ВІСНИК ЛЬВІВ. УН-ТУ VISNYK OF L’VIV UNIV. Серія біологічна. 2002. Вип.29. С. 52-61 Biology Series. 2002. Is.29. P. 52-61 Голуб Н., Козаревська І., Черник Я., 2002 УДК 575. 24: 595. 77Х. 4 ІНДУКЦІЯ НІТРОЗОЕТИЛСЕЧОВИНОЮ ДОМІНАНТНИХ ЛЕТАЛЬНИХ МУТАЦІЙ ТА Х-ЗЧЕПЛЕНИХ ВИДИМИХ МУТАЦІЙ У DROSOPHILA MELANOGASTER Н. Голуб, І. Козаревська, Я. Черник Львівський національний університет імені Івана Франка, вул. Грушевського, 4, м. Львів 79005, Україна, e-mail: [email protected] Досліджено вплив нітрозоетилсечовини на індукцію домінантних летальних мутацій, частоту появи видимих мутантів та мутантів зі змінами структур головно- го мозку по Х-хромосомі у Drosophila melanogaster. З’ясовано, що нітрозоетилсе- човина у концентрації 0,5 мкг/мл (LD 50 для лінії y 2 w a4 ) призводить до збільшення частоти виникнення домінантних летальних мутацій у 6 разів порівняно з контро- лем. У цій дозі нітрозоетилсечовина зумовлює виникнення видимих мутантів по Х-хромосомі з частотою 2,6х10 -4 -4,3х10 -4 протягом двох поколінь. У концентрації 0,4 мкг/мл цей мутаген індукує появу з високою частотою змін у структурах голов- ного мозку дрозофіли, які можуть виникати в різні періоди життя імаго (у 5-, 10- та 30-денному віці). Усім мутантним культурам з нейродегенеративними змінами мозку властиві швидке відмирання особин та знижені показники середньої трива- лості життя. Ключові слова: дрозофіла, нітрозоетилсечовина, домінантні летальні мутації, мута- генез, нейродегенерації. Багато сучасних праць присвячено вивченню наслідків впливу різних хіміч- них факторів на живі об’єкти. Особливу увагу приділяють алкілувальним сполу- кам. Алкілувальні агенти є сильними мутагенами, оскільки спричинюють мутації майже у всіх організмах. Результатом дії алкілувальних мутагенів є заміна азотис- тих основ [1, 2, 3]. Вони можуть також зумовлювати різні хромосомні перебудови: великі і дрібні делеції, інверсії, транслокації та ін. [4]. Нітрозоетилсечовина (НЕС) та етилметансульфонат (ЕМС) є супермутагенами, оскільки мутації у випадку іх- ньої дії в різних тест-системах виникають з високою частотою. Г.Р. Щербата та Д.В. Максимів [5] довели, що НЕС індукує транспозиції МГЕ copia у генетично нестабільної лінії white Drosophila melanogaster. А. Пастнік зі співавторами [6] піс- ля впливу НЕС на дрозофілу проаналізували молекулярні механізми виникнення 36 мутацій у локусі white і довели, що більшість мутацій зумовлена замінами пар основ і перебудовами всередині гена розміром не більше 50-100 н.п. Мутагенну дію НЕС досліджували на ссавцях, зокрема, на мишах і хом’яках [7, 8, 9, 10]. З’я-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ВІСНИК ЛЬВІВ. УН-ТУ VISNYK OF L’VIV UNIV. Серія біологічна. 2002. Вип.29. С. 52-61 Biology Series. 2002. Is.29. P. 52-61
Голуб Н., Козаревська І., Черник Я., 2002
УДК 575. 24: 595. 77Х. 4
ІНДУКЦІЯ НІТРОЗОЕТИЛСЕЧОВИНОЮ ДОМІНАНТНИХ ЛЕТАЛЬНИХ МУТАЦІЙ ТА Х-ЗЧЕПЛЕНИХ ВИДИМИХ
МУТАЦІЙ У DROSOPHILA MELANOGASTER
Н. Голуб, І. Козаревська, Я. Черник Львівський національний університет імені Івана Франка,
вул. Грушевського, 4, м. Львів 79005, Україна, e-mail: [email protected]
Досліджено вплив нітрозоетилсечовини на індукцію домінантних летальних
мутацій, частоту появи видимих мутантів та мутантів зі змінами структур головно-го мозку по Х-хромосомі у Drosophila melanogaster. З’ясовано, що нітрозоетилсе-човина у концентрації 0,5 мкг/мл (LD50 для лінії y2wa4) призводить до збільшення частоти виникнення домінантних летальних мутацій у 6 разів порівняно з контро-лем. У цій дозі нітрозоетилсечовина зумовлює виникнення видимих мутантів по Х-хромосомі з частотою 2,6х10-4-4,3х10-4 протягом двох поколінь. У концентрації 0,4 мкг/мл цей мутаген індукує появу з високою частотою змін у структурах голов-ного мозку дрозофіли, які можуть виникати в різні періоди життя імаго (у 5-, 10- та 30-денному віці). Усім мутантним культурам з нейродегенеративними змінами мозку властиві швидке відмирання особин та знижені показники середньої трива-лості життя.
