Short communications α-Glucosidase and α-amylase inhibitory constituents of Tinospora crispa: Isolation and chemical profile confirmation by ultra-high performance liquid chromatography-quadrupole time-of-flight/mass spectrometry H.A. Hamid, M.M. Yusoff *, M. Liu, M.R. Karim Faculty of Industrial Sciences & Technology, Universiti Malaysia Pahang, Lebuhraya Tun Razak, 26300 Gambang Kuantan, Pahang, Malaysia ARTICLE INFO Article history: Received 24 December 2014 Received in revised form 10 April 2015 Accepted 16 April 2015 Available online ABSTRACT Tinospora crispa has been used in Malaysia to treat diabetes mellitus. We evaluated the effects of compounds isolated from T. crispa vines on the starch digestive enzymes, α-glucosidase and α-amylase. Borapetoside C was the most potent α-glucosidase inhibitor (IC50 = 0.0527 ± 0.008 mg/ml), followed by 4-hydroxy benzaldehyde (IC50 = 0.557 ± 0.004 mg/ml), and the alkaloids lysicamine and liriodenine (IC50 = 0.562 ± 0.003 mg/ml). Borapetoside C also potently inhibited α-amylase inhibitory activity (IC50 = 0.775 ± 0.005 mg/ml). The rela- tionship between compound activity and content in the extracts was determined by ultra- high performance liquid chromatography-quadrupole time-of-flight/mass spectrometry (UPLC- QToF/MS).These results suggest that borapetoside C is the main primary active component of T. crispa extracts, mediating α-glucosidase and α-amylase inhibition. © 2015 Elsevier Ltd. All rights reserved. Keywords: Fine chemicals Tinospora crispa Ultra-performance liquid chromatography α-amylase α-glucosidase 1. Introduction Diabetes is a metabolic disorder characterized by high plasma glucose levels that can be classified as eitherType 1 orType 2 (Mitrakou et al., 1992; Porte, Jr., & Kahn, 2001). InType 1 or insulin- dependent diabetes, the pancreas fails to secrete insulin. In contrast,Type 2 also called non-insulin-dependent diabetes or diabetes mellitus (DM) is caused by an imbalance between blood sugar absorption and insulin secretion, leading to disability due * Corresponding author. Faculty of Industrial Sciences & Technology, Universiti Malaysia Pahang, Lebuhraya Tun Razak, 26300 Gambang Kuantan, Pahang, Malaysia. Tel.: +60 9 5492767; fax: +60 9 5492766. E-mail address: [email protected] (M.M. Yusoff). Chemical compounds: Borapetoside C (PubChem CID: 15934414); 4-Hydroxybenzaldehyde (PubChem CID: 126); β-Sitosterol (PubChem CID: 222284); Liriodenine (PubChem CID: 10144); Lysicamine (PubChem CID: 122691); Columbamine (PubChem CID: 72310); Magnoflorine (PubChem CID: 73337); N-trans-Feruloyltyramine (PubChem CID: 5280537). http://dx.doi.org/10.1016/j.jff.2015.04.011 1756-4646/© 2015 Elsevier Ltd. All rights reserved. journal of functional foods 16 (2015) 74–80 Available online at www.sciencedirect.com ScienceDirect journal homepage: www.elsevier.com/locate/jff

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Short communications
α-Glucosidase and α-amylase inhibitoryconstituents of Tinospora crispa: Isolation andchemical profile confirmation by ultra-highperformance liquid chromatography-quadrupoletime-of-flight/mass spectrometry
H.A. Hamid, M.M. Yusoff *, M. Liu, M.R. KarimFaculty of Industrial Sciences & Technology, Universiti Malaysia Pahang, Lebuhraya Tun Razak, 26300Gambang Kuantan, Pahang, Malaysia
A R T I C L E I N F O
Article history:
Received 24 December 2014
Received in revised form 10 April
2015
Accepted 16 April 2015
Available online
A B S T R A C T
Tinospora crispa has been used in Malaysia to treat diabetes mellitus. We evaluated the
effects of compounds isolated from T. crispa vines on the starch digestive enzymes,
α-glucosidase and α-amylase. Borapetoside C was the most potent α-glucosidase inhibitor
(IC50 = 0.0527 ± 0.008 mg/ml), followed by 4-hydroxy benzaldehyde (IC50 = 0.557 ± 0.004 mg/ml),
and the alkaloids lysicamine and liriodenine (IC50 = 0.562 ± 0.003 mg/ml). Borapetoside C
also potently inhibited α-amylase inhibitory activity (IC50 = 0.775 ± 0.005 mg/ml). The rela-
tionship between compound activity and content in the extracts was determined by ultra-
high performance liquid chromatography-quadrupole time-of-flight/mass spectrometry (UPLC-
QToF/MS). These results suggest that borapetoside C is the main primary active component
of T. crispa extracts, mediating α-glucosidase and α-amylase inhibition.
