13-13 Copyright ©The McGraw-Hill Companies, Inc. Permission required for reproduction or display

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

13-13Copyright ©The McGraw-Hill Companies, Inc. Permission required for reproduction or display

Copyright ©The McGraw-Hill Companies, Inc. Permission required for reproduction or display
Special codons: AUG (which specifies methionine) = start codon
AUG specifies additional methionines within the coding sequence UAA, UAG and UGA = termination, or stop, codons
The code is degenerate More than one codon can specify the same amino acid
For example: GGU, GGC, GGA and GGG all code for lysine In most instances, the third base is the degenerate base
It is sometime referred to as the wobble base
The code is nearly universal Only a few rare exceptions have been noted
Refer to Table 13.3
13-14

13-16Figure 13.2
Figure 13.2 provides an overview of gene expression

In the 1950s, Francis Crick & Mahon Hoagland proposed the adaptor hypothesis
tRNAs play a direct role in the recognition of codons in the mRNA
Structure and Function of tRNA
Proline anticodon

Structure of tRNAFigure 13.10
Found in all tRNAstRNA 2º Structure
The modified bases are: I = inosine mI = methylinosine T = ribothymidine D= dihydrouridine m2G = dimethylguanosine = pseudouridine
D loopD
D
TψC loop
loop

3º Structure of tRNA

aminoacyl-tRNA synthetases The enzymes that attach amino acids to tRNAs There are >20 types
One for each amino acid Ones for isoacceptor tRNAs put same a.a. on different tRNAs
Aminoacyl-tRNA synthetases catalyze a two-step reaction 1- adenylation of amino acid 2- aminoacylation of tRNA
Charging of tRNAs

Figure 13.11
Aminoacyl tRNA Synthetase
Function
The amino acid is attached to the 3’ OH
by an ester bond

The genetic code is degenerate There are >20 but < 64 tRNAs How does the same tRNA bind to different codons?
Francis Crick proposed the wobble hypothesis in 1966 to explain the pattern of degeneracy, 1st two bases of the codon-anticodon pair strictly by
Watson-Crick rules The 3rd position can wobble This movement allows alternative H-bonding between
bases to form non-WC base paring
tRNAs and the Wobble Rule

Wobble position and base pairing rulesFigure 13.12
tRNAs charged with the same amino acid, but that recognize multiple codons are
termed isoacceptor tRNAs
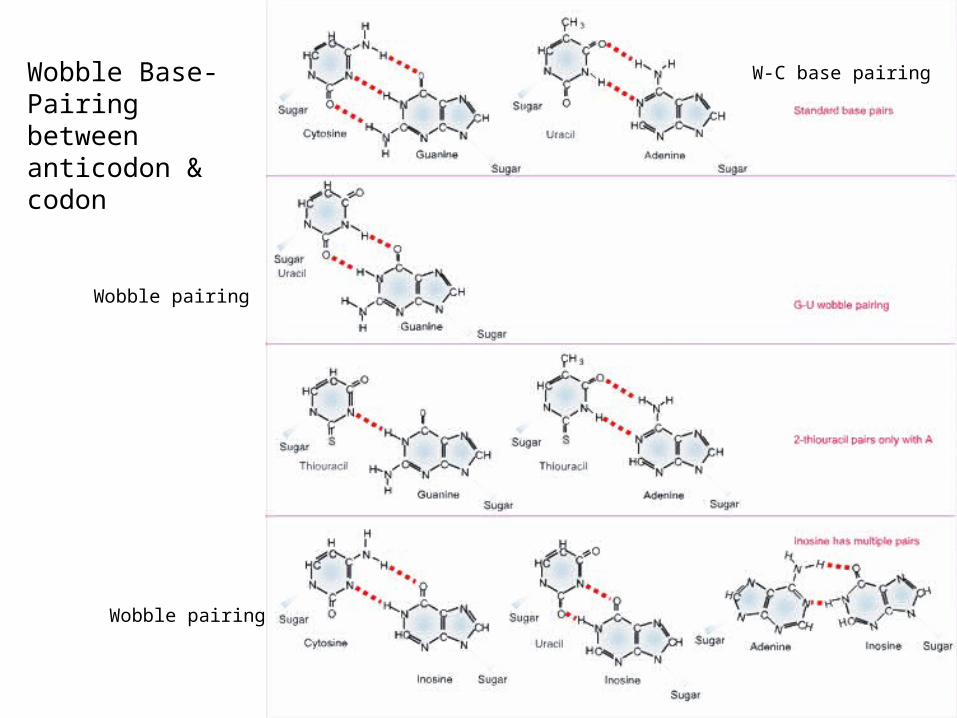
Wobble Base-Pairing between anticodon & codon
W-C base pairing
Wobble pairing
Wobble pairing

