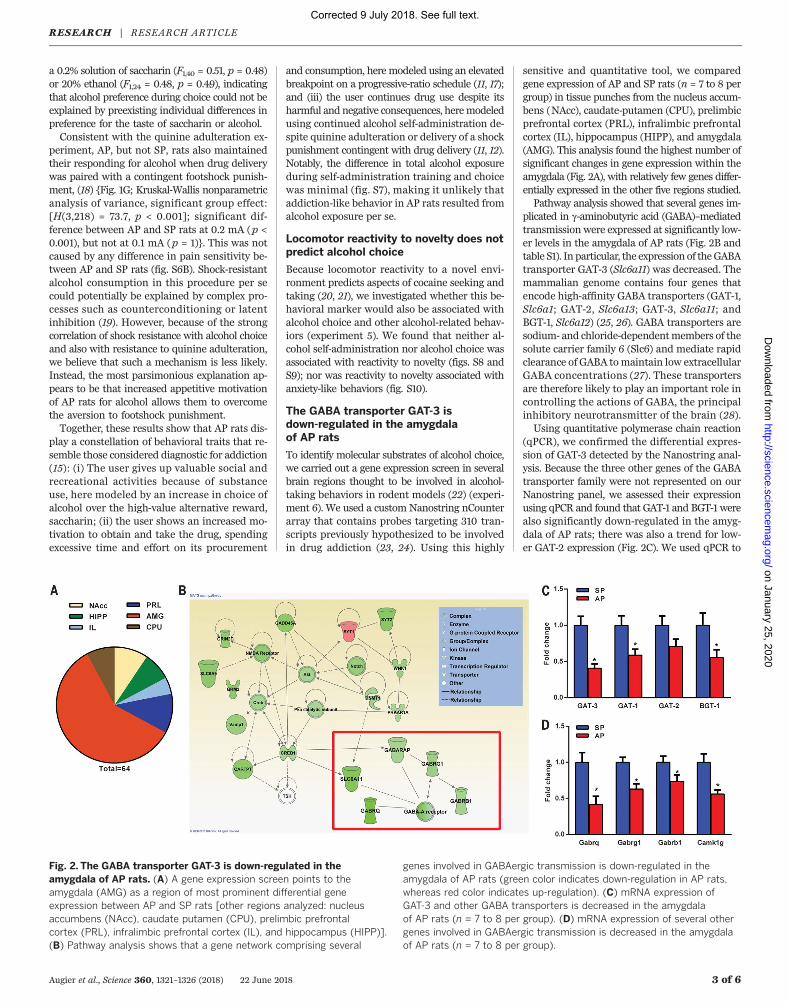
sensitive and quantitative tool, we comparedgene expression of AP and SP rats (n = 7 to 8 pergroup) in tissue punches from the nucleus accum-bens (NAcc), caudate-putamen (CPU), prelimbicprefrontal cortex (PRL), infralimbic prefrontalcortex (IL), hippocampus (HIPP), and amygdala(AMG). This analysis found the highest number ofsignificant changes in gene expression within theamygdala (Fig. 2A), with relatively few genes differ-entially expressed in the other five regions studied.Pathway analysis showed that several genes im-
plicated in g-aminobutyric acid (GABA)–mediatedtransmission were expressed at significantly low-er levels in the amygdala of AP rats (Fig. 2B andtable S1). In particular, the expression of theGABAtransporter GAT-3 (Slc6a11) was decreased. Themammalian genome contains four genes thatencode high-affinity GABA transporters (GAT-1,Slc6a1; GAT-2, Slc6a13; GAT-3, Slc6a11; andBGT-1, Slc6a12) (25, 26). GABA transporters aresodium- and chloride-dependentmembers of thesolute carrier family 6 (Slc6) and mediate rapidclearance of GABA tomaintain low extracellularGABA concentrations (27). These transportersare therefore likely to play an important role incontrolling the actions of GABA, the principalinhibitory neurotransmitter of the brain (28).Using quantitative polymerase chain reaction
(qPCR), we confirmed the differential expres-sion of GAT-3 detected by the Nanostring anal-ysis. Because the three other genes of the GABAtransporter family were not represented on ourNanostring panel, we assessed their expressionusing qPCR and found that GAT-1 and BGT-1 werealso significantly down-regulated in the amyg-dala of AP rats; there was also a trend for low-er GAT-2 expression (Fig. 2C). We used qPCR to
Augier et al., Science 360, 1321–1326 (2018) 22 June 2018 3 of 6
Fig. 2. The GABA transporter GAT-3 is down-regulated in theamygdala of AP rats. (A) A gene expression screen points to theamygdala (AMG) as a region of most prominent differential geneexpression between AP and SP rats [other regions analyzed: nucleusaccumbens (NAcc), caudate putamen (CPU), prelimbic prefrontalcortex (PRL), infralimbic prefrontal cortex (IL), and hippocampus (HIPP)].(B) Pathway analysis shows that a gene network comprising several
