Mesenchymal stroma cells trigger early attraction of M1 macrophages and endothelial cells into fibrin hydrogels, stimulating long bone healing without long-term engraftment Elisabeth Seebach, Holger Freischmidt, Jeannine Holschbach 1 , Jörg Fellenberg, Wiltrud Richter ⇑ Research Centre for Experimental Orthopaedics, Orthopaedic University Hospital Heidelberg, Schlierbacher Landstrasse 200a, 69118 Heidelberg, Germany article info Article history: Received 22 May 2014 Received in revised form 10 July 2014 Accepted 14 July 2014 Available online 22 July 2014 Keywords: Bone healing Mesenchymal stroma cells Fibrin hydrogel Host cell recruitment Macrophage abstract Implantation of mesenchymal stroma cells (MSCs) is an attractive approach to stimulate closure of large bone defects but an optimal carrier has yet to be defined. MSCs may display trophic and/or immunomod- ulatory features or stimulate bone healing by their osteogenic activity. The aim of this study was to unra- vel whether fibrin hydrogel supports early actions of implanted MSCs, such as host cell recruitment, immunomodulation and tissue regeneration, in long bone defects. Female rats received cell-free fibrin or male MSCs embedded in a fibrin carrier into plate-stabilized femoral bone defects. Removed callus was analyzed for host cell invasion (day 6), local cytokine expression (days 3 and 6) and persistence of male MSCs (days 3, 6, 14 and 28). Fibrin–MSC composites triggered fast attraction of host cells into the hydrogel while cell-free fibrin implants were not invaded. A migration front dominated by M1 macrophages and endothelial progenitor cells formed while M2 macrophages remained sparse. Only MSC-seeded fibrin hydrogel stimulated early tissue maturation and primitive vessel formation at day 6 in line with significantly higher VEGF mRNA levels recorded at day 3. Local TNF-a, IL-1b and IL-10 expres- sion indicated a balanced immune cell activity independent of MSC implantation. Implanted MSCs persisted until day 14 but not day 28. Our results demonstrate that fibrin hydrogel is an attractive carrier for MSC implantation into long bone defects, supporting host cell attraction and pro-angiogenic activity. By this angiogenesis, implant integration and tissue maturation was stimulated in long bone healing independent of long-term engraftment of implanted MSCs. Ó 2014 Acta Materialia Inc. Published by Elsevier Ltd. All rights reserved. 1. Introduction The closure of large bone defects secondary to trauma or tumor resection remains a challenge in orthopedic surgery. One promis- ing approach to support natural bone healing is the implantation of bone marrow-derived mesenchymal stroma cells (MSCs) in a suitable carrier [1]. So far, sponge-like scaffolds have mainly been used for cell implantation although these have disadvantages, such as limited cell seeding, unequal cell distribution and non-homoge- neous matrix deposition. Hydrogels are attractive vehicles for cell transplantation because of their easy adaptation to any shape of defect, homogeneous cell distribution, even extracellular matrix deposition and beneficial interaction with the host tissue [2–4]. High biocompatibility, biodegradation and the adhesiveness of fibrin glue [5] have rendered fibrin hydrogel a common carrier for autologous cell transplantation in cartilage repair [6,7]. Surpris- ingly, fibrin has so far attracted little attention as a carrier for MSC- based treatment of bone defects and little is known as to whether fibrin hydrogels support early actions of MSCs in stimulating long bone healing. MSCs can easily be isolated from bone marrow [8], are characterized by fast proliferation in culture, and express CD90, CD105 and vascular cell adhesion molecule 1 (VCAM1, CD106) on their surface in the absence of CD34 and CD45 [9,10]. After expansion culture, MSCs can be induced to differentiate into chondrogenic, adipogenic and osteogenic lineages in vitro [11]. Moreover, MSCs provide a supportive cellular microenvironment by secreting a variety of molecules including growth factors and cytokines which potentially mediate host cell recruitment, angio- genesis and immune modulation [12,13]. MSCs further possess a significant osteogenic potential in vivo and have successfully been used to induce repair of bone defects in animals [14,15] and in humans [16,17]. http://dx.doi.org/10.1016/j.actbio.2014.07.017 1742-7061/Ó 2014 Acta Materialia Inc. Published by Elsevier Ltd. All rights reserved. ⇑ Corresponding author. Tel.: +49 (0) 6221 5629254; fax: +49 (0) 6221 5629288. E-mail address: [email protected] (W. Richter). 1 Current address: Merck KGaA, Frankfurter Strasse 250, 64293 Darmstadt, Germany. Acta Biomaterialia 10 (2014) 4730–4741 Contents lists available at ScienceDirect Acta Biomaterialia journal homepage: www.elsevier.com/locate/actabiomat

1 s2.0-s1742706114003110-main(4)
Jul 22, 2015
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Acta Biomaterialia 10 (2014) 4730–4741
Contents lists available at ScienceDirect
Acta Biomaterialia
journal homepage: www.elsevier .com/locate /ac tabiomat
Mesenchymal stroma cells trigger early attraction of M1 macrophagesand endothelial cells into fibrin hydrogels, stimulating long bone healingwithout long-term engraftment
http://dx.doi.org/10.1016/j.actbio.2014.07.0171742-7061/� 2014 Acta Materialia Inc. Published by Elsevier Ltd. All rights reserved.
⇑ Corresponding author. Tel.: +49 (0) 6221 5629254; fax: +49 (0) 6221 5629288.E-mail address: [email protected] (W. Richter).
1 Current address: Merck KGaA, Frankfurter Strasse 250, 64293 Darmstadt,Germany.
Elisabeth Seebach, Holger Freischmidt, Jeannine Holschbach 1, Jörg Fellenberg, Wiltrud Richter ⇑Research Centre for Experimental Orthopaedics, Orthopaedic University Hospital Heidelberg, Schlierbacher Landstrasse 200a, 69118 Heidelberg, Germany
a r t i c l e i n f o
Article history:Received 22 May 2014Received in revised form 10 July 2014Accepted 14 July 2014Available online 22 July 2014
Keywords:Bone healingMesenchymal stroma cellsFibrin hydrogelHost cell recruitmentMacrophage
a b s t r a c t
Implantation of mesenchymal stroma cells (MSCs) is an attractive approach to stimulate closure of largebone defects but an optimal carrier has yet to be defined. MSCs may display trophic and/or immunomod-ulatory features or stimulate bone healing by their osteogenic activity. The aim of this study was to unra-vel whether fibrin hydrogel supports early actions of implanted MSCs, such as host cell recruitment,immunomodulation and tissue regeneration, in long bone defects. Female rats received cell-free fibrinor male MSCs embedded in a fibrin carrier into plate-stabilized femoral bone defects. Removed calluswas analyzed for host cell invasion (day 6), local cytokine expression (days 3 and 6) and persistence ofmale MSCs (days 3, 6, 14 and 28). Fibrin–MSC composites triggered fast attraction of host cells intothe hydrogel while cell-free fibrin implants were not invaded. A migration front dominated by M1macrophages and endothelial progenitor cells formed while M2 macrophages remained sparse. OnlyMSC-seeded fibrin hydrogel stimulated early tissue maturation and primitive vessel formation at day 6in line with significantly higher VEGF mRNA levels recorded at day 3. Local TNF-a, IL-1b and IL-10 expres-sion indicated a balanced immune cell activity independent of MSC implantation. Implanted MSCspersisted until day 14 but not day 28. Our results demonstrate that fibrin hydrogel is an attractive carrierfor MSC implantation into long bone defects, supporting host cell attraction and pro-angiogenic activity.By this angiogenesis, implant integration and tissue maturation was stimulated in long bone healingindependent of long-term engraftment of implanted MSCs.
� 2014 Acta Materialia Inc. Published by Elsevier Ltd. All rights reserved.
1. Introduction
The closure of large bone defects secondary to trauma or tumorresection remains a challenge in orthopedic surgery. One promis-ing approach to support natural bone healing is the implantationof bone marrow-derived mesenchymal stroma cells (MSCs) in asuitable carrier [1]. So far, sponge-like scaffolds have mainly beenused for cell implantation although these have disadvantages, suchas limited cell seeding, unequal cell distribution and non-homoge-neous matrix deposition. Hydrogels are attractive vehicles for celltransplantation because of their easy adaptation to any shape ofdefect, homogeneous cell distribution, even extracellular matrixdeposition and beneficial interaction with the host tissue [2–4].High biocompatibility, biodegradation and the adhesiveness of
fibrin glue [5] have rendered fibrin hydrogel a common carrierfor autologous cell transplantation in cartilage repair [6,7]. Surpris-ingly, fibrin has so far attracted little attention as a carrier for MSC-based treatment of bone defects and little is known as to whetherfibrin hydrogels support early actions of MSCs in stimulating longbone healing. MSCs can easily be isolated from bone marrow [8],are characterized by fast proliferation in culture, and expressCD90, CD105 and vascular cell adhesion molecule 1 (VCAM1,CD106) on their surface in the absence of CD34 and CD45 [9,10].After expansion culture, MSCs can be induced to differentiate intochondrogenic, adipogenic and osteogenic lineages in vitro [11].Moreover, MSCs provide a supportive cellular microenvironmentby secreting a variety of molecules including growth factors andcytokines which potentially mediate host cell recruitment, angio-genesis and immune modulation [12,13]. MSCs further possess asignificant osteogenic potential in vivo and have successfully beenused to induce repair of bone defects in animals [14,15] and inhumans [16,17].

