JOURNAL OF CRUSTACEAN BIOLOGY, 33(3), 309-316, 2013 A COMPARATIVE ANALYSIS OF SETAE ON THE PEREIOPODS OF REPRODUCTIVE MALE AND FEMALE ORCONECTES RUSTICUS (DECAPODA: ASTACIDAE) Rachelle M. Belanger 1,2,∗ and Paul A. Moore 1 1 Laboratory for Sensory Ecology, J. P. Scott Center for Neuroscience, Mind & Behavior, Department of Biological Sciences, Bowling Green State University, Bowling Green, OH 43403, USA 2 Biology Department, University of Detroit Mercy, Detroit, MI 48221, USA ABSTRACT The pereiopods (walking legs) of crustaceans have been shown to have both mechano- and chemosensory functions including: detecting various odors, mechanical stimuli, as well as being used for grooming of gills and brood care. Using scanning electron microscopy, we examined the pereiopods of reproductive male (form I) and reproductive female (glair) crayfish Orconectes rusticus Girard, 1852 and found that the distal portion (dorsal surface) of the propodus of pereiopods 2 through 5 contain smooth, plumose, serrate and cuspidate setae. Various studies have implicated both smooth and serrate setae as being important for grooming; therefore, we used scanning electron microscopy to examine and compare the distribution of smooth and serrate setae on the pereiopods of reproductive male and female crayfish in order to determine if there were differences between sexes. We found that there were no differences in the number of tufts (pockets) or number of smooth setae on the second and third pereiopods (chelipeds) between male and female crayfish. Further, the second and third pereiopods of both male and female crayfish had significantly more smooth setae than the fourth and fifth pereiopods. When the distribution of serrate setae was compared, we found that reproductive female crayfish possess significantly more serrate setae on their fifth pereiopods than their fourth pereiopods and both pereiopods 4 and 5 of form I male crayfish. Overall, increases in serrate setae on the fifth pereiopods of reproductive female crayfish are significant. We suggest that female crayfish may use serrate setae of the fifth pereiopods extensively for grooming eggs and brood care during reproduction. KEY WORDS: crayfish, grooming, Orconectes rusticus, pereiopods, serrate setae, simple setae DOI: 10.1163/1937240X-00002144 I NTRODUCTION Decapod crustaceans sense both chemical and mechanical information from their external environment via the use of sensory setae, located on the cuticle of chephalothoracic ap- pendages (Derby, 1982, 1989; Laverack, 1988). These ap- pendages, containing both mechano- and chemosensory se- tae, include antennae, antennules, maxillipeds (mouthparts), and pereiopods (major chelae and walking legs). Clawed lobsters and crayfish contain five pairs of pereiopods, which include a pair of major chelae (pereiopod 1) and four anatomically distinct pairs of walking legs (pereiopods 2- 5). The first two pairs of walking legs (pereiopods 2 and 3) are also chelipeds, but they contain smaller chelae than those found on the first pereiopods. The fourth and fifth pereiopods have the same overall structure as second and third pereiopods, but they are non-chelate (Pond, 1975; Lavalli and Factor, 1995; Holdich, 2002). Several different types of setae have been found on the pereiopods of crus- taceans including: smooth (simple), cuspidate (also referred to as toothed or fringed setae), plumose (feathered hairs), squamous (serrulate) and serrate setae as well as type I and II peg sensillum (Shelton and Laverack, 1968; Thomas, 1970; Derby, 1982, 1989; Lavalli and Factor, 1995; Belanger et al., 2008). The type, distribution and abundance of setae found on the pereiopods may also vary depending on the pereio- ∗ Corresponding author; e-mail: [email protected] pod they are located on and the sex and reproductive status of the individual (Thomas, 1981; Lavalli and Factor, 1995; Belanger et al., 2008). Furthermore, Thomas (1981) showed that there were no differences in the types of setae found on the pereiopods of crayfish Austropotamobius pallipes (Lere- boullet, 1858), but the abundance and distribution of the se- tae types between the males and females was not been com- pared. The pereiopods of crayfish are used in agonistic en- counters, predator defense, locomotion and escape, feeding, grooming/antifouling and reproduction (Bovbjerg, 1956; Hodgson, 1958; Pond, 1975; Stein, 1976; Ameyaw-Akumfi, 1977; Cooke and Macmillan, 1985; Jones and Lester, 1996; Bauer, 1998; Hazlett and Schoolmaster, 1998; Nyström and Pérez, 1998; Batang and Suzuki, 1999, 2000; Hazlett, 1999; Holdich, 2002). The setae, found on pereiopods 2 through 5 (the walking legs), have been shown to respond to both mechano- and chemosensory information and are also used for antifouling the gills (Hodgson, 1958; Bauer et al., 1981; Bauer, 1998, 2002; Corotto and O’Brien, 2002). Chemosen- sory receptors, located on the pereiopods, have also been implicated as potential sources of information for distance orientation when locating food sources (Moore et al., 1991; Keller et al., 2003). The second and third pereiopods are chelipeds that are capable of acquiring and holding food © The Crustacean Society, 2013. Published by Brill NV, Leiden DOI:10.1163/1937240X-00002144

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

JOURNAL OF CRUSTACEAN BIOLOGY, 33(3), 309-316, 2013
A COMPARATIVE ANALYSIS OF SETAE ON THE PEREIOPODS OF REPRODUCTIVEMALE AND FEMALE ORCONECTES RUSTICUS (DECAPODA: ASTACIDAE)
Rachelle M. Belanger 1,2,∗ and Paul A. Moore 1
1 Laboratory for Sensory Ecology, J. P. Scott Center for Neuroscience, Mind & Behavior, Department ofBiological Sciences, Bowling Green State University, Bowling Green, OH 43403, USA
2 Biology Department, University of Detroit Mercy, Detroit, MI 48221, USA
A B S T R A C T
The pereiopods (walking legs) of crustaceans have been shown to have both mechano- and chemosensory functions including: detectingvarious odors, mechanical stimuli, as well as being used for grooming of gills and brood care. Using scanning electron microscopy, weexamined the pereiopods of reproductive male (form I) and reproductive female (glair) crayfish Orconectes rusticus Girard, 1852 andfound that the distal portion (dorsal surface) of the propodus of pereiopods 2 through 5 contain smooth, plumose, serrate and cuspidatesetae. Various studies have implicated both smooth and serrate setae as being important for grooming; therefore, we used scanning electronmicroscopy to examine and compare the distribution of smooth and serrate setae on the pereiopods of reproductive male and femalecrayfish in order to determine if there were differences between sexes. We found that there were no differences in the number of tufts(pockets) or number of smooth setae on the second and third pereiopods (chelipeds) between male and female crayfish. Further, the secondand third pereiopods of both male and female crayfish had significantly more smooth setae than the fourth and fifth pereiopods. When thedistribution of serrate setae was compared, we found that reproductive female crayfish possess significantly more serrate setae on their fifthpereiopods than their fourth pereiopods and both pereiopods 4 and 5 of form I male crayfish. Overall, increases in serrate setae on the fifthpereiopods of reproductive female crayfish are significant. We suggest that female crayfish may use serrate setae of the fifth pereiopodsextensively for grooming eggs and brood care during reproduction.
