ELSEVTER Journal of Neuroscience Methods 57 (1995) 67-75 A modification of biotinylated dextran amine histochemisty for labeling the developing mammalian brain Song-Lin Ding, Andrea J. Elberger * Department ofAnatomy and Neurobiology, The University of Tennessee at Memphis, 8.55 Monroe At:enue, Memphis, TN 1816.1~ VS.1 Received 11 March1994; revised 11 July1994; accepted 21July1994 Abstract Biotinyiated dextran amine (BDA) hasproven to be an excellent anterograde tracer in adult mammalian brains,having some advantages over other anterograde tracers suchas Phaseolus vulgaris-leucoagglutinin (PHA-L) and biocytin. However, results are inferior when BDA is used in neonatal mammals. To improve the sensitivity and quality of BDA labeling in neonatal mammalian brains, the tetramethylbenzidine-sodium tungstate (TMB-ST) method for horseradish peroxidase (WRP) histochemistrywas modified and used in BDA histochemistry. After BDA application to the visual cortex of neonatal rat and cat, contralateral and ipsilateral cortical and subcortical regions were examined for BDA-labeled axons and terminals. The modified BDA histochem- istry produced corpus callosum (CC) axons in neonatal rat and cat that were heavily and continuously labeled, The distribution, trajectories, branching and termination of individual CC axons, and even possible axon-axon contacts, were clearly identified in exquisite detail, even at low magnification. The quality of BDA labeling in the ipsilateral lateral geniculate nucleus and superior colliculus was similar to that of the CC axonal labeling. These results indicate that the modified BDA histochemistry provides a very sensitive and reliable approach to revealing the detailed distribution and morphology of projecting axonsand terminals in the developing mammalian nervous system. Keywords: Biotinylated dextran amine; Tetramethylbenzidine; Mammal; Development; Axonal morphology; Histochemistry: Anterograde: Corpus callosum 1. Introduction The majority of the information on anatomical orga- nization of different pathways has come from studies using retrograde and anterograde tracing techniques. The detailed morphology of pathway origins, including cell bodies and their dendrites, in neonates and adults has been revealed using in vivo methods of combined retrograde tracing and intracellular injections (Buhl et al., 1989; Vercelli and Innocenti, 1993) and co-injection of biotinylated dextran amine (BDA) or biocytin and N-methyl-o,L-aspartic acid (NMDA) (Jiang et al., 1993), or using in vitro methods of retrograde tracing with DiI (Koester and O’Leary, 1993; Elberger et al., 1994). In comparison, the detailed morphology of axons and terminals in adults has been extensively investigated * Corresponding author. E-mail: AELBERGER@UTMEMl. UTMEM.EDU. Tel.: (901) 448-4101: Fax:(901) 448-7193. O165-0270/95/$09.50 0 1995 Elsevier Science B.V. All rights reserved SSDI 0165-0270(94)00127-8 using in vivo Phaseolus vulgar&leucoagglutinin (PHA- L) (Gerfen and Sawchenko, 1984), and more recently biocytin or neurobiotin King et al., 1989; Izzo, 1991; Kita and Armstrong, 1991; Lapper and Bolam, 1991); in vitro studies using DiI have revealed detailed mor- phology of developing axons (e.g., Godement et al., 1987; O’Leary and Terashima, 1988; Elberger, 1993, 1994). However, it is generally accepted that most of the available in vivo anterograde tracers are less effec- tive when used in developing brains than in adult brains (Gerfen and Sawchenko, 1984; Payne et al.. 1988; Weber and Amaral, 1988). To date, only a few anterograde tracing studies exist using PHA-L, or bio- cytin in developing brain (e.g., Lord Plummer and Behan, 1992; Bruce, 1993). Moreover, the rate of suc- cessful use of PHA-L in developing brain is reported to be around 30%, far lower than in adult brain (Weber and Amaral, 1988). Thus, the detailed morphology of axons, especially long-distance projecting axons such as those of the corpus callosum (CC), in developing ani-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ELSEVTER Journal of Neuroscience Methods 57 (1995) 67-75
A modification of biotinylated dextran amine histochemisty for labeling the developing mammalian brain
Song-Lin Ding, Andrea J. Elberger * Department ofAnatomy and Neurobiology, The University of Tennessee at Memphis, 8.55 Monroe At:enue, Memphis, TN 1816.1~ VS.1
Received 11 March 1994; revised 11 July 1994; accepted 21 July 1994
Abstract
Biotinyiated dextran amine (BDA) has proven to be an excellent anterograde tracer in adult mammalian brains, having some advantages over other anterograde tracers such as Phaseolus vulgaris-leucoagglutinin (PHA-L) and biocytin. However, results are inferior when BDA is used in neonatal mammals. To improve the sensitivity and quality of BDA labeling in neonatal mammalian brains, the tetramethylbenzidine-sodium tungstate (TMB-ST) method for horseradish peroxidase (WRP) histochemistry was modified and used in BDA histochemistry. After BDA application to the visual cortex of neonatal rat and cat, contralateral and ipsilateral cortical and subcortical regions were examined for BDA-labeled axons and terminals. The modified BDA histochem- istry produced corpus callosum (CC) axons in neonatal rat and cat that were heavily and continuously labeled, The distribution, trajectories, branching and termination of individual CC axons, and even possible axon-axon contacts, were clearly identified in exquisite detail, even at low magnification. The quality of BDA labeling in the ipsilateral lateral geniculate nucleus and superior colliculus was similar to that of the CC axonal labeling. These results indicate that the modified BDA histochemistry provides a very sensitive and reliable approach to revealing the detailed distribution and morphology of projecting axons and terminals in the developing mammalian nervous system.
