1 A CENTRAL ROLE FOR NEURONAL AMP ACTIVATED PROTEIN KINASE (AMPK) IN CANCER-INDUCED ANOREXIA Short title: AMPK activation reverses anorexia in rats. Eduardo R. Ropelle, José R. Pauli, Karina G. Zecchin, Mirian Ueno, Cláudio T. de Souza, Joseane Morari, Marcel C. Faria, Lício A. Velloso, Mario J. A. Saad, José B. C. Carvalheira. Department of Internal Medicine, FCM, State University of Campinas (UNICAMP), Campinas, SP, Brazil. Please address correspondence to: José B. C. Carvalheira, MD. Department of internal medicine FCM - State University of Campinas (UNICAMP) 13081-970 - Campinas, SP, Brazil. Fax: + 55 19 3521-8950 E-mail: [email protected] Disclosure statement: The authors have nothing to disclose. Endocrinology. First published ahead of print August 23, 2007 as doi:10.1210/en.2007-0381 Copyright (C) 2007 by The Endocrine Society

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1
A CENTRAL ROLE FOR NEURONAL AMP ACTIVATED PROTEIN KINASE (AMPK) IN CANCER-INDUCED ANOREXIA
Short title: AMPK activation reverses anorexia in rats.
Eduardo R. Ropelle, José R. Pauli, Karina G. Zecchin, Mirian Ueno, Cláudio T. de Souza,
Joseane Morari, Marcel C. Faria, Lício A. Velloso, Mario J. A. Saad, José B. C.
Carvalheira.
Department of Internal Medicine, FCM, State University of Campinas (UNICAMP),
Campinas, SP, Brazil.
Please address correspondence to:
José B. C. Carvalheira, MD.
Department of internal medicine
FCM - State University of Campinas (UNICAMP)
13081-970 - Campinas, SP, Brazil.
Fax: + 55 19 3521-8950
E-mail: [email protected]
Disclosure statement: The authors have nothing to disclose.
Endocrinology. First published ahead of print August 23, 2007 as doi:10.1210/en.2007-0381
Copyright (C) 2007 by The Endocrine Society

2
ABSTRACT
The pathogenesis of cancer anorexia is multifactorial and is associated with
disturbances of the central physiological mechanisms controlling food intake. However the
neurochemical mechanisms responsible for cancer induced-anorexia are unclear. Here, we
show that chronic infusion of 5-amino-4imidazolecarboxamide-riboside (AICAR) into the
third cerebral ventricle and a chronic peripheral injection of 2 deoxy-D-glucose (2DG)
promotes hypothalamic AMPK activation, increases food intake and prolongs the survival
of anorexic tumor bearing (TB) rats. In parallel, the pharmacological activation of
hypothalamic AMPK in TB animals, markedly reduced the hypothalamic production of
inducible nitric oxide synthase (iNOS), interleukin-1β (IL-1β ) and tumor necrosis factor-
alpha (TNF-α) and modulated the expression of POMC, a hypothalamic neuropeptide that
is involved in the control of energy homeostasis. Furthermore, the daily oral and i.c.v.
treatment with biguanide anti diabetic drug metformin also induced AMPK
phosphorylation in the central nervous system and increased food intake and life span in
anorexic TB rats. Collectively, the findings of this study suggest that hypothalamic AMPK
activation reverses cancer anorexia by inhibiting the production of proinflammatory
molecules and controlling the neuropeptide expression in the hypothalamus, reflecting in a
prolonged life span in TB rats. Thus, our data indicate that hypothalamic AMPK activation
presents an attractive opportunity for the treatment of cancer-induced anorexia.

3
INTRODUCTION
Approximately 50% of cancer patients report abnormalities in eating behavior upon
diagnosis (1). Anorexia and reduced energy intake also negatively affect quality of life,
which is an important endpoint in the management of patients with cancer and in the design
of clinical trials (2). Accumulating evidences indicate that cancer anorexia is multifactorial
in its pathogenesis, and most of the hypothalamic neuronal signaling pathways modulating
energy intake are likely to be involved (3-6).
Tumor growth is frequently associated with the development of anorexia. Evidence
suggests that multiple proinflammatory-cytokines [Interleukin-1β (IL-1β), Interleukin-6
(IL-6) and tumor necrosis factor-alpha (TNF-α)] in the central nervous system are
mediators of anorexia in tumor-bearing hosts (7). Several studies have sought to explain
how peripheral cytokines enter the brain and affect neuronal pathways involved in food
intake regulation (8, 9). This could be done by persistent stimulation of anorexigenic
neuropeptides, such as corticotropin-releasing factor, as well as by inhibition of the
neuropeptide Y (NPY) orexigenic network (10, 11).
Analogous to leptin, pro-opiomelanocortin (POMC)-containing neurons in the ARH
also contain receptors to cytokines such as IL-1β and leukemia inhibitory factor, and the
injections of each of these molecules into the third ventricle up-regulates POMC, and
decreases feeding behavior (12, 13). In addition, the high levels of IL-1β, IL-6, and TNF-α
interfere with NPY release and neurotransport to a greater degree than gene expression (7,
14). Some studies have reported that hypothalamic mRNA and peptide levels of orexigen
NPY are decreased in different models of anorectic tumor-bearing rats (11, 15).

4
Multiple hypothalamic neuronal signaling pathways and the cross-talk between
these pathways are involved in the control of energy intake (16, 17). Recently,
hypothalamic AMP-activated protein kinase (AMPK) signaling has become an important
focus of interest in the control of food intake. It has been proposed that changes in neuronal
energy status affect AMPK phosphorylation and neuropeptide expression, leading to
changes in food intake (18-20). AMPK is the central component of a protein kinase cascade
that plays a key role in the regulation of energy control. AMPK is activated in response to
ATP depletion, which causes an increase in the AMP:ATP ratio (21). AMPK activation
occurs through the phosphorylation of a specific threonine residue (Thr-172), catalyzed by
an upstream kinase that has been identified as LKB1 (22). In addition, recent studies
identified calmodulin-dependent protein kinase kinase (CaMKK) as an upstream kinase of
AMPK (23-25).
Growing experimental evidences suggests the anti-inflammatory effects of AMPK;
since, the pharmacological activation of AMPK with 5-amino-4-imidazole carboxamide
ribose (AICAR), blocks LPS-induced inflammatory processes by blocking the expression
of proinflammatory cytokines, iNOS, COX-2 and MnSOD genes in astroglial cells (26, 27).�
Furthermore, the activation of AMPK inhibits LPS-induced expression of�proinflammatory
cytokines in primary rat astrocytes, microglia and peritoneal macrophages (26, 28).
Although growing evidence demonstrates the importance of AMPK activity in regulating
the inflammatory process (in vitro and in vivo) (26) and food intake and body weight in
different models of animals (18, 19, 29), the role of neuronal AMPK activity in cancer-
induced anorexia is not known.
The present data demonstrate that different pharmacological activators of AMPK
signaling, including oral administration of the anti diabetic drug metformin, increase

5
AMPK phosphorylation in the central nervous system, reverse cancer anorexia and increase
the life span in TB rats through the inhibition of the production of proinflammatory
molecules and by controlling neuropeptide expression in the hypothalamus.

