Ecological Modelling 152 (2002) 145 – 168 A model study of the coupled biological and physical dynamics in Lake Michigan Changsheng Chen a, *, Rubao Ji d , David J. Schwab b , Dmitry Beletsky b , Gary L. Fahnenstiel c , Mingshun Jiang d , Thomas H. Johengen b , Henry Vanderploeg b , Brian Eadie b , Judith Wells Budd e , Marie H. Bundy f , Wayne Gardner g , James Cotner h , Peter J. Lavrentyev i a School for Marine Science and Technology, Uniersity of Massachusetts -Dartmouth, 706 South Rodney French Bouleard, New Bedford, MA 02744 -1221, USA b NOAA/GLERL, 2205 Commonwealth Bouleard, Ann Arbor, MI 48105 -2945, USA c The Lake Michigan Field Station, NOAA/GLERL, 1431 Beach Street, Muskegon, MI 49441, USA d Department of Marine Sciences, The Uniersity of Georgia, Athens, GA 30602, USA e Department of Geological Engineering and Sciences, Michigan Technological Uniersity, Houghton, MI 49931, USA f Academy of Natural Sciences, Estuarine Research Center, St. Leonard, MD 20005, USA g Department of Marine Sciences, Uniersity of Texas at Austin, Austin, TX 78712, USA h Department of Ecology Eolution and Behaior, Uniersity of Minnesota, St. Paul, MN 55108, USA i Department of Biology, Uniersity of Akron, Akron, OH 44325, USA Received 24 January 2001; received in revised form 22 October 2001; accepted 31 October 2001 Abstract A coupled physical and biological model was developed for Lake Michigan. The physical model was the Princeton ocean model (POM) driven directly by observed winds and net surface heat flux. The biological model was an eight-component, phosphorus-limited, lower trophic level food web model, which included phosphate and silicate for nutrients, diatoms and non-diatoms for dominant phytoplankton species, copepods and protozoa for dominant zooplankton species, bacteria and detritus. Driven by observed meteorological forcings, a 1-D modeling experiment showed a controlling of physical processes on the seasonal variation of biological variables in Lake Michigan: diatoms grew significantly in the subsurface region in early summer as stratification developed and then decayed rapidly in the surface mixed layer when silicate supplied from the deep stratified region was reduced as a result of the formation of the thermocline. The non-diatoms subsequently grew in mid and late summer under a limited-phosphate environment and then declined in the fall and winter as a result of the nutrient consumption in the upper eutrophic layer, limitation of nutrients supplied from the deep region and meteorological cooling and wind mixing. The flux estimates suggested that the microbial loop had a significant contribution in the growth of microzooplankton and hence, to the lower-trophic level food web system. The model results agreed with observations, suggesting that the www.elsevier.com/locate/ecolmodel * Corresponding author. Tel.: +1-508-910-6388; fax: +1-508-910-6371. E-mail address: [email protected] (C. Chen). 0304-3800/02/$ - see front matter © 2002 Published by Elsevier Science B.V. PII: S0304-3800(02)00026-1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Ecological Modelling 152 (2002) 145–168
A model study of the coupled biological and physicaldynamics in Lake Michigan
Changsheng Chen a,*, Rubao Ji d, David J. Schwab b, Dmitry Beletsky b,Gary L. Fahnenstiel c, Mingshun Jiang d, Thomas H. Johengen b,
Henry Vanderploeg b, Brian Eadie b, Judith Wells Budd e, Marie H. Bundy f,Wayne Gardner g, James Cotner h, Peter J. Lavrentyev i
a School for Marine Science and Technology, Uni�ersity of Massachusetts-Dartmouth, 706 South Rodney French Boule�ard,New Bedford, MA 02744-1221, USA
b NOAA/GLERL, 2205 Commonwealth Boule�ard, Ann Arbor, MI 48105-2945, USAc The Lake Michigan Field Station, NOAA/GLERL, 1431 Beach Street, Muskegon, MI 49441, USA
d Department of Marine Sciences, The Uni�ersity of Georgia, Athens, GA 30602, USAe Department of Geological Engineering and Sciences, Michigan Technological Uni�ersity, Houghton, MI 49931, USA
f Academy of Natural Sciences, Estuarine Research Center, St. Leonard, MD 20005, USAg Department of Marine Sciences, Uni�ersity of Texas at Austin, Austin, TX 78712, USA
h Department of Ecology E�olution and Beha�ior, Uni�ersity of Minnesota, St. Paul, MN 55108, USAi Department of Biology, Uni�ersity of Akron, Akron, OH 44325, USA
Received 24 January 2001; received in revised form 22 October 2001; accepted 31 October 2001
Abstract
A coupled physical and biological model was developed for Lake Michigan. The physical model was the Princetonocean model (POM) driven directly by observed winds and net surface heat flux. The biological model was aneight-component, phosphorus-limited, lower trophic level food web model, which included phosphate and silicate fornutrients, diatoms and non-diatoms for dominant phytoplankton species, copepods and protozoa for dominantzooplankton species, bacteria and detritus. Driven by observed meteorological forcings, a 1-D modeling experimentshowed a controlling of physical processes on the seasonal variation of biological variables in Lake Michigan:diatoms grew significantly in the subsurface region in early summer as stratification developed and then decayedrapidly in the surface mixed layer when silicate supplied from the deep stratified region was reduced as a result of theformation of the thermocline. The non-diatoms subsequently grew in mid and late summer under a limited-phosphateenvironment and then declined in the fall and winter as a result of the nutrient consumption in the upper eutrophiclayer, limitation of nutrients supplied from the deep region and meteorological cooling and wind mixing. The fluxestimates suggested that the microbial loop had a significant contribution in the growth of microzooplankton andhence, to the lower-trophic level food web system. The model results agreed with observations, suggesting that the
www.elsevier.com/locate/ecolmodel
* Corresponding author. Tel.: +1-508-910-6388; fax: +1-508-910-6371.E-mail address: [email protected] (C. Chen).
0304-3800/02/$ - see front matter © 2002 Published by Elsevier Science B.V.
PII: S0304-3800(02)00026-1

C. Chen et al. / Ecological Modelling 152 (2002) 145–168146
model was robust to capture the basic seasonal variation of the ecosystem in Lake Michigan. © 2002 Published byElsevier Science B.V.
Keywords: Ecosystem; Coupled biological and physical model; Phytoplankton growth; Microbial processes; Lake Michigan
1. Introduction
Lake Michigan is characterized as a typicalphosphorus-limited lake ecosystem (Eadie et al.,1984; McCormick and Tarapchak, 1984; Schelskeet al., 1986). Previous biological and chemicalobservations showed that the annual averagedconcentrations of total inorganic nitrogen andphosphate in the lake have been increasing dra-matically (Brooks and Edington, 1994). Nitrogen,a requisite component for the growth of phyto-plankton in the coastal ocean, is not a limitingfactor in Lake Michigan (Neilson et al., 1994).The phosphorus in Lake Michigan is supplied andmaintained mainly through four processes: (1)external loading; (2) internal recycling; (3) sedi-ment flux; and (4) nutrient release of suspendedsediments. Recent measurements reveal a net in-crease of the total phosphorus during the springphytoplankton bloom. Of this increment, 20% isaccounted for by the external phosphorus loading(Brooks and Edington, 1994) and only 4% issupplied from the bottom sediment flux (Conleyet al., 1988). Although the previous interpretationof phosphorus loading and flux is somewhat inquestion, there is no doubt that a large portion ofthe net increase of phosphorus is due to thenutrient releases from suspended sediment andinternal recycling (Eadie et al., 1984; Johengen etal., 1994).
Silicon also plays an important role in regulat-ing planktonic communities in Lake Michigan.The collapse of the spring diatom bloom in south-ern Lake Michigan generally occurs in summer assilicon is rapidly depleted in the euphotic zonedue to the restriction of nutrient supplies from thedeep region below the thermocline (Scavia andFahnenstiel, 1987). Unlike phosphorus, the siliconin the water column is hardly released from thedetrital pool and only 21% of it is accounted forby the sediment flux at the bottom (Conley et al.,1988).
In early spring, the planktonic community inLake Michigan is dominated by diatoms. In gen-eral, there are two types of abundant diatoms inthe lake: large net ones (Melosira, Asterionella,etc.) and small centrics (5–10 �m). When thewind mixing-induced thermocline is established, itacts like a barrier to restrict the silicon flux fromthe deep water. As a result, the biomass of di-atoms decreases dramatically in late spring as thesilicon in the photic zone is depleted (Scavia andFahnenstiel, 1987). The non-diatoms, which com-prise of large flagellates (Cryptophytes) and abunch of small flagellates (3–10 �m, Chryso-phytes, Haptophytes and some Prasinophytes,etc.), grow up gradually under conditions of lim-ited phosphorus in late spring through summer(Sherr et al., 1988). Therefore, because of thesilicon limitation, the growth of phytoplankton inLake Michigan features two distinct patterns: thediatom bloom in early spring and then non-di-atoms in late spring through summer.
The major consumers of phytoplankton inLake Michigan are large zooplankton (copepods)and microzooplankton (Ciliates, Daphnia spp. andCalanoida, etc.). A high abundance of copepodsoccurs in early spring, which is consistent with arapid growth of diatoms as stratification develops.The dominant species shift to the microzooplank-ton as diatoms are depleted and non-diatomsgrow up. The microbial food web (MFW) isbelieved to be a major contributor to the abun-dance of microzooplankton in Great Lakes(Gardner et al., 1986; Carrick and Fahnenstiel,1990; Carrick et al., 1991; Valiela, 1995). This isevident from the recent observation that the het-erotrophic bacteria can account for �22% of thetotal plankton biomass and heterotrophic proto-zoa (a major component of the MFW) can ac-count for �32% of the total heterotrophicmicro-organisms in St. Lawrence Great Lakes(Fahnenstiel et al., 1998). Protozoan ciliates con-stituted �30% of the zooplankton biomass in thesouthern part of the lake in a 1-year survey

