Developmental Cell Article A Double-Assurance Mechanism Controls Cell Cycle Exit upon Terminal Differentiation in Drosophila Laura A. Buttitta, 1 Alexia J. Katzaroff, 1,2 Carissa L. Perez, 1,2 Aida de la Cruz, 1 and Bruce A. Edgar 1, * 1 Division of Basic Sciences, Fred Hutchinson Cancer Research Center, Seattle, WA 98109, USA 2 Molecular and Cellular Biology Program, University of Washington, Seattle, WA 98195, USA *Correspondence: [email protected] DOI 10.1016/j.devcel.2007.02.020 SUMMARY Terminal differentiation is often coupled with permanent exit from the cell cycle, yet it is un- clear how cell proliferation is blocked in differ- entiated tissues. We examined the process of cell cycle exit in Drosophila wings and eyes and discovered that cell cycle exit can be pre- vented or even reversed in terminally differen- tiating cells by the simultaneous activation of E2F1 and either Cyclin E/Cdk2 or Cyclin D/Cdk4. Enforcing both E2F and Cyclin/Cdk ac- tivities is required to bypass exit because feed- back between E2F and Cyclin E/Cdk2 is in- hibited after cells differentiate, ensuring that cell cycle exit is robust. In some differentiating cell types (e.g., neurons), known inhibitors in- cluding the retinoblastoma homolog Rbf and the p27 homolog Dacapo contribute to parallel repression of E2F and Cyclin E/Cdk2. In other cell types, however (e.g., wing epithelial cells), unknown mechanisms inhibit E2F and Cyclin/ Cdk activity in parallel to enforce permanent cell cycle exit upon terminal differentiation. INTRODUCTION During development, terminal differentiation is coupled with permanent exit from the cell cycle in G1. A loss of co- ordination between these processes can lead to aberrant tissue development and tumorigenesis. Despite the im- portance of this process, it remains unclear how prolif- eration is so potently blocked in differentiated tissues. Current models of cell cycle exit invoke repression of E2F/DP transcription factor complexes, or inhibition of the G1 Cyclin/Cyclin-dependent kinase (Cdk) complexes Cyclin E/Cdk2 and Cyclin D/Cdk4 (reviewed in Zhu and Skoultchi, 2001). Cyclin E/Cdk2 (CycE/Cdk2) kinase activ- ity controls S phase initiation, while E2F/DP transcriptional activity controls the expression of genes required for DNA replication and mitosis, including many of the Cyclins and Cdks (reviewed in Stevaux and Dyson, 2002). E2F/DP complexes act as either transcriptional activators or repressors depending upon the E2F subunit used and whether it is associated with the retinoblastoma tumor suppressor proteins (RBs), which convert E2F/DP com- plexes to repressors (Dyson, 1998). CycD/Cdk4 and CycE/Cdk2 promote E2F activity indirectly by phosphory- lating and inhibiting RBs (reviewed in Du and Pogoriler, 2006), while E2F promotes CycE/Cdk2 activity via tran- scriptional activation (Dimova and Dyson, 2005). Impor- tantly, the potential for positive feedback between E2F and Cyclin/Cdk activities predicts that inhibition of either activity should repress the other and therefore be suffi- cient to arrest the cell cycle in G1. Consistent with this, two groups of negative cell cycle regulators are thought to be important for cell cycle exit: the RB-type proteins and the Cdk inhibitors (CKIs), which inhibit CycE/Cdk2 and CycD/Cdk4 activity (Cobrinik, 2005; Vidal and Koff, 2000). Loss of RBs leads to ectopic cell cycle progression in certain cell types, while CKI mutants exhibit ectopic proliferation in many tissues (re- viewed in Vidal and Koff, 2000; Zhang, 1999). However, determining the roles of RB/E2F-mediated repression or CKI-mediated inhibition of G1 Cyclin/Cdks in cell cycle exit upon terminal differentiation has been difficult, be- cause mammals have several paralogs that partially com- pensate for each other, including 8 E2Fs, 3 RBs, and 7 Cip/Kip- or Ink-type CKIs (Attwooll et al., 2004; Vidal and Koff, 2000). Drosophila has been useful for studies of cell cycle exit, in part because it has fewer paralogs of each cell cycle regulator. Flies lack the Ink-type CKIs, which inhibit CycD/Cdk4, and have a single Cip/Kip-type CKI, Dacapo (Dap), which selectively inhibits CycE/Cdk2 (de Nooij et al., 1996; Lane et al., 1996; Meyer et al., 2000). Dro- sophila contain two E2F subunits (dE2F1 and dE2F2), one DP subunit (dDP), and two RB homologs (Rbf1 and Rbf2) (Dyson, 1998). dE2F1/DP complexes act as activa- tors (referred to as E2F activity) unless associated with Rbf1, which converts them to repressors. In contrast, dE2F2/DP complexes act as repressors and associate with either Rbf1 or Rbf2 (Stevaux et al., 2002; Stevaux and Dyson, 2002). As rbf2 and de2f2 null mutants are viable, Rbf1 and dE2F1 are thought to be the major con- tributors to cell cycle regulation in the somatic tissues of Drosophila (Cayirlioglu et al., 2001; Stevaux et al., 2005). Findings in both Drosophila and mammals suggest that simple models of cell cycle exit invoking RBs or CKIs are insufficient. For example, Dap is not absolutely es- sential for cell cycle exit in Drosophila embryos or eyes Developmental Cell 12, 631–643, April 2007 ª2007 Elsevier Inc. 631

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Developmental Cell
Article
ADouble-AssuranceMechanismControlsCellCycleExit upon Terminal Differentiation in DrosophilaLaura A. Buttitta,1 Alexia J. Katzaroff,1,2 Carissa L. Perez,1,2 Aida de la Cruz,1 and Bruce A. Edgar1,*1 Division of Basic Sciences, Fred Hutchinson Cancer Research Center, Seattle, WA 98109, USA2 Molecular and Cellular Biology Program, University of Washington, Seattle, WA 98195, USA
*Correspondence: [email protected] 10.1016/j.devcel.2007.02.020
SUMMARY
Terminal differentiation is often coupled withpermanent exit from the cell cycle, yet it is un-clear how cell proliferation is blocked in differ-entiated tissues. We examined the process ofcell cycle exit in Drosophila wings and eyesand discovered that cell cycle exit can be pre-vented or even reversed in terminally differen-tiating cells by the simultaneous activation ofE2F1 and either Cyclin E/Cdk2 or CyclinD/Cdk4. Enforcing both E2F and Cyclin/Cdk ac-tivities is required to bypass exit because feed-back between E2F and Cyclin E/Cdk2 is in-hibited after cells differentiate, ensuring thatcell cycle exit is robust. In some differentiatingcell types (e.g., neurons), known inhibitors in-cluding the retinoblastoma homolog Rbf andthe p27 homolog Dacapo contribute to parallelrepression of E2F and Cyclin E/Cdk2. In othercell types, however (e.g., wing epithelial cells),unknown mechanisms inhibit E2F and Cyclin/Cdk activity in parallel to enforce permanentcell cycle exit upon terminal differentiation.
INTRODUCTION
During development, terminal differentiation is coupled
with permanent exit from the cell cycle in G1. A loss of co-
ordination between these processes can lead to aberrant
tissue development and tumorigenesis. Despite the im-
portance of this process, it remains unclear how prolif-
eration is so potently blocked in differentiated tissues.
Current models of cell cycle exit invoke repression of
E2F/DP transcription factor complexes, or inhibition of
the G1 Cyclin/Cyclin-dependent kinase (Cdk) complexes
Cyclin E/Cdk2 and Cyclin D/Cdk4 (reviewed in Zhu and
Skoultchi, 2001). Cyclin E/Cdk2 (CycE/Cdk2) kinase activ-
ity controls S phase initiation, while E2F/DP transcriptional
activity controls the expression of genes required for DNA
replication and mitosis, including many of the Cyclins and
Cdks (reviewed in Stevaux and Dyson, 2002). E2F/DP
complexes act as either transcriptional activators or
repressors depending upon the E2F subunit used and
Develo
whether it is associated with the retinoblastoma tumor
suppressor proteins (RBs), which convert E2F/DP com-
plexes to repressors (Dyson, 1998). CycD/Cdk4 and
CycE/Cdk2 promote E2F activity indirectly by phosphory-
lating and inhibiting RBs (reviewed in Du and Pogoriler,
2006), while E2F promotes CycE/Cdk2 activity via tran-
scriptional activation (Dimova and Dyson, 2005). Impor-
tantly, the potential for positive feedback between E2F
and Cyclin/Cdk activities predicts that inhibition of either
activity should repress the other and therefore be suffi-
cient to arrest the cell cycle in G1.
Consistent with this, two groups of negative cell cycle
regulators are thought to be important for cell cycle exit:
the RB-type proteins and the Cdk inhibitors (CKIs), which
inhibit CycE/Cdk2 and CycD/Cdk4 activity (Cobrinik,
2005; Vidal and Koff, 2000). Loss of RBs leads to ectopic
cell cycle progression in certain cell types, while CKI
mutants exhibit ectopic proliferation in many tissues (re-
viewed in Vidal and Koff, 2000; Zhang, 1999). However,
determining the roles of RB/E2F-mediated repression or
CKI-mediated inhibition of G1 Cyclin/Cdks in cell cycle
exit upon terminal differentiation has been difficult, be-
cause mammals have several paralogs that partially com-
pensate for each other, including 8 E2Fs, 3 RBs, and 7
Cip/Kip- or Ink-type CKIs (Attwooll et al., 2004; Vidal and
Koff, 2000).
Drosophila has been useful for studies of cell cycle exit,
in part because it has fewer paralogs of each cell cycle
regulator. Flies lack the Ink-type CKIs, which inhibit
CycD/Cdk4, and have a single Cip/Kip-type CKI, Dacapo
(Dap), which selectively inhibits CycE/Cdk2 (de Nooij
et al., 1996; Lane et al., 1996; Meyer et al., 2000). Dro-
sophila contain two E2F subunits (dE2F1 and dE2F2),
one DP subunit (dDP), and two RB homologs (Rbf1 and
Rbf2) (Dyson, 1998). dE2F1/DP complexes act as activa-
tors (referred to as E2F activity) unless associated with
Rbf1, which converts them to repressors. In contrast,
dE2F2/DP complexes act as repressors and associate
with either Rbf1 or Rbf2 (Stevaux et al., 2002; Stevaux
and Dyson, 2002). As rbf2 and de2f2 null mutants are
viable, Rbf1 and dE2F1 are thought to be the major con-
tributors to cell cycle regulation in the somatic tissues of
Drosophila (Cayirlioglu et al., 2001; Stevaux et al., 2005).