6
MATERIALS AND METHODS
Reagents for sodium dodecyl sulfate polyacrylamide gel electrophoresis and
immunoblotting were from Bio-Rad. Tris[hydroxymethyl]amino-methane (Tris), aprotinin,
ATP, dithiothreitol, phenylmethylsulfonyl fluoride, Triton X-100, Tween 20, glycerol, and
bovine serum albumin (fraction V) were from Sigma Chemical Co. (St.Louis, MO).
Nitrocellulose paper (BA85, 0.2mm) was from Schleicher & Schuell (Keene, NH).
Ketamine hydrochloride was from Cristália (Itapira SP, Brazil). Antiphospho-[Ser79] ACC
was from Upstate Biotechnology (Charlottesville, VA, USA). Anti-ACC, anti-iNOS, anti-
α−tubulin, anti-IL-1β and anti-TNF-α were from Santa Cruz Biotechnology, Inc.
Antiphospho-[Thr172] AMPK and anti-AMPK were from Cell Signaling Technology
(Beverly, MA, USA). 5-amino-4imidazolecarboxamide-riboside (AICAR), 2-deoxy-D-
glucose (2DG) and metformin (1,1-dimethylbiguanide hydrochloride) were obtained
Sigma- Aldrich, (St. Louis, MO, USA) and were dissolved in sterile saline. The doses
administrated in each experimental group are given below.
Animals and Surgical Procedures
Adult male Wistar rats (250–300 g) were used in all of the experiments in
accordance with the guidelines of the Brazilian College for Animal Experimentation
(COBEA); the ethics committee at the State University of Campinas approved experiments.
Room temperature was maintained (28±1 °C), and rats were housed in individual cages,
subjected to a standard light-dark cycle (6:00 a.m. to 6:00 p.m./6:00 p.m. to 6:00 a.m.), and
provided with standard rodent chow and water ad libitum. After an overnight fast, the rats
were anesthetized with ketamine hydrochloride (100 mg/kg, i.p.) and positioned on a
Stoelting stereotaxic apparatus. The implantation of an intracerebroventricular (i.c.v.)

7
catheter into the third ventricle has been previously described (16). After five days recovery
period, cannula placement was confirmed by a positive drinking response after
administration of angiotensin II (40 ng/2 μl), and animals that did not drink 5 ml of water
within 15 minutes after treatment were not included in the experiments.
The Walker 256 tumor cell line (originally obtained from the Christ Hospital Line,
National Cancer Institute Bank, Cambridge, USA) is currently maintained frozen in liquid
nitrogen in our laboratory. Walker 256 tumor cells were obtained from the ascitic fluid of
the peritoneal cavity, 5 days after the intraperitoneal injection of 20 × 106 carcinoma cells.
After cell harvesting, the percentage of viable cells was determined by using 1% Trypan
blue solution in a Neubauer chamber. Tumor cells (approximately 2 x 106 cells in 1 ml
saline solution) were then subcutaneously injected in the right flank after the surgical
implantation of the i.c.v. cannula.
Definition of cancer anorexia and treatments
The metabolic and feeding experiments were performed 1 week after complete
recovery from the stereotaxic surgery according to the protocol depicted in figure 1. Each
animal’s individual baseline 24 h food intake was defined as the average daily food intake
over a period of 3 consecutives days. Subsequent food intake data are expressed as
individual percentage, baseline daily food intake. In TB animals, cancer anorexia was
defined as a single value below 70% of baseline occurring after a steady decline of at least
3 d duration.
When criteria for anorexia had been met, TB animals were daily treated with: i.p.
injection of vehicle or 2DG (500mg/kg); i.c.v. injection (3 μl bolus injection) of AICAR
(0.5 or 2.0 mM), metformin (1mM), anti-IL1β antibody, anti-TNF-α antibody or vehicle; or

8
oral gavage with metformin (500 mg/kg). These treatments were performed during 4 days
prior to Western Blott and cumulative food intake analysis and the daily chronic treatment
were conducted for survival curve detemination. In preliminary experiments we determined
water intake in TB animals that received i.c.v. AICAR, i.p. 2DG and oral and i.c.v.
metformin. Water intake was not altered by these treatments compared to the respective
TB+saline group.
Physiological and metabolic parameters
After six hours of fasting, rats were submitted to an insulin tolerance test (ITT;
1U/kg body weight of insulin). After 30 minutes of i.c.v. AICAR, 180 minutes of i.p. 2DG
and 180 minutes of oral metformin administration, the rats were injected with insulin and
the blood samples were collected at 0, 4, 8, 12 and 16 minutes from the tail for serum
glucose determination. The rate constant for plasma glucose disappearance (Kitt) was
calculated using the formula 0.693/ (t1/2). The plasma glucose t1/2 was calculated from the
slope of last square analysis of the plasma glucose concentration during the linear phase of
decline (30). Plasma glucose was determined using a glucose meter (Roche Diagnostic,
Rotkreuze, Switzerland) and RIA was employed to measure serum insulin, according to a
previous description (31). Leptin concentrations were determined using a commercially
available Enzyme Linked Immuno Sorbent Assay (ELISA) kit (Crystal Chem Inc, Chicago,
IL). Rats were killed after 4-d of the different treatments and the body, tumor weight, and
spleen index were obtained. The spleen index was calculated as follows (32):
spleen wt/body wt TB rats without tumorspleen wt/body wt normal rat
Proinflammatory-cytokine determination.
Spleen index =

9
Cytokine levels (IL1β and TNF-α) were determined in samples of hypothalamic
protein extracts (2.0 mg/ml) as well as serum TNF-α using ELISA kits (Pierce Endogen,
Rockford, IL, USA), according to manufacturer's instructions.
Real-time quantitative PCR
NPY and POMC neuropeptides were measured in the hypothalami of controls,
treated and untreated TB animals rats under fasting conditions (12 hours). AICAR (2.0
mM) or vehicle were injected 6 hours before the hypothamic extraction. Six rats were
anesthetized, decapitated, and their hypothalamus immediately dissected on ice and snap
frozen in liquid nitrogen. Frozen samples were immersed in TRIzol and homogenized for
30 sec at maximum speed. Total RNA was isolated according to the manufacturer’s
guidelines and quantified by a spectrophotometer. The integrity of RNA was verified on
ethidium bromide-stained 1% agarose gel, and the fluorescence intensity ratio of 28S/18S
rRNA was determined (Eagle Eye; Stratagene, La Jolla, CA). Only samples that met the
criteria of quality (both 260/280 nm and 28S/18S > 1.8) were included in the experiments.
RNA samples were DNAse treated at 37ºC for 30 min and the enzyme inactivated at 65°C
for 10 min. All samples were column purified, and 1.6 μg of total RNA were reverse
transcribed at the same time using a master mix containing oligo (dT) primer and
SuperScript III in a final volume of 20 μl. Intron-skipping primers were optimized for
mRNA encoding NPY, POMC and Gapdh and are listed below: NPY: forward, 5'
accaggcagagatatggcaaga 3'; reverse, 5' ggacattttctgtgctttctctcatta 3'; POMC: forward,
5'cgctcctactctatggagcactt 3'; reverse, 5' tcacctaccagctccctcttg 3'; Gapdh: forward, 5'
aacgaccccttcattgac 3'; reverse, 5' tccacgacatactcagcac 3'. The specificities of the products
were confirmed by BLAST analyses and electrophoresis on an ethidium bromide-stained

10
3% agarose gel. Real-time PCR analysis of gene expression was carried out in an ABI
Prism 7700 sequence detection system (Applied Biosystems). The optimal concentration of
cDNA and primers, as well as the maximum efficiency of amplification were obtained
through five-point, 2-fold dilution curve analysis for each gene. Each PCR contained 2.5 or
5.0 ng of reverse-transcribed RNA (depending on the gene), 200 nM of each specific
primer, SYBR Green PCR master mix, and RNase free water to a 20 μl final volume.
cDNA samples from hypothalamus were processed at the same time in triplicate for each
gene and the negative controls included for each primer. The PCR conditions were 10 min
at 95° C, followed by 40 cycles at 95°C for 15 sec and 60° C for 60 sec, and a melting step
(dissociation curve) was performed after each run to further confirm the specificity of the
products and the absence of primer dimers. Real-time data were analyzed using the
Sequence Detector System 1.7 (Applied Biosystems).
Immunoblotting
Four days after the respective treatments, rats were anaesthetized with sodium
amobarbital (15 mg/kg body weight, i.p.) and used as soon as anesthesia was assured by the
loss of pedal and corneal reflexes. The rats were killed and the basal diencephalon,
including the preoptic area and the hypothalamus, were removed at the time-points
indicated, minced coarsely, and homogenized immediately in solubilization buffer
containing (mM) 100 Tris (pH 7.6), 1% Triton X-100, 10 Na3VO4, 100 NaF, 10 Na4P2O7, 4
EDTA, 150 NaCl, 0.1 mg aprotinin and 35 mg phenylmethylsulfonyl fluoride/mL, using a
polytron PTA 20S generator (Model PT 10/35, Brinkmann Instruments, Westbury, NY)
operated at maximum speed for 30 seconds and clarified by centrifugation. Hypothalami
(200 μg of protein) were used for immunoblotting followed by Western blot analysis with

11
the indicated antibodies. Blots were exposed to pre-flashed Kodak XAR film with Cronex
Lightning Plus intensifying screens at -80 °C for 12–48 h. Band intensities were quantitated
by optical densitometry (Scion Image software, ScionCorp).
Statistical Analysis
The survival curves were estimated using Kaplan-Meier’s estimates, and curves
were compared using the log-rank test and the level of significance was set at p < 0.001.
Where appropriate, results are expressed as the means ± S.E.M. accompanied by the
indicated number of rats used in experiments. Comparisons among groups were made using
parametric one-way ANOVA; where F ratios were significant, further comparisons were
made using the Bonferroni test. The level of significance was set at p < 0.05. The results of
blots are presented as direct comparisons of bands in autoradiographs and quantified by
densitometry.

