doi:10.1111/j.1365-2052.2004.01161.x A high-resolution radiation hybrid map of porcine chromosome 6 H. Cao*, J. A. B. Robinson*, Z. Jiang † , J. S. Melville*, S. P. Golovan*, M. W. Jones* and A. M. Verrinder Gibbins* *Department of Animal and Poultry Science, University of Guelph, Guelph, Ont., Canada. † Department of Animal Science, Washington State University, Pullman, WA, USA Summary A high-resolution comprehensive map was constructed for porcine chromosome (SSC) 6, where quantitative trait loci (QTL) for reproduction and meat quality traits have been reported to exist. A radiation hybrid (RH) map containing 105 gene-based markers and 15 microsatellite markers was constructed for this chromosome using a 3000-rad porcine/ hamster RH panel. In total, 40 genes from human chromosome (HSA) 1p36.3-p22, 29 from HSA16q12-q24, 17 from HSA18p11.3-q12 and 19 from HSA19q13.1-q13.4 were assigned to SSC6. All primers for these gene markers were designed based on porcine gene or EST sequences, and the orthologous status of the gene markers was confirmed by direct sequencing of PCR products amplified from separate Meishan and Large White genomic DNA pools. The RH map spans SSC6 and consists of six linkage groups created by using a LOD score threshold of 4. The boundaries of the conserved segments between SSC6 and HSA1, 16, 18 and 19 were defined more precisely than previously reported. This represents the most comprehensive RH map of SSC6 reported to date. Polymorphisms were detected for 38 of 105 gene-based markers placed on the RH map and these are being exploited in ongoing chromosome wide scans for QTL and eventual fine mapping of genes associated with prolificacy in a Meishan · Large White multigenerational commercial population. Keywords porcine chromosome 6, radiation hybrid map, reproduction, sow productivity. Introduction Radiation hybrid (RH) mapping, a somatic cell hybrid technique, was developed as a general approach for con- structing high-resolution maps of mammalian chromo- somes (Cox et al. 1990). This method holds several advantages over other mapping procedures, the most important being that polymorphism is not required and thus the mapping of both polymorphic and monomorphic markers is facilitated (Amarante et al. 2000). Recently, linkage between economically important traits and specific chromosomal regions has been analysed intensively in swine populations. SSC6 has been reported to harbour QTL associated with reproductive (Wilkie et al. 1999; Cassady et al. 2001) and meat quality traits (de Koning et al. 1999; Ovilo et al. 2000; Grindflek et al. 2001), but previous maps of SSC6 lacked the resolution required for further fine mapping of any QTL regions to enable final identification of the underlying genes. Exploi- tation of data from the much more detailed human and murine maps is essential in accelerating the improvement of livestock maps. Accordingly, numerous genes have been mapped on the porcine genome to provide a more detailed correspondence with the human genome (http://www.toulouse.inra.fr/lgc/pig/compare/HSA.htm). As a result, a number of breaks in conservation of synteny between HSA chromosomes and SSC6 have been demon- strated (Pinton et al. 2000; Karnuah et al. 2001; Sun et al. 2001; Jiang et al. 2002; Kiuchi et al. 2002; Rink et al. 2002). Nevertheless, to facilitate the identification of can- didate genes within small chromosome segments, the homologous regions need to be defined more accurately. Radiation hybrid mapping was utilized in this study to link both microsatellite and gene-based markers into a comprehensive high-resolution map with as few break points as possible. The current draft sequence of the human genome (http://www.ncbi.nlm.nih.gov/genome/guide/ human/) and the previous comparative mapping between the human and pig genomes performed by our group (Jiang Address for correspondence J. A. B. Robinson, Department of Animal and Poultry Science, University of Guelph, Guelph, Ont., Canada N1G 2W1. E-mail: [email protected] Accepted for publication 26 May 2004 Ó 2004 International Society for Animal Genetics, Animal Genetics, 35, 367–378 367

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

doi:10.1111/j.1365-2052.2004.01161.x
A high-resolution radiation hybrid map of porcine chromosome 6
H. Cao*, J. A. B. Robinson*, Z. Jiang†, J. S. Melville*, S. P. Golovan*, M. W. Jones*
and A. M. Verrinder Gibbins*
*Department of Animal and Poultry Science, University of Guelph, Guelph, Ont., Canada. †Department of Animal Science, Washington
State University, Pullman, WA, USA
Summary A high-resolution comprehensive map was constructed for porcine chromosome (SSC) 6,
where quantitative trait loci (QTL) for reproduction and meat quality traits have been
reported to exist. A radiation hybrid (RH) map containing 105 gene-based markers and
15 microsatellite markers was constructed for this chromosome using a 3000-rad porcine/
hamster RH panel. In total, 40 genes from human chromosome (HSA) 1p36.3-p22,
29 from HSA16q12-q24, 17 from HSA18p11.3-q12 and 19 from HSA19q13.1-q13.4 were
assigned to SSC6. All primers for these gene markers were designed based on porcine gene
or EST sequences, and the orthologous status of the gene markers was confirmed by direct
sequencing of PCR products amplified from separate Meishan and Large White genomic
DNA pools. The RH map spans SSC6 and consists of six linkage groups created by using a
LOD score threshold of 4. The boundaries of the conserved segments between SSC6 and
HSA1, 16, 18 and 19 were defined more precisely than previously reported. This represents
the most comprehensive RH map of SSC6 reported to date. Polymorphisms were detected for
38 of 105 gene-based markers placed on the RH map and these are being exploited in
ongoing chromosome wide scans for QTL and eventual fine mapping of genes associated
with prolificacy in a Meishan · Large White multigenerational commercial population.
Keywords porcine chromosome 6, radiation hybrid map, reproduction, sow productivity.
Introduction
Radiation hybrid (RH) mapping, a somatic cell hybrid
technique, was developed as a general approach for con-
structing high-resolution maps of mammalian chromo-
somes (Cox et al. 1990). This method holds several
advantages over other mapping procedures, the most
important being that polymorphism is not required and thus
the mapping of both polymorphic and monomorphic
markers is facilitated (Amarante et al. 2000).
Recently, linkage between economically important traits
and specific chromosomal regions has been analysed
intensively in swine populations. SSC6 has been reported to
harbour QTL associated with reproductive (Wilkie et al.
1999; Cassady et al. 2001) and meat quality traits
(de Koning et al. 1999; Ovilo et al. 2000; Grindflek et al.
2001), but previous maps of SSC6 lacked the resolution
required for further fine mapping of any QTL regions to
enable final identification of the underlying genes. Exploi-
tation of data from the much more detailed human and
murine maps is essential in accelerating the improvement
of livestock maps. Accordingly, numerous genes have
been mapped on the porcine genome to provide a
more detailed correspondence with the human genome
(http://www.toulouse.inra.fr/lgc/pig/compare/HSA.htm).
As a result, a number of breaks in conservation of synteny
between HSA chromosomes and SSC6 have been demon-
strated (Pinton et al. 2000; Karnuah et al. 2001; Sun et al.
2001; Jiang et al. 2002; Kiuchi et al. 2002; Rink et al.
2002). Nevertheless, to facilitate the identification of can-
didate genes within small chromosome segments, the
homologous regions need to be defined more accurately.
Radiation hybrid mapping was utilized in this study to
link both microsatellite and gene-based markers into a
comprehensive high-resolution map with as few break
points as possible. The current draft sequence of the human
genome (http://www.ncbi.nlm.nih.gov/genome/guide/
human/) and the previous comparative mapping between
the human and pig genomes performed by our group (Jiang
Address for correspondence
J. A. B. Robinson, Department of Animal and Poultry Science,
University of Guelph, Guelph, Ont., Canada N1G 2W1.
E-mail: [email protected]
Accepted for publication 26 May 2004
� 2004 International Society for Animal Genetics, Animal Genetics, 35, 367–378 367

et al. 2002) provide a basis for the selection of new markers
from HSA1, 16, 18 and 19 for mapping on SSC6.
Materials and methods
Marker selection and PCR primer design
Orthologous genes were selected at approximately 3 Mb
intervals based on homologous regions of HSA1, 16, 18
and 19 as previously defined using a comparative mapping
approach (Jiang et al. 2002). The selected genes span a
region of approximately 0–93 Mb on HSA1, 39–81 Mb on
HSA16, 0–34 Mb on HSA18 and 34–60 Mb on HSA19.