genes involved in GABAergic transmission is down-regulated in theamygdala of AP rats (green color indicates down-regulation in AP rats,whereas red color indicates up-regulation). (C) mRNA expression ofGAT-3 and other GABA transporters is decreased in the amygdalaof AP rats (n = 7 to 8 per group). (D) mRNA expression of several othergenes involved in GABAergic transmission is decreased in the amygdalaof AP rats (n = 7 to 8 per group).
RESEARCH | RESEARCH ARTICLE
Corrected 9 July 2018. See full text. on January 25, 2020
http://science.sciencemag.org/
Dow
nloaded from
confirm the down-regulation of additional genesinvolved in GABA-mediated transmission identi-fied by the Nanostring screen. The expression ofgenes encoding several GABAA receptors subunits,Gabrq, Gabrg1, Gabrb1, was also lower in AP rats,presumably reflecting adaptations to an increasedGABA tone due to down-regulated expression oftransporters that clear extracellular GABA (Fig.2D; for detailed statistics, see table S2). Resultsof the Nanostring and qPCR analyses were high-ly correlated (fig. S11). Thus, up-regulated GABA-mediated transmission in the amygdala representsa candidate mechanism for mediating alcoholchoice, which is in agreement with and expandson prior work (29, 30).In further support of a role for GAT-3 in pre-
clinical models of dependence, this transcriptwas also selectively down-regulated in the amyg-dala of the Indiana alcohol preferring rats P-rats(compared to their nonpreferring counterpart;p < 0.01, fig. S12). This strain has been selectivelybred from Wistar rats for high alcohol drinkingand has been proposed to model aspects of hu-man alcoholism (31, 32).
AP rats show increased tonic inhibitionin central amygdala (CeA)
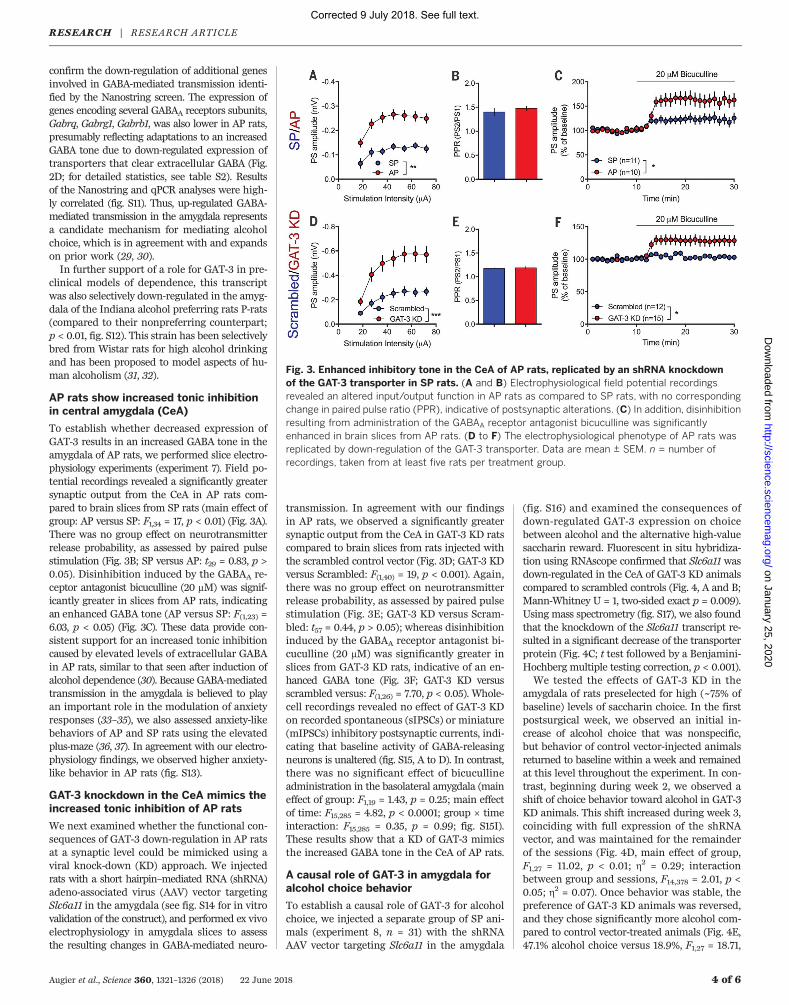
To establish whether decreased expression ofGAT-3 results in an increased GABA tone in theamygdala of AP rats, we performed slice electro-physiology experiments (experiment 7). Field po-tential recordings revealed a significantly greatersynaptic output from the CeA in AP rats com-pared to brain slices from SP rats (main effect ofgroup: AP versus SP: F1,34 = 17, p < 0.01) (Fig. 3A).There was no group effect on neurotransmitterrelease probability, as assessed by paired pulsestimulation (Fig. 3B; SP versus AP: t29 = 0.83, p >0.05). Disinhibition induced by the GABAA re-ceptor antagonist bicuculline (20 mM) was signif-icantly greater in slices from AP rats, indicatingan enhanced GABA tone (AP versus SP: F(1,23) =6.03, p < 0.05) (Fig. 3C). These data provide con-sistent support for an increased tonic inhibitioncaused by elevated levels of extracellular GABAin AP rats, similar to that seen after induction ofalcohol dependence (30). Because GABA-mediatedtransmission in the amygdala is believed to playan important role in the modulation of anxietyresponses (33–35), we also assessed anxiety-likebehaviors of AP and SP rats using the elevatedplus-maze (36, 37). In agreement with our electro-physiology findings, we observed higher anxiety-like behavior in AP rats (fig. S13).
GAT-3 knockdown in the CeA mimics theincreased tonic inhibition of AP rats
We next examined whether the functional con-sequences of GAT-3 down-regulation in AP ratsat a synaptic level could be mimicked using aviral knock-down (KD) approach. We injectedrats with a short hairpin–mediated RNA (shRNA)adeno-associated virus (AAV) vector targetingSlc6a11 in the amygdala (see fig. S14 for in vitrovalidation of the construct), and performed ex vivoelectrophysiology in amygdala slices to assessthe resulting changes in GABA-mediated neuro-
transmission. In agreement with our findingsin AP rats, we observed a significantly greatersynaptic output from the CeA in GAT-3 KD ratscompared to brain slices from rats injected withthe scrambled control vector (Fig. 3D; GAT-3 KDversus Scrambled: F(1,40) = 19, p < 0.001). Again,there was no group effect on neurotransmitterrelease probability, as assessed by paired pulsestimulation (Fig. 3E; GAT-3 KD versus Scram-bled: t57 = 0.44, p > 0.05); whereas disinhibitioninduced by the GABAA receptor antagonist bi-cuculline (20 mM) was significantly greater inslices from GAT-3 KD rats, indicative of an en-hanced GABA tone (Fig. 3F; GAT-3 KD versusscrambled versus: F(1,26) = 7.70, p < 0.05). Whole-cell recordings revealed no effect of GAT-3 KDon recorded spontaneous (sIPSCs) or miniature(mIPSCs) inhibitory postsynaptic currents, indi-cating that baseline activity of GABA-releasingneurons is unaltered (fig. S15, A to D). In contrast,there was no significant effect of bicucullineadministration in the basolateral amygdala (maineffect of group: F1,19 = 1.43, p = 0.25; main effectof time: F15,285 = 4.82, p < 0.0001; group × timeinteraction: F15,285 = 0.35, p = 0.99; fig. S15I).These results show that a KD of GAT-3 mimicsthe increased GABA tone in the CeA of AP rats.