Ключові слова: дрозофіла, нітрозоетилсечовина, домінантні летальні мутації, мута-генез, нейродегенерації.
Багато сучасних праць присвячено вивченню наслідків впливу різних хіміч-них факторів на живі об’єкти. Особливу увагу приділяють алкілувальним сполу-кам. Алкілувальні агенти є сильними мутагенами, оскільки спричинюють мутації майже у всіх організмах. Результатом дії алкілувальних мутагенів є заміна азотис-тих основ [1, 2, 3]. Вони можуть також зумовлювати різні хромосомні перебудови: великі і дрібні делеції, інверсії, транслокації та ін. [4]. Нітрозоетилсечовина (НЕС) та етилметансульфонат (ЕМС) є супермутагенами, оскільки мутації у випадку іх-ньої дії в різних тест-системах виникають з високою частотою. Г.Р. Щербата та Д.В. Максимів [5] довели, що НЕС індукує транспозиції МГЕ copia у генетично нестабільної лінії white Drosophila melanogaster. А. Пастнік зі співавторами [6] піс-ля впливу НЕС на дрозофілу проаналізували молекулярні механізми виникнення 36 мутацій у локусі white і довели, що більшість мутацій зумовлена замінами пар основ і перебудовами всередині гена розміром не більше 50-100 н.п. Мутагенну дію НЕС досліджували на ссавцях, зокрема, на мишах і хом’яках [7, 8, 9, 10]. З’я-

ІНДУКЦІЯ НІТРОЗОЕТИЛСЕЧОВИНОЮ… 53
совано, що НЕС зумовлює збільшення кількості мутаційних подій у 10 разів порів-няно зі спонтанним рівнем у соматичних (клітинах кісткового мозку) та генератив-них (сперматозоїдах) клітинах мишей [7]. У деяких працях на мишах [11, 12, 13] доведена канцерогенна дія НЕС. Виявлено, що НЕС також має здатність індукува-ти утворення лімфом у людини [12].
Мутагенез алкілувальними сполуками дає перспективи для аналізу поведінко-вих мутацій та фізіологічних фенотипів хребетних, зумовлених генетичними змі-нами. Наприклад, сьогодні актуальні дослідження захворювань нервової системи людини, що зумовлені генетичними порушеннями. Тому важливими є досліджен-ня, спрямовані на вивчення генів людини, які відповідають за формування струк-тур нервової системи та їхнє функціювання. Зручним об’єктом для вивчення цієї проблеми є Drosophila melanogaster, у якої процеси диференціації і дегенерації нервової системи подібні до таких же процесів у ссавців, крім того, у неї висока гомологічність генів (до 70%), що контролюють нейродегенеративні зміни [14, 15]. Вивчення механізмів генетичних процесів можливе за наявності групи мутантів, у яких порушені досліджувані функції. Оптимальним підходом для отримання му-тантів зі змінами структур головного мозку є застосування хімічних мутагенів, перш за все алкілувальних агентів, які здатні спричинювати точкові мутації в різ-них генах, в тому числі й тих, які відповідають за функціювання нервової системи.
Наша мета – дослідити вплив НЕС на індукцію домінантних летальних мута-цій (ДЛМ) та частоту появи видимих мутантів по Х-хромосомі; отримати та про-аналізувати індуковані НЕС мутанти по Х-хромосомі зі змінами структур головно-го мозку у Drosophila melanogaster.
У дослідженнях використано лінії Drosophila melanogaster: дикий тип Oregon; лабораторну лінію y2wa4 з морфологічними маркерами по Х-хромосомі.
НЕС вносили в середовище для личинкового згодовування. Для обліку види-мих мутацій в Х-хромосомі самців використовували самок лінії C(1)DX, yf зі зчеп-леними Х-хромосомами [16]. Тест на виявлення ДЛМ виконували згідно з працею [17]. Для отримання препаратів головного мозку досліджуваних культур дрозофі-ли виготовляли парафінові зрізи за методом,. Описаним у [18]. Побудову кривих виживання (КВ), аналіз середньої тривалості життя (СТЖ) і максимальної трива-лості життя (МТЖ) проводили згідно з [19]. Дрозофіл утримували в темноті при температурі 250С на стандартному поживному середовищі [20].
Для НЕС LD50 ми визначали у стабільної лабораторної лінії y2wa4. Для цього готували розчини таких концентрацій: 0,5; 1,0; 5,0; 10,0; 50,0; 100,0 мкг/мл, і зго-довували личинкам. З’ясовано, що напівлетальною дозою НЕС для лінії y2wa4 є концентрація 0,5 мкг/мл, яку ми й використали у подальшій роботі з цією лінією.
