FIELD EXPERIMENTS SHOW THAT ACOUSTIC PINGERS REDUCE MARINE MAMMAL BYCATCH IN THE CALIFORNIA DRIFT GILL NET FISHERY JAY BARLOW GRANT A. CAMERON 1 Southwest Fisheries Science Center, National Marine Fisheries Service, 8604 La Jolla Shores Drive, La Jolla, California 92037, U.S.A. E-mail: [email protected] ABSTRACT A controlled experiment was carried out in 1996–1997 to determine whether acoustic deterr ent devi ces (pi nger s) reduce marine mammal bycatch in the California drift gill net fishery for swordfish and sharks. Using Fisher’s exact test, bycatc h rates with pingers were signifi cantly less for all cetacean species combined ( P , 0.001) and for all pinniped species combined ( P ¼ 0.003). For species tested separately with this test, bycatch reduction was statistically significant for short- beaked common dolphins ( P ¼ 0.001) and California sea lions ( P ¼ 0.02). Bycatch reduct ion is not statistica lly significant for the other species tested separately, but sample sizes and statis tic al power wer e low, and byc atc h rat es wer e lower in pingered nets for six of the eight other cetacean and pinniped species. A log-linear model relating the mean rate of entanglement to the number of pingers deployed was fit to the data for three gr oups : shor t- beaked common dolphins, ot her cetacea ns, and pinnipeds. For a net with 40 pingers, the models predict approximately a 12- fold decrease in entanglement for short-beaked common dolphins, a 4-fold decrease for other cetaceans, and a 3-fold decrease for pinnipeds. No other variables were found that could explain this effect. The pinger experiment ended when regulations were enacted to make pingers mandatory in this fishery. Key words: bycatch, fishery, pinger, cetacean, dolphin, pinniped, Delphinus delphis, Zalophus californianus, short-beaked common dolphin, California sea lion. Acoustic deterrent dev ices (pi ngers) reduced the bycatc h of har bor por poi se ( Phocoena phocoena) in bottom-set gill nets during controlled experiments: in the Gulf of Maine (Kraus et al. 1997), in the Bay of Fundy (Trippel et al. 1999), along the Olympic Peninsula (Gearin et al. 2000), and in the North Sea. 2 In all cases 1 Current addre ss: Scrip ps Institution of Ocea nogr aphy , UCSD, 9500 Gilman Drive, La Joll a, California 92093, U.S.A. 2 Larsen, F. 1997. Effekten af akustiske alarmer pa ˚ bifangst af marsvin i garn. Report number 44-97 (unp ublis hed) . Ava ilab le from the Dani sh Insti tute for Fish eries Resea rch, Jæge rsbor gvej 64-6 6, DK-2800 Kgs. Lyngby, Denmark. 265 MARINE MAMMAL SCIENCE, 19(2):265–283 (April 2003) 2003 by the Society for Marine Mammalogy

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 1/19
FIELD EXPERIMENTS SHOW THAT ACOUSTICPINGERS REDUCE MARINE MAMMAL BYCATCHIN THE CALIFORNIA DRIFT GILL NET FISHERY
JAY BARLOW
GRANT A. CAMERON1
Southwest Fisheries Science Center,National Marine Fisheries Service,
8604 La Jolla Shores Drive, La Jolla, California 92037, U.S.A.E-mail: [email protected]
ABSTRACT
A controlled experiment was carried out in 1996–1997 to determine whetheracoustic deterrent devices (pingers) reduce marine mammal bycatch in theCalifornia drift gill net fishery for swordfish and sharks. Using Fisher’s exact test,bycatch rates with pingers were significantly less for all cetacean species combined( P , 0.001) and for all pinniped species combined ( P ¼ 0.003). For species testedseparately with this test, bycatch reduction was statistically significant for short-
beaked common dolphins ( P ¼ 0.001) and California sea lions ( P ¼ 0.02). Bycatchreduction is not statistically significant for the other species tested separately, butsample sizes and statistical power were low, and bycatch rates were lower inpingered nets for six of the eight other cetacean and pinniped species. A log-linearmodel relating the mean rate of entanglement to the number of pingers deployedwas fit to the data for three groups: short-beaked common dolphins, other cetaceans,and pinnipeds. For a net with 40 pingers, the models predict approximately a 12-fold decrease in entanglement for short-beaked common dolphins, a 4-fold decreasefor other cetaceans, and a 3-fold decrease for pinnipeds. No other variables werefound that could explain this effect. The pinger experiment ended when regulationswere enacted to make pingers mandatory in this fishery.
Key words: bycatch, fishery, pinger, cetacean, dolphin, pinniped, Delphinus delphis,
Zalophus californianus, short-beaked common dolphin, California sea lion.
Acoustic deterrent devices (pingers) reduced the bycatch of harbor porpoise( Phocoena phocoena) in bottom-set gill nets during controlled experiments: in theGulf of Maine (Kraus et al. 1997), in the Bay of Fundy (Trippel et al. 1999), alongthe Olympic Peninsula (Gearin et al. 2000), and in the North Sea.2 In all cases
1 Current address: Scripps Institution of Oceanography, UCSD, 9500 Gilman Drive, La Jolla,
California 92093, U.S.A.2 Larsen, F. 1997. Effekten af akustiske alarmer pa bifangst af marsvin i garn. Report number 44-97
(unpublished). Available from the Danish Institute for Fisheries Research, Jægersborgvej 64-66,DK-2800 Kgs. Lyngby, Denmark.
265
MARINE MAMMAL SCIENCE, 19(2):265–283 (April 2003) 2003 by the Society for Marine Mammalogy

8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 2/19
a large (approximately 77%–90%) decrease in harbor porpoise mortality wasachieved in short-term experiments. The mechanisms are not well understood(Kraus et al. 1997), but in field trials and in captive studies, the sounds produced by
pingers appear to be aversive to harbor porpoises (Kastelein et al. 1995, 2000; Laakeet al.;3 Culik et al. 2001). Another pinger experiment was conducted in 1994 ona drift gill net fishery for swordfish along the U.S. east coast whose bycatchincluded a wide variety of cetaceans. Results of that experiment were somewhatequivocal: in paired tests pingered nets had lower bycatch, but both pingered andunpingered nets in the experiment had higher bycatch than unpingered nets in therest of the fleet.4 Prior to these recent successes, the use of active or passive acousticdeterrents showed little or no effect on net entanglement of Dall’s porpoises( Phocoenoides dalli) (Hatakeyama et al. 1994), and there was little optimism in thescientific community that such approaches would work with other species (Dawson1994, Perrin et al. 1994, Jefferson and Curry 1996). The recent success of pingers in
reducing harbor porpoise entanglements in bottom set gill nets prompted a re-evaluation of their potential to reduce mortality of other cetacean species in otherfisheries.5 In this paper we describe an experiment to evaluate the effectiveness of pingers to reduce cetacean mortality in the drift gill net fishery for swordfish andsharks along the coasts of California and Oregon.
This drift gill net fishery typically operates 37–370 km offshore from southernCalifornia to northern California and, in some years, to Oregon (Fig. 1). Theprimary season for broadbill swordfish (Xiphias gladius) is between 15 August and31 January, but some vessels fish for sharks (primarily common thresher, Alopiusvulpinas, and shortfin mako, Isurus oxyrinchus) between 15 May and 15 August.There were approximately 130 vessels actively fishing in 1995.6 Vessels aretypically 9–23 m in length, and each vessel fishes at night with one multifilamentgill net (stretched mesh size of 43–56 cm) with a maximum length of 1,830 m.Nets are suspended completely below the surface by float lines which werea minimum of 11 m in length. Previous bycatch included a wide assortment of cetacean species (Julian and Beeson 1998) including delphinids (common dolphins,Pacific white-sided dolphins, northern right whale dolphins, Risso’s dolphins, pilotwhales, bottlenose dolphins, and killer whales), beaked whales (Cuvier’s beakedwhales, Baird’s beaked whales, and Mesoplodon spp.), dwarf sperm whales, spermwhales, and humpback whales (see Table 2 for scientific names). Based on the
3 Laake, J., D. Rugh and L. Baraff. 1998. Observations of harbor porpoise in the vicinity of acoustic
alarms on a set gill net. U.S. Department of Commerce, NOAA Technical Memorandum NMFS-AFSC-
84 (unpublished). 40 pp. Available from the National Marine Mammal Laboratory, 7600 Sand Point
Way NE, Seattle, WA 98115, U.S.A.4 DeAlteris, J., E. Williams and K. Castro. 1994. Results of an experiment using acoustic devices to
reduce the incidental take of marine mammals in the swordfish drift gillnet fishery in the Northwest
Atlantic Ocean. Unpublished report. 10 pp. Available from the University of Rhode Island, Kingston,
RI 02881, U.S.A.5 Reeves, R. R., R. J. Hofman, G. K. Silber and D. Wilkinson. 1996. Acoustic deterrence of harmful
marine mammal-fishery interactions. Proceedings of a workshop held in Seattle, Washington, 20–22
March 1996. U.S. Department of Commerce, NOAA Technical Memorandum NMFS-OPR-10
(unpublished). 70 pp. Available from the NMFS Office of Protected Resources, 1335 East/West
Highway, Silver Springs, MD 20910, U.S.A.6 Barlow, J., K. A. Forney, P. S. Hill, R. L. Brownell, Jr., J. V. Carretta, D. P. DeMaster, F. Julian, M.
S. Lowry, T. Ragen and R. R. Reeves. 1997. U.S. Pacific Marine Mammal Stock Assessments: 1996.NOAA Technical Memorandum NOAA-TM-NMFS-SWFSC-248. 223 pp.
266 MARINE MAMMAL SCIENCE, VOL. 19, NO. 2, 2003