Keywords: Biotinylated dextran amine; Tetramethylbenzidine; Mammal; Development; Axonal morphology; Histochemistry: Anterograde: Corpus callosum
1. Introduction
The majority of the information on anatomical orga- nization of different pathways has come from studies using retrograde and anterograde tracing techniques. The detailed morphology of pathway origins, including cell bodies and their dendrites, in neonates and adults has been revealed using in vivo methods of combined retrograde tracing and intracellular injections (Buhl et al., 1989; Vercelli and Innocenti, 1993) and co-injection of biotinylated dextran amine (BDA) or biocytin and N-methyl-o,L-aspartic acid (NMDA) (Jiang et al., 1993), or using in vitro methods of retrograde tracing with DiI (Koester and O’Leary, 1993; Elberger et al., 1994). In comparison, the detailed morphology of axons and terminals in adults has been extensively investigated
* Corresponding author. E-mail: AELBERGER@UTMEMl. UTMEM.EDU. Tel.: (901) 448-4101: Fax: (901) 448-7193.
O165-0270/95/$09.50 0 1995 Elsevier Science B.V. All rights reserved SSDI 0165-0270(94)00127-8
using in vivo Phaseolus vulgar&leucoagglutinin (PHA- L) (Gerfen and Sawchenko, 1984), and more recently biocytin or neurobiotin King et al., 1989; Izzo, 1991; Kita and Armstrong, 1991; Lapper and Bolam, 1991); in vitro studies using DiI have revealed detailed mor- phology of developing axons (e.g., Godement et al., 1987; O’Leary and Terashima, 1988; Elberger, 1993, 1994). However, it is generally accepted that most of the available in vivo anterograde tracers are less effec- tive when used in developing brains than in adult brains (Gerfen and Sawchenko, 1984; Payne et al.. 1988; Weber and Amaral, 1988). To date, only a few anterograde tracing studies exist using PHA-L, or bio- cytin in developing brain (e.g., Lord Plummer and Behan, 1992; Bruce, 1993). Moreover, the rate of suc- cessful use of PHA-L in developing brain is reported to be around 30%, far lower than in adult brain (Weber and Amaral, 1988). Thus, the detailed morphology of axons, especially long-distance projecting axons such as those of the corpus callosum (CC), in developing ani-

68 S.-L. Ding, A.J. Elberger/Journal of Neuroscience Methods 57 (1995) 67-75
mals has been difficult to study and so is not well kIlOWll.
Recently, BDA was reported to be a reliable and sensitive anterograde tracer, revealing detailed mor- phology of labeled axons and terminals in adult pigeon, rat and monkey, as well as in chick embryo (Brandt and Apkarian, 1992; Veenman et al., 1992). To study the detailed morphology, distribution and ‘point-to- point’ connnectivity of developing CC axons in cats and rats, BDA was selected since it has a shorter experi- mental endpoint than PI-IA-L, its histochemistry is simpler than that of PHA-L (Brandt and Apkarian, 1992; Veenman et al., 1992) and it is catabolized much more slowly than biocytin or neurobiotin (King et al., 1989; Izzo, 1991; Kita and Armstrong, 1991; Lapper and Bolam, 1991; Brandt and Apkarian, 1992; Veen- man et al., 1992). Our pilot data showed that BDA was an effective anterograde tracer for the CC axons of neonatal rat and cat, however with the original histo- chemistry method using a nickel-enhanced diami- nobenzidine (DAB) reaction to visualize the BDA (Brandt and Apkarian, 1992; Veenman et al., 19921, few labeled CC axons were found and the labeling was weak. Tetramethylbenzidine (TMB) has been shown to be a very sensitive chromogen for horseradish peroxi- dase (HRP) histochemistry (Mesulam and Rosene, 1979). Recently, sodium tungstate (ST) was introduced as a very effective stabilizer for the HRP-TMB reac- tion product (TMB-ST method) (Gu et al., 1990,1992). In attempting to improve upon our pilot results, we modified and simplified the original TMB-ST method, and applied it to BDA histochemistry. By using this modified method when BDA was applied to one hemi- sphere of neonatal rat or cat, the modification of the TMB-ST method revealed CC axons that were heavily and continuously labeled, as well as extensive termi- nals, in both contralateral cortex and related ipsilateral subcortical structures.
2. Materials and methods
2.1. Surgical procedure
Twenty neonatal rats aged 2,4 and 8 postnatal days (PND), and 5 neonatal cats aged 0 and 3 PND (day of birth = 0 PND) were used for this study. In pilot exper- iments, two methods for applying BDA to cortex were tested in neonatal rat and cat: pressure injection of 20% BDA in 0.9% saline (Veenman et al., 1992), and implantation of BDA-soaked units of filter paper. The second method is simpler and produced more long-dis- tance labeled elements, and all the elements were more clearly labeled; therefore the second method was used. Pieces of standard #2 grade filter paper were soaked in a solution of 20% BDA 10,000 MW (Molecu-
lar Probes, Eugene, OR) in 0.9% saline 2 h before use, and then air dried. When dry, the paper was cut into small units of 0.5 X 1.5 mm (for rat) or 1 X 3 mm (for cat). The rat/cat was anesthetized with sodium pento- barbital (40/33 mg/kg, i.p.1. Under aseptic conditions and microscopic guidance, the scalp was incised, a window was opened in the skull, and the dura was incised. A total of 12 units of BDA filter paper were implanted into the posterior part of the lateral and suprasylvian gyri in each cat, and l-2 units of BDA filter paper were implanted into the cortical region corresponding to presumed visual cortex in each rat. The BDA filter paper was left in place and the scalp was sutured. The animals recovered from the anesthe- sia and were returned to their mothers for extended care.