12
RESULTS
Physiological and metabolic parameters
Table 1 shows comparative data for controls and tumor bearing (TB) groups with
respective treatments. As previously shown (33, 34), four days after the criteria of anorexia
had been met, all the TB groups showed weight loss, reduction in the 6 hour-fasting serum
insulin and leptin levels and insulin resistance when compared to age-matched controls.
When the tumor weights were excluded, treatments with intracerebroventricular (i.c.v.)
AICAR, intraperitoneal (i.p.) 2DG and oral metformin were observed to increase the body
weight, when compared to saline treatment in TB animals. All the TB groups presented an
increased serum TNF-α and spleen index and, four days after the beginning of the different
treatments, there was a significant reduction in serum TNF-α and splenomegaly compared
to the TB+saline group. In addition, AICAR, 2DG and metformin increased the 6 hour-
fasting serum insulin and leptin and improved insulin sensitivity, compared with saline
treatment in TB rats. No significant variations were found in 6 hour-fasting serum glucose
levels between the groups and the tumor weights were similar between TB groups.
Central infusion of AICAR, peripheral injection of 2DG or oral administration of
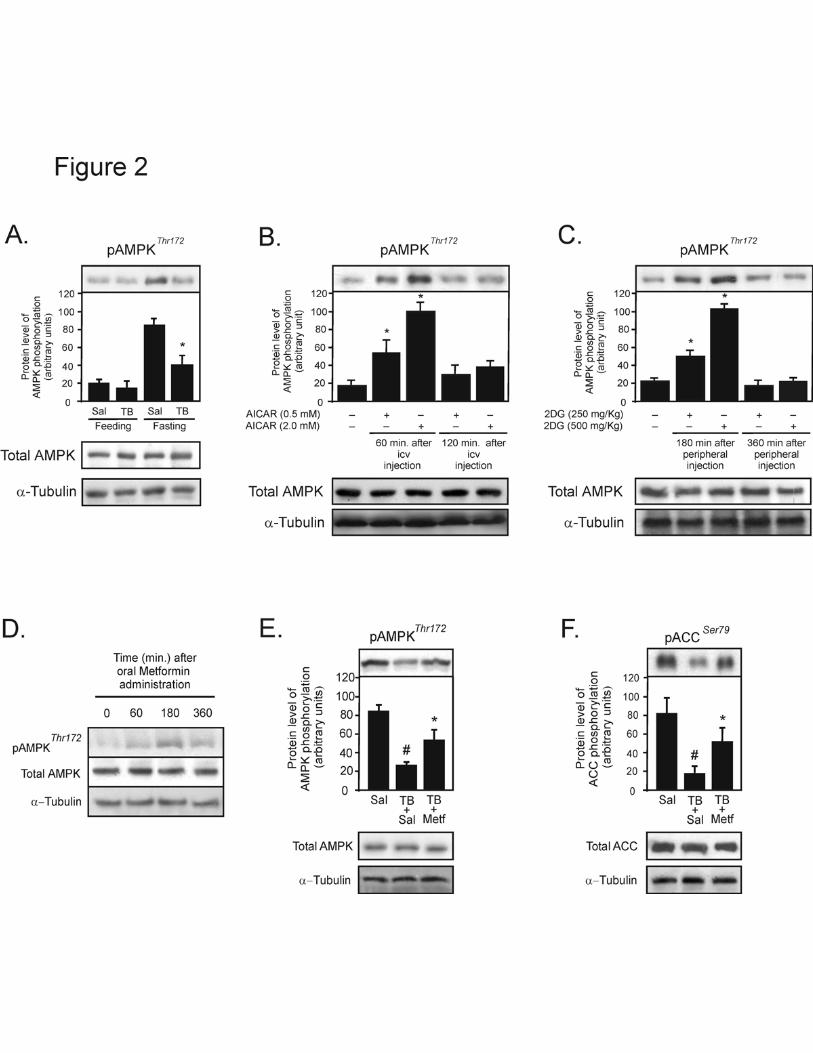
metformin increase hypothalamic AMPK phosphorylation in TB rats.
We first examined the hypothalamic AMPK phosphorylation in tumor bearing (TB)
and control rats under feeding and 12 hour fasting states. Under feeding conditions, AMPK
phosphorylation was similar in the hypothalamus of TB and control rats; however, in the 12
hour fasting state the hypothalamic AMPK phosphorylation in the TB animals was reduced
by 2.1- fold when compared to the control group (Fig. 2A-upper panel). The protein

13
expression of AMPK and α-tubulin were similar in the hypothalamus of controls and TB
rats in both conditions (Fig. 2A-middle and lower panels).
The i.c.v. microinfusion of AICAR and i.p. injection of 2DG in TB rats after 12-
hour fast caused an increase in hypothalamic AMPK phosphorylation in a dose- and time-
manner dependent (Fig. 2B and C). The i.c.v. infusion of AICAR (0.5 and 2.0mM)
increased AMPK phosphorylation in the hypothalamus of TB rats by 2.2- and 4.9-fold at 60
minutes, respectively, when compared to vehicle (Fig. 2B-upper panel). The 2DG (250 and
500 mg/kg) i.p. injection increased the hypothalamic AMPK phosphorylation by 1.2- and
3.6-fold, respectively, in the hypothalamus of TB rats at 180 minutes, when compared to
vehicle (Fig. 2C-upper panel). The AMPK and α-tubulin protein levels did not differ
between the groups after AICAR or 2DG administration (Fig. 2B and C-middle and lower
panels).
To determine whether the anti diabetic drug, metformin, induces hypothalamic
AMPK phosphorylation in TB rats, we administered metformin using oral gavage during
cancer-induced anorexia in rats after 12-hour fast. The oral administration of metformin
(500 mg/kg) in TB rats caused an increase in hypothalamic AMPK phosphorylation in a
time-manner dependent, with maximal response at 180 minutes after oral gavage (Fig. 2D-
upper panel). The total AMPK and α-tubulin protein levels were not different between the
groups after metformin administration (Fig. 2D-middle and lower panels).
Next, we explored the effects of oral gavage of metformin on hypothalamic AMPK
and ACC phosphorylation in the TB animal. After 12 hour fasting conditions, AMPK and
ACC phosphorylation were reduced by 3.1- and 4.6-fold in the hypothalamus of TB rats
when compared to the control group, respectively. Metformin administration increased