E. Seebach et al. / Acta Biomaterialia 10 (2014) 4730–4741 4731
According to the concept of stem cell-based bone regeneration,implanted MSCs contribute physically to the repair tissue by gen-erating new bone tissue, as has been reported in ectopic implanta-tion models in immune-deficient mice in which fibrin hydrogelformed part of the carrier [18,19]. The role and fate of MSCsimplanted into a bone defect is, however, unclear and long-termpersistence, especially in immune competent animals, has beendiscussed [20–22]. Fast attraction of a large number of diverse hostcells to the defect site, acceleration of new vessel formation andmodulation of the early inflammatory reaction may be importanttrophic roles of MSCs in stimulation of bone healing which shouldbe supported by the carrier used for implantation. In an orthotopicsetting, these mechanisms are potentially more important forimproved bone regeneration than enhancing the number of localbone-forming cells by implantation of additional osteogenic pro-genitors. Whether fibrin hydrogel is an attractive carrier for MSCimplantation in this respect remains to be determined.
A highly potent MSC population has to be produced in a stan-dardized fashion from each donor to allow optimal healing resultsin the patient. In view of the changing paradigm of MSC function[23], the therapeutically most potent MSC subpopulations may,however, be characterized by pronounced trophic and/or immuno-modulatory features displayed in the early callus instead of a highosteogenic differentiation capacity. Thus, it is essential to betterunderstand relevant actions of MSC composites in a bone defectin order to optimize MSC-based bone tissue engineeringapproaches and to design an optimal application strategy for clin-ical regeneration of large bone defects.
The aim of this study was to assess whether fibrin hydrogel isan attractive carrier for MSC implantation supporting early boneregeneration with regard to host cell attraction, angiogenesis,immunomodulation and physical contribution of implanted MSCsto newly formed bone tissue. In order to follow up implanted MSCswithout extra manipulation by a labelling reaction, our approachwas to implant sex-mismatched MSCs in a fibrin hydrogel into along bone defect in rats. Callus was removed from female recipi-ents at distinct time points after surgery and analyzed for host cellrecruitment, the presence of inflammatory cytokines and the per-sistence of male MSCs to pinpoint the dominant actions offibrin–MSC composites in early bone regeneration.
2. Materials and methods
2.1. Animals and study design
Sprague–Dawley rats (n = 46; 17–23 weeks of age, CharlesRiver, Sulzfeld, Germany) were included in the animal study. Thehind limbs in the growing rat are skeletally mature at around15 weeks of age [24]. The experimental protocol was approvedby the local animal experimental ethics committee and all proce-dures were performed according to the European Laboratory Ani-mal Science Guidelines. Male rats were used for MSC donationand female rats served as MSC recipients. A total of 43 animalsreceived a non-critical 2 mm bone defect in the right femur. Twoexperimental groups were defined. Rats of group 1 received anempty fibrin clot and served as controls (n = 20), while rats ofgroup 2 were treated with 1 � 106 MSCs in a fibrin clot (n = 23).Animals were killed 3, 6, 14 or 28 days after surgery. Histologicalevaluation of callus was performed at day 6 (five animals pergroup). Gene expression analysis and detection of the sex-deter-mining region of Y-Gen (SRY)-specific DNA were assessed in thesame sample at days 3 and 6 from the callus of five animals percondition. Visualization of new bone formation by micro-com-puted tomography (lCT) and SRY-DNA detection was done onthe same animals with n = 3 animals for the cell-free and n = 4
animals for the MSC-treated group at day 14, and with n = 2animals for the cell-free and n = 4 animals for the MSC-treatedgroup at day 28.
2.2. MSC isolation and expansion
MSCs were isolated from bone marrow of three male rats using amodified centrifugation method [25]. Animals were killed byasphyxiation with CO2 and MSCs were extracted from tibiae andfemora by centrifugation at 750g for 2 min. All cells were pooled,washed with phosphate-buffered saline (PBS) and seeded into0.1% gelatin-coated flasks in a high-glucose Dulbecco’s modifiedEagle’s medium (DMEM) containing 12.5% fetal calf serum (FCS),2 mM L-glutamine, 1% non-essential amino acids, 0.1% 2-mercap-toethanol (Invitrogen, Karlsruhe, Germany), 100 U ml–1 penicillin,100 lg ml–1 streptomycin (Biochrom, Berlin, Germany) and4 ng ml–1 recombinant human fibroblast growth factor-2 (ActiveBioscience, Hamburg, Germany) at a density of 1.25 � 105
cells cm–2. After 5 days under standard culturing conditions (37 �Cand 6% CO2) non-adherent cells were removed by washing. Mediumwas changed every 2 days and cells were split at 80% confluency andreplated at a density of 4–6 � 103 cells cm–2. All cells were frozenin passage 1 in liquid nitrogen and recultivated at a density of6 � 103 cells cm–2 3 days before implantation.
2.3. Flow cytometry
At the time of implantation cells were harvested with trypsinand 1 � 105 cells were labelled with anti-CD45 (1:250), anti-CD68 (1:100), anti-CD90-FITC (1:25; Abcam, Cambridge, UK) andanti-CD106-PE (1:50; BD, Heidelberg, Germany) antibodies,respectively. For CD45 and CD68 staining cells were incubatedwith a secondary anti-mouse FITC-conjugated antibody (1:100;Jackson ImmunoResearch Laboratories, Suffolk, UK). Surface mar-ker positive cells were quantified in a FACSCalibur (BD, Heidelberg,Germany).
2.4. Osteogenic, adipogenic and chondrogenic in vitro differentiation
For osteogenesis, quadruplicates of 3.5 � 104 MSCs were seededper well of a 24-well plate in osteogenic medium consistingof high-glucose DMEM supplemented with 10% FCS, 0.1 lMdexamethasone, 0.17 mM ascorbic acid 2-phosphate, 10 mMb-glycero-phosphate (Sigma-Aldrich, Taufkirchen, Germany),100 U ml–1 penicillin and 100 lg ml–1 streptomycin. At days 1, 7and 14 the alkaline phosphatase (ALP) enzyme activity of celllysate was measured as described previously [18]. For adipogenicdifferentiation cells were seeded in duplicates at 3.5 � 104 cellsper well of a 24-well plate in high-glucose DMEM supplementedwith 10% FCS, 0.01 lM dexamethasone, 5 lg ml–1 insulin (Sanofi-Aventis, Frankfurt, Germany), 100 U ml–1 penicillin and100 lg ml–1 streptomycin [26]. Formation of lipid droplets wasevaluated by Oil Red O staining (Chroma, Münster, Germany) afterfixing the cells in 4% paraformaldehyde (PFA, Merck, Darmstadt,Germany). Chondrogenesis was performed with 5 � 105 cells in a3D pellet-culture in high-glucose DMEM supplemented with5 lg ml–1 transferrin, 5 ng ml–1 sodium selenite, 0.1 lM dexameth-asone, 0.17 mM ascorbic acid 2-phosphate, 1 mM sodium pyru-vate, 0.35 mM proline, 1.25 mg ml–1 BSA (Sigma-Aldrich,Taufkirchen, Germany), 100 U ml–1 penicillin, 100 lg ml–1 strepto-mycin, 5 lg ml–1 insulin and 10 ng ml–1 transforming growth fac-tor-beta 3 (TGFb-3; Miltenyi Biotec GmbH, Bergisch Gladbach,Germany). Differentiation was evaluated on day 14 by Safranin Ostaining (Chroma, Münster, Germany) for proteoglycan depositionand collagen type II immunohistochemistry as described previ-ously [27].

4732 E. Seebach et al. / Acta Biomaterialia 10 (2014) 4730–4741
2.5. Preparation of fibrin-MSC composites for implantation
MSCs were prepared separately for every animal, to limit thetime period between preparing the fibrinogen cell suspensionand implantation to �20 min. The fibrinogen solution (Baxter,Wien, Austria) was diluted 1:2 and the thrombin (Baxter, Wien,Austria) was diluted 1:50 in PBS containing protease inhibitorcocktail (cOmplete Mini: Roche, Mannheim, Germany). 1 � 106
MSCs were resuspended in 40 ll of the fibrinogen solution, keptat room temperature until 40 ll of the thrombin solution wasadded. The solidifying gel was transferred into the bone defect.Fibrin clots for the control animals were prepared without cells.
Five additional fibrin–MSC clots were frozen at �80 �C and usedfor gene expression analysis of MSCs before implantation. Threeadditional fibrin(–MSC) clots were used for histological examina-tion of non-implanted samples.
2.6. Implantation of fibrin–MSC composites in the bone defect
Rats were anaesthetized by intraperitoneal injection ofketamine hydrochloride (Ketamin: 100 mg kg–1 body weight,Orion Pharma, Hamburg, Germany) and medetomidine hydrochlo-ride (Domitor: 0.2 mg kg–1 body weight, Orion Pharma, Hamburg,Germany). A longitudinal incision (30 mm) of skin and fascia ofthe right hind leg was performed. The vastus lateralis and bicepsfemoris muscles were carefully separated and the anterolateralside of the femoral shaft was exposed. The attached muscles werestripped from the bone and a polyether ether ketone (PEEK) plate(Rat Fix, AO, Davos, Switzerland) was adjusted to the femur. ThePEEK plate was held in position and pilot holes were drilledthrough both cortices along the predrilled holes of the plate. Theplate was fixed to the bone by three screws (Rat Fix, AO, Davos,Switzerland) at each side. A 2 mm bone defect was created witha sawing wire (Rat Fix, AO, Davos, Switzerland). Bleeding was pre-vented by pressing a gelatin sponge (Gelita-Spon: Gelita Medical,Amsterdam, the Netherlands) into the bone defect for 2 min. Thepolymerizing fibrin clot was quickly transferred into the bonegap via a pipette and allowed to solidify for 3 min. Muscles, deepfascia, subcutaneous fascia and skin were closed in a standard fash-ion. Animals were treated once with 0.02 ml gentamicin sulfate(Refobacin: 80 mg, Merck, Darmstadt, Germany) for antibiosisand buprenorphine hydrochloride (Temgesic: 0.01 mg kg–1 bodyweight, RB Pharmaceuticals, UK) was given subcutaneously foranalgesia. The animals were allowed full activity in their cagesand were medicated with buprenorphine hydrochloride (Temge-sic: 0.01 mg kg–1 body weight) for the first two postoperative days.At days 3, 6, 14 or 28 after surgery animals were narcotized withCO2 and killed by exsanguination via heart puncture. Femora weredissected by opening the sutures and careful stripping off of thesurrounding tissue. Callus was carefully isolated from bone defectand either directly frozen at �80 �C for MSC detection and geneexpression analysis or fixed in 4% PFA for histology.
