Pergamon Psychoneuroendocrinology, Vol. 21, No. 2, pp. 173-187, 1996 Copyright © 1996 Elsevier Science Ltd. All rights reserved Printed in Great Britain 0306-4530/96 $15.00 + .00 0306-4530(95)00040-2 DYNAMICAL ENTROPY IS CONSERVED DURING COCAINE- INDUCED CHANGES IN FETAL RAT MOTOR PAT-FERNS William P. Smotherman, 1 Karen A. Selz2 and Arnold J. Mandell3 ILaboratory of Perinatal Neuroethology, Center for Developmental Psychobiology, Department of Psychology, Binghamton University, Binghamton, NY 13902-6000, USA; 2Clinical Counseling Center and Department of Mathematics, Florida Atlantic University, Boca Raton, FL 33431-0991, USA; and 3Departments of Mathematics, Physics and Psychology, Florida Atlantic University, Boca Raton, FL 33431-0991, USA SUMMARY Our previous studies demonstrated that the intra-cisternal (IC) administration of cocaine to fetal rats increased motor activity and decreased responsiveness to perioral stimulation. One explanation for these observations comes from the behavioral pharmacology of stimulant drugs: increased motor activity is often associated with a decrease in its variety. Previous power spectral transformation of this data suggests an alternative explanation: cocaine-induced hyperactivity fixates a new behavioral pattern with complexity equal to that of saline controls. We explore these possibilities using statistical techniques derived from studies of nonlinear dynamical systems, examining patterns of the total motor activity of the individual fetus as counts per 5 s interval on either gestational day E20 or E21 for 20 min following IC injections of saline, 2.5 or 10 mg/kg of cocaine. The results are consistent with a state in which increased spontaneous activity is associated with the emergence of a new dynamical pattern which conserves entropy and provides experimental support for a fundamental conservation-variational relation, hr ~ ~l × Dn, that has been proven for abstract models of chaotic dynamical systems. A multivariate analysis of variance (MANOVA) followed by appropriate analyses of variance (ANOVAs) and pairwise comparisons revealed that, whereas cocaine induced increases in the total amount of motor activity, the rate of increase in the variety of new sequences in activity counts over time did not change with treatment and age conditions. This invariant is quantified by an absence of change in topological entropy, Ahr=0. The analyses also showed that, in order to maintain hr values, compensatory changes took place in the leading Lyapounov characteristic exponent, ;'-1 (the distance between sequential values 'stretched' along the increasing amplitudes of the variations) such that A21 > 0, and the correlation dimension, DR (the hierarchical range of possible values, 'complicated clustering') was reduced, so that ADR < 0. Our findings are consistent with the idea that the association between cocaine-induced increases in activity and decreases in adaptive response are not due to the dynamical entropy loss of decreased behavioral variety, but are rather the result of competitive interference by a drug-induced, equally complex, new pattern of spontaneous behavior. Copyright © 1996 Elsevier Science Ltd. Keywords--Cocaine; Fetal behavior; Entropy; Lyapounov exponent; Fractal dimension; Chaos. INTRODUCTION Maternal use of cocaine during pregnancy has been correlated with adverse effects on development in the human infant. Deficits in motor, sensory and cognitive performance have Address correspondence and reprint requests to: William P. Smotherman, Laboratory of Perinatal Neuroethology, Center for Developmental Psychobiology, Department of Psychology, Binghamton University, Binghamton, NY 13902-6000, USA. 173

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Pergamon Psychoneuroendocrinology, Vol. 21, No. 2, pp. 173-187, 1996
Copyright © 1996 Elsevier Science Ltd. All rights reserved Printed in Great Britain
0306-4530/96 $15.00 + .00
0306-4530(95)00040-2
D Y N A M I C A L E N T R O P Y IS C O N S E R V E D D U R I N G C O C A I N E -
I N D U C E D C H A N G E S IN F E T A L R A T M O T O R PAT-FERNS
William P. Smotherman, 1 Karen A. Selz 2 and Arnold J. Mandell 3 I Laboratory of Perinatal Neuroethology, Center for Developmental Psychobiology, Department of Psychology, Binghamton University, Binghamton, NY 13902-6000, USA; 2Clinical Counseling
Center and Department of Mathematics, Florida Atlantic University, Boca Raton, FL 33431-0991, USA; and 3Departments of Mathematics, Physics and Psychology, Florida Atlantic University,
Boca Raton, FL 33431-0991, USA
SUMMARY
Our previous studies demonstrated that the intra-cisternal (IC) administration of cocaine to fetal rats increased motor activity and decreased responsiveness to perioral stimulation. One explanation for these observations comes from the behavioral pharmacology of stimulant drugs: increased motor activity is often associated with a decrease in its variety. Previous power spectral transformation of this data suggests an alternative explanation: cocaine-induced hyperactivity fixates a new behavioral pattern with complexity equal to that of saline controls. We explore these possibilities using statistical techniques derived from studies of nonlinear dynamical systems, examining patterns of the total motor activity of the individual fetus as counts per 5 s interval on either gestational day E20 or E21 for 20 min following IC injections of saline, 2.5 or 10 mg/kg of cocaine. The results are consistent with a state in which increased spontaneous activity is associated with the emergence of a new dynamical pattern which conserves entropy and provides experimental support for a fundamental conservation-variational relation, hr ~ ~l × Dn, that has been proven for abstract models of chaotic dynamical systems. A multivariate analysis of variance (MANOVA) followed by appropriate analyses of variance (ANOVAs) and pairwise comparisons revealed that, whereas cocaine induced increases in the total amount of motor activity, the rate of increase in the variety of new sequences in activity counts over time did not change with treatment and age conditions. This invariant is quantified by an absence of change in topological entropy, Ahr=0. The analyses also showed that, in order to maintain hr values, compensatory changes took place in the leading Lyapounov characteristic exponent, ;'-1 (the distance between sequential values 'stretched' along the increasing amplitudes of the variations) such that A21 > 0, and the correlation dimension, DR (the hierarchical range of possible values, 'complicated clustering') was reduced, so that ADR < 0. Our findings are consistent with the idea that the association between cocaine-induced increases in activity and decreases in adaptive response are not due to the dynamical entropy loss of decreased behavioral variety, but are rather the result of competitive interference by a drug-induced, equally complex, new pattern of spontaneous behavior. Copyright © 1996 Elsevier Science Ltd.
Keywords--Cocaine; Fetal behavior; Entropy; Lyapounov exponent; Fractal dimension; Chaos.
