A human homolog of mouse Lbh gene, hLBH, expresses in heart and activates SRE and AP-1 mediated MAPK signaling pathway Jianping Ai Yuequn Wang Kunrong Tan Yun Deng Na Luo Wuzhou Yuan Zequn Wang Yongqing Li Ying Wang Xiaoyang Mo Chuanbing Zhu Zhaochu Yin Mingyao Liu Xiushan Wu Received: 20 November 2006 / Accepted: 26 February 2007 / Published online: 28 March 2007 ȑ Springer Science+Business Media B.V. 2007 Abstract It has been reported that mouse Lbh (limb-bud and heart) can regulate cardiac gene expression by modu- lating the combinatorial activities of key cardiac tran- scription factors, as well as their individual functions in cardiogenesis. Here we report the cloning and character- ization of the human homolog of mouse Lbh gene, hLBH, from a human embryonic heart cDNA library. The cDNA of hLBH is 2927 bp long, encoding a protein product of 105 amino acids. The protein is highly conserved in evolution across different species from zebra fish, to mouse, to hu- man. Northern blot analysis indicates that a 2.9 kb tran- script specific for hLBH is most abundantly expressed in both embryonic and adult heart tissue. In COS-7 cells, hLBH proteins are localized to both the nucleus and the cytoplasm. hLBH is a transcription activator when fused to Gal-4 DNA-binding domain. Deletion analysis indicates that both the N-terminal containing proline-dependent serine/threonine kinase group and the C-terminal contain- ing ERK D-domain motif are required for transcriptional activation. Overexpression of hLBH in COS-7 cells acti- vates the transcriptional activities of activator protein-1 (AP-1) and serum response element (SRE). These results suggest that hLBH proteins may act as a transcriptional activator in mitogen-activated protein kinase signaling pathway to mediate cellular functions. Keywords hLBH Á Transcription factor Á Gene regulation Á SRE Á AP-1 Abbreviations DMEM Dulbecco’s Modified Eagle Medium DAPI 4¢,6¢-diamidino-2- phenylindole hydrochloride MAPK Mitogen-activated protein kinase MAPKK, MKK or MEK MAPK kinase MAPKKK or MEKK A MAPKK kinase or MEK kinase SRE Serum response element AP-1 Activation protein 1 Much progress has been made toward a physiological and clinical understanding of cardiac development and heart disease. However, the molecular mechanisms underlying these processes are still not completely understood [1]. Cardiogenesis requires precise regulation of gene expression in a temporal or tissue specific manner. Many transcriptional factors play a key role in these complex processes [1–3]. Mouse Lbh (limb-bud and heart) is a member of a highly conserved family of small acidic proteins in vertebrates, which displays a unique spatiotemporal gene expression pattern during early mouse heart development. Lbh has transcriptional activation activity in mammalian cells and could act as a transcriptional coactivator involved in The authors Jianping Ai and Yuequn Wang have contributed equally to the work. J. Ai Á Y. Wang Á K. Tan Á Y. Deng Á N. Luo Á W. Yuan Á Z. Wang Á Y. Li Á Y. Wang Á X. Mo Á C. Zhu Á Z. Yin Á M. Liu (&) Á X. Wu (&) The Center For Heart Development, Key Lab of MOE for Development Biology and Protein Chemistry, College of Life Sciences, Hunan Normal University, Changsha, Hunan 410081, Peoples’ Republic of China e-mail: [email protected] X. Wu email:[email protected] 123 Mol Biol Rep (2008) 35:179–187 DOI 10.1007/s11033-007-9068-4

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A human homolog of mouse Lbh gene, hLBH, expresses in heartand activates SRE and AP-1 mediated MAPK signaling pathway
Jianping Ai Æ Yuequn Wang Æ Kunrong Tan Æ Yun Deng Æ Na Luo ÆWuzhou Yuan Æ Zequn Wang Æ Yongqing Li Æ Ying Wang ÆXiaoyang Mo Æ Chuanbing Zhu Æ Zhaochu Yin Æ Mingyao Liu ÆXiushan Wu
Received: 20 November 2006 / Accepted: 26 February 2007 / Published online: 28 March 2007
� Springer Science+Business Media B.V. 2007
Abstract It has been reported that mouse Lbh (limb-bud
and heart) can regulate cardiac gene expression by modu-
lating the combinatorial activities of key cardiac tran-
scription factors, as well as their individual functions in
cardiogenesis. Here we report the cloning and character-
ization of the human homolog of mouse Lbh gene, hLBH,
from a human embryonic heart cDNA library. The cDNA
of hLBH is 2927 bp long, encoding a protein product of 105
amino acids. The protein is highly conserved in evolution
across different species from zebra fish, to mouse, to hu-
man. Northern blot analysis indicates that a 2.9 kb tran-
script specific for hLBH is most abundantly expressed in
both embryonic and adult heart tissue. In COS-7 cells,
hLBH proteins are localized to both the nucleus and the
cytoplasm. hLBH is a transcription activator when fused to
Gal-4 DNA-binding domain. Deletion analysis indicates
that both the N-terminal containing proline-dependent
serine/threonine kinase group and the C-terminal contain-
ing ERK D-domain motif are required for transcriptional
activation. Overexpression of hLBH in COS-7 cells acti-
vates the transcriptional activities of activator protein-1
(AP-1) and serum response element (SRE). These results
suggest that hLBH proteins may act as a transcriptional
activator in mitogen-activated protein kinase signaling
pathway to mediate cellular functions.