Findings in both Drosophila and mammals suggest that
simple models of cell cycle exit invoking RBs or CKIs
are insufficient. For example, Dap is not absolutely es-
sential for cell cycle exit in Drosophila embryos or eyes
pmental Cell 12, 631–643, April 2007 ª2007 Elsevier Inc. 631
Developmental Cell
Redundant Mechanisms Ensure Cell Cycle Exit
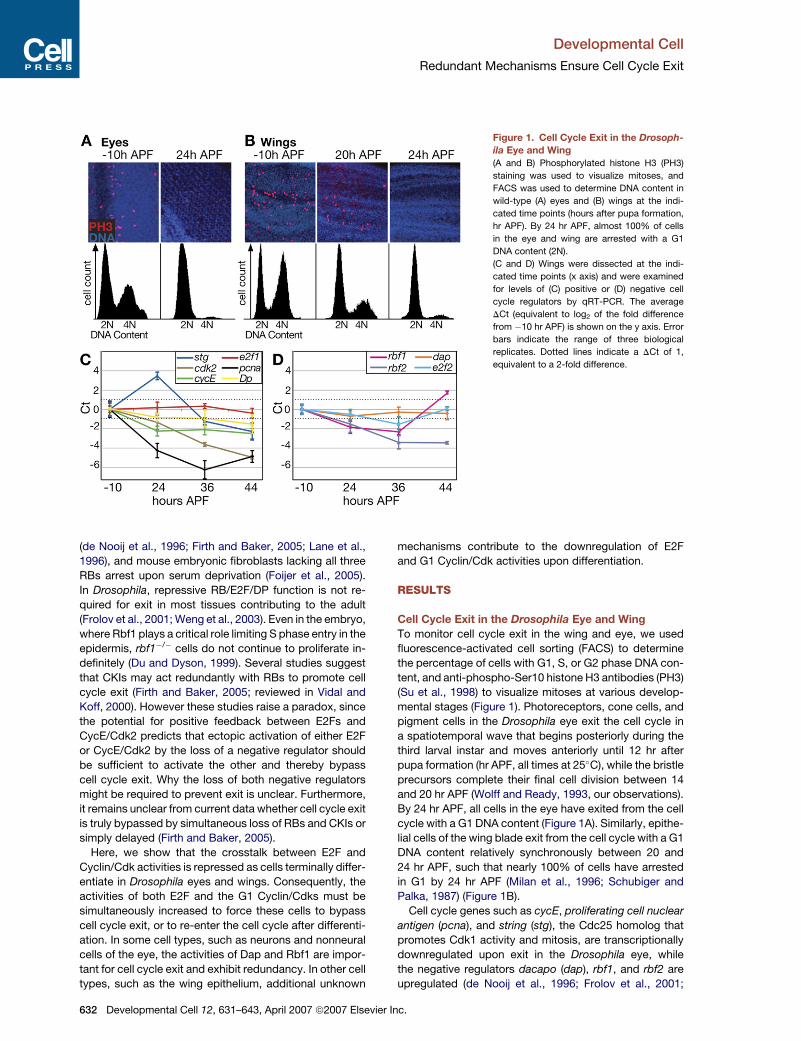
Figure 1. Cell Cycle Exit in the Drosoph-
ila Eye and Wing
(A and B) Phosphorylated histone H3 (PH3)
staining was used to visualize mitoses, and
FACS was used to determine DNA content in
wild-type (A) eyes and (B) wings at the indi-
cated time points (hours after pupa formation,
hr APF). By 24 hr APF, almost 100% of cells
in the eye and wing are arrested with a G1
DNA content (2N).
(C and D) Wings were dissected at the indi-
cated time points (x axis) and were examined
for levels of (C) positive or (D) negative cell
cycle regulators by qRT-PCR. The average
DCt (equivalent to log2 of the fold difference
from �10 hr APF) is shown on the y axis. Error
bars indicate the range of three biological
replicates. Dotted lines indicate a DCt of 1,
equivalent to a 2-fold difference.
(de Nooij et al., 1996; Firth and Baker, 2005; Lane et al.,
1996), and mouse embryonic fibroblasts lacking all three
RBs arrest upon serum deprivation (Foijer et al., 2005).
In Drosophila, repressive RB/E2F/DP function is not re-
quired for exit in most tissues contributing to the adult
(Frolov et al., 2001; Weng et al., 2003). Even in the embryo,
where Rbf1 plays a critical role limiting S phase entry in the
epidermis, rbf1�/� cells do not continue to proliferate in-
definitely (Du and Dyson, 1999). Several studies suggest
that CKIs may act redundantly with RBs to promote cell
cycle exit (Firth and Baker, 2005; reviewed in Vidal and
Koff, 2000). However these studies raise a paradox, since
the potential for positive feedback between E2Fs and
CycE/Cdk2 predicts that ectopic activation of either E2F
or CycE/Cdk2 by the loss of a negative regulator should
be sufficient to activate the other and thereby bypass
cell cycle exit. Why the loss of both negative regulators
might be required to prevent exit is unclear. Furthermore,
it remains unclear from current data whether cell cycle exit
is truly bypassed by simultaneous loss of RBs and CKIs or
simply delayed (Firth and Baker, 2005).
Here, we show that the crosstalk between E2F and
Cyclin/Cdk activities is repressed as cells terminally differ-
entiate in Drosophila eyes and wings. Consequently, the
activities of both E2F and the G1 Cyclin/Cdks must be
simultaneously increased to force these cells to bypass
cell cycle exit, or to re-enter the cell cycle after differenti-
ation. In some cell types, such as neurons and nonneural
cells of the eye, the activities of Dap and Rbf1 are impor-
tant for cell cycle exit and exhibit redundancy. In other cell
types, such as the wing epithelium, additional unknown
632 Developmental Cell 12, 631–643, April 2007 ª2007 Elsevier
mechanisms contribute to the downregulation of E2F
and G1 Cyclin/Cdk activities upon differentiation.
RESULTS
Cell Cycle Exit in the Drosophila Eye and Wing
To monitor cell cycle exit in the wing and eye, we used
fluorescence-activated cell sorting (FACS) to determine
the percentage of cells with G1, S, or G2 phase DNA con-
tent, and anti-phospho-Ser10 histone H3 antibodies (PH3)
(Su et al., 1998) to visualize mitoses at various develop-
mental stages (Figure 1). Photoreceptors, cone cells, and
pigment cells in the Drosophila eye exit the cell cycle in
a spatiotemporal wave that begins posteriorly during the
third larval instar and moves anteriorly until 12 hr after
pupa formation (hr APF, all times at 25�C), while the bristle
precursors complete their final cell division between 14
and 20 hr APF (Wolff and Ready, 1993, our observations).
By 24 hr APF, all cells in the eye have exited from the cell
cycle with a G1 DNA content (Figure 1A). Similarly, epithe-
lial cells of the wing blade exit from the cell cycle with a G1
DNA content relatively synchronously between 20 and
24 hr APF, such that nearly 100% of cells have arrested
in G1 by 24 hr APF (Milan et al., 1996; Schubiger and
Palka, 1987) (Figure 1B).
Cell cycle genes such as cycE, proliferating cell nuclear
antigen (pcna), and string (stg), the Cdc25 homolog that
promotes Cdk1 activity and mitosis, are transcriptionally
downregulated upon exit in the Drosophila eye, while
the negative regulators dacapo (dap), rbf1, and rbf2 are
upregulated (de Nooij et al., 1996; Frolov et al., 2001;
Inc.

Developmental Cell
Redundant Mechanisms Ensure Cell Cycle Exit
Keller et al., 2005; Kylsten and Saint, 1997; Richardson
et al., 1995). To determine whether similar changes in
cell cycle gene expression occur in the wing, we examined
mRNA expression in wild-type wings by quantitative
RT-PCR (qRT-PCR) at time points before and after exit.
We compared gene expression at three time points during
and after exit (24, 36, and 44 hr APF) to a proliferative
stage (�10 hr APF). As reported for the eye, we observed
repression of cycE, cdk2, and the well-characterized
target of E2F1, pcna, in the differentiating wings (Fig-
ure 1C). However, levels of stg pulsed at 24 hr APF before
dropping at 44 hr APF. We did not observe significant
changes in e2f1 or Dp levels at any time points assayed.
In contrast to the eye, we did not see strong upregulation
of negative cell cycle regulators upon cell cycle exit in
the wing (Figure 1D), or at two time points prior to exit
(Figure S9; see the Supplemental Data available with this
article online). Levels of dap and e2f2 remained relatively
constant after exit, but dap levels peaked with a 2.4-fold
increase before exit at 18 hr APF (Figure S9). We observed
an increase in rbf1 specifically at 44 hr APF, but rbf1 levels
at 24 APF, when cells establish G1 arrest, were low. rbf2
levels did not rise, but actually decreased after prolonged
exit. These data suggest that cell cycle exit in the wing at
24 hr APF is not due to transcriptional induction of known
negative cell cycle regulators. Moreover, the distinct tran-
scriptional profiles exhibited by stg, cycE, pcna, and dap,
all of which have been implicated as E2F targets (Dimova
et al., 2003; Duronio et al., 1996; Neufeld et al., 1998; Reis
and Edgar, 2004), suggest that E2F-independent mecha-
nisms control the temporal expression of some of these
genes during differentiation.
Ectopic E2F or Cyclin/Cdk Activity Delays, but Does
Not Prevent, Cell Cycle Exit
Increased E2F or Cyclin/Cdk activity can delay cell cycle
exit of some cells in the eye and wing (Asano et al.,
1996; Datar et al., 2000; Du et al., 1996b; Milan et al.,
1996; Richardson et al., 1995), but the duration of this
effect is unknown. To characterize this effect more pre-
cisely, we used the heat shock (hs)-FLP tub>Gal4/UAS
system in combination with a temperature-sensitive tub-
Gal80 (Gal80TS) to conditionally express cell cycle regula-
tors in cell clones in differentiating pupal eyes and wings.
Using temperature shifts to confine gene expression to the
pupal stages, we were able to circumvent the deleterious
effects of overproliferation and compensatory apoptosis
earlier in development.