© 2015 Elsevier Ltd. All rights reserved.
Keywords:
Fine chemicals
Tinospora crispa
Ultra-performance liquid
chromatography
α-amylase
α-glucosidase
1. Introduction
Diabetes is a metabolic disorder characterized by high plasmaglucose levels that can be classified as either Type 1 or Type 2
(Mitrakou et al., 1992; Porte, Jr., & Kahn, 2001). InType 1 or insulin-dependent diabetes, the pancreas fails to secrete insulin. Incontrast,Type 2 also called non-insulin-dependent diabetes ordiabetes mellitus (DM) is caused by an imbalance between bloodsugar absorption and insulin secretion, leading to disability due
* Corresponding author. Faculty of Industrial Sciences & Technology, Universiti Malaysia Pahang, Lebuhraya Tun Razak, 26300 GambangKuantan, Pahang, Malaysia. Tel.: +60 9 5492767; fax: +60 9 5492766.
E-mail address: [email protected] (M.M. Yusoff).Chemical compounds: Borapetoside C (PubChem CID: 15934414); 4-Hydroxybenzaldehyde (PubChem CID: 126); β-Sitosterol (PubChem
CID: 222284); Liriodenine (PubChem CID: 10144); Lysicamine (PubChem CID: 122691); Columbamine (PubChem CID: 72310); Magnoflorine(PubChem CID: 73337); N-trans-Feruloyltyramine (PubChem CID: 5280537).http://dx.doi.org/10.1016/j.jff.2015.04.0111756-4646/© 2015 Elsevier Ltd. All rights reserved.
j o u rna l o f f un c t i ona l f o od s 1 6 ( 2 0 1 5 ) 7 4 – 8 0
Available online at www.sciencedirect.com
ScienceDirect
journal homepage: www.elsevier.com/ locate / j ff

to vascular and neurological complications (Porte, Jr., & Kahn,2001). Controlling plasma glucose level is essential to delay orprevent DM. One possible way to decrease the rate of bloodsugar absorption from the small intestine is to slow or inter-rupt the digestion of dietary starch, the major dietary sourceof glucose (Josse et al., 2003).
The inhibition of enzymes that digest dietary starch, suchas α-amylase and α-glucosidase has been studied as a methodto control blood sugar levels (Ali, Houghton, & Soumyanath,2006; Svensson, Kristoffersen, Oskarsson, & Bensch, 2004).α-Amylase catalyzes the hydrolysis of α-(1,4)-glucosidic link-ages to produce maltose and glucose, whereas α-glucosidasereleases glucose from maltose and/or sucrose (Sogaard-Andersen& Valentin-Hansen, 1993; Teeri, 1991). α-Glucosidase is amembrane-bound enzyme located at the epithelium of the smallintestine, which catalyzes the cleavage of glucose from disac-charides and oligosaccharides. Absorption of glucose into thebloodstream can be delayed by inhibiting these enzymes, thereby,ameliorating DM symptoms such as hyperglycemia.
