Journal of Cerebral Blood Flow and Metabolism 14:1021029 © 1994 The International Society of Cerebral Blood Flow and Metabolism Published by Raven Press, Ltd., New York 3-Acetylpyridine Produces Age-Dependent Excitotoxic Lesions in Rat Striatum Jorg B. Schulz, D. Ross Henshaw, *Bruce G. Jenkins, tRobert J. Ferrante, tNeil W. Kowall, *Bruce R. Rosen, and M. Flint Beal Neurochemist Laborato, Neurology Service, and *MGH-NMR Center, Department of Radiology, Massachusetts General Hospital and Harvard Medical School, and tGeriatric Research Education and Clinical Center, Bedford VA Medical Center, and Departments of Neurology and Pathology, Boston University School of Medicine, Boston, Massachusetts, U.S.A. Summary: The effects of 3-acetylpyridine (3-AP) were studied in rat striatum. Striatal injections of 3-AP pro- duced dose-dependent lesions. The lesion size was signif- icantly increased in 4- and 12-month-old rats compared to l-month-old rats. Coinjection of the competitive N-meth- yl-D-aspartate (NMDA) antagonist 2-amino-5- phosphonovaleric acid (APV) or systemic administration of the noncompetitive NMDA antagonist MK-801, the competitive NMDA antagonist L Y274614, or the gluta- mate release inhibitor lamotrigine partially but signifi- cantly attenuated striatal lesion volume. Consistent with an NMDA receptor-mediated excitotoxic effect, histo- logic studies showed that 3-AP lesions result in relative sparing of NADPH-diaphorase neurons. Using freeze clamp, 3-AP resulted in a marked depletion of ATP. Two- 3-Acetylpyridine (3-AP), a niacinamide antago- nist, is a potent neurotoxin when administered to laboratory animals by single intraperitoneal injec- tions. It induces chronic complex motor and behav- ioral abnormalities, which may be seen as hyperk- inesias, ataxia, tremor of the front legs, tonic cramps, and spontaneous convulsions (Herken, 1968). 3-AP has been reported to induce degenera- tion of the inferior olivary neurons, which give rise Received August 26, 1993; final revision received April 22, 1994; accepted April 25, 1994. Address correspondence and reprint requests to Dr. M. F. Beal, Warren 408, Neurology Service, Massachusetts General Hospital, Boston, MA 02114, U.S.A. Abbreviations used: ANOV A, one-way analysis of variance; 3-AP, 3-acetylpyridine; APV, 2-amino-5-phosphonovaleric acid; MPP+, I-methyl-4-phenylpyridinium; NMDA, N-methyl-D- aspartate; PLSD, protected least significant difference; TE, echo time; TR, relaxation time; TTC, 2,3, 5-triphenyltetrazolium chlo- ride monohydrate; 1024 dimensional water-suppressed proton chemical shiſt mag- netic resonance imaging showed a striatal depletion of the neuronal marker N-acetylaspartate but no focal increase in lactate during the first 3 h aſter intrastriatal 3-AP in- jections. Pretreatment with fructose-l,6-biphosphate at- tenuated the lesion volume significantly, which may be due to its ability to serve as a substrate for glycolytic metabolism, with resulting ATP production. The results of the present studies support the hypothesis that 3-AP produces an impairment of energy metabolism due to its substitution for niacinamide in the formation of NAD(P). Furthermore, 3-AP toxicity may involve a secondary ex- citotoxic mechanism mediated by NMDA receptors. Key Words: 3-Acetylpyridine-Aging-Excitotoxicity-N- Methy 1- D-aspartate-Neurodegeneration. to the climbing fiber innervation of the cerebellum (Desclin, 1974; Desclin and Escubi, 1974; Balaban, 1985). Therefore, 3-AP has been used extensively to study cerebellar pathophysiology. The 3-AP- induced degeneration of the inferior olive and cer- ebellum is similar to the pathology seen in olivop- ontocerebellar atrophies, a heterogeneous group of disorders, although the neurotoxin does not cause degeneration of the basis pontis. Other target struc- tures of 3-AP toxicity include the hippocampal for- mation, pars compacta of the substantia nigra with subsequent degeneration of the nigrostriatal projec- tions, nucleus ambiguus, hypoglossal nucleus, dor- sal motor nucleus of the vagus nerve, interpedun- cular nucleus, supraoptic and paraventricular nu- clei, and horizontal limb of the nucleus of the diagonal band (Hicks, 1955; Desclin and Escubi, 1974; Balaban, 1985; Deutsch et aI., 1989). The neurotoxicity of 3-AP is antagonized by prior

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Cerebral Blood Flow and Metabolism 14:1024-1029 © 1994 The International Society of Cerebral Blood Flow and Metabolism Published by Raven Press, Ltd., New York
3-Acetylpyridine Produces Age-Dependent Excitotoxic
Lesions in Rat Striatum
Jorg B. Schulz, D. Ross Henshaw, *Bruce G. Jenkins, tRobert J. Ferrante, tNeil W. Kowall, *Bruce R. Rosen, and M. Flint Beal
Neurochemistry Laboratory, Neurology Service, and *MGH-NMR Center, Department of Radiology, Massachusetts
General Hospital and Harvard Medical School, and tGeriatric Research Education and Clinical Center, Bedford VA
Medical Center, and Departments of Neurology and Pathology, Boston University School of Medicine,
Boston, Massachusetts, U.S.A.