14
hypothalamic AMPK and ACC phosphorylation by 2.0- and 3.9-fold, when compared to
saline-treated TB rats (Fig. 2E and F-upper panels). The AMPK, ACC and α-tubulin
protein levels were not different between the groups after metformin administration (Fig. 2
E and F-middle and lower panels).
�
AMPK activation attenuates cancer anorexia and increases survival in tumor bearing
rats.
To determine whether the hypothalamic AMPK activation increases food intake and
survival in TB animals, we administered i.c.v. AICAR or i.p. 2DG during the period of
anorexia in TB rats. Figures 3A and D show that the tumor development markedly reduced
food intake during 24 hours in TB rats after saline treatment when compared to control
animals. The central or peripheral administration of AMPK activators in TB rats caused an
increase in food intake that was apparent 4 hours after treatment and lasted for 12 and 24
hours (Fig. 3A and D). When criteria for anorexia had been met, we administered a daily
injection of icv AICAR or ip 2DG during 4 days in TB rats to evaluate the cumulative food
intake. In TB animals the cumulative food intake was reduced by about 75% when
compared to respective control groups; however, the chronic treatment with AICAR (i.c.v.)
or 2DG (i.p.) increased the cumulative food intake by about 25 and 35%, respectively,
when compared to TB rats treated with i.c.v. or i.p. saline (Fig. 3B and E). Because AMPK
is the only enzyme known to be activated by both AICAR and 2DG, these data suggest that
tumor induced anorexia is dependent on the activation of AMPK.
In addition, we investigated whether the hypothalamic AMPK activation increases
survival in TB animals after chronic AICAR or 2DG treatment. As shown in the Kaplan-
Maier’s graphs, a daily central infusion of AICAR or a peripheral injection of 2DG,

15
statistically prolonged the survival in TB animals, whereas, the median survival of TB rats
after chronic administration of saline via i.c.v. or i.p. was ~5.5 days and chronic
administration of AICAR or 2DG increased the median survival to 9.5 and 8.0 days,
respectively (Fig. 3C and F).
Figure 3G shows that the tumor development markedly reduced food intake during
24 hours in TB rats after saline treatment when compared to control animals. Furthermore,
oral administration of metformin in TB rats, caused an increase in food intake that was
apparent 4 hours after treatment and lasted for 12 and 24 hours (Fig. 3G). When criteria for
anorexia had been met, the cumulative food intake was evaluated after a daily oral gavage
treatment with saline or metformin during 4 days in TB animals. The cumulative food
intake was reduced by about 75% when compared to control animals, however, the daily
oral treatment with metformin, increased the cumulative food intake by about 28%, when
compared to TB rats treated with oral saline (Fig. 3H).
We also investigated whether the hypothalamic AMPK activation increases the
survival in TB animals after chronic oral treatment with metformin. Figure 3I shows, a
daily oral administration of metformin significantly increases survival, whereas the median
survival of oral saline-treated TB rats was 5.5 days and chronic administration of oral
metformin increased the median survival to 8.5 days.
To explore the specific action of metformin in the hypothalamus, we injected i.c.v.
metformin (1mM) in TB rats to evaluate food intake and survival. Figure 3J shows that the
tumor development markedly reduced food intake during 24 hours in TB rats after saline
treatment, when compared to control animals. The i.c.v. administration of metformin in TB
rats, caused an increase in food intake that was apparent 4 hours after treatment and lasted
for 12 and 24 hours (Fig. 3J). When criteria for anorexia had been met, the cumulative food

16
intake was evaluated after a daily i.c.v. treatment with saline or metformin during 4 days in
TB animals. The cumulative food intake was reduced by about 75% when compared to
control animals; however, the daily treatment with metformin increased the cumulative
food intake by about 32%, when compared to TB rats treated with i.c.v. saline (Fig. 3K).
We finally investigated whether the hypothalamic AMPK activation increases the
survival in TB animals after chronic i.c.v. treatment with metformin. Figure 3L shows that
daily i.c.v. administration of metformin significantly increases survival, whereas the
median survival of oral saline-treated TB rats was 5.5 days and chronic administration of
i.c.v. metformin increased the median survival to 9 days.
Hypothalamic AMPK activation decreases the expression of proinflammatory
molecules in TB rats.
We next investigated the anti-inflammatory properties of AICAR in the
hypothalamus of TB rats. To test the hypothesis that AMPK activation reduces the�
production of proinflamatory cytokines in TB rats, we injected the AMPK activator and
evaluated the hypothalamic expression of iNOS, IL-1β and TNF-α. The protein levels of
iNOS increased by 3.8-fold in the hypothalamus of TB rats when compared to control
animals, and the micro infusion of AICAR reduced iNOS expression by 1.5-fold in the
hypothalamus of TB rats, when compared to saline-treated TB rats (Fig. 4A - upper panel).
The α-tubulin protein levels were not different between the groups after AICAR
administration (Fig. 4A - lower panel). In addition, the protein levels of IL-1β and TNF-α
were increased by 7.9 and 2.0-fold, respectively, in the hypothalamus of TB rats when
compared to the control group, and AMPK activation reduced the hypothalamic expression

17
of IL-1β by 1.9-fold and TNF-α by 1.5-fold, when compared to TB rats treated with i.c.v.
saline (Fig. 4B and C, respectively).
To explore the hypothesis that the reduction in inflammatory molecules in the
hypothalamus attenuates anorexia in the of TB rats, we injected a specific anti-IL-1β or
anti-TNFα antibodies via i.c.v in TB animals. In TB rats, the cumulative food intake was
reduced by about 75% when compared to the respective control group; however, the
chronic treatment with anti-IL-1β antibody (i.c.v.) increased the cumulative food intake by
about 72%, when compared to TB rats treated with i.c.v. saline (Fig. 4D). Chronic i.c.v.
treatment with anti-TNFα antibody produced a slight increase in cumulative food intake,
but we did not observe a significative difference when compared to saline i.c.v. treatment in
TB animals (Fig. 4D).
Central infusion of AICAR reduced POMC mRNA levels in the hypothalamus of TB
rats.
To explore the mechanism(s) by which AMPK activation regulates food intake in
TB rats, we examined the expression of hypothalamic neuropeptides involved in the control
of energy homeostasis in TB animals. After 12 hours of fasting, the neuropeptide Y (NPY)
mRNA levels were reduced by 1.7-fold in the hypothalamus of TB rats when compared to
the control group, and the i.c.v. microinfusion of AICAR was not able to increase
hypothalamic NPY mRNA levels in TB rats when compared to saline-treated TB rats (Fig.
5A). On the other hand, pro-opiomenalnocortin (POMC) mRNA levels were increased by
2.2-fold in the hypothalamus of TB rats when compared to the control group, and the i.c.v.

18
microinfusion of AICAR decreased hypothalamic POMC mRNA levels by 1.4-fold in TB
rats when compared to saline-treated TB rats (Fig. 5B).

19
DISCUSSION
In the present study, we demonstrate that central AMPK activation can attenuate
cancer-induced anorexia, whereas, the activation of neuronal AMPK inhibits the
hypothalamic expression of proinflammatory molecules, suppresses POMC mRNA levels
in the hypothalamus of anorectic tumor bearing rats. Interestingly, different
pharmacological activators of AMPK signaling, including intracerebroventricular
microinfusion of AICAR, intraperitoneal injection of 2DG and oral or i.c.v. administration
of metformin, activate AMPK in the central nervous system, increase food consumption
and prolong life span in the model of cancer-induced anorexia.
An increased expression of cytokines during tumour growth prevents the
hypothalamus from responding appropriately to peripheral signals by persistently activating
anorexigenic systems and inhibiting prophagic pathways (7). Convincing evidence suggests
that cytokines may across the blood brain barrier and have a vital role, triggering the
complex neurochemical cascade, leading to the onset of cancer anorexia (7, 8). In addition,
cytokine production in brain regions distant from the brain tumor site may also be involved
in this clinical manifestation (35). In patients with cancer, it is probable that cytokines and
anorexia are connected, since the biological effects of cytokines are largely mediated by
paracrine and autocrine influences (7). In accordance with our data demonstrating that the
reduction in IL-1β cause the increased appetite; in the Fischer rat/MCA sarcoma model,
brain IL-1β concentrations inversely correlate with food intake (36) and intrahypothalamic
microinjections of an interleukin-1-receptor antagonist increases energy intake (37). Tumor
necrosis factor-α (TNF-α) is one of the mediators of the hypothalamic anorexigenic signals
that participate in the induction of the cachexia syndromes present in advanced stage cancer

