ORIGINAL PAPER A method for measuring the settling velocity distribution of large biotic particles B. Loubet N. Jarosz S. Saint-Jean L. Huber Received: 24 March 2006 / Accepted: 29 January 2007 / Published online: 19 June 2007 Ó Springer Science+Business Media B.V. 2007 Abstract A simple method for measuring the settling velocity (V s ) distribution of pollen and spores 30–100 mm in diameter is detailed and evaluated. The method is called the ‘settling tower’ and consists in taking sequential pictures of particles falling under gravity in calm air. The scene is illuminated by a cold light source, while a camera takes 15 pictures per second. Between 20,000 and 100,000 images are analysed to obtain the distribution of V s for a given set of particles. The method was validated using two standard particles with mean diameters of 68 and 108 mm, respectively, as well as Lycopodium spores, with a mean diameter of 35 mm. For each set of particles, the theoretical V s distribution was estimated from the particle diameter distribution and the volu- metric mass using a non-Stokian law, as the Reynolds numbers of the particles were large. The mean V s was measured with the ‘settling tower’ with less than 12% error, while the standard deviation of the V s distribu- tion was estimated with less than 51% error. The maximum error on the mean V s was 12% for the Lycopodium spores and less than 2% for the two larger particles. The mean V s of Lycopodium spores was 4.2 cm s 1 , and its standard deviation was 0.7 cm s 1 . The reason for the small overestimation of V s for Lycopodium spores by the ‘settling tower’ method is discussed. Preliminary measurements shows that, the ‘settling tower’ could be of great practical interest for measuring the distribution of V s of maize pollen as well as other types of pollen or spores. Keywords Falling speed Lycopodium spores Maize pollen Particle standards Particle tracking List of symbols Symbol Signification Unit Value a (range) Roman A T Greyscale threshold for converting 8-bit images to a binary 15–21 (0–255) C D Drag coefficient of the particle – – d p Diameter of the particle m 30–200 · 10 6 d G Diameter of the Gaussian filtering pixel 1–3 f Frequency of image acquisition images s 1 15 g Acceleration due to gravity ms 2 9.81 B. Loubet (&) N. Jarosz S. Saint-Jean L. Huber National Institute of Agronomic Research (INRA), UMR EGC, 78850 Thiverval-Grignon, France e-mail: [email protected] N. Jarosz National Institute of Agronomic Research (INRA), UR EPHYSE, BP 81, 33883 Villenave d’Ornon, France 123 Aerobiologia (2007) 23:159–169 DOI 10.1007/s10453-007-9054-2

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL PAPER
A method for measuring the settling velocity distributionof large biotic particles
B. Loubet Æ N. Jarosz Æ S. Saint-Jean Æ L. Huber
Received: 24 March 2006 / Accepted: 29 January 2007 / Published online: 19 June 2007
� Springer Science+Business Media B.V. 2007
Abstract A simple method for measuring the
settling velocity (Vs) distribution of pollen and spores
30–100 mm in diameter is detailed and evaluated. The
method is called the ‘settling tower’ and consists in
taking sequential pictures of particles falling under
gravity in calm air. The scene is illuminated by a cold
light source, while a camera takes 15 pictures per
second. Between 20,000 and 100,000 images are
analysed to obtain the distribution of Vs for a given set
of particles. The method was validated using two
standard particles with mean diameters of 68 and
108 mm, respectively, as well as Lycopodium spores,
with a mean diameter of 35 mm. For each set of
particles, the theoretical Vs distribution was estimated
from the particle diameter distribution and the volu-
metric mass using a non-Stokian law, as the Reynolds
numbers of the particles were large. The mean Vs was
measured with the ‘settling tower’ with less than 12%
error, while the standard deviation of the Vs distribu-
tion was estimated with less than 51% error. The
maximum error on the mean Vs was 12% for the
Lycopodium spores and less than 2% for the two larger
particles. The mean Vs of Lycopodium spores was
4.2 cm s�1, and its standard deviation was 0.7 cm s�1.
The reason for the small overestimation of Vs for
Lycopodium spores by the ‘settling tower’ method is
discussed. Preliminary measurements shows that, the
‘settling tower’ could be of great practical interest for
measuring the distribution of Vs of maize pollen as
well as other types of pollen or spores.
Keywords Falling speed � Lycopodium spores �Maize pollen � Particle standards � Particle tracking
List of symbols
Symbol Signification Unit Valuea
(range)
Roman
AT Greyscale threshold
for converting 8-bit
images to a binary
15–21 (0–255)
CD Drag coefficient
of the particle
– –
dp Diameter
of the particle
m 30–200 · 10�6
dG Diameter of the
Gaussian filtering
pixel 1–3
f Frequency of image
acquisition
images
s�115
g Acceleration due
to gravity
m s�2 9.81
B. Loubet (&) � N. Jarosz � S. Saint-Jean � L. Huber
National Institute of Agronomic Research (INRA),
UMR EGC, 78850 Thiverval-Grignon, France
e-mail: [email protected]
N. Jarosz
National Institute of Agronomic Research (INRA),
UR EPHYSE, BP 81, 33883 Villenave d’Ornon, France
123
Aerobiologia (2007) 23:159–169
DOI 10.1007/s10453-007-9054-2

continued
Symbol Signification Unit Valuea
(range)
L Length of a trajectory
let by a particle
during time s
pixel –
Rep Reynolds number
of the particle:
Rep = Vs dp / ma
– –
Vs Settling velocity
of the particle
m s�1 –
xI Location of
trajectories
on the i-th
following image
xI = I Vs / f + x0
m –
x0 Location of a
trajectory on
the first image
of a serie
m –
q Ratio of minimum-to-
maximum diameter
of spores
– 1.15
Greek
a Number of pixels
per metre
pixel m�1 [151 · 102] –
[155 · 102]
e Pixel tolerance for
tracking a particle
pixel 1–4
s Integration time
of the electronic
shutter of the
camera
s 42.67 · 10�3
(10�5 – 2)
sp Particle inertial time
scale sp = Vs / gs –
qa Volumetric mass of
air
kg m�3 1.20b
qp Volumetric mass
of the particles
kg m�3 1030–1050
la Dynamic viscosity
of air
kg m�1 s�1 1.82 · 10�5b
ma Kinematic viscosity
of air : ma = la / qa
m2 s�1 1.51 · 10�5b
a Value taken in the study if not otherwise statedb Values at 208C and 105 Pa
1 Introduction
Pollen and spores are dispersal agents of genes and
diseases in the atmosphere (e.g. McCartney 1994). In
the context of an increasing use of genetically
modified (GM) crops as well as the prospect of
diminishing pesticides inputs, it is essential to be able
to characterise well the transport of such particles at
several scales (Aylor and Irwin 1999; Aylor 2003).