INTRODUCTION
Maternal use of cocaine during pregnancy has been correlated with adverse effects on development in the human infant. Deficits in motor, sensory and cognitive performance have
Address correspondence and reprint requests to: Will iam P. Smotherman, Laboratory of Perinatal Neuroethology, Center for Developmental Psychobiology, Department of Psychology, Binghamton University, Binghamton, NY 13902-6000, USA.
173

174 W.P. Smotherman et al.
been reported in human neonates exposed to cocaine in utero (Dow-Edwards, 1991; Gingras et al., 1992; Singer et al., 1991, 1992; Spear, 1995). By what structural and/or dynamical mechanisms prenatal exposure to cocaine influences fetal development remains an open question. Our investigations examined the effects of cocaine in an experimental animal model that permits drug administration to individual fetal subjects followed by quantitative observation of their motor behaviors over time (Simonik et al., 1993, 1994). These studies have shown that cocaine administered into the peripheral circulation or directly into the central nervous system of individual fetal rats produces three- to four-fold increases in motor activity in the forelimbs and hindlimbs lasting several minutes. Simonik et al.'s power spectral analyses of these data also suggested an increase in the prominence of fast frequency modes. Studies also have found that cocaine reduced responsiveness to cutaneous stimulation with an artificial nipple and disrupted coordination in upper limb aversive responses (e.g., facial wiping movements) to punctate perioral stimulation (Simonik et al., 1993, 1994). These findings are consistent with many studies of the patterns of spontaneous behavior in response to increasing doses of stimulant drugs, which indicate that an increase in the amount of activity is associated with a decrease in behavioral variety (Lyons & Robbins, 1975).
Our recent adaptations of invariant statistical 'ergodic' measures have been useful in the characterization of the phase space behavior of chaotic dynamical systems as represented by temporal-spatial motor activity in animals and man. These techniques have added new subtlety and power with which to examine the nature of drug-induced changes in the time- dependent aspects of these observables (Mandell, 1983; Mandell et al., 1982; Paulus et al., 1990; Selz, 1992; Selz & Mandell, 1993; Selz et al., in press).
Because normative patterns of spontaneous motor activity by the fetus appear to be essential for healthy neuroanatomical and behavioral development (Moessinger, 1988; Oppenheim, 1982; Smotherman & Robinson, 1987, 1988), the drug-induced combination of increased time-dependent limb activity associated with decreased and disordered responsiveness to oral stimulation constitute an interesting combination of statistical dynamical changes in motor activity (Simonik et al., 1993, 1994; Smotherman & Robinson, 1987, 1988). The counter-intuitive findings that increased general motor activity in the forelimbs was apparently less available for use in at least some kinds of adaptive responding, suggested that countervailing influences were invoked, of the sort that may occur in response to the perturbation of dynamical systems when an underlying property is being conserved. It has been suggested that this conserved property in signal sensitive biological systems, such as the brain and heart, is dynamical complexity, the rate of generation of new possibilities with time (Mandell, 1983, 1987; Mandell et al., 1982; Selz, 1992).
Onsager's classical models of biologically relevant, 'irreversible' statistical mechanical systems demonstrated relationships between patterns of spontaneous activity and responses to perturbation (Kittel, 1958). This suggests that there may be indications in the statistical properties of the cocaine-induced changes in spontaneous motor activity of the fetuses that auger the drug-induced loss of responsiveness to perioral stimulation. In the analyses presented here, we explore this possibility using measures developed for the statistical study of time-dependent complexity in nonlinear dynamical systems (e.g., Bowen, 1978; Smale, 1981).
An emphasis on the time-dependence of motor activity over development is natural because one of the most experimentally accessible and distinctive features of fetal behaviors is their temporal structure. Fetal rats (Smotherman et al., 1988), sheep (Robertson & Bacher,

Conservation of Dynamical Entropy 175
1995) and humans (Robertson, 1985) exhibit patterns of recurrent motor activity with a broadly distributed power spectral peak in the 1-min range. Autonomously recurrent motor activity patterns of this sort are robust and persist for several months into the postnatal period in humans (Robertson, 1987). It has been suggested that these patterns reflect a fundamental temporal property of the developing nervous system (Aschoff, 1981; Corner, 1977; Robertson, 1988). It has been demonstrated that they can be altered by changes in maternal physiology as well as the fetal environment. For example, maternal diabetes may disrupt the normal dynamical patterns in fetal movement by flattening the record, making the pattern more irregular or shifting the dominant broad-band frequency (Robertson & Dierker, 1986). These studies, along with other evidence that intra-uterine physiological changes can influence the time-dependent patterns of fetal motor activity, motivated our studies of their dynamical characteristics as influenced by prenatal exposure to cocaine.
Most research in neuropsychobiological behavioral dynamics has, as a first approxima- tion, treated phenomena such as these recurrent fetal motoric patterns with broadly distributed, average frequencies as 'cycles' (e.g., Robertson, 1988). Under the impetus of modern dynamical systems theory, a more detailed examination of brain and behavioral patterns has shown them to be more complicated and subtle than simple periodic behavior, and more rich with respect to their potential for encoding and transport of information (Mandell et al., 1982; Mandell & Selz, 1993a, b; Paulus et al., 1990; Selz & Mandell, 1991, 1992).
This new view suggests that the averaged regularity manifested by patterns of recurrence that are often regarded as 'periods'--as in our motoric observables--may be analogous to frequency or amplitude-modulated 'carrier waves', with their information-bearing function reflected in the fine structure that results in the broadening of the power spectral peaks. Although the amount of motor activity may increase and decrease in time in a roughly regular manner, a closer examination reveals that these patterns of recurrence possess addi- tional dynamical properties that are of particular relevance to development. The combination of fluctuations around an only roughly cyclic pattern, and the nonlinear dependence on the current value of the observable in any nonlinear mechanism with respect to its production of the next one results in the continual generation of new durations (reflected also in the number of events in a unit of observation) and return times of the observable. One can easily imagine that psychobiological processes might benefit by this continuing generation of new temporal possibilities, dynamical complexity, during fetal development.
We transform data series into geometric objects on a plane by plotting their value at one time against their value at some previous time or in a three dimensional box by adding the dimension of an additional previous time. When this is done, we see that nearby points in these 'phase spaces' often separate exponentially in time and then, because the values in psychobiological systems are bounded, fold back into the global recurrent pattern in an unpredictable way. Taken together, the stretching and folding of temporal distances between individual points in a recurrent process result in the specific sequence of points getting out of order (in a process called mixing), even though they follow recurrent pathways. Instead of autocorrelation functions being infinite as they are in strictly periodic motion, those associated with real psychobiological data streams that mix, decay with a variety of experimental condition-dependent rates.
