Molecular Cell, Vol. 17, 603–609, February 18, 2005, Copyright ©2005 by Elsevier Inc. DOI 10.1016/j.molcel.2005.01.015 Short Article A Flexible Domain Is Essential for the Large Step Size and Processivity of Myosin VI arm length (number of IQ motifs) and the size of the working stroke (Warshaw et al., 1998; Purcell et al., 2002; Sakamoto et al., 2003). Surprisingly, myosin VI takes 30–36 nm steps, similar in size to those of myosin V (Rock et al., 2001; Nishikawa et al., 2002; Purcell et al., Ronald S. Rock, 1,3 Bhagavathi Ramamurthy, 2 Alexander R. Dunn, 1 Sara Beccafico, 2 Bhadresh R. Rami, 1 Carl Morris, 2 Benjamin J. Spink, 1 Clara Franzini-Armstrong, 2 James A. Spudich, 1, * and H. Lee Sweeney 2 1 Department of Biochemistry 2002), despite having a shorter lever arm. The large variability in step size suggests that myosin Stanford University School of Medicine Stanford, California 94305 VI uses a larger diffusive search during stepping than other characterized myosins. Therefore, myosin VI should 2 Department of Physiology University of Pennsylvania School of Medicine possess a flexible element that allows the diffusive movement of the heads. Recent results suggest that if 3700 Hamilton Walk Philadelphia, Pennsylvania 19104 such a flexible element exists, it must lie beyond the IQ motif (toward the C terminus) (Okten et al., 2004; Yildiz et al., 2004). In fact, the region immediately following the IQ motif (the proximal tail) has a low probability of Summary forming a stable coiled coil and is thus a likely candidate for flexibility. Here, we present EM and single molecule Myosin VI moves processively along actin with a larger studies that demonstrate that the proximal tail is the step size than expected from the size of the motor. flexible element responsible for the large and variable Here, we show that the proximal tail (the 80-residue step size of myosin VI. segment following the IQ domain) is not a rigid struc- ture but, rather, a flexible domain that permits the Results and Discussion heads to separate. With a GCN4 coiled coil inserted in the proximal tail, the heads are closer together in The Two Heads Are Widely Separated electron microscopy (EM) images, and the motor takes in Myosin VI HMM shorter processive steps. Single-headed myosin VI S1 We examined the myosin VI construct (M6HMM) that constructs take nonprocessive 12 nm steps, sug- was used in our previous study (Rock et al., 2001) by gesting that most of the processive step is covered rotary shadowing EM. In this construct, the native tail by a diffusive search for an actin binding site. Based sequence that contains predicted coiled-coil regions on these results, we present a mechanical model that (see below) is truncated at Arg992 and followed by a describes stepping under an applied load. GCN4-p1 coiled coil (Lumb et al., 1994), YFP, and a FLAG affinity tag. All M6HMM images are heavy-chain Introduction dimers, with two distinct heads connected by narrow stalks to a common tail (Figure 1A). The heads have the Myosin VI is unique in the myosin superfamily with sev- typical globular appearance of myosin S1 domains. The eral fascinating characteristics. It consists of an N-ter- C-terminal region has an additional globular mass that minal motor domain, a 53-residue unique insert and a is often resolvable into two distinct components, pre- single IQ motif (both of which bind calmodulin [Bahloul sumably the YFP moieties. In some molecules, the stalks et al., 2004]), a putative coiled-coil heptad repeat region, converge after a short distance, and the two heads are and a C-terminal tail domain (Hasson and Mooseker, close together; in others, the heads are separated by a 1994; Sellers and Goodson, 1995). It was the first myosin much larger distance. shown to move in the opposite direction on actin fila- The observed distance between the centers of the ments as compared to myosin II (Wells et al., 1999). two heads of M6HMM is 27 6 nm (SD, n 37). This Consistent with an earlier report (Lister et al., 2004), we value is greater than expected if the two heads were have seen that full-length myosin VI is monomeric (data held together at the C terminal end of the IQ/CaM do- not shown). However, single molecules of dimeric myo- main, where the heads should span no more than 18 sin VI take multiple steps along an actin filament (Rock nm. Flexibility was also observed in the EM images of et al., 2001; Nishikawa et al., 2002), consistent with the Nishikawa et al. (2002), where the last three panels of kinetic properties of the motor (De La Cruz et al., 2001). their Figure 1B show head separations of 20–30 nm. Myosin superfamily members contain a variable num- ber of IQ motifs that immediately follow the motor do- The Proximal Tail Is Not Predicted main. These IQ motifs form the calmodulin/light chain to be a Coiled Coil binding sites that comprise the myosin lever arm Given the small size of the motor domain and lever arm, (Holmes and Geeves, 2000). For myosins other than my- some portion of myosin VI must extend to allow the osin VI, there is good correspondence between the lever large head-to-head separation observed in the EM im- ages. The tail sequence is predicted to form a coiled coil, *Correspondence: [email protected] although the boundaries of the coiled coil vary among 3 Present address: Department of Biochemistry and Molecular Biol- prediction methods. We examined the tail sequence by ogy, University of Chicago, 920 E. 58th Street, Chicago, Illinois 60637. using the PAIRCOIL algorithm (Berger et al., 1995),