20
and in severe infectious (38, 39). Although we did not observe a significant increase in
cumulative food intake with anti-TNFα treatment, our data demonstrated that the reversal
effect of AMPK on cancer induced-anorexia is associated with decreased concentrations of
iNOS, IL-1β and TNF-α in the hypothalamus of tumor-bearing rats. The anti-inflammatory
response produced by AMPK activation in the CNS is in accordance with several studies,
suggesting that AICAR inhibits production of proinflammatory mediators (TNF-α, IFNγ,
and NO) in brain glial cells, primary astrocytes, microglia, and peritoneal macrophages (26,
28). In addition, the pharmacological activation of AMPK also inhibited the production of
proinflammatory molecules in serum and their expression in the CNS of rats injected with a
sublethal dose of LPS by intraperitoneal injection (26).
The mechanisms involved in the proinflammatory cytokine-dependent modulation
of food intake and energy expenditure may also involve the control of hypothalamic
neurotransmitter expression (40, 41), in situ stimulation of the expression of other
cytokines, particularly IL-1β (42, 43), and the activation of anorexigenic leptin-like signal
transduction in the hypothalamus (44). IL-1β has been clearly associated with the induction
of anorexia (39), by blocking neuropeptide Y (NPY)-induced feeding. The levels of this
molecule are reduced in anorectic tumor-bearing rats (11), and a correlation between food
intake and brain IL-1 has been found in anorectic rats with cancer. The mechanism
involved in the attenuation of NPY activity by cytokines may be related to an inhibition of
cell firing rates or to an inhibition of NPY synthesis or an attenuation of its postsynaptic
effects (45). On the other hand, it has been demonstrated that POMC-containing cells
contain cytokine receptors, and that these neurons are activated during acute and chronic
inflammation (13). This discovery demonstrates that proinflammatory cytokines such as

21
TNF-α, IL-1β and IL-6 can bind to cytokine receptors on POMC-containing cells, causing
these cells to increase their signaling to second-order neurons that affect outputs related to
anorexia, loss of lean body mass and increased energy expenditure. In our model of cancer-
induced anorexia, we observed high levels of iNOS, IL-1β and TNF-α in the hypothalamus
of Walker tumor-bearing rats and, after 12 hours of fasting, the mRNA levels of NPY were
lower and POMC were much higher than in control animals. Notably, we also observed
defective 172 threonine AMPK phosphorylation in the hypothalamus of anorexic tumor-
bearing rats under 12-hour fasting conditions, when compared to normal rats, suggesting
that the neuronal AMPK could be involved in cancer-mediated anorexia.
AMPK is expressed in the hypothalamic neurons involved in the regulation of food
intake (46), whereas, the central pharmacological activation of AMPK by AICAR leads to
increased food intake and decreased energy expenditure in normal mice (19). Furthermore,
acute intraperitoneal administration of 2DG increased food intake and both the expression
of AgRP and NPY in the arcuate nucleus (47). These findings were further explored in
different studies, demonstrating that increasing hypothalamic AMPK activity increased the
expression of orexigen neuropeptides in the arcuate nucleus of the hypothalamus, leading to
increased food intake (29), moreover, preventing activity of the melanocortin system, using
either genetic deletions of the MC4 receptor or small-molecule antagonists of this receptor,
is effective in improving appetite and lean body mass in small animal models of cachexia in
chronic disease (48-50). We demonstrate that the microinfusion of AICAR activates AMPK
in neuronal cells in a time- and dose-dependent manner and that the higher dose of AICAR
(2mM) (although it does not change NPY mRNA levels) partially restores POMC mRNA
levels in the hypothalamus of anorectic rats after a 12-hour fast. Moreover, both central

22
microinfusion of AICAR and intraperitoneal injection of 2DG increased food intake and
prolonged survival in tumor-bearing animals.
Several groups have used the adenosine analogue AICAR to activate AMPK.
AICAR is taken up into the cell by adenosine transporters (51) and converted by adenosine
kinase into the monophosphorylated nucleotide, ZMP, which mimics all of the effects of
AMP on the AMPK system (52). Moreover, recent evidence demonstrates that
CaMKK/AMPK axis is phosphorylated by 2DG (53). These data taken together with our
findings that tumor’s inhibitory effect on food intake is blocked by either of two AMPK
activators reduces the possibility of non-specific effects of these drugs and supports an
important patophysiological role for this intracellular signaling pathway.
In addition to the increased food intake, other possibilities beneficial effects on the
survival of animals may occur after these treatments. First, AMPK activation is related to
inhibition of the mTOR cascade, which decreases cell proliferation (54). In this regard, we
may not have seen any differences in tumor size after the different treatments, possibly due
to the short duration of the treatments (4 days); moreover, AICAR and metformin i.c.v.
infusion probably does not directly modulate the AMPK/mTOR pathway in the tumor and
tumor growth. Secondly, our results show that targeting the brain improves the metabolic
milieu, as shown by the i.c.v. infusion of AICAR, suggesting that the brain may control
some of the insulin resistance characteristics. Finally, it is also possible that AMPK
activators are also acting as immune suppressing agents and decreasing the overall
inflammatory response to the tumor. Although we found a close correlation between the
intensity of hypothalamic inflammation, AMPK activity and life span, survival did not
correspond to serum TNF-α and to the intensity of the inflammatory response in the spleen.

23
Direct hypothalamic activation with AICAR reproduces the effect of oral treatment with
metformin and intraperitoneal treatment with 2DG on food intake and survival, despite a
distinct serum TNF-α and spleen index between the intracerebroventricular and systemic
treatment. Thus, we can not exclude the possibility that AMPK activation in the
hypothalamus decreases some of whole body inflammatory response to the tumor and that
this contributes to the increased survival; these results are intriguing and deserve further
investigation.
Efforts have been made to implicate leptin as a modulator of food intake in tumor-
induced anorexia, however, lower leptin levels were found in patients with gastrointestinal
cancers, regardless of their degree of weight loss and in colon cancer patients who had no
weight loss at all (55-57). In contrast, an association between leptin levels and weight loss
was noted in a cohort of lung (58) and pancreatic cancer patients (59). Similarly, we found
an association between leptin levels and cancer-induced weight loss. It was recently shown
that both i.p. and i.c.v. administrations of the melanocortin agonist, MTII, increase IL1β
mRNA expression in the mediobasal hypothalamus (60). Thus IL1β signaling may be
downstream of melanocortin signaling. Moreover, physiological changes in leptin appear to
alter signaling in only a small subset of POMC neurons in the rostral portion of the ARC
(61). Taken together these data suggest that tumor-induced anorexia is independent of the
levels of leptin.
AMPK provides a candidate target, capable of mediating the beneficial metabolic
effects of metformin (62). Metformin lowers blood glucose and blood lipid contents, and
these effects are thought to be at least partially responsible for its therapeutic benefits (63,
64). Metformin decreases the leptin concentration in morbidly obese subjects (65, 66) and

24
in normal-weight healthy men (67). Metformin has been suggested to act through the
stimulation of AMPK in peripheral tissues; however, few studies have demonstrated
whether metformin modulates AMPK activity in the neuronal cell. In Zucker rats, a single
subcutaneous dose of metformin (300 mg/kg) reduced food intake only in obese animals,
while the same dose of metformin given orally did not affect food intake in either lean or
obese animals (68). In an acute study, metformin treatment increased the anorexic effect of
leptin, and this was accompanied by increased levels of phosphorylated signal transducer
and activator of transcription 3 (STAT3) in the hypothalamus of high-fat-fed obese rats
(69). Metformin also inhibits AMPK activation and prevents increases in NPY expression
in cultured hypothalamic neurons (70). On the other hand, our results indicated that a high
dose of oral metformin (500 mg/kg) increases the phosphorylation of the AMPK/ACC
pathway in a time-dependent manner in the hypothalamus of rodents in cancer-induced
anorexia after 12-hour fast. A daily oral or i.c.v. administration of metformin blockaded the
anorectic response, leading to increased food intake and life span in tumor-bearing rats.
These apparent contradictory effects of metformin in the hypothalamus may be related to
the different physiological and metabolic profiles observed in these studies. In addition, we
used a prolonged fasting period (12h) to evaluate the hypothalamic AMPK
phosphorylation. Furthermore, we performed the same experiments with an oral
administration of metformin (500 mg/kg) and i.c.v metformin (1mM) in normal rats and we
did not observe changes in either food intake or body weight (data not shown), suggesting
that the physiological and metabolic parameters are essential to metformin-induced changes
in food intake and body weight in rodents.
In summary, the findings of this study suggest that neuronal AMPK activation
reverses cancer anorexia by inhibiting the production of proinflammatory molecules and

25
controlling the expression of POMC, reflecting in the prolonged life span of tumor-bearing
rats. Thus, our data indicate that hypothalamic AMPK activation presents an attractive
opportunity for the treatment of cancer-induced anorexia, whereas, the restoration of
appetite in cancer patients is likely to improve quality of life and might also improve
overall patient survival.