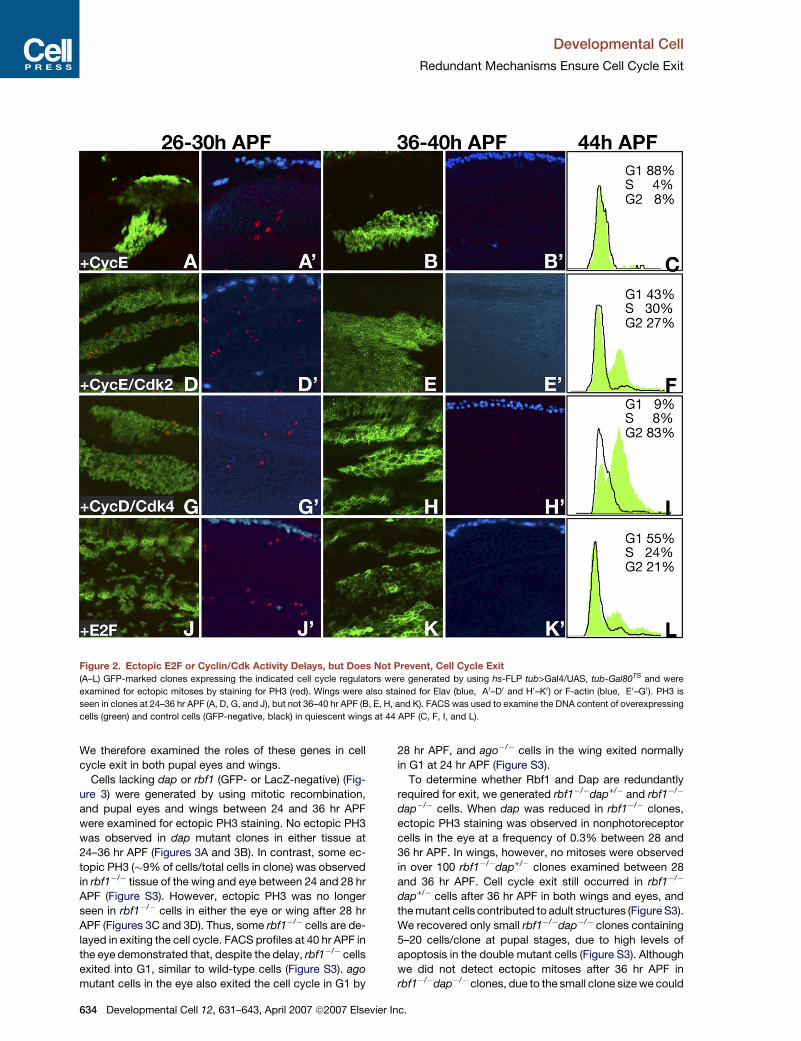
GFP-marked cell clones capable of overexpressing
Drosophila CycE, CycE+Cdk2, CycD+Cdk4, E2F1+DP
(hereafter referred to as E2F), or other cell cycle regulators
(Table S1) were generated early in development (48–72 hr
after embryo deposition), with Gal4 activity silenced by
Gal80TS. Gal4-mediated gene expression was induced
in the clones at 0 hr APF by inactivation of Gal80TS at
29�C, and effects on cycle exit were examined at later
stages by scoring ectopic mitoses by using the PH3 anti-
body. For each genotype noted above, ectopic PH3 was
evident in GFP-positive clones at 24–36 hr APF in both
Develo
the wing blade (26–30 hr APF shown) (Figures 2A, 2D,
2G, and 2J) and eye (Figure S1), whereas neighboring
GFP-negative cells, or clones expressing Gal4 and GFP
alone (not shown), were quiescent. By 36–40 hr APF how-
ever, no further ectopic mitoses were detected in any of
these genotypes (Figures 2B, 2E, 2H, and 2K; Figure S1).
Antibody and reagent penetrance was confirmed in late-
stage wings and eyes by staining for the neural marker
Elav (blue, Figures 2A–2D0 and 2H–2K0) or actin (blue, Fig-
ures 2E–2G0). We also confirmed that Gal4-mediated gene
expression continued in clones at late stages (Figure S2).
We conclude that ectopic Cyclin/Cdk or E2F activity is
sufficient to delay cell cycle exit until 36 hr APF in the
eye and wing, but that it does not eliminate exit.
To determine how many extra cell cycles the overex-
pressing cells went through, we took advantage of the
relatively synchronous exit in the wing to perform a clonal
analysis of the cell divisions. Nonoverlapping, GFP-
marked clones expressing cell cycle regulators were in-
duced at 0 hr APF by using the hs-FLP act>Gal4/UAS sys-
tem. Wings were fixed at 40 hr APF, when mitoses were
no longer evident, and cells per clone were counted blind
for at least 100 clones in the wing blade. Clonal cell counts
of control cells averaged 2.3 ± 0.5 cells/clone, while clones
expressing CycE+Stg, CycD+Cdk4, or E2F averaged
4.0 ± 0.6, 3.9 ± 1.0, and 4.2 ± 0.4 cells/clone, respectively.
This indicates that the observed delay in exit leads to one
extra cell cycle in the wing before additional mechanisms
override high Cyclin/Cdk or E2F activity.
We next used FACS to determine the DNA content of
cells overexpressing these cell cycle regulators (green)
and control cells (GFP-negative, black) in quiescent wings
(Figures 2C, 2F, 2I, and 2L) and eyes (Figure S1) at 44 hr
APF. Cells expressing CycE exited the cell cycle with a
G1 DNA content at this later stage (Figure 2C), while cells
expressing both CycE and Cdk2 arrested in G1, S, or G2
(Figure 2F). Many cells expressing CycD/Cdk4 also ar-
rested with a G2 DNA content (Figure 2I). E2F overexpres-
sion had a milder effect, with a smaller percentage of cells
arresting in S and G2 phases. (Figure 2L). While we cannot
rule out the possibility that some cells continue slowly
through S phase (lasting >8 hr), the arrest or prolonged
S phase of some cells suggests that factors essential for
DNA replication are not fully activated despite sustained
E2F or Cyclin/Cdk activity. Likewise, the arrest of some
cells in G2 indicates that factors critical for mitosis are
not maintained by sustained E2F or G1 Cyclin/Cdk activity
alone.
Requirements for Negative Cell Cycle
Regulators Are Tissue Specific
Negative regulators of E2F and CycE/Cdk2 have been
shown to be important for cell cycle exit in the Drosophila
eye. These include Rbf1 in combination with Dap (Firth
and Baker, 2005), and the F-box protein Archipelago,
which mediates CycE degradation (Ago/Fbw7) (Moberg
et al., 2001). Yet, it was unknown how many extra cell
cycles occur in cells lacking these regulators, or whether
they are important for exit in tissues other than the eye.
pmental Cell 12, 631–643, April 2007 ª2007 Elsevier Inc. 633
Developmental Cell
Redundant Mechanisms Ensure Cell Cycle Exit
Figure 2. Ectopic E2F or Cyclin/Cdk Activity Delays, but Does Not Prevent, Cell Cycle Exit
(A–L) GFP-marked clones expressing the indicated cell cycle regulators were generated by using hs-FLP tub>Gal4/UAS, tub-Gal80TS and were
examined for ectopic mitoses by staining for PH3 (red). Wings were also stained for Elav (blue, A0–D0 and H0–K0) or F-actin (blue, E0–G0). PH3 is
seen in clones at 24–36 hr APF (A, D, G, and J), but not 36–40 hr APF (B, E, H, and K). FACS was used to examine the DNA content of overexpressing
cells (green) and control cells (GFP-negative, black) in quiescent wings at 44 APF (C, F, I, and L).
We therefore examined the roles of these genes in cell
cycle exit in both pupal eyes and wings.
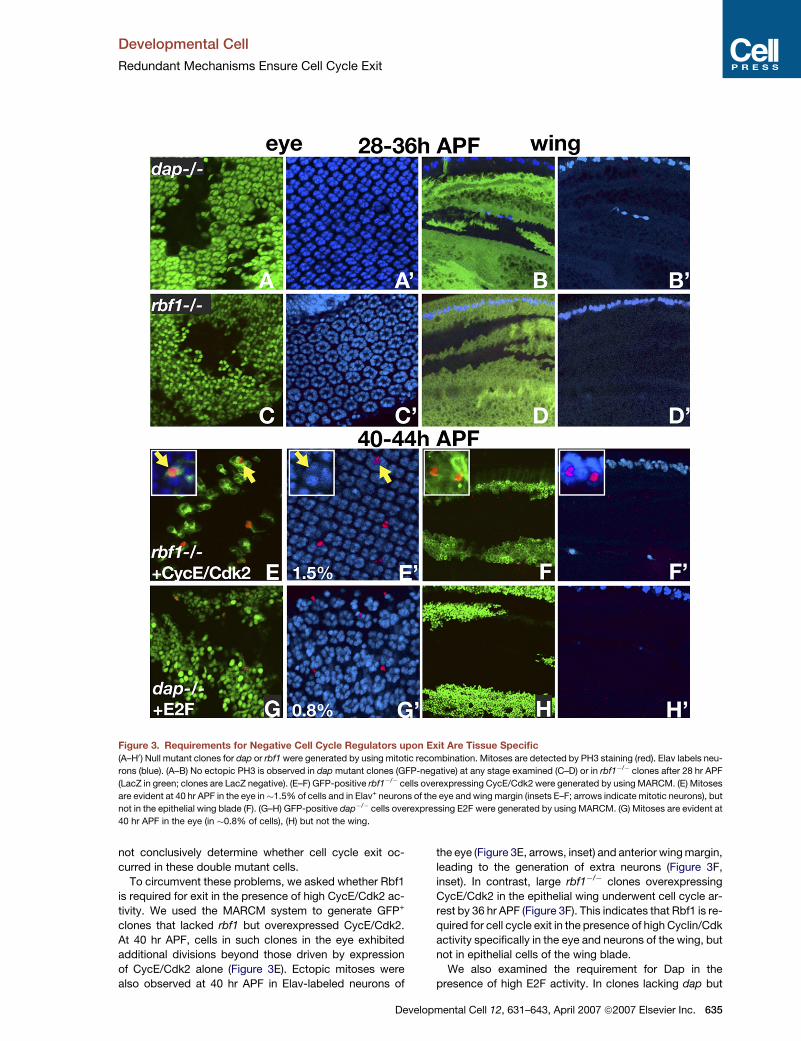
Cells lacking dap or rbf1 (GFP- or LacZ-negative) (Fig-
ure 3) were generated by using mitotic recombination,
and pupal eyes and wings between 24 and 36 hr APF
were examined for ectopic PH3 staining. No ectopic PH3
was observed in dap mutant clones in either tissue at
24–36 hr APF (Figures 3A and 3B). In contrast, some ec-
topic PH3 (�9% of cells/total cells in clone) was observed
in rbf1�/� tissue of the wing and eye between 24 and 28 hr
APF (Figure S3). However, ectopic PH3 was no longer
seen in rbf1�/� cells in either the eye or wing after 28 hr
APF (Figures 3C and 3D). Thus, some rbf1�/� cells are de-
layed in exiting the cell cycle. FACS profiles at 40 hr APF in
the eye demonstrated that, despite the delay, rbf1�/� cells
exited into G1, similar to wild-type cells (Figure S3). ago
mutant cells in the eye also exited the cell cycle in G1 by
634 Developmental Cell 12, 631–643, April 2007 ª2007 Elsevier
28 hr APF, and ago�/� cells in the wing exited normally
in G1 at 24 hr APF (Figure S3).
To determine whether Rbf1 and Dap are redundantly
required for exit, we generated rbf1�/�dap+/� and rbf1�/�
dap�/� cells. When dap was reduced in rbf1�/� clones,
ectopic PH3 staining was observed in nonphotoreceptor
cells in the eye at a frequency of 0.3% between 28 and
36 hr APF. In wings, however, no mitoses were observed
in over 100 rbf1�/�dap+/� clones examined between 28
and 36 hr APF. Cell cycle exit still occurred in rbf1�/�
dap+/� cells after 36 hr APF in both wings and eyes, and
the mutant cells contributed to adult structures (Figure S3).
