A cDNA microarray for the three-spined stickleback, Gasterosteus aculeatus L., and analysis of the interactive effects of oestradiol and dibenzanthracene exposures F. GEOGHEGAN*, I. KATSIADAKI †, T. D. WILLIAMS*‡ AND J. K. CHIPMAN* *School of Biosciences, The University of Birmingham, Edgbaston, Birmingham B15 2TT, U.K. and †Centre for Environment, Fisheries and Aquaculture Science, Cefas Weymouth Laboratory, The Nothe, Barrack Road, Weymouth, Dorset DT4 8UB, U.K. The stickleback is a useful model species for aquatic toxicology and endocrinology and with the recent sequencing of its genome, ecotoxicogenomics. The effects of treatment on male and female sticklebacks with the polycyclic aromatic hydrocarbon (PAH), dibenzanthracene (DbA), the female hormone, 17b-oestradiol (E2) and a binary mixture of the two were investigated. Quantitative RT-PCR assays were carried out for cytochrome P450 1A (cyp1A), vitellogenin (VTG) and oestrogen receptor alpha, and cyp1A enzyme activity was assessed by measurement of ethoxyresorufin-o-deethylase (EROD) activity , VTG and spiggin protein by enzyme-linked immunosorbent assay. A stickleback cDNA microarray consisting of 9692 clones was developed and used to assess gene expression responses to the treatments. Induction of cyp1A mRNA and EROD activity was seen in both sexes in response to DbA and a further induction of cyp1A found with the binary treatment. VTG mRNA was induced in male fish exposed to E2 and the binary mixture, while for females, statistically significant induction was seen only with the binary mixture. Similar behaviour was found with zona pellucida and chorion protein mRNA using the microarray. ERa mRNA was induced with E2 and the binary mixture in both sexes. The microarray showed additional transcripts that were differentially regulated by these treatments, which provide novel candidates for biomarker development and mechanistic studies. # 2008 The Authors Journal compilation # 2008 The Fisheries Society of the British Isles Key words: cyp1A; ecotoxicogenomics; microarray; oestradiol; stickleback; vitellogenin. INTRODUCTION The three-spined stickleback Gasterosteus aculeatus L. is widely distributed across Northern Europe, America and Asia and inhabits both freshwater and marine environments. It has long been used as a model species in ethology and evolutionary studies, and its genome has recently been sequenced. There is a particularly high interest in its use as a sentinel species for monitoring the effects of environmental pollutants, especially endocrine disruptors (Katsiadaki et al., 2002a, 2006). ‡Author to whom correspondence should be addressed. Tel.: þ44 (0)121 414 3393, þ44 (0)7796 321103; fax: þ44 (0)121 414 5925; email: [email protected] Journal of Fish Biology (2008) 72, 2133–2153 doi:10.1111/j.1095-8649.2008.01859.x, available online at http://www.blackwell-synergy.com 2133 # 2008 The Authors Journal compilation # 2008 The Fisheries Society of the British Isles

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A cDNA microarray for the three-spined stickleback,Gasterosteus aculeatus L., and analysis of the interactiveeffects of oestradiol and dibenzanthracene exposures
F. GEOGHEGAN*, I. KATSIADAKI†, T. D. WILLIAMS*‡AND J. K. CHIPMAN*
*School of Biosciences, The University of Birmingham, Edgbaston, Birmingham B15 2TT,U.K. and †Centre for Environment, Fisheries and Aquaculture Science, Cefas Weymouth
Laboratory, The Nothe, Barrack Road, Weymouth, Dorset DT4 8UB, U.K.
The stickleback is a useful model species for aquatic toxicology and endocrinology and with
the recent sequencing of its genome, ecotoxicogenomics. The effects of treatment on male and
female sticklebacks with the polycyclic aromatic hydrocarbon (PAH), dibenzanthracene (DbA),
the female hormone, 17b-oestradiol (E2) and a binary mixture of the two were investigated.
Quantitative RT-PCR assays were carried out for cytochrome P450 1A (cyp1A), vitellogenin
(VTG) and oestrogen receptor alpha, and cyp1A enzyme activity was assessed by measurement
of ethoxyresorufin-o-deethylase (EROD) activity , VTG and spiggin protein by enzyme-linked
immunosorbent assay. A stickleback cDNAmicroarray consisting of 9692 clones was developed
and used to assess gene expression responses to the treatments. Induction of cyp1A mRNA and
EROD activity was seen in both sexes in response to DbA and a further induction of cyp1A
found with the binary treatment. VTG mRNA was induced in male fish exposed to E2 and the
binary mixture, while for females, statistically significant induction was seen only with the
binary mixture. Similar behaviour was found with zona pellucida and chorion protein mRNA
using the microarray. ERa mRNA was induced with E2 and the binary mixture in both sexes.
The microarray showed additional transcripts that were differentially regulated by these
treatments, which provide novel candidates for biomarker development and mechanistic
studies. # 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles
Key words: cyp1A; ecotoxicogenomics; microarray; oestradiol; stickleback; vitellogenin.
INTRODUCTION
The three-spined stickleback Gasterosteus aculeatus L. is widely distributedacross Northern Europe, America and Asia and inhabits both freshwaterand marine environments. It has long been used as a model species in ethologyand evolutionary studies, and its genome has recently been sequenced. There isa particularly high interest in its use as a sentinel species for monitoring theeffects of environmental pollutants, especially endocrine disruptors (Katsiadakiet al., 2002a, 2006).
‡Author to whom correspondence should be addressed. Tel.: þ44 (0)121 414 3393, þ44 (0)7796 321103;
fax: þ44 (0)121 414 5925; email: [email protected]
Journal of Fish Biology (2008) 72, 2133–2153
doi:10.1111/j.1095-8649.2008.01859.x, available online at http://www.blackwell-synergy.com
2133# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles

Gene expression biomarkers, among others, can be used to indicate exposureto or effects of toxicants, thus acting as sensitive, early warning sensors (Malins& Ostrander, 1994). Exposure of organisms to compounds, either as individualchemicals or as mixtures, may result in complex changes at the levels of geneexpression. In the three-spined stickleback, biomarkers indicating endocrinedisruption include the egg yolk protein vitellogenin (VTG) produced in res-ponse to oestrogenic compounds (Katsiadaki et al., 2002a; Hahlbeck et al.,2004; Andersson et al., 2007) and the glue protein spiggin synthesized in res-ponse to androgenic compounds (Katsiadaki et al., 2002b, 2006).However, the expression of single biomarker genes cannot provide an inte-
grated assessment of the effects of different chemical classes. Transcriptomicsoffers the potential to assess the expression of thousands of genes simulta-neously, thereby providing a detailed profile of biological response to a com-pound or mixture. Key to this process is the comparison of the responses oforganisms experimentally exposed to individual toxicants or mixtures withthe responses found in environmentally sampled organisms. Transcript profil-ing of controlled experimental exposures provides opportunities to determinethe genes and pathways affected by a treatment, assisting in the determinationof the modes of action of chemicals on biological systems and potentially aid-ing the risk assessment of chemicals and assessment of the effects of chemicalpollutants on organisms in the wild (Ankley et al., 2006). The open nature ofthe experimental platform permits the discovery of unanticipated responses andprovides the flexibility necessary to assess effects of multiple stressors and theirmixtures. The design and development of the stickleback microarray was facil-itated by the information and materials derived from the stickleback genomesequencing project including genomic DNA sequence, annotation and resourcessuch as cDNA libraries (Kingsley et al., 2004).In this study, the authors initiated stickleback toxicogenomics by develop-
ment of a G. aculeatus cDNA microarray. Fish were exposed to dibenzanthra-cene (DbA) and 17b-oestradiol (E2) as single and binary treatments prior toanalysis of hepatic gene expression by microarray and quantitative RT-PCR(qPCR). The latter analyses of selected genes, such as cytochrome P450 1A(cyp1A), were complemented by measurement of ethoxyresorufin-o-deethylase(EROD) activity and VTG and spiggin protein.DbAis a polycyclic aromatic hydrocarbon (PAH) that induces the expression
of cyp1A (a phase I detoxification enzyme) and associated EROD activity.Potential effects on VTG protein production have also been reported with fieldstudies showing a decrease in levels of VTG (Casillas et al., 1991; Pereira et al.,1992) in fish at contaminated sites, suggesting that PAHs could affect variouspoints in the endocrine system, although it was found that at low levels of con-tamination, VTG was apparently increased (Pereira et al., 1992). The ability ofcompounds to induce cyp1A expression has since been suggested to depend onlevels of circulating oestradiol (E2) (Anderson et al., 1996).Circulating E2 has also been shown to have an effect on the expression
of hepatic cyp1A with the steroid showing an inhibitory effect on catalyticactivity and also on levels of protein and mRNA expression. Whether or notinduction of cyp1A by xenobiotics such as PAHs or dioxins such as 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) can be overridden by high levels of E2