Summary: The effects of 3-acetylpyridine (3-AP) were studied in rat striatum. Striatal injections of 3-AP produced dose-dependent lesions. The lesion size was significantly increased in 4- and 12-month-old rats compared to l-month-old rats. Coinjection of the competitive N-methy l -D - a s p a r t a t e (N M D A ) a n t a g o n i s t 2 - a m i n o - 5-phosphonovaleric acid (APV) or systemic administration of the noncompetitive NMDA antagonist MK-801, the competitive NMDA antagonist L Y274614, or the glutamate release inhibitor lamotrigine partially but significantly attenuated striatal lesion volume. Consistent with an NMDA receptor-mediated excitotoxic effect, histologic studies showed that 3-AP lesions result in relative sparing of NADPH-diaphorase neurons. Using freeze clamp, 3-AP resulted in a marked depletion of ATP. Two-
3-Acetylpyridine (3-AP), a niacinamide antagonist, is a potent neurotoxin when administered to laboratory animals by single intraperitoneal injections. It induces chronic complex motor and behavioral abnormalities, which may be seen as hyperkinesias, ataxia, tremor of the front legs, tonic cramps, and spontaneous convulsions (Herken, 1968). 3-AP has been reported to induce degeneration of the inferior olivary neurons, which give rise
Received August 26, 1993; final revision received April 22, 1994; accepted April 25, 1994.
Address correspondence and reprint requests to Dr. M. F. Beal, Warren 408, Neurology Service, Massachusetts General Hospital, Boston, MA 02114, U.S.A.
Abbreviations used: ANOV A, one-way analysis of variance; 3-AP, 3-acetylpyridine; APV, 2-amino-5-phosphonovaleric acid; MPP+, I-methyl-4-phenylpyridinium; NMDA, N-methyl-Daspartate; PLSD, protected least significant difference; TE, echo time; TR, relaxation time; TTC, 2,3, 5-triphenyltetrazolium chloride monohydrate;
1024
dimensional water-suppressed proton chemical shift magnetic resonance imaging showed a striatal depletion of the neuronal marker N-acetylaspartate but no focal increase in lactate during the first 3 h after intrastriatal 3-AP injections. Pretreatment with fructose-l,6-biphosphate attenuated the lesion volume significantly, which may be due to its ability to serve as a substrate for glycolytic metabolism, with resulting ATP production. The results of the present studies support the hypothesis that 3-AP produces an impairment of energy metabolism due to its substitution for niacinamide in the formation of NAD(P). Furthermore, 3-AP toxicity may involve a secondary excitotoxic mechanism mediated by NMDA receptors. Key Words: 3-Acetylpyridine-Aging-Excitotoxicity-NMethy 1-D-aspartate-N eurodegeneration.
to the climbing fiber innervation of the cerebellum (Desclin, 1974; Desclin and Escubi, 1974; Balaban, 1985). Therefore, 3-AP has been used extensively to study cerebellar pathophysiology. The 3-APinduced degeneration of the inferior olive and cerebellum is similar to the pathology seen in olivopontocerebellar atrophies, a heterogeneous group of disorders, although the neurotoxin does not cause degeneration of the basis pontis. Other target structures of 3-AP toxicity include the hippocampal formation, pars compacta of the substantia nigra with subsequent degeneration of the nigrostriatal projections, nucleus ambiguus, hypoglossal nucleus, dorsal motor nucleus of the vagus nerve, interpeduncular nucleus, supraoptic and paraventricular nuclei, and horizontal limb of the nucleus of the diagonal band (Hicks, 1955; Desclin and Escubi, 1974; Balaban, 1985; Deutsch et aI. , 1989).
The neurotoxicity of 3-AP is antagonized by prior

3-ACETYLPYRIDINE PRODUCES EXCITOTOXIC LESIONS 1025
or concurrent administration of niacinamide in vivo (Desclin and Escubi, 1974; Simantov et aI., 1976) or in vitro (Weller et aI., 1992), suggesting that 3-AP toxicity is caused by competition of 3-AP with niacinamide in the formation of NAD and NADP (Herken, 1968; Weller et aI., 1992). The functional depletion of niacinamide leads to a disturbance of hydride ion transfer occurring between enzymes and substrates using NADH and NADPH as a cofactor. This particularly affects many dehydrogenases (e.g., malate dehydrogenase, lactate dehydrogenase, isocitrate dehydrogenase, a-ketoglutarate dehydrogenase, glucose-6-phosphate dehydrogenase, and glutamate dehydrogenase) and complex I of the mitochondrial electron transport chain. 3-AP may therefore impair energy metabolism due both to effects on the tricarboxylic acid cycle and a depletion of substrate for the electron transport chain.