Porcine gene or EST sequences were selected based on
their potential orthology with genes on HSA1, 16, 18 and
19, as predicted by BLASTN. Polymerase chain reaction
(PCR) primer pairs were designed to amplify regions with
least similarity to the murine orthologous gene sequences,
using the online oligonucleotide design tool Primer 3
(http://www.genome.wi.mit.edu/cgi-bin/primer/primer3_www.
cgi) and were purchased from Laboratory Services Division,
University of Guelph (Guelph, Ont., Canada). A common
tail, 5¢-CCTCGACTGCGTACCAATTCC-3¢, was added to one
of the primers in each pair to simplify direct sequencing of
the PCR products. The primer sequences and their related
information are shown in Table 1. The PCR amplification
products were sequenced and compared with original
sequences in GenBank to confirm orthologous status. These
primer pairs were also tested using hamster DNA, to avoid
using primers that would co-amplify host homologous genes
in the pig/hamster RH panel. Primers for microsatellite
markers previously assigned to SSC6 were designed based
on information from http://www.genome.iastate.edu/maps/
marcmap.html.
Radiation hybrid panel analysis
A whole genome pig/hamster RH 3000 panel that con-
tained 94 clones was purchased from Research Genetics
(Huntsville, AL, USA).
Amplification of the RH clones was carried out in a
10 ll reaction containing 50 ng RH DNA, 7.5 ng of each
primer, 150 nmol of dNTPs, 1.5–2.5 mM MgCl2 and
0.75 units of Taq Gold polymerase (Applied Biosystems,
Foster City, CA, USA). For multiplex PCR reactions, up to
three primer pairs were added to the same reaction
mixture. The PCR conditions were as follows: 95 �C for
10 min, and 32 cycles of 94 �C for 30 s, annealing
temperature (specific for each primer pair; Table 1) for
30 s, and 72 �C for 30 s, followed by a 5-min extension
at 72 �C. PCR products were analysed by electrophoresis
through 2% agarose gels containing ethidium bromide
(0.4 lg/ml) in 1x TBE buffer. Each locus was scored as
either present (1) or absent (0) for each hybrid line. Each
ambiguous locus was re-analysed.
The RHMAP software package (version 3.0, Boehnke
et al. 1991) was used to analyse the data. Markers were
assigned to linkage groups on achieving a minimum two
point LOD score of 4.0, using the RH2PT routine of RHMAP
3.0. A LOD score of 4.0 was used to be consistent with the
objective of minimizing artificial breakpoints. Multipoint
analysis and ordering of markers was performed with the
RHMAXLIK routine (Lunetta et al. 1996). The marker order
with the greatest likelihood was accepted. The RH map
was constructed using Genetic Map Creator (http://www.
wesbarris.com/mapcreator/).
Distances (D), in Rays, were calculated as D ¼ )ln(1 ) h), where h is the breakage frequency.
Polymorphism discovery
Primers designed for gene-based markers were used to
amplify two pools of genomic DNA. The pools were created
by combining equal amounts of DNA from13 Large White
pigs or 16 Meishan pigs, including animals from three
generations (F0, F1 and F2) in each case. Direct sequencing
of these PCR products was performed using an ABI 377
automatic sequencer (Applied Biosystems, Foster City, CA,
USA) by Laboratory Service Division (University of Guelph).
Results and discussion
Radiation hybrid map construction
Initially, 181 orthologous genes were selected from HSA1,
16, 18 and 19 based on available information for pig gene
or EST sequences in GenBank. Primers were designed to
amplify unique sequences of these genes and were tested
using pig and hamster DNA as templates; 17% of the primer
pairs amplified both pig and hamster genomic DNA, and
15% of the primer pairs failed to provide specific PCR
products. Thus, 68% of the primer pairs were used for RH
panel typing with the result that 105 gene-based markers
and 15 microsatellite markers were assigned to the SSC6
RH map. The marker symbols, names, corresponding loca-
tions, and start and stop distances on HSA1, 16, 18 or 19
are shown in Table 1.
The complete data set was analysed with the RHMAP
software package version 3.0. The mean retention fre-
quency for gene-based markers was 23.7% (range 5.3–
54.3%), and for microsatellite markers was 57% (range
11.7–68.1%). Using a two-point LOD score of 4.0, two-point
analyses of the 3000-rad panel revealed six linkage groups,
with 23, 7, 20, 14, 33 and 23 loci, respectively (Fig. 1).
These data represent the largest single assignment of
gene-based markers to SSC6. As well as the six linkage
groups shown in Fig. 1, the two-point analysis also revealed
two small linkage groups with only two loci each, leaving
just eight unlinked markers (not shown in Fig. 1 or
Table 1). Nine of the 15 microsatellites examined had been
� 2004 International Society for Animal Genetics, Animal Genetics, 35, 367–378
Cao et al.368

Tab
le1
SSC6RH
map
markers,withdatab
aseinform
ation,regional
localizationofHSA
homologousloci,primer
inform
ation,product
size
andmap
location.
Locus
symbol
Gen
enam
eHSA
location
HSA
distance
(start/stop;Mb)
Accession
no.
Primer
sequen
ces
Tm
(�C)
Product
size
(bp)
LG(AD)
CDH1
Cadherin1,type1,
E-cadherin(epithelial)
16q22.1
59.46/59.55
AW431113,BE2
35473,
BE0
13620an
dBM659150
F:ACCCCACTGCCCCACCTTAT
R:TCCTCACCGCCTCCGTACAT
61
181
1(0.000)
NFAT5
Nuclearfactorof
activatedTcells5,
tonicity-responsive
16q22.1
60.28/60.42
AJ005160
F:CCTCGACTGCGTACCAATTCCGAGCTATGGAACAAGGATG
R:AGCAACCAGAATACCATGGCTACAA
61
254
1(0.589)
ATBF1
AT-binding
transcriptionfactor1
16q22.3-q23.1
63.71/63.76
BI337553,AW785662
andBE2
35417
F:CCTCGACTGCGTACCAATTCCCCCTTCAAGAGTCAGCAAC
R:GATGCAATCATATCTGCCAGCTTCT
61
436
1(1.089)
HPR
Haptoglobin-related
protein
16q22.1
62.99/63.00
AF4
92467
F:CCTCGACTGCGTACCAATTCCGGAGATTGAGAAGGTGATT
R:CAGATGCTCCGTAAAGTTGAGGTTG
61
196
1(1.437)
DHODH
Dihydroorotate
dehydrogenase
16q22
62.94/62.95
BI343408an
dAW
486848
F:CCTCGACTGCGTACCAATTCCGGAGAGGGATGCCTTGAA
R:ACTGTGGTGTTCGTGACAATCAGTC
61
1264
1(1.867)
VPS4A
Vacuolarprotein
sorting4A
(yeast)
16q23.1
60.03/60.04
AW785058,BM484481
andBI346401
F:CCTCGACTGCGTACCAATTCCGCGAATCTATATCCCGTTG
R:GTGAGTTGCTGACTGCACTTTCCTC
61
202
1(2.566)
KATNB1
Katanin
p80
(WD40-containing)
subunitB1
16q13
48.17/48.19
BF1
89878,AW435616,
BF0
78237,BE0
13827
andBF7
10054
F:CCTCGACTGCGTACCAATTCCCCAAGGAGGCAGCAAAG
R:GAGGCCTTCAGCCCGATG
61
309
1(3.009)
GOT2
Glutamic-oxaloacetic
transaminase
2,
mitochondrial(aspartate
aminotransferase
2)
16q21
49.14/49.16
M11732
F:CCTCGACTGCGTACCAATTCCGTCTCTGCCAGTCCTATGC
R:CGTTGACAGGAGGGTTGGAATACAT
61
�1086
1(3.471)
MT3
Metallothionein
3
[growth
inhibitory
Pfactor(neurotrophic)]
16q13
47.01/47.01
BF1
91454
F:CCTCGACTGCGTACCAATTCCTGCGAGGGATGCAAATG
R:AGCTGCACTTCTCTTCCTCAGCTTC
61
788
1(4.002)
KIAA0872
KIAA0872protein
16q22.1
68.15/68.16
BF1
98919
andAW785756
F:CCTCGACTGCGTACCAATTCCTTTCTTCTATGCCTACGTG
R:GATGTCCAGCGGCTTATAGAGGAAG
61
259
1(4.862)
CASPR4
Cellrecognition
protein
CASPR4
16q23.1
67.27/67.52
BF1
90030
andAW415146
F:CCTCGACTGCGTACCAATTCCGTGCATCCATCATTTACAA
R:AAAGGAGCTCACGTAGAGCAGCAAG
61
164
1(5.345)
WW
OX
WW
domain
containing
oxidoreductase
16q23.3-q24.1
69.06/70.18
BC014716
F:GTCCAAGCTCTGCAACATCCTCTT
R:CCTCGACTGCGTACCAATTCCACTTGGTGAAGGGTCTGG
61
167
1(5.551)
DC13
DC13protein
16q23.3
71.94/71.97
BE0
14487
R:CCTCGACTGCGTACCAATTCCAGAAGAGGAACAAGAGCAG
F:ACTCCAAGGATTCTGGAGACCAAGT
61
180
1(6.077)
LOC124359
Sim
ilarto
RIKEN
cDNA
4930453I21
16q23.3
71.57/71.68
AW
360379
F:AGAAACTCTCGCACAGACCCTCAG
R:CCTCGACTGCGTACCAATTCCCCTGTAGGACACCGTCTTG
61
155
1(6.768)
HSD17B2
Hydroxysteroid
(17-beta)
dehydrogenase
2
16q24.1-q24.2
72.91/72.97
AW785511
andAW312396
F:CCTCGACTGCGTACCAATTCCATGCTTTGTCCAAGTATC
R:CATCTCTTATCTGCTGCGAGTTGGT
61
156
1(7.137)
MLYCD
Malonyl-CoA
decarboxylase
16q24
74.78/74.85
BF1
89220,AW415837
andBI343470
F:CCTCGACTGCGTACCAATTCCAAGTCGAAAACCAAGGAG
R:GGTGATGCCCTTGAGGCTGAC
61
309
1(7.655)
� 2004 International Society for Animal Genetics, Animal Genetics, 35, 367–378
High-resolution radiation hybrid map of porcine chromosome 6 369

Tab
le1
Continued
Locus
symbol
Gen
enam
e
HSA
location
HSA
distance
(start/stop;Mb)
Accession
no.