2.2. BDA histochemistry
After survival periods of 3, 4 and 5 days for 2,4 and 8 PND rats and 6 and 7 days for 0 and 3 PND cats, respectively, the rats/cats were given an overdose of sodium pentobarbital (loo/SO mg/kg, i.p.) and trans- cardially perfused with 0.9% saline followed by a fixa- tive of 4% paraformaldehyde/O.l% glutaraldehyde in phosphate-buffered saline (PBS, pH 7.2). The brains were removed from the skulls and kept overnight in fresh fixative. On the next day, the brains were blocked and rinsed in PBS for 30 min. Coronal sections 100 pm thick were cut on a Vibratome and collected in PBS. After 3 x 10 min rinses in PBS, sections were pre-in- cubated in PBS containing 0.2% Triton X-100 for 30 min and then incubated in an avidin-biotin HRP com- plex solution (Vector) diluted 1: 200 in PBS for 60 min. After 5 x 5 min rinses in PBS, the BDA conjugated with the HRP complex in the sections was visualized by the original TMB-ST reaction (Gu et al., 1990, 1992) or the modified TMB-ST reaction in addition to DAB or nickel-enhanced DAB reaction as in the original BDA method (Veenman et al., 1992).
2.3. Modified TMB-ST reaction
In the modified TMB-ST reaction, (1) the original protocol’s concentrations of TMB, ST and H,O, were reduced by approximately 50%, (2) the original proto- col was simplified by removing HCl from the TMB-ST incubation medium, and (3) the original protocol was simplified by adding all of the H,O, at one time instead of adding the total H,O, in several steps. In addition, the pre-incubation and incubation times in the TMB-ST medium were shortened. The details of the modified TMB-ST reaction are as follows.
A stock solution of 0.2 M PB (pH 5.0-5.4) was made by dissolving 26.6 g of sodium phosphate monobasic (NaH,PO, * H,O; MW: 137.99) and 2.58 g of sodium
phosphate dibasic (Na,HPO, . 12 H,O; MW: 358.14) in 1000 ml of dH,O (Gu et al., 1990, 1992). A TMB solution was freshly prepared by combining 4 mg of TMB (Cr,H,N,; MW: 240.3, Sigma) in 0.5 ml of absolute acetone or ethanol and shaking until the TMB was dissolved. Each 100 ml of TMB-ST incubation medium was prepared as follows: 0.5 g of ST (Na,WO, .2 H,O; MW: 329.86, Sigma) was dissolved in 50 ml of dH,O and filtered. Then 50 ml of 0.2 M PB was added, after which 0.5 ml of a freshly prepared TMB solution was stirred in. The BDA histochemical reac- tion was begun with a short rinse in dH,O (30-60 s), after which sections were pre-incubated in freshly pre- pared TMB-ST medium for 5 min at room tempera- ture and then reacted for 5-8 min in a TMB-ST medium containing 0.005% H,O,. The TMB-ST reac- tion was stopped by rinsing sections 5 x 2 min in 0.05 M PB (pH < 6.5). Then some sections were mounted on gelatin-coated glass slides and counterstained with 1% Neutral Red for 3-5 min. Most sections were reacted for 15-30 min in 0.05 M Tris buffer (pH 7.4, 37°C) containing 0.05% DAB, 0.4% nickel ammonium sulfate and 0.01% H,O, to further stabilize the reac-
tion product (Gu et al., 1991). Sections were mounted on glass slides, dehydrated, cleared and coverslipped.
3. Results
3.1. BDA application site in developing cerebral cortex