There is an on-going interest in evaluating the effects ofplants and crude drugs on starch digestive enzymes as po-tential sources of novel oral hypoglycemic compounds anddietary supplements (Braithwaite et al., 2014; Sang, 2014). Moststudies have focused on α-glucosidase inhibition because thisenzyme regulates glucose release from maltose and/or sucrose.Although the inhibition of α-amylase also decreases glucoserelease, its complete inhibition is not desired, because it couldcause intestinal disorders due to the undigested starch beingutilized by gut micro flora for gas production (Cho, Han, & You,2011). However, partial inhibition of α-amylase may help modu-late the rate of glucose release from starch.The aqueous extractof Tinospora crispa is taken orally to treat DM in Malaysia (Noor& Ashcroft, 1989). In this paper, we report the isolation andstructural elucidation of 12 known compounds from T. crispaand evaluate the effects of the isolated compounds onα-glucosidase and α-amylase.
2. Materials and methods
2.1. General
Ultraviolet (UV) spectra were recorded on a Genesys 10S UV-VIS (Thermo Fisher Scientific, Madison, WI, USA) visiblespectrophotometer in methanol solution. Infrared (IR) spectrawere recorded on a Perkin Elmer 100 FT-IR (Beaconsfield, UK)spectrophotometer as KBr pellets or thin films. NMR spectrawere recorded on a Bruker Avance 500 Spectrometer (Faelanden,Switzerland), using 500 MHz for 1H and 125 MHz for 13C. Deu-terated chloroform (CDCl3) and deuterated methanol (MeOD)were used as solvents (Merck, Darmstadt, Germany).Electrospray ionization mass spectra (ESIMS) were obtainedusing a JEOL JMS-HX-110 (Peabody, MA, USA) mass spectrometer.Silica gel (70–230 mesh) was used for column chromatogra-phy. Thin-layer chromatography (TLC) was performed on aMerck pre-coated silica gel 60 F254+365. α-Glucosidase (Saccha-romyces cerevisiae), α-amylase (sourced from porcine pancreas),ρ-nitrophenyl α-D-glucopyranoside (ρ-NPG) and acarbose wereprocured from Sigma Aldrich Co. (St. Louis, MO, USA). Chemi-cals were purchased from Merck unless otherwise stated.
2.2. Plant materials
T. crispa vines were collected in Bentong, Pahang and identi-fied by Berhaman Ahmad (Botanist, Universiti Malaysia Sabah).A voucher specimen (FRI54832) was deposited at the Forest Re-search Institute of Malaysia (KEP).
2.3. Extraction and determination of the activecomposition using ultra-high performance liquidchromatography-quadrupole time-of-flight/massspectrometry (UPLC-QToF/MS)
Powdered, dried T. crispa vines (5.0 kg) were defatted by per-colation with hexane and filtration (fractionA).The plant materialwas dried, extracted by three cycles of sonication (30 min each)with MeOH–H2O (4:1), and filtered to obtain a brown extract (frac-tion B), which was concentrated under low pressure. Theconcentrated extract was acidified with H2SO4 to pH 2, and thenpartitioned with CHCl3.The CHCl3 phase was collected and con-centrated, leaving a brown gummy residue (fraction C) containingnon-alkaloidal components. The aqueous phase was basifiedwith NH4OH to pH 10, and extracted with MeOH–CHCl3 (3:1) toyield a dark brown gummy residue (fraction D) most likely, con-taining alkaloids. A small amount of fractions C and D weredissolved in methanol and subjected to UPLC-QToF/MS usingan HSS BEH C18 column (2.1 × 100 mm, 1.8 µm). HPLC data wereobtained on a Waters Xevo G2-S QToF. The mobile phase wasa mixture of water with 0.1% formic acid (A) and a mixture ofMeOH with 0.1% formic acid (B). The polarity was changed byadjusting the ratio of MeOH.The injection volume was 2 µl, andthe flow rate was set at 0.4 ml/min. The Q-ToF MS was oper-ated in the range 100–1500 Da, in both the positive and negativemodes. All operations, acquisitions, and data analyses weremonitored by the Natural Products Application Solution withUNIFI featuring the Traditional Medicine Library.