A causal role of GAT-3 in amygdala foralcohol choice behavior
To establish a causal role of GAT-3 for alcoholchoice, we injected a separate group of SP ani-mals (experiment 8, n = 31) with the shRNAAAV vector targeting Slc6a11 in the amygdala
(fig. S16) and examined the consequences ofdown-regulated GAT-3 expression on choicebetween alcohol and the alternative high-valuesaccharin reward. Fluorescent in situ hybridiza-tion using RNAscope confirmed that Slc6a11 wasdown-regulated in the CeA of GAT-3 KD animalscompared to scrambled controls (Fig. 4, A and B;Mann-Whitney U = 1, two-sided exact p = 0.009).Using mass spectrometry (fig. S17), we also foundthat the knockdown of the Slc6a11 transcript re-sulted in a significant decrease of the transporterprotein (Fig. 4C; t test followed by a Benjamini-Hochberg multiple testing correction, p < 0.001).We tested the effects of GAT-3 KD in the
amygdala of rats preselected for high (~75% ofbaseline) levels of saccharin choice. In the firstpostsurgical week, we observed an initial in-crease of alcohol choice that was nonspecific,but behavior of control vector-injected animalsreturned to baseline within a week and remainedat this level throughout the experiment. In con-trast, beginning during week 2, we observed ashift of choice behavior toward alcohol in GAT-3KD animals. This shift increased during week 3,coinciding with full expression of the shRNAvector, and was maintained for the remainderof the sessions (Fig. 4D, main effect of group,F1,27 = 11.02, p < 0.01; h2 = 0.29; interactionbetween group and sessions, F14,378 = 2.01, p <0.05; h2 = 0.07). Once behavior was stable, thepreference of GAT-3 KD animals was reversed,and they chose significantly more alcohol com-pared to control vector-treated animals (Fig. 4E,47.1% alcohol choice versus 18.9%, F1,27 = 18.71,
Augier et al., Science 360, 1321–1326 (2018) 22 June 2018 4 of 6
Fig. 3. Enhanced inhibitory tone in the CeA of AP rats, replicated by an shRNA knockdownof the GAT-3 transporter in SP rats. (A and B) Electrophysiological field potential recordingsrevealed an altered input/output function in AP rats as compared to SP rats, with no correspondingchange in paired pulse ratio (PPR), indicative of postsynaptic alterations. (C) In addition, disinhibitionresulting from administration of the GABAA receptor antagonist bicuculline was significantlyenhanced in brain slices from AP rats. (D to F) The electrophysiological phenotype of AP rats wasreplicated by down-regulation of the GAT-3 transporter. Data are mean ± SEM. n = number ofrecordings, taken from at least five rats per treatment group.