C. Chen et al. / Ecological Modelling 152 (2002) 145–168 147
conducted from December 1986 to November1987 (Carrick and Fahnenstiel, 1990).
Previous observations and laboratory experi-ments have shown a clear schematic of the lowertrophic level food web of Lake Michigan, whichprovide a foundation for modeling exploration.Based on biological measurements, Scavia (1979,1980) developed a food web model for Lake On-tario. The model was driven by observed biologi-cal distributions with inclusion of a specifiedone-dimensional (1-D) temperature profile andempirically determined vertical diffusion. Thismodel was modified and applied to Lake Michi-gan (Scavia et al., 1988). The updated version ofthe biological food web model consisted of threephytoplankton components (diatoms, flagellatesand blue greens), two nutrients (silicon and phos-phorus) and two crustaceans (Diaptomu andDaphnia). Numerical experiments based on thisbiological model revealed the basic characteristicsof the transformation and fluxes between nutri-ents, phytoplankton and zooplankton in GreatLakes ecosystems. A similar model was also de-veloped for Saginaw Bay, Lake Huron by Biemanand Dolan (1981). All these biological modelswere driven by idealized simple physical fields,which could not resolve the 3-D or even 2-D timeand spatial distributions of biological fields inGreat Lakes.
There has not been a lower trophic level foodweb model available for Lake Michigan until ourpresent modeling studies. Although previous ob-servations have provided us with some insightsinto the structure of the biological community inLake Michigan, the impact of the physical pro-cesses on the seasonal variation of the food websystem has not been well examined. How dostratification and wind mixing control the sea-sonal variation and species regulation of theplankton in Lake Michigan? What is the role ofmicrobial food web in the ecosystem energy bal-ance in Lake Michigan under a realistic physicalcondition? Could we develop a fully coupledphysical and biological model to simulate theseasonal variation of the ecosystem in this lake?
As one component of the modeling projects ofthe Episodic Events Great Lake Experiments (EE-GLE), a fully coupled physical and biological
model was developed for Lake Michigan. Thephysical part was the well-calibrated Lake Michi-gan circulation model that was originally modifiedfrom the Princeton ocean model (POM) by theNOAA Great Lake Laboratory modeling group(Beletsky and Schwab, 1998; Beletsky et al., 2000;Schwab et al., 2000). The biological part consistedof an eight-component, phosphorus-controlled,lower trophic level food web model with inclusionof phosphate and silicate for nutrients, diatomsand non-diatoms for dominant phytoplanktonspecies, copepod and heterotrophic flagellate fordominant zooplankton species, bacteria and de-tritus. This coupled model has been tested andcalibrated under the realistic lake environment of1994–1995 and a spring plume event of 1998.
The objectives of this paper are (1) to describein detail the development of this coupled model;and (2) to use this model to examine the impact ofphysical processes on the seasonal variation of theplankton and the interaction between biologicalvariables in a food web system in this lake. Mod-eling studies were focused on a 1-D numericalsimulation of the plankton at a long-term moni-toring station (shown in Fig. 1) for 1994–1995. A3-D experiment, with focus of the impact of sea-sonally recurring coastal suspended sedimentplume on the spatial distribution of the planktonin southern Lake Michigan, is presented in aseparate paper in this volume (Ji et al., 2002).
The remaining sections are organized as fol-lows. The coupled biological and physical modeland the design of numerical experiments are de-scribed in Section 2. The model results of the 1-Dexperiments are presented in Section 3. The im-pacts of physical and microbial processes on thefood web are examined in Section 4. The sensitiv-ity analysis of biological parameters is given inSection 5 and finally, conclusions are summarizedin Appendix B.
2. The coupled biological and physical model
2.1. Biological model
The biological model is based on the observedfeatures of the lower trophic level food web in

C. Chen et al. / Ecological Modelling 152 (2002) 145–168148
Lake Michigan (Scavia and Fahnenstiel, 1987). Itis a phosphorus-controlled model with eight inde-pendent variables (Fig. 2). The governing equa-tions are given as:
dPL
dt−
�
�z�
Ah
�PL
�z�
=LP(uptake)−LP(mortality)−LZLP(grazing)
−LP(sinking) (1)
dPS
dt−
�
�z�
Ah
�PS
�z�
=SP(uptake)−SP(mortality)−SZSP(grazing)(2)
dZL
dt−
�
�z�
Ah
�ZL
�z�
=�ZL�PLZLP(grazing)+�
ZLSLZSZ(grazing)
−LZ(mortality) (3)
dZS
dt−
�
�z�
Ah
�ZS
�z�
=�ZSSZSP(grazing)−LZSZ(grazing)
+�BSZB(grazing)−SZ(mortality) (4)
dBdt
−�
�z�
Ah
�B�z�
=DB(decomposition)+BP(uptake)
−SZB(grazing)−B(mortality) (5)
dDP
dt−
�
�z�
Ah
�DP
�z�
= (1−�zL)�PLZLP(grazing)
+ (1−�ZS)SZSP(grazing)
+ (1−�B)SZB(grazing)
+ (1−�ZLS)LZSZ(grazing)
−DB(decomposition)−DP(sinking)
−DP(remineralization)+�PLP(mortality)
+SP(mortality)+LZ(mortality)
+SZ(mortality)+B(mortality) (6)
dDS
dt−
�
�z�
Ah
�DP
�z�
=�SLZLP(grazing)+�SLP(mortality)
−DS(remineralization) (7)
dPdt
−�
�z�
Ah
�P�z�
= −�PLP(uptake)−SP(uptake)−BP(uptake)
+DP(remineralization)+PQ (8)
dSi
dt−
�
�z�
Ah
�Si
�z�
= −�SLP(uptake)+DS(remineralization)+SQ(9)
Fig. 1. Bathmetry of Lake Michigan and the location of theGrand Haven monitoring station chosen for the 1-D modelexperiment. The contour of the water depth was plotted withan interval of 50 m.

C. Chen et al. / Ecological Modelling 152 (2002) 145–168 149
Fig. 2. Schematic of the lower trophic level food web model in Lake Michigan. The definition for each symbol is given in the text.
where PL, PS, ZL, ZS, B, P and Si are the largesize phytoplankton, small size phytoplankton,large size zooplankton, small size zooplankton,bacteria, phosphorus, and silicon, respectively. DP
and DS are the phosphate- and silica-related com-ponents of detritus. Ah is the thermal diffusioncoefficient that is calculated using the Mellor andYamada level 2.5 turbulent closure scheme incor-porated in the physical model.
ddt
=�
�t+u
�
�x+�
�
�y+w
�
�z
is the derivative operator; x, y and z are theeastward, northward, and vertical axes of theCartesian coordinate, and u, � and w are the x, y
and z components of the velocity. The definitionof parameters �
ZL, �ZLS, �
ZS, �B, �S and �P aregiven in Table 1. PQ and SQ are the phosphorusand silicon fluxes from suspended sediments. Themathematical formula of LP (uptake), LZLP(grazing), SP (uptake), SZSP (grazing), LZSZ(grazing), SZB (grazing), DB (decomposition), BP(uptake), DP (remineralization), DS (remineral-ization), LP (sinking), DP (sinking), LP (mortal-ity), SP (mortality), LZ (mortality), SZ(mortality) and B (mortality) are given in detail inAppendix A.
In the model, phosphorus and silicon are twolimiting nutrients that control the primary pro-duction. Nitrogen is not included in the model

