A Janus Splicing Regulatory Element Modulates HIV-1 tat and rev mRNA Production by Coordination of hnRNP A1 Cooperative Binding Virginie Marchand 1 , Agne `s Me ´reau 1 , Sandrine Jacquenet 1 Denise Thomas 1 , Annie Mougin 1 , Renata Gattoni 2 , James Ste ´venin 2 and Christiane Branlant 1 * 1 Laboratoire de Maturation des ARN et Enzymologie Mole ´culaire, UMR 7567 UHP-CNRS, Universite ´ Henri Poincare ´ Nancy 1, Boulevard des Aiguillettes, BP239, 54506 Vandoeuvre-le `s-Nancy Cedex France 2 IGBMC, 1 rue Laurent Fries BP 163, Parc d’ innovation 67404 Illkirch Cedex, France Retroviral protein production depends upon alternative splicing of the viral transcript. The HIV-1 acceptor site A7 is required for tat and rev mRNA production. Production of the Tat transcriptional activator is highly controlled because of its apoptotic properties. Two silencer elements (ESS3 and ISS) and two enhancer elements (ESE2 and ESE3/ (GAA)3) were previously identified at site A7. hnRNP A1 binds ISS and ESS3 and is involved in the inhibitory process, ASF/SF2 activates site A7 utilisation. Here, by using chemical and enzymatic probes we established the 2D structure of the HIV-1 BRU RNA region containing site A7 and identified the RNA segments protected in nuclear extract and by purified hnRNP A1. ISS, ESE3/(GAA)3 and ESS3 are located in three distinct stem-loop structures (SLS1, 2 and 3). As expected, hnRNP A1 binds sites 1, 2 and 3 of ISS and ESS3b, and oligomerises on the polypurine sequence upstream of ESS3b. In addition, we discovered an unidentified hnRNP A1 binding site (AUAGAA), that overlaps ESE3/(GAA)3. On the basis of competition experiments, hnRNP A1 has a stronger affinity for this site than for ESS3b. By insertion of (GAA)3 alone or preceded by the AUA tri- nucleotide in a foreign context, the AUAGAA sequence was found to modulate strongly the (GAA)3 splicing enhancer activity. Cross-linking experiments on these heterologous RNAs and the SLS2-SLS3 HIV-1 RNA region, in nuclear extract and with recombinant proteins, showed that binding of hnRNP A1 to AUA(GAA)3 strongly competes the association of ASF/SF2 with (GAA)3. In addition, disruption of AUA(GAA)3 demon- strated a key role of this sequence in hnRNP A1 cooperative binding to the ISS and ESS3b inhibitors and hnRNP A1 oligomerisation on the polypur- ine sequence. Thus, depending on the cellular context ([ASF/SF2]/ [hnRNP A1] ratio), AUA(GAA)3 will activate or repress site A7 utilisation and can thus be considered as a Janus splicing regulator. q 2002 Elsevier Science Ltd. All rights reserved Keywords: virus HIV-1; splicing regulation; Tat protein; ASF/SF2; hnRNP A1 *Corresponding author Introduction As a unique transcript is produced by transcrip- tion of the integrated proviral cDNA of retro- viruses, regulation of expression of the retroviral proteins occurs at the splicing and/or translation step(s). Regulation of splicing is very important for retroviruses as part of the transcript has to be transported to the cytoplasm in an intact form, to serve as genome for the production of new virions. To achieve such equilibrium between spliced and unspliced transcripts, retroviruses generally have RNA with 3 0 -splice sites (3 0 ss) of low efficiency 0022-2836/02/$ - see front matter q 2002 Elsevier Science Ltd. All rights reserved Present addresses: S. Jacquenet, Unite ´ de Virologie humaine (U412 INSERM), Ecole Normale Supe ´rieure de Lyon, 46 alle ´e d’Italie, 69007 Lyon, France; A. Mougin, Laboratoire de Biologie Mole ´culaire Eucaryote UMR5099-CNRS, Universite ´ Paul Sabatier, 118 route de Narbonne, 31062 Toulouse Cedex 4, France. E-mail address of the corresponding author: [email protected] Abbreviations used: HIV-1, human immunodeficiency virus type 1; 3 0 ss, 3 0 splice site; ISS, intronic splicing silencer; ESS, exonic splicing silencer; hnRNP, heterogeneous ribonucleoprotein; 5 0 ss, 5 0 splice site; SLS, stem-loop structure; PPT, polypyrimidine tract. doi:10.1016/S0022-2836(02)00967-1 available online at http://www.idealibrary.com on B w J. Mol. Biol. (2002) 323, 629–652

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A Janus Splicing Regulatory Element Modulates HIV-1tat and rev mRNA Production by Coordination ofhnRNP A1 Cooperative Binding
Virginie Marchand1, Agnes Mereau1, Sandrine Jacquenet1
Denise Thomas1, Annie Mougin1, Renata Gattoni2, James Stevenin2
and Christiane Branlant1*
1Laboratoire de Maturation desARN et EnzymologieMoleculaire, UMR 7567UHP-CNRS, Universite HenriPoincare Nancy 1, Boulevarddes Aiguillettes, BP239, 54506Vandoeuvre-les-Nancy CedexFrance
2IGBMC, 1 rue Laurent FriesBP 163, Parc d’ innovation67404 Illkirch Cedex, France
Retroviral protein production depends upon alternative splicing of theviral transcript. The HIV-1 acceptor site A7 is required for tat and revmRNA production. Production of the Tat transcriptional activator ishighly controlled because of its apoptotic properties. Two silencerelements (ESS3 and ISS) and two enhancer elements (ESE2 and ESE3/(GAA)3) were previously identified at site A7. hnRNP A1 binds ISS andESS3 and is involved in the inhibitory process, ASF/SF2 activates site A7utilisation. Here, by using chemical and enzymatic probes we establishedthe 2D structure of the HIV-1BRU RNA region containing site A7 andidentified the RNA segments protected in nuclear extract and by purifiedhnRNP A1. ISS, ESE3/(GAA)3 and ESS3 are located in three distinctstem-loop structures (SLS1, 2 and 3). As expected, hnRNP A1 binds sites1, 2 and 3 of ISS and ESS3b, and oligomerises on the polypurine sequenceupstream of ESS3b. In addition, we discovered an unidentified hnRNP A1binding site (AUAGAA), that overlaps ESE3/(GAA)3. On the basis ofcompetition experiments, hnRNP A1 has a stronger affinity for this sitethan for ESS3b. By insertion of (GAA)3 alone or preceded by the AUA tri-nucleotide in a foreign context, the AUAGAA sequence was found tomodulate strongly the (GAA)3 splicing enhancer activity. Cross-linkingexperiments on these heterologous RNAs and the SLS2-SLS3 HIV-1 RNAregion, in nuclear extract and with recombinant proteins, showed thatbinding of hnRNP A1 to AUA(GAA)3 strongly competes the associationof ASF/SF2 with (GAA)3. In addition, disruption of AUA(GAA)3 demon-strated a key role of this sequence in hnRNP A1 cooperative binding to theISS and ESS3b inhibitors and hnRNP A1 oligomerisation on the polypur-ine sequence. Thus, depending on the cellular context ([ASF/SF2]/[hnRNP A1] ratio), AUA(GAA)3 will activate or repress site A7 utilisationand can thus be considered as a Janus splicing regulator.
q 2002 Elsevier Science Ltd. All rights reserved
Keywords: virus HIV-1; splicing regulation; Tat protein; ASF/SF2; hnRNP A1*Corresponding author
Introduction
As a unique transcript is produced by transcrip-tion of the integrated proviral cDNA of retro-viruses, regulation of expression of the retroviralproteins occurs at the splicing and/or translationstep(s). Regulation of splicing is very importantfor retroviruses as part of the transcript has to betransported to the cytoplasm in an intact form, toserve as genome for the production of new virions.To achieve such equilibrium between spliced andunspliced transcripts, retroviruses generally haveRNA with 30-splice sites (30ss) of low efficiency
0022-2836/02/$ - see front matter q 2002 Elsevier Science Ltd. All rights reserved
Present addresses: S. Jacquenet, Unite de Virologiehumaine (U412 INSERM), Ecole Normale Superieure deLyon, 46 allee d’Italie, 69007 Lyon, France; A. Mougin,Laboratoire de Biologie Moleculaire EucaryoteUMR5099-CNRS, Universite Paul Sabatier, 118 route deNarbonne, 31062 Toulouse Cedex 4, France.
E-mail address of the corresponding author:[email protected]
Abbreviations used: HIV-1, human immunodeficiencyvirus type 1; 30ss, 30 splice site; ISS, intronic splicingsilencer; ESS, exonic splicing silencer; hnRNP,heterogeneous ribonucleoprotein; 50ss, 50 splice site; SLS,stem-loop structure; PPT, polypyrimidine tract.
doi:10.1016/S0022-2836(02)00967-1 available online at http://www.idealibrary.com onBw
J. Mol. Biol. (2002) 323, 629–652

(suboptimal polypyrimidine tracts (PPT) andbranch-site sequences (BS), intronic or exonic splic-ing silencer elements (ISS or ESS, respectively)).1 – 8
Transcripts of the complex HIV-1 retrovirus con-tain four 50-splice sites (50ss) and eight 30ss, whichare used in combination to produce about 40 dis-tinct mRNAs.9 Among the eight HIV 30ss, site A7plays a very important role, since its utilisation incombination with the D4 50ss is required for pro-duction of the rev and tat mRNAs. Site A7 delimitsthe third exon of Tat and Rev proteins. Expressionof the Tat transcriptional activator and the Rev pro-tein that is implicated in the transport of unsplicedor singly spliced viral RNAs is necessary for virusmultiplication.10,11 The Tat protein is required fortranscription of full-length HIV-1 RNA,9 that isused as messenger RNAs or as genomic RNA.However, the Tat protein has apoptotic propertiesand can be exported out of the infected cell maybeafter their apoptosis.10,12,13 Tat is responsible for alarge part of the HIV-1 destructive activity againstthe host organism.14 Thus, HIV-1 has to limit tatmRNA production. This is done in part at thesplicing step through downregulation of site A3and A7 utilisation.6,7,15 – 19
Several studies have already been performed onsite A7 regulation. Two cis inhibitory elementshave been described: a bipartite one, ESS3, that islocated in the second exon,6,7 and the ISS element,which is located in the intron 30-terminal part.8
Heterogeneous ribonucleoprotein (hnRNP) A1was found to bind both silencer elements, and asynergistic binding to the two silencer elementswas proposed.8,18 hnRNP A1 binds the ESS3bmotif of ESS3, but the nuclear partner(s) of ESS3amotif has not yet been identified.18 Utilisation ofpoorly efficient 30ss is generally activated byenhancer elements that bind SR proteins. By theirRS domain, SR proteins increase the efficiency ofassembly of the spliceosome components at theregulated sites.20 – 22 At the HIV-1 A7 30ss, the SRprotein SC35 was proposed to bind the segmentlinking ESS3a to ESS3b.23 In addition, two enhancerelements ESE2 and (GAA)3 (previously designatedESE3) have also been described for site A76,18,19 andthe ASF/SF2 SR protein was found to activate theutilisation of site A7 in vitro.18,19,23,24 However, noclear relationship between the two enhancerelements and ASF/SF2 was established yet, andthe ESE3 element seemed more complex thanexpected.19
hnRNP A1 is one of the more abundant hnRNPproteins.25 It was already shown to be implicatedin the regulation of the alternative utilisation ofnumerous 50 and 30ss.16,26 – 32 In several cases,protein hnRNP A1 was found to have one or twostrong binding site(s) in the vicinity of the regu-lated splicing site and sometimes hnRNP A1 wasshown to multimerise along the RNA region con-taining the regulatory site(s).18,26,27 When the twobinding sites are located far apart in the RNAmolecule, as is the case for the hnRNP A1 pre-mRNA, an RNA loop can be generated by inter-
action of two hnRNP A1 proteins bound at thetwo distant sites.31 Regulation of alternative 50ssutilisation by hnRNP A1 generally results from acompetition between hnRNP A1 and SR proteinsfor binding to the 50ss.27,33 However, the mechan-isms of inhibition of 30ss utilisation by hnRNP A1are much less understood. hnRNP A1 plays a veryimportant role in the regulation of HIV-1 RNAsplicing, since it is implicated in the down-regulation of the utilisation of three of the 30ss(A2, A3 and A7).8,16,18,26
In an attempt to get further information on themechanism of inhibition of the utilisation of theHIV-1 A7 30ss by hnRNP A1 and with a view toestablish relationships between the opposite effectsof hnRNP A1 and ASF/SF2 at site A7, we first per-formed an experimental analysis of the secondarystructure of the RNA region containing site A7and all its regulatory elements. Then, we identifiedthe hnRNP A1 binding sites on the establishedstructure using footprinting experiments, gel-shiftand cross-linking assays. These experiments wereperformed on wild-type (WT) or mutated HIV-1RNA fragments. New hnRNP A1 binding siteswere discovered by this approach, one of themoverlaps the ESE3/(GAA)3 enhancer element andmodulates its activity. As elimination of thishnRNP A1 binding site strongly decreased hnRNPA1 binding at the other sites, this newly identifiedbinding site seems to play a very important role inthe downregulation of site A7 activity.
Results
The HIV-1 RNA region containing site A7 andits regulatory elements forms three stem-loop structures
Because RNA secondary structure may modu-late the activity of splicing cis regulatoryelements,34 – 37 we studied the secondary structureof the HIV-1BRU RNA region that contains theacceptor site A7 and its cis regulatory elements(segment from position 7887 to 8072 in the HIV-1BRU RNA) (Figure 1). As HIV-1 RNA is too largefor a direct experimental analysis of its secondarystructure, experiments were performed on RNAfragments. To be convinced that the identifiedstructural elements were not artifacts generated bythe RNA fragmentation, the analyses were per-formed on overlapping fragments of differentlengths. Four transcripts were studied (Figure 1):RNA L47-8072 (HIV-1BRU RNA segments 5577–5676 and 7692–8072 joined together) contained amini D4-A7 intron and all the described regulatoryelements acting on site A7 (ISS upstream of A7,8
ESE2, ESE3/(GAA)3 and ESS3 downstream fromA7).6,7,15,18,19 RNA L47-8043 and RNA L47-8490also contained the mini D4-A7 intron, but RNAL47-8043 was lacking ESS3, whereas RNA L47-8490 had an extra 30-terminal segment. RNA A7SLS1,2,3 (positions 7903–8170) started upstream of
630 Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss

ISS in the mini D4-A7 intron and contained all theA7 cis regulatory elements.
We used two chemical reagents acting atWatson–Crick positions of residues located insingle-stranded segments, namely, dimethylsulfate(DMS) (N1-A and N3-C) and 1-cyclohexyl-3-(2-morpholinoethyl)-carbodiimide-metho-p-toluenesulfonate (CMCT) (N1-G and N3-U). We also used:T1 and T2 RNases in conditions where they prefer-entially cleave single-stranded regions, the single-strand-specific S1 nuclease, as well as V1 RNase,which cleaves preferentially double-stranded andstacked RNA regions. It was important to studyRNAs in conditions used for in vitro splicing assays(buffer D at 3.125 mM Mg2þ, see Materials andMethods). Thus, one series of reactions includingDMS modification, T1, T2 and V1 RNase digestionswas performed in these conditions. However, dueto the poor efficiency of CMCT and S1 nuclease inthese conditions,38 a second series of analyses,including CMCT, DMS modifications, V1 and S1
nuclease digestions was performed in the presenceof 10 mM Mg2þ using classical conditions for RNAsecondary structure analysis.35 Positions ofcleavages and modifications were identified byprimer extension analyses using three differentprimers (O-2156, O-609 and O-2382), which aredescribed in Table 1. These experiments wererepeated several times for each RNA and repro-ducible results were obtained. Figure 2(a) and (b)shows representative examples of analyses per-formed on the L47-8043 RNA using classical con-ditions for RNA secondary structure analysis
(10 mM Mg2þ) and on the A7 SLS1,2,3 RNA inbuffer D, respectively. The data obtained wereused to predict RNA secondary structure with theMfold software of the GCG package. One second-ary structure model was compatible with the twosets of data. It contains three stem-loop structures,that we denoted SLS1, SLS2 and SLS3 (Figure 2(c)and (d)). Their calculated stability (Mfold software,see Materials and Methods) is of 212.6 kcal/mol,217.5 kcal/mol and 27.7 kcal/mol, respectively.Data of probing experiments illustrated in Figure2(a) (L47-8043 RNA, 10 mM Mg2þ) and Figure 2(b)(A7 SLS1,2,3 RNA, 3.125 mM Mg2þ) are schemati-cally represented on the proposed secondary struc-ture in Figure 2(c) and (d), respectively. WhereasSLS3 shows a rather classical helical structurewith only one G·U base-pair and an internal ACpair, SLS1 and SLS2 have peculiar characteristicfeatures. SLS1 contains a block of three non-Watson–Crick U·G pairs, which form a helicalstructure on the basis of the observed V1 RNasecleavages. Some instability of the U·G pair (7911–7949) is shown by the mild T1 RNase cleavage 30
to G7949. The enzymatic probing on SLS2 suggeststhe presence of several GA and AG sheared pairsin addition to the Watson–Crick base-pairs.Indeed, the V1 RNase cleavages found 30 to G8008and the absence of T1 RNase cleavages 30 to this Gresidue and G residue 7989 (Figure 2(d)) stronglysuggest the formation of two successive GA andAG sheared pairs in the prolongation of the threeG-C pairs (7991–7993, 8005–8007). Similarly, thevery limited level of T1 RNase cleavages in the
Figure 1. Schematic representation of the HIV-1 genome and the RNAs used in this study. The HIV-1 genome isshown in the upper part of the Figure. The 50ss (D1–D4) and the 30ss (A1–A7) are indicated. Boxes represent thedifferent open reading frames. The various RNA transcripts used here are represented below, with their names andutilisation on the left. Numbering of HIV-1BRU RNA sequences is as by Ratner et al.51 Exon sequences are representedby rectangles, introns by thin lines, sequences from plasmid pBluescript KSIIþ at the beginning of the transcripts areshown by thick horizontal lines. The exonic cis regulatory elements (ESE2, ESE3/(GAA)3, ESS3) are symbolised byvertical rectangles (dark grey, grey and white, respectively), the intronic ISS element is shown by a white circle.
Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss 631
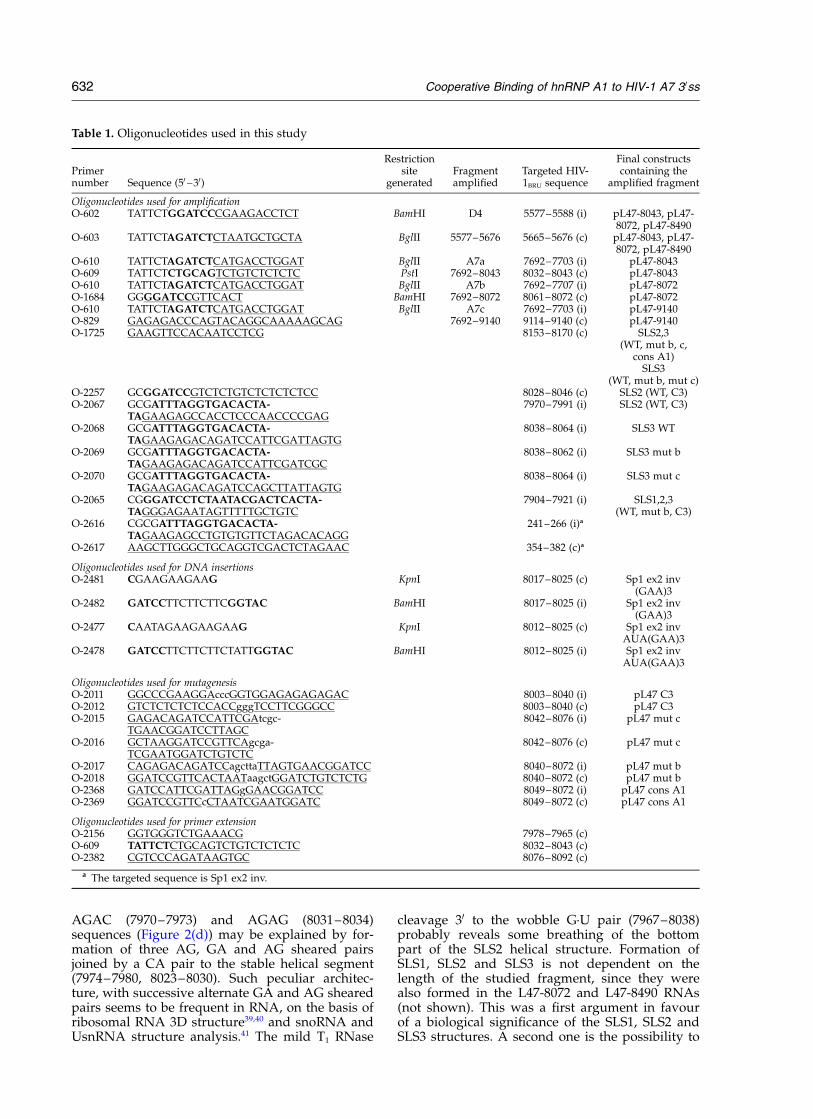
AGAC (7970–7973) and AGAG (8031–8034)sequences (Figure 2(d)) may be explained by for-mation of three AG, GA and AG sheared pairsjoined by a CA pair to the stable helical segment(7974–7980, 8023–8030). Such peculiar architec-ture, with successive alternate GA and AG shearedpairs seems to be frequent in RNA, on the basis ofribosomal RNA 3D structure39,40 and snoRNA andUsnRNA structure analysis.41 The mild T1 RNase
cleavage 30 to the wobble G·U pair (7967–8038)probably reveals some breathing of the bottompart of the SLS2 helical structure. Formation ofSLS1, SLS2 and SLS3 is not dependent on thelength of the studied fragment, since they werealso formed in the L47-8072 and L47-8490 RNAs(not shown). This was a first argument in favourof a biological significance of the SLS1, SLS2 andSLS3 structures. A second one is the possibility to
Table 1. Oligonucleotides used in this study
Primernumber Sequence (50 –30)
Restrictionsite
generatedFragmentamplified
Targeted HIV-1BRU sequence
Final constructscontaining the
amplified fragment
Oligonucleotides used for amplificationO-602 TATTCTGGATCCCGAAGACCTCT BamHI D4 5577–5588 (i) pL47-8043, pL47-
8072, pL47-8490O-603 TATTCTAGATCTCTAATGCTGCTA BglII 5577–5676 5665–5676 (c) pL47-8043, pL47-
8072, pL47-8490O-610 TATTCTAGATCTCATGACCTGGAT BglII A7a 7692–7703 (i) pL47-8043O-609 TATTCTCTGCAGTCTGTCTCTCTC PstI 7692–8043 8032–8043 (c) pL47-8043O-610 TATTCTAGATCTCATGACCTGGAT BglII A7b 7692–7707 (i) pL47-8072O-1684 GGGGATCCGTTCACT BamHI 7692–8072 8061–8072 (c) pL47-8072O-610 TATTCTAGATCTCATGACCTGGAT BglII A7c 7692–7703 (i) pL47-9140O-829 GAGAGACCCAGTACAGGCAAAAAGCAG 7692–9140 9114–9140 (c) pL47-9140O-1725 GAAGTTCCACAATCCTCG 8153–8170 (c) SLS2,3
(WT, mut b, c,cons A1)
SLS3(WT, mut b, mut c)
O-2257 GCGGATCCGTCTCTGTCTCTCTCTCC 8028–8046 (c) SLS2 (WT, C3)O-2067 GCGATTTAGGTGACACTA-
TAGAAGAGCCACCTCCCAACCCCGAG7970–7991 (i) SLS2 (WT, C3)
O-2068 GCGATTTAGGTGACACTA-TAGAAGAGACAGATCCATTCGATTAGTG
8038–8064 (i) SLS3 WT
O-2069 GCGATTTAGGTGACACTA-TAGAAGAGACAGATCCATTCGATCGC
8038–8062 (i) SLS3 mut b
O-2070 GCGATTTAGGTGACACTA-TAGAAGAGACAGATCCAGCTTATTAGTG
8038–8064 (i) SLS3 mut c
O-2065 CGGGATCCTCTAATACGACTCACTA-TAGGGAGAATAGTTTTTGCTGTC
7904–7921 (i) SLS1,2,3(WT, mut b, C3)
O-2616 CGCGATTTAGGTGACACTA-TAGAAGAGCCTGTGTGTTCTAGACACAGG
241–266 (i)a
O-2617 AAGCTTGGGCTGCAGGTCGACTCTAGAAC 354–382 (c)a
Oligonucleotides used for DNA insertionsO-2481 CGAAGAAGAAG KpnI 8017–8025 (c) Sp1 ex2 inv
(GAA)3O-2482 GATCCTTCTTCTTCGGTAC BamHI 8017–8025 (i) Sp1 ex2 inv
(GAA)3O-2477 CAATAGAAGAAGAAG KpnI 8012–8025 (c) Sp1 ex2 inv
AUA(GAA)3O-2478 GATCCTTCTTCTTCTATTGGTAC BamHI 8012–8025 (i) Sp1 ex2 inv
AUA(GAA)3
Oligonucleotides used for mutagenesisO-2011 GGCCCGAAGGAcccGGTGGAGAGAGAGAC 8003–8040 (i) pL47 C3O-2012 GTCTCTCTCTCCACCgggTCCTTCGGGCC 8003–8040 (c) pL47 C3O-2015 GAGACAGATCCATTCGAtcgc-
TGAACGGATCCTTAGC8042–8076 (i) pL47 mut c
O-2016 GCTAAGGATCCGTTCAgcga-TCGAATGGATCTGTCTC
8042–8076 (c) pL47 mut c
O-2017 CAGAGACAGATCCagcttaTTAGTGAACGGATCC 8040–8072 (i) pL47 mut bO-2018 GGATCCGTTCACTAATaagctGGATCTGTCTCTG 8040–8072 (c) pL47 mut bO-2368 GATCCATTCGATTAGgGAACGGATCC 8049–8072 (i) pL47 cons A1O-2369 GGATCCGTTCcCTAATCGAATGGATC 8049–8072 (c) pL47 cons A1
Oligonucleotides used for primer extensionO-2156 GGTGGGTCTGAAACG 7978–7965 (c)O-609 TATTCTCTGCAGTCTGTCTCTCTC 8032–8043 (c)O-2382 CGTCCCAGATAAGTGC 8076–8092 (c)
a The targeted sequence is Sp1 ex2 inv.
632 Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss

form structures showing common features with theBRU SLS1, 2 and 3 in RNAs of HIV-1 strains of thevarious clades of group M (Figure 3). Due to a highdegree of primary structure conservation, SLS1 ishighly conserved. The few base substitutions com-pared to BRU RNA (red nucleotides in Figure3(b)) that we observed in the collection of studiedstrains do not disturb the stability of the helices.This high sequence conservation of SLS1 shouldreflect a functional importance, since this con-served part of HIV-1 RNA codes for only oneprotein, Env, whereas the more variable SLS2 andSLS3 regions code for three proteins (Tat, Rev andEnv). In SLS2 and SLS3, the only highly conservedelements are: the CAGACCC sequence containingsite A7, the GAAGAAG sequence in the ESE3/(GAA)3 regulatory element, the CCGACAGGsequence that forms the terminal part of SLS2 andwhich is contained in a segment previously pro-posed to be an enhancer element19 and the ESS3sequence of the inhibitory element. However, thepossibility to form a short SLS3 structure and anirregular SLS2 structure with site A7 in an internalloop, two GAA triplets of the (GAA)3 motif in alarge internal loop and a GACA tetraloop closedby two G-C pairs, is rather well preserved ingroup M (Figure 3(b)). Hence, experimental dataand phylogenetic comparisons support the ideathat, in the group M of HIV-1 strains, the A7 30ssis located in an irregular stem-loop structure SLS2,with an upstream stem-loop structure SLS1 thatcontains the branch-site sequences and the intronicinhibitory element ISS8 (Figure 3(a)). The exonicinhibitory element ESS3, which is composed oftwo sub-elements ESS3a and ESS3b that bind dis-tinct inhibitory components,7 is located in a smallstem-loop structure (SLS3). ESS3a is involved information of the helix, whereas ESS3b is in theterminal loop (Figure 3(a)). The two activatorelements (ESE2 and ESE3/(GAA)3), that were pre-viously described for site A7,6,19 are located in theterminal structure and the large internal loop ofSLS2, respectively (Figure 3(a)). Finally, the poly-pyrimidine tract (PPT) of site A7 is only partiallybase-paired (bottom helix of SLS2).
Nuclear extract and purified hnRNP A1 protectlarge parts of SLS1, SLS2 and SLS3
Our structural analysis showed that several ofthe sequence elements found to regulate site A7utilisation are single-stranded in the naked A7RNA region (part of ISS, part of ESE2, two GAAmotifs, ESS3b). It was thus interesting to testwhether the sensitivity to T1 RNase of some ofthese single-stranded segments was abolished orlimited in a nuclear extract, as a result of the bind-ing of nuclear components. For an estimation ofthe protein/RNA ratio that had to be used for T1
RNase footprinting experiments in nuclear extract,we first performed gel-shift assays using nuclearextract and a small RNA, that contained stem-loopstructures 2 and 3 only (SLS2,3 RNA) (Figure 1).
As illustrated in Figure 4(a), in the conditionsused (see Materials and Methods), the 30-endlabelled RNA was totally shifted at a [protein]/[SLS2,3 RNA] ratio of 10.4 (mg/pmol). Takingthese data into account, a [protein]/[SLS1,2,3RNA] ratio of 46 (mg/pmol) was selected to probethe complexes formed by SLS1,2,3 RNA and com-ponents of the nuclear extract. After T1 RNasetreatment of the naked RNA and RNP complexes(see conditions in Materials and Methods), pos-itions and yields of cleavages were estimated byextension of primer 2382 with reverse transcriptase(Figure 4(b)). As schematically represented inFigure 4(b) and (c), the strongest protections werefound in: (i) the terminal loop of SLS3 (ESS3binhibitory element), (ii) the ESE3/(GAA)3 elementconsidered as an activator element, and (iii) theterminal loop of SLS1 (ISS inhibitory element).More limited protections were also detected in theterminal loop of SLS2 (ESE2 activator element)and the purine-rich sequence that is located fromposition 8036 to 8044. Especially, the cleavageoccurring 30 to G8038 was markedly reduced(Figure 4(b) and (c)). A faint protection of the Gresidue 7965 in the polypyrimidine tract was alsodetected (Figure 4(b) and (c)). Taken together,these data demonstrated that the three splicingregulatory elements (ISS, ESE3/(GAA)3 andESS3b) constitute strong binding sites for nuclearcomponents. Site A7, the PPT, and ESE2 have alower affinity for nuclear components. The conser-vation of the patterns of T1 RNase accessibility inthe long helical region of SLS1, most of the helicalregions of SLS2 (except the bottom helix) and theSLS3 helical region, show that binding of nuclearcomponents does not alter RNA secondary struc-ture. Concerning the bottom helix of SLS2, bothstrands were protected by components of nuclearextract, suggesting that they remain associatedupon binding of nuclear component.
Using cross-linking experiments and gel-shiftassays, protein hnRNP A1 was previously shownto interact with the ISS and ESS3 inhibitoryelements.8,18 To determine which parts of the pro-tections observed in nuclear extract were due tohnRNP A1, we produced a soluble recombinanthnRNP A1 protein without any tag sequence andprobed A7 SLS1,2,3 RNA/hnRNP A1 complexes.To get an idea of the [hnRNP A1]/[SLS1,2,3 RNA]molar ratio that had to be used for footprintingexperiments, gel-shift assays were first performedwith increasing amounts of hnRNP A1 and asmall RNA (A7 SLS2,3 RNA) (Figure 1), that con-tains an hnRNP A1 binding site in ESS3.18 RNA/protein complexes with decreasing electrophoreticmobilities were obtained upon increasing thehnRNP A1 concentration. This suggests the pre-sence of several hnRNP A1 binding sites withdifferent affinities for hnRNP A1 in SLS2,3 RNA,that were progressively saturated upon increasingthe protein concentration. In the conditions used(see Materials and Methods), a complete shift ofthe 30-end labelled RNA was obtained for a 20-fold
Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss 633
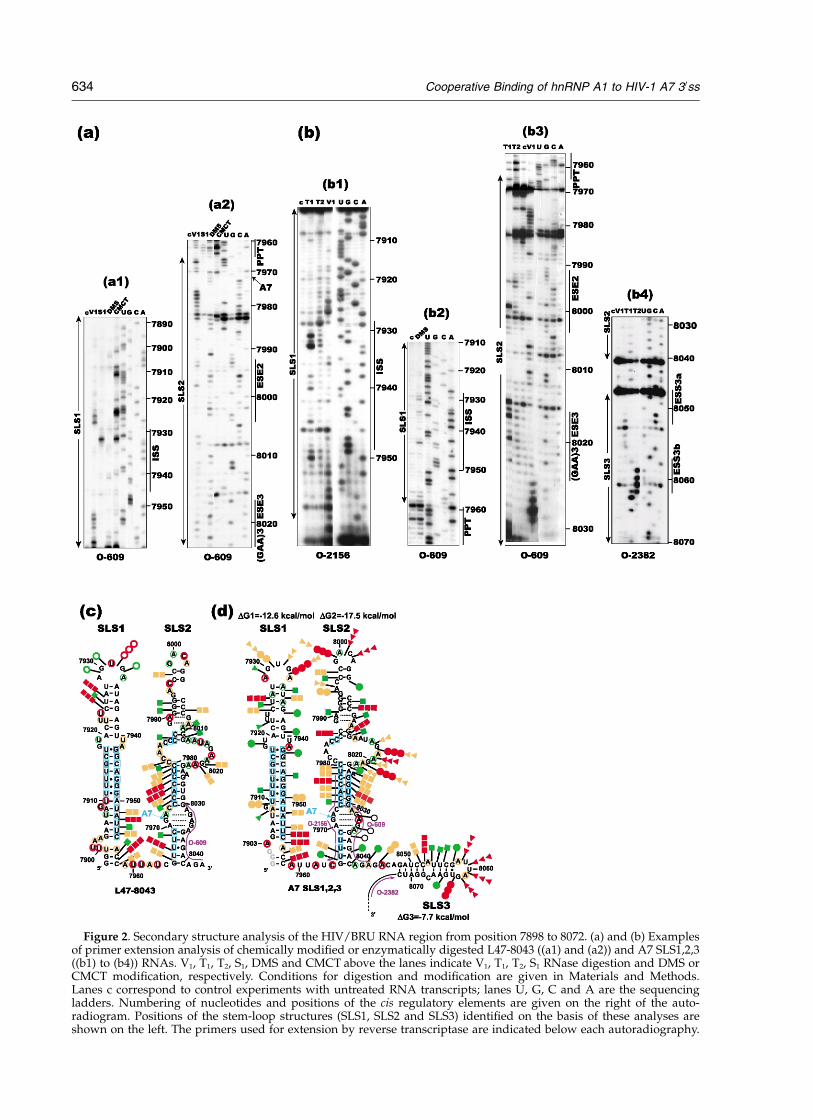
Figure 2. Secondary structure analysis of the HIV/BRU RNA region from position 7898 to 8072. (a) and (b) Examplesof primer extension analysis of chemically modified or enzymatically digested L47-8043 ((a1) and (a2)) and A7 SLS1,2,3((b1) to (b4)) RNAs. V1, T1, T2, S1, DMS and CMCT above the lanes indicate V1, T1, T2, S1 RNase digestion and DMS orCMCT modification, respectively. Conditions for digestion and modification are given in Materials and Methods.Lanes c correspond to control experiments with untreated RNA transcripts; lanes U, G, C and A are the sequencingladders. Numbering of nucleotides and positions of the cis regulatory elements are given on the right of the auto-radiogram. Positions of the stem-loop structures (SLS1, SLS2 and SLS3) identified on the basis of these analyses areshown on the left. The primers used for extension by reverse transcriptase are indicated below each autoradiography.
634 Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss

molar excess of protein compared to RNA (Figure5(a)). Taking these data into consideration, RNPcomplexes between hnRNP A1 and the SLS1,2,3RNA were formed at a [hnRNP A1]/[RNA] ratioof 45 and footprinting experiments with T1 RNasewere performed as described in Materials andMethods. Oligonucleotides 2382 and 2156 wereused for primer extension analyses. Large parts ofthe A7 SLS1,2,3 RNA were found to be protectedby hnRNP A1. The SLS3 terminal loop was
strongly protected, which confirmed the bindingof hnRNP A1 to the ESS3b inhibitory element.18
The SLS1 terminal loop and G residues 7937, 7939and 7944 in the SLS1 30-strand were partially pro-tected, which is in agreement with the binding ofhnRNP A1 to the putative sites 1, 2 and 3 pre-viously proposed in the ISS element.8 Interestingly,in SLS2, three G residues in the purine-richsequence (positions 8038, 8042 and 8044) and thesecond and third G residues (positions 8020 and
Figure 3. Conservation of SLS1, SLS2 and SLS3 stem-loop structures in the group M of HIV-1 strains. (a) The second-ary structure proposed for the HIV-1BRU RNA region from positions 7898 to 8072. Positions of nucleotides in HIV-1BRU
RNA are given. The splicing cis regulatory elements are indicated by nucleotides in coloured circles. The A7 30ss isindicated by an arrow, the three proposed branch sites residues (BP1, BP2 and BP3)8 are circled in red. The polypyrimi-dine tract (PPT) is indicated by squared nucleotides. (b) SLS1, 2 and 3 are drawn for strains of the group M of HIV-1that belong to the A, C, D, F, G, H and J clades. Nucleotide sequences are from the HIV sequence database (Los AlamosNational Laboratory, Los Alamos, NM: http://hiv.web.lanl.gov/). The HIV-1 RNA regions were identified byalignment of the HIV-1BRU RNA sequence with sequences from other strains using the ClustalW program.58 The basesubstitutions as compared to HIV-1BRU RNA are indicated by red nucleotides, respectively. The nucleotides from theISS, PPT, A7 site, ESE2, ESE3/(GAA)3, ESS3a and ESS3b motifs, which are conserved in the studied strains comparedto the BRU strain, are shown in bold characters. The triplet 50 to ESE3/(GAA)3 that modulates ESE3 activity in theBRU strain is indicated in a rectangle.
(c) and (d) Schematic representation of the experimental data on the secondary structure model proposed for L47-8043(c) and A7 SLS1,2,3 (d) RNAs. Cleavages of phosphodiester bonds by V1 RNase ((c) and (d)), S1 nuclease (c), T1 RNaseand T2 RNase (d) are represented by arrows surmounted on squares, open circles (c), filled coloured or white circles (d)and triangles (d), respectively. Three red squares, circles or triangles indicate a strong cleavage, two orange a mediumcleavage, one green a low cleavage. In the case of T1 RNase, black open circles (d) show very low cleavages. Nucleo-tides modified by DMS or CMCT are circled and colours of circles (red, orange and green) indicate the intensity ofmodification as above. Where we got a clear evidence of the absence of chemical modification of a given residue, thisresidue is indicated with a blue square. Stem-loop structures are numbered. The hybridisation sites of the oligonucleo-tides used for primer extension are indicated. The free energy of the proposed stem-loop structures at 37 8C in 1 MNaCl was calculated with the Mfold software. In (d), the three 50-terminal G residues coloured in grey were generatedby T7 RNA polymerase transcription. Analysis with chemical probe was not done for SLS2 from positions 7965 to 7999.
Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss 635
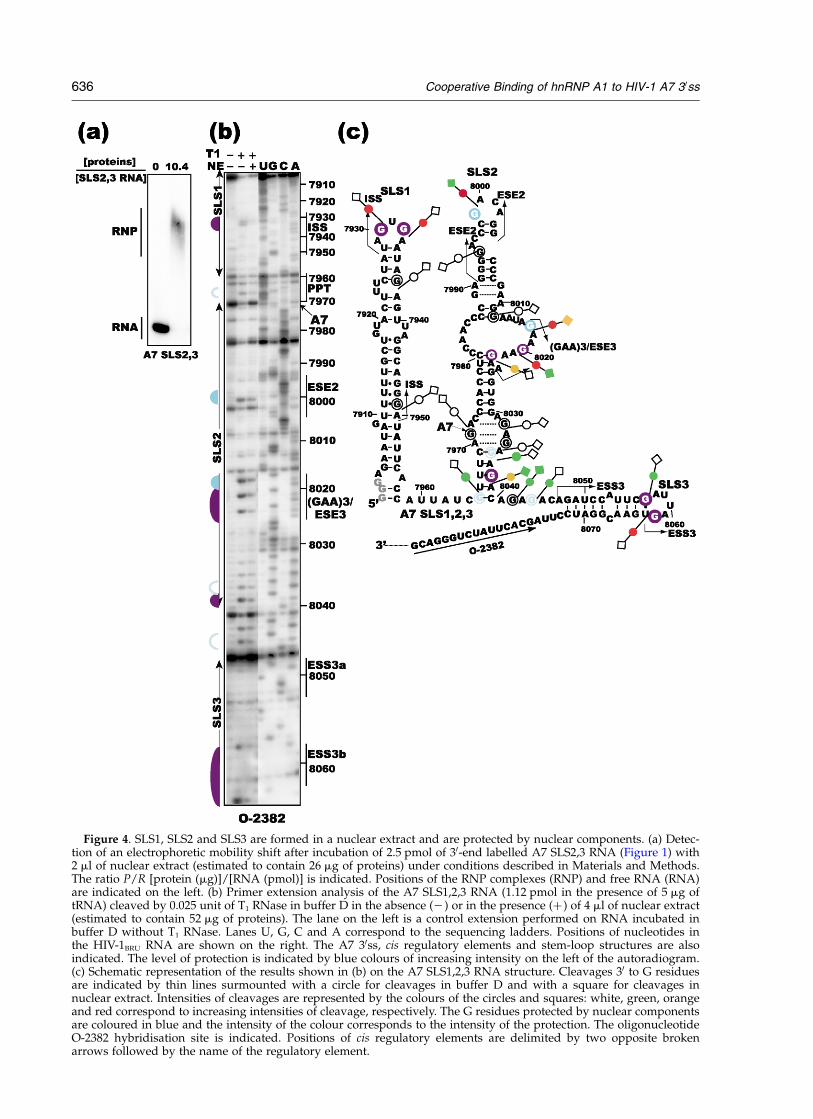
Figure 4. SLS1, SLS2 and SLS3 are formed in a nuclear extract and are protected by nuclear components. (a) Detec-tion of an electrophoretic mobility shift after incubation of 2.5 pmol of 30-end labelled A7 SLS2,3 RNA (Figure 1) with2 ml of nuclear extract (estimated to contain 26 mg of proteins) under conditions described in Materials and Methods.The ratio P/R [protein (mg)]/[RNA (pmol)] is indicated. Positions of the RNP complexes (RNP) and free RNA (RNA)are indicated on the left. (b) Primer extension analysis of the A7 SLS1,2,3 RNA (1.12 pmol in the presence of 5 mg oftRNA) cleaved by 0.025 unit of T1 RNase in buffer D in the absence (2) or in the presence (þ ) of 4 ml of nuclear extract(estimated to contain 52 mg of proteins). The lane on the left is a control extension performed on RNA incubated inbuffer D without T1 RNase. Lanes U, G, C and A correspond to the sequencing ladders. Positions of nucleotides inthe HIV-1BRU RNA are shown on the right. The A7 30ss, cis regulatory elements and stem-loop structures are alsoindicated. The level of protection is indicated by blue colours of increasing intensity on the left of the autoradiogram.(c) Schematic representation of the results shown in (b) on the A7 SLS1,2,3 RNA structure. Cleavages 30 to G residuesare indicated by thin lines surmounted with a circle for cleavages in buffer D and with a square for cleavages innuclear extract. Intensities of cleavages are represented by the colours of the circles and squares: white, green, orangeand red correspond to increasing intensities of cleavage, respectively. The G residues protected by nuclear componentsare coloured in blue and the intensity of the colour corresponds to the intensity of the protection. The oligonucleotideO-2382 hybridisation site is indicated. Positions of cis regulatory elements are delimited by two opposite brokenarrows followed by the name of the regulatory element.
636 Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss

8023) in the (GAA)3 motif were markedly pro-tected. Protections of the purine-rich sequence con-firmed the prediction that binding of hnRNP A1 toESS3b is followed by a cooperative binding of theprotein on the upstream RNA region.18 However,being considered as an activator element and onthe basis of its sequence, the ESE3/(GAA)3 motifwas expected to bind the SR protein ASF/SF2rather than hnRNP A1. G residue position 7965 inthe PPT was partially protected, which is in accord-ance with the protection observed in the 30-strandof the helix. Comparison of the protectionsdetected after incubation of the A7 SLS1,2,3 RNAin a nuclear extract and with purified hnRNP A1(Figures 4(b) and 5(b)), suggests that large parts ofthe protections detected in nuclear extract,especially from position 8017 to 8062, can beattributed to hnRNP A1. Note that, in contrast tothe ESE3/(GAA)3 activator, the ESE2 activatorelement was not protected by the purified hnRNP
A1 protein (G residue 7999) (Figure 5(b) and (c)).Also, as found for nuclear extract, the pattern ofT1 RNase digestion observed for the SLS1,2,3RNA/hnRNP A1 complex suggests that RNA sec-ondary structure is not altered upon hnRNP A1binding.
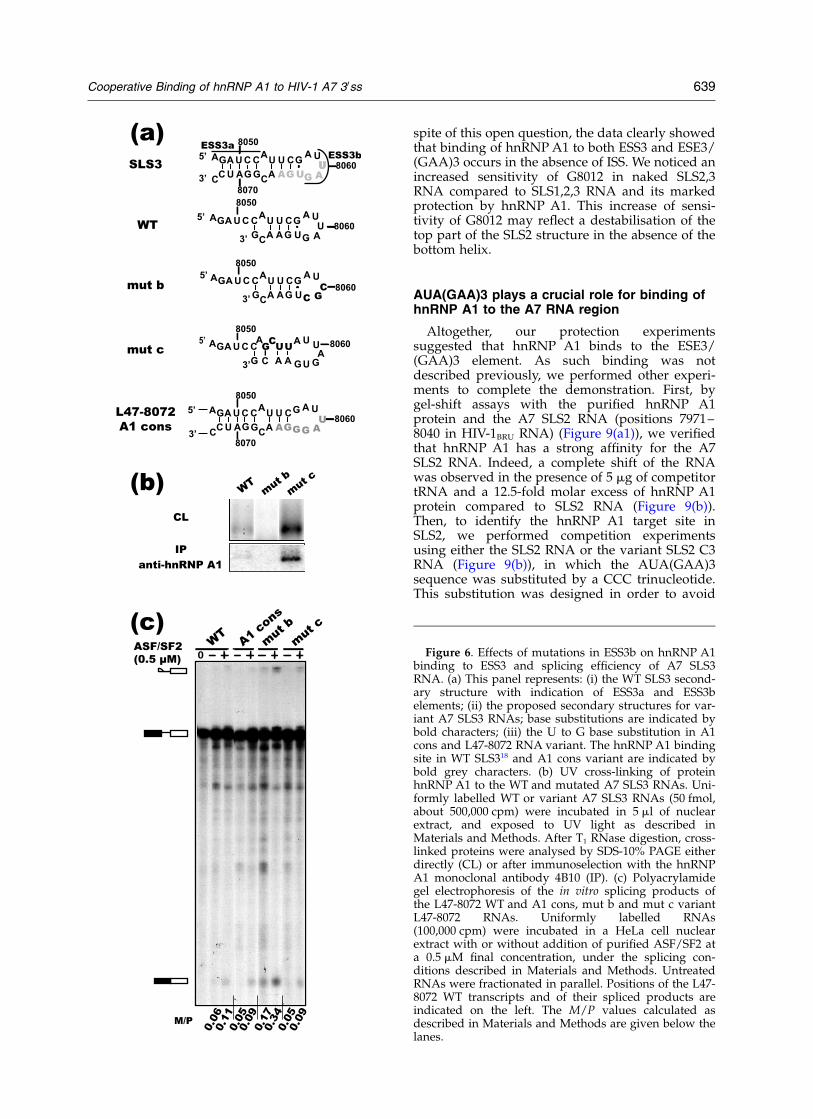
In spite of a low affinity for hnRNP A1 ofisolated SLS3, ESS3b has a marked negativeeffect on site A7 efficiency
Our probing data confirmed the presence of anhnRNP A1 binding site in SLS3. On the basis ofthe sequence of previously identified hnRNP A1binding sites, the UAGUGA sequence (positions8060–8065) of SLS3 is expected to be the hnRNPA1 target site and this is in agreement with therecent data of Zhu et al.18 However, this sequenceshows one U to G substitution compared to thewinner sequence obtained in a SELEX experiment
Figure 5. hnRNP A1 binds SLS1,SLS2 and SLS3. (a) Detection of anelectrophoretic mobility shift afterincubation of 2.5 pmol of 30-endlabelled A7 SLS2,3 RNA (Figure 1)with increasing amounts of hnRNPA1 (0, 12.5, 25, 50 pmol) under con-ditions described in Materials andMethods. For each assay, the ratioP/R [hnRNP A1]/[RNA] is indi-cated. Positions of free RNA andRNP complexes are indicated.(b) Primer extension analysis of A7SLS1,2,3 RNA (1.12 pmol in the pre-sence of 5 mg of tRNA) cleaved with0.025 unit of T1 RNase (lanesmarked by þ) in buffer D in theabsence (2) or in the presence (þ )of 50 pmol hnRNP A1 ([hnRNPA1]/[RNA] ¼ 45). As a control,primer extensions were made withthe intact RNA transcript incubatedin buffer D (without or with50 pmol of hnRNP A1). Otherindications are as in Figure 4(b).(c) Schematic representation of theexperimental data show in (b) onthe A7 SLS1,2,3 secondary struc-ture. Cleavages 30 to G residues areindicated by thin lines surmountedwith a circle for cleavages in theabsence of hnRNP A1 and with asquare for cleavages in the presenceof hnRNP A1. Intensities ofcleavages are indicated by coloursof circles and squares with thesame rules as in Figure 4(c). G resi-dues protected by hnRNP A1 arecircled in purple and blue (purple,dark blue, light blue represent astrong, medium and low level ofprotection, respectively) and theintensity is proportional to thestrength of the protection.
Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss 637