KEY WORDS: crayfish, grooming, Orconectes rusticus, pereiopods, serrate setae, simple setae
DOI: 10.1163/1937240X-00002144
INTRODUCTION
Decapod crustaceans sense both chemical and mechanicalinformation from their external environment via the use ofsensory setae, located on the cuticle of chephalothoracic ap-pendages (Derby, 1982, 1989; Laverack, 1988). These ap-pendages, containing both mechano- and chemosensory se-tae, include antennae, antennules, maxillipeds (mouthparts),and pereiopods (major chelae and walking legs). Clawedlobsters and crayfish contain five pairs of pereiopods, whichinclude a pair of major chelae (pereiopod 1) and fouranatomically distinct pairs of walking legs (pereiopods 2-5). The first two pairs of walking legs (pereiopods 2 and3) are also chelipeds, but they contain smaller chelae thanthose found on the first pereiopods. The fourth and fifthpereiopods have the same overall structure as second andthird pereiopods, but they are non-chelate (Pond, 1975;Lavalli and Factor, 1995; Holdich, 2002). Several differenttypes of setae have been found on the pereiopods of crus-taceans including: smooth (simple), cuspidate (also referredto as toothed or fringed setae), plumose (feathered hairs),squamous (serrulate) and serrate setae as well as type I and IIpeg sensillum (Shelton and Laverack, 1968; Thomas, 1970;Derby, 1982, 1989; Lavalli and Factor, 1995; Belanger et al.,2008). The type, distribution and abundance of setae foundon the pereiopods may also vary depending on the pereio-
∗ Corresponding author; e-mail: [email protected]
pod they are located on and the sex and reproductive statusof the individual (Thomas, 1981; Lavalli and Factor, 1995;Belanger et al., 2008). Furthermore, Thomas (1981) showedthat there were no differences in the types of setae found onthe pereiopods of crayfish Austropotamobius pallipes (Lere-boullet, 1858), but the abundance and distribution of the se-tae types between the males and females was not been com-pared.
The pereiopods of crayfish are used in agonistic en-counters, predator defense, locomotion and escape, feeding,grooming/antifouling and reproduction (Bovbjerg, 1956;Hodgson, 1958; Pond, 1975; Stein, 1976; Ameyaw-Akumfi,1977; Cooke and Macmillan, 1985; Jones and Lester, 1996;Bauer, 1998; Hazlett and Schoolmaster, 1998; Nyström andPérez, 1998; Batang and Suzuki, 1999, 2000; Hazlett, 1999;Holdich, 2002). The setae, found on pereiopods 2 through5 (the walking legs), have been shown to respond to bothmechano- and chemosensory information and are also usedfor antifouling the gills (Hodgson, 1958; Bauer et al., 1981;Bauer, 1998, 2002; Corotto and O’Brien, 2002). Chemosen-sory receptors, located on the pereiopods, have also beenimplicated as potential sources of information for distanceorientation when locating food sources (Moore et al., 1991;Keller et al., 2003). The second and third pereiopods arechelipeds that are capable of acquiring and holding food
© The Crustacean Society, 2013. Published by Brill NV, Leiden DOI:10.1163/1937240X-00002144

310 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 33, NO. 3, 2013
items and are used in locomotion and stabilization (Stein,1976; Cooke and Macmillan, 1985; Brown, 1995; Holdich,2002). These pereiopods have been shown to contain bi-modal smooth (simple) setae arranged in tufts over thechelae and cuspidate setae that line the ventral margins ofthe dactyli, used for seizing objects (Thomas, 1970; Hatt,1986). The second and third pereiopods also contain se-tobranch setae, located on the coxa, which are used forgill cleaning and antifouling (Bauer, 1981, 1998). Electro-physiological and behavioral studies that examine the sec-ond and third pereiopods of Cambarus bartonii scioten-sis (Rhoades, 1944), Austropotamobius torrentium (Schrank,1803), Orconectes limosus (Rafinesque, 1815) and Procam-barus clarkii (Girard, 1852) demonstrate that they bear setaecapable of serving as mechanoreceptors or contact chemore-ceptors (Hodgson, 1958; Bauer et al., 1981; Altner et al.,1983; Corotto and O’Brien, 2002). By covering the dactylsof second and third pereiopods in P. clarkii, Ameyaw-Akumfi (1977) also demonstrated behaviorally that theycontain both mechano- and chemosensory setae, importantfor food detection. Crayfish, Cherax quadricarinatus (vonMartens, 1868), have also been shown to groom their bod-ies with the second and third chelipeds and female crayfish,Pacifastacus trowbridgi (Stimpson, 1857), also use thesechelipeds for mechanically grasping, turning and clean-ing eggs (Mason, 1970b; Jones and Lester, 1996; Holdich,2002). Given that female crayfish use the chelae of the sec-ond and third pereiopods for egg grooming and maintenanceand that the tufts of setae on the chelae of these pereiopodsmay be used for grooming (Bauer, 1998), differences in thenumber of tufts, setae abundance, and distribution on thesepereiopods may exist between reproductive male and femalecrayfish.
Pereiopods 2 through 5 all have shown to be important forbody grooming/antifouling and egg grooming and mainte-nance. Along with using pereiopods 2 and 3 for egg groom-ing as in C. quadricarinatus, A. pallipes carefully cleans thepleopods with serrate, smooth, and cuspidate setae locatedon the propods and dactyls of the fourth and fifth pereiopodsof the crayfish before egg laying (Mason, 1970a; Thomas,1970, 1977; Jones and Lester, 1996; Holdich, 2002). Fur-ther, long bouts of grooming also occur between copula-tion and egg-laying in the crayfish P. trowbridgii with thefourth and fifth pereiopods (Mason, 1970a). In fact, foulingof the body increases in crayfish (C. quadricarinatus) whengrooming limbs are incapacitated (Jones and Lester, 1996).The fourth and fifth pereiopods of Orconectes rusticus (Gi-rard, 1852) and Procambarus fallax (Hagen, 1870) are posi-tioned in very close proximity to clutches of eggs, attachedto pleopods (Fig. 1; also see Fig. 1 in Vogt and Tolley, 2004).Overall, it has been concluded that setae on pereiopods 4and 5 are used predominantly for grooming, especially byreproductive females, prior to and after egg laying. Quanti-fying and examining the arrangement of setae types on thepereiopods will lead and clearer understanding of their func-tion and determine if sexual dimorphisms with respect to se-tae quantity and distribution exist between male and femalecrayfish.
The aim of this study is to identify, quantify and comparethe setae found on the propodus of the walking legs
Fig. 1. The ventral surface of an egg-bearing female O. rusticus. Thesecond through fifth pereiopods may be used to groom the eggs (arrow)that are attached to the pleopods on the last abdominal segment. Scale baris 2 cm. Photo by H. Evan Spencer.