C. Chen et al. / Ecological Modelling 152 (2002) 145–168150
Table 1Bio-parameters
Ranges SourcesParameter Definition Value used
0.8–6 d−1Maximum growth rate for PL (Bieman and Dolan, 1981; Scavia et1.6 d−1VmaxPL
al., 1988)Vmax
PS 1.2 d−1 0.8–2 d−1 (Bieman and Dolan, 1981; Scavia etMaximum growth rate for PS
al., 1988)0.8–6 d−1 Various sources1.2 d−1Vmax
S Maximum Si uptake rate by PL
0.05 d−1Maximum P uptake rate by B ?VmaxB
VmaxDOP 23–144 d−1Maximum DOP uptake rate by B (Bentzen et al., 1992)5 d−1
0.2 �mol P l−1 0.07–0.4 �mol PHalf-saturation constant for the P (Bieman and Dolan, 1981; Tilman etkP L
l−1uptake by PL al., 1982)0.05 �mol PHalf-saturation constant for the PkP S
0.015–? �mol P (Bieman and Dolan, 1981)l−1l−1uptake by PS
5.0 �mol SiHalf-saturation constant the SikS 3.5–3.57 �mol (Bieman and Dolan, 1981; JorgensenSi l−1 et al., 1991)uptake by PL l−1
kB 0.2 �mol P l−1Half-saturation constant for the P 0.02–0.2 �mol P (Cotner and Wetzel, 1992)uptake by B l−1
0.005–0.02 �mol0.1 �mol P l−1Half-saturation constant for thekDOP (Bentzen et al., 1992)DOP uptake by B P l−1
GmaxZL 0.2–0.86 d−1Maximum PL grazing rate by ZL (Scavia et al., 1988; Jorgensen et al.,0.4 d−1
1991)0.1 d−1Maximum PS grazing rate by ZS (Bieman and Dolan, 1981)0.2 d−1Gmax
ZS
GmaxB 3.5 d−1 3.5 d−1 (Hamilton and Preslon, 1970)Maximum B grazing rate by ZS
0.4 d−1Maximum ZS grazing rate by ZL ?GmaxZLS
0.001–1 l �mol−1 (Scavia et al., 1988; Jorgensen et al.,0.06 l �mol−1kZL Ivlev constant for ZL grazing1991)
0.011 lIvlev constant for Ps grazing by Zs (Bieman and Dolan, 1981)kZS 0.02 l �mol−1
�mol−1
0.03 l �mol−1Ivlev constant for the B grazing by 0.022 l (Hamilton and Preslon, 1970)kB
Zs �mol−1
kZLS Ivlev constant for the Zs grazing by 0.07 ?ZL
Assimilation efficiency of ZL 0.35 0.15–0.5 (Jorgensen et al., 1991)�ZL
?0.3�ZS Assimilation efficiency of Zs
0.3�B ?Assimilation efficiency of B grazingby Zs
0.6 ?Assimilation efficiency of the Zs by�ZLS
ZL
�ZL 0.02 d−1 0.01–0.05 d−1 (Bieman and Dolan, 1981; JorgensenMortality rate of ZL
et al., 1991)Mortality rate of Zs 0.03 d−1 0.1 d−1 (Bieman and Dolan, 1981)�
ZS
0.5–5.9 d−1 (Jorgensen et al., 1991)0.5 d−1�B Mortality rate of B0.08 m−1 0.12–0.17 m−1 (Scavia et al., 1986)k0 Photosynthetic attenuation
coefficient0.02 0.1–0.58Proportionality of DOP from the (Valiela, 1995)�D
detrital P�P L
0.6 m d−1 0.5–9 m d−1 (Scavia et al., 1988; Jorgensen et al.,Sinking velocity of PL
1991)�P S
0.01–3 m d−1Sinking velocity of Ps (Fahnenstiel and Scavia, 1987b)0.3 m d−1
0.5–1 m d−1 (Jorgensen et al., 1991)0.6 m d−1Sinking velocity of D�D
0.15 d−1Remineralization rate of detrital P 0.05 d−1 (Fasham et al., 1990)eP
Remineralization rate of detrital Si 0.03 d−1 ?eS
0.069 (Parsons et al., 1984)0.069� Temperature dependence coefficient35Ratio of carbon (C) to chlorophyll 23–79 (Parsons et al., 1984)�C:Chli80Ratio of C to P ? (Parsons et al., 1984)�C:P

C. Chen et al. / Ecological Modelling 152 (2002) 145–168 151
since it is always in sufficient supply. This foodweb model has the advantage of avoiding thecomplex ammonia dynamics (Fasham et al.,1990).
Two sizes of phytoplankton are considered inthe model. One is diatom (range: 13–312 �m) andthe other is small non-diatom (flagellates with asize smaller than 10 �m). In Lake Michigan, thephytoplankton is dominated by the large net di-atom in early spring and is then followed by thegrowth of phytoflagellates and cyanobacteria orgreen alge (Fahnenstiel and Scavia, 1987b) insummer and autumn. The existence of these twodistinct phase patterns is associated with the sea-sonal variation in silicon levels, which decreasesrapidly as the thermocline develops. Dividing thephytoplankton species into two size groups isbased on the purpose of capturing the basic sea-sonal pattern of the phytoplankton with the sim-plest food web model.
In Lake Michigan, zooplankton is dominatedby copepods and cladocera all year around(Scavia and Fahnenstiel, 1987). The abundantspecies for copepods are Diaptomus spp. and forcladocera is Daphnia galeata mendota. Micro-zooplankton are dominated by ciliates and het-eroflagellates. The ciliates contribute to theheterotrophic plankton and their abundant spe-cies vary significantly with seasons (Carrick andFahnenstiel, 1990), while the heteroflagellates gen-erally account for a small fraction of micro-zooplankton abundance in the lake. Based on thegrazing characteristics of different sizes ofzooplankton, we group the zooplankton into twocategories: large and small zooplankton. The largezooplankton represents copepod and cladoceraand small zooplankton includes microzoo-plankton.
The most preferable food for copepods in LakeMichigan is ciliates other than diatoms (Vander-ploeg et al., 1988; Vanderploeg, 1994). Based onsize and shape selectivity, the freshwater planktonwith a size range of 3–30 �m (the size of mostciliates) is preferred by calanoid copepods (Van-derploeg, 1981) and filter-feeding cladocera (Gli-wicz, 1980). The linkage between copepods andciliates in the lake is taken into account in the
model by adding the grazing process of largezooplankton over small zooplankton. We alsoassume that copepods do not digest the diatomfrustuler and thus, the silica parts of diatoms aredirectly deposited into detrital pool (Scavia et al.1988). In some previous modeling studies for thereservoirs, diatoms are allowed to grow withoutinclusion of zooplankton grazing (Thebault andSalencon, 1993; Salencon and Thebault, 1996).Our experiments suggest that this assumptionprobably fails to simulate the collapse of diatomsin late autumn and caused the earlier occurrenceof the diatoms’ bloom in the subsequent year inLake Michigan.
Bacteria contribute to the lower trophic levelfood web mainly through uptake of nutrients,decomposition of detritus and grazing by micro-zooplankton (Gardner et al. 1986). The mortalityof bacteria directly deposits into the detritus pool,which indirectly influences the detritus remineral-ization to dissolved nutrients and decompositionback to bacteria. Following the procedure ofFasham et al. (1990), the total amount of bacteriais included in the model, with no separation of theattached bacteria from free bacteria.
Detritus refers to fecal pellets, dead phyto-plankton, zooplankton and bacteria. In LakeMichigan, the large egestion of zooplankton andlow assimilation efficiency contribute to theamount of detritus, especially in summer afterstratification develops (Scavia and Fahnenstiel,1987). The sediment flux from the bottom de-creases by a factor of 10 from early spring to latesummer after the thermocline is established (Eadieet al., 1984). This change accounts for part of thealgal losses in Lake Michigan during summer.Since the POC remains almost unchanged in thelake in early spring through summer, the sedimen-tation during that period is dominated by detritallosses. This pattern is evident from previous ob-servations, which show that after the springbloom, the net decrease of total phosphorus isproportional in the epillimnion to sedimentationfluxes that are attributable to detritus (Brooksand Edington, 1994).
In our model, the remineralization, decomposi-tion and sinking of detritus are taken into accountin the food web system. It is difficult to resolve

C. Chen et al. / Ecological Modelling 152 (2002) 145–168152
ratio of silicon to phosphorus from the totalconcentration of detritus, since this ratio varieswith multiple factors related to the mortality ofphytoplankton, zooplankton and bacteria and as-similation rate of zooplankton. For example, thegrowth of large phytoplankton is limited by bothphosphorus and silicon, while only phosphorus isneeded for small phytoplankton (non-diatoms).As a result, the dead diatoms contain both phos-phorus and silicon, but dead small phytoplanktonhas only phosphorus. Similar processes occur forthe grazing of large and small zooplankton, inwhich the unassimilated part of large zooplanktoncontributes to both phosphorus and silicon pools,but that of the small zooplankton contributes tophosphorus only. Because the ratio of phosphorusto silicon in the total detritus depends on multipletime-dependent factors, it is almost impossible toderive a simple mathematical formula to separateone from another if only total concentration ofdetritus is calculated. For this reason, we techni-cally divide detritus into two components: DP andDS, with DP including all phosphorus-related de-tritus and DS all silica-related detritus. This sepa-ration is only valid with an assumption that ratiosof phosphorus to silicon in large phytoplanktonand zooplankton are well-defined.
The effects of upper trophic level predators,such as large fishes, human fishing and birds, arenot included in the present model. As Valiela(1995) suggested, most of these higher trophiclevel effects can be simulated by adding an addi-tional sinking rate of the detrital pool. Since ourinterests are in the physical and biological interac-tion on the lower trophic food web in the lake, weneglect these higher trophic level terms based onan assumption that they have only a secondarycontribution to the food web system in LakeMichigan.
The model includes 37 biological parameters.These parameters are selected according to previ-ous observations and literature values (see Table1). Since these parameters vary over a wide rangein time and space, we first ran the model with aninitial setup of parameters and then carried out asensitivity analysis over the given parameterranges. A detailed description of our initial setupof biological parameters is given in Appendix A.
2.2. Physical model
The physical model used in this study is thePrinceton ocean model (POM) developed origi-nally by Blumberg and Mellor (1987). The modelincorporates the Mellor and Yamada (1982) level2.5 turbulent closure scheme, with a modificationby Galperin et al. (1988) to provide a time- andspace-dependent parameterization of vertical tur-bulent mixing (MY2.5). The estimation of surfacemixing length scale is improved by linking it withmixing intensity and the first model layer thick-ness (Melsom, 1996). This improvement providesa more realistic surface mixing which is criticallyimportant for simulating the thermocline andhence for nutrient supply and phytoplanktongrowth. The POM has been widely used in thecoastal ocean and Great Lake studies.
The POM has been configured for Lake Michi-gan geometry (Beletsky and Schwab, 1998; Belet-sky et al. 2000; Schwab et al., 2000). A uniformgrid with a resolution of 10 km is used in thehorizontal and 21-� levels in the vertical. Themodel is driven directly by the observed meteoro-logical forces, including winds and heat flux. Thewind stress is calculated using the GLERLmethod from Liu and Schwab (1987). The heatforcing included two parts: (1) the surface netheat flux; and (2) the penetrated short-wave irra-diance (Beletsky and Schwab, 1998; Chen et al.,2001; Zhu et al., 2001).
2.3. Boundary conditions
The surface and bottom boundary conditionsfor the momentum equations are given by
��u��
,��
��
�=
D�oAm
(�0x, �0y); =0; and
�T��
=D
�ocPAh
(Q−Io), at �=0 (10)
��u��
,��
��
�=
D�oAm
(�bx, �by); =0; and
�T��
=0, at �= −1 (11)