The dispersal of pollen and spores can be charac-
terised by experiments based on physical measure-
ments (Aylor and Taylor 1983; Jarosz et al. 2003,
2005) or by using tracers (Haskell and Dow 1951; Luna
et al. 2001). The dispersal of biotic particles can also be
modelled using either statistical approaches (Lavigne
et al. 1996, 1998, 2002) or mechanistic models (Klein
et al. 2003; Jarosz et al. 2004; Dupont et al. 2006). The
latter require less experimental input and may, as such,
be of great interest for predicting the dispersal of biotic
particles in the environment under various conditions.
One essential parameter of those models is the settling
velocity of the particle (Vs), defined as the terminal
velocity of the particle in steady air. Each type of
pollen and spore has a distribution of sizes, densities
and shapes. Consequently, biotic particles have a
distributed Vs (e.g. Renoux and Boulaud 1998;
Seinfeld and Pandis 1998). Since only a small fraction
of the particles can be responsible for major effects
(disease spread, transfer of the genetic material of GM
organisms), it is therefore essential to know the
distribution of Vs rather than only an average value
of Vs for a given specie.
The settling velocity Vs is an aerodynamic charac-
teristic of heavy particles, which is well parameterised
for ideal spherical particles in the atmosphere (Fuchs
1964; Renoux and Boulaud 1998; Seinfeld and Pandis
1998; Aylor 2002). However, these parameterisations
require prior knowledge of the volumetric mass and
size distributions of the particles, both of which are
difficult to measure for pollen and spores; in addition,
these characteristics can evolve very quickly in pollen
and spores in the atmosphere due to drying (Aylor
et al. 2003). Hence, for these biotic particles it may be
easier to directly measure Vs. Several techniques are
currently used to determine Vs, including:
– The Laser Doppler Velocimetry (LDV), which
consists in monitoring the frequency of the inter-
ference pattern created by particles crossing two
shifted-phased beams of one laser. The frequency
is linked to the velocity by the so-called Doppler
relationship (e.g. Rambert et al. 1998);
– The Particle Image Velocimetry (PIV), based on
calculating the autocorrelation of successive
images of particles obtained with a fast video
160 Aerobiologia (2007) 23:159–169
123

camera in a scene illuminated by a powerful laser
source (e.g. Adrian 1991);
– The Time-Of-Flight particle Spectrometer
(TOFS), which consists in measuring the time
spent by a particle to travel a given distance
between two laser beams after being accelerated
in a calibrate nozzle (Fields 2002);
– Time of Fall Under Gravity (TFUG) method,
which is based on measuring the time taken by a
particle to fall under gravity through a given
distance in steady air (Aylor 2002; Di-Giovanni
et al. 1995; Ferrandino and Aylor 1984; Malcom
and Raupach 1991; Sawyer et al. 1994).
The LDV and the PIV methods are reference
techniques; however, they both are very expensive
due to the use of a powerful laser and a fast camera,
and the accuracy of the former may be questionable
when the particle shape is not spherical (e.g. Rambert
et al. 1998). The TOFS method can also be consid-
ered as a reference method; however, it also is
expensive, and it requires the particles to be main-
tained in a small volume and injected separately,
which may modify their water content. The TFUG
method is conceptually much simpler and requires
less expensive equipment, but the results are often
associated with uncertainty due to difficulties in
determining the time at which the particles start to
settle (Di-Giovanni et al. 1995; Ferrandino and Aylor
1984). The method presented by (Aylor 2002) avoids
this limitation but is time-consuming as it requires
that the time of flight of each pollen grain be recorded
by hand in order that a whole distribution of Vs be
estimated. The method presented by Sawyer et al.
(1994) is comparable to that of Aylor (2002) but uses
a camera with 60 images per second and a tracking
algorithm. This method has proved to be very well
adapted for particles smaller than 30 mm but is
limited by the number of particles that can be
analysed due to the small size of the field of view.
In this study, we present an automated method for
measuring the settling velocity distribution of pollen
and spores having a diameter ranging from 30 to
100 mm, based on the time of fall under gravity. The
method is evaluated against standard parameterisa-
tions of Vs and calibrated particles. An example
distribution of maize pollen Vs is then presented and
the potential implication for pollen dispersal is briefly
discussed.
2 Material and methods
2.1 The ‘settling tower’ apparatus
The scheme and dimensions of the ‘settling tower’
apparatus are given in Fig. 1. In this approach, the
particles are released from a brush placed at a few
millimeters over the top of a 1-m-high stainless steel
tube of a rectangular section. The bottom of the tube
is placed on the top of a wood photographic
chamber with dimensions such as to avoid any
significant wall effects (Table 1). Within the cham-
ber, a window allows filming, while two fibre-optic
light sources illuminate the scene from the sides.
The use of fibre-optic light sources (150 W in total)
avoids overheating of the chamber. The camera
(XCD-X700, Sony, Japan) is equipped with a zoom
(ICA30, 12.5–75 mm, macro, 1:1.8). The tempera-
ture is measured with a mercury thermometer inside
the photographic chamber; in our experiments the
temperature never exceeded 308C. The photographic
chamber is enclosed in a larger box that is
completely sealed and isolated with polystyrene
plates (width: 5 cm). The inside of the photographic
chamber is painted with a black body painting to
avoid any unwanted light reflection. The camera is
connected to a computer for image acquisition with
a frequency f = 15 images s�1. The field of view is
typically 5 cm.
A decametre placed on the side of the film was
used to determine the number of pixels per metre (a).