2134 F. GEOGHEGAN ET AL .
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153

has been the subject of several studies with some showing continued induction(O’Hare et al., 1995; Navas & Segner, 2001) and others showing an inhibitionby E2 (Elskus et al., 1992).Subsequent work has revealed an inhibitory effect of induced aromatic hydro-
carbon receptor (AhR) on expression of E2 responsive genes (Safe & Wormke,2003) involving the oestrogen receptors (ERs) (Navas & Segner, 2001). AhR hassome degree of control over ER expression and oestrogenic response and isreciprocated with E2-activated oestrogen receptor alpha (ERa) having beenshown to interact directly with AhR–ARNT (aryl hydrocarbon receptor/arylhydrocarbon receptor nuclear translocator heterodimer) result in a repressionof dioxin-induced transcription (Beischlag & Perdew, 2005). Crosstalk may becell, tissue or species specific and could be the result of several mechanisms thathave yet to be fully described. The potential interactive effects of DbA and E2were therefore assessed in the liver of sticklebacks.
MATERIALS AND METHODS
EST LIBRARIES
The cDNA clones used for microarray construction were mainly derived from a singlecDNA library but were supplemented with a small number of clones derived fromsuppression, subtractive hybridization (SSH) and individually cloned genes.
The majority of clones were provided by the stickleback ‘CDA’ cDNA library madefrom heads and mixed internal organs of wild adult stickleback in pBK-CMV vector.This library, consisting of 9600 cDNA clones, was a kind gift from D. M. Kingsley(Stanford, U.S.A.). These clones had already been 59- and 39-sequenced using anABI3730xl sequencer with quality assessment as calculated by Phred (Ewing & Green,1998; Ewing et al., 1998). Clustering of sequence data suggested that 4780 distinct geneclusters, comprising multiple or single sequences, were represented in this library(Kingsley et al., 2004). All sequences had been deposited in GenBank (accession num-bers CD492707–CD51029). A stickleback SSH hepatic cDNA library was produced byforward suppressive, subtractive hybridization (PCR-Select System; BD Biosciences,Franklin Lakes, NJ, U.S.A.) between male fish exposed for 3 days to a mixture ofE2 (50 ng l�1), methyltestosterone (300 ng l�1), hexabromocyclododecane (300 ng l�1)and DbA (40 mg l�1) or a solvent control in a flow-through system. Products werecloned into pCR2.1 vector using a TA cloning kit according to the manufacturer’s in-structions (Invitrogen) and re-amplified from culture with Taq polymerase (Bioline,Taunton, MA, U.S.A.) in a 2720 Thermal Cycler (Applied Biosystems, Foster City,CA, U.S.A.) or Mastercycler Gradient (Eppendorf, Cambridge, U.K.) using M13forward and reverse primers (Stratagene, La Jolla, Ca, U.S.A.). Amplicons were 59and 39-sequenced at the NERC–ESF (Edinburgh, U.K.) with M13 forward and reverseprimers using an ABI3730 instrument (Applied Biosystems). Sequences were base-calledusing Phred (Ewing et al., 1998) and trimmed to remove low quality and vector regionsusing Lucy2 (Li & Chou, 2004).
ANNOTATION
Two approaches were taken to sequence annotation. First, all sequences, both 59and 39, were BLAST-ed v. the GenBank protein, nucleotide and ‘EST-others’ data-bases using Blast2GO (Conesa et al., 2005). Second, all sequences were comparedby megablast with the stickleback genome predicted cDNA set from Ensemblhttp://www.ensembl.org/Gasterosteus_aculeatus/index.html (Assembly BROAD S1).Highly similar predicted cDNAs were downloaded from BioMart (Durinck et al.,
STICKLEBACK MICROARRAY 2135
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153

2005) together with annotation from their Ensembl fugu Takifugu rubipres (Temminckand Schlegel), zebrafish Danio rerio (Hamilton) and human putative orthologs. Thesepredicted stickleback cDNAs were also used for database searching by Blast2GO.CAP3 (Huang & Madan, 1999) was used to cluster sequences. Subsequently, annota-tion from all sources was combined to annotate the stickleback microarray clones.Where annotation was inconsistent between the sources, such as 59 and 39 sequencesmatching to different genes, clones were excluded from further analysis. Gene ontology(GO) terms were annotated using Blast2GO.
MICROARRAY CONSTRUCTION
Bacterial cultures of stickleback clones in plasmid vectors were directly amplifiedwith T7- and M13-reverse primers. Polymerase chain reaction (PCR) was carriedout in 96-well format (Eppendorf Mastercycler or ABI2720) with verification by 96-well agarose gels. Failed or multi-product reactions were not removed in order toavoid the possibility of cross-contamination, but associated data were later removedin silico. PCR products were purified (Nucleofast 96; BD Biosciences) and roboticallytransferred to 27 � 384-well plates (Genetix) before resuspension in Pronto spottingbuffer (Corning Inc., Corning, NY, U.S.A.). This set consisted of 9692 sticklebackclones with remaining wells filled by spotting buffer. An additional 384-well platewas added comprising Lucidea calibration, ratio, utility and negative controls (Amer-sham, Little Chalfont, U.K.), plasmid DNA and polylinker amplicons from pRL(Promega, Madison, WI, U.S.A.), pBluescriptII SKþ (Stratagene), pCR2.1(Invitrogen, Carlsbad, CA, U.S.A.) and pTriplEx2 (BD Biosciences) and Renilla lucif-erase and b-lactamase amplicons. These were used as negative controls in this exper-iment. Microarray slides (Corning Ultra-GAPS, Corning Inc.) were printed atBirmingham Functional Genomics Laboratory with an MGII robot (Biorobotics,Cambridge, U.K.) using a 48� split-pin tool. Each amplicon was printed in duplicatefor a total of 21 504 spots. Slides were baked at 80° C for 2 h post-printing.
EXPOSURE TO COMPOUNDS
Five male and five female sticklebacks from a laboratory stock bred and maintainedat CEFAS Weymouth were exposed to the single compounds DbA (catalogue numberD-31400; Sigma-Aldrich, St Louis, MO, U.S.A.) (40 mg l�1 DbA in 0�0125% acetone)and oestradiol (E2, catalogue number E-8875, Sigma) (50 ng l�1 E2 in 0�0125% ace-tone) or binary DbA þ E2 (40 mg l�1 DbA þ 50 ng l�1 E2 in 0�0125% acetone) in addi-tion to a solvent control (acetone, 0�0125%) over a period of 48 h. Adult fish weremaintained in a 12D:12L cycle and were fed daily with frozen bloodworm, salinitywas 32, pH 7–8 and dissolved oxygen >70%. The delivery of compounds took placeby means of a peristaltic pump at a rate of 200 � 5 ml min�1, while the delivery of dilu-tion water was achieved by fine flow meters attached to each tank at a rate of 200 � 5ml min�1. The relationship between dilution water and compound was therefore 1 mlml�1. Both flow meters and peristaltic pumps were calibrated prior to the start ofthe experiment and their rates checked on a daily basis during the exposure. Samplingwas performed at day 3 at which time livers were removed and snap frozen in liquidnitrogen for parallel preparation of microsomes and isolation of total RNA.
PREPARATION OF MICROSOMES AND QUANTIFICATIONOF PROTEIN (BRADFORD ASSAY)
Microsomes were prepared by homogenizing livers from individual fish in 200 ml ofbuffer [0�1 M phosphate buffer (pH 7�8), 20% glycerol, proteinase inhibitor cocktail(Sigma)] before centrifuging at 10 000 g for 15 min at 4° C. Supernatant was removedand snap frozen before storing at �80° C. Protein concentrations were determinedusing the Bradford assay (Bradford, 1976)
2136 F. GEOGHEGAN ET AL .
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153

ETHOXYRESORUFIN-O -DEETHYLASE
Hundred micrograms of protein and 1 ml of 0�1 M phosphate buffer (pH 7�8) weremixed followed by the addition of 1 ml NADPH (25 mg resuspended in 600 ml phos-phate buffer) and finally 1 ml of ethoxyresorufin (8 mM in dimethylsulfoxide, Sigma).Tubes were inverted then incubated at 20° C for 30 min before terminating the reactionby addition of 1 ml ethanol. Excitation was measured at 547 nm and emission at 585 nmfor both samples and standards. Activity was calculated as pmol of product formed permin per mg of protein. Average values for each treatment group were calculated anddisplayed � S.D. Statistical significance was determined using Student’s t-test.