C. Chen et al. / Ecological Modelling 152 (2002) 145–168 153
where (�ox, �ox) and (�bx, �by)=Cd�u2+�2 (u2+�2) are the x and y components of surface windand bottom stresses. The drag coefficient Cd atthe bottom is determined by matching a logarith-mic bottom layer to the model at a height zab
above the bottom, i.e.
Cd=max�
k2/ln�zab
z0
�2
, 0.0025n
(12)
where k=0.4 is the Karman’s constant and z0 isthe bottom roughness parameter, which is takenas 0.001 m in this study. Am is the vertical eddyviscosity; D, the total water depth; T, the watertemperature; cP, the specific heat of seawater; �o,the reference density; Qn, the net surface heat flux;and Io, the incident irradiance at the sea surface.
The surface and bottom boundary conditionsfor biological variables are given by
�
��(PS, ZL, ZS, B, DS, P, S)=0;
�PL
��=
DAh
wP LPL; and
�DP
��=
DAh
wDDP,
at �=0,−1 (13)
where wP Land wD are the sinking velocities of PL
and DP.
2.4. Design of numerical experiments
Because of the limitation of 3-D interdisci-plinary data in Lake Michigan, we started ournumerical experiments at a 1-D site located at theGrand Haven station (43°00� N and 86°24� W, seeFig. 1). This site is a GLERL/NOAA and EPAecosystem monitoring station with a long-termrecord of both hydrological and biological data.The water depth at this station is �100 m andthe location is considered to be a transitional sitefor sediment accumulation (Cahill, 1981). Thesefacts suggest that little sediment resuspensionwould occur at this station, even during strongplume events. For this reason, we did not includethe sediment term in the numerical experiments.The 3-D physical model experiments have shownthat the horizontal advections and upwelling aregenerally one order of magnitude smaller offshorethan near the coastal region. A 1-D approxima-
tion, as a first step, is a practical starting point totest and calibrate the biological model.
The initial distribution of temperature was spe-cified based on the climatological conditions inwinter of 1994, which were vertically uniformeverywhere. From January 1 to April 1 the watertemperature was held constant at 2 °C in themodel by eliminating surface heat flux for thisperiod. During this period, the lake was slightlystratified with up to 80% ice coverage and coldersurface water temperature (�2 °C) in open waterareas. Since we did not have information aboutthe vertical temperature structure during this pe-riod, we chose to use uniform 2 °C until April 1when the satellite imagery showed surface watertemperature was nearly uniform at 2 °C. At thattime, surface heat flux was restored in the model.This approximation should not have a significanteffect on model results.
The initial distributions of biological variablesalso were specified using the climatological condi-tions in winter of 1994, in which phosphorus: 0.1�mol P l−1; silicon: 27 �mol Si l−1; large phyto-plankton: 0.5 �mol C l−1; small phytoplankton:0.5 �mol C l−1; large zooplankton: 0.7 �mol Cl−1; small zooplankton: 0.5 �mol C l−1; bacteria:3.2 �mol C l−1; phosphorus-related detritus: 6.5�mol C l−1 and silicon-related detritus: 1.0 �molC l−1. Since the water was vertically well mixed inJanuary 1994, the uniform initial values used forbiological variables were a good approximationwith little influence on the seasonal simulationresults. The ratio of C :P used in the model was80, which was the same value used by Parsons etal. (1984).
The coupled model was run using the 3-D codewith five grid points in which all physical andbiological variables were specified uniform in thehorizontal. The numerical integration was con-ducted prognostically over a period of 2 yearsstarting at the beginning of January 1, 1994.
3. Model results
3.1. Physical structures
The 1-D physical model captured the basicpattern of the seasonal variation of temperature

C. Chen et al. / Ecological Modelling 152 (2002) 145–168154
Fig. 3. Time series of the real-time wind speed (upper) andsurface net heat flux (middle) and vertical distribution ofmodel-predicted temperature (lower) in 1994 and 1995 at theGrand Haven monitoring station. A 40-h low-passed filter wasused to treat the wind and heat flux data. The temperatureinterval was 1 °C.
creased considerably in mid-August though earlySeptember and then disappeared in late October.A similar seasonal pattern also was found in 1995,except that the onset of thermal stratificationoccurred �10 days earlier.
The model-predicted temperature is in goodagreement with the observations taken at depthsof 12, 25, 37 and 67 m (Fig. 4). Based on theseagreements, the model results suggest that 1995had a warmer winter and summer compared with1994. The difference in temperature between these2 years was �2 °C in winter and 5 °C in sum-mer. It is not surprising that the model-predictedtemperature curve is much smoother than theobserved temperature curve, since the model isonly 1-D and the model predicted field is
Fig. 4. Comparison between observed (heavy solid) andmodel-predicted (thin-solid) temperatures at selected depths of12, 25, 37 and 67 m.
at Grand Haven station (Fig. 3). In 1994, stratifi-cation developed in early June as a result ofsurface heating. A continuous warming tendencycaused the water in the upper 40 m to be wellstratified in late June. The wind-induced mixedlayer, defined as a layer in which vertical tempera-ture difference is �2.5 °C, formed in early Julyand then gradually deepened with time throughsummer. Cold-front events with strong wind mix-ing and cooling occurred episodically in autumn,which caused a rapid increase of the thickness ofthe mixed layer. Correspondingly, a well-definedthermocline formed at depths of 15–20 m in earlyJune and then deepened gradually to �25 m inlate August. The intensity of the thermocline de-

C. Chen et al. / Ecological Modelling 152 (2002) 145–168 155
Fig. 5. Time sequences of the vertical distributions of phos-phorus, silicon, small phytoplankton and large phytoplanktonat the Grand Haven monitoring station over 1994–1995.
annual variation of stratification, as mentionedabove.
Model-predicted large phytoplankton PL grewrapidly in the upper 40 m in early May as stratifi-cation developed. A maximum value of PL oc-curred near the surface in late May and thenshifted to a depth of 30 m below the surface inlate June. When the thermocline formed, PL in theupper 40 m collapsed rapidly in a short period,starting near the surface and then extended down-ward. Subsequently, small phytoplankton PS grewnear the surface in late May, with remarkablevariation in the mixed layer during summer. Insummer through autumn, the biomass of PL be-low the thermocline remained at a relatively high,decreasing slightly with time. An opposite struc-ture was found in PS, which had a relatively lowbiomass and was vertically well mixed in the deepregion just below the thermocline. Similar pat-
Fig. 6. Time sequences of the vertical distributions of smallzooplankton, large zooplankton, bacteria and total detritus atthe Grand Haven monitoring station over 1994–1995.
smoothed each time step to ensure the numericalstability.
3.2. Seasonal distribution of biological �ariables
The model-predicted seasonal variation of bio-logical variables is closely associated with theseasonal distribution of temperature (Figs. 5 and6). In 1994, both phosphate and silica concentra-tions in the upper 20 m decreased rapidly in earlyMay as stratification developed and then re-mained minimum within the mixed layer duringsummer through autumn as the thermoclineformed. Relatively large values of phosphate andsilica were found below the thermocline in latesummer and then concentrations vertically wellmixed again in winter. This seasonal pattern wasrepeated in 1995, with slight variability due to the