8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 3/19
management scheme used in the United States, the estimated bycatch in 1992–1996 exceeded the PBR (Potential Biological Removal) for some marine mammal
species and may not be sustainable.6
Concern about these bycatch levels promptedthe formation of the Pacific Offshore Cetacean Take Reduction Team to identifypotential solutions to this problem. The experiment described here was amongtheir first recommendations.
METHODS
Experimental Design
The experiment was designed to maximize statistical power and minimize bias.Each set was assigned randomly as either an experimental set (with pingers) or
a control set (without pingers). The experiment was carried out only on those 20%–25% of fishing trips that carried National Marine Fisheries Service bycatchobservers. Prior to a trip, observers were given packets of 10 sealed and numberedenvelopes. Prior to each set, observers would open the envelope with the numbercorresponding to the sequential set number for that trip and would read a cardwhich would indicate whether that set was to be ‘‘experimental’’ or ‘‘control.’’Randomized within each packet of ten envelopes were five cards labeled ‘‘pingers’’and five labeled ‘‘no pingers.’’ If the number of sets per trip exceeded 10, a newpacket of envelopes was used starting with set number 11. To minimize thepotential for experimental manipulation, the selection of experimental and controlsets was made after the skipper had identified a fishing location and immediately
prior to setting the net. A double-blind experimental design (such as that used byKraus et al. 1997 and Larsen2) was logistically infeasible.
Figure 1. Geographic distribution of sets with pingers (left) and without pingers (right)that were included in analyses.
267BARLOW AND CAMERON: PINGERS REDUCE BYCATCH

8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 4/19
8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 5/19
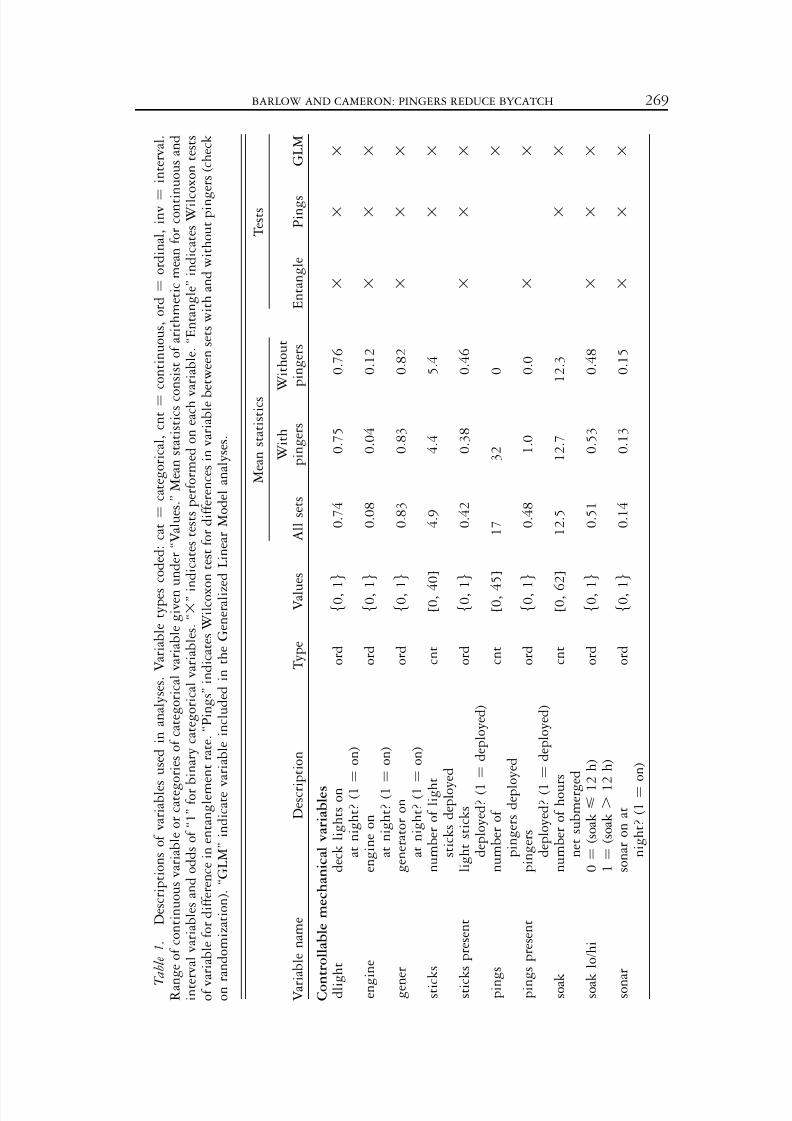
T a b l e 1 .
D e s c r i p t i o n s o f v a r i a b l e s u s e d i n a n a l y s e s . V a r i a b l e t y p e s
c o d e d : c a t ¼
c a t e g o r i c a l , c n t ¼
c o n t i n u o u s , o r d ¼
o r d i n a l , i n v ¼
i n t e r v a l .
R a n
g e o f c o n t i n u o u s v a r i a b l e o r c a t e g o r i e s o f c a t e g o r i c a l v a r i a b l e g i v e n u n d e r ‘ ‘ V a l u e s . ’ ’ M e a n s t a t i s t i c s c o
n s i s t o f a r i t h m e t i c m e a n f o r c o n t i n u o u s a n d
i n t e
r v a l v a r i a b l e s a n d o d d s o f ‘ ‘ 1 ’ ’ f o r b i n a r y c a t e g o r i c a l v a r i a b l e s . ‘ ‘ 3 ’ ’ i n d i c a t e s t e s t s p e r f o r m e d o n e a c h v a r i a b l e . ‘ ‘ E n t a n g l e ’ ’ i n d i c a t e s W i l c o x o n t e s t s
o f v a r i a b l e f o r d i f f e r e n c e i n e n t a n g l e m e n t r a t e . ‘ ‘ P i n g s ’ ’ i n d i c a t e s W i l c o x o n t e s t f o r d i f f e r e n c e s i n v a r i a b l e b e t w e e n s e t s w i t h a n d w i t h o u t p i n g
e r s ( c h e c k
o n r a n d o m i z a t i o n ) . ‘ ‘ G L M ’ ’ i n d i c a t e v
a r i a b l e i n c l u d e d i n t h e G e n e r a l i z e d L i n e a r M o d e l a n a l y s e s .
M e a n s t a t i s t i c s
T e s t s
V a r i a b l e n a m e
D e s c r i p t i o n
T y p e
V a l u
e s
A l l s e t s
W i t h
p i n g e r s
W i t h o u t
p i n g e r s
E n t a n g l e
P i n g s
G L M
C o n
t r o l l a b l e m e c h a n i c a l v a r i a b l e s
d l i g
h t
d e c k l i g h t s o n
a t n i g h t ? ( 1 ¼
o n )
o r d
f 0 , 1 g
0 . 7 4
0 . 7 5
0 . 7 6
3
3
3
e n g
i n e
e n g i n e o n
a t n i g h t ? ( 1 ¼
o n )
o r d
f 0 , 1 g
0 . 0 8
0 . 0 4
0 . 1 2
3
3
3
g e n
e r
g e n e r a t o r o n
a t n i g h t ? ( 1 ¼
o n )
o r d
f 0 , 1 g
0 . 8 3
0 . 8 3
0 . 8 2
3
3
3
s t i c k s
n u m b e r o f l i g h t
s t i c k s d e p l o y
e d
c n t
[ 0 , 4
0 ]
4 . 9
4 . 4
5 . 4
3
3
s t i c k s p r e s e n t
l i g h t s t i c k s
d e p l o y e d ? ( 1
¼
d e p l o y e d )
o r d
f 0 , 1 g
0 . 4 2
0 . 3 8
0 . 4 6
3
3
3
p i n g s
n u m b e r o f
p i n g e r s d e p l o y e d
c n t
[ 0 , 4
5 ]
1 7
3 2
0
3
p i n g s p r e s e n t
p i n g e r s
d e p l o y e d ? ( 1
¼
d e p l o y e d )
o r d
f 0 , 1 g
0 . 4 8
1 . 0
0 . 0
3
3
s o a k
n u m b e r o f h o u r s
n e t s u b m e r g e d
c n t
[ 0 , 6
2 ]
1 2 . 5
1 2 . 7
1 2 . 3
3
3
s o a k l o / h i
0 ¼
( s o a k < 1 2 h )
o r d
f 0 , 1 g
0 . 5 1
0 . 5 3
0 . 4 8
3
3
3
1 ¼
( s o a k . 1 2 h )
s o n a r
s o n a r o n a t
n i g h t ? ( 1 ¼
o n )
o r d
f 0 , 1 g
0 . 1 4
0 . 1 3
0 . 1 5
3
3
3
269BARLOW AND CAMERON: PINGERS REDUCE BYCATCH