This mixing process and its associated 'exponential or power law decay of correlations', following Fourier transformation, broadens the amplitude peaks of time series and their

176 W.P. Smotherman et al.
power spectral representations as 'broad-band cycles' (Ruelle, 1989; Shlesinger, 1989). However, these broad prominences in the power spectra are not decomposable into cycles.
In these experiments, cocaine was administered in a single dose into the central nervous systems of individual fetal rats. The effects of different dosages of cocaine on fetal motor behavior were examined on the last 2 days of gestation (i.e., E20 and E21) over 20 min, with motor activity summed and then partitioned into 5-s bins. Three measures made on the sequence of observables derived from the studies of abstract stretching, folding and mixing processes serve to discriminate between motoric patterns under changing experimental conditions. We used these measures computed on the time series of motor activity of the individual fetal subjects in order to evaluate time-dependent behavioral changes associated with low (2.5 mg/kg) and high (10 mg/kg) cocaine exposure compared with the saline treated, SAL, group over a small range, 2 days, of developmental time (i.e., on E20 and E21). The size of the experimental sample, as well as the precision and reliability of the experimental procedures, matched genetic and litter developmental controls and observable counts made possible our cross-sectional statistical approaches to relatively short sample lengths, using measures that usually require individual time series of psychobiologically unrealistic lengths for asymptotic stability.
We found that a dimensionless variational relation, hT ~ 2~ x DR, exists between these dependent measures of the system's statistical dynamics. These measures, the topological entropy, characteristic (leading Lyapounov) exponent and correlation dimension, are described below in some detail. A remarkable aspect of these findings is that this relation has required entirely unrealistic conditions, such as uniform expansivity, in order to be proven as a mathematical theorem, true for maximally chaotic abstract dynamical systems (Manning, 1981; Ledrappier & Young, 1985). That this 'law' could also be found to apply in real psychobiological systems was unexpected.
This relationship between measures suggests an explanation for the results of Smotherman & Robinson (1987, 1988) and Simonik et al. (1993, 1994). It might explain how it is that in the cocaine-induced dynamical condition, increased amounts of spontaneous activity can be associated with decreased responsiveness to perioral perturbation, and do so without invoking ideas associating the loss of dynamical complexity with stimulant-drug induced reduction in motoric variety.
METHOD
Subjects A total of 48 rat fetuses from 16 litters served as subjects on day E20 and a total of 42
fetuses from 20 litters served as subjects on gestational day E21. Fetal subjects were the offspring of time-mated Sprague-Dawley rats (Charles River Laboratories, Wilmington, MA, USA) bred in the Laboratory of Perinatal Psychobiology at Binghamton University - SUNY. Female rats were housed in groups of three in plastic breeding cages (36 × 47 x 20 cm) with one male during a 4-day breeding period. Daily vaginal smears were taken to date pregnancies, with the detection of sperm designated as day 0 (i.e., E0) of gestation. Pregnant females were housed together until experimentation on E20 or E21. Purina laboratory chow and tap water were available ad lib. Rats were maintained on a 12:12 h light-dark cycle (lights on at 0830h), in a temperature (22°C) controlled colony room, in accordance with the guidelines for animal care established by the National Institutes of Health (1986) (PHS publication 86-23). Pregnant rats and fetuses were killed by

Conservation of Dynamical Entropy 177
rapid cervical dislocation and decapitation, respectively, at the completion of the experiment.
Preparation of Pregnant Rats and Fetal Subjects Each pregnant rat was prepared on E20 or E21 of gestation to permit observation of
individual fetal subjects. The female was placed under ether anesthesia to perform a chemomyelotomy. This procedure involved the injection of 100% ethanol (100/A) into the spinal column between the L1 and L2 vertebrae. Chemomyelotomy results in an irreversible spinal blockade in the lower thoracic and lumbosacral regions. The prepared female was placed in a plastic holding apparatus that was partly submerged in a warm bath (37.5~C) containing buffered isotonic saline. The uterus was externalized into the bath through a midline abdominal incision. Subject fetuses were delivered into the bath through a small incision in the uterus. The amniotic and chorionic membranes surrounding the subject fetus were removed prior to experimental manipulation. These procedures preserve the umbilical- placental and placental-uterine connections, allowing the fetuses to remain fully oxy- genated, as indicated by their pink coloration. The prepared female and fetuses were allowed to recover after removal from the ether anesthetic and adjust to the bath for 20 rain prior to experimentation (Smotherman & Robinson, 1991).
Administration of Cocaine to Individual Fetal Subjects Fetal subjects were assigned to one of three drug groups on day E20 or E21: (a) SAL
(saline control); (b) 2.5 mg/kg cocaine (COC); or (c) 10.0 mg/kg cocaine (COC). Cocaine hydrochloride (Research Biochemicals Inc., Natick, MA, USA) was dissolved in an isotonic saline vehicle. The two dosages of cocaine were calculated according to a standard body weight (4.4 #g for E20 fetuses and 5.5 pg for E21 fetuses) and a fixed volume of injection (1.0/tl), using the method described in Simonik et al. (1993, 1994). The pH of all solutions was neutralized to 7.0 using sodium hydroxide. Cocaine and vehicle solutions were administered by intra-cisternal (IC) injection. The shank of a 30 gauge hypodermic needle was attached to a length of polyethylene tubing (PE-10) and the drug solution was loaded into the distal section of the tubing. The needle then was inserted under visual guidance into the foramen magnum between the occipital bone and the first cervical vertebra, with the tip placed in the cisterna magna (Varlinskaya et al., 1994). Successful placement of the needle into the cisterna magna was confirmed by the appearance of a small volume of cerebrospinal fluid in the tubing. Cocaine solutions and isotonic saline were administered at fetal body temperature (37.5°C) in an 8--10 s pulse using a 0.2 ml micrometer glass syringe (Gilmont Instruments, Barrington, IL, USA). A 1 ~1 isotonic saline injection was used to control for the effects of IC injection. Rat fetuses treated by administration of isotonic saline into the central nervous system do not differ from sham-injected or untreated control fetuses (Petrov et al., 1994; Simonik et al., 1994; Varlinskaya et al., 1994). Care was taken to ensure that the subject, except for the site of the injection, remained submerged in the bath during the injection.
All three drug groups were represented at least once in a litter when feasible, with no more than two subject fetuses from the same litter assigned to the same treatment condition, to circumvent the potential confounding of litter effects with treatment effects (Holson & Pearce, 1992). In addition, the order of testing for the different drug groups within a pregnancy was counterbalanced between pregnancies. Sixteen fetuses were tested in each of

178 w.P. Smotherman et al.
the three treatment groups on E20. On E21, 14, 15 and 13 fetuses were tested in the SAL, 2.5 mg/kg cocaine and 10 mg/kg cocaine treatment groups, respectively.