2.7. lCT
New bone formation of day 14 and 28 samples was evaluatedusing a Sky-Scan 1076 in vivo X-ray microtomograph (Skyscan,Antwerpen, Belgium). Femora were scanned with surroundingmuscles wrapped in plastic foil using the 0.025 mm titan filter,with the following settings: voxel size 17.7 lm, voltage 65 kV, cur-rent 139 lA, exposure time 280 ms, frame averaging 3. Data wererecorded every 0.4� rotation step through 180�. Reconstruction wasperformed using NRecon� software (version 1.6.3.2, Skyscan,Antwerp, Belgium). 3-D pictures were made with CTan�/CTVol�
software (Skyscan, Antwerp, Belgium).
2.8. DNA isolation and SRY-gene-specific PCR
Genomic DNA from day 3 and 6 callus was extracted along withtotal RNA according to the manufacturer’s protocol using the peq-GOLD TriFast technique (peqlab, Erlangen, Germany). Day 14 or 28callus was digested in lysis buffer with 100 ng ml–1 proteinase K(Fermentas, St Leon-Rot, Germany) at 56 �C overnight and DNAwas extracted by ethanol precipitation. A SRY-gene-specific DNAfragment was amplified in a standard PCR using Taq DNA Polymer-ase (Invitrogen, Karlsruhe, Germany). For detection of implantedmale MSCs in defined regions, day 6 callus was separated by lasermicrodissection, and DNA was isolated using the QIAmp DNAMicro Kit (Qiagen GmbH, Hilden, Germany) according to the man-ufacturer’s protocol. DNA analogously isolated from equivalentsections of the control samples was used as negative control.SRY-gene-specific DNA was amplified by nested PCR. PCR productswere visualized on a 2% agarose gel and their specificity was con-firmed by DNA sequencing.
2.9. Gene expression analysis
Callus was homogenized in peqGOLD TriFast and total RNAwas extracted according to the manufacturer’s protocol (peqlab,Erlangen, Germany). cDNA synthesis was performed with 1 lgtotal RNA using Omniscript reverse transcriptase (0.2 U ll–1), oli-go(dT) primer (1 lM, both Qiagen GmbH, Hilden, Germany) anda ribonuclease inhibitor (RNaseOUT: 40 U ll–1, Invitrogen, Kar-lsruhe, Germany) according to the manufacturer’s instructions.Expression levels of individual genes were analyzed by quantita-tive PCR (Stratagene Mx3000P; Agilent Technologies, Böblingen,Germany). cDNA were amplified using gene-specific primer sets(Table 1) obtained from Eurofins (Ebersberg, Germany) and real-time fluorimetric intensity of SYBR green I (Thermo Scientific,Rockford, USA) was monitored. The apparent threshold cycles(Ct) of the genes of interest were compared to the Ct of the refer-ence gene hypoxanthine-guanine phosphoribosyltransferase-1(HPRT-1) in the same cDNA sample. When indicated, the percent-age of the reference gene expression levels shown in the figuresrefer to relative levels as a percentage of HPRT-1. Melting curvesand agarose gel electrophoresis of the PCR products were usedfor quality control.
2.10. Histology
Callus was isolated from the bone defect and subjected to his-tology without proximal and distal bone ends to allow preparationof the samples without decalcification. In brief, samples were fixedin 4% PFA, dehydrated in a graded alcohol series and embedded inparaffin. In vivo orientation of the callus was maintained through-out the embedding process and callus was cut along this axis. Forhistological investigations, sections (5 lm) from the callus centerwere used and stained with haematoxylin and eosin (H&E:Chroma, Münster, Germany) or Movat’s Pentachrome (all colours:Chroma, Münster, Germany) in a standard procedure.
For CD31 and CD68 immunohistochemistry, digestion with20 ng ml–1 proteinase K (Fermentas, St Leon-Rot, Germany) for20 min at 37 �C was used for antigen retrieval. Sections forCD163 staining were pre-treated with 1 mg ml–1 pronase (Roche,Mannheim, Germany) for 30 min at 37 �C. No antigen retrievalwas required for CCR7 staining. Unspecific binding was blockedfor 30 min with 5% BSA for CD31, CD68 and CD163, and with 5%human serum for CCR7, and then sections were incubated withthe primary antibodies CD31 (1:200; Abbiotec, San Diego, CA,USA), CD68 (1:100; Abcam, Cambridge, UK), CD163 (1:100, AbDSerotec, Oxford, UK) and CCR7 (1:250; Epitomics, Burlingame, CA,USA) overnight at 4 �C. After washing sections were incubated with

Table 1List of oligonucleotides used for qRT-PCR analysis and SRY-gene-specific PCR.
Gene GenBank no. Forward primer Reverse primer
BSP [NM_012587.2] ACGCTGGAAAGTTGGAGTTAG GACCTGCTCATTTTCATCCACD45 [NM_001109890.1] GCATGCATCAATCCTAGTCC GGCCATGATGTCATAGAGGAHPRT-1 [NM_012583.2] GCCAGACTTTGTTGGATTTG CACTTTCGCTGATGACACAAIL-1b [NM_031512.2] GACAAGCAACGACAAAATCC ACCGCTTTTCCATCTTCTTCIL-2 [NM_053836.1] AGCGTGTGTTGGATTTGACT TCTCCTCAGAAATTCCACCAIL-6 [NM_012589.2] AGCCAGAGTCATTCAGAGCA AGTTGGATGGTCTTGGTCCTIL-10 [NM_012854.2] GACGCTGTCATCGATTTCTC TTCATGGCCTTGTAGACACCMIP-2 [NM_053647.1] TGAAGTTTGTCTCAACCCTGA GGTGCAGTTCGTTTCTTTTCTOC [NM_013414.1] AGGGCAGTAAGGTGGTGAAT CTAAACGGTGGTGCCATAGAPPAR-c [NM_013124.3] ATAAAGTCCTTCCCGCTGAC ATCTCTTGCACAGCTTCCACSRY [X89730.1] CTTTCGGAGCAGTGACAGTT CACTGATATCCCAGCTGCTTSRY nested [X89730.1] CTTTCGGAGCAGTGACAGTT CATGCTGGGATTCTGTTGATNF-a [NM_012675.3] TCTACTGAACTTCGGGGTGA CCACCAGTTGGTTGTCTTTGVEGFa [NM_031836.3] CAATGATGAAGCCCTGGA CTATGCTGCAGGAAGCTCAT
E. Seebach et al. / Acta Biomaterialia 10 (2014) 4730–4741 4733
the respective biotinylated secondary antibodies (CD31 and CCR7:anti-rabbit IgG; CD68 and CD163: anti-mouse IgG; 1:500; bothDianova, Hamburg, Germany), followed by ALP–streptavidin (Vec-tastain; Vector Laboratories, Burlingame, CA, USA) and Fast Red(Roche, Mannheim, Germany). The nuclei were counterstainedwith haematoxylin (Chroma) and slides were covered in Aquatex(Merck, Darmstadt, Germany). Negative controls were performedby omitting the respective primary antibody. Histomorphometricevaluation was performed on H&E-stained sections of two zonesof every callus respectively (n = 10 per group). Total callus areawas marked black and the area infiltrated by host cells was labelledgrey using Photoshop 7.0 (Adobe Systems Inc., USA). ImageJ 1.44p(Wayne Rasband, National Institutes of Health, USA) was used toassess the total area and the infiltrated area. Cell invasion is statedas the percentage of infiltrated area related to total area.
2.11. Laser microdissection
To evaluate persistence of implanted MSCs next to invadinghost cells we searched for possible SRY-signals in the infiltratedarea of the callus vs. the non-infiltrated part of the MSC implant.Appropriate parts of the tissue section were separated by lasermicrodissection. In brief, sections (10 lm) of the MSC-containingcallus were taken up on manually foil-coated slides. After H&Estaining, the area infiltrated by host cells was separated from theremaining implant using a PALM MicroBeam Laser MicrodissectionSystem (Carl Zeiss AG, Jena, Germany). To obtain enough materialfor nested PCR, tissue collected from the infiltrated area (two sec-tions per sample from five animals, n = 10) or of the remainingfibrin clot were pooled, respectively (n = 10) and subjected toDNA extraction. Analogously, tissue collected from the animalsreceiving cell-free fibrin (n = 10) were pooled and further used asfemale control.
