Page | 1 Thesis for the degree Doctor of Philosophy By Yoav Politi Advisor: Prof. Eli Arama January, 2018 Submitted to the Scientific Council of the Weizmann Institute of Science Rehovot, Israel הרס המיטוכונדריה האבהית לאחר הפריה מתווך על ידי מסלול אנדוציט י ו אוטופאגי משותף בדרוזופילהPaternal mitochondrial destruction after fertilization is mediated by a common endocytic and autophagic pathway in Drosophila לתואר)תזה( עבודת גמר דוקטור לפילוסופיה מאת יואב פוליתי טבת, תשע"ח מוגשת למועצה המדעית של מכון ויצמן למדע רחובות, ישראל מנח ה: פרופ' אלי ארמה

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

P a g e | 1
Thesis for the degree
Doctor of Philosophy
By
Yoav Politi
Advisor:
Prof. Eli Arama
January, 2018
Submitted to the Scientific Council of the
Weizmann Institute of Science
Rehovot, Israel
י מתווך על ידי מסלול אנדוציט הרס המיטוכונדריה האבהית לאחר הפריה
בדרוזופילהמשותף אוטופאגי ו
Paternal mitochondrial destruction after fertilization is mediated by
a common endocytic and autophagic pathway in Drosophila
עבודת גמר )תזה( לתואר
דוקטור לפילוסופיה
מאת
יואב פוליתי
תשע"ח ,טבת
מוגשת למועצה המדעית של
מכון ויצמן למדע
רחובות, ישראל
:המנח
פרופ' אלי ארמה

P a g e | 2
Acknowledgements
First and foremost I wish to express my deepest thanks to my mentor Eli Arama. Boss, I am not sure
if you remember but at first you didn't have room for me in the lab for the 1st and 2nd rotation. But
something inside me told me to insist and I am thankful for that. Thank you for giving me the
opportunity to prove myself. Thank you for moving me to Kalifa's project and for letting me and
Liron lead the project to publication and beyond. Thanks for giving me the chance to explore, gain
new skills and find other areas of interest. Thanks for never holding me back, method or money-wise.
Thank you for allowing me to take part in your academic career, at its critical point around your
tenure promotion. Thank you for the time with you during conferences, trips, group meetings and
other occasions. I can't think of a better place to do my Ph.D., meet such great people and acquire
lifetime friends. I have to say here that at first I wasn’t so sure about my skills and ability to do
Masters, to do it at the Weizmann, and I am not even speaking about Ph.D. But here I am, 8 years
later, and about to finish my Ph.D., with three lovely daughters, and less hair on my head. I am
grateful from the bottom of my heart for the chance to take the ride of my life. When I doubt myself
from time to time I remember the Seinfeld episode where George start working in a new office, and
his boss is not sure that he is made of the “Penske material”. So, maybe a little out of the episode’s
context, I am glad to say that today I fill made of “Arama material”. I wish you many more years of
making meaningful science, self-fulfillment and good health above all.
I am grateful for Yossi Kalifa, who took me under his supervision and basically taught me everything
I needed to continue with this project, and practically handed over a work at its peak. Thank you for
the good advices throughout the years, not only considering our project, but also in other aspects of
my life, and for keeping in touch despite the physical distance.
To my partner in the lab Liron Gal. Lironch, my endless thanks to you, for completing me as a
scientist, for taking me under your wings as a rotation student when I just came to the lab, for
persuading me to join Yossi Kalifa. Thank you for jumping on the PMD wagon without too many
hesitations. For showing how motivation can lead you to the moon and back (if you only wanted).
Thanks for shaping most of my time in the Weizmann as it was. Thanks for your laughter and smile
and for being a great friend. All the luck in which summit you will decide to conquer!
To my wonderful aisle body Keren Yacobi-Sharon. Karnun, thank you for being my best friend in
the lab – much more than just a lab-neighbor. Thank you for sharing with me these years, your

P a g e | 3
thoughts and emotions, I can't think of a better compliment than that! Thank you for your calmness,
your good spirit and realistic view of the world.
To the generation of giants, three Ph.D. musketeers who were there when it all started: the legendary
Yossi Kaplan, whose reputation precedes him, master of confocal, cloning, Western blot and
generally comprehensive work, thanks for your cynicism, Yiddish lessons and for serving as a role
model. Anat Florentin, who've been a PI already during her Ph.D. Thanks for your good advices and
enormous knowledge. Liat Ravid-Lustig, both sarcastic and sensitive (she can laugh and cry in a
single sound – Springsteen), thank you for all your help and support, and for being a good friend
throughout the years. Best of luck to the founders wherever you are abroad, keep exceling and make
us proud.
To former lab members that I joyfully spent many years in the lab:
Lior Aram, thank you for being there from my second day in the lab back then as a rotation student.
For being an example of how science should be done – marking targets and hitting them straight
between the eyes. For showing me how to bring skills to perfection. And for being a good friend, as
both of us entered the lab as newlywed and came out with lovely girls.
Carmel Braverman, thank you for your smile, your laughter and your wonderful spirit. Thank you
for coming back to the lab after maternity leave, and I wish you lots of success in the rest of your
Ph.D., and generally in life.
To all my beloved current lab members, for being the faces I enjoy meeting every day:
Anna Gorelick Ashkenazi, whom we run neck to neck throughout our Ph.D. Thank you for your
endless care and constant thoughtfulness, showing how things can be done with lots of thought and
close consideration.
Tslil Braun, which I am proud of for being my successor as a rotation student that continued to
Masters and for Ph.D., in the lab. Thank you for your easy going spirit, calmness and smile. Best of
luck in the following years.
Alina Kolpakova, thank you for always willing to help, assists in all sorts of requests and for being a
good friend outside of the lab, sharing the affection for gardening and cats.
Ron Weiss, thank you for showing me the meaning of hard work, for your friendship and for sharing
with me (and Eli) the men side of the lab.
Lama Tarayrah, thank you for sharing the embryonal side of the lab with me and for the nice
conversations. Thanks also for your realistic yet optimistic way of looking at science and life in
general.

P a g e | 4
Lena Sapozhnikov, thank you for being a great lab-roommate for the past months, for your calm
mood and for being a true animal lover.
Agur Wiscott, thank you for bringing fresh spirit to the lab, for your dedicated and thorough work.
Always willing to help and learn new stuff. I hope you will proceed in the academia, not as a
technician, but rather as a masters student.
To Nechama Rakow and Jordana Lindner-Ovadia, thank you for making the right decision and
coming to the Arama group. I wish you lots of success and long years at the Weizmann.
To Raz Eliav and Eitan Waltman, for sharing our Master’s period together, for the support and good
vibe.
I would like to thank my precious former-rotation students (despite the fact they didn't stay for
Masters in the lab), Bat-Chen Tamim-Yecheskel, Hanoch Templehof, Maya Ron and Mirit Biton.
Thank you for assisting my project, for forcing me to improve and for letting me to be a part of your
academic career. All the best and good luck in the future.
To my Ph.D. committee members, Ami Navon and Michael Glickman, thank you for your time and
attention, for your good perspective and fruitful advices.
Warm thanks for Shmuel Pietrokovsky for data analyses, good perspective and endless will to assist.
Many thanks to Shari Carmon for willing to share her tremendous knowledge, experience and also
reagents with me.
My sincere gratitude to the electron microscopy unit in the WIS, for serving as a second lab for me in
numerous periods during my Ph.D studies. Especially I wish to thank Vera Shinder for her close
guidance and practically enabling me to conduct EM work from A to Z. I also thank Smadar
Zaidman, Eyal Shimoni, Katya Rechav, Tali Dadosh, Hancoch and Talmon for occasional intimate
work, technical support and lots of patience.
I would like to thank Yoav Peleg, Shira Albeck and Josef Jacobovitch from the Center for structural
proteomics (ISPC), for their immense assistance in cloning and protein expression procedures, for
their willingness to help although I was officially a running project in the unit.
I thank Yishai Levin and his staff from the mass spectrometry unit in the G-INCPM, for performing
our mass spec' experiments fast and with professionalism.
Many thanks for Ugo Mayor and his lab for their patience, close attention and professionalism. I am
sure this collaboration will bear many fruits in the future.

P a g e | 5
I would like to thank all our colleagues who kindly shared fly lines and antibodies with us: Uri Adbu,
Ben-Zion Shilo, Avraham Yaron, Maya Schuldiner, Oren Schuldiner, Orly Reiner, Yael
Heifetz, Lilach Gilboa, Uri Abdu, Gabor Juhasz, Hugo Bellen, Marie-He´le`ne Bre´, Aaron
DiAntonio, Se´bastien Gaumer, Thomas P. Neufeld, Ioannis P. Nezis, Leo J. Pallanck, Kim
Finley, Tor-Erik Rusten, Hubert J. Amrein, Timothy T. Weil, Pernille Rörth, Fumiko
Kawasaki, Troy J. Littleton, Lily and Yuh Nung Jan, Sebastian Rümpf, Suzanne Eaton, Clive
Wilson, Junichi Nakai, Masamichi Ohkura, Ronald Kühnlein,
We thank the TRiP at Harvard Medical School (NIH/National Institute of General Medical Sciences
grant R01-GM084947) for providing the transgenic RNAi fly stocks used in this study. We thank the
Bloomington Drosophila stock center, Vienna Drosophila resource center (VDRC) and Kyoto stock
center (DGRC) for fly lines and plasmids.
Last but not least, I wish to thank my beloved family. First, my beautiful girls, Tamari, Aya and Libi –
my most precious thing in the world, for your hugs and smiles, and for taking care of me as much as I
take care of you. To my parents Ofra and Chaim, to my siblings Yael and Ori, to my grandparents
Shaul, Hanna and Tmima. Thank you for your constant support and for providing carrying
surroundings. Finally, to my wife Maya, the love of my life, the one that always puts sense in
everything and organizes my thoughts. You never had even the slightest doubt in me. You lifted me to
where I am today, not only educationally, but in all life aspects. The thing that make you so great and
admired person by your surroundings, and the fact that I am your partner in life, actually means as a
consequence a lot about me, and that is the biggest compliment I have ever received.

P a g e | 6
Declaration
I declare that this thesis summarizes my work. Some parts were done in collaboration with other
people, as described in the following. First, this work has been done in collaboration with Dr. Liron
Gal, a former graduate student in the lab. The data shown in sections 5.1 to 5.9 was published with
Liron being a co-first author (Politi et al., 2014). Dr. Yossi Kalifa was involved in the initial steps of
the project, including the establishment of the live imaging assay and the early electron microscope
analyses. Dr. Liat Ravid-Lustig generated the genetic system for inactivation of Parkin during late
spermatogenesis. For the electron microscopy work, I was assisted by Dr. Vera Shinder, Dr. Smadar
Zaidman, Dr Katya Rechav and Dr. Eyal Shimoni in the Irving and Cherna Moskowitz Center for
Nano and Bio-Nano Imaging at the WIS. Matlab code for live imaging quantifications was written by
Ofra Golani from the biological services unit at the WIS. Initial K63 ubiquitin staining was performed
together with Bat-Chen Tamim-Yecheskel (a former rotation student in the lab). Atg2 knockout
experiments were performed together with Hanoch Tempelhof (a former rotation student in the lab).
For the Mitotracker labeling procedures I was assisted by Maya Ron (a former rotation student in the
lab). For the Mul1 and Parkin live imaging experiments I was assisted by Mirit Biton (a former
rotation student in the lab). SNARE knockdown experiments were performed together with Ilana
Weinstein and Dominik Etter (summer students). Mass spectrometry for the detection of sperm
mitochondrial proteins in early fertilized eggs was done by Dr. Yishai Levin in the proteomics unit at
the G-INCPM, WIS. Mass spectrometry of extracted MVBs was performed by Prof. Arie Admon at
the Technion. The Ub6-BirA project is performed in collaboration with the lab of Prof. Ugo Mayor,
CIC Biogune, Spain. Pre proteomics data analyzing was done by Prof. Shmuel Pietrokovsky, WIS.

P a g e | 7
Contents
1. Abbreviations ............................................................................................................................................... 9
2. Abstract ...................................................................................................................................................... 10
3. Introduction ............................................................................................................................................... 12
3.1. Mitochondrial inheritance ................................................................................................................ 12
3.2. Mitochondrial turnover .................................................................................................................... 13
3.3. Autophagy .......................................................................................................................................... 14
3.4. Multivesicular bodies (MVBs) .......................................................................................................... 14
4. Materials and methods .............................................................................................................................. 16
4.1. Fly strains ........................................................................................................................................... 16
4.2. Cloning ............................................................................................................................................... 17
4.3. Ultrastructural studies ...................................................................................................................... 21
4.4. Live imaging studies .......................................................................................................................... 22
4.5. Immunostaining ................................................................................................................................. 22
4.6. Western blots ..................................................................................................................................... 23
4.7. Gastrulation assay ............................................................................................................................. 23
4.8. RNA isolation and RT-PCR.............................................................................................................. 24
4.9. Quantification and statistics analyses of PMD kinetics .................................................................. 24
4.10. Mass Spectrometry ........................................................................................................................ 24
4.11. MVB purifications ......................................................................................................................... 25
5. Results ......................................................................................................................................................... 27
5.1. Ultrastructure of the PMD process in Drosophila .......................................................................... 27
5.2. The PMD kinetics .............................................................................................................................. 31
5.3. Autophagy-related vesicles associate with the sperm MD soon after fertilization ...................... 33
5.4. PMD is mediated by a network of vesicles displaying markers common to the autophagic and
endocytic pathways ........................................................................................................................................ 35
5.5. Inactivating mutations in the autophagic and endocytic pathways attenuate PMD ................... 37
5.6. The paternal MD is ubiquitinated through lysine-63-linked polyubiquitin side chains soon after
fertilization ..................................................................................................................................................... 42
5.7. The Ub-binding autophagy receptor p62 is recruited to the paternal MD .................................. 46
5.8. The ubiquitin/p62 pathway is involved in PMD ............................................................................. 46
5.9. The sperm MD is depolarized........................................................................................................... 47
5.10. Loss of Mul1 and Parkin significantly attenuates PMD ............................................................ 51
5.11. Prohibitin, a major MD ubiquitination substrate after fertilization ........................................ 58

P a g e | 8
5.12. Investigate the role of the MVB-like vesicles in the destruction of the paternal mitochondria
64
5.13. The role of the mitochondria fusion/fission machinery on PMD .............................................. 73
6. Discussion ................................................................................................................................................... 75
6.1. A working model for PMD, hypotheses and open questions ......................................................... 75
6.2. The significance of paternal mitochondrial ubiquitination and association with p62 and
LC3/Atg8 ........................................................................................................................................................ 77
6.3. The role of the MVBs and their microvesicles in PMD .................................................................. 78
6.4. Why is the paternal mitochondria eliminated after fertilization? ................................................ 79

P a g e | 9
1. Abbreviations
Mitochondrial derivative (MD), multivesicular bodies (MVBs), intraluminal vesicles (ILVs), paternal
mitochondrial destruction (PMD), deubiquitinating enzyme (DUB), immunoprecipitation (IP), K63
(Lysine 63), mtDNA (mitochondrial DNA), PCR (polymerase chain reaction), AEL (after egg laying),
MTS (mitochondrial targeting signal), RF (restriction free), TEV (tobacco etch virus), DDW
(deionized distilled water), EM (electron microscope), TEM (transmission electron microscope), SEM
(scanning electron microscope), FIB (focused ion beam), PBS (phosphate buffer solution), Axo
(axoneme), DJ (Don-juan), WIS (Weizmann Institute of Science), MVC (multivesicular cluster),
MLB (multilamellar body), MF (mitochondrial fragments), GFP (green fluorescent protein), ESCRT
(endosomal sorting complexes required for transport), YFP (yellow fluorescent protein), RNAi (RNA
interference), TRiP (transgenic RNAi project), shRNA (short hairpin RNA), RT-PCR (reverse
transcriptase PCR), DN (dominant negative), LAP (LC3 associated phagocytosis), TP (time point),
polyUb (poly ubiquitin), UAS (upstream activation sequence), LC3 (light chain 3), LIR (LC3
interaction region), Atg (autophagy related gene), UVrag (UV radiation resistance associated), Rab
(Ras-related GTP binding protein), VPS (vacuolar protein sorting), Ref(2)p (refractory to sigma P),
Hrs (hepatocyte growth factor-regulated tyrosine kinase substrate), Stam (signal transducing adaptor
molecule), Hdac6 (histone deacetylase 6), Key (Kenny), Chip (carboxyl terminus of Hsc70-interacting
protein), Sina (Seven in absensia), Traf6 (TNF (Tumor necrosis factor) receptor associated factor 6),
Vti1 (vps 10 (ten) interacting 1), Snap29 (synaptosome associated protein 29), Vamp7 (vesicle
associated membrane protein 7), Tsg101 (tumor susceptibility gene 101), Dor (deep orange), Car
(carnation), Uba (ubiquitin like modifier activating enzyme), VCP (valosin containing protein),
tbc1d15/17 (TBC1 (Tre2, Bub2, Cdc16) domain family member 15/17), Drp1 (dynamin related
protein 1), Fzo (fuzzy onions), Marf (mitochondrial assembly regulatory factor), UBAN (ubiquitin
binding in Abin and Nemo), SpnF (spindle F), dBruce (Drosophila BIR (Baculovirus inhibition of
apoptosis protein repeat) repeat containing ubiquitin conjugating enzyme), CRISPR (clustered
regularly interspaced short palindromic repeats), NS (no significance), UTR (untranslated region),
BAC (bacterial artificial chromosome), GAP (GTPase activation protein), kDa (kilo Dalton), MS
(mass spectrometry), SNARE (SNAP (soluble N-ethylmaleimide-sensitive fusion (NSF)) attachment
protein receptors), GCD (germ cell death), IAP (inhibitor of apoptosis protein), TMRE
(Tetramethylrhodamine, Ethyl Ester), MTR (mitotracker), EPS (embryo permeabilization solvent),
SRM (selective reaction monitoring), SILAC (stable isotope labeling by/with amino acids in cell
culture), ND (not detected), GO (gene ontology), EV (extracellular vesicle).

P a g e | 10
2. Abstract
Almost all animals contain mitochondria of maternal origin only, but the exact mechanisms
underlying this phenomenon are still vague. We investigated the fate of the Drosophila paternal
mitochondria after fertilization. We demonstrate that the sperm mitochondrial derivative (MD) is
rapidly eliminated in a stereotypical process, which we dubbed paternal mitochondrial destruction
(PMD). PMD is initiated by a network of vesicles, resembling multivesicular bodies and displaying
common features of the endocytosis and autophagy pathways. These vesicles associate with the sperm
tail and mediate the disintegration of its plasma membrane. Subsequently, the MD separates from the
axoneme and breaks into smaller fragments, which are then sequestered by autophagosomes for
degradation in lysosomes. We further provide evidence for the involvement of the ubiquitin pathway
and the autophagy receptor p62 in this process. We show that the ubiquitin E3 ligases, Parkin and
Mul1, are both involved in PMD, and that their source is both paternal and maternal. We provide
genetic evidence that the abundant inner mitochondrial membrane protein, Prohibitin1 (Phb1), could
be a target for ubiquitination on the sperm mitochondrial derivative after fertilization. In parallel, we
also take a proteomics approach aiming to detect additional MD ubiquitinated proteins involved in
targeting this organelle for destruction. Furthermore, we devise a combined method to isolate intra-
egg MVBs for proteomics analysis. Collectively, this work sheds light on the highly conserved
process of paternal mitochondrial destruction in organisms with flagellated sperm. Considering the
high molecular and conceptual similarity between PMD and other (mainly stress-induced) paradigms
of selective organelle destruction, our work also provides a framework for the discovery of new
factors that could be involved in the selectivity and elimination (recycling) of damaged organelles
during normal cell homeostasis.

P a g e | 11
תקציר
מיטוכונדריה שלהם מהאם, אך המנגנונים המדויקים המסבירים תופעה זו ה מקבלים אתבעלי החיים למעשה כל
בזבוב לאחר ההפריה של המיטוכונדריה האבהית במהלך עבודת הדוקטורט, חקרתי את תהליך ההיעלמות . עדיין אינם ברורים
מובנה שקראנו לובמהירות בתהליך תמסולקשל הזרע ה המוארכתשהמיטוכונדרימצאנו דרוזופילה מלנוגסטר. התסיסה,
paternal mitochondrial destruction אוPMD .בעלות מאפיינים משותפים למערכות שלפוחיותכשתהליך מתחיל ה
זנב ממברנת במגע עם (, באותMVBsאו multivesicular bodiesהאנדוציטית והאוטופאג'ית, ומקורן בביצית )מכונות גם
הסרת הממברנה הפלסמתית של הזרעלתוך הזרע. תהליך זה מוביל ל (microvesicles) התוכן שלהןאת ומפרישות הזרע
לחתיכות תנשברהמוארכת ההמיטוכונדרי .האקסונים - של הזרעהשילדי המוארך מהאברון ההמיטוכונדרי והיפרדות של
מעורבות של מערכת היוביקוויטין מצאנו בנוסף, בליזוזום. לבסוף מפורקיםאלו על ידי אוטופאגוזומים ו נאספותאשר ותקטנ
בהרס המיטוכונדריה האבהית. הראנו שליגאזות , p62 ליוביקוויטין, אוטופאג'יהמתווך בין מערכת הושל הקולטן
. סיפקנו עדויות בזרעוגם בביצית ןבהרס המיטוכונדריה האבהית, ושמקור ותמעורבגם הן ,Mul1 ו Parkinהיוביקוויטין
המצוי ברמות גבוהות בממברנה הפנימית של מיטוכונדריות, יכול להוות מטרה Prohibitin1גנטיות לכך שהחלבון
,. במקביל, סמן המאפשר הרס ספציפי של המיטוכונדריה האבהיתשל הזרע לאחר הפרייההמיטוכונדריה ליוביקוויטינציה על
עוברים יוביקוויטינציה הו תהאבהי הלזהות חלבונים נוספים השייכים למיטוכונדרי בכדיגישה פרוטאומית בגם נוקטים אנו
מתוך הביצית המופרית MVBsה שיטה לבידוד פיתחנו את האברון הזה לפירוק. זאת ועוד, לסמן במטרהלאחר ההפריה
ריה האבהית לאחר הפריה לשם ניתוח פרוטאומי של תכולתם. לסיכום, עבודה זו שופכת אור על תהליך פירוק המיטוכונד
והעקרוני יצורים בהם יש זרע בעל שוטון. בהתחשב בדמיון המולקולרי בדרוזופילה היכול לשמש כפרוטוטיפ של תהליך זה ב
עקה(, בעקבות )בעיקר בתא של אברונים םסלקטיביומחזור הרס תהליכיהרס המיטוכונדריה האבהית לבין תהליך בין
לגילוי מרכיבים נוספים אשר עשויים להיות מעורבים בסלקטיביות גם מודל מחקר בדרוזופילה מהווה המיטוכונדריה האבהית
תאים.ב הומאוסטזיס על שמירהובפירוק של אברונים פגומים במהלך

P a g e | 12
3. Introduction
3.1. Mitochondrial inheritance
Almost all eukaryotic organisms inherit their mitochondria, the primary cellular power plants from
the maternal parent. Although this renowned phenomenon has been widely accepted among
geneticists and developmental biologists, the striking variation among different organisms in the
extent and pattern of uniparental inheritance has led to a diverse number of proposed mechanistic
models for this phenomenon and thus has been the cause of much uncertainty (Ankel-Simons and
Cummins, 1996; Birky, Jr., 2001). Until recently, the prevailing mechanistic explanation for maternal
mitochondrial inheritance has been a passive model of simple dilution of the paternal mitochondria by
an excess copy number of the egg mitochondria (Gyllensten et al., 1991; Birky, Jr., 2001). This
explanation is mainly based on a study which used unique mitochondrial (mt)DNA nucleotide
identifiers to demonstrate that paternally inherited mtDNA molecules can be detected in low
frequency in hybrid mice, suggesting that paternal contribution exist, but that it may often be
overshadowed in mature mice to a point beyond the limits of detection by conventional PCR analyses
(Gyllensten et al., 1991). On the other hand, recent studies in C. elegans have reported the active
involvement of autophagy in this process. As opposed to organisms with flagellated sperm, in which
the mitochondria undergo unique structural remodeling to become part of the flagellum, C. elegans
produces non-flagellated amoeboid sperm with mitochondria of much simpler morphology (Al Rawi
S. et al., 2011; Sato and Sato, 2011; Levine and Elazar, 2011). Furthermore, recent findings show that
the autophagy receptor, p62, and the ubiquitin-like modifier of autophagy LC3 colocalize to the sperm
tail after fertilization in mice (Al Rawi S. et al., 2011). More recent reports on the mouse model
further added to this controversy, suggesting that the sperm mitochondria are not degraded by
autophagy, an observation that was based on a single autophagy gene mutant, and that the sperm
mtDNA is already eliminated before fertilization, while the paternal mitochondria are unevenly
distributed in a mosaic fashion in the early embryo (Luo et al., 2013). However, this study left
unexplained the observation showing that p62 and LC3 are localized on the sperm mitochondria soon
after fertilization (Luo et al., 2013). In contrast, a more recent study demonstrated that PMD in mouse
is an active process involving specific E3 ubiquitin ligases, the autophagy adaptor p62 and loss of
sperm mitochondrial membrane potential after fertilization (Rojansky et al., 2016). Intriguingly,
several studies in Drosophila, fish and mouse models, showed that the sperm mitochondrial DNA
(mtDNA) is degraded already before or soon after fertilization (Nishimura et al., 2006; DeLuca and
O'Farrell, 2012; Luo et al., 2013). Since preventing heteroplasmy (the presence of more than one type