Primer
sequen
ces
Tm
(�C)
Product
size
(bp)
LG(AD)
S0035
DNA
segment,
microsatellite
F:GGCCGTCTTATACTCTCAGCATA
R:CCAAATAAACAGCAGGCAGCCT
57
186
1(8.277)
MC1R
Melanocortin
1receptor
(alphamelanocyte
stim
ulatinghorm
one
receptor)
16q24.3
81.30/81.31
AF3
26520
F:CCTCGACTGCGTACCAATTCCCCATGTACTACTTCGTCTG
R:GTGTGGTGGTAGTAGGCGATGAAGA
61
343
1(8.767)
CYBA
Cytochromeb-245,
alphapolypeptide
16q24
80.15/80.15
U02477
F:GTGTTGGTCTGCCTGCTGGAGTA
R:CCTCGACTGCGTACCAATTCCCCCGGATGTAGTAGTTCCT
61
255
1(9.242)
APRT
Adenine
phosphoribosyltransferase
16q24
80.44/80.45
BI342595an
dBI343520
F:CCTCGACTGCGTACCAATTCCCTGGAGATCCAGAGAGA
R:ACTCGAACTGCAGAAGGGAGAAGAA
61
358
1(9.559)
DPEP1
Dipeptidase
1(renal)
16q24.3
81.01/81.03
X53730
F:CCTCGACTGCGTACCAATTCCCACTCCATCGACAGCAG
R:CGTGTCCACCAGCCAGTTGT
61
213
1(9.910)
ZDHHC7
Zincfinger,DHHC
domain
containing7
16q23.3
75.55/75.59
BI185968an
dBI184335
F:AGTGGACTGAGTGCAGTGGTTTCTC
R:CCTCGACTGCGTACCAATTCCCACTCTTGAGCCTCTCGAT
61
653
1(10.490)
NOC4
NeighbourofCOX4
16q24
77.03/77.06
AW
312595
F:CCTCGACTGCGTACCAATTCCCGAACACGATGAGAACAA
R:GAATGTCATCCAGGTGGTTATCAAA
61
662
1(10.683)
TNRC9
Trinucleotiderepeat
containing9
16q12.2
42.90/43.00
F15073
F:CTCGGGGATGAGGAGTTTGAAATAC
R:CCTCGACTGCGTACCAATTCCAGATTCCTTGAGATGGTAA
61
191
2(0.000)
RBL2
retinoblastoma-like2
(p130)
16q12.2
43.88/43.94
BI304691,BI132957
andBI404578
F:TGCTATAGGACTCAGCCACAAGC
R:CCTCGACTGCGTACCAATTCCACTATCACTGCTGCCAGAAT
61
492
2(0.188)
LYPLA
3Lysophospholipase
3
(lysosomal
phospholipase
A2)
16q22.2
58.97/58.99
BF4
42148
F:CCTCGACTGCGTACCAATTCCCAACCGTATCCCGGTCAT
R:ATGAGCCAGCCATCTTTGAAGC
61
195
2(1.032)
LCAT
Lecithin-cholesterol
acyltransferase
16q22.1
58.66/58.67
BG384679,AW
346632
andAW
786443
F:CCTCGACTGCGTACCAATTCCGGCTGGTACATGTGGCTA
R:AAAACCATATTGAGGTGCTGTGTCC
61
278
2(1.336)
CDH5
Cadherin5,type2,
VE-cadherin
(vascularepithelium)
16q22.1
56.98/57.02
Y13919
F:CCTCGACTGCGTACCAATTCCTCTCCCTCCTCGTGTACCT
R:CAAAGATATGCAGCGTGTCGTAGG
61
351
2(1.794)
CDH11
Cadherin11,type2,
OB-cadherin(osteoblast)
16q22.1
55.36/55.47
BF0
78663
F:CGAGGAAGAAGATGTCCGTGAGA
R:CCTCGACTGCGTACCAATTCCTGATGAAATCATCCACATC
61
212
2(2.269)
SW
R2149
DNA
segment,
microsatellite
AF2
53918
F:TAACCTGCTGAGCCACAATG
R:GGGGGTTAAAAGTCCCTATTC
56
171
2(2.670)
U2AF65
U2sm
allnuclear
ribonucleoprotein
auxiliary
factor(65kD)
19q13.43
57.04/57.07
BI182545
F:CCTCGACTGCGTACCAATTCCAAGAAGAAGGTCCGCAAGT
R:AGTGATGCCAAAGGGGATGTTG
61
429
3(0.000)
S0300
DNA
segment,
microsatellite
U11544
F:TCCTCGGAAAGATGTAACTGC
R:CTGTCACCAACAGGGCACTA
56
160
3(0.737)
� 2004 International Society for Animal Genetics, Animal Genetics, 35, 367–378
Cao et al.370
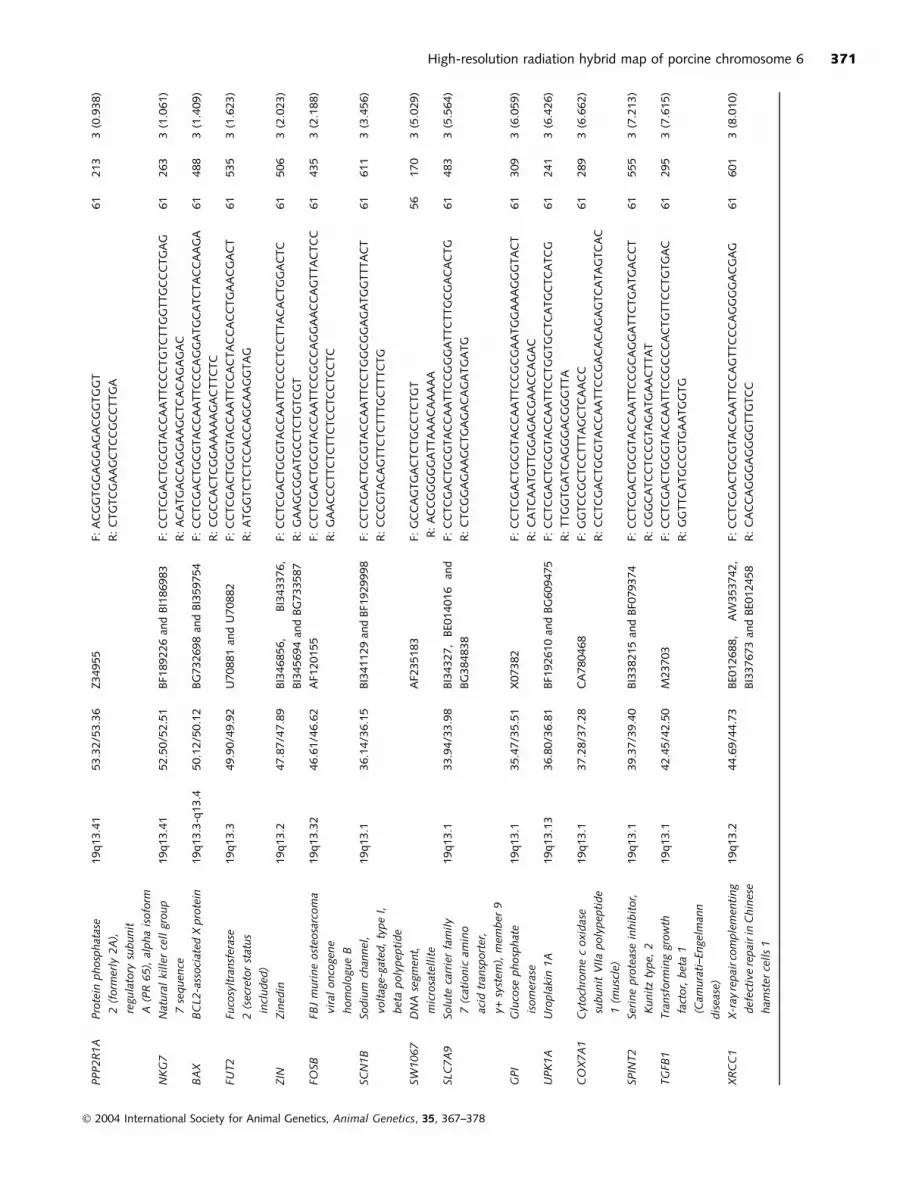
PPP2R1A
Protein
phosphatase
2(form
erly2A),
regulatory
subunit
A(PR65),alphaisoform