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Molecular Cell, Vol. 17, 603–609, February 18, 2005, Copyright ©2005 by Elsevier Inc. DOI 10.1016/j.molcel.2005.01.015
Short ArticleA Flexible Domain Is Essentialfor the Large Step Sizeand Processivity of Myosin VI
arm length (number of IQ motifs) and the size of theworking stroke (Warshaw et al., 1998; Purcell et al., 2002;Sakamoto et al., 2003). Surprisingly, myosin VI takes30–36 nm steps, similar in size to those of myosin V(Rock et al., 2001; Nishikawa et al., 2002; Purcell et al.,
Ronald S. Rock,1,3 Bhagavathi Ramamurthy,2
Alexander R. Dunn,1 Sara Beccafico,2
Bhadresh R. Rami,1 Carl Morris,2 Benjamin J. Spink,1
Clara Franzini-Armstrong,2 James A. Spudich,1,*and H. Lee Sweeney2
1Department of Biochemistry 2002), despite having a shorter lever arm.The large variability in step size suggests that myosinStanford University School of Medicine
Stanford, California 94305 VI uses a larger diffusive search during stepping thanother characterized myosins. Therefore, myosin VI should2 Department of Physiology
University of Pennsylvania School of Medicine possess a flexible element that allows the diffusivemovement of the heads. Recent results suggest that if3700 Hamilton Walk
Philadelphia, Pennsylvania 19104 such a flexible element exists, it must lie beyond the IQmotif (toward the C terminus) (Okten et al., 2004; Yildizet al., 2004). In fact, the region immediately followingthe IQ motif (the proximal tail) has a low probability ofSummaryforming a stable coiled coil and is thus a likely candidatefor flexibility. Here, we present EM and single moleculeMyosin VI moves processively along actin with a largerstudies that demonstrate that the proximal tail is thestep size than expected from the size of the motor.flexible element responsible for the large and variableHere, we show that the proximal tail (the �80-residuestep size of myosin VI.segment following the IQ domain) is not a rigid struc-
ture but, rather, a flexible domain that permits theResults and Discussionheads to separate. With a GCN4 coiled coil inserted
in the proximal tail, the heads are closer together inThe Two Heads Are Widely Separatedelectron microscopy (EM) images, and the motor takesin Myosin VI HMMshorter processive steps. Single-headed myosin VI S1We examined the myosin VI construct (M6HMM) thatconstructs take nonprocessive 12 nm steps, sug-was used in our previous study (Rock et al., 2001) bygesting that most of the processive step is coveredrotary shadowing EM. In this construct, the native tailby a diffusive search for an actin binding site. Basedsequence that contains predicted coiled-coil regionson these results, we present a mechanical model that(see below) is truncated at Arg992 and followed by adescribes stepping under an applied load.GCN4-p1 coiled coil (Lumb et al., 1994), YFP, and aFLAG affinity tag. All M6HMM images are heavy-chainIntroductiondimers, with two distinct heads connected by narrowstalks to a common tail (Figure 1A). The heads have theMyosin VI is unique in the myosin superfamily with sev-typical globular appearance of myosin S1 domains. Theeral fascinating characteristics. It consists of an N-ter-C-terminal region has an additional globular mass thatminal motor domain, a 53-residue unique insert and ais often resolvable into two distinct components, pre-single IQ motif (both of which bind calmodulin [Bahloulsumably the YFP moieties. In some molecules, the stalkset al., 2004]), a putative coiled-coil heptad repeat region,converge after a short distance, and the two heads areand a C-terminal tail domain (Hasson and Mooseker,close together; in others, the heads are separated by a1994; Sellers and Goodson, 1995). It was the first myosinmuch larger distance.shown to move in the opposite direction on actin fila-
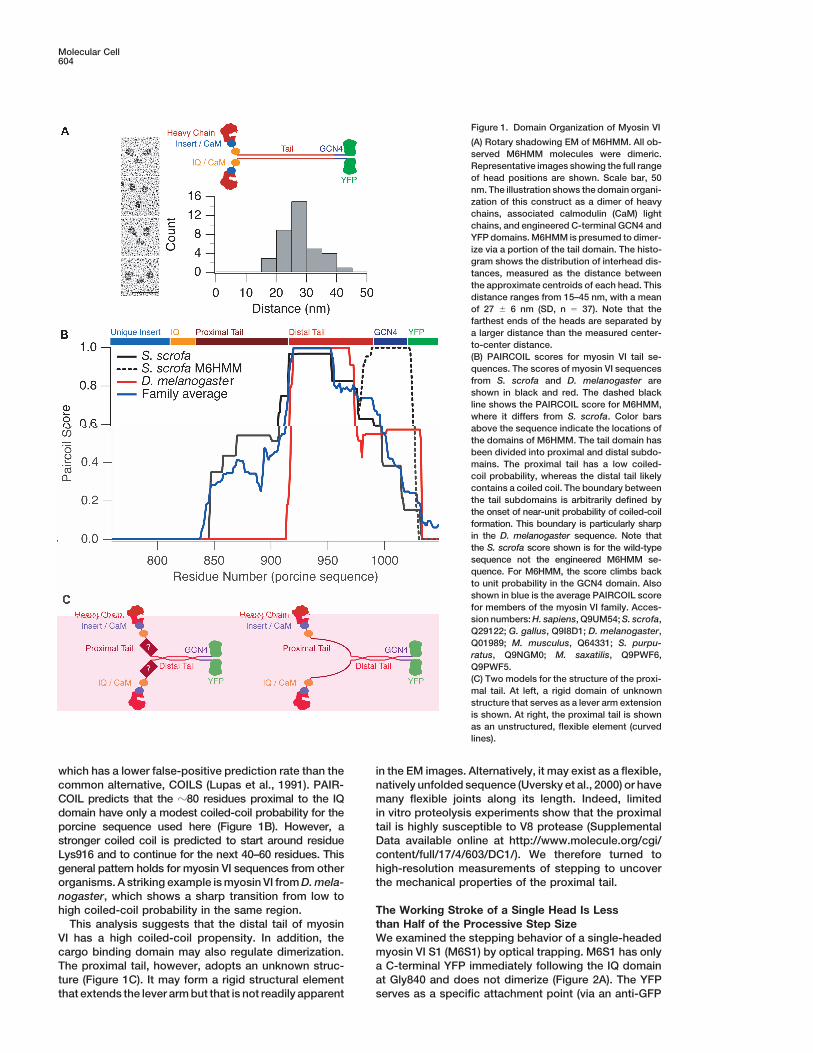
The observed distance between the centers of thements as compared to myosin II (Wells et al., 1999).two heads of M6HMM is 27 � 6 nm (SD, n � 37). ThisConsistent with an earlier report (Lister et al., 2004), wevalue is greater than expected if the two heads werehave seen that full-length myosin VI is monomeric (dataheld together at the C terminal end of the IQ/CaM do-not shown). However, single molecules of dimeric myo-main, where the heads should span no more than �18sin VI take multiple steps along an actin filament (Rocknm. Flexibility was also observed in the EM images ofet al., 2001; Nishikawa et al., 2002), consistent with theNishikawa et al. (2002), where the last three panels ofkinetic properties of the motor (De La Cruz et al., 2001).their Figure 1B show head separations of �20–30 nm.Myosin superfamily members contain a variable num-
ber of IQ motifs that immediately follow the motor do-The Proximal Tail Is Not Predictedmain. These IQ motifs form the calmodulin/light chainto be a Coiled Coilbinding sites that comprise the myosin lever armGiven the small size of the motor domain and lever arm,(Holmes and Geeves, 2000). For myosins other than my-some portion of myosin VI must extend to allow theosin VI, there is good correspondence between the leverlarge head-to-head separation observed in the EM im-ages. The tail sequence is predicted to form a coiled coil,
