3D dental microwear texture analysis of feeding habits of sympatric ruminants in the Białowie _ za Primeval Forest, Poland Gildas Merceron a,⇑ , Emilia Hofman-Kamin ´ ska b , Rafał Kowalczyk b a iPHEP, UMR 7262, CNRS & University of Poitiers, France b Mammal Research Institute, Polish Academy of Sciences, Białowie _ za, Poland article info Article history: Received 18 December 2013 Received in revised form 23 May 2014 Accepted 24 May 2014 Keyword: Bison bonasus Cervus elaphus Capreolus capreolus Alces alces Diet Food niche overlap abstract With four species of ruminants, including red deer, roe deer, moose, and European bison, the Białowie _ za Primeval Forest is unique on the European continent, where only one to three ungulate species are usu- ally found. The present study is the first effort to explore the dietary overlap of a European community of sympatric ruminants using 3D dental microwear texture analysis. Results obtained for ungulates from the Białowie _ za Forest were compared with those of four species with well known differences in diet (semi-wild Heck cattle, African buffaloes, giraffes, and yellow-backed duikers). These ruminants frame the spectrum of expected 3D dental microwear textures among ruminants: C3 and C4 grazers share high anisotropy and low to intermediate complexity while browsers display intermediate to high complexity associated with low anisotropy. No significant differences between browsers (moose and roe deer) and mixed feeders (red deer) were detected by this analysis supporting a significant overlap in diet found in Białowie _ za. The results on the fourth ruminant are unequivocal: the European bison is not grazer but instead is highly engaged in browsing. Furthermore, through 3D dental microwear texture analysis, the high plasticity in feeding behavior of the European bison can be tracked depending on the seasons and on available access to feeding supplements during the winter. Ó 2014 Published by Elsevier B.V. 1. Introduction Evidence of overlap in resource use by sympatric species is essential for the understanding of interspecific competition (Mysterud, 2000). Often, resource use by one species reduces avail- ability of resources for another species or leads to species displace- ment (Putman, 1996; Ferretti et al., 2008). Competition for resource exploitation is one of the main mechanisms for interac- tion among large ungulates. Such interaction may occur when ungulates share similar habitats, an overlap in food consumed and when resources are limited (Gordon and Illius, 1989; Mysterud, 2000). Evidence of interactions among ungulates and other groups of species in natural ecosystems is limited (Putman, 1996). The Białowie _ za Primeval Forest, located on the Polish-Belarus- sian border is one of the best preserved forests in Europe (Je ˛ drzejewska and Je ˛ drzejewski, 1998). With five species of ungulates (including red deer Cervus elaphus, roe deer Capreolus capreolus, moose Alces alces, European bison Bison bonasus and wild boar Sus scrofa), it is unique on the European continent, largely inhabited by only one to three ungulate species (Okarma, 1995). Ungulates coexist here with a high diversity of carnivores, includ- ing wolves and lynx. These predators hunt roe and red deer almost exclusively (Je ˛ drzejewska et al., 1997; Je ˛ drzejewska and Je ˛ drzejewski, 1998). Deciduous and mixed forests with rich under- growth and herb layers offer good conditions for ungulates and may limit possible competition among them. In historical times, high densities of ungulates maintained for hunting purposes, together with cattle grazing in the forest (Samojlik and Kuijper, 2013), had a strong impact on tree stands and resulted in species competition. The two largest herbivores, the bison and the moose, were especially shaped by intra- and inter-specific competition for food resources (Je ˛ drzejewska et al., 1997; Je ˛ drzejewska and Je ˛ drzejewski, 1998). Nowadays, populations of ungulates in Białowie _ za occur in moderate densities and their numbers are increasing (Borowik et al., 2013; Kowalczyk et al., 2013). Cervids and European bison – the only bovidae species in the Białowie _ za Forest – are characterized by different feeding habits (Hoffman, 1989). The moose (A. alces) is the largest cervid. Its diet is mainly based on browsing trees and shrubs (birches, willows, rowan etc.) and herbaceous dicots; grasses and sedges being very rare in this diet (Morow, 1976; Franzmann, 1981). The red deer occupies various habitats from forest to moorland. In Europe, its http://dx.doi.org/10.1016/j.foreco.2014.05.041 0378-1127/Ó 2014 Published by Elsevier B.V. ⇑ Corresponding author. Tel.: +33 (0)5 49 36 63 05. E-mail address: [email protected] (G. Merceron). Forest Ecology and Management 328 (2014) 262–269 Contents lists available at ScienceDirect Forest Ecology and Management journal homepage: www.elsevier.com/locate/foreco

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Forest Ecology and Management 328 (2014) 262–269
Contents lists available at ScienceDirect
Forest Ecology and Management
journal homepage: www.elsevier .com/ locate/ foreco
3D dental microwear texture analysis of feeding habits of sympatricruminants in the Białowie _za Primeval Forest, Poland
http://dx.doi.org/10.1016/j.foreco.2014.05.0410378-1127/� 2014 Published by Elsevier B.V.
⇑ Corresponding author. Tel.: +33 (0)5 49 36 63 05.E-mail address: [email protected] (G. Merceron).
Gildas Merceron a,⇑, Emilia Hofman-Kaminska b, Rafał Kowalczyk b
a iPHEP, UMR 7262, CNRS & University of Poitiers, Franceb Mammal Research Institute, Polish Academy of Sciences, Białowie _za, Poland
a r t i c l e i n f o a b s t r a c t
Article history:Received 18 December 2013Received in revised form 23 May 2014Accepted 24 May 2014
Keyword:Bison bonasusCervus elaphusCapreolus capreolusAlces alcesDietFood niche overlap
With four species of ruminants, including red deer, roe deer, moose, and European bison, the Białowie _zaPrimeval Forest is unique on the European continent, where only one to three ungulate species are usu-ally found. The present study is the first effort to explore the dietary overlap of a European community ofsympatric ruminants using 3D dental microwear texture analysis. Results obtained for ungulates fromthe Białowie _za Forest were compared with those of four species with well known differences in diet(semi-wild Heck cattle, African buffaloes, giraffes, and yellow-backed duikers). These ruminants framethe spectrum of expected 3D dental microwear textures among ruminants: C3 and C4 grazers share highanisotropy and low to intermediate complexity while browsers display intermediate to high complexityassociated with low anisotropy. No significant differences between browsers (moose and roe deer) andmixed feeders (red deer) were detected by this analysis supporting a significant overlap in diet foundin Białowie _za. The results on the fourth ruminant are unequivocal: the European bison is not grazerbut instead is highly engaged in browsing. Furthermore, through 3D dental microwear texture analysis,the high plasticity in feeding behavior of the European bison can be tracked depending on the seasonsand on available access to feeding supplements during the winter.
� 2014 Published by Elsevier B.V.
1. Introduction
Evidence of overlap in resource use by sympatric species isessential for the understanding of interspecific competition(Mysterud, 2000). Often, resource use by one species reduces avail-ability of resources for another species or leads to species displace-ment (Putman, 1996; Ferretti et al., 2008). Competition forresource exploitation is one of the main mechanisms for interac-tion among large ungulates. Such interaction may occur whenungulates share similar habitats, an overlap in food consumedand when resources are limited (Gordon and Illius, 1989;Mysterud, 2000). Evidence of interactions among ungulates andother groups of species in natural ecosystems is limited (Putman,1996).