We recovered only small rbf1�/�dap�/� clones containing
5–20 cells/clone at pupal stages, due to high levels of
apoptosis in the double mutant cells (Figure S3). Although
we did not detect ectopic mitoses after 36 hr APF in
rbf1�/�dap�/� clones, due to the small clone size we could
Inc.
Developmental Cell
Redundant Mechanisms Ensure Cell Cycle Exit
Figure 3. Requirements for Negative Cell Cycle Regulators upon Exit Are Tissue Specific
(A–H0) Null mutant clones for dap or rbf1 were generated by using mitotic recombination. Mitoses are detected by PH3 staining (red). Elav labels neu-
rons (blue). (A–B) No ectopic PH3 is observed in dap mutant clones (GFP-negative) at any stage examined (C–D) or in rbf1�/� clones after 28 hr APF
(LacZ in green; clones are LacZ negative). (E–F) GFP-positive rbf1�/� cells overexpressing CycE/Cdk2 were generated by using MARCM. (E) Mitoses
are evident at 40 hr APF in the eye in�1.5% of cells and in Elav+ neurons of the eye and wing margin (insets E–F; arrows indicate mitotic neurons), but
not in the epithelial wing blade (F). (G–H) GFP-positive dap�/� cells overexpressing E2F were generated by using MARCM. (G) Mitoses are evident at
40 hr APF in the eye (in �0.8% of cells), (H) but not the wing.
not conclusively determine whether cell cycle exit oc-
curred in these double mutant cells.
To circumvent these problems, we asked whether Rbf1
is required for exit in the presence of high CycE/Cdk2 ac-
tivity. We used the MARCM system to generate GFP+
clones that lacked rbf1 but overexpressed CycE/Cdk2.
At 40 hr APF, cells in such clones in the eye exhibited
additional divisions beyond those driven by expression
of CycE/Cdk2 alone (Figure 3E). Ectopic mitoses were
also observed at 40 hr APF in Elav-labeled neurons of
Develo
the eye (Figure 3E, arrows, inset) and anterior wing margin,
leading to the generation of extra neurons (Figure 3F,
inset). In contrast, large rbf1�/� clones overexpressing
CycE/Cdk2 in the epithelial wing underwent cell cycle ar-
rest by 36 hr APF (Figure 3F). This indicates that Rbf1 is re-
quired for cell cycle exit in the presence of high Cyclin/Cdk
activity specifically in the eye and neurons of the wing, but
not in epithelial cells of the wing blade.
We also examined the requirement for Dap in the
presence of high E2F activity. In clones lacking dap but
pmental Cell 12, 631–643, April 2007 ª2007 Elsevier Inc. 635
Developmental Cell
Redundant Mechanisms Ensure Cell Cycle Exit
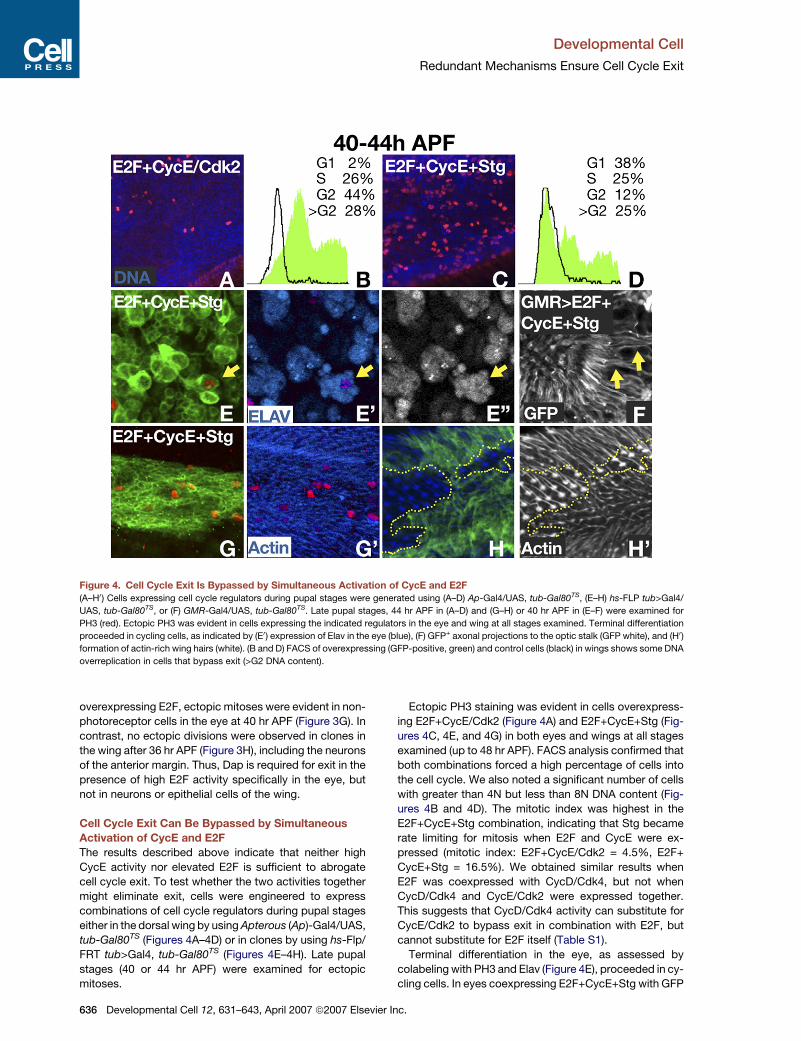
Figure 4. Cell Cycle Exit Is Bypassed by Simultaneous Activation of CycE and E2F
(A–H0) Cells expressing cell cycle regulators during pupal stages were generated using (A–D) Ap-Gal4/UAS, tub-Gal80TS, (E–H) hs-FLP tub>Gal4/
UAS, tub-Gal80TS, or (F) GMR-Gal4/UAS, tub-Gal80TS. Late pupal stages, 44 hr APF in (A–D) and (G–H) or 40 hr APF in (E–F) were examined for
PH3 (red). Ectopic PH3 was evident in cells expressing the indicated regulators in the eye and wing at all stages examined. Terminal differentiation
proceeded in cycling cells, as indicated by (E0) expression of Elav in the eye (blue), (F) GFP+ axonal projections to the optic stalk (GFP white), and (H0)
formation of actin-rich wing hairs (white). (B and D) FACS of overexpressing (GFP-positive, green) and control cells (black) in wings shows some DNA
overreplication in cells that bypass exit (>G2 DNA content).
overexpressing E2F, ectopic mitoses were evident in non-
photoreceptor cells in the eye at 40 hr APF (Figure 3G). In
contrast, no ectopic divisions were observed in clones in
the wing after 36 hr APF (Figure 3H), including the neurons
of the anterior margin. Thus, Dap is required for exit in the
presence of high E2F activity specifically in the eye, but
not in neurons or epithelial cells of the wing.
Cell Cycle Exit Can Be Bypassed by Simultaneous
Activation of CycE and E2F
The results described above indicate that neither high
CycE activity nor elevated E2F is sufficient to abrogate
cell cycle exit. To test whether the two activities together
might eliminate exit, cells were engineered to express
combinations of cell cycle regulators during pupal stages
either in the dorsal wing by using Apterous (Ap)-Gal4/UAS,
tub-Gal80TS (Figures 4A–4D) or in clones by using hs-Flp/
FRT tub>Gal4, tub-Gal80TS (Figures 4E–4H). Late pupal
stages (40 or 44 hr APF) were examined for ectopic
mitoses.
636 Developmental Cell 12, 631–643, April 2007 ª2007 Elsevie
Ectopic PH3 staining was evident in cells overexpress-
ing E2F+CycE/Cdk2 (Figure 4A) and E2F+CycE+Stg (Fig-
ures 4C, 4E, and 4G) in both eyes and wings at all stages
examined (up to 48 hr APF). FACS analysis confirmed that
both combinations forced a high percentage of cells into
the cell cycle. We also noted a significant number of cells
with greater than 4N but less than 8N DNA content (Fig-
ures 4B and 4D). The mitotic index was highest in the
E2F+CycE+Stg combination, indicating that Stg became
rate limiting for mitosis when E2F and CycE were ex-
pressed (mitotic index: E2F+CycE/Cdk2 = 4.5%, E2F+
CycE+Stg = 16.5%). We obtained similar results when
E2F was coexpressed with CycD/Cdk4, but not when
CycD/Cdk4 and CycE/Cdk2 were expressed together.
This suggests that CycD/Cdk4 activity can substitute for
CycE/Cdk2 to bypass exit in combination with E2F, but
cannot substitute for E2F itself (Table S1).
Terminal differentiation in the eye, as assessed by
colabeling with PH3 and Elav (Figure 4E), proceeded in cy-
cling cells. In eyes coexpressing E2F+CycE+Stg with GFP
r Inc.
Developmental Cell
Redundant Mechanisms Ensure Cell Cycle Exit
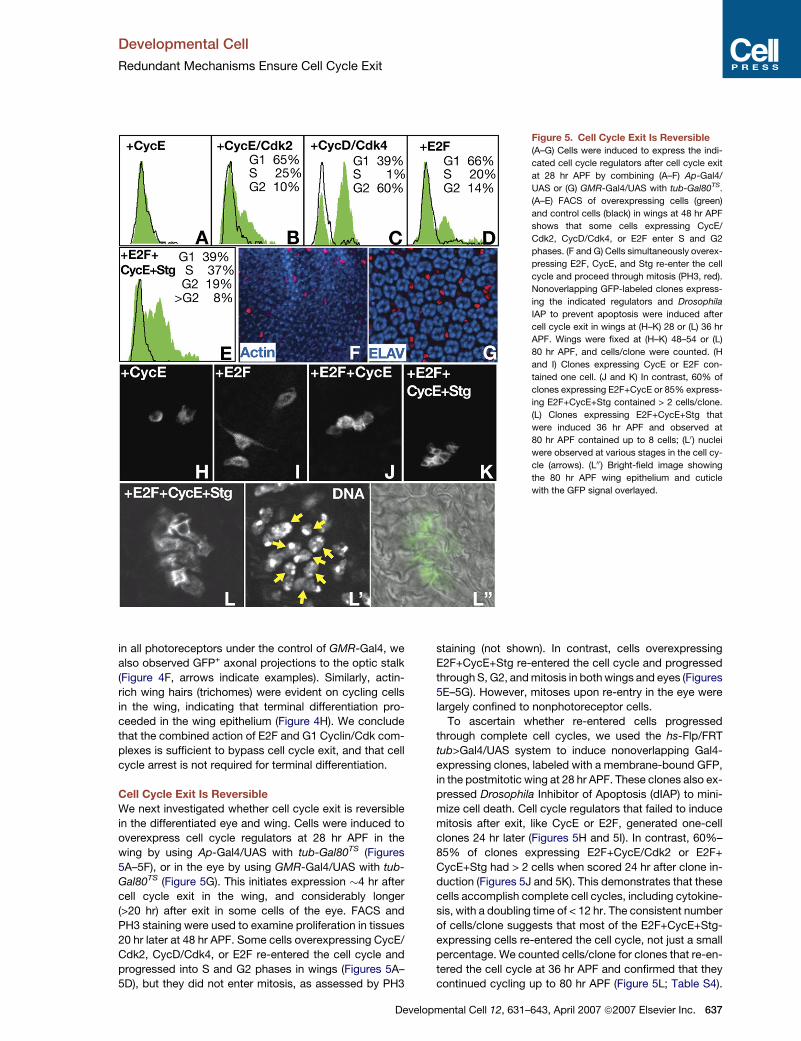
Figure 5. Cell Cycle Exit Is Reversible
(A–G) Cells were induced to express the indi-
cated cell cycle regulators after cell cycle exit
at 28 hr APF by combining (A–F) Ap-Gal4/
UAS or (G) GMR-Gal4/UAS with tub-Gal80TS.