*Correspondence: [email protected] the boundaries of the coiled coil vary among3 Present address: Department of Biochemistry and Molecular Biol-prediction methods. We examined the tail sequence byogy, University of Chicago, 920 E. 58th Street, Chicago, Illinois
60637. using the PAIRCOIL algorithm (Berger et al., 1995),
Molecular Cell604
Figure 1. Domain Organization of Myosin VI
(A) Rotary shadowing EM of M6HMM. All ob-served M6HMM molecules were dimeric.Representative images showing the full rangeof head positions are shown. Scale bar, 50nm. The illustration shows the domain organi-zation of this construct as a dimer of heavychains, associated calmodulin (CaM) lightchains, and engineered C-terminal GCN4 andYFP domains. M6HMM is presumed to dimer-ize via a portion of the tail domain. The histo-gram shows the distribution of interhead dis-tances, measured as the distance betweenthe approximate centroids of each head. Thisdistance ranges from 15–45 nm, with a meanof 27 � 6 nm (SD, n � 37). Note that thefarthest ends of the heads are separated bya larger distance than the measured center-to-center distance.(B) PAIRCOIL scores for myosin VI tail se-quences. The scores of myosin VI sequencesfrom S. scrofa and D. melanogaster areshown in black and red. The dashed blackline shows the PAIRCOIL score for M6HMM,where it differs from S. scrofa. Color barsabove the sequence indicate the locations ofthe domains of M6HMM. The tail domain hasbeen divided into proximal and distal subdo-mains. The proximal tail has a low coiled-coil probability, whereas the distal tail likelycontains a coiled coil. The boundary betweenthe tail subdomains is arbitrarily defined bythe onset of near-unit probability of coiled-coilformation. This boundary is particularly sharpin the D. melanogaster sequence. Note thatthe S. scrofa score shown is for the wild-typesequence not the engineered M6HMM se-quence. For M6HMM, the score climbs backto unit probability in the GCN4 domain. Alsoshown in blue is the average PAIRCOIL scorefor members of the myosin VI family. Acces-sion numbers: H. sapiens, Q9UM54; S. scrofa,Q29122; G. gallus, Q9I8D1; D. melanogaster,Q01989; M. musculus, Q64331; S. purpu-ratus, Q9NGM0; M. saxatilis, Q9PWF6,Q9PWF5.(C) Two models for the structure of the proxi-mal tail. At left, a rigid domain of unknownstructure that serves as a lever arm extensionis shown. At right, the proximal tail is shownas an unstructured, flexible element (curvedlines).
which has a lower false-positive prediction rate than the in the EM images. Alternatively, it may exist as a flexible,natively unfolded sequence (Uversky et al., 2000) or havecommon alternative, COILS (Lupas et al., 1991). PAIR-
COIL predicts that the �80 residues proximal to the IQ many flexible joints along its length. Indeed, limitedin vitro proteolysis experiments show that the proximaldomain have only a modest coiled-coil probability for the
porcine sequence used here (Figure 1B). However, a tail is highly susceptible to V8 protease (SupplementalData available online at http://www.molecule.org/cgi/stronger coiled coil is predicted to start around residue
Lys916 and to continue for the next 40–60 residues. This content/full/17/4/603/DC1/). We therefore turned tohigh-resolution measurements of stepping to uncovergeneral pattern holds for myosin VI sequences from other
organisms. A striking example is myosin VI from D. mela- the mechanical properties of the proximal tail.nogaster, which shows a sharp transition from low tohigh coiled-coil probability in the same region. The Working Stroke of a Single Head Is Less
than Half of the Processive Step SizeThis analysis suggests that the distal tail of myosinVI has a high coiled-coil propensity. In addition, the We examined the stepping behavior of a single-headed
myosin VI S1 (M6S1) by optical trapping. M6S1 has onlycargo binding domain may also regulate dimerization.The proximal tail, however, adopts an unknown struc- a C-terminal YFP immediately following the IQ domain
at Gly840 and does not dimerize (Figure 2A). The YFPture (Figure 1C). It may form a rigid structural elementthat extends the lever arm but that is not readily apparent serves as a specific attachment point (via an anti-GFP
Myosin VI Proximal Tail605
on average, the diffusive search covers twice the dis-placement of the working stroke. In spite of the wide-ranging diffusive search, myosin VI steps predominantlyin one direction (only one out of seven steps is backwardat 1.7 pN load) (Rock et al., 2001; Altman et al., 2004).A potential explanation for this feat is that the 12 nmstroke is sufficient to bias the stepping in the forwarddirection and that there are a limited number of accessi-ble actin binding sites of the proper azimuthal orienta-tion for the free head to bind in the reverse direction.Indeed, the helical geometry of an actin filament seemsideally suited to allow such biased diffusive searchesfor both myosin V and myosin VI. A 12 nm working strokewill place the free head near actin sites that are at anunfavorable azimuth so that additional diffusion towardthe pointed end must occur to reach the most favoredbinding-sites (Rock et al., 2001).