8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 6/19
T a b l e 1
.
C o n t i n u e d .
M e a n s t a t i s t i c s
T e s t s
V a r i a b l e n a m e
D e s c r i p t
i o n
T y p e
V a l u e s
A l l s e t s
W i t h
p i n g e r s
W i t h o u t
p i n g e r s
E n t a n g l e
P i n g s
G L M
E n v
i r o n m e n t v a r i a b l e s
b c l d
c l o u d c o v e r a t s t a r
t o f s e t :
i n v
f 0 , 1 , . .
.
, 9 g
3 . 7 3
3 . 6 1
3 . 8 5
3
l i n e a r s c a l e 0 ¼
0 % ,
8 ¼
1 0 0 % , 9 ¼
t o o d a r k
b c l d
l o / h i
0 ¼
c l e a r ( b c l d ,
5 )
o r d
f 0 , 1 g
0 . 3 6
0 . 3 5
0 . 3 6
3
3
3
1 ¼
c l o u d y ( b c l d >
5 )
e c l d
c l o u d c o v e r a t e n d
o f s e t :
i n v
f 0 , . . . , 9
g
5 . 2
5 . 1
5 . 3
3
l i n e a r s c a l e 0 ¼
0 % ,
8 ¼
1 0 0 % , 9 ¼
t o o d a r k
e c l d
l o / h i
0 ¼
c l e a r ( e c l d ,
5 )
o r d
f 0 , 1 g
0 . 3 0
0 . 3 1
0 . 3 0
3
3
3
1 ¼
c l o u d y ( e c l d >
5 )
b b e a u l o / h i
B e a u f o r t s e a s t a t e
a t s t a r t o f s e t
o r d
f 0 , 1 g
0 . 4 9
0 . 4 9
0 . 4 8
3
3
3
0 ¼
c a l m ( , 3 ) ,
1 ¼
r o u g h ( > 3
)
e b e a
u l o / h i
B e a u f o r t s e a s t a t e
a t e n d o f s e t
o r d
f 0 , 1 g
0 . 4 4
0 . 4 5
0 . 4 3
3
3
3
0 ¼
c a l m ( , 3 ) ,
1 ¼
r o u g h ( > 3
)
s e a s o n
0 ¼
M a y – O c t , 1 ¼
N o v – A p r
c a t
f 0 , 1 g
0 . 5 6
0 . 5 5
0 . 5 6
3
3
3
d e p t h
W a t e r d e p t h a t t i m e
o f n e t r e t r i e v a l ( f a t h o m s )
c n t
[ 0 , 2 , 7 0
0 ]
1 , 1 5 0
1 , 1 6 7
1 , 1 3 5
3
3
d e p t h l o / h i
0 ¼
s h a l l o w ( , 1 , 0 0 0
f a t h o m s )
o r d
f 0 , 1 g
0 . 4 6
0 . 4 8
0 . 4 4
3
3
3
1 ¼
d e e p ( . 1 , 0 0 0 f a t h o m s )
N e t
v a r i a b l e s
e x t e
n d l o / h i
0 ¼
( e x t e n d , 3 7
f t )
c a t
f 0 , 1 g
0 . 2 5
0 . 2 7
0 . 2 4
3
3
3
1 ¼
( e x t e n d > 3 7
f t )
270 MARINE MAMMAL SCIENCE, VOL. 19, NO. 2, 2003

8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 7/19
T a b l e 1 .
C o n t i n u e d .
M e a n s t a t i s t i
c s
T e s t s
V a r i a b l e n a m e
D e s c r i p t i o n
T y p e
V a l u e s
A l l s e t s
W i t h
p i n g e r s
W i t h o u t
p i n g e r s
E n t a n g l e
P i n g s
G L M
e x t e n d
d i s t a n c e b e t w e e n c o r k l i n e
a n d s u r f a c e fl o a t s ( f t )
c n t
[ 1 2 , 7 8 ]
3 8 . 2
3 7 . 3
3 8 . 2
3
3
m e s h
s t r e t c h e d m e s h s i z e ( i n . )
c n t
[ 1 5 , 2 2 ]
2 0 . 3
2 0 . 4
2 0 . 3
3
3
m e s h l o / h i
0 ¼
( m e s h < 2 0 )
o r d
f 0 , 1 g
0 . 5 1
0 . 5 1
0 . 5 0
3
3
3
1 ¼
( m e s h . 2 0 )
n t c o l o r
c o l o r o f n e t
c a t
f g r e e n , r e d ,
b l u e ,
b r o w n ,
o t h e r g
2 2 2 , 2 4 , 4 ,
6 4 , 3 0
1 0 9 , 1 3 , 2 ,
3 1 , 1 1
1 1 3 , 1 1 ,
2 , 3 3 ,
1 9
3
n e t d p t h
n u m b e r o f m e s h e s , c o r k l i n e
t o l e a d l i n e
c n t
[ 3 6 , 1 , 0 5 0 ]
1 2 8 . 5
1 2 8 . 2
1 2 8 . 8
3
3
n e t l e n
l e n g t h o f n e t ( f a t h
o m s )
c n t
[ 2 2 2 , 1 , 0 0 0 ]
9 5 0 . 7
9 4 9 . 3
9 5 1 . 7
3
3
s l a c
k
p e r c e n t s l a c k : c a l c u l a t e d
f r o m n u m b e r m
e s h e s
h a n g i n g a n d h a n g i n g l e n g t h
c n t
[ 0 , 5 0 ]
4 2 . 1
4 2 . 2
4 2
3
3
L o c
a t i o n / s e a s o n v a r i a b l e s
r e g i o n
0 ¼
s o u t h o f 3 4 . 5
8 N ,
c a t
f 0 , 1 g
0 . 4 5
0 . 4 6
0 . 4 4
3
3
3
1 ¼
n o r t h o f 3 4 . 5
8 N
l a t
l a t i t u d e a t s t a r t o f
s e t
c n t
[ 3 0 , 4 7 ]
3 4 . 7 9
3 4 . 8 7
3 4 . 7 1
3
l o n g
l o n g i t u d e a t s t a r t o f s e t
c n t
[ 1 1 7 , 1 2
6 ]
1 2 0 . 6
1 2 0 . 9
1 2 0 . 4
3
271BARLOW AND CAMERON: PINGERS REDUCE BYCATCH