2.4. Isolation of active compounds
Fraction C (35.0 g) was chromatographed using a column withsilica gel slurry packed in CHCl3. Elution was initiated withCHCl3, with progressively increasing concentrations of MeOHin CHCl3, until reaching 100% MeOH.This protocol yielded com-pounds (1) (4.5 mg), (2) (5.5 mg) and (3) (100 mg). Fraction D(25.0 g) was chromatographed using the same conditions de-scribed above, yielding nine sub-fractions (F1–F9). F2 (2.31 g)was chromatographed using SiO2 (100 g) with hexane-ethylacetate as an eluent to yield 78 sub-fractions. Sub-fractions withsimilar TLC profiles were combined to obtain four new sub-fractions, labeled F2A–F2D. F2C (110 mg) was chromatographedby silica gel column using hexane-ethyl acetate as an eluent,yielding 30 sub-fractions. Those with similar TLC profiles werecombined and labeled as F2Ca–F2Cc. Sub-fractions 25–30 labeledformed F2Cc (70 mg), and were re-chromatographed withhexane:EtOAc (9:1) (Rf: 0.40), yielding compound (5) (8.0 mg; ayellowish powder). Sub-fractions 18–24 from F2Ca and F2Cbwere combined and re-chromatographed using preparative TLCwith hexane:ethyl acetate (8:2) to obtain compound (4) (3.5 mg),as a light brown powder with an Rf of 0.48. Subfraction 5was subjected to silica gel column chromatography with MeOH–CHCl3 (6:4) elution to yield compounds (6) (5.5 mg) and
75j o u rna l o f f un c t i ona l f o od s 1 6 ( 2 0 1 5 ) 7 4 – 8 0

(7) (10 mg). Subfraction 8 was eluted with MeOH–CHCl3 gradu-ally. After reaching 50%, a few drops of NH4OH were added toyield compound (8) (8 mg). Subfraction 2 was eluted with EtOAc–Hex (7:3) to yield compounds (9) (3.5 mg) and (10) (45 mg).Subfraction 3 was eluted with CHCl3–MeOH (3:1) to yield com-pound (11) (40 mg).
2.5. α-Glucosidase inhibition
Inhibition of α-glucosidase activity was evaluated by a previ-ously reported method (Kim, Kwon, & Son, 2000; Subramaniam,Asmawi, & Sadikun, 2008) The reaction mixture consisted of0.1 M phosphate buffer (pH 6.9, 50 µl), 0.5 mM 4-nitrophenyl α-D-glucopyranoside (PNPG) in 0.1 M phosphate buffer (pH 6.9, 25 µl),test sample (10 µl), and 0.1 U/ml α-glucosidase solution in 0.01 Mphosphate buffer (pH 6.0, 25 µl), and was incubated at 37 °C for20 min. The reaction was quenched by the addition of 0.1 Msodium carbonate solution (100 µl). The enzymatic hydrolysisof the substrate was monitored by the amount of p-nitrophenolreleased in the reaction mixture at 400 nm. Individual blankscomprising buffer only were prepared to correct for back-ground absorbance. Controls were conducted in an identicalmanner, replacing the plant extracts with methanol. Acarbosewas used as a positive control. All experiments were carriedout in triplicate. The inhibitory activity of α-glucosidase wasassessed by the following formula:
Inhibitory activity A A Acontrol sample control% = −( ) ×Δ Δ Δ 100
2.6. α-Amylase inhibition
Inhibition of α-amylase activity was evaluated using a previ-ously reported method by Shu et al. (2009) with minormodifications. The reaction mixture consisted of test sample(10 µl) dissolved in DMSO to obtain varying concentrations and0.1 U/ml α-amylase solution in 0.01 M phosphate buffer (pH 7.0,50 µl). The reaction mixture was pre-incubated for 20 min at37 °C. Potato starch solution (50 µl) was added and incubatedfor 20 min at 37 °C. The reaction was terminated by the addi-tion 1% 3,5-dinitrosalicylic acid (DNS) and 12% sodiumpotassium tartrate in 0.4 M NaOH (100 µl).The reaction mixturewas heated for 15 min at 100 °C and cooled to room tempera-ture. The α-amylase activity was determined by measuring theabsorbance at 540 nm. Individual blanks were prepared by re-placing the enzymes with buffer to correct for the backgroundabsorbance. Controls were conducted in an identical manner,replacing the samples with DMSO as a negative control andacarbose as a positive control. All experiments were carriedout in triplicate.