26
ACKNOWLEDGMENTS
The authors thank Mr. Luiz Janeri, Jósimo Pinheiro, Leandro Macow and Márcio A. da
Cruz for technical assistance. This study was supported by grants from FAPESP and CNPq.

27
REFERENCES���
�
1. Dewys WD, Begg C, Lavin PT, Band PR, Bennett JM, Bertino JR, Cohen MH, Douglass HO, Jr., Engstrom PF, Ezdinli EZ, Horton J, Johnson GJ, Moertel CG, Oken MM, Perlia C, Rosenbaum C, Silverstein MN, Skeel RT, Sponzo RW, Tormey DC 1980 Prognostic effect of weight loss prior to chemotherapy in cancer patients. Eastern Cooperative Oncology Group. Am J Med 69:491-7
2. Apolone G 2000 The state of research on multipurpose severity of illness scoring systems: are we on target? Intensive Care Med 26:1727-9
3. Marks DL, Ling N, Cone RD 2001 Role of the central melanocortin system in cachexia. Cancer Res 61:1432-8.
4. Plata-Salaman CR 1991 Immunoregulators in the nervous system. Neurosci Biobehav Rev 15:185-215.
5. Plata-Salaman CR 1997 Anorexia during acute and chronic disease: relevance of neurotransmitter-peptide-cytokine interactions. Nutrition 13:159-60.
6. Wisse BE, Frayo RS, Schwartz MW, Cummings DE 2001 Reversal of cancer anorexia by blockade of central melanocortin receptors in rats. Endocrinology 142:3292-301.
7. Laviano A, Meguid MM, Rossi-Fanelli F 2003 Cancer anorexia: clinical implications, pathogenesis, and therapeutic strategies. Lancet Oncol 4:686-94
8. Hosoi T, Okuma Y, Nomura Y 2002 The mechanisms of immune-to-brain communication in inflammation as a drug target. Curr Drug Targets Inflamm Allergy 1:257-62
9. Nonaka N, Hileman SM, Shioda S, Vo TQ, Banks WA 2004 Effects of lipopolysaccharide on leptin transport across the blood-brain barrier. Brain Res 1016:58-65
10. Inui A 1999 Cancer anorexia-cachexia syndrome: are neuropeptides the key? Cancer Res 59:4493-501
11. Chance WT, Balasubramaniam A, Dayal R, Brown J, Fischer JE 1994 Hypothalamic concentration and release of neuropeptide Y into microdialysates is reduced in anorectic tumor-bearing rats. Life Sci 54:1869-74
12. Scarlett JM, Jobst EE, Enriori PJ, Bowe DD, Batra AK, Grant WF, Cowley MA, Marks DL 2007 Regulation of central melanocortin signaling by interleukin-1{beta}. Endocrinology
13. Deboer MD, Marks DL 2006 Cachexia: lessons from melanocortin antagonism. Trends Endocrinol Metab 17:199-204

28
14. Gayle D, Ilyin SE, Romanovitch AE, Peloso E, Satinoff E, Plata-Salaman CR1999 Basal and IL-1beta-stimulated cytokine and neuropeptide mRNA expression in brain regions of young and old Long-Evans rats. Brain Res Mol Brain Res 70:92-100
15. Ramos EJ, Suzuki S, Meguid MM, Laviano A, Sato T, Chen C, Das U 2004 Changes in hypothalamic neuropeptide Y and monoaminergic system in tumor-bearing rats: pre- and post-tumor resection and at death. Surgery 136:270-6
16. Carvalheira JB, Siloto RM, Ignacchitti I, Brenelli SL, Carvalho CR, Leite A, Velloso LA, Gontijo JA, Saad MJ 2001 Insulin modulates leptin-induced STAT3 activation in rat hypothalamus. FEBS Lett 500:119-24.
17. Schwartz MW, Porte D, Jr. 2005 Diabetes, obesity, and the brain. Science 307:375-9
18. Kim MS, Park JY, Namkoong C, Jang PG, Ryu JW, Song HS, Yun JY, Namgoong IS, Ha J, Park IS, Lee IK, Viollet B, Youn JH, Lee HK, Lee KU2004 Anti-obesity effects of alpha-lipoic acid mediated by suppression of hypothalamic AMP-activated protein kinase. Nat Med 10:727-33
19. Andersson U, Filipsson K, Abbott CR, Woods A, Smith K, Bloom SR, Carling D, Small CJ 2004 AMP-activated protein kinase plays a role in the control of food intake. J Biol Chem 279:12005-8
20. Lee K, Li B, Xi X, Suh Y, Martin RJ 2005 Role of neuronal energy status in the regulation of adenosine 5'-monophosphate-activated protein kinase, orexigenic neuropeptides expression, and feeding behavior. Endocrinology 146:3-10
21. Hardie DG, Salt IP, Hawley SA, Davies SP 1999 AMP-activated protein kinase: an ultrasensitive system for monitoring cellular energy charge. Biochem J 338 (Pt 3):717-22
22. Shaw RJ, Kosmatka M, Bardeesy N, Hurley RL, Witters LA, DePinho RA, Cantley LC 2004 The tumor suppressor LKB1 kinase directly activates AMP-activated kinase and regulates apoptosis in response to energy stress. Proc Natl Acad Sci U S A 101:3329-35
23. Woods A, Dickerson K, Heath R, Hong SP, Momcilovic M, Johnstone SR, Carlson M, Carling D 2005 Ca2+/calmodulin-dependent protein kinase kinase-beta acts upstream of AMP-activated protein kinase in mammalian cells. Cell Metab 2:21-33
24. Hawley SA, Pan DA, Mustard KJ, Ross L, Bain J, Edelman AM, Frenguelli BG, Hardie DG 2005 Calmodulin-dependent protein kinase kinase-beta is an alternative upstream kinase for AMP-activated protein kinase. Cell Metab 2:9-19
25. Hurley RL, Anderson KA, Franzone JM, Kemp BE, Means AR, Witters LA2005 The Ca2+/calmodulin-dependent protein kinase kinases are AMP-activated protein kinase kinases. J Biol Chem 280:29060-6