З метою дослідження впливу НЕС на ранні стадії онтогенезу дрозофіли вико-нано тест на ДЛМ, який дає змогу реєструвати як генні мутації, так і хромосомні аберації, враховувати частоту ранньої (РЕЗ) та пізньої ембріональної загибелі (ПЕЗ). У цьому разі домінантні мутації виявляються як у гомозиготному, так і в ге-терозиготному станах, що дає змогу з високою точністю визначити кількість мута-ційних пошкоджень геному (табл. 1). Як видно з даних табл. 1, у лінії y2wa4 по-
Н. Голуб, І. Козаревська, Я. Черник 54
казник частоти виникнення ДЛМ становив 3,97% і статистично не відрізнявся від такого значення для лінії дикого типу Oregon (3,74%). На підставі аналізу загаль-ної частоти появи ДЛМ, індукованих НЕС, виявлено майже шестиразове збіль-шення цього показника (19,46%) порівняно з контролем. Таке збільшення відбу-валося, головно, завдяки зростанню частоти РЕЗ (до 17,89%), тоді як частота ПЕЗ не перевищувала значень контролю і становила 1,58%. Отже, за наявності 0,5 мкг/мл НЕС виявлено значні порушення ембріогенезу дрозофіли, зумовлені ви-соким порівняно з контролем показником виникнення ДЛМ.
У подальших експериментах досліджено вплив НЕС дозою LD50 на частоту появи видимих мутантів по Х-хромосомі у лінії y2wa4. Лінія y2wa4 є стабільною ла-бораторною лінією, у якої частота появи мутантів становить 8,9х10-5, що не пере-вищує значення природного рівня мутування. Серед 11 222 самців ми виявили од-ного мутанта по гену yellow (y2→y+). Унаслідок обробки личинок НЕС частота ви-никнення мутантів була на порядок вищою порівняно зі спонтанним рівнем. На-приклад, в F1 частота мутування становила 2.6х10-4, а в F2 – 4.3х10-4, що в 1,6 раза перевищувало показник першого покоління (табл. 2).
Надалі НЕС використано для індукції мутацій у Х-хромосомі, які б приводи-ли до появи нейродегенеративних змін у структурах головного мозку дрозофіли. Мутаген вносили для личинкового згодовування лінії Oregon у різних концентра-ціях: 0,4; 2,0; 4,0 мкг/мл. З огляду на те, що сперматогенез у дрозофіли відбуваєть-ся двома хвилями, схрещування проводили методом брудів, який дає змогу врахо-вувати чутливість різних стадій сперматогенезу до дії мутагенного фактора. Для
Таблиця 1 Частота виникнення ДЛМ, індукованих НЕС (0,5 мкг/мл) у лінії y2wa4, %
Частота Дослід РЕЗ ПЕЗ ДЛМ
Коефіцієнт Стьюдента
Oregon 3,02 0,72 3,74 0,85 y2wa4 (контроль) 2,06 1,91 3,97 Y2wa4 (НЕС) 17,89 1,58 19,46 26,16* * р≥ 0,95 щодо контролю
Таблиця 2 Частота появи та генотипи мутантів, отриманих при впливі НЕС (0,5 мкг/мл)
на лінію y2wa4
Дослід Поколі-ння
Кількість проаналізо-ваних самців
Генотип і кількість мутантів
Частота появи
мутантів
t, коефіціент Стьюдента
y2wa4 (контроль) 11222 1 y+wa4 8,9x10-5
y2wa4 (НЕС) F1 7497 1 y1w+ 1 y2wa4sc1 2,7х10-4 0,82 F2
2326 y2wa4sc1 4,3x10-4 0,96
ІНДУКЦІЯ НІТРОЗОЕТИЛСЕЧОВИНОЮ… 55
одержання мутантних культур по Х-хромосомі використовували самок музейної лінії FM4 (In(1)FM4/y31dsс8dmB), яка містить велику інверсію та запирачі кросин-говеру [20]. Мутантних самців схрещували з віргінільними самками FM4.
Оскільки нейродегенеративні зміни у головному мозку виникають, головно, протягом пізнього онтогенезу, то синхронізовані культури було поставлено на ста-ріння і виготовлено гістологічні препарати з мозку 30-денних імаго. Серед 132 отриманих індивідуальних культур виявлено 25 культур зі змінами структур го-ловного мозку (табл. 3). Як видно з табл. 3, оптимальною в наших експериментах була концентрація 0,4 мкг/мл. Наприклад, у бруді при затравці цією концентра-цією отримано 14 культур зі змінами в мозковій тканині, у другому і третьому – по дві, тоді як при концентрації 2 мкг/мл отримано в першому бруді 4, а в другому та третьому - по одній культурі зі змінами структур мозку. За концентрації 4 мкг/мл одержано всього 12 культур, з яких в однієї виявлено нейродегенеративні зміни (за даними першого бруду). Зазначимо, що збідьшення концентрації мутагену не сприяло збільшенню частоти появи мутантів зі змінами структур мозку. На підста-ві даних табл. 3 можна зробити висновок, що найчутливішими до дії мутагену є сперматогонії та ранні сперматоцити, оскільки найбільшу кількість пошкоджень зареєстровано за даними першого бруду. Для підтвердження отриманих даних і усунення артефактів у подальших дослідженнях культури були повторно синхро-нізовані, поставлені на старіння і з мозку імаго були виготовлені гістологічні пре-парати. Зміни підтвердилися в 17 із 25 відібраних культур.