P a g e | 13
of mitochondrial genome within a cell) is the prevailing reason, among researchers in the field, as for
why the paternal mitochondria is targeted for destruction after fertilization, these findings raise
interesting questions regarding the reason and the need to also target the vacuolated (mtDNA-less)
paternal mitochondria. Furthermore, the controversy about the passive and active mechanisms in the
mouse brings up the need for additional studies of PMD in other, genetically amenable, organisms
with flagellated sperm, such as the Drosophila.
3.2. Mitochondrial turnover
The ability of cells to monitor the quality of the mitochondria and control their turnover is
increasingly recognized as an essential element in maintaining mitochondrial homeostasis (de Castro
et al., 2010; Weber and Reichert, 2010). Mitochondrial quality control depends upon a balance
between biogenesis, dynamics (fusion and fission), and selective turnover (Schafer and Reichert,
2009; Twig et al., 2008; Ashrafi and Schwarz, 2013). However, despite its important role in quality
control, the molecular mechanisms underlying selective mitochondrial destruction are still not well
understood (Wang and Klionsky, 2011). Accumulating data suggest the involvement of the ubiquitin
pathway in the selective recognition and autophagy-mediated destruction of cytosolic microbes,
protein aggregates and damaged organelles (Johansen and Lamark, 2011; Knaevelsrud and Simonsen,
2010; Kirkin et al., 2009b; Narendra et al., 2008; Randow, 2011). However, the function of the
autophagic machinery and the ubiquitin pathway in normal mitochondrial turnover (a process also
dubbed mitophagy) and their roles during development are still vague (Ashrafi and Schwarz, 2013).
Whereas in some examples, the mammalian sperm mitochondria were shown to be ubiquitinated after
fertilization, (Sutovsky et al., 1999; Sutovsky et al., 2000; Al Rawi S. et al., 2011; Luo et al., 2013),
other studies in C. elegans and mice have suggested that paternal mitochondria are either not
ubiquitinated (Sato and Sato, 2011) or that the ubiquitination was not restricted to the sperm
mitochondria only (Luo et al., 2013), respectively. The realization that some neurodegenerative
diseases, such as Parkinson’s disease, might be caused by the failure to normally degrade and recycle
damaged mitochondria, raised new interest in the research of organelle turnover (Franco-Iborra et al.,
2016; Celardo et al., 2014). However, while highly informative, most of the investigated paradigms
involve the induction of massive damage to the cell mitochondria in culture, which raise justified
questions regarding their relevance to normal cell homeostasis and pathology (Narendra et al., 2008;
Narendra et al., 2010b). The establishment of a genetically amenable system to monitor and study

P a g e | 14
selective mitochondrial destruction under physiological conditions shall therefore help to fill in the
gaps in our understanding of this important process.
3.3. Autophagy
Autophagy is a highly regulated membrane-mediated intracellular degradation process, ubiquitous
in eukaryotic cells. During autophagy, double-membrane vesicles, called autophagosomes, engulf
cytoplasmic materials, including proteins and organelles, and deliver them to the lysosome for
degradation (Nakatogawa et al., 2009; Rubinsztein et al., 2012; He and Klionsky, 2009). The recycled
products are subjected to cellular metabolism for production of energy and to build new proteins and
membranes (Rabinowitz and White, 2010). In addition to its fundamental role in metabolic adaptation
during cell homeostasis, autophagy also functions in cell growth, survival, cell death, and as an
intracellular quality control system (Neufeld and Baehrecke, 2008). More recent studies have
highlighted the importance of selective autophagy in mediating the turnover of specific unwanted
cargo, including damaged organelles (Mizushima and Komatsu, 2011; Weidberg et al., 2011;
Sumpter, Jr. and Levine, 2011). However, whereas selective clearance of organelles resembles an
autophagy-related process at the anatomical level, genetic, molecular and cellular studies have
indicated some divergence from the core autophagic machinery (Xie and Klionsky, 2007; Manjithaya
et al., 2010; Farre et al., 2009; Zhang et al., 2009).
3.4. Multivesicular bodies (MVBs)
Endocytosis is a form of active transport in which the cell transports molecules into its inner part
by engulfing them in an energy-dependent process. Plasma membrane proteins, such as receptors, that
are destined to degrade, are incorporated into intraluminal vesicles (ILVs), giving rise to the formation
of late endosome compartments also known as multivesicular bodies (MVBs), which can then fuse
with lysosomes, leading to the degradation of the ILVs and their contents (Piper 2010). The
biogenesis of the MVBs is dependent on complexes called ESCRTs (endosomal sorting complexes
required for transport). Each one of the five ESCRT complexes is composed of 2 to 4 subunits,
including ubiquitin binding protein, that can recognize ubiquitinated cargos and in turn recruit other
complex components to facilitate membrane invagination and closure of ILVs (Filimonenko et al.,
2007; Schmidt and Teis, 2012). Together with their role as intracellular cargo sorters from various
membranes to the lysosome or back to their origin, the MVBs can also fuse to the plasma membrane
and release their micro-vesicles to the extracellular domain as exosomes, thereby releasing cargo from

P a g e | 15
the inner milieu of the cell to its surroundings (Piper and Katzmann, 2007). Endocytosis and
autophagy are often seen as parallel pathways; however these processes sometimes share several
features, such as some protein components and the lysosome as the terminal destination. In addition,
the source of the autophagic membranes is thought to mainly reside in the endocytic membranes
(Lamb et al., 2013a). Moreover, autophagic vesicles can sometimes fuse with MVBs prior to fusion of
the former with the lysosomes, thus generating intermediate vesicles called amphisomes, albeit the
significance of these vesicles is still unclear (Tooze et al., 2014).

P a g e | 16
4. Materials and methods
4.1. Fly strains
All strains were grown at 25°C. yw flies were used as wild-type controls. Fly mutant and transgenic
alleles used in this study are as follows: [atg7d14] and [atg7d77] (Juhasz et al., 2007); [FRT82B,
Atg13Δ81], [ref(2)Pod2], [ref(2)Pod3] and [UASp-eGFP-mCherry-DrAtg8a] (Nezis et al., 2010); [park25]
and [parkZ3-472] (Greene et al., 2003); [UASt-UBP2] (DiAntonio et al., 2001); [UASp-mCherry-Atg8a]
and [UASp-eGFP-Atg8a] (Rusten et al., 2007); [DJ-GFP] (Bazinet and Rollins, 2003); [Δ64a-f] (Slone
et al., 2007); [UASp-IK2-GFP], [UASp-SpnF-GFP], [spnFAA] and [spnF3325] (Dubin-Bar et al., 2008;
Bitan et al., 2010); [FRT42D, ovoD1-18] (gift from Rörth lab, IMCB, Singapore); [FRTG13, uba1]
(gift from Schuldiner lab, WIS, Israel) ; [dbrucee101] (Bader et al., 2010); [UASt-dBruce] (Arama et
al., 2003; Kaplan et al., 2010); [cathD1] (Yacobi-Sharon et al., 2013).
For maternal ectopic expression we used the driver lines: P[matα4-GAL-VP16]V37 (Bloomington
Stock #7063) and P[matα4-GAL-VP16]V2H (#7062). For male knockdown or overexpression
experiments we used the driver: [Bam-Gal4;;UAS-Dicer,Bam-Gal4] (Aram et al., 2016). Other
Bloomington stocks used in this study are as follows: [FRT82B, ovoD1-18] (#2149); [Df(3L)Pc-MK]
(#3068); [parkDf] (#26831); [FRTG13, ovoD1-18] (#4434); [UASp-YFP.Rab7] (#23270); [UASp-
YFP.Rab7DN] (#9778); [UASp-YFP.Rab5] (#24616); [UASp-YFP.Rab11] (#9790); [phb14] (#3395);
[phb123] (#5390); [phb211345] (#11345); [phb224459] (#24459); [phb2Df] (#7895); [phb2Ri] (#40835);
[mul1EY12156] (#20832); [atg8a1] (#10107); [atg8a2] (#14639); [atg2EP3697] (#17156); [atg2Df]
(#8976); [ik21] (#5322); [ik25] (#5324);
For male knockdown experiments we used VDRC (Vienna Drosophila Resource Center) lines:
[mul1v109808] (#v109808); [CG7656Ri1] (#v100791); [CG7656Ri2] (#v26881); [CG2241Ri1] (#v105002);
[CG2241Ri2] (#v49244); [CG3752Ri] (#v21707); [CG43691Ri1] (#v32837); [CG43691Ri2] (#v32839);
[CG6094Ri1] (#v106144); [CG6094Ri2] (#v48721); [CG4836Ri1] (#v109692); [CG4836Ri2] (#v109999);
[CG4836Ri3] (#v41431); [CG14540Ri1] (#v102874); [CG14540Ri2] (#v48539); [CG14546Ri1]
(#v100374); [CG14546Ri2] (#v45592); [CG5653Ri1] (#v104121); [CG5653Ri2] (#v14064);
[CG12313Ri1] (#v100361); [CG12313Ri2] (#v4075); [CG6691Ri] (#v3951).
For the knockdown experiments, the triple maternal driver MTD (Bloomington #31777) was crossed
to the following shRNA lines from the TRiP collection at Harvard Medical School: atg7shR (#34369),
atg6shR (#35741), atg1shR (#35177), fip1200shR (#36918), uvragshR (#34368), stamshR (#35016), atg2shR
(#34719), atg9shR (#34901), hdac6shR (#34072), keyshR (#35572), chipshR1 (#34017), chipshR2 (#33938),
sinashR (#40842), traf6shR (#33931), vti1shR (#38526), ykt6shR1 (#38314), ykt6shR2 (#50937), snap29shR

P a g e | 17
(#51983), vamp7shR (#43543), tsg101shR (#35710), dorshR (#54460), carshR (#34007), tbc1d15/17shR
(#34859), drp1shR (#51483), prosα6shR1 (#53974), prosα6shR2 (#55243), prosβ1shR (#34824), prosβ4shR
(#32390).
For CRISPR inactivation of mul1 we drove Cas9 expression in the early embryos using the NIG-FLY
line - y2 cho2 v1; attP40(nos-Cas9)/CyO.
Embryos were staged by developmental hours at 25°C in a humid incubator, in hours after egg laying
(AEL). Females can sometimes retain embryos for a longer period before they lay them due to being
fed on limited diet or to being too old (more than 7 days old). Therefore, we used young females
which were fed on a reach yeast diet. Under these conditions, embryogenesis is synchronized and lasts
21 ± 1 hour at 25°C.
4.2. Cloning
4.2.1. MD-DsRed and MD-Venus
To generate the MTS-DsRed and MTS-Venus flies, a pDsRed2 or a Venus gene were fused
to an N-terminal mitochondrial targeting signal (MTS) from the subunit VIII of human
cytochrome c oxidase (mito-DsRed; Clontech), PCR amplified and cloned into a pCaSpeR4
vector downstream of a don juan promoter and upstream of the cyt-c-d 3’UTR (Blumer et al.,
2002; Santel et al., 1998; Bader et al., 2011). Transgenic flies were generated by micro-
injection into embryos using standard procedures (performed by Genetic Services Inc,
Sudbury, MA).
4.2.2. MD-tdTomato
To generate the MTS-tdTomato we amplified tdTomato CDS from pRSETB-tdTomato
vector using the forward primer:
CATTCGTTGGGGGATCCACCGGTCGCCACCATGGTGAGCAAGGGCGAGGAGG
and reverse primer:
ATCTTGTTGTTTCGCAGGCGATTGCGGCCGTTACTTGTACAGCTCGTCCATGC.
Then, by the RF (restriction free) technique we replaced the DsRed CDS in the MTS-DsRed
vector with tdTomato CDS. From this clone onward all the transgene injections were carried
out by Bestgene Inc.
4.2.3. t-Parkin and dj-Tev
To inactivate parkin during late spermatogenesis (see also the scheme in Figure 14A), we
first cloned a genomic fragment of parkin (g-parkin) encompassing the 2nd intron and the 3rd

P a g e | 18
exon of the gene (isoform RB of the CG10523 gene) into the pL452 vector (a gift from
Stephen P. Creekmore, NCI-Frederick) using the BamHI and SacII restriction sites. This
genomic fragment was amplified from the BACPAC genomic clone CH322-72M15
(BACPAC resources) using the following forward primer:
CGGCGGATCCGAAGTTAACTTTTACTGTCTTC and reverse primer:
AGTCCGCGGTATTCAGACGTCCCTGGAAATAAAGATTTTCGAGCTCTCCTTGGAA
GTAAAGGTTTTCAGAAGCACCCTGAAAATACAAATTCTCCTCGCTTTCCAGCTGC
A. The reverse primer contained 3 tandem arrays of the TEV-recognition sequence
ENLYFQG. Recombineering of the “targeting (PCR) fragment” (which contains the
kanamycin resistant gene flanked by two LoxP sites and followed by the g-parkin fragment)
with the parkin BACPAC (which is cloned in attB-P[acman]-CmR-BW) was performed as
previously described (Venken et al., 2006). The following primers were used to amplify the
“targeting (PCR) fragment” from the pL452 plasmid, which also added two flanking parkin
homology sequences 50 bp and 70 bp, respectively, for the subsequent recombineering
process: forward primer:
CGATGCCACGACAATAGAGGTAAGAAGTTTCTTTAATGTATCCGCTACATGCAGC
CCAATTCCGATCATATTC; reverse primer:
GGGTTTCGTTAAAAAAAAAAAAAACAATAAAAACTAATTGTTGCCTACCTTCAT
CGGTTATATTCAGACGTCCCTGGAAA. Recombineering followed by floxing (in a Cre
producing bacteria) resulted in the insertion of the Tev recognition sequences into parkin 3rd
exon, while leaving one LoxP site inside the 2nd intron of parkin. This t-parkin construct
was then injected into embryos (containing the attP-VK2 landing site; BestGene Inc) to
generate transgenic t-parkin flies. The single LoxP site in the t-parkin transgene is then
removed by conventional splicing, thus ultimately promoting the generation of t-parkin
mRNA which contains the Tev recognition sites within its ORF. The dj-Tev transgene was
generated by PCR amplification of the TEV protease sequence from a plasmid (a gift from
Kim Nasmyth, University of Oxford) and its subsequent cloning into the pCaSpeR4 vector
downstream of a don juan promoter and upstream of the cyt-c-d 3’UTR (see above), using
the BamHI and NotI restriction sites. Transgenic flies were generated using standard
procedures as indicated above.
4.2.4. UASp-BirA and UASp-Ub6-BirA
To identify the ubiquitinated substrates we used the UASp-BirA and UASp-Ub6-BirA fly
lines that were generated by amplified the UASt-BirA and the UASt-Ub6-BirA constructs

P a g e | 19
that were received from our collaborator, Ugo Mayor and sub-cloned them into the pUASp2
plasmid (see also (Franco et al., 2011)). The BirA CDS was amplified using the following
forward primer: GGCGGTACCGCTCTTCGGTATGAAGGATAACA and reverse primer:
GGGACTAGTCTATTATTTTTCTGCAC and cloned using the restriction enzymes: KnpI
and SpeI for the insert, KnpI and XbaI for the pUASp2 vector. The Ub6-BirA CDS was
amplified using the following forward primer:
CCTGGGTACCTAACAGATCTGCTCTTCGGT and reverse primer:
GGGACTAGTCTATTATTTTTCTGCAC and cloned using the restriction enzymes: KnpI
and SpeI for the insert, KnpI and XbaI for the pUASp2 vector.
4.2.5. Phb1 rescue
The Phb1 rescue construct, l(2)37cc-GFPR-BAC was generating using recombineering
technique as previously described (Venken et al., 2006). “Targeting (PCR) fragment” was
inserted before the stop codon of Phb1 in the CH322-116M5 BACPAC. The following
primers were used to amplify the “targeting (PCR) fragment” from the pL452 C-EGFP
plasmid, which also added two flanking Prohibitin homology sequences 50 bp for the
subsequent recombineering process: forward primer:
TGCCCAGCGGACAGAGCACGCTGCTCAATCTGCCATCGACCATCGCGCAGGCAG
CCCAATTCCGATCATATTC, reverse primer:
GTAAATGCTATAGGTAGTTACAACTTAACGGAACTAGATGCACCCAGCTA
TTACTTGTACAGCTCGTCCATG. Recombineering followed by floxing (in a Cre
producing bacteria) resulted in the insertion of in frame EGFP at the C’ terminal of Phb1.
The above transgenic flies were generated using the φC31-mediated site specific transgenesis
technique, which allows insertion of transgenes into known sites of the Drosophila genome.
The clones were inserted into the attP2 site on chromosome 3L estimate cyto site 68A4 in y[1]
w[67c23]; y[+t7.7]=CaryP attP2 flies.
4.2.6. UASp-UBP2
We generated transgenic flies that overexpress the yeast deubiquitinating enzyme UBP2
under UASp element according to the CDS used by DiAntonio et al. (DiAntonio et al., 2001).
The UBP2 CDS was PCR amplified from pGP564-UBP2 vector (a gift from Maya
Schuldiner’s lab, WIS, Israel)using the forward primer:
TAACGGTACCATGCCGAACGAAGATAATGAACTTCAAAAAGC and the reverse
primer: TATCGGCCGCTACTTTAGAATTCTTTTCAATGGC and cloned using the
restriction enzymes: KpnI and EagI for the insert and KpnI and NotI for the pUASp2 vector.

P a g e | 20
4.2.7. UASp-Marf
We generated transgenic flies that overexpress the mitochondrial fusion protein Marf under
UASp element. The Marf CDS was PCR amplified from whole fly mRNA extractions which
were reverse transcribed using Invitrogen RT-PCR kit. The PCR amplification performed by
using the forward primer: AACGGTACCATGGCGGCCTACTTGAACC and the reverse
primer: CGCTCTAGACTACTGCGGCGATATATAGTTGTGC and cloned using the
restriction enzymes: KpnI and XbaI for the insert and KpnI and XbaI for the pUASp2 vector.
4.2.8. UASp-Fzo
We generated transgenic flies that overexpress the testis specific mitochondrial fusion protein
Fzo under UASp element. The Fzo CDS was PCR amplified from whole fly mRNA
extractions which were reverse transcribed using Invitrogen RT-PCR kit. The PCR
amplification performed by using the forward primer:
GCGGGTACCATGGCGGAATCTGACT and the reverse primer:
ACTACTAGTTCAGCTCTTGGTC and cloned using the restriction enzymes: KpnI and
SpeI for the insert and KpnI and XbaI for the pUASp2 vector.
4.2.9. UASp-EGFP-CD63
We generated transgenic flies that overexpress the human extracellular vesicles’ marker
CD63 fused with EGFP and under UASp element. The EGFP-CD63 CDS was PCR
amplified a UASt-EGFP-CD63 plasmid (kindly shared by Suzanne Eaton, the Max Plank
Institute of Molecular Cell Biology and Genetics, Germany). The PCR amplification
performed by using the forward primer:
CCGCGCGGCCGCATGGTGAGCAAGGGCGAGGAGCTG and the reverse primer:
GCGGACTAGTCTACATCACCTCGTAGCCAC and cloned using the restriction enzymes:
NotI and SpeI for the insert and NotI and XbaI for the pUASp2 vector
4.2.10. Mul1 CRISPR
To generate precise deletion of Mul1's coding region we used the CRIPSR protocol described
by the Ueda group (Kondo and Ueda, 2013). In short, two gRNA target sites were designed
by the NIG-FLY Cas9 target finder, to target Cas9 mediated excision 100 base pairs after
Mul1's start codon and 1015 bases after Mul1's start codon resulting in amino acid... For each
site the following oligos were ordered: CTTCGCGAACTGCCAAGGTGCTGA and
AAACTCAGCACCTTGGCAGTTCGC for the upstream site;
CTTCGTGCTCCACCAATCCCAAGG and AAACCCTTGGGATTGGTGGAGCAC for
downstream site. Then the oligos were ligated one to another and cloned into pre-cut pBFv-

P a g e | 21
U6.2B and pBFv-U6.2 (in accordance) by the restriction enzyme BbsI. Both vectors were cut
again by EcoRI and NotI, to unite the two gRNAs into the vector pBFv-U6.2B. The clones
were inserted into the attP40 site on chromosome 2L with estimated cyto site 25C6.
After receiving the gRNA expressing transgenic flies, we crossed them flies expressing Cas9
at early embryonic stages (y2 cho2 v1; attP40(nos-Cas9)/CyO) and screened the progeny for
the predicted excision in Mul1's coding region.
4.3. Ultrastructural studies
4.3.1. TEM
Fertilized eggs were collected for 10 minutes and either immediately dechorionated or aged
at 25°C for 20 minutes, and then dechorionated by hand-peeling on a double-sided adhesive
tape. The eggs were fixed for 20 minutes in 1:1 volume of heptane and 2.5% glutaraldehyde
(diluted in 0.1M Cacodylate buffer - CaCo, pH=7.4). Fixed eggs were hand-devitellinized on
a metal lattice, re-fixed in 2.5% glutaraldehyde, and kept at 4°C. Preserved eggs were washed
with 0.1M CaCo buffer, post fixed in 1% osmium-tetroxide (diluted in 0.1M CaCo buffer)
for 2 hours without shaking, and washed with 2% uranylacetate in DDW. Samples were
dehydrated in graded ethanols, embedded in Epon 812, and hardened at 60°C for 2 days.
Ultrathin sections (70-90 nm thickness) were prepared with ultramicrotome Leica UCT
(Leica), analyzed under 120kV Transmission Electron Microscope (TEM) Tecnai 12, and
digitized with EAGLE CCD camera using TIA software (FEI, Eindhoven). Testes were
prepared for TEM analysis as described in (Arama et al., 2006).
4.3.2. FIB
We use the dual beam microscope (FIB–SEM) and the Serial Surface View method (SSV) to
investigate the organization of multivesicular aggregates associated with paternal
mitochondrial destruction in 3D. The method involves sequentially exposing new surfaces of
an embedded sample using the focused beam of gallium ions and imaging the exposed face
with the electron beam (Heymann et al., 2006). In this way, a stack of serial surface images
with nanometric resolution in all three orthogonal directions is obtained.
An ion beam current of 0.92 NA at 30 kV was used for milling slices. The lateral resolution
and the slice thickness are both of the order of 10 nm and the field of view is around 15 × 15
microns. ImageJ software (National Institutes of Health) was used to align images using the