The light saturation of the camera charge-coupled
device (CCD) sensor was set to the maximum, and
the electronic shutter was set to 42.67 ± 0.005 ms
integration time (s). By doing so, the particles falling
during the time s left a ‘track’ on each image. The
tracks are hereafter called ‘trajectories’. Seven video
files of 1 min were successively recorded as AVI
using SONYCAP software (Sony, Japan), while the
brush was gently tapped.
2.2 Image analysis and data processing
The recorded video files were split into 100-Mo
pieces and analysed with IMAGE J (http://rsb.info.
nih.gov/ij/) with the following sequence of analysis:
– Converting the image to 8 bits with greyscale
from levels 0 to 255;
Aerobiologia (2007) 23:159–169 161
123

– Cropping the image to withdraw the edges of the
window;
– Gaussian filtering with radius dG,;
– Convolving the image with a matrix of five
identical rows (each row [�1; �1; 4; �1; �1]) to
maximise vertical structures (particle trajectories);
X . Y . Z positioning
05
032
059
300600
1200
20
5
SIDE VIEW
Sony XCD-X700digital camera
to computer
70
Hole for the optic fiber
Optics ICA3Optics ICA3
032
059
230
30
Fibber-optics
FRONT VIEW
150 W
Window50 x 50
Nylon mesh
150 W
Fibber-optics
TOP VIEW
Small hole (5 x 30) to force the particles in the focal plane
Camera
Zoom
150 W150 W
003
230
150 W
3
150 W150 W150 W
Fig. 1 Scheme of the
‘settling tower’. The
dimensions are given in
millimetres. Particles were
released at the top of the
tower, passing through a
small hole before entering
the ‘photographic chamber’
illuminated by two fibre-
optic light sources. The
particle trajectories were
recorded on a video with
a digital camera equipped
with a zoom (ICA3). Two
rolls of nylon meshes
(1-mm grid,
5 mm · 23 mm) were
placed on the sides of the
‘photographic chamber’ to
scatter the light from the
fibre-optic light sources and
to isolate the scene from the
small amount of heat
generated by these light
sources. A thin hole at the
top of the box forced the
particles into the narrow
focal plane of the camera
Table 1 Particle characteristics used for the calibration of the ‘settling tower’
Particle Material Diameter (mm) Volumetric
mass
(kg m�3)
Vs
(cm s�1)
Rep Wall
effect
(cm)
AT
(0–255)
dG
(pixel)
e(pixel)
Mean SD
Lycopodium Spore 35.4 ± 0.4 2.3 ± 0.2 1030 ± 90 3.8 0.09 0.79 21 1 1
Dri-Cal 70 Polysterene divinylbenzene
(DVB)(4–8%)
68.3 ± 3 3.7 1050 13.0 0.59 0.23 17 1 2
Dri-Cal 100 Polysterene DVB
(4–8%)
108 ± 4 9.8 1050 28.7 2.07 0.10 15 3 3
Particle characteristics: the mean and standard deviation (SD) of the diameters of the Dri-Cal particles (70 and 100 mm, respectively)
are certified, while the mean diameter of Lycopodium spores was fitted with a Gaussian curve to obtain the mean and standard
deviation of the spores. The theoretical settling velocity Vs is also given as calculated using Eqs. (3) and (4) as well as the Reynolds
number of the particle (Rep) and the distance at which the wall effect may be important (20 dp/Rep; see Sawyer et al. 1994). The
greyscale threshold AT, the Gaussian filter diameter dG and the pixel tolerance (e) used in the image analysis are also given for each
particle type. Data are given for 208C and 105 Pa. The air humidity is ignored. The mean and standard deviation of the Lycopodiumspores estimated using a microscope was 36.8 ± 4.2 mm. This measurement is much less precise than that obtained using the Coulter
apparatus
162 Aerobiologia (2007) 23:159–169
123

– Gaussian filtering with radius dG,;
– Converting the 8-bit image to a binary image
using a greyscale threshold level AT (over 255
levels);
– Identifying each object in the image with the
macro ANALYZE PARTICLES, based on the use of the
WAND method.
This image analysis, illustrated in Fig. 2, gave the
size and location of the trajectories left by each
particle on each image. The following trajectories
were removed: (1) trajectories with a width larger
than 4 pixels due to overlapping trajectories and (2)
trajectories located at less than 50 pixels from any
side of the image to avoid cut-trajectories. The length
of each trajectory was diminished by its width to
account for the diameter of the particle. The resulting
distribution of trajectory lengths (L, in pixels) was
then used to process the distribution of the settling
velocity by simply using the following equation:
Vs ¼ L=ða� sÞ ð1Þ
The data obtained by this method are hereafter
called ‘uncorrected’. A correction was then applied in
order to eliminate trajectories too close to each other
and to keep only particles with trajectories identified
on successive images. A recursive algorithm was
built to estimate the uppermost location xI of a given
particle trajectory on the next images, knowing its
actual location (x0), its settling velocity Vs, as
estimated with Eq. (1) and the frequency of image
acquisition (f):
xI ¼ x0 þ IVs=f ð2Þ
where I is the image number. A tolerance (e) of
1–4 pixels corresponding roughly to 0.07–0.27 mm
(or equivalently an error on the settling velocity of
0.16�0.63 cm s�1) was allowed to track the trajec-
tories from one image to the next. The resulting
distribution of Vs is hereafter denoted ‘corrected’.
The number of particle trajectories before correction
ranged between 19,400 and 99,200; after correction,
the number of trajectories varied between 130 and
640, which is less than 1% of the number of
trajectories recorded.
2.3 Calibrated particles
In order to test and calibrate the method described
above, three sets of spherical particles with different
mean diameters were used (see Table 1 for charac-
teristics). These particles were chosen to have similar
characteristics to pollen or large spores: a volumetric
mass close to 1000 kg m�3 and a high refractive
index. Two sets consisted of standard particles made
of perfectly spherical polysterene DVB microspheres
[Dri-Cal 70 (70 mm), lot No. 27127 and Dri-Cal 100
(100 mm), lot No 28000; Duke Scientific, USA] with
certified diameters and known volumetric mass; one
set consisted of almost spherical Lycopodium spores
(Lot No. 62701; ADP Laboratoire Pharmaceutique,
France) with the diameter and volumetric mass
determined prior to the experiment.