У літературі [22, 23, 24] описані мутації, які призводять до появи нейродеге-неративних змін у структурах мозку дрозофіли в процесі старіння. Доведено [22], що у мутантів swiss cheese (sws) мозкові дефекти виявляються у формуванні ваку-
Таблиця 3 Результати індукції НЕС змін у структурі головного мозку в лінії Oregon
Drosophila melanogaster Кількість препаратів зі змінами структур
мозку Концентрація НЕС, мкг/мл
Номер бруду
Кількість отриманих культур
Кількість гістологічних препаратів n %
0,4 1 60 30 14 46,7 2 20 15 2 13,3 3 10 6 2 33,3
2 1 20 15 4 26,7 2 6 5 1 20,0 3 4 4 1 25,0
4 1 7 5 1 20,0 2 3 2 0 0 3 2 1 0 0

Н. Голуб, І. Козаревська, Я. Черник 56
олей у всіх мозкових структурах. Ген swiss cheese локалізований у Х-хромосомі в районі 7D-1. Автори припускають, що продукт цього гена регулює гліальне обгор-тання нейронів, а мутація sws призводить до гіпермієлінізації. Друга досліджена нейродегенеративна мутація Vacuolar medulla (Vam) спричинює дегенерацію клі-тин глії, що веде до відмирання відповідних нейронів і формування вакуолей в окремих структурах головного мозку, зокрема в медулі [24]. У гомозигот Vam/Vam вакуолізація мозку постежується вже через 5 хв. після вилуплення. Серйозні пове-дінкові дефекти і дегенерація мозку, що виявляються через кілька днів після ви-льоту імаго, зумовлені Х-зчепленою рецесивною мутацією drop-dead [23]. Унаслі-док дослідження мозку дорослих мутантів виявлено, що багато гліальних клітин мають вкорочені відростки, тоді як морфологія нейрона є, головно, нормальною.
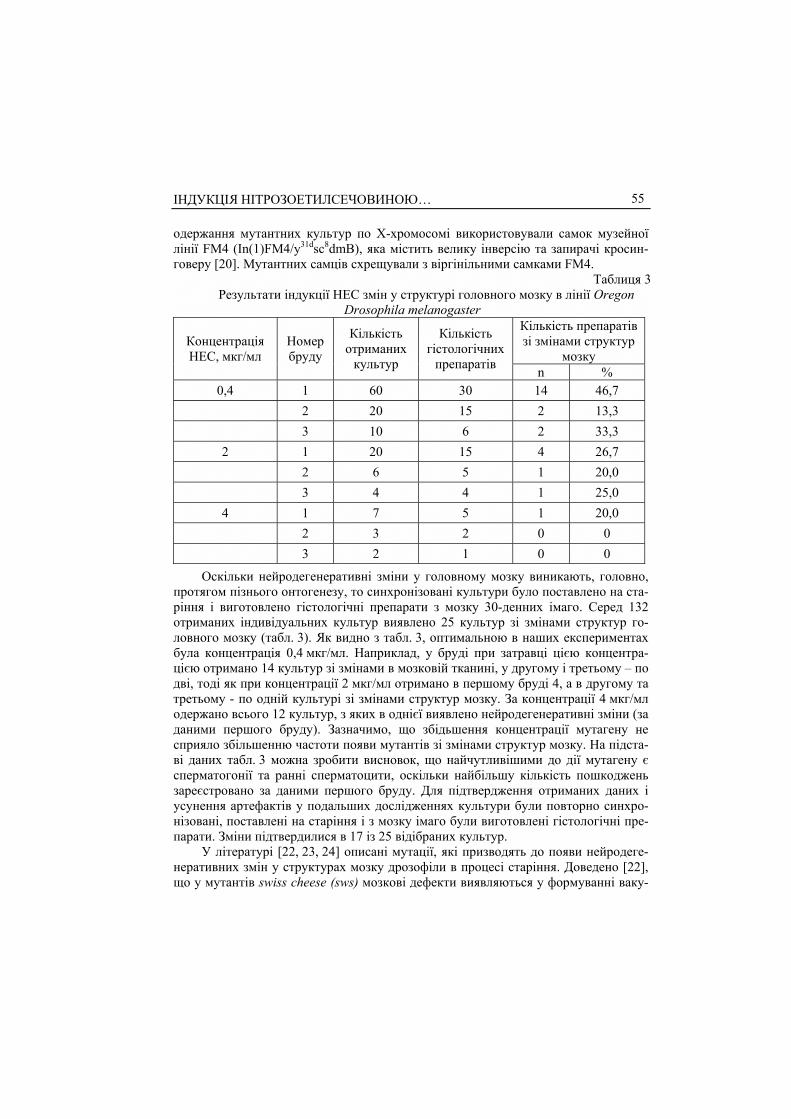
Отримані мутантні лінії були згруповані за характером розташування і розмі-ром вакуолей, що виникли у головному мозку дрозофіли (табл. 4, рис. 1, а-г).