(pereiopods 2-5) of reproductive (form I) male crayfishand reproductive (glair) female O. rusticus. Given theimportance and diversity of functions of setae on thepereiopods, it is important to further investigate any sex-specific differences that might be attributed to reproductiveroles and uses of these setae. Because the pereiopods havebeen shown to be used for body grooming in both sexes andfor egg grooming behaviors in females (Mason, 1970b; Hatt,1986; Jones and Lester, 1996; Bauer, 2002; Holdich, 2002),we hypothesize that reproductive female crayfish will havemore setae on the propodus of the pereiopods. Specifically,we expect that because the chelipeds are used for eggturning, grooming and cleaning (Jones and Lester, 1996),reproductive females will have more tufts or an increasein the abundance of smooth setae, contained in thosetufts, when compared to form I males. Moreover, becausepereiopods 4 and 5 contain serrate setae used for cleaningand that reproductive female grooms her pleon extensivelybefore laying eggs (Mason, 1970a; Bauer, 2002); we expectthat reproductive females will contain more serrate setaeon these pereiopods when compared to males. Overall,because the setae on pereiopods 2 through 5 are importantfor grooming and antifouling of eggs, reproductive femaleshould have more setae on these appendages. Conversely,because both male and female crustaceans spend a lotof time performing general body grooming (Bauer, 1981,1989), differences in setae abundance and distribution maynot exist. Furthermore, because smooth setae are bimodal,and have been shown to be contact chemoreceptors (Hatt,1986), male and female crayfish may have a similar numberof these setae.
MATERIALS AND METHODS
Crayfish Collection
Reproductive male (form I) and reproductive female O. rusticus werecollected from the Portage River near Bowling Green State University,Bowling Green, OH, USA. Intermolt male crayfish used in all imagingexperiments were housed in population tanks in an environmental chamber

BELANGER AND MOORE: SETAE ON CRAYFISH PEREIOPODS 311
(23°C, 14 light : 10 dark). Crayfish were fed a diet of rabbit pellets threetimes per week. Crayfish mass, carapace length, and chelae length (mean ±S.D.) were measured for five males and five females (males: 14.2 ± 2.9 g;3.4 ± 0.2 cm carapace length; females: 15.0 ± 2.4 g; 3.7 ± 0.1 cm carapacelength). We used a 1-way MANOVA with the single factor being sex of thecrayfish with the two dependent variables being weight and carapace length.Results indicate that there was an overall significance (F(2,7,0.05) = 5.680,P = 0.034), although a Tukey-HSD post-hoc showed that there wasno difference between male and female weights or carapace length. Forall imaging experiments, pereiopods were dissected from the body of thecrayfish after they were anesthetized in an ice bath for at least 30 min.We used a 2-way ANOVA with a Tukey-HSD post-hoc test to comparethe length of the propodus for each pereiopod was measured (mean ± S.D.)for the male and female crayfish used in this study (males: 3.3 ± 0.3 cmpereiopod 1, 0.9 ± 0.1 cm pereiopod 2, 1.3 ± 0.1 cm pereiopod 3, 1.0 ±0.1 pereiopod 4 and 1.0 ± 0.04 cm pereiopod 5; females: 2.6 ± 0.3 cmpereiopod 1, 0.9 ± 0.03 cm pereiopod 2; 1.1 ± 0.1 cm pereiopod 3; 1.0 ±0.01 cm pereiopod 4 and 1.0 ± 0.04 cm pereiopod 5). There was an overallsignificant difference (F(4,40,0.05) = 10.1, P < 0.0001) between maleand female pereiopod length; however when individual pereiopods werecompared, only the major chelae (pereiopod 1) were significantly different(P < 0.05) between males and females.
Form I males were identified by examining their stylets (Crocker andBarr, 1968). Form I males were identified as having relatively long whitestylets that extend to the base of the second pereiopods. Females wereclassified as reproductive when glair glands could be visualized. Glairglands appear as whitened tissue on the underside of the female’s tail(Holdich, 2002).
Scanning Electron Microscopy
To examine the structure and setae distribution on the dorsal surface of thepropodus of the walking legs (pereiopods 2, 3, 4 and 5), scanning electronmicroscopy was performed using methods similar to Belanger et al. (2008).The pereiopods of our crayfish were dissected (N = 5 of each sex) andplaced in a sonicator containing 0.1 M phosphate-buffered saline (PBS,pH 7.4) to remove debris from the setae. Following sonication, fixation ofthe pereiopods occurred in 2.5% gluteraldehyde/3% paraformaldehyde in0.1 M PBS for at least 48 h. The pereiopods were then rinsed for 10 minin 0.1 M PBS, post-fixed in 1% osmium tetroxide for 2 h, and rinsed againfor 10 min with 0.1 M PBS. Following rinsing, pereiopods were dehydratedin an ethanol series (40-60-80-95-100-100-100%; 15 min each) and storedfor final drying in 100% ethanol. A graded hexamethyldislizane (ElectronMicroscopy Sciences, Fort Washington, PA, USA) and ethanol series wasused for drying the appendages (Nation, 1983). After the last change of100% hexamethyldislizane was complete, the pereiopods were allowed toair-dry overnight and were subsequently mounted onto stubs with doublesided tape and graphite paint and sputter-coated with gold/palladium. Thepereiopods were examined using a scanning electron microscope (SEM;Hitachi S-2700) where setae counts were performed and images werecaptured. The person identifying and counting the setae numbers was notaware of the sex of the crayfish being examined. Further, setae counts wereperformed twice on random samples to ensure accuracy (P > 0.05).