2.7. Statistical analyses
All results are expressed as mean ± SD. IC50 values were onlydetermined for the plant extracts with inhibition ≥50% at5 mg/ml by plotting a percent inhibition versus concentra-tion curve, in which the concentration of sample required for50% inhibition was determined and expressed as IC50 value.
3. Results and discussion
3.1. Isolation of active compounds and structuraldetermination
Eleven pure compounds were isolated from the non-alkaloidal(fraction C) and alkaloidal (fraction D) as described above andidentified as borapetoside C (1), 4-hydroxy benzaldehyde (2),β-sitosterol (3), liriodenine (4), lysicamine (5),dihydrodiscretamine(6), columbamine (7), magnoflorine (8), N-formylannonaine (9),N-formylnornuciferine (10), and N-trans-feruloyltyramine (11)(Fig. 1). NMR data for compounds (6)–(11) were reported in ourprevious paper (Yusoff, Hamid, & Houghton, 2014).
Compound (1), colorless needles. 1H NMR (CDCl3, 500 MHz):δ 1.16 (3H, s, 20-CH3), 1.44 (3H, s, 19-CH3), 3.75 (3H, s, OCH3), 4.45(1H, d, 7.6, Sugar), 4.68 (1H, d, 4.55,6-H), 5.84 (1H, dd, 12.15,1.55,12-H), 6.56 (1H, d, 1.05, 14-H), 6.84 (1H, t, 3.65, 3-H), 7.52 (1H,t, 1.65, 15-H), 7.62 (1H, s, 16-H). 13C NMR (500 MHz, CDCl3): 26.1(C1), 18.2 (C-2), 141.2 (C-3), 137.1 (C-4), 40.1 (C-5), 77.3 (C-6), 28.1(C-7), 46.3 (C-8), 37.2 (C-9), 28.3 (C-10), 44.1 (C-11), 70.2 (C-12),126.5 (C-13), 109.3 (C-14), 139 (C-15), 144.2 (C-16), 176.1 (C-17),168.4 (C-18), 37.4 (C-19), 30.1 (C-20), 52.6 (COOCH3), 105.2 (Sugar-1′), 74.8 (Sugar-2′), 75.2 (Sugar-3′), 69.8 (Sugar-4′), 74.8 (Sugar-5′), 61.8 (Sugar-6′).Yield (50.0 mg, 0.001%). ESIMS m/z 535 [M-H]−.The NMR data are consistent with those reported by Fukuda,Yonemitsu, and Kimura (1993). Compound (1) was identifiedas borapetoside C.
Compound (2), pale brown solid. 1H NMR (CDCl3, 500 MHz):6.70 (1H, d, J = 8.70, H-2), 6.94 (2H, d, J = 8.55, H-6), 7.25 (d, J = 8.30,H-3), 7.809 (D, J = 8.75, H-5), 9.287 (OH, s). 13C NMR (500 MHz,CDCl3): 114.0 (C-2), 115.4 (C-6), 128.2 (C-4), 132.4 (C-3, C-5), 149(C-4), 162 (C-1). ESIMS m/z 123 [M+]. This compound was re-ported in this plant for the first time. Compound (2) wasidentified as 4-hydroxy benzaldehyde.
Compound (3): white powder, 1H NMR (CDCl3, 500 MHz) spec-trum displayed 3.55 (1H, m), 5.38 (1H, d, J = 5.40 Hz), 0.70 (3H,s), 1.03 (3H, s), 0.95 (3H, d, J = 6.55 Hz), 0.85 (3H, d, J = 4.40 Hz),0.87 (3H, d, J = 1.60 Hz), 0.83 (3H, t, J = 1.45 Hz), 1.07–2.36 (22H,m). ESIMS m/z 275 [M+]. The MS and NMR data are identicalwith those in the literature (Khaleque, Maith, Huq, & Basar,1970). Compound (3) was identified as β-sitosterol.