Previous EM reconstructions of myosin VI decoratedactin filaments revealed that, in the presumed rigorpoststroke state, the myosin VI lever arm is orientednearly parallel to the actin axis. Given a 12 nm workingstroke of M6S1 and an approximately 10 nm long leverarm, the prepowerstroke orientation of the lever armshould be nearly perpendicular to the actin filament.Indeed, EM images of full-length myosin VI in the ADP•Pistate show the lever arm in what would be a roughlyperpendicular orientation to the actin filament (Lister etal., 2004). Larger steps of 18 nm have been measuredwith full-length myosin VI (Lister et al., 2004), possiblybecause regions C terminal to the calmodulin bindingdomain somehow contribute to the functional length ofthe lever arm (Lister et al., 2004).
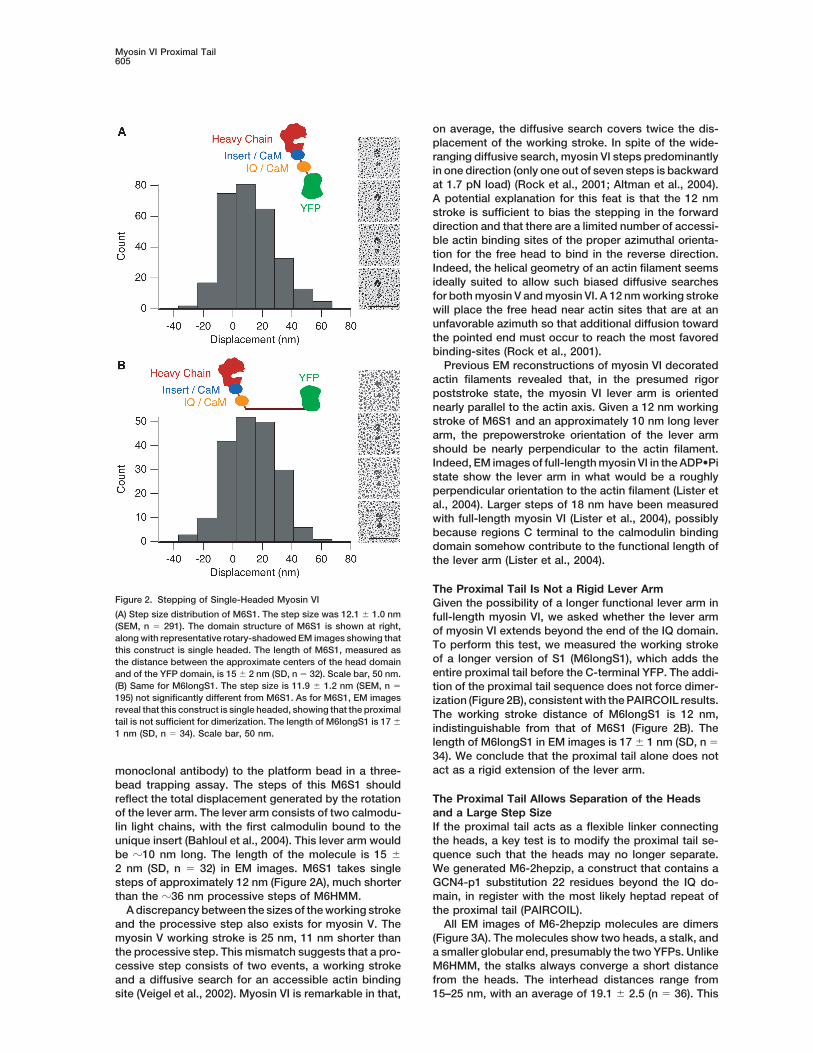
The Proximal Tail Is Not a Rigid Lever ArmFigure 2. Stepping of Single-Headed Myosin VI Given the possibility of a longer functional lever arm in(A) Step size distribution of M6S1. The step size was 12.1 � 1.0 nm full-length myosin VI, we asked whether the lever arm(SEM, n � 291). The domain structure of M6S1 is shown at right, of myosin VI extends beyond the end of the IQ domain.along with representative rotary-shadowed EM images showing that
To perform this test, we measured the working strokethis construct is single headed. The length of M6S1, measured asof a longer version of S1 (M6longS1), which adds thethe distance between the approximate centers of the head domainentire proximal tail before the C-terminal YFP. The addi-and of the YFP domain, is 15 � 2 nm (SD, n � 32). Scale bar, 50 nm.
(B) Same for M6longS1. The step size is 11.9 � 1.2 nm (SEM, n � tion of the proximal tail sequence does not force dimer-195) not significantly different from M6S1. As for M6S1, EM images ization (Figure 2B), consistent with the PAIRCOIL results.reveal that this construct is single headed, showing that the proximal The working stroke distance of M6longS1 is 12 nm,tail is not sufficient for dimerization. The length of M6longS1 is 17 �
indistinguishable from that of M6S1 (Figure 2B). The1 nm (SD, n � 34). Scale bar, 50 nm.length of M6longS1 in EM images is 17 � 1 nm (SD, n �34). We conclude that the proximal tail alone does notact as a rigid extension of the lever arm.monoclonal antibody) to the platform bead in a three-
bead trapping assay. The steps of this M6S1 shouldreflect the total displacement generated by the rotation The Proximal Tail Allows Separation of the Heads
and a Large Step Sizeof the lever arm. The lever arm consists of two calmodu-lin light chains, with the first calmodulin bound to the If the proximal tail acts as a flexible linker connecting
the heads, a key test is to modify the proximal tail se-unique insert (Bahloul et al., 2004). This lever arm wouldbe �10 nm long. The length of the molecule is 15 � quence such that the heads may no longer separate.
We generated M6-2hepzip, a construct that contains a2 nm (SD, n � 32) in EM images. M6S1 takes singlesteps of approximately 12 nm (Figure 2A), much shorter GCN4-p1 substitution 22 residues beyond the IQ do-
main, in register with the most likely heptad repeat ofthan the �36 nm processive steps of M6HMM.A discrepancy between the sizes of the working stroke the proximal tail (PAIRCOIL).