Setae Identification, Quantification and Data Analysis
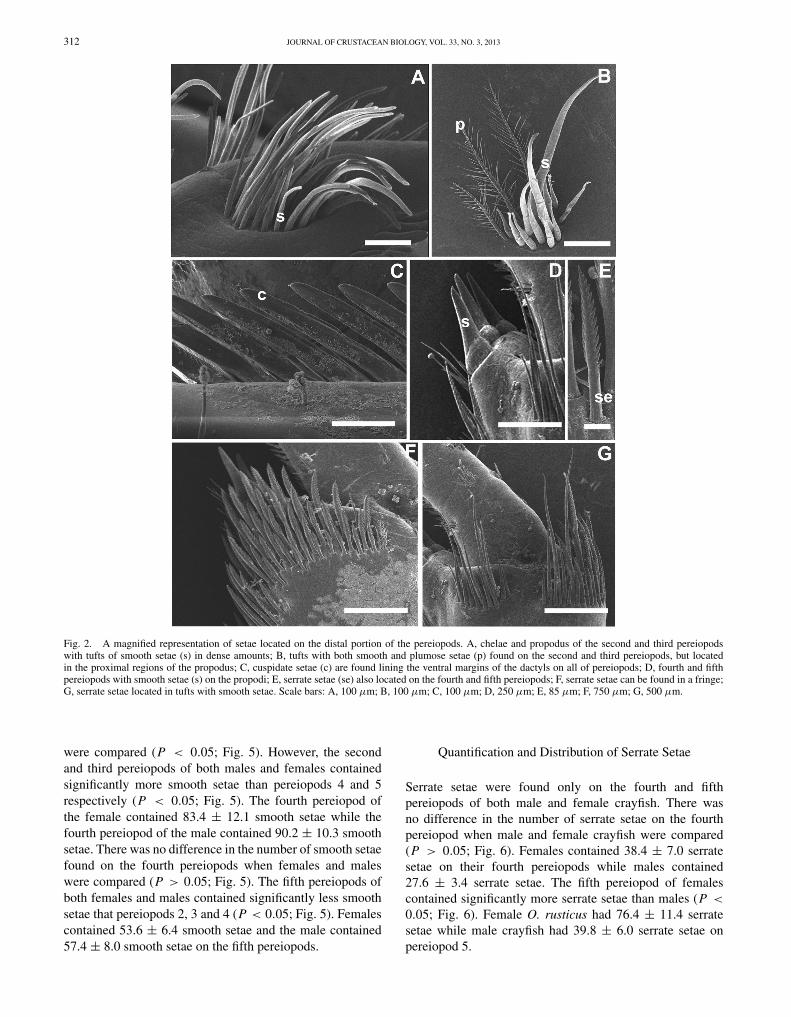
While examining video images of the pereiopods on the scanning electronmicroscope, smooth, plumose, toothed, serrate, and cuspidate setae presenton the dorsal surface of the distal portion of the propodus were identifiedand images were captured (see Fig. 2). Smooth and plumose setae wereidentified using descriptions from Thomas (1970) where smooth setae wereshown to have a smooth outline with pointed tips (Fig. 2A) and plumosesetae were feather-like, with long setules (Fig. 2B). Garm and Watling(2013) describe cuspidate setae as having a broad base and taper graduallytoward the somewhat rounded tip (Fig. 2C). Cuspidate setae are located onthe ventral margins of the dactyls on pereiopods 2 through 5 and on thedorsal margins of the propodial projection of pereiopods 2 and 3. Smoothsetae are found in the position opposite of the propodial projection and arevery robust with a length: width ratio of >15 (Fig. 2D). They have a broadbase and taper gradually towards the somewhat rounded tip (Thomas, 1970;Garm, 2004). Serrate setae, as described by Garm (2004), have a nakedproximal half with denticles that are distal to the annulus which may befound in a fringe or in patches (Fig. 2E).
In order to determine if there are differences in the number of groomingsetae on the pereiopods of reproductive male and female O. rusticus, we
used a SEM to examine the distal portion of the pereiopods and quantifiedand compared setae numbers between sexes. Because it has been suggestedthat crayfish use tufts of simple setae on the second and third pereiopods forgrooming (Bauer, 1998), we counted the total number of smooth setae andthe number of tufts on the chelipeds. Further, we quantified and comparedthe smooth and serrate setae on the fourth and fifth pereiopods of male andfemale O. rusticus. Serrate setae have been shown to be used for cleaningthe pleopods before egg-laying and for preening of the eggs (Thomas, 1970,1977; Holdich, 2002). A 1-way ANOVA (number of tufts with respect to thepereiopod they were location on) was used to compare the overall numberof setae tufts between male and female crayfish. We used a 3-way ANOVAwith a Fisher-LSD post-hoc test to identify changes in setae number withrespect to sex (factor 1), pereiopod number (factor 2), as well as setae type(factor 3).
RESULTS
Pereiopod and Setae Descriptions
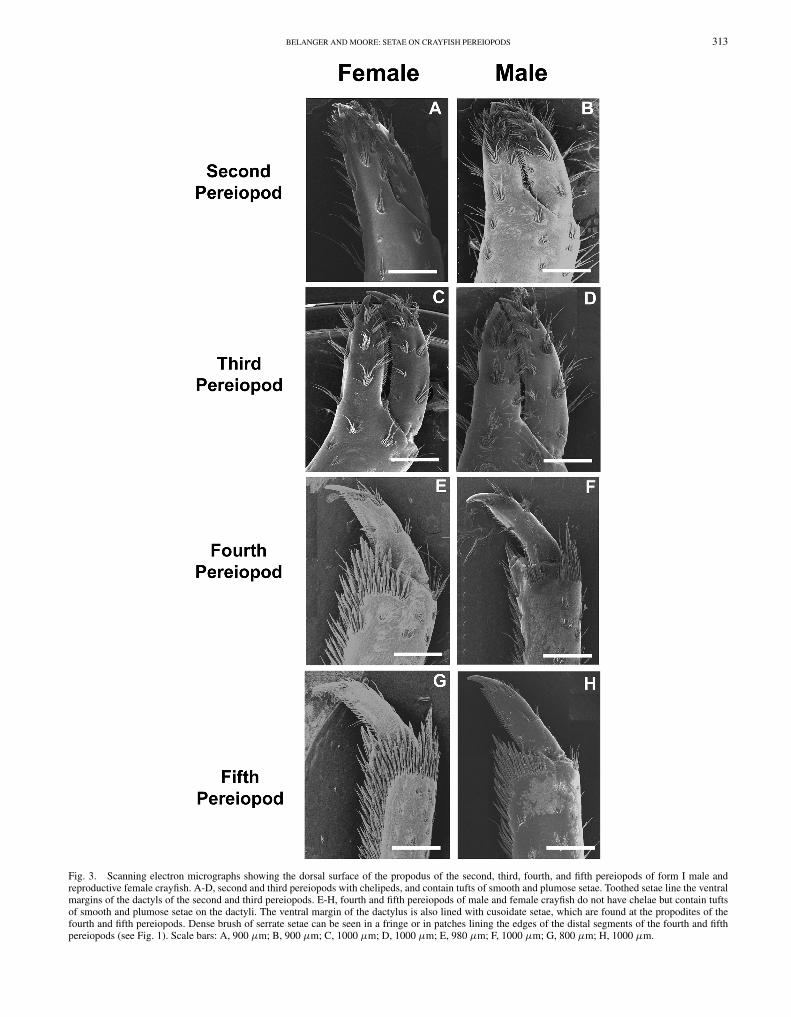
Scanning electron micrographs of the dorsal surface ofthe distal portion of the propodus of pereiopods (walkinglegs) of both reproductive (glair) female and reproductive(form I) male O. rusticus were examined (Fig. 3). Thesecond and third pereiopods of both females and malesare chelipeds containing tufts of setae (Fig. 3A-D). Thesetufts consist mainly of smooth setae distally but may alsocontain small numbers of plumose setae in tufts located moreproximally (Fig. 2A, B). Cuspidate setae line the ventral anddorsal margins of the dactyls and propodial projections ofpereiopods 2 and 3 of both sexes (Fig. 2C). Pereiopods 4and 5 are long and slender and do not contain chelae, butonly contain propodial projections (Fig. 3E-H). The marginsof propodial projections are lined with cuspidate setae inboth sexes (Fig. 2C). Smooth setae are found in the positionopposite of the projection of the propodus (Fig. 2D). In bothfemales and males, the propodi of pereiopods 4 and 5 containsome smooth and plumose setae. The propodi of pereiopods4 and 5 are lined with both smooth and serrate setae in boththe female and male crayfish (Fig. 2F, G).