Keywords hLBH � Transcription factor �Gene regulation � SRE � AP-1
Abbreviations
DMEM Dulbecco’s Modified Eagle
Medium
DAPI 4¢,6¢-diamidino-2-
phenylindole hydrochloride
MAPK Mitogen-activated protein
kinase
MAPKK, MKK or MEK MAPK kinase
MAPKKK or MEKK A MAPKK kinase or MEK
kinase
SRE Serum response element
AP-1 Activation protein 1
Much progress has been made toward a physiological and
clinical understanding of cardiac development and heart
disease. However, the molecular mechanisms underlying
these processes are still not completely understood [1].
Cardiogenesis requires precise regulation of gene expression
in a temporal or tissue specific manner. Many transcriptional
factors play a key role in these complex processes [1–3].
Mouse Lbh (limb-bud and heart) is a member of a highly
conserved family of small acidic proteins in vertebrates,
which displays a unique spatiotemporal gene expression
pattern during early mouse heart development. Lbh has
transcriptional activation activity in mammalian cells and
could act as a transcriptional coactivator involved in
The authors Jianping Ai and Yuequn Wang have contributed
equally to the work.
J. Ai � Y. Wang � K. Tan � Y. Deng � N. Luo �W. Yuan � Z. Wang � Y. Li � Y. Wang � X. Mo �C. Zhu � Z. Yin � M. Liu (&) � X. Wu (&)
The Center For Heart Development, Key Lab of MOE for
Development Biology and Protein Chemistry, College of Life
Sciences, Hunan Normal University, Changsha, Hunan 410081,
Peoples’ Republic of China
e-mail: [email protected]
X. Wu
email:[email protected]
123
Mol Biol Rep (2008) 35:179–187
DOI 10.1007/s11033-007-9068-4

molecular pathways that pattern the limb and the heart [4].
Lbh deregulation interferes with normal cardiac develop-
ment, in part through the attenuation of Nkx2.5 and Tbx5
transcription factor function and the Lbh transgene also
interfers with Gata4-dependent pathways. Lbh might play
an important role in transcriptional control during normal
cardiogenesis [5].
Partial trisomy 2p syndrome includes a spectrum of con-
genital heart disease (CHD) that is characterized by complex
malformations of the outflow and inflow tracts, defects in
cardiac septation, heart position, as well as abnormal ven-
tricular development. The human LBH (hLBH) maps to
chromosome 2p23, a genomic region related to CHD in
partial trisomy 2p syndrome. Previous studies have impli-
cated hLBH as a candidate gene for CHD associated with
partial trisomy 2p syndrome [5]. To specifically study the
function of the human homolog of mouse Lbh, hLBH, in
heart development and disease, we have cloned the hLBH
gene from a human embryonic heart cDNA library. Northern
blot analysis demonstrated that hLBH is most abundantly
expressed in heart at both adult and embryonic stages, which
implies that hLBH might play a role in heart development.
hLBH has a predicted 105 amino acid open reading frame
(ORF), encoding a putative protein that has a molecular
weight of approximately 16.2 kDa. The N-terminal region of
hLBH protein contains a proline-dependent serine/threonine
kinase group, and its C-terminal contains an ERK D-domain
motif (a binding site of ERK), suggesting that hLBH may
have a potential role in cell signaling.
Mitogen-activated protein kinases (MAPK) are major
components of pathways controlling embryogenesis, cell
differentiation, cell proliferation, and cell death. One of the
most explored functions of MAPK signaling is the regu-
lation of gene expression by direct or indirect phosphory-
lation and subsequent activation of transcription factors [6].
Recent studies suggest that MAPK pathways are critical
not only to the response of cardiovascular cells to extra-
cellular stress but also to developmental cues that regulate
cardiovascular development [7, 8]. In mammals, MAPK
pathways are involved in multiple cellular processes
through phosphorylation of specific endpoint targets, such
as ELK-1 and SRE, which together with SRF compose a
ternary complex that induces expression of c-fos and other
early growth response genes controlling the transition from
quiescence to proliferation [7]. The c-fos products hetero-
dimerize with c-Jun proteins to form AP-1 complexes [8].
Activation of AP-1 involves the direct phosphorylation/
dephosphorylation of AP-1 components as well as the
phosphorylation and activation of transcription factors that
induce elevated expression of c-Jun or c-fos. hLBH is a
transcription activator when fused to Gal-4 DNA-binding
domain. Overexpression of hLBH in COS-7 cells activates
the transcriptional activities of activator protein-1 (AP-1)
and serum response element (SRE), suggesting that hLBH
proteins may act as a transcriptional activator in mitogen-
activated protein kinase signaling pathway to mediate
cellular functions.
Materials and methods
Construction of cDNA library of human embryonic
heart
The 20-week human embryonic heart cDNA library was
constructed as reported previously [9]. Briefly, 5 lg
mRNA was purified from 500 lg total human embryonic
heart RNA using Rapid mRNATM purification Kit (Am-
resco). Reverse transcription reactions were performed
with the purified embryonic heart mRNA and Oligo dT-RA
primer according to cDNA Synthesis kit protocol (Takara).
After Cassette Adaptor Ligation reactions using cDNA
PCR Library Kit, cDNA amplification reactions were per-
formed with RA primer (5¢-CTGATCTAGACCTGCA
GGCTC-3), CA primer (5¢-CGTGGTACCATGGT CTA
GAGT-3), and Ex Taq (Takara).