З літератури відомо [23, 24] про різний час появи змін структур мозку протя-гом онтогенезу. Тому ми проаналізували виникнення таких змін у 5- та 10-денних мутантних особин. Дані наведені в табл. 5. З табл. 5 видно, що у ліній 5.1.1, 17.1.1, 31.3.2, 102.3.4 зміни відбуваються вже на п’ятий день після вильоту імаго і зберіга-ються у 10- та 30-денному віці. У ліній 4.1.1, 9.1.1, 45.1.1 поодинокі вакуолі почи-нають виникати у 10-денном віці. В особин ліній 33.1.1, 50.1.1, 60.1.1, 58.3.4 струк-турні зміни головного мозку з’являються лише на пізніх стадіях онтогенезу.
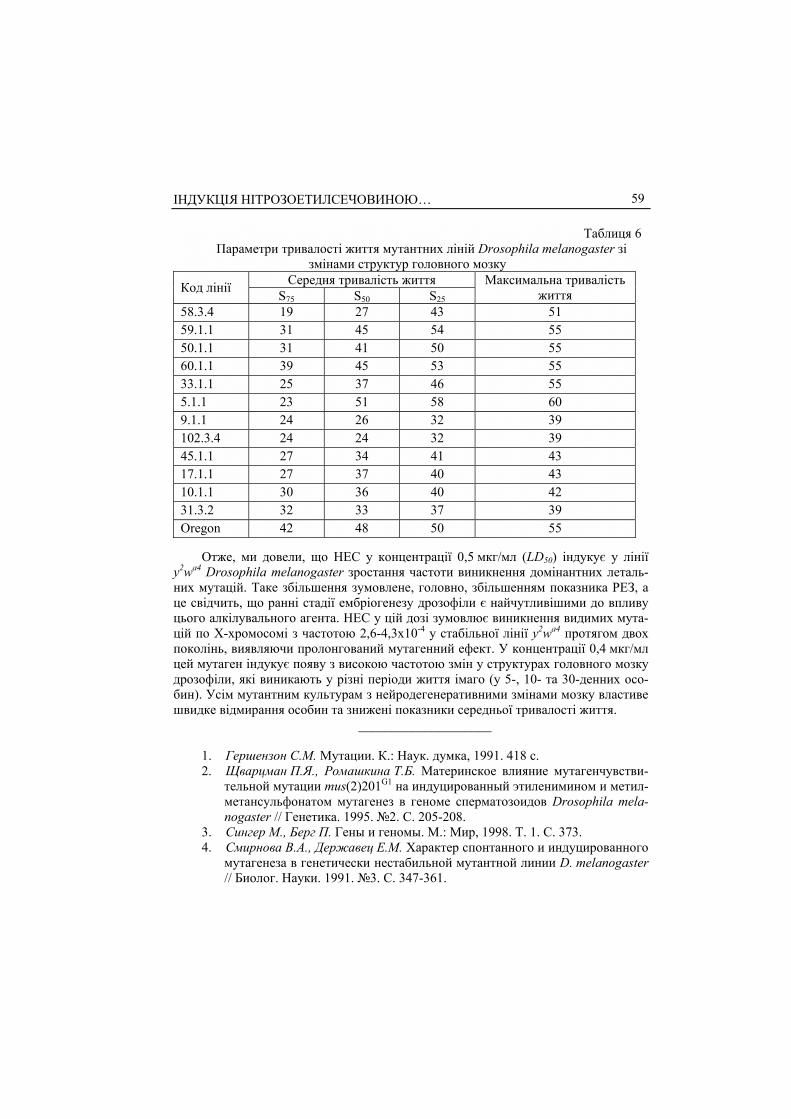
Відомо [22], що нейродегенеративні зміни спричиняють прискорений процес старіння. Тому цікаво було виявити кореляцію між часом появи змін і параметра-ми середньої тривалості життя мутантних ліній. Ми побудували криві виживання та проаналізували середню і максимальну тривалість життя мутантних ліній (табл. 6, рис. 2, а, б). Контролем слугувала КВ лінії дикого типу Oregon, параметри якої прийнято за нормальну диманіку старіння дрозофіли. Протягом 30 днів чи-сельність мух на КВ перевищувала 90%; з 30-го дня зафіксовано спад кривої, який свідчив про інтенсивне відмирання особин. Значення S75 і S50 становили 42 і 48 днів, відповідно; 25% живих мух було на 50-й день. Максимальна тривалість жит-тя дорівнювала 55 днів.