C. Chen et al. / Ecological Modelling 152 (2002) 145–168156
terns of PL and PS appeared in 1995, suggesting aclosely linkage with the seasonal variation ofstratification.
Seasonal distributions of large and smallzooplankton (ZL and ZS) were cohered well withseasonal patterns of PL and bacteria (B). In 1994and 1995, B abundance grew significantly in thesubsurface in late May and June as stratificationdeveloped and remained lower within the mixedlayer in summer through autumn. The abundancebecame vertically uniform in winter as a result ofwind mixing and surface cooling. Similar seasonalpatterns were also found for ZL and ZS, eventhough the maximum biomass of these two vari-ables was about four times smaller.
Total detritus (D) was vertically well mixed inthe winter of 1994. It significantly increased inearly May as stratification developed. Similar toother biological variables, the total D remained ata minimum in the mixed layer in summer throughautumn. Two extreme values of total D werefound over seasons: one was at the depth of 35 min late June and the other near the bottom in lateAugust. Tracing back to two separate compo-nents of detritus, we found that the subsurfacemaximum was dominated by the phosphorus-re-lated component and the bottom extreme detritusmainly came from the silicon-related component.
3.3. Mean biomass and flux
The model-predicted mean biomass of each bio-logical variable in the euphotic zone was esti-mated in the three dynamical phases which weredefined based on the temporal variation of thethermocline. Phase I: a period with a rapid devel-opment of the thermocline; phase II: the durationwith a fully developed thermocline and a near-constant mixed layer depth; and phase III: acollapse period of the thermocline. The meanconcentrations of P and Si were 0.11 �mol P l−1
and 14.76 �mol Si l−1 in phase I, 0.09 �mol P l−1
and 9.38 �mol Si l−1 in phase II and 0.08 �mol Pl−1 and 9.36 �mol Si l−1 in phase III (Fig. 7a,b).The mean biomasses of PL and PS were 6.33 and3.79 �mol C l−1 in phase I, 2.28 and 3.08 �mol Cl−1 in phase II and 0.31 and 3.42 in phase III(Fig. 7c,d). Similarly, the mean biomasses of ZL,
ZS, B and D were 1.41, 1.50, 2.84 and 5.54 �molC l−1 in phase I, 1.52, 1.52, 2.53 and 3.80 �mol Cl−1 in phase II and 1.29, 1.28, 2.27 and 1.91 �molC l−1 in phase III, respectively (Fig. 7e–h). Themodel results suggested significant seasonal varia-tions of large phytoplankton and detritus, but notfor small phytoplankton, small and largezooplankton. The model-predicted bacteria hadits maximum in spring and then gradually andslowly decreased over summer through autumn,which is consistent with recent finding from thedirect measurement (Cotner et al., 2000; Biddandaet al., 2001).
The averaged phosphate-based mean flux ofeach biological process in the euphotic zone forthe three phases is indicated in Fig. 8. The model-predicted mean flux of phosphorus to PS exhibitsa small variation from spring through autumn,with the values of 1.47×10−3 umol P l−1 perday in phase I, 1.22×10−3 umol P l−1 per day inphase II and 1.51×10−3 umol P l−1 per day inphase III. These values are about two or threetimes larger than the sum of the mean fluxes to PL
and B in all three phases, implying that the up-take of the phosphorus by small phytoplanktonwas a major consumer of phosphorus. The meanflux to ZL varied with season, coming comparablyfrom PL and ZS in phase I and being dominatedby ZS in phases II and III. There was a net fluxfrom D to B, which was about eight to nine timeslarger than the flux from phosphorus to B. Mostof the flux from detritus and phosphorus weretaken by ZS and a net flux in the D and P to Band to ZS implied that B increased in phase I,decreased in phase II and then slightly increasedin phase III. The decrease of phosphorus in theeuphotic zone was evident in the net loss ofphosphorus flux after compensation by the rem-ineralization process from detritus, though themodel showed that the total P in the watercolumn remained constant.
Linking all these fluxes together suggest thatafter the thermocline was fully developed the sec-ondary production could be attributed mainly tothe detritus–bacteria–small zooplankton– largezooplankton loop, with little influences from theprimary production of phytoplankton. Thismeans that in summer and fall the zooplankton

C. Chen et al. / Ecological Modelling 152 (2002) 145–168 157
Fig. 7. The averaged biomasses of phosphorus, silicon, large phytoplankton and small phytoplankton, large zooplankton, smallzooplankton, bacteria and total detritus in the euphotic layer during phases I–III.
processes could be separated from phytoplanktonprocesses in southern Lake Michigan as a firstorder approximation. This model result is consis-tent with the suggestion from previous observa-tions (Carrick et al., 1991), which showed that alarger carbon flux from heterotrophic bacteria toprotozoans and copepods gained more carbon
from ciliates than from diatoms. Our modelingexperiments agree that the microbial food webplays a critical role in the ecosystem in LakeMichigan. These results are also consistent withrecent measurements taken in the springtimeplume in southern Lake Michigan (Cotner et al.,2000).

C. Chen et al. / Ecological Modelling 152 (2002) 145–168158
3.4. Comparison with obser�ations
The model-predicted vertical and seasonal dis-tributions of nutrients and phytoplankton agreewith the field measurement taken in 1994–1995 atthe Grand Haven station (Fig. 9). Silicon wasvertically uniform in winter, rapidly decreased inthe mixed layer in summer and decreased in thedeep region in late summer and fall and thenmixed vertically again in late fall and subsequentwinter. These features are captured in our 1-Dmodeling experiment. The model predicts a sharpvertical gradient of silicon in the thermocline area,which was unresolved in the observational databecause of a coarse sampling resolution in thevertical. The observations also show that chloro-
phyll-a was uniform vertically in winter, grewsignificantly and reached a maximum at the sub-surface in early June and was then depleted insummer. These vertical and seasonal features alsoare evident in the model-predicted total phyto-plankton biomass.
The model-predicted, depth-averaged siliconand chlorophyll concentrations coincide well withthe seasonal distribution of silicon and chloro-phyll-a (Fig. 10). The model results show thatdepth-averaged concentration of silicon reached aminimum level of 16.5 umol Si l−1 in late June forboth 1994 and 1995 and a maximum level of 25umol Si l−1 in January, which matched the sea-sonal distribution of observed silicon over 2 years.The model-predicted, depth-averaged chlorophyll
Fig. 8. The averaged fluxes of each biological variables within the lower trophic level food web system in the euphotic layer duringphases I–III. The unit is 10−3 �mol P l−1 per day.

C. Chen et al. / Ecological Modelling 152 (2002) 145–168 159
Fig. 9. Comparison between the vertical distributions of model-predicted and observed silicon and phytoplankton at the GrandHaven station over 1994–1995.
reached a maximum level of 3.9 ug l−1 in mid-June and a minimum level of 1.4 ug l−1 inearly November in 1994 and then again a maxi-mum level of 3.7 ug l−1 in mid-June in thesubsequent 1995. This model-predicted seasonaldistribution of phytoplankton matches the ob-served chlorophyll-a concentrations, especially in1995.
The model results also are consistent withprevious measurements taken in southern LakeMichigan. Chlorophyll-a concentration at a sta-
tion 12.9 km offshore the Grand River was �3.5 ug l−1 during summer and 2.4 ug l−1 inautumn (Moll and Brache, 1986). The depth-av-eraged chlorophyll-a in 1986–1989 varied ina range of 0.5–2.5 ug l−1 from winter to sum-mer (Brooks and Edington, 1994). Taking theannual variation of stratification into account,the model-predicted silicon and phytoplanktonrepresent the general seasonal pattern of nutri-ents and phytoplankton in southern LakeMichigan.

C. Chen et al. / Ecological Modelling 152 (2002) 145–168160
A deep chlorophyll layer (DCL) which devel-oped after the onset of seasonal stratificationwas discovered in 1984–1985 (Fahnenstiel andScavia, 1987a). The DCL initially occurred in15–30 m below the surface, deepened to 25–50
m in July and then to 40–70 m in August.About 50% of primary production in summeroccurred below the epillimnion. Our 1-D modelclearly show that the maximum layer of phyto-plankton began to develop as stratification de-veloped. It was near the surface at thebeginning, gradually deepened to 30 m in lateJune and then to 45 m in late August.
A sensitivity study was conducted to test themodel reliability to uncertainties in bio-parame-ters. For a given light attenuation, the measureindices of sensitivity for all bio-parameters were�0.5 regarding the influence on the meanbiomass. This suggests that the model-predictedseasonal pattern of biological variables were ro-bust. A detailed discussion of the sensitivityanalysis is given in Appendix B.
4. Discussion
Our model reveals that the large phytoplank-ton in the euphotic layer grew rapidly in springand early summer and was then depletedthrough summer through autumn. This seasonalpattern was associated with the formation andcollapse of the seasonal thermocline. The thick-ness of the well-defined mixed layer was gener-ally smaller than the euphotic layer in southernLake Michigan (Fig. 11). This suggests the exis-tence of a growth favorable layer (GFL) be-tween the bottom of the mixed layer and thelower base of the euphotic layer. Based on thefield measurements taken in southern LakeMichigan, Fahnenstiel and Scavia (1987a) sug-gested that the GFL worked as an area of nu-trients, which directly supported the occurrenceof deep chlorophyll-a layer (DCL) in the ther-mocline. To examine the relative importance ofbiological and physical processes in the seasonalvariation of large phytoplankton in southernLake Michigan, we estimated the total flux ofeach term in large phytoplankton and siliconequations in the euphotic layer and deep regionover spring through autumn of 1994. In a 1-Dcase, for example, Eq. (1) and Eq. (9) in theeuphotic layer can be rewritten as integratedforms as follows
Fig. 10. Comparisons between the model-predicted and ob-served depth-averaged silicon (a) and phytoplankton (b) over1994–1995.
Fig. 11. The temporal distribution of the surface mixed layer,growth favorable layer and euphotic layer over phase I–III.