Compound (4), light brown powder, 1H NMR (CDCl3, 500 MHz):6.39 (2H, s, OCH2O), 7.23 (1H, s, H-3), 7.61 (1H, t, J = 7.40, H-9),7.78 (1H, m, H-10), 7.81 (1H, d, J = 5.00, H-4), 8.60 (1H, d, J = 8.15,H-8), 8.70 (1H, d, J = 8.30, H-11), 8.91 (1H, d, J = 5.00, H-5). 13C NMR(CDCl3, 500 MHz); 101.84 (OCH2O), 103.41 (C-3), 108.74 (C-11b),122.62 (C-11c), 124.50 (C-4), 127.72 (C-11), 128.08 (C-8), 128.87(C-9), 130.95 (C-7a), 132.57 (C-11a), 134.23 (C-10), 135.46 (C-6a),145.32 (C-5), 145.83 (C-3a), 148.03 (C-1), 151.93 (C-2), 182.46 (C-7);ESIMS m/z 275 [M+]. The NMR data were consistent with thosein the literature (Husain, Jamal, & Jalil, 2012). Compound (4)was identified as liriodenine.
Compound (5), yellow powder, 1H NMR (CDCl3, 500 MHz): 4.05(3H, s, 1-OCH3), 4.14 (3H, s, 2-OCH3), 7.27 (1H, s, H-3), 7.60 (1H,t, J = 8.0, H-9), 7.80 (1H, dd, J = 8.5,1.5, H-10), 7.86 (1H, d, J = 5.20,H-4), 8.63 (1H, d, J = 8.0, H-11), 8.96 (1H, d, J = 5.20, H-5), 9.22 (1H,d, J = 8.0, H-11). 13C NMR (CDCl3, 500 MHz): 56.24 (2-OCH3), 60.69(1-OCH3), 106.49 (C-3), 119.95 (C-11c), 122.26 (C-3a), 123.63 (C-9),124.49 (C11b), 128.48 (C-8), 128.85 (C-9), 123.95 (C-10), 130.88
76 j o u rna l o f f un c t i ona l f o od s 1 6 ( 2 0 1 5 ) 7 4 – 8 0
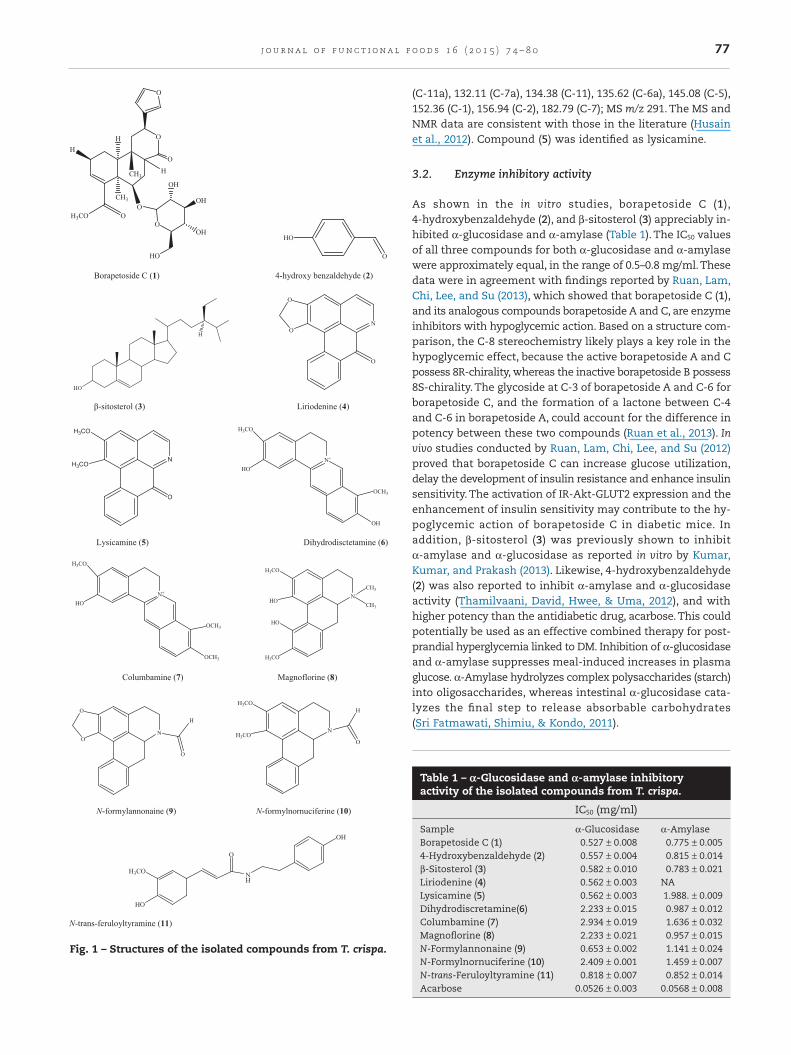
(C-11a), 132.11 (C-7a), 134.38 (C-11), 135.62 (C-6a), 145.08 (C-5),152.36 (C-1), 156.94 (C-2), 182.79 (C-7); MS m/z 291. The MS andNMR data are consistent with those in the literature (Husainet al., 2012). Compound (5) was identified as lysicamine.
3.2. Enzyme inhibitory activity