Таблиця 4 Групи мутантів за характером змін у структурі головного мозку дрозофіли
Код лінії Характер змін 9.1.1, 33.1, 4.1.1, 5,1.1, 50.1.1 Дрібні вакуолі по всій структурі мозку медулі 17.1.1, 45.1.1, 60.1.1 Поодинокі вакуолі по всій структурі мозку і
великих розмірів 31.3.2, 58.3.4, 102.3.4 Поодинокі великі вакуолі в різних ділянках
головного мозку
ІНДУКЦІЯ НІТРОЗОЕТИЛСЕЧОВИНОЮ… 57
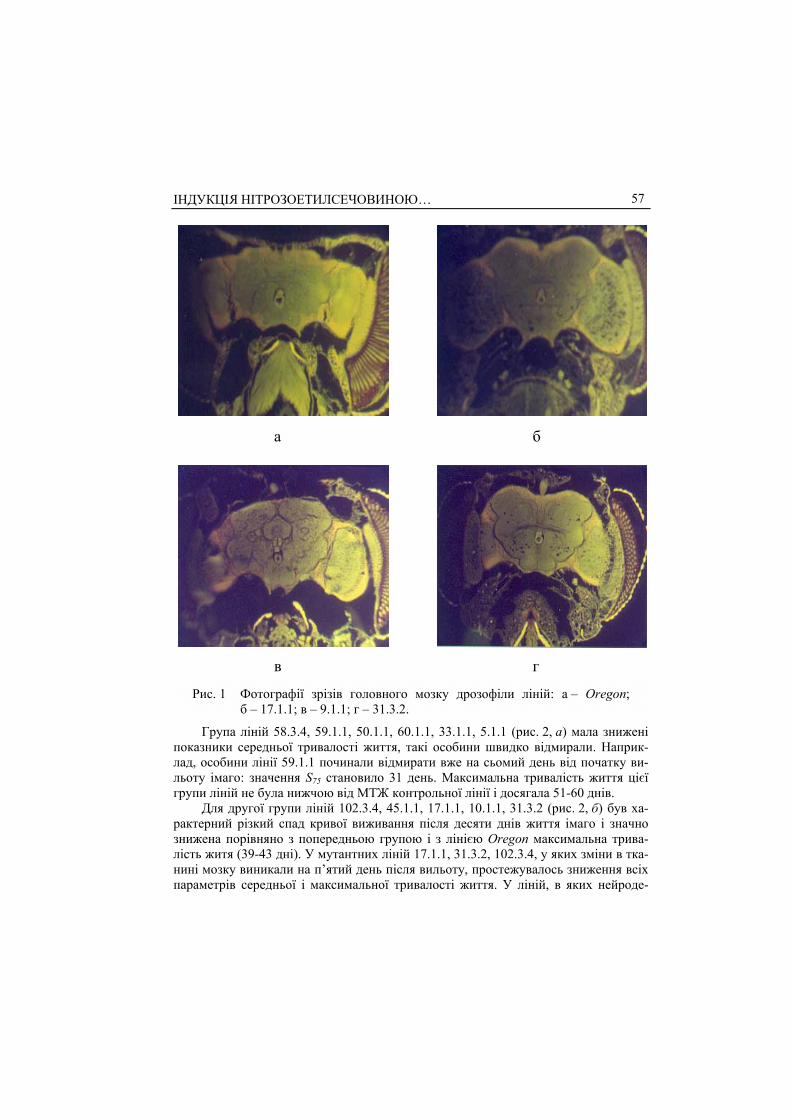
Група ліній 58.3.4, 59.1.1, 50.1.1, 60.1.1, 33.1.1, 5.1.1 (рис. 2, а) мала знижені показники середньої тривалості життя, такі особини швидко відмирали. Наприк-лад, особини лінії 59.1.1 починали відмирати вже на сьомий день від початку ви-льоту імаго: значення S75 становило 31 день. Максимальна тривалість життя цієї групи ліній не була нижчою від МТЖ контрольної лінії і досягала 51-60 днів.
Для другої групи ліній 102.3.4, 45.1.1, 17.1.1, 10.1.1, 31.3.2 (рис. 2, б) був ха-рактерний різкий спад кривої виживання після десяти днів життя імаго і значно знижена порівняно з попередньою групою і з лінією Oregon максимальна трива-лість житя (39-43 дні). У мутантних ліній 17.1.1, 31.3.2, 102.3.4, у яких зміни в тка-нині мозку виникали на п’ятий день після вильоту, простежувалось зниження всіх параметрів середньої і максимальної тривалості життя. У ліній, в яких нейроде-
а б
в г
Рис. 1 Фотографії зрізів головного мозку дрозофіли ліній: а – Oregon; б – 17.1.1; в – 9.1.1; г – 31.3.2.