C. Chen et al. / Ecological Modelling 152 (2002) 145–168 161
PL�t−PL�t o
=� t
t o
� 0
−hE
LP(uptake)dzdt �
−� t
t o
� 0
−hE
LP(mortality)dzdt �
−� t
t o
� 0
−hE
LZLP(grazing)dzdt �− (WsPL)�−hE
−Ah
�PL
�z�−hE
(14)
Si �t−Si �t o= −�S
� t
t o
� 0
−hE
LP(uptake)dzdt �
+� t
t o
� 0
−hE
DS (remineralization)dzdt �
−Ah
�Si
�z�−hE
(15)
Similarly, we can derive the integrated forms of PL
and Si in the deep layer with an upper boundaryat the base of the euphotic layer as
PL�t−PL�t o
=� t
t o
� −hE
−H
LP(uptake)dzdt �
−� t
t o
� −hE
−H
LP(mortality)dzdt �
−� t
t o
� −hE
−H
LZLP(grazing)dzdt �+ (WsPL)�−hE
+Ah
�PL
�z�−hE
(16)
Si �t−Si �t o= −�S
� t
t o
� −hE
−H
LP(uptake)dzdt �
+� t
t o
� −hE
−H
DS(remineralization)dzdt �
+Ah
�Si
�z�−hE
(17)
In the euphotic layer, in phase I, PL grew rapidlyin May through mid-June at the onset of thermalstratification and then depleted quickly in late Juneand July as the thermocline strengthened anddeepened. Although the growth rate of PL in-creased significantly in this phase, it was smallerthan the total loss caused by phytoplankton mor-
tality, sinking and zooplankton grazing after lateJune. The temporal variation of PL with time wasmainly dominated by internal biological processeswith very little contribution from vertical diffusion(Figs. 12(a) and 13). PL continued to decrease inphase II due to a net loss through phytoplanktonmortality, sinking and zooplankton grazing againsta small growth. In phase III, the change of PL wasslow down and vertical diffusion became a firstorder contributor. The rapid decrease of PL in theeuphotic layer in the late past of phase I and entirephase II was related to a rapid decrease of Si inphase I and limited abundance in phase II. In phaseI, phytoplankton uptake was much larger thannutrient regeneration from detritus remineraliza-tion and supply rates from the deep layer throughvertical diffusion and was the dominant sink for Si.In phases II and III, Si remained consistently lowbecause the uptake rate of nutrient, remineraliza-tion and vertical diffusion were all small. Unlikephytoplankton, vertical diffusion played the samerole as remineralization in the temporal variationof nutrients in the euphotic layer.
In the deep layer, PL decreased gradually inphases II and III after the thermocline developed.This phenomenon was mainly caused by a net lossthrough phytoplankton mortality and zooplanktongrazing via phytoplankton sinking and verticaldiffusion. A relatively high nutrient pool formed inthe deep region in phases II and III was caused byparticle sedimentation and subsequent nutrientremineralization. The existence of a large detrituspool in the deep layer resulted from materials leftafter zooplankton grazing and death of large phy-toplankton.
The integrated structure of PL and Si in theeuphotic zone also represented the general fea-tures of PL and Si in the favorable growth layer(FGL) since the flux through the mixed layer tothe FGL was much smaller. In our model experi-ment, the formation of DCL in the thermoclineduring early summer was mainly caused by arapid increase of the phytoplankton uptake andphytoplankton sinking and it disappeared in latesummer due to the suppression of silicon suppliesfrom the deep region as the thermocline devel-oped. The physical diffusion process was impor-tant for the nutrient supply from the deep regionto the euphotic layer, but it could be ignored in

C. Chen et al. / Ecological Modelling 152 (2002) 145–168162
Fig. 12. Time series of each term in Eq. (14) and Eq. (15) over phases I–III for the euphotic layer.
the temporal variation of phytoplankton as a firstorder approximation.
Our 1-D model suggested that the microbialfood web might play an important role in thelower trophic food web system in southern LakeMichigan. A much larger flux from D to B thanfrom P to B and from B to ZS than from PS
to ZS, suggested a detritus–bacteria–micro-zooplankton loop that was decoupled from phy-toplankton and nutrients. Also, there was arelatively large grazing rate of ZS by ZL in phasesII and III after the thermocline developed, imply-ing that zooplankton dynamics might be decou-pled from phytoplankton and nutrients in summerand autumn in southern Lake Michigan. Thesemodeling findings are qualitatively consistent with
previous observations in Lake Michigan (Carricket al., 1991) and recent EEGLE field measure-ments taken in southern Lake Michigan. We un-derstand that the reliability of these suggestionsdepends on the choice of biological parameters.Sensitivity analysis conducted in Appendix Bshowed that changing biological parameters hadno significant impact on the biomass of eachbiological variable, and secondary production wasnot sensitive to the maximum nutrient uptakerates of large phytoplankton and bacteria, whichto a certain extent supports a decoupled food websystem between nutrients–phytoplankton andbacteria–micozooplankton–zooplankton. In ad-dition, a good agreement between model-pre-dicted and observed nutrients and phytoplankton
C. Chen et al. / Ecological Modelling 152 (2002) 145–168 163
distributions over 1994–1995, indirectly impliedthat those parameters chosen in the model wasreasonable.
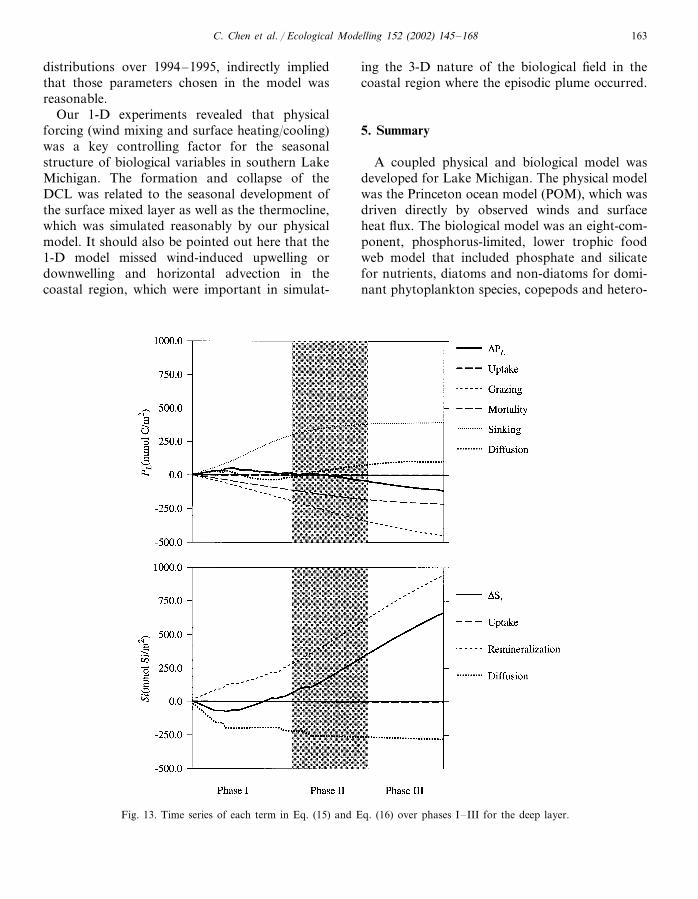
Our 1-D experiments revealed that physicalforcing (wind mixing and surface heating/cooling)was a key controlling factor for the seasonalstructure of biological variables in southern LakeMichigan. The formation and collapse of theDCL was related to the seasonal development ofthe surface mixed layer as well as the thermocline,which was simulated reasonably by our physicalmodel. It should also be pointed out here that the1-D model missed wind-induced upwelling ordownwelling and horizontal advection in thecoastal region, which were important in simulat-
ing the 3-D nature of the biological field in thecoastal region where the episodic plume occurred.
5. Summary
A coupled physical and biological model wasdeveloped for Lake Michigan. The physical modelwas the Princeton ocean model (POM), which wasdriven directly by observed winds and surfaceheat flux. The biological model was an eight-com-ponent, phosphorus-limited, lower trophic foodweb model that included phosphate and silicatefor nutrients, diatoms and non-diatoms for domi-nant phytoplankton species, copepods and hetero-
Fig. 13. Time series of each term in Eq. (15) and Eq. (16) over phases I–III for the deep layer.

C. Chen et al. / Ecological Modelling 152 (2002) 145–168164
trophic flagellates for dominant zooplankton spe-cies, bacteria and detritus.
Driven by observed meteorological forcings, a1-D modeling experiment was conducted at theGrand Haven monitoring station for a 2-yearperiod from 1994 to 1995. The model showed thatthe large phytoplankton (diatoms) significantlygrew in the subsurface region in early summer asstratification developed and then decayed rapidlyin the mixed layer with the reduction of silicatesupplied from the deep stratified region as a resultof the formation of the thermocline. The smallphytoplankton (non-diatoms) subsequently grewin mid and late summer under a limited-phos-phate environment and then reduced rapidly inthe fall and winter as results of the nutrientconsumption in the upper eutrophic layer, limita-tion of nutrients supplied from the deep region,and meteorological cooling and wind mixing. Theflux estimates suggested that the microbial loophad a significant contribution to the growth ofmicrozooplankton. The food web system might bedivided into two decoupled loops: (1) detritus–bacteria–microzooplankton– large zooplankton;and (2) nutrient–phytoplankton–detritus.
Sensitivity analysis suggested that changing thebiological parameters had no significant impactson the biomass of each biological parameter. Themost sensitive parameters were Vmax
PL and kP Lfor
primary production, and VmaxDOP, kDOP and �B for
secondary production. These parameters must beestimated accurately from the field measurementsor laboratory experiments in order to make themodel robust and applicable to Lake Michigan.
Acknowledgements
This research was supported by the NSF/NOAA EGGLE Program under the NSF grantnumber OCE-9712869 for Changsheng Chen,OCE-9712872 for J.W. Budd, NOAA CoastalOcean Program grant for D.J. Schwab, D. Belet-sky, G.L. Fahnenstiel, J. Cotner, T.H. Johengen,B. Eadie, H. Vanderploeg, W. Gardner and P.J.Lavrentyev and under the NSF grant numberOCE-9730416 for M.H. Bundy. R. Ji and M.Jiang are supported by Chen’s NSF grant.
Appendix A. The biological model
The mathematical expression for each term inthe biological model shown in Eqs. (1)– (9) aregiven below:
LP(uptake)
=min�
VmaxPL
PkP L
+P, Vmax
S SkS+S
�f(I)PL (A1)
LZLP(grazing)=GmaxZL (1−e−kPLPL)ZL (A2)
SP(uptake)=VmaxPs
PkP L
+Pf(I)Ps (A3)
SZSP(grazing)=GmaxZs (1−e−kPsPs)Zs (A4)
LZSZ(grazing)=GmaxZLS(1−e−kZLSZs)ZL (A5)
SZB(grazing)=GmaxB (1−e−kBB)Zs (A6)
BP(uptake)=VmaxB P
kB+PB (A7)
DB(decomposition)=VmaxDOP �DD
kDOP+�DD(A8)
SZB(grazing)=GmaxB (1−e−kBB)Zs (A9)
DP(remineralization)=ePDP (A10)
DS(remineralization)=esDs (A11)
LP(sinking)=wP L
�PL
�z(A12)
LP(mortality) =�PLPL
2 (A13)
SP(mortality)=�PsP s
2 (A14)
LZ(mortality)=�ZLZL
2 (A15)
SZ(mortality)=�ZsZ s
2 (A16)
B(mortality)=�BB2 (A17)
where the definition for each parameter used inthe above equations is given in Table 1. �P and �s
are the phosphorus and silica fractions of largephytoplankton (diatom) contained in the totalamount of unassimilated zooplankton grazing, re-spectively and �P+�s=1. The value of �P wasmade according to the observed ratios of carbonto phosphorus in general plants and diatom andthen �s was directly derived by 1−�P.