(A–E) FACS of overexpressing cells (green)
and control cells (black) in wings at 48 hr APF
shows that some cells expressing CycE/
Cdk2, CycD/Cdk4, or E2F enter S and G2
phases. (F and G) Cells simultaneously overex-
pressing E2F, CycE, and Stg re-enter the cell
cycle and proceed through mitosis (PH3, red).
Nonoverlapping GFP-labeled clones express-
ing the indicated regulators and Drosophila
IAP to prevent apoptosis were induced after
cell cycle exit in wings at (H–K) 28 or (L) 36 hr
APF. Wings were fixed at (H–K) 48–54 or (L)
80 hr APF, and cells/clone were counted. (H
and I) Clones expressing CycE or E2F con-
tained one cell. (J and K) In contrast, 60% of
clones expressing E2F+CycE or 85% express-
ing E2F+CycE+Stg contained > 2 cells/clone.
(L) Clones expressing E2F+CycE+Stg that
were induced 36 hr APF and observed at
80 hr APF contained up to 8 cells; (L0) nuclei
were observed at various stages in the cell cy-
cle (arrows). (L00) Bright-field image showing
the 80 hr APF wing epithelium and cuticle
with the GFP signal overlayed.
in all photoreceptors under the control of GMR-Gal4, we
also observed GFP+ axonal projections to the optic stalk
(Figure 4F, arrows indicate examples). Similarly, actin-
rich wing hairs (trichomes) were evident on cycling cells
in the wing, indicating that terminal differentiation pro-
ceeded in the wing epithelium (Figure 4H). We conclude
that the combined action of E2F and G1 Cyclin/Cdk com-
plexes is sufficient to bypass cell cycle exit, and that cell
cycle arrest is not required for terminal differentiation.
Cell Cycle Exit Is Reversible
We next investigated whether cell cycle exit is reversible
in the differentiated eye and wing. Cells were induced to
overexpress cell cycle regulators at 28 hr APF in the
wing by using Ap-Gal4/UAS with tub-Gal80TS (Figures
5A–5F), or in the eye by using GMR-Gal4/UAS with tub-
Gal80TS (Figure 5G). This initiates expression �4 hr after
cell cycle exit in the wing, and considerably longer
(>20 hr) after exit in some cells of the eye. FACS and
PH3 staining were used to examine proliferation in tissues
20 hr later at 48 hr APF. Some cells overexpressing CycE/
Cdk2, CycD/Cdk4, or E2F re-entered the cell cycle and
progressed into S and G2 phases in wings (Figures 5A–
5D), but they did not enter mitosis, as assessed by PH3
Deve
staining (not shown). In contrast, cells overexpressing
E2F+CycE+Stg re-entered the cell cycle and progressed
through S, G2, and mitosis in both wings and eyes (Figures
5E–5G). However, mitoses upon re-entry in the eye were
largely confined to nonphotoreceptor cells.
To ascertain whether re-entered cells progressed
through complete cell cycles, we used the hs-Flp/FRT
tub>Gal4/UAS system to induce nonoverlapping Gal4-
expressing clones, labeled with a membrane-bound GFP,
in the postmitotic wing at 28 hr APF. These clones also ex-
pressed Drosophila Inhibitor of Apoptosis (dIAP) to mini-
mize cell death. Cell cycle regulators that failed to induce
mitosis after exit, like CycE or E2F, generated one-cell
clones 24 hr later (Figures 5H and 5I). In contrast, 60%–
85% of clones expressing E2F+CycE/Cdk2 or E2F+
CycE+Stg had > 2 cells when scored 24 hr after clone in-
duction (Figures 5J and 5K). This demonstrates that these
cells accomplish complete cell cycles, including cytokine-
sis, with a doubling time of < 12 hr. The consistent number
of cells/clone suggests that most of the E2F+CycE+Stg-
expressing cells re-entered the cell cycle, not just a small
percentage. We counted cells/clone for clones that re-en-
tered the cell cycle at 36 hr APF and confirmed that they
continued cycling up to 80 hr APF (Figure 5L; Table S4).
lopmental Cell 12, 631–643, April 2007 ª2007 Elsevier Inc. 637
Developmental Cell
Redundant Mechanisms Ensure Cell Cycle Exit
We also obtained re-entry with E2F+CycD/Cdk4 overex-
pression, but not with CycD/Cdk4+CycE/Cdk2 (Table S1).
As in our previous experiments, this demonstrates that
CycD/Cdk4 activity can substitute for CycE/Cdk2 after
exit, but cannot activate endogenous E2F to levels suffi-
cient to drive the cell cycle.
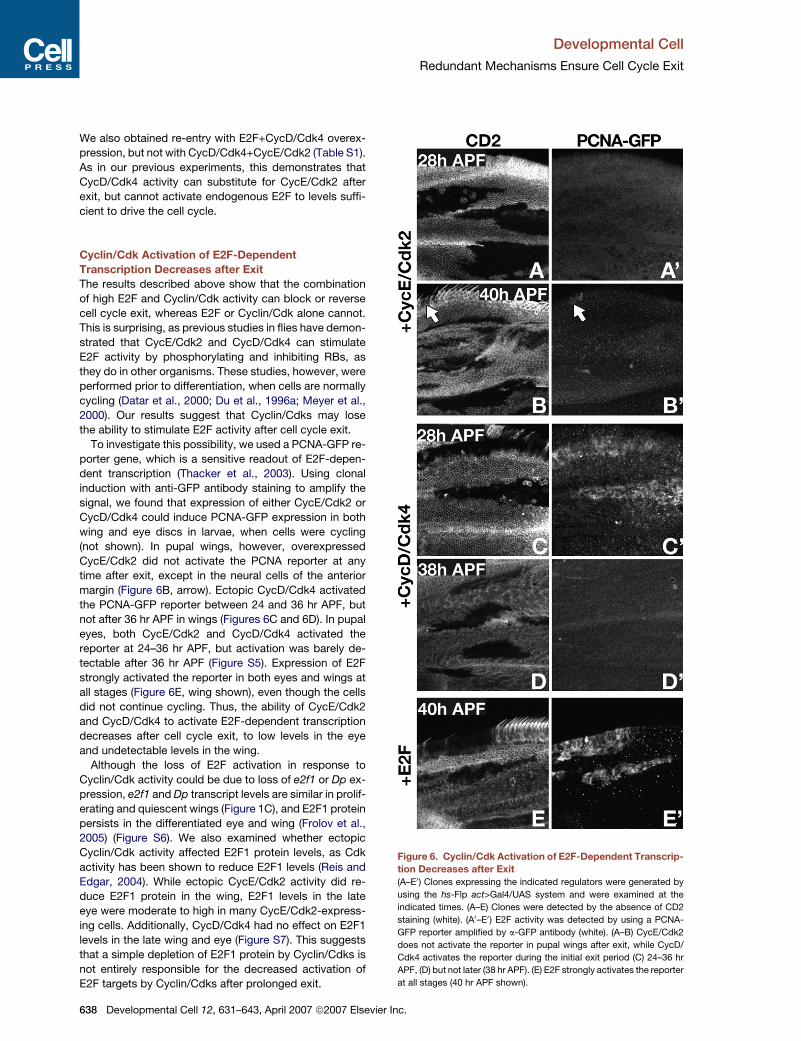
Cyclin/Cdk Activation of E2F-Dependent
Transcription Decreases after Exit
The results described above show that the combination
of high E2F and Cyclin/Cdk activity can block or reverse
cell cycle exit, whereas E2F or Cyclin/Cdk alone cannot.
This is surprising, as previous studies in flies have demon-
strated that CycE/Cdk2 and CycD/Cdk4 can stimulate
E2F activity by phosphorylating and inhibiting RBs, as
they do in other organisms. These studies, however, were
performed prior to differentiation, when cells are normally
cycling (Datar et al., 2000; Du et al., 1996a; Meyer et al.,
2000). Our results suggest that Cyclin/Cdks may lose
the ability to stimulate E2F activity after cell cycle exit.
To investigate this possibility, we used a PCNA-GFP re-
porter gene, which is a sensitive readout of E2F-depen-
dent transcription (Thacker et al., 2003). Using clonal
induction with anti-GFP antibody staining to amplify the
signal, we found that expression of either CycE/Cdk2 or
CycD/Cdk4 could induce PCNA-GFP expression in both
wing and eye discs in larvae, when cells were cycling
(not shown). In pupal wings, however, overexpressed
CycE/Cdk2 did not activate the PCNA reporter at any
time after exit, except in the neural cells of the anterior
margin (Figure 6B, arrow). Ectopic CycD/Cdk4 activated
the PCNA-GFP reporter between 24 and 36 hr APF, but
not after 36 hr APF in wings (Figures 6C and 6D). In pupal
eyes, both CycE/Cdk2 and CycD/Cdk4 activated the
reporter at 24–36 hr APF, but activation was barely de-
tectable after 36 hr APF (Figure S5). Expression of E2F
strongly activated the reporter in both eyes and wings at
all stages (Figure 6E, wing shown), even though the cells
did not continue cycling. Thus, the ability of CycE/Cdk2
and CycD/Cdk4 to activate E2F-dependent transcription
decreases after cell cycle exit, to low levels in the eye
and undetectable levels in the wing.
Although the loss of E2F activation in response to
Cyclin/Cdk activity could be due to loss of e2f1 or Dp ex-
pression, e2f1 and Dp transcript levels are similar in prolif-
erating and quiescent wings (Figure 1C), and E2F1 protein
persists in the differentiated eye and wing (Frolov et al.,
2005) (Figure S6). We also examined whether ectopic
Cyclin/Cdk activity affected E2F1 protein levels, as Cdk
activity has been shown to reduce E2F1 levels (Reis and
Edgar, 2004). While ectopic CycE/Cdk2 activity did re-
duce E2F1 protein in the wing, E2F1 levels in the late
eye were moderate to high in many CycE/Cdk2-express-
ing cells. Additionally, CycD/Cdk4 had no effect on E2F1
levels in the late wing and eye (Figure S7). This suggests
that a simple depletion of E2F1 protein by Cyclin/Cdks is
not entirely responsible for the decreased activation of
E2F targets by Cyclin/Cdks after prolonged exit.