The diameter distribution of the Lycopodium
spores was measured with an automatic cell counter
(Coulter Multisizer III; Beckman, USA) equipped
with a 240-mm-diameter orifice. Between 5000 and
8000 Lycopodium spores were suspended in an
Fig. 2 Example of araw
image (a), threshold image
(b) and particle contour (c)
following image analysis
for Dri-Cal 70 (mm)
particles. Note that the raw
image can be of bad quality
without altering the
trajectory detection
pathway
Aerobiologia (2007) 23:159–169 163
123

electrolyte solution (Coulter Isoton, Beckman) and
counted within the next minute. The operation was
repeated 20 times. The diameter distribution was
then fit with a Gaussian curve to obtain the mean
and standard deviation (Table 1). We calculated the
mean diameter of the Lycopodium spores to be
35.4 mm, which is similar to the 35.6 mm reported
by Rambert et al. (1998) using both a phase-doppler
method and a laser-diffraction spectrometer method.
However, it is larger than the 32.8 mm reported by
Ferrandino and Aylor (1984). Visual control of the
spores with a microscope showed that the spores
were tetrahedral, as expected, with a smaller
dimension of 34.4 mm and a larger dimension of
39.3 mm, giving a ratio of larger to smaller
dimension q = 1.15; the corresponding average
and standard deviation of the diameter is 36.8 and
4.2 mm, respectively.
The Lycopodium spores were suspended in the
electrolyte solution for less than 1 min in order to
avoid changes in their diameter. Preliminary exper-
iments had proven that 1 min is sufficiently short to
avoid the spores absorbing electrolyte solution and
consequently increasing their diameter; in contrast,
the diameter of the spores could increase from 35 to
38 mm over a 30-min period in the electrolyte
solution. Consequently, the Lycopodium spores were
kept in a dry environment (laboratory).
The volumetric mass of the three sets of
spherical particles was measured by weighing a
10- to 20-mg sample of particles on a precision
balance (precision: 0.1 mg). The sample was then
put into a known volume of the electrolyte solution.
The volume of particles contained in 1 ml of the
solution was determined ten times within a very
short time period (less than 1 min) using the
automatic counter. This procedure was repeated five
times. The uncertainty of the method was estimated
from the uncertainty of the balance and the standard
deviation of the counted volume of particles over
50 repetitions.
2.4 Theoretical settling velocity
The theoretical settling velocity of calibrated parti-
cles was calculated using the parameterisation given
by Aylor (2002) and taken from Fuchs (1964), over a
wide range of Reynolds number Rep (from
0.001 to 200):
V2s ¼
4gdpqp
3CDqa
ð3Þ
CD ¼24
Rep
1þ 0:158Re2=3p
� �ð4Þ
where dp and qp are particle diameter and volumetric
mass, respectively and qa is the air volumetric mass.
The particle inertial time scale sp is defined as sp = Vs
g�1. The characteristic distance required for the
particle velocity to reach its terminal velocity was
estimated in a simplified way as Z = sp Vs = Vs2 g�1.
For all particles, it was smaller than 1 cm. The height
of the ‘settling tower’ was, however, high enough to
minimise the influence of the external turbulence
inside the photographic chamber.
2.5 Determination of the image analysis
parameters
Three parameters were adjusted to optimise the image
analysis and data processing: the Gaussian filter
diameter dG, the threshold AT and the tolerance e.The standard set of parameters used in this study is:
dG = 1 pixel, AT = 17 and e = 2 pixels. Since the choice
of these parameters is sensitive to the set-up of the light
source, the position of the camera as well as the settling
velocity, the sensitivity to dG and AT was assessed for
each type of particle (Table 1) varying dG from 1 to 3
and AT from 5 to 40. Figure 3 shows the number of
trajectories found on two images of settling Dri-Cal 70.
The correct number of trajectories, determined by eye,
were found for dG = 1 and AT = 17. Small changes
around the optimum AT did not affect the number of
trajectories found. As a rule of thumb, the less the
trajectories are contrasted (the harder to see), the larger
the dG and the smaller the AT. The last parameter e was
determined empirically around a value equal to 2 · dG.
3 Results and discussion
3.1 Settling velocity distributions of calibrated
particle
Figure 4 shows the uncorrected and corrected settling
velocity distributions of the three calibrated particles:
uncorrected Vs are seen to give wider distributions
than corrected Vs, and additional modes are generally
164 Aerobiologia (2007) 23:159–169
123
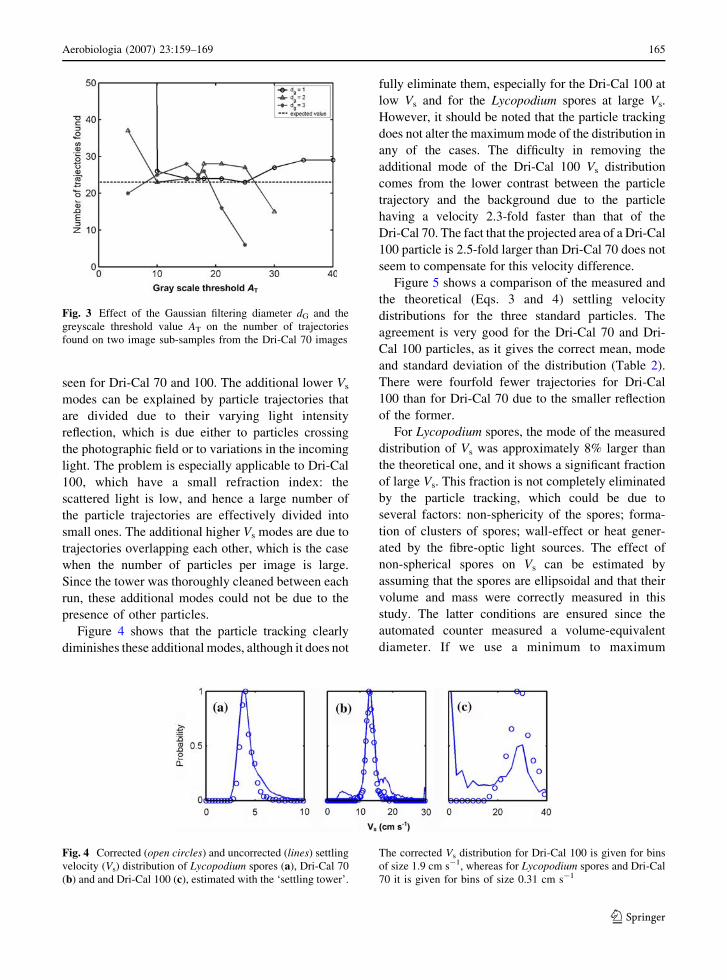
seen for Dri-Cal 70 and 100. The additional lower Vs
modes can be explained by particle trajectories that
are divided due to their varying light intensity
reflection, which is due either to particles crossing
the photographic field or to variations in the incoming
light. The problem is especially applicable to Dri-Cal
100, which have a small refraction index: the
scattered light is low, and hence a large number of
the particle trajectories are effectively divided into
small ones. The additional higher Vs modes are due to
trajectories overlapping each other, which is the case
when the number of particles per image is large.