As shown in the in vitro studies, borapetoside C (1),4-hydroxybenzaldehyde (2), and β-sitosterol (3) appreciably in-hibited α-glucosidase and α-amylase (Table 1). The IC50 valuesof all three compounds for both α-glucosidase and α-amylasewere approximately equal, in the range of 0.5–0.8 mg/ml.Thesedata were in agreement with findings reported by Ruan, Lam,Chi, Lee, and Su (2013), which showed that borapetoside C (1),and its analogous compounds borapetoside A and C, are enzymeinhibitors with hypoglycemic action. Based on a structure com-parison, the C-8 stereochemistry likely plays a key role in thehypoglycemic effect, because the active borapetoside A and Cpossess 8R-chirality, whereas the inactive borapetoside B possess8S-chirality. The glycoside at C-3 of borapetoside A and C-6 forborapetoside C, and the formation of a lactone between C-4and C-6 in borapetoside A, could account for the difference inpotency between these two compounds (Ruan et al., 2013). Invivo studies conducted by Ruan, Lam, Chi, Lee, and Su (2012)proved that borapetoside C can increase glucose utilization,delay the development of insulin resistance and enhance insulinsensitivity. The activation of IR-Akt-GLUT2 expression and theenhancement of insulin sensitivity may contribute to the hy-poglycemic action of borapetoside C in diabetic mice. Inaddition, β-sitosterol (3) was previously shown to inhibitα-amylase and α-glucosidase as reported in vitro by Kumar,Kumar, and Prakash (2013). Likewise, 4-hydroxybenzaldehyde(2) was also reported to inhibit α-amylase and α-glucosidaseactivity (Thamilvaani, David, Hwee, & Uma, 2012), and withhigher potency than the antidiabetic drug, acarbose. This couldpotentially be used as an effective combined therapy for post-prandial hyperglycemia linked to DM. Inhibition of α-glucosidaseand α-amylase suppresses meal-induced increases in plasmaglucose. α-Amylase hydrolyzes complex polysaccharides (starch)into oligosaccharides, whereas intestinal α-glucosidase cata-lyzes the final step to release absorbable carbohydrates(Sri Fatmawati, Shimiu, & Kondo, 2011).