2.12. Statistical analysis
For comparison of gene expression between the MSC groupsand control groups of days 3 and 6 a Kruskal–Wallis test withpost-hoc Mann–Whitney U-tests was conducted. Only if the Krus-kal–Wallis tests indicated significance were data analyzed post hocand corrected with the Bonferroni test. The Mann–Whitney U-testwas furthermore applied for histomorphometric evaluation of theMSC group and the control group at day 6 and for comparison ofgenomic DNA content between the MSC group and the controlgroup at day 14. A two-tailed significance value of P < 0.05 wasconsidered statistically significant. Data analysis was performedwith SPSS for Windows 16.0 (SPSS Inc., Chicago, IL, USA).
3. Results
3.1. Enhanced attraction of host cells into fibrin–MSC composites
One pool of MSCs from long bones of three male rats was pro-duced and extensively characterized before application in allexperiments. Cells were strongly positive for the rat surface mark-ers CD90 (97%) and CD106 (35%) while almost negative for CD45(1.8%) and CD68 (1.7%) as expected for MSC populations(Fig. 1A). During in vitro osteogenesis, cells showed a strong up-regulation of the osteogenic markers bone sialoprotein (BSP) andosteocalcin (OC) and an increasing ALP enzyme activity (Fig. 1B).During adipogenic induction cells up-regulated mRNA levels ofthe adipogenic marker peroxisome proliferator-activated recep-tor-gamma (PPAR-c and deposited lipid droplets as visualized byOil Red O staining (Fig. 1C). Chondrogenic differentiation in high-density pellets resulted in deposition of a proteoglycan- and colla-gen type II-rich extracellular matrix (Fig. 1D).
MSCs were seeded in fibrin hydrogel allowing a standardizedimplantation of a defined concentration of cells into the 2 mmplate-stabilized femur defect of female rats. The hydrogel wasdirectly transferred after starting the polymerization and solidifiedwithin the defect, enabling a complete filling of the bone gap bythe implant with intimate contact to the open bone endings(Fig. 2A). Six days after implantation cell-seeded and non-seededfibrin implants were still present in the bone defect (Fig. 2B). Thein vivo orientation was maintained for histology (Fig. 2C). Asshown in Fig. 3, staining of parallel non-implanted fibrin–MSCcomposites showed a homogeneous distribution of the seeded cellswithin the fibrin hydrogel (Fig. 3H inset). Six days post-implanta-tion, overview staining of control samples revealed little infiltra-tion of host cells into the MSC-free implant but indicated somedissolution of the fibrin from the proximal side of the bone defectin two out of five samples (Fig. 3A). This appeared to create spacefor accumulation of some densely packed host cells in a callusaside from the fibrin (Fig. 3B), while the fibrin matrix itselfremained almost empty (Fig. 3C, D). In contrast, large numbers ofhost cells infiltrated the implant in the MSC group, especially fromthe proximal bone ending (Fig. 3E). Interestingly, in three out offive samples, a dense migration front was formed consisting ofpacked cells which apparently moved through the defect fromthe proximal to the distal side (Fig. 3F, G). The majority of MSCslocated in front of this border looked viable (Fig. 3H). Behind themigration front, the first signs of tissue maturation and extracellu-lar matrix deposition were observed (Fig. 3F) with evidence ofsome proteoglycan deposition in the three most advancedsamples according to pentachrome staining (Fig. 3I). A comparable

Fig. 1. Characterization of rat MSCs. (A) Male rat MSCs pooled from three donors were expanded for two passages, labelled with the indicated antibodies and analyzed byflow cytometry. Representative histograms are shown with the stained population in black and the respective controls (no primary antibody) as grey lines. (B–D) MSCs weresubjected to osteogenic (B), adipogenic (C) or chondrogenic conditions (D) for 2 weeks. (B) For osteogenesis, expression of bone sialoprotein (BSP) and osteocalcin (OC) wasquantified by RT-qPCR and ALP-enzyme activity was measured (n = 2 experiments). Results are expressed as mean ± standard deviation. (C) Adipogenesis was confirmed bygene expression of peroxisome proliferator-activated receptor-gamma (PPAR-c) and Oil Red O staining of formed lipid droplets. (D) MSCs underwent chondrogenicdifferentiation according to deposition of proteoglycans (Safranin O staining, left side) and collagen type II (immunohistochemistry, right side). Staining is representative forthree replicates. Scale bar: 100 lm.
Fig. 2. Implantation of fibrin–MSC composites into long bone defects in rats and processing of callus for histology. A plate-stabilized 2 mm bone defect was created in theright femur of 42 female rats. Either 1 � 106 male MSCs embedded in a fibrin gel or empty fibrin gel (80 ll) were implanted into the defect. After 3, 6, 14 or 28 days femorawere removed and early callus was lifted out from the defect for further analysis. (A) View directly after implantation of the fibrin gel with MSCs. (B) Early callus formation6 days after MSC implantation. (C) For histology, day 6 callus was separated by a scalpel from the bone endings and lifted out of the bone defect. In vivo orientation wasrecorded throughout the embedding process and during histological evaluation.
4734 E. Seebach et al. / Acta Biomaterialia 10 (2014) 4730–4741

Fig. 3. Histological and histomorphometric evaluation of day 6 callus. (A–H) Paraffin sections of formalin-fixed tissue were stained with H&E by a standard procedure. (A, E)Overview pictures of the callus at day 6 with the proximal explant side in the upper left and distal explant side in the lower right corner. Scale bar: 500 lm. Enlarged areas asdefined in (A) and (E) are shown in (B)–(D) and (F)–(H). (B) Host-derived callus; (C) contact zone between host tissue and implant; (D) empty fibrin clot. Inset: H&E-stainedempty fibrin clot before implantation. (F) Highly infiltrated area from (E); (G) the migration front; (H) remaining MSC-seeded fibrin clot. Inset: H&E stained MSC-seeded fibrinclot before implantation. Representative pictures of five replicates are shown in (A)–(H). Scale bar: 100 lm. (I) Movat’s Pentachrome staining: the infiltrated area of the threemost mature samples of the MSC group showed deposition of glycosaminoglycan (light blue staining) next to fibrous tissue (red staining). Scale bar: 200 lm. (J) Forhistomorphometric evaluation, callus was divided into infiltrated area (dark grey) and unaffected implant (light grey). (K) Histomorphometric evaluation of the areainfiltrated by host cells in relation to the total callus area (%, n = 10 sections per group, two per each of the five animals). Boxes represent the 25th and 75th percentile, medianis given as horizontal line and whiskers are maximal and minimal values. Significant difference (P < 0.01) between with and without MSCs is designated by two asterisks.
E. Seebach et al. / Acta Biomaterialia 10 (2014) 4730–4741 4735
maturation tissue was not seen in the fibrin control group.Histomorphometric assessment of the host cell-infiltrated vs. non-infiltrated areas (Fig. 3J) revealed a significantly larger invadedarea in the fibrin–MSC composites compared to control samples(P = 0.01; Fig. 3K).
3.2. M1 macrophages and endothelial progenitor cells as maininvaders
Immunohistochemical staining revealed that the fibrin–MSCcomposites were mainly infiltrated by CD68- and CD31-positivecells indicative of macrophages and endothelial cells (Fig. 4).CD68-positive macrophages were present throughout the infil-trated area and were enriched in the migration front (Fig. 4B).Among ‘‘pioneer cells’’ migrating ahead of the more crowded zoneand penetrating deep into the fibrin clot, many cells stained CD68positive (Fig. 4B, lower right corner). In several areas, elongatedCD31-positive cells apparently migrated from the proximal to thedistal part of the implant forming primitive structures consistentwith the formation of immature microvessels (Fig. 4D, blackarrows).
Macrophages can occur in a predominantly pro-inflammatorysubtype M1 characterized by CCR7 expression and a predomi-nantly anti-inflammatory subtype M2 characterized by CD163expression [28,29]. Staining for M1 and M2 macrophage markers
at day 6 revealed that CCR7 signals were dominant in cell-denseareas. CCR7-positive cells also surrounded elongating structures,presumably areas of primitive vessel formation and tissue matura-tion (Fig. 4F, black arrows). In contrast, rather few CD163-positivecells were present in the host cell infiltrated area. Only in the sam-ple with the most advanced tissue regeneration had CD163-positive cells accumulated, especially in the cell-dense migrationfront (Fig. 4H). In the control group, the host tissue-derived calluscontained CD31-, CD68- and/or CCR7-positive cells but hardly anycells were present in the fibrin hydrogel (Fig. 4A, C, E). CD163-positive (M2) cells, however, were not observed (Fig. 4G).