All EM images of M6-2hepzip molecules are dimersand the processive step also exists for myosin V. Themyosin V working stroke is 25 nm, 11 nm shorter than (Figure 3A). The molecules show two heads, a stalk, and
a smaller globular end, presumably the two YFPs. Unlikethe processive step. This mismatch suggests that a pro-cessive step consists of two events, a working stroke M6HMM, the stalks always converge a short distance
from the heads. The interhead distances range fromand a diffusive search for an accessible actin bindingsite (Veigel et al., 2002). Myosin VI is remarkable in that, 15–25 nm, with an average of 19.1 � 2.5 (n � 36). This
Molecular Cell606
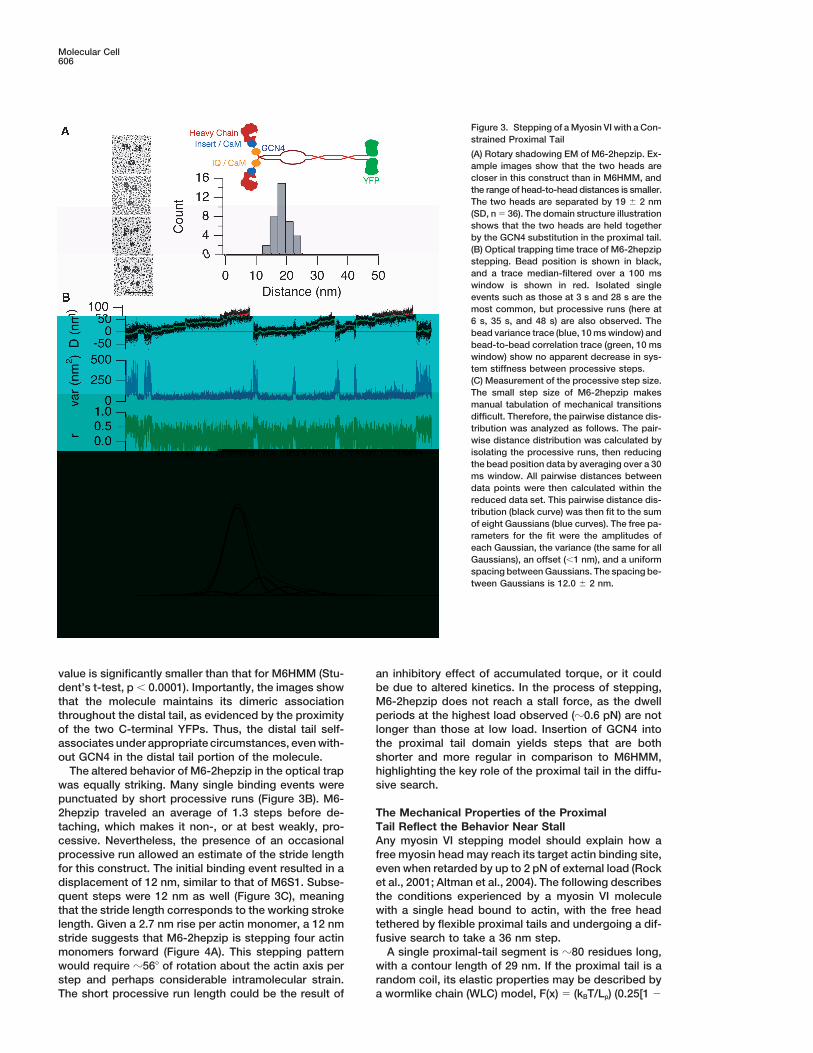
Figure 3. Stepping of a Myosin VI with a Con-strained Proximal Tail
(A) Rotary shadowing EM of M6-2hepzip. Ex-ample images show that the two heads arecloser in this construct than in M6HMM, andthe range of head-to-head distances is smaller.The two heads are separated by 19 � 2 nm(SD, n � 36). The domain structure illustrationshows that the two heads are held togetherby the GCN4 substitution in the proximal tail.(B) Optical trapping time trace of M6-2hepzipstepping. Bead position is shown in black,and a trace median-filtered over a 100 mswindow is shown in red. Isolated singleevents such as those at 3 s and 28 s are themost common, but processive runs (here at6 s, 35 s, and 48 s) are also observed. Thebead variance trace (blue, 10 ms window) andbead-to-bead correlation trace (green, 10 mswindow) show no apparent decrease in sys-tem stiffness between processive steps.(C) Measurement of the processive step size.The small step size of M6-2hepzip makesmanual tabulation of mechanical transitionsdifficult. Therefore, the pairwise distance dis-tribution was analyzed as follows. The pair-wise distance distribution was calculated byisolating the processive runs, then reducingthe bead position data by averaging over a 30ms window. All pairwise distances betweendata points were then calculated within thereduced data set. This pairwise distance dis-tribution (black curve) was then fit to the sumof eight Gaussians (blue curves). The free pa-rameters for the fit were the amplitudes ofeach Gaussian, the variance (the same for allGaussians), an offset (�1 nm), and a uniformspacing between Gaussians. The spacing be-tween Gaussians is 12.0 � 2 nm.