8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 8/19
T a b l e 1
.
C o n t i n u e d .
M e a n s t a t i s t i c s
T e s t s
V a r i a b l e n a m e
D e s c r i p
t i o n
T y p e
V a l u e
s
A l l s e t s
W i t h
p i n g e r s
W i t h o u t
p i n g e r s
E n t a n g l e
P i n g s
G L M
a r e a
F i v e fi s h i n g r e g i o n
s .
R e g i o n s 1 , 3 , 4 , 5
s e p a r a t e d b y l a t i t u d e s 3 3 . 8 3 8 ,
3 4 . 3 3 8 , a n d 4 2 . 0 0 8 N . R e g i o n 2
c o m p o s e d o f s m
a l l
d i s j o i n t a r e a s s u
r r o u n d i n g
C h a n n e l I s l a n d s .
c a t
f 1 , . . . , 5 g
3 1 5 , 1 3 ,
7 , 2 5 3 ,
2 1
1 6 1 , 1 0 , 5 , 1 2 8 ,
1 0
1 5 4 , 3 ,
2 , 1 2 5 ,
1 1
m o n
t h
m o n t h o f s e t
c a t
f 1 , . . . , 1 2 g
6 9 , 0 ,
0 , 0 ,
0 , 0 ,
0 , 5 4 ,
1 3 3 ,
1 5 7 ,
1 0 3 ,
9 3
3 6 , 0 ,
0 , 0 ,
0 , 0 ,
0 , 2 6 , 6 6 , 9 0 , 4 8 , 4 8
3 3 , 0 , 0 ,
0 , 0 ,
0 , 0 ,
2 8 , 6 7 , 6 7 , 5 5 , 4 5
3
272 MARINE MAMMAL SCIENCE, VOL. 19, NO. 2, 2003
8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 9/19
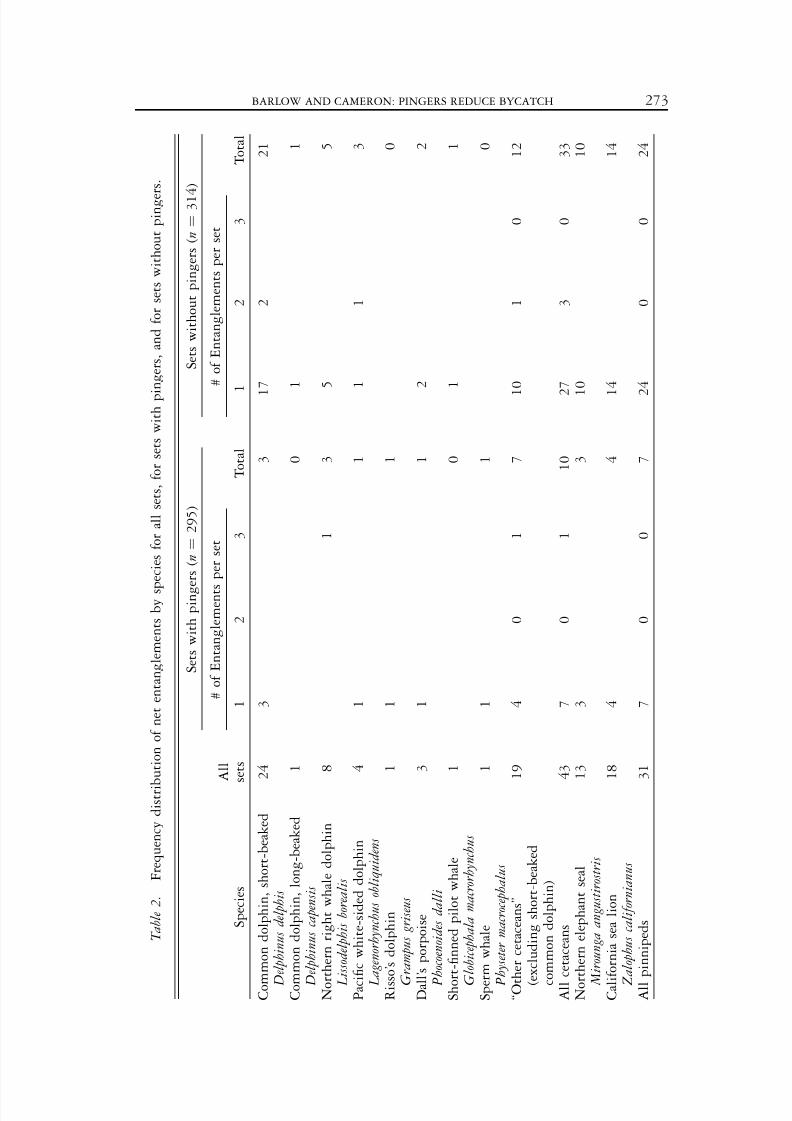
T a b l e 2 .
F r e q u e n c y d i s t r i b u t i o n o f n e t e n t a n g l e m e n t s b y s p e c i e s f o r a l l s e t s , f o r s e t s w i t h p i n g e r s , a n d f o r s e t s w i t h o u t p i n g e r s .
S e t s w i t h p i n g e r s ( n ¼
2 9 5 )
S e t s w i t h o u t p i n g e r s ( n ¼
3 1 4 )
A l l
# o f E n t a n g l e m e n t s
p e r s e t
# o f E n t a n g l e m e n t s p e r s e t
S p e c i e s
s e t s
1
2
3
T o t a l
1
2
3
T o t a l
C o m
m o n d o l p h i n , s h o r t - b e a k e d
2 4
3
3
1
7
2
2 1
D
e l p h i n u s d e l p h i s
C o m
m o n d o l p h i n , l o n g - b e a k e d
1
0
1
1
D
e l p h i n u s c a p e n s i s
N o r t h e r n r i g h t w h a l e d o l p h i n
8
1
3
5
5
L i s s o d e l p h i s b o r e a l i s
P a c i fi c w h i t e - s i d e d d o l p h i n
4
1
1
1
1
3
L a g e n o r h y n c h u s o b l i q u i d e n s
R i s s o ’ s d o l p h i n
1
1
1
0
G
r a m p u s g r i s e u s
D a l l ’ s p o r p o i s e
3
1
1
2
2
P h o c o e n o i d e s d a l l i
S h o r t - fi n n e d p i l o t w h a l e
1
0
1
1
G
l o b i c e p h a l a m a c r o r h y n c h u s
S p e r m w h a l e
1
1
1
0
P h y s e t e r m a c r o c e p h a l u s
‘ ‘ O t h e r c e t a c e a n s ’ ’
1 9
4
0
1
7
1
0
1
0
1 2
( e x c l u d i n g s h o r t - b e a k e d
c o m m o n d o l p h i n )
A l l
c e t a c e a n s
4 3
7
0
1
1 0
2
7
3
0
3 3
N o r t h e r n e l e p h a n t s e a l
1 3
3
3
1
0
1 0
M
i r o u n g a a n g u s t i r o s t r i s
C a l i f o r n i a s e a l i o n
1 8
4
4
1
4
1 4
Z
a l o p h u s c a l i f o r n i a n u s
A l l
p i n n i p e d s
3 1
7
0
0
7
2
4
0
0
2 4
273BARLOW AND CAMERON: PINGERS REDUCE BYCATCH