19q13.41
53.32/53.36
Z34955
F:ACGGTGGAGGAGACGGTGGT
R:CTGTCGAAGCTCCGCCTTGA
61
213
3(0.938)
NKG7
Naturalkillercellgroup
7sequence
19q13.41
52.50/52.51
BF1
89226an
dBI186983
F:CCTCGACTGCGTACCAATTCCCTGTCTTGGTTGCCCTGAG
R:ACATGACCAGGAAGCTCACAGAGAC
61
263
3(1.061)
BAX
BCL2-associatedXprotein
19q13.3-q13.4
50.12/50.12
BG732698an
dBI359754
F:CCTCGACTGCGTACCAATTCCCAGGATGCATCTACCAAGA
R:CGCCACTCGGAAAAAGACTTCTC
61
488
3(1.409)
FUT2
Fucosyltransferase
2(secretorstatus
included)
19q13.3
49.90/49.92
U70881an
dU70882
F:CCTCGACTGCGTACCAATTCCACTACCACCTGAACGACT
R:ATGGTCTCTCCACCAGCAAGGTAG
61
535
3(1.623)
ZIN
Zinedin
19q13.2
47.87/47.89
BI346856,
BI343376,
BI345694an
dBG733587
F:CCTCGACTGCGTACCAATTCCCCTCCTTACACTGGACTC
R:GAAGCGGATGCCTCTGTCGT
61
506
3(2.023)
FOSB
FBJmurineosteosarcoma
viraloncogene
homologueB
19q13.32
46.61/46.62
AF1
20155
F:CCTCGACTGCGTACCAATTCCGCCAGGAACCAGTTACTCC
R:GAACCCTTCTCTTCTCCTCCTCCTC
61
435
3(2.188)
SCN1B
Sodium
channel,
voltage-gated,typeI,
beta
polypeptide
19q13.1
36.14/36.15
BI341129an
dBF1
929998
F:CCTCGACTGCGTACCAATTCCTGGCGGAGATGGTTTACT
R:CCCGTACAGTTCTCTTTGCTTTCTG
61
611
3(3.456)
SW
1067
DNA
segment,
microsatellite
AF2
35183
F:GCCAGTGACTCTGCCTCTGT
R:ACCGGGGGATTAAACAAAAA
56
170
3(5.029)
SLC7A9
Solute
carrierfamily
7(cationic
amino
acidtransporter,
y+system),member9
19q13.1
33.94/33.98
BI34327,BE0
14016
and
BG384838
F:CCTCGACTGCGTACCAATTCCGGGATTCTTGCGACACTG
R:CTCGGAGAAGCTGAGACAGATGATG
61
483
3(5.564)
GPI
Glucose
phosphate
isomerase
19q13.1
35.47/35.51
X07382
F:CCTCGACTGCGTACCAATTCCGCGAATGGAAAGGGTACT
R:CATCAATGTTGGAGACGAACCAGAC
61
309
3(6.059)
UPK1A
Uroplakin
1A
19q13.13
36.80/36.81
BF1
92610an
dBG609475
F:CCTCGACTGCGTACCAATTCCTGGTGCTCATGCTCATCG
R:TTGGTGATCAGGGACGGGTTA
61
241
3(6.426)
COX7A1
Cytochromecoxidase
subunitVIIapolypeptide
1(m
uscle)
19q13.1
37.28/37.28
CA780468
F:GGTCCGCTCCTTTAGCTCAACC
R:CCTCGACTGCGTACCAATTCCGACACAGAGTCATAGTCAC
61
289
3(6.662)
SPINT2
Serineprotease
inhibitor,
Kunitztype,2
19q13.1
39.37/39.40
BI338215an
dBF0
79374
F:CCTCGACTGCGTACCAATTCCGCAGGATTCTGATGACCT
R:CGGCATCCTCCGTAGATGAACTTAT
61
555
3(7.213)
TGFB1
Transform
inggrowth
factor,beta
1
(Camurati–Engelm
ann
disease)
19q13.1
42.45/42.50
M23703
F:CCTCGACTGCGTACCAATTCCGCCCACTGTTCCTGTGAC
R:GGTTCATGCCGTGAATGGTG
61
295
3(7.615)
XRCC1
X-rayrepaircomplementing
defectiverepairin
Chinese
hamstercells1
19q13.2
44.69/44.73
BE0
12688,
AW
353742,
BI337673an
dBE0
12458
F:CCTCGACTGCGTACCAATTCCAGTTCCCAGGGGACGAG
R:CACCAGGGAGGGGTTGTCC
61
601
3(8.010)
� 2004 International Society for Animal Genetics, Animal Genetics, 35, 367–378
High-resolution radiation hybrid map of porcine chromosome 6 371

Tab
le1
Continued
Locus
symbol
Gen
enam
e
HSA
location
HSA
distance
(start/stop;Mb)
Accession
no.