VTG AND SPIGGIN
VTG protein concentration was determined in stickleback plasma and spiggin pro-tein determined in stickleback kidney tissue using the enzyme-linked immunosorbentassay methods as previously shown (Andersson et al., 2007; Katsiadaki et al., 2002a, b,2006). Concentrations were calculated as mg per ml plasma for VTG and arbitraryunits per g body mass for spiggin. Average values for each treatment group were cal-culated and displayed with S.D. Statistical significance was determined using Student’st-test.
ISOLATION OF TOTAL RNA
RNA was isolated using the SV Total RNA isolation kit incorporating on-columngenomic DNA digestion (Promega), and integrity of RNA was verified by running 9 mlof the sample on a 1% agarose gel and quantified by spectrophotometry.
REVERSE TRANSCRIPTION FOR QUANTITATIVERT-PCR ANALYSIS
Reverse transcription was performed with Superscript II reverse transcriptase (Invi-trogen) by adding 300 ng of random primers and 1 ml of 10 mM deoxynucleotide tri-phosphates to 400 ng of total RNA from individual fish. After purification usinga QIA-Quick PCR purification kit (Qiagen, Crawley, U.K.), samples were diluted 1/10before use in quantitative RT-PCR analysis.
QUANTITATIVE RT-PCR ANALYSIS
Quantitative RT-PCR analysis of selected genes used a master mix consisting of1� SYBR green master mix (SensiMix; Quantace, London, U.K.), 10 mmol primersand 0�8 ml cDNA. Cycling parameters were used where 95° C for 10 min was usedto activate the hot start Taq enzyme followed by 40 cycles of 95° C for 15 s (denaturingstep) and 60° C for 1 min (combined annealing and extension). Ct values were determinedby first setting the baseline values where no amplification was occurring using the linearplot of fluorescence. Threshold values were then set manually by identification of expo-nential amplification using the log plot of fluorescence. Primers used were: for 18S rRNA,18S-L 59-TAGCACATCCAAAGAAGGCA-39, 18S-R 59-TCGATCCCGAGATC-CAACTA-39; ribosomal protein L8, L8-F 59-CGACCCGTACCGCTTCAAGAA-39,L8-R 59-GGACATTGCCAATGTTCAGCTGA-39; cyp1A, cyp1A-L 59-ACGTGCA-GATGTCAGACGAG-39, cyp1A-R 59-TTGGGTTTGTCGGAGAGAAG-39; VTG,sVTG-F 59-TTGGGACCCAAACAATCCCC-39, sVTG-R 59-AACGTCCCCAATC-TGTGCCT-39; ERa, ERa-F 59-CTGCCAGGCTTGCCGTCTTAG-39, ERa-R 59-AGTGCGTCTCTTGTCCCGCC-39.
Dissociation curves were plotted for all primer pairs to ensure that single productswere produced. Absolute fluorescence values were used to determine PCR efficiencies
STICKLEBACK MICROARRAY 2137
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153

for each well using the LinReg programme (Ramakers et al., 2003). Output from theprogramme was used to filter for efficiencies >1�45 so that outliers could be removed.PCR efficiency was averaged for each gene and used to normalize CT values followedby normalization using the geometric mean of two housekeeping genes: 18S and ribo-somal protein L8 before proceeding with the delta-delta Ct method of relative quanti-fication (Livak & Schmittgen, 2001). Statistical significance was determined usingStudent’s t-test.
MICROARRAY EXPERIMENTS
In this preliminary microarray experiment, RNA was pooled from four males or fourfemales for each sex–exposure group. Pools of treated fish samples were compared withthe relevant solvent controls. Dye swaps were carried out such that a total of 12 micro-arrays were analysed. Total RNA was prepared from 12 mg of pooled RNA from fourindividual fish for control or exposed samples. A similar protocol to that used withflounder microarrays was employed (Williams et al., 2003, 2006, 2007). Twelve micro-grams of total RNA was treated to remove DNA contamination using a DNA-free kit(Ambion, Huntingdon, U.K.), then reverse transcribed using 200 U Superscript II(Invitrogen) with oligo-dT (12–18) primers before repurification with a QIA-QuickPCR purification kit. A Bioanalyser 2100 Biosizing total RNA nano assay (Agilent,Stockport, U.K.) was used to assess the integrity of selected total RNA samples. cDNA(500 ng) was labelled with Cy5-dCTP or Cy3-dCTP (Amersham) using 40 U Klenowpolymerase (Invitrogen) with random primers. Labelled cDNA was purified by QIA-Quick and fluorophore incorporation measured by spectrophotometry at A650 forCy5 and A550 for Cy3. Slides were pre-hybridized in 25% formamide, 5� SSC (Ausubel,1995), 0�1% SDS, 10 mg ml�1 bovine serum albumin fraction V (Sigma) for 2 h at 42° C,followed two dips in deionized water and one in ethanol before drying by centrifugationat 2000 rpm in a Mistral 4L centrifuge (MSE, London, U.K.) with a swing-out rotor atroom temperature. Cy5- and Cy3- dye-labelled cDNA (60 pmol incorporated) werehybridized to the array in a total volume of 60 ml in a buffer containing 25% formamide,5� SSC, 0�1% SDS, 0�8 mg ml�1 Poly-A, 4 mg ml�1 yeast tRNA (Sigma) undera 25�60I Lifterslip (VWR, Lutterworth, U.K.) at 42° C for 18 h. Wash solutions werefiltered, slides were washed once in 2� SSC/0�1% SDS at 42° C, once in 0�1� SSC/0�1%SDS at room temperature and four times in 0�1� SSC at room temperature for 5 mineach followed two dips in deionized water and one in ethanol before drying by centrifu-gation as above. All slides were washed and dried in separate 50 ml tubes (Falcon, FosterCity, CA, U.S.A.). Slides were scanned with an Axon Genepix 4000A array scanner(Axon Instruments, Fort Worth, TX, U.S.A.) at 100% laser power with a photomultipliertube voltage of 600 in both channels. Genepix software (Axon Instruments) was used toassess the quality of individual spots and extract data for analysis; spots with low inten-sity or poor morphology were automatically flagged by Genepix software, and in addi-tion, all spots were manually checked and flagged, allowing removal of untrustworthydata in Genespring (Agilent, Santa Clara, CA, U.S.A.). The data used in furtheranalyses consisted of local background-subtracted median intensities. The array designis available in ArrayExpress as accession A-MAXD-39 and the experimental data asE-MAXD-39.
ANALYSIS OF MICROARRAY DATA
Genespring v7.2 software was used for analysis. Data were normalized using pergene, per chip, intensity dependent (Lowess) normalization and filtered to removelow-intensity and highly variable spots. Filtering employed an intensity cutoff derivedfrom the base–proportional score calculated by Genespring, followed by removal ofdata where S.D. between replicate spots was >1�4. Statistically significant gene expres-sion changes were identified by Student’s t-test (P < 0�05) with Benjamini & Hochberg(1995) multiple testing correction, resulting in a predicted false discovery rate of 0�05.
2138 F. GEOGHEGAN ET AL .
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153

This test was carried out to find clones that are reproducibly differentially expressed indye-swapped technical replicates, whereas to assess the effects of interindividualvariation, biological replicates would be required. Fold changes were determined bythe mean expression ratio of test:control. All clones significantly, and at least two-fold,altered in expression in comparison with controls in any treatment were clustered bycondition to determine the relationships among the gene expression levels of samples.The similarity measure used for clustering was Spearman correlation, and this was car-ried out in Genespring. All arrays, including dye swap experiments, were clustered indi-vidually. Duplicate clones of the same genes and clones showing no similarity to knownproteins were removed for clarity.