Behavioral Observation Observation of fetal behavior began within 5 s of IC injection, and continued for the
20 min test session. Fetal motor behavior was observed by an experimenter who was blind to the drug treatment assignment of the individual subject fetus. This experimenter reported behavioral categories to an assistant who recorded the data using an event recording program on a Macintosh computer, allowing real-time recording of the frequency and timing of fetal subjects' movements (Smotherman & Robinson, 1991). Five behavioral categories were distinguished, which provided an exhaustive record of all fetal movement: (a) forelimb (a movement of any part of one or both forelimbs); (b) hindlimb (a movement of any part of one or both hindlimbs); (c) head (a movement of the head on any axis relative to body orientation); (d) oral (opening and closing of the mouth or protrusion of the tongue); and (e) facial wipe (movement of one or both of the forelimbs in a direction from the ear towards the mouth, along the side of the face, with the forelimb stroke making contact with the face). The sum of fetal movements in all five behavioral categories provided the measure of overall fetal motor activity examined here.
Data Analysis The time series of total fetal movements, summed over the five behavioral categories and
partitioned into 5-s bins for all the 20-rain sessions, generated 240 sequential observations per subject. Transformations of these time series were chosen to examine the dimensionless entropy conservation-variational relation developed in the introduction, hr ,~ 21 x DR.
The topological entropy, hr, is roughly a logarithmic estimate of the rate of generation of new patterns of total motions over time. The leading Lyapounov characteristic exponent, 2~, can be thought of as a logarithmic estimate of the largest rate of stretching and folding of total activity values between sequential observations, sometimes called 'sensitivity to initial conditions'. The correlation dimension, DR, is a measure of the system's 'support', the hierarchical space inscribed as the irregularity in the geometry of the parametric surfaces of the actions (the manifolds of behavioral possibilities). We also examine group differences in each fetus's mean total (mean) and maximum (max) activity per 5-s partitiofi over the 20 min sessions.
We eschew the frequently used applied dynamical systems procedure of generating surrogate data sets as reference sequences for comparison (via randomization of either the primary sequences or the coefficients of their Fourier transformations). The goal of the surrogate data procedure, the discrimination of 'deterministic chaos' from 'randomness', is of no relevance to our problem, which is directed towards the statistical discrimination of observables from experimentally defined states.
As is frequently the case with technically difficult, finite length biological series, no single animal's times series of total motions, here n = 240 points in time, is sufficient to yield a stable individual estimate of any of our measures, hr, 21 or DR. For example, it is generally believed that the sample length required to compute the Dn as an estimate of the number of independent parameters in the (unknown) deterministic differential equation representing the process that generated the observables, ranges from n = 10 °/z+l to n -- 10 °. This would mean that one of the drug-free population means of DR ~ 6, if observed in a single animal, would require a time series of 104-106 points, weeks beyond the duration of action of an injection

Conservation of Dynamical Entropy ~79
of cocaine. Eckmann et al. (1986) and others have emphasized the importance of an adequate time of observation rather than the number of observations with respect to encompassing the phenomena being observed adequately. Our observation time is satisfactory with respect to this issue.
With respect to the necessary dimension of the embedding space for the computation of these measures, a recent theorem and associated numerical experiments (Ding et al., 1993) have demonstrated that the phase space reconstruction of the time series, 'the delay map', does not have to be one-to-one, and it is sufficient for its dimension, d,,, = Dn. We have found that a standard delay space dimension of 5, for both DR and 21, in the neighborhood of that required by these findings, yields the variational relations we seek when these relations are studied using statistically significant differences among population means. Whereas the hr ~ 2~ x DR relation held across a range of delay space embedding dimensions, we chose a constant dm= 5. The reliability of the estimates of changes in hr, 2~ and Dn induced by the experimental conditions derives from statistical inferences from group averages with 13--16 animals per group and under conditions of remarkably reliable experimental procedures.
Topological Entropy. The hr of some time-dependent process, q)t(x), hr(cpt(x)), is an estimate of how many different orbits cpt(x) is generating over time. In this computation, we use a natural 'moment partition' of the amplitude space of x~ = responses~5 s into six bins bounded by the mean _1, 2 or 3 SD, and follow the amplitude-over-time orbits from partition to partition using a 6 x 6 transition matrix across the 240 points. From the transition matrix we generate an incidence matrix of 0s and ls (making this matrix slightly sparse by counting any box with ~< 0.0375n - 0 and >0.0375n = 1, i.e. if any of the cells was visited nine times or less it was considered empty). The asymptotic growth rate of the trace (sum of the elements of the major diagonal) of this matrix as it is exponentiated reflects the growth rate of new orbits of all periods in the system (Bowen, 1975), and serves as an estimate of its largest eigenvalue, the logarithm of which is called the topological entropy, hr (Fried, 1986; Parry & Pollicott, 1983). The counting function most commonly used for this purpose has been called the Artin-Mazur (Smale, 1967) or the Ruelle zeta function (Polli- cott, 1985) and is usually written: ~(t) ~ exp(~((N,, t~)/n), where N,, is the number of points of period n and t is a dummy variable. This and other algorithmic expressions of hT are presented with examples in MandeU and Selz (1993b).
Leading Lyapounov Exponent. /~1 is a measure of the average exponential rate of divergence of nearby initial conditions (x and y), such that any, even infinitesimal, initial distance between them, e, dlx-y l <~ e, increases under the (putative) action of qo t ~ exp(2t), such that dlw(x)~'-~0(y)~'l>~ >> e for any ~. We compute )q over the data series as reconstructed by plotting its value xt against xt_r, xt_2r, Xt-3T, and xt~r in a five-dimensional phase space, following the local linear dynamics on this frame of perpendicular vectors. We chose neighboring pairs, xi and Yi, computing the evolution of the separation between them as an average within an n-dimensional hypersphere at five successive time steps, then iteratively repeating the process choosing another xi and some neighboring point, Yi, to obtain an average rate of separation of xis and yis that is roughly equivalent to the directional derivatives of the process. We chose the leading (largest, most positive) 21, representing the rate of separation along the vectorial dimension of the greatest dynamical stretching process. We compute an error term for 21 as 2.5 times its standard deviation divided by the square root of the number of xi, Yi distance trajectories examined (see Wolf et al., 1985). ,~l is reported as an empirical measure.