P a g e | 22
Stackreg plugin as previously described (Schindelin et al., 2012). Imaris (Visage Imaging)
was used for segmentation, rendering, and visualization of the data.
The electron microscopy studies were conducted at the Irving and Cherna Moskowitz Center for Nano
and Bio-Nano Imaging at the WIS.
4.4. Live imaging studies
4.4.1. For kinetics experiments, fertilized eggs were collected 0-5 minutes AEL and hand-
dechorionated as described above. Dechorionated eggs were transferred to a glass slide
coated with a thin adhesive layer (extracted by heptane from a double sided adhesive tape,
dispersed on a slide, dried, and bordered by hydrophobic pen), mounted in a drop of PBS and
visualized under the confocal microscope (Zeiss LSM 510). For each embryo, 12 Z-stacks of
4 µm each were taken every 4 minutes for 25 rounds. Frame series were converted into
movies using Imaris software. Fluorescence intensity was quantified by Matlab with the help
of the Department of Veterinary Resources at the WIS.
4.4.2. For 0h, 1h, 2h kinetics experiments fertilized eggs were collected as described in 4.4.1,
and visualized under the confocal microscope (Zeiss LSM 710), but weren't scanned. Instead,
each fertilized egg was given a mark and scored "Strong", "Weak", or "None", according to
its MD-tdTomato intensity. The fertilized eggs were left to develop inside a humidity
chamber and scored again twice, in 1 hour and 2 hours after egg laying. All the data was
summed and plotted on a graph using excel.
4.4.3. For live imaging of eggs expressing mCherry-Atg8a, YFP-Rab7 and EGFP-CD63,
fertilized eggs were collected as described above, and for each embryo, adjusted z-stacks of 2
µm each were taken every 20 minutes for consecutive time points.
4.5. Immunostaining
Fertilized eggs were collected for 10 minutes, aged at 25°C for 10, 30, or 50 minutes and processed
for whole mount antibody staining using standard techniques (Ashburner, 1989). The primary
antibodies used in this study are human monoclonal anti-Ub-K63 antibody (anti-ubiquitin lys63-
specific clone Apu3; 1:100; Genetech) (Newton et al., 2008), mouse monoclonal anti-polyglycylated
tubulin antibody (AXO 49; 1:5000) (Bre et al., 1996), rabbit polyclonal anti-Drosophila p62 antibody

P a g e | 23
(anti-ref(2)P; 1:100) (Wyers et al., 1995), and guinea-pig polyclonal anti-Hrs antibody (1:100) (Lloyd
et al., 2002). All secondary antibodies were used in a dilution of 1:100 (Jackson ImmunoReaserch).
Eggs were mounted in Fluoromount G (SouthernBiotech) and observed under the confocal
microscope (Zeiss LSM 510 and Zeiss LSM710).
To preserve the endogenous fluorescence of the DJ-GFP transgene after fixation, we performed the
hand devitellinization of embryo procedure, which is methanol free, as described in (Rothwell and
Sullivan, 2007). Fixation was performed in 4% paraformaldehyde. This procedure, however,
preserved the immunoreactivity of the AXO 49 antibody, but not that of the anti-Ub-K63 and anti-p62
antibodies.
4.6. Western blots
4.6.1. Parkin Tev inactivation validations
Proteins were extracted from testes and seminal vesicles from 1 and 5 days old male flies,
respectively. Samples were run in SDS-PAGE and transferred to nitroglycerin membrane,
which was reacted with rabbit anti-Parkin antibody (1:5000) (Greene et al., 2005) and mouse
anti-β-tubulin (clone E7, Hybridoma Bank; 1:1000).
4.6.2. First MVBs isolation strategy – ultracentrifugation and sucrose gradient
Each fraction was determined by refractive index measurements, loaded and run in SDS-
PAGE, and reacted with guinea-pig polyclonal anti-HRS (1:500) (Lloyd et al., 2002) anti-
GFP (Abcam290 1;1000), rabbit anti GAPDH (IMGENEX 1:500), mouse anti-
ATPsynthase-α (anti-ATP5A, MitoScience 1:1000).
4.6.3. Second MVBs isolation strategy – gradual centrifugations and marker based IP
Each fraction from the fractionation assay was determined by refractive index measurements,
loaded and run in SDS-PAGE, and reacted with anti-GFP (Abcam290 1;1000), anti-mCherry
(Abcam 1C51, 1:2000).
4.7. Gastrulation assay
Fertilized eggs were collected for 30 minutes, allowed to develop for 3 hours at 25°C, fixed, and
mounted. Embryos were then monitored for gastrulation using phase contrast or digital interference
contrast (DIC or Numarski) microscopy.

P a g e | 24
4.8. RNA isolation and RT-PCR
Total RNA was extracted from 10-20 unfertilized eggs using the PureLinkTM Total RNA Purification
System (Invitrogen) according to the manufacturer’s recommendations. Purified RNA was reversed
transcribed using Qiagen OneStep RT–PCR kit (Qiagen). cDNA was then PCR amplified for 25-30
cycles using specific primers.
4.9. Quantification and statistics analyses of PMD kinetics
4.9.1. For kinetics experiments, twenty-five projections (one for each time point) from 12 z-
stack images per each time point of live dechorionated fertilized eggs (beginning at 5-10
minutes AEL and lasting for about 2 hours) were analyzed by a dedicated Matlab script
designed to quantify the fluorescence intensity of the MD. The first time point of each
embryo was set as 1, and the values of the ensuing time points were expressed as percentage
of the first time point. Fluorescence intensity levels are represented as the mean ± SEM of all
the examined embryos at each time point. Statistical analysis was performed using the two-
tailed unpaired Student’s t test. Values of P < 0.05 were accepted as statistically significant.
4.9.2. For 0h, 1h, 2h experiments, each MD-tdTomato intensity was represented by a
numerical score (strong=2, weak=1, none=0). For each genotype and in each time point the
scores were summed and checked for significance using the StatView software. Statistical
analysis was performed using the two-tailed unpaired Student’s t test, Fiscer's test and
Scheffe test. A strict approach was taken saying only values of P<0.05 or lower that were
found in all three tests were accepted as statistically significant.
4.10. Mass Spectrometry
4.10.1. Targeted mass-spectrometry analyses of selected genes
Fertilized eggs were collected for 20 minutes, aged at 25°C for either 10, 30, 50, 70, and 90
min (a short protocol) or 10, 70, 130, 190 and 250 minutes (a long protocol). These eggs
were then dechorionized in 50% bleach, dried and rapidly frozen in liquid nitrogen and stored
in a -80°C freezer. The collected eggs from each time point were combined and lysed on ice
in detergent free lysis buffer (25 mM Hepes pH=7.4, 2.5 mM MgCl2, 0.5 mM EDTA, 150
mM NaCl) mixed with protease inhibitor (1:1000 Leupeptin, 1:1000 Aprotinin, 1:100 PMSF;
Sigma), and then subjected to high speed centrifugation in 4°C for 20 minutes. Supernatant

P a g e | 25
collected and subjected to mass spectrometry analysis (Dr. Yishai Levin, the Mass Spec unit
at the Weizmann Institute of Science [WIS]). Protein concentration measurements and full
digestion of lysates by trypsin were performed. The, digested samples were analyzed against
desired in-silico digested protein list (composed with the help of Shmuel Pietrokovsky, at the
WIS) using the Xevo TQ-S tandem quadrupole mass spectrometer.
4.10.2. Full discovery mass-spectrometry of early fertilized eggs
Eggs were collected according to the following methods; First, wild-type fertilized eggs were
collected for 30 minutes and aged at 25°C for 90 minutes or not aged at all. Second, wild-
type eggs, fertilized by DJ-GFP sperm were collected for 30 minutes. As a control, non-
fertilized eggs were collected for 2 hours. All of these eggs were then dechorionized in 50%
bleach, dried and rapidly frozen in liquid nitrogen and stored in a -80°C freezer. The
collected eggs from each time point were combined and lysed on ice in SDT-lysis buffer: (4
% (w/v) SDS, 100mM Tris/HCl pH 7.6, 0.1M DTT) and then subjected to high speed
centrifugation in 4°C for 20 minutes. Supernatant collected and subjected to mass
spectrometry analysis (Dr. Yishai Levin, the Mass Spec unit at the Weizmann Institute of
Science [WIS]). Protein concentration measurements and full digestion of lysates by trypsin
were performed. The, digested samples were run in 2D-LC-MS/MS. The raw data was
searched against the Drosophila melanogaster protein database as downloaded from Uniprot.
Clusters were made using the DAVID algorithm, for hits that exhibit infinite or up to 10 fold
ratio (the ratio was calculated by the division of protein levels in the fertilized samples by the
protein levels in non-fertilized samples).
4.11. MVB purifications
4.11.1. Gradual centrifugations, ultracentrifugation and fractionation
Hundred µl of embryos at 0-2 hours AEL were bleached and then crushed in a PBSX1 buffer
(137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 2 mM KH2PO4) supplemented with
protease inhibitor (Sigma), followed by differential centrifugation at 2,000 x g and 10,000 x g
before pelleting the vesicles at 100,000 x g in a TLA 120.2 rotor for 70 minutes (Beckman).
This procedure is further described in (Gross et al., 2012). The pellet was washed with
PBSX1 and again centrifuged at 100,000 x g for 3 hours. Supernatant was discarded and the
vesicles were suspended in 200µl PBSX1, layered on a continuous sucrose density gradient
(0.8-2 M sucrose generated by a gradient maker in 12 ml ultraclear SW40 centrifuge tubes),

P a g e | 26
and centrifuged for 3 hours at 100,000 × g (SW41 rotor; Beckman Instruments). Twelve
fractions of 1 ml were collected by Fraction collector (Gilson) from the top (low density) to
the bottom of the tube (high density) of the sucrose gradient using Gradient fractionator
(Piston).
4.11.2. Gradual centrifugations, ultracentrifugation and IP
To obtain vesicles from MVBs using the combined method from early fertilized eggs we first
collected 0-1h AEL fertilized eggs and bleached them before deep froze in liquid nitrogen
and kept in -80°C. We used yw fertilized eggs as control, in addition with eggs which
overexpress EGFP-CD63 and/or mCherry-Atg8a or GFP-Atg8a. For vesicle isolation, frozen
samples were slowly warmed on ice for at least one hour. Then crushed in cell dissociation
buffer (Sigma) supplemented with protease inhibitor (Sigma) using dounce homogenizer
(Sigma), followed by differential centrifugation starting at 300 x g (2*10min), 2,000 x g
(10min) and 10,000 x g (30min). Then supernatant was filtered using 0.45μm filter
(Sartorius) and concentrated using Amicon filter with a cutoff of 100kDa (Milipore), before
pelleting the vesicles at 100,000 x g in a TLA 120.2 rotor for low volumes (up to 1ml) or
type 50 TI rotor for larger volumes (1-5ml), all for 3 hours (Beckman ultracentrifuges).

P a g e | 27
5. Results
5.1. Ultrastructure of the PMD process in Drosophila
In the fruit fly, Drosophila melanogaster, mitochondrial remodeling initiates immediately after
meiosis in the haploid round spermatid with the fusion of all the mitochondria into a giant sphere
called the Nebenkern. The Nebenkern is then resolved into two mitochondrial derivatives (MDs),
which unfold and elongate alongside and in close association with the growing axoneme to the length
of the sperm flagellum (nearly 2 mm long; Figure 1A, I-II; Figure 1B). During advanced spermatid
elongation stages, one MD (“minor”) constantly loses volume and shrivels, whereas the other MD
(“major”) accumulates dark amorphic material of an unknown nature termed the “paracrystalline”
material (Figure 1A, III; Figure 1C) (Lindsley and Tokuyasu, 1980; Fuller, 1993).

P a g e | 28
Figure 1. Visualization of the paternal MD under the electron and confocal microscopes
(A) Schematic representation of Drosophila sperm during three consecutive maturation stages.
(B, C and F-H) Electron micrographs of cross sections through (B and C) developing Drosophila sperm cells and (F-H) anterior regions
of fertilized eggs at 10-20 min AEL.
(B and C) Micrographs corresponding to stages II and III in A, respectively.
(F) Part of the sperm flagellum (Fm) is shown among several maternal mitochondria (mM). Note the presence of the axoneme (AXO)
and the MD
(G and H) Higher magnifications of cross- and transverse sections through the sperm flagellum, respectively.
(B, C, G and H) The colors indicated were manually added using Adobe Photoshop to correspond with the colored organelles in (A).
(C and G) The asterisks indicate the remnants of the minor MD.
(D and E, I-N) Confocal images of mature sperm cells from (D and E) seminal vesicles and (I-N) inside fertilized eggs at 15-30 minutes
AEL.
(D and E) Insets are enlargements of the boxed areas.
(D, E and I-N) The MD marked by DJ-GFP transgene (green), colocalizes with the MTS-DsRed transgene (D and I-K) (red) but not
with the axoneme (E and L-N) (AXO; red; labeled with the AXO 49 antibody). Anterior is to the left.
(D, E, K and N) Note the distinction between the MD and the axoneme (E and N), whereas the two MD markers completely overlap (D
and K).
Scale bars represent 200 nm (B, C, and F-H), 10µm (D, E, and I-K) and 5 μm (L-N). See also Figure 4.

P a g e | 29
Upon fertilization, a single sperm cell completely penetrates the Drosophila egg through a
specialized structure, the micropyle, a single hole in the chorion located at the anterior end of the egg
(Gilbert and Richmond, 1981; Karr, 1991). To uncover the fate of the paternal MD after fertilization,
we first used transmission EM (TEM) of ultrathin-sections prepared from the anterior part of early
fertilized eggs. At 15-30 minutes after egg laying (AEL), the intact sperm flagellum could be readily
distinguished from the egg’s organelles by virtue of its unique organellar contents (i.e. the axoneme
and the attached MD; Figure 1F-H). The still largely intact flagellum is approached by globular
clusters of microvesicles (also referred to as multivesicular clusters; MVCs; Figure 2A and A’) which
then associate with the sperm plasma membrane (Figure 2A’-Figure 2B’; also note the microvesicles
which are attached to the sperm plasma membrane). The MVCs are also abundant in unfertilized eggs,
suggesting that the developing egg may already be prepared for sperm penetration (Figure 3A-A’). In
addition to the MVCs, other large vesicular organelles reminiscent of either multivesicular bodies
(MVBs) or multilamellar bodies (MLBs), are completely wrapped around some regions of the
flagellum were also detected (Figure 2C and Figure 2C’, respectively). For convenience, we will
hereafter refer to the MVCs, MVBs, and MLBs as MVB-like vesicles. At a later stage, the MD
separates from the axoneme (Figure 2D and 2D’), a process also readily visualized under the
fluorescent microscope (Figure 1L-N; Figure 4). Subsequently, still within the preblastoderm (mitotic
cycles 1-9), the MD breaks into oval fragments (MFs) with diameters between 0.1-0.4 µm, which are
then enveloped by autophagosome-like double membranes (Figure 2E and 2E’). The MFs are
eventually sequestered to lysosomes where they are completely degraded and assimilated into the egg
cytoplasm (Figure 2F and Figure 2F’; see also Figure 3C, Figure 3E for autolysosomes that are in the
process of digesting the MF’s), and they are no longer detectable by the cellular blastoderm stage
(130-180 minutes AEL). We conclude that immediately after fertilization, the paternal MD is
degraded by a network of vesicles originating in the egg and reminiscent of elements of the endocytic
(MVB-like vesicles) and autophagic pathways. Because this is a stereotypical active process, we
hereafter refer to this process as paternal mitochondrial destruction or PMD.

P a g e | 30
Figure 2. Ultrastructural characterization of the PMD process
Electron micrographs of cross-sections through fertilized eggs at 10-60 minutes AEL. (A and A’) Soon after fertilization,
multivesicular clusters approach and associate with the flagellum (detected in six sections of three embryos). Note that the sperm
plasma membrane is readily visualized (light green arrow). mv, microvesicles. (B and B’) The microvesicles specifically attach to the
sperm membrane, but not the maternal mitochondria (detected in 14 different sections of three embryos). (C and C’) Multivesicular
bodies (C) and Multilamellar bodies (C’), which completely enwrap some flagellar regions, are also detected (detected in two different
sections in one embryos). Note that the MD and the axoneme are still enveloped by the sperm plasma membrane, as indicated by the
double membrane of the MD and the surrounding plasma membrane within the MVB (light green arrow), and that the highly ordered 9
+ 2 microtubule structure of the axoneme is disrupted (11 white dotes). (D and D’) The MD is separating from the axoneme (detected
in different sections of one embryo). (E and E’) Autophagy-like double membranes, reminiscent of preautophagosomal structures
(PAS), phagophores, and autophagosomes, are formed around and envelope MD fragments, which can be readily recognized by their
dark paracrystalline content (detected in three different sections of two embryos). (F and F’) The MFs are sequestered to large
lysosomes, where they are disintegrated (F) and assimilated into the ooplasm (F’) (detected in eight different sections of three
embryos). The scale bars represent 500 nm (A-B, F and F’) and 200 nm (B’, and C-E’). See also Figure 3.

P a g e | 31
5.2. The PMD kinetics
In order to investigate the mechanisms underlying PMD, we first established a fluorescence-based
imaging system to monitor the sperm MD within early fertilized eggs. We generated transgenic flies
carrying mitochondrial targeted red (MTS-DsRed) and yellow (MTS-Venus) fluorescent proteins
under the regulatory regions of the sperm-specific gene, don juan (dj). The red and yellow (but not
green) fluorescent transgenes were preferentially selected because of the large amount of masking
green autofluorescence coming from the egg yolk. By crossing these flies to transgenic flies
expressing the sperm-specific mitochondrial protein marker, DJ-GFP (Santel et al., 1998; Bazinet and
Rollins, 2003), we validated that these transgenes specifically label the sperm MD (Figure 1D and
Figure 1I-K). Furthermore, staining the DJ-GFP-expressing sperm with the anti-polyglycylated
tubulin antibody (AXO 49), which specifically detects the mature axoneme (Bre et al., 1998; Arama et
al., 2007), revealed a clear distinction between these two organelles despite their close alignment
throughout the sperm flagellum (Figure 1E and Figure 1L-N). Transgenic male flies carrying MTS-
Figure 3. The unfertilized egg already contains numerous MVCs
(A and the enlargements in A’) Electron micrographs of a cross section through an unfertilized egg soon AEL. The area confined by
black square was magnified and presented in separated micrographs(A’). (B) Live confocal imaging of an early unfertilized. The egg
maternally expressed the single-labeled autophagy reporter, mCherry-Atg8 (red). Atg8-positive vacuoles are dispersed throughout the
egg. (C-E) Electron micrographs of cross sections through fertilized eggs at 10-60 minutes AEL. Note the presence of autolysosomes
which still contain MD fragments (MF) together with degraded material. MVC, multivesicular clusters; Fm, flagellum; mM, maternal
mitochondria. Scale bars: A, 1 μm; A’, 500 nm; B, 20µm; C-E, 2 µm.
P a g e | 32
DsRed, MTS-Venus, and DJ-GFP were fertile and gave rise to normal progeny (Figure 8C and Figure
8E and data not shown), and can thus serve for further analyses of the PMD process.
To determine the kinetics of the PMD process, we monitored the elimination rate of the sperm MD
by live imaging of eggs fertilized by the MTS-DsRed sperm. Beginning at 10 minutes AEL, time-
lapse sequences of z projection images with a 4 min interval were captured for about 2 hr. These
analyses revealed that the MD almost completely disappears at 80-90 minutes AEL (i.e. at the onset of
the syncytial blastoderm stage; see Movie 1 and still images from this 2 hours movie at the top of
Figure 7A, and the quantification of the fluorescence intensities in Figure 7B). It is noteworthy that
the disappearance of the fluorescent signal does not occur in a uniform fashion throughout the sperm
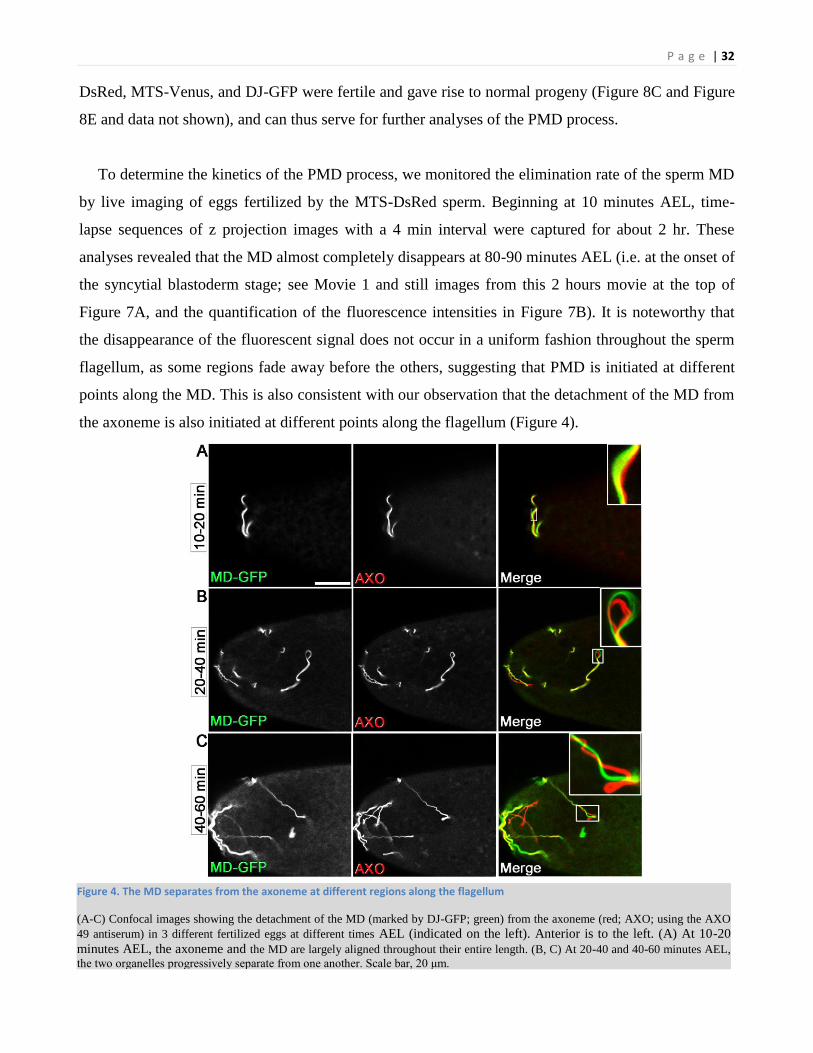
flagellum, as some regions fade away before the others, suggesting that PMD is initiated at different
points along the MD. This is also consistent with our observation that the detachment of the MD from
the axoneme is also initiated at different points along the flagellum (Figure 4).
Figure 4. The MD separates from the axoneme at different regions along the flagellum
(A-C) Confocal images showing the detachment of the MD (marked by DJ-GFP; green) from the axoneme (red; AXO; using the AXO
49 antiserum) in 3 different fertilized eggs at different times AEL (indicated on the left). Anterior is to the left. (A) At 10-20
minutes AEL, the axoneme and the MD are largely aligned throughout their entire length. (B, C) At 20-40 and 40-60 minutes AEL,
the two organelles progressively separate from one another. Scale bar, 20 μm.

P a g e | 33
5.3. Autophagy-related vesicles associate with the sperm MD soon after fertilization
To further characterize the molecular nature of the MVB-like vesicles and the autophagosome-like
vesicles, we used fluorescent protein markers. Males expressing the MTS-Venus sperm were first
crossed to females maternally expressing a genetic reporter of autophagy, eGFP-mCherry-Atg8a. This
double-labeled reporter can reveal autophagosomes and autolysosomes by virtue of the Atg8 ability to
localize to phagophore-assembly sites and thus fluorescently label autophagosomes in green and red
(i.e. yellow), and autolysosomes in red only because of the quenching of the eGFP signal in the acidic
environment (Nezis et al., 2010). Significantly, as opposed to classical autophagosomes which are
usually smaller than 0.5-0.7 µm in diameters, this reporter labeled much larger vesicles in the range of
0.5-1.8 µm in diameters, which were specifically associated with the MD (an embryo at 40-60 min
AEL, are shown in Figure 5A). These vesicles were all detected in both the green and red fluorescent
channels, suggesting that they were not fused with lysosomes. In addition, the MD was still detected
even when the embryo reached the cellular blastoderm stage (2 hours AEL; data not shown),
suggesting that this reporter may abrogate the function of the vesicles and thus attenuate the PMD
process.
We considered the possibility that the double-labeled Atg8 reporter may be somewhat harmful to
the function of the vesicles than the singly-labeled counterparts, and hence we instead used a singly-
labeled reporter, mCherry-Atg8a. Indeed, although the single-labeled reporter still caused a mild
attenuation in the PMD process (data not shown), we could clearly and reproducibly follow the
dynamics of the Atg8-positive vesicles, concomitant with the destruction of the MD (Figure 5C-E). At
15-20 minutes AEL, only small scattered mCherry-Atg8-positive vesicles were detected in the
anterior region of the egg, without any specific association with the MD (Figure 5B). However, at
about 30 minutes AEL onward, mCherry-Atg8 started accumulating in the vicinity of and became
associated with the MD, which in turn displayed gradually increasing fragmented morphology (Figure
5C-E). Interestingly, these vesicles gradually increased in size, becoming unusually large, reminiscent
of the size range of the vesicles detected with the double-labeled reporter as well as the sizes of the
MVB-like vesicles detected by the EM (Figure 5D-E). Furthermore, similar to the MVBs and MLBs
which entirely encapsulated some flagellar regions/pieces, large mCherry-Atg8-positive vesicles
encapsulating fragments of the MD and which were already detached from the main MD piece were
also detected (Figure 5E). Taken together, these observations suggest an unconventional role of the
autophagy pathway during PMD.