The Białowie _za Primeval Forest, located on the Polish-Belarus-sian border is one of the best preserved forests in Europe(Jedrzejewska and Jedrzejewski, 1998). With five species ofungulates (including red deer Cervus elaphus, roe deer Capreoluscapreolus, moose Alces alces, European bison Bison bonasus and wildboar Sus scrofa), it is unique on the European continent, largely
inhabited by only one to three ungulate species (Okarma, 1995).Ungulates coexist here with a high diversity of carnivores, includ-ing wolves and lynx. These predators hunt roe and red deer almostexclusively (Jedrzejewska et al., 1997; Jedrzejewska andJedrzejewski, 1998). Deciduous and mixed forests with rich under-growth and herb layers offer good conditions for ungulates andmay limit possible competition among them. In historical times,high densities of ungulates maintained for hunting purposes,together with cattle grazing in the forest (Samojlik and Kuijper,2013), had a strong impact on tree stands and resulted in speciescompetition. The two largest herbivores, the bison and the moose,were especially shaped by intra- and inter-specific competition forfood resources (Jedrzejewska et al., 1997; Jedrzejewska andJedrzejewski, 1998). Nowadays, populations of ungulates inBiałowie _za occur in moderate densities and their numbers areincreasing (Borowik et al., 2013; Kowalczyk et al., 2013).
Cervids and European bison – the only bovidae species in theBiałowie _za Forest – are characterized by different feeding habits(Hoffman, 1989). The moose (A. alces) is the largest cervid. Its dietis mainly based on browsing trees and shrubs (birches, willows,rowan etc.) and herbaceous dicots; grasses and sedges being veryrare in this diet (Morow, 1976; Franzmann, 1981). The red deeroccupies various habitats from forest to moorland. In Europe, its

G. Merceron et al. / Forest Ecology and Management 328 (2014) 262–269 263
diet includes different proportions of browse and grass dependingon the season, region and habitat (Gebert and Verheyden-Tixier,2001). Roe deer - the most widely distributed European ungulatespecies (Linnell et al., 1998) – is recognized as being a selectivefeeder (browser) occupying a large set of habitats from opencultivated areas to dense forest (Tixier and Duncan, 1996). TheEuropean bison, the largest terrestrial mammal in Europe, restoredfrom captive survivors following its extinction in the wild at thebeginning of the 20th century, is found mainly in forest habitats(Krasinska and Krasinski, 2007; Kerley et al., 2012). It is assumedto be a mixed feeder or grazer (Hoffman, 1989; Krasinska andKrasinski, 2007; Kowalczyk et al., 2011), however its diet differsstrongly from summer to winter due to the supplementary feedingwidely used for the management of species conservation (Kerleyet al., 2012). In the last two decades, however, agricultural landproviding crops of rapeseed and cereals or hay left by farmers inthe meadows has been more frequently used in winter and someseasonal migrations can therefore be observed (Kowalczyk et al.,2011; Hofman-Kaminska and Kowalczyk, 2012).
In the present study, we explore the dietary preferences of thefour sympatric ruminants from the Białowie _za Forest through 3Ddental microwear texture analysis (3D-DMTA). The 3D-DMTA isbased on the automated quantification of 3D surfaces by using ascale-sensitive fractal analysis. It has proved to be very efficientin discriminating dietary differences between species of primatesboth extant and extinct (Merceron et al., 2009; Ungar et al.,2010; Scott et al., 2012) as well as ungulates (Ungar et al., 2007;Scott, 2012) and carnivores (DeSantis et al., 2012; Stynder et al.,2012). Because it only provides information for a timescale rangingfrom a few days to a few weeks (Teaford and Oyen, 1989; Schulzet al., 2013), the dental microwear texture is an appropriate proxyto detect seasonal variations in diet and to pinpoint the exploita-tion of fallback foods (Merceron et al., 2010), i.e. food items thatare consumed when preferred food is not available anymore.Through this 3D dental microwear texture analysis, we will explorethe resource partitioning or overlapping among the four sympatricruminants.
We aimed to: (i) establish a first comparative dataset of wildEuropean ungulates for 3D-DMTA, (ii) explore the ecological nichepartitioning among a guild of wild and sympatric ungulates includ-ing bovids and cervids inhabiting the very same area, and (iii) andthen assess the foraging habits of European bison recognizedrecently as a refugee species in European forests (Kerley et al.,2012). More specifically, we aim to assess the portion of abrasivegrasses in the bison’s diet all year round, notably in winter whencertain bison herds have access to supplementary hay fodder. Thus,results of this study might have implications for future manage-ment strategies for European bison conservation.
2. Material and methods
2.1. Material
2.1.1. The Białowie_za forestThis study was conducted in the Polish part of the Białowie _za
Primeval Forest (52�350–52�550N, 23�300–24�000E; 625 km2), oneof the best preserved temperate lowland forests in Europe. Thedominating habitats found there are mixed coniferous and mixeddeciduous forest (pine Pinus silvestris, spruce Picea abies, oak Quer-cus robur, with admixtures of birch Betula spp., and aspen Populustremula), which covers 39.3% of the area and rich deciduous forest(oak Q. robur, hornbeam Carpinus betulus, lime Tilia cordata, andmaple Acer platanoides) covering 34.9%. Wet alder-ash forest (blackalder Alnus glutinosa, ash Fraxinus excelsior) covers 12.6% of thearea, coniferous forest (mainly pine P. silvestris, and spruce P. abies)
covers 6.2%, and open habitats within the forest cover 7%(Sokołowski, 2004). Most of habitats are covered with rich under-growth dominated by hornbeam (37%) and Norway spruce (22%)(Podgórski et al., 2008; Kowalczyk et al., 2011). The Forest issurrounded by agricultural areas with meadows, wastelands andarable land interspersed with small woodlands. The study areaincluded both Białowie _za National Park (17%) and the exploitedforests (83%, of which 34% is protected). The latter are exploitedby clear cutting small areas for natural regeneration or replanting.
The climate of Białowie _za Primeval Forest is transitional,between the Atlantic and continental types, with clearly markedcold and warm seasons. The average annual temperature is 7 �C.The coldest month is January (average daily temperature is�4.8 �C), and the warmest is July (18.4 �C). Snow cover persistsfor 60–96 days per year with a maximum recorded depth of95 cm (Jedrzejewska and Jedrzejewski, 1998).
The densities of the four species of ruminants are as follows:European bison �0.7 individual per km2, red deer (C. elaphus)�6.0 individual per km2, roe deer (C. capreolus) �2.0 individualper km2 and moose (A. alces) �0.08 individual per km2. As wellas this, the density of wild boar (S. scrofa) reaches 5.4 individualper km2 (Borowik et al., 2013).
2.1.2. The Białowie_za ruminants3D dental microwear texture analysis is applied on 86 speci-
mens of ruminants from the Białowie _za Forest, including 19 roedeer, 14 red deer, 8 moose, and 45 European bison. All specimenswere living free before being culled or found dead in theBiałowie _za Forest and their skulls are stored in the mammal collec-tion of the Mammal Research Institute, Białowie _za (Poland). Formost of them, date of death and gender are known. All mooseand roe deer were culled in winter. The red deer sample is moreheterogeneous including individuals shot in winter and spring,and 6 individuals over 14 red deer for which date of death isunknown. However, they were shot during the hunting period infall and winter (Appendix 1).
Among the 45 individuals of European bison, 39 can be clus-tered in four groups defined by period of death and access to sup-plementary hay fodder in winter (Table 1). The first group of bison(Bison #1; N = 18) is composed of individuals sampled from springto summer (from April to September). During that span of time, theunderstory vegetation in the primary forest of Białowie _za offersabundant resources for bison (Falinski, 1986; Jedrzejewska et al.,1997). The groups Bison #2 (N = 8), Bison #3 (N = 7), and Bison#4 (N = 6) consist of individuals sampled in late autumn and win-ter (from November to early March; Table 1). Bison #2 group iscomposed of individuals from populations intensively fed withhay (3–5 times a week); Bison #3 with individuals less intensivelyfed with hay (usually once a week). Non-fed individuals usingmainly agricultural areas in winter compose the fourth group(Bison #4; Table 1; see also Kowalczyk et al., 2011). The rest ofthe bison sample (6 individuals) were sampled in October. Wemade the choice to exclude them from the intra-population analy-sis in order to keep samples with sharp differences in vegetalresources access (see Appendix 1).