RESULTS
EST LIBRARIES, ANNOTATION AND MICROARRAYDEVELOPMENT
SSH resulted in 88 clones that were successfully sequenced, these sequenceswere submitted to dbEST at GenBank and were assigned accession numbersEG365008–EG365117. These included lipid transporters such as apolipoproteinAI and fatty acid-binding protein, ribosomal proteins and immune-function-related genes such as complement component C3 and chemotaxin.The size range of PCR-amplified clone inserts varied from <100 bp to >2 kb,
as shown by the non-overlap of some 59- and 39-sequences, which aligned to thesame gene. Of 9692 stickleback clones, 616 possessed no good quality sequence,920 were successfully sequenced but were not identifiable (by BLAST at E <10�6) with predicted cDNA sequences from the stickleback genome, ESTs orproteins from other organisms. The remaining 8156 clones were identified withknown sequences, 8046 from the GenBank nr protein database. Of these, 6864were annotated with one or more GO terms. An illustration of the diversityof GO terms assigned by Blast2GO is shown in Fig. 7. Thirty-eight clones wererejected because of inconsistent annotation, indicating potential chimerism orcross-contamination. Amplification of the spotted probes was 78�5% efficient asassessed by agarose gel electrophoresis. Sequence clustering by CAP3 suggestedthat 4972 distinct EST clusters were represented on the microarray, of these 4122were identifiable by BLASTx at E < 10�6 v. the GenBank non-redundant proteindatabase, while 850 displayed no such similarity to known proteins.In microarray optimization experiments (data not shown), the median S.D.
between three technical replicates was 0�067, the median S.D. between three bio-logical replicates was 0�247, the correlation between a forward-labelled andreverse-labelled sample in dye-reverse experiments was �0�857.
ETHOXYRESORUFIN-O -DEETHYLASE
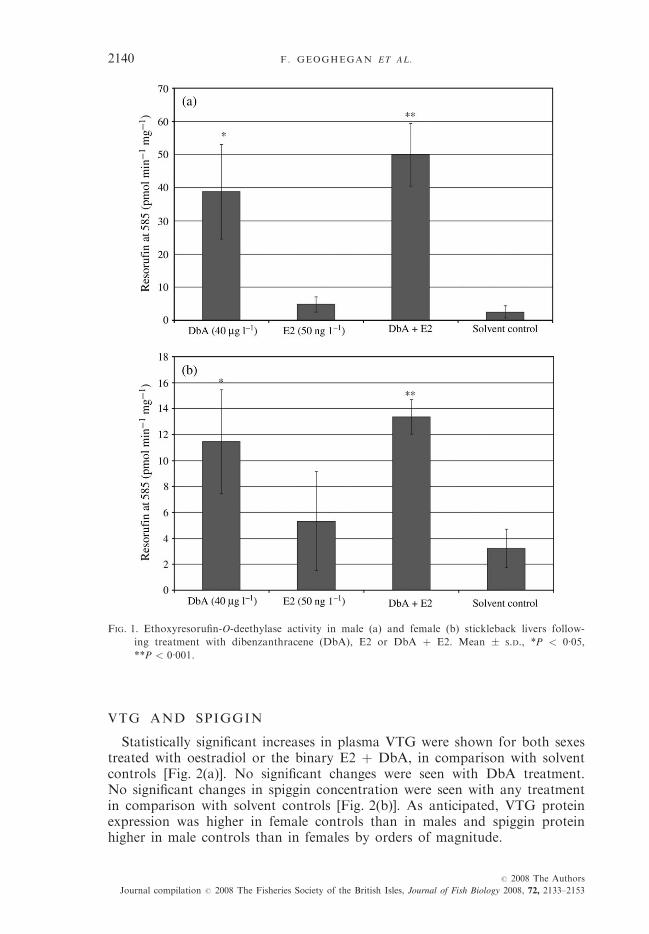
Although there was interindividual variation between fish under the sameconditions, mean values showed a range of responses in fish compared withcontrol. A significant induction of EROD activity was observed in both femaleand male fish following exposure to DbA and the binary DbA/E2 with no sig-nificant difference between the single and binary treatment [Fig. 1(a), (b)]. Nosignificant response was observed in the solvent control or following treatmentwith E2 (Fig. 1).
STICKLEBACK MICROARRAY 2139
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153
VTG AND SPIGGIN
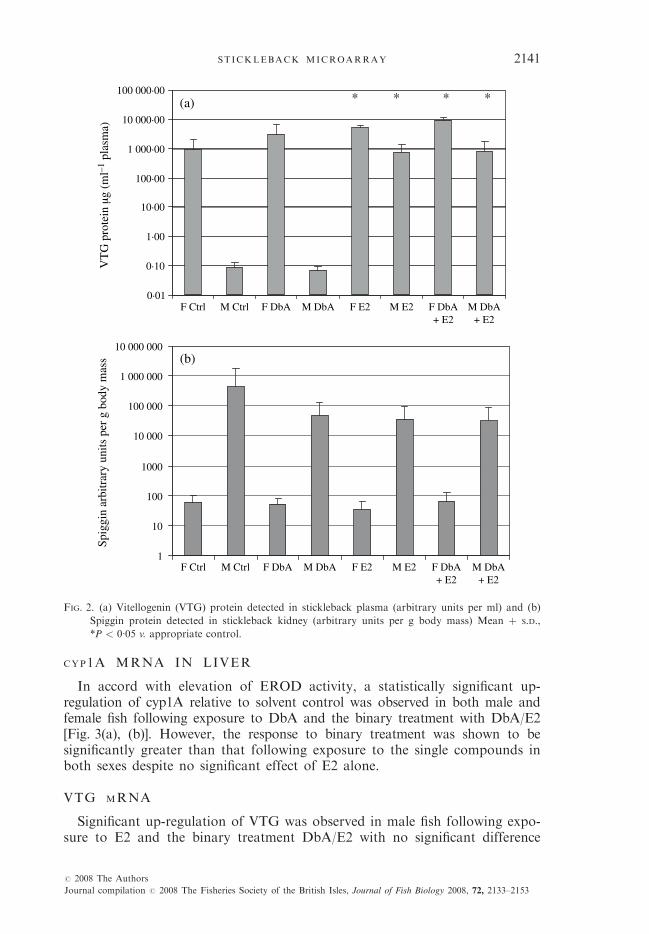
Statistically significant increases in plasma VTG were shown for both sexestreated with oestradiol or the binary E2 þ DbA, in comparison with solventcontrols [Fig. 2(a)]. No significant changes were seen with DbA treatment.No significant changes in spiggin concentration were seen with any treatmentin comparison with solvent controls [Fig. 2(b)]. As anticipated, VTG proteinexpression was higher in female controls than in males and spiggin proteinhigher in male controls than in females by orders of magnitude.
FIG. 1. Ethoxyresorufin-O-deethylase activity in male (a) and female (b) stickleback livers follow-
ing treatment with dibenzanthracene (DbA), E2 or DbA þ E2. Mean � S.D., *P < 0�05,**P < 0�001.
2140 F. GEOGHEGAN ET AL .
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153
CYP1A MRNA IN LIVER
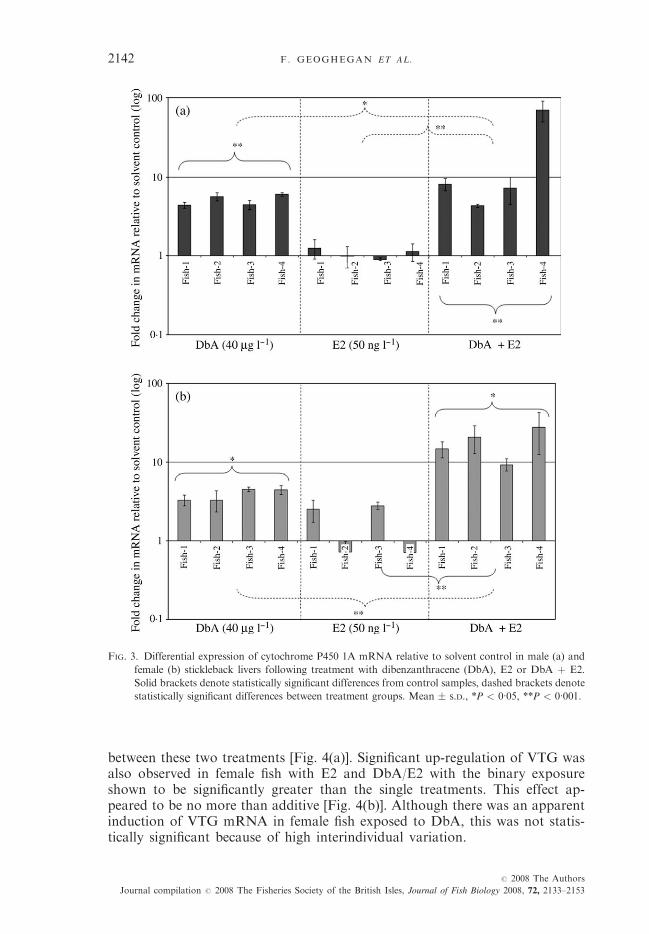
In accord with elevation of EROD activity, a statistically significant up-regulation of cyp1A relative to solvent control was observed in both male andfemale fish following exposure to DbA and the binary treatment with DbA/E2[Fig. 3(a), (b)]. However, the response to binary treatment was shown to besignificantly greater than that following exposure to the single compounds inboth sexes despite no significant effect of E2 alone.