Primer
sequen
ces
Tm
(�C)
Product
size
(bp)
LG(AD)
SIRT2
Sirtuin
(silentmatingtype
inform
ationregulation
2homologue)2
(S.cerevisiae)
19q13
39.98/40.00
AW
657450,AW
346910,
BE2
34094an
dBI184249
F:CCTCGACTGCGTACCAATTCCAAGAAGGAGCTGGAGGAC
R:GAGCTCCCCAGAAAAGAGCTGTAAG
61
255
3(8.791)
SW
193
DNA
segment,
microsatellite
AF2
35233
F:GCAATTTGCCTGATGCTCTT
R:TTCACTCTGAGGGGTCCTGA
57
184
3(9.327)
SUPT5H
SuppressorofTy5
homologue(S.cerevisiae)
19q13
40.58/40.61
AW
619853,AW
785911,
BI343372an
dBI400129
F:CCTCGACTGCGTACCAATTCCCAGGTCAACCCACAGTA
R:GCGAGGGTGTGGGGTGATAG
61
316
3(9.555)
STK13
Serine/threoninekinase
13(aurora/IPL1-like)
19q13.43
58.64/58.65
AW477959an
d
CA778485
F:CCTCGACTGCGTACCAATTCCATAGAGAAGGAAGGACTGG
R:GCATTTCTGCAGTTCCTTGTAGAGC
61
814
4(0.000)
ISG15
Interferon-stimulated
protein,15kDa
1p36.33
0.48/0.48
BI183574
F:CCTCGACTGCGTACCAATTCCGGTGCAAAGCTTCAGAGAC
R:GGTGCACATAGGCTTGAGGTCATAC
61
220
4(0.620)
BC-2
Putativebreast
adenocarcinoma
marker(32kD)
19q
59.96/59.96
BI338403
F:CCTCGACTGCGTACCAATTCCGCTCAAGTCTAACAACTCC
R:CATCGTTCATCATCTCCTCCTTCAT
61
235
4(0.956)
ARPM2
Actin-relatedprotein
M2
1p36.3
2.38/2.38
BF1
94602an
d
AW
430875
F:CCTCGACTGCGTACCAATTCCAGGTCTTGCACCTGCACT
R:AGGGAGTAGCCCTCGAAGATGG
61
325
4(1.388)
ICMT
Isoprenylcysteine
carboxylmethyltransferase
1p36.21
5.86/5.87
BE0
32550
F:GCAGATCACCTGGCTCAGTGC
R:CCGGAACCAGGCGTAGACTC
61
166
4(2.089)
ENO1
Enolase
1(alpha)
1p36.3-p36.2
8.54/8.56
AW
619583
F:AAGGCTGGCGCTGTTGAGAAG
R:CCTCGACTGCGTACCAATTCCCCGTATTTCTCCTTGATG
61
593
4(2.858)
RERE
Arginine-glutamic
acid
dipeptide(RE)repeats
1p36.1-p36.2
7.99/8.14
Z84142
F:CCAACCCCATGGAGCATTTC
R:CCTCGACTGCGTACCAATTCCTCCGGGTAGCTCATTTCT
61
159
4(3.008)
SW
4DNA
segment,
microsatellite
AF2
53862
F:CCTGGGGCACACTCACACCT
R:CCGTCGTGTGTGCCAAAGAT
61
144
4(3.357)
NPPA
Natriureticpeptide
precursorA
1p36.21
11.75/1175
X54669
F:AGGCTATGCCCCCACAAGTACTAAG
R:CCTCGACTGCGTACCAATTCCCAGCCCAGTCCACTCTGTG
61
271
4(3.836)
NPPB
Natriureticpeptide
precursorB
1p36.2
11.98/11.98
M23596
F:GAGCGGACGGACCTGGAG
R:CCAAAGCAGCCAGAGTCACG
61
164
4(3.899)
MTHFR
5,10-m
ethylenetetra
hydrofolate
reductase
1p36.3
11.69/11.71
BG835531,BG384323
andAW436969
F:CCTCGACTGCGTACCAATTCCATCATCACCCAGCTTTTCT
R:CTCGATCACGTCCTTGATCTGCT
61
1035
4(4.037)
SW
122
DNA
segment,
microsatellite
AF2
35206
F:TTAGAAGACAAATTTTAGAAGCTCTTT
R:AAAAAGGCAAAAGATTGACACA
56
201
4(4.408)
SDR1
Short-chain
dehydrogenase/reductase
1
1p36.1
12.66/12.71
BG834113
F:AATGGCTGGGAGCGTTGGTA
R:CGCGTTCTGCGAACTCCCTA
61
174
4(4.674)
PGD
Phosphogluconate
dehydrogenase
1p36.3-p36.13
10.42/10.44
X16638
F:CCTCGACTGCGTACCAATTCCAGTCTTTGCTCGATGCTTA
R:CATGAAGCCCTGAGTGTAGGAGATG
61
348
4(5.045)
� 2004 International Society for Animal Genetics, Animal Genetics, 35, 367–378
Cao et al.372

CAC-1
CAC-1
1p35-p34
33.18/33.19
BF1
89480
F:CCTCGACTGCGTACCAATTCCGTGTTGTCACGCTACCT
R:ACTCTCTGCCACGCAGAACAGTAA
61
557
5(0.000)
GJB5
Gapjunctionprotein,beta
5
(connexin
31.1)
1p35.1
35.04/35.05
AW786361
andBE2
35692
F:CCTCGACTGCGTACCAATTCCCTGCTACGACGAGTTCTT
R:GAGGGTGTACTTGGGGTAGAACAGG
61
286
5(0.507)
MTF1
Metal-regulatory
transcrip-
tionfactor1
1p33
38.11/38.16
Z84162,BE2
35677,
AW312370an
dZ84098
F:CCTCGACTGCGTACCAATTCCACAGTCCAGACAACAGCAT
R:AGTGAGGGTTGCACCTTCGATATTT
61
362
5(1.014)
PABPC4
Poly(A)bindingprotein,
cytoplasm
ic4(inducible
form
)
1p32-p36
39.94/39.96
AW436678an
d
BI185581
F:ACTCATCCAGACCATGCACTCAAAC
R:CCTCGACTGCGTACCAATTCCTGGCATGATGAGCCTGTAG
61
388
5(1.554)
MC5R
Melanocortin
5receptor
18p11.2
13.92/13.92
AF1
33793
F:CCTCGACTGCGTACCAATTCCGAGAACATCTTGGTCATTG
R:AGTGCGTAGAAGATGGTCACGTAGC
61
287
5(1.843)
MC2R
Melanocortin
2receptor
(adrenocorticotropic
hor-
mone)
18p11.2
13.96/13.96
AF0
64077
F:CCTCGACTGCGTACCAATTCCGCCTTCACAGCACTGTTC
R:AAGGGGTCGATGATAGCATTACACA
61
290
5(2.054)
PPT1
Palm
itoyl-protein
thioesterase
1
(ceroid-lipofuscinosis,
neuronal1,infantile)
1p32
40.57/40.59
BE2
34609an
dBI340400
F:CCTCGACTGCGTACCAATTCCTGTAATCCCTTAAGCATGG
R:TGGGAGTTGACATTCAAAAAGAAGC
61
317
5(2.538)
GALNAC4ST-2
GalNAc-4-sulphotransf-
erase
2
18q11.2
24.43/24.70
BI337921
andAW
670338
F:CCTCGACTGCGTACCAATTCCCTACATTAGCTACAACTGC
R:ATAAAAAGCCATTGCCTCATCTGGT
61
290
5(3.936)
DSG2
Desm
oglein
218q12.1
29.10/29.15
BE2
32087,BG382012
andBG733433
F:CCTCGACTGCGTACCAATTCCCAAACCTTTGCCTGAAGTA
R:AAGGAGGAAGCTGGGGCATAGAC
61
249
5(5.290)
TTR
Transthyretin(pre-albumin,
amyloidosistypeI)
18q12.1
29.19/29.20
X82258
F:CCTCGACTGCGTACCAATTCCGCTTAGCCCCTACTCTTAC
R:TGTTTTATTGTCTCTGCCCGAGTTT
61
168
5(5.751)
SS18
Synovialsarcomatransloca-
tion,chromosome18
18q11.2
23.57/23.64
BU946845,BU946839
andBF7
02330
F:CCTCGACTGCGTACCAATTCCTCTGCCTGATTGTTTTAGC
R:CAAATTTATGGCACCCCTTCTGTTT
61
232
5(6.366)
OAZ
OLF-1/EBF-associatedzinc
fingergene
16q12
39.94/40.18
BI184377an
d
AU059448
F:CCTCGACTGCGTACCAATTCCGCACACCTTTGTCTTCTTT
R:GAAGGCTTTGCTACAGAACTTGCAG
61
307
5(6.669)
LAMA3
Laminin,alpha3[nicein
(150kD),kalinin
(165kD),
BM600(150kD),epilegrin]
18q11.2
21.34/21.43
AW
787099,BG385235
andBE2
32484
F:CCTCGACTGCGTACCAATTCCGGAACTTTCAGTTGGATCT
R:CTTCCAATGTGAATGAGGATTCCAG
61
340
5(7.228)
GATA6
GATA
bindingprotein
618q11.1-q11.2
19.63/19.66
AF2
95687an
dBI181578
F:CCTCGACTGCGTACCAATTCCGAGGGAATTCAGACCAGGA
R:GCTGCAATCGTCTGAGTTAGAAGAGG
61
221
5(8.054)
CETN1
Centrin,EF-handprotein,1
18p11.32
0.86/0.87
BF1
91379
F:CCTCGACTGCGTACCAATTCCGGAGGAGATGAAGAGGAT
R:CCACGCGTTTTAGATTTTTGAATGA
61
191
5(9.232)
ROCK1
Rho-associated,coiled-coil
containingprotein
kinase
1
18q11.1
18.39/18.57
D89493
F:ATTAAAGTCTCAAGGTGGCGATGGT
R:CCTCGACTGCGTACCAATTCCAAGAATCTGCATAAAATGG
61
428
5(9.948)
ADCYAP1
Adenylate
cyclase
activating
polypeptide1(pituitary)
18p11
0.89/0.90
AF0
47007
F:CCTCGACTGCGTACCAATTCCGAAACAAATGGCTGTTAAG
R:TTTGGATAGAACACATGAGCGATGA
61
195
5(10.253)
EMILIN-2
Extracellularglycoprotein
EMILIN-2
precursor
18p11.3
3.12/3.14
BI340897an
d
AW
479974
F:CCTCGACTGCGTACCAATTCCGCGCTACCTGATCACAG
R:CTTCAAGTGCACGACCAGGTG
61
194
5(10.727)
EPB41L3
Erythrocyte
membrane
protein
band4.1-like3
18p11.32
5.43/5.55
CA779909
F:CCTCGACTGCGTACCAATTCCCCGAATCAGCATCACTAAA
R:AATAAGCTACGAAAGGCATGGATTG
61
180
5(11.202)
High-resolution radiation hybrid map of porcine chromosome 6 373
� 2004 International Society for Animal Genetics, Animal Genetics, 35, 367–378

Tab
le1
Continued
Locus
symbol
Gen
enam
e
HSA
location
HSA
distance
(start/stop;Mb)
Accession
no.