P a g e | 34
Figure 5. Unconventionally large Atg8-positive vesicles specifically associate with the sperm soon after fertilization
(A-E) Live confocal imaging of early fertilized eggs. Anterior is to the left. The insets are enlargements of the respective regions in the
small white squares. The female and male symbols indicate the origins of transgenes’ expression, egg or sperm, respectively.
The scale bar represent 20µm.
(A) An egg at 40-60 minutes AEL fertilized by the MTS-Venus transgenic sperm (yellow; MD) and maternally expressing the double-
labeled autophagy reporter, eGFP-mCherry-Atg8 (yellow vesicles; arrows in the inset point at a region of the MD and attached selected
vesicles with the indicated diameters).
(B-E) An egg fertilized by the DJ-GFP transgenic sperm (green; MD) and maternally expressing the singly-labeled autophagy reporter,
mCherry-Atg8 (red). The time points AEL are indicated to the left of each panel. AF, after fertilization

P a g e | 35
5.4. PMD is mediated by a network of vesicles displaying markers common to the autophagic
and endocytic pathways
Existing evidence suggest that under certain conditions, the endocytic and autophagic pathways
may converge, giving rise to hybrid vesicular organelles termed amphisomes (Berg et al., 1998;
Stromhaug and Seglen, 1993; Fader et al., 2008; Manil-Segalen et al., 2012; Filimonenko et al., 2007;
Lamb et al., 2013b). Amphisomes are believed to constitute prelysosomal intermediate vesicles
involved in autophagosomal maturation. The facts that Atg8 was localized to unusually large vesicles
during PMD, and that MVBs and MLBs, but not autophagosomes, are the only vesicles of a similar
size range associated with the sperm during this process (the EM data), raised the hypothesis that the
Atg8-positive vesicles may constitute hybrid organelles of the endocytic and autophagic pathways. To
test this idea, we used available markers of the endosomal pathway. First, we stained early fertilized
eggs with the anti-Hrs antibody, which detects the endosome-associated, ubiquitin-binding protein,
Hrs/Vps27, a member of the endosomal sorting complexes required for transport 0 (ESCRT-0) (Lloyd
et al., 2002). Whereas this antibody faintly stained large vesicles (1.3-1.9 µm) found in the vicinity of
the sperm flagellum (visualized by the AXO 49 antibody), vesicles which were already wrapped
around flagellar material displayed a stronger signal, suggesting that the MVBs may be highly active
during and/or after the encapsulation process (white and yellow arrows in Figure 6B). To further test
this possibility, we used an antibody which specifically recognizes the nonclassical K63-linked
ubiquitin side chains ((Newton et al., 2008); see also the text for Figure 9). Large ubiquitin-positive
vesicles (1.4-3.5 µm) were detected in the vicinity of the flagellum, some of which completely
enwrapped flagellar material (arrows in Figure 6C). Interestingly, ubiquitin-positive vesicles which
were still wrapped around some regions of the main flagellar piece could also be detected, suggesting
that the encapsulation process may promote subsequent clipping of these flagellar regions (upper
arrow in the inset of Figure 6C). Taken together, these results are consistent with the idea that at least
some of the large vesicles associated with the sperm flagellum are related to the endocytic (MVB)
pathway.
Rab GTPases function as master regulators of endocytic trafficking, and thus can serve as protein
markers for specific steps in the endocytic pathway (Figure 6A) (Pfeffer, 2013). To directly test
whether the Atg8-positive vesicles may also contain features of the endocytic pathway, we examined
early fertilized eggs expressing the mCherry-Atg8 reporter and either Rab 5, 7, or 11 fused to the
yellow fluorescent protein, YFP. Indeed, YFP-Rab7, a protein marker for mature endosomes/MVBs,

P a g e | 36
co-localized with the large Atg8-positive vesicles associated with the sperm flagellum (Figure 6D-E).
In contrast, the respective markers for early and recycling endosomes, YFP-Rab5 and YFP-Rab11,
marked unrelated vesicles in the egg (data not shown). Therefore, the large vesicles associated with
the sperm flagellum display features common to the endocytic and autophagic pathways.
Figure 6. Vesicular elements of the endocytic and autophagic pathways overlap during PMD
(A) A simplified diagram depicting the endocytic pathway and the relative vesicular locations of the proteins examined in this study.
(B and C) Early fertilized eggs stained to visualize the axoneme (green) and (B) the Hrs protein (red) or (C) ubiquitinated proteins with
K63-polyUb (red). Anterior is to the left. The diameters of selected vesicles are indicated by arrows in the insets. The scale bar
represents 20µm.
(D and E) Live confocal imaging of early fertilized eggs maternally expressing the autophagy reporter, mCherry-Atg8 (red), and the late
endosome marker, YFP-Rab7 (green).
All insets are enlargements of the respective regions in the small white squares.

P a g e | 37
5.5. Inactivating mutations in the autophagic and endocytic pathways attenuate PMD
To examine the functional involvement/requirement of the autophagic and endocytic pathways in
PMD, we next inactivated key components in the two pathways in the eggs, and monitored the effects
on the kinetics of PMD using the live imaging assay. To compromise the autophagic pathway, we first
analyzed eggs from females carrying two atg7 null alleles in trans to one another (atg7d14/d77; see also
complex #3a in Figure 8A). These mutant flies complete metamorphosis and survive to adulthood,
despite showing some severe defects in autophagy (Juhasz et al., 2007; McPhee and Baehrecke,
2009). Furthermore, atg7 maternal mutant embryos develop with normal kinetics at least until the end
of the cellularization stage and beginning of gastrulation, a stage when the MD is already completely
eliminated in wild-type counterparts (Figure 8D-E). Critically, atg7 mutant eggs displayed a
significant delay in the clearance of the MD, showing about 60% reduction in the fluorescence
intensity of the MD at 45 minutes AEL as compared with 85% reduction in the wild-type counterparts
(compare upper panels with lower panels in Figure 7A and the quantifications in Figure 7B). A similar
effect was also observed following egg inactivation of Atg8a, another important autophagy protein
(for details see also paragraph 5.3; Figure 8A and Figure 9) (Simonsen et al., 2008)). These findings
that there is a significant but still partial effect is consistent with the idea that autophagy does not play
an orthodox role during the initial stages of PMD. On the other hand, the final stages of PMD, namely
the clearance of the MD fragments by classical autophagosomes, were also not completely blocked in
this mutant. The latter is mainly attributed to a technical limitation of our system to quantify the
fluorescence of the small MD fragments, as they are dispersed with time throughout the egg until they
become too dispersed to be distinguished from the background auto-fluorescence.
Another possibility for the observed incomplete block of the clearance of the MD fragments in the
atg7 mutant eggs may be the evolvement of an Atg7-independent mechanism of autophagy (Zhang et
al., 2009; Chang et al., 2013). To test this idea, we thus explored the involvement of other key
components in the autophagic pathway using available transgenic RNAi lines, which were designed to
induce efficient knockdowns in the oocyte (the Drosophila transgenic RNAi project [TRiP] collection
of shRNAs at Harvard Medical School). Indeed, RT-PCR analysis of total RNA extracted from eggs
maternally expressing RNAi against atg7 (a positive control reference), as well as atg6 and uvrag,
which promote autophagosome nucleation and maturation, respectively (complex #2 in Figure 8A,
note that both components are also involved in the endocytic pathway), revealed highly efficient
knockdown of all these genes (atg7shR, atg6shR and uvragshR; Figure 8G). Significantly, knockdown of

P a g e | 38
all three genes attenuated the clearance of the MD with similar kinetics as those detected in the atg7
mutant eggs (Figure 7B). Therefore, the attenuation effect detected in the atg7 mutants may indeed
reflect the maximal level of involvement of the autophagic pathway in PMD, thus making it less likely
that Atg7-independent mechanisms of autophagy are involved in this process. Furthermore, maternal
expression of a dominant-negative form of Rab7 (Rab7-DN), as well as maternal knockdown of
stam/hse1 (using a stamshR; Figure 8G), which encodes for the Hrs/Vps27 partner in the ESCRT-0
complex (Williams and Urbe, 2007), also attenuated PMD with almost identical kinetics, which was
not further enhanced in the Rab7-DN and atg7shR or the Rab7-DN and stamshR double mutants (Figure
7B). We also examined TBC1D15, the mitochondrial GAP for Rab7. TBC1D15 homologs were
shown to be involved in proper autophagic encapsulation of mitochondria, binding the mitochondrial
receptor Fis1 and mediating mitochondrial morphological changes, and promoting mitophagy and
PMD in mice upstream of p62 (Yamano et al., 2014; Onoue et al., 2013; Rojansky et al., 2016). Egg
knockdown of tbc1d15 resulted in a significant delay in PMD as compared to wild-type, further
demonstrating the importance of the endocytic pathway in this process (Figure 214B). Our results
suggest that our assay mainly measures the effects on the initial stages of PMD (rather than the final
clearance of the MD fragments), during which the endocytic and autophagic pathways converge in a
common nonorthodox pathway. In support of this idea, maternal knockout of atg7 or knockdown of
uvrag dramatically affected the biogenesis of more than 50% of the MVCs, displaying MVCs that are
largely devoid of micorvesicles (Figure 7C-E). In addition, monitoring PMD in eggs from atg13,
vps34 and atg2 mutant females, as well as maternal knockdown of atg1, fip200, atg2 and atg9
(Figure 8G and Figure 9), which operate in different autophagy complexes (Figure 8A), revealed no
(atg13Δ81GLC, atg2EP3697/Df and atg9shR) or mild (atg1shR, fip200shR and atg2shR) effects on the
clearance of the MD (Figure 8F and Figure 9), suggesting that the initiation signal of the
autophagic/endocytic pathway in PMD might be distinct from that of classical autophagy.
Interestingly, another noncanonical autophagy form, called LC3-associated phagocytosis (LAP), also
proceeds in the absence of the initiation complex, suggesting some possible similarities between PMD
and LAP (Sanjuan et al., 2007; Florey et al., 2011).

P a g e | 39
Figure 7. The clearance kinetics of the MD is attenuated upon inactivation of components in the autophagic and endocytic pathways
(A) Selected frames from two 2hr movies of paternal mitochondrial elimination after fertilization in eggs from a wild-type female (top)
and an atg7 mutant female (bottom) fertilized by males with the mito-DsRed sperm. The windows of time AEL are indicated on top of
the panels. Anterior is to the left.
(B) Quantification of fluorescence intensity in 4 min intervals in live fertilized eggs mutant for the indicated genes/proteins. Error bars
represent standard errors. The number (N) of examined embryos from each genotype is indicated in parenthesis. p values of the
differences in the fluorescence intensities between the mutants and the wild-type were calculated for each time point (TP) as follows:
p≤0.005 for atg7d14/d77 and stamshR (second TP onward), atg7shR, atg6shR, uvragshR, and Rab7-DN (fourth TP onward). For the other TPs
P<0.05.
(C-E) Electron micrographs of cross-sections through fertilized at 10-40 min AEL showing (C) a wild-type MVC and (D and E)
biogenesis-deformed MVCs. See also Figure 8.

P a g e | 40
Figure 8. Compromised autophagy does not affect fertilized egg progression through the early embryonic stages
(A) Schematic illustration of the four main autophagic machinery units and complexes. The proteins which make each unit are depicted
as rectangles or circles. The bold (proteins) names correspond to the genes examined in this study.
(B-D) Dissecting scope images of embryos from the indicated genotypes and crosses (top). Note the presence of the cephalic furrow
(yellow arrows) in all embryos, which marks the gastrulation stage. Anterior is to the left.
(E) Quantification of the results described in (B-D). Note that although the atg7 mutant females lay fewer eggs than wild-type females,
fertilized atg7 mutant eggs develop normally at least until the gastrulation stage, at which time the MD is normally already eliminated.
(F) Quantification of fluorescence intensity in 4-minute intervals in live fertilized eggs mutant for the indicated genes/proteins. Error
bars represent standard error. The number (N) of examined embryos from each genotype is indicated in parenthesis.
(G) Validation of the potency of the different shRNA transgenes to efficiently knockdown the indicated genes in the egg. Reverse
transcriptase (RT-PCR) analyses on RNA from unfertilized eggs of the indicated genotypes are presented. The tubulin (tub) gene was
also amplified from each RNA sample to control for the presence and levels of RNAs.

P a g e | 41
Figure 9. Atg8 knock out attenuates PMD but other autophagy related genes don't
Quantification of fluorescence intensity in 4 min intervals in live fertilized eggs mutant for the indicated genes/proteins. Error bars
represent standard errors. The number (N) of examined embryos from each genotype is indicated in parenthesis. p values of the
differences in the fluorescence intensities between the mutants and the wild-type were calculated for each time point (TP) as follows:
p≤0.05 for atg81/2 (9th TP onward), atg2shR, atg9shR (second TP to 12th and 6th in accordance. For the other TP p>0.05). For atg2EP3697/Df,
vps34Δm22GLC p>0.05 for all TP.

P a g e | 42
5.6. The paternal MD is ubiquitinated through lysine-63-linked polyubiquitin side chains
soon after fertilization
Recent studies suggest that the ubiquitin system may be involved in the target specificity of cargo
cleared by selective autophagy (Kirkin et al., 2009b). In these paradigms, the targets are surface
ubiquitinated through non-classical ubiquitin side chains, such as lysine 63-linked polyubiquitin (K63-
polyUb) chains, which may preferentially bind to autophagy receptors, thus linking the ubiquitin
pathway with the autophagy pathway. To explore the possibility that the paternal MD may also be
ubiquitinated, fertilized eggs were stained with a linkage-specific antibody that recognizes
polyubiquitin chains joined through K63 of ubiquitin (Newton et al., 2008), and with the AXO 49
antiserum which detects the axoneme. Interestingly, already at 10-20 minutes AEL, one of the two
elongated sperm tail organelles was entirely decorated by K63-polyUb (Figure 10A and Figure 11A).
Although for technical reasons we could not detect both the K63-polyUb staining and the fluorescent
MD sperm at the same time (due to the insufficient fluorescence of the MD sperm after fixation), the
ubiquitinated organelle clearly corresponds to the MD, as evident in flagellar regions where the
ubiquitinated organelle (i.e. the MD) starts separating from the axoneme (Figure 10A, inset; see also
Figure 4, Figure 11B-C).

P a g e | 43
Figure 10. The MD is associated with lysine 63-linked polyubiquitin chains and p62 soon after fertilization
(A and B) Fertilized wild-type eggs at 10-30 minutes AEL were double stained to visualize K63-polyUb (red) and (A) the axoneme
(green) or (B) p62 (green). Anterior is to the left. Insets are enlargements of the boxed areas. Arrows point at (A) a region where the
MD was separated from the axoneme and (B) an MD fragment soon after the detachment from the main MD, whereas arrowheads point
at MD fragments distant from the main MD. The scale bars represent 50 µm.
(C) Quantification of fluorescence intensity in live fertilized eggs mutants for p62, eggs expressing the yeast DUB, UBP2 and eggs
compromised for hdac6 or key. The number of examined embryos from each genotype is indicated in parenthesis. All calculations were
performed as in Figure 7B. p-values are as follows: p62od2/od3: second to fifth TP, p<0.05, sixth TP onward p<0.005; UBP2: fourth TP,
p=0.05; fifth TP onward, p<0.05. hdac6shR: second to nineth TP, p<0.05, tenth onward p>0.05; keyshR: second to 14th TP, p<0.05, 15th
TP onwards, p>0.05.

P a g e | 44
The finding raised the question of whether the ubiquitination occurs before or after egg penetration.
Although staining of the testes revealed a strong K63-polyUb signal in elongating spermatids, this
signal completely disappeared during a later stage when the spermatids removed their bulk
cytoplasmic contents (i.e. the “individualization” stage; Figure 11D). Consistently, no staining was
detected from the male storage organ (i.e., the seminal vesicle) and the female storage organs (i.e.,
seminal receptacle and paired spermathecae) (Figure 11E, F). We therefore conclude that
ubiquitination of the MD occurs after egg penetration.

P a g e | 45
Figure 11. Ubiquitination of the MD occurs soon after fertilization and declines with time during the PMD process
(A-C) Confocal images of three wild-type eggs at the indicated times AEL (indicated on the left) stained to visualize the axoneme
(green; AXO; the AXO 49 antiserum) and lysine 63-linked polyubiquitin chains (anti-Ub-K63; red), which decorate the MD. Anterior is
to the left. Arrows are pointing at regions were the MD is completely detached from the axoneme. Note the gradual decrease in the level
of ubiquitination as PMD progresses. (D-F) Ubiquitination of the MD does not occur on late differentiating spermatids or mature sperm
before fertilization. (D) Terminally differentiation spermatids in the testis were double stained to visualize lysine 63-linked
polyubiquitin chains (anti-Ub-K63; red) and the individualization complex (IC) found in the waste bags (WB) and marks the late stage
of spermatid individualization (F-actin; green). Note the massive K63-polyUb staining before the individualization stage and its
elimination from individualized spermatid regions (arrows pointing at post-individualized regions). (E) Mature sperm from the male
seminal vesicle (SV) expressing the MD marker, DJ-GFP (green), do not display K63-polyUb staining (red). (F) Mature sperm in the
female storage organs (spermathecae and seminal receptacle) expressing the MD marker, MTS-DsRed (red), do not display K63-
polyUb staining (green). Scale bars: A-C, 100 μm; D-E, 50 μm.

P a g e | 46
5.7. The Ub-binding autophagy receptor p62 is recruited to the paternal MD
Autophagy receptor/adaptor proteins, such as p62/SQSTM1 and its structurally related protein,
NBR1, can interact with both polyubiquitin-tagged proteins and Atg8/LC3 (Pankiv et al., 2007; Kirkin
et al., 2009a; Bjorkoy et al., 2005). In Drosophila, the single p62 ortholog, called refractory to Sigma
P (ref(2)P; CG10360) (Nezis et al., 2008; Carre-Mlouka et al., 2007), was recently shown to mediate
the clearance by autophagy of mitochondria with enhanced levels of misfolded proteins (Pimenta, I et
al., 2012). To examine whether p62 may be recruited to the paternal MD, early fertilized eggs (10-30
minutes AEL) were double labeled with anti-Ub-K63 and anti-Drosophila p62 antibodies.
Importantly, p62 was specifically associated with the ubiquitinated MD, as well as some of the MD
fragments which were detached from the main derivative, suggesting a direct binding between the
ubiquitinated targets on the surface of the MD and p62 (Figure 10B).
5.8. The ubiquitin/p62 pathway is involved in PMD
Next, we wanted to examine the consequence of this ubiquitination on the PMD process. For this,
we maternally expressed the yeast deubiquitinating protease (DUB) UBP2, which has been previously
shown to antagonize some ubiquitination pathways in Drosophila neurons (DiAntonio et al., 2001)
(Baker et al., 1992). DUB-expressing eggs under a somatic expression element (UASt) displayed a
moderate yet significant delay in the clearance of the MD compared to wild-type eggs, suggesting that
ubiquitination is indeed involved in this process (Figure 10C). Unfortunately, the full effect could not
be examined, as increased expression levels of the DUB by using the oocyte and egg specific control
element, UASp, caused female sterility, likely due to a deleterious effect on egg development (data
not shown). Notably, other types of ubiquitinations of the MD surface proteins (i.e. through other
lysine side chains) could also be involved in PMD, hence would be less efficiently antagonized by
overexpression of the yeast UBP2.
Monitoring PMD in eggs from p62 mutant mothers (Nezis et al., 2008; Wyers et al., 1995) also
revealed only a moderate delay in the clearance of the MD (Figure 10C), as compared with the
autophagy and endocytic mutants (compare with Figure 7B). Although we used a viable
transheterozygous combination of two mutant p62 alleles, ref(2)Pod2 and ref(2)Pod3, they were shown
to be strong alleles of this gene, producing stable truncated p62 proteins (Nezis et al., 2008).
Therefore, the relatively weak effect may be attributed to the fact that other adaptor proteins could
also be involved in this process (Narendra et al., 2010a; Kirkin et al., 2009a; Wild et al., 2011;

P a g e | 47
Schweers et al., 2007). For example, the non-canonical histone deacetylase 6 (Hdac6) was shown to
bind ubiquitinated protein cargos with preference to K63-linked polyubiquitin chains while in parallel
also binding to dynein (Kirkin et al., 2009b). Indeed, Hdac6-deprived eggs caused some delay in
PMD, suggesting a possible role for the egg cytoskeleton in PMD (Figure 10).
Another pathway that could play a role in the recognition of ubiquitinated mitochondrial fragments
is the TBK1/Optineurin pathway, which was previously shown to promote xenophagy of Salmonella
in macrophages (Wild et al., 2011). To test this possibility, we first examined the upstream kinase
ortholog of TBK1 in Drosophila, Ik2. Since ik2 loss-of-function was lethal at the larval stage, we
overexpressed a GFP-tagged Ik2 in the egg, but detected no significant accumulation of GFP near the
sperm flagellum (data not shown). Optineurin has no known ortholog in Drosophila, but it's paralog,
the IκB kinase complex member IKKγ/NEMO (Zhu et al., 2007), has a fly ortholog called Kenny
(Key). Optineurin, NEMO and Kenny possess an N-terminal NEMO motif and a C-terminal
polyubiquitin binding element, UBAN (Shirinian et al., 2015). Interestingly, maternal knockdown of
key resulted in PMD attenuation, an effect that was even stronger than that of the hdac6 knockdown,
but somewhat weaker than that of the p62 mutant (Figure 10). Collectively, these findings suggest that
multiple ubiquitin-autophagy adaptor proteins may operate during PMD.
5.9. The sperm MD is depolarized
Our and other results suggest that ubiquitination of the sperm mitochondrial proteins after
fertilization may determine the specificity of the egg toward targeting the paternal and not its own
mitochondria. However, why the sperm and not the egg mitochondria are ubiquitinated after
fertilization still not understood. One possibility is that the sperm mitochondria lose its membrane
potential at a certain point before fertilization, thus specifying this organelle for ubiquitination and
subsequent degradation. Indeed, although immediately after fertilization in mouse the mitochondria
are marked by the membrane potential dye TMRE, gradual loss of membrane potential was
demonstrated at 30-48 hours post-fertilization, a time when the sperm mitochondria are readily
present and only begin to disperse in the egg (mitochondria in mice are eliminated at 60-84h post-
fertilization)(Rojansky et al., 2016).
To examine whether the sperm MD might also loses its membrane potential before or early after
fertilization, we labeled fertilized eggs with Mitotrackers (MTR, ThermoFishcer). To allow efficient
penetration of these probes to the live eggs, we gently permeabilized the eggs using surfactants

P a g e | 48
extracted from fruits, which removed the thin waxy layer that resides between the chorion and the
vitelline membrane (Rand et al., 2010; Rand, 2014). We then labeled the eggs with MTR green, which
binds to mitochondria regardless of their membrane potential (MP), and MTR red that labels
mitochondria with intact MP. However, the MD was not labeled by both MTRs, although the MTRs
efficiently penetrated the eggs, as revealed by the clear labeling of the maternal mitochondria (Figure
12A', A'' Figure 13A', A''). Similarly, negative labeling was also observed after labeling the sperm in
the female storage organs, the seminal receptacle and the spermathecae (Figure 12B, Figure 13B', B'').
In contrast, both MTRs readily labeled the MD in sperm from the male storage organ, the seminal
vesicle (Figure 12C', C'' for MTR green, Figure 13C', C'' for MTR red). We conclude that during the
transition from the male to the female, the sperm MD losses it membrane potential, thus when the
sperm enters the egg, the MD is already depolarized, which might trigger its subsequent
ubiquitination-mediated destruction.