180 W.P. Smotherman et al.
Correlation Dimension. DR also is computed using a phase space reconstruction of dimen- sion dm= 5. We pass repeatedly through the data series, xi, each time centering a dm- dimensional hypersphere with radius r on each point. We count the number of data points n(xi) that fall within the n-sphere as r ~ 0 and graph log(n(xi)) vs. log(r). DR is the average slope of the center 1/4 (most likely to be linearly scaling) part of the vertical axis of the graph. The associated error term is equal to half of the difference of the maximum and minimum local slopes in this interval. DR is well defined if the graph log(n(xi)) vs. log(r) has a linear region and a plot of its value vs. the phase space embedding dimension used in its computation, dm, has a plateau region. Remarkably for these relatively short time series, perhaps in part due to their relatively small range of integer values and the 'infinite precision' of a simple counting procedure, it was almost always the case that DR plateaued at a dm value only a little delayed from the DR observed (see Ding et al., 1993; Grassberger & Procaccia, 1983).
A MANOVA using the dependent measures mean, max, hr, 2z and DR was followed, when appropriate, by factorial ANOVAs (2 x 3--age by treatment), weighted for unequal cell sizes, to determine the presence of univariate age by treatment interaction effects and main effects of treatment (SAL, 2.5 mg/kg cocaine; COC, 10 mg/kg COC) or age (E20, E21). Post-hoc pairwise contrasts were performed to clarify the origins of any significant interaction or main effects of treatment.
As a result of the postulated conservation-variational relation of hr, ~1 and DR and the natural relation of the mean and max values of the data series, dependent measures might be expected to show strong intercorrelations. Whereas this is empirically not always the case, it has been our experience that the measure spaces generated in this way tend to be oblique. We have, in other contexts, dealt with this issue by the construction of an orthogonal component space based on these measures (Selz, 1992), but in this case the inter-relations of the dependent measures are of direct interest to us, and the creation of abstract dependent components would interfere with interpretation.
RESULTS
Cocaine Effects on Fetal Motor Activity Representative individual examples of the total motor behavior of control and cocaine-
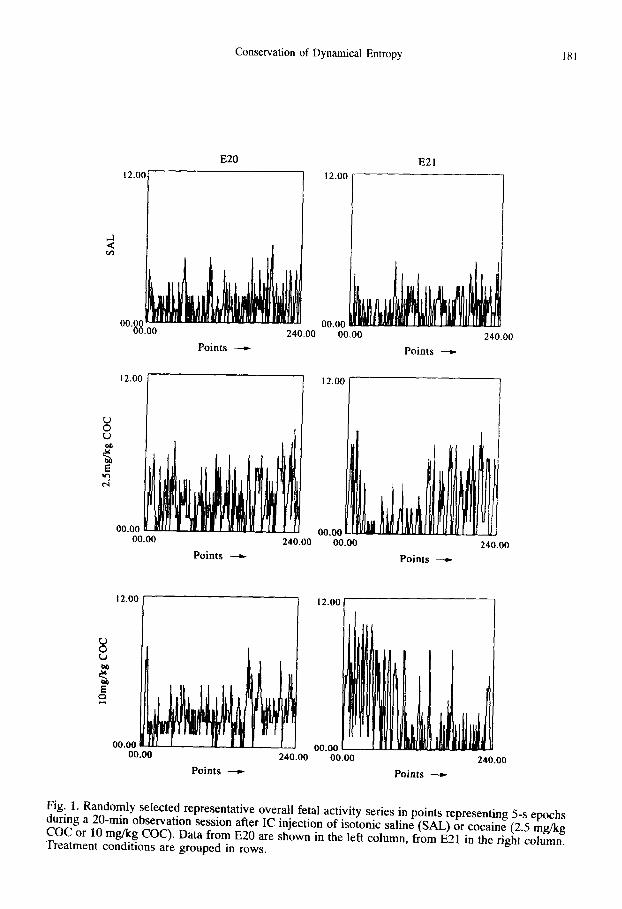
treated fetuses are graphed for gestational days E20 and E21 in Fig. 1, with each point indicating total motor movements per 5-s period over the 20 min of observation. As is qualitatively obvious in the examples in Fig. 1, and represented quantitatively for the groups in Fig. 2 (upper left box plot), COC produced significant, but not dose-dependent elevations in mean total fetal activity above control levels over the 20 rain of observation. This effect was particularly prominent in the E21 fetuses (see Fig. 1) during the first 5 min after IC injection. This pattern of cocaine effects on fetal motor behavior has been reported previously (Simonik et al., 1993, 1994) as has power spectral evidence for the increased prominence of some faster frequency modes by COC.
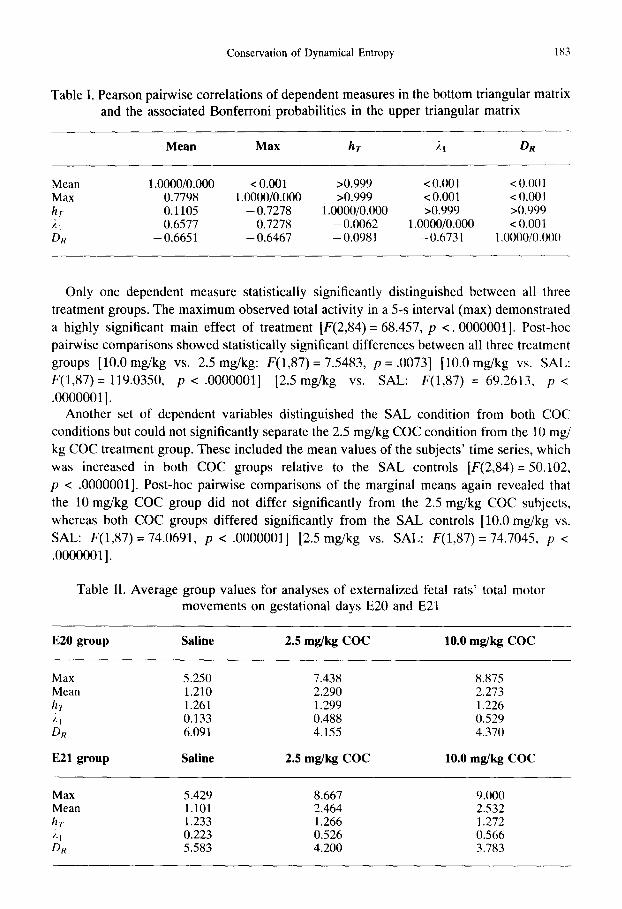
As mentioned above, many of our dependent measures might be expected to show strong intercorrelations. Table I lists the intercorrelations of the dependent measures discussed above, and their associated Bonferroni adjusted probabilities. Note that whereas hr is not correlated with either 21 or DR, 21 and DR demonstrate a strong inverse relationship.