VTG MRNA
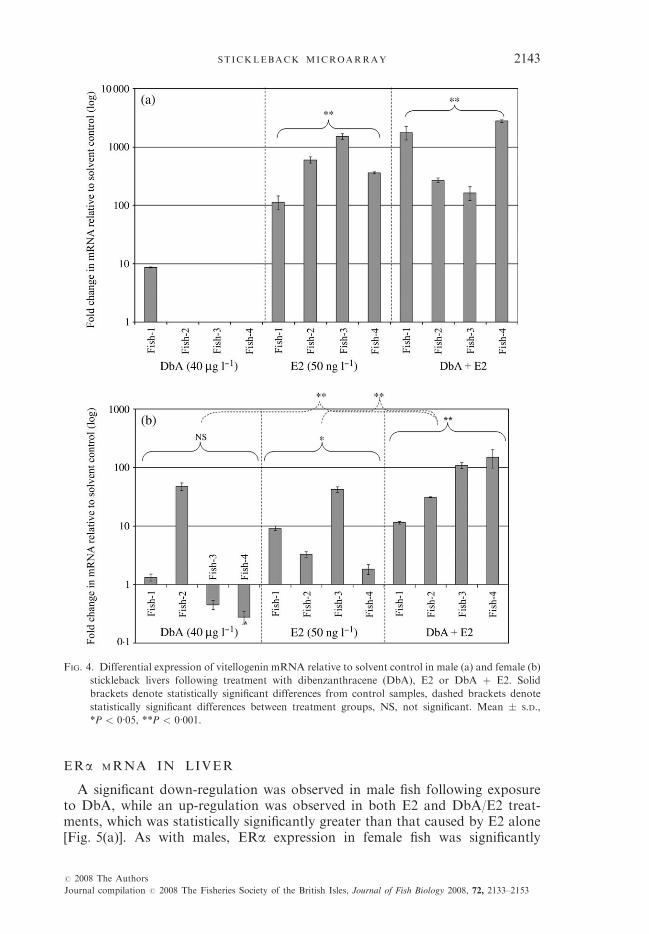
Significant up-regulation of VTG was observed in male fish following expo-sure to E2 and the binary treatment DbA/E2 with no significant difference
1
10
100
1000
10 000
100 000
1 000 000
10 000 000
Spig
gin
arbi
trar
y un
its p
er g
bod
y m
ass
0·01
0·10
1·00
10·00
100·00
1 000·00
10 000·00
100 000·00
F Ctrl M Ctrl F DbA F DbA+ E2
M DbA+ E2
M DbA F E2 M E2
F Ctrl M Ctrl F DbA F DbA+ E2
M DbA+ E2
M DbA F E2 M E2
VT
G p
rote
in µ
g (m
l–1 p
lasm
a)* * * *
(b)
(a)
FIG. 2. (a) Vitellogenin (VTG) protein detected in stickleback plasma (arbitrary units per ml) and (b)
Spiggin protein detected in stickleback kidney (arbitrary units per g body mass) Mean þ S.D.,
*P < 0�05 v. appropriate control.
STICKLEBACK MICROARRAY 2141
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153
between these two treatments [Fig. 4(a)]. Significant up-regulation of VTG wasalso observed in female fish with E2 and DbA/E2 with the binary exposureshown to be significantly greater than the single treatments. This effect ap-peared to be no more than additive [Fig. 4(b)]. Although there was an apparentinduction of VTG mRNA in female fish exposed to DbA, this was not statis-tically significant because of high interindividual variation.
FIG. 3. Differential expression of cytochrome P450 1A mRNA relative to solvent control in male (a) and
female (b) stickleback livers following treatment with dibenzanthracene (DbA), E2 or DbA þ E2.
Solid brackets denote statistically significant differences from control samples, dashed brackets denote
statistically significant differences between treatment groups. Mean � S.D., *P < 0�05, **P < 0�001.
2142 F. GEOGHEGAN ET AL .
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153
ERa MRNA IN LIVER
A significant down-regulation was observed in male fish following exposureto DbA, while an up-regulation was observed in both E2 and DbA/E2 treat-ments, which was statistically significantly greater than that caused by E2 alone[Fig. 5(a)]. As with males, ERa expression in female fish was significantly
FIG. 4. Differential expression of vitellogenin mRNA relative to solvent control in male (a) and female (b)
stickleback livers following treatment with dibenzanthracene (DbA), E2 or DbA þ E2. Solid
brackets denote statistically significant differences from control samples, dashed brackets denote
statistically significant differences between treatment groups, NS, not significant. Mean � S.D.,
*P < 0�05, **P < 0�001.
STICKLEBACK MICROARRAY 2143
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153
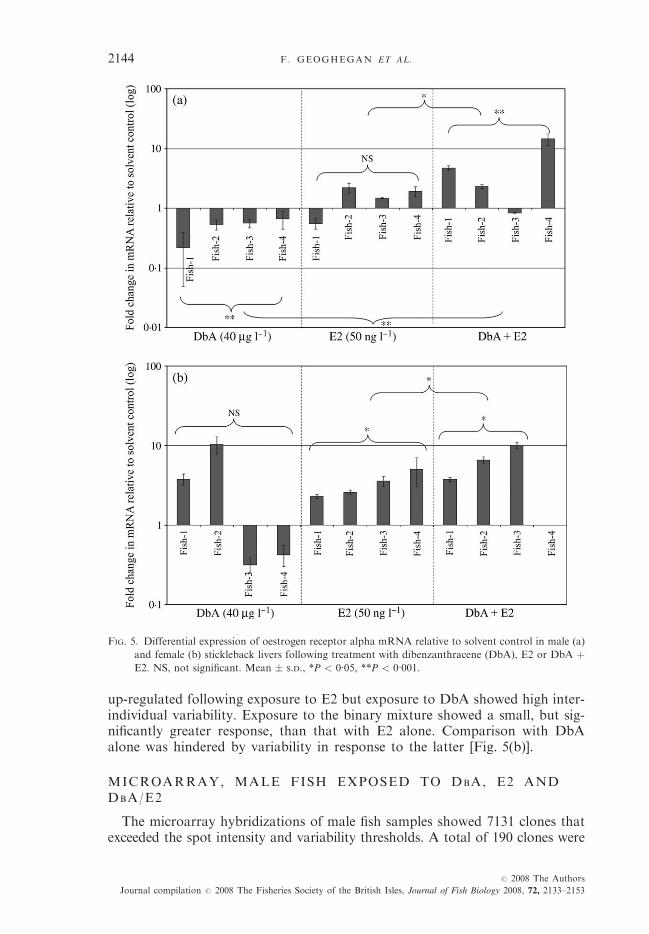
up-regulated following exposure to E2 but exposure to DbA showed high inter-individual variability. Exposure to the binary mixture showed a small, but sig-nificantly greater response, than that with E2 alone. Comparison with DbAalone was hindered by variability in response to the latter [Fig. 5(b)].
MICROARRAY, MALE FISH EXPOSED TO DBA, E2 ANDDBA/E2
The microarray hybridizations of male fish samples showed 7131 clones thatexceeded the spot intensity and variability thresholds. A total of 190 clones were
FIG. 5. Differential expression of oestrogen receptor alpha mRNA relative to solvent control in male (a)
and female (b) stickleback livers following treatment with dibenzanthracene (DbA), E2 or DbA þE2. NS, not significant. Mean � S.D., *P < 0�05, **P < 0�001.