2.1.3. A comparative datasetOur analysis includes 58 individuals representing four species
of ruminants with clearly defined and well known differences indiet to serve as reference points and frame the spectrum ofexpected 3D dental microwear textures among ruminants(Table 1). The African buffaloes (Syncerus caffer; N = 8) from openlandscapes in Central and Eastern Africa and the semi-wild,non-fed Heck cattle (Bos taurus; N = 8) from Oostvaardersplassen(Netherlands) are grazers; the former mostly foraging on C4 andthe latter on C3 monocotyledonous (Sinclair, 1977; Estes, 1991;
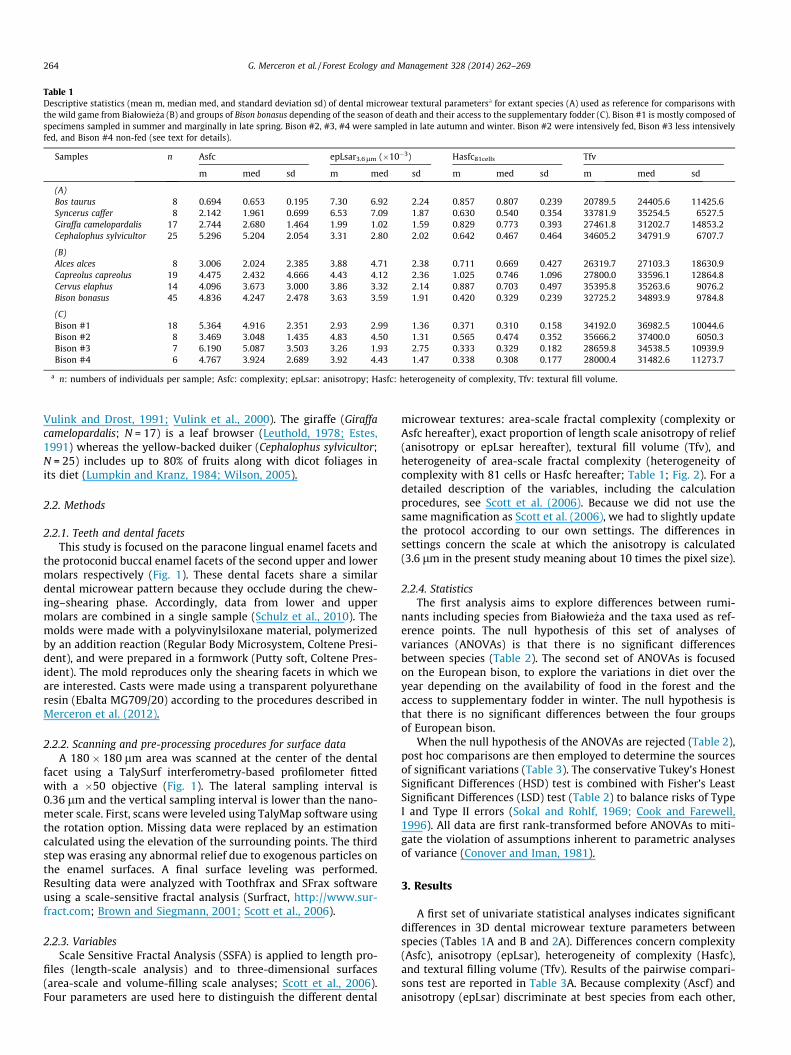
Table 1Descriptive statistics (mean m, median med, and standard deviation sd) of dental microwear textural parametersa for extant species (A) used as reference for comparisons withthe wild game from Białowie _za (B) and groups of Bison bonasus depending of the season of death and their access to the supplementary fodder (C). Bison #1 is mostly composed ofspecimens sampled in summer and marginally in late spring. Bison #2, #3, #4 were sampled in late autumn and winter. Bison #2 were intensively fed, Bison #3 less intensivelyfed, and Bison #4 non-fed (see text for details).
Samples n Asfc epLsar3.6lm (�10�3) Hasfc81cells Tfv
m med sd m med sd m med sd m med sd
(A)Bos taurus 8 0.694 0.653 0.195 7.30 6.92 2.24 0.857 0.807 0.239 20789.5 24405.6 11425.6Syncerus caffer 8 2.142 1.961 0.699 6.53 7.09 1.87 0.630 0.540 0.354 33781.9 35254.5 6527.5Giraffa camelopardalis 17 2.744 2.680 1.464 1.99 1.02 1.59 0.829 0.773 0.393 27461.8 31202.7 14853.2Cephalophus sylvicultor 25 5.296 5.204 2.054 3.31 2.80 2.02 0.642 0.467 0.464 34605.2 34791.9 6707.7
(B)Alces alces 8 3.006 2.024 2.385 3.88 4.71 2.38 0.711 0.669 0.427 26319.7 27103.3 18630.9Capreolus capreolus 19 4.475 2.432 4.666 4.43 4.12 2.36 1.025 0.746 1.096 27800.0 33596.1 12864.8Cervus elaphus 14 4.096 3.673 3.000 3.86 3.32 2.14 0.887 0.703 0.497 35395.8 35263.6 9076.2Bison bonasus 45 4.836 4.247 2.478 3.63 3.59 1.91 0.420 0.329 0.239 32725.2 34893.9 9784.8
(C)Bison #1 18 5.364 4.916 2.351 2.93 2.99 1.36 0.371 0.310 0.158 34192.0 36982.5 10044.6Bison #2 8 3.469 3.048 1.435 4.83 4.50 1.31 0.565 0.474 0.352 35666.2 37400.0 6050.3Bison #3 7 6.190 5.087 3.503 3.26 1.93 2.75 0.333 0.329 0.182 28659.8 34538.5 10939.9Bison #4 6 4.767 3.924 2.689 3.92 4.43 1.47 0.338 0.308 0.177 28000.4 31482.6 11273.7
a n: numbers of individuals per sample; Asfc: complexity; epLsar: anisotropy; Hasfc: heterogeneity of complexity, Tfv: textural fill volume.
264 G. Merceron et al. / Forest Ecology and Management 328 (2014) 262–269
Vulink and Drost, 1991; Vulink et al., 2000). The giraffe (Giraffacamelopardalis; N = 17) is a leaf browser (Leuthold, 1978; Estes,1991) whereas the yellow-backed duiker (Cephalophus sylvicultor;N = 25) includes up to 80% of fruits along with dicot foliages inits diet (Lumpkin and Kranz, 1984; Wilson, 2005).
2.2. Methods
2.2.1. Teeth and dental facetsThis study is focused on the paracone lingual enamel facets and
the protoconid buccal enamel facets of the second upper and lowermolars respectively (Fig. 1). These dental facets share a similardental microwear pattern because they occlude during the chew-ing–shearing phase. Accordingly, data from lower and uppermolars are combined in a single sample (Schulz et al., 2010). Themolds were made with a polyvinylsiloxane material, polymerizedby an addition reaction (Regular Body Microsystem, Coltene Presi-dent), and were prepared in a formwork (Putty soft, Coltene Pres-ident). The mold reproduces only the shearing facets in which weare interested. Casts were made using a transparent polyurethaneresin (Ebalta MG709/20) according to the procedures described inMerceron et al. (2012).
2.2.2. Scanning and pre-processing procedures for surface dataA 180 � 180 lm area was scanned at the center of the dental
facet using a TalySurf interferometry-based profilometer fittedwith a �50 objective (Fig. 1). The lateral sampling interval is0.36 lm and the vertical sampling interval is lower than the nano-meter scale. First, scans were leveled using TalyMap software usingthe rotation option. Missing data were replaced by an estimationcalculated using the elevation of the surrounding points. The thirdstep was erasing any abnormal relief due to exogenous particles onthe enamel surfaces. A final surface leveling was performed.Resulting data were analyzed with Toothfrax and SFrax softwareusing a scale-sensitive fractal analysis (Surfract, http://www.sur-fract.com; Brown and Siegmann, 2001; Scott et al., 2006).