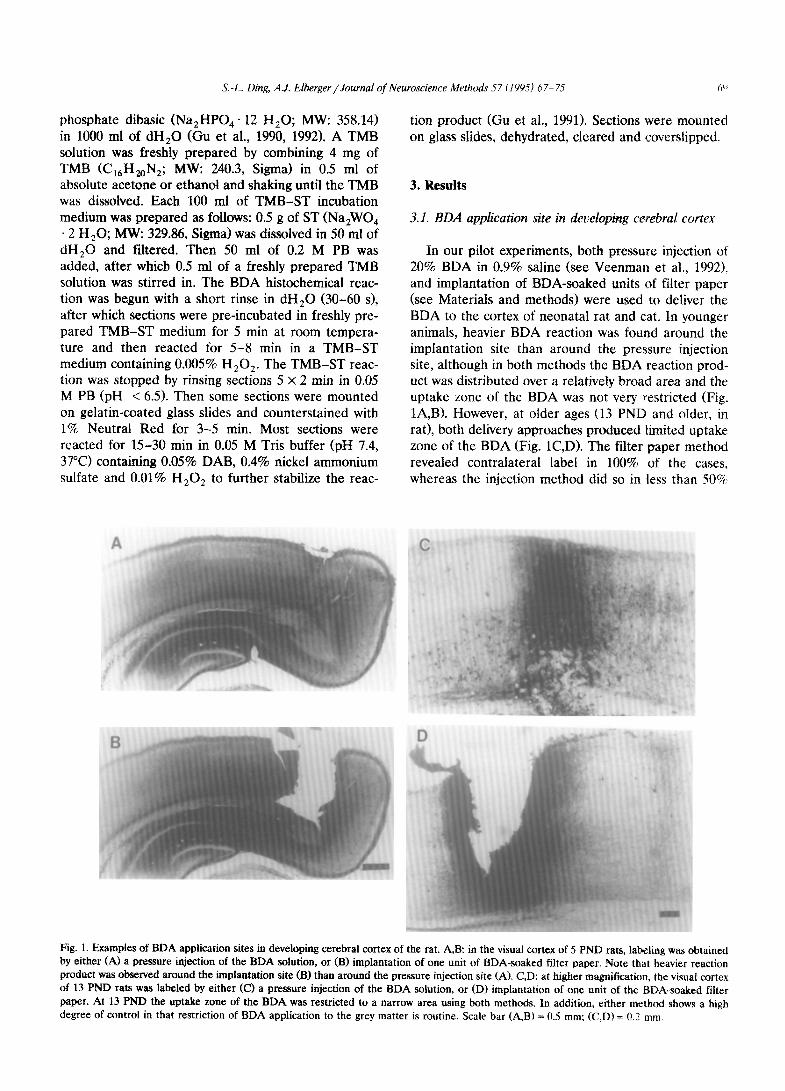
In our pilot experiments, both pressure injection of 20% BDA in 0.9% saline (see Veenman et al., 1992). and implantation of BDA-soaked units of filter paper (see Materials and methods) were used to deliver the BDA to the cortex of neonatal rat and cat. In younger animals, heavier BDA reaction was found around the implantation site than around the pressure injection site, although in both methods the BDA reaction prod- uct was distributed over a relatively broad area and the uptake zone of the BDA was not very restricted (Fig. lA,B). However, at older ages (13 PND and older, in rat), both delivery approaches produced limited uptake zone of the BDA (Fig. lC,D). The filter paper method revealed contralateral label in 100% of the cases, whereas the injection method did so in less than 50%
S.-L. Ding, A.J. Elberger/JournaI of Neuroscience Methods 57 (1995) 67-75 ($i
Fig. 1. Examples of BDA application sites in developing cerebral cortex of the rat. A,B: in the visual cortex of 5 PND rats, labeling was obtained by either (A) a pressure injection of the BDA solution, or (B) implantation of one unit of BDA-soaked filter paper. Note that heavier reaction product was observed around the implantation site (B) than around the pressure injection site (A). CD: at higher magnification, the visual cortex of 13 PND rats was labeled by either (0 a pressure injection of the BDA solution, or (D) implantation of one unit of the BDA-soaked filter paper. At 13 PND the uptake zone of the BDA was restricted to a narrow area using both methods. In addition, either method shows a high degree of control in that restriction of BDA application to the grey matter is routine. Scale bar (A,B) = 0.5 mm: fC,D) = 0.2 mm.
70 S.-L. Ding, A.J. Elberger/Joumal of Neuroscience Methods 57 (1995) 67-75
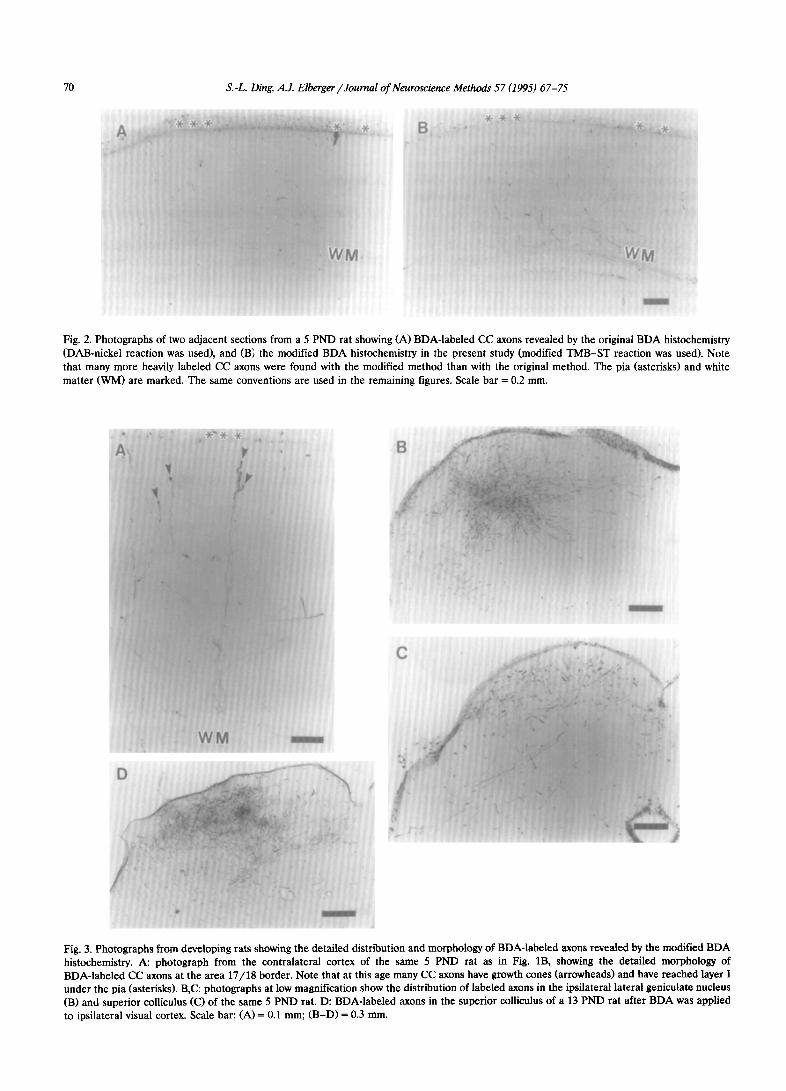
Fig. 2. Photographs of two adjacent sections from a 5 PND rat showing (A) BDA-labeled CC axons revealed by the original BDA histochemistry (DAB-nickel reaction was used), and (B; the modified BDA histochemistty in the present study (modified TMB-ST reaction was used). Note that many more heavily labeled CC axons were found with the modified method than with the original method. The pia (asterisks) and white matter (WM) are marked. The same conventions are used in the remaining figures. Scale bar = 0.2 mm.
Fig. 3. Photographs from developing rats showing the detailed distribution and morphology of BDA-labeled axons revealed by the modified BDA histochemistry. A: photograph from the contralateral cortex of the same 5 PND rat as in Fig. lB, showing the detailed morphology of BDA-labeled CC axons at the area 17/H border. Note that at this age many CC axons have growth cones (arrowheads) and have reached layer I under the pia (asterisks). B,C: photographs at low magnification show the distribution of labeled axons in the ipsilateral lateral geniculate nucleus (B) and superior colliculus (C) of the same 5 PND rat. D: BDA-labeled axons in the superior colliculus of a 13 PND rat after BDA was applied to ipsilateral visual cortex. Scale bar: (A) = 0.1 mm; (B-D) = 0.3 mm.