2144 F. GEOGHEGAN ET AL .
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153

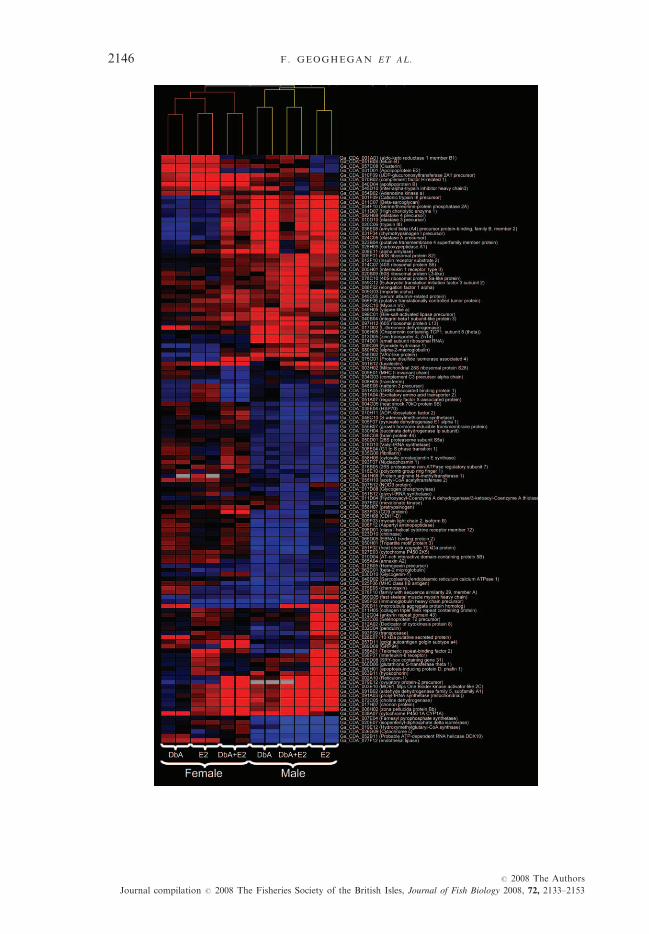
statistically significantly up- or down-regulated more than two-fold in res-ponse to DbA, E2 or the binary mixture. Figure 6 shows differentially expres-sed clones observed following treatment of both male and female fish, theexpression values for these clones are shown in Table SI.Clones down-regulated in all treatments included sarcoplasmic/endoplasmic
reticulum calcium ATPase 1, ATP-dependent RNA helicase DDX10, three genesinvolved in lipid metabolism, endothelial lipase, isopentenyl-diphosphate deltaisomerase and farnesyl pyrophosphate synthetase and cytochrome c, the mito-chondrial electron transport protein and apoptosis effector. The molecular chap-erone heat-shock protein 70 was also found down-regulated in all treatments.The biomarker cyp1A was highly up-regulated in both DbA and binary ex-
posures, mirroring the results of the quantitative RT-PCR experiments. Alsoinduced in DbA and binary exposures were VAV-like protein, a possible onco-protein (Schuebel et al., 1998) and integrin b1 subunit 3, a receptor involved incell adhesion and recognition.Clones involved in oestrogenic responses such as zona pellucida/chorion pro-
tein were found highly up-regulated with both E2 and DbA/E2 but not follow-ing DbA treatment (Fig. 6); additionally, a transcript encoding ovulatoryprotein-2 precursor was induced. Protein trafficking-associated genes reticulon1 and importin were similarly induced, as were the mitochondrial transcriptsprolyl-tRNA synthetase and aldehyde dehydrogenase 5A1. Hyosophorin wassignificantly up-regulated only following exposure to E2, as were SRY-boxcontaining gene 31, apoptosis-inducing gene phafin and glutathione-S-transferasetheta 1. Genes induced only in response to the binary treatment included thoserelating to translation, such as ribosomal proteins and eukaryotic translationinitiation and elongation factors.
MICROARRAY, FEMALE FISH EXPOSED TO DBA, E2 ANDDBA/E2
The microarray hybridizations of female fish samples showed 7258 clonesthat exceeded the spot intensity and variability cutoff thresholds. Of these,a total of 94 clones were statistically significantly up- or down-regulated morethan two-fold in response to DbA, E2 or the binary mixture. Figure 6 showsdifferentially expressed clones observed following treatment of both male andfemale fish; the expression values for these clones are shown in Table SI.cyp1A was the only gene up-regulated following exposure to DbA and the
binary treatment in both males and females, and apolipoprotein B was inducedwith all treatments in females. E2 treatment of females resulted in few geneexpression changes, with only the apolipoproteins being induced more thanthree-fold. However, the female binary treatment resulted in induction of anumber of genes in common with males. These included zona pellucida andchorion proteins, fucolectin, prolyl-tRNA synthetase, aldehyde dehydrogenase5A1, choline dehydrogenase, MOB1, ovulatory protein-2 and reticulon 1.UDP-glucuronosyltransferase 2A1 was only induced in females, while hyoso-phorin was induced only in males (Fig. 6).
STICKLEBACK MICROARRAY 2145
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153
2146 F. GEOGHEGAN ET AL .
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153

Genes involved in the immune response such as the complement factor H-related I were found up-regulated in both E2 and binary treatments, whileMHC IIA antigen, the MHCII variant chain, immunoglobin light chain vari-able region were found down-regulated following exposure to the binary treat-ment only with the exception of the immunoglobin heavy chain precursor,which was found down-regulated in E2 and the DbA/E2 exposures (Fig. 6).
DISCUSSION
These results indicate statistically significant increases in both cyp1A mRNAexpression and EROD activity in both male and female fish following exposureto DbA and following the binary exposure to DbA and E2. An up-regulation ofcyp1A was also detected for both sexes following use of the microarray. A sig-nificant difference between single and binary exposures was found following theqPCR assay in which the binary treatment resulted in a significantly higherexpression of cyp1A. This is in contrast to studies that have shown that in roachRutilus rutilus (L.), E2 exposure attenuated EROD induction following exposureto a PAH, suggesting an inhibitory effect (O’Hare et al., 1995). The relative con-centrations are likely to be major determinants of the response.Previous work with rainbow trout Oncorhynchus mykiss (Walbaum) hepato-
cytes has suggested that exposure to E2 results in decreased basal levels of cyp1A(Navas & Segner, 2001) in terms of both EROD activity and mRNA expressionand that the oestrogen receptor was suggested to be directly involved. In the cur-rent study, exposure of the three-spined stickleback to E2 resulted in no signif-icant change in EROD activity or cyp1A expression in either male or female fish,suggesting that in this case, E2 does not have an inhibitory effect.Down-regulation of ERa mRNA was observed in male fish following expo-
sure to DbA, which suggests an inhibitory effect of the PAH (although VTGexpression was unchanged). An inhibitory effect of the AhR-agonist TCDDon both ERa gene expression and protein level has been previously observed(Tian et al., 1998; Wormke et al., 2000, 2003), AhR is known to interactdirectly with the oestrogen receptor; however, the mechanism by which ERais decreased has yet to be fully elucidated although degradation by proteo-somes has been demonstrated (Wormke et al., 2000). No significant changein ERa expression was observed in female fish exposed to DbA, which couldbe because of relatively high endogenous levels of E2.VTG mRNA and VTG serum protein were highly induced in both sexes in
both the E2 and binary treatments. Results from the microarray experimentsindicated an up-regulation of genes associated particularly with vitellogenesisin female fish following exposure to the binary DbA and E2, while the singleE2 exposure resulted in no great change in expression, likely because of high
FIG. 6. Spearman cluster diagram and heat-map showing all clones statistically significant, and at least
two-fold, altered in expression in comparison with controls in any treatment. Each column
represents one microarray, including dye swaps. Duplicate clones of the same genes and clones
showing no similarity to known proteins were removed for clarity. Increased expression relative to
the relevant controls is indicated in red, decreased expression in blue and no data in grey. Intensity of
colour indicates the magnitude of change to a maximum intensity at three-fold.
STICKLEBACK MICROARRAY 2147
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153
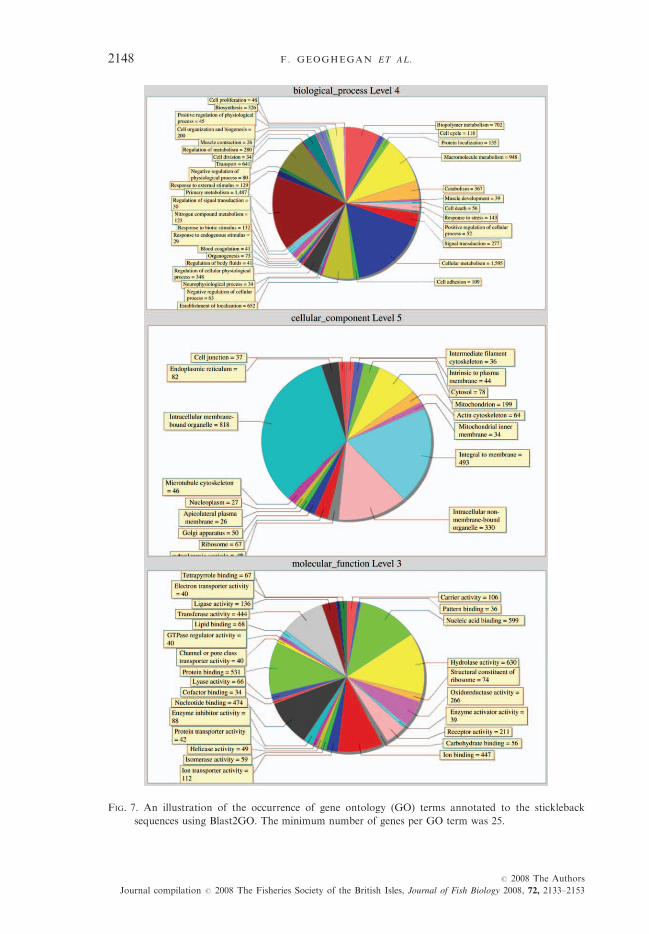
FIG. 7. An illustration of the occurrence of gene ontology (GO) terms annotated to the stickleback
sequences using Blast2GO. The minimum number of genes per GO term was 25.