Full-length hLBH cDNA cloning and bioinformatics
analysis
The nucleic acid sequence of mouse Lbh was obtained
from NCBI (http://www.ncbi.nlm.nih.gov) and used to
search human EST database with the BLAST searching
program (http://www.ncbi.nlm.nih.gov/blast). Through a
combined BLAST search as previously described [10], a
number of ESTs presenting the same gene (hLBH) were
identified in our database search. The first forward primer
in BM543364 and the reverse primer in BF718797
(POUT1 and POUT2, Table 1), and the second forward
primer in BM543364 and the reverse primer in BF718797
(PIN3 and PIN4, Table 1) were designed using Primer
Premier 5.0 to perform standard PCR. hLBH sequences of
ORF were confirmed by PCR amplification with a pair of
primers (PORF1 and PORF2, Table 1). The 5¢-gene spe-
cific primers (PS and PAS, Table 1) were designed for 5¢-rapid amplification of cDNA ends (RACE) according to
standard PCR methods [9, 11]. 5¢-RACE was performed
using SMARTTM RACE cDNA Amplification Kit
(Clonetech). All the PCR products were then cloned into
pMD18-T-vector (Takara) and sequenced with 377 DNA
Sequencer (ABI PRISM). Sequence analysis was per-
formed using the DNAstar program and BLAST program
from NCBI. Blastn program was used to identify the locus
of genes and to look for exons and introns. BLASTn and
Pfam 9.0 were used to analyze genomic structure and the
protein domain, respectively. The homologues of hLBH
were found with BLASTp, and the sequence alignment and
180 Mol Biol Rep (2008) 35:179–187
123

phylogenetic tree analysis were performed with MegAlign
program (DNAstar).
RNA isolation and Northern blot hybridization
Human tissues from therapeutically aborted fetuses were
obtained under the approval of Changsha Women and
Children’s Hospital, People’s Republic of China, with the
consent of the patients and in accordance with university
policy. Total RNA was isolated from various tissues
(muscle, cerebellum, cerebrum, tongue, lung, adrenal,
heart, intestine, liver from 17-week stage, and lung, liver,
muscle, kidney, brain, heart, intestine, pancreas from 25-
week stage) using standard methods [9]. 20 lg samples of
each tissue were separated by electrophoresis through
formaldehyde-agarose gel. The embryo multiple tissues
Northern blots were prepared as described in previous
studies [9]. A commercially available Northern blot con-
taining mRNA from eight adult tissues was purchased
from Clontech. The adult human Multiple Tissue North-
ern blot and the two multiple embryonic tissue blots were
hybridized with cDNA probe of hLBH. The hLBH cDNA
was labeled with [a-32p] dCTP using a Random Primer
Labeling Kit (Takara). The blots were then exposed to
X-ray films at –80�C for several days and the films were
developed. The blots were then stripped and probed with
radiolabeled b-actin cDNA (Clontech) as indicator of
mRNA loading.
Plasmid construction
The following plasmids were constructed and used for
mammalian cell transfection. (i) Construction of pEGFP-
N1-hLBH: to generate a fusion protein of hLBH with
enhanced green fluorescent protein (EGFP), the coding
region of hLBH was subcloned into the XhoI and HindIII
sites of pEGFP-N1 vector in-frame with the AAG instead
of the TAG stop codon at the end of hLBH coding se-
quence. (ii) Construction of pCMV-BD-hLBH-AS1
(pGAL4-hLBH, amino acids 1–105), and two deletion
mutations: pCMV-BD-hLBH-AS2 (amino acids 1–58 with
proline-dependent serine/threonine kinase group) and
pCMV-BD-hLBH-AS3 (amino acids 54–105 with ERK
D-domain motif): the DNA fragment containing the
whole coding region of hLBH was amplified with primers
P1 and P2 containing EcoRI and SalI sites, respectively
(Table 1). The amplified DNA fragment was cloned into
pMD18-T-vector, and then the EcoRI and SalI fragment
of hLBH was subcloned into a pCMV-BD expression
vector that contained the GAL4 DNA-binding domain
(DBD) to create a fusion protein between GAL4 DBD
and hLBH ORF. The fragments of AS2 and AS3 were
amplified separately by PCR from the pMD18T-hLBH
plasmid with two pairs of primers P1 and P4, P3 and P2.