2.2.3. VariablesScale Sensitive Fractal Analysis (SSFA) is applied to length pro-
files (length-scale analysis) and to three-dimensional surfaces(area-scale and volume-filling scale analyses; Scott et al., 2006).Four parameters are used here to distinguish the different dental
microwear textures: area-scale fractal complexity (complexity orAsfc hereafter), exact proportion of length scale anisotropy of relief(anisotropy or epLsar hereafter), textural fill volume (Tfv), andheterogeneity of area-scale fractal complexity (heterogeneity ofcomplexity with 81 cells or Hasfc hereafter; Table 1; Fig. 2). For adetailed description of the variables, including the calculationprocedures, see Scott et al. (2006). Because we did not use thesame magnification as Scott et al. (2006), we had to slightly updatethe protocol according to our own settings. The differences insettings concern the scale at which the anisotropy is calculated(3.6 lm in the present study meaning about 10 times the pixel size).
2.2.4. StatisticsThe first analysis aims to explore differences between rumi-
nants including species from Białowie _za and the taxa used as ref-erence points. The null hypothesis of this set of analyses ofvariances (ANOVAs) is that there is no significant differencesbetween species (Table 2). The second set of ANOVAs is focusedon the European bison, to explore the variations in diet over theyear depending on the availability of food in the forest and theaccess to supplementary fodder in winter. The null hypothesis isthat there is no significant differences between the four groupsof European bison.
When the null hypothesis of the ANOVAs are rejected (Table 2),post hoc comparisons are then employed to determine the sourcesof significant variations (Table 3). The conservative Tukey’s HonestSignificant Differences (HSD) test is combined with Fisher’s LeastSignificant Differences (LSD) test (Table 2) to balance risks of TypeI and Type II errors (Sokal and Rohlf, 1969; Cook and Farewell,1996). All data are first rank-transformed before ANOVAs to miti-gate the violation of assumptions inherent to parametric analysesof variance (Conover and Iman, 1981).
3. Results
A first set of univariate statistical analyses indicates significantdifferences in 3D dental microwear texture parameters betweenspecies (Tables 1A and B and 2A). Differences concern complexity(Asfc), anisotropy (epLsar), heterogeneity of complexity (Hasfc),and textural filling volume (Tfv). Results of the pairwise compari-sons test are reported in Table 3A. Because complexity (Ascf) andanisotropy (epLsar) discriminate at best species from each other,
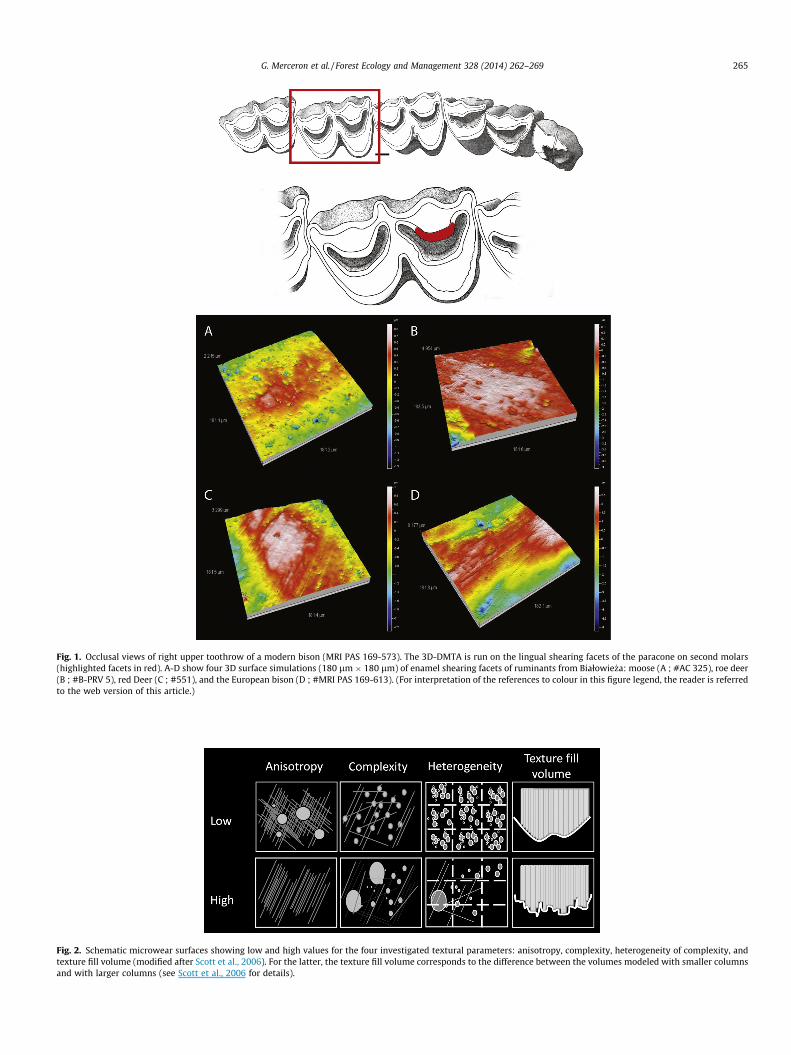
Fig. 1. Occlusal views of right upper toothrow of a modern bison (MRI PAS 169-573). The 3D-DMTA is run on the lingual shearing facets of the paracone on second molars(highlighted facets in red). A-D show four 3D surface simulations (180 lm � 180 lm) of enamel shearing facets of ruminants from Białowie _za: moose (A ; #AC 325), roe deer(B ; #B-PRV 5), red Deer (C ; #551), and the European bison (D ; #MRI PAS 169-613). (For interpretation of the references to colour in this figure legend, the reader is referredto the web version of this article.)
Fig. 2. Schematic microwear surfaces showing low and high values for the four investigated textural parameters: anisotropy, complexity, heterogeneity of complexity, andtexture fill volume (modified after Scott et al., 2006). For the latter, the texture fill volume corresponds to the difference between the volumes modeled with smaller columnsand with larger columns (see Scott et al., 2006 for details).
G. Merceron et al. / Forest Ecology and Management 328 (2014) 262–269 265
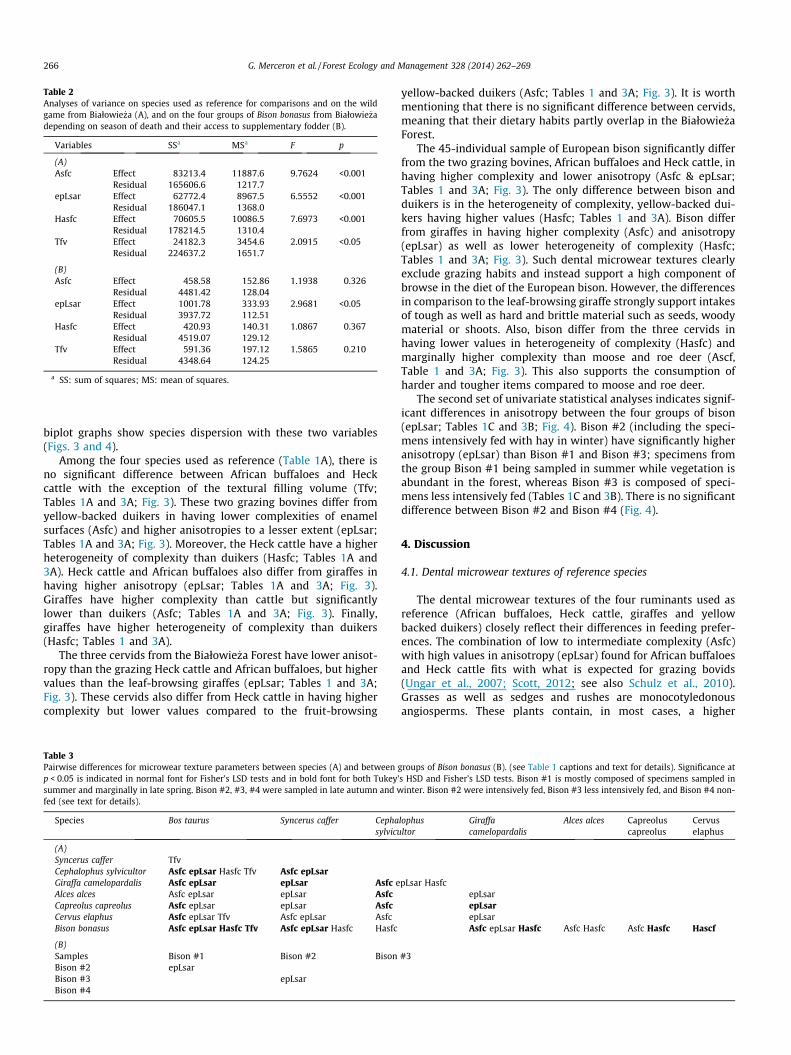
Table 2Analyses of variance on species used as reference for comparisons and on the wildgame from Białowie _za (A), and on the four groups of Bison bonasus from Białowie _zadepending on season of death and their access to supplementary fodder (B).