2148 F. GEOGHEGAN ET AL .
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153

endogenous levels of E2. This is in contrast to the VTG-associated genesfound differentially regulated in male fish in which marked up-regulationwas observed in the single E2 treatment in addition to binary exposure. Inaddition to the zona pellucida and chorion proteins, ovulatory protein-2was induced in both male and female binary treatments, and hyosophorinwas up-regulated by E2 only in male fish. Ovulatory protein-2 precursor(Garczynski & Goetz, 1997) is a homologue of mammalian WAP four sul-phide core domain 3, a likely extracellular protease inhibitor; however, thesegenes are currently poorly characterized. Hyosophorin is a polysialoglycopro-tein and is a component of cortical granules, which will be exocytosed intothe perivitelline space during the cortical reaction. It is suggested to beinvolved in water accumulation, and its ability to bind calcium suggests a roleas a calcium reservoir (Tsao et al., 1999). Hyosophorin has been cloned fromcarp Cyprinus carpio L. and sequence analysis has demonstrated a series ofrepeat units that will be cleaved following exocytosis so that it consists of justone repeat unit. No previous studies have demonstrated changes in expressionof this gene following exposure to oestrogens. These two genes provide can-didate oestrogenic response biomarkers, and the fact that hyosophorin induc-tion was only found in male fish is interesting and may imply oestrogenreceptor-mediated induction, potentially its high expression in male fish mightbe related to the increased need for water excretion in conditions of unantic-ipated VTG accumulation.While it would be premature to propose detailed mechanistic actions based
on this limited microarray study, certain gene expression changes highlightadditional processes that may benefit from further examination. These includechanges in expression of transcripts encoding lipid metabolic and transportingproteins, a VAV-like protein (Bustelo, 2001) and a b-integrin (Wu, 2006) thatcould potentially affect carcinogenesis and those involved in translation, zinctransport and immune response. For example, the immune response genesfound differentially expressed included those associated with specific immunitysuch as the major histocompatibility complex in addition to those involvedwith non-specific–innate immunity using the complement system. Previousstudies have demonstrated the effects of PAH exposure on specific immunity,reviewed in Reynaud & Deschaux (2006), with suppression observed in res-ponse to some compounds such as TCDD (Arkoosh et al., 1994); in this study,a down-regulation was observed in response to the binary DbA/E2 treatmentwith some down-regulation also observed in male fish but not in female fishfollowing exposure to DbA. These differences between male and female fishare likely because of the differences in endogenous levels of hormones, whichare known to affect the immune response of fish (Harris & Bird, 2000).This microarray experiment represented a proof-of-principle, which, as
well as providing validation of the microarray by comparison with qPCRdata, provided further information on the molecular responses of the sticklebackto these binary toxicant treatments. Clustering showed segregation of male andfemale fish and replicate dye-swap hybridizations clustered together in all cases.However, for more comprehensive analyses of gene expression, the authors arecurrently investigating responses in individual fish, with a greater number ofbiological replicates in order to maintain the statistical power of the analyses.
STICKLEBACK MICROARRAY 2149
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153

The stickleback microarray is currently being updated with additionalhepatic cDNA libraries, two normalized and one subtractive, in order toincrease representation of liver-associated clones, such as VTG, which wasabsent from this array. These libraries have been constructed and sequenced(Brown et al., in press) and will result in an increase to c. 15 000 clones onthe microarray. Annotation will also be periodically updated as further infor-mation becomes available on the stickleback genome. In addition, cDNAclones are now available from the stickleback genome consortium, such thatany especially interesting genes not represented in the libraries may be added.The updated stickleback cDNA microarray and the flounder Platichthys
flesus (L.) array (Williams et al., 2006, 2007) are being used in a multi-centrefish toxicogenomics project in which the aim was to relate transcriptomics, pro-teomics, metabolomics, genetics and individual biomarkers towards identifyingand defining the bases of individual and population susceptibility and adapta-tion to environmental pollutants in fish. Additionally, this technology is beingused to profile stickleback responses to nanoparticles.This microarray will be made available to the scientific community subse-
quent to first publication of data from the authors’ current stickleback experi-ments. The microarray, because of the multi-tissue basis of its construction,could be applied to a variety of investigations involving G. aculeatus. Althoughthe authors’ research has focused on liver, there is the potential for transcrip-tomic investigation of other organs, such as gill, kidney, brain and gonads orwhole juvenile fish. Additionally, the flounder microarray has been used in anassessment of cross-species hybridization efficiency (Cohen et al., 2007) where itwas demonstrated that within-order cross-hybridization is likely to be effective,which for order Gasterosteiformes includes, as well as sticklebacks, the sea-horsesand pipefishes Syngnathiae. The advantage of a cDNA microarray platform is itsflexibility, within a range of species, to address any problem requiring geneexpression analysis.
SUPPLEMENTARY MATERIAL
TABLE SI. Gene expression and P values for clones whose expression is illus-trated in Fig. 6.
This work was funded by a Biotechnology & Biological Sciences Research Council–Centre for Environment, Fisheries & Aquaculture Science (BBSRC-CEFAS) joint student-ship (F. G.) and U.K. Natural Environment Research Council (NERC) post-genomic andproteomic programme grant NE/C507661/1, Birmingham functional genomics facilities byBBSRC grant 6/JIF 13209 and bioinformatics byMRC infrastructure grant G.4500017withadditional facilities provided by NERC. We thank D. Kingsley and J. Grimwood for theirkind provision of stickleback clones, Marion Sebire for assistance with fish exposures andA. Jones and L. Klovrza for assistance with genomics.
References
Anderson, M. J., Miller, M. R. & Hinton, D. E. (1996). In vitro modulation of 17-beta-estradiol-induced vitellogenin synthesis: effects of cytochrome P4501A1 inducingcompounds on rainbow trout (Oncorhynchus mykiss) liver cells. Aquatic Toxicology34, 327–350.
2150 F. GEOGHEGAN ET AL .
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153

Andersson, C., Katsiadaki, I., Lundstedt-Enkel, K. & Orberg. J. (2007). Effects of 17a-ethynylestradiol on EROD activity, spiggin and vitellogenin in three-spinedstickleback (Gasterosteus aculeatus). Aquatic Toxicology 83, 33–42.
Ankley, G. T., Daston, G. P., Degitz, S. J., Denslow, N. D., Hoke, R. A., Kennedy,S. W., Miracle, A. L., Perkins, E. J., Snape, J., Tillitt, D. E., Tyler, C. R. & Versteeg,D. (2006). Toxicogenomics in regulatory ecotoxicology. Environment Science andTechnology 40, 4055–4065.
Arkoosh, M. R., Clemons, E., Myers, M. & Casillas, E. (1994). Suppression of B-cellmediated immunity in juvenile Chinook salmon (Oncorhynchus tshawytscha) afterexposure to either a polycyclic aromatic hydrocarbon or to polychlorinatedbiphenyls. Immunopharmacology and Immunotoxicology 16, 293–314.
Ausubel, F. M. (1995). Short Protocols in Molecular Biology. Chichester: Wiley.Beischlag, T. V. & Perdew, G. H. (2005). ER alpha-AHR-ARNT protein-protein
interactions mediate estradiol-dependent transrepression of dioxin-inducible genetranscription. Journal of Biological Chemistry 280, 21607–21611.
Benjamini, Y. & Hochberg, Y. (1995). Controlling the false discovery rate: a practical andpowerful approach to multiple testing. Journal of the Royal Statistical Society B 57,289–300.
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgramquantities of protein utilizing the principle of protein-dye binding. AnalyticalBiochemistry 72, 248–254.