638 Developmental Cell 12, 631–643, April 2007 ª2007 Elsevie
Figure 6. Cyclin/Cdk Activation of E2F-Dependent Transcrip-
tion Decreases after Exit
(A–E0) Clones expressing the indicated regulators were generated by
using the hs-Flp act>Gal4/UAS system and were examined at the
indicated times. (A–E) Clones were detected by the absence of CD2
staining (white). (A0–E0) E2F activity was detected by using a PCNA-
GFP reporter amplified by a-GFP antibody (white). (A–B) CycE/Cdk2
does not activate the reporter in pupal wings after exit, while CycD/
Cdk4 activates the reporter during the initial exit period (C) 24–36 hr
APF, (D) but not later (38 hr APF). (E) E2F strongly activates the reporter
at all stages (40 hr APF shown).
r Inc.
Developmental Cell
Redundant Mechanisms Ensure Cell Cycle Exit
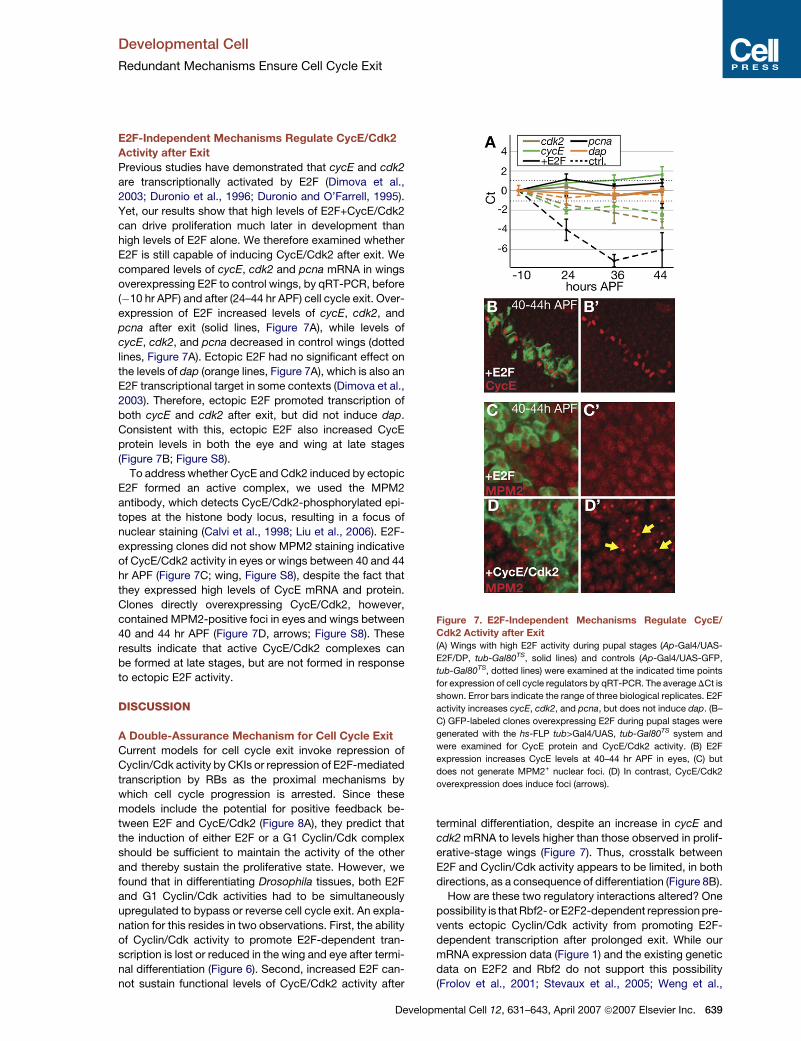
E2F-Independent Mechanisms Regulate CycE/Cdk2
Activity after Exit
Previous studies have demonstrated that cycE and cdk2
are transcriptionally activated by E2F (Dimova et al.,
2003; Duronio et al., 1996; Duronio and O’Farrell, 1995).
Yet, our results show that high levels of E2F+CycE/Cdk2
can drive proliferation much later in development than
high levels of E2F alone. We therefore examined whether
E2F is still capable of inducing CycE/Cdk2 after exit. We
compared levels of cycE, cdk2 and pcna mRNA in wings
overexpressing E2F to control wings, by qRT-PCR, before
(�10 hr APF) and after (24–44 hr APF) cell cycle exit. Over-
expression of E2F increased levels of cycE, cdk2, and
pcna after exit (solid lines, Figure 7A), while levels of
cycE, cdk2, and pcna decreased in control wings (dotted
lines, Figure 7A). Ectopic E2F had no significant effect on
the levels of dap (orange lines, Figure 7A), which is also an
E2F transcriptional target in some contexts (Dimova et al.,
2003). Therefore, ectopic E2F promoted transcription of
both cycE and cdk2 after exit, but did not induce dap.
Consistent with this, ectopic E2F also increased CycE
protein levels in both the eye and wing at late stages
(Figure 7B; Figure S8).
To address whether CycE and Cdk2 induced by ectopic
E2F formed an active complex, we used the MPM2
antibody, which detects CycE/Cdk2-phosphorylated epi-
topes at the histone body locus, resulting in a focus of
nuclear staining (Calvi et al., 1998; Liu et al., 2006). E2F-
expressing clones did not show MPM2 staining indicative
of CycE/Cdk2 activity in eyes or wings between 40 and 44
hr APF (Figure 7C; wing, Figure S8), despite the fact that
they expressed high levels of CycE mRNA and protein.
Clones directly overexpressing CycE/Cdk2, however,
contained MPM2-positive foci in eyes and wings between
40 and 44 hr APF (Figure 7D, arrows; Figure S8). These
results indicate that active CycE/Cdk2 complexes can
be formed at late stages, but are not formed in response
to ectopic E2F activity.
DISCUSSION
A Double-Assurance Mechanism for Cell Cycle Exit
Current models for cell cycle exit invoke repression of
Cyclin/Cdk activity by CKIs or repression of E2F-mediated
transcription by RBs as the proximal mechanisms by
which cell cycle progression is arrested. Since these
models include the potential for positive feedback be-
tween E2F and CycE/Cdk2 (Figure 8A), they predict that
the induction of either E2F or a G1 Cyclin/Cdk complex
should be sufficient to maintain the activity of the other
and thereby sustain the proliferative state. However, we
found that in differentiating Drosophila tissues, both E2F
and G1 Cyclin/Cdk activities had to be simultaneously
upregulated to bypass or reverse cell cycle exit. An expla-
nation for this resides in two observations. First, the ability
of Cyclin/Cdk activity to promote E2F-dependent tran-
scription is lost or reduced in the wing and eye after termi-
nal differentiation (Figure 6). Second, increased E2F can-
not sustain functional levels of CycE/Cdk2 activity after
Develo
terminal differentiation, despite an increase in cycE and
cdk2 mRNA to levels higher than those observed in prolif-
erative-stage wings (Figure 7). Thus, crosstalk between
E2F and Cyclin/Cdk activity appears to be limited, in both
directions, as a consequence of differentiation (Figure 8B).
How are these two regulatory interactions altered? One
possibility is that Rbf2- or E2F2-dependent repression pre-
vents ectopic Cyclin/Cdk activity from promoting E2F-
dependent transcription after prolonged exit. While our
mRNA expression data (Figure 1) and the existing genetic
data on E2F2 and Rbf2 do not support this possibility
(Frolov et al., 2001; Stevaux et al., 2005; Weng et al.,
Figure 7. E2F-Independent Mechanisms Regulate CycE/
Cdk2 Activity after Exit(A) Wings with high E2F activity during pupal stages (Ap-Gal4/UAS-
E2F/DP, tub-Gal80TS, solid lines) and controls (Ap-Gal4/UAS-GFP,
tub-Gal80TS, dotted lines) were examined at the indicated time points
for expression of cell cycle regulators by qRT-PCR. The average DCt is
shown. Error bars indicate the range of three biological replicates. E2F
activity increases cycE, cdk2, and pcna, but does not induce dap. (B–
C) GFP-labeled clones overexpressing E2F during pupal stages were
generated with the hs-FLP tub>Gal4/UAS, tub-Gal80TS system and
were examined for CycE protein and CycE/Cdk2 activity. (B) E2F
expression increases CycE levels at 40–44 hr APF in eyes, (C) but
does not generate MPM2+ nuclear foci. (D) In contrast, CycE/Cdk2
overexpression does induce foci (arrows).
pmental Cell 12, 631–643, April 2007 ª2007 Elsevier Inc. 639

Developmental Cell
Redundant Mechanisms Ensure Cell Cycle Exit
2003), we have not yet tested the roles of Rbf2 or E2F2
in the presence of continued Cyclin/Cdk activity. There-
fore, transcriptional repression of E2F targets by Rbf2 or
E2F2 remains an important issue to address in future
experiments.
More enigmatic is the inability of the ectopic CycE/Cdk2
provided by overexpressed E2F to promote cell cycle pro-
gression (Figure 7). One plausible explanation for this is
that novel inhibitors of CycE are expressed with the onset
of differentiation, and that these raise the threshold of
Cyclin/Cdk activity required to promote cell cycle progres-
sion. Such inhibitors might make the critical substrates of
CycE/Cdk2, which reside on chromatin in DNA-replication
and -transcription initiation complexes, less accessible or
otherwise recalcitrant to activation. The notion of an in-
creased Cdk threshold is consistent with our observation
that the >10-fold increase in CycE/Cdk2 provided by
direct overexpression of the kinase bypassed cell cycle
exit in conjunction with E2F, while the �4-fold increase
provided indirectly by ectopic E2F was insufficient to drive
the cell cycle (Figure 7A; Figure S9). Although a >10-fold
increase in Cdk activity as applied in our experiments is
far above the normal physiological range, (Figure S9),
such dramatic deregulation of cell cycle genes may be
Figure 8. A Double-Assurance Mechanism for Cell Cycle Exit
(A) Feedback between E2F and CycE (B) is lost after terminal differen-
tiation (gray dotted arrows), ensuring that cell cycle exit induced by
inhibition of CycE or E2F is robust.
640 Developmental Cell 12, 631–643, April 2007 ª2007 Elsevie
physiologically relevant to cancers, in which gene expres-
sion can be greatly amplified.
RB- and CKI-Independent Mechanisms Mediate Cell
Cycle Exit in the Wing
Recent studies of cycle exit in larval Drosophila eyes have
concluded that Rbf1 and Dap are required to inhibit E2F
and CycE/Cdk2 in differentiating photoreceptors (Firth
and Baker, 2005; Escudero and Freeman, 2007). Other
studies document the roles of Ago/Fbw7 and components
of the Hippo/Warts-signaling pathway in downregulating
CycE for cell cycle exit in nonneural cells in the eye (Edgar,
2006; Moberg et al., 2001). Although our data are consis-
tent with these studies in the eye, we found that Ago and
the Hippo/Warts pathway are dispensable for cell cycle
exit in the wing (Figures S3 and S4; Table S2). Further-
more, deletion of Rbf1 did not prevent cell cycle exit in
the epithelial wing, even when high levels of CycE/Cdk2
were provided. Conversely, deletion of Dap was not suffi-
cient to keep wing cells cycling, even when excessive
E2F activity was provided (Figure 3). These observations
suggest that unknown inhibitors of E2F and Cyclin/Cdk
activity mediate cell cycle exit in specific contexts, such
as the wing.