RESEARCH | RESEARCH ARTICLE
Corrected 9 July 2018. See full text. on January 25, 2020
http://science.sciencemag.org/
Dow
nloaded from
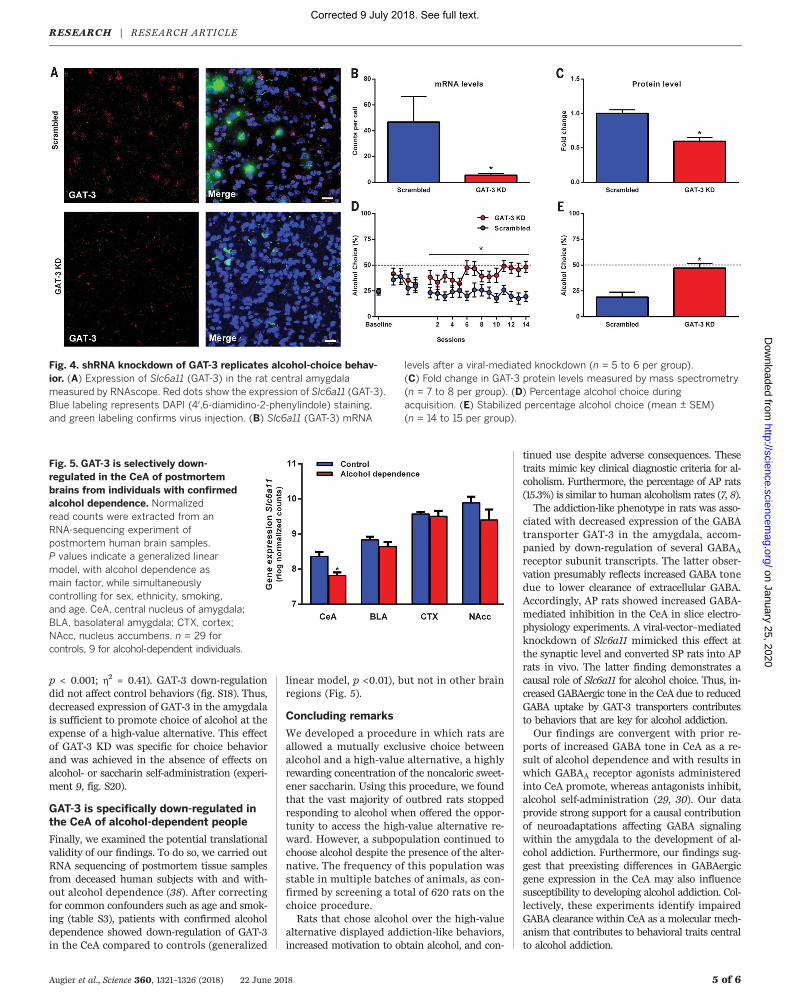
p < 0.001; h2 = 0.41). GAT-3 down-regulationdid not affect control behaviors (fig. S18). Thus,decreased expression of GAT-3 in the amygdalais sufficient to promote choice of alcohol at theexpense of a high-value alternative. This effectof GAT-3 KD was specific for choice behaviorand was achieved in the absence of effects onalcohol- or saccharin self-administration (experi-ment 9, fig. S20).
GAT-3 is specifically down-regulated inthe CeA of alcohol-dependent people
Finally, we examined the potential translationalvalidity of our findings. To do so, we carried outRNA sequencing of postmortem tissue samplesfrom deceased human subjects with and with-out alcohol dependence (38). After correctingfor common confounders such as age and smok-ing (table S3), patients with confirmed alcoholdependence showed down-regulation of GAT-3in the CeA compared to controls (generalized
linear model, p <0.01), but not in other brainregions (Fig. 5).