29
26. Giri S, Nath N, Smith B, Viollet B, Singh AK, Singh I 2004 5-aminoimidazole-4-carboxamide-1-beta-4-ribofuranoside inhibits proinflammatory response in glial cells: a possible role of AMP-activated protein kinase. J Neurosci 24:479-87
27. Ayasolla KR, Singh AK, Singh I 2005 5-aminoimidazole-4-carboxamide-1-beta-4-ribofuranoside (AICAR) attenuates the expression of LPS- and Abeta peptide-induced inflammatory mediators in astroglia. J Neuroinflammation 2:21
28. Jhun BS, Jin Q, Oh YT, Kim SS, Kong Y, Cho YH, Ha J, Baik HH, Kang I2004 5-Aminoimidazole-4-carboxamide riboside suppresses lipopolysaccharide-induced TNF-alpha production through inhibition of phosphatidylinositol 3-kinase/Akt activation in RAW 264.7 murine macrophages. Biochem Biophys Res Commun 318:372-80
29. Minokoshi Y, Alquier T, Furukawa N, Kim YB, Lee A, Xue B, Mu J, Foufelle F, Ferre P, Birnbaum MJ, Stuck BJ, Kahn BB 2004 AMP-kinase regulates food intake by responding to hormonal and nutrient signals in the hypothalamus. Nature 428:569-74
30. Bonora E, Moghetti P, Zancanaro C, Cigolini M, Querena M, Cacciatori V, Corgnati A, Muggeo M 1989 Estimates of in vivo insulin action in man: comparison of insulin tolerance tests with euglycemic and hyperglycemic glucose clamp studies. J Clin Endocrinol Metab 68:374-8
31. Scott AM, Atwater I, Rojas E 1981 A method for the simultaneous measurement of insulin release and B cell membrane potential in single mouse islets of Langerhans. Diabetologia 21:470-5
32. Schrier DJ, Ripani LM, Katzenstein AL, Moore VL 1982 Role of angiotensin-converting enzyme in Bacille Calmette-Guerin-induced granulomatous inflammation. Increased angiotensin-converting enzyme levels in lung lavage and suppression of inflammation with captopril. J Clin Invest 69:651-7
33. Gambardella A, Paolisso G, D'Amore A, Granato M, Verza M, Varricchio M1993 Different contribution of substrates oxidation to insulin resistance in malnourished elderly patients with cancer. Cancer 72:3106-13
34. Gomez-Ambrosi J, Zabalegui N, Bing C, Tisdale MJ, Trayhurn P, Williams G2002 Weight loss in tumour-bearing mice is not associated with changes in resistin gene expression in white adipose tissue. Horm Metab Res 34:674-7
35. Ilyin SE, Gayle D, Gonzalez-Gomez I, Miele ME, Plata-Salaman CR 1999 Brain tumor development in rats is associated with changes in central nervous system cytokine and neuropeptide systems. Brain Res Bull 48:363-73
36. Opara EI, Laviano A, Meguid MM, Yang ZJ 1995 Correlation between food intake and CSF IL-1 alpha in anorectic tumor bearing rats. Neuroreport 6:750-2

30
37. Laviano A, Gleason JR, Meguid MM, Yang ZJ, Cangiano C, Rossi Fanelli F2000 Effects of intra-VMN mianserin and IL-1ra on meal number in anorectic tumor-bearing rats. J Investig Med 48:40-8
38. Matthys P, Billiau A 1997 Cytokines and cachexia. Nutrition 13:763-70
39. Plata-Salaman CR 2000 Central nervous system mechanisms contributing to the cachexia-anorexia syndrome. Nutrition 16:1009-12
40. Bernardini R, Kamilaris TC, Calogero AE, Johnson EO, Gomez MT, Gold PW, Chrousos GP 1990 Interactions between tumor necrosis factor-alpha, hypothalamic corticotropin-releasing hormone, and adrenocorticotropin secretion in the rat. Endocrinology 126:2876-81
41. Wisse BE, Schwartz MW, Cummings DE 2003 Melanocortin signaling and anorexia in chronic disease states. Ann N Y Acad Sci 994:275-81
42. Bristulf J, Bartfai T 1995 Interleukin-1 beta and tumour necrosis factor-alpha stimulate the mRNA expression of interleukin-1 receptors in mouse anterior pituitary AtT-20 cells. Neurosci Lett 187:53-6
43. Zhai QH, Futrell N, Chen FJ 1997 Gene expression of IL-10 in relationship to TNF-alpha, IL-1beta and IL-2 in the rat brain following middle cerebral artery occlusion. J Neurol Sci 152:119-24
44. Rizk NM, Stammsen D, Preibisch G, Eckel J 2001 Leptin and tumor necrosis factor-alpha induce the tyrosine phosphorylation of signal transducer and activator of transcription proteins in the hypothalamus of normal rats in vivo. Endocrinology 142:3027-32
45. King PJ, Widdowson PS, Doods H, Williams G 2000 Effect of cytokines on hypothalamic neuropeptide Y release in vitro. Peptides 21:143-6
46. Turnley AM, Stapleton D, Mann RJ, Witters LA, Kemp BE, Bartlett PF 1999 Cellular distribution and developmental expression of AMP-activated protein kinase isoforms in mouse central nervous system. J Neurochem 72:1707-16
47. Sergeyev V, Broberger C, Gorbatyuk O, Hokfelt T 2000 Effect of 2-mercaptoacetate and 2-deoxy-D-glucose administration on the expression of NPY, AGRP, POMC, MCH and hypocretin/orexin in the rat hypothalamus. Neuroreport 11:117-21
48. Cheung W, Yu PX, Little BM, Cone RD, Marks DL, Mak RH 2005 Role of leptin and melanocortin signaling in uremia-associated cachexia. J Clin Invest 115:1659-65
49. Markison S, Foster AC, Chen C, Brookhart GB, Hesse A, Hoare SR, Fleck BA, Brown BT, Marks DL 2005 The regulation of feeding and metabolic rate and the prevention of murine cancer cachexia with a small-molecule melanocortin-4 receptor antagonist. Endocrinology 146:2766-73

31
50. Marks DL, Butler AA, Turner R, Brookhart G, Cone RD 2003 Differential role of melanocortin receptor subtypes in cachexia. Endocrinology 144:1513-23
51. Gadalla AE, Pearson T, Currie AJ, Dale N, Hawley SA, Sheehan M, Hirst W, Michel AD, Randall A, Hardie DG, Frenguelli BG 2004 AICA riboside both activates AMP-activated protein kinase and competes with adenosine for the nucleoside transporter in the CA1 region of the rat hippocampus. J Neurochem 88:1272-82
52. Corton JM, Gillespie JG, Hawley SA, Hardie DG 1995 5-aminoimidazole-4-carboxamide ribonucleoside. A specific method for activating AMP-activated protein kinase in intact cells? Eur J Biochem 229:558-65
53. Tzatsos A, Tsichlis PN 2007 Energy depletion inhibits phosphatidylinositol 3-kinase/Akt signaling and induces apoptosis via AMP-activated protein kinase-dependent phosphorylation of IRS-1 at Ser-794. J Biol Chem 282:18069-82
54. Shaw RJ, Bardeesy N, Manning BD, Lopez L, Kosmatka M, DePinho RA, Cantley LC 2004 The LKB1 tumor suppressor negatively regulates mTOR signaling. Cancer Cell 6:91-9
55. Arpaci F, Yilmaz MI, Ozet A, Ayta H, Ozturk B, Komurcu S, Ozata M 2002 Low serum leptin level in colon cancer patients without significant weight loss. Tumori 88:147-9
56. Bolukbas FF, Kilic H, Bolukbas C, Gumus M, Horoz M, Turhal NS, Kavakli B2004 Serum leptin concentration and advanced gastrointestinal cancers: a case controlled study. BMC Cancer 4:29
57. Dulger H, Alici S, Sekeroglu MR, Erkog R, Ozbek H, Noyan T, Yavuz M 2004 Serum levels of leptin and proinflammatory cytokines in patients with gastrointestinal cancer. Int J Clin Pract 58:545-9
58. Simons JP, Schols AM, Campfield LA, Wouters EF, Saris WH 1997 Plasma concentration of total leptin and human lung-cancer-associated cachexia. Clin Sci (Lond) 93:273-7
59. Brown DR, Berkowitz DE, Breslow MJ 2001 Weight loss is not associated with hyperleptinemia in humans with pancreatic cancer. J Clin Endocrinol Metab 86:162-6
60. Wisse BE, Ogimoto K, Schwartz MW 2006 Role of hypothalamic interleukin-1beta (IL-1beta) in regulation of energy homeostasis by melanocortins. Peptides 27:265-73
61. Schwartz MW, Seeley RJ, Woods SC, Weigle DS, Campfield LA, Burn P, Baskin DG 1997 Leptin increases hypothalamic pro-opiomelanocortin mRNA expression in the rostral arcuate nucleus. Diabetes 46:2119-23