In the present study we examined the mechanisms of 3-AP toxicity. Based on the attractive hypothesis that an impairment of energy metabolism may secondarily result in slow excitotoxic neuronal death (Albin and Greenamyre, 1992; Beal et aI., 1993c) and the finding that mitochondrial toxins can cause excitotoxic lesions as a result of impairment of oxidative metabolism (Beal et aI., 1991b, 1993a, b; Storey et aI., 1992), we examined whether 3-AP causes an energy depletion and whether 3-AP produces lesions by an excitotoxic mechanism.
METHODS
Male Sprague-Dawley rats (Charles River, Wilmington, MA, U.S.A.) weighing 175-200 g were anesthetized with pentobarbital (50 mg/kg i.p.) 3-AP, 9.1 and 18.2 !-lmol (Sigma, St. Louis, MO, U.S.A.), pH adjusted to 7.4, was injected into the left striatum in a volume of 1 and 2 !-ll, respectively. Injections were made with a 1O-!-l1 Hamilton syringe fitted with a 30-gauge blunt-tipped needle (Beal et aI., 1991a). All injections were made over 1 min and the needle was left in place for a further 2 min before being slowly withdrawn. To study neuroprotective effects animals were treated with either normal saline, MK-801 (5 mg kg-I; Research Biochemical Incorporated, Natick, MA, U.S.A.), lamotrigine (16 mg kg-I; Burroughs Wellcome), L Y274614 (20 mg kg -I; Lilly Pharmaceuticals, Indianapolis, IN, U.S.A.), or fructose-l,6-biphosphate (500 mg kg-I; Sigma) i.p. MK-801 and fructose-l,6-biphosphate was given 0.5 h before and 2.5 h after the striatal injection. Lamotrigine and LY274614 were administered only 0.5 h before the surgery. In one group of animals, a coinjection of 18.2 !-lmol of 3-AP and 240 nmol of 2-amino-5-phosphonovaleric acid (APV; Sigma) was performed. To study the age dependence of the lesion size, 18.2 !-lmol of 3-AP was injected into the striatum of I-month-old (100--125 g; n = 7), 4-month-old (375-400 g, n = 7), and 12-month-old (50{}'-750 g; n = 4) rats. One week after the striatal injection, the animals were sacrificed by decapitation, and the brains were rapidly re-
moved, placed in cold saline for 10 min, and sectioned coronally into slices at 2-mm intervals. Slices were stained with a 2% 2,3,5-triphenyltetrazolium chloride monohydrate (TTC; Sigma) solution at room temperature in the dark for 30 min followed by fixation in 4% formaldehyde (Bederson et aI., 1986). The lesion volumes were determined by computer-assisted planimetry. The lesioned area (noted by pale staining) was measured on the posterior surface of each section, on which a lesion was detectable. The lesion volume was calculated by multiplying the lesioned area of each slice by the thickness of the slice (2 mm) and adding the volumes for every slice (Bioquant IV image analysis system, EVEREX computer, Mti Videocamera, Sony Monitor System, and Olympus SZH microscope). The person measuring the lesions was blinded to the specific treatment of the animals.
For histological evaluation four animals with intraperitoneal saline injection and three animals with MK-801 pretreatment were deeply anesthetized 1 week after the intrastriatal injection of 18.2 !-lmol of 3-AP and perfused with ice-cold saline, followed by phosphate-buffered 4% paraformaldehyde (Beal et aI., 1991a). Brains were sectioned at 50-!-lm intervals on a freezing microtome. Combined staining for Nissl substance and NADPHdiaphorase was performed. Quantitation of striatal neurons was carried out in 450-!-lm2 fields centered around the lesion at a magnification of 250 x, using an eyepiece graticule. In each animal counts were made in five sections at different striatal levels and in 8-10 fields per section and were averaged. Fields of the lesioned striatum were compared with fields in the same location in the contralateral unlesioned striatum.
We examined the effects of intrastriatal injection of 18.2 !-lmol 3-AP on ATP concentrations 3 h after intrastriatal injection in six animals using the freeze-clamp technique. Rats were deeply anesthetized and the skull surface was exposed. After decapitation the heads were rapidly frozen in liquid nitrogen and subsequently stored at -70°C. The striata were dissected from a 2-mm slice frontal to and adjacent to the anterior commissure on a cold plate. The corpus callosum, the septal complex, and the anterior commissure were used as dorsal and lateral, medial, and ventral boundaries. ATP was measured using the luciferase-luciferin assay (Lust et aI., 1981). The effects of intrastriatal injection of 3-AP were assessed in vivo using magnetic resonance imaging (Brouillet et aI., 1993). Rats were imaged under halothane anesthesia with a 4.7-T GE Omega CSI Imager with a 30-mm birdcage coil. The parameters for the T2-weighted imaging were relaxation time (TR)/echo time (TE) = 3,200/80 ms and a 2.5-mm slice. Diffusion-weighted imaging parameters were TRITE = 2,200/70 ms, with a diffusion time of 15 ms, a diffusion gradient time of 15 ms, and symmetric x,
y, and z diffusion gradients of 3.1 G/cm. Images of lactate and N-acetylaspartate were made using three-dimensional water-suppressed chemical shift imaging (Jenkins et aI., 1991). Parameters were TRiTE/T
I = 2,200/272/210
ms. The field of view was 35 x 35 mm, and the slice thickness was 8 mm.