Quantification of Setae Tufts
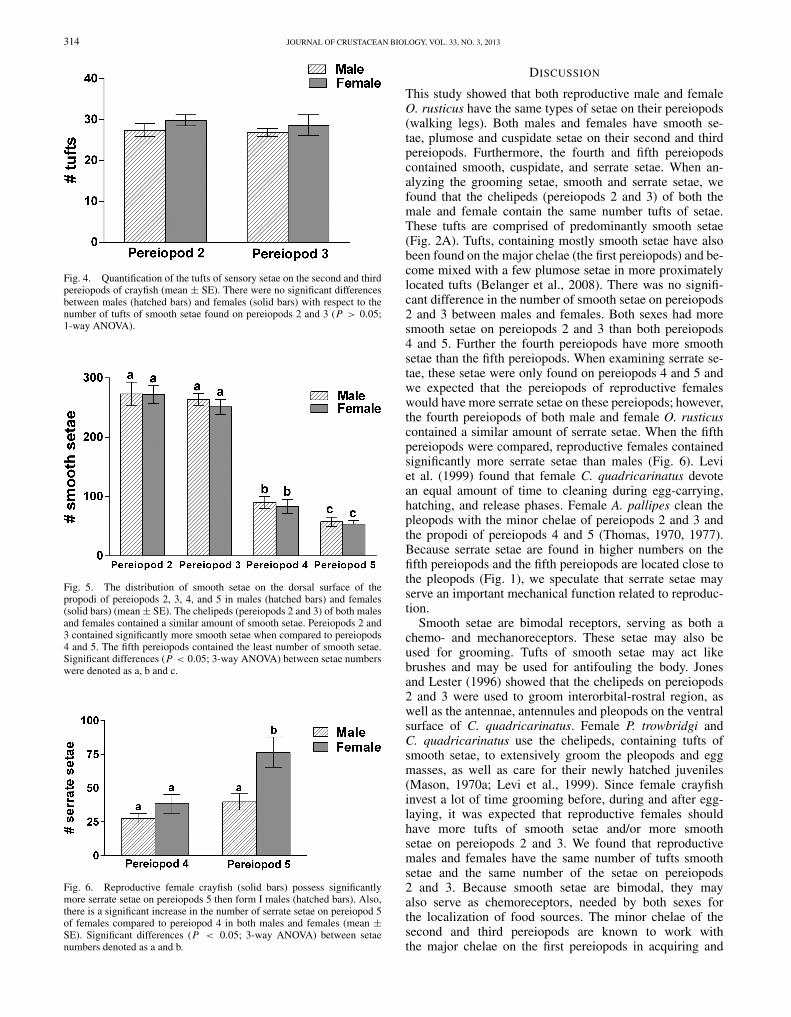
Setae tufts, containing predominately smooth setae, foundon pereiopods 2 and 3 of the female and male werequantified and compared. Females contain 29.8 ± 1.4 tuftsof setae on pereiopod 2 and 28.6 ± 2.5 on pereiopod 3(mean ± SE). Males have 27.4 ± 1.6 tufts of setae onpereiopods 2 and 27.0 ± 0.9 on pereiopod 3. There wasno significant difference in the number of tufts of setaefound on pereiopods 2 and 3 between females and males(F(3,16,0.05) = 0.61, P = 0.62; Fig. 4).
Quantification and Distribution of Smooth Setae
Overall there were differences in the distribution of setae onthe pereiopods on males and females (F(6,6,0.05) = 165.6,P < 0.0001; Figs. 5 and 6). Dense pockets of predominatelysmooth setae were found in tufts distally on the minorchelae of pereiopods 2 and 3 of both males and females.Female O. rusticus contain 272.4 ± 15.3 smooth setae onthe second pereiopod and 251.2 ± 12.7 smooth setae on thethird pereiopods. Males contain 273 ± 19.9 smooth setae onpereiopod 2 and 263.6 ± 10.5 smooth setae on pereiopod3. There was no difference in the number of smooth setaefound on pereiopods 2 and 3 when both the male and female
312 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 33, NO. 3, 2013
Fig. 2. A magnified representation of setae located on the distal portion of the pereiopods. A, chelae and propodus of the second and third pereiopodswith tufts of smooth setae (s) in dense amounts; B, tufts with both smooth and plumose setae (p) found on the second and third pereiopods, but locatedin the proximal regions of the propodus; C, cuspidate setae (c) are found lining the ventral margins of the dactyls on all of pereiopods; D, fourth and fifthpereiopods with smooth setae (s) on the propodi; E, serrate setae (se) also located on the fourth and fifth pereiopods; F, serrate setae can be found in a fringe;G, serrate setae located in tufts with smooth setae. Scale bars: A, 100 μm; B, 100 μm; C, 100 μm; D, 250 μm; E, 85 μm; F, 750 μm; G, 500 μm.
were compared (P < 0.05; Fig. 5). However, the secondand third pereiopods of both males and females containedsignificantly more smooth setae than pereiopods 4 and 5respectively (P < 0.05; Fig. 5). The fourth pereiopod ofthe female contained 83.4 ± 12.1 smooth setae while thefourth pereiopod of the male contained 90.2 ± 10.3 smoothsetae. There was no difference in the number of smooth setaefound on the fourth pereiopods when females and maleswere compared (P > 0.05; Fig. 5). The fifth pereiopods ofboth females and males contained significantly less smoothsetae that pereiopods 2, 3 and 4 (P < 0.05; Fig. 5). Femalescontained 53.6 ± 6.4 smooth setae and the male contained57.4 ± 8.0 smooth setae on the fifth pereiopods.
Quantification and Distribution of Serrate Setae
Serrate setae were found only on the fourth and fifthpereiopods of both male and female crayfish. There wasno difference in the number of serrate setae on the fourthpereiopod when male and female crayfish were compared(P > 0.05; Fig. 6). Females contained 38.4 ± 7.0 serratesetae on their fourth pereiopods while males contained27.6 ± 3.4 serrate setae. The fifth pereiopod of femalescontained significantly more serrate setae than males (P <
0.05; Fig. 6). Female O. rusticus had 76.4 ± 11.4 serratesetae while male crayfish had 39.8 ± 6.0 serrate setae onpereiopod 5.