C. Chen et al. / Ecological Modelling 152 (2002) 145–168 165
The dependence of the phytoplankton growthrate on incident irradiance intensity f(I) is givenas
f(I)=e−k0z (A18)
where k0 is the diffuse attenuation coefficient. Eq.(A18) was normalized using the surface incidentirradiance intensity. In general, the response ofphytoplankton to light intensity varies accordingto different species. For many phytoplankton spe-cies, the photosynthesis reaches its saturation levelat a certain level of light intensity and is theninhibited as light intensity continues to bestronger. The linear assumption of ln f(I), usedwidely in previous lower trophic level food webmodels, does not consider saturation and inhibi-tion of photosynthesis via light. It was found tobe a good approximation in a mixed region(Franks and Chen, 1996; Chen et al. 1997, 1999),but it should be aware of its limitation to repro-duce the vertical profile of primary productionwhich normally exhibits a maximum value atsubsurface.
Instead of using a constant mortality rate, weconcurred in assuming that the organism mortal-ity was proportional to its biomass. A sensitivityanalysis of biological model without inclusion ofphysical forcing has revealed that this mortalityrate was robust to capture a conservative biologi-cal system under a condition with no extrasources and sinks.
Bacteria assimilation for dissolved organicphosphorus (DOP) was also assumed to followthe Michaelis–Menten function, in which DOPwas proportional to the total detritus. This as-sumption was similar to make the bacteria grazedetritus directly with a half-saturation constant ofkDOP/�D. A large portion of bacterial ingestion,which may be excreted into particular organicpool, was taken into account in our numericalexperiments by assuming a larger mortality rate ofbacteria (Cotner and Wetzel, 1992).
Values of biological parameters used in ournumerical experiments were listed in Table 1.These values were obtained from previous fieldmeasurements taken in Lake Michigan and theliterature. Since biological parameters varied in awide range with time and space, we first ran the
model with an initial setup of parameters andthen carried out a sensitivity analysis of parameterranges. A detailed description of this sensitivityanalysis is given in Appendix B.
Appendix B. Sensitivity analysis
The most difficult issue in the development of abiological model is to determine the bio-parame-ters, since they vary in a wide range with time andspace for different species. To qualify the model-predicted biological fields, a series of sensitivityanalysis was conducted to test the model reliabil-ity to uncertainties in bio-parameters. The objec-tive of this analysis, at first, was to find the mostsensitive parameters for primary production (PP),secondary production (SP), and mean biomass,and then to evaluate if the model-predicted sea-sonal patterns of biological variables were robust.
The sensitivity of bio-parameters in our 1-Dexperiments was estimated by
S� = � �F/F�Parameter/Parameter
�(A19)
where S� is a measure index of sensitivity, F is theconcentration of a biological variable in themodel run with a standard set of biologicalparameters and �F is the change of F caused byvarying the model parameter. �Parameter isvaried by 1% from the standard value. Thismethod was the exact same as that used in Franksand Chen (1996), Fasham et al. (1990) and Chenet al. (1999). According to the definition used inthose previous studies, one parameter is deter-mined to be sensitive as its sensitivity index S� isequal to or larger than 0.5.
The resulting sensitivity indices for all bio-parameters are listed in Table 2. For a given lightattenuation k0, S� for all bio-parameters were �0.5 regarding the influence on the mean biomass.This result suggests that the model-predicted sea-sonal patterns of biological variables were robust.As long as the primary production was concerned,the most sensitive bio-parameters were Vmax
PL andkP L
. This conclusion implied that the model mightnot provide a robust quantitative result of pri-mary production due to the photosynthesis of

C. Chen et al. / Ecological Modelling 152 (2002) 145–168166
Table 2Sensitivity indices for bio-parameters
Testing value PercentBio-parameter PrimaryStandard value Secondary Mean biomassproductionproduction
1.3VmaxPL 0.0831.2 0.84 0.16 0.42
0.8 0.143 0.09VmaxPS 0.840.7 0.06
1.3 0 0.041.2 0.04VmaxS 0
0.05VmaxB 0.06 2 0.11 0.15 0
5VmaxDOP 6 0.2 0.46 1.84 0.16
0.15 0.25 0.510.2 0.03kP L0.22
0.05kP S0.06 0.2 0.01 0.29 06 0.2 0.025 0.02kS 0
0.2kB 0.25 0.25 0.04 0.09 00.1kDOP 0.12 0.2 0.31 1.07 0.08
0.35 0.125 0.380.4 0.18GmaxZL 0.27
0.2GmaxZS 0.25 0.25 0.02 0.04 0.01
3 0.143 0.203.5 0.50GmaxB 0.02
0.35 0.125 0.03GmaxLS 0.310.4 0.08
0.05 0.167 0.330.06 0.10kZL 0.210.025 0.25 0.02 0.04 0.01kZS 0.020.02 0.333 0.230.03 0.48kB 0.02
0.07kLS 0.06 0.143 0.03 0.29 0.080.35�
ZL 0.3 0.143 0.03 0.06 0.030.35 0.167 0.030.3 0.04�
ZS 00.35 0.167 0.25�B 0.350.3 00.5 0.167 0.150.6 0.13�LS 0.08
0.003�PL 0.0025 0.167 0.11 0.08 0.10
0.02�PS 0.015 0.25 0.04 0.12 0.08
0.025 0.25 0.180.02 0.13�ZL 0.09
0.035�ZS 0.1670.03 0.41 0.01 0.14
0.6 0.2 0.390.5 0.58�B 0.100.2�P S
0.3 0.5 0.05 0.19 0.04�P L
0.70.6 0.167 0.16 0.05 0.150.7 0.167 0.13 0.400.6 0.09�D
large phytoplankton since the model-predictedvalues were sensitive to the maximum growth rateand half-saturation constant in the uptake of nu-trients by large phytoplankton. A similar analysiswas also made for the secondary production,which showed three sensitive bio-parameters:Vmax
DOP, kDOP and �B. These results indicated that inour 1-D model, the variation of secondary pro-duction was dominantly controlled by the maxi-mum growth rate of bacteria by taking DOP inthe detritus pool and its half-saturation constantvia bacteria mortality rate. Since the flux to mi-crozooplankton was one order of magnitudelarger from bacteria than from small phytoplank-ton when a standard set of bio-parameters were
used and also mean biomass was not affectedsignificantly with these bio-parameters, the impor-tance of microbial food web in the ecosystem ofsouthern Michigan Lake was qualitativelymeaningful.
In summary, we conclude that the seasonalvariation pattern of phytoplankton and nutrientspredicted by our 1-D model is robust. The moreaccurate estimation for the maximum growth rateand half-saturation constant for diatoms andmaximum DOP uptake rate, half-saturation con-stant for the DOP uptake and mortality rate forbacteria must be made in order to provide moreaccurate simulation of primary and secondaryproductions.