Variables SSa MSa F p
(A)Asfc Effect 83213.4 11887.6 9.7624 <0.001
Residual 165606.6 1217.7epLsar Effect 62772.4 8967.5 6.5552 <0.001
Residual 186047.1 1368.0Hasfc Effect 70605.5 10086.5 7.6973 <0.001
Residual 178214.5 1310.4Tfv Effect 24182.3 3454.6 2.0915 <0.05
Residual 224637.2 1651.7
(B)Asfc Effect 458.58 152.86 1.1938 0.326
Residual 4481.42 128.04epLsar Effect 1001.78 333.93 2.9681 <0.05
Residual 3937.72 112.51Hasfc Effect 420.93 140.31 1.0867 0.367
Residual 4519.07 129.12Tfv Effect 591.36 197.12 1.5865 0.210
Residual 4348.64 124.25
a SS: sum of squares; MS: mean of squares.
266 G. Merceron et al. / Forest Ecology and Management 328 (2014) 262–269
biplot graphs show species dispersion with these two variables(Figs. 3 and 4).
Among the four species used as reference (Table 1A), there isno significant difference between African buffaloes and Heckcattle with the exception of the textural filling volume (Tfv;Tables 1A and 3A; Fig. 3). These two grazing bovines differ fromyellow-backed duikers in having lower complexities of enamelsurfaces (Asfc) and higher anisotropies to a lesser extent (epLsar;Tables 1A and 3A; Fig. 3). Moreover, the Heck cattle have a higherheterogeneity of complexity than duikers (Hasfc; Tables 1A and3A). Heck cattle and African buffaloes also differ from giraffes inhaving higher anisotropy (epLsar; Tables 1A and 3A; Fig. 3).Giraffes have higher complexity than cattle but significantlylower than duikers (Asfc; Tables 1A and 3A; Fig. 3). Finally,giraffes have higher heterogeneity of complexity than duikers(Hasfc; Tables 1 and 3A).
The three cervids from the Białowie _za Forest have lower anisot-ropy than the grazing Heck cattle and African buffaloes, but highervalues than the leaf-browsing giraffes (epLsar; Tables 1 and 3A;Fig. 3). These cervids also differ from Heck cattle in having highercomplexity but lower values compared to the fruit-browsing
Table 3Pairwise differences for microwear texture parameters between species (A) and betweenp < 0.05 is indicated in normal font for Fisher’s LSD tests and in bold font for both Tukey’summer and marginally in late spring. Bison #2, #3, #4 were sampled in late autumn and wfed (see text for details).
Species Bos taurus Syncerus caffer Cephasylvicu
(A)Syncerus caffer TfvCephalophus sylvicultor Asfc epLsar Hasfc Tfv Asfc epLsarGiraffa camelopardalis Asfc epLsar epLsar Asfc eAlces alces Asfc epLsar epLsar AsfcCapreolus capreolus Asfc epLsar epLsar AsfcCervus elaphus Asfc epLsar Tfv Asfc epLsar AsfcBison bonasus Asfc epLsar Hasfc Tfv Asfc epLsar Hasfc Hasfc
(B)Samples Bison #1 Bison #2 BisonBison #2 epLsarBison #3 epLsarBison #4
yellow-backed duikers (Asfc; Tables 1 and 3A; Fig. 3). It is worthmentioning that there is no significant difference between cervids,meaning that their dietary habits partly overlap in the Białowie _zaForest.
The 45-individual sample of European bison significantly differfrom the two grazing bovines, African buffaloes and Heck cattle, inhaving higher complexity and lower anisotropy (Asfc & epLsar;Tables 1 and 3A; Fig. 3). The only difference between bison andduikers is in the heterogeneity of complexity, yellow-backed dui-kers having higher values (Hasfc; Tables 1 and 3A). Bison differfrom giraffes in having higher complexity (Asfc) and anisotropy(epLsar) as well as lower heterogeneity of complexity (Hasfc;Tables 1 and 3A; Fig. 3). Such dental microwear textures clearlyexclude grazing habits and instead support a high component ofbrowse in the diet of the European bison. However, the differencesin comparison to the leaf-browsing giraffe strongly support intakesof tough as well as hard and brittle material such as seeds, woodymaterial or shoots. Also, bison differ from the three cervids inhaving lower values in heterogeneity of complexity (Hasfc) andmarginally higher complexity than moose and roe deer (Ascf,Table 1 and 3A; Fig. 3). This also supports the consumption ofharder and tougher items compared to moose and roe deer.
The second set of univariate statistical analyses indicates signif-icant differences in anisotropy between the four groups of bison(epLsar; Tables 1C and 3B; Fig. 4). Bison #2 (including the speci-mens intensively fed with hay in winter) have significantly higheranisotropy (epLsar) than Bison #1 and Bison #3; specimens fromthe group Bison #1 being sampled in summer while vegetation isabundant in the forest, whereas Bison #3 is composed of speci-mens less intensively fed (Tables 1C and 3B). There is no significantdifference between Bison #2 and Bison #4 (Fig. 4).