BELANGER AND MOORE: SETAE ON CRAYFISH PEREIOPODS 313
Fig. 3. Scanning electron micrographs showing the dorsal surface of the propodus of the second, third, fourth, and fifth pereiopods of form I male andreproductive female crayfish. A-D, second and third pereiopods with chelipeds, and contain tufts of smooth and plumose setae. Toothed setae line the ventralmargins of the dactyls of the second and third pereiopods. E-H, fourth and fifth pereiopods of male and female crayfish do not have chelae but contain tuftsof smooth and plumose setae on the dactyli. The ventral margin of the dactylus is also lined with cusoidate setae, which are found at the propodites of thefourth and fifth pereiopods. Dense brush of serrate setae can be seen in a fringe or in patches lining the edges of the distal segments of the fourth and fifthpereiopods (see Fig. 1). Scale bars: A, 900 μm; B, 900 μm; C, 1000 μm; D, 1000 μm; E, 980 μm; F, 1000 μm; G, 800 μm; H, 1000 μm.
314 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 33, NO. 3, 2013
Fig. 4. Quantification of the tufts of sensory setae on the second and thirdpereiopods of crayfish (mean ± SE). There were no significant differencesbetween males (hatched bars) and females (solid bars) with respect to thenumber of tufts of smooth setae found on pereiopods 2 and 3 (P > 0.05;1-way ANOVA).
Fig. 5. The distribution of smooth setae on the dorsal surface of thepropodi of pereiopods 2, 3, 4, and 5 in males (hatched bars) and females(solid bars) (mean ± SE). The chelipeds (pereiopods 2 and 3) of both malesand females contained a similar amount of smooth setae. Pereiopods 2 and3 contained significantly more smooth setae when compared to pereiopods4 and 5. The fifth pereiopods contained the least number of smooth setae.Significant differences (P < 0.05; 3-way ANOVA) between setae numberswere denoted as a, b and c.
Fig. 6. Reproductive female crayfish (solid bars) possess significantlymore serrate setae on pereiopods 5 then form I males (hatched bars). Also,there is a significant increase in the number of serrate setae on pereiopod 5of females compared to pereiopod 4 in both males and females (mean ±SE). Significant differences (P < 0.05; 3-way ANOVA) between setaenumbers denoted as a and b.
DISCUSSION
This study showed that both reproductive male and femaleO. rusticus have the same types of setae on their pereiopods(walking legs). Both males and females have smooth se-tae, plumose and cuspidate setae on their second and thirdpereiopods. Furthermore, the fourth and fifth pereiopodscontained smooth, cuspidate, and serrate setae. When an-alyzing the grooming setae, smooth and serrate setae, wefound that the chelipeds (pereiopods 2 and 3) of both themale and female contain the same number tufts of setae.These tufts are comprised of predominantly smooth setae(Fig. 2A). Tufts, containing mostly smooth setae have alsobeen found on the major chelae (the first pereiopods) and be-come mixed with a few plumose setae in more proximatelylocated tufts (Belanger et al., 2008). There was no signifi-cant difference in the number of smooth setae on pereiopods2 and 3 between males and females. Both sexes had moresmooth setae on pereiopods 2 and 3 than both pereiopods4 and 5. Further the fourth pereiopods have more smoothsetae than the fifth pereiopods. When examining serrate se-tae, these setae were only found on pereiopods 4 and 5 andwe expected that the pereiopods of reproductive femaleswould have more serrate setae on these pereiopods; however,the fourth pereiopods of both male and female O. rusticuscontained a similar amount of serrate setae. When the fifthpereiopods were compared, reproductive females containedsignificantly more serrate setae than males (Fig. 6). Leviet al. (1999) found that female C. quadricarinatus devotean equal amount of time to cleaning during egg-carrying,hatching, and release phases. Female A. pallipes clean thepleopods with the minor chelae of pereiopods 2 and 3 andthe propodi of pereiopods 4 and 5 (Thomas, 1970, 1977).Because serrate setae are found in higher numbers on thefifth pereiopods and the fifth pereiopods are located close tothe pleopods (Fig. 1), we speculate that serrate setae mayserve an important mechanical function related to reproduc-tion.
Smooth setae are bimodal receptors, serving as both achemo- and mechanoreceptors. These setae may also beused for grooming. Tufts of smooth setae may act likebrushes and may be used for antifouling the body. Jonesand Lester (1996) showed that the chelipeds on pereiopods2 and 3 were used to groom interorbital-rostral region, aswell as the antennae, antennules and pleopods on the ventralsurface of C. quadricarinatus. Female P. trowbridgi andC. quadricarinatus use the chelipeds, containing tufts ofsmooth setae, to extensively groom the pleopods and eggmasses, as well as care for their newly hatched juveniles(Mason, 1970a; Levi et al., 1999). Since female crayfishinvest a lot of time grooming before, during and after egg-laying, it was expected that reproductive females shouldhave more tufts of smooth setae and/or more smoothsetae on pereiopods 2 and 3. We found that reproductivemales and females have the same number of tufts smoothsetae and the same number of the setae on pereiopods2 and 3. Because smooth setae are bimodal, they mayalso serve as chemoreceptors, needed by both sexes forthe localization of food sources. The minor chelae of thesecond and third pereiopods are known to work withthe major chelae on the first pereiopods in acquiring and

BELANGER AND MOORE: SETAE ON CRAYFISH PEREIOPODS 315
holding food items (Brown, 1995). Chemosensory setae,including smooth setae, are used for food odor detectionand orientation (Hodgson, 1958; Bauer et al., 1981; Altneret al., 1983; Moore et al., 1991; Corotto and O’Brien,2002; Keller et al., 2003). Electrophysiological studies showthat the second and third pereiopods of Orconectes limosus(Rafinesque, 1815), P. clarkii and C. bartonii sciotensisrespond to a variety of amino acids and pereiopod probingoccurs after the introduction of food odors in P. clarkii andO. rusticus (Hodgson, 1958; Bauer et al., 1981; Steele et al.,1999; Corotto and O’Brien, 2002). Because both males andfemales have the same amount and distribution of smoothsetae on their pereiopods, these setae must be important inboth sexes. Pereiopods 4 and 5 contain significantly lesssmooth setae than pereiopods 2 and 3 (Fig. 3). This decreasein amount smooth setae and the presence of serrate setaesuggests that the fourth and fifth pereiopods may be usedmore for grooming than for odor detection.