Subsequently, EcoRI and SalI fragments were subcloned
in-frame into expression vector pCMV-BD to construct
pCMV-BD-hLBH-AS2, pCMV-BD-hLBH-AS3. (iii)
Table 1 PCR primers,
programs and cyclesPrimers Nucleotide sequences Programs Cycles
POUT1 5¢-GAGTTGTGTCCACCTTGCCGAC-3¢ 94�C, 30 s; 32
POUT2 5¢-AGGAGGAGGCCAGATAGGAGCT-3¢ 54�C, 30 s;
72�C, 2 min 30 s
PIN3 5¢-GGGACGCAGGGACCGTTTTTAA-3¢ 94�C, 30 s; 30
PIN4 5¢-GGCCAGATAGGAGCTAAGCCAT-3¢ 56�C, 30 s;
72�C, 2 min
PORF1 5¢-CCTCGAGCTTCATGTCTATATATTTCCC-3¢ 94�C, 30 s; 30
PORF2 5¢-GGA AGCTTCTGCTCTTTATTTTCTTTCG-3¢ 56.5�C, 30 s;
72�C, 1 min 30 s
PS 5¢-AGATACCCCACCAAACCCA-3¢ 94�C, 30 s; 32
PAS 5¢-GGGGCAGTGAATGGGGAAA-3¢ 51�C, 30 s;
72�C, 1 min 30 s
P1 5¢-CCCGAATTCTTCATGTCTATATATTTCC-3¢ 94�C, 30 s; 30
P2 5¢-GGAAGCTTCTGCTCTTTATTTTCTTTCG-3¢ 58�C, 30 s;
72�C, 1 min
P1 5¢-CCCGAATTCTTCATGTCTATATATTTCC-3¢ 94�C, 30 s; 30
P4 5¢-CCTAAGCTTAGCAGCAGCGGTCAAAATC-3¢ 58�C, 30 s;
72�C, 1 min
P3 5¢-GATGAATTCTCCTACCAGATCTTCCCAG-3¢ 94�C, 30 s; 30
P2 5¢-GGAAGCTTCTGCTCTTTATTTTCTTTCG-3¢ 58�C, 30 s;
72�C, 45 s
Mol Biol Rep (2008) 35:179–187 181
123

Construction of pCMV-Tag2B-hLBH: the expression
plasmid for FLAG epitopetagged hLBH, pCMV-Tag2B-
hLBH, was constructed by inserting hLBH ORF down-
stream of the FLAG epitope sequence (MDYKDDDDK)
in a pCMV-Tag2B expression vector.
Cell culture, transient transfection, and subcellular
localization analysis
COS-7 cells were maintained and passaged according to
standard methods in Dulbecco’s Modified Eagle Medium
(DMEM, Gibco BRL) supplemented with 10% fetal calf
serum (FCS) in an humidified atmosphere of 95% air and
5% CO2. Cells were transfected with pEGFP-N1-hLBH
using LipofectAMINE (Invitrogen) according to the
method described previously [12]. Forty-eight hours after
transfection, cells were fixed with 4% paraformaldehyde
for 15 min and washed with PBS three times and nuclei
were stained with 4¢, 6¢-diamidino-2-phenylindole hydro-
chloride (DAPI). Subcellular localization of the EGFP-
hLBH fusion protein was detected using an inverted fluo-
rescence microscopy (Nikon, E400).
Fig. 1 Nucleotide sequence
and deduced protein sequence
of the hLBH gene. hLBHencodes a polypeptide of 105
amino acids. Both nucleotides
and amino acids are numbered
at the left and right side of each
line, respectively. The initiation
ATG and TGA stop codons are
boxed. The putative
polyadenylation signal sequence
aataaa is underlined and in gray.
A proline-dependent serine/
threonine kinase group (PMEE
IGLSPRKDGLS) and an ERK
D-domain (LKDR
LPSIVVEPTEG) were
underlined
182 Mol Biol Rep (2008) 35:179–187
123

Transient expression reporter gene assay and deletion
analysis
To understand potential transcriptional activity of hLBH,
the pCMV-BD-hLBH-AS1, pCMV-BD-hLBH-AS2,
pCMV-BD-hLBH-AS3 and pCMV-BD were transiently
co-transfected into COS-7 cells along with the pL8G5-Luc
reporter and pLexA-VP16 using LipofectAMINE as de-
scribed above [12]. Forty-eight hours later, the luciferase
activity assay was performed according to the protocols of
Stratagene [12]. The luciferase activity was normalized for
transfection efficiency by co-transfection with pCMV-lacZ
and spectrophotometry analysis. The data are the mean of
three repeats in a single transfection experiment after
normalization for b-galactosidease activity. Each experi-
ment was repeated at least three times.
Luciferase was assayed in COS-7 cells by cotransfecting
pCMV-Tag2B-hLBH, and the luciferase reporter plasmid
pAP-1-Luc (or pSRE-Luc). Forty-eight hours later, lucif-
erase activity was measured [12]. Each experiment was
performed in triplicate and each assay was repeated at least
three times. The means of the data from three individual
transfected wells are presented after normalization for b-
galactosidase activity.
Results and discussion
Molecular characterization and evolutionary
conservation of the hLBH gene
The mouse Lbh regulates cardiac gene expression by
modulating the combinatorial activities of key cardiac
transcription factors, as well as their individual functions in
cardiogenesis [5]. To specifically study the function of the
human homolog of mouse Lbh, hLBH, in heart develop-
ment and cardiogenesis, we cloned the hLBH gene through
combined BLAST search and PCRs analysis as previously
Table 2 Genomic structure of the hLBH gene
Extron number Extron size (bp) Splice donor site Intron size (bp) Splice acceptor site
I 211 gtgagtaccctgcg 1340 gggtttcttggcag
II 103 gtgagtaagtcctg 22925 tgcttttctttcag
III 2591 ttccactctggtct
Summary of the size of the extrons and introns of the hLBH gene based on comparison of the cDNA and the genomic sequence. Invariant
nucleotides (gt/ag) are in boldface type
Fig. 2 hLBH is conserved
during evolution. (A) The
phylogenetic tree analysis of
hLBH proteins and its
homologs. The amino acid
sequences of H. sapiens hLBH
and its homologues in Pongopygmaeus (CAH90344), Musmusculus (NP_084275.3),
Gallus gallus (XP-419367.1),
Tetraodon nigroviridis(CAG10379), Danio rerio(NP_956814.1) are used in
analysis. (B) Comparison of the
amino acid sequences of hLBH,
mouse LBH and Danio reriovXcl2. Residues that are highly
conserved among these
sequences are indicated within
the top majority sequence. The
identical amino acid residues
are boxed and shaded in dark
Mol Biol Rep (2008) 35:179–187 183
123

described [10]. We screened the human EST database with
the nucleotide sequence of the mouse LBH gene. To confirm
the cDNA sequences identified from the database, two pairs
of primers (POUT1/POUT2 and PIN3/PIN4, Table 1) based
on the sequences of four overlapping ESTs (AL530914,
BM472620, BU191949 and BF718797) were used to carry
out standard PCR using the human embryonic heart cDNA
library as template. A PCR product fragment of 2608 bp was
obtained and confirmed to be the cDNA sequences of hLBH.