Н. Голуб, І. Козаревська, Я. Черник 58
генеративні зміни починалися на десятий день життя імаго (4.1.1, 9.1.1, 45.1.1), ха-рактер кривої виживання та параметри тривалості життя такі ж, як і в ліній, у яких зміни в структурах мозку з’являлись у пізньому онтогенезі.
0
10
20
30
40
50
60
70
80
90
100
0 10 20 30 40 50 60дні
% – 1
– 2
– 3
– 4
– 5
– 6
– 7
Рис. 2. Криві виживання мутантних ліній дрозофіли зі змінами структур мозку: 1 – 58.3.4; 2 – 59.1.1; 3 – 50.1.1; 4 – 60.1.1; 5 – 33.1.1; 6 – 5.1.1; 7 – Oregon.
Таблиця 5 Динаміка виникнення індукованих НЕС змін у структурах головного мозку
лінії Oregon Drosophila melanogaster Вік імаго, дні Код лінії
5 10 4.1.1 - + 5.1.1 + + 9.1.1 - +
17.1.1 + + 33.1.1 - - 45.1.1 - + 50.1.1 - - 60.1.1 - - 31.3.2 + + 58.3.4 + -
102.3.4 + +
ІНДУКЦІЯ НІТРОЗОЕТИЛСЕЧОВИНОЮ… 59
Отже, ми довели, що НЕС у концентрації 0,5 мкг/мл (LD50) індукує у лінії y2wa4 Drosophila melanogaster зростання частоти виникнення домінантних леталь-них мутацій. Таке збільшення зумовлене, головно, збільшенням показника РЕЗ, а це свідчить, що ранні стадії ембріогенезу дрозофіли є найчутливішими до впливу цього алкілувального агента. НЕС у цій дозі зумовлює виникнення видимих мута-цій по Х-хромосомі з частотою 2,6-4,3х10-4 у стабільної лінії y2wa4 протягом двох поколінь, виявляючи пролонгований мутагенний ефект. У концентрації 0,4 мкг/мл цей мутаген індукує появу з високою частотою змін у структурах головного мозку дрозофіли, які виникають у різні періоди життя імаго (у 5-, 10- та 30-денних осо-бин). Усім мутантним культурам з нейродегенеративними змінами мозку властиве швидке відмирання особин та знижені показники середньої тривалості життя.
____________________
1. Гершензон С.М. Мутации. К.: Наук. думка, 1991. 418 c. 2. Щварцман П.Я., Ромашкина Т.Б. Материнское влияние мутагенчувстви-
тельной мутации mus(2)201G1 на индуцированный этиленимином и метил-метансульфонатом мутагенез в геноме сперматозоидов Drosophila mela-nogaster // Генетика. 1995. №2. С. 205-208.
3. Сингер М., Берг П. Гены и геномы. М.: Мир, 1998. Т. 1. С. 373. 4. Смирнова В.А., Державец Е.М. Характер спонтанного и индуцированного
мутагенеза в генетически нестабильной мутантной линии D. melanogaster // Биолог. Науки. 1991. №3. С. 347-361.
Таблиця 6 Параметри тривалості життя мутантних ліній Drosophila melanogaster зі
змінами структур головного мозку Середня тривалість життя Код лінії S75 S50 S25
Максимальна тривалість життя
58.3.4 19 27 43 51 59.1.1 31 45 54 55 50.1.1 31 41 50 55 60.1.1 39 45 53 55 33.1.1 25 37 46 55 5.1.1 23 51 58 60 9.1.1 24 26 32 39 102.3.4 24 24 32 39 45.1.1 27 34 41 43 17.1.1 27 37 40 43 10.1.1 30 36 40 42 31.3.2 32 33 37 39 Oregon 42 48 50 55

Н. Голуб, І. Козаревська, Я. Черник 60
5. Щербата Г.Р., Максымив Д.В. Молекулярно-генетическая природа мута-ций по локусу white, индуцированных химическими веществами у Droso-phila melanogaster // Генетика. 1997. Т. 33. №1. C. 19-24.
6. Pastnik A., Vreeken C., Vogel E.W. The nature of N-ethyl-N-nitrosourea induc-ed mutations at the white locus of Drosophila melanogaster // Mutat. Res. 1988. Vol. 199. №1. P. 47-53.
7. Rinchik E.M., Carpenter D.A, Selby P.B. A strategy for fine-structure function-al analysis of a 6-to 11-centimorgan region of mouse chromosome 7 by high-efficiency mutagenesis // Proc. Nat. Acad. Sci. USA. 1990. Vol. 87. P. 896-900.