Data are expressed as means ± SD. The statistical significance of differences in lesion volume between groups was determined by Student's t test (two tailed) or oneway analysis of variances (ANOVA) followed by Fisher's protected least significant difference (PLSD) post hoc test to compare group means.
J Cereb Blood Flow Metab, Vol. 14, No. 6, 1994
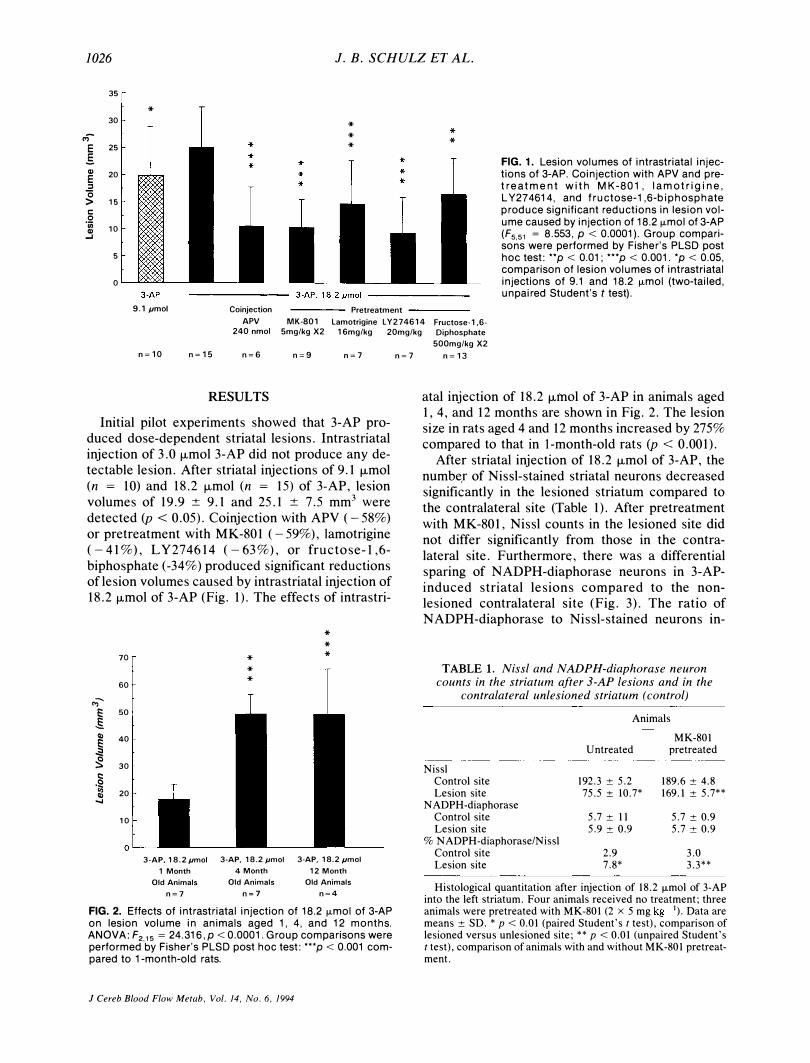
1026 1. B. SCHULZ ET AL.
35
* 30 *
* M- * *
E 25 *
.§ FIG. 1. Lesion volumes of intrastriatal injec-Q) 20 tions of 3-AP. Coinjection with APV and pre-E
t r e a t m e n t w i t h MK- 801, lamotrigine, ::s '0 L Y274614, and fructose-1,6-biphosphate > 15
produce significant reductions in lesion vol-e:: ume caused by injection of 18.2 ILmol of 3-AP 0
'iii 10 (F551 = 8.553, P < 0.0001). Group com pari-Q) --'
sons were performed by Fisher's PLSD post 5 hoc test: **p < 0.01; ***p < 0.001. *p < 0.05,
comparison of lesion volumes of intrastriatal 0 injections of 9.1 and 18.2 ILmol (two-tailed,
unpaired Student's t test). 9.1 pmol Coinjection Pretreatment
APV MK-801 Lamotrigine LY274614 Fructose-1,6-240 nmol Smg/kg X2 16mg/kg 20mg/kg
n�10 n�lS n�6 n�9 n�7
RESULTS
Initial pilot experiments showed that 3-AP produced dose-dependent striatal lesions. Intrastriatal injection of 3.0 /-lmol 3-AP did not produce any detectable lesion. After striatal injections of 9.1 /-lmol (n = 10) and 18.2 fLmol (n = 15) of 3-AP, lesion volumes of 19.9 ± 9.1 and 25.1 ± 7.5 mm3 were detected (p < 0.05). Coinjection with APV (-58%) or pretreatment with MK -801 (-59%), lamotrigine (-41%), LY274614 (-63%), or fructose-l,6-biphosphate (-34%) produced significant reductions of lesion volumes caused by intrastriatal injection of 18.2 /-lmol of 3-AP (Fig. 1). The effects of intrastri-
70
60
M---E: so
,§. Q) 40 E: � � 30 c:
.� 20 Q) .....