3.3. Cytokine expression in the early callus
In order to analyze the trophic and immune modulatory effectsof MSCs in the early callus, implants were harvested 3 and 6 daysafter surgery, respectively, to assess mRNA levels by quantitativePCR for a panel of pro- and anti-inflammatory mediators (Fig. 5).At the time of implantation, MSCs expressed vascular endothelialgrowth factor (VEGF), interleukin-6 (IL-6) and macrophage inflam-matory protein-2 (MIP-2, Fig. 5A). In vivo, median VEGF expressionper cell increased from day 0 to day 3, reaching significantly higherlevels in the MSC group vs. controls at day 3 (3.7-fold; P = 0.009)suggesting an enriched pro-angiogenic environment (Fig. 5D).IL-6 and MIP-2 were also expressed in vivo (Fig. 5E, F). Negligible
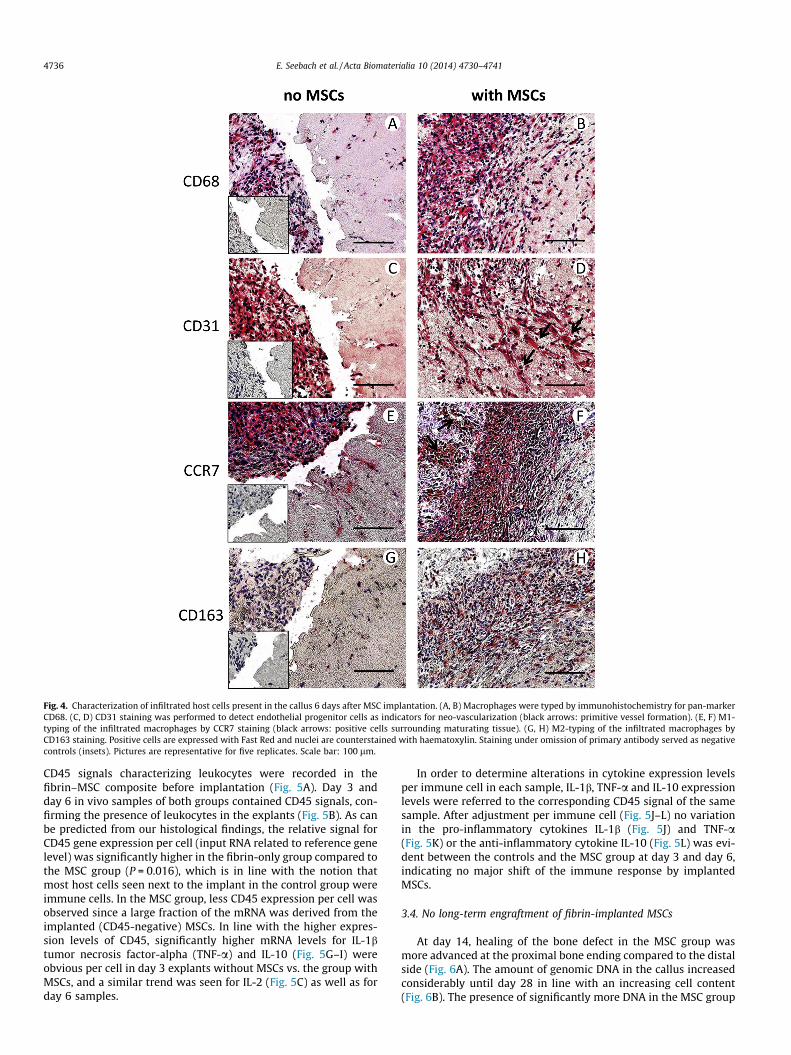
Fig. 4. Characterization of infiltrated host cells present in the callus 6 days after MSC implantation. (A, B) Macrophages were typed by immunohistochemistry for pan-markerCD68. (C, D) CD31 staining was performed to detect endothelial progenitor cells as indicators for neo-vascularization (black arrows: primitive vessel formation). (E, F) M1-typing of the infiltrated macrophages by CCR7 staining (black arrows: positive cells surrounding maturating tissue). (G, H) M2-typing of the infiltrated macrophages byCD163 staining. Positive cells are expressed with Fast Red and nuclei are counterstained with haematoxylin. Staining under omission of primary antibody served as negativecontrols (insets). Pictures are representative for five replicates. Scale bar: 100 lm.
4736 E. Seebach et al. / Acta Biomaterialia 10 (2014) 4730–4741
CD45 signals characterizing leukocytes were recorded in thefibrin–MSC composite before implantation (Fig. 5A). Day 3 andday 6 in vivo samples of both groups contained CD45 signals, con-firming the presence of leukocytes in the explants (Fig. 5B). As canbe predicted from our histological findings, the relative signal forCD45 gene expression per cell (input RNA related to reference genelevel) was significantly higher in the fibrin-only group compared tothe MSC group (P = 0.016), which is in line with the notion thatmost host cells seen next to the implant in the control group wereimmune cells. In the MSC group, less CD45 expression per cell wasobserved since a large fraction of the mRNA was derived from theimplanted (CD45-negative) MSCs. In line with the higher expres-sion levels of CD45, significantly higher mRNA levels for IL-1btumor necrosis factor-alpha (TNF-a) and IL-10 (Fig. 5G–I) wereobvious per cell in day 3 explants without MSCs vs. the group withMSCs, and a similar trend was seen for IL-2 (Fig. 5C) as well as forday 6 samples.
In order to determine alterations in cytokine expression levelsper immune cell in each sample, IL-1b, TNF-a and IL-10 expressionlevels were referred to the corresponding CD45 signal of the samesample. After adjustment per immune cell (Fig. 5J–L) no variationin the pro-inflammatory cytokines IL-1b (Fig. 5J) and TNF-a(Fig. 5K) or the anti-inflammatory cytokine IL-10 (Fig. 5L) was evi-dent between the controls and the MSC group at day 3 and day 6,indicating no major shift of the immune response by implantedMSCs.
3.4. No long-term engraftment of fibrin-implanted MSCs
At day 14, healing of the bone defect in the MSC group wasmore advanced at the proximal bone ending compared to the distalside (Fig. 6A). The amount of genomic DNA in the callus increasedconsiderably until day 28 in line with an increasing cell content(Fig. 6B). The presence of significantly more DNA in the MSC group
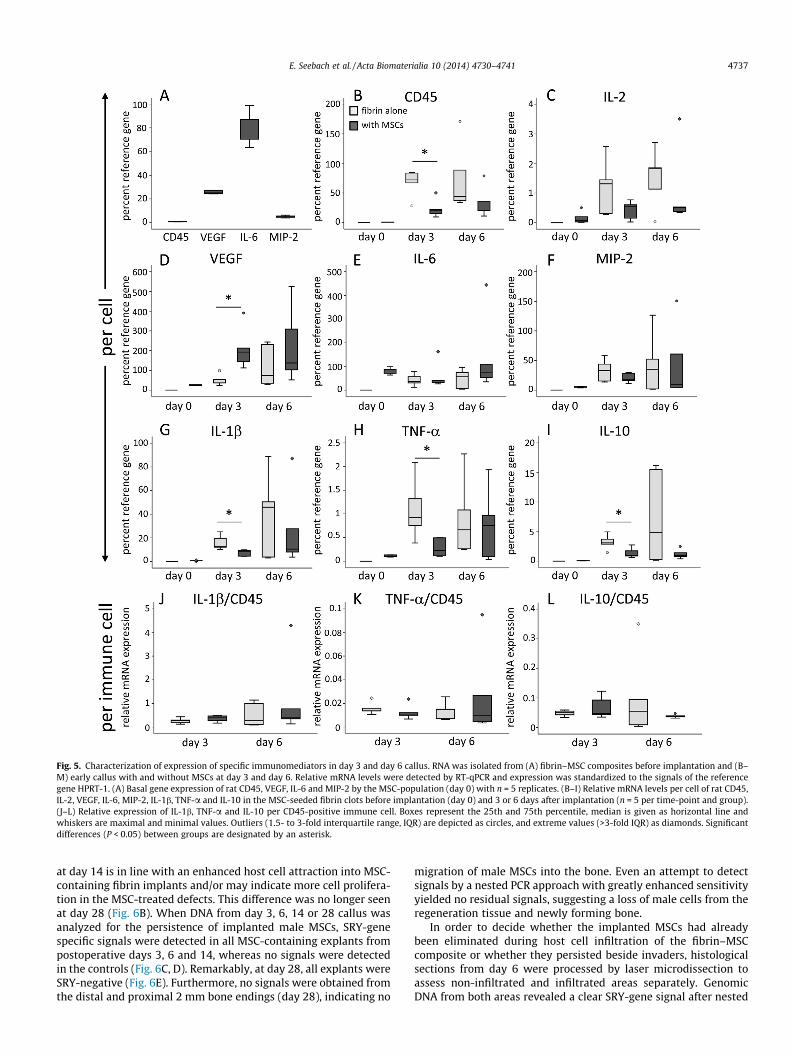
Fig. 5. Characterization of expression of specific immunomediators in day 3 and day 6 callus. RNA was isolated from (A) fibrin–MSC composites before implantation and (B–M) early callus with and without MSCs at day 3 and day 6. Relative mRNA levels were detected by RT-qPCR and expression was standardized to the signals of the referencegene HPRT-1. (A) Basal gene expression of rat CD45, VEGF, IL-6 and MIP-2 by the MSC-population (day 0) with n = 5 replicates. (B–I) Relative mRNA levels per cell of rat CD45,IL-2, VEGF, IL-6, MIP-2, IL-1b, TNF-a and IL-10 in the MSC-seeded fibrin clots before implantation (day 0) and 3 or 6 days after implantation (n = 5 per time-point and group).(J–L) Relative expression of IL-1b, TNF-a and IL-10 per CD45-positive immune cell. Boxes represent the 25th and 75th percentile, median is given as horizontal line andwhiskers are maximal and minimal values. Outliers (1.5- to 3-fold interquartile range, IQR) are depicted as circles, and extreme values (>3-fold IQR) as diamonds. Significantdifferences (P < 0.05) between groups are designated by an asterisk.
E. Seebach et al. / Acta Biomaterialia 10 (2014) 4730–4741 4737
at day 14 is in line with an enhanced host cell attraction into MSC-containing fibrin implants and/or may indicate more cell prolifera-tion in the MSC-treated defects. This difference was no longer seenat day 28 (Fig. 6B). When DNA from day 3, 6, 14 or 28 callus wasanalyzed for the persistence of implanted male MSCs, SRY-genespecific signals were detected in all MSC-containing explants frompostoperative days 3, 6 and 14, whereas no signals were detectedin the controls (Fig. 6C, D). Remarkably, at day 28, all explants wereSRY-negative (Fig. 6E). Furthermore, no signals were obtained fromthe distal and proximal 2 mm bone endings (day 28), indicating no
migration of male MSCs into the bone. Even an attempt to detectsignals by a nested PCR approach with greatly enhanced sensitivityyielded no residual signals, suggesting a loss of male cells from theregeneration tissue and newly forming bone.