C. Chen et al. / Ecological Modelling 152 (2002) 145–168 167
References
Beletsky, D., Schwab, D.J., 1998. Modeling thermal structureand circulation in Lake Michigan. Proceedings of theEstuarine and Coastal Modeling Fifth International Con-ference, October 22–24, 1997, Alexandria, VA, 511–522.
Beletsky, D., Schwab, D.J., McCormick, M.J., Miller, G.S.,Saylor, J.H., Roebber, P.J., 2000. Hydrodynamic modelingfor the 1998 Lake Michigan coastal turbidity plume event.Proceedings of the Estuarine and Coastal Modeling Con-ference, American Society of Civil Engineers, November3–5, 1999, New Orleans, LA, 597–613.
Bentzen, E., Taylor, W.D., Millard, E.S., 1992. The impor-tance of dissolved organic phosphorus to phosphorus up-take by limnetic plankton. Limnol. Oceanogr. 37, 217–231.
Biddanda, B., Ogdahl, M., Cotner, J.B., 2001. Variable contri-bution of heterotrophic bacteria to planktonic biomass andrespiration in natural waters: relationship to system pro-ductivity. Limnol. Oceanogr. (In press).
Bieman, V.J., Dolan, D.M., 1981. Modeling of phytoplank-ton-nutrient dynamics in Saginaw bay, Lake Huron. J.Great Lake Res. 7 (4), 409–439.
Blumberg, A.F., Mellor, G.L., 1987. A description of a three-dimensional coastal ocean circulation model. Coast. Es-tuar. Sci. 4, 1–6 In: Heaps, N.S. (Ed.), Three-DimensionalCoastal Ocean Model.
Brooks, A.S., Edington, D.N., 1994. Biogeochemical controlof phosphate cycling and primary production in LakeMichigan. Limnol. Oceanogr. 39 (4), 961–968.
Cahill, R.A., 1981. Geochemistry of recent Lake Michigansediments. III. Geol. Survey, Champaign, IL, Circ. 517,94p.
Carrick, H.J., Fahnenstiel, G.L., 1990. Planktonic protozoanin Lake Huron and Michigan: seasonal abundance andcomposition of ciliates and dinoflagellates. J. Great LakeRes. 16, 319–329.
Carrick, H.J., Fahnenstiel, G.L., Stoermer, F.E., Wetzel,R.G., 1991. The importance of zooplankton–protozoantrophic couplings in Lake Michigan. Limnol. Oceanogr.36, 1335–1345.
Chen, C., Wiesenburg, L.D.A., Xie, L., 1997. Influences ofriver discharge on biological production in the inner shelf:a coupled biological and physical model of the Louisiana–Texas shelf. J. Mar. Res. 55, 293–320.
Chen, C., Ji, R., Zheng, L., Zhu, M., Rawson, M., 1999.Influences of physical processes on the ecosystem inJiaozhou Bay: a coupled physical and biological modelexperiment. J. Geophys. Res. 104, 925–929.
Chen, C., Zhu, J., Ralph, E., Green, S.A., Budd, J.W., 2001.Prognostic modeling studies of the Keweenaw current inLake Superior. Part I: formation and evolution. J. Phys.Oceanogr. 31, 379–395.
Conley, D.J., Quigley, M.A., Schelske, C.L., 1988. Silica andphosphorus flux from sediments: importance of internalrecycling in Lake Michigan. Can. J. Fish. Aquat. Sci. 45,1030–1035.
Cotner, J.B., Wetzel, R.G., 1992. Uptake of dissolved inor-ganic and organic phosphorus compounds by phytoplank-ton and bacterioplankton. Limnol. Oceanogr. 37, 232–243.
Cotner, J.B., Johengen, T.H., Biddanda, B.A., 2000. Intensewinter heterotrophic production stimulated by benthic re-suspension. Limnol. Oceanogr. 45, 1672–1676.
Eadie, B.J., Chambers, R.L., Gardner, W.S., Bell, G.W., 1984.Sediment trap studies in Lake Michigan: resuspension andchemical fluxes in the southern basis. J. Great Lake Res.10, 307–321.
Fahnenstiel, G.L., Scavia, D., 1987a. Dynamics of Lake Mich-igan phytoplankton: primary production and growth. Can.J. Fish. Aquat. Sci. 44, 499–508.
Fahnenstiel, G.L., Scavia, D., 1987b. Dynamics of Lake Mich-igan phytoplankton: recent changes in surface and deepcommunities. Can. J. Fish. Aquat. Sci. 44, 509–514.
Fahnenstiel, G.L., Krause, A.E., McCormick, M.J., Carrick,H.J., Schelske, C.L., 1998. The structure of the planktonicfood-web in the St. Lawrence Great Lakes. J. Great LakeRes. 24 (3), 531–554.
Fasham, M.J.R., Duklow, H.W., Mckelvie, S.M., 1990. Anitrogen-based model of plankton dynamics in the oceanicmixed layer. J. Mar. Res. 48, 591–639.
Franks, P.J.S., Chen, C., 1996. Plankton production in tidalfronts: a model of Georges Bank in summer. J. Mar. Res.54, 631–651.
Galperin, B., Kantha, L.H., Hassid, S., Rosati, A., 1988. Aquasi-equilibrium turbulent energy model for geophysicalflows. J. Atmos. Sci. 45, 55–62.
Gardner, W.S., Chandler, J.F., Laird, G.A., Scavia, D., 1986.Microbial response to amino acid additions in Lake Mich-igan: grazer control and substrate limitation of bacterialpopulations. J. Great Lakes Res. 12 (3), 161–174.
Gliwicz, Z.M., 1980. Filtering rates, food size selection andfeeding rates in cladocerans: another aspect of interspecificcompetition in filter-feeding zooplankton. Am. Soc. Lim-nol. Oceanogr. Spec. Symp. 3, 282–291.
Hamilton, R.D., Preslon, J.E., 1970. Observations on thecontinuous culture of a planktonic phagotrophic proto-zoan. J. Exp. Mar. Biol. 5, 94–104.
Ji, R., Chen, C., Schwab, D.J., Beletsky, B., Fahnenstiel, G.L.,Johengen, T.H., Vanderploeg, H., Eadie, B., Bundy, M.,Gardner, W., Cotner, J., 2002. Influences of suspendedsediments on the ecosystem in Lake Michigan: a 3-Dcoupled bio-physical modeling experiment. Ecol. Model.152(2-3), 169–190.
Johengen, T.H., Johannsson, O.E., Pernie, G.L., Millard, E.S.,1994. Temporal and seasonal trends in nutrient dynamicsand biomass measures in Lake Michigan and Ontario inresponse to phosphorus control. Can. J. Fish. Aquat. Sci.51, 2570–2578.
Jorgensen, S.E., Nielsen, S.N., Jorge, L.A., 1991. Handbookof Ecological Parameters and Ecotoxicology. Elsevier,Amsterdam.
Liu, P.C., Schwab, D.J., 1987. A comparison of methods forestimating U* from given Uz and air–sea temperaturedifferences. J. Geophys. Res. 92 (C6), 6488–6494.

C. Chen et al. / Ecological Modelling 152 (2002) 145–168168
McCormick, M.J., Tarapchak, S.J., 1984. Uncertainty analysisof calculated nutrient regeneration rates in Lake Michigan.Can. J. Fish. Aquat. Sci. 41, 206–211.
Mellor, G.L., Yamada, T., 1982. Development of a turbulentclosure model for geophysical fluid problem. Rev.Geophys. Space Phys. 20, 851–875.
Melsom, A., 1996. Realistic near-surface momentum mixing inthe Princeton Ocean Model. The Norwegian Meteorologi-cal Institute Research Report, No. 41, 25pp.
Moll, R., Brache, M., 1986. Seasonal and spatial distributionof bacteria, chlorophyll, and nutrients in nearshore LakeMichigan. J. Great Lake Res. 12 (1), 52–62.
Neilson, M., L’Italien, S., Glumac, V., Williams, D., Bertram,P., 1994. Nutrients: trends and ecosystem response. Reportfrom the State of the Lakes Ecosystem Conference. Envi-ronment Canada, Ottawa, Ont.
Parsons, T.R., Takahashi, M., Hargrave, B., 1984. BiologicalOceanographic Processes, third ed. Pergamon Press, NewYork.
Salencon, M.J., Thebault, J.M., 1996. Simulation model of amesotrophic reservoir (Lac de Pareloup, France): MELO-DIA, an ecosystem reservoir management model. Ecol.Model. 84, 163–187.
Scavia, D., 1979. Examination of phosphorus cycling andcontrol of phytoplankton dynamics in Lake Ontario withan ecological model. J. Fish. Res. Board Can. 36, 1336–1346.
Scavia, D., 1980. An ecological model of Lake Ontario. Ecol.Model. 8, 49–78.
Scavia, D., Fahnenstiel, G.L., 1987. Dynamics of Lake Michi-gan phytoplankton: mechanisms controlling epilimneticcommunities. J. Great Lake Res. 13 (2), 103–120.
Scavia, D., Fahnenstiel, G.L., Evans, M.S., Jude, D.L., Leh-man, J.T., 1986. Influence of salmonine predation andweather on long-term water quality trends in Lake Michi-gan. Can. J. Fish. Aquat. Sci. 43, 435–443.
Scavia, D., Lang, G.A., Kitchell, J.F., 1988. Dynamics ofLake Michigan plankton: a model evaluation of nutrientloading, competition, and predation. Can. J. Fish. Aquat.Sci. 45, 165–177.
Schelske, C.L., Stoermer, E.F., Fahnenstiel, G.L., Halbach,M., 1986. Phosphorus enrichment, silica utilization, andbiogeochemical silica depletion in the Great Lakes. Can. J.Fish. Aquat. Sci. 43, 407–415.
Schwab, D.J., Beletsky, D., Lou, J., 2000. The 1998 coastalturbidity plume in Lake Michigan. Estuar. Coast. ShelfSci. 50, 49–58.
Sherr, B.F., Sherr, E.B., Hopkinson, C.S., 1988. Trophic inter-actions within pelagic microbial communities: indicationsof feedback regulation of carbon flow. Hydrobiologia 159,19–26.
Thebault, J.M., Salencon, M.J., 1993. Simulation model of amesotrophic reservoir (Lac de Pareloup, France): biologi-cal model. Ecol. Model. 65, 1–30.
Tilman, D., Kilham, S.S., Kilham, P., 1982. Phytoplanktoncommunity ecology: the role of limiting nutrients. Annu.Rev. Ecol. Sys. 13, 349–372.
Valiela, I., 1995. Marine Ecological Processes. Springer-Ver-lag, New York 686pp.
Vanderploeg, H.A., 1981. Seasonal particle selection by Diap-tomus sicilis in offshore Lake Michigan. Can. J. Fish.Aquat. Sci. 38, 504–517.
Vanderploeg, H.A., 1994. Zooplankton particle selection andfeeding mechanism. In: Wotton, R.S. (Ed.), The Biology ofParticles in Aquatic Systems. Lewis, New York.
Vanderploeg, H.A., Paffenhofer, G.A., Liebig, J.R., 1988.Diaptomus vs net phytoplankton: effects of algal size andmorphology on selectivity of a behaviorally flexiblem, om-nivorous copepod. Bull. Mar. Sci. 43 (3), 377–394.
Zhu, J., Chen, C., Ralph, E., Green, S.A., Budd, J.W., 2001.Prognostic modeling studies of the Keweenaw current inLake Superior. Part II: simulation. J. Phys. Oceanogr. 31,396–410.
Related Documents