10
3-AP, 18.2 pmol
1 Month
Old Animals
n=7
* * *
3-AP. 18.2 pmol
4 Month
Old Animals
n=7
* * *
3-AP. 18.2 pmol
12 Month
Old Animals
n�4
FIG. 2. Effects of intrastriatal injection of 18.2 ILmol of 3-AP on lesion volume in animals aged 1, 4, and 12 months. ANOVA: F215 = 24.316, P < 0.0001. Group comparisons were performed by Fisher's PLSD post hoc test: ***p < 0.001 compared to 1-month-old rats.
J Cereb Blood Flow Metab, Vol. 14, No.6, 1994
n�7
Diphosphate
SOOmg/kg X2
n�13
atal injection of 18.2 /-lmol of 3-AP in animals aged 1 4 and 12 months are shown in Fig. 2. The lesion size
'in rats aged 4 and 12 months increased by 275%
compared to that in I-month-old rats (p < 0.001). After striatal injection of 18.2 /-lmol of 3-AP, the
number of Nissl-stained striatal neurons decreased signifi�antly in the lesioned striatum compared to the contralateral site (Table 1). After pretreatment with MK-801, Nissl counts in the lesioned site did not differ significantly from those in the contralateral site. Furthermor�, there was a differential sparing of NADPH-diaphorase neurons in 3-APinduced striatal lesions compared to the nonlesioned contralateral site (Fig. 3). The ratio of NADPH-diaphorase to Nissl-stained neurons in-
TABLE 1. Nissl and NADPH-diaphorase neuron counts in the striatum after 3-AP lesions and in the
contralateral unlesioned striatum (control)
Animals
MK-801 Untreated pretreated
Nissl Control site 192.3 ± 5.2 189.6 ± 4.8 Lesion site 75.5 ± 10.7* 169.1 ± 5.7**
NAOPH-diaphorase Control site 5.7 ± 11 5.7 ± 0.9 Lesion site 5.9 ± 0.9 5.7 ± 0.9
% NAOPH-diaphorase/Nissl Control site 2.9 3.0 Lesion site 7.8* 3.3**
Histological quantitation after injection of 18.2 ILmol of 3-AP into the left striatum. Four animals received no treatment; three animals were pretreated with MK-801 (2 x 5 mg kg- I). Data are means ± SO. * p < 0.01 (paired Student's t test), comparison of lesioned versus unlesioned site; ** p < 0.01 (unpaired Student's ttest), comparison of animals with and without MK-801 pretreatment.
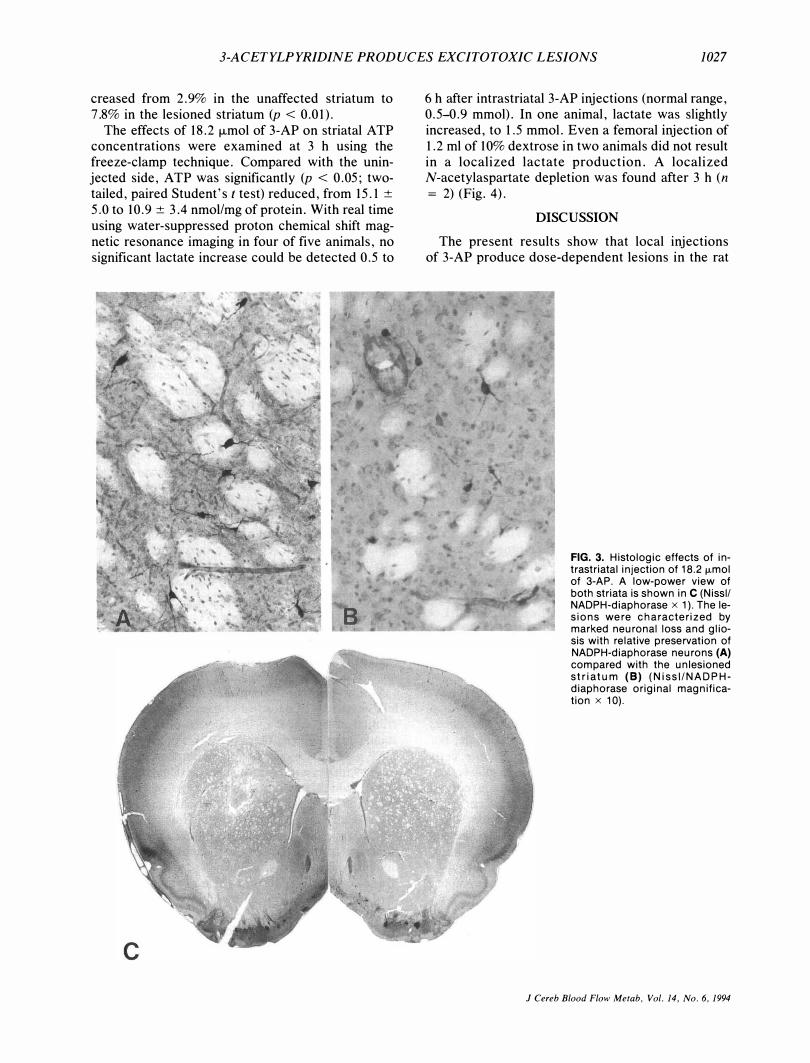
3-ACETYLPYRIDINE PRODUCES EXCITOTOXIC LESIONS 1027
creased from 2.9% in the unaffected striatum to 7.8% in the lesioned striatum (p < 0.0l).