Since the tower was thoroughly cleaned between each
run, these additional modes could not be due to the
presence of other particles.
Figure 4 shows that the particle tracking clearly
diminishes these additional modes, although it does not
fully eliminate them, especially for the Dri-Cal 100 at
low Vs and for the Lycopodium spores at large Vs.
However, it should be noted that the particle tracking
does not alter the maximum mode of the distribution in
any of the cases. The difficulty in removing the
additional mode of the Dri-Cal 100 Vs distribution
comes from the lower contrast between the particle
trajectory and the background due to the particle
having a velocity 2.3-fold faster than that of the
Dri-Cal 70. The fact that the projected area of a Dri-Cal
100 particle is 2.5-fold larger than Dri-Cal 70 does not
seem to compensate for this velocity difference.
Figure 5 shows a comparison of the measured and
the theoretical (Eqs. 3 and 4) settling velocity
distributions for the three standard particles. The
agreement is very good for the Dri-Cal 70 and Dri-
Cal 100 particles, as it gives the correct mean, mode
and standard deviation of the distribution (Table 2).
There were fourfold fewer trajectories for Dri-Cal
100 than for Dri-Cal 70 due to the smaller reflection
of the former.
For Lycopodium spores, the mode of the measured
distribution of Vs was approximately 8% larger than
the theoretical one, and it shows a significant fraction
of large Vs. This fraction is not completely eliminated
by the particle tracking, which could be due to
several factors: non-sphericity of the spores; forma-
tion of clusters of spores; wall-effect or heat gener-
ated by the fibre-optic light sources. The effect of
non-spherical spores on Vs can be estimated by
assuming that the spores are ellipsoidal and that their
volume and mass were correctly measured in this
study. The latter conditions are ensured since the
automated counter measured a volume-equivalent
diameter. If we use a minimum to maximum
Fig. 3 Effect of the Gaussian filtering diameter dG and the
greyscale threshold value AT on the number of trajectories
found on two image sub-samples from the Dri-Cal 70 images
Fig. 4 Corrected (open circles) and uncorrected (lines) settling
velocity (Vs) distribution of Lycopodium spores (a), Dri-Cal 70
(b) and and Dri-Cal 100 (c), estimated with the ‘settling tower’.
The corrected Vs distribution for Dri-Cal 100 is given for bins
of size 1.9 cm s�1, whereas for Lycopodium spores and Dri-Cal
70 it is given for bins of size 0.31 cm s�1
Aerobiologia (2007) 23:159–169 165
123
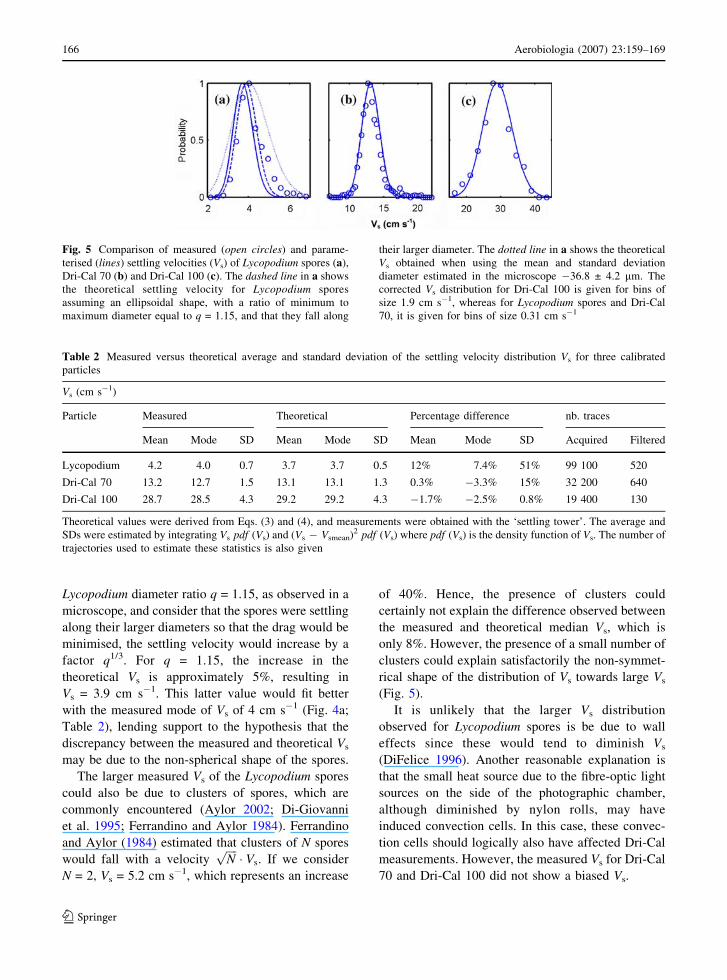
Lycopodium diameter ratio q = 1.15, as observed in a
microscope, and consider that the spores were settling
along their larger diameters so that the drag would be
minimised, the settling velocity would increase by a
factor q1/3. For q = 1.15, the increase in the
theoretical Vs is approximately 5%, resulting in
Vs = 3.9 cm s�1. This latter value would fit better
with the measured mode of Vs of 4 cm s�1 (Fig. 4a;
Table 2), lending support to the hypothesis that the
discrepancy between the measured and theoretical Vs
may be due to the non-spherical shape of the spores.