Concluding remarks
We developed a procedure in which rats areallowed a mutually exclusive choice betweenalcohol and a high-value alternative, a highlyrewarding concentration of the noncaloric sweet-ener saccharin. Using this procedure, we foundthat the vast majority of outbred rats stoppedresponding to alcohol when offered the oppor-tunity to access the high-value alternative re-ward. However, a subpopulation continued tochoose alcohol despite the presence of the alter-native. The frequency of this population wasstable in multiple batches of animals, as con-firmed by screening a total of 620 rats on thechoice procedure.Rats that chose alcohol over the high-value
alternative displayed addiction-like behaviors,increased motivation to obtain alcohol, and con-
tinued use despite adverse consequences. Thesetraits mimic key clinical diagnostic criteria for al-coholism. Furthermore, the percentage of AP rats(15.3%) is similar to human alcoholism rates (7, 8).The addiction-like phenotype in rats was asso-
ciated with decreased expression of the GABAtransporter GAT-3 in the amygdala, accom-panied by down-regulation of several GABAA
receptor subunit transcripts. The latter obser-vation presumably reflects increased GABA tonedue to lower clearance of extracellular GABA.Accordingly, AP rats showed increased GABA-mediated inhibition in the CeA in slice electro-physiology experiments. A viral-vector–mediatedknockdown of Slc6a11 mimicked this effect atthe synaptic level and converted SP rats into APrats in vivo. The latter finding demonstrates acausal role of Slc6a11 for alcohol choice. Thus, in-creased GABAergic tone in the CeA due to reducedGABA uptake by GAT-3 transporters contributesto behaviors that are key for alcohol addiction.Our findings are convergent with prior re-
ports of increased GABA tone in CeA as a re-sult of alcohol dependence and with results inwhich GABAA receptor agonists administeredinto CeA promote, whereas antagonists inhibit,alcohol self-administration (29, 30). Our dataprovide strong support for a causal contributionof neuroadaptations affecting GABA signalingwithin the amygdala to the development of al-cohol addiction. Furthermore, our findings sug-gest that preexisting differences in GABAergicgene expression in the CeA may also influencesusceptibility to developing alcohol addiction. Col-lectively, these experiments identify impairedGABA clearance within CeA as a molecular mech-anism that contributes to behavioral traits centralto alcohol addiction.
Augier et al., Science 360, 1321–1326 (2018) 22 June 2018 5 of 6
Fig. 4. shRNA knockdown of GAT-3 replicates alcohol-choice behav-ior. (A) Expression of Slc6a11 (GAT-3) in the rat central amygdalameasured by RNAscope. Red dots show the expression of Slc6a11 (GAT-3).Blue labeling represents DAPI (4′,6-diamidino-2-phenylindole) staining,and green labeling confirms virus injection. (B) Slc6a11 (GAT-3) mRNA
levels after a viral-mediated knockdown (n = 5 to 6 per group).(C) Fold change in GAT-3 protein levels measured by mass spectrometry(n = 7 to 8 per group). (D) Percentage alcohol choice duringacquisition. (E) Stabilized percentage alcohol choice (mean ± SEM)(n = 14 to 15 per group).
Fig. 5. GAT-3 is selectively down-regulated in the CeA of postmortembrains from individuals with confirmedalcohol dependence. Normalizedread counts were extracted from anRNA-sequencing experiment ofpostmortem human brain samples.P values indicate a generalized linearmodel, with alcohol dependence asmain factor, while simultaneouslycontrolling for sex, ethnicity, smoking,and age. CeA, central nucleus of amygdala;BLA, basolateral amygdala; CTX, cortex;NAcc, nucleus accumbens. n = 29 forcontrols, 9 for alcohol-dependent individuals.
RESEARCH | RESEARCH ARTICLE
Corrected 9 July 2018. See full text. on January 25, 2020
http://science.sciencemag.org/
Dow
nloaded from

REFERENCES AND NOTES
1. H. A. Whiteford et al., Lancet 382, 1575–1586 (2013).2. D. J. Nutt, L. A. King, L. D. Phillips; Independent Scientific
Committee on Drugs, Lancet 376, 1558–1565 (2010).3. M. Heilig, D. H. Epstein, M. A. Nader, Y. Shaham, Nat. Rev.
Neurosci. 17, 592–599 (2016).4. M. Heilig, W. H. Sommer, R. Spanagel, Curr. Top. Behav.