made with hnRNP A1.42 In addition, a part of thissequence is involved in the formation of the SLS3helix. We produced two variant SLS3 RNAs (HIV-1BRU RNA from position 8041 to 8068): in variantmut b the 50-terminal UAG triplet of ESS3b wassubstituted for a CGC triplet and in variant mut cthe UUCG sequence linking ESS3a to ESS3b wassubstituted by a GCUU sequence. This four-basesubstitution strongly impaired the possibility toform a helix (Figure 6(a)). Then, we tested by UVcross-linking experiments in nuclear extract, therelative affinity of the SLS3 variant and WT RNAsfor hnRNP A1. Levels of RNA/protein cross-linkswere estimated by SDS-PAGE analysis, after T1
RNase digestion of the RNA, without (CL in Figure6(b)) or with (IP in Figure 6(b)) selection on amonoclonal hnRNP A1 antibody (mAb 4B10)bound to Sepharose beads. Both approachesrevealed that, in nuclear extract, only a low levelof hnRNP A1 was cross-linked to the WT SLS3RNA. As expected no cross-link was observed forthe mut b SLS3 RNA. In contrast, a considerablyhigher level of cross-linking was observed for themut c SLS3 RNA, suggesting that the RNA helixclosing ESS3b in the WT RNA strongly decreasesESS3b accessibility to hnRNP A1. The mut b andmut c base substitutions were then generated inthe L47-8072 RNA. A U to G substitution atposition 8063, that converted ESS3b into anhnRNP A1 consensus binding site (Figure 6(a)),was also generated in this RNA. The in vitrosplicing efficiency of the mini D4-A7 intron in theWT and variant L47-8072 RNAs were tested usinga HeLa cell nuclear extract supplemented or notwith the recombinant ASF/SF2 protein (Figure6(c)). Abolition of hnRNP A1 binding to ESS3b(mut b variant) increased splicing efficiency by afactor of 3 both in the absence and the presence ofpurified ASF/SF2 (Figure 6(c)). Conversion ofESS3b into a consensus hnRNP A1 binding site(A1 cons variant) and the base substitutions in thesegment linking ESS3a to ESS3b (mut c) found toincrease hnRNP A1 binding, both had very limitedeffects on splicing efficiency in the presence orabsence of purified ASF/SF2 (Figure 6(c)).Altogether, the data suggested that the WT SLS3stem-loop structure, when taken individually, hasa limited affinity for hnRNP A1. However, withinHIV-1 RNA, this affinity is sufficient to ensurethe inhibitory property of ESS3b. A possibleexplanation was an increased affinity of hnRNPA1 for ESS3b in the entire HIV-1 RNA due to acooperative binding of hnRNP A1 to its numeroustarget sites located in the SLS1, SLS2 and SLS3structures.8,18
Binding of hnRNP A1 to ESS3b influences itsbinding to the entire A7 RNA region
To test for a possible cooperative binding ofhnRNP A1 to its various binding sites locatedaround site A7, we performed T1 RNase footprint-ing on the A7 SLS1,2,3 mut b RNA, using the
same conditions as for the A7 SLS1,2,3 RNA(Figure 5(b)). When the protections found for themut b RNA variant (Figure 7(a) and (b)) were com-pared to those detected for the A7 SLS1,2,3 RNA(Figure 5(b)), we observed that: (i) as expected, themutated terminal loop remained accessible to T1
RNase; (ii) in agreement with the prediction thatafter association with ESS3b, the binding ofhnRNP A1 propagates along the downstreamsequence,18 the protection of the purine-rich seg-ment from position 8030 to 8044 was abolished inthe mut b RNA; (iii) protection of the ESE3/(GAA)3 sequence was different compared to WTRNA: residue G8017 was more protected, whereasG residues 8020 and 8023 were less protected; (iv)finally, in accord with the proposed cooperativebinding of hnRNP A1 to ESS3 and ISS,8 the yieldof protection of the ISS terminal loop in the mut bRNA was slightly decreased. However, protectionof site 3 in the 30-strand of ISS was not modified tothe same extent as protection of sites 1 and 2. Wenoticed a very limited protection of the G residue7971 at the A7 site (Figure 7(a) and (b)), that wasnot detected for the WT RNA. Altogether, the datademonstrated a strong influence of the disruptionof ESS3b on the binding of purified hnRNP A1 toseveral of its target sites in the A7 HIV-1 RNAregion. However, protection of residue G8017shows that hnRNP A1 binding to AUAGAA inESE3 occurs in the absence of ESS3b, whereas bind-ing to the 30-terminal GAAGAA sequence of ESE3disappeared in this case. Binding at the ISS site 3also seems to be independent of ESS3b. Ourprobing data were confirmed by cross-linkingexperiments performed with the WT and mut bSLS2,3 RNAs in a nuclear extract (Figure 7(c)).Indeed, a twofold decrease of the level of hnRNPA1 cross-link was observed upon disruption ofESS3b, whereas only one hnRNP A1 binding siteamong six to eight (depending on the number ofmolecules bound to the polypurine region) wasdestroyed.
Absence of ISS does not reduce the protectionof ESS3b and ESE3/(GAA)3 by hnRNP A1
To test whether the presence of ISS is requiredfor efficient binding of hnRNP A1 to the ESS3band ESE3/(GAA)3 elements, the complexesformed between hnRNP A1 and the SLS2,3 RNA,that is missing the SLS1 structure containing theISS element, were probed with T1 RNase using thesame conditions as for the SLS1,2,3 RNA. Compari-son of data in Figures 5(b) and 8(a) reveals thatESS3b and ESE3/(GAA)3 remained as much pro-tected in the SLS2,3 RNA as in the SLS1,2,3 RNA.In the purine-rich sequence protection was lowerin SLS2,3 compared to SLS1,2,3 RNA, only the Gresidue 8044 was markedly protected. However,as the 50-strand of the bottom helix is missing, thedecreased protection observed for the purine-rich sequence could be attributed either to itssingle-stranded state or to the absence of ISS. In
638 Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss
spite of this open question, the data clearly showedthat binding of hnRNP A1 to both ESS3 and ESE3/(GAA)3 occurs in the absence of ISS. We noticed anincreased sensitivity of G8012 in naked SLS2,3RNA compared to SLS1,2,3 RNA and its markedprotection by hnRNP A1. This increase of sensi-tivity of G8012 may reflect a destabilisation of thetop part of the SLS2 structure in the absence of thebottom helix.
AUA(GAA)3 plays a crucial role for binding ofhnRNP A1 to the A7 RNA region
Altogether, our protection experimentssuggested that hnRNP A1 binds to the ESE3/(GAA)3 element. As such binding was notdescribed previously, we performed other experi-ments to complete the demonstration. First, bygel-shift assays with the purified hnRNP A1protein and the A7 SLS2 RNA (positions 7971–8040 in HIV-1BRU RNA) (Figure 9(a1)), we verifiedthat hnRNP A1 has a strong affinity for the A7SLS2 RNA. Indeed, a complete shift of the RNAwas observed in the presence of 5 mg of competitortRNA and a 12.5-fold molar excess of hnRNP A1protein compared to SLS2 RNA (Figure 9(b)).Then, to identify the hnRNP A1 target site inSLS2, we performed competition experimentsusing either the SLS2 RNA or the variant SLS2 C3RNA (Figure 9(b)), in which the AUA(GAA)3sequence was substituted by a CCC trinucleotide.This substitution was designed in order to avoid
Figure 6. Effects of mutations in ESS3b on hnRNP A1binding to ESS3 and splicing efficiency of A7 SLS3RNA. (a) This panel represents: (i) the WT SLS3 second-ary structure with indication of ESS3a and ESS3belements; (ii) the proposed secondary structures for var-iant A7 SLS3 RNAs; base substitutions are indicated bybold characters; (iii) the U to G base substitution in A1cons and L47-8072 RNA variant. The hnRNP A1 bindingsite in WT SLS318 and A1 cons variant are indicated bybold grey characters. (b) UV cross-linking of proteinhnRNP A1 to the WT and mutated A7 SLS3 RNAs. Uni-formly labelled WT or variant A7 SLS3 RNAs (50 fmol,about 500,000 cpm) were incubated in 5 ml of nuclearextract, and exposed to UV light as described inMaterials and Methods. After T1 RNase digestion, cross-linked proteins were analysed by SDS-10% PAGE eitherdirectly (CL) or after immunoselection with the hnRNPA1 monoclonal antibody 4B10 (IP). (c) Polyacrylamidegel electrophoresis of the in vitro splicing products ofthe L47-8072 WT and A1 cons, mut b and mut c variantL47-8072 RNAs. Uniformly labelled RNAs(100,000 cpm) were incubated in a HeLa cell nuclearextract with or without addition of purified ASF/SF2 ata 0.5 mM final concentration, under the splicing con-ditions described in Materials and Methods. UntreatedRNAs were fractionated in parallel. Positions of the L47-8072 WT transcripts and of their spliced products areindicated on the left. The M/P values calculated asdescribed in Materials and Methods are given below thelanes.
Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss 639

the generation of a new hnRNP A1 binding site inthe mutated RNA and to preserve RNA secondarystructure (Figure 9(a2)). Interestingly, the SLS2RNA but not the variant C3 RNA could displacehnRNP A1 from the complex formed with radio-
labelled SLS2 RNA (Figure 9(b)). This was a strongindication for an essential role of the AUA(GAA)3sequence in the binding of hnRNP A1 to SLS2. Toverify that SLS2 WT but not SLS2 C3, is bindinghnRNP A1 in a nuclear extract, UV cross-linking
Figure 7. Mutation of ESS3b decreases hnRNP A1 binding to ESS3 and ISS. (a) Primer extension analysis of A7SLS1,2,3 mut b RNA (1.12 pmol in the presence of 5 mg of tRNA) (Figure 6(a)) cleaved with 0.025 unit of T1 RNase inthe presence (þ ) or in the absence (2) of 50 pmol of hnRNP A1 ([hnRNP A1]/[RNA] ¼ 45). Oligonucleotides O-2156and O-2382 were used as primers. Same legend as for Figure 5(b). (b) Schematic representation of the experimentaldata shown in (a) on the A7 SLS1,2,3 mut b RNA structure. Levels of protections of G residues are indicated by purpleand blue colours as in Figure 5(c), * and % indicate a decreased and an increased protection compared to dataobtained with the A7 SLS1,2,3 RNA (Figure 5(c)), respectively. (c) UV cross-linking of hnRNP A1 to WT or mut bSLS2,3 RNAs. Uniformly labelled RNAs (50 fmol, about 500,000 cpm) were incubated in the presence of 5 ml of nuclearextract, and exposed to UV light as described in Materials and Methods. After T1 RNase digestion, cross-linkedproteins were immunoselected with the monoclonal antibody 4B10 directed against hnRNP A1. The eluted materialwas fractionated by SDS-10% PAGE and radiolabelled proteins hnRNP A1 were detected by autoradiography andshown in the upper part (IP). As a control of hnRNP A1 immunoselection, the proteins in the polyacrylamide gelwere transferred to a nitrocellulose membrane, probed with anti-hnRNP A1 antibodies followed by ECL detectionand shown in the lower part (Western blot). The ratio between radioactivity of the cross-linked hnRNP A1 (IP)measured by a PhosphorImager and the amount of immunoselected hnRNP A1 measured by scanning the Westernblot membrane is given below each lane.
640 Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss

experiments were performed in nuclear extract,using the uniformly labelled SLS2 WT or SLS2 C3RNAs (Figure 9(c1)). As expected, after T1 RNasedigestion, the level of radioactivity associated withhnRNP A1 was severely decreased for the SLS2 C3RNA compared to the SLS2 WT RNA. As in thetwo experiments the same amounts of hnRNP A1were retained on the antibody-coated Sepharosebeads (Western blot), the data demonstrated theprimary role of the AUA(GAA)3 sequence for thebinding of hnRNP A1 to SLS2 in splicing con-ditions. Interestingly, a very similar decrease ofhnRNP A1 binding to SLS2,3 RNA was alsoobserved when these cross-linking experimentswere repeated with the SLS2,3 WT and C3 RNAs(Figure 9(c2)). This strongly suggested that disrup-tion of the AUA(GAA)3 sequence also has an effecton the binding of hnRNP A1 to ESS3b and thepurine-rich sequence. To test for this hypothesis,we performed T1 RNase probing experiments onthe complexes formed between recombinanthnRNP A1 and the SLS1,2,3 C3 RNA using thesame conditions as for the SLS1,2,3 WT RNA. Asshown in Figure 10(a) and (b), binding of hnRNPA1 to the remaining part of SLS2 in the C3 variant,especially the purine-rich region from position8030 to 8046, was almost abolished. In addition,the protections observed for ESS3b were markedlyreduced. In SLS1, protection of sites 1 in theterminal loop and 2 in the stem were nearlyabolished. However, a protection of site 3 was stilldetected. This demonstrated that the AUA(GAA)3
sequence plays a key role for the binding ofhnRNP A1 to ESS3, the purine-rich sequence andalso sites 1 and 2 of ISS. Only binding at site 3 inISS seems to be rather independent of the ESE3/(GAA)3 presence.
ESS3b has a lower affinity for hnRNP A1compared to ESE3/(GAA)3
Having shown the importance of the ESS3b andESE3/(GAA)3 sequences for binding of hnRNP A1to the entire A7 RNA region, it was interesting tocompare the relative affinity of these two hnRNPA1 binding sites. To this end, RNA/protein com-plexes were formed with radiolabelled SLS3, SLS2and SLS2,3 WT RNAs and hnRNP A1 at increasingconcentrations, in the absence of competitortRNAs. After electrophoresis in non-denaturingconditions, no complex was detected for the SLS3WT RNA, even at an hnRNP A1/RNA ratio of 20(Figure 11(a1)), whereas SLS2 and SLS2,3 RNAswere almost totally shifted at RNA/protein ratiosof 15 and 8.4, respectively (Figure 11(a2) and (a3)).This clearly demonstrated the stronger affinity ofhnRNP A1 for SLS2 compared to SLS3 RNA. How-ever, as shown by the higher affinity of SLS2,3RNA compared to SLS2 RNA, the presence of thelow-affinity ESS3b binding site markedlyreinforces hnRNP A1 affinity through the coopera-tive binding of hnRNP A1, polymerisation to ESE3and ESS3b and its oligomerisation on the poly-purine-rich sequence. A second confirmation of
Figure 8. Protection of SLS2 andSLS3 by hnRNP A1 in the absenceof SLS1. (a) Primer extensionanalysis of A7 SLS2,3 RNA(1.51 pmol in the presence of 5 mgof tRNA) cleaved with 0.025 unit ofT1 RNase (lanes marked by þ) inbuffer D in the absence (2) or inthe presence (þ ) of 50 pmol ofhnRNP A1 ([hnRNP A1]/[RNA] ¼ 33). As controls, primerextensions were made with theintact RNA transcript incubated inbuffer D (without or with 50 pmolof hnRNP A1). Other indicationsare as in Figure 5(b). (b) Schematicrepresentation of the experimentaldata show in (a) on the A7 SLS2,3secondary structure. The protectedresidues are shown (purple andblue for strong and medium protec-tions, respectively).
Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss 641
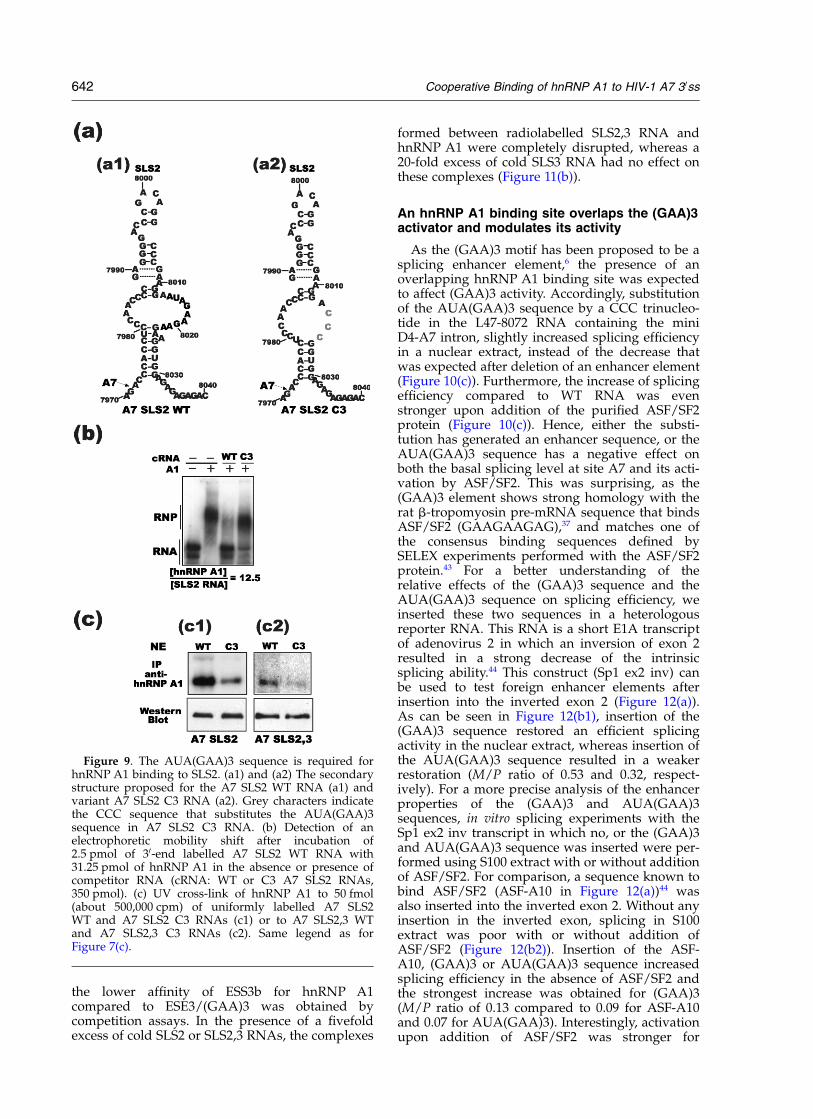
the lower affinity of ESS3b for hnRNP A1compared to ESE3/(GAA)3 was obtained bycompetition assays. In the presence of a fivefoldexcess of cold SLS2 or SLS2,3 RNAs, the complexes
formed between radiolabelled SLS2,3 RNA andhnRNP A1 were completely disrupted, whereas a20-fold excess of cold SLS3 RNA had no effect onthese complexes (Figure 11(b)).
An hnRNP A1 binding site overlaps the (GAA)3activator and modulates its activity
As the (GAA)3 motif has been proposed to be asplicing enhancer element,6 the presence of anoverlapping hnRNP A1 binding site was expectedto affect (GAA)3 activity. Accordingly, substitutionof the AUA(GAA)3 sequence by a CCC trinucleo-tide in the L47-8072 RNA containing the miniD4-A7 intron, slightly increased splicing efficiencyin a nuclear extract, instead of the decrease thatwas expected after deletion of an enhancer element(Figure 10(c)). Furthermore, the increase of splicingefficiency compared to WT RNA was evenstronger upon addition of the purified ASF/SF2protein (Figure 10(c)). Hence, either the substi-tution has generated an enhancer sequence, or theAUA(GAA)3 sequence has a negative effect onboth the basal splicing level at site A7 and its acti-vation by ASF/SF2. This was surprising, as the(GAA)3 element shows strong homology with therat b-tropomyosin pre-mRNA sequence that bindsASF/SF2 (GAAGAAGAG),37 and matches one ofthe consensus binding sequences defined bySELEX experiments performed with the ASF/SF2protein.43 For a better understanding of therelative effects of the (GAA)3 sequence and theAUA(GAA)3 sequence on splicing efficiency, weinserted these two sequences in a heterologousreporter RNA. This RNA is a short E1A transcriptof adenovirus 2 in which an inversion of exon 2resulted in a strong decrease of the intrinsicsplicing ability.44 This construct (Sp1 ex2 inv) canbe used to test foreign enhancer elements afterinsertion into the inverted exon 2 (Figure 12(a)).As can be seen in Figure 12(b1), insertion of the(GAA)3 sequence restored an efficient splicingactivity in the nuclear extract, whereas insertion ofthe AUA(GAA)3 sequence resulted in a weakerrestoration (M/P ratio of 0.53 and 0.32, respect-ively). For a more precise analysis of the enhancerproperties of the (GAA)3 and AUA(GAA)3sequences, in vitro splicing experiments with theSp1 ex2 inv transcript in which no, or the (GAA)3and AUA(GAA)3 sequence was inserted were per-formed using S100 extract with or without additionof ASF/SF2. For comparison, a sequence known tobind ASF/SF2 (ASF-A10 in Figure 12(a))44 wasalso inserted into the inverted exon 2. Without anyinsertion in the inverted exon, splicing in S100extract was poor with or without addition ofASF/SF2 (Figure 12(b2)). Insertion of the ASF-A10, (GAA)3 or AUA(GAA)3 sequence increasedsplicing efficiency in the absence of ASF/SF2 andthe strongest increase was obtained for (GAA)3(M/P ratio of 0.13 compared to 0.09 for ASF-A10and 0.07 for AUA(GAA)3). Interestingly, activationupon addition of ASF/SF2 was stronger for
Figure 9. The AUA(GAA)3 sequence is required forhnRNP A1 binding to SLS2. (a1) and (a2) The secondarystructure proposed for the A7 SLS2 WT RNA (a1) andvariant A7 SLS2 C3 RNA (a2). Grey characters indicatethe CCC sequence that substitutes the AUA(GAA)3sequence in A7 SLS2 C3 RNA. (b) Detection of anelectrophoretic mobility shift after incubation of2.5 pmol of 30-end labelled A7 SLS2 WT RNA with31.25 pmol of hnRNP A1 in the absence or presence ofcompetitor RNA (cRNA: WT or C3 A7 SLS2 RNAs,350 pmol). (c) UV cross-link of hnRNP A1 to 50 fmol(about 500,000 cpm) of uniformly labelled A7 SLS2WT and A7 SLS2 C3 RNAs (c1) or to A7 SLS2,3 WTand A7 SLS2,3 C3 RNAs (c2). Same legend as forFigure 7(c).
642 Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss

(GAA)3 and ASF-A10, than for AUA(GAA)3 (M/Pratio of 0.21 for (GAA)3 compared to 0.1 for AUA-(GAA)3). This indicated that the 50-flanking AUAnucleotides have a negative effect on the (GAA)3element. One possibility was that hnRNP A1 bindsto the first half of the sequence (AUAGAAGA)and competes the binding of ASF/SF2 to the(GAA)3 motif. To test for this hypothesis directly,we performed gel-shift and UV cross-linkingexperiments in the presence of equimolar amountsof purified ASF/SF2 and hnRNP A1 recombinantproteins, using RNAs corresponding to the secondexon of the reporter Inv Ex2 RNAs (Figure 12(c)).The same experiment was repeated with nuclearextract without any addition of exogenous hnRNP
A1 and ASF/SF2 (Figure 12(c)). Each cross-linkedprotein was detected with specific antibodies(anti-ASF/SF2 (c1), anti-hnRNP A1 (c2)). Bothwith purified proteins and with nuclear extract,we showed that the (GAA)3-containing RNA ispreferentially recognised by ASF/SF2, whereas theAUA(GAA)3 containing RNA is primarily recog-nised by hnRNP A1, which almost completelycompetes the binding of ASF/SF2 to this RNA.
It was also interesting to test how hnRNP A1modifies the binding of ASF/SF2 to the SLS2 HIV-1 RNA region. To tackle this problem, we per-formed supershift assays on complexes formed bythe WT and variant C3 SLS2,3 RNAs (Figures 1and 9(a)) in a nuclear extract (Figure 13(a)). Upon
Figure 10. Deletion of AUA(GAA)3 diminishes binding of hnRNP A1 to the entire A7 RNA region. (a) Primerextension analysis of A7 SLS1,2,3 C3 transcript (1.16 pmol) cleaved with 0.025 or 0.1 unit of T1 RNase (lanes markedþ and þþ , respectively) in the presence (þ ) or absence (2 ) of 50 fmol hnRNP A1 ([hnRNP A1]/[RNA] ¼ 43). Oligo-nucleotides O-2156 and O-2382 were used as primers. Same legend as for Figure 5(b). (b) Schematic representation ofthe experimental data shown in (a) on the A7 SLS1,2,3 C3 RNA secondary structure. Same symbols as for Figure7(b). (c) Polyacrylamide gel electrophoresis of the in vitro splicing products of the L47-8072 WT and C3 RNAs in thepresence or absence of recombinant ASF/SF2 at a 0.5 mM concentration. Same legend as for Figure 6(c).
Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss 643
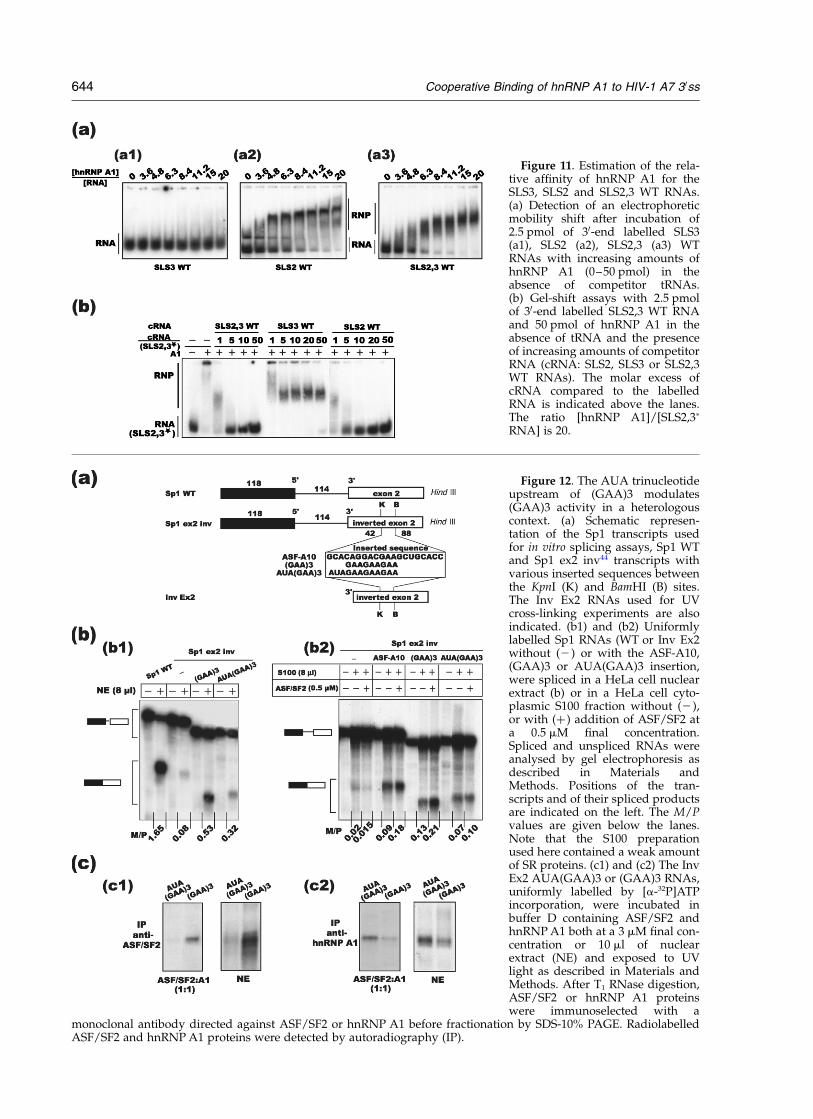
Figure 11. Estimation of the rela-tive affinity of hnRNP A1 for theSLS3, SLS2 and SLS2,3 WT RNAs.(a) Detection of an electrophoreticmobility shift after incubation of2.5 pmol of 30-end labelled SLS3(a1), SLS2 (a2), SLS2,3 (a3) WTRNAs with increasing amounts ofhnRNP A1 (0–50 pmol) in theabsence of competitor tRNAs.(b) Gel-shift assays with 2.5 pmolof 30-end labelled SLS2,3 WT RNAand 50 pmol of hnRNP A1 in theabsence of tRNA and the presenceof increasing amounts of competitorRNA (cRNA: SLS2, SLS3 or SLS2,3WT RNAs). The molar excess ofcRNA compared to the labelledRNA is indicated above the lanes.The ratio [hnRNP A1]/[SLS2,3p
RNA] is 20.
Figure 12. The AUA trinucleotideupstream of (GAA)3 modulates(GAA)3 activity in a heterologouscontext. (a) Schematic represen-tation of the Sp1 transcripts usedfor in vitro splicing assays, Sp1 WTand Sp1 ex2 inv44 transcripts withvarious inserted sequences betweenthe KpnI (K) and BamHI (B) sites.The Inv Ex2 RNAs used for UVcross-linking experiments are alsoindicated. (b1) and (b2) Uniformlylabelled Sp1 RNAs (WT or Inv Ex2without (2) or with the ASF-A10,(GAA)3 or AUA(GAA)3 insertion,were spliced in a HeLa cell nuclearextract (b) or in a HeLa cell cyto-plasmic S100 fraction without (2),or with (þ ) addition of ASF/SF2 ata 0.5 mM final concentration.Spliced and unspliced RNAs wereanalysed by gel electrophoresis asdescribed in Materials andMethods. Positions of the tran-scripts and of their spliced productsare indicated on the left. The M/Pvalues are given below the lanes.Note that the S100 preparationused here contained a weak amountof SR proteins. (c1) and (c2) The InvEx2 AUA(GAA)3 or (GAA)3 RNAs,uniformly labelled by [a-32P]ATPincorporation, were incubated inbuffer D containing ASF/SF2 andhnRNP A1 both at a 3 mM final con-centration or 10 ml of nuclearextract (NE) and exposed to UVlight as described in Materials andMethods. After T1 RNase digestion,ASF/SF2 or hnRNP A1 proteinswere immunoselected with a
monoclonal antibody directed against ASF/SF2 or hnRNP A1 before fractionation by SDS-10% PAGE. RadiolabelledASF/SF2 and hnRNP A1 proteins were detected by autoradiography (IP).
644 Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss
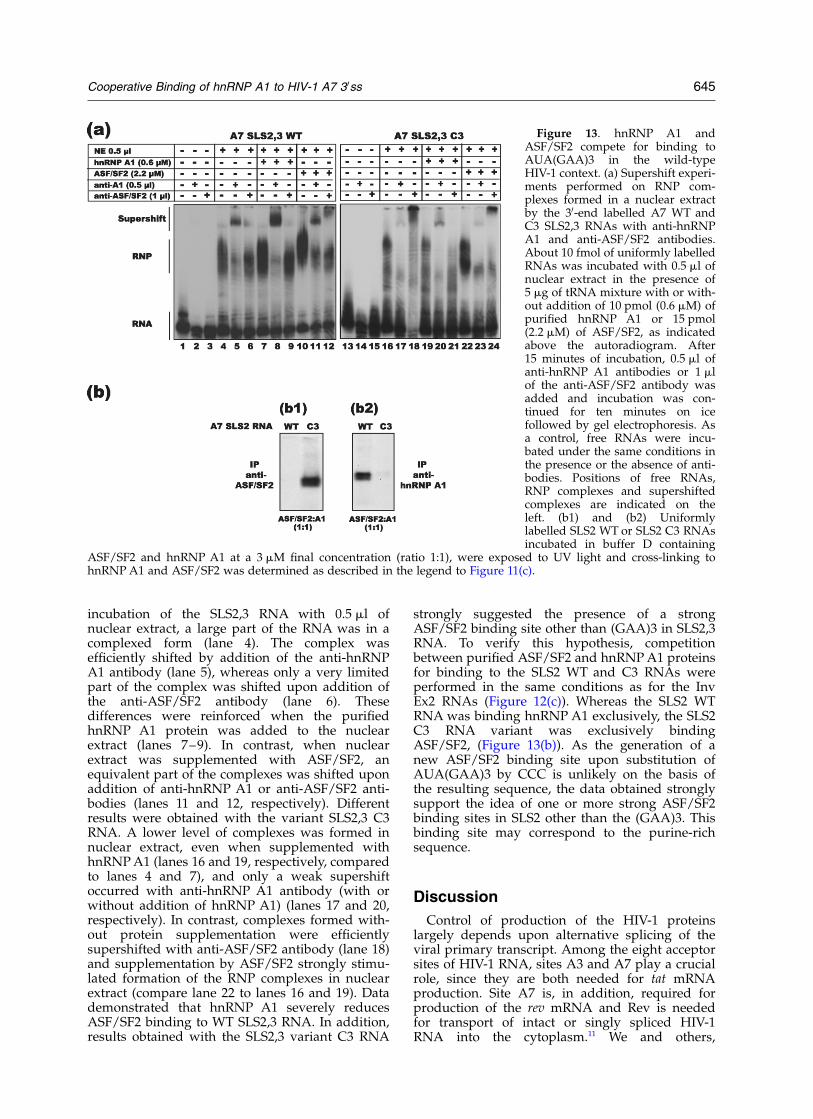
incubation of the SLS2,3 RNA with 0.5 ml ofnuclear extract, a large part of the RNA was in acomplexed form (lane 4). The complex wasefficiently shifted by addition of the anti-hnRNPA1 antibody (lane 5), whereas only a very limitedpart of the complex was shifted upon addition ofthe anti-ASF/SF2 antibody (lane 6). Thesedifferences were reinforced when the purifiedhnRNP A1 protein was added to the nuclearextract (lanes 7–9). In contrast, when nuclearextract was supplemented with ASF/SF2, anequivalent part of the complexes was shifted uponaddition of anti-hnRNP A1 or anti-ASF/SF2 anti-bodies (lanes 11 and 12, respectively). Differentresults were obtained with the variant SLS2,3 C3RNA. A lower level of complexes was formed innuclear extract, even when supplemented withhnRNP A1 (lanes 16 and 19, respectively, comparedto lanes 4 and 7), and only a weak supershiftoccurred with anti-hnRNP A1 antibody (with orwithout addition of hnRNP A1) (lanes 17 and 20,respectively). In contrast, complexes formed with-out protein supplementation were efficientlysupershifted with anti-ASF/SF2 antibody (lane 18)and supplementation by ASF/SF2 strongly stimu-lated formation of the RNP complexes in nuclearextract (compare lane 22 to lanes 16 and 19). Datademonstrated that hnRNP A1 severely reducesASF/SF2 binding to WT SLS2,3 RNA. In addition,results obtained with the SLS2,3 variant C3 RNA
strongly suggested the presence of a strongASF/SF2 binding site other than (GAA)3 in SLS2,3RNA. To verify this hypothesis, competitionbetween purified ASF/SF2 and hnRNP A1 proteinsfor binding to the SLS2 WT and C3 RNAs wereperformed in the same conditions as for the InvEx2 RNAs (Figure 12(c)). Whereas the SLS2 WTRNA was binding hnRNP A1 exclusively, the SLS2C3 RNA variant was exclusively bindingASF/SF2, (Figure 13(b)). As the generation of anew ASF/SF2 binding site upon substitution ofAUA(GAA)3 by CCC is unlikely on the basis ofthe resulting sequence, the data obtained stronglysupport the idea of one or more strong ASF/SF2binding sites in SLS2 other than the (GAA)3. Thisbinding site may correspond to the purine-richsequence.
Discussion
Control of production of the HIV-1 proteinslargely depends upon alternative splicing of theviral primary transcript. Among the eight acceptorsites of HIV-1 RNA, sites A3 and A7 play a crucialrole, since they are both needed for tat mRNAproduction. Site A7 is, in addition, required forproduction of the rev mRNA and Rev is neededfor transport of intact or singly spliced HIV-1RNA into the cytoplasm.11 We and others,
Figure 13. hnRNP A1 andASF/SF2 compete for binding toAUA(GAA)3 in the wild-typeHIV-1 context. (a) Supershift experi-ments performed on RNP com-plexes formed in a nuclear extractby the 30-end labelled A7 WT andC3 SLS2,3 RNAs with anti-hnRNPA1 and anti-ASF/SF2 antibodies.About 10 fmol of uniformly labelledRNAs was incubated with 0.5 ml ofnuclear extract in the presence of5 mg of tRNA mixture with or with-out addition of 10 pmol (0.6 mM) ofpurified hnRNP A1 or 15 pmol(2.2 mM) of ASF/SF2, as indicatedabove the autoradiogram. After15 minutes of incubation, 0.5 ml ofanti-hnRNP A1 antibodies or 1 mlof the anti-ASF/SF2 antibody wasadded and incubation was con-tinued for ten minutes on icefollowed by gel electrophoresis. Asa control, free RNAs were incu-bated under the same conditions inthe presence or the absence of anti-bodies. Positions of free RNAs,RNP complexes and supershiftedcomplexes are indicated on theleft. (b1) and (b2) Uniformlylabelled SLS2 WT or SLS2 C3 RNAsincubated in buffer D containing
ASF/SF2 and hnRNP A1 at a 3 mM final concentration (ratio 1:1), were exposed to UV light and cross-linking tohnRNP A1 and ASF/SF2 was determined as described in the legend to Figure 11(c).
Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss 645