P a g e | 49
Figure 12. Sperm mitochondrial derivative shows positive mitotracker green staining only in male stoage organ
(A-C'') Live labeling of embryos, female storage organs and male storage organ using the mitochondrial membrane potential
independent dye mitotracker green. (A'-A'') Two embryos fertilized by the mito-tdTomato sperm and gently peeled using the EPS
protocol to stain with mitotracker green. Anterior to the left.
(B) Dissected and torn female storage organ from (A), stained by mitotracker green.
(C'-C'') Mito-tdTomato filled sperm storage organ does stain by mitotracker green.
Scale bars as indicated in images.

P a g e | 50
Figure 13. Sperm mitochondrial derivative shows positive mitotracker red staining only in male stoage organ
(A-C'') Live labeling of embryos, female storage organs and male storage organ using the mitochondrial membrane potential
dependent dye mitotracker red. (A'-A'') Two embryos fertilized by the DJ-GFP sperm and gently peeled using the EPS protocol to
stain with mitotracker red. Anterior to the left.
(B) Dissected and torn female storage organs from (A), stained by mitotracker red.
(C'-C'') DJ-GFP filled sperm storage organ does stain by mitotracker red.
Scale bars as indicated in images.

P a g e | 51
5.10. Loss of Mul1 and Parkin significantly attenuates PMD
Genetic studies in Drosophila have placed the two fly orthologs of the Parkinson’s disease-
associated genes, pink1 and parkin, in a similar pathway: dispensable for the fly’s viability but
required for normal mitochondrial remodeling during flight muscle specification and spermatid
differentiation (Clark et al., 2006; Park et al., 2006; Greene et al., 2003). More recent studies in
mammalian cells uncovered a role of the Pink1/Parkin pathway in the turnover of damaged
mitochondria (Narendra et al., 2008; Narendra et al., 2010b). According to this model, Pink1 is
stabilized on the surface of damaged mitochondria, which then promotes the recruitment of Parkin to
the site and the subsequent clearance of these organelles by autophagy. Recent advancement in
understanding the role of Pink1 and Parkin in mitophagy came from the lab of Matsuda showing that
Pink1 phosphorylates Ubiquitin directly on Ser65 and this leads to Parkin activation and recruitment
to depolarized mitochondria (Koyano et al., 2014; Okatsu et al., 2015). It has also been reported that
Parkin mainly induces K63-linked chain ubiquitination (Lim et al., 2005; Olzmann et al., 2007;
Geisler et al., 2010). Therefore, we wanted to test the hypothesis that Parkin may be involved in the
process of PMD (possibly through ubiquitination of the paternal MD). Because the source of Parkin
can be either maternal or paternal, parkin mutant females (park25/Df, a null allelic combination (Greene
et al., 2003; Greene et al., 2003); see also lane 3 in Figure 14B) were first crossed with males
producing the MTS-DsRed sperm, and the clearance of the MD was monitored using the live imaging
assay. However, no significant effect on the clearance of the paternal MD was detected in these
mutant eggs (Figure 14C), suggesting that if Parkin is involved in this process it may originate in the
sperm.
We next setup to test for a possible effect on the kinetics of PMD when Parkin is inactivated in
the males. As opposed to the fertile females, parkin mutant males are sterile, displaying severe
spermatid differentiation defects, which hinder the accumulation of mature sperm in the seminal
vesicle (this study and (Greene et al., 2003)). To overcome this technical obstacle, we devised a
complex genetic scheme to allow inducible inactivation of Parkin at a late spermatid developmental
stage, thus still enabling normal spermatid differentiation and sperm production (Figure 14A). For
this, we modified a genomic clone containing the parkin gene, such that the engineered gene, t-parkin,
contains TEV protease cleavage sites within its open reading frame, and generated transgenic flies
carrying this construct in the background of the endogenous parkin mutant (park25/Z3-472). The ability
of t-parkin to restore functional sperm production and fertility in these mutant males was validated

P a g e | 52
(data not shown). Subsequently, another transgene consisting of the TEV protease gene under the
regulatory regions of the dj gene was generated and crossed to flies of the parkin rescue background.
The late cleavage of the t-Parkin protein was validated by Western blotting of testis and seminal
vesicle extracts (Figure 14B). Importantly, efficient inactivation/elimination of t-Parkin could be
detected in the seminal vesicle extracts which contain only the mature sperm (Figure 14B, lane 5).
The MTS-DsRed transgene was also crossed to these flies to mark the sperm MD, and these males
(i.e. with the following genotype: dj-Tev/t-park; park25/Z3-472, dj-MTS-DsRed) were shown to be fertile,
giving rise to viable progeny (data not shown). Finally, monitoring PMD in eggs which were fertilized
by the Parkin-devoid sperm revealed no significant effect on the clearance of the MD (Figure 14C).
Taken together, we conclude that at least by itself, Parkin does not play a major role in PMD.

P a g e | 53
Figure 14. Maternal and paternal Parkin are not involved in PMD
(A) Schematic view of the recombineering-based approach used to inactivate Parkin during late spermatogenesis. The relative region of
the inserted 3 x TEV-protease recognition sites is indicated in gray. parkin DNA and RNA coding exon sequences are shown in light
green bars, UTR regions in light blue bars, and introns in solid dark lines. The numbers of exons appear inside the bars. The targeting
(PCR) fragment which also includes the kanamycin resistant gene (KanR; red bar) flanked by LoxP sites (light blue rectangles) is
indicated without the flanking homology sequences. A light blue rectangle (a LoxP site) inside a yellow bar indicates the remaining
sequences after floxing. Parkin protein sequences are indicated by dark green bars and the relative locations of its different domains are
indicated in purple. (B) Cleavage of the t-Parkin protein was assessed by Western blotting of protein extracts from dissected testes
(denoted by blue text) or seminal vesicles (denoted by red text) of the indicated genotypes using the anti-Parkin antibody (top panels).
Note that the t-Parkin (lane 4) is larger than the endogenous Parkin (endo; lanes 1 and 2) because of the inserted TEV-protease
recognition sites in t-Parkin. Also note that no endogenous Parkin is detected in the park25/Df background (lane 3), and that after
expression of the TEV-protease, t-Parkin is cleaved (cParkin; middle panel; lane 4) and both t-Parkin and cParkin are almost completely
eliminated from mature sperm (lane 5). It is noteworthy that despite the cleavage of t-Parkin, the uncleaved form was still readily
detected in the testis: this is due to the fact that in addition to individualizing spermatids, the testes also contain germ cells of various
(earlier) stages, which express t-Parkin but still do not express the TEV protease, as the latter is not expressed until the onset of
spermatid individualization stage (lane 4). The same membrane was reblotted with the anti–β-tub antibody for loading control (bottom
panels). wt, wild-type. Molecular mass is indicated in kilodaltons.
(C) Quantification of fluorescence intensity in live fertilized eggs mutants for maternal and paternal parkin. The number (N) of
examined embryos from each genotype is indicated in parenthesis. All calculations were performed as in Figure 7B. The PMD kinetics
of the wild-type (wt) and mutant eggs were not statistically significant.

P a g e | 54
To identify other possible E3 ubiquitin ligases that could be involved in PMD, we tested
Drosophila orthologs of ubiquitin ligases that were previously shown to be involved in autophagy,
localized to the mitochondria, and/or involved in the turnover of unfolded proteins. We used specific
shRNAs against CHIP (carboxyl terminus of Hsc70-interacting protein), an important ubiquitin ligase
for selective autophagy in Drosophila muscles (Arndt et al., 2010), Sina (Seven in absensia), which
promotes Parkinson's disease progression by monoubiqutinating α-synuclein in mouse dopaminergic
neurons (Cai et al., 2015), and Traf6 (TNF receptor associated factor 6), which acts together with p62
to promote NFκB signaling and xenophagy of the invading bacteria (Dupont et al., 2009). However,
compromising these genes alone resulted in either very minor effect or no effect at all on the
attenuation of PMD (Figure 15).
A recent report revealed a Parkin-independent mitochondrial integrity pathway that could act in
parallel and compensate for the loss of Parkin in flies (Yun et al., 2014). The main effector protein in
this pathway is the mitochondrial E3 ubiquitin ligase Mul1 (also known as Mulan or MAPL). It was
shown that loss of Mul1 only mildly affected mitochondrial function, but the loss of Mul1 and either
Pink1 or Parkin resulted in a much higher level of mitochondrial dysfunction (Yun et al., 2014)(Figure
16A). A more recent work showed that Parkin, together with Pink1 and Mul1, has redundant function
in eliminating depolarized mitochondria in cell culture and during PMD after fertilization in mouse
Figure 15. Investigating more potential E3 ubiquitin ligases
Quantification of fluorescence intensity in live fertilized eggs compromised for chip, sina, and traf6 or wild-type eggs fertilized by
males with sperm compromised for mul1. The number of examined embryos from each genotype is indicated in parenthesis. All
calculations were performed as in Figure 7B. p-values are as follows: chipshR1: second to 15th TP, p<0.05, 16th TP onward p>0.05;
chipshR2: second to fifth TP, p<0.05; sixth TP onward, p>0.05; sinashR: second to seventh TP, p<0.05, eighth onward p>0.05; traf6shR:
All TP, p>0.05; mul1v109808
: All TP p>0.05; mul1EY12158: All TP p>0.05.

P a g e | 55
(Rojansky et al., 2016). We therefore looked in FlyBase for the distribution of the Drosophila Mul1
ortholog in the different tissues, and found that the mul1 mRNA is highly expressed in the testis.
To investigate whether Mul1 alone or in combination with Parkin could be involved in PMD, we
deleted the entire mul1 gene using the CRISPR/Cas9 procedure (Kondo and Ueda, 2013; Bassett et
al., 2013) (Figure 16B, lanes 10.3, 10.7 and Blast analyses in Figure 16B’). To avoid possible
unrelated background effects, we used a transheterozygous allelic combination of the mul1 CRISPR
allele (the Mul110.3) over an independently generated deficiency line (mul1Δ64) that carries a deletion
of the complete mul1 genomic locus (Slone et al., 2007) (Figure 16B’, Δ64 lane and Figure 16B’’).
We also used a more sensitive protocol to measure the kinetics of the PMD process, which includes a
modified confocal imaging procedure, aiming to reduce fluorescence bleaching by scanning in fewer
time points, using reduced gain and pinhole parameters, and extending the total measuring time.
Furthermore, we also generated a transgenic fly line carrying a brighter mitochondrially-targeted red
protein, the tandem dimer Tomato (tdTomato)(Shaner et al., 2005) (Figure 16C, D). The fluorescence
intensity was manually determined by the parallel observation of several embryos and subsequent
simple classification of the signal to either "strong", "weak" or "none" (Figure 16E).
Using this setup, we first examined possible effects on the PMD when either the females or the
males, or both were mutant for mul1 (Figure 16F, lanes 2,3 and Figure 16G, lane 1). Wild-type
females crossed to males expressing a single copy of the mito-tdTomato transgene were used as
negative control (Figure 16F, lane 1). For positive control, we crossed uvrag knockdown females
(described in section 5.5) with the mito-tdTomato transgenic males (Figure 16F, lane 6). Whereas
neither females nor males mul1 mutants exhibited PMD attenuation, a significant attenuation effect
was detected when both the males and the females were mutant for mul1, suggesting that the Mul1
source in the early zygote is from both the sperm and the egg (Figure 16F lanes 2,3 and Figure 16G,
lane 1).
Encouraged by the Mul1 effect on PMD, we then repeated the experiments with the parkin mutants
using the new settings. Similar to mul1 mutants, no effect on PMD was observed when either the
females or the males were compromised for Parkin (Figure 16F, lanes 3,4). However, when both the
males and the females were devoid of parkin, a significant attenuation effect was observed, indicating
that like Mul1, both the sperm and the egg contribute to the pool of Parkin in the early zygote (Figure
16G, lane 2).

P a g e | 56
Since both Mul1 and Parkin exhibited similar effects on PMD, we asked whether they function in
parallel or in a similar pathway during PMD. Females or males double mutant for both mul1 and
parkin displayed significant PMD attenuation when crossed to wild-type counterparts (Figure 16G,
lanes 5,6; note that the double mutant females displayed stronger attenuation than the double mutant
males). The attenuation levels observed in these double mutants were similar to those obtained in the
single mutants (when both the males and females were compromised for the same gene). Furthermore,
the attenuation level was not further aggravated when both the males and females were double
mutants (Figure 16G, lane 7). These findings suggest that Mul1 and Parkin may function in a similar
pathway to promote PMD, and that the source of these genes in the zygote is both the sperm and the
egg.
The finding that in the zygote mutations in at least 2 copies of either of these genes (regardless of
their origin in the male or the female) could lead to PMD attenuation, implies for redundancy between
Mul1 and Parkin. To determine what is the predominant origin of each of these genes, we examined
embryos derived from mul1 mutant females and parkin mutant males, and vice versa. Whereas both
combinations displayed significant PMD attenuation, embryos derived from mul1 mutant males and
parkin mutant females exhibited the highest PMD attenuation, even as compared to other mutant
combinations (Figure 16G, lanes 3,4). Collectively, these findings are consistent with the idea that the
origin of Parkin, which is primarily cytosolic, is mainly the egg, whereas the origin of Mul1, which is
located to the mitochondria, is the sperm mitochondrial derivative. Furthermore, the fact that the level
of the strongest PMD attenuation effect obtained with mul1 and parkin mutants is still considerably
lower than that obtained in the atg7 and uvrag mutants, could suggest that ubiquitination of the
paternal MD is not the major event in the PMD process, but rather that it may be required for the
second destruction stage, for the clearance of the MD fragments.

P a g e | 57
Figure 16. Mul1 as a possible E3 ubiquitin ligase important for PMD
(A) The E3 ubiquitin ligase Mul1 functions in parallel to Pink1/Parkin pathway (Yun et al., 2014). (B-B’’) Two Mul1 mutant alleles
that were used by us. Analyses of CRISPR mutant lines resulting from initial cross of gRNA and Cas9. Lanes 3, 6, are the F1 progeny
(marked by a the numbers 10,15 in accordance plus the word “genomic”), while each two lanes on the left of “genomic” lane are single
lines, without a balancer (B). Sequencing the Mul1 CRISPR deletion lines (10.3, 10.7) shows an exact cut of 914 bases resulting in
truncated Mul1 protein composed of its first 33 amino acids (B’). Mul1 deletion Δ64 that was used in trans with Mul1 CRISPR line
(Mul1 genomic location is marked with asterisk (CG1134), B’’). (C-D) Comparing MD-DsRed and two MD-tdTomato in early
fertilized eggs under light microscopy (C) and by using live imaging kinetics assay (D). The scale bars represent 50µm. (E) Three states
of MD-tdTomato that were scored in accordance – "Strong", Weak", None". (F) A graph showing 6 genotypes of wild-type, mul1 and
park mutant combinations in 3 time points 0,1 and 2 hours after egg laying. Each sample was examined under confocal microscope and
was given a score of “Strong” (=2 points), “Weak” (=1 point) and “None” (=0 points) according to examples shown in (E). Number of
embryos counted is marked in 0h lanes. To determine significance, each embryo's score at 2 hours after egg laying got a numerical score
as follows: Strong=2 points, Weak=1 point, None=0 points. According to this calculation there is ignificance (NS) between lanes 1 to 5,
but lane 6 was significantly changed compared to lanes 1-5 by p-value<0.001. (G) A graph showing 7 genotypes of mul1 and park
double-mutant combinations in 3 time points 0,1 and 2 hours after egg laying, as described in (F). Significance calculations shows that
accept sample 3 that is statistically significant than sample 6, all other samples are NS between each other.

P a g e | 58
5.11. Prohibitin, a major MD ubiquitination substrate after fertilization
To identify potential protein substrates for ubiquitination on the MD after fertilization, we have
been using two approaches, a candidate approach and an unbiased mass spectrometry approach. For
the first approach, the mitochondrially abundant chaperon, Prohibitin (PHB), was previously shown to
be ubiquitinated in both bull and monkey sperm mitochondria, albeit the relevance of this
ubiquitination for the PMD process was not determined (Thompson et al., 2003). To test whether
PHD ubiquitination is involved in PMD, we first identified the Drosophila orthologs of the two
mammalian Phb genes, Phb1 and Phb2, named l(2)37Cc and l(2)03709, respectively. The phb1 locus
encodes for two phb1 RNA isoforms, RA and RB, which only differ in their 5’ untranslated regions
(UTRs)(a third transcript has recently been added to the constantly updated version of FlyBase, called
transcript RD. RD contains the 5' UTR of RB but with a larger 3' UTR than RA and RB). Using RT-
PCR analysis on germ-less testes, we determined that the RA isoform is mainly expressed in the testis
(Figure 17A, B). Several phb1 mutant alleles have been reported and deposited in FlyBase (Stathakis
et al., 1995). Most of these alleles were recessive lethal during embryogenesis or early first instar
larvae, but we were able to also identify a hypomorphic allelic combination which is both viable and
male fertile. In this line, flies carry the weak phb123 allele over the strong phb14 allele. Whereas the
lesion in the phb14 mutant was shown (and confirmed by us) to carry an A-to-T transversion, resulting
in the elimination of the original first methionine, there was no record for the nature of the lesion in
the phb123 mutant, and we could not identify any lesion associated with the coding region or the
exons-intron junctions, suggesting that the lesion might lie within a control element of the gene.
We next crossed phb14/23 mutant males to wild-type females and for effects on the PMD process, as
well as on the ubiquitination state of MD after fertilization. Indeed, we recorded a moderate but still
significant PMD attenuation, which was not detected with the heterozygote males (phb14/+ and
phb123/+) (Figure 17C). In addition, the level of the MD ubiquitination after fertilization was
significantly reduced with the phb14/23 mutant males as compared with control males (Figure 17D, E).
Importantly, re-introducing a genomic BAC clone, encompassing the intact phb1 locus, to these
mutant males, restored normal PMD kinetics and MD ubiquitination level, suggesting that these
effects are directly related to the compromised phb1 (probably levels) on the MD. We conclude that
the paternal Phb1 is at least in part required for proper PMD and for the major ubiquitination events
on the MD after fertilization (Figure 17C, E).

P a g e | 59
Whereas the work by Sutovsky and colleagues suggested that Phb1 is the major ubiquitination
target on the sperm mitochondria (Thompson et al., 2003) (Artal-Sanz and Tavernarakis, 2009)), a
very recent work showed that it is the paralog protein, Phb2, that is a key mediator of both mitophagy
and allophagy (the specific elimination of the paternal mitochondria after fertilization in C. elegans)
(Wei et al., 2017). In this work, Phb2 was shown to act as a mitophagy receptor by directly binding to
LC3 upon mitochondria depolarization and proteasome-dependent outer membrane rupture. To test
whether Phb2 might also be involved in PMD in flies, we inactivated the second phb gene, l(2)03709.
Knockdown of phb2 resulted in male sterility, which hindered analysis of post-fertilization effects.
However, similar to the phb1 hypomorphic mutant combination, we identified an allelic mutant
combination, phb211345/Df, which despite displaying clear reduction in mRNA expression, gave rise to
viable and fertile males (Figure 18A, B). These flies carry the phb211345 allele, which carries a
transposable element in the 5'UTR of Phb2, over a deficiency allele that lacks the entire phb2 locus.
Nevertheless, no effect on PMD kinetics was recorded when these mutant males were crossed to wild-
type females. These results could be attributed to the mild nature of the mutant severity or to the fact
that both Phb1 and Phb2 function in a complex, and thus could exhibit some redundancy. Further
analysis of double mutant/knockdown combinations could shed light on the requirement of the
specific Phb proteins for PMD.

P a g e | 60
Figure 17. The mitochondrial chaperon Phb1 may serve as a substrate for ubiquitination on the paternal mitochondrion after fertilization
(A) Isoforms of l(2)37Cc. (B) RT-PCR of different tissue’s extracts showing that only the RA isoform of l(2)37Cc is expressed in
testes. (C) Quantification of fluorescence intensity in live fertilized wild type eggs and mutant sperm combinations for prohibitins. The
number (N) of examined embryos from each genotype is indicated in parenthesis. All calculations were performed as in Figure 7B.
Only l(2)37Cc4/23 combination is statistically significant. (D) Wild-type eggs fertilized by wild-type sperm and prohibitin mutant sperm
at 10-30 minutes AEL were double stained to visualize K63-polyUb (red) and the axoneme (green). Anterior is to the left. (E)
Quantification of biological repeats according to 3 criteria of K63-polyUb staining intensity: Non, Weak, and Normal staining.

P a g e | 61
Another, unbiased approach to identify ubiquitinated proteins on the mitochondria after
fertilization is to specifically isolate these ubiquitinated proteins and subject them to mass
spectrometry (MS) analysis. For this, we initiated a collaboration with Ugo Mayor (CIC Biogune,
Spain), who developed a technique to isolate ubiquitin conjugated proteins from late Drosophila
embryos (Figure 19A) (Franco et al., 2011). Concisely, the technique is based on a transgenic
expression of a construct comprised of the bacterial biotinylation enzyme, BirA, preceded by six
tandem copies of ubiquitin moieties, each ubiquitin is tagged with a biotinylation recognition site for
BirA. Once expressed, the BirA and the ubiquitin moieties are immediately separated from each other
by endogenous deubiquitinating enzymes (DUBs), which cleave after the BirA and between the
ubiquitin moieties. These tagged ubiquitin moieties are then incorporated to the growing ubiquitin
chains during normal endogenous ubiquitination events, while the BirA biotinilates the tagged
moieties, thus facilitating efficient immuno-precipitation of these ubiquitinated proteins with
streptavidin beads.
Figure 18. The 2nd Prohibitin homolog Phb2 is not involved in PMD
(A) Different allelic combinations of phb2 mutants examined by RT-PCR, (B) As opposed to phb1, compromised phb2 does not lead
to a delay in PMD

P a g e | 62
To facilitate expression of the Ub6-BirA construct during very early embryonic stages, we first
subcloned this construct under the UASp regulatory region and generated transgenic flies (Figure
19B, C). The expression of Ub6-BirA in fertilized embryos required more than 1gr’ of total protein to
obtain a robust biotinylation signal (Figure 19D-E). However, although a similar amount of protein
has been collected for non-fertilized eggs (served as control, Figure 19F), we could not detect a
biotinylation signal, indicating that most of the egg ubiquitination events are triggered only after
fertilization, thus cannot serve as proper negative controls in this experiment. We are therefore setting
up another strategy to specifically label the proteins that originate in the sperm by using the stable
isotope labelling by amino acids in cell culture (SILAC) technique that was also adopted to flies (Sury
et al., 2010). Concisely, wild-type males fed on isotope labelled yeast would be crossed to Ub6-BirA
females fed on regular yeast, thus allowing differentiation of the maternally versus paternally
originated ubiquitinated proteins. This approach shall identify other sperm mitochondrial proteins,
besides the Phbs, of which their ubiquitination after fertilization could promote efficient removal of
the sperm MD in Drosophila.

P a g e | 63
Figure 19. A proteomic approach to identify ubiquitinated sperm mitochondrial proteins
(A) Schematic illustration of the strategy for the in vivo biotinylation of ubiquitin (adapted from Franco et al., 2011). Digestion of the
Ub6-BirA precursor by endogenous DUBs is followed by BirA-mediated biotinylation of the ubiquitin chains tagged with the BirA
recognition sequence. The Ub6-BirA can then be incorporated by endogenous ubiquitin-conjugating enzymes (E1, E2, and E3) into their
protein substrates. (B) The scheme depicts the two constructs we generated for specific expression in the early fertilized eggs. (C)
Western blot analysis of wild-type (yw), BirA-expressing ((vp16-GAL4, UASp-birA)/TM6b) and Ub6-BirA-expressing ((vp16-GAL4,
UASp-Ub6-BirA/CyO) fertilized eggs at 1 AEL using an anti-biotin antibody. One strong endogenously biotinylated protein is
prominent both in wild-type and BirA extracts, but no additional bands are observed in the BirA lane. Biotinylated ubiquitin conjugates
appear as a smear in the Ub6-BirA lane, confirming that e Ub6-BirAis incorporated efficiently into different substrates. (D) Western blot
analyses performed on ~330 milligram of embryos from wild-typet (yw), Ub6-BirA fertilized and non-fertilized background, after
pulled-down, using anti Biotin antibody. A smear of biotinylated proteins appears only in the fertilized Ub6-BirA lane after IP. (E)
Western blot analyses performed on ~1200 milligram of embryos from wild-typet (yw), and triplicates of Ub6-BirA fertilized and non-
fertilized background, before (left) and after (right) pulled-down, using anti Biotin antibody. A smear of biotinylated proteins appears
only in the fertilized Ub6-BirA lanes before and after IP. (F) Silver stain of matching samples as described in (E) showing same loading
amounts of proteins in all lanes.