8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 10/19
program. Observers opportunistically recorded data on marine mammal sightingsduring the day as the vessel traveled from one location to another.
Data Selection
Experimental protocols were not followed on every set. Sometimes skippers chosenot to employ pingers in rough seas (18 cases), during the first set of a season or thefirst set with an inexperienced crew (7 cases), when pingers were causing problems (2cases), or for other reasons (20 cases). Occasionally, skippers chose to employ pingerseven when the protocol called for none (because marine mammals were known to bepresent, 5 cases). For analyses presented here, we excluded every set which did notfollow the experimental protocols. To prevent experimental manipulation of results,we also excluded all the sets from trips that were judged to be substantially out of compliance with experimental protocols (more than one-third of sets not following
protocols). Of the 713 sets that were observed during the experiment, 104 wereexcluded, resulting in 609 sets that we included in our analyses.
Statistical Analyses
Descriptions and summary statistics for variables that are likely to affect marinemammal entanglement are given in Table 1. We use abbreviated variable names(Table 1) throughout this report. Some continuous variables and categoricalvariables with multiple states were collapsed to two-state categorical variables forsome analyses; for example, the number of chemical light sticks (‘‘sticks’’) wasincluded as a continuous variable and as the categorical variable ‘‘sticks present.’’
The random distribution of net and set variables in pingered and unpingeredsets was tested using the two-sample Wilcoxon rank sum test (two-tailed). Thereduction in marine mammal bycatch when pingers were present was tested usinga one-tailed Fisher’s exact test using a 2 3 2 contingency table (no entanglementsvs. one or more entanglements per set). Reduction in the number of entanglementsper set was tested with a non-parametric Wilcoxon rank sum test (one-tailed test).The distributions of fish catch were far from Poisson or normal; therefore, thereduction in the number of target and non-target fish caught was tested only withthe Wilcoxon rank sum test (one-tailed).
Multivariate tests of the effect of pingers and other variables on marine mammalentanglement were conducted using a Generalized Linear Modelling (GLM)framework (McCullagh and Nelder 1989). A logarithmic link function was used toapproximate a Poisson error structure:
lnðE½Y iÞ ¼ b0 þX
Xij b j
where Y i is the number of entanglements for observation i, (for a species or speciesgroup); Xij is the value of predictor variable j for observation i, which may includemain effects and interaction terms; b j is the model coefficient for predictor variable j ; and b0 is the coefficient for a constant term. The error structure was actuallyallowed to vary as
varðY iÞ ¼ r2 E½Y i
where the dispersion parameter, r
2
, can be estimated from the residuals toaccommodate deviations from Poisson expectations (r2 ¼ 1.0). Maximum
274 MARINE MAMMAL SCIENCE, VOL. 19, NO. 2, 2003

8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 11/19
likelihood estimates of the coefficients, b j , were computed using iterativelyreweighted least squares using SPLUS software. According to likelihood theory,these parameters are asymptotically normal for known variance, hence, a t -test was
used to determine whether an estimated coefficient is significantly different fromzero.
Three pinger response variables (entanglements of ‘‘short-beaked commondolphin,’’ ‘‘other cetaceans,’’ and ‘‘pinnipeds’’) were modeled as linear functions of predictor variables including the number of pingers (‘‘pings’’), the number of pingers squared (‘‘pings squared’’), and each variable indicated under the ‘‘GLM’’column of Table 1. A ‘‘net volume’’ term, the product of soak time, net length,and net depth, was included by adding soak time, net length, and net depthsimultaneously in a single model. Preliminary multivariate models were built usingan approximate stepwise approach implemented in SPLUS. These models were thenpruned by sequentially removing the least significant variable until all remaining
variables were statistically significant using a test for a reduction in overall deviance(a ¼ 0.05). For Poisson-distributed entanglements, a chi-square test was used formodel selection, and for over-dispersed models, an F-test was used (McCullagh andNelder 1989).
RESULTS
Entanglements
A total of 74 marine mammals (43 cetaceans and 31 pinnipeds) was entangled inthe 609 sets during the experiment (Table 2). Short-beaked common dolphins were
the most common species and accounted for over half of the cetaceanentanglements. Pinniped entanglements included northern elephant seals (Mir-ounga angustirostris) and California sea lions (Zalophus californianus) in roughly equalnumbers. For both cetaceans and pinnipeds, entanglement rates in nets withpingers were approximately one-third the rates in nets without pingers (Table 3).
Most marine mammal entanglements consisted of single individuals; however,three northern right whale dolphins ( Lissodelphis borealis) were found entangled ina single net (with 24 pingers). The empirical distributions of the number of entanglements per set for ‘‘short-beaked common dolphins,’’ ‘‘other cetaceans,’’ and‘‘pinnipeds’’ did not differ significantly from the Poisson distribution (chi-squaregoodness of fit, a ¼ 0.05).
Possible Confounding Factors
There were no significant differences between pingered and unpingered nets forany of the variables tested except for the number of light sticks (‘‘sticks’’ and ‘‘stickspresent’’). Geographic distributions of sets showed no obvious differences betweenpingered and unpingered sets (Fig. 1). Only two variables other than the number of pingers were related to entanglement rates. Entanglement of short-beaked commondolphins was significantly related to the number of common dolphins sightings onthat trip (‘‘cdsight,’’ Wilcoxon rank sum test, P ¼ 0.0008). Entanglement of ‘‘othercetaceans’’ was not significantly related to any other variables. Entanglement of
pinnipeds was significantly related to the cloud cover at the end of the set (‘‘ecld lo/hi,’’ Wilcoxon rank sum test, P ¼ 0.04). Using a Bonferroni correction for multiple
275BARLOW AND CAMERON: PINGERS REDUCE BYCATCH