To obtain the full-length of cDNA, 5¢-RACE were performed
using 5¢-gene specific primers (PS and PAS) as described
previously [9, 11]. The procedure yielded a 216 bp DNA
fragment for 5¢-RACE. The products were then cloned and
sequenced. A sequence of 2927 bp constituting the full-
length novel gene was assembled, which was named hLBH
as approved by the HUGO Nomenclature Committee. The
nucleotide sequence reported here is available in GenBank
with accession number EF025586.
The hLBH gene consists of an ORF of 318 bp long
extending from the first ATG codon at nucleotide 186 to a
TGA stop codon at 503, a 185 bp 5¢-untranslated region
(UTR), and a 2424 bp 3¢-UTR with a consensus poly-
adenylation signal (aataaa) (Fig. 1). The deduced hLBH
protein has 105 amino acids (Fig. 1) with a calculated
molecular mass of approximately 16.2 kDa. Comparison of
the hLBH sequence with the genomic sequence shows that
hLBH is mapped to chromosome 2p23 and spans approx-
imately 28.5 kb on the genome. Interestingly, chromo-
somal band 2p23 is frequently triplicated in partial trisomy
2p syndrome and has been related to CHD associated with
this syndrome [5].
The hLBH gene consists of three exons and two introns.
The exon–intron boundaries conform to the consensus
splicing signals, where there are a gt and an ag dinucleo-
tide at the 5¢-donor and 3¢-acceptor site, respectively (Ta-
ble 2).
We then identified the homologues of Homo sapiens
hLBH in Pongo pygmaeus (CAH90344), Mus musculus
(NP_084275.3), Gallus gallus (XP-419367.1), Tetraodon
nigroviridis (CAG10379), Danio rerio (NP_956814.1).
The amino acid sequence of hLBH was aligned with its
homologues and an evolutionary relationship among these
proteins was examined using phylogenetic tree analysis.
hLBH is one of the conserved proteins during evolution
across different species (Fig. 2A) and hLBH protein shares
more than 95% amino acid sequence identity with mouse
Lbh protein (Fig. 2B).
Analysis of hLBH protein using the Motif Scan program
(http://scansite.mit.edu) indicates that the protein product
of hLBH contains a proline-dependent serine/threonine
kinase group (amino acids 27–41, PMEE IGLSPRKDGLS)
and an ERK D-domain (amino acids 57–71, LKDR
LPSIVVEPTE G) (Fig. 1).
hLBH is strongly expressed in heart at embryonic and
adult stages
To characterize the transcript size and expression patterns
of hLBH, adult and fetal multiple tissue northern blot were
performed using hLBH cDNA as the probe. A 2.9 kb
transcript of hLBH was most abundantly detected in human
adult heart with varying low level of expression in pla-
centa, lung, skeletal muscle, kidney and liver (Fig. 3A).
hLBH was detected mainly in heart tissues of both 25 and
17 weeks human embryo (Fig. 3B and C). The results
suggest that hLBH protein plays a role in human embryo
heart development.
Subcellular localization of hLBH protein
We examined the subcellular localization of hLBH to
determine whether it could function as a transcriptional
regulator. pEGFP-N1-hLBH was transfected into COS-7
Fig. 3 Expression of hLBH in human adult and embryonic tissues