Translation occurs on the surface of a large macromolecular complex termed the ribosome
Prokaryotic cells 1 type of ribosome located in the cytoplasm
Eukaryotic cells 2 types of ribosomes 1 found in the cytoplasm 2nd found in organelles -Mitochondria; Chloroplasts
These are like prokaryotic ribosomes
Ribosome Structure and Assembly

Figure 13.13
(a) Bacterial cell
Prokaryotic Ribosomes

Figure 13.13
Eukaryotic Ribosomes

Copyright ©The McGraw-Hill Companies, Inc. Permission required for reproduction or display
During bacterial translation, the mRNA lies on the surface of the 30S subunit As a polypeptide is being synthesized, it exits through a
hole within the 50S subunit
Ribosomes contain three discrete sites Peptidyl site (P site) Aminoacyl site (A site) Exit site (E site)
Ribosomal structure is shown in Figure 13.14
Functional Sites of Ribosomes
13-57

Figure 13.14

Stages of Translation
Initiation Elongation Termination

Release factors
Initiator tRNA
Stages of Translation
Figure 13.15

Components mRNA, initiator tRNA, Initiation factors ribosomal subunits
The initiator tRNA In prokaryotes, this tRNA is designated tRNAi
fmet It carries a methionine modified to N-formylmethionine
In eukaryotes, this tRNA is designated tRNAimet
It carries an unmodified methionine In both cases the initiator tRNA is different from a tRNAmet
that reads an internal AUG codon
Translation Initiation

16S rRNA binds to an mRNA at the ribosomal-binding site or Shine-Dalgarno box
16S rRNAFigure 13.17
Prokaryotic Ribosome-mRNA Recognition
7 nt

(actually 9 nucleotides long)
Figure 13.16
Prokaryotic Translation Initiation

Figure 13.16
The tRNAiMet is
positioned in the P site
All other tRNAs enter the A site
Prokaryotic Translation Initiation

Copyright ©The McGraw-Hill Companies, Inc. Permission required for reproduction or display
In eukaryotes, the assembly of the initiation complex is similar to that in bacteria However, additional factors are required
Note that eukaryotic Initiation Factors are denoted eIF
Refer to Table 13.7
The initiator tRNA is designated tRNAmet It carries a methionine rather than a formylmethionine
Eukaryotic mRNA-Ribosoime Recognition
13-65

The consensus sequence for optimal start codon recognition is show here
Start codon
G C C (A/G) C C A U G G
-6 -5 -4 -3 -2 -1 +1 +2 +3 +4
Most important positions for codon selection
This sequence is called Kozak’s consensus after Marilyn Kozak who first determined it
Eukaryotic Ribosome Binding

Eukaryotic Translation Initiation Initiation factors bind to the 5’ cap in mRNA &
to the pA tail These recruit the 40S subunit, tRNAi
met
The entire assembly scans along the mRNA until reaching a Kozak’s consensus
Once right AUG found, the 60S subunit joins Translation intitiates

During this stage, the amino acids are added to the polypeptide chain, one at a time
The addition of each amino acid occurs via a series of steps outlined in Figure 13.18
This process, though complex, can occur at a remarkable rate In bacteria 15-18 amino acids per second In eukaryotes 6 amino acids per second
Translation Elongation