previously described a complex regulation of theactivity of the A3 acceptor site.7,16,17,38 Two cis actinginhibitory elements, that bind proteins hnRNP Hand hnRNP A1, respectively, were found to down-regulate site A3 utilisation. Several cis regulatoryelements were also previously found to controlsplicing at site A7.6,7,18,19 Here, we show that thedifferent splicing regulatory elements acting onsite A7 (ISS, ESE3/(GAA)3 and ESS3) are each con-tained in a distinct stem-loop structure and thathnRNP A1 orchestrates their activity by itscooperative binding to these numerous regulatoryelements and some of their linking sequences.Globally, by wrapping large parts of the RNAregion downstream and upstream of site A7,hnRNP A1 competes the binding of ASF/SF2 tosite A7.
The previously identified ESE3/(GAA)3 splicingenhancer binds hnRNP A1
Our probing and cross-linking experiments onthe HIV-1 SLS2,3 RNA region, in nuclear extractand with purified hnRNP A1, clearly demonstratethe binding of hnRNP A1 to the ESE3/(GAA)3element (Figures 4, 5 and 9). This sequence waspreviously identified as a splicing enhancerelement.6 Why was the ambivalence of the ESE3/(GAA)3 element not previously detected? Evidencefor a splicing enhancer property of the ESE3/(GAA)3 element was obtained by deletion of the(GAA)3 sequence6 and upon this deletion, anhnRNP A1 binding site with a sequence similar toESS3b (UAGUGG) was created. Hence, in theseprevious experiments, the intrinsic enhanceractivity of (GAA)3 was eliminated by the deletion,whereas the capacity to bind hnRNP A1 was con-served. Our CCC RNA variant was designated inorder to avoid generation of an hnRNP A1 siteand the CCC substitution was not expected tocreate a new ASF/SF2 specific site. In accord witha splicing inhibitory property of the AUA(GAA)3sequence, the C3 variant RNA with a deletion ofthe AUA(GAA)3 sequence was spliced more effi-ciently than the WT RNA (Figure 10). hnRNP A1binding to the AUA(GAA)3 was also evidencedby previous cross-linking experiments by Zhuet al.18 Indeed, upon using an HIV-1 RNA fragmentlabelled by incorporation of [a-32P]UTP from pos-ition 7972 to 8027 for cross-linking experiments,these authors got radiolabelled hnRNP A1 afterRNase digestion. The only uridine residue in thelabelled fragment was that in the AUA(GAA)3sequence, showing that this sequence bindshnRNP A1. However, the authors did not make alink between these data and the possibility thatthe AUA(GAA)3 sequence is an hnRNP A1 bind-ing site. Thus, our data and previous data, are ingood agreement with the presence of an up tonow not described hnRNP A1 binding site inESE3/(GAA)3 and this site modulates theenhancer activity of this element.
A central role of the AUA(GAA)3 hnRNP A1binding site for cooperative binding of hnRNPA1 to the HIV-1 A7 RNA region
Our gel shift assays (Figure 11(a1) and (b)), andcross-linking data (Figure 6(b)), clearly show thatthe previously identified hnRNP A1 binding site(ESS3b), that is located in a terminal loop, has alower affinity for hnRNP A1 compared to theAUA(GAA)3 sequence located in a large internalloop. Indeed, the SLS3 stem-loop structure contain-ing ESS3b is not able to displace hnRNP A1 boundto the SLS2 stem-loop structure, even in largeexcess (Figure 11(b)). However, in the SLS1,2,3RNA, hnRNP A1 binding to ESS3b is stronglyreinforced by the presence of AUA(GAA)3 (Figures5, 10 and 11), showing a synergistic binding ofhnRNP A1 to AUA(GAA)3 and ESS3b. This prob-ably explains why, in spite of its low intrinsicaffinity for hnRNP A1, ESS3b has a markedsplicing silencer property (Figure 6(c)). Accord-ingly, the inhibitory activity of ESS3 inserted in aforeign context was previously found to be highlydependent upon the sequence of this foreigncontext,6 and when the entire segment containingAUA(GAA)3 and ESS3 was transported into aforeign context, the splicing activity was augmen-ted upon increasing the distance between the twoelements.6
In agreement with the previously proposedoligomerisation of hnRNP A1 upstream of ESS3bafter binding to ESS3b,18 disruption of ESS3babolished the protection by hnRNP A1 of the entiresegment that links ESS3b and the AUAGAA50-terminal sequence of ESE3 (Figure 7). However,we showed that oligomerisation of hnRNP A1 onthe polypurine sequence also depends upon thepresence of ESE3/(GAA)3 (Figure 10). Disruptionof ESS3b or AUA(GAA)3 also affects the bindingof hnRNP A1 to the ISS sites 1 and 2 (Figures 7and 10) and this is in agreement with previousdata, suggesting a cooperative binding of hnRNPA1 to ESS3b and ISS.8 Our data show that the pre-sence of AUA(GAA)3 is more determinant forbinding of hnRNP A1 to the ISS sites 1 and 2 thanthe presence of ESS3b (Figures 7 and 10). Interest-ingly, hnRNP A1 binding to the third ISS site (site3), that blocks the access to the branch sitesequence numbered 2,8 is less dependent upon thepresence of ESS3b and ESE3 (Figures 7 and 10).Site 1 in ISS, the AUAGAA site of ESE3 and ESS3bare all located in terminal or internal loops thatflank irregular helical structures. This RNA archi-tecture probably has enough flexibility to ensure acooperative binding of hnRNP A1 to these threedistant sites. A cooperative binding of hnRNP A1to these different sites and to the segment linkingESS3b and ESE3 is in agreement with the previousdata showing that addition of the entire recombi-nant hnRNP A1, but not its N-terminal domain,which lacks the protein–protein interactioncapacity, restored splicing inhibition in a hnRNPA1 depleted nuclear extract.18
646 Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss

Cooperative binding of hnRNP A1 upstreamand downstream of site A7 competes ASF/SF2 binding
Taken together, our cross-linking experiments innuclear extract and with purified recombinanthnRNP A1 and ASF/SF2 proteins demonstrate thatbinding of hnRNP A1 to AUA(GAA)3 competes thebinding of ASF/SF2 to the enhancer sequence(Figure 13(b)). Accordingly, the supershift assaysthat we performed on complexes assembled on theSLS2,3 RNA in nuclear extract (Figure 13(a)), demon-strated that in splicing conditions, only a verylimited amount of SLS2,3 RNA is associated withASF/SF2 compared to hnRNP A1. As a second con-firmation, whereas the (GAA)3 sequence had astrong enhancer activity in a heterologous contextand was binding ASF/SF2 efficiently, the AUA-(GAA)3 sequence transported in the same heter-ologous context had a limited enhancer activity andwas predominantly bound by hnRNP A1 comparedto ASF/SF2 (Figure 12). A second ASF/SF2 bindingsite was previously proposed to be present in theRNA portion from positions 7992 to 8004. This RNAsegment was found to have a splicing enhanceractivity in a foreign context.19 According to our sec-ondary structure analysis, this RNA region formsthe terminal part of SLS2. Interestingly the CCGA-CGAG sequence of this SLS2 terminal part wasfound to be highly conserved in all the strains thatwe compared (Figure 3), suggesting a high func-tional importance. Furthermore, the SLS2 terminalloop was protected in nuclear extract, but not in thepresence of purified hnRNP A1 (Figures 4 and 5).Thus, one or several component(s) of nuclear extractbind(s) to this terminal loop. This is probably notASF/SF2, since no binding of ASF/SF2 was detectedon the SLS2 stem-loop structure in the presence ofhnRNP A1, whereas hnRNP A1 does not bind theSLS2 terminal loop (Figure 13(b)). This does notexclude an indirect binding by protein–protein inter-actions. Furthermore, our supershift assays on thevariant SLS2,3 C3 RNA reveal the presence of anASF/SF2 binding site other than (GAA)3 in theSLS2,3 RNA region (Figure 13(a)). This ASF/SF2target site may be the AGACAGAGAC purine-richelement (8037–8045), whose sequence fits some ofthe sequences previously identified as ASF/SF2binding sites.43 This sequence is buried by boundhnRNP A1 protein in the WT RNA, but is not associ-ated with hnRNP A1 in the C3 RNA (Figures 5 and10). Thus, binding of ASF/SF2 to the AGACAG-AGAC sequence in the C3 SLS2,3 RNA is possible.Taking into account the presence of two possibleASF/SF2 binding sites in the A7 downstreamsequence, binding of ASF/SF2 to this region mightalso be cooperative.
The proposed model of A7 site regulation andits functional implications
Taking into consideration all our data, we pro-pose the following model (Figure 14): hnRNP A1
binds cooperatively to three distinct segments ofthe HIV-1 A7 RNA region: the terminal part ofSLS1, AUAGAA in ESE3 and ESS3b. In addition,hnRNP A1 oligomerises along the purine-richsequence when sites ESS3b and AUAGAA areoccupied. This wrapping of the A7 RNA region byhnRNP A1 occludes the access of ASF/SF2 to itstarget site (GAA)3 and may be to the AGACAG-AGAC sequence. At the same time, access to thebranch point sequence number 2 identified byTange et al.8 is buried by hnRNP A1 binding tothis site. According to this model, when ASF/SF2competes hnRNP A1 for binding to the AUA-(GAA)3 sequence, the multiple hnRNP A1protein–protein interactions should be disrupted.This decompaction of the RNA structure and thepresence of ASF/SF2, likely favour binding ofspliceosomal components. We can also imaginethat a simultaneous and cooperative binding ofASF/SF2 to the (GAA)3 and AGACAGAGACsequences reinforces the capacity of ASF/SF2 tocompete hnRNP A1.
Several alternatively used splicing sites werefound to be regulated by combination of enhancerand silencer elements located in close proximity.20
Here we found a peculiar architecture of the A7splicing regulatory elements, with on the onehand, a silencer element overlapping an enhancerelement and, on the other hand, a complex arrayof hnRNP A1 binding sites, which, by interactionof the associated proteins, blocks the access ofsplicing factors to their target sites. Muhlemannet al.45 recently described an adenovirus splicingregulatory element that showed activator propertyin normal HeLa cell nuclear extract and inhibitoryproperty in adenovirus-infected cells. Thiselement with opposite function depending uponthe cellular context was designated as a Janussplicing regulatory element by the authors. TheHIV-1 AUA(GAA)3 motif can also be consideredas a Janus splicing regulatory element, that maybe an activator or an inhibitor, depending uponthe relative concentrations of hnRNP A1 andASF/SF2 in the cell and also upon their levels ofphosphorylation. It might be that both proteinconcentrations and phosphorylation levels vary asa function of the stage of the HIV-1 infection.Indeed, variations of hnRNP A1 concentrations inthe course of infection by virus HTLV-1 wererecently found to be involved in the regulation ofthe expression of the viral proteins.46 The AUA-(GAA)3 motif may be a key motif in the control oftat and rev mRNA production by sensingvariations of the hnRNP A1 and ASF/SF2 ratioor their degree of phosphorylation. Interestingly,the (GAA)3 sequence was also recently found tobe required for efficient transport of unsplicedRNA in the cytoplasm by the Rev-associatedmechanism.47 This reinforces the idea that theAUA(GAA)3 sequence is a multifunctionalelement that plays a key role in the regulationof HIV-1 protein production by alternativesplicing.
Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss 647