Neurosci. 28, 151–171 (2016).5. S. H. Ahmed, Neurosci. Biobehav. Rev. 35, 172–184 (2010).6. F. A. Wagner, J. C. Anthony, Neuropsychopharmacology 26,
479–488 (2002).7. J. C. Anthony, in Neuropsychopharmacology: The Fifth
Generation of Progress, K. L. Davis, D. Charney, J. T. Coyle,C. Nemeroff, Eds. (Lippincott Williams and Wilkins,Philadelphia, PA, 2002), chap. 109, pp. 1557–1573.
8. B. F. Grant et al., JAMA Psychiatry 72, 757–766 (2015).9. J. R. Weeks, Science 138, 143–144 (1962).10. T. E. Robinson, Science 305, 951–953 (2004).11. V. Deroche-Gamonet, D. Belin, P. V. Piazza, Science 305,
1014–1017 (2004).12. L. J. Vanderschuren, B. J. Everitt, Science 305, 1017–1019
(2004).13. M. Lenoir, F. Serre, L. Cantin, S. H. Ahmed, PLOS ONE 2, e698
(2007).14. L. Cantin et al., PLOS ONE 5, e11592 (2010).15. Diagnostic and Statistical Manual of Mental Disorders (American
Psychiatric Association, Washington, DC, ed. 5, 2013).16. K. C. Berridge, M. L. Kringelbach, Psychopharmacology (Berl.)
199, 457–480 (2008).17. W. Hodos, Science 134, 943–944 (1961).18. T. Seif et al., Nat. Neurosci. 16, 1094–1100 (2013).19. A. Dickinson, J. M. Pearce, Psychol. Bull. 84, 690–711 (1977).
20. P. V. Piazza, J. M. Deminière, M. Le Moal, H. Simon, Science245, 1511–1513 (1989).
21. D. Belin, A. C. Mar, J. W. Dalley, T. W. Robbins, B. J. Everitt,Science 320, 1352–1355 (2008).
22. R. Spanagel, Physiol. Rev. 89, 649–705 (2009).23. E. Barbier et al., Mol. Psychiatry 22, 1746–1758 (2017).24. G. K. Geiss et al., Nat. Biotechnol. 26, 317–325 (2008).25. J. Guastella et al., Science 249, 1303–1306 (1990).26. Q. R. Liu, B. López-Corcuera, S. Mandiyan, H. Nelson,
N. Nelson, J. Biol. Chem. 268, 2106–2112 (1993)[corrected].
27. F. Conti, A. Minelli, M. Melone, Brain Res. Brain Res. Rev. 45,196–212 (2004).
28. J. Bormann, Trends Pharmacol. Sci. 21, 16–19 (2000).29. N. W. Gilpin, M. A. Herman, M. Roberto, Biol. Psychiatry 77,
859–869 (2015).30. M. Roberto, S. G. Madamba, D. G. Stouffer, L. H. Parsons,
G. R. Siggins, J. Neurosci. 24, 10159–10166 (2004).31. T. K. Li, L. Lumeng, W. J. McBride, M. B. Waller, J. M. Murphy,
NIDA Res. Monogr. 66, 41–49 (1986).32. W. J. McBride, T. K. Li, Crit. Rev. Neurobiol. 12, 339–369
(1998).33. P. Botta et al., Nat. Neurosci. 18, 1493–1500 (2015).34. K. M. Tye et al., Nature 471, 358–362 (2011).35. P. Nuss, Neuropsychiatr. Dis. Treat. 11, 165–175 (2015).36. S. Pellow, P. Chopin, S. E. File, M. Briley, J. Neurosci. Methods
14, 149–167 (1985).