The larger measured Vs of the Lycopodium spores
could also be due to clusters of spores, which are
commonly encountered (Aylor 2002; Di-Giovanni
et al. 1995; Ferrandino and Aylor 1984). Ferrandino
and Aylor (1984) estimated that clusters of N spores
would fall with a velocityffiffiffiffiNp� Vs. If we consider
N = 2, Vs = 5.2 cm s�1, which represents an increase
of 40%. Hence, the presence of clusters could
certainly not explain the difference observed between
the measured and theoretical median Vs, which is
only 8%. However, the presence of a small number of
clusters could explain satisfactorily the non-symmet-
rical shape of the distribution of Vs towards large Vs
(Fig. 5).
It is unlikely that the larger Vs distribution
observed for Lycopodium spores is be due to wall
effects since these would tend to diminish Vs
(DiFelice 1996). Another reasonable explanation is
that the small heat source due to the fibre-optic light
sources on the side of the photographic chamber,
although diminished by nylon rolls, may have
induced convection cells. In this case, these convec-
tion cells should logically also have affected Dri-Cal
measurements. However, the measured Vs for Dri-Cal
70 and Dri-Cal 100 did not show a biased Vs.
Fig. 5 Comparison of measured (open circles) and parame-
terised (lines) settling velocities (Vs) of Lycopodium spores (a),
Dri-Cal 70 (b) and Dri-Cal 100 (c). The dashed line in a shows
the theoretical settling velocity for Lycopodium spores
assuming an ellipsoidal shape, with a ratio of minimum to
maximum diameter equal to q = 1.15, and that they fall along
their larger diameter. The dotted line in a shows the theoretical
Vs obtained when using the mean and standard deviation
diameter estimated in the microscope �36.8 ± 4.2 mm. The
corrected Vs distribution for Dri-Cal 100 is given for bins of
size 1.9 cm s�1, whereas for Lycopodium spores and Dri-Cal
70, it is given for bins of size 0.31 cm s�1
Table 2 Measured versus theoretical average and standard deviation of the settling velocity distribution Vs for three calibrated
particles
Vs (cm s�1)
Particle Measured Theoretical Percentage difference nb. traces
Mean Mode SD Mean Mode SD Mean Mode SD Acquired Filtered
Lycopodium 4.2 4.0 0.7 3.7 3.7 0.5 12% 7.4% 51% 99 100 520
Dri-Cal 70 13.2 12.7 1.5 13.1 13.1 1.3 0.3% �3.3% 15% 32 200 640
Dri-Cal 100 28.7 28.5 4.3 29.2 29.2 4.3 �1.7% �2.5% 0.8% 19 400 130
Theoretical values were derived from Eqs. (3) and (4), and measurements were obtained with the ‘settling tower’. The average and
SDs were estimated by integrating Vs pdf (Vs) and (Vs � Vsmean)2 pdf (Vs) where pdf (Vs) is the density function of Vs. The number of
trajectories used to estimate these statistics is also given
166 Aerobiologia (2007) 23:159–169
123

Others studies have estimated settling velocities
of Lycopodium spores: using a laser Doppler
method, Ferrandino and Aylor (1984) found a mean
Vs of 1.94 ± 0.02 cm s�1, while Rambert et al.
(1998) found Vs = 1.8 cm s�1; Gregory (1973) also
found 1.7 cm s�1, with diameters similar to ours or
slightly smaller. These results, obtained with vari-
ous methods, still give smaller values of Vs than our
measurements. However, these results also give
smaller Vs than what is expected from Eqs. (3) and
(4): with dp = 32.8 mm, as reported by Ferrandino
and Aylor (1984), Eqs. (3) and (4) give
Vs = 3.17 cm s�1, which is much larger than the
reported value Vs = 1.94 cm s�1. The ‘honey-
combed’ surface of some Lycopodium spores may
explain the small Vs reported by other studies.
Indeed, a ‘honeycombed’ surface contains air,
which leads to a diminished apparent volumetric
mass of the falling spores. Using Eqs. (3) and (4) to
reconcile the measured dp and Vs by Ferrandino and
Aylor (1984), we can estimate that the effective
volumetric mass of the spore should be 600 kg m�3
rather than 1000 kg m�3. This would mean that the
holes of the ‘honeycombed’ surface were 0.2 dp
deep. The fact that our Vs measurements are in
agreement with the results of Eqs. (3) and (4)
suggests that our Lycopodium spores had smaller
‘honeycombs’. As also noticed, it is unlikely that
convection currents can explain the differences of
more than 1 cm s�1 between other studies and this
study, since it would also have biased the measured
on Dri-Cal 70 and 100, which is not the case (see
Table 2).
The comparison of the measured and theoretical
mean Vs (Table 2) shows that the percentage differ-
ence is smaller than 1% for the Dri-Cal particles and
smaller than 12% for the Lycopodium spores. The
mode is even better estimated for the Lycopodium
spores with a 7.4% error. For Dri-Cal 70 and 100, the
estimated mode is more uncertain due to the smaller
number of particles. The standard deviations of the
distributions are always overestimated by the mea-
surements. However, the error expressed in percent-
age of the theoretical standard deviation is small and
is 51, 15 and 0.8% for the Lycopodium spores and the
Dri-Cal 70 (70 mm) and Dri-Cal 100 (100 mm)
particles, respectively. These differences become
only 6, 2 and 0.4%, respectively, when expressed as
a percentage of the mode of the distribution.
The absolute error between the measured and the
theoretical mean Vs ranges from 0.5 cm s�1 for
Lycopodium spores to 0.1 cm s�1 for Dri-Cal 100.
The accuracy of the method is roughly 0.25 cm s�1
when estimated as the standard deviation of the linear
regression between the theoretical and measured
mean Vs.
3.2 Example of settling velocity of maize pollen
Well-hydrated maize pollen is spherical and its mean
diameter is 90 mm (Di-Giovanni et al. 1995). The
settling velocity of maize pollen was measured using
the ‘settling tower’ in order to evaluate the method
with another biotic particle and to see whether the Vs
distribution is monodispersed. Maize tassels of vars.
Meribel and DK315 were sampled in a field near the
laboratory at different hours of the day. They were
quickly (less than 10 min) brought back to the lab and
gently taped above the settling tower.