In order to decide whether the implanted MSCs had alreadybeen eliminated during host cell infiltration of the fibrin–MSCcomposite or whether they persisted beside invaders, histologicalsections from day 6 were processed by laser microdissection toassess non-infiltrated and infiltrated areas separately. GenomicDNA from both areas revealed a clear SRY-gene signal after nested
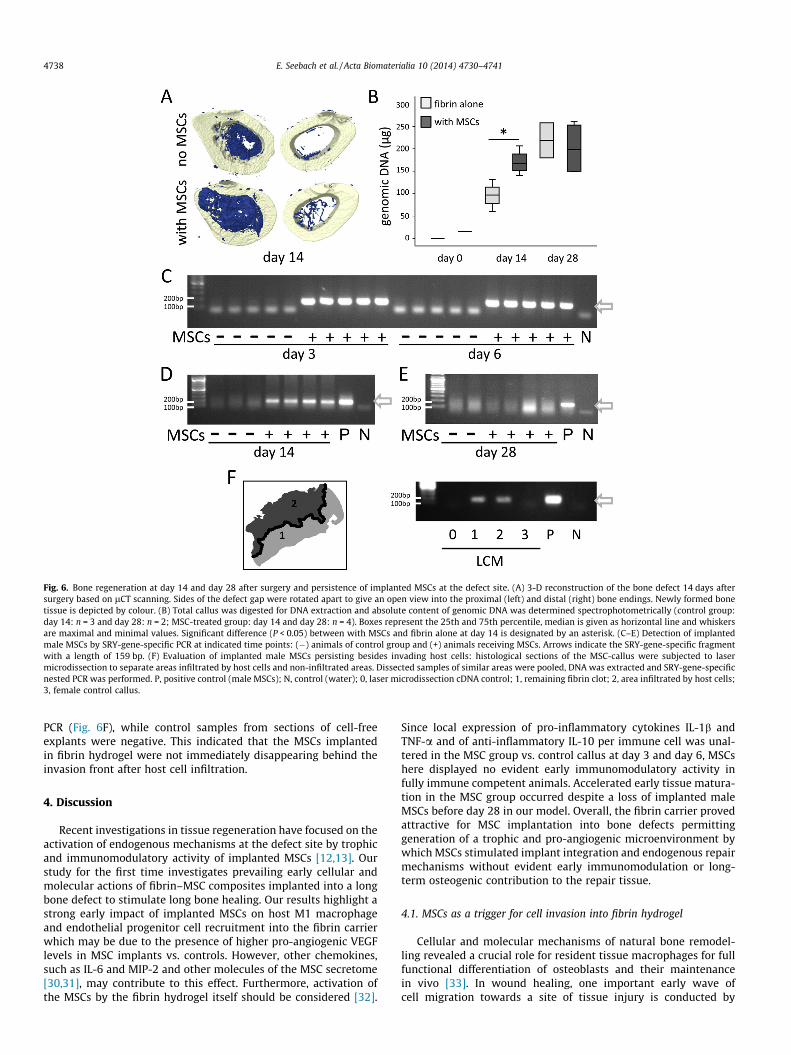
Fig. 6. Bone regeneration at day 14 and day 28 after surgery and persistence of implanted MSCs at the defect site. (A) 3-D reconstruction of the bone defect 14 days aftersurgery based on lCT scanning. Sides of the defect gap were rotated apart to give an open view into the proximal (left) and distal (right) bone endings. Newly formed bonetissue is depicted by colour. (B) Total callus was digested for DNA extraction and absolute content of genomic DNA was determined spectrophotometrically (control group:day 14: n = 3 and day 28: n = 2; MSC-treated group: day 14 and day 28: n = 4). Boxes represent the 25th and 75th percentile, median is given as horizontal line and whiskersare maximal and minimal values. Significant difference (P < 0.05) between with MSCs and fibrin alone at day 14 is designated by an asterisk. (C–E) Detection of implantedmale MSCs by SRY-gene-specific PCR at indicated time points: (�) animals of control group and (+) animals receiving MSCs. Arrows indicate the SRY-gene-specific fragmentwith a length of 159 bp. (F) Evaluation of implanted male MSCs persisting besides invading host cells: histological sections of the MSC-callus were subjected to lasermicrodissection to separate areas infiltrated by host cells and non-infiltrated areas. Dissected samples of similar areas were pooled, DNA was extracted and SRY-gene-specificnested PCR was performed. P, positive control (male MSCs); N, control (water); 0, laser microdissection cDNA control; 1, remaining fibrin clot; 2, area infiltrated by host cells;3, female control callus.
4738 E. Seebach et al. / Acta Biomaterialia 10 (2014) 4730–4741
PCR (Fig. 6F), while control samples from sections of cell-freeexplants were negative. This indicated that the MSCs implantedin fibrin hydrogel were not immediately disappearing behind theinvasion front after host cell infiltration.
4. Discussion
Recent investigations in tissue regeneration have focused on theactivation of endogenous mechanisms at the defect site by trophicand immunomodulatory activity of implanted MSCs [12,13]. Ourstudy for the first time investigates prevailing early cellular andmolecular actions of fibrin–MSC composites implanted into a longbone defect to stimulate long bone healing. Our results highlight astrong early impact of implanted MSCs on host M1 macrophageand endothelial progenitor cell recruitment into the fibrin carrierwhich may be due to the presence of higher pro-angiogenic VEGFlevels in MSC implants vs. controls. However, other chemokines,such as IL-6 and MIP-2 and other molecules of the MSC secretome[30,31], may contribute to this effect. Furthermore, activation ofthe MSCs by the fibrin hydrogel itself should be considered [32].
Since local expression of pro-inflammatory cytokines IL-1b andTNF-a and of anti-inflammatory IL-10 per immune cell was unal-tered in the MSC group vs. control callus at day 3 and day 6, MSCshere displayed no evident early immunomodulatory activity infully immune competent animals. Accelerated early tissue matura-tion in the MSC group occurred despite a loss of implanted maleMSCs before day 28 in our model. Overall, the fibrin carrier provedattractive for MSC implantation into bone defects permittinggeneration of a trophic and pro-angiogenic microenvironment bywhich MSCs stimulated implant integration and endogenous repairmechanisms without evident early immunomodulation or long-term osteogenic contribution to the repair tissue.
4.1. MSCs as a trigger for cell invasion into fibrin hydrogel
Cellular and molecular mechanisms of natural bone remodel-ling revealed a crucial role for resident tissue macrophages for fullfunctional differentiation of osteoblasts and their maintenancein vivo [33]. In wound healing, one important early wave ofcell migration towards a site of tissue injury is conducted by

E. Seebach et al. / Acta Biomaterialia 10 (2014) 4730–4741 4739
macrophages around 2–6 days after injury [34]. Macrophage-secreted cytokines affect endothelial cell proliferation, angiogene-sis and collagen synthesis [34], suggesting that they may act inconcert with other resident cells to stimulate vascularization. Ourmodel took advantage of an empty and a MSC-seeded fibrinhydrogel clot, allowing us to distinguish early arriving and possiblyfaster migrating cells penetrating deep into the implant, from laterarriving and/or later migrating cells that are still closer to theimplant edges on day 6. We consistently observed earlier host cellinfiltration from the proximal defect side which is possibly causedby a relative shortage of blood-borne invading host cells at the dis-tal defect ending after disruption of blood flow by defect creation.Remarkably, host cell infiltration was only initiated in fibrin–MSCcomposites while host cells apparently showed little interest ininvading a cell-free fibrin hydrogel in a bone defect. This demon-strated that signals from viable MSCs represented an importanttrigger for implant integration and remodelling, emphasizing theimportance of—most likely—soluble factors as trophic mediatorsof MSC action. Most importantly, although the local number ofosteoprogenitor cells in the bone cavity is high and may sufficeto initiate bone healing, defect filling with empty fibrin hydrogelalone cannot be recommended due to its rather inert persistencewithout evident cell invasion.
A large panel of trophic factors is secreted by MSCs [30,31] ofwhich IL-6, VEGF and the neutrophil-attracting factor MIP-2 werehere shown to be expressed by the fibrin-embedded MSCs beforeimplantation. This first demonstration of significantly higher VEGFmRNA levels in vivo and differential expression patterns betweenfibrin–MSC composites and cell-free controls suggest VEGF as animportant candidate molecule for endothelial cell attraction, angi-ogenesis, implant remodelling and bone regeneration in line withthe literature [35–37]. Whether the higher VEGF mRNA levelsresulted from an up-regulation of VEGF in MSCs or is contributedby attracted macrophages and endothelial cells remains to bedetermined. However, MSCs not only secrete growth factors andchemokines but also produce matrix-degrading proteases andextracellular matrix molecules which might act on angiogenesisand help in cell migration [38,39]. Overall our study, thus, demon-strates that fibrin supports MSCs in organizing a beneficial envi-ronment for bone regeneration by facilitating host macrophageinvasion relevant for tissue remodelling and enhancing endothelialprogenitor cell migration indispensible for new blood vesselformation.