4. Discussion
4.1. Dental microwear textures of reference species
The dental microwear textures of the four ruminants used asreference (African buffaloes, Heck cattle, giraffes and yellowbacked duikers) closely reflect their differences in feeding prefer-ences. The combination of low to intermediate complexity (Asfc)with high values in anisotropy (epLsar) found for African buffaloesand Heck cattle fits with what is expected for grazing bovids(Ungar et al., 2007; Scott, 2012; see also Schulz et al., 2010).Grasses as well as sedges and rushes are monocotyledonousangiosperms. These plants contain, in most cases, a higher
groups of Bison bonasus (B). (see Table 1 captions and text for details). Significance ats HSD and Fisher’s LSD tests. Bison #1 is mostly composed of specimens sampled in
inter. Bison #2 were intensively fed, Bison #3 less intensively fed, and Bison #4 non-
lophusltor
Giraffacamelopardalis
Alces alces Capreoluscapreolus
Cervuselaphus
pLsar HasfcepLsarepLsarepLsarAsfc epLsar Hasfc Asfc Hasfc Asfc Hasfc Hascf
#3
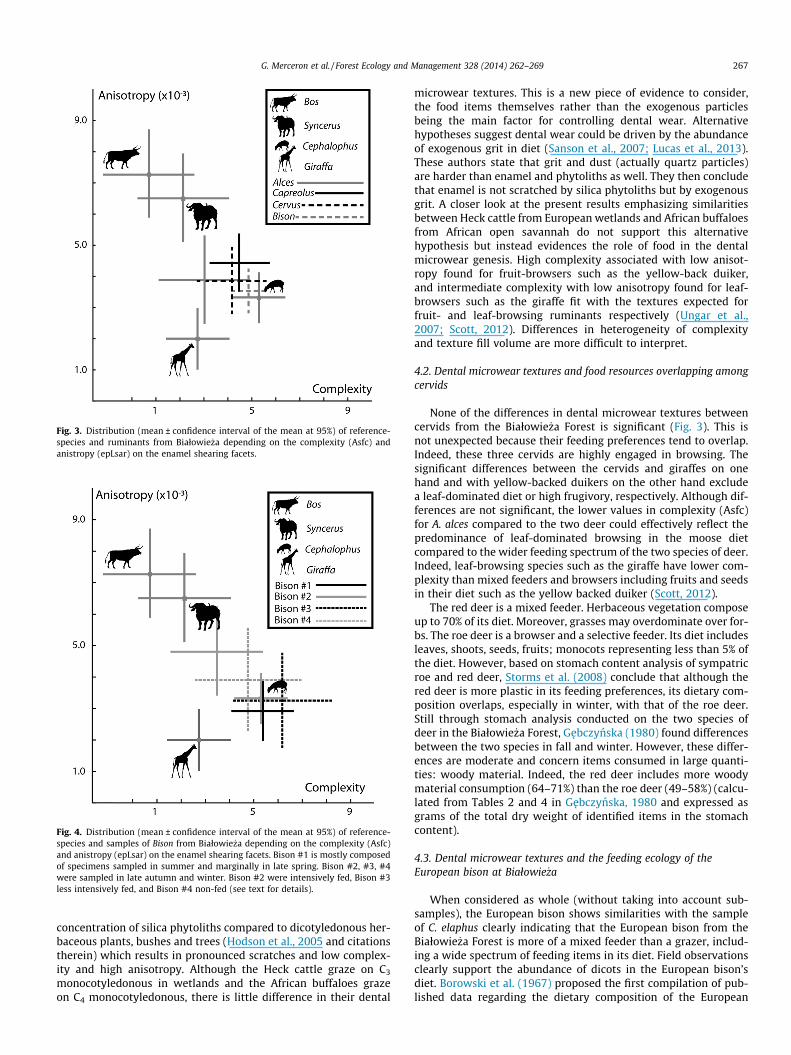
Fig. 3. Distribution (mean ± confidence interval of the mean at 95%) of reference-species and ruminants from Białowie _za depending on the complexity (Asfc) andanistropy (epLsar) on the enamel shearing facets.
Fig. 4. Distribution (mean ± confidence interval of the mean at 95%) of reference-species and samples of Bison from Białowie _za depending on the complexity (Asfc)and anistropy (epLsar) on the enamel shearing facets. Bison #1 is mostly composedof specimens sampled in summer and marginally in late spring. Bison #2, #3, #4were sampled in late autumn and winter. Bison #2 were intensively fed, Bison #3less intensively fed, and Bison #4 non-fed (see text for details).
G. Merceron et al. / Forest Ecology and Management 328 (2014) 262–269 267
concentration of silica phytoliths compared to dicotyledonous her-baceous plants, bushes and trees (Hodson et al., 2005 and citationstherein) which results in pronounced scratches and low complex-ity and high anisotropy. Although the Heck cattle graze on C3
monocotyledonous in wetlands and the African buffaloes grazeon C4 monocotyledonous, there is little difference in their dental
microwear textures. This is a new piece of evidence to consider,the food items themselves rather than the exogenous particlesbeing the main factor for controlling dental wear. Alternativehypotheses suggest dental wear could be driven by the abundanceof exogenous grit in diet (Sanson et al., 2007; Lucas et al., 2013).These authors state that grit and dust (actually quartz particles)are harder than enamel and phytoliths as well. They then concludethat enamel is not scratched by silica phytoliths but by exogenousgrit. A closer look at the present results emphasizing similaritiesbetween Heck cattle from European wetlands and African buffaloesfrom African open savannah do not support this alternativehypothesis but instead evidences the role of food in the dentalmicrowear genesis. High complexity associated with low anisot-ropy found for fruit-browsers such as the yellow-back duiker,and intermediate complexity with low anisotropy found for leaf-browsers such as the giraffe fit with the textures expected forfruit- and leaf-browsing ruminants respectively (Ungar et al.,2007; Scott, 2012). Differences in heterogeneity of complexityand texture fill volume are more difficult to interpret.
4.2. Dental microwear textures and food resources overlapping amongcervids
None of the differences in dental microwear textures betweencervids from the Białowie _za Forest is significant (Fig. 3). This isnot unexpected because their feeding preferences tend to overlap.Indeed, these three cervids are highly engaged in browsing. Thesignificant differences between the cervids and giraffes on onehand and with yellow-backed duikers on the other hand excludea leaf-dominated diet or high frugivory, respectively. Although dif-ferences are not significant, the lower values in complexity (Asfc)for A. alces compared to the two deer could effectively reflect thepredominance of leaf-dominated browsing in the moose dietcompared to the wider feeding spectrum of the two species of deer.Indeed, leaf-browsing species such as the giraffe have lower com-plexity than mixed feeders and browsers including fruits and seedsin their diet such as the yellow backed duiker (Scott, 2012).
The red deer is a mixed feeder. Herbaceous vegetation composeup to 70% of its diet. Moreover, grasses may overdominate over for-bs. The roe deer is a browser and a selective feeder. Its diet includesleaves, shoots, seeds, fruits; monocots representing less than 5% ofthe diet. However, based on stomach content analysis of sympatricroe and red deer, Storms et al. (2008) conclude that although thered deer is more plastic in its feeding preferences, its dietary com-position overlaps, especially in winter, with that of the roe deer.Still through stomach analysis conducted on the two species ofdeer in the Białowie _za Forest, Gebczynska (1980) found differencesbetween the two species in fall and winter. However, these differ-ences are moderate and concern items consumed in large quanti-ties: woody material. Indeed, the red deer includes more woodymaterial consumption (64–71%) than the roe deer (49–58%) (calcu-lated from Tables 2 and 4 in Gebczynska, 1980 and expressed asgrams of the total dry weight of identified items in the stomachcontent).
4.3. Dental microwear textures and the feeding ecology of theEuropean bison at Białowie_za
When considered as whole (without taking into account sub-samples), the European bison shows similarities with the sampleof C. elaphus clearly indicating that the European bison from theBiałowie _za Forest is more of a mixed feeder than a grazer, includ-ing a wide spectrum of feeding items in its diet. Field observationsclearly support the abundance of dicots in the European bison’sdiet. Borowski et al. (1967) proposed the first compilation of pub-lished data regarding the dietary composition of the European

268 G. Merceron et al. / Forest Ecology and Management 328 (2014) 262–269
bison in the wild. More than 200 species are eaten and amongthose 5–25 plants are preferred. Almost 29% of the plants eatenare monocotyledonous (grasses, sedges). Forbs (herbaceous dicots)and trees/bushes represent 31% and 39% of the eaten species,respectively. In terms of dry matter weight in stomach contents,forbs together with grasses represent 50% of the food bolus, therest being composed of tree and bush foliages, shoots and fine bark(Borowski et al., 1967 and citations therein). Later, Borowski andKossak (1972) concluded that monocotyledonous and dicotyledon-ous herbs (forbs) collectively represent 67% of the dietary bolus. Allthese ecological data are in keeping with what the 3D-DMTAextracts from the enamel surface analysis. Grasses and grass-likeplants such as rushes and sedges do not constitute a major elementbut a secondary food resource. The European bison in Białowie _za isnot a grazer (not even a variable grazer sensu Gagnon and Chew,2000) but a mixed feeder.