Primer
sequen
ces
Tm
(�C)
Product
size
(bp)
LG(AD)
PTPRM
Protein
tyrosine
phosphatase,receptor
type,M
18p11.2
8.57/8.68
BI183364,
AW656688
andBE0
13858
F:CCTCGACTGCGTACCAATTCCAACTCCAGCCAGATTAAA
R:ATGAGAGCGGCGTTGATGTAGTT
61
847
5(11.571)
PPP4R1
Protein
phosphatase
4,
regulatory
subunit1
18p11.21
9.65/9.72
BF1
89573an
d
AW
436288
F:CCTCGACTGCGTACCAATTCCCCTGCTGGATCAGTATC
R:CGTAGGTCTCCCTGAGGCAGTG
61
146
5(11.909)
HMCS
Molybdenum
cofactor
sulphurase
18q12
33.80/33.88
BI403162,BI403428an
d
BG384067
F:CCTCGACTGCGTACCAATTCCTGGGAGTCGGTTCTGTTA
R:GGAGCAGTACGATTATTCACCAGCA
61
428
5(12.837)
MAPRE2
Microtubule-associated
protein,RP/EBfamily,
member2
18q12.1
32.59/32.76
BE0
13298an
d
AW
657451
F:CCTCGACTGCGTACCAATTCCATAGGAGCCTAAGCCCAAG
R:GGCGAAGATTTGGAAAGATACTAAGA
57
258
5(13.125)
LAPTM5
Lysosomal-associated
multispanningmembrane
protein-5
1p34
31.00/31.02
BI304807an
dBE2
36206
F:CCTCGACTGCGTACCAATTCCTGAGCACTCAGTGGAAGTG
R:TGATGACAAAGAGCATGGTGATGAG
61
1010
5(13.778)
SFRS4
Splicingfactor,arginine/
serine-rich4
1p35.2
29.48/29.51
BF7
10794an
dBI400329
F:CCTCGACTGCGTACCAATTCCAGCCGTAGCACAAGTGATT
R:CCATCCAGGTACTGAGCTCCTTGTA
61
252
5(14.073)
SECP43
tRNA
selenocysteine-
associatedprotein
1p35.2
28.89/28.91
BE0
31048an
d
BG609066
F:CCTCGACTGCGTACCAATTCCTAAAACCGGTGGAATATAG
R:GCATGGTGCTCTGAGTATAGCCGTA
61
153
5(14.347)
FGR
Gardner-Rasheedfeline
sarcomaviral(v-fgr)
oncogenehomologue
1p36.2-p36.1
27.51/27.53
AW670343an
d
BI335994
F:CCTCGACTGCGTACCAATTCCACTGAACTCATCAACAAGG
R:CCAGAAAGGACTGCAGATACTCGAA
61
303
5(14.553)
SW2505
DNA
segment,
microsatellite
AF2
25168
F:CTGCAAGGAGGCAGGGGATT
R:CCTGCAGAGGCACAGTTTGG
61
207
5(14.936)
DJ465N24.2.1
Hypotheticalprotein
dJ465N24.2.1
1p36.13-p35.1
25.09/25.10
BI399694,BF7
04492an
d
BE0
30569
F:CCTCGACTGCGTACCAATTCCTACGTGAACGATCTGTGG
R:GCGCGAGTAGGACCTCGAGTAG
61
256
5(15.092)
HTR1D
5-hydroxytryptamine
(serotonin)receptor1
1p36.3-p34.3
23.09/23.09
AF1
17655
F:CCTCGACTGCGTACCAATTCCCCACAGAAACTCCAAAAG
R:GCGTTTACTATACTCCAGGGCATCC
61
383
5(15.692)
WNT4
Wingless-typeMMTV
integrationsite
family,
member4
1p36.23-p35.1
22.02/22.04
AW
415026an
d
CA997682
F:CCTCGACTGCGTACCAATTCCACTCAAGGAGAAGTTTGAC
R:CAATGGCCTTGGACGTCTTGTT
61
212
5(15.820)
S0444
F:GACAGTGACTTGCAGGGTGA
R:GCCCACAATCTTGAGCCTAT
56
219
5(16.330)
PLA2G2A
Phospholipase
A2,groupIIA
(platelets,synovialfluid)
1p35
19.87/19.88
BI359586
F:CCTCGACTGCGTACCAATTCCTGGCAGTGATCATGGCATT
R:TAGCAACCATAGAAGGCGTAATTGG
61
324
5(17.043)
PRKACB
Protein
kinase,
cAMP-dependent,
catalytic,
beta
1p36.1
84.94/85.10
F:CCTCGACTGCGTACCAATTCCAAAAGAGTTTTGCGAATTT
R:CCTTCCAATGTCACAGAGGCATAAC
61
392
6(0.000)
CTBS
Chitobiase,di-N-acetyl-
1p22
85.52/85.54
AW
619352an
d
BE2
32595
F:CCTCGACTGCGTACCAATTCCGTTGGCCATAAAACTTGGA
R:GGCCTGCTTAATTTGTTGAGCTATC
61
831
6(0.199)
Cao et al.374
� 2004 International Society for Animal Genetics, Animal Genetics, 35, 367–378
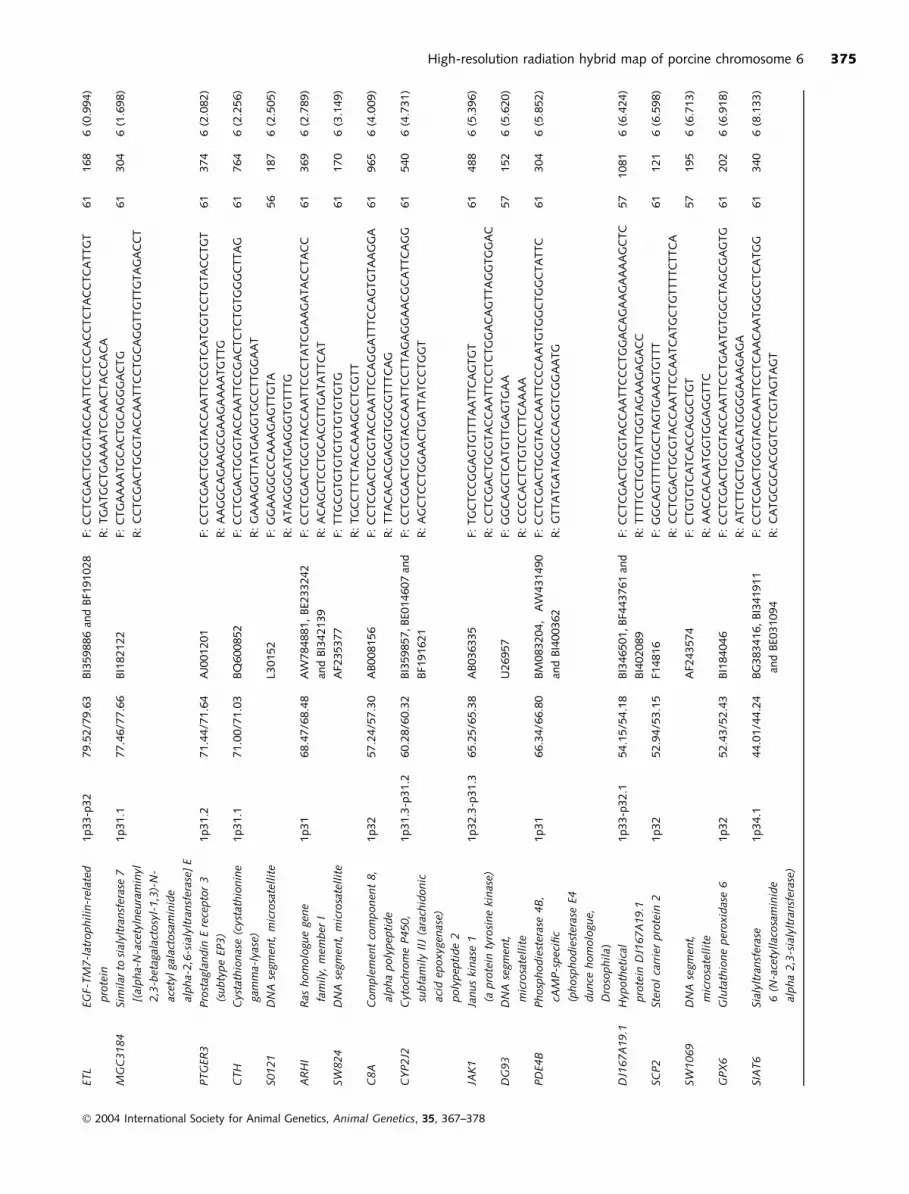
ETL
EGF-TM7-latrophilin-related
protein
1p33-p32
79.52/79.63
BI359886an
dBF1
91028
F:CCTCGACTGCGTACCAATTCCTCCACCTCTACCTCATTGT
R:TGATGCTGAAAATCCAACTACCACA
61
168
6(0.994)
MGC3184
Sim
ilarto
sialyltransferase
7
[(alpha-N
-acetylneuraminyl
2,3-betagalactosyl-1,3)-N-
acetylgalactosaminide
alpha-2,6-sialyltransferase]E
1p31.1
77.46/77.66
BI182122
F:CTGAAAATGCACTGCAGGGACTG
R:CCTCGACTGCGTACCAATTCCTGCAGGTTGTTGTAGACCT
61
304
6(1.698)
PTGER3
Prostaglandin
Ereceptor3
(subtypeEP3)
1p31.2
71.44/71.64
AJ001201
F:CCTCGACTGCGTACCAATTCCGTCATCGTCCTGTACCTGT
R:AAGGCAGAAGCGAAGAAAATGTTG
61
374
6(2.082)
CTH
Cystathionase
(cystathionine
gamma-lyase)
1p31.1
71.00/71.03
BQ600852
F:CCTCGACTGCGTACCAATTCCGACTCTCTGTGGGCTTAG
R:GAAAGGTTATGAGGTGCCTTGGAAT
61
764
6(2.256)
S0121
DNA
segment,microsatellite
L30152
F:GGAAGGCCCAAAGAGTTGTA
R:ATAGGGCATGAGGGTGTTTG
56
187
6(2.505)
ARHI
Rashomologuegene
family,memberI
1p31
68.47/68.48
AW784881,BE2
33242
andBI342139
F:CCTCGACTGCGTACCAATTCCCTATCGAAGATACCTACC
R:ACAGCTCCTGCACGTTGATATTCAT
61
369
6(2.789)
SW824
DNA
segment,microsatellite
AF2
35377
F:TTGCGTGTGTGTGTGTGTGTG
R:TGCCTTCTACCAAAGCCTCGTT
61
170
6(3.149)
C8A
Complementcomponent8,
alphapolypeptide
1p32
57.24/57.30
AB008156