4.2. The cascade of invading cells
Pioneering cells penetrating deep into the implant were mainlyCD68-positive macrophages accompanied and followed by CD31-positive endothelial progenitor cells. To our knowledge this is thefirst study demonstrating a cell-dense migration front as a domi-nant structure of in vivo integration and remodelling of a fibrin–MSC composite. This cell-dense structure developing always fromthe proximal bone ending may thus be a special feature of longbone healing or be related to the fibrin hydrogel. At ectopic sitesin immune-competent [40,41] and immune-deficient host animals[42,43] no cell-dense invasion front of host cells into tissue engi-neering constructs has been observed. This structure may so far,however, have escaped detection either because only late observa-tion time points were chosen or because no histology wasperformed at the early time points.
In many stress situations monocytes enter the damaged areaand differentiate into a spectrum of mononuclear phagocytes.These newly recruited cells usually exhibit pro-inflammatoryaction, and in the tissue inflammatory mediators such as IL-1,IL-12 and TNF-a must be balanced with the need to protect tissueintegrity [44]. Our data for the first time provide evidence that
pro-inflammatory M1 macrophages also prevailed early duringlong bone healing, while CD163-positive ‘‘anti-inflammatory’’ M2macrophages [45] remained rare at day 6. Macrophages remainresponsive to further stimuli after their initial activation, are byno means restricted to only these two categories [46], and M1 mac-rophages can convert themselves into anti-inflammatory macro-phages with an M2 wound-healing phenotype [47]. Furtherstudies at later time points are important to fully understand thecascade of invading cells and their fate during long bone healing.
4.3. Immunomodulation by fibrin-based MSC implantation
To analyze the effect of fibrin-implanted MSCs on the inflam-matory phase of bone healing, we looked for gene expression ofCD45 as well as IL-1b, TNF-a and IL-10 which represent relevantimmune mediators during early bone regeneration [48,49]. Thenatural callus adjacent to cell-free fibrin hydrogel in the controlgroup contained a higher concentration of CD45-positive immunecells than MSC-seeded implants and thus contained higher mRNAlevels of pro- and anti-inflammatory cytokines at day 3. The pres-ence of MSCs in the cell-seeded fibrin hydrogel did not only dilutethe inflammatory cytokine-producing immune cells of the earlycallus, but can also be expected to actively contribute with a broadpanel of trophic factors. These beneficial effects ultimately pro-duced a stimulatory microenvironment evident from histology,organizing remodelling of the fibrin hydrogel. Suppressive actionsof MSCs towards immune cells, as reported in other settingsin vitro and in vivo [50], were not apparent at this stage, sincethe average levels of IL-1b, TNF-a and IL-10 per CD45-positiveimmune cell were unaltered.
Although Sprague–Dawley rats have frequently been used forstudying outcome after cell and tissue transplantation betweenanimals with no evidence for strain-intern rejection [51–53], welooked for local gene expression of the T-cell mediator IL-2 as anearly rejection marker after allogeneic implantation of MSCs. IL-2mRNA levels were not increased in the MSC group at day 3 andday 6, indicating that no acute T-cell-mediated rejection of theimplanted MSCs occurred within the fibrin hydrogel within thefirst 6 days, in line with the findings in a syngeneic ectopic mousemodel [40]. Overall, this argues in favour of a microenvironment inMSC-seeded fibrin hydrogel without early silencing of hostimmune cells at days 3 and 6.
4.4. Tissue maturation and the fate of fibrin-implanted MSCs
According to detection of the implanted MSCs in our histologi-cal sections at day 6, fibrin hydrogel was stable and cell supportiveenough to enable a local persistence of implanted MSCs for at least6 days in a bone defect without connection to the vasculature. Fur-thermore, according to detection of male DNA in laser-dissectedtissue areas behind the migration front, arriving immune cellsdid not immediately degrade implanted MSCs during invasion.However, since all cells were lost within 4 weeks after implanta-tion, either their limited lifespan, a failure of re-adaptation tochanging needs during tissue maturation or other weaknessesmay have caused their ultimate replacement by host-derivedosteoprogenitor cells localized in adjacent bone marrow. In ectopicmodels a mineral component such as beta-tricalcium phosphate(b-TCP) or hydroxyapatite-tricalcium phosphate (HA/TCP) isrequired for long-term MSC engraftment and MSC-mediated boneformation [18,54]. Due to our interest in the early immunomodu-latory actions of implanted MSCs, we decided to use a fibrin carrierwithout ceramic particles since some inflammatory activity hasbeen reported for b-TCP [55,56] which would bias our results. Inour setting it remains unclear whether or not fibrin-implantedMSCs have physically contributed to the maturing osteogenic

4740 E. Seebach et al. / Acta Biomaterialia 10 (2014) 4730–4741
tissue of the MSC group until day 14 before replacement by hostbone. Although sex-mismatched animal models are well estab-lished for studying MSC-mediated bone formation [21,57] and ele-gantly permit the detection of persisting male cells in femalerecipients without a need for cell labelling [58], a long-term rejec-tion of male cells by the female immune system may have contrib-uted to our result. The transient role of fibrin-implanted MSCs seenhere is, however, in line with data from genetically engineeredASCs, which effectively stimulated femoral bone healing in rabbits,despite some induction of a humoral- and cell-mediated immuneresponse and limited cell persistence over 4 weeks [59].
5. Conclusions
In summary, by using fibrin hydrogel—known for its biocom-patibility, biodegradability and cell support for implantation ofMSCs into a long bone defect—we identified attraction of M1 mac-rophages and endothelial progenitor cells into the implant as amain early beneficial action of fibrin–MSC composites, promotingimplant integration, angiogenesis and tissue maturation duringbone regeneration. No evidence for an early immune suppressionby fibrin-implanted MSCs was obtained, as expression levels ofthe pro-inflammatory cytokines IL-1b and TNF-a as well as theanti-inflammatory cytokine IL-10 by the invading immune cellswere unaltered. Trophic and pro-angiogenic stimulation duringbone regeneration occurred independently of a long-term engraft-ment of the implanted MSCs at the defect site. Early recruitment ofhost repair cells into fibrin hydrogel was most likely stimulatedthrough the secretion of bioactive factors including VEGF whoseelevated levels may have generated a pro-angiogenic environmentbeneficial for bone regeneration. With this advanced understand-ing of the mechanisms involved in fibrin hydrogel-based MSC-sup-ported bone regeneration, we suggest fibrin hydrogel can serve asan attractive carrier for MSC-based tissue engineering approachesin long bone repair, supporting early trophic and pro-angiogenicactivities to improve clinical treatment of atrophic pseudarthrosis,bone injury or tumor resection.
Disclosure
The authors indicate no potential conflicts of interest.
Acknowledgements
We thank Svenja Schäfer and Viviana Grajales for their supportwith the surgery and with animal care. Furthermore we thankNicole Buchta and Birgit Frey for their help with the experimentsand Simone Gantz for her statistical support. This study (grant RI707/8-1) was funded by the priority program SPP1468 ‘‘Immuno-bone’’ of the German Research Foundation (DFG).
Appendix A. Figures with essential color discrimination
Certain figures in this article, particularly Figs. 1, 2 and 6 aredifficult to interpret in black and white. The full color images canbe found in the on-line version, at http://dx.doi.org/10.1016/j.actbio.2014.07.017.
References
[1] Patterson TE, Kumagai K, Griffith L, Muschler GF. Cellular strategies forenhancement of fracture repair. J Bone Joint Surg Am 2008;90(Suppl. 1):111–9.
[2] Wu X, Ren J, Li J. Fibrin glue as the cell-delivery vehicle for mesenchymalstromal cells in regenerative medicine. Cytotherapy 2012;14:555–62.
[3] Bensaid W, Triffitt JT, Blanchat C, Oudina K, Sedel L, Petite H. A biodegradablefibrin scaffold for mesenchymal stem cell transplantation. Biomaterials2003;24:2497–502.
[4] Dickhut A, Gottwald E, Steck E, Heisel C, Richter W. Chondrogenesis ofmesenchymal stem cells in gel-like biomaterials in vitro and in vivo. FrontBiosci 2008;13:4517–28.
[5] Sierra DH. Fibrin sealant adhesive systems: a review of their chemistry,material properties and clinical applications. J Biomater Appl 1993;7:309–52.
[6] Steck E, Fischer J, Lorenz H, Gotterbarm T, Jung M, Richter W. Mesenchymalstem cell differentiation in an experimental cartilage defect: restriction ofhypertrophy to bone-close neocartilage. Stem Cells Dev 2009;18:969–78.
[7] Jung M, Kaszap B, Redohl A, Steck E, Breusch S, Richter W, et al. Enhanced earlytissue regeneration after matrix-assisted autologous mesenchymal stem celltransplantation in full thickness chondral defects in a minipig model. CellTransplant 2009;18:923–32.
[8] Friedenstein AJ, Petrakova KV, Kurolesova AI, Frolova GP. Heterotopic of bonemarrow. Analysis of precursor cells for osteogenic and hematopoietic tissues.Transplantation 1968;6:230–47.
[9] Sacchetti B et al. Self-renewing osteoprogenitors in bone marrow sinusoids canorganize a hematopoietic microenvironment. Cell 2007;131:324–36.
[10] Dominici M et al. Minimal criteria for defining multipotent mesenchymalstromal cells. The International Society for Cellular Therapy positionstatement. Cytotherapy 2006;8:315–7.
[11] Pittenger MF et al. Multilineage potential of adult human mesenchymal stemcells. Science 1999;284:143–7.
[12] Meirelles Lda S, Fontes AM, Covas DT, Caplan AI. Mechanisms involved in thetherapeutic properties of mesenchymal stem cells. Cytokine Growth FactorRev 2009;20:419–27.