The fourth and fifth pereiopods are anatomically dis-tinct when compared to the second and third pereiopods.Pereiopods 4 and 5 are not chelipeds and contain only adactylus (Holdich, 2002). We found that the fourth andfifth pereiopods are lined with predominantly serrate se-tae. When reproductive male and female crayfish were com-pared, we found that there was no difference in the numberof serrate setae on the fourth pereiopods. However, repro-ductive female crayfish have significantly more serrate se-tae on their fifth pereiopods when compared to reproduc-tive males (Fig. 6). Like smooth setae, serrate setae havealso been found to be chemically and mechanically sensitive(Derby, 1982). Furthermore, the presence of serrate setae onpereiopods 4 and 5 suggests a grooming function (Derby,1982). Male and female C. quadricarinatus, P. trowbridgi,A. pallipes and P. clarkii have been shown to use the brushesof serrate setae on pereiopods 4 and 5 are used for gen-eral body grooming and to scrape the lateral surfaces of thecephalothorax (Mason, 1970a; Thomas, 1970; Bauer, 1981,1989; Jones and Lester, 1996; Batang and Suzuki, 2000;Holdich, 2002). When the pereiopods of C. quadricarinatuswere removed, significant increases in infestation of temno-cephalid occurred (Jones and Lester, 1996). In addition tobeating the pleopods and fanning the pleon, females groom,preen, and roll the eggs and embryos with the chelipeds onpereiopods 2 and 3 and setae brushes on pereiopods 4 and 5to keep them clean (Bauer, 1989; Holdich, 2002). Levi et al.(1999) showed that reproductive female C. quadricarinatususe the chelipeds of pereiopods 2 and 3 and the setae of thepropodus of pereiopods 4 and 5 to groom the pleopods be-fore, during and after egg laying. Because serrate setae arebimodal, the combined grooming and chemosensory func-tion of serrate setae may be important for distinguishing eggsfrom fouling material (Derby, 1982). Reproductive femalecrayfish are known to clean extensively as part of the fe-male maternal behavior. When we compared reproductivemales and females, we found that reproductive females hadsignificantly more serrate setae on their fifth pereiopods.Brushes of serrate setae are used for general body groomingand preening (Mason, 1970a; Thomas, 1970, 1977; Bauer,2002), but we suggest that the increase in these setae on thefifth pereiopods of reproductive females is a dimorphic trait,
used for cleaning of the pleopods containing eggs and/or ju-veniles. The fifth pereiopods are located in close proximityto the pleopods where the eggs are deposited and close tothe juveniles when they hatch (Fig. 1). As freshwater cray-fish are among the few decapods with extended parental care(Hazlett, 1983), they use their pereiopods and pleopods toclean and aerate the eggs and they clean consistently duringall three phases of reproduction (egg-carrying, hatching andrelease phases). A sexually dimorphic increase in serrate se-tae on the fifth pereiopods of reproductive females indicatesthat they serve a reproductive function and may be exten-sively used for grooming eggs and brood care.
Overall, the results of this study suggest that reproductiveO. rusticus have a similar distribution and amount of groom-ing setae (smooth and serrate setae) with the exception ofthe number of serrate setae found on the fifth pereiopods.Smooth setae are arranged in tufts, used as brushes forgrooming (Thomas, 1970; Hatt, 1986; Levi et al., 1999;Belanger et al., 2008). The minor chelae containing thesetufts are used for egg turning and cleaning during repro-duction in crayfish (Mason, 1970a; Jones and Lester, 1996;Levi et al., 1999; Holdich, 2002); however, both sexes mayuse smooth setae for general body grooming. Pereiopods2 through 5 contain smooth setae and pereiopods 2 and 3have significantly more smooth setae than pereiopods 4 and5. Pereiopods 2 and 3 are chelipeds that contain contactchemoreceptors, which are important for the detection offood odors and distance orientation (Hodgson, 1958; Baueret al., 1981; Altner et al., 1983; Moore et al., 1991; Corottoand O’Brien, 2002; Keller et al., 2003). Detecting prey isimportant for survival in both sexes of crayfish and thereforethe use of tufts of smooth setae for grooming may be a sec-ondary function. Further behavioral observations comparingmale, and reproductive and non-reproductive female O. rus-ticus will allow us to quantify time spent performing varioustasks with the pereiopods.
ACKNOWLEDGEMENTS
The authors are indebted to Dr. M. Cayer at the Electron MicroscopeFacility at Bowling Green State University for her generosity and expertiseas well as co-workers in the Laboratory for Sensory Ecology for assistancein crayfish collection. We are grateful for the assistance of Dr. A.Garm (University of Copenhagen) for email communications on setaemorphology and identification. The authors would also like to thank theUniversity of Detroit Mercy and the J. P. Scott Center for Neuroscience,Mind & Behavior at Bowling Green State University for grants to R.M.B.and N.S.F. (IBN No. 0131320) for a grant to P.A.M.
REFERENCES
Altner, H., H. Hatt, and I. Altner. 1983. Structural and functional propertiesof the mechanoreceptors and chemoreceptors in the anterior esophagealsensilla of the crayfish, Astacus astacus. Cell and Tissue Research 228:357-374.
Ameyaw-Akumfi, C. 1977. Feeding chemoreceptor sites in the crayfishProcambarus clarkii (Girard). Crustaceana 33: 259-264.
Batang, Z. B., and H. Suzuki. 1999. Gill-cleaning mechanisms of the mudlobster Thalassina anomala (Decapoda: Thalassinidea: Thalassinidae).Journal of Crustacean Biology 19: 671-683.
, and . 2000. Gill structure and gill-cleaning mechanismsof the redclaw crayfish Cherax quadricarinatus (Decapoda, Astacidea,Parastacidae). Journal of Crustacean Biology 20: 699-714.
Bauer, R. T. 1981. Grooming behavior and morphology in the DecapodCrustacea. Journal of Crustacean Biology 1: 153-173.

316 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 33, NO. 3, 2013
. 1989. Decapod crustacean grooming: functional morphology,adaptive value, and phylogenetic significance, pp. 49-73. In, B. E. Fel-genhauer, L. Watling, and A. A. Thistle (eds.), Functional Morphologyof Feeding and Grooming in Crustacea. Crustacean Issues 6. Balkema,Rotterdam.
. 1998. Gill-cleaning mechanisms of the crayfish Procambarusclarkii (Astacidea: Cambaridae): experimental testing of setobranchfunction. Invertebrate Biology 117: 129-143.
. 2002. The ineffectiveness of grooming in prevention of bodyfouling in the red swamp crayfish, Procambarus clarkii. Aquaculture208: 39-49.
Bauer, U., J. Dudel, and H. Hatt. 1981. Characteristics of single chemore-ceptive units sensitive to amino acids and related substances in the cray-fish leg. Journal of Comparative Physiology 144: 67-74.
Belanger, R., X. Ren, K. McDowell, S. Chang, P. Moore, and B. Zielinski.2008. Sensory setae on the major chelae of male crayfish, Orconectesrusticus (Decapoda, Astacidae): impact of reproductive state on functionand distribution. Journal of Crustacean Biology 28: 27-36.
Bovbjerg, R. V. 1956. Some factors affecting aggressive behavior incrayfish. Physiological Zoology 29: 127-136.
Brown, P. B. 1995. Physiological adaptations in the gastrointestinal tract ofcrayfish. American Zoologist 35: 20-27.
Cooke, I. R. C., and D. L. Macmillan. 1985. Further studies of crayfishescape behaviour I. The role of the appendages and the stereotyped natureof non-giant escape swimming. Journal of Experimental Biology 118:351-365.
Corotto, F. S., and M. R. O’Brien. 2002. Chemosensory stimuli for thewalking legs of the crayfish Procambarus clarkii. Journal of ChemicalEcology 28: 1117-1130.
Crocker, D. W., and D. W. Barr. 1968. Handbook of the Crayfish of Ontario.University of Toronto Press, Toronto, ON.
Derby, C. D. 1982. Structure and function of cuticular sensilla of the lobsterHomarus americanus. Journal of Crustacean Biology 2: 1-21.