A MANOVA revealed no significant multivariate (or univariate) age by treatment interaction effects on the dependent measures at an ct ~< 0.05 level. Similarly, the MANOVA
Conservation of Dynamical Entropy 181
,.-1 < r~
0 E
t'N
E20 12.001
008800 240.00
Points ~
12.00
00.00 00~00
E21
240.00
Points
12.00
00.00 00.00
( 12.00
00.00 Lr|vur~~~ 240.00 00.00 240.00
Points ~ Points
r..) O
uo
O
12.00
00.00 O0.Oq Points - - ~
240.00
12.00
00.00 00.00
Points
hJlJ 240.00
Fig. 1. Randomly selected representative overall fetal activity series in points representing 5-s epochs during a 20-min observation session after IC injection of isotonic saline (SAL) or cocaine (2.5 mg/kg COC or 10 mg/kg COC). Data from E20 are shown in the left column, from E21 in the right column. Treatment conditions are grouped in rows.
182 W.P. Smotherman et al.
3 ~ J
2
1.5
1.4
1.3 1 -
1.2
1.1
1.0
i i i
10mg/kg 2.5mg/kg SAL
! | w
i i I
10mg/kg 2.5mg/kg SAL
8
15
a ~
10
0
0.8
0.6
0.4i
0.2
0.0
0.2
i i i
I I i
10mg/kg 2.5mg/kg SAL
i
i A I
10mg/kg 2.5mg/kg SAL
Ij J I I I
10mg/kg 2.5mg/kg SAL
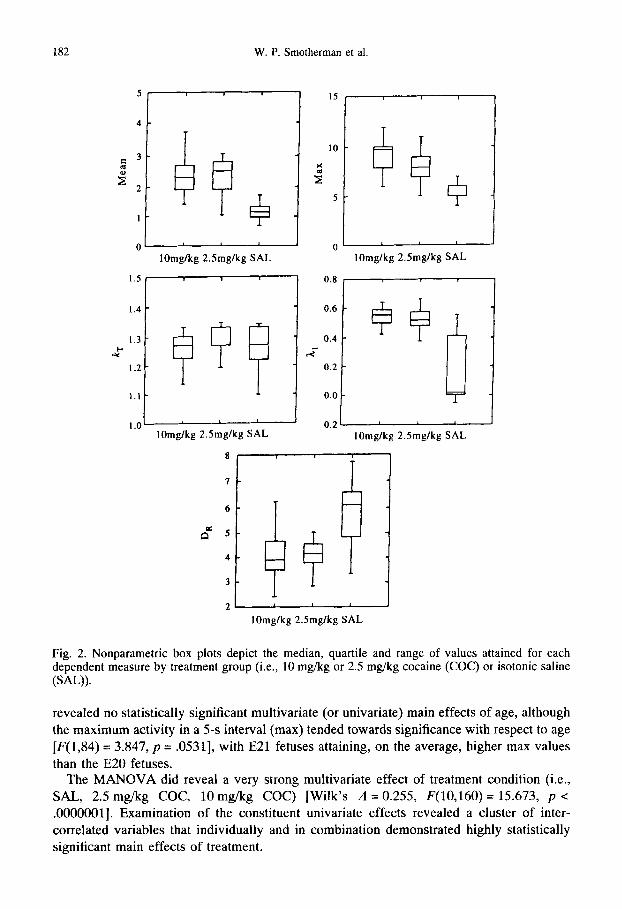
Fig. 2. Nonparametric box plots depict the median, quartile and range of values attained for each dependent measure by treatment group (i.e., 10 mg/kg or 2.5 mg/kg cocaine (COC) or isotonic saline (SAL)).
revealed no statistically significant multivariate (or univariate) main effects of age, although the maximum activity in a 5-s interval (max) tended towards significance with respect to age [F(1,84) = 3.847, p = .0531], with E21 fetuses attaining, on the average, higher max values than the E20 fetuses.
The MANOVA did reveal a very strong multivariate effect of treatment condition (i.e., SAL, 2.5 mg/kg COC, 10mg/kg COC) [Wilk's A =0.255, F(10,160)= 15.673, p < .0000001]. Examination of the constituent univariate effects revealed a cluster of inter- correlated variables that individually and in combination demonstrated highly statistically significant main effects of treatment.
Conservation of Dynamical Entropy 183
Table I. Pearson pairwise correlations of dependent measures in the bottom triangular matrix and the associated Bonferroni probabilities in the upper triangular matrix
Mean Max hr 2t DR
Mean 1.0000/0.000 < 0.001 >0.999 < 0.001 < 0.001 Max 0.7798 1.0000/0.000 >0.999 < 0.001 < 0.001 hi 0.1105 - 0.7278 1.0000/0.000 >0.999 >0.999 ,;~l 0.6577 0.7278 - 0•0062 1.0000/0.000 < 0.001 Dn - 0.6651 -- 0•6467 - 0.0981 - 0.6731 1.0000/0.000
Only one dependent measure statistically significantly distinguished between all three treatment groups• The maximum observed total activity in a 5-s interval (max) demonstrated
a highly significant main effect of treatment [F(2,84) = 68.457, p <. 0000001]. Post-hoc pairwise comparisons showed statistically significant differences between all three treatment
groups [10.0 mg/kg vs. 2.5 mg/kg: F(1,87) = 7.5483, p = .0073] [10.0 mg/kg vs. SAL: F(1,87)= 119.0350, p < .0000001] [2.5 mg/kg vs. SAL: F(1,87) = 69.2613, p <
.00000011. Another set of dependent variables distinguished the SAL condition from both COC
conditions but could not significantly separate the 2.5 mg/kg COC condition from the 10 mg/ kg COC treatment group. These included the mean values of the subjects' time series, which
was increased in both COC groups relative to the SAL controls [F(2,84)= 50.102,
p < .0000001]. Post-hoc pairwise comparisons of the marginal means again revealed that the 10 mg/kg COC group did not differ significantly from the 2.5 mg/kg COC subjects,
whereas both COC groups differed significantly from the SAL controls [10.0 mg/kg vs. SAL: F(1,87)= 74.0691, p < .0000001] [2.5 mg/kg vs. SAL: F(1,87)=74.7045, p < • 0000001 ].
Table II. Average group values for analyses of externalized fetal rats' total motor movements on gestational days E20 and E21
E20 group Saline 2.5 mg/kg COC 10.0 mg/kg COC
Max 5.250 7.438 8.875 Mean 1.210 2.290 2.273 hr 1.261 1.299 1.226 ),1 0.133 0.488 0.529 D R 6.091 4.155 4.370
E21 group Saline 2.5 mg/kg COC 10.0 mg/kg COC
Max 5.429 8.667 9.000 Mean 1.101 2.464 2.532 hv 1.233 1.266 1.272 )-I 0.223 0.526 0.566 DR 5.583 4.200 3.783

184 w.P. Smotherman et al.