[13] Caplan AI, Correa D. The MSC: an injury drugstore. Cell Stem Cell 2011;9:11–5.[14] Kadiyala S, Jaiswal N, Bruder SP. Culture-expanded, bone marrow-derived
mesenchymal stem cells can regenerate a critical-sized segmental bone defect.Tissue Eng 1997;3:173–85.
[15] Bruder SP, Kraus KH, Goldberg VM, Kadiyala S. The effect of implants loadedwith autologous mesenchymal stem cells on the healing of canine segmentalbone defects. J Bone Joint Surg Am 1998;80:985–96.
[16] Quarto R et al. Repair of large bone defects with the use of autologous bonemarrow stromal cells. N Engl J Med 2001;344:385–6.
[17] Marcacci M et al. Stem cells associated with macroporous bioceramics for longbone repair: 6- to 7-year outcome of a pilot clinical study. Tissue Eng2007;13:947–55.
[18] Janicki P, Boeuf S, Steck E, Egermann M, Kasten P, Richter W. Prediction ofin vivo bone forming potency of bone marrow-derived human mesenchymalstem cells. Eur Cell Mater 2011;21:488–507.
[19] Janicki P, Kasten P, Kleinschmidt K, Luginbuehl R, Richter W. Chondrogenicpre-induction of human mesenchymal stem cells on beta-TCP: enhanced bonequality by endochondral heterotopic bone formation. Acta Biomater2010;6:3292–301.
[20] Giannoni P, Scaglione S, Daga A, Ilengo C, Cilli M, Quarto R. Short-time survivaland engraftment of bone marrow stromal cells in an ectopic model of boneregeneration. Tissue Eng Part A 2010;16:489–99.
[21] Boukhechba F et al. Fate of bone marrow stromal cells in a syngenic model ofbone formation. Tissue Eng Part A 2011;17:2267–78.
[22] Zimmermann CE, Gierloff M, Hedderich J, Acil Y, Wiltfang J, Terheyden H.Survival of transplanted rat bone marrow-derived osteogenic stem cellsin vivo. Tissue Eng Part A 2011;17:1147–56.
[23] Prockop DJ. Repair of tissues by adult stem/progenitor cells (MSCs):controversies, myths, and changing paradigms. Mol Ther 2009;17:939–46.
[24] Hughes PC, Tanner JM. The assessment of skeletal maturity in the growing rat.J Anat 1970;106:371–402.
[25] Dobson KR, Reading L, Haberey M, Marine X, Scutt A. Centrifugal isolation ofbone marrow from bone: an improved method for the recovery andquantitation of bone marrow osteoprogenitor cells from rat tibiae andfemurae. Calcif Tissue Int 1999;65:411–3.
[26] Meirelles Lda S, Nardi NB. Murine marrow-derived mesenchymal stem cell:isolation, in vitro expansion, and characterization. Br J Haematol2003;123:702–11.
[27] Winter A et al. Cartilage-like gene expression in differentiated human stemcell spheroids: a comparison of bone marrow-derived and adipose tissue-derived stromal cells. Arthritis Rheum 2003;48:418–29.
[28] Mantovani A, Sica A, Sozzani S, Allavena P, Vecchi A, Locati M. The chemokinesystem in diverse forms of macrophage activation and polarization. TrendsImmunol 2004;25:677–86.
[29] Brown BN, Valentin JE, Stewart-Akers AM, McCabe GP, Badylak SF.Macrophage phenotype and remodeling outcomes in response to biologicscaffolds with and without a cellular component. Biomaterials2009;30:1482–91.
[30] Wagner W et al. Molecular and secretory profiles of human mesenchymalstromal cells and their abilities to maintain primitive hematopoieticprogenitors. Stem Cells 2007;25:2638–47.
[31] Kinnaird T, Stabile E, Burnett MS, Lee CW, Barr S, Fuchs S, et al. Marrow-derived stromal cells express genes encoding a broad spectrum of arteriogeniccytokines and promote in vitro and in vivo arteriogenesis through paracrinemechanisms. Circ Res 2004;94:678–85.
[32] Huang NF, Chu J, Lee RJ, Li S. Biophysical and chemical effects of fibrin onmesenchymal stromal cell gene expression. Acta Biomater 2010;6:3947–56.
[33] Raggatt LJ, Partridge NC. Cellular and molecular mechanisms of boneremodeling. J Biol Chem 2010;285:25103–8.
[34] Park JE, Barbul A. Understanding the role of immune regulation in woundhealing. Am J Surg 2004;187:11S–6S.

E. Seebach et al. / Acta Biomaterialia 10 (2014) 4730–4741 4741
[35] Geiger F et al. VEGF producing bone marrow stromal cells (BMSC) enhancevascularization and resorption of a natural coral bone substitute. Bone2007;41:516–22.
[36] Kumar S, Wan C, Ramaswamy G, Clemens TL, Ponnazhagan S. Mesenchymalstem cells expressing osteogenic and angiogenic factors synergisticallyenhance bone formation in a mouse model of segmental bone defect. MolTher 2010;18:1026–34.
[37] Lin CY et al. Long-term tracking of segmental bone healing mediated bygenetically engineered adipose-derived stem cells: focuses on boneremodeling and potential side effects. Tissue Eng Part A 2014;20:1392–402.
[38] Choi YA et al. Secretome analysis of human BMSCs and identification of SMOC1as an important ECM protein in osteoblast differentiation. J Proteome Res2010;9:2946–56.
[39] Polacek M, Bruun JA, Elvenes J, Figenschau Y, Martinez I. The secretory profilesof cultured human articular chondrocytes and mesenchymal stem cells:implications for autologous cell transplantation strategies. Cell Transplant2011;20:1381–93.
[40] Tasso R et al. Recruitment of a host’s osteoprogenitor cells using exogenousmesenchymal stem cells seeded on porous ceramic. Tissue Eng Part A2009;15:2203–12.
[41] Tasso R, Fais F, Reverberi D, Tortelli F, Cancedda R. The recruitment of twoconsecutive and different waves of host stem/progenitor cells during thedevelopment of tissue-engineered bone in a murine model. Biomaterials2010;31:2121–9.
[42] Tortelli F, Tasso R, Loiacono F, Cancedda R. The development of tissue-engineered bone of different origin through endochondral andintramembranous ossification following the implantation of mesenchymalstem cells and osteoblasts in a murine model. Biomaterials 2010;31:242–9.
[43] Ghanaati S et al. Scaffold vascularization in vivo driven by primary humanosteoblasts in concert with host inflammatory cells. Biomaterials2011;32:8150–60.
[44] Murray PJ, Wynn TA. Protective and pathogenic functions of macrophagesubsets. Nat Rev Immunol 2011;11:723–37.
[45] Ambarus CA et al. Systematic validation of specific phenotypic markers forin vitro polarized human macrophages. J Immunol Methods 2012;375:196–206.
[46] Novak ML, Koh TJ. Macrophage phenotypes during tissue repair. J Leukoc Biol2013;93:875–81.
[47] Arnold L et al. Inflammatory monocytes recruited after skeletal muscle injuryswitch into antiinflammatory macrophages to support myogenesis. J Exp Med2007;204:1057–69.
[48] Mountziaris PM, Mikos AG. Modulation of the inflammatory response forenhanced bone tissue regeneration. Tissue Eng Part B Rev 2008;14:179–86.
[49] Kolar P et al. The early fracture hematoma and its potential role in fracturehealing. Tissue Eng Part B Rev 2010;16:427–34.
[50] Ghannam S, Bouffi C, Djouad F, Jorgensen C, Noel D. Immunosuppression bymesenchymal stem cells: mechanisms and clinical applications. Stem Cell ResTher 2010;1:2.
[51] Larsson J, Juliusson B, Holmdahl R, Ehinger B. MHC expression in syngeneicand allogeneic retinal cell transplants in the rat. Graefes Arch Clin ExpOphthalmol 1999;237:82–5.
[52] Troum S, Dalton Jr ML. Osteogenesis in a rat model: use of bone marrow cellsand biodegradable gelatin matrix carrier. J South Orthop Assoc 2001;10:37–43.
[53] Yang Y, Rossi FM, Putnins EE. Periodontal regeneration using engineered bonemarrow mesenchymal stromal cells. Biomaterials 2010;31:8574–82.
[54] Krebsbach PH, Kuznetsov SA, Satomura K, Emmons RV, Rowe DW, Robey PG.Bone formation in vivo: comparison of osteogenesis by transplanted mouseand human marrow stromal fibroblasts. Transplantation 1997;63:1059–69.
[55] Lange T, Schilling AF, Peters F, Haag F, Morlock MM, Rueger JM, et al.Proinflammatory and osteoclastogenic effects of beta-tricalciumphosphateand hydroxyapatite particles on human mononuclear cells in vitro.Biomaterials 2009;30:5312–8.
[56] Curran JM, Gallagher JA, Hunt JA. The inflammatory potential of biphasiccalcium phosphate granules in osteoblast/macrophage co-culture.Biomaterials 2005;26:5313–20.
[57] Nakamura A et al. Cell sheet transplantation of cultured mesenchymal stemcells enhances bone formation in a rat nonunion model. Bone 2010;46:418–24.
[58] An J, Beauchemin N, Albanese J, Abney TO, Sullivan AK. Use of a rat cDNA probespecific for the Y chromosome to detect male-derived cells. J Androl1997;18:289–93.
[59] Lin CY et al. Immune responses during healing of massive segmental femoralbone defects mediated by hybrid baculovirus-engineered ASCs. Biomaterials2012;33:7422–34.
Related Documents