. 1989. Physiology of sensory neurons in morphologically identifiedcuticular sensilla of crustaceans, pp. 27-47. In, B. E. Felgenhauer,L. Watling, and A. A. Thistle (eds.), Functional Morphology of Feedingand Grooming in Crustacea. Crustacean Issues 6. Balkema, Rotterdam.
Garm, A. 2004. Revising the definition of the crustacean seta and setalclassification systems based on examinations of the mouthpart setae ofseven species of decapods. Zoological Journal of the Linnean Society142: 233-252.
, and L. Watling. 2013. The crustacean integument: setae, setulesand other ornamentation, pp. 160-190. In, L. Watling and M. Thiel (eds.),Functional Morphology and Diversity. Oxford University Press, NewYork, NY.
Girard, C. 1852. A revision of the North American Astici, with observationson their habits and geographic distribution. Proceedings of the Academyof Natural Sciences of Philadelphia 6: 87-91.
Hagen, H. A. 1870. Monograph of the North American Astacidae. Illus-trated Catalogue of the Museum of Comparative Zoology at Harvard Col-lege 3: i-viii, 1-109.
Hatt, H. 1986. Responses of a bimodal neuron (chemo- and vibration-sensitive) on the walking legs of the crayfish. Journal of ComparativePhysiology A 159: 611-617.
Hazlett, B. A. 1983. Parental behavior in decapod crustacean, pp.171-193.In, S. Rebach and D. W. Dunham (eds.), Studies in Adaptation. TheBehavior of Higher Crustacea. John Wiley & Sons, New York, NY.
. 1999. Responses to multiple chemical cues by the crayfishOrconectes virilis. Behaviour 136: 161-177.
, and D. R. Schoolmaster. 1998. Responses of cambarid crayfish topredator odor. Journal of Chemical Ecology 24: 1757-1770.
Hodgson, E. S. 1958. Electrophysiological studies of arthropod chemore-ception, 3. Chemoreceptors of terrestrial and freshwater arthropods. Bio-logical Bulletin 115: 114-125.
Holdich, D. M. 2002. Background and functional morphology, pp. 3-29. In,D. M. Holdich (ed.), Biology of Freshwater Crayfish. Blackwell Science,Oxford.
Jones, T. C., and R. J. G. Lester. 1996. Factors influencing populations ofthe ectosymbiont Diceratocephala boschmai (Platyhelminthes: Temno-
cephalida), on the redclaw crayfish Cherax quadricarinatus maintainedunder laboratory conditions. Aquaculture 143: 233-243.
Keller, T. A., I. Powell, and M. J. Weissburg. 2003. Identifying the role ofolfactory appendages in chemically mediated orientation of blue crabs.Marine Ecology Progress Series 261: 217-231.
Lavalli, K. L., and J. R. Factor. 1995. The feeding appendages, pp. 349-393.In, J. R. Factor (ed.), The Biology of the Lobster, Homarus americanus.Academic Press, New York, NY.
Laverack, M. S. 1988. The diversity of chemoreceptors, pp. 287-317. In,J. Atema, R. R. Fay, A. N. Popper, and W. N. Tavolga (eds.), SensoryBiology of Aquatic Animals. Springer, New York, NY.
Lereboullet, A. 1858. Description des deux nouvelles especies d’écrevissesde nos rivieres. Memoires de la Societe des Sciences Naturelles 5: 1-11.
Levi, T., A. Barki, G. Hulata, and I. Karplus. 1999. Mother-offspringrelationships in the red claw crayfish Cherax quadricarinatus. Journalof Crustacean Biology 19: 477-484.
Mason, J. C. 1970a. Egg-laying in the western North American Crayfish,Pacifastacus trowbridgii (Stimpson) (Decapoda, Astacidae). Crustaceana19: 37-44.
. 1970b. Maternal-offspring behavior of the crayfish, Pacifastacustrowbridgi (Stimpson). American Midland Naturalist 84: 463-473.
Moore, P. A., N. Scholz, and J. Atema. 1991. Chemical orientation ofthe lobster, Homarus americanus in a turbulent odor plume. Journal ofChemical Ecology 17: 1293-1307.
Nation, J. L. 1983. A new method using hexamethyldisilazane for prepara-tion of soft insect tissues for scanning electron microscopy. Stain Tech-nology 58: 347-351.
Nyström, P., and J. R. Pérez. 1998. Crayfish predation on the common pondsnail (Lymnaea stagnalis): the effect of habitat complexity and snail sizeon foraging efficiency. Hydrobiologia 368: 201-208.
Pond, C. M. 1975. The role of the “walking legs” in aquatic and terrestriallocomotion of the crayfish Austropotamobius pallipes (Lereboullet).Journal of Experimental Biology 62: 447-454.
Rafinesque, C. S. 1815. Analyse de la Nature ou Tableau de l’Univers.Palerme.
Rhoades, R. 1944. The crayfishes of Kentucky, with notes on variation,distribution, and descriptions of new species and subspecies. AmericanMidland Naturalist 31: 111-149.
Schrank, F. 1803. Fauna boica. Durchgedachte geschichte der in Baierneinheimischen und zahmen Thiere. Steischen buchandlung. Nürnberg. 3,vii+. 272 pp.
Shelton, R. G. J., and M. S. Laverack. 1968. Observations on a redescribedcrustacean cuticular sense organ. Comparative Biochemistry and Physi-ology 25: 1049-1059.
Steele, C., C. Skinner, C. Steele, P. Alberstadt, and C. Mathewson.1999. Organization of chemically activated food search behavior inProcambarus clarkii Girard and Orconectes rusticus Girard crayfishes.Biological Bulletin 196: 295-302.
Stein, R. A. 1976. Sexual dimorphism in crayfish chelae: functional signifi-cance linked to reproductive activities. Canadian Journal of Zoology 54:220-227.
Stimpson, W. 1857. The Crustacea and Echinodermata of the Pacific shoresof North America. Boston Journal of Natural History 6: 444-532.
Thomas, W. J. 1970. The setae of Austropotamobius pallipes (Crustacea:Astacidae). Journal of Zoology (London) 160: 91-142.
. 1977. The biting edges of the chelae and pereiopods of Austropota-mobius pallipes. Cellular and Molecular Life Sciences 33: 1037-1039.
. 1981. Dimorphism in the British crayfish Austropotamobiuspallipes (Lereboullet), pp. 12-17. In, C. R. Goldman (ed.), FreshwaterCrayfish V. AVI, Westport, CT.
Vogt, G., and L. Tolley. 2004. Brood care in freshwater crayfish andrelationship with offspring’s sensory deficiencies. Journal of Morphology262: 566-582.
von Martens, E. 1868. Ueber einige ostasiatische Süsswasserthiere. Archivfür Naturgeschichte 34: 1-67, pl. 1.
RECEIVED: 21 September 2012.ACCEPTED: 11 January 2013.AVAILABLE ONLINE: 23 February 2013.
Related Documents