F:CCTCGACTGCGTACCAATTCCAGGATTTCCAGTGTAAGGA
R:TTACACACGAGGTGGCGTTTCAG
61
965
6(4.009)
CYP2J2
CytochromeP450,
subfamilyIIJ(arachidonic
acidepoxygenase)
polypeptide2
1p31.3-p31.2
60.28/60.32
BI359857,BE0
14607an
d
BF1
91621
F:CCTCGACTGCGTACCAATTCCTTAGAGGAACGCATTCAGG
R:AGCTCCTGGAACTGATTATCCTGGT
61
540
6(4.731)
JAK1
Januskinase
1
(aprotein
tyrosinekinase)
1p32.3-p31.3
65.25/65.38
AB036335
F:TGCTCCGGAGTGTTTAATTCAGTGT
R:CCTCGACTGCGTACCAATTCCTCTGGACAGTTAGGTGGAC
61
488
6(5.396)
DG93
DNA
segment,
microsatellite
U26957
F:GGCAGCTCATGTTGAGTGAA
R:CCCCACTCTGTCCTTCAAAA
57
152
6(5.620)
PDE4B
Phosphodiesterase
4B,
cAMP-specific
(phosphodiesterase
E4
dunce
homologue,
Drosophila)
1p31
66.34/66.80
BM083204,
AW431490
andBI400362
F:CCTCGACTGCGTACCAATTCCCAATGTGGCTGGCTATTC
R:GTTATGATAGGCCACGTCGGAATG
61
304
6(5.852)
DJ167A19.1
Hypothetical
protein
DJ167A19.1
1p33-p32.1
54.15/54.18
BI346501,BF4
43761an
d
BI402089
F:CCTCGACTGCGTACCAATTCCCTGGACAGAAGAAAAGCTC
R:TTTTCCTGGTATTGGTAGAAGAGACC
57
1081
6(6.424)
SCP2
Sterolcarrierprotein
21p32
52.94/53.15
F14816
F:GGCAGTTTTGGCTAGTGAAGTGTTT
R:CCTCGACTGCGTACCAATTCCAATCATGCTGTTTTCTTCA
61
121
6(6.598)
SW
1069
DNA
segment,
microsatellite
AF2
43574
F:CTGTGTCATCACCAGGCTGT
R:AACCACAATGGTGGAGGTTC
57
195
6(6.713)
GPX6
Glutathioneperoxidase
61p32
52.43/52.43
BI184046
F:CCTCGACTGCGTACCAATTCCTGAATGTGGCTAGCGAGTG
R:ATCTTGCTGAACATGGGGAAAGAGA
61
202
6(6.918)
SIAT6
Sialyltransferase
6(N
-acetyllacosaminide
alpha2,3-sialyltransferase)
1p34.1
44.01/44.24
BG383416,BI341911
andBE0
31094
F:CCTCGACTGCGTACCAATTCCTCAACAATGGCCTCATGG
R:CATGCGCACGGTCTCGTAGTAGT
61
340
6(8.133)
High-resolution radiation hybrid map of porcine chromosome 6 375
� 2004 International Society for Animal Genetics, Animal Genetics, 35, 367–378

mapped previously to SSC6 (INRA-UMN RH map; http://
www.toulouse.inra.fr/lgc/pig/RH/Menuchr.htm), but the
order of some of these loci differed slightly in our study. In
addition, in our results, linkage group III includes loci
SW193, S0300 and SW1067 as part of a single linkage
group, which were previously reported to map to two small
groups (INRA-UMN RH map).
Comparative map
Of the 105 gene-based markers, 40 spanned a region of
approximately 0–86 Mb on HSA1p36.3-p22, 29 spanned
40–81 Mb on HSA16q12-q24, 17 spanned 1–34 Mb on
HSA18p11.3-q12 and 19 spanned 34–60 Mb on
HSA19q13.1-q13.4. The previous mapping of 29 of the 120
markers to SSC6 by cytogenetic mapping (http://
www.toulouse.inra.fr/lgc/pig/compare/SSC.htm and Lah-
bib-Mansais et al. 2000) and of 16 of the 120 by linkage
mapping (http://www.toulouse.inra.fr/lgc/pig/RH/Menuchr.
htm) was used in determining linkage group order and
orientation (Fig. 1).
While large regions of conserved segments have been
reported between SSC6 and HSA1, 16, 18 and 19 (Goureau
et al. 1996), one potential inconsistency was found. The
human genome position for OAZ is reported to be on
HSA16q12 (accession no. NM_015069), therefore this gene
was expected to map to SSC6p in linkage group I or II. In
our RH map, OAZ is located in group V on SSC6q2.8–3.1.
Repeated analyses of this marker confirmed our results; no
other inconsistencies have emerged.
Sixty-four genes expressed in a28-day-old pig embryohave
been assigned to an RH map (Karnuah et al. 2001), nine of
the genes being placed on SSC6. Our results support the
homologies reported between SSC6 and HSA1, 16 or 18 for
six of these genes. Although RH mapping can provide higher
resolution, of the 62 genes or ESTs physicallymapped to SSC6
(http://www.toulouse.inra.fr/lgc/pig/cyto/gene/chromo/
SSCG6.htm) only RYR1, UOX and HMG17 have been map-
ped previously using an RH panel (IMpRH panel; Hawken
et al. 1999; Karnuah et al. 2001). Thus, overall the assign-
ment of the 105 genes/ESTs in the present study significantly
increases the density of type I markers on the SSC6 RH map.
Polymorphism discovery and QTL location
A total of 137 gene-based markers were amplified from
Large White and Meishan pools and sequenced. Polymor-
Tab
le1
Continued
Locus
symbol
Gen
enam
e
HSA
location
HSA
distance
(start/stop;Mb)
Accession
no.