8. Buzard G., Enomoto T., Perantoli A. et al. Neu-mutation in schamonas induced transplacentally in Syrian golden hamsters by N-nitrosoethylurea: high incid-ence but low allelic representation // Journal of Cancer Research and Clinical Oncology. 1999. Vol. 125. Is. 10. P. 529-540.
9. Rinchik E.M. Chemical mutagenesis and fine-structure functional analysis of the mouse genome // Trends Genet. 1991. Vol. 7. P. 15-21.
10. Itoh S., Miura M., Shimada H. Lack mutagenicity of levofloxacin in lacZ transgenic mice // Mutagenesis. 1998. Vol. 13. P. 51-55.
11. Dobrovolsky V.N., Casciano D.A., Heflich R.H. Mouse tk+/- model for in vivo mutagenicity study // Science Forum Poster Abstracts. FDA. 1997. Abstract b 32. P. 89.
12. Herns Anton J.M. et al. Methods for determining oncogenic potencial of che-mical compounds. // GenPharm. International, Inc. 1992. Vol. 29. P. 5, 174, 986.
13. Kessel M., Gruss P. Murine developmental control genes // Science. 1990. Vol. 249. P. 374-379.
14. Hayasaka K., Himoro M., Sato W. et al. Charcot-Marie-Tooth neuropathy ty-pe-1b is assotiated with mutations of the myelin-P(0) gene // Nat. Genet. 1993. Vol. 5. P. 31-34.
15. Ferrus A. Neurogenetics of Drosophila // Discussion in Neuroscience. 1992. Vol. 9. №1. P. 11-52.
16. Медведев В.П. Практическая генетика. М.: Наука, 1968. 294 c. 17. Ватти К В., Джапаридзе Л.А. Сравнительное изучение мутабильности
особей разных полов: РСПЛ и ДЛМ у Drosophila melanogaster // Генетика. 1980. №8. С. 1389-1395.
18. Lints F.A., Stoll J., Gruzer G. et al. An attemp to select for increased longevity in Drosophila melanogaster // Gerontology (Schweiz). 1979. Vol. 25. №4. P. 192-204.
19. Bergoffen J., Wang S., Scott M.O., et al. Connexin mutations in X-linked Char-cot-Marie-Tooth disease // Science. 1993. Vol. 262. P. 2039-2042.
20. Ashburner M. Drosophila. A laboratory handbook // Cold Spring Harbor Labo-ratory Press, Cold Spring Harbor. NY USA. 1989. Vol. 1331.

ІНДУКЦІЯ НІТРОЗОЕТИЛСЕЧОВИНОЮ… 61
21. Tosal l., Comendados M., Sierra L. N-ethyl-N-nitrosourea predominantly indu-ces mutations at the AT base pairs in pre-meiotic germ cells of Drosophila ma-les // Mutagenesis. 1998. Vol. 13. P. 375-380.
22. Kretzschmar D., Hasan G., Heisenberg M., Benzer S. The swiss cheese mutant glial hyperwrapping and brain degeneration in Drosophila // The Journal of Neuroscience. 1997. Vol. 17. № 19. P. 7425-7432.
23. Buchanan R.L., Benzer S. Defective glia in the Drosophila brain degeneration mutant drop-dead // Neuron. 1993. Vol. 10. P. 839-859.
24. Coombe P.E., Heisenberg M. The structural brain mutant Vacuollar medulla of Drosophila melanogaster with specific behavioral defects and cell degenera-tion in the adult // Neurogenetics. 1986. Vol. 3. P. 135-158.
INDUCTION OF DOMINANT LETHAL MUTATIONS AND X-LINKED VISIBLE MUTATIONS BY NITROSOETHYL UREA IN
DROSOPHILA MELANOGASTER
N. Holub, I. Kozarevska, Ya. Chernyk Ivan Franko National University of L’viv, Hrushevskogo St. 4, L’viv 79005, Ukraine,
e-mail: [email protected]
The influence of nitrosoethylurea (ENU) on induction of dominant lethal mutations (DLM), appearance of visible mutants and mutants with changes in brain structures link-ed to X-chromosome of Drosophila melanogaster have been investigated. It was shown that ENU in dose of 0,5 mkg/ml (LD50 for laboratory strain y2wa4) leads to 6 times incre-asing the frequency of DLM appearance comparing with control. This concentration of ENU causes the appearance of visible mutants in X-chromosome with 2,6x10-4-4,3x10-4 frequency within two generations. In concentration 0,4 mkg/ml mutagene induces the high frequency of changes in brain structures of Drosophila which appeare in different periods of imago life (in the age of 5, 10 and 30 days). All mutants cultures with neuro-degenerative brain changes are characterized by flies rapid dying off and reduced inde-xes of average life span.
Key words: drosophila, nitrosoethylurea, DLM, mutagenesis, neurodegeneration.
Стаття надійшла до редколегії 25.12.2001 Прийнята до друку 01.02.2002
Related Documents