.5-. -I-. Ding, A.J. Eiberger /Journal of Neumscience Meihds 5 7 ( I WS) h7- 75 ‘7 !
of the cases in the youngest neonates. Furthermore, the contralateral labeling resulting from the injection of BDA was discontinuous, and had a fainter appear- ante. Based on these pilot results, we adopted the implantation approach since this method, in compari- son with the pressure injection, is simpler, less time- consuming and produces better results in neonates.
3.2. Optimizing the chromogen reaction ,for BDA histo- chemistry in neonates
In the original BDA histochemical procedure, a DAB or nickel-enhanced DAB reaction was used to
visualize the BDA; this successfully revealed the de- tailed morphology of labeled axons and terminals in adult mammals (Brandt and Apkarian, 1492; Veenman et al., 1992). However, in our initial experiment. when the nickel-enhanced DAB reaction (Veenman et al., 1992) was used to visualize the BDA in neonatal rat and cat, few labeled CC axons were found and the labeling was weak (Fig. 2A). Therefore, a more sensi- tive method, the TMB-ST method developed for HRP histochemistry (Gu et al., 1990, 1992 J was directly applied to BDA histochemistry for tracing CC axons in neonates: many more CC axons were observed. and the reaction product was so dense that individual f’C’ axons
Fig. 4. Detailed distribution and trajectories of individual CC axons in neonatal cats revealed by the modified RD.4 histochemistry. A: Distribution and morphology of labeled CC axons at area 18/19 border of a 10 PND cat. Note that tangential (short arrows) and radial (long arrows) trajectories and growth cones (arrowheads) of individual CC axons are clearly identified. B: an example from a 6 PND cat showing that 1 CC axon (arrows) in the grey matter stemmed from its parent axon in the white matter (WM), then extended to layer I where it branched into terminal arbors. C: an example from a 10 PND cat showing a parent CC axon (arrowhead) in the deep WM giving rise to 2 latrral collaterals. one of which (arrows) was followed into superficial layers of the cortex. Scale bar: (A.B) = (I I mm: CC) = 0.2 mm.

72 S.-L. Ding, A.J. Elberger/Joumal of Neuroscience Methods 57 (1995) 67-75
and terminals were continuously labeled. Unfortu- nately, the reaction occurred so rapidly that the la- beled CC axons and terminals were visible almost immediately, and heavy non-specific background stain- ing also resulted. In order to reduce the background staining and the reaction speed, the concentrations of TMB, ST and H,O, in the original TMB-ST method (Gu et al., 1990, 1992) were halved. Using the modified TMB-ST method the reaction speed was reduced so that 5-8 min were needed for the TMB-ST reaction; while the background remained clear, the quality of BDA-labeled CC axons and terminals in neonates was
the same as with more concentrated solutions (Figs. 2B, 3-5). Furthermore, the protocol for the TMB-ST reaction was simplified as described in the Materials and methods section, yet results were not compro- mised.
3.3. Quality of the modified BDA histochemical reaction: tracing neonatal axons and terminals
After the BDA-soaked units of filter paper were implanted in the visual cortex of one hemisphere of neonatal rat and cat, contralateral and ipsilateral cor-
Fig. 5. Exquisite details of the labeled CC axons and terminals in layer I of neonatal cats. A: an CC axon (arrowhead) terminates in a clustered series of varicosities; another CC axon (arrow) forms several separate terminal arbors in deep layer I. B: ‘dusty’-like terminal labeling, i.e., dense terminal arbors (arrowhead) of CC axons mixed together. C: 2 CC axons (large arrowheads) tipped with growth cones are extending toward layer I. One CC axon (filled arrow) gives off several terminal arbors which have reached deep layer I. Note that some of the terminal arbors are tipped with growth cones and 1 CC growth cone appears to be making contact with another CC axon (open arrow). All growth cones are indicated by small arrowheads. D: 1 CC axon (arrows) tipped with a growth cone extends tangentially for a long distance within layer I. The open arrow indicates a close contact of a growth cone with another CC axon in layer I. Growth cones (small arrowheads) are marked. Scale bar (A-D) = 0.1 mm.

S.-L. Ding, A.J. Elberger /Journal of Neuroscience Methods 57 (149.5) 67-Z 71
tex, as well as ipsilateral subcortical structures, were labeled. In all cases studied, the morphology of LabeIed axons could be examined in great detail because the axons were heavily and continuously labeled. For ex- ample, at the earliest age studied, 1 unit of BDA filter paper was implanted in a 2 PND rat and the animal was killed at 5 PND; at low magnification, even with a 2.5 x objective and a 2.5 X photo-eyepiece, labeled ax- ons were clearly observed in contralateral cortex (i.e., CC axons) and in ipsilateral subcortical structures such as the dorsal lateral geniculate nucleus and superior colliculus (Figs. 2B, 3A-Cl. At older ages, the quality of axonal labeling was even better (Fig. 3D). At a higher magnification, the distribution and morphology of labeled axons could be observed in exquisite detail. For instance, at the area 18/19 border of a 10 PND cat, many labeled CC axons were followed from the white matter to different cortical layers; some of them ended in growth cones (Fig. 4A). Precise trajectories and branchings of individual CC axons could be de- tected in all cases (Fig. 4B,C). In addition, labeled axon terminals were well defined, as shown in layer I of 10 PND cats (Fig. 51. The detailed terminal branchings of CC axons (Fig. 5A-Cl and the tangential extention of CC axons within layer I were clearly revealed (Figs. 4A, 5B,D). Interestingly, possible axon-axon contacts were also clearly visible at the light microscope level using the present modification (Fig. 5C,D).