This compilation does not take into account the seasonalchanges in diet. A closer look at the variations among sub-sampleswill help to draw a more detailed picture. The only significant dif-ference between the four groups of the European bison concernsanisotropy (EpLsar), a variable positively correlated to the amountof abrasive monocotyledonous (Ungar et al., 2007; Schulz et al.,2010; Scott, 2012). Bison #1 composed of specimens which diedin the summer differs from the intensively-fed bison (Bison #2).The latter tend more to be grazers due to their access to intensivesupplementary feed and hay. Due to the higher amount of silica ingrasses, hay is more abrasive than browse and is responsible of thehigher anisotropy. Bison #1 group has access to huge amounts ofvegetation in the forest during the summer: fresh grasses but alsofoliage, twigs, shoots, buds and fruits. The less intensively fedgroup, Bison #3 does not differ from Bison #1 but does differ fromBison #2 suggesting a higher amount of browse compared to Bison#2. This result was expected, the assumption being that restrictedaccess to supplementary fodder encourages bison to forage forother foods available in the forest during winter: tree shoots, barkand dry vegetation available in more productive habitats and forestgaps. These elements are not as abrasive as monocotyledonous butsome of them like bark are tougher than the softer foliage availablein summer and spring and thus require more processes to be bro-ken down before ingestion. Bison #4 is composed of specimens for-aging mostly outside the forest during the winter. Theseindividuals forage mainly on hay stored by farmers and meadowsmaintained by EU subsidies (Hofman-Kaminska and Kowalczyk,2012). Occasionally they also forage on winter crops of rapeseedand cereals accessible under the snow cover and shrubby vegeta-tion available on forest edges (Kowalczyk et al., 2011). Such a diet,mixing monocotyledonous, herbaceous dicots and woody vegeta-tion, explains why this group has an intermediate value in anisot-ropy as well as in complexity between Bison #1 and Bison #2.
5. Conclusion
This study is the first attempt to explore the ecological nichepartitioning between member of a unique guild of sympatric rumi-nants in Europe through 3D dental microwear textural analysis.Being an observer-free approach, the 3D-DMTA allows the objec-tive exploration of differences in the feeding habits of a given spe-cies or population.
The present 3D-DMTA supports previous pioneering studies onungulates (Ungar et al., 2007; Schulz et al., 2010; Scott, 2012).Indeed, the results on the four species used as reference points toframe the model are clearly defined. Grazers significantly differfrom browsers mostly in having higher anisotropy and lower com-plexity. Among browsers, fruit browsers have higher complexitythan leaf browsers. The 3D-DMTA on ruminants from Białowie _za
clearly supports the hypothesis that cervids overlap in their feed-ing preferences, which is not unexpected for browsers (roe deerand moose) and mixed feeders (red deer) exploiting the same hab-itat. The high rate of overlap in food habits may indicate anabsence of competition and sharing of common resources(Gordon and Illius, 1989; Putman, 1996; Mysterud, 2000). Long-term data analysis by Jedrzejewska et al. (1997) showed that thefeeding habits of ungulates in the Białowie _za Forest were shapedby intra- and interspecific competition for food at high densities,especially at the beginning of 20th century, which was an effectof extensive supplementary feeding and the extermination of largepredators. Much lower densities in latter decades allowed theshared use of the rich food resources of the Białowie _za Forest with-out competitive interactions. This probably resulted in a higheroverlap of the diet and microwear patterns.
The results of the 3D-DMTA on the Białowie _za bovid areunequivocal: the European bison is highly involved in browsing.Tree leaves and shoots during spring and summer are more easilydigestible than grass, which may explain mixed feeding foraging bybison. The 3D-DMTA on the different sub-samples also track thehigh plasticity in feeding behavior for the European bison. Indeed,specimens sampled during periods where dense understory vege-tation was available within the forest differ in dental microweartexture from those sampled in winter at the close vicinity of thesupplementary hay fodders. When the access to supplementaryhay fodder is reduced or even removed, the European bison eithercompletes its diet with browse in the forest or broadens its homerange out of the forest to exploit the hay left by farmers and otherfarm crops (Hofman-Kaminska and Kowalczyk, 2012).
Acknowledgments
This study was financed by the Polish National Science Centre(Grant No N N304 301940) and the Project ANR TRIDENT (ANR-13-JSV7-0008-01, PI: G. Merceron). We are also grateful to the sup-port of the Mammal Research Institute PAS in Białowie _za andiPHEP (UMR 7262, CNRS & University of Poitiers). We are gratefulto the former director of the Mammal Research Institute for theaccess to mammalian collection and to N. Brunetière from PprimeInstitut at the University of Poitiers for the access to the TaylorHobson 3D non contact surface Profilometer. We also would liketo thank Dr. L. Sönnichsen and T. Borowik for the access to addi-tional mandibles of red deer and roe deer from Białowie _za region.Finally, we thank Seth Ramdarshan (http://srtranslations.word-press.com/) for improving the English.
Appendix A. Supplementary material
Supplementary data associated with this article can be found,in the online version, at http://dx.doi.org/10.1016/j.foreco.2014.05.041.
References
Borowik, T., Cornulier, T., Jedrzejewska, B., 2013. Environmental factors shapingungulate abundances in Poland. Acta Theriol. 58, 403–413.
Borowski, S., Kossak, S., 1972. The natural food preferences of the European bison inseasons free of snow cover. Acta Theriol. 17, 151–169.
Borowski, S., Krasinski, Z.A., Miłkowski, L., 1967. Food and role of the Europeanbison in forest ecosystems. Acta Theriol. 12, 367–376.
Brown, C.A., Siegmann, S., 2001. Fundamental scales of adhesion and area scalefractal analysis. Int. J. Mach. Tools Manu. 41, 1927–1933.
Conover, W.J., Iman, R.L., 1981. Rank transformations as a bridge betweenparametric and nonparametric statistics. Am. Stat. 35, 124–129.
Cook, R.J., Farewell, V.T., 1996. Multiplicity considerations in the design and analysisof clinical trials. J. Roy. Stat. Soc. 159, 93–110.
DeSantis, L.R.G., Schubert, B.W., Scott, J.R., Ungar, P.S., 2012. Implications of diet forthe extinction of Saber-toothed cats and American lions. PLoS ONE 7, e52453.

G. Merceron et al. / Forest Ecology and Management 328 (2014) 262–269 269
Estes, R.D., 1991. Behavior Guide to African Mammals. The University of CaliforniaPress, Los Angeles.
Falinski, J.B., 1986. Vegetation Dynamics in Temporate Lowland Prmeval Forests.Ecological Studies in Białowie _za Forest. Dr W. Junk Publishers, Dordrecht.
Ferretti, F., Myers, R.A., Serena, F., Lotze, H.K., 2008. Loss of large predatory sharksfrom the Mediterranean Sea. Conserv. Biol. 22, 952–964.
Franzmann, A.W., 1981. Alces alces. Mamm. Species 154, 1–7.Gagnon, M., Chew, A.E., 2000. Dietary preferences in extant African Bovidae. J.
Mammal. 8, 490–511.Gebczynska, Z., 1980. Food of the roe deer and red deer in the Białowie _za Primeval
Forest. Acta Theriol. 25, 487–500.Gebert, C., Verheyden-Tixier, H., 2001. Variations of diet composition of red deer
(Cervus elaphus L.) in Europe. Mammal Rev. 31, 189–201.Gordon, U., Illius, A.W., 1989. Resource partitioning by ungulates on the Isle of
Rhum. Oecologia 79, 383–389.Hodson, M.J., White, P.J., Mead, A., Broadley, M.R., 2005. Phylogenetic variation in
the silicon composition of plants. Ann. Bot. 96, 1027–1046.Hoffman, R.R., 1989. Evolutionary steps of ecophysiological adaptation and
diversification view of their digestive system. Oecologia 78, 443–457.Hofman-Kaminska, E., Kowalczyk, R., 2012. Farm crops depredation by European
bison (/Bison bonasus/) in the vicinity of forest habitats in northeastern Poland.Environ. Manage. 50, 530–541.