Figure 13.18
Translation Elongation – tRNA Entry
A charged tRNA binds to the A site
EF-1 facilitates tRNA entry
Peptidyl transferase catalyzes peptide bond formation
The polypeptide is transferred to the aminoacyl-tRNA in the A site
The 23S rRNA (a component of the large subunit) is the actual
peptidyl transferase
Thus, the ribosome is a ribozyme!
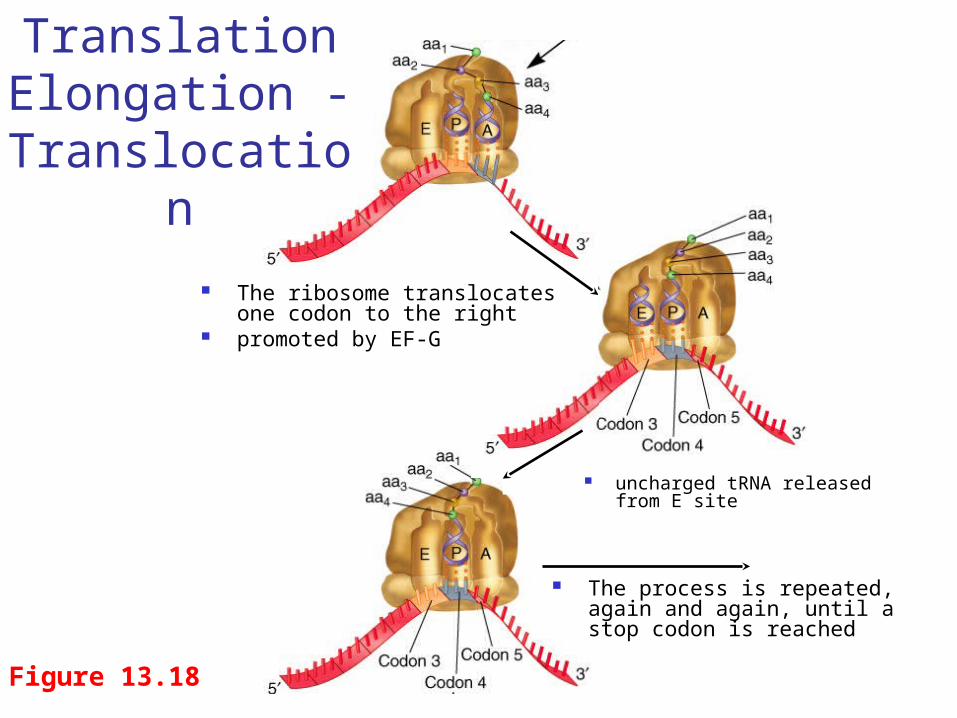
Figure 13.18
The ribosome translocates one codon to the right
promoted by EF-G
Translation Elongation -
Translocation
uncharged tRNA released from E site
The process is repeated, again and again, until a stop codon is reached

Occurs when a stop codon is reached in the mRNA Three stop or nonsense codons
UAG UAA UGA
Recognized by proteins called release factors – NOT tRNAs
Translation Termination

Bacteria have three release factors RF1 - recognizes UAA and UAG RF2 - recognizes UAA and UGA RF3 - binds GTP and facilitates termination process
Eukaryotes only have one release factor
eRF1 - recognizes all three stop codons
Translation Termination

Ribosomal subunits & mRNA dissociate
Figure 13.19
Translation Termination

Translation begins at 5’ end of mRNA 5’3’
Peptide bonds are formed directionally
Peptide bond is formed between the COO- of the previous amino acid in the chain and the NH2 of the amino acid being added
Polypeptides Have Directionality

Carboxyl group Amino group
Peptide Bond Formation
Figure 13.20

Figure 13.20
N terminal C terminal
Colinearity of DNA, mRNA, & Protein Sequence

Figure 13.4
The amino acid sequence of the
enzyme lysozyme
129 amino acids long
Within the cell, the protein will not be found in this linear state It will adapt a
compact 3-D structure
Indeed, this folding can begin during translation
The progression from the primary to the 3-D structure is dictated by the amino acid sequence within the polypeptide

Figure 13.6
A protein subunit

Molecular Basis of Phenotype
Related Documents