value is significantly smaller than that for M6HMM (Stu- an inhibitory effect of accumulated torque, or it couldbe due to altered kinetics. In the process of stepping,dent’s t-test, p � 0.0001). Importantly, the images show
that the molecule maintains its dimeric association M6-2hepzip does not reach a stall force, as the dwellperiods at the highest load observed (�0.6 pN) are notthroughout the distal tail, as evidenced by the proximity
of the two C-terminal YFPs. Thus, the distal tail self- longer than those at low load. Insertion of GCN4 intothe proximal tail domain yields steps that are bothassociates under appropriate circumstances, even with-
out GCN4 in the distal tail portion of the molecule. shorter and more regular in comparison to M6HMM,highlighting the key role of the proximal tail in the diffu-The altered behavior of M6-2hepzip in the optical trap
was equally striking. Many single binding events were sive search.punctuated by short processive runs (Figure 3B). M6-2hepzip traveled an average of 1.3 steps before de- The Mechanical Properties of the Proximal
Tail Reflect the Behavior Near Stalltaching, which makes it non-, or at best weakly, pro-cessive. Nevertheless, the presence of an occasional Any myosin VI stepping model should explain how a
free myosin head may reach its target actin binding site,processive run allowed an estimate of the stride lengthfor this construct. The initial binding event resulted in a even when retarded by up to 2 pN of external load (Rock
et al., 2001; Altman et al., 2004). The following describesdisplacement of 12 nm, similar to that of M6S1. Subse-quent steps were 12 nm as well (Figure 3C), meaning the conditions experienced by a myosin VI molecule
with a single head bound to actin, with the free headthat the stride length corresponds to the working strokelength. Given a 2.7 nm rise per actin monomer, a 12 nm tethered by flexible proximal tails and undergoing a dif-
fusive search to take a 36 nm step.stride suggests that M6-2hepzip is stepping four actinmonomers forward (Figure 4A). This stepping pattern A single proximal-tail segment is �80 residues long,
with a contour length of 29 nm. If the proximal tail is awould require �56� of rotation about the actin axis perstep and perhaps considerable intramolecular strain. random coil, its elastic properties may be described by
a wormlike chain (WLC) model, F(x) � (kBT/Lp) (0.25[1 �The short processive run length could be the result of

Myosin VI Proximal Tail607
x/L]�2 � 0.25 � x/L), where x is the extension, L isthe contour length, kBT is thermal energy, and Lp is thepersistence length (Bustamante et al., 1994). The persis-tence length of unfolded polypeptides is approximately0.9 nm at the low forces considered here (Schwaiger etal., 2004).
A simple mechanical description of the myosin VI mo-tor is shown in Figures 4B and 4C. In this model, eachhead and lever arm are coupled to the distal tail via WLCelastic elements, the proximal tails. Given the 12 nmworking stroke, the proximal tails would need to extendto �24 nm for both heads to bind 36 nm apart (Figure4C). The calculated stiffness (k) of both proximal tailsegments (WLC of 160 residues total, LP � 0.9 nm) is�0.125 pN•nm�1 and is nearly constant over a 24 nmextension. This stiffness yields a relaxation time, � ��/k � 0.48 s, for a myosin head with a viscous dragcoefficient � of 60 pN•s/m (Howard, 2001). To diffuse24 nm, the free head must overcome an energy barrierof 36 pN•nm, or 8.8 kBT, under zero load. The Kramersfirst-passage time (Figure 4D) across this distance is �1ms, meaning that such diffusion is not rate limiting atsaturating ATP conditions (Kramers, 1940; Howard,2001).
In the presence of a constant 2.2 pN load sufficientto stall the motor, the situation is quite different. Now,the equilibrium position of the free head is shifted back-ward by �8.8 nm, as only one of the two proximal tailsegments is under load (a single segment is twice asstiff as the full proximal tail; 0.25 pN/nm). The free headmust therefore diffuse forward 32.8 nm to take a 36 nmstep, corresponding to an energy barrier of 67 pN•nm.The Kramers first-passage time under these conditionsis 1.4 s, sufficiently large to be considered stalled. Atlower loads, the first-passage time drops rapidly, lead-ing to an abrupt decrease in dwell time (Figure 4D). Thesharp transition to stall around 2 pN broadly recapitu-lates the kinetics measured earlier (Altman et al., 2004),suggesting that deformation of the proximal tail maycontribute to stalling under high load. In this model, themotor stalls at high load with the free head in a detachedstate, or the free head reattaches to yield a short step.Both of these possibilities will presumably block theallosteric communication recently observed by Robblee
Figure 4. Structural Model of Myosin VI Stepping et al. (2004), where high intramolecular strain acceler-(A) Model for M6HMM and M6-2hepzip, each stepping from right ates nucleotide binding to the trailing head. The pre-to left. For M6HMM (left), the unstructured proximal tails allow a dicted stall behavior depends sensitively on the proxi-broad diffusive search. The lever arm swing biases the diffusive mal tail stiffness, and our estimate may be improved bysearch 12 nm toward the pointed end. The long-pitch helix of the
direct measurement of this stiffness.actin filament provides an additional bias toward sites that are at aThe above analyses present a highly simplified modelnearby azimuth. For M6-2hepzip (right), the coiled-coil insert im-
of a complex system. The proximal tail has a highposes a significant constraint upon the diffusive search. The twoheads span four actin monomers, with regular steps. Such a step-ping pattern requires tight spiraling around the actin filament.(B) Mechanical model of M6HMM, with the trailing head (T) attachedto actin and the leading head (L) free. The 12 nm working stroke of 36 nm step as a function of load. The equilibrium position of thethe trailing head is indicated. The proximal tails are shown as free head is given by �F/2k, where k is the combined stiffness ofsprings. Dark circles indicate junctions that are expected to act as both segments of proximal tail (0.12 pN/nm). In this model, thefree swivels. proximal-tail stiffness remains constant, but as the external load(C) Mechanical model of M6HMM after the lead head binds. To span increases, the distance traversed by the free head also increases.36 nm, the proximal tails must together extend 24 nm. The head The free head must therefore diffuse over an energy barrier givenseparations before and after a step do not have to be equal, as is by U0 � k(24 � F/2k)2/2. This energy barrier is used to find a first-assumed in this illustration. Note that the observed step size is half passage time as �/k(/4)1/2(kbT/U0)1/2exp(U0/kbT), where � is the vis-the distance traversed by the trailing head when it becomes the cous drag coefficient, k is the stiffness, and kbT is thermal energy.new leading head. The dwell time curve adds a zero-load, rate-limiting transition at �3(D) Calculated first-passage time (black) and dwell time (red) for a s�1 under these conditions (Altman et al., 2004).