analyzed using Northern blot. A commercially available Northern blot
membrane containing multiple human adult tissues and two
membranes containing multiple human embryonic tissues were
hybridized with a-32P random-labeled hLBH cDNA probe. b-actin
was used as a control for the amount of loaded mRNA (total RNA). A
transcript of ~2.9 kb specific for hLBH was detected in adult (A) and
embryo membranes of 25 weeks (B) and 17 weeks (C)
184 Mol Biol Rep (2008) 35:179–187
123
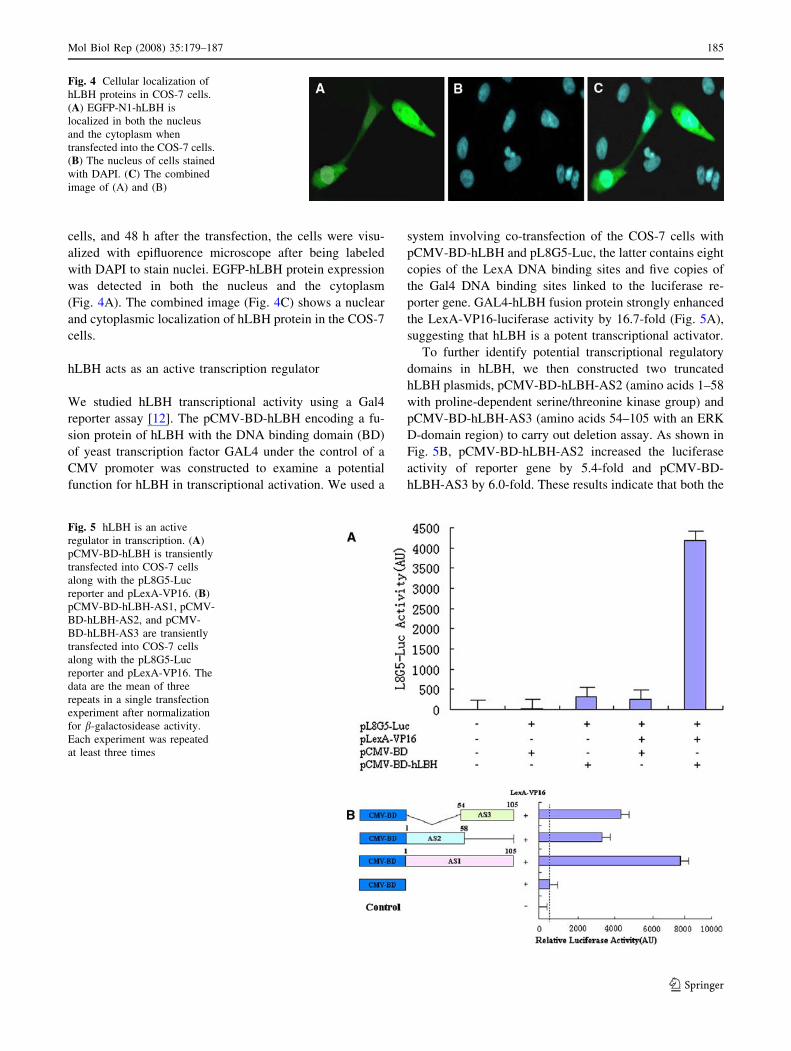
cells, and 48 h after the transfection, the cells were visu-
alized with epifluorence microscope after being labeled
with DAPI to stain nuclei. EGFP-hLBH protein expression
was detected in both the nucleus and the cytoplasm
(Fig. 4A). The combined image (Fig. 4C) shows a nuclear
and cytoplasmic localization of hLBH protein in the COS-7
cells.
hLBH acts as an active transcription regulator
We studied hLBH transcriptional activity using a Gal4
reporter assay [12]. The pCMV-BD-hLBH encoding a fu-
sion protein of hLBH with the DNA binding domain (BD)
of yeast transcription factor GAL4 under the control of a
CMV promoter was constructed to examine a potential
function for hLBH in transcriptional activation. We used a
system involving co-transfection of the COS-7 cells with
pCMV-BD-hLBH and pL8G5-Luc, the latter contains eight
copies of the LexA DNA binding sites and five copies of
the Gal4 DNA binding sites linked to the luciferase re-
porter gene. GAL4-hLBH fusion protein strongly enhanced
the LexA-VP16-luciferase activity by 16.7-fold (Fig. 5A),
suggesting that hLBH is a potent transcriptional activator.
To further identify potential transcriptional regulatory
domains in hLBH, we then constructed two truncated
hLBH plasmids, pCMV-BD-hLBH-AS2 (amino acids 1–58
with proline-dependent serine/threonine kinase group) and
pCMV-BD-hLBH-AS3 (amino acids 54–105 with an ERK
D-domain region) to carry out deletion assay. As shown in
Fig. 5B, pCMV-BD-hLBH-AS2 increased the luciferase
activity of reporter gene by 5.4-fold and pCMV-BD-
hLBH-AS3 by 6.0-fold. These results indicate that both the
Fig. 4 Cellular localization of
hLBH proteins in COS-7 cells.
(A) EGFP-N1-hLBH is
localized in both the nucleus
and the cytoplasm when
transfected into the COS-7 cells.
(B) The nucleus of cells stained
with DAPI. (C) The combined
image of (A) and (B)
Fig. 5 hLBH is an active
regulator in transcription. (A)
pCMV-BD-hLBH is transiently
transfected into COS-7 cells
along with the pL8G5-Luc
reporter and pLexA-VP16. (B)
pCMV-BD-hLBH-AS1, pCMV-
BD-hLBH-AS2, and pCMV-
BD-hLBH-AS3 are transiently
transfected into COS-7 cells
along with the pL8G5-Luc
reporter and pLexA-VP16. The
data are the mean of three
repeats in a single transfection
experiment after normalization
for b-galactosidease activity.
Each experiment was repeated
at least three times
Mol Biol Rep (2008) 35:179–187 185
123
proline-dependent serine/threonine kinase group at N-ter-
minal and the ERK D-domain region at C-terminal con-
tribute to the transcriptional activation of hLBH.