37. S. L. Handley, S. Mithani, Naunyn Schmiedebergs Arch.Pharmacol. 327, 1–5 (1984).
38. A. S. Warden, R. D. Mayfield, Neuropharmacology 122, 161–174(2017).
ACKNOWLEDGMENTS
We thank Y. Shaham, G. McNally, P.-V. Piazza, and M. Roberto forvaluable comments. Funding: This work was supported by theSwedish Research Council (grant 2013-7434 to M.H.). L.A. wassupported by the Stiftelsen Psykiatriska Forskningsfonden. Authorcontributions: E.A. and M.H. jointly conceptualized and designedthe studies. E.A. designed and supervised experiments andanalyzed and interpreted results. E.A., R.S.D., and G.A. performedthe behavioral experiments. E.A., E.B., R.S.D., G.A., E.D., andR.B. performed the molecular analyses. E.B., G.A., and E.D.performed surgeries. L.A. and V.L. performed electrophysiologicalrecordings. L.A. supervised electrophysiological experimentsand assisted in manuscript preparation. S.F. and R.D.M. providedIllumina Sequencing data for the human postmortem analysis.D.N. performed bioinformatics analysis. E.A. and M.H. drafted themanuscript. All authors have read and approved the manuscript.Competing interests: The authors report no biomedical financialinterests or potential conflicts of interest. Data and materialsavailability: All data are available in the main text or thesupplementary materials.
SUPPLEMENTARY MATERIALS
www.sciencemag.org/content/360/6395/1321/suppl/DC1Materials and MethodsFigs. S1 to S20Tables S1 to S3References (39–54)
20 June 2017; resubmitted 28 February 2018Accepted 1 May 201810.1126/science.aao1157
Augier et al., Science 360, 1321–1326 (2018) 22 June 2018 6 of 6
RESEARCH | RESEARCH ARTICLE
Corrected 9 July 2018. See full text. on January 25, 2020
http://science.sciencemag.org/
Dow
nloaded from

A molecular mechanism for choosing alcohol over an alternative reward
Daniel Nätt, R. Dayne Mayfield, Louise Adermark and Markus HeiligEric Augier, Estelle Barbier, Russell S. Dulman, Valentina Licheri, Gaëlle Augier, Esi Domi, Riccardo Barchiesi, Sean Farris,
DOI: 10.1126/science.aao1157 (6395), 1321-1326.360Science
, this issue p. 1321; see also p. 1298SciencePostmortem tissue analysis supported the possibility of a similar pathology in human alcoholism.
-aminobutyric acid) clearance in the central amygdala.γadverse consequences. The cause was impaired GABA (traits resembling the human clinical condition, including a high motivation to obtain alcohol and continued use despite high-value alternative (such as sugar) was available. That minority displayed a remarkable constellation of behavioralPerspective by Spanagel). They found that a minority of outbred rats continued to self-administer alcohol even when a
established a choice procedure that begins to address alcohol addiction in rats (see theet al.healthy alternatives. Augier of people confronted with opportunities to consume alcohol involves numerous choices between this drug reward and
''Only'' about 10 to 15% of people exposed to alcohol develop alcohol-related problems. The behavioral repertoireFinding the vulnerable minority
ARTICLE TOOLS http://science.sciencemag.org/content/360/6395/1321
MATERIALSSUPPLEMENTARY http://science.sciencemag.org/content/suppl/2018/06/20/360.6395.1321.DC1
CONTENTRELATED
http://stm.sciencemag.org/content/scitransmed/5/170/170ra14.fullhttp://stm.sciencemag.org/content/scitransmed/5/170/170fs3.fullhttp://stm.sciencemag.org/content/scitransmed/4/116/116ra6.fullhttp://stm.sciencemag.org/content/scitransmed/3/107/107ra109.fullhttp://science.sciencemag.org/content/sci/360/6395/1298.full
REFERENCES
http://science.sciencemag.org/content/360/6395/1321#BIBLThis article cites 50 articles, 12 of which you can access for free
PERMISSIONS http://www.sciencemag.org/help/reprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAAS.ScienceScience, 1200 New York Avenue NW, Washington, DC 20005. The title (print ISSN 0036-8075; online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science. No claim to original U.S. Government WorksCopyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of
on January 25, 2020
http://science.sciencemag.org/
Dow
nloaded from
Related Documents