8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 12/19
T a b l e 3 .
B y c a t c h r a t e s a n d o n e - t a
i l e d s t a t i s t i c a l t e s t s o f d e c r e a s e s i n e n t a n g l e m e n t s i n s e t s w i t h p i n g e r s c o m p a r e d t o s e t s w i t h o u t p i n g e r s .
B y c a t c h r a t e s ( t o t a l b y c a t c h / t o t a l s e t s )
S t a t i s t i c a l t e s t r e s u l t s ( P - v a l u e )
S p e c i e s
S e t s w i t h p i n g e r s
S e t s w i t h o u t p i n g e r s
W i l c o x o n r a n k s u m t e s t
F i s h e r ’ s
e x a c t t e s t
C o m
m o n d o l p h i n , s h o r t - b e a k e d
0 . 0 1 0
0 . 0 6 7
0 . 0 0 1
0
. 0 0 1
D
e l p h i n u s d e l p h i s
C o m
m o n d o l p h i n , l o n g - b e a k e d
0 . 0 0 0
0 . 0 0 6
0 . 2 2 7
0
. 2 5 8
D
e l p h i n u s c a p e n s i s
N o r
t h e r n r i g h t w h a l e d o l p h i n
0 . 0 1 0
0 . 0 1 6
0 . 0 7 0
0
. 1 2 4
L i s s o d e l p h i s b o r e a l i s
P a c i fi c w h i t e - s i d e d d o l p h i n
0 . 0 0 3
0 . 0 1 0
0 . 3 1 7
0
. 3 2 9
L a g e n o r h y n c h u s o b l i q u i d e n s
R i s s o ’ s d o l p h i n
0 . 0 0 3
0 . 0 0 0
0 . 7 8 9
0
. 4 8 5
G
r a m p u s g r i s e u s
D a l l ’ s p o r p o i s e
0 . 0 0 3
0 . 0 0 6
0 . 3 1 8
0
. 3 3 0
P h o c o e n o i d e s d a l l i
S h o r t - fi n n e d p i l o t w h a l e
0 . 0 0 0
0 . 0 0 3
0 . 2 2 7
0
. 2 5 8
G
l o b i c e p h a l a m a c r o r h y n c h u s
S p e r m w h a l e
0 . 0 0 3
0 . 0 0 0
0 . 2 2 7
0
. 4 8 5
P h y s e t e r m a c r o c e p h a l u s
‘ ‘ O t h e r c e t a c e a n s ’ ’
0 . 0 2 4
0 . 0 4 1
0 . 0 8 7
0
. 1 2 7
( e x c l u d i n g s h o r t - b e a k e d
c o m m o n d o l p h i n )
A l l
c e t a c e a n s
0 . 0 3 4
0 . 1 1 0
, 0 . 0 0 1
, 0
. 0 0 1
N o r
t h e r n e l e p h a n t s e a l
0 . 0 1 0 0
0 . 0 3 2
0 . 0 3 6
0
. 0 5 6
M
i r o u n g a a n g u s t i r o s t r i s
C a l i f o r n i a s e a l i o n
0 . 0 1 4
0 . 0 4 5
0 . 0 1 3
0
. 0 2 0
Z
a l o p h u s c a l i f o r n i a n u s
A l l
p i n n i p e d s
0 . 0 2 2
0 . 0 7 6
0 . 0 0 3
0
. 0 0 3
276 MARINE MAMMAL SCIENCE, VOL. 19, NO. 2, 2003

8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 13/19
testing (a ¼ 0.05/19 ¼ 0.002), only one variable (the number of common dolphinsightings) remained significantly related to entanglements.
Pinger Effects on Entanglements of Short-beaked Common Dolphins
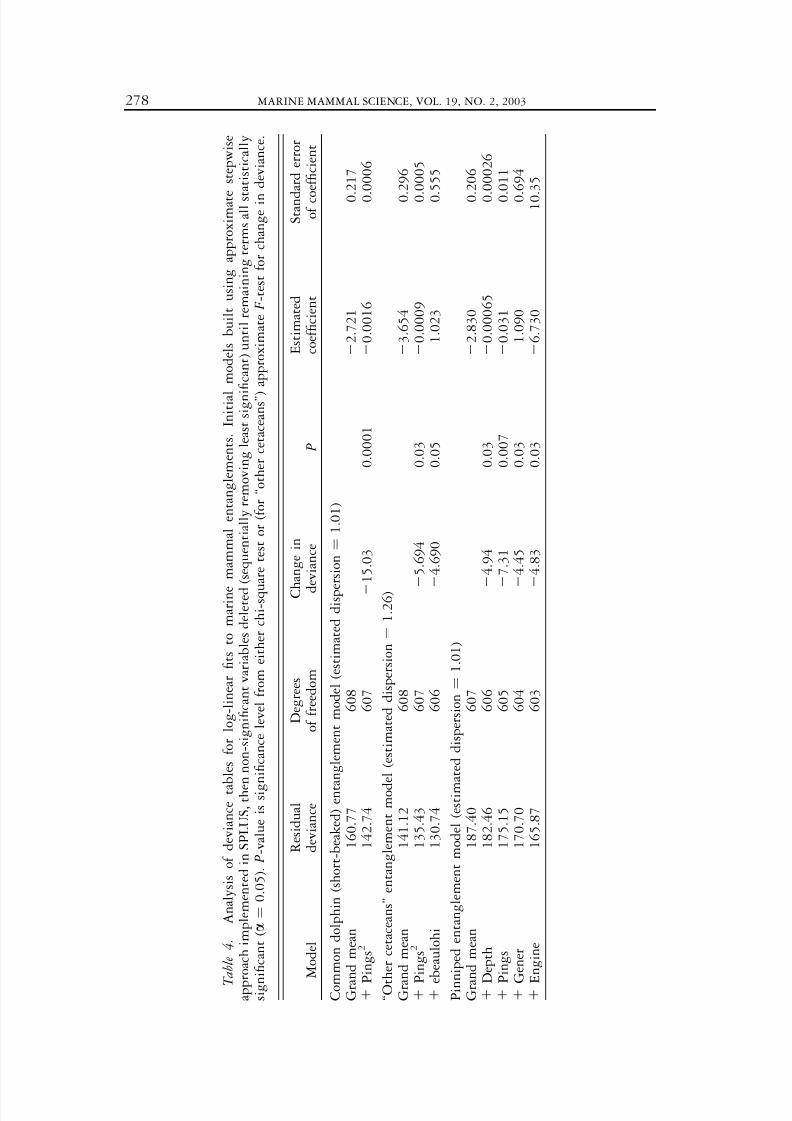
The bycatch of short-beaked common dolphins was significantly lower in netswith pingers ( P ¼ 0.001, for both the one-tailed Wilcoxon rank sum test and theFisher exact test, Table 3). The only other variable that appeared to be statisticallysignificant was the number of common dolphin sightings on a trip ( P , 0.001).The only variable selected in the stepwise log-linear model was the number of pingers squared ( P ¼ 0.0001, Table 4, Fig. 2).
Pinger Effects on Entanglements of Other Cetaceans
The bycatch of ‘‘other cetaceans’’ (other than short-beaked common dolphins)
was not significantly related to pinger use in univariate tests ( P ¼ 0.08 and P ¼0.13 using the one-tailed Wilcoxon rank sum test and the Fisher exact test,respectively) (Table 3). However, when the number of pingers used was included ina GLM model (as number of pingers squared), the pinger effect was statisticallysignificant ( P ¼ 0.03, Table 4, Fig. 3). The only other significant variable in theGLM model was the Beaufort sea state at the end of the set. Pingers were notsignificantly related to entanglement rates for any of the other species testedseparately, but sample sizes were low in all cases (only one to eight totalentanglements per species). Entanglement rates were lower in pingered nets for fiveout of the seven other cetacean species.
Pinger Effects on Entanglements of Pinnipeds
Pinniped bycatch was also significantly lower in pingered nets ( P ¼ 0.003 or0.003, one-tailed Wilcoxon rank sum test or the Fisher exact test, respectively)(Table 3). For individual species tested alone, bycatch reduction was significant forCalifornia sea lions ( P ¼ 0.01 or 0.02, respectively) and marginally significant fornorthern elephant seals ( P ¼ 0.04 or 0.06, respectively). The number of pingers(‘‘pings’’) was one of four significant variables selected in the stepwise building of a GLM model for pinniped entanglement ( P ¼ 0.007, Table 4, Fig. 4). The othersignificant variables in the GLM model were water ‘‘depth,’’ ‘‘gener,’’ and ‘‘engine.’’In univariate tests the only significant variable in explaining pinnipedentanglement was cloud cover (‘‘ecldlohi’’). This variable is not correlated withpinger use and cannot be used to explain the effect of pingers on entanglement.
Pinger Effects on Catch
There were no significant differences in the catch rates for the three target fishspecies (broadbill swordfish, common thresher shark, and shortfin mako shark)(one-tailed Wilcoxon rank sum test, Table 5). The catch rates of the non-target fishspecies were also not significantly related to pinger use (Table 5).
DISCUSSION
Pingers significantly reduced total cetacean and pinniped entanglement in driftgill nets without significantly affecting swordfish or shark catch. Results also
277BARLOW AND CAMERON: PINGERS REDUCE BYCATCH
8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 14/19
T a b l e 4 .
A n a l y s i s o f d e v i a n c e t a b l e s f o r l o g - l i n e a r fi t s t o m a r i n e
m a m m a l e n t a n g l e m e n t s . I n i t i a l
m o d e l s b u i l t u s i n g a p p r o x i m a t e
s t e p w i s e
a p p r o a c h i m p l e m e n t e d i n S P L U S , t h e n n o n - s i g n i fi c a n t v a r i a b l e s d e l e t e d
( s e q u e n t i a l l y r e m o v i n g l e a s t s i g n i fi c a n t ) u n t i l r e m a i n i n g t e r m s a l l s t
a t i s t i c a l l y
s i g n
i fi c a n t ( a ¼
0 . 0 5 ) . P - v a l u e i s s i g n i fi c a n c e l e v e l f r o m e i t h e r c h i - s q u
a r e t e s t o r ( f o r ‘ ‘ o t h e r c e t a c e a n s ’ ’ )
a p p r o x i m a t e F - t e s t f o r c h a n g e i n
d e v i a n c e .
M o d e l
R e s i d u a l
d e v i a n c e
D e g r e e s
o f f r e e d o m
C h
a n g e i n
d e v i a n c e
P
E s t i m a t e d
c o e f fi c i e n t
S t a n d
a r d e r r o r
o f c o
e f fi c i e n t
C o m
m o n d o l p h i n ( s h o r t - b e a k e d ) e n t a
n g l e m e n t m o d e l ( e s t i m a t e d d i s p e
r s i o n ¼
1 . 0 1 )
G r a n d m e a n
1 6 0 . 7 7
6 0 8
2 2 . 7 2 1
0 . 2 1 7
1 P
i n g s 2
1 4 2 . 7 4
6 0 7
2
1 5 . 0 3
0 . 0 0 0 1
2 0 . 0 0 1 6
0 . 0 0 0 6
‘ ‘ O t h e r c e t a c e a n s ’ ’ e n t a n g l e m e n t m o d e l ( e s t i m a t e d d i s p e r s i o n ¼
1 . 2 6 )
G r a n d m e a n
1 4 1 . 1 2
6 0 8
2 3 . 6 5 4
0 . 2 9 6
1 P
i n g s 2
1 3 5 . 4 3
6 0 7
2 5 . 6 9 4
0 . 0 3
2 0 . 0 0 0 9
0 . 0 0 0 5
1 e
b e a u l o h i
1 3 0 . 7 4
6 0 6
2 4 . 6 9 0
0 . 0 5
1 . 0 2 3
0 . 5 5 5
P i n n i p e d e n t a n g l e m e n t m o d e l ( e s t i m a t e d d i s p e r s i o n ¼
1 . 0 1 )
G r a n d m e a n
1 8 7 . 4 0
6 0 7
2 2 . 8 3 0
0 . 2 0 6
1 D
e p t h
1 8 2 . 4 6
6 0 6
2 4 . 9 4
0 . 0 3
2 0 . 0 0 0 6 5
0 . 0 0 0 2 6
1 P
i n g s
1 7 5 . 1 5
6 0 5
2 7 . 3 1
0 . 0 0 7
2 0 . 0 3 1
0 . 0 1 1
1 G
e n e r
1 7 0 . 7 0
6 0 4
2 4 . 4 5
0 . 0 3
1 . 0 9 0
0 . 6 9 4
1 E
n g i n e
1 6 5 . 8 7
6 0 3
2 4 . 8 3
0 . 0 3
2 6 . 7 3 0
1 0 . 3 5
278 MARINE MAMMAL SCIENCE, VOL. 19, NO. 2, 2003