P a g e | 64
5.12. Investigate the role of the MVB-like vesicles in the destruction of the paternal
mitochondria
Our work suggested an important role for the MVB-like vesicles in targeting the sperm and
triggering the initial destruction of the MD after fertilization. Although MVBs display similar
morphology in many cell types, the contents of their microvesicles may be distinct from one cell to
another according to their specific functions. Some MVBs secrete their microvesicles to the
extracellular domain as exosomes (Corrigan et al., 2014). By way of similarity, the MVB-like vesicles
in the fly egg secrete their microvesicles outside of the egg, but in this case, into the sperm.
Interestingly, when we examined electron microscopy images of fertilized mammalian eggs in the
published literature, we noticed the presence of MVBs in the vicinity of the sperm after fertilization in
hamster, cow, and monkeys, suggesting that the role of the MVBs in PMD might be conserved in
mammals (Hiraoka and Hirao, 1988; Sutovsky et al., 2000; Politi et al., 2014) (Figure 20A-C).
To start investigating the specific role of the MVBs in PMD, we first examined the association of
these vesicles with the sperm plasma membrane along almost the entire 2 mm long sperm tail. This
was performed using the FIB (focused ion beam) technique, which enables high quality 3D analysis of
EM data. We took hundreds of consecutive SEM images in a chosen area of an early fertilized egg
and created a 3D reconstitution image/video of that area. Importantly, this analysis revealed that the
sperm tail is associated with several MVBs in parallel, and that in these areas of association, the sperm
plasma membrane appears highly swelled and irregular, suggesting that the MVBs may initiate the
disintegration of the sperm plasma membrane (Movie 2).
Figure 20. Approaching of MVBs to the sperm flagellum after fertilization is evolutionary conserved
(A-C) Representing electron micrographs showing accumulation of MVBs and lysosomes (B’) near and on the sperm
flagellum after fertilization in; hamster (A), cow (B) and fruit fly (C).

P a g e | 65
We further characterized the MVBs and their interacting components with the sperm after
fertilization. First, we examined fertilized eggs derived from males expressing a plasma membrane-
tethered GFP (CD8-GFP) and showed that the sperm enters the egg when it is still enveloped in an
intact plasma membrane (Figure 21A). Since we found that the MVBs fuse to the sperm plasma
membrane and secret the microvesicles contents to the sperm, we hypothesized that this process might
require the function of SNARE proteins, the known mediators of vesicles fusion with the plasma
membrane. Furthermore, a recent report also suggested a role of several SNAREs in the fusion of
autophagy vesicles to lysosome in Drosophila (Takats et al., 2013). Out of four different egg
knockdowns genes we tested, only two has not resulted in female sterility, and one these gene's
mutant eggs exhibited a significant PMD attenuation, indicating an important role of SNAREs in
PMD (Figure 21B, C).

P a g e | 66
Figure 21. SNARE proteins in PMD
(A) A frame from a two 2hr movie of egg from a wild-type female fertilized by a male with CD8-Venus sperm. Anterior is to the left.
(B) Quantification of fluorescence intensity in live fertilized eggs compromised for vamp7 and ykt6. The number of examined embryos
from each genotype is indicated in parenthesis. All calculations were performed as in Figure 7B. p-values are as follows: vamp7shR1:
second to fifth TP, p<0.05, sixth TP onward p>0.05; ykt6shR1: second to 20th TP, p<0.05; 21st TP onward, p>0.05.
(C) Validation of the potency of the different shRNA transgenes to efficiently knockdown the indicated genes in the egg. Reverse
transcriptase (RT-PCR) analyses on RNA from unfertilized eggs of the indicated genotypes are presented. The tubulin (tub) gene was
also amplified from each RNA sample to control for the presence and levels of RNAs.

P a g e | 67
To further characterize the role of these special MVBs for PMD, we inactivated several upstream
and downstream components in the endocytic pathway. Knockdown of the ESCRT-I associated
protein tsg101/vps23 (which follows ESCRT-0, see 5.5), the lysosomal biogenesis related protein,
Deep-orange (Dor/Vps18; a component of the homotypic fusion and vacuole protein sorting - HOPS -
complex), and a mutant of the endocytic trafficking protein, SpnF (which is located at the minus-end
of the oocyte microtubule network and is important for proper bristle formation (Abdu et al., 2006;
Dubin-Bar et al., 2008; Bitan et al., 2010)), all significantly attenuated PMD (Figure 22). Importantly,
overexpression of SpnF-GFP in the early embryo showed a remarkable accumulation of GFP near and
in association with the sperm MD, although non-sperm associated SpnF-GFP vesicles were also
detected (Figure 22A). In contrast to Tsg101, Dor and SpnF, no major effect on the PMD process was
detected in knockdown of another component of the lysosomal HOPS complex Carnation
(Car/Vps33A), as well as a mutant of a major lysosomal hydrpolase, CathepsinD (CathD), suggesting
some divergence from the canonical role of the lysosomes in autophagy (Figure 22). Collectively,
these results are consistent with our previous results (i.e. Stam/Vps27 and UVrag), suggesting an
important role of the endocytic pathway in PMD.
Figure 22. Diverse endocytosis related genes participitate in PMD
Quantification of fluorescence intensity in live fertilized eggs compromised for tsg101, dor, car and ykt6 or egg knockout for cathD and
spnF. The number of examined embryos from each genotype is indicated in parenthesis. All calculations were performed as in Figure
7B. p-values are as follows: tsg101shR: second to 17th TP, p<0.05, 18th TP onward p>0.05; dorshR: second to 15th TP, p<0.05; 16st TP
onward, p≈0.05; carshR: second TP to third, p<0.05, fourth TP onward p>0.05; cathD1: All TP, p>0.05; spnFAA/3325: second TP to 21st
p<0.05, 22nd TP onward p>0.05.

P a g e | 68
Our findings that the endocytic pathway is important for PMD and that at least in two of the
mutants that we examined (i.e. atg7 and uvrag), the MVBs fail to generate microvesicles, imply that
the content of the microvesicles could be instrumental for the PMD process. To test this idea, we
aimed to isolate the MVBs and devised two approaches to do so. The first approach involves
modifications to classical protocols for extracellular vesicles (exosomes) isolation (Gross et al., 2012).
This procedure includes gradual centrifugations of extracted proteins, followed by
ultracentrifugation and separation of the fractions by sucrose gradient (Figure 23A). Western blot
analysis was performed to estimate the presence of extracellular vesicles (Figure 23B). Another
marker that was used, the protein GAPDH, was previously shown to be localized to some MVBs, in
addition to its cytoplasmic localization (Sahu et al., 2011). Both Atg8 and Rab7 positive fractions
were eluted in the same sucrose concentration (Figure 23B, lanes 2 and 10).
To assess the validity of our procedure and its ability to identify relevant MVB content proteins,
we sent the isolated sample to the MS unit at the Technion. 18 different proteins were identified by 2
or more unique peptides. These include 7 proteins that are known to be involved in protein
degradation processes, which we further examined for possible involvement in PMD (Figure 23C).
Perhaps the most interesting protein in this list is dBruce, which also appeared in a preliminary
analysis of the ubiquitinated mitochondrial proteins (data not shown). dBruce is an E2/E3 Ub ligase of
the inhibitor of apoptosis (IAP) protein family and, among other roles, is required for late
spermatogenesis stages in flies (Arama et al., 2003; Kaplan et al., 2010). In addition, during
oogenesis, dBruce is degraded by autophagy to promote caspase activation and DNA fragmentation in
nurse cells (Nezis et al., 2010). Interestingly, whereas males carrying a hypomorphic allele of dbruce
(dbrucee101) barely affect PMD when crossed to wild-type females, overexpression of dbruce in the
males resulted in enhanced PMD (Figure 24A). On the other hand, knockdown of dbruce in the egg
showed no effect compared to control when estimated in the 0,1,2h assay (Figure 24B). Staining early
fertilized embryos with an anti-dBruce antibody (Kaplan et al., 2010), revealed a specific expression
pattern on the sperm axoneme (Figure 24C-E). In non-fertilized eggs, dBruce was localized at the
anterior part of the egg (Figure 24F). We conclude that although dBruce does not appear to be an
executioner of PMD (having a direct role in the destruction of the MD), it might promote the specific
degradation process through its own ubiquitination.
In addition to dBruce, the other 6 proteins have been also examined, however, egg knockdown of
the proteasomal related genes prosα6, prosβ1 and prosβ4 resulted in female sterility, thus hindering
further analysis of specific effects on PMD (data not shown and text in 5.11). In contrast, egg

P a g e | 69
knockdown of Poe (an E3 Ubiquitin Ligase protein), Rpn9 (non-ATPase regulatory part of the
proteasome) and USP7 (ubiquitin specific protease), produced fertile female and are currently under
examination.
Figure 23. First MVBs isolation strategy – ultracentrifugation and sucrose gradient
(A) Differential centrifugation protocol for the isolation of extracellular vesicles from cell culture supernatants (adapted from Gross et al.
2012). One asterisk marks extracellular vesicle fraction pre-sucrose gradient stage and two asterisks mark the sucrose gradient fractions.
(B) Western blot analysis of wild-type (yw), YFP-Rab7 expressing and GFP-mCehrry-Atg8a fertilized eggs during 2 hours AEL. Using
an anti GFP antibody that detects both Rab7 and Atg8a (markers of late endosomes, black arrow. See also Figure 6D and 6E). The
GAPDH antibody marks the cytosol but also the extracellular vesicles themselves. Lanes 1-3 represent the extracellular vesicles after
100,000g centrifugation. Lanes 4-8 and 9-11 represent fractions from sucrose gradient of Atg8a and Rab7 (respectively).
(C) ) A narrowed list of selected proteins found after mass-spec analyses of extracellular vesicles enriched fraction as described in B.

P a g e | 70
Figure 21. dBruce plays an illusive role in PMD
(A) Quantification of fluorescence intensity in live fertilized wild-type eggs and males with either mutant dbruce or overexpression of
dBruce.. The number of examined embryos from each genotype is indicated in parenthesis. All calculations were performed as in
Figure 7B. p-values are as follows: dbrucee101: All TP, p>0.05; dBruceoverexpression: second to sixth TP, p>0.05, seventh TP onward,
p<0.05.
(B) A graph showing 3 genotypes of wild-type, dbruce and tbc1d15 female knockdown in 3 time points 0,1 and 2 hours after egg
laying, like described in figure 14F. According to this calculation there is no significance (NS) between lanes 1 to 2, but lane 3 was
significantly changed compared to lanes 1 and 2 by p-value<0.05.
(C-F) Confocal images of wild-type eggs stained to visualize the axoneme (green; AXO; the AXO 49 antiserum) and dBruce (anti-
dBruce; red). Anterior is to the left. Various antibody fractions were used to stain dBruce in fertilized eggs showing strongest
immunogenicity in serum fraction (C), weaker in 2nd blead fraction (D) or none in pre-serum fraction (E). (F) Non-fertilized eggs
stained as in (C), showing no axoneme (green) staining and scattered dBruce staining (red).

P a g e | 71
Our second approach to isolate the MVBs from the eggs combined gradual centrifugations (as
described above) with immunoprecipitation of the MVBs/microvesicles. For this, we first examined
several markers of the microvesicles. Human CD63 (hCD63) is a known marker of exosomes in
cultured human cells. Recently, it was shown that an EGFP-hCD63 transgene can specifically mark
MVBs and their microvesicles in some Drosophila tissues (Corrigan et al., 2014; Panakova et al.,
2005). We thus generated transgenic flies expressing a construct comprising the EGFP-hCD63 under
the UASp regulatory region for efficient maternal expression (see also Figure 19B). Consistent with
our previous findings that the MVBs associate with the sperm flagellum, when expressed in the
fertilized eggs, the EGFP-hCD63 was localized to vesicles that accumulated near and on the sperm
flagellum (Figure 22B). Co-expression of EGFP-hCD63 and Atg8-mCherry in the fertilized egg
revealed co-localization of both proteins in the vesicles at the vicinity of the sperm flagellum (Figure
22C). To functionally characterize the CD63 positive vesicles and Atg8 positive vesicles, we extracted
protein from mCherry-Atg8 and EGFP-CD63 co-expressing eggs (as in Figure 22C), followed by
gradual centrifugations and ultracentrifugation (as described in the above protocol). This sample was
subjected to Western blotting using an anti-GFP or anti-mCherry antibody. The band representing
CD63 or Atg8 was clearly visualized in some of the lanes, but in EGFP-CD63 it seems that the
majority of CD63 positive vesicles is found in pellet fractions and faintly in the last ultra-pellet
fraction, as opposed to the Atg8 positive vesicles that seemed, at least in part, to reside in supernatant
fractions until precipitated in the desired ultra-pellet fraction (Figure 22D and E). We further analyzed
the pellet of the ultracentrifugation by the NanoSight device (NS300, Malvern), which evaluates the
size distribution and concentration measurements of particles in liquid suspension. Importantly, the
samples examined were mostly composed of small vesicles with the size of about 150 nm, which is in
agreement with the accepted size of microvesicles (Figure 22F right; compare with middle and left
graphs). In summary, despite co-localization of CD63 positive vesicles with the outer layer of MVBs,
PMD related vesicles are best represented by Atg8. Currently, we collect high amounts of embryos
from each genotype in order to perform the immunoprecipitation and eventually proteomics
evaluation of the vesicles' contents.

P a g e | 72
Figure 22. Second MVBs isolation strategy – gradual centrifugations and marker based IP
(A-C) Live imaging confocal images of egg expressing SpnF-GFP (A), EGFP-hCD63 alone (B) or together with mCherry-Atg8 (C) and
fertilized with the MD-DsRed sperm (A, B) or no-label sperm (C). Anterior to the left.
(D-E) Evaluating the extraction efficiency of vesicles by Western blotting of the matching genotype and extraction method as written i n
each lane’s title using the anti-GFP antibody (D) or anti-mCherry antibody (E). Note that EGFP-hCD63 band is 52 kDa and mCherry-
Atg8 is 42kD. S1 – supernatant after 1st centrifugation, P1 – pellet of the same fraction (S2, P2, S3 and P3 are matching lanes for 2nd and
3rd centrifugation in accordance). Ultra sup – supernatant after 3hour ultracentrifugation in 100,00 g, ultra pellet – resuspended sample
from pellet of above described ultracentrifugation. Molecular mass is indicated in kilodaltons.
(F) Analyzing 1:1000 diluted portions of Atg8 positive vesicles using NanoSight. Left – post supernatant 3, middle – after filtration and
before ultracentrifugation, right – pellet of ultracentrifugation The score in the bottom is a summary of 5 movies, 1 minute long each, in
which the machine measures all the movements of the particles run in the frame.

P a g e | 73
5.13. The role of the mitochondria fusion/fission machinery on PMD
Selective autophagy requires prior fission of the mitochondria presumably in order to facilitate
enwrapping of smaller fragments by autophagosomes. This may be particularly relevant in the case of
the paternal MD in Drosophila, which is 1.9 mm long and which we found to undergo fragmentation
prior to clearance by autophagosomes. Therefore, we aimed to genetically alter the balance between
the pro-fusion and pro-fission mitochondrial gene expressions, as was previously reported in other
systems, in order to examine possible involvement of this system in PMD. For this, we first
overexpressed fuzzy onions (fzo), the testis specific mitochondrial pro-fusion gene, in the early egg
using a weak maternal driver, as stronger expression of Fzo and Marf (the somatic mitochondrial
fusion protein) caused female sterility probably due to extreme fusion of the maternal mitochondria.
However, we could not detect any significant effect on PMD, which could be attributed to the weak
expression of this factor (Figure 23A, B). We then tried to unbalance the other side of the equation,
namely knocking down the major fission gene, dynamin related protein (drp1), which was previously
reported to be stabilized on damaged mitochondria by Mul1, and in addition, serves as a mitophagy
promoting agent through PINK1/Parkin pathway (Deng et al., 2008; Braschi et al., 2009). However,
compromising drp1 in the eggs resulted in some PMD attenuation that was not consistent (Figure
23A). These results suggest that the canonical mitochondrial fusion/fission machinery may not drive
the breaking of the MD after fertilization. To complete these studies, we are now also examining
possible involvement of TBC1D15 and Fis1, two mitochondrial proteins involved in shaping
mitochondrial morphology (Onoue et al., 2013)(See also 5.5).

P a g e | 74
Figure 23. A mild effect for drp1 knockdown maternaly in PMD
(A) Quantification of fluorescence intensity in live fertilized eggs with either drp1 knockdown or overexpression of Fzo. The number
of examined embryos from each genotype is indicated in parenthesis. All calculations were performed as in Figure 7B. p-values are as
follows: drp1shR: second to 15th TP, p<0.05, 16th TP onward, p>0.05; Fzooverexpression: second to nineth TP, p<0.05; 10th TP onward,
p>0.05.
(B) Validation of the efficiency of ectopic Fzo expression in the egg. Reverse transcriptase (RT-PCR) analyses on RNA from
unfertilized eggs of the indicated genotypes are presented. The tubulin (tub) gene was also amplified from each RNA sample to control
for the presence and levels of RNAs.

P a g e | 75
6. Discussion
6.1. A working model for PMD, hypotheses and open questions
Our results thus far demonstrate that the paternal MD in Drosophila melanogaster is rapidly
eliminated after fertilization by active mechanisms. Our study supports a multistep model for this
process which we also termed PMD. After sperm penetration of the egg (Figure 27I), a network of
MVB-like vesicles originating in the egg and displaying molecular features of both the endocytic and
autophagic pathways associates with the sperm tail (yellow arrows in Figure 27II). These vesicles are
involved in the initial recognition and disintegration of the sperm plasma membrane after fertilization,
and may also mediate the clipping of the sperm flagellum at different regions along the tail, as
vesicles that completely encapsulate flagellar regions (containing the sperm plasma membrane,
axoneme and MD) are detected by both electron and confocal microscopy (white arrow in Figure
27II). MVCs making direct contacts with sperm plasma membrane were often detected under the EM,
and may also contribute to the membrane disintegration and the subsequent MD separation from the
axoneme, as they were only detected prior to the separation of these two organelles (arrowhead in
Figure 27II).
Another event that occurs before the separation of the MD from the axoneme is the specific
ubiquitination of the MD throughout its length and recruitment of p62, although it is unclear whether
the ubiquitination occurs before or immediately after the engagement of the vesicles with the sperm
(Figure 27II). The ensuing stages initiate at different points along the flagellum and proceed in a
stepwise manner. The MD separates from the axoneme and breaks into small fragments (Figure
27III), which are then enwrapped by autophagosomes (Figure 27IV) and sequestered for degradation
in lysosomes (Figure 27V). The mechanism which facilitates mitochondrial fragmentation is
unknown; however, it may involve the mitochondrial fusion/fission machinery, as it has been
previously implicated in promoting the unique structural organization of the spermatid mitochondria
(Aldridge et al., 2007; Hales and Fuller, 1997). It is noteworthy that during this relatively rapid
process, the axoneme remains largely intact (Figure 27). The elimination of this structure in D.
melanogaster appears to largely occur via its sequestration to the developing gut region (Pitnick and
Karr, 1998).

P a g e | 76
Figure 27. A model of the distinct steps of the PMD process
Sequential schemes of the different steps during the PMD process in Drosophila. Refer to the main text for detailed explanations of
each step

P a g e | 77
6.2. The significance of paternal mitochondrial ubiquitination and association with p62 and
LC3/Atg8
Decoration of the paternal MD by K63-linked polyubiquitin chains and the subsequent association
with p62 and Atg8 occur early during the PMD process at a stage when the MD is still much too large
to be enwrapped by autophagosomes, suggesting that these events may serve another role different
than marking the mitochondria for selective autophagy. Consistent with this idea, our findings support
an unconventional role of a common autophagic and endocytic vesicular pathway in promoting the
initial disintegration of the sperm tail and separation between the MD and the axoneme, both of which
occur at a stage when the MD is already ubiquitinated. Furthermore, we demonstrated that Parkin
alone, which has been shown to be sufficient for targeting damaged mitochondria for selective
autophagy in both mammals and Drosophila (Pimenta, I et al., 2012; Narendra et al., 2008) does not
play a major role during PMD. Instead, we showed that Parkin acts synergistically with another E3
ubiquitin ligase, Mul1, and together their inactivation caused a significant delay in PMD when
compromised both in males and females, with the strongest PMD attenuation observed when the
maternal Parkin and male Mul1 were compromised. Interestingly, Mul1 was previously also shown to
compensate for the loss of Pink1/Parkin in both Drosophila and mammals, suggesting a redundancy
between these two ubiquitin ligases (Yun et al., 2014). Moreover, a very recent work showed that both
Parkin and Mul1 are both required for mitophagy in mouse cell culture and during paternal
mitochondrial elimination in mouse embryos (Rojansky et al., 2016).
Ubiquitination of sperm mitochondria via K63-linked polyubiquitin chains, and p62 and LC3
association were also reported to occur soon after fertilization in several mammalian systems. This is
an important point, since a recent report suggested a passive model of mitochondrial dispersal to
explain paternal mitochondrial elimination after fertilization in mice (Luo et al., 2013), even though
they also noted an early process of sperm tail disintegration which facilitates the subsequent dispersal
of the mitochondria (Al Rawi S. et al., 2011; Sutovsky et al., 2003; Sutovsky et al., 2004; Sutovsky et
al., 1999; Sutovsky et al., 2000; Luo et al., 2013). Indeed, this may also be in agreement with their
finding that atg5 knockout mice displayed no effect on the kinetics of the sperm mitochondrial
dispersal, implying a nonorthodox role of some elements in the autophagy system but not others
during this process in mammals (Luo et al., 2013). Alternatively, this may also be attributed to
technical issues as mouse embryos deprived of core autophagy components fail to develop beyond the
4-cell stage (Rojansky et al., 2016). Furthermore, in organisms which produce non-flagellated sperm
with already dispersed mitochondria, such as C. elegnas, this pathway does not exist, and the

P a g e | 78
mitochondria are directly targeted for clearance by autophagy (Al Rawi S. et al., 2011; Sato and Sato,
2011). Moreover, the Sutovsky lab showed that in addition to LC3 and p62, the ubiquitin binding
dislocase, valosin-containing protein (VCP), mediates the presentation of ubiquitinated sperm
mitochondrial proteins to the 26S proteasome system to facilitate sperm mitophagy after fertilization
in mammals (Song et al., 2016). Therefore, and especially in the case of the giant MD in Drosophila,
ubiquitinated paternal mitochondrial proteins might not only serve as a signalling tag for the
destruction machinery, but could also function as plugs that must be removed to allow proper PMD
progression.
Which sperm mitochondrial proteins are ubiquitinated after fertilization? Our results suggest that
the mitochondrial chaperon, Phb1, may be a major candidate, albeit it may not be the sole substrate
for ubiquitination on the MD. Fertilized eggs derived from males that carry a mutant allelic
combination for phb1, exhibited low levels of K63-type ubiquitin side chains on the MD after
fertilization. However, Phb1 could also indirectly affect PMD by virtue of its function as a chaperon,
which might modulate the correct folding of other mitochondrial proteins, including some
ubiquitination substrates. Consistent with this idea, our (preliminary) proteomics analysis of
ubiquitinated MD proteins failed to identify Phb1. Among the two Phb paralogs, Phb2 was recently
shown to act as a major autophagy receptor, directly binding to LC3. This binding occur both
following mitochondrial depolarization that precedes Parkin recruitment to the mitochondria, and
during the elimination of the paternal mitochondria after fertilization in C. elegans (Wei et al., 2017).
In these setups, Phb1 does not directly bind to LC3, but rather indirectly associates with LC3 through
its binding to Phb2 (Wei et al., 2017). Further examination of the specific ubiquitination states of
Phb1 and Phb2 on the paternal mitochondria by mass spectrometry shall help revealing whether Phb is
required directly or indirectly for mitochondrial protein ubiquitination, and which Phb paralog (or
maybe both) is important for PMD.
6.3. The role of the MVBs and their microvesicles in PMD
In the past few years there is growing interest in understanding the roles of extracellular vesicles
(EVs)(Yuana et al., 2013; Buzas et al., 2014). EVs diverge in size and function and were shown to be
involved in a wide range of cell-cell communication systems, in addition to their well characterized
role as neurotransmitter carriers in synapses (Budnik et al., 2016). EVs were found to carry diverse
substances, including protein receptors, miRNAs, and even fragments of genomic DNA, and these