In attempts to identify upstream factors regulating cell
cycle exit, we have manipulated a variety of growth and
patterning signals in the pupal wing and eye, and we ex-
amined their effects on cell cycle exit. Surprisingly, signals
that act as potent inducers of proliferation in wings and
eyes at earlier stages did not prevent or even delay cell cy-
cle exit upon terminal differentiation (Tables S2 and S3).
Thus, an important focus for future studies will be the na-
ture of the signals upstream of E2F and CycE that mediate
cell cycle exit. These could be novel signals, or combina-
tions of known signals delivered in unappreciated ways.
How General Is Double Assurance?
Studies of cell cycle exit in mammals do not offer a consis-
tent answer to this question. We show that S phase re-
entry can be achieved in differentiated cells by activating
E2F, CycE/Cdk2, or CycD/Cdk4 alone, but this does not
lead to cell division or continued proliferation. Several
studies with mammalian cells in vivo have shown that
neither increased E2F nor Cyclin/Cdk activity alone is
sufficient to fully reverse differentiation-associated quies-
cence (Camarda et al., 2004; Chandrasekaran et al., 1996;
Latella et al., 2001; Pajalunga et al., 1999; Sacco et al.,
2003), consistent with the double-assurance model we
propose here. Also consistent with this model is the ability
of proteins from DNA tumor viruses, such as adenovirus
E1A, SV40 LargeT, and HPV E6 and E7, to fully reverse dif-
ferentiation-associated cell cycle exit in many cell types
(Crescenzi et al., 1995; Derer et al., 2002; Endo and Na-
dal-Ginard, 1998; Kim et al., 1994; Latella et al., 2000).
These viral onco-proteins stimulate cell cycle progression
by targeting multiple cell cycle factors, which ultimately in-
crease both E2F and G1 Cyclin/Cdk activities simulta-
neously. For example, LargeT and E1A inhibit both RBs
r Inc.

Developmental Cell
Redundant Mechanisms Ensure Cell Cycle Exit
and CKIs, such as p21Cip1 and p27Kip1 (reviewed in Felsani
et al., 2006; Funk and Galloway, 1998).
There are some instances, however, in which differenti-
ation-associated cell cycle exit has been bypassed, not
just delayed, by the deletion of CKIs or RBs. In one such
case, p19Ink4d and p27Kip1 were knocked out in the mouse
brain, and ectopic mitoses were documented in neuronal
cells weeks after they normally become quiescent (Zindy
et al., 1999). Similar results have been obtained with hair
and support cells in the mouse inner ear, where deletion
of p19Ink4d, p27Kip1, or pRB can bypass developmentally
programmed cell cycle exit (reviewed in Chen, 2006). In
light of these findings, it is interesting to speculate that
certain differentiated tissues may retain some ability to re-
pair or regenerate by maintaining the capacity for positive
feedback between E2F and CycE/Cdk2 activity. Inner-ear
hair cells may be such an example, since in many verte-
brates they are capable of regeneration, although this abil-
ity has been lost in mammals (Corwin and Oberholtzer,
1997). Although the mammalian brain has a very limited
capacity for regeneration, the cell cycle can be reactivated
in the brains of other vertebrates, such as fish, in response
to injury (Zupanc, 2006). Thus, the retention of crosstalk
between E2F and Cyclin/Cdk activities in the evolutionary
descendents of regeneration-competent cells might
explain some of the tissue-specific sensitivities to loss of
CKIs or RBs observed in mammals.
EXPERIMENTAL PROCEDURES
Fly Strains and Antibodies
A complete list of the strains and antibodies used is available in Sup-
plemental Experimental Procedures.
Histology
Pupae, staged from white pre-pupae (0 hr after pupa formation, hr
APF) at 25�C, were dissected and fixed as described (Reis and Edgar,
2004). Pupal cuticle was removed from wings postfixation. Tissues 24–
36 hr APF were blocked in PBS + 0.1% Triton X-100 + 1% BSA for 1 hr.
Tissues 36–44 hr APF were blocked in PBS + 0.3% Triton X-100 + 1%
BSA (PAT) overnight, and tissues from pupae older than 44 hr APF
were incubated for 20 min at 25�C in a mixture of 50 mg/ml chitinase,
100 mg/ml collagenase, and 250 mg/ml trypsin, then blocked in PAT
overnight. Note that the wing hinge and notum were excluded from
our analyses. F-actin was stained with Alexa 633- or 568-conjugated
phalloidin (1:200, Molecular Probes). Hoechst 33258 (Molecular
Probes, 1 mg/ml) labels nuclei. Confocal sections were collected on a
Zeiss LSM 510 microscope. Single representative sections (2.2 mm)
are shown.
Flow Cytometry
Dissociation of cells from staged, dissected pupae and FACS were
carried out as described (Neufeld et al., 1998). Secretion of cuticle im-
peded the dissociation of wings after 36 hr APF; therefore, cell counts
per experiment were�10,000 (Figure 1),�5,000 (Figures 2 and 4), and
�2,000 (Figure 5). All experiments were carried out at least three times;
representative examples are shown.
Clonal Analysis
We used the heat shock (hs)-Flp/FRT tub>CD2>Gal4 method, with
tubulin temperature-sensitive Gal80 (tub-Gal80TS), for clonal overex-
pression (McGuire et al., 2004). By phenotypic analyses and GFP visu-
alization, we find complete inhibition of Gal4 in all lines used here with
Dev
tub-Gal80TS at 18�C, and we detect activation of target genes within 6
hr of shifting to 29�C. Development at 29�C proceeds 1.2 times faster
than at 25�C, and 2.2 times more slowly at 18�C (Ashburner, 1989). All
incubation times were adjusted accordingly. Hours APF are presented
as the equivalent time at 25�C for simplicity. Details of experiments
with Gal80TS are available in Supplemental Experimental Procedures.
Clones for cell/clone analysis were induced by hs-Flp/FRT
act>CD2>Gal4 recombination (Pignoni and Zipursky, 1997) at 0 hr
APF. Pupae were heat shocked for 2 min at 37�C, and wings were dis-
sected and fixed at 40 hr APF. GFP-positive cells per clone were
scored blind on a Leica DMR microscope.
Loss-of-function clones were generated by using mitotic recombi-
nation (Xu and Rubin, 1993) or MARCM (Lee and Luo, 2001). Larvae
were heat shocked for 45 min at 37�C at 48–72 hr after egg deposition,
collected for staging at 0 hr APF, aged at 25�C, and dissected at the
indicated times.
Quantitative Real-Time RT-PCR
RNA was isolated from 20 pupal wings by using Trizol (Invitrogen) and
RNAeasy columns (QIAGEN). Approximately one-third of total RNA
was used for cDNA synthesis in a 40 ml reaction. PCR products were
monitored by using SYBR-Green (Molecular Probes) and were ana-
lyzed on a MyIQ iCycler (BioRad). Primer sequences and additional
details are available in Supplemental Experimental Procedures.
Supplemental Data
Supplemental Data include nine figures and four tables, lists of
Drosophila strains and oligonucleotide primers used, and additional
Experimental Procedures and are available at http://www.
developmentalcell.com/cgi/content/full/12/4/631/DC1/.
ACKNOWLEDGMENTS
We thank D. O’Keefe for help with Gal80TS, members of the B.A.E. Lab
and M. Welcker for discussions, and Drs. C. Moens and M. Van Gilst
for use of their equipment. We thank Drs. W. Du, C. Lehner, D. Pan,
T. Orr-Weaver, G. Halder, L. Firth, N. Baker, I. Hariharan, H. Richard-
son, the Bloomington Stock Center, and the Developmental Studies
Hybridoma Bank for flies and/or antibodies. L.A.B. is supported by
a Damon Runyon postdoctoral fellowship (DRG#1838-04). Develop-
mental Biology Training Grant T32 HDO7183 supports A.J.K. This
work was supported by National Institutes of Health grants
GM51186 (2003-2006) and GM070887 (2006-2007) to B.A.E.
Received: December 20, 2006
Revised: February 13, 2007
Accepted: February 27, 2007
Published: April 9, 2007
REFERENCES
Asano, M., Nevins, J.R., and Wharton, R.P. (1996). Ectopic E2F ex-
pression induces S phase and apoptosis in Drosophila imaginal discs.
Genes Dev. 10, 1422–1432.
Ashburner, M. (1989). Drosophila: A Laboratory Handbook (Cold
Spring Harbor, NY: Cold Spring Harbor Press).
Attwooll, C., Lazzerini Denchi, E., and Helin, K. (2004). The E2F family:
specific functions and overlapping interests. EMBO J. 23, 4709–4716.
Calvi, B.R., Lilly, M.A., and Spradling, A.C. (1998). Cell cycle control of
chorion gene amplification. Genes Dev. 12, 734–744.
Camarda, G., Siepi, F., Pajalunga, D., Bernardini, C., Rossi, R., Monte-
cucco, A., Meccia, E., and Crescenzi, M. (2004). A pRb-independent
mechanism preserves the postmitotic state in terminally differentiated
skeletal muscle cells. J. Cell Biol. 167, 417–423.
Cayirlioglu, P., Bonnette, P.C., Dickson, M.R., and Duronio, R.J.
(2001). Drosophila E2f2 promotes the conversion from genomic DNA
elopmental Cell 12, 631–643, April 2007 ª2007 Elsevier Inc. 641

Developmental Cell
Redundant Mechanisms Ensure Cell Cycle Exit
replication to gene amplification in ovarian follicle cells. Development
128, 5085–5098.
Chandrasekaran, C., Coopersmith, C.M., and Gordon, J.I. (1996). Use
of normal and transgenic mice to examine the relationship between
terminal differentiation of intestinal epithelial cells and accumulation
of their cell cycle regulators. J. Biol. Chem. 271, 28414–28421.
Chen, Z.Y. (2006). Cell cycle, differentiation and regeneration: where
to begin? Cell Cycle 5, 2609–2612.
Cobrinik, D. (2005). Pocket proteins and cell cycle control. Oncogene
24, 2796–2809.
Corwin, J.T., and Oberholtzer, J.C. (1997). Fish n’ chicks: model rec-
ipes for hair-cell regeneration? Neuron 19, 951–954.
Crescenzi, M., Soddu, S., and Tato, F. (1995). Mitotic cycle reactivation
in terminally differentiated cells by adenovirus infection. J. Cell. Phys-
iol. 162, 26–35.
Datar, S.A., Jacobs, H.W., de la Cruz, A.F., Lehner, C.F., and Edgar,
B.A. (2000). The Drosophila cyclin D-Cdk4 complex promotes cellular
growth. EMBO J. 19, 4543–4554.
de Nooij, J.C., Letendre, M.A., and Hariharan, I.K. (1996). A cyclin-
dependent kinase inhibitor, Dacapo, is necessary for timely exit from
the cell cycle during Drosophila embryogenesis. Cell 87, 1237–1247.