4. Discussion
The modification of BDA histochemistry in the pre- sent study has preserved the advantages of BDA label- ing observed in adult mammals, namely the exquisite detailing of trajectories, branchings and termination of individual CC axons, to be replicated in neonatal mam- mals. This indicates that the modified method is a very sensitive and reliable approach for examining the pre- cise distribution and morphology of projecting axons and terminals in developing brains. One of the original BDA studies showed clear IabeIing of periphera1 ner- vous system (PNS) in chick embryo (Veenman et al., 1992), but our pilot experiments using the same method showed only weak labeling of CC axons in neonatal rat and cat. This discrepancy could be due to a number of factors, including differences in length of pathways studied, animal class (Aves vs. Mammals), neuron or axon type (PNS vs. CNS), and/or methodology (use of Tyrode for injection and sucrose for cryoprotection vs. neither chemical). In the latter case, sucrose has been shown to affect benzidine substances when applied to HRP reactions (Elberger et al., 1992). In any case, even in adult mammals, our modified method showed better BDA labeling of CC axons and terminals than did the original method (Ding and Elberger, 1994b). Since the
fine morphology of long projecting axons, especially those in neocortex, have been difficutt to investigate with other methods (Rockland, 19921, the modified BDA method of the present study should prove to be a useful alternative for in vivo studies of developing pathways. For example, developing CC axons were previously labeled in vivo by radioactive amino acids, HRP, or wheat-germ agglutinin conjugated to HRP (WGA-HRPl (Wise and Jones, 1976; Goldman and Nauta, 1977; Innocenti, 1981; Ivy and Killackey, 1981; Innocenti and Clarke, 1984; Olavarria and Van Sluyters, 1985). These tracers produced punctate or precipitate labeling of presumed CC axons and termi- nals so that main axons could not be identified. In contrast, the detailed morphology of CC: axons in the visual cortex of neonatal rat and cat was clearly demonstrated with the modified in vivo method in the present study. In addition, the present modified method has revealed the existence of transitory CC axons in area 17 of neonatal rat and cat (Ding and Elberger, 1993, 1994a,b) which was not revealed with other in vivo tracers in previous studies (Innocenti, 1981; Inno- centi and Clarke, 1984; Olavarria and Van Sluyters. 1985), but was only demonstrated with the in vitro tracer DiI (Elberger, 1993, 1994).
The TMB-ST reaction product has been shown to be stable for HRP (Gu et al., 1990, 1992). Further- more, this product could be enhanced and further stabilized with DAB-cobalt or DAB-nickel reaction; at the ultrastructural level, the combination of the TMB-ST reaction and DAB-cobalt reaction showed good tissue preservation and the reaction product ap- peared as characteristic crystal-like structures which were extremely electron dense and often aggregated into clumps (Gu et al., 1990, 1992). The present BDA histochemistry with the modified TMB-ST reaction is likely to be effective for EM analysis, and is expected to maintain the advantages of the original BDA histo- chemistry shown when combined with other labeling methods (Veenman et al., 19921 and when analyzed with the EM (Wouterlood and Jorritsma-Byham, 1993).
The increased sensitivity of the modified BDA his- tochemistry in the present study relied on the use of a very sensitive chromogen, TMB (Mesularn and Rosene, 1979). Thus, it is possible that the sensitivity of other anterograde tracers such as WGA-HRP, PHA-L, bio- cytin or neurobiotin can also be improved with the TMB-ST reaction. Even with such an improvement, BDA has several advantages over these other tracers. For instance, PI-IA-L is applied iontophoreticalty, re- quires a 2- or 3-step immunohistochemical procedure (Gerfen and Sawchenko, 19841, and was reported by some authors to be unreliabie in cats and monkeys (e.g., Brandt and Apkarian, 1992) and in developing brains (e.g., Weber and Amaral, 1988). Therefore. use of PHA-1, is more labor-intensive, requires special

14 S.-L. Ding, A.J. Elberger/Journal of Neuroscience Methods 57 (1995) 67-75
equipment, and is successful in a limited population. A disadvantage for WGA-HRP is that trans-synaptic la- beling can result with high concentrations and long survival times (Apkarian and Hodge, 1989). Biocytin or neurobiotin appears to be degraded quickly (King et al., 1989; Izzo, 1991; Kita and Armstrong, 1991; Lapper and Bolam, 1991; Norita et al., 1991); for example, the quality of labeling with these tracers deteriorates after a 24 h survival time (Kita and Armstrong, 1991; Lapper and Bolam, 1991). Thus, WGA-HRP, biocytin and neurobiotin might not be very effective and reliable for tracing long-projecting axons, especially in developing brains. In contrast, BDA can be used with survival times ranging from several days for rat pups to several weeks for adult monkeys, yet good labeling quality is maintained while no trans-synaptic labeling is observed (Brandt and Apkarian, 1992; Veenman et al., 1992). In addition, the BDA application process is simpler, since it can be applied by pressure injection (Brandt and Apkarian, 1992; Veenman et al., 1992) or implanting BDA soaked units of filter paper (present study). Nev- ertheless, it should be pointed out that WGA-HRP, PHA-L, biocytin or neurobiotin might also be useful in tracing developing axons, especially relatively short-dis- tance projecting axons, when the TMB-ST reaction is used.