Molecular Cell608
10 mM DTT, and 4 mM MgCl2), with variable ATP (20 M unless�-helical propensity, which may tend to increase theotherwise noted), an ATP regeneration system (100 g•mL�1 cre-persistence length and lower the stiffness. This wouldatine phosphokinase, and 1 mM creatine phosphate), and an oxygenin turn lower the energetic barrier and facilitate the diffu-scavenging system (25 g•mL�1 glucose oxidase, 45 g•mL�1 cata-
sive search. In addition, the proximal tail is highly lase, and 1% glucose). Assays were performed at 23�C. Three-beadcharged (pI 10.2, net charge of �5), which may function optical trapping methods have been previously described (Mehta
et al., 1998; Rock et al., 2001). Briefly, 10–50 ng•mL�1 of the YFP-as a means of electrostatic steering of an unbound headtagged myosin VI was surface attached with anti-GFP monoclonaland might even provide binding of the flexible linkerantibodies (Quantum, 3E6, 0.05 mg•mL�1). Two optically trapped,to the actin filament in the poststroke state. A similar1 m diameter polystyrene beads were attached to each end of anelectrostatic interaction has been proposed for the neckactin filament via a streptavidin-biotin link, and the resulting assem-
region of kinesin (Thorn et al., 2000). Finally, the ob- bly was stretched to 1–2 pN of tension. The actin filament wasserved stalling behavior of myosin VI may also be influ- moved near surface bound, nitrocellulose-coated silica spheres that
served as platforms for single myosin VI molecules. Both polystyreneenced by the load-dependent kinetic properties of thebeads were tracked with nm resolution at 10 kHz. Myosin concentra-catalytic domain.tions were set so only 10% of the platforms tested showed anWe currently disfavor an alternative model, where theactin interaction.proximal tail of the bound head is docked to the head
Binding events were identified in data records by using Page’sor neck in some rigid structure to support the load, that test on the bead correlation signal (Mehta et al., 1997; Knight et al.,we had entertained earlier (Altman et al., 2004). The 2001). Event durations were tabulated, along with event amplitudes
(relative to the mean equilibrium position for detached periods total-above analyses indicate that such a docking is not nec-ing 1 s before and after the event). Amplitudes were corrected foressary for processive movement against a load. Further-series compliance by using the method of correlated thermal diffu-more, in this head-docking model, the entire diffusivesion (Mehta et al., 1997). Analyzed datasets were aggregates of fivesearch involves extension of only the free head’s proxi-molecules in the case of M6S1, one molecule for M6longS1, and
mal tail. Such a 24 nm extension is nearly the entire two molecules of M6-2hepzip.contour length of a single proximal tail, requiring forcesin excess of 120 pN. Acknowledgments
Both myosin V and myosin VI use a combination of aThe authors would like to thank Matthias Rief and David Altman forworking stroke and a diffusive search to step pro-helpful discussions. R.S.R. is supported by a Burroughs Wellcomecessively. Why has myosin VI evolved such that diffusionCareer Award at the Scientific Interface. A.R.D. is a Jane Coffin
is the major component of the step unlike myosin V? Childs Postdoctoral Fellow. C.F.-A., J.A.S., and H.L.S. are supportedPerhaps the flexible coupling is necessary for the load- by grants from the National Institutes of Health.induced switching from motor activity to an anchor (Alt-
Received: August 24, 2004man et al., 2004). Another possibility is that this motorRevised: December 17, 2004is designed to straddle neighboring actin filaments inAccepted: January 7, 2005its role in the inner ear or in endocytosis, and thereforePublished: February 17, 2005needs unusual flexibility.
ReferencesExperimental Procedures
Altman, D., Sweeney, H.L., and Spudich, J.A. (2004). The mechanismProtein Constructs and Expressionof myosin VI translocation and its load-induced anchoring. CellThe construction of porcine double-headed myosin VI (M6HMM)116, 737–749.containing GCN4 and YFP was described previously (Rock et al.,Bahloul, A., Chevreux, G., Wells, A.L., Martin, D., Nolt, J., Yang, Z.,2001). A double-headed construct where the sequence followingChen, L.Q., Potier, N., Van Dorsselaer, A., Rosenfeld, S., et al. (2004).the IQ motif formed a stable coiled coil (M6-2hepzip) was made byThe unique insert in myosin VI is a structural calcium-calmodulinreplacing amino acids Val858 to Ser888 with the first 30 residuesbinding site. Proc. Natl. Acad. Sci. USA 101, 4787–4792.of GCN4-p1 (Trybus et al., 1997). The native pig myosin VI continued
through Arg992, followed by a glycine, YFP, and a FLAG tag (GDY Berger, B., Wilson, D.B., Wolf, E., Tonchev, T., Milla, M., and Kim,KDDDK). P.S. (1995). Predicting coiled coils by use of pairwise residue corre-
The two single-headed myosin VI constructs (M6S1 and lations. Proc. Natl. Acad. Sci. USA 92, 8259–8263.M6longS1) were generated by truncating the cDNA coding for pig Bustamante, C., Marko, J.F., Siggia, E.D., and Smith, S. (1994). En-myosin VI after the codons corresponding to Gly840 and Lys918. tropic elasticity of lambda-phage DNA. Science 265, 1599–1600.YFP and a FLAG tag were appended to the C terminus of the coding
De La Cruz, E.M., Ostap, E.M., and Sweeney, H.L. (2001). Kineticsequences. The generation of recombinant baculovirus, expression
mechanism and regulation of myosin VI. J. Biol. Chem. 276, 32373–in SF9 cells, and protein purification followed previously published
32381.procedures (Sweeney et al., 1998; De La Cruz et al., 2001).
Hasson, T., and Mooseker, M.S. (1994). Porcine myosin-VI: charac-terization of a new mammalian unconventional myosin. J. Cell Biol.EM127, 425–440.Myosin constructs were stored in a solution containing 50 mM KCl,
1 mM MgCl2, 1mM EGTA, 1mM DTT, 10 mM imidazole•HCl (pH Holmes, K.C., and Geeves, M.A. (2000). The structural basis of mus-7.0), 200 M ADP, and 50% (v/v) glycerol. The protein suspensions cle contraction. Philos. Trans. R. Soc. Lond. B Biol. Sci. 355,containing approximately 30 g•mL�1 of protein were finely sprayed 419–431.on freshly split mica (Tyler and Branton, 1980; Winkelmann and Howard, J. (2001). Mechanics of Motor Proteins and the Cytoskele-Lowey, 1986). The droplets were dried, shadowed with platinum at ton (Sunderland, MA: Sinauer Associates, Inc.).7�, and replicated with carbon in a freeze-fracture apparatus (model
Knight, A.E., Veigel, C., Chambers, C., and Molloy, J.E. (2001). Analy-BFA 400; Balzers S.p.A., Hudson, NH). The replicas were photo-
sis of single-molecule mechanical recordings: application to acto-graphed in a Philips 410 electron microscope.
myosin interactions. Prog. Biophys. Mol. Biol. 77, 45–72.