Primer
sequen
ces
Tm
(�C)
Product
size
(bp)
LG(AD)
SW607
DNAsegment,
microsatellite
AF2
35332
F:CATCTTGCTCATTGCTGCAT
R:GGACCTTCCAGGCTCTTAGC
56
242
6(8.344)
SW
2466
DNAsegment,
microsatellite
AF2
53824
F:CCTTCCCCCACACAATCTC
R:TGAGTGCAACCGTGGTTAGG
56
150
6(8.477)
SLC2A1(G
LUT1)
Solute
carrier
family2(facilitatedglucose
transporter),member1
1p35-p31.3
43.38/43.42
X17058
F:CCTCGACTGCGTACCAATTCCCTGCTGGCCTTCATATCT
R:ATGAGGATGCCGACGACGAT
61
209
6(8.554)
GUCA2B
Guanylate
cyclase
activator2B(uroguanylin)
1p34-p33
42.65/42.65
Z83746
F:CTTCCGGGTCCAGCTGAAAT
R:TCAAGGCCTGGAAGATGCTG
61
179
6(8.656)
LG,linkagegroup;AD,accumulative
distance
(inrays).
Figure 1 A comprehensive RH map for SSC6 with 120 markers in six
linkage groups. On the left of the SSC6 RH map are shown the
cytogenetic locations of 29 and linkage assignments of 16 previously
published markers; on the right of the SSC6 RH map are the human RH
(GB4: http://www.ncbi.nlm.nih.gov/genemap99/) and human cyto-
genetic positions of the genes mapped using the pig/hamster RH panel.
Cao et al.376
� 2004 International Society for Animal Genetics, Animal Genetics, 35, 367–378

High-resolution radiation hybrid map of porcine chromosome 6 377
� 2004 International Society for Animal Genetics, Animal Genetics, 35, 367–378

phisms were detected for 52 of these markers, and included
SNPs (single nucleotide polymorphisms) and indels (inser-
tion deletion polymorphisms) for 45 and 10 of these
markers, respectively, with three genes having both SNP
and indel polymorphisms. Of these polymorphisms, 23 were
in the coding region of the gene, 36 in non-coding region
with seven genes having polymorphisms in both coding and
non-coding regions. Thirty-eight of these polymorphic loci
are located on the RH map.
The Chinese Meishan breed is one of the most prolific pig
breeds known, with the genetic potential to enhance pro-
duction efficiency of European breeds (Haley & Lee 1993). A
putative QTL for the number of piglets born per litter was
identified on SSC6 at position 104 cM (P-value ¼ 0.0075)
in a Meishan · Yorkshire population (Wilkie et al. 1999).
Evidence also exists for a QTL associated with the number
of nipples, which has been mapped to SSC6 at position
145 cM (P < 0.10) in a Meishan · Large White population
(Cassady et al. 2001). Figure 1 shows that these two QTL
could be associated with markers in linkage group V or VI.
In summary, we have constructed a high-resolution RH
map of SSC6, which we are using currently in a chromo-
some-wide analysis for QTL and candidate genes associated
with prolificacy traits in a Meishan · Large White multi-
generational commercial population.
Acknowledgements
This research was supported by the Genex Swine Group
(Canada), the Natural Sciences and Engineering Research
Council of Canada (NSERC), the Food System Biotechnology
Centre (FSBC) at the University of Guelph, and the Ontario
Ministry of Agriculture and Food (OMAF).
References
Amarante M.R.V., Yang Y.P., Kata S.R., Lopes C.R. & Womack J.E.
(2000) RH maps of bovine chromosomes 15 and 29: conserva-
tion of human chromosomes 11 and 5. Mammalian Genome 11,
364–8.
Boehnke M., Lange K. & Cox D.R. (1991) Statistical methods for
multipoint radiation hybrid mapping. The American Journal of
Human Genetics 49, 1174–88.
Cassady J.P., Johnson R.K., Pomp D., Rohrer G.A., Van Vleck L.D.,
Spiegel E.K. & Gilson K.M. (2001) Identification of quantitative
trait loci affecting reproduction in pigs. Journal of Animal Science
79, 623–33.
Cox D.R., Burmeister M., Price E.R., Kim S. & Myers R.M. (1990)
Radiation hybrid mapping: a somatic cell genetic method for
constructing high-resolution maps of mammalian chromosomes.
Science 250, 245–50.
de Koning D.J., Janss L.L.G., Rattink A.P., Van Oers P.A.M., de Vries
B.J., Groenen M.A.M., van der Poel J.J., de Groot P.N., Brascamp
E.W. & van Arendonk J.A.M. (1999) Detection of quantitative
trait loci for backfat thickness and intramuscular fat content in
pigs (Sus scrofa). Genetics 152, 1679–90.
Goureau A., Yerle M., Schmitz A., Riquet J., Milan D., Pinton P.,
Frelat G. & Gellin J. (1996) Human and porcine correspondence
of chromosome segments using bi-directional chromosome
painting. Genomics 36, 252–62.
Grindflek E., Szyda J., Liu Z. & Lien S. (2001) Detection of quanti-
tative trait loci for meat quality in a commercial slaughter pig
cross. Mammalian Genome 12, 299–304.
Haley C.S. & Lee G.J. (1993) Genetic basis of prolificacy in
Meishan pigs. Journal of Reproduction and Fertility (Suppl.) 48,
247–59.
Hawken R.J., Murtaugh J., Flickinger G.H., Yerle M., Robic A.,
Milan D., Gellin J., Beattie C.W., Schook L.B. & Alexander L.J.
(1999) A first-generation porcine whole-genome radiation hybrid
map. Mammalian Genome 10, 824–30.
Jiang Z., Melville J.S., Cao H., Kumar S., Filipski A. & Gibbins A.M.V.
(2002) Measuring conservation of contiguous sets of autosomal
markers on bovine and porcine genomes in relation to the map of
the human genome. Genome 45, 1–7.
Karnuah A.B., Uenishi H., Kiuchi S., Kojima M., Onishi A., Yasue H.
& Mitsuhashi T. (2001) Assignment of 64 genes expressed in 28-
day-old pig embryo to radiation hybrid map. Mammalian. Genome
12, 518–23.
Kiuchi S., Chen Y.Z., Uenishi H., Hayashi Y., Soeda E. & Yasue H.
(2002) Construction of a dense comparative map between hu-
man chromosome 1p36-p35 and swine chromosome 6 by using
human sequence-tagged sites. Cytogenetic and Genome Research
98, 67–70.
Lahbib-Mansais Y., Leroux S., Milan D., Yerle M., Robic A., Jiang Z.
& Gellin J. (2000) Comparative mapping between humans and
pigs: localization of 58 anchorage markers (TOASTs) by use of
porcine somatic cell and radiation hybrid panels. Mammalian
Genome 11, 1098–106.
Lunetta K.L., Boehnke M., Lange K. & Cox D.R. (1996) Selected
locus and multiple panel models for radiation hybrid mapping.
The American Journal of Human Genetics 59, 717–25.
Ovilo C., Perez-Enciso M., Barragan C., Clop A., Rodriguez C. Oliver
M.A., Toro M.A. & Noguera J.L. (2000) A QTL for intramuscular
fat and backfat thickness is located on porcine chromosome 6.
Mammalian Genome 11, 344–6.
Pinton P., Schibler L., Cribiu E., Gellin J. & Yerle M. (2000) Loca-
lization of 113 anchor loci in pigs: improvement of the com-
parative map for humans, pigs, and goats. Mammalian Genome
11, 306–15.
Rink A., Santschi E.M., Eyer K.M., Roelofs B., Hess M. Godfrey M.,
Karajusuf E.K., Yerle M., Milan D. & Beattie C.W. (2002) A first-
generation EST EH comparative map of the porcine and human
genome. Mammalian Genome 13, 578–87.
Sun H.S., Tuggle C.K., Goureau A., Fitzsimmons C.J., Pinton P.,
Chardon P. & Yerle M. (2001) Precise mapping of breakpoints in
conserved synteny between human chromosome 1 and pig
chromosomes 4, 6 and 9. Animal Genetics 33, 91–6.
Wilkie P.J., Paszek A.A., Beattie C.W., Alexander L.J., Wheeler M.B.
& Schook L.B. (1999) A genomic scan of porcine reproductive
traits reveals possible quantitative trait loci (QTLs) for number of
corpora lutea. Mammalian Genome 10, 573–8.
Cao et al.378
� 2004 International Society for Animal Genetics, Animal Genetics, 35, 367–378
Related Documents