hLBH activates SRE and AP-1-mediated transcriptional
activation
To investigate the role of hLBH proteins, we examined
whether hLBH was directly or indirectly involved in the
regulation of transcription factors. MAPK signal trans-
duction pathways are the most widespread mechanisms of
eukaryotic cell regulation [13]. To examine the effect of
hLBH on this specific cell-signaling pathway, we per-
formed pathway-specific reporter gene assays to measure
the transcription of AP-1 and SRE by hLBH in the COS-7
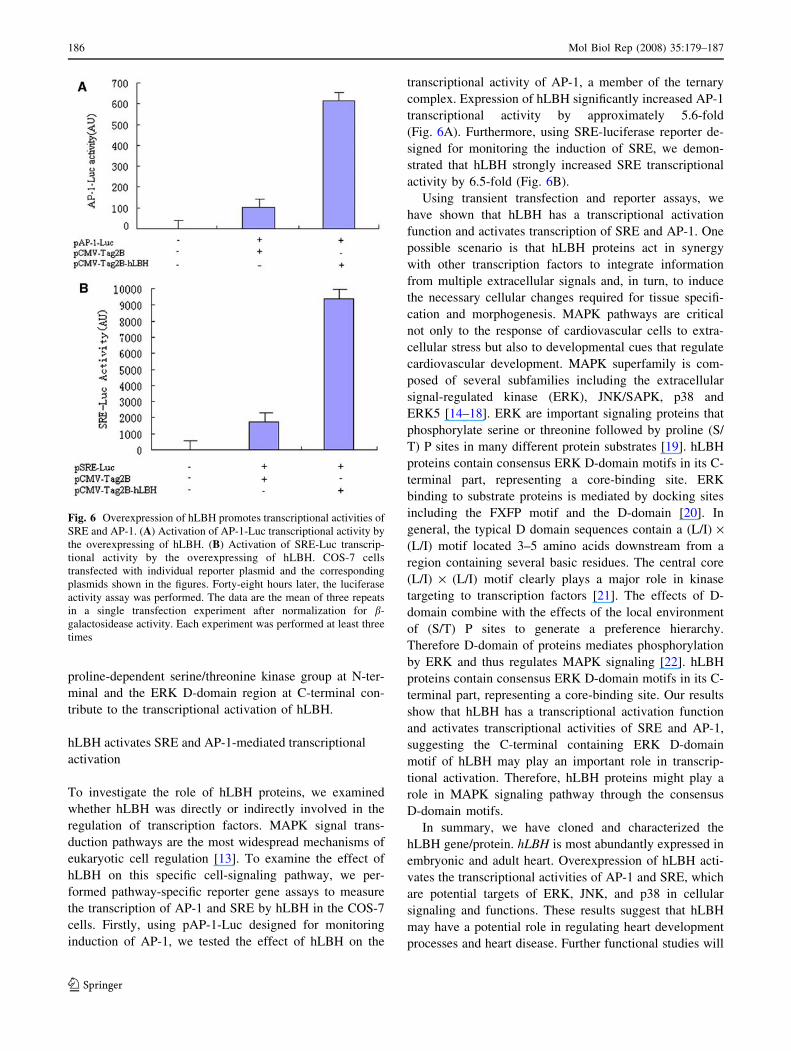
cells. Firstly, using pAP-1-Luc designed for monitoring
induction of AP-1, we tested the effect of hLBH on the
transcriptional activity of AP-1, a member of the ternary
complex. Expression of hLBH significantly increased AP-1
transcriptional activity by approximately 5.6-fold
(Fig. 6A). Furthermore, using SRE-luciferase reporter de-
signed for monitoring the induction of SRE, we demon-
strated that hLBH strongly increased SRE transcriptional
activity by 6.5-fold (Fig. 6B).
Using transient transfection and reporter assays, we
have shown that hLBH has a transcriptional activation
function and activates transcription of SRE and AP-1. One
possible scenario is that hLBH proteins act in synergy
with other transcription factors to integrate information
from multiple extracellular signals and, in turn, to induce
the necessary cellular changes required for tissue specifi-
cation and morphogenesis. MAPK pathways are critical
not only to the response of cardiovascular cells to extra-
cellular stress but also to developmental cues that regulate
cardiovascular development. MAPK superfamily is com-
posed of several subfamilies including the extracellular
signal-regulated kinase (ERK), JNK/SAPK, p38 and
ERK5 [14–18]. ERK are important signaling proteins that
phosphorylate serine or threonine followed by proline (S/
T) P sites in many different protein substrates [19]. hLBH
proteins contain consensus ERK D-domain motifs in its C-
terminal part, representing a core-binding site. ERK
binding to substrate proteins is mediated by docking sites
including the FXFP motif and the D-domain [20]. In
general, the typical D domain sequences contain a (L/I) ·(L/I) motif located 3–5 amino acids downstream from a
region containing several basic residues. The central core
(L/I) · (L/I) motif clearly plays a major role in kinase
targeting to transcription factors [21]. The effects of D-
domain combine with the effects of the local environment
of (S/T) P sites to generate a preference hierarchy.
Therefore D-domain of proteins mediates phosphorylation
by ERK and thus regulates MAPK signaling [22]. hLBH
proteins contain consensus ERK D-domain motifs in its C-
terminal part, representing a core-binding site. Our results
show that hLBH has a transcriptional activation function
and activates transcriptional activities of SRE and AP-1,
suggesting the C-terminal containing ERK D-domain
motif of hLBH may play an important role in transcrip-
tional activation. Therefore, hLBH proteins might play a
role in MAPK signaling pathway through the consensus
D-domain motifs.
In summary, we have cloned and characterized the
hLBH gene/protein. hLBH is most abundantly expressed in
embryonic and adult heart. Overexpression of hLBH acti-
vates the transcriptional activities of AP-1 and SRE, which
are potential targets of ERK, JNK, and p38 in cellular
signaling and functions. These results suggest that hLBH
may have a potential role in regulating heart development
processes and heart disease. Further functional studies will
Fig. 6 Overexpression of hLBH promotes transcriptional activities of
SRE and AP-1. (A) Activation of AP-1-Luc transcriptional activity by
the overexpressing of hLBH. (B) Activation of SRE-Luc transcrip-
tional activity by the overexpressing of hLBH. COS-7 cells
transfected with individual reporter plasmid and the corresponding
plasmids shown in the figures. Forty-eight hours later, the luciferase
activity assay was performed. The data are the mean of three repeats
in a single transfection experiment after normalization for b-
galactosidease activity. Each experiment was performed at least three
times
186 Mol Biol Rep (2008) 35:179–187
123

elucidate the roles of hLBH proteins in development and
identify the signals to which they respond.