The effects of 18.2 !-Lmol of 3-AP on striatal ATP concentrations were examined at 3 h using the freeze-clamp technique. Compared with the uninjected side, ATP was significantly (p < 0.05; twotailed, paired Student's t test) reduced, from 15.1 ±
5.0 to 10.9 ± 3.4 nmol/mg of protein. With real time using water-suppressed proton chemical shift magnetic resonance imaging in four of five animals, no significant lactate increase could be detected 0.5 to
6 h after intrastriatal 3-AP injections (normal range, 0.5-0.9 mmol). In one animal, lactate was slightly increased, to 1.5 mmol. Even a femoral injection of 1.2 ml of 10% dextrose in two animals did not result in a localized lactate production. A localized N-acetylaspartate depletion was found after 3 h (n
= 2) (Fig. 4).
DISCUSSION
The present results show that local injections of 3-AP produce dose-dependent lesions in the rat
FIG. 3. Histologic effects of intrastriatal injection of 18.2 fLmol of 3-AP. A low-power view of both striata is shown in C (Nissll NADPH-diaphorase x 1). The lesions were characterized by marked neuronal loss and gliosis with relative preservation of NADPH-diaphorase neurons (A) compared with the unlesioned striatum (8) (NissI/NADPHdiaphorase original magnification x 10).
J Cereb Blood Flow Metab, Vol. 14, No.6, 1994
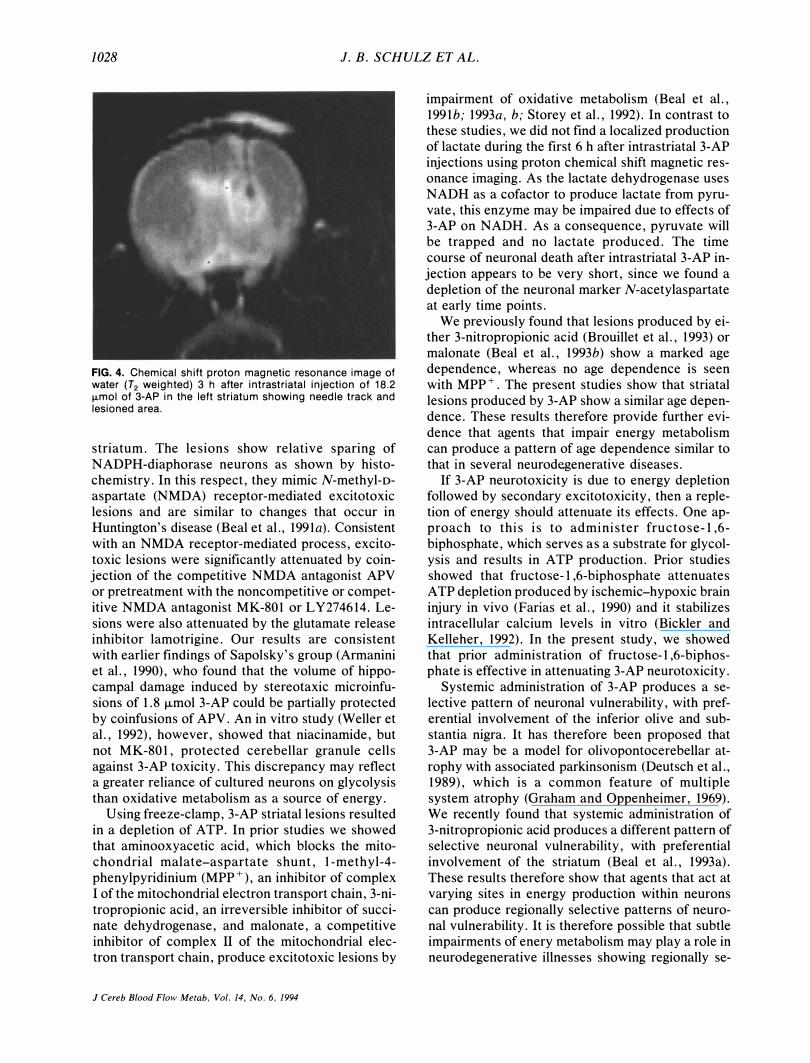
1028 1. B. SCHULZ ET AL.