However, in spite of its great functional import-ance, the AUA(GAA)3 sequence is not universalin HIV-1 strains. The AUA(GAA)3 sequence isfound in the BRU, related HXB2 and NL4-3 strains,which are frequently encountered in European andAmerican infected patients. It is also present in thehighly virulent MAL strain isolated in Gabon. It isreplaced by the AUC(GAA)3 sequence in severalother HIV-1 strains†. It should be noticed that inthe HIV-1 BRU, HXB2, NL4-3 and MAL strainsthe presence of UAG in the AUA(GAA)3 motifgenerates a termination stop in the Tat open readingframe.48 The Tat proteins produced by these strainsare 85 or 86 amino acid residues long. In strainswith the AUC(GAA)3 sequence, the Tat terminationcodon is the UAG triplet contained in the ESS3bhnRNP A1 binding site and the Tat proteins pro-duced are 101 amino acid residues long.48 Hence,this opens the following question: is there a necessityfor a tighter control of tat mRNA productiondepending upon the type of Tat protein produced?
No specific function has been attributed to the 15amino acid residue extension in Tat 101. However,comparative analyses of the in vivo properties of thetwo proteins are limited up to now.
Materials and Methods
Plasmid construction
Plasmid pBRU349 was used as the source of cDNAsequences from the HIV-1 BRU/LAI strain (GenBankaccession number K02013). Pieces of HIV-1 cDNA werePCR amplified using conditions of Nour et al.50 A HIV-1BRU cDNA fragment (positions 5577–5676 in theHIV-1BRU genome, numbering as by Ratner et al.51) wasdesignated D4, as it contained the D4 50ss. It was ampli-fied with an antisense primer that generated a BglII site49 nt downstream from site D4 in the intronic sequence(Table 1). A series of HIV-1 cDNA segments containingthe A7 30ss, was amplified with generation of a BglIIsite 279 nt upstream of the site A7 in the intronicsequence (A7 fragments). They differed one from theother by the length of the exonic sequence that they con-tained (Table 1). The D4 and A7 fragments were digestedby the BglII nuclease. Then, mini-genes with a miniD4-A7 intron were built by ligation of D4 fragment withone of the A7 fragments. The four A7 fragments thatwere produced corresponded to the following HIV-1BRU
RNA regions (Table 1): fragment A7a (positions 7692–8043), fragment A7b (positions 7692–8072), fragmentA7c (positions 7692–9140), and fragment A7d (positions7692–8490), respectively. Fragment A7d was obtainedby digestion of the A7c fragment with the XhoI nuclease.Plasmid pBluescript KSIIþ was used as the host vectorfor all the constructs. Methods for recombinant DNAproduction were as described by Sambrook et al.52
The Sp1 ex2 inv (GAA)3 and the Sp1 ex2 inv AUA-(GAA)3 constructs were obtained by insertion betweenthe unique KpnI and BamHI restriction sites of theAdenovirus Sp1 ex2 inv plasmid44 of short fragments ofHIV-1BRU cDNA (positions 8017–8025) for the (GAA)3construct and (positions 8012–8025) for the AUA(GAA)3construct (see Table 1).
The cDNA matrices for in vitro transcription of theSLS2, SLS3 or SLS2,3 WT RNA were PCR amplifiedfrom pL47-8490 plasmid using the antisense primerO-1725 or O-2257 (Table 1) and sense primers thatgenerated a SP6 RNA polymerase promoter. Themutated versions of pL47-8490 plasmid were used asmatrices for the production of variant SLS2, SLS3 andSLS2,3, SLS1,2,3 RNAs. The sense primer O-2067 wasused to amplify fragments SLS2 WT, SLS2 C3, SLS2,3WT, SLS2,3 C3, SLS2,3 mut b, SLS2,3 mut c and thesense primers O-2068, O-2070 and O-2069 (Table 1) wereused to amplify fragments SLS3 WT, SLS3 mut b andSLS3 mut c, respectively. The sense primer O-2065 wasused to amplify SLS1,2,3 WT, SLS1,2,3 mut b, SLS1,2,3C3. The amplified cDNA fragments were purified byphenol-extraction followed by ethanol-precipitation,dissolved in distilled water, and used for in vitro tran-scription as described below.
Site-directed mutagenesis
Site-directed mutageneses were performed by the PCRmethod, using the Stratagene Quickchangee Site-DirectedMutagenesis kit and pL47-8072 plasmid as the matrix. The
Figure 14. Model of hnRNP A1 binding to the HIV-1A7 RNA region. A schematic representation of hnRNPA1 bound to its various target sites that we identifiedby T1 RNase footprinting experiments is made on thesecondary structure established for the A7 RNA region.Double arrows indicate the cooperative interaction thatwe detected between hnRNP A1 molecules bound to itsdifferent target sites. The level of cooperativity is shownby the thickness of the double arrow. Competitionbetween hnRNP A1 and ASF/SF2 for binding to theAUA(GAA)3 sequence is also represented. The potentialsecond ASF/SF2 binding site in the purine-rich sequenceis shown. X in a blue circle indicates the unidentifiednuclear component bound to the terminal loop of SLS2.
† HIV sequence database: http://hiv.web.lanl.gov/
648 Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss

oligonucleotides used for: (i) substitution of the AUA-(GAA)3 sequence of the ESE3/(GAA)3 motif (variant C3),(ii) mutation of ESS3b (variant mut b) motif, (iii) mutationof the sequence located between the ESS3 a and b elements(variant mut c), and (iv) substitution of the ESS3b hnRNPA1 binding site by an hnRNP A1 consensus binding site(variant A1 cons) are given in Table 1. The presence of theexpected mutations in the mutated pL47-8072 plasmidwas verified by DNA sequencing.
In vitro transcription
Prior to in vitro transcription, each plasmid waslinearised with the appropriate restriction nuclease.Synthesis of cold transcripts for RNA secondarystructure analysis were carried out in a 30 ml mixturecontaining 2 mg of linearised plasmid, as described.38
RNA was dissolved in milliQ water and concentrationwas estimated by A260 nm measurement. For splicingassays, uniformly labelled capped transcripts wereproduced by T7 RNA polymerase transcription asdescribed.53 For UV cross-linking reactions and immuno-precipitations, uniformly radiolabelled RNAs wereproduced by SP6 RNA polymerase transcription usingsuitable PCR products as matrices (Table 1). Transcrip-tions were carried out on 2 pmol of PCR product in an18 ml reaction mixture containing 20 mM MgCl2,80 mg/ml of PEG8000, 0.01% (v/v) Triton X-100, 1 mMspermidine, 40 mM Tris–HCl (pH 7.9), 10 mM DTT,0.1 mg/ml of BSA, ten units of RNasin, 1 mM eachrUTP (or rATP), rCTP, rGTP, 5 mM rATP (or rUTP),20 mCi of [a-32P]ATP (or [a-32P]UTP) (800 Ci/mmol)(ICN) and 40 units of SP6 RNA polymerase (USBPharmacia Biotech). For gel-shift assays, 300 pmol ofcold RNA transcript was 30-end labelled with[50-32P]pCp as described.54
RNA secondary structure analysis
RNA secondary structure analyses were all performedon about 100 ng of cold RNA transcript prepared asdescribed above, either in classical conditions in thepresence of 10 mM Mg2þ35 or in buffer D.38
For analysis in classical conditions, V1 RNase(prepared from Naja oxiana venom,55 and S1 nuclease(Amersham Pharmacia Biotech) digestions were per-formed in the presence of 1.25 mg and 2.5 mg of yeasttRNA mixture (Roche Diagnostics), respectively. RNAwas pre-incubated in 50 ml of a 50 mM KCl, 10 mMMgCl2, 10 mM Tris–HCl (pH 7.5) solution (V1 RNase)and in 50 ml of a 50 mM KCl, 1 mM ZnCl2, 10 mMMgCl2, 25 mM sodium acetate (pH 4.5) solution (S1
nuclease) for 30 minutes at room temperature prior toaddition of the enzyme (0.1 unit of V1 RNase and threeunits of S1 nuclease). Digestion was for 12 minutes at20 8C. V1 RNase action was stopped by addition of100 mM EDTA, followed by phenol-extraction. S1
nuclease digestions were stopped by adding 20 mg oftRNA, chilling on ice and by phenol-extraction.
Dimethylsulfate (DMS) and 1-cyclohexyl-3-(2-mor-pholinoethyl)-carbodiimide-metho-p-toluene sulfonate(CMCT) modifications were performed in the presenceof 10 mg of yeast tRNA mixture. RNA was pre-incubatedfor 20 minutes at 20 8C in 50 ml of a 50 mM KCl, 10 mMMgCl2, 100 mM sodium cacodylate (pH 7.5) solution(DMS) or a 50 ml of 50 mM KCl, 10 mM MgCl2, 100 mMsodium borate (pH 8) solution (CMCT). Incubation withDMS (1 ml of a 25% DMS–ethanol) or CMCT (3.2 mg/
300 ml assay) was for 30 minutes at 20 8C. DMS reactionswere stopped by addition of 3 ml of 6 M b-mercapto-ethanol followed by ethanol-precipitation and CMCTreactions were stopped by ethanol-precipitation.
Both enzymatic and chemical probing in buffer Dwithout polyvinyl alcohol (PVA) were performed in thepresence of 5 mg of tRNA mixture. For enzymaticprobing, RNA was pre-incubated for five minutes at65 8C in 15 ml of a 73 mM KCl, 3.125 mM MgCl2,0.15 mM EDTA, 0.35 mM DTT, 0.18 mM PMSF, 15%glycerol, 15 mM Hepes (pH 7.9) solution. After slowcooling at 30 8C, 0.025 or 0.1 unit of T1 RNase (RocheDiagnostics) (conditions indicated by þ or þþ, respect-ively, in Figures), 0.6 unit of T2 RNase (Gibco BRL) or0.15 unit of V1 RNase were added and incubation wasfor ten minutes at 30 8C. V1 RNase digestions werestopped as above, T1 and T2 RNase digestions werestopped by addition of 20 mg of tRNA followed byphenol-extraction and ethanol-precipitation.
For DMS probing in buffer D, RNA was pre-incubatedfor five minutes at 65 8C in 15 ml of the following sol-ution: 73 mM KCl, 3.125 mM MgCl2, 0.15 mM EDTA,0.35 mM DTT, 0.18 mM PMSF, 15% glycerol, 15 mMHepes (pH 7.9) buffer. After slow cooling at 30 8C,sodium cacodylate was added to a 50 mM final concen-tration to obtain efficient DMS modification. Then, 1 mlof a 25% DMS–ethanol solution was added and incu-bation was for ten minutes at 30 8C. The reaction wasstopped by addition of 3 ml of 6 M b-mercaptoethanolfollowed by ethanol-precipitation.
Identification of modification and cleavage positionsand secondary structure prediction
Positions of chemical modifications and enzymaticcleavages were identified by primer extension analysiswith the avian myeloblastosis virus reverse transcriptase(Appligene), using 50-end labelled O-2156, O-609, O-2382primers (Table 1). The 50-end labellings with [g-32P]ATP3000 Ci/mmol (Amersham Pharmacia Biotech) were asdescribed by Sambrook et al.52 Annealing of primers andprimer extensions were performed as described.35
Predictions of secondary structure were made with theMfold program of the GCG software, version 8.1 Unix(1995), that is on the basis of the thermodynamic valuesproposed by Jaeger et al.56 for an RNA in solution in1 M NaCl buffer at 37 8C. The results of the experimentalanalysis were introduced in the computer search, inorder to define the secondary structure, that had thebest fit with the experimental data.
The GenBank accession numbers of HIV-1 or relatedHIV-1 sequences used in the comparative analyses are:K02013 (HIV-1BRU), AF069670 (HIV-1SE-SE7253), AF067159(HIV-111246), K03454 (HIV-1ELI), AF075703 (HIV-1FIN9363),AF0616142 (HIV-1SE6165), AF190127 (HIV-1VI991) andAF082394 (HIV-1SE92809).
HnRNP A1 and ASF/SF2 production and purification
Full-length hnRNP A1 protein was expressed inEscherichia coli from plasmid pET11a-hnRNPA1.57
Transformed E. coli BL21 (DE3) cells were grown inLB/Amp medium. IPTG induction was made at an A600
of 0.6. After 20 hours growth, cells were sonicated inlysis buffer (50 mM Hepes (pH 7.6), 1 mM EDTA,500 mM KCl, 10% glycerol, 1 mM DTT, 1 mM PMSF,10 mM Na2S2O5). Cell extract was clarified by centri-fugation, diluted with three volumes of column buffer
Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss 649

A (50 mM Hepes (pH 7.6), 1 mM EDTA, 10% glycerol)and loaded onto a 50 ml Q-Sepharose (Pharmacia) FPLCcolumn. The column was washed with buffer A with250 mM KCl. The flow-through was diluted twice inbuffer A and loaded onto a 5 ml Hitrap-HeparinSepharose (Pharmacia) FPLC column. The column waswashed with buffer A containing 125 mM KCl andbound proteins were eluted with a linear 125 mM–1 MKCl gradient. Fractions containing hnRNP A1 (eluted at200 mM KCl) were dialysed against buffer D (100 mMKCl, 0.2 mM EDTA, 0.5 mM DTT, 0.25 mM PMSF, 20%glycerol, 20 mM Hepes (pH 7.9)). Final preparation wasjudged to be homogeneous by SDS-PAGE and hnRNPA1 concentration was determined by Bradford proteinassay. The recombinant ASF/SF2 protein, which is histi-dine-tagged at the N-terminal part, was produced andpurified from baculovirus-infected cells as described.44
T1 RNase probing of RNP complexes
To form RNP complexes, 100 ng of cold RNA tran-script and 5 mg of yeast tRNA mixture prepared in 14 mlof 25 mM KCl, 4.5 mM MgCl2, 0.05 mM EDTA, 0.12 mMDTT, 0.06 mM PMSF, 5% glycerol, 5 mM Hepes (pH 7.9)were incubated for five minutes at 65 8C. After slowcooling at room temperature, 4 ml of nuclear extract(Computer Cell Culture Center S.A., Belgium) that corre-spond to 52 mg of proteins or 1.7 mg of purified hnRNPA1 (corresponding to 50 pmol of hnRNP A1) was addedand complexes were formed by incubation for ten minutesat 30 8C. Then, T1 RNase (0.025 or 0.1 unit, conditions indi-cated (þ) or (þþ) in the Figures showing the primer exten-sion analyses) was added and incubation was for tenminutes at 30 8C. The reaction was stopped by addition of20 mg of tRNA mixture and cooling at 0 8C, immediatelyfollowed by phenol-extraction. As a control, enzymaticdigestions were performed with the same amount of RNAtranscript and tRNAs incubated in the same conditionsexcept that nuclear extract or hnRNP A1 addition wasreplaced by addition of 4 ml of buffer D.
In vitro splicing assays
In vitro splicing assays were performed with HeLa cellnuclear or cytoplasmic S100 extracts using 100,000 cpmof uniformly labelled RNA transcript per assay, asdescribed.53 To test for splicing activation by SR proteins,nuclear extract or cytoplasmic S100 extract was sup-plemented with 300 ng of recombinant ASF/SF2 protein(0.5 mM) expressed in baculovirus. The splicing reactionmixture was prepared on ice and then incubated at30 8C for 120 minutes, except for the transcripts derivedfrom the adenovirus Sp1 ex2 inv, which were incubatedfor 75 minutes. Splicing products were deproteinisedwith proteinase K, phenol-extracted and analysed on a5% (w/v) polyacrylamide sequencing gel, as described.53
In vitro splicing experiments were repeated three timesfor a given construct using different preparations of thetranscript and splicing efficiencies were determined foreach experiment by scanning the gel with a MolecularDynamics PhosphorImager using the Image Quant Soft-ware, version 3.3. The M/P ratio (amount of mature RNAversus the amount of residual precursor) was calculatedfor each transcript, taking into account the measured radio-activity and the number of uracil residues per molecule.
Electrophoretic mobility shift sssays (EMSA) andsupershift assays
EMSAs were performed with 2.5 pmol (about5000 cpm) of 30-end labelled transcript. RNA–proteincomplex formation was in buffer D without PVA con-taining 3 mM MgCl2, in the presence or absence of 5 mgof competitor tRNAs. Prior to RNP complex formation,RNAs were incubated in 6 ml of buffer D containing3 mM MgCl2 for five minutes at 65 8C, followed by aslow cooling at room temperature. Then, 5 ml of bufferD containing different amounts of purified ASF/SF2 orhnRNP A1 protein dialysed against buffer D or nuclearextract was added. Incubation was for 15 minutes onice. For competition experiments with cold RNAs,incubation of labelled RNA with hnRNP A1 wasfollowed by incubation with a single amount (140-foldexcess ¼ 350 pmol) or various increasing amounts (from2.5 to 50 pmol) of cold competitor RNA. For the assaysperformed with nuclear extract, heparin (5 mg/ml) wasadded after the end of the 15 minutes incubation on iceand incubation was continued for ten minutes at roomtemperature. For supershift experiments, after incubationwith nuclear extract, 0.5 ml of the anti-hnRNP A1 mono-clonal antibody 4B10 (a generous gift from G. Dreyfuss,University of Pennsylvania) or 1 ml of the anti-ASF/SF2monoclonal antibody was added before the heparin treat-ment and incubation was continued for ten minutes at4 8C. RNP complexes formed for shift or supershift assayswere then fractionated onto a 6% polyacrylamide (38:2(w/w) acrylamide/bis-acrylamide) gel with 5% (v/v) gly-cerol in 1 mM EDTA, 45 mM Tris–borate (pH 8.3). Thefree and bound RNAs were visualised by autoradiography.
UV cross-linking reactions and immunoprecipitations
UV cross-linking experiments were performed asdescribed.17 Briefly, radiolabelled RNA (50 fmol, about500,000 cpm) were incubated for 15 minutes at 30 8Cwith nuclear extract, purified hnRNP A1 or ASF/SF2proteins. Formation of RNP complexes was verified byEMSA and the reaction mixture was transferred to 96-well plates for irradiation at 4 8C with 254 nm UV light1 cm from the source for ten minutes. RNA componentswere then digested by addition of 50 units of T1 RNase,and incubation was for 30 minutes at 50 8C.
Selection of hnRNP A1 on protein G-Sepharose beadscoated with anti-hnRNP A1 monoclonal 4B10 antibody(a generous gift from G. Dreyfuss) was performed asdescribed.17 The same procedure was also applied forselection of ASF/SF2 on protein A-Sepharose beadscoated with anti-ASF/SF2 antibody. Each sample ofeluted proteins was divided in two parts: one part wasfractionated on a gel used for autoradiography; theother part was fractionated on a gel used for immuno-blotting. Immunoblotting with anti-hnRNP A1 and anti-ASF/SF2 antibodies was performed as described above.
Acknowledgements
G. Dreyfuss is thanked for his generous gift ofhnRNP A1 specific antibodies and plasmidpET11a-hnRNPA1. H. Hallay is strongly thankedfor her participation in hnRNP A1 production andpurification. V. Igel is acknowledged for her excel-lent technical assistance. V.M. is a fellow of the
650 Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss

French Ministere de la Recherche et de l’Enseigne-ment Superieur. This work was supported by theFrench Agence Nationale de la Recherche sur leSIDA (ANRS), the SIDACTION foundation, theCentre National de la Recherche Scientifique(CNRS), the Institut National de la Sante et de laRecherche Medicale (INSERM), the FrenchMinistere de la Recherche et de l’EnseignementSuperieur, and local funds of the Region Lorraine.
References
1. Smith, C. W. & Valcarcel, J. (2000). Alternative pre-mRNA splicing: the logic of combinatorial control.Trends Biochem. Sci. 25, 381–388.
2. O’Reilly, M. M., McNally, M. T. & Beemon, K. L.(1995). Two strong 50 splice sites and competing, sub-optimal 30 splice sites involved in alternative splicingof human immunodeficiency virus type 1 RNA.Virology, 213, 373–385.
3. Dyhr-Mikkelsen, H. & Kjems, J. (1995). Inefficientspliceosome assembly and abnormal branch siteselection in splicing of an HIV-1 transcript in vitro.J. Biol. Chem. 270, 24060–24066.
4. Swanson, A. K. & Stoltzfus, C. M. (1998). Over-lapping cis sites used for splicing of HIV-1 env/nefand rev mRNAs. J. Biol. Chem. 273, 34551–34557.
5. Staffa, A. & Cochrane, A. (1994). The tat/rev intronof human immunodeficiency virus type 1 is ineffi-ciently spliced because of suboptimal signals in the30 splice site. J. Virol. 68, 3071–3079.
6. Staffa, A. & Cochrane, A. (1995). Identification ofpositive and negative splicing regulatory elementswithin the terminal tat-rev exon of human immuno-deficiency virus type 1. Mol. Cell. Biol. 15, 4597–4605.
7. Si, Z. H., Rauch, D. & Stoltzfus, C. M. (1998). Theexon splicing silencer in human immunodeficiencyvirus type 1 Tat exon 3 is bipartite and acts early inspliceosome assembly. Mol. Cell. Biol. 18, 5404–5413.
8. Tange, T. O., Damgaard, C. K., Guth, S., Valcarcel, J.& Kjems, J. (2001). The hnRNP A1 protein regulatesHIV-1 tat splicing via a novel intron silencer element.EMBO J. 20, 5748–5758.
9. Purcell, D. F. & Martin, M. A. (1993). Alternativesplicing of human immunodeficiency virus type 1mRNA modulates viral protein expression, replica-tion, and infectivity. J. Virol. 67, 6365–6378.
10. Karn, J. (1999). Tackling Tat. J. Mol. Biol. 293,235–254. doi:10.1006/jmbi.1999.3060.
11. Boris-Lawrie, K., Roberts, T. M. & Hull, S. (2001).Retroviral RNA elements integrate components ofpost-transcriptional gene expression. Life Sci. 69,2697–2709.
12. Bonavia, R., Bajetto, A., Barbero, S., Albini, A.,Noonan, D. M. & Schettini, G. (2001). HIV-1 Tatcauses apoptotic death and calcium homeostasisalterations in rat neurons. Biochem. Biophys. Res.Commun. 288, 301–308.
13. Li, C. J., Friedman, D. J., Wang, C., Metelev, V. &Pardee, A. B. (1995). Induction of apoptosis inuninfected lymphocytes by HIV-1 Tat protein.Science, 268, 429–431.
14. Piguet, V. & Trono, D. (2001). Living in oblivion: HIVimmune evasion. Semin. Immunol. 13, 51–57.
15. Amendt, B. A., Si, Z. H. & Stoltzfus, C. M. (1995).Presence of exon splicing silencers within humanimmunodeficiency virus type 1 tat exon 2 and tat-
rev exon 3: evidence for inhibition mediated bycellular factors. Mol. Cell. Biol. 15, 4606–4615.
16. Caputi, M., Mayeda, A., Krainer, A. R. & Zahler,A. M. (1999). hnRNP A/B proteins are required forinhibition of HIV-1 pre-mRNA splicing. EMBO J. 18,4060–4067.
17. Jacquenet, S., Mereau, A., Bilodeau, P. S., Damier, L.,Stoltzfus, C. M. & Branlant, C. (2001). A secondexon splicing silencer within human immuno-deficiency virus type 1 tat exon 2 represses splicingof Tat mRNA and binds protein hnRNP H. J. Biol.Chem. 276, 40464–40475.
18. Zhu, J., Mayeda, A. & Krainer, A. R. (2001). Exonidentity established through differential antagonismbetween exonic splicing silencer-bound hnRNP A1and enhancer-bound SR proteins. Mol. Cell, 8,1351–1361.
19. Tange, T. O. & Kjems, J. (2001). SF2/ASF binds to asplicing enhancer in the third HIV-1 tat exon andstimulates U2AF binding independently of the RSdomain. J. Mol. Biol. 312, 649–662. doi:10.1006/jmbi.2001.4971.
20. Hastings, M. L. & Krainer, A. R. (2001). Pre-mRNAsplicing in the new millennium. Curr. Opin. Cell.Biol. 13, 302–309.
21. Valcarcel, J. & Green, M. R. (1996). The SR proteinfamily: pleiotropic functions in pre-mRNA splicing.Trends Biochem. Sci. 21, 296–301.
22. Zhu, J. & Krainer, A. R. (2000). Pre-mRNA splicing inthe absence of an SR protein RS domain. Genes Dev.14, 3166–3178.
23. Mayeda, A., Screaton, G. R., Chandler, S. D., Fu, X. D.& Krainer, A. R. (1999). Substrate specificities of SRproteins in constitutive splicing are determined bytheir RNA recognition motifs and composite pre-mRNA exonic elements. Mol. Cell. Biol. 19,1853–1863.
24. Fu, X. D., Mayeda, A., Maniatis, T. & Krainer, A. R.(1992). General splicing factors SF2 and SC35 haveequivalent activities in vitro, and both affect alterna-tive 50 and 30 splice site selection. Proc. Natl Acad.Sci. USA, 89, 11224–11228.
25. Gorlach, M., Burd, C. G., Portman, D. S. & Dreyfuss,G. (1993). The hnRNP proteins. Mol. Biol. Rep. 18,73–78.
26. Bilodeau, P. S., Domsic, J. K., Mayeda, A., Krainer,A. R. & Stoltzfus, C. M. (2001). RNA splicing athuman immunodeficiency virus type 1 30 splice siteA2 is regulated by binding of hnRNP A/B proteinsto an exonic splicing silencer element. J. Virol. 75,8487–8497.
27. Eperon, I. C., Makarova, O. V., Mayeda, A., Munroe,S. H., Caceres, J. F., Hayward, D. G. & Krainer, A. R.(2000). Selection of alternative 50 splice sites: role ofU1 snRNP and models for the antagonistic effects ofSF2/ASF and hnRNP A1. Mol. Cell. Biol. 20,8303–8318.
28. Mayeda, A. & Krainer, A. R. (1992). Regulation ofalternative pre-mRNA splicing by hnRNP A1 andsplicing factor SF2. Cell, 68, 365–375.
29. Mayeda, A., Helfman, D. M. & Krainer, A. R. (1993).Modulation of exon skipping and inclusion byheterogeneous nuclear ribonucleoprotein A1 andpre-mRNA splicing factor SF2/ASF. Mol. Cell. Biol.13, 2993–3001.
30. Siebel, C. W., Kanaar, R. & Rio, D. C. (1994).Regulation of tissue-specific P-element pre-mRNAsplicing requires the RNA-binding protein PSI.Genes Dev. 8, 1713–1725.
Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss 651

31. Blanchette, M. & Chabot, B. (1999). Modulation ofexon skipping by high-affinity hnRNP A1-bindingsites and by intron elements that repress splice siteutilization. EMBO J. 18, 1939–1952.
32. Del Gatto, F., Plet, A., Gesnel, M. C., Fort, C. &Breathnach, R. (1997). Multiple interdependentsequence elements control splicing of a fibroblastgrowth factor receptor 2 alternative exon. Mol. Cell.Biol. 17, 5106–5116.
33. Hanamura, A., Caceres, J. F., Mayeda, A., Franza,B. R., Jr & Krainer, A. R. (1998). Regulated tissue-specific expression of antagonistic pre-mRNAsplicing factors. RNA, 4, 430–444.
34. Watakabe, A., Inoue, K., Sakamoto, H. & Shimura, Y.(1989). A secondary structure at the 30 splice siteaffects the in vitro splicing reaction of mouseimmunoglobulin m chain pre-mRNAs. Nucl. AcidsRes. 17, 8159–8169.
35. Mougin, A., Gregoire, A., Banroques, J., Segault, V.,Fournier, R., Brule, F. et al. (1996). Secondarystructure of the yeast Saccharomyces cerevisiae pre-U3A snoRNA and its implication for splicingefficiency. RNA, 2, 1079–1093.
36. Libri, D., Piseri, A. & Fiszman, M. Y. (1991). Tissue-specific splicing in vivo of the beta-tropomyosingene: dependence on an RNA secondary structure.Science, 252, 1842–1845.
37. Muro, A. F., Caputi, M., Pariyarath, R., Pagani, F.,Buratti, E. & Baralle, F. E. (1999). Regulation of fibro-nectin EDA exon alternative splicing: possible role ofRNA secondary structure for enhancer display. Mol.Cell. Biol. 19, 2657–2671.
38. Jacquenet, S., Ropers, D., Bilodeau, P. S., Damier, L.,Mougin, A., Stoltzfus, C. M. & Branlant, C. (2001).Conserved stem-loop structures in the HIV-1 RNAregion containing the A3 30 splice site and its cis-regulatory element: possible involvement in RNAsplicing. Nucl. Acids Res. 29, 464–478.
39. Winkler, W. C., Grundy, F. J., Murphy, B. A. &Henkin, T. M. (2001). The GA motif: an RNA elementcommon to bacterial antitermination systems, rRNA,and eukaryotic RNAs. RNA, 7, 1165–1172.
40. Klein, D. J., Schmeing, T. M., Moore, P. B. & Steitz,T. A. (2001). The kink-turn: a new RNA secondarystructure motif. EMBO J. 20, 4214–4221.
41. Watkins, N. J., Segault, V., Charpentier, B., Nottrott,S., Fabrizio, P., Bachi, A. et al. (2000). A commoncore RNP structure shared between the smallnucleoar box C/D RNPs and the spliceosomal U4snRNP. Cell, 103, 457–466.
42. Burd, C. G. & Dreyfuss, G. (1994). RNA bindingspecificity of hnRNP A1: significance of hnRNP A1high-affinity binding sites in pre-mRNA splicing.EMBO J. 13, 1197–1204.
43. Tacke, R. & Manley, J. L. (1995). The human splicingfactors ASF/SF2 and SC35 possess distinct, function-ally significant RNA binding specificities. EMBO J.14, 3540–3551.
44. Cavaloc, Y., Bourgeois, C. F., Kister, L. & Stevenin, J.(1999). The splicing factors 9G8 and SRp20 transacti-
vate splicing through different and specificenhancers. RNA, 5, 468–483.
45. Muhlemann, O., Yue, B. G., Petersen-Mahrt, S. &Akusjarvi, G. (2000). A novel type of splicingenhancer regulating adenovirus pre-mRNA splicing.Mol. Cell. Biol. 20, 2317–2325.
46. Duc Dodon, M., Hamaia, S., Martin, J. & Gazzolo, L.(2002). Heterogeneous nuclear ribonucleoprotein A1interferes with the binding of the human T cellleukemia virus type 1 Rex regulatory protein to itsresponse element. J. Biol. Chem. 277, 18744–18752.
47. Pongoski, J., Asai, K. & Cochrane, A. (2002). Positiveand negative modulation of human immuno-deficiency virus type 1 Rev function by cis and transregulators of viral RNA splicing. J. Virol. 76,5108–5120.
48. Peloponese, J. M., Jr, Collette, Y., Gregoire, C., Bailly,C., Campese, D., Meurs, E. F. et al. (1999). Full pep-tide synthesis, purification, and characterization ofsix Tat variants. Differences observed between HIV-1 isolates from Africa and other continents. J. Biol.Chem. 274, 11473–11478.
49. Charneau, P., Alizon, M. & Clavel, F. (1992). Asecond origin of DNA plus-strand synthesis isrequired for optimal human immunodeficiencyvirus replication. J. Virol. 66, 2814–2820.
50. Nour, M., Naimi, A., Beck, G. & Branlant, C. (1995).16S-23S and 23S-5S intergenic spacer regions ofStreptococcus thermophilus and Streptococcus salivarius,primary and secondary structure. Curr. Microbiol. 31,270–278.
51. Ratner, L., Haseltine, W., Patarca, R., Livak, K. J.,Starcich, B., Josephs, S. F. et al. (1985). Completenucleotide sequence of the AIDS virus, HTLV-III.Nature, 313, 277–284.
52. Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989).Molecular Cloning: A Laboratory Manual, 2nd edit.,Cold Spring Harbor Laboratory Press, Cold SpringHarbor, NY.
53. Damier, L., Domenjoud, L. & Branlant, C. (1997). TheD1-A2 and D2-A2 pairs of splice sites from humanimmunodeficiency virus type 1 are highly efficientin vitro, in spite of an unusual branch site. Biochem.Biophys. Res. Commun. 237, 182–187.
54. England, T., Bruce, A. & Uhlenbeck, O. (1980).Specific labeling of 30 termini of RNA with T4 ligase.Methods Enzymol. 65, 65–74.
55. Vassilenko, S. K. & Babkina, G. T. (1965). Isolationand properties of ribonuclease isolated from cobravenom. Biokhymiya, 30, 705–712.
56. Jaeger, J. A., Turner, D. H. & Zuker, M. (1989).Improved predictions of secondary structures forRNA. Proc. Natl Acad. Sci. USA, 86, 7706–7710.
57. Portman, D. S. & Dreyfuss, G. (1994). RNA annealingactivities in HeLa nuclei. EMBO J. 13, 213–221.
58. Thompson, J. D., Higgins, D. G. & Gibson, T. J.(1994). CLUSTAL W: improving the sensitivity ofprogressive multiple sequence alignment throughsequence weighting, position-specific gap penaltiesand weight matrix choice. Nucl. Acids Res. 22,4673–4680.
Edited by J. Karn
(Received 20 May 2002; received in revised form 4 September 2002; accepted 5 September 2002)
652 Cooperative Binding of hnRNP A1 to HIV-1 A7 30ss
Related Documents