Similarly, and in keeping with our postulated conservation-variational relation, the Lyapounov characteristic exponent (21) [F(2,84) --- 55.310, p < .0000001] was increased in either COC group relative to the SAL controls [10.0 mg/kg vs. SAL: F(1,87)= 90.8978, p < .00000001] [2.5 mg/kg vs. SAL: F(1,87)= 75.3817, p < .0000001], but could not distinguish between the 2.5 mg/kg COC and 10mg/kg COC treatment groups. The correlation dimension (DR) of the subjects' activity series were also decreased in both COC conditions as compared to the SAL control series' values [F(2,84) = 31.044, p < .0000001]. Pairwise comparisons demonstrated that the two COC groups did not differ significantly from each other, whereas both COC groups differed significantly from the SAL controls [10.0mg/kg vs. SAL: F(1,87)=47.0682, p < .0000001] [2.5 mg/kg vs. SAL: F(1,87) = 44.8604, p < .0000001]. This supports the relation 21 ~ DR 1.
The group means of all these variables are shown in Table II, and the effects depicted in Fig. 2.
Also, as predicted by the dynamical entropy conservation hypothesis, the topological entropy (hr) did not show any significant effects of age or treatment conditions. Given the relation hr ~ 21 × DR, 21 and DR should vary inversely to each other if hr is conserved across treatment groups.
DISCUSSION
The patterns of measures generated by the transformations of the time series of cocaine- influenced vs. control spontaneous total fetal motor activity revealed a dimensionless variational relation among statistical dynamical indicators associated with the conservation of the behavior's global dynamical entropy. These results demonstrate that, with respect to group means, the system's rate of generation of different patterns of spontaneous motor movement, quantified by its topological entropy, can be conserved, AhT= 0, if, when constrained to a lower fractal dimension, quantified here by a lower correlation dimension, ADR < 0, it mixes faster in time via a stretching and folding process reflected in an associated increase in its most positive characteristic exponent, A21 > 0.
From the standpoint of standard psychopharmacological theory (Lyons & Robbins, 1975), these stimulant drug-induced effects fail to indicate that the increased spontaneous motor activity is associated with a decrement in its variety. Rather, cocaine appears to generate a new dynamical pattern characterized by an expansion of the relative distance between consecutive data points and a relative decrease in the complexity of the patterns of their nearest-neighbor relations, which, taken together, leave the sequence-dependent behavioral variety unchanged.
These and other techniques of discrimination between patterns of behavior are part of an emerging 'tool box' of useful new measures being derived from basic studies of the properties of chaotic dynamical systems. When compared with distributions and power spectral transformation, they allow increased subtlety of characterization without the loss of statistical power.
As noted above, this dimensionless conservation-variational relation, hr ~ 21 × DR has been proved as a mathematical theorem for suitably conditioned, abstract chaotic dynamical systems (Ledrappier & Young, 1985; Manning, 1981). In less formal terms, these analyses suggest that cocaine induces a more 'workaholic' or 'compulsive' pattern in fetal motor movements, characterized by a complexity-conserving, variational relation associated with increasing busyness in time and reduced hierarchical parametric degrees of freedom.

Conservation of Dynamical Entropy 185
To extend this close analogy further, failure to respond adaptively to biologically relevant new stimuli, such as oral stimulation, may result from being 'just as busy but with something else'. If normal fetal development requires dynamical flexibility with respect to responding to temporally critical trophic factors, it may be that 'preoccupation' is as potentially damaging as the pathological simplification with entropy loss implicit in a 'lost variety theory' of stimulant drug action.
This pattern of cocaine-induced dynamical change is consistent with the experimental findings of Smotherman & Robinson (1987, 1988) and Simonik et al. (1993, 1994) in which increased motoric activity was associated with increases in the power of broad-band fast frequencies in their power spectral representations and with the loss of normal responsivity to perioral perturbation. It is perhaps relevant that, unlike amphetamine, cocaine induces suppression of the P300 orienting response to novel stimuli in behaviorally activated human subjects who are performing highly demanding tasks (Jones, 1984).
Acknowledgements: WPS is supported by MERIT award HD 16102 and by NICHD grants HD 28231 and 28014. AJM and KAS are supported by the Office of Naval Research, sections on Physics and Biological Intelligence.
REFERENCES
Aschoff J (1981) Handbook of Behavioral Neurobiology, Vol 4, Biological Rhythms. Plenum, New York.
Bowen R (1975) Equilibrium States and the Ergodic Theory of Anosov Diffeomorphisms. Lect. Notes Math, Am Math Soc, Providence, RI.
Bowen R (1978) On Axiom A Diffeomorphisms 35. Am Math Soc, Providence, RI. Corner MA (1977) Sleep and the beginnings of behavior in the animal kingdom - - studies of ultradian
motility cycles in early life. Progr Neurobiol 8:278-295. Ding M, Grebogi C, Ott E, Sauer T, Yorke J (1993) Plateau onset for correlation dimension: When
does it occur? Phys Rev Lett 70:3872-3875. Dow-Edwards DL (1991) Cocaine effects on fetal development: A comparison of clinical and animal
research findings. Neurotox Teratol 13:347-352. Eckmann JP, Kampharst SO, Ruelle D, Ciliberto S (1986) Lyapounov exponents from time series.
Phys Rev A34:4971-4978. Fried D (1986) The zeta function of Ruelle and Selberg I. Ann Sci Ec Norm Sup 71:213-242. Gingras JL, Weese-Mayer DE, Hume RF, O'Donnell KJ (1992) Cocaine and development:
Mechanisms of fetal toxicity and neonatal consequences of prenatal cocaine exposure. Early Hum Dev 31:1-24.
Grassberger P, Procaccia I (1983) Measuring the stangeness of strange attractors. Physica 9D:189- 208.
Holson RR, Pearce B (1992) Principles and pitfalls in the analysis of prenatal effects in multiparous species. Neurotox Teratol 14:221-228.
Jones RT (1984) P-300 studies with cocaine in man. In: Grabowski J (Ed) Cocaine: Pharmacology, Effects and Treatment of Abuse. NIDA Res Monographs 50:34--54.
Kittel C (1958) Elementary Statistical Physics. Wiley, New York. Ledrappier F, Young LS (1985) The metric entropy of diffeomorphisms Part II: Relations between
entropy, exponents and dimension. Ann Math 122:540-574. Lyons M, Robbins TW (1975) The action of central nervous system stimulant drugs. In: Essman W,
Valzelli L (Eds) Current Developments in Psychopharmacoiogy, Vol. 2. Spectrum, New York, pp 79-163.