Brown, M. M., Williams, T. D., Chipman, J. K., Katsiadaki, I., Sanders, M. & Craft,J. A. (in press). Construction of subtracted EST and normalized cDNA librariesfrom liver of chemical-exposed three-spined stickleback (Gasterosteus aculeatus)containing pollutant responsive genes as a resource for transcriptome analysis.Marine Environmental Research. doi: 10.1016/j.marenvres.2008.02.043
Bustelo, X. R. (2001). Vav proteins, adaptors and cell signaling. Oncogene 20, 6372–6381.Casillas, E., Misitano, D., Johnson, L., Rhodes, L., Collier, T. K., Stein, J. E., McCain,
B. B. & Varanasi, U. (1991). Inducibility of spawning and reproductive success offemale English sole (Parophrys vetulus) from urban and nonurban areas of Pugetsound, Washington. Marine Environmental Research 31, 99–122.
Cohen, R., Chalifa-Caspi, V., Williams, T. D., Auslander, M., George, S. G., Chipman,J. K. & Tom, M. (2007). Estimating the efficiency of fish cross-species cDNAmicroarray hybridization. Marine Biotechnology New York 9, 491–499.
Conesa, A., Gotz, S., Garcia-Gomez, J. M., Terol, J., Talon, M. & Robles, M. (2005).Blast2GO: a universal tool for annotation, visualization and analysis in functionalgenomics research. Bioinformatics 21, 3674–3676.
Durinck, S., Moreau, Y., Kasprzyk, A., Davis, S., De Moor, B., Brazma, A. & Huber, W.(2005). BioMart and bioconductor: a powerful link between biological databasesand microarray data analysis. Bioinformatics 21, 3439–3440.
Elskus, A. A., Pruell, R. & Stegeman, J. J. (1992). Endogenously-mediated, pretransla-tional suppression of cytochrome P4501A in PCB-contaminated flounder. MarineEnvironmental Research 32, 97–101.
Ewing, B. & Green, P. (1998). Base-calling of automated sequencer traces using phred. II.Error probabilities. Genome Research 8, 186–194.
Ewing, B., Hillier, L., Wendl, M. C. & Green, P. (1998). Base-calling of automatedsequencer traces using phred. I. Accuracy assessment. Genome Research 8,175–185.
Garczynski, M. A. & Goetz, F. W. (1997). Molecular characterization of a ribonucleicacid transcript that is highly up-regulated at the time of ovulation in the brooktrout (Salvelinus fontinalis) ovary. Biology of Reproduction 57, 856–864.
Hahlbeck, E., Katsiadaki, I., Mayer, I., Adolfsson-Erici, M., James, J. D. & Bengtsson,B.-E. (2004). The juvenile three-spined stickleback (Gasterosteus aculeatus L.) asa model organism for endocrine disruption II - kidney hypertrophy, vitellogeninand spiggin induction. Aquatic Toxicology 70, 311–326.
Harris, J. & Bird, D. J. (2000). Modulation of the fish immune system by hormones.Veterinary Immunology and Immunopathology 77, 163–176.
STICKLEBACK MICROARRAY 2151
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153

Huang, X. & Madan, A. (1999). CAP3: a DNA sequence assembly program. GenomeResearch 9, 868–877.
Katsiadaki, I., Scott, A. P. & Mayer, I. (2002a). The potential of the three-spinedstickleback (Gasterosteus aculeatus L.) as a combined biomarker for oestrogens andandrogens in European waters. Marine Environmental Research 54, 725–728.
Katsiadaki, I., Scott, A. P., Hurst, M. R., Matthiessen, P. & Mayer, I. (2002b). Detectionof environmental androgens: a novel method based on enzyme-linked immuno-sorbent assay of spiggin, the stickleback (Gasterosteus aculeatus) glue protein.Environmental Toxicology and Chemistry 21, 1946–1954.
Katsiadaki, I., Morris, S., Squires, C., Hurst, M. R., James, J. D. & Scott, A. P. (2006). Asensitive, in vivo test for the detection of environmental anti-androgens, using thethree-spined stickleback (Gasterosteus aculeatus). Environmental Health Perspectives114 (Suppl. 1) 115–121.
Kingsley, D. M., Zhu, B., Osoegawa, K., De Jong, P. J., Schein, J., Marra, M., Peichel,C., Amemiya, C., Schulter, D., Balabhadra, S., Friedlander, B., Cha, Y. M.,Dickson, M., Grimwood, J., Schmutz, J., Talbot, W. S. & Myers, R. (2004). Newgenomic tools for molecular studies of evolutionary change in threespine stickle-backs. Behaviour 141, 1331–1344.
Li, S. & Chou, H. H. (2004). LUCY2: an interactive DNA sequence quality trimming andvector removal tool. Bioinformatics 20, 2865–2866.
Livak, K. J. & Schmittgen, T. D. (2001). Analysis of relative gene expression data usingreal-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25,402–408.
Malins, D. C. & Ostrander, G. K. (1994). Aquatic Toxicology: Molecular and CellularPerspectives. Boca Raton: Lewis.
Navas, J. M. & Segner, H. (2001). Estrogen-mediated suppression of cytochrome P4501A(cyp1A) expression in rainbow trout hepatocytes: role of estrogen receptor.Chemico-Biological Interactions 138, 285–298.
O’Hare, D., Siddall, R., Robotham, P. W. J. & Gill, R. A. (1995). Influence of oestradiolon induction of EROD activity in roach (Rutilus rutilus L.). Chemosphere 30,1423–1428.
Pereira, J. J., Ziskowski, J., Mercaldo-Allen, R., Kuropat, C., Luedke, D. & Gould, E.(1992). Vitellogenin in winter flounder (Pleuronectes americanus) from Long IslandSound and Boston Harbor. Estuaries 15, 289–297.
Ramakers, C., Ruijter, J. M., Deprez, R. H. & Moorman, A. F. (2003). Assumption-freeanalysis of quantitative real-time polymerase chain reaction (PCR) data. Neuro-science Letters 339, 62–66.
Reynaud, S. & Deschaux, P. (2006). The effects of polycyclic aromatic hydrocarbons onthe immune system of fish: a review. Aquatic Toxicology 77, 229–238.
Safe, S. & Wormke, M. (2003). Inhibitory aryl hydrocarbon receptor-estrogen receptoralpha cross-talk and mechanisms of action. Chemical Research in Toxicology 16,807–816.
Schuebel, K. E., Movilla, N., Rosa, J. L. & Bustelo, X. R. (1998). Phosphorylation-dependent and constitutive activation of Rho proteins by wild-type and oncogenicVav-2. EMBO Journal 17, 6608–6621.
Tian, Y., Ke, S., Thomas, T., Meeker, R. J. & Gallo, M. A. (1998). Transcriptionalsuppression of estrogen receptor gene expression by 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Journal of Steroid Biochemistry and Molecular Biology 67, 17–24.
Tsao, C. C., Huang, F. L. & Chang, Y. S. (1999). Purification, characterization, andmolecular cloning of carp hyosophorin. Molecular Reproduction and Development52, 288–296.
Williams, T. D., Gensberg, K., Minchin, S. D. & Chipman, J. K. (2003). A DNAexpression array to detect toxic stress response in European flounder (Platichthysflesus). Aquatic Toxicology 65, 141–157.
Williams, T. D., Diab, A. M., George, S. G., Godfrey, R. E., Sabine, V., Conesa, A.,Minchin, S. D., Watts, P. C. & Chipman, J. K. (2006). Development of theGENIPOL European flounder (Platichthys flesus) microarray and determination of
2152 F. GEOGHEGAN ET AL .
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153

temporal transcriptional responses to cadmium at low dose. Environment Scienceand Technology 40, 6479–6488.
Williams, T. D., Diab, A. M., George, S. G., Sabine, V. & Chipman, J. K. (2007). Geneexpression responses of European flounder (Platichthys flesus) to 17-beta estradiol.Toxicology Letters 168, 236–248.
Wormke, M., Stoner, M., Saville, B. & Safe, S. (2000). Crosstalk between estrogenreceptor alpha and the aryl hydrocarbon receptor in breast cancer cells involvesunidirectional activation of proteasomes. FEBS Letters 478, 109–112.
Wormke, M., Stoner, M., Saville, B., Walker, K., Abdelrahim, M., Burghardt, R. & Safe,S. (2003). The aryl hydrocarbon receptor mediates degradation of estrogenreceptor alpha through activation of proteasomes. Molecular and Cellular Biology23, 1843–1855.
Wu, W. S. (2006). The signaling mechanism of ROS in tumor progression. Cancer andMetastasis Reviews 25, 695–705.
STICKLEBACK MICROARRAY 2153
# 2008 The Authors
Journal compilation # 2008 The Fisheries Society of the British Isles, Journal of Fish Biology 2008, 72, 2133–2153
Related Documents