FIG. 4. Chemical shift proton magnetic resonance image of water (T2 weighted) 3 h after intrastriatal injection of 18. 2 IJ-mol of 3-AP in the left striatum showing needle track and lesioned area.
striatum. The lesions show relative sparing of NADPH-diaphorase neurons as shown by histochemistry. In this respect, they mimic N-methyl-oaspartate (NMDA) receptor-mediated excitotoxic lesions and are similar to changes that occur in Huntington's disease (Beal et aI., 1991a). Consistent with an NMDA receptor-mediated process, excitotoxic lesions were significantly attenuated by coinjection of the competitive NMDA antagonist APV or pretreatment with the noncompetitive or competitive NMDA antagonist MK-801 or LY274614. Lesions were also attenuated by the glutamate release inhibitor lamotrigine. Our results are consistent with earlier findings of Sapolsky's group (Armanini et aI., 1990), who found that the volume of hippocampal damage induced by stereotaxic microinfusions of 1.8 f..Lmol 3-AP could be partially protected by coinfusions of APV. An in vitro study (Weller et aI., 1992), however, showed that niacinamide, but not MK-801, protected cerebellar granule cells against 3-AP toxicity. This discrepancy may reflect a greater reliance of cultured neurons on glycolysis than oxidative metabolism as a source of energy.
Using freeze-clamp, 3-AP striatal lesions resulted in a depletion of A TP. In prior studies we showed that aminooxyacetic acid, which blocks the mitochondrial malate-aspartate shunt, 1-methyl-4-phenylpyridinium (MPP+), an inhibitor of complex I of the mitochondrial electron transport chain, 3-nitropropionic acid, an irreversible inhibitor of succinate dehydrogenase, and malonate, a competitive inhibitor of complex II of the mitochondrial electron transport chain, produce excitotoxic lesions by
J Cereb Blood Flow Metab, Vol. 14, No.6, 1994
impairment of oxidative metabolism (Beal et aI., 1991b; 1993a, b; Storey et aI., 1992). In contrast to these studies, we did not find a localized production of lactate during the first 6 h after intrastriatal 3-AP injections using proton chemical shift magnetic resonance imaging. As the lactate dehydrogenase uses NADH as a cofactor to produce lactate from pyruvate, this enzyme may be impaired due to effects of 3-AP on NADH. As a consequence, pyruvate will be trapped and no lactate produced. The time course of neuronal death after intrastriatal 3-AP injection appears to be very short, since we found a depletion of the neuronal marker N-acetylaspartate at early time points.
We previously found that lesions produced by either 3-nitropropionic acid (Brouillet et aI., 1993) or malonate (Beal et aI., 1993b) show a marked age dependence, whereas no age dependence is seen with MPP+. The present studies show that striatal lesions produced by 3-AP show a similar age dependence, These results therefore provide further evidence that agents that impair energy metabolism can produce a pattern of age dependence similar to that in several neurodegenerative diseases,
If 3-AP neurotoxicity is due to energy depletion followed by secondary excitotoxicity, then a repletion of energy should attenuate its effects. One approach to this is to administer fructose-1,6-biphosphate, which serves as a substrate for glycolysis and results in A TP production. Prior studies showed that fructose-l,6-biphosphate attenuates A TP depletion produced by ischemic-hypoxic brain injury in vivo (Farias et aI., 1990) and it stabilizes intracellular calcium levels in vitro (Bickler and Kelleher, 1992). In the present study, we showed that prior administration of fructose-1,6-biphosphate is effective in attenuating 3-AP neurotoxicity.
Systemic administration of 3-AP produces a selective pattern of neuronal vulnerability, with preferential involvement of the inferior olive and substantia nigra. It has therefore been proposed that 3-AP may be a model for olivopontocerebellar atrophy with associated parkinsonism (Deutsch et aI., 1989), which is a common feature of multiple system atrophy (Graham and Oppenheimer, 1969). We recently found that systemic administration of 3-nitropropionic acid produces a different pattern of selective neuronal vulnerability, with preferential involvement of the striatum (Beal et aI., 1993a), These results therefore show that agents that act at varying sites in energy production within neurons can produce regionally selective patterns of neuronal vulnerability. It is therefore possible that subtle impairments of enery metabolism may play a role in neurodegenerative illnesses showing regionally se-

3-ACETYLPYRIDINE PRODUCES EXCITOTOXIC LESIONS 1029
lective neuronal vulnerability such as multiple system atrophy, Parkinson's disease, and Huntington's disease.
Acknowledgment: We thank Lilly and Burroughs Wellcome Pharmaceuticals for generously supplying us with LY276416 and lamotrigine, respectively. This work was supported by NINDS Grants 16367 and NS 10828 and the Hereditary Disease Foundation. J.B.S. is supported by a fellowship of the Deutsche Forschungsgemeinschaft (Schu 932/1-1).