32
62. Shaw RJ, Lamia KA, Vasquez D, Koo SH, Bardeesy N, Depinho RA, Montminy M, Cantley LC 2005 The kinase LKB1 mediates glucose homeostasis in liver and therapeutic effects of metformin. Science 310:1642-6
63. Khan AS, McLoughney CR, Ahmed AB 2006 The effect of metformin on blood glucose control in overweight patients with Type 1 diabetes. Diabet Med 23:1079-84
64. El-Batran SA, Abdel-Salam OM, Nofal SM, Baiuomy AR 2006 Effect of rosiglitazone and nateglinide on serum glucose and lipid profile alone or in combination with the biguanide metformin in diabetic rats. Pharmacol Res 53:69-74
65. Kay JP, Alemzadeh R, Langley G, D'Angelo L, Smith P, Holshouser S 2001 Beneficial effects of metformin in normoglycemic morbidly obese adolescents. Metabolism 50:1457-61
66. Glueck CJ, Fontaine RN, Wang P, Subbiah MT, Weber K, Illig E, Streicher P, Sieve-Smith L, Tracy TM, Lang JE, McCullough P 2001 Metformin reduces weight, centripetal obesity, insulin, leptin, and low-density lipoprotein cholesterol in nondiabetic, morbidly obese subjects with body mass index greater than 30. Metabolism 50:856-61
67. Fruehwald-Schultes B, Oltmanns KM, Toschek B, Sopke S, Kern W, Born J, Fehm HL, Peters A 2002 Short-term treatment with metformin decreases serum leptin concentration without affecting body weight and body fat content in normal-weight healthy men. Metabolism 51:531-6
68. Rouru J, Pesonen U, Koulu M, Huupponen R, Santti E, Virtanen K, Rouvari T, Jhanwar-Uniyal M 1995 Anorectic effect of metformin in obese Zucker rats: lack of evidence for the involvement of neuropeptide Y. Eur J Pharmacol 273:99-106
69. Kim YW, Kim JY, Park YH, Park SY, Won KC, Choi KH, Huh JY, Moon KH2006 Metformin restores leptin sensitivity in high-fat-fed obese rats with leptin resistance. Diabetes 55:716-24
70. Chau-Van C, Gamba M, Salvi R, Gaillard RC, Pralong FP 2007 Metformin inhibits adenosine 5'-monophosphate-activated kinase activation and prevents increases in neuropeptide Y expression in cultured hypothalamic neurons. Endocrinology 148:507-11

33
FIGURE LEGENDS
Figure 1- Schematic representation of the experimental procedures. Surgical implantation
of ICV cannula was performed ~3 weeks before the in vivo study. Full recovery of body
weight and food intake was achieved after 7 days. When criteria for anorexia had been met,
ICV infusions were started and blood chemistries, food intake and/or survival analysis were
assessed.
Figure 2- The basal levels of hypothalamic AMPK phosphorylation and the time-course
and dose-response after central infusion of AICAR, peripheral administration of 2DG or
oral metformin in tumor-bearing rats. Representative Western blots demonstrating: (A) The
effect of 12 hours fasting on hypothalamic AMPK phosphorylation in control and tumor-
bearing animals. (B) The hypothalamic AMPK phosphorylation in a time- and dose-
dependent manner after central infusions of AICAR, and after intraperitoneal injections of
2DG in tumor-bearing rats (C). Representative Western blots demonstrating the effect of
oral metformin on hypothalamic AMPK phosphorylation in a time-dependent manner (D).
Representative Western blots demonstrating the effect of oral metformin on hypothalamic
AMPK and ACC phosphorylation (E and F). Tissue extracts were immunoblotted (IB) with
anti- phospho AMPK antibody and anti- phospho ACC (upper panels), anti- AMPKα, anti-
ACC and anti-α-tubulin antibodies (lower pannels). The results of scanning densitometry
were expressed as arbitrary units. Bars represent means ± SEM of six to eight rats. #p<0.05,
vs. control fasted and *p<0.05, vs. respective vehicles.
Figure 3- The effects of intracerebroventricular infusion of AICAR, intraperitoneal 2DG,
oral or i.c.v. metformin on food intake and on survival in tumor-bearing (TB) rats.

34
Determination of 24- hours of food intake after AICAR microinfusion (A), after 2DG
injection (D), after oral metformin (G) and after i.c.v. metformin (J). Determination of 4
days of cumulative food intake after AICAR (B), 2DG treatment (E), oral metformin (H) or
i.c.v. metformin (K). Results represent means ± SEM of six to eight rats. #p<0.05, vs.
saline and *p<0.05, vs.TB rats + saline. Representative survival curves demonstrating the
effect of chronic administration of AICAR (C), 2DG (F), oral metformin (I) or i.c.v.
metformin (L) on life span of tumor-bearing animals. The survival curves were estimated
using Kaplan-Meier’s estimates, and curves were compared using the log-rank test, p<0.001
(n=10).
Figure 4- Intracerebroventricular infusion of AICAR attenuated the production of
proinflammatory molecules in the hypothalamus of tumor-bearing rats. Representative
Western blots demonstrating the effects of AICAR on iNOS (A) production in the
hypothalamus of tumor bearing rats. Bars represent means ± SEM of six rats. Tissue
extracts were immunoblotted (IB) with anti – iNOS antibody (upper panel) and with anti-α-
tubulin antibody (lower panel). ELISA assay demonstrated the anti- inflammatory
proprieties of AICAR on IL-1β (B), and TNF-α (C) expression in the hypothalamus of
tumor-bearing rats. Determination of cumulative food intake after chronic treatment with
i.c.v. anti-IL-1β or anti-TNFα antibodies (D), (n =5 per group). #p<0.05, vs. saline and
*p<0.05, vs. TB rats + saline.
Figure 5- The effect of central infusion of AICAR on NPY and POMC mRNA levels in the
hypothalamus of tumor bearing rats after 12h fast. The real-time PCR demonstrated the

35
NPY (A) and POMC (B) mRNA levels after AICAR microinfusion. Results for real-time
PCR are presented as transcript amount per hypothalamus. Bars represent means ± SEM of
six rats. #p<0.05, vs. saline and *p<0.05, vs. TB rats + saline.
�

36
TABLE 1. Characteristics of tumor bearing (TB) rats and their age-matched controls.
Groups (n=6) Control TB+Saline TB+AICAR TB+2DG TB+Metformin
Body weight (g) 369.3±5.3 346.5±7.6* 355.5±3.7* # 361.6±7.0* # 356.3±3.3* # Body weight excluding tumor (g) 369.3±5.3 310.9±12.2* 314.3±5.5* # 329.2±9.1* # 318.4±6.3* #
Tumor weight (g) _____ 35.6±3.6 41.3±2.3 32.4±2.7 35.0±2.5 Spleen index 1.00±0.09 6.60±0.74* 4.13±0.20*# 2.64±0.31*#$ 2.74±0.90*#$ Serum TNF-αααα (pg/ml) 10.7±2.6 36.2±2.4* 24.2±2.3*# 16.5±2.6*#$ 16.7±1.6*#$ Fasting glucose (mg dl-1) 80.2±2.6 85.4±4.0 81.8±4.9 81.7±7.8 85.5±2.8 Fasting insulin (ng ml–1) 1.05±0.17 0.06±0.01* 0.41±0.1* # 0.16±0.01* # 0.64±0.17* # Fasting leptin (ng ml–1) 5.62±1.4 0.21±0.04* 0.39±0.03* # 0.48±0.06* 1.11±0.05* # Kitt (% min–1) 4.53±0.6 1.93±0.4* 2.84±0.5* # 2.54±0.3* # 3.32±0.4* #
* p< 0.05, vs. control group. # p< 0.05, vs. TB+saline group. $ p< 0.05, vs. TB+AICAR group.




Related Documents


![Resveratrol suppresses glial activation and alleviates ...€¦ · sphate (AMP)-activated protein kinase (AMPK) [19]. Mul-tiple lines of evidence have indicated that AMPK is emerging](https://static.cupdf.com/doc/110x72/60f875cb55012f48d821e488/resveratrol-suppresses-glial-activation-and-alleviates-sphate-amp-activated.jpg)