Recently DiI has been used to trace long-projecting axons (e.g., Godement et al., 1987; O’Leary and Terashima, 1988; Elberger, 1993, 1994). Two major advantages of DiI are (1) it is very sensitive in develop- ing brains, and (2) it can be used in fiied tissue, eliminating the effect of post-surgical survival times for in vivo tracers on the results in developmental studies; length of survival times creates difficulty in interpreting some developmental results. Two major limitations of DiI are (1) it is difficult to use to study ‘point-to-point’ connectivity because application to regions such as neocortex produces unsatisfactory diffusion (Elberger, unpublished data), and (2) diffusion to completely label connections needs a relatively long time. Therefore, the complementary advantages of in vitro DiI and in vivo BDA can be used when applied to the same investigations in developmental studies.
In the present study, BDA-soaked units of filter paper were used for BDA delivery to the cortex in the neonate; this method obtained better results than using pressure injections of the BDA. This is likely due to 2 factors. Firstly, the pressure injection may produce leaking of a significant amount of BDA along the needle track in the cortex of the neonate, while the filter paper is kept in situ to reduce the leakage of BDA. Secondly, the filter paper allows BDA to be released more slowly than does the pressure injection and thus more BDA is taken up over time at the application site, compared to the injection of BDA solution. However, our preliminary results showed that
at older ages (13 PND onward in the rat), both pres- sure injection and the filter paper implantation re- sulted in satisfactory labeling of CC axons (not shown). Therefore, application of BDA in non-superficial structures by pressure injection or iontophoresis could also produce enhanced results with the modified BDA histochemistry, but this is more likely to occur in older animals. In the younger animal, it should be noted that the specific zone of uptake of the BDA delivered by pressure injection or filter paper is not very defined because the neonate has much more extracellular space which allows the tracer to diffuse over a relatively large region around the site of application (Payne et al., 1988). Thus, for investigations of ‘point-to-point’ pro- jections in the neonatal cortex, an alternative ap- proach, ionotophoretic application of BDA might be desirable since iontophoresis could produce very re- stricted injection sites (Gerfen and Sawchenko, 1984; Veenman et al., 1992). Nevertheless, at older ages (13 PND onward in rat), specific zones of uptake of the BDA (see Fig. 1) are able to be achieved by both pressure injection and the filter paper; therefore these two approaches may also be used for revealing ‘point- to-point’ projections in developing brains. In any case, the modified BDA histochemistry in the present study should be able to improve the quality of labeling in developing brains.
In conclusion, the modification for BDA histochem- istry detailed in the present study creates a simple, reliable and very sensitive technique for investigating the detailed distribution, trajectories, branchings and terminations of axons, especially long-projecting ones, in the CNS of neonate through adult.
Acknowledgements
This work was supported by NIH grant EY08466 (A.J.E.). Partial support (S.L.D.) came via a Postdoc- toral Fellowship from the State of Tennessee Neuro- science Center of Excellence.
References
Apkarian, A.V. and Hodge, C.J. (1989) The primate spinothalamic pathways. III. Thalamic terminations of the dorsolateral and ventral spinothalamic pathways, J. Comp. Neurol., 288: 493-511.
Brandt, H.M. and Apkarian, A.V. (1992) Biotin-dextran: a sensitive anterograde tracer for neuroanatomic studies in rat and monkey, J. Neurosci. Methods, 45: 35-40.
Bruce, L.L. (1993) Postnatal development and specification of the cat’s visual corticotectal projection: efferents from the postero- medial lateral suprasylvian area, Dev. Brain Res., 73: 47-61.
Buhl, E.H., Schwerdtfeger, W.K., Germroth, P. and Singer, W. (1989) Combining retrograde tracing, intracellullar injection, an-

S -I>. Ding, A.J. Elberger / Journal of Neurosciewe Methods 57 i 199.5) h7- ?i 7..,
terograde degeneration and electron microscopy to reveal synap- tic links, J. Neurosci. Methods, 19: 241-250.
Ding, S.-L. and Elberger, A.J. (1993) Existence of transitory corpus callosum axons throughout visual cortex in neonatal cat and rat confirmed using in vivo biotinylated dextran amine, Sot. Neu- rosci. Abst., 19: X92.
Ding, S.-L. and Elberger, A.J. (1994a) Confirmation of the existence of transitory corpus callosum axons in area 17 of neonatal cat: an anterograde tracing study using biotinylated dextran amine. Neu- rosci. Lett.. 177: 66-70.
Ding, S.-L. and Elberger, A.J. (1994b) Postnatal development of corpus callosum axons projecting to the visual cortex of the rat: an anterograde tracing study with biotinylated dextran amine. submitted.
Elbergrr. A.J. (1993) Distribution of transitory corpus callosum ax- ons projecting to developing cat visual cortex revealed by Dil, J. (Jnmp. Neural.. 333: 326-342.
Elbergcr. A.J. (1994) Distribution of transitory corpus callosum ax- ons projecting to developing rat visual cortex revealed by Dil, (.‘errb. Cortex. 4: 279-299.
Elberger. A.J.. Hester. M.M. and Stiles, H. (1992) HRP reacted with the chromogen o-tolidine produces whole-cell reaction product at light and electron microscope levels: negative effects of sucrose and Golgi staining on henzidine reactions. J. Neurosci. Methods, 45 ‘27-73X.
Elberger. A.J.. Murphree. V.J., Ding. S.-L. and Seaton, D. (1994) Developmental changes in the distribution and morphology of cell5 in ciaual cortex projecting through the corpus callosum. submitted.
Gerfen. C.R. and Sawchenko, P.E. (1984) An anterograde neu- roanatomical tracing method that shows the detailed morphology elf neurons. their axons and terminals: immunohistochemical lo- calization of an axonally transported plant lectin, Phaseolus WI- xar;s-leucoagglutinin, Brain Res.. 290: 219-238.
Godement. P.. Vanselow. J.. Thanos, S. and Bonhoeffer, F. (1987) A study in developing visual systems with a new method of staining neur(ms and their processes in fixed tissue, Development, 101: 097 713
Goldman, P.S. and Nauta, W.J.H. (1977) Columnar distribution of corttcocortical fibers in the frontal, association, limbic and motor cortex ot the developing rhesus monkey, Brain Res., 122: 393-413.