Acknowledgments We are grateful to all members of the Center for
Heart Development, College of Life Sciences in Hunan Normal
University for their excellent technical assistance and encouragement.
This study was supported in part by the National Natural Science
Foundation of China (No. 90508004, 30470867, 30570934,
30571048), PCSIRT of Education Ministry of China (IRT0445),
National Basic Research Program of China (2005CB522505), the
Foundation of Hunan Province (No. 05FJ2007, 06JJ4120), and Sci-
entific and Research Fund of Hunan Provincial Education Department
(04C327).
References
1. Wu X (2002) Molecular mechanism of heart development. Hunan
Publishing House, Changsha
2. Wu X, Park M, Golden K, Axelrod JD, Bodmer R (1996) The
wingless signaling pathway is directly involved in Drosophilaheart development. Dev Biol 177:104–116
3. Wu X, Golden K, Bodmer R (1995) Heart development in Dro-sophila requires the polarity gene wingless. Dev Biol 169:619–
628
4. Briegel KJ, Joyner AL (2001) Identification and characterization
of Lbh, a novel conserved nuclear protein expressed during early
limb and heart development. Dev Biol 233:291–304
5. Briegel KJ, Baldwin HS, Epstein JA, Joyner AL (2005) Con-
genital heart disease reminiscent of partial trisomy 2p syndrome
in mice transgenic for the transcription factor Lbh. Development
132:3305–3316
6. Whitmarsh AJ, Davis RJ (2000) Regulation of transcription factor
function by phosphorylation. Cell mol Life Sci 57:1172–1183
7. Aggeli IS, Gaitanaki C, Lazou A, Beis I (2002) Hyperosmotic
and thermal stresses activate p38-MAPK in the perfused
amphibian heart. J Exp Biol 205:443–454
8. Tanoue T, Nishida E (2003) Molecular recognitions in the MAP
kinase cascades. Cell Signal 15:455–462
9. Zhu Y, Wang Y, Xia C, Li D, Li Y, Zeng W, Yuan W, Liu H, Zhu
C, Wu X, Liu M (2004) WDR26, a novel G-beta-like protein,
suppresses MAPK signaling pathway. J Cell Biochem 93:579–
587
10. Huang C, Wang Y, Li D, Li Y, Luo J, Yuan W, Ou Y, Zhu C,
Zhang Y, Wang Z, Wu X, Liu M (2004) Inhibition of tran-
scriptional activities of AP-1 and c-Jun by a new zinc-finger
protein ZNF394. Biochem Biophys Res Commun 320:1298–1305
11. Wang Y, Li Y, Zeng W, Zhu C, Xiao J, Yuan W, Wang Y, Cai Z,
Zhou J, Liu M, Wu X (2004) IXL, a new subunit of the mam-
malian mediator complex, functions as a transcriptional sup-
pressor. Biochem Biophys Res Commun 325:1330–1338
12. Wang Y, Li Y, Qi X, Yuan W, Ai J, Zhu C, Cao L, Yang H, Liu
F, Wu X, Liu M (2004) TRIM45, a novel human RBCC/TRIM
protein, inhibits transcriptional activities of ElK-1 and AP-1.
Biochem Biophys Res Commun 323:9–16
13. Reszka AA, Seger R, Diltz CD, Krebs EG, Fischer EH (1995)
Association of mitogen-activated protein kinase with the micro-
tubule cytoskeleton. Proc Natl Acad Sci USA 92:8881–8885
14. English J, Pearson G, Wilsbacher J, Swantek J, Karandikar M, Xu
S, Cobb MH (1999) New insights into the control of MAP kinase
pathways. Exp Cell Res 253:255–270
15. Pawson T, Nash P (2000) Protein–protein interactions define
specificity in signal transduction. Genes Dev 4:1027–1047
16. Treisman R (1996) Regulation of transcription by MAP kinase
cascades. Curr Opin Cell Biol 8:205–215
17. Gille H, Kortenjann M, Thomae O, Moomaw C, Slaughter C,
Cobb MH, Shaw PE (1995) ERK phosphorylation potentiates
Elk-1-mediated ternary complex formation and transactivation.
EMBO J 14(5):951–962
18. Kyriakis JM, Avruch J (2001) Mammalian mitogen-activated
protein kinase signal transduction pathways activated by stress
and inflammation. Physiol Rev 81:807–869
19. Fantz DA, Jacobs D, Glossip D, Kornfeld K (2001) Docking sites
on substrate proteins direct extracellular signal regulated kinase to
phosphorylate specific residues. J Biol Chem 276:27256–27265
20. Sharrocks AD, Yang SH, Galanis A (2000) Docking domains and
substrate-specificity determination for MAP kinases. Trends
Biochem Sci 25:448–453
21. Yang SH, Whitmarsh AJ, Davis RJ, Sharrocks AD (1998) Dif-
ferential targeting of MAP kinases to the ETS-domain tran-
scription factor Elk-1. EMBO J 17:1740–1749
22. Buchwalter G, Gross C, Wasylyk B (2004) Ets ternary complex
transcription factors. Gene 324:1–14
Mol Biol Rep (2008) 35:179–187 187
123
Related Documents