Mandell AJ (1983) From intermittency to transitivity in neuropsychobiological flows. Am J Physiol 245 :R484-R494.
Mandell AJ (1987) Dynamical complexity and pathological order in the cardiac monitoring problem. Physica D27:1235-1243.

186 W.P. Smotherman et al.
Mandell AJ, Russo PV, Knapp S (1982) Strange stability in hierarchically coupled neuropsychobio- logical systems. In: Haken H (Ed) Evolution of Order and Chaos. Springer, New York, pp 270--286.
Mandell AJ, Selz KA (1993a) Brain stem neuronal noise and neocortical resonance. J Stat Phys 70:355-373.
Mandell AJ, Selz KA (1993b) The new statistical dynamics: An informal look at invariant measures of psychological time series. In: Vallacher RR, Nowak AJ (Eds) Dynamical Systems in Social Psychology. Academic Press, New York, pp 55-69.
Manning A (1981) A relation between Lyapounov exponents, Hausdorff dimension and entropy. Erg Theory Dynam Sys 1:451-459.
Moessinger AC (1988) Morphological consequences of depressed or impaired fetal activity. In: Smotherman WP, Robinson SR (Eds) Behavior of the Fetus. Telford Press, Caldwell, New Jersey, pp 163-173.
National Institutes of Health (1986) Guide for the Care and Use of Laboratory Animals (DHEW Pub. No. 86-23). US Government Printing Office, Washington DC.
Oppenheim RW (1982) The neuroembryological study of behavior: Progress, problems, perspectives. In: Hunt RK (Ed) Current Topics in Developmental Biology, Vol. 17, Neural Development. Academic Press, New York, pp 257-309.
Parry W, Pollicott M (1983) An analogue of the prime number theorem for closed orbits of Axiom A flows. Ann Math 118:573-591.
Paulus MP, Geyer MA, Gold LH, Mandell AJ (1990) Ergodic measures of complexity in rat exploratory behavior. Proc Natl Acad Sci 87:723-727.
Petrov ES, Varlinskaya El, Robinson SR, Smotherman WP, Costa BR de, Rice KC (1994) Kappa opioid effects on fetal behavior: Central administration of U50,488. Physiol Behav 56:175-182.
Pollicott M (1985) On the rate of mixing of Axiom A flows. Invent Math 81:413-426. Robertson SS (1985) Cyclic motor activity in the human fetus after midgestation. Dev Psychobiol
18:411-419. Robertson SS (1987) Human cyclic motility: Fetal-newborn continuities and newborn state
differences. Dev Psychobiol 20:425-442. Robertson SS (1988) Mechanism and function of cyclicity in spontaneous movement. In: Smotherman
WP, Robinson SR (Eds) Behavior of the Fetus. Telford Press, Caldwell, New Jersey, pp 77-94. Robertson SS, Bacher LF (1995) Oscillation and chaos in fetal activity. In: Lecanuet J-P, Krasnegor
NA, Fifer WP, Smotherman WP (Eds) Fetal Development: A Psychobiological Perspective. Lawrence Erlbaum, New York, pp 169-190.
Robertson SS, Dierker LJ (1986) The development of cyclic motility in fetus of diabetic mothers. Dev Psychobiol 19:223-234.
Ruelle D (1989) Chaotic Evolution and Strange Attractors. Cambridge University Press, New York. Selz KA (1992) Mixing Properties in Human Behavioral Style and Time Dependencies in Behavior
Identification: The Modeling of a Universal Dynamical Law. University Microfilms International, Ann Arbor, Michigan.
Selz KA, Mandell AJ (1991) The Bernoulli partition-equivalence of intermittent neuronal discharge patterns. Int J Bifurc Chaos 1:717-722.
Selz KA, Mandell AJ (1992) Critical coherence and characteristic times in brain stem neuronal discharge patterns. In: McKenna T, Davis J, Zornetzer S (Eds) Single Neuron Computation. Academic, New York, pp 525-550.
Selz KA, Mandell AJ (1993) Style as mechanism: From man to a map of the interval and back. SPIE 2036:174---182.
Selz KA, Mandell AJ, Anderson CM, Smotherman WP, Teicher MH Distribution of local Mandelbrot- Hurst exponents: Motor activity in cocaine treated fetal rats and manic-depressive patients. Fractals (In press).
Shlesinger MF (1989) Fractal time in condensed matter. Ann Rev Phys Chem 39:269-290. Simonik DK, Robinson SR, Smotherman WP (1993) Cocaine alters behavior in the rat fetus. Behav
Neurosci 107:867-875. Simonik DK, Robinson SR, Smotherman WP (1994) Cocaine alters cyclic motor activity in the fetal
rat. Dev Psychobiol 27:489-501. Singer L, Farkas K, Kliegman R (1992) Childhood medical and behavioral consequences of maternal
cocaine use. J Pediatr Psych 17:389-406.

Conservation of Dynamical Entropy 187
Singer L, Garber R, Kliegman R (1991) Neurobehavioral sequelae of fetal cocaine exposure. J Pediatrics 119:667-672.
Smale S (1967) Differential dynamical systems. Bull Am Math Soc 73:747-817. Smale S (1981) The Mathematics of Time. Springer, New York. Smotherman WP, Robinson SR (1987) Prenatal influences on development: Behavior is not a trivial
aspect of fetal life. Dev Behav Pediatr 8:171-176. Smotherman WP, Robinson SR (1988) The uterus as environment: The ecology of fetal experience.
In: Blass EM (Ed) Handbook of Behavioral Neurobiology, Voi. 9, Developmental Psychobiology and Behavioral Ecology. Plenum, New York, pp 149-196.
Smotherman WP, Robinson SR (1991) Accessibility of the rat fetus for psychobiological investigation. In: Shair HN, Barr GA, Hofer MA (Eds) Developmental Psychobiology: New Methods and Changing Concepts. Oxford University Press, New York, pp 148-163.
Smotherman WP, Robinson SR, Robertson, SS (1988) Cyclic motor activity in the fetal rat (Rattus norvegicus). J Comp Psych 102:78-82.
Spear LP (1995) Neurobehavioral consequences of gestational cocaine exposure: A companttive analysis. Adv Inf Res 9:55-105.
Varlinskaya EI, Petrov ES, Robinson SR, Smotherman WP (1994). Behavioral effects of centrally administered arginine vasopressin in the rat fetus. Behav Neurosci 108:395409.
Wolf A, Swift JB, Swinney HL, Vastano JA (1985) Determining Lyapounov spectra from a time series. Physica 16D:285-317.
Related Documents