Gu. Y.. (‘hen, Y. and Ye, 1,. (1990) A new highly sensitive HRP-TMB method using sodium tungstate as a stabilizer. I. Light micro- scopic study, Chin. J. Neuroanat.. 6: 121-127.
Gu, Y. Chen, Y. and Ye, L. (1992) Electron microscopical demon- stration of horseradish peroxidase by use of tetramethylbenzidine as chromogen and sodium tungstate as stabilizer (TMB-ST method): a tracing method with high sensitivity and well pre- sel,ed ultrastructural tissue, J. Neurosci. Methods, 42: l-10.
Gu, \ Xie. >‘. and Chen. Y. (1991) The study on the posttreating procedures for the tetramethylbenzidine-sodium tungstate (TMB-ST) reaction product, Guangdong Anat. Bull., 13: 14-19.
Innocenti, G.M. (1981) Growth and reshaping of axons in the estab- Irshment of visual callosal connections. Science, 212: 824-827.
Innocenti, G.M. and Clarke. S. (1984) The organization of immature cailosal connections, J. Comp. Neural.. 230: 287-309.
Ivy, G t ). and Killackey. H.P. (1981) The ontogeny of the distribution of cdllosal projection neurons in the rat parietal cortex, J. Comp. Neural.. 195: 367-3X9.
IZZO. P.U ( 1991 i /\ note on the USC of hiocytin in anterograde tracing
studies in the central nervous system: application at both light and electron microscopic level, J. Neurosci. Methods. 36: 1% 166.
Jiang, X., Johnson, R.R. and Burkhalter, A. f 1993) Visualization of dendritic morphology of cortical projection neunlm by retrograde axonal tracing, J. Neurosci. Methods, 511: 45-60
King. M.A., Louis. P.M.. Hunter, BE. and Walker. D.W. (1989i Biocytin: d versatile anterograde neuroanatomicnl tracr-tracing alternative. Brain Res.. 497: 361- 367.
Kita. f-1. and Armstrong, W. (1991) A bir)tin-cont;lining cr>mpound N-(2.aminoethyl) biotinamide for intracellular i&cling and neu ronal tracing studies: comparison with bioc\tin. J Yeurnhcl. Methods. 37: 131.-150.
Koestrr. S.E. and O‘Leary. D.D.M. (19%) Fun::tional classes of cortical projection neurons develop dendritlc distinctions hy class-specific hculpting 11l’ an early common pat:erci. .l_ Neuroscl. 12: 13X’-- 1.393.
Lapper, S.R. and Bolam. J.P. (1991) The anterograde and retrograde transport of neurobiotin in the central nervous system of the rat: comparison with hiocytin. J. Neurosci. Method?,. 3Y: 16.i- 175.
Lord Plummer. Ii. and Brhan. M. (1992) Postnatal development of the corticotectal projection in cath. J. Camp. heurc~l .ilt;: 17X 1’39.
Mesulam. M.-M. and Rosenr, D.L. i. 1979) Sensitn ity m hurheradish peroxidase neurohistochemistry: a comparative and quantitatlvc study of nine methods, J. Histochem. <‘ytocheni.. 27: 763.-773.
Norita. M., McHaffie, J.G.. Shimizu. H. and Stein. 13.E. (1991) The corticostriatal and corticotectal projections of :hc feline lateral suprasylrian cortex demonstrated with anterograde biocytin and retrograde fluorescent techniques, Neurosci. Rt:s.. LO: 11%155.
Olavarria. J. and Van Sluyters. R.C. (1985) Organization and postna- tal development of callosal connections in the \.sual coriex of thr. rat. J. Comp. Neural.. 339: l--26.
O‘Leary. D.D.M. and Terashima, T. (19x8) (‘ortlc;d axons ilranch to multiple subcortical targets by interstitial axon budding: implica tions for target recognition and ‘wailing per.t)di’ Neuron. 1. 901-911~.
Payne. B.. Pearson, II. and Cornwell. I”. i 19%) I.~rvelopmenl ol visual and auditory cortical connections in the car. In: :\. Peter> and E.G. Jones (Eds.), Cerebral Cortex. Vol. 7, I.>evcloptnent and Maturation of’ Cerebral Cortex. Plenum Press hcu Z’i)rk. pp 309-3x9.
Rockland. KS. ( 1992) Configuration. m \erlal 1 ccoastruction. 01 individual axons projecting from area VZ IO Vi *F the macaque monkey, C‘ereh. Cortex, 2: 353-374.
Veenman. C.L.. Reiner. A. and Honig. M.G cI’r92) Biotinylared dextran amine as an anterograde tracer ior sirtyir- and double- labeling studies. J. Neurosci. Methods. 41: 239 2%.
Vercelli, A. and Innocenti. G.M. (1993) Morphoiogy ot visual callosai neurons with different locations, contralateral t;trgc:!~ or patterns of development, Exp. Brain Res., 94: 393.-404.
Weber. J. and Amaral, D.(G. (1988) The USC‘ of PXI/t-L for tracing connections in the immature rai brain. Sot. hi:~rr,sci. ,\bst., 14. 549.
Wise. S-P. and Jones, E.G. (1976) The rtrpanizatmn And postnatal development of the commissural projectlon 01’ the rat somatic sensory cortex, J. Comp Neuroi., 16X: 3 I-344
Wouterlood. F.G. and Jorritsma-Byham, B. t 1993) The antarogradc neuroanatomical tracer biotinylated dextran amine comparison with the tracer Phuseolus l,ulgaris-leucoagglutininirl preparation3 for electron microscopy. J. Neurosci. Methods. t? ‘:i .-~1
Related Documents