8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 15/19
indicate a greater reduction with a greater number of pingers. These results aresimilar to results of previous experiments that showed a significant reduction inharbor porpoise bycatch when pingers were used on set gill nets (Kraus et al. 1997,Larsen2, Trippel 1999, Gearin et al. 2000). Our experiment is, however, the firstunequivocal demonstration that pingers are correlated with a significant reductionin the bycatch for a delphinid cetacean (short-beaked common dolphin) and fora pinniped (California sea lion). The significant reduction in total cetacean bycatchwas largely driven by the reduction in bycatch of short-beaked common dolphins.Bycatch reduction was not statistically significant for any other cetacean species(although, bycatch was lower for most). An impractically large sample would berequired to find a statistically significant result for rare species, even if their
response was the same as for common dolphins.Because of the potential importance of these results in reducing marine mammal
bycatch worldwide, it is important to investigate potential spurious causes of thesepatterns. One potential concern is the lack of a true double-blind control in ourexperimental protocol. We cannot tell whether the observed pinger effect wascaused by the sound produced by the pingers or by the presence of something novelhanging from the net. We believe that the visual enhancement caused by thepresence of the pingers at night is trivial and that the sounds they emit almostcertainly caused the reduction in bycatch; however, our design does not allow us todistinguish between these hypotheses. A more serious concern is the possible director inadvertent manipulation of the results by the observers or the fishermen. The
observers had no direct role in the design or analysis of the experiment and wouldnot directly benefit by manipulating the results (other than the common human
Figure 2. Predicted bycatch per set of short-beaked common dolphins as function of number of pingers based on GLM. Dotted lines show approximate 95% confidence intervals.
279BARLOW AND CAMERON: PINGERS REDUCE BYCATCH

8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 16/19
desire for successful outcomes). Fishermen knew that their industry was undergrowing scrutiny and that, if bycatch were not reduced, they might face additionalregulations or even closure; therefore, fishermen had a strong incentive to show thatpingers worked. The ability for fishermen to manipulate results was limited becausethe fishermen had already chosen a location before a set was determined to be‘‘pingered’’ or ‘‘unpingered.’’ Sets were eliminated from analysis when this protocolwas not followed. Once a net is set in a given location, there is little that a fishermancan do to affect marine mammal bycatch. Of the variables that are under a captain’scontrol (‘‘dlight,’’ ‘‘engine,’’ ‘‘gener,’’ ‘‘sticks,’’ ‘‘soak,’’ and ‘‘sonar’’), only ‘‘sticks’’ was
significantly correlated with pinger use, and none were significantly correlated withcetacean bycatch. The effect of pingers on bycatch was greater than the effects of anyother variables (except number of common dolphin sightings), and it would beimpossible to contrive such a strong pinger effect by subtle experimentalmanipulations. Additional analyses (including classification and regression trees,CART) were conducted to look for other variables that might explain patterns of entanglements,12 and pingers also emerged as an important explanatory variable inthose studies.
Figure 3. Predicted bycatch per set of ‘‘other cetaceans’’ (other than short-beakedcommon dolphins) as function of number of pingers based on GLM. Dotted lines showapproximate 95% confidence intervals.
12 Cameron, G. 1999. Report on the effect of acoustic warning devices (pingers) on cetacean and
pinniped bycatch in the California drift gillnet fishery. Administrative Report LJ-99-08C
(unpublished). 71 pp. Available from the Southwest Fisheries Science Center, 8604 La Jolla ShoresDrive, La Jolla, CA 92037, U.S.A.
280 MARINE MAMMAL SCIENCE, VOL. 19, NO. 2, 2003