Although maize pollen Vs was as large as Dri-Cal
100 particles, the obtained films showed a much better
contrast between the particle trajectory of the former
and the background due to a good light refraction by
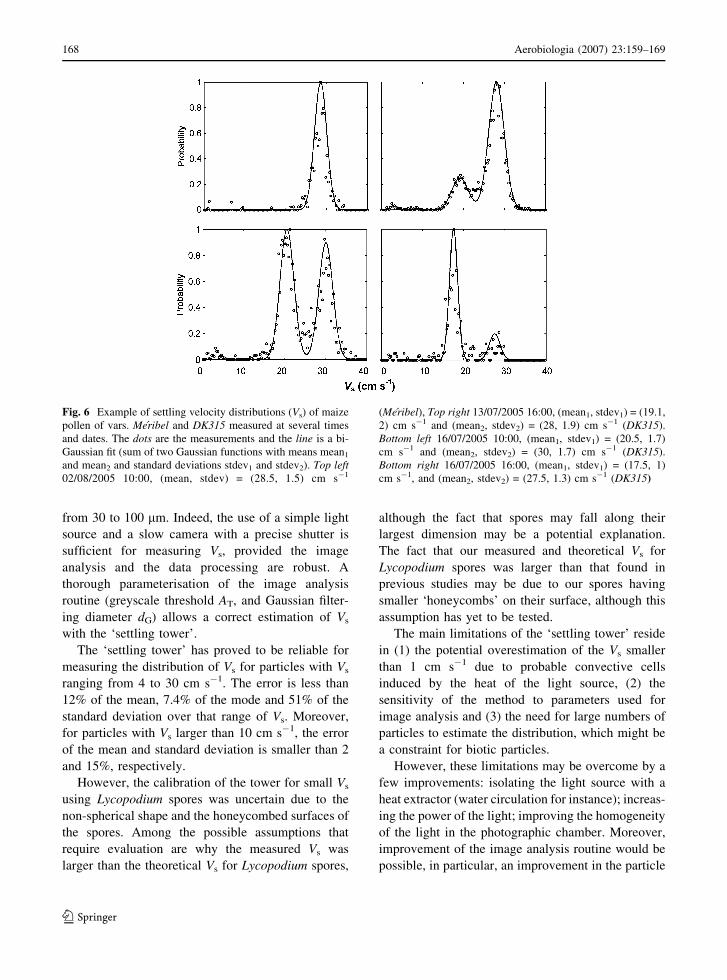
the maize pollen. Figure 6 shows an example of four Vs
distributions of maize pollen sampled at different
times and dates and, therefore, with varying mean
water content. It clearly shows two populations of Vs
present in different proportions. From the top left to
the bottom right of Fig. 6, both a range in the
proportions of the two populations and a shift in
the mode of the largest Vs distribution are observed.
The validation of the method over a range of Vs larger
than that observed in Fig. 6 enables any errors in the
measurements to be discarded. Figure 6 points out the
potential of using the detailed distribution of Vs to
identify the populations of pollen emitted and there-
fore improve our understanding of the effects of the
environmental conditions on the emission processes.
The work of Aylor (2002, 2003) suggests that these
modes correspond to dry and wet pollen, respectively.
However, more research is needed to interpret the two
modes observed on these maize pollen distributions.
4 Concluding remarks
The method presented here for measuring Vs has been
shown to be adaptable to the measurement of unit
volumetric mass particles with a diameter ranging
Aerobiologia (2007) 23:159–169 167
123
from 30 to 100 mm. Indeed, the use of a simple light
source and a slow camera with a precise shutter is
sufficient for measuring Vs, provided the image
analysis and the data processing are robust. A
thorough parameterisation of the image analysis
routine (greyscale threshold AT, and Gaussian filter-
ing diameter dG) allows a correct estimation of Vs
with the ‘settling tower’.
The ‘settling tower’ has proved to be reliable for
measuring the distribution of Vs for particles with Vs
ranging from 4 to 30 cm s�1. The error is less than
12% of the mean, 7.4% of the mode and 51% of the
standard deviation over that range of Vs. Moreover,
for particles with Vs larger than 10 cm s�1, the error
of the mean and standard deviation is smaller than 2
and 15%, respectively.
However, the calibration of the tower for small Vs
using Lycopodium spores was uncertain due to the
non-spherical shape and the honeycombed surfaces of
the spores. Among the possible assumptions that
require evaluation are why the measured Vs was
larger than the theoretical Vs for Lycopodium spores,
although the fact that spores may fall along their
largest dimension may be a potential explanation.
The fact that our measured and theoretical Vs for
Lycopodium spores was larger than that found in
previous studies may be due to our spores having
smaller ‘honeycombs’ on their surface, although this
assumption has yet to be tested.
The main limitations of the ‘settling tower’ reside
in (1) the potential overestimation of the Vs smaller
than 1 cm s�1 due to probable convective cells
induced by the heat of the light source, (2) the
sensitivity of the method to parameters used for
image analysis and (3) the need for large numbers of
particles to estimate the distribution, which might be
a constraint for biotic particles.
However, these limitations may be overcome by a
few improvements: isolating the light source with a
heat extractor (water circulation for instance); increas-
ing the power of the light; improving the homogeneity
of the light in the photographic chamber. Moreover,
improvement of the image analysis routine would be
possible, in particular, an improvement in the particle
Fig. 6 Example of settling velocity distributions (Vs) of maize
pollen of vars. Meribel and DK315 measured at several times
and dates. The dots are the measurements and the line is a bi-
Gaussian fit (sum of two Gaussian functions with means mean1
and mean2 and standard deviations stdev1 and stdev2). Top left02/08/2005 10:00, (mean, stdev) = (28.5, 1.5) cm s�1
(Meribel), Top right 13/07/2005 16:00, (mean1, stdev1) = (19.1,
2) cm s�1 and (mean2, stdev2) = (28, 1.9) cm s�1 (DK315).
Bottom left 16/07/2005 10:00, (mean1, stdev1) = (20.5, 1.7)
cm s�1 and (mean2, stdev2) = (30, 1.7) cm s�1 (DK315).