Jedrzejewska, B., Jedrzejewski, W., 1998. Predation in Vertebrate Communities. TheBiałowie _za Primeval Forest as a Case Study. Springer-Verlag, Berlin.
Jedrzejewska, B., Jedrzejewski, W., Bunevich, A.N., Miłkowski, L., Krasinski, Z.A.,1997. Factors shaping population densities and increase rates of ungulates inBiałowie _za Primeval Forest (Poland and Belarus) in the 19th and 20th centuries.Acta Theriol. 42, 399–451.
Kerley, G.I.H., Kowalczyk, R., Cromsigt, J.P.G.M., 2012. Conservation implications ofthe refugee species concept and the European bison: king of the forest orrefugee in a marginal habitat? Ecography 35, 519–529.
Kowalczyk, R., Taberlet, P., Coissac, E., Valentini, A., Miquel, C., Kaminski, T., Wójcik,J.M., 2011. Influence of management practices on large herbivore diet – case ofEuropean bison in Białowie _za Primeval Forest (Poland). For. Ecol. Manage. 261,821–828.
Kowalczyk, R., Krasinska, M., Kaminski, T., Górny, M., Strus, P., Hofman-Kaminska,E., Krasinski, Z.A., 2013. Movements of European bison (Bison bonasus) beyondthe Białowie _za Forest (NE Poland): range expansion or partial migrations? ActaTheriol. 58, 391–401.
Krasinska, M., Krasinski, Z.A., 2007. European Bison: A Nature Monograph. MammalResarch Institute PAS, Białowie _za.
Leuthold, B.M., 1978. Ecology of the giraffe in Tsavo East National Park, Kenya. EastAfr. Wildlife J. 16, 1–20.
Linnell, J.D.C., Duncan, P., Andersen, R., 1998. The European roe deer: a portrait of asuccessful species. In: Andersen, R., Duncan, P., Linnell, J.D.C. (Eds.), TheEuropean Roe Deer: The Biology of Success. Scandinavian University Press, Oslo,pp. 11–22.
Lucas, P.W., Omar R., Al-Fadhalah, K., Almusallam, A.S., Henry, A.G., Michael, S., Thai,L.A., Watzke, J., Strait, D.S., Atkins, A.G., 2013. Mechanisms and causes of wear intooth enamel: implications for hominin diets. J. Roy. Soc. Interface http://dx.doi.org/10:20120923.
Lumpkin, S., Kranz, K.R., 1984. Cephalophus sylvicultor. Mamm. Species 225, 1–7.Merceron, G., Scott, J., Scott, R.S., Geraads, D., Spassov, N., Ungar, P.S., 2009. Folivory
or fruit/seed predation for Mesopithecus, an earliest colobine from the LateMiocene of Eurasia? J. Hum. Evol. 57, 732–738.
Merceron, G., Escarguel, G., Angibault, J.-M., Verheyden-Tixier, H., 2010. Can dentalmicrowear textures record dietary inter-individual dietary variations? PLoSONE 5 (3), e9542.
Merceron, G., Costeur, L., Maridet, O., Ramdarshan, A., Göhlich, U.B., 2012. Multi-proxy approach detects heterogeneous habitats for primates during theMiocene climatic optimum in Central Europe. J. Hum. Evol. 63, 150–161.
Morow, K., 1976. Food habits of moose from Augustów Forest. Acta Theriol. 21,101–116.
Mysterud, A., 2000. Diet overlap among ruminants in Fennoscandia. Oecologia 124,130–137.
Okarma, H., 1995. The trophic ecology of wolves and their predatory role inungulate communities of forest ecosystems in Europe. Acta Theriol. 40, 335–386.
Podgórski, T., Schmidt, K., Kowalczyk, R., Gulczynska, A., 2008. Microhabitatselection by Eurasian lynx and its implications for species conservation. ActaTheriol. 53, 97–110.
Putman, R.J., 1996. Competition and Resource Partitioning in Temperate UngulateAssemblies. Chapman & Hall, London.
Samojlik, T., Kuijper, D., 2013. Grazed wood pasture versus browsed high forests –impact of ungulates on forest landscapes from the perspective of the Białowie _zaPrimeval Forest. In: Rotherham, I.D. (Ed.), Trees, Forested Landscapes andGrazing Animals – A European Perspective on Woodlands and GrazedTreescapes. Routledge, New York, pp. 143–162.
Sanson, G.D., Kerr, S.A., Gross, K.A., 2007. Do silica phytoliths really mammalianteeth? J. Archaeol. Sci. 34, 526–531.
Schulz, E., Calandra, I., Kaiser, T.M., 2010. Applying tribology to teeth of hoofedmammals. Scanning 32, 162–182.
Schulz, E., Piotrowski, V., Clauss, M., Mau, M., Merceron, G., Kaiser, T.M., 2013.Dietary abrasiveness determines variability in microwear and dental surfacetexture in rabbits. PLoS ONE 8, e56167.
Scott, J.R., 2012. Dental microwear texture analysis of extant African Bovidae.Mammalia 76, 157–174.
Scott, R.S., Ungar, P., Bergstrom, T.S., Brown, C.A., Childs, B.E., Teaford, M.F., Walker,A., 2006. Dental microwear texture analysis: technical considerations. J. Hum.Evol. 51, 339–349.
Scott, R.S., Teaford, M.F., Ungar, P.S., 2012. Dental microwear texture andanthropoid diets. Am. J. Phys. Anthropol. 147, 551–579.
Sinclair, A.R.E., 1977. The African Buffalo: A Study Of Resource Limitations OfPopulations. The University of Chicago Press, Chicago.
Sokal, S.R., Rohlf, F.J., 1969. Biometry. W.E. Freeman and Company, New York.Sokołowski, A.W., 2004. Lasy Puszczy Białowieskiej [Woodlands of Białowie _za
Primeval Forest]. State Forests Information Centre, Warsaw, pp. 364.Storms, D., Aubry, P., Hamann, J.-L., Saïd, S., Fritz, H., Saint-Andrieux, C., Klein, F.,
2008. Seasonal variation in diet composition and similarity of sympatric reddeer Cervus elaphus and roe deer Capreolus capreolus. Wildlife Biol. 14, 237–250.
Stynder, D.D., Ungar, P.S., Scott, J.R., Schubert, B.W., 2012. A dental microweartexture analysis of the Mio-Pliocene hyaenids from Langebaanweg, SouthAfrica. Acta Paleontol. Pol. 57, 485–496.
Teaford, M.F., Oyen, O.J., 1989. In vivo and in vitro turnover in dental microwear. Am.J. Phys. Anthropol. 80, 447–460.
Tixier, H., Duncan, P., 1996. Are European roe deer browsers? A review of variationsin the composition of their diets. Revue d’Ecol. (Terre Vie) 51, 3–17.
Ungar, P.S., Merceron, G., Scott, R.S., 2007. Dental microwear texture analysis ofVarswater bovids and Early Pliocene paleoenvironments of Langebaanweg,Western Cape Province, South Africa. J. Mammal. Evol. 14, 163–181.
Ungar, P.S., Scott, R.S., Grine, F.E., Teaford, M.F., 2010. Molar microwear textures andthe diets of Australopithecus anamensis and Au. afarensis. Philos. Trans. Roy. Soc.Lond. B: Biol. Sci. 365, 3345–3354.
Vulink, T.J., Drost, J.T., 1991. Nutritional characteristics of cattle forage plants in theeutrophic nature-reserve Oostvaardersplassen, Netherlands. Neth. J. Agric. Sci.39, 263–272.
Vulink, T.J., Drost, J.T., Jans, L., 2000. The influence of different grazing regimes onPhragmites- and shrub vegetation in the well-drained zone of a eutrophicwetland. Appl. Veg. Sci. 3, 73–80.
Wilson, V.J., 2005. Duikers of Africa (Masters of the African Forest Floor). ZimbiBooks, Pretoria.
Related Documents