8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 17/19
T a b l e 5 .
C a t c h ( n u m b e r
o f fi s h ) a n d o n e - t a i l e d s t a t i s t i c a l
t e s t s f o r d e c r e a s e s i n c a t c h r a t e s f o r s e t s w i t h a n d w i t h o u t p i n g e r s .
T o t a l c a t c h
S e t s w i t h
p i n g e r s
S e t s w i t h
o u t p i n g e r s
W i l c o x o n
r a n k s u m
S p e c i e s
( # o f fi s h )
C a t c h
C a t c h / s e t
C a t c h
C a t c h / s e t
P - v a l u e
T a r g e t
S
w o r d fi s h , b r o a d b i l l
1 , 0 7 5
5 1 3
1 . 7 4
5 6 2
1 . 7 9
0 . 4 6
X i p h i a s g l a d i u s
S
h a r k , C o m m o n t h r e s h e r
4 6 2
1 7 0
0 . 5 8
2 9 2
0 . 9 3
0 . 2 4
A l o p i u s v u l p i n a s
S
h a r k , m a k o
8 1 5
4 1 8
1 . 4 2
3 9 7
1 . 2 6
0 . 5 3
I s u r u s o x y r i n c h u s
N o n - t a r g e t
M
o l a , c o m m o n
2 , 1 6 2
1 , 0 1 2
3 . 4 3
1 , 1 5 0
3 . 6 6
0 . 4 3
M o l a m o l a
O
p a h
6 0 7
3 0 6
1 . 0 4
3 0 1
0 . 9 6
0 . 3 0
L a m p r i s g u t t a t u s
S
h a r k , b i g e y e t h r e s h e r
6 9
2 5
0 . 0 9
4 4
0 . 1 4
0 . 3 2
A l o p i u s s u p e r s i l i o s u s
S
h a r k , b l u e
2 , 1 1 9
1 , 0 6 6
3 . 6 1
1 , 0 5 3
3 . 3 5
0 . 7 1
P r i o n a c e g l a u c a
T
u n a , a l b a c o r e
1 , 1 1 7
6 9 6
2 . 3 6
4 2 1
1 . 3 4
0 . 4 6
T h u n n u s a l a l u n g a
T
u n a , b l u e fi n
5 7 2
2 9 5
1 . 0 0
2 7 7
0 . 8 8
0 . 3 7
T h u n n u s t h y n n u s
T
u n a , s k i p j a c k
5 8 0
2 7 4
0 . 9 3
3 0 6
0 . 9 7
0 . 3 2
K a t s u w o n u s p e l a m i s
281BARLOW AND CAMERON: PINGERS REDUCE BYCATCH

8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 18/19
Additional work is needed to determine the optimal number and placement of pingers on drift gill nets. Log-linear models indicate that mortality rate is stilldecreasing with number of pingers within the range of 30–40 pingers (Fig. 2–4);however, there were few data during this experiment within the range of 1–20pingers, so there is considerable uncertainty about the shape of this response curvein that region. The GLM model identified Beaufort sea state, engine noise, andgenerator noise as possible explanatory variables in some analyses. All threevariables are sources of noise that might mask the sounds produced by pingers;however, engine and generator noise could also act to alert animals to the presenceof the net. Water depth is another explanatory variable for pinnipeds; this might beexpected because California sea lions forage only in the shallower, inshore portion of the operational range of drift gill net vessels.
The reduction we see in pinniped entanglements is particularly surprising becauseothers have predicted that pinnipeds might be attracted to nets to feed on thecaptured fish (the ‘‘dinner bell’’ effect). However, in an experimental study of theresponse of captive California sea lions to pingers, Anderson (2000) showed that theyinitially responded with a start followed by avoidance (five of six sea lions left thewater). This response helps explain the reduction we noted in sea lion entanglements.
Although pingers appear to reduce bycatch for a large range of marine mammalspecies, we echo the concerns that have been expressed by many other authors thatanimals may habituate to pingers. Given the relatively small number of nets andthe huge area fished, habituation may be less of a concern for the California driftgill net fishery than for intensive, localized set gill net fisheries in the Gulf of Maine
and in the North Sea. We believe that pingers are unlikely to reduce the bycatch of all cetacean species or all pinniped species.
Figure 4. Predicted bycatch per set of pinnipeds as function of number of pingers basedon GLM. Dotted lines show approximate 95% confidence intervals.
282 MARINE MAMMAL SCIENCE, VOL. 19, NO. 2, 2003

8/11/2019 0912f50a6962c1327c000000
http://slidepdf.com/reader/full/0912f50a6962c1327c000000 19/19
ACKNOWLEDGMENTS
We thank Tim Price and the hard-working fishery observers for collecting these data. Wethank the California drift gill net fishermen who have become actively involved in theprocess of improving their fishery. We are grateful to Rand Rasmussen for editing andmaintaining a reliable data base and Peter Perkins, Fred Julian and Cleridy Lennert for freelysharing their statistical expertise. Jim Carretta helped prepare Figure 1. Finally we wouldlike to thank all others who planned, funded, and helped carry out the pinger experimentwithin the California drift gill net monitoring program.
LITERATURE CITED
ANDERSON, R. C. 2000. Responses of captive California sea lions (Zalophus californianus) tonovel stimuli and the effects of motivational state. Master’s thesis, University of SanDiego, San Diego, CA. 192 pp.
CULIK, B. M., S. KOSCHINSKI, N. TREGENZA AND G. M. ELLIS. 2001. Reactions of harborporpoises Phocoena phocoena and herring Clupea harengus to acoustic alarms. MarineEcology Progress Series 211:255–260.
DAWSON, S. M. 1994. The potential for reducing entanglement of dolphins and porpoiseswith acoustic modifications to gillnets. Reports of the International WhalingCommission (Special Isssue 15):573–578.
GEARIN, P. J., M. E. GOSHO, J. L. LAAKE, L. COOKE, R. DELONG AND K. M. HUGHES. 2000.Experimental testing of acoustic alarms (pingers) to reduce bycatch of harbourporpoise, Phocoena phocoena, in the state of Washington. Journal Cetacean Research andManagement 2:1–9.
HATAKEYAMA, Y., K. ISHII, T. AKAMATSU, H. SOEDA, T. SHIMAMURA AND T. KOJIMA. 1994. Areview of studies on attempts to reduce the entanglement of the Dall’s porpoise,
Phocoenoides dalli, in the Japanese salmon gillnet fishery. Reports of the InternationalWhaling Commission (Special Isssue 15):549–563.
JEFFERSON, T. A., AND B. E. CURRY . 1996. Acoustic methods of reducing or eliminatingmarine mammal-fishery interactions: do they work? Ocean and Coastal Management31:41–70.
JULIAN, F., AND M. BEESON. 1998. Estimates of marine mammal, turtle, and seabirdmortality for two California gillnet fisheries: 1990–95. Fishery Bulletin, U.S. 96:271–284.
KASTELEIN, R. A., D. DE HAAN, C. STAAL, S. H. NIEUWSTRATEN AND W. C. VERBOOM. 1995.Entanglement of harbour porpoises ( Phocoena phocoena) in fishing nets. Pages 91–156 inP. E. Nachtigall, J. Lien, W. W. L. Au and A. J. Read, eds. Harbour porpoises—laboratory studies to reduce bycatch. De Spil Publishers, Woerden, The Netherlands.
KASTELEIN, R. A., H. T. RIPPE, N. VAUGHAN, N. M. SCHOONEMAN, W. C. VERBOOM AND
D. DE HAAN. 2000. The effects of acoustic alarms on the behavior of harbor porpoises( Phocoena phocoena) in a floating pen. Marine Mammal Science 16:46–64.
KRAUS, S . , A . J . READ, A . SOLOW, K . BALDWIN, T. SPRADLIN, E . ANDERSON AND
J. WILLIAMSON. 1997. Acoustic alarms reduce porpoise mortality. Nature 388:525.MCCULLAGH, P., AND J. A. NELDER. 1989. Generalized linear models. Chapman and Hall,
New York, NY.PERRIN, W. F., G. P. DONOVAN AND J. BARLOW, EDS. 1994. Gillnets and cetaceans.Reports of
the International Whaling Commission Special Issue 15.TRIPPEL, E. A., M. B. STRONG, J. M. TERHUNE AND J. D. CONWAY . 1999. Mitigation of
harbour porpoise ( Phocoena phocoena) by-catch in the gillnet fishery in the lower Bayof Fundy. Canadian Journal of Fisheries and Aquatic Science 56:113–123.
Received: 13 February 2002
Accepted: 1 August 2002
283BARLOW AND CAMERON: PINGERS REDUCE BYCATCH