Derer, W., Easwaran, H.P., Leonhardt, H., and Cardoso, M.C. (2002). A
novel approach to induce cell cycle reentry in terminally differentiated
muscle cells. FASEB J. 16, 132–133.
Dimova, D.K., and Dyson, N.J. (2005). The E2F transcriptional network:
old acquaintances with new faces. Oncogene 24, 2810–2826.
Dimova, D.K., Stevaux, O., Frolov, M.V., and Dyson, N.J. (2003). Cell
cycle-dependent and cell cycle-independent control of transcription
by the Drosophila E2F/RB pathway. Genes Dev. 17, 2308–2320.
Du, W., and Dyson, N. (1999). The role of RBF in the introduction of G1
regulation during Drosophila embryogenesis. EMBO J. 18, 916–925.
Du, W., and Pogoriler, J. (2006). Retinoblastoma family genes. Onco-
gene 25, 5190–5200.
Du, W., Vidal, M., Xie, J.E., and Dyson, N. (1996a). RBF, a novel RB-
related gene that regulates E2F activity and interacts with cyclin E in
Drosophila. Genes Dev. 10, 1206–1218.
Du, W., Xie, J.E., and Dyson, N. (1996b). Ectopic expression of dE2F
and dDP induces cell proliferation and death in the Drosophila eye.
EMBO J. 15, 3684–3692.
Duronio, R.J., and O’Farrell, P.H. (1995). Developmental control of the
G1 to S transition in Drosophila: cyclin E is a limiting downstream target
of E2F. Genes Dev. 9, 1456–1468.
Duronio, R.J., Brook, A., Dyson, N., and O’Farrell, P.H. (1996). E2F-
induced S phase requires cyclin E. Genes Dev. 10, 2505–2513.
Dyson, N. (1998). The regulation of E2F by pRB-family proteins. Genes
Dev. 12, 2245–2262.
Edgar, B.A. (2006). From cell structure to transcription: hippo forges
a new path. Cell 124, 267–273.
Endo, T., and Nadal-Ginard, B. (1998). Reversal of myogenic terminal
differentiation by SV40 large T antigen results in mitosis and apoptosis.
J. Cell Sci. 111, 1081–1093.
Escudero, L.M., and Freeman, M. (2007). Mechanism of G1 arrest in
the Drosophila eye imaginal disc. BMC Dev. Biol. 7, 13.
Felsani, A., Mileo, A.M., and Paggi, M.G. (2006). Retinoblastoma family
proteins as key targets of the small DNA virus oncoproteins. Oncogene
25, 5277–5285.
Firth, L.C., and Baker, N.E. (2005). Extracellular signals responsible for
spatially regulated proliferation in the differentiating Drosophila eye.
Dev. Cell 8, 541–551.
Foijer, F., Wolthuis, R.M., Doodeman, V., Medema, R.H., and te Riele,
H. (2005). Mitogen requirement for cell cycle progression in the ab-
sence of pocket protein activity. Cancer Cell 8, 455–466.
642 Developmental Cell 12, 631–643, April 2007 ª2007 Elsevie
Frolov, M.V., Huen, D.S., Stevaux, O., Dimova, D., Balczarek-Strang,
K., Elsdon, M., and Dyson, N.J. (2001). Functional antagonism be-
tween E2F family members. Genes Dev. 15, 2146–2160.
Frolov, M.V., Moon, N.S., and Dyson, N.J. (2005). dDP is needed for
normal cell proliferation. Mol. Cell. Biol. 25, 3027–3039.
Funk, J.O., and Galloway, D.A. (1998). Inhibiting CDK inhibitors: new
lessons from DNA tumor viruses. Trends Biochem. Sci. 23, 337–341.
Keller, S.A., Ullah, Z., Buckley, M.S., Henry, R.W., and Arnosti, D.N.
(2005). Distinct developmental expression of Drosophila retinoblas-
toma factors. Gene Expr. Patterns 5, 411–421.
Kim, S.H., Roth, K.A., Coopersmith, C.M., Pipas, J.M., and Gordon, J.I.
(1994). Expression of wild-type and mutant simian virus 40 large tumor
antigens in villus-associated enterocytes of transgenic mice. Proc.
Natl. Acad. Sci. USA 91, 6914–6918.
Kylsten, P., and Saint, R. (1997). Imaginal tissues of Drosophila mela-
nogaster exhibit different modes of cell proliferation control. Dev. Biol.
192, 509–522.
Lane, M.E., Sauer, K., Wallace, K., Jan, Y.N., Lehner, C.F., and
Vaessin, H. (1996). Dacapo, a cyclin-dependent kinase inhibitor, stops
cell proliferation during Drosophila development. Cell 87, 1225–1235.
Latella, L., Sacchi, A., and Crescenzi, M. (2000). Long-term fate of
terminally differentiated skeletal muscle cells following E1A-initiated
cell cycle reactivation. Cell Death Differ. 7, 145–154.
Latella, L., Sacco, A., Pajalunga, D., Tiainen, M., Macera, D., D’Angelo,
M., Felici, A., Sacchi, A., and Crescenzi, M. (2001). Reconstitution of
cyclin D1-associated kinase activity drives terminally differentiated
cells into the cell cycle. Mol. Cell. Biol. 21, 5631–5643.
Lee, T., and Luo, L. (2001). Mosaic analysis with a repressible cell
marker (MARCM) for Drosophila neural development. Trends Neuro-
sci. 24, 251–254.
Liu, J.L., Buszczak, M., and Gall, J.G. (2006). Nuclear bodies in the
Drosophila germinal vesicle. Chromosome Res. 14, 465–475.
McGuire, S.E., Mao, Z., and Davis, R.L. (2004). Spatiotemporal gene
expression targeting with the TARGET and gene-switch systems in
Drosophila. Sci. STKE 2004, pl6.
Meyer, C.A., Jacobs, H.W., Datar, S.A., Du, W., Edgar, B.A., and
Lehner, C.F. (2000). Drosophila Cdk4 is required for normal growth
and is dispensable for cell cycle progression. EMBO J. 19, 4533–4542.
Milan, M., Campuzano, S., and Garcia-Bellido, A. (1996). Cell cycling
and patterned cell proliferation in the Drosophila wing during metamor-
phosis. Proc. Natl. Acad. Sci. USA 93, 11687–11692.
Moberg, K.H., Bell, D.W., Wahrer, D.C., Haber, D.A., and Hariharan,
I.K. (2001). Archipelago regulates Cyclin E levels in Drosophila and is
mutated in human cancer cell lines. Nature 413, 311–316.
Neufeld, T.P., de la Cruz, A.F., Johnston, L.A., and Edgar, B.A. (1998).
Coordination of growth and cell division in the Drosophila wing. Cell 93,
1183–1193.
Pajalunga, D., Tognozzi, D., Tiainen, M., D’Angelo, M., Ferrantelli, F.,
Helin, K., Sacchi, A., and Crescenzi, M. (1999). E2F activates late-G1
events but cannot replace E1A in inducing S phase in terminally differ-
entiated skeletal muscle cells. Oncogene 18, 5054–5062.
Pignoni, F., and Zipursky, S.L. (1997). Induction of Drosophila eye
development by decapentaplegic. Development 124, 271–278.
Reis, T., and Edgar, B.A. (2004). Negative regulation of dE2F1 by
cyclin-dependent kinases controls cell cycle timing. Cell 117, 253–264.
Richardson, H., O’Keefe, L.V., Marty, T., and Saint, R. (1995). Ectopic
cyclin E expression induces premature entry into S phase and disrupts
pattern formation in the Drosophila eye imaginal disc. Development
121, 3371–3379.
Sacco, A., Siepi, F., and Crescenzi, M. (2003). HPV E7 expression in
skeletal muscle cells distinguishes initiation of the postmitotic state
from its maintenance. Oncogene 22, 4027–4034.
r Inc.

Developmental Cell
Redundant Mechanisms Ensure Cell Cycle Exit
Schubiger, M., and Palka, J. (1987). Changing spatial patterns of DNA
replication in the developing wing of Drosophila. Dev. Biol. 123, 145–
153.
Stevaux, O., and Dyson, N.J. (2002). A revised picture of the E2F tran-
scriptional network and RB function. Curr. Opin. Cell Biol. 14, 684–691.
Stevaux, O., Dimova, D., Frolov, M.V., Taylor-Harding, B., Morris, E.,
and Dyson, N. (2002). Distinct mechanisms of E2F regulation by
Drosophila RBF1 and RBF2. EMBO J. 21, 4927–4937.
Stevaux, O., Dimova, D.K., Ji, J.Y., Moon, N.S., Frolov, M.V., and
Dyson, N.J. (2005). Retinoblastoma family 2 is required in vivo for the
tissue-specific repression of dE2F2 target genes. Cell Cycle 4, 1272–
1280.
Su, T.T., Sprenger, F., DiGregorio, P.J., Campbell, S.D., and O’Farrell,
P.H. (1998). Exit from mitosis in Drosophila syncytial embryos requires
proteolysis and cyclin degradation, and is associated with localized
dephosphorylation. Genes Dev. 12, 1495–1503.
Thacker, S.A., Bonnette, P.C., and Duronio, R.J. (2003). The contribu-
tion of E2F-regulated transcription to Drosophila PCNA gene function.
Curr. Biol. 13, 53–58.
Vidal, A., and Koff, A. (2000). Cell-cycle inhibitors: three families united
by a common cause. Gene 247, 1–15.
Deve
Weng, L., Zhu, C., Xu, J., and Du, W. (2003). Critical role of active
repression by E2F and Rb proteins in endoreplication during Drosophila
development. EMBO J. 22, 3865–3875.
Wolff, T., and Ready, D. (1993). Pattern formation in the Drosophila
retina. In The Development of Drosophila melanogaster, M. Bate and
A. Arias, eds. (Cold Spring Harbor, NY: Cold Spring Harbor Press),
pp. 1277–1326.
Xu, T., and Rubin, G.M. (1993). Analysis of genetic mosaics in develop-
ing and adult Drosophila tissues. Development 117, 1223–1237.
Zhang, P. (1999). The cell cycle and development: redundant roles of
cell cycle regulators. Curr. Opin. Cell Biol. 11, 655–662.
Zhu, L., and Skoultchi, A.I. (2001). Coordinating cell proliferation and
differentiation. Curr. Opin. Genet. Dev. 11, 91–97.
Zindy, F., Cunningham, J.J., Sherr, C.J., Jogal, S., Smeyne, R.J., and
Roussel, M.F. (1999). Postnatal neuronal proliferation in mice lacking
Ink4d and Kip1 inhibitors of cyclin-dependent kinases. Proc. Natl.
Acad. Sci. USA 96, 13462–13467.
Zupanc, G.K. (2006). Neurogenesis and neuronal regeneration in the
adult fish brain. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav.
Physiol. 192, 649–670.
lopmental Cell 12, 631–643, April 2007 ª2007 Elsevier Inc. 643
Related Documents