Bottom right 16/07/2005 16:00, (mean1, stdev1) = (17.5, 1)
cm s�1, and (mean2, stdev2) = (27.5, 1.3) cm s�1 (DK315)
168 Aerobiologia (2007) 23:159–169
123

tracking between successive images would increase
the robustness of the method and allow the lower
trajectories to be rejected. We also recommend that a
systematic sensitivity study should be performed to
define the optimum parameters of the image analysis
for each new setting of the tower.
As suggested by preliminary measurements, the
‘settling tower’ could be of great practical interest for
measuring the distribution of Vs of maize pollen.
Moreover, given that a large number of pollen and
spores have a diameter between 30 and than 100 mm,
this method could be applied to most other types of
pollen or spores. For these particles, Vs measurements
would allow their potential for dispersal in the
atmosphere to be estimated and provide input for
dispersal models, such as those of Aylor et al. (2003)
and Jarosz et al. (2004). Moreover, Vs may be used as
a manner to characterize the presence of several
populations of pollens within a sample, such as the
number of dehydrated pollen or clusters of pollen.
References
Adrian, R. J. (1991). Particle-imaging techniques for experi-
mental fluid mechanics. Annual Review of FluidMechanics, 23, 261–304.
Aylor, D. E. (2002). Settling speed of corn (Zea mays) pollen.
Journal of Aerosol Science, 33, 1601–1607.
Aylor, D. E. (2003). Rate of dehydration of corn (Zea mays L.)
pollen in the air. Journal of Experimental Botany, 54,
2307–2312.
Aylor, D. E., & Taylor, G. S. (1983). Escape of PeronosporaTabacina spores from a field of diseased Tobacco plants.
Phytopathology, 73, 525–529.
Aylor, D. E., & Irwin, M. E. (1999). Aerial dispersal of pests
and pathogens: implications for integrated pest manage-
ment. Agricultural and Forest Meteorology, 97, 233–234.
Aylor, D. E., Schultes, N. P., & Shields, E. J. (2003). An aero-
biological framework for assessing cross-pollination in
maize. Agricultural and Forest Meteorology, 119, 111–129.
DiFelice, R. (1996). A relationship for the wall effect on the
settling velocity of a sphere at any flow regime. Interna-tional Journal of Multiphase Flow, 22, 527–533.
Di-Giovanni, F., Kevan, P. G., & Nasr, M. E. (1995). The
variability in settling velocities of some pollen and spores.
Grana, 34, 39–44.
Dupont, S., Brunet, Y., & Jarosz, N. (2006). Eulerian modelling
of pollen dispersal over heterogenous vegetation canopies.
Agricultural and Forest Meteorology, 141, 82–104.
Ferrandino, F. J., & Aylor, D. E. (1984). Settling speed of
clusters of spores. Phytopatholgy, 74, 969–972.
Fields, T. B. (2002). Aerodynamic time-of-flight particle size
measurement. Journal of Dispersion Science and Tech-nology, 23, 729–735.
Fuchs, N. A. (1964). The mechanics of aerosols. New York:
Macmillan.
Gregory, P. H. (1973). Microbiology of the atmosphere. New
York: John Wiley & Sons.
Haskell, G., & Dow, P. (1951). Studies with sweet corn V.
Seed-settings with distances from pollen source. EmpireJournal of Experimental Agriculture, 19, 45–50.
Jarosz, N., Loubet, B., Durand, B., McCartney, H. A., Fouei-
llassar, X., & Huber, L. (2003). Field measurements of
airborne concentration and deposition of maize pollen.
Agricultural and Forest Meteorology, 119, 37–51.
Jarosz, N., Loubet, B., & Huber, L. (2004). Modelling airborne
concentration and deposition rate of maize pollen. Atmo-spheric Environment, 38, 5555–5566.
Jarosz, N., Loubet, B., Durand, B., Foueillassar, X., & Huber,
L. (2005). Variations in maize pollen emission and
deposition in relation to microclimate. EnvironmentalScience and Technology, 39, 4377–4384.
Klein, E. K., Lavigne, C., Foueillassar, X., Gouyon, P. H., &
Laredo, C. (2003). Corn pollen dispersal: Quasi-mecha-
nistic models and field experiments. Ecological Mono-graphs, 73, 131–150.
Lavigne, C., Godelle, B., Reboud, X., & Gouyon, P. H. (1996).
A method to determine the mean pollen dispersal of
individual plants growing within a large pollen source.
Theoretical and Applied Genetics, 93, 1319–1326.
Lavigne, C., Klein, E. K., Vallee, P., Pierre, J., Godelle, B., &
Renard, M. (1998). A pollen-dispersal experiment with
transgenic oilseed rape. Estimation of the average pollen
dispersal of an individual plant within a field. Theoreticaland Applied Genetics, 96, 886–896.
Lavigne, C., Klein, E. K., & Couvet, D. (2002). Using seed
purity data to estimate an average pollen mediated gene
flow from crops to wild relatives. Theoretical and AppliedGenetics, 104, 139–145.
Luna, V. S., Figueroa M. J., Baltazar M. B., Gomez L. R.,
Townsend R., & Schoper, J. B. (2001). Maize pollen
longevity and distance isolation requirements for effective
pollen control. Crop Science, 41, 1551–1557.
Malcolm, L. P., & Raupach, M. R. (1991). Measurements in an
air settling tube of the terminal velocity distribution of soil
material. Journal of Geophysical Research, 96, 15275–
15286.
McCartney, H. A. (1994). Dispersal of spores and pollen from
crops. Grana, 33, 76–80.
Rambert, A., Huber, L., & Gougat., P. (1998). Laboratory
study of fungal spore movement using Laser Doppler
Velocimetry. Agricultural and Forest Meteorology, 92,
43–53.
Renoux, A., & Boulaud, D. (1998). Les aerosols. Physique etMetrologie. Paris: Lavoisier.
Sawyer, A. J., Griggs, M. H., & Wayne, R. (1994). Dimen-
sions, density, and settling velocity of entomophthoralean
Conidia : implications for aerial dissemination of spores.
Journal of Invertebrate Pathology, 63, 43–55.
Seinfeld, J. H., & Pandis, S. N. (1998). Atmospheric chemistryand physics. From air pollution to climate change. New
York: Wiley-Interscience.
Aerobiologia (2007) 23:159–169 169
123
Related Documents