REFERENCES
Albin RL, Greenamyre JT (1992) Alternative excitotoxic hypotheses. Neurology 42:733-738
Armanini MP, Hutchins C, Stein BA, Sapolsky RM (1990) Glucocorticoid endangerment of hippocampal neurons is NMDA-receptor dependent. Brain Res 532:7-12.
Balaban CD (1985) Central neurotoxic effects of intraperitoneally administered 3-acetylpyridine, harmaline and niacinamide in Sprague-Dawley and Long-Evans rats: a critical review of central 3-acetylpyridine toxicity. Brain Res Rev 9:21-42
Beal MF, Ferrante RJ, Swartz KJ, Kowall NW (l991a) Chronic quinolinic acid lesions in rats closely resemble Huntington's disease. J Neurosci 11:1,649-1,659
Beal MF, Swartz KJ, Hyman BT, Storey E, Finn SF, Koroshetz W (l991b) Aminooxyacetic acid results in excitotoxin lesions by a novel indirect mechanism. J Neurochem 57:1,068-1,073
Beal MF, Brouillet E, Jenkins BG, Ferrante RJ, Kowall NW, Miller JM, Storey E, Srivastava R, Rosen BR, Hyman BT (1993a) Neurochemical and histological characterization of striatal excitotoxic lesions produced by the mitochondrial toxin 3-nitropropionic acid. J Neurosci 13:4181-4192
Beal MF, Brouillet E, Jenkins B, Henshaw R, Rosen R, Hyman BT (1993b) Age-dependent striatal excitotoxic lesions produced by the endogenous mitochondrial inhibitor malonate. J Neurochem 61:1,147-1,150 '
Beal MF, Hyman BT, Koroshetz WJ (1993c) Do defects in mitochondrial energy metabolism underlie the pathology of neurodegenerative diseases. Trends Neurosci 16:125-131
Bederson JB, Pitts LH, Germano SM, Nishimura MC, Davis RL, Bartkowski HM (1986) Evaluation of 2,3,5-triphenyltetrazolium chloride as a stain for detection and quantification of experimental cerebral infarction in rats. Stroke 17: I ,304-1 , 308
Bickler PE, Kelleher JA (1992) Fructose-l,6-biphosphate stabilizes brain intracellular calcium during hypoxia in rats. Stroke 23:1,617-1,622
Brouillet EP, Jenkins BG, Hyman BT, Ferrante RJ, Kowall NW, Srivastava R, Roy SD, Rosen BR, Beal MF (1993) Agedependent vulnerability of the striatum to the mitochondrial toxin 3-nitroproprionic acid. J Neurochem 60:356-359
Desclin JC (1974) Histological evidence supporting the inferior olive as the major source of cerebellar climbing fibers in the rat. Brain Res 77:365-384
Desclin JC, Escubi J (1974) Effects of 3-acetylpyridine on the central nervous system of the rat, as demonstrated by silver methods. Brain Res 77:349-364
Deutsch A Y, Rosin DL, Goldstein M, Roth RH (1989) 3-acetylpyridine-induced degeneration of the nigrostriatal dopamine system: an animal model of olivopontocerebellar atrophy-associated parkinsonism. Exp Neurol 105: 1-9
Farias LA, Smith EE, Markow AK (1990) Prevention of ischemic-hypoxic brain injury and death in Rabbits with fructose-I,6-diphosphate. Stroke 21 :606-613
Graham JG, Oppenheimer DR (1969) Orthostatic hypotension and nicotine sensitivity in a case of multiple system atrophy. J Neuro/ Neurosurg Psychiatry 32:28-34
Herken H (1968) Functional disorders of the brain induced by synthesis of nucleotides containing 3-acetylpyridine. Z Klin Chem Klin Biochem 6:357-472
Hicks SP (1955) Pathologic effects of antimetabolites. I. Acute lesions in the hypothalamus, peripheral ganglia, and adrenal medulla caused by 3-acetylpyridine and prevented by nicotinamide. Am J Pathol 31:189-199
Jenkins BG, Storey E, Beal MF, Rosen BR (1991) Chemical shift imaging of focal neurochemical lesions in rat brain. Proc Soc Magn Reson Med 1:437
Lust WD, Feussner GK, Barbehenn EK, Passoneau JV (1981) The enzymatic measurements of adenine nucleotides and P-creatinine in picomole amounts. Anal Biochem 110:258-266
Simantov R, Snyder SH, Oster-Granite ML (1976) Harmalineinduced tremor in the rat: abolition by 3-acetylpyridine destruction of cerebellar climbing fibers. Brain Res 114: 144-151
Storey E, Hyman BT, Jenkins B, Brouillet E, Miller JM, Rosen BR, Beal MF (1992) I-Methyl-4-phenylpyridinium produces excitotoxic lesions in rat striatum as a result of impairment of oxidative metabolism. J Neurochem 58: I , 975-1 , 978
Weller M, Marini AM, Paul SM (1992) Niacinamide blocks 3-acetylpyridine toxicity of cerebellar granule cells in vitro. Brain Res 594:160-164
J Cereb Blood Flow Metab, Vol. 14, No. 6, 1994
Related Documents