P a g e | 79
occur in a variety of processes, such as tumorigenesis and metastasis (Melo et al., 2014; Hoshino et
al., 2015), Malaria parasites cross talk (Regev-Rudzki et al., 2013), alga-viral infection (Schatz et al.,
2014), and during fertilization in mammals (Al-Dossary et al., 2015). Our finding that MVBs
associate with the sperm immediately after fertilization and secrete their microvesicles to the
flagellum suggests an important role for these microvesicles in the initial steps of the PMD process. In
addition, we showed that the autophagy and endocytic pathway mutants which caused a significant
attenuation in PMD dynamics, also displayed sever defects in the biogenesis of these microvesicles,
further suggesting the importance of these microvesicles and their contents for this process. Our
preliminary proteomics analysis on the extracted MVBs suggests an interesting role for the
proteasome in PMD, perhaps in the local removal of specific proteins on the MD, which facilitates the
fragmentation of the MD before the clearance by autophagy. Other possible protein contents may
include additional proteins that are involved in PMD, such as E3 ligases, mitochondrial fission
proteins etc. Large scale and targeted isolation of these vesicles by centrifugations and IP shall shed
light of their role in promoting the PMD process.
6.4. Why is the paternal mitochondria eliminated after fertilization?
Several hypotheses have been proposed to address this question, but the exact answer still remains
a mystery. One idea is that the egg may regard the paternal mitochondria as a potentially dangerous
trespasser, similar to the way somatic cells react against intrusive bacteria. Indeed, the mitochondria
are believed to originate from a bacterial endosymbiont (Andersson et al., 1998). Furthermore,
microbes that invade cells during infection are selectively disposed by a specialized form of
autophagy called xenophagy (Levine and Deretic, 2007; Schmid and Munz, 2007). However, the fact
that maternal mitochondrial inheritance has been conserved throughout eukaryotic evolution, strongly
suggest that this phenomenon is not merely a preserved remnant of a primitive defense mechanism,
but rather reflects a crucial need to prevent the delivery of certain paternal mitochondrial factors to the
developing embryo. Along these lines, it is widely believed that elimination of the paternal
mitochondria may be important to prevent mtDNA heteroplasmy, which in extreme cases may cause
mitochondrial disorders (Kraytsberg et al., 2004; Montoya et al., 2009). Furthermore, inducing
mtDNA heteroplasmy in mice was recently reported to result in significant physiological, cognitive,
and behavioral complications (Kraytsberg et al., 2004; Montoya et al., 2009; Sharpley et al., 2012). It
is still not fully understood why mtDNA heteroplasmy can cause such deleterious effects to the
organism, but it is known that mitochondrial DNA has a much higher mutation rate than nuclear DNA

P a g e | 80
because it lacks protective histones, is readily exposed to damage from ROS production and lacks
certain DNA repair mechanisms. Secondary mitochondrial dysfunction results from the influence of
external mechanisms such as environmental or pharmacologic toxins that can damage the mtDNA.
Mitochondria can protect themselves from the accumulation of damage through various quality
control mechanisms (e.g., fission and fusion) however, if these mechanisms are altered, mitochondrial
dysfunction can lead to disease (Filler et al., 2014). Also, it has been suggested that sperm mtDNA has
a higher mutation rate than the egg mtDNA, and that spermatozoa even from normal men may contain
significant levels of mtDNA deletions (Cummins, 2000). In addition, sperm mtDNA damage was also
attributed to reactive oxygen species which are involved in fertilization (Ford, 2004). However, the
possibility of increased accumulation of mutations in the sperm mtDNA may not be the only reason
for the deleterious effects of heteroplasmy, as even under conditions in which the two mixed mtDNA
were normal, induction of heteroplasmy was shown to be genetically unstable and to produce adverse
physiological effects (Sharpley et al., 2012).
Prevention of mtDNA heteroplasmy by itself may not be the entire story, as paternal mitochondrial
factors other than mtDNA may be also detrimental to the developing embryo. Indeed, the estimated
number of maternal mtDNA copies in the animal egg far exceeds that of sperm by factor of 103-106,
indicating that most of the sperm mtDNA is already excluded during sperm differentiation (Birky, Jr.,
2001). Furthermore, elimination of the sperm mtDNA in the Japanese medaka fish has been reported
to happen through unknown mechanisms in two steps, during spermatogenesis and immediately after
fertilization, but before the destruction of the vacuolar mitochondria (Nishimura et al., 2006).
Moreover, recent works in Drosophila, C. elegans, and mouse, showed that the mtDNA is actively
removed before or immediately after fertilization. In Drosophila, it was demonstrated that essentially
all of the sperm mtDNA is already degraded during late spermatogenesis by the actions of both the
mitochondrial endonuclease, EndoG, and the nuclearly encoded mitochondrial DNA polymerase,
Tamas (DeLuca and O'Farrell, 2012) (Yu et al., 2017). In C. elegans, CPS-6, the EndoG ortholog, was
found to relocate from the intermembrane lumen of the sperm mitochondria to the matrix
compartment in order to facilitate mtDNA elimination prior to the degradation of the sperm
mitochondria via autophagy (Zhou et al., 2016). In mouse, it was reported that most of the mtDNA is
already eliminated prior to fertilization in the majority of the sperm cells (Luo et al., 2013). In light of
the idea that the paternal mitochondria lose membrane potential before or soon after fertilization (the
current work and work of others in mouse), it is possible that the cause for the paternal mitochondrial
damage might be the elimination of the mtDNA. Taken together with the idea that active mechanisms

P a g e | 81
in the egg operate to eliminate the mtDNA devoid paternal mitochondria, it is attractive to speculate
that the degradation of the mtDNA could trigger the chain of events that eventually results in PMD
(i.e. mtDNA degradation - gradual depolarization - ubiquitination - degradation).
Another way of thinking about PMD is that the paternal mitochondria may not necessarily be
deleterious to the developing embryo, but rather serve as a source of nutrients or signals to the
developing embryo, such that its recycling may be required to gain missing and/or rate-limiting
substances. Indeed, during starvation or stress conditions, autophagy is induced in order to enhance
recycling of organelles and cellular contents to provide nutrients for cell survival. It is interesting that
in Drosophila a positive correlation was found between the male sperm size and its chances for
mating. Drosophila females evolved reproductive tracts that selectively bias paternity in favor of
males with longer sperm (Miller and Pitnick, 2002). Collectively, we propose the intriguing
hypotheses that either other detrimental factors or, alternatively, some beneficial factors may also
exist, which may provide evolutionary pressure for eggs to clear and recycle the paternal vacuolar
(mtDNA-less) mitochondria.
In conclusion, our studies have uncovered an autophagic/endocytic pathway, which might have
evolved in organisms with flagellated sperm to efficiently eliminate paternal mitochondria.
Understanding the mechanisms that govern PMD is not only of academic interest, as sophisticated
human in vitro fertilization (IVF) technologies are being widely used in the clinic and the risk
attributed to paternal factors has been the subject of ongoing debate (Verpoest and Tournaye, 2006).
An understanding of the paternally and maternally derived pathways and factors that mediate PMD
may be critical to determining possible risks associated with new IVF and cloning technologies.

P a g e | 82
Reference List
Abdu,U., Bar,D., and Schupbach,T. (2006). spn-F encodes a novel protein that affects oocyte
patterning and bristle morphology in Drosophila. Development 133, 1477-1484.
Al Rawi S., Louvet-Vallee,S., Djeddi,A., Sachse,M., Culetto,E., Hajjar,C., Boyd,L., Legouis,R., and
Galy,V. (2011). Postfertilization Autophagy of Sperm Organelles Prevents Paternal Mitochondrial
DNA Transmission. Science 334, 1144-1147.
Al-Dossary,A.A., Bathala,P., Caplan,J.L., and Martin-DeLeon,P.A. (2015). Oviductosome-Sperm
Membrane Interaction in Cargo Delivery: DETECTION OF FUSION AND UNDERLYING
MOLECULAR PLAYERS USING THREE-DIMENSIONAL SUPER-RESOLUTION
STRUCTURED ILLUMINATION MICROSCOPY (SR-SIM). J. Biol. Chem. 290, 17710-17723.
Aldridge,A.C., Benson,L.P., Siegenthaler,M.M., Whigham,B.T., Stowers,R.S., and Hales,K.G.
(2007). Roles for Drp1, a dynamin-related protein, and milton, a kinesin-associated protein, in
mitochondrial segregation, unfurling and elongation during Drosophila spermatogenesis. Fly. (Austin.
) 1, 38-46.
Andersson,S.G., Zomorodipour,A., Andersson,J.O., Sicheritz-Ponten,T., Alsmark,U.C.,
Podowski,R.M., Naslund,A.K., Eriksson,A.S., Winkler,H.H., and Kurland,C.G. (1998). The genome
sequence of Rickettsia prowazekii and the origin of mitochondria. Nature 396, 133-140.
Ankel-Simons,F. and Cummins,J.M. (1996). Misconceptions about mitochondria and mammalian
fertilization: implications for theories on human evolution. Proc. Natl. Acad. Sci U. S. A 93, 13859-
13863.
Aram,L., Braun,T., Braverman,C., Kaplan,Y., Ravid,L., Levin-Zaidman,S., and Arama,E. (2016). A
Krebs Cycle Component Limits Caspase Activation Rate through Mitochondrial Surface Restriction
of CRL Activation. Dev. Cell 37, 15-33.
Arama,E., Agapite,J., and Steller,H. (2003). Caspase activity and a specific cytochrome C are required
for sperm differentiation in Drosophila. Dev. Cell 4, 687-697.
Arama,E., Bader,M., Rieckhof,G.E., and Steller,H. (2007). A Ubiquitin Ligase Complex Regulates
Caspase Activation During Sperm Differentiation in Drosophila. PLoS Biol 5, e251.
Arama,E., Bader,M., Srivastava,M., Bergmann,A., and Steller,H. (2006). The two Drosophila
cytochrome C proteins can function in both respiration and caspase activation. EMBO J. 25, 232-243.
Arndt,V., Dick,N., Tawo,R., Dreiseidler,M., Wenzel,D., Hesse,M., Furst,D.O., Saftig,P., Saint,R.,
Fleischmann,B.K., Hoch,M., and Hohfeld,J. (2010). Chaperone-assisted selective autophagy is
essential for muscle maintenance. Curr. Biol. 20, 143-148.
Artal-Sanz,M. and Tavernarakis,N. (2009). Prohibitin and mitochondrial biology. Trends Endocrinol.
Metab 20, 394-401.
Ashburner,M. (1989). Drosophila: A laboratory manual. (New York: Cold Spring Harbor Laboratory
Press).

P a g e | 83
Ashrafi,G. and Schwarz,T.L. (2013). The pathways of mitophagy for quality control and clearance of
mitochondria. Cell Death. Differ. 20, 31-42.
Bader,M., Arama,E., and Steller,H. (2010). A novel F-box protein is required for caspase activation
during cellular remodeling in Drosophila. Development 137, 1679-1688.
Bader,M., Benjamin,S., Wapinski,O.L., Smith,D.M., Goldberg,A.L., and Steller,H. (2011). A
conserved F box regulatory complex controls proteasome activity in Drosophila. Cell 145, 371-382.
Baker,R.T., Tobias,J.W., and Varshavsky,A. (1992). Ubiquitin-specific proteases of Saccharomyces
cerevisiae. Cloning of UBP2 and UBP3, and functional analysis of the UBP gene family. J. Biol.
Chem. 267, 23364-23375.
Bassett,A.R., Tibbit,C., Ponting,C.P., and Liu,J.L. (2013). Highly efficient targeted mutagenesis of
Drosophila with the CRISPR/Cas9 system. Cell Rep. 4, 220-228.
Bazinet,C. and Rollins,J.E. (2003). Rickettsia-like mitochondrial motility in Drosophila
spermiogenesis. Evol. Dev. 5, 379-385.
Berg,T.O., Fengsrud,M., Stromhaug,P.E., Berg,T., and Seglen,P.O. (1998). Isolation and
characterization of rat liver amphisomes. Evidence for fusion of autophagosomes with both early and
late endosomes. J. Biol. Chem. 273, 21883-21892.
Birky,C.W., Jr. (2001). The inheritance of genes in mitochondria and chloroplasts: laws, mechanisms,
and models. Annu. Rev. Genet. 35, 125-148.
Bitan,A., Guild,G.M., Bar-Dubin,D., and Abdu,U. (2010). Asymmetric microtubule function is an
essential requirement for polarized organization of the Drosophila bristle. Mol. Cell Biol. 30, 496-507.
Bjorkoy,G., Lamark,T., Brech,A., Outzen,H., Perander,M., Overvatn,A., Stenmark,H., and
Johansen,T. (2005). p62/SQSTM1 forms protein aggregates degraded by autophagy and has a
protective effect on huntingtin-induced cell death. J. Cell Biol. 171, 603-614.
Blumer,N., Schreiter,K., Hempel,L., Santel,A., Hollmann,M., Schafer,M.A., and Renkawitz-Pohl,R.
(2002). A new translational repression element and unusual transcriptional control regulate expression
of don juan during Drosophila spermatogenesis. Mech. Dev. 110, 97-112.
Braschi,E., Zunino,R., and McBride,H.M. (2009). MAPL is a new mitochondrial SUMO E3 ligase
that regulates mitochondrial fission. EMBO Rep. 10, 748-754.
Bre,M.H., Redeker,V., Quibell,M., rmanaden-Delorme,J., Bressac,C., Cosson,J., Huitorel,P.,
Schmitter,J.M., Rossler,J., Johnson,T., Adoutte,A., and Levilliers,N. (1996). Axonemal tubulin
polyglycylation probed with two monoclonal antibodies: widespread evolutionary distribution,
appearance during spermatozoan maturation and possible function in motility. J. Cell Sci. 109 ( Pt 4),
727-738.
Bre,M.H., Redeker,V., Vinh,J., Rossier,J., and Levilliers,N. (1998). Tubulin polyglycylation:
differential posttranslational modification of dynamic cytoplasmic and stable axonemal microtubules
in paramecium. Mol. Biol. Cell 9, 2655-2665.

P a g e | 84
Budnik,V., Ruiz-Canada,C., and Wendler,F. (2016). Extracellular vesicles round off communication
in the nervous system. Nat. Rev. Neurosci. 17, 160-172.
Buzas,E.I., Gyorgy,B., Nagy,G., Falus,A., and Gay,S. (2014). Emerging role of extracellular vesicles
in inflammatory diseases. Nat. Rev. Rheumatol. 10, 356-364.
Cai,Z.L., Xu,J., Xue,S.R., Liu,Y.Y., Zhang,Y.J., Zhang,X.Z., Wang,X., Wu,F.P., and Li,X.M. (2015).
The E3 ubiquitin ligase seven in absentia homolog 1 may be a potential new therapeutic target for
Parkinson's disease. Neural Regen. Res. 10, 1286-1291.
Carre-Mlouka,A., Gaumer,S., Gay,P., Petitjean,A.M., Coulondre,C., Dru,P., Bras,F., Dezelee,S., and
Contamine,D. (2007). Control of sigma virus multiplication by the ref(2)P gene of Drosophila
melanogaster: an in vivo study of the PB1 domain of Ref(2)P. Genetics 176, 409-419.
Celardo,I., Martins,L.M., and Gandhi,S. (2014). Unravelling mitochondrial pathways to Parkinson's
disease. Br. J. Pharmacol. 171, 1943-1957.
Chang,T.K., Shravage,B.V., Hayes,S.D., Powers,C.M., Simin,R.T., Wade,H.J., and Baehrecke,E.H.
(2013). Uba1 functions in Atg7- and Atg3-independent autophagy. Nat. Cell Biol. 15, 1067-1078.
Clark,I.E., Dodson,M.W., Jiang,C., Cao,J.H., Huh,J.R., Seol,J.H., Yoo,S.J., Hay,B.A., and Guo,M.
(2006). Drosophila pink1 is required for mitochondrial function and interacts genetically with parkin.
Nature.
Corrigan,L., Redhai,S., Leiblich,A., Fan,S.J., Perera,S.M., Patel,R., Gandy,C., Wainwright,S.M.,
Morris,J.F., Hamdy,F., Goberdhan,D.C., and Wilson,C. (2014). BMP-regulated exosomes from
Drosophila male reproductive glands reprogram female behavior. J. Cell Biol. 206, 671-688.
Cummins,J.M. (2000). Fertilization and elimination of the paternal mitochondrial genome. Hum.
Reprod. 15 Suppl 2, 92-101.
de Castro,I., Martins,L.M., and Tufi,R. (2010). Mitochondrial quality control and neurological
disease: an emerging connection. Expert. Rev. Mol. Med. 12, e12.
DeLuca,S.Z. and O'Farrell,P.H. (2012). Barriers to male transmission of mitochondrial DNA in sperm
development. Dev. Cell 22, 660-668.
Deng,H., Dodson,M.W., Huang,H., and Guo,M. (2008). The Parkinson's disease genes pink1 and
parkin promote mitochondrial fission and/or inhibit fusion in Drosophila. Proc. Natl. Acad. Sci. U. S.
A 105, 14503-14508.
DiAntonio,A., Haghighi,A.P., Portman,S.L., Lee,J.D., Amaranto,A.M., and Goodman,C.S. (2001).
Ubiquitination-dependent mechanisms regulate synaptic growth and function. Nature 412, 449-452.
Dubin-Bar,D., Bitan,A., Bakhrat,A., Kaiden-Hasson,R., Etzion,S., Shaanan,B., and Abdu,U. (2008).
The Drosophila IKK-related kinase (Ik2) and Spindle-F proteins are part of a complex that regulates
cytoskeleton organization during oogenesis. BMC. Cell Biol. 9, 51.
Dupont,N., Lacas-Gervais,S., Bertout,J., Paz,I., Freche,B., Van Nhieu,G.T., van der Goot,F.G.,
Sansonetti,P.J., and Lafont,F. (2009). Shigella phagocytic vacuolar membrane remnants participate in

P a g e | 85
the cellular response to pathogen invasion and are regulated by autophagy. Cell Host. Microbe 6, 137-
149.
Fader,C.M., Sanchez,D., Furlan,M., and Colombo,M.I. (2008). Induction of autophagy promotes
fusion of multivesicular bodies with autophagic vacuoles in k562 cells. Traffic 230-250.
Farre,J.C., Krick,R., Subramani,S., and Thumm,M. (2009). Turnover of organelles by autophagy in
yeast. Curr. Opin. Cell Biol. 21, 522-530.
Filimonenko,M., Stuffers,S., Raiborg,C., Yamamoto,A., Malerod,L., Fisher,E.M., Isaacs,A., Brech,A.,
Stenmark,H., and Simonsen,A. (2007). Functional multivesicular bodies are required for autophagic
clearance of protein aggregates associated with neurodegenerative disease. J. Cell Biol. 179, 485-500.
Filler,K., Lyon,D., Bennett,J., McCain,N., Elswick,R., Lukkahatai,N., and Saligan,L.N. (2014).
Association of Mitochondrial Dysfunction and Fatigue: A Review of the Literature. BBA. Clin. 1, 12-
23.
Florey,O., Kim,S.E., Sandoval,C.P., Haynes,C.M., and Overholtzer,M. (2011). Autophagy machinery
mediates macroendocytic processing and entotic cell death by targeting single membranes. Nat. Cell
Biol. 13, 1335-1343.
Ford,W.C. (2004). Regulation of sperm function by reactive oxygen species. Hum. Reprod. Update.
10, 387-399.
Franco,M., Seyfried,N.T., Brand,A.H., Peng,J., and Mayor,U. (2011). A novel strategy to isolate
ubiquitin conjugates reveals wide role for ubiquitination during neural development. Mol. Cell
Proteomics. 10, M110.
Franco-Iborra,S., Vila,M., and Perier,C. (2016). The Parkinson Disease Mitochondrial Hypothesis:
Where Are We at? Neuroscientist. 22, 266-277.
Fuller,M.T. (1993). Spermatogenesis in Drosophila. In The Development of Drosophila
melanogaster, M.Bate and A.M.Arias, eds. Cold Spring Harbor Laboratory Press), pp. 71-147.
Geisler,S., Holmstrom,K.M., Skujat,D., Fiesel,F.C., Rothfuss,O.C., Kahle,P.J., and Springer,W.
(2010). PINK1/Parkin-mediated mitophagy is dependent on VDAC1 and p62/SQSTM1. Nat. Cell
Biol 12, 119-131.
Gilbert,D.G. and Richmond,R.C. (1981). Studies of esterase 6 in Drosophila melanogaster. VI.
ejaculate competitive abilities of males having null or active alleles. Genetics 97, 85-94.
Greene,J.C., Whitworth,A.J., Andrews,L.A., Parker,T.J., and Pallanck,L.J. (2005). Genetic and
genomic studies of Drosophila parkin mutants implicate oxidative stress and innate immune responses
in pathogenesis. Hum. Mol. Genet. 14, 799-811.
Greene,J.C., Whitworth,A.J., Kuo,I., Andrews,L.A., Feany,M.B., and Pallanck,L.J. (2003).
Mitochondrial pathology and apoptotic muscle degeneration in Drosophila parkin mutants. Proc. Natl.
Acad. Sci. U. S. A 100, 4078-4083.
Gross,J.C., Chaudhary,V., Bartscherer,K., and Boutros,M. (2012). Active Wnt proteins are secreted
on exosomes. Nat. Cell Biol. 14, 1036-1045.

P a g e | 86
Gyllensten,U., Wharton,D., Josefsson,A., and Wilson,A.C. (1991). Paternal inheritance of
mitochondrial DNA in mice. Nature 352, 255-257.
Hales,K.G. and Fuller,M.T. (1997). Developmentally regulated mitochondrial fusion mediated by a
conserved, novel, predicted GTPase. Cell 90, 121-129.
He,C. and Klionsky,D.J. (2009). Regulation mechanisms and signaling pathways of autophagy. Annu.
Rev. Genet. 43, 67-93.
Heymann,J.A., Hayles,M., Gestmann,I., Giannuzzi,L.A., Lich,B., and Subramaniam,S. (2006). Site-
specific 3D imaging of cells and tissues with a dual beam microscope. J. Struct. Biol. 155, 63-73.
Hiraoka,J. and Hirao,Y. (1988). Fate of sperm tail components after incorporation into the hamster
egg. Gamete Res. 19, 369-380.
Hoshino,A., Costa-Silva,B., Shen,T.L., Rodrigues,G., Hashimoto,A., Tesic,M.M., Molina,H.,
Kohsaka,S., Di,G.A., Ceder,S., Singh,S., Williams,C., Soplop,N., Uryu,K., Pharmer,L., King,T.,
Bojmar,L., Davies,A.E., Ararso,Y., Zhang,T., Zhang,H., Hernandez,J., Weiss,J.M., Dumont-
Cole,V.D., Kramer,K., Wexler,L.H., Narendran,A., Schwartz,G.K., Healey,J.H., Sandstrom,P.,
Labori,K.J., Kure,E.H., Grandgenett,P.M., Hollingsworth,M.A., de,S.M., Kaur,S., Jain,M., Mallya,K.,
Batra,S.K., Jarnagin,W.R., Brady,M.S., Fodstad,O., Muller,V., Pantel,K., Minn,A.J., Bissell,M.J.,
Garcia,B.A., Kang,Y., Rajasekhar,V.K., Ghajar,C.M., Matei,I., Peinado,H., Bromberg,J., and
Lyden,D. (2015). Tumour exosome integrins determine organotropic metastasis. Nature 527, 329-335.
Johansen,T. and Lamark,T. (2011). Selective autophagy mediated by autophagic adapter proteins.
Autophagy. 7, 279-296.
Juhasz,G., Erdi,B., Sass,M., and Neufeld,T.P. (2007). Atg7-dependent autophagy promotes neuronal
health, stress tolerance, and longevity but is dispensable for metamorphosis in Drosophila. Genes Dev.
21, 3061-3066.
Kaplan,Y., Gibbs-Bar,L., Kalifa,Y., Feinstein-Rotkopf,Y., and Arama,E. (2010). Gradients of a
Ubiquitin E3 Ligase Inhibitor and a Caspase Inhibitor Determine Differentiation or Death in
Spermatids. Dev. Cell 19, 160-173.
Karr,T.L. (1991). Intracellular sperm/egg interactions in Drosophila: a three-dimensional structural
analysis of a paternal product in the developing egg. Mech. Dev. 34, 101-111.
Kirkin,V., Lamark,T., Sou,Y.S., Bjorkoy,G., Nunn,J.L., Bruun,J.A., Shvets,E., McEwan,D.G.,
Clausen,T.H., Wild,P., Bilusic,I., Theurillat,J.P., Overvatn,A., Ishii,T., Elazar,Z., Komatsu,M.,
Dikic,I., and Johansen,T. (2009a). A role for NBR1 in autophagosomal degradation of ubiquitinated
substrates. Mol. Cell 33, 505-516.
Kirkin,V., McEwan,D.G., Novak,I., and Dikic,I. (2009b). A role for ubiquitin in selective autophagy.
Mol. Cell 34, 259-269.
Knaevelsrud,H. and Simonsen,A. (2010). Fighting disease by selective autophagy of aggregate-prone
proteins. FEBS Lett. 584, 2635-2645.
Kondo,S. and Ueda,R. (2013). Highly improved gene targeting by germline-specific Cas9 expression
in Drosophila. Genetics 195, 715-721.