Kramers, H.A. (1940). Brownian motion in a field of force and theOptical Trappingdiffusion model of chemical reactions. Phys. 7, 284–304.All motility assays were conducted in assay buffer (AB) (25 mM
imidazole HCl, [pH 7.4], 25 mM KCl, 5 M calmodulin, 1 mM EGTA, Lister, I., Schmitz, S., Walker, M., Trinick, J., Buss, F., Veigel, C.,

Myosin VI Proximal Tail609
and Kendrick-Jones, J. (2004). A monomeric myosin VI with a large mechanism with its lever arm undergoing fluctuations whenattached to actin. J. Biol. Chem. 279, 37223–37226.working stroke. EMBO J. 23, 1729–1738.
Lumb, K.J., Carr, C.M., and Kim, P.S. (1994). Subdomain folding ofthe coiled coil leucine zipper from the bZIP transcriptional activatorGCN4. Biochemistry 33, 7361–7367.
Lupas, A., Van Dyke, M., and Stock, J. (1991). Predicting coiled coilsfrom protein sequences. Science 252, 1162–1164.
Mehta, A.D., Finer, J.T., and Spudich, J.A. (1997). Detection of single-molecule interactions using correlated thermal diffusion. Proc. Natl.Acad. Sci. USA 94, 7927–7931.
Mehta, A.D., Finer, J.T., and Spudich, J.A. (1998). Use of opticaltraps in single-molecule study of nonprocessive biological motors.Methods Enzymol. 298, 436–459.
Nishikawa, S., Homma, K., Komori, Y., Iwaki, M., Wazawa, T., Hiki-koshi Iwane, A., Saito, J., Ikebe, R., Katayama, E., Yanagida, T., andIkebe, M. (2002). Class VI myosin moves processively along actinfilaments backward with large steps. BBRC 290, 311–317.
Okten, Z., Churchman, L.S., Rock, R.S., and Spudich, J.A. (2004).Myosin VI walks hand-over-hand along actin. Nat. Struct. Mol. Biol.11, 884–887.
Purcell, T.J., Morris, C., Spudich, J.A., and Sweeney, H.L. (2002).Role of the lever arm in the processive stepping of myosin V. Proc.Natl. Acad. Sci. USA 99, 14159–14164.
Robblee, J.P., Olivares, A.O., and de la Cruz, E.M. (2004). Mechanismof nucleotide binding to actomyosin VI: evidence for allosteric head-head communication. J. Biol. Chem. 279, 38608–38617.
Rock, R.S., Rice, S.E., Wells, A.L., Purcell, T.J., Spudich, J.A., andSweeney, H.L. (2001). Myosin VI is a processive motor with a largestep size. Proc. Natl. Acad. Sci. USA 98, 13655–13659.
Sakamoto, T., Wang, F., Schmitz, S., Xu, Y., Xu, Q., Molloy, J.E.,Veigel, C., and Sellers, J.R. (2003). Neck length and processivity ofmyosin V. J. Biol. Chem. 278, 29201–29207.
Schwaiger, I., Kardinal, A., Schleicher, M., Noegel, A.A., and Rief, M.(2004). A mechanical unfolding intermediate in an actin-crosslinkingprotein. Nat. Struct. Mol. Biol. 11, 81–85.
Sellers, J.R., and Goodson, H.V. (1995). Motor proteins 2: myosin.Protein Profile 2, 1323–1423.
Sweeney, H.L., Rosenfeld, S.S., Brown, F., Faust, L., Smith, J., Xing,J., Stein, L.A., and Sellers, J.R. (1998). Kinetic tuning of myosin viaa flexible loop adjacent to the nucleotide binding pocket. J. Biol.Chem. 273, 6262–6270.
Thorn, K.S., Ubersax, J.A., and Vale, R.D. (2000). Engineering theprocessive run length of the kinesin motor. J. Cell Biol. 151, 1093–1100.
Trybus, K.M., Freyzon, Y., Faust, L.Z., and Sweeney, H.L. (1997).Spare the rod, spoil the regulation: necessity for a myosin rod. Proc.Natl. Acad. Sci. USA 94, 48–52.
Tyler, J.M., and Branton, D. (1980). Rotary shadowing of extendedmolecules dried from glycerol. J. Ultrastruct. Res. 71, 95–102.
Uversky, V.N., Gillespie, J.R., and Fink, A.L. (2000). Why are “nativelyunfolded” proteins unstructured under physiologic conditions? Pro-teins 41, 415–427.
Veigel, C., Wang, F., Bartoo, M.L., Sellers, J.R., and Molloy, J.E.(2002). The gated gait of the processive molecular motor, myosinV. Nat. Cell Biol. 4, 59–65.
Warshaw, D.M., Hayes, E., Gaffney, D., Lauzon, A.M., Wu, J., Ken-nedy, G., Trybus, K., Lowey, S., and Berger, C. (1998). Myosin confor-mational states determined by single fluorophore polarization. Proc.Natl. Acad. Sci. USA 95, 8034–8039.
Wells, A.L., Lin, A.W., Chen, L.Q., Safer, D., Cain, S.M., Hasson, T.,Carragher, B.O., Milligan, R.A., and Sweeney, H.L. (1999). MyosinVI is an actin-based motor that moves backwards. Nature 401,505–508.
Winkelmann, D.A., and Lowey, S. (1986). Probing myosin head struc-ture with monoclonal antibodies. J. Mol. Biol. 188, 595–612.
Yildiz, A., Park, H., Safer, D., Yang, Z., Chen, L.Q., Selvin, P.R.,and Sweeney, H.L. (2004). Myosin VI steps via a hand-over-hand

Related Documents