P a g e | 87
Koyano,F., Okatsu,K., Kosako,H., Tamura,Y., Go,E., Kimura,M., Kimura,Y., Tsuchiya,H.,
Yoshihara,H., Hirokawa,T., Endo,T., Fon,E.A., Trempe,J.F., Saeki,Y., Tanaka,K., and Matsuda,N.
(2014). Ubiquitin is phosphorylated by PINK1 to activate parkin. Nature 510, 162-166.
Kraytsberg,Y., Schwartz,M., Brown,T.A., Ebralidse,K., Kunz,W.S., Clayton,D.A., Vissing,J., and
Khrapko,K. (2004). Recombination of human mitochondrial DNA. Science 304, 981.
Lamb,C.A., Dooley,H.C., and Tooze,S.A. (2013a). Endocytosis and autophagy: Shared machinery for
degradation. Bioessays 35, 34-45.
Lamb,C.A., Yoshimori,T., and Tooze,S.A. (2013b). The autophagosome: origins unknown,
biogenesis complex. Nat. Rev. Mol. Cell Biol. 14, 759-774.
Levine,B. and Deretic,V. (2007). Unveiling the roles of autophagy in innate and adaptive immunity.
Nat. Rev. Immunol. 7, 767-777.
Levine,B. and Elazar,Z. (2011). Development. Inheriting maternal mtDNA. Science 334, 1069-1070.
Lim,K.L., Chew,K.C., Tan,J.M., Wang,C., Chung,K.K., Zhang,Y., Tanaka,Y., Smith,W.,
Engelender,S., Ross,C.A., Dawson,V.L., and Dawson,T.M. (2005). Parkin mediates nonclassical,
proteasomal-independent ubiquitination of synphilin-1: implications for Lewy body formation. J.
Neurosci. 25, 2002-2009.
Lindsley,D. and Tokuyasu,K.T. (1980). Spermatogenesis. In Genetics and Biology of Drosophila,
M.Ashburner and T.R.Wright, eds. (New York: Academic Press), pp. 225-294.
Lloyd,T.E., Atkinson,R., Wu,M.N., Zhou,Y., Pennetta,G., and Bellen,H.J. (2002). Hrs regulates
endosome membrane invagination and tyrosine kinase receptor signaling in Drosophila. Cell 108,
261-269.
Luo,S.M., Ge,Z.J., Wang,Z.W., Jiang,Z.Z., Wang,Z.B., Ouyang,Y.C., Hou,Y., Schatten,H., and
Sun,Q.Y. (2013). Unique insights into maternal mitochondrial inheritance in mice. Proc. Natl. Acad.
Sci. U. S. A 110, 13038-13043.
Manil-Segalen,M., Lefebvre,C., Culetto,E., and Legouis,R. (2012). Need an ESCRT for
autophagosomal maturation? Commun. Integr. Biol. 5, 566-571.
Manjithaya,R., Nazarko,T.Y., Farre,J.C., and Subramani,S. (2010). Molecular mechanism and
physiological role of pexophagy. FEBS Lett. 584, 1367-1373.
McPhee,C.K. and Baehrecke,E.H. (2009). Autophagy in Drosophila melanogaster. Biochim. Biophys.
Acta 1793, 1452-1460.
Melo,S.A., Sugimoto,H., O'Connell,J.T., Kato,N., Villanueva,A., Vidal,A., Qiu,L., Vitkin,E.,
Perelman,L.T., Melo,C.A., Lucci,A., Ivan,C., Calin,G.A., and Kalluri,R. (2014). Cancer exosomes
perform cell-independent microRNA biogenesis and promote tumorigenesis. Cancer Cell 26, 707-721.
Miller,G.T. and Pitnick,S. (2002). Sperm-female coevolution in Drosophila. Science 298, 1230-1233.
Mizushima,N. and Komatsu,M. (2011). Autophagy: renovation of cells and tissues. Cell 147, 728-
741.

P a g e | 88
Montoya,J., Lopez-Gallardo,E., ez-Sanchez,C., Lopez-Perez,M.J., and Ruiz-Pesini,E. (2009). 20 years
of human mtDNA pathologic point mutations: carefully reading the pathogenicity criteria. Biochim.
Biophys. Acta 1787, 476-483.
Nakatogawa,H., Suzuki,K., Kamada,Y., and Ohsumi,Y. (2009). Dynamics and diversity in autophagy
mechanisms: lessons from yeast. Nat. Rev. Mol. Cell Biol 10, 458-467.
Narendra,D., Kane,L.A., Hauser,D.N., Fearnley,I.M., and Youle,R.J. (2010a). p62/SQSTM1 is
required for Parkin-induced mitochondrial clustering but not mitophagy; VDAC1 is dispensable for
both. Autophagy. 6, 1090-1106.
Narendra,D., Tanaka,A., Suen,D.F., and Youle,R.J. (2008). Parkin is recruited selectively to impaired
mitochondria and promotes their autophagy. J Cell Biol. 183, 795-803.
Narendra,D.P., Jin,S.M., Tanaka,A., Suen,D.F., Gautier,C.A., Shen,J., Cookson,M.R., and Youle,R.J.
(2010b). PINK1 is selectively stabilized on impaired mitochondria to activate Parkin. PLoS. Biol 8,
e1000298.
Neufeld,T.P. and Baehrecke,E.H. (2008). Eating on the fly: function and regulation of autophagy
during cell growth, survival and death in Drosophila. Autophagy. 4, 557-562.
Newton,K., Matsumoto,M.L., Wertz,I.E., Kirkpatrick,D.S., Lill,J.R., Tan,J., Dugger,D., Gordon,N.,
Sidhu,S.S., Fellouse,F.A., Komuves,L., French,D.M., Ferrando,R.E., Lam,C., Compaan,D., Yu,C.,
Bosanac,I., Hymowitz,S.G., Kelley,R.F., and Dixit,V.M. (2008). Ubiquitin chain editing revealed by
polyubiquitin linkage-specific antibodies. Cell 134, 668-678.
Nezis,I.P., Shravage,B.V., Sagona,A.P., Lamark,T., Bjorkoy,G., Johansen,T., Rusten,T.E., Brech,A.,
Baehrecke,E.H., and Stenmark,H. (2010). Autophagic degradation of dBruce controls DNA
fragmentation in nurse cells during late Drosophila melanogaster oogenesis. J Cell Biol 190, 523-531.
Nezis,I.P., Simonsen,A., Sagona,A.P., Finley,K., Gaumer,S., Contamine,D., Rusten,T.E.,
Stenmark,H., and Brech,A. (2008). Ref(2)P, the Drosophila melanogaster homologue of mammalian
p62, is required for the formation of protein aggregates in adult brain. J. Cell Biol. 180, 1065-1071.
Nishimura,Y., Yoshinari,T., Naruse,K., Yamada,T., Sumi,K., Mitani,H., Higashiyama,T., and
Kuroiwa,T. (2006). Active digestion of sperm mitochondrial DNA in single living sperm revealed by
optical tweezers. Proc. Natl. Acad. Sci U. S. A 103, 1382-1387.
Okatsu,K., Koyano,F., Kimura,M., Kosako,H., Saeki,Y., Tanaka,K., and Matsuda,N. (2015).
Phosphorylated ubiquitin chain is the genuine Parkin receptor. J. Cell Biol. 209, 111-128.
Olzmann,J.A., Li,L., Chudaev,M.V., Chen,J., Perez,F.A., Palmiter,R.D., and Chin,L.S. (2007).
Parkin-mediated K63-linked polyubiquitination targets misfolded DJ-1 to aggresomes via binding to
HDAC6. J Cell Biol 178, 1025-1038.
Onoue,K., Jofuku,A., Ban-Ishihara,R., Ishihara,T., Maeda,M., Koshiba,T., Itoh,T., Fukuda,M.,
Otera,H., Oka,T., Takano,H., Mizushima,N., Mihara,K., and Ishihara,N. (2013). Fis1 acts as a
mitochondrial recruitment factor for TBC1D15 that is involved in regulation of mitochondrial
morphology. J. Cell Sci. 126, 176-185.

P a g e | 89
Panakova,D., Sprong,H., Marois,E., Thiele,C., and Eaton,S. (2005). Lipoprotein particles are required
for Hedgehog and Wingless signalling. Nature 435, 58-65.
Pankiv,S., Clausen,T.H., Lamark,T., Brech,A., Bruun,J.A., Outzen,H., Overvatn,A., Bjorkoy,G., and
Johansen,T. (2007). p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of
ubiquitinated protein aggregates by autophagy. J Biol Chem. 282, 24131-24145.
Park,J., Lee,S.B., Lee,S., Kim,Y., Song,S., Kim,S., Bae,E., Kim,J., Shong,M., Kim,J.M., and Chung,J.
(2006). Mitochondrial dysfunction in Drosophila PINK1 mutants is complemented by parkin. Nature.
Pfeffer,S.R. (2013). Rab GTPase regulation of membrane identity. Curr. Opin. Cell Biol. 25, 414-419.
Pimenta,d.C., I, Costa,A.C., Lam,D., Tufi,R., Fedele,V., Moisoi,N., Dinsdale,D., Deas,E., Loh,S.H.,
and Martins,L.M. (2012). Genetic analysis of mitochondrial protein misfolding in Drosophila
melanogaster. Cell Death. Differ.
Piper,R.C. and Katzmann,D.J. (2007). Biogenesis and function of multivesicular bodies. Annu. Rev.
Cell Dev. Biol 23, 519-547.
Pitnick,S. and Karr,T.L. (1998). Paternal products and by-products in Drosophila development. Proc.
Biol. Sci. 265, 821-826.
Politi,Y., Gal,L., Kalifa,Y., Ravid,L., Elazar,Z., and Arama,E. (2014). Paternal mitochondrial
destruction after fertilization is mediated by a common endocytic and autophagic pathway in
Drosophila. Dev. Cell 29, 305-320.
Rabinowitz,J.D. and White,E. (2010). Autophagy and metabolism. Science 330, 1344-1348.
Rand,M.D. (2014). A method of permeabilization of Drosophila embryos for assays of small molecule
activity. J. Vis. Exp.
Rand,M.D., Kearney,A.L., Dao,J., and Clason,T. (2010). Permeabilization of Drosophila embryos for
introduction of small molecules. Insect Biochem. Mol. Biol. 40, 792-804.
Randow,F. (2011). How cells deploy ubiquitin and autophagy to defend their cytosol from bacterial
invasion. Autophagy. 7, 304-309.
Regev-Rudzki,N., Wilson,D.W., Carvalho,T.G., Sisquella,X., Coleman,B.M., Rug,M., Bursac,D.,
Angrisano,F., Gee,M., Hill,A.F., Baum,J., and Cowman,A.F. (2013). Cell-cell communication
between malaria-infected red blood cells via exosome-like vesicles. Cell 153, 1120-1133.
Rojansky,R., Cha,M.Y., and Chan,D.C. (2016). Elimination of paternal mitochondria in mouse
embryos occurs through autophagic degradation dependent on PARKIN and MUL1. Elife. 5.
Rothwell,W.F. and Sullivan,W. (2007). Fixation of Drosophila embryos. CSH. Protoc. 2007, db.
Rubinsztein,D.C., Shpilka,T., and Elazar,Z. (2012). Mechanisms of autophagosome biogenesis. Curr.
Biol. 22, R29-R34.

P a g e | 90
Rusten,T.E., Vaccari,T., Lindmo,K., Rodahl,L.M., Nezis,I.P., Sem-Jacobsen,C., Wendler,F.,
Vincent,J.P., Brech,A., Bilder,D., and Stenmark,H. (2007). ESCRTs and Fab1 regulate distinct steps
of autophagy. Curr. Biol. 1817-1825.
Sahu,R., Kaushik,S., Clement,C.C., Cannizzo,E.S., Scharf,B., Follenzi,A., Potolicchio,I., Nieves,E.,
Cuervo,A.M., and Santambrogio,L. (2011). Microautophagy of cytosolic proteins by late endosomes.
Dev. Cell 20, 131-139.
Sanjuan,M.A., Dillon,C.P., Tait,S.W., Moshiach,S., Dorsey,F., Connell,S., Komatsu,M., Tanaka,K.,
Cleveland,J.L., Withoff,S., and Green,D.R. (2007). Toll-like receptor signalling in macrophages links
the autophagy pathway to phagocytosis. Nature 450, 1253-1257.
Santel,A., Blumer,N., Kampfer,M., and Renkawitz-Pohl,R. (1998). Flagellar mitochondrial
association of the male-specific Don Juan protein in Drosophila spermatozoa. J Cell Sci. 111, 3299-
3309.
Sato,M. and Sato,K. (2011). Degradation of Paternal Mitochondria by Fertilization-Triggered
Autophagy in C. elegans Embryos. Science 334, 1141-1144.
Schafer,A. and Reichert,A.S. (2009). Emerging roles of mitochondrial membrane dynamics in health
and disease. Biol Chem. 390, 707-715.
Schatz,D., Shemi,A., Rosenwasser,S., Sabanay,H., Wolf,S.G., Ben-Dor,S., and Vardi,A. (2014).
Hijacking of an autophagy-like process is critical for the life cycle of a DNA virus infecting oceanic
algal blooms. New Phytol. 204, 854-863.
Schindelin,J., Arganda-Carreras,I., Frise,E., Kaynig,V., Longair,M., Pietzsch,T., Preibisch,S.,
Rueden,C., Saalfeld,S., Schmid,B., Tinevez,J.Y., White,D.J., Hartenstein,V., Eliceiri,K.,
Tomancak,P., and Cardona,A. (2012). Fiji: an open-source platform for biological-image analysis.
Nat. Methods 9, 676-682.
Schmid,D. and Munz,C. (2007). Innate and adaptive immunity through autophagy. Immunity. 27, 11-
21.
Schmidt,O. and Teis,D. (2012). The ESCRT machinery. Curr. Biol. 22, R116-R120.
Schweers,R.L., Zhang,J., Randall,M.S., Loyd,M.R., Li,W., Dorsey,F.C., Kundu,M., Opferman,J.T.,
Cleveland,J.L., Miller,J.L., and Ney,P.A. (2007). NIX is required for programmed mitochondrial
clearance during reticulocyte maturation. Proc. Natl. Acad. Sci. U. S. A 104, 19500-19505.
Sehgal,P.B. and Lee,J.E. (2011). Protein trafficking dysfunctions: Role in the pathogenesis of
pulmonary arterial hypertension. Pulm. Circ. 1, 17-32.
Shaner,N.C., Steinbach,P.A., and Tsien,R.Y. (2005). A guide to choosing fluorescent proteins. Nat.
Methods 2, 905-909.
Sharpley,M.S., Marciniak,C., Eckel-Mahan,K., McManus,M., Crimi,M., Waymire,K., Lin,C.S.,
Masubuchi,S., Friend,N., Koike,M., Chalkia,D., MacGregor,G., Sassone-Corsi,P., and Wallace,D.C.
(2012). Heteroplasmy of mouse mtDNA is genetically unstable and results in altered behavior and
cognition. Cell 151, 333-343.

P a g e | 91
Shirinian,M., Kambris,Z., Hamadeh,L., Grabbe,C., Journo,C., Mahieux,R., and Bazarbachi,A. (2015).
A Transgenic Drosophila melanogaster Model To Study Human T-Lymphotropic Virus Oncoprotein
Tax-1-Driven Transformation In Vivo. J. Virol. 89, 8092-8095.
Simonsen,A., Cumming,R.C., Brech,A., Isakson,P., Schubert,D.R., and Finley,K.D. (2008).
Promoting basal levels of autophagy in the nervous system enhances longevity and oxidant resistance
in adult Drosophila. Autophagy. 4, 176-184.
Slone,J., Daniels,J., and Amrein,H. (2007). Sugar receptors in Drosophila. Curr. Biol. 17, 1809-1816.
Song,W.H., Yi,Y.J., Sutovsky,M., Meyers,S., and Sutovsky,P. (2016). Autophagy and ubiquitin-
proteasome system contribute to sperm mitophagy after mammalian fertilization. Proc. Natl. Acad.
Sci. U. S. A 113, E5261-E5270.
Stathakis,D.G., Pentz,E.S., Freeman,M.E., Kullman,J., Hankins,G.R., Pearlson,N.J., and Wright,T.R.
(1995). The genetic and molecular organization of the Dopa decarboxylase gene cluster of Drosophila
melanogaster. Genetics 141, 629-655.
Stromhaug,P.E. and Seglen,P.O. (1993). Evidence for acidity of prelysosomal autophagic/endocytic
vacuoles (amphisomes). Biochem. J. 291 ( Pt 1), 115-121.
Sumpter,R., Jr. and Levine,B. (2011). Selective autophagy and viruses. Autophagy. 7, 260-265.
Sury,M.D., Chen,J.X., and Selbach,M. (2010). The SILAC fly allows for accurate protein
quantification in vivo. Mol. Cell Proteomics. 9, 2173-2183.
Sutovsky,P., McCauley,T.C., Sutovsky,M., and Day,B.N. (2003). Early degradation of paternal
mitochondria in domestic pig (Sus scrofa) is prevented by selective proteasomal inhibitors lactacystin
and MG132. Biol Reprod. 68, 1793-1800.
Sutovsky,P., Moreno,R.D., Ramalho-Santos,J., Dominko,T., Simerly,C., and Schatten,G. (1999).
Ubiquitin tag for sperm mitochondria. Nature 402, 371-372.
Sutovsky,P., Moreno,R.D., Ramalho-Santos,J., Dominko,T., Simerly,C., and Schatten,G. (2000).
Ubiquitinated sperm mitochondria, selective proteolysis, and the regulation of mitochondrial
inheritance in mammalian embryos. Biol. Reprod. 63, 582-590.
Sutovsky,P., van,L.K., McCauley,T., Day,B.N., and Sutovsky,M. (2004). Degradation of paternal
mitochondria after fertilization: implications for heteroplasmy, assisted reproductive technologies and
mtDNA inheritance. Reprod. Biomed. Online. 8, 24-33.
Takats,S., Nagy,P., Varga,A., Pircs,K., Karpati,M., Varga,K., Kovacs,A.L., Hegedus,K., and
Juhasz,G. (2013). Autophagosomal Syntaxin17-dependent lysosomal degradation maintains neuronal
function in Drosophila. J. Cell Biol. 201, 531-539.
Thompson,W.E., Ramalho-Santos,J., and Sutovsky,P. (2003). Ubiquitination of prohibitin in
mammalian sperm mitochondria: possible roles in the regulation of mitochondrial inheritance and
sperm quality control. Biol Reprod. 69, 254-260.
Tooze,S.A., Abada,A., and Elazar,Z. (2014). Endocytosis and autophagy: exploitation or cooperation?
Cold Spring Harb. Perspect. Biol. 6, a018358.

P a g e | 92
Twig,G., Hyde,B., and Shirihai,O.S. (2008). Mitochondrial fusion, fission and autophagy as a quality
control axis: the bioenergetic view. Biochim. Biophys. Acta 1777, 1092-1097.
Venken,K.J., He,Y., Hoskins,R.A., and Bellen,H.J. (2006). P[acman]: a BAC transgenic platform for
targeted insertion of large DNA fragments in D. melanogaster. Science 314, 1747-1751.
Wang,K. and Klionsky,D.J. (2011). Mitochondria removal by autophagy. Autophagy. 7, 297-300.
Weber,T.A. and Reichert,A.S. (2010). Impaired quality control of mitochondria: aging from a new
perspective. Exp Gerontol. 45, 503-511.
Wei,Y., Chiang,W.C., Sumpter,R., Jr., Mishra,P., and Levine,B. (2017). Prohibitin 2 Is an Inner
Mitochondrial Membrane Mitophagy Receptor. Cell 168, 224-238.
Weidberg,H., Shvets,E., and Elazar,Z. (2011). Biogenesis and cargo selectivity of autophagosomes.
Annu. Rev. Biochem. 80, 125-156.
Wild,P., Farhan,H., McEwan,D.G., Wagner,S., Rogov,V.V., Brady,N.R., Richter,B., Korac,J.,
Waidmann,O., Choudhary,C., Dotsch,V., Bumann,D., and Dikic,I. (2011). Phosphorylation of the
autophagy receptor optineurin restricts Salmonella growth. Science 333, 228-233.
Williams,R.L. and Urbe,S. (2007). The emerging shape of the ESCRT machinery. Nat. Rev. Mol. Cell
Biol. 8, 355-368.
Wyers,F., Petitjean,A.M., Dru,P., Gay,P., and Contamine,D. (1995). Localization of domains within
the Drosophila Ref(2)P protein involved in the intracellular control of sigma rhabdovirus
multiplication. J. Virol. 69, 4463-4470.
Xie,Z. and Klionsky,D.J. (2007). Autophagosome formation: core machinery and adaptations. Nat.
Cell Biol. 9, 1102-1109.
Yacobi-Sharon,K., Namdar,Y., and Arama,E. (2013). Alternative germ cell death pathway in
Drosophila involves HtrA2/Omi, lysosomes, and a caspase-9 counterpart. Dev. Cell 25, 29-42.
Yamano,K., Fogel,A.I., Wang,C., van der Bliek,A.M., and Youle,R.J. (2014). Mitochondrial Rab
GAPs govern autophagosome biogenesis during mitophagy. Elife. 3, e01612.
Yu,Z., O'Farrell,P.H., Yakubovich,N., and DeLuca,S.Z. (2017). The Mitochondrial DNA Polymerase
Promotes Elimination of Paternal Mitochondrial Genomes. Curr. Biol. 27, 1033-1039.
Yuana,Y., Sturk,A., and Nieuwland,R. (2013). Extracellular vesicles in physiological and pathological
conditions. Blood Rev. 27, 31-39.
Yun,J., Puri,R., Yang,H., Lizzio,M.A., Wu,C., Sheng,Z.H., and Guo,M. (2014). MUL1 acts in parallel
to the PINK1/parkin pathway in regulating mitofusin and compensates for loss of PINK1/parkin.
Elife. 3, e01958.
Zhang,J., Randall,M.S., Loyd,M.R., Dorsey,F.C., Kundu,M., Cleveland,J.L., and Ney,P.A. (2009).
Mitochondrial clearance is regulated by Atg7-dependent and -independent mechanisms during
reticulocyte maturation. Blood 114, 157-164.

P a g e | 93
Zhou,Q., Li,H., Li,H., Nakagawa,A., Lin,J.L., Lee,E.S., Harry,B.L., Skeen-Gaar,R.R., Suehiro,Y.,
William,D., Mitani,S., Yuan,H.S., Kang,B.H., and Xue,D. (2016). Mitochondrial endonuclease G
mediates breakdown of paternal mitochondria upon fertilization. Science.
Zhu,G., Wu,C.J., Zhao,Y., and Ashwell,J.D. (2007). Optineurin negatively regulates TNFalpha-
induced NF-kappaB activation by competing with NEMO for ubiquitinated RIP. Curr. Biol. 17, 1438-
1443.
Related Documents