O
O
OH
O
H
H3CO O
H
CH3
OH
OH
OHO
HO
CH3
O
HO
Borapetoside C (1) 4-hydroxy benzaldehyde (2)
HO
H
N
O
O
O
β-sitosterol (3) Liriodenine (4)
N
H3CO
H3CO
O
N+
H3CO
HO
OCH3
OH
Lysicamine (5) Dihydrodisctetamine (6)
N+
H3CO
HO
OCH3
OCH3
N+
CH3
H3CO
HO
HO
H3CO
CH3
Columbamine (7) Magnoflorine (8)
N
O
H
O
ON
O
HH3CO
H3CO
N-formylannonaine (9) N-formylnornuciferine (10)
NH
O
OH
H3CO
HO
N-trans-feruloyltyramine (11)
Fig. 1 – Structures of the isolated compounds from T. crispa.
Table 1 – α-Glucosidase and α-amylase inhibitoryactivity of the isolated compounds from T. crispa.
IC50 (mg/ml)
Sample α-Glucosidase α-AmylaseBorapetoside C (1) 0.527 ± 0.008 0.775 ± 0.0054-Hydroxybenzaldehyde (2) 0.557 ± 0.004 0.815 ± 0.014β-Sitosterol (3) 0.582 ± 0.010 0.783 ± 0.021Liriodenine (4) 0.562 ± 0.003 NALysicamine (5) 0.562 ± 0.003 1.988. ± 0.009Dihydrodiscretamine(6) 2.233 ± 0.015 0.987 ± 0.012Columbamine (7) 2.934 ± 0.019 1.636 ± 0.032Magnoflorine (8) 2.233 ± 0.021 0.957 ± 0.015N-Formylannonaine (9) 0.653 ± 0.002 1.141 ± 0.024N-Formylnornuciferine (10) 2.409 ± 0.001 1.459 ± 0.007N-trans-Feruloyltyramine (11) 0.818 ± 0.007 0.852 ± 0.014Acarbose 0.0526 ± 0.003 0.0568 ± 0.008
77j o u rna l o f f un c t i ona l f o od s 1 6 ( 2 0 1 5 ) 7 4 – 8 0
The activity of the isolated alkaloids, (6)–(11) againstα-glucosidase and α-amylase are shown in Table 1. Three al-kaloids, liriodenine, lysicamine,and N-formylannonaine,showedstrong inhibitory activity against α-glucosidase, with IC50 valuesranging from 0.5 to 0.8 mg/ml. The effect of alkaloids againstα-amylase varied.N-trans-feruloyltyramine,dihydrodiscretamine,and magnoflorine,exhibited significant inhibitory activity againstα-amylase,with IC50 values ranging from 0.8 to 0.9 mg/ml.Almostall of the isolated alkaloids formed A-rings, B-rings, and C-ringswith sp2 hybridization, allowing these atoms to form a largeconjugated-system necessary for interaction with α-glucosidaseand α-amylase (Wang, Du, & Song, 2010). However, some showedweak inhibitory activity, likely due to the different functionalgroups attached to the alkaloids.We deduced that the presenceof methylene dioxide in compounds (4) and (5) plays an importantrole in α-glucosidase inhibition, while the two hydroxyl groupsin (6) and (8) are favorable for α-amylase inhibition. N-trans-Feruloyltyramine, which lacks rings A, B, and C, possesses
significant inhibitory activity.Structure–activity relationship analy-sis indicates that the phenyl hydroxyl group is fundamental forthe activity of these compounds (Borges de Melo, Da SilveiraGomes, & Carvalho, 2006). Fan, Terrier, Hay, Marston, andHostettmann (2010) showed that N-trans-feruloyltyramine hasstronger activity than the positive control acarbose(IC50 = 0.135 mg/ml). The enzyme inhibitory effects of most ofisolated alkaloids in the studied concentration range were notas potent as acarbose but it shows promising activity. Thesestudies will help us determine the functional groups respon-sible for the bioactivity of the compounds, allowing for therationale design of new lead compounds for these biologicaltargets.
A number of alkaloids from natural sources have been provenefficacious in curing various ailments. Indole, carbazole,isoquinoline and aporphine alkaloids are several examples whichare useful in treatment of diabetes mellitus (Patel & Mishra, 2011).As a traditional medicinal herb, T. crispa was studied fre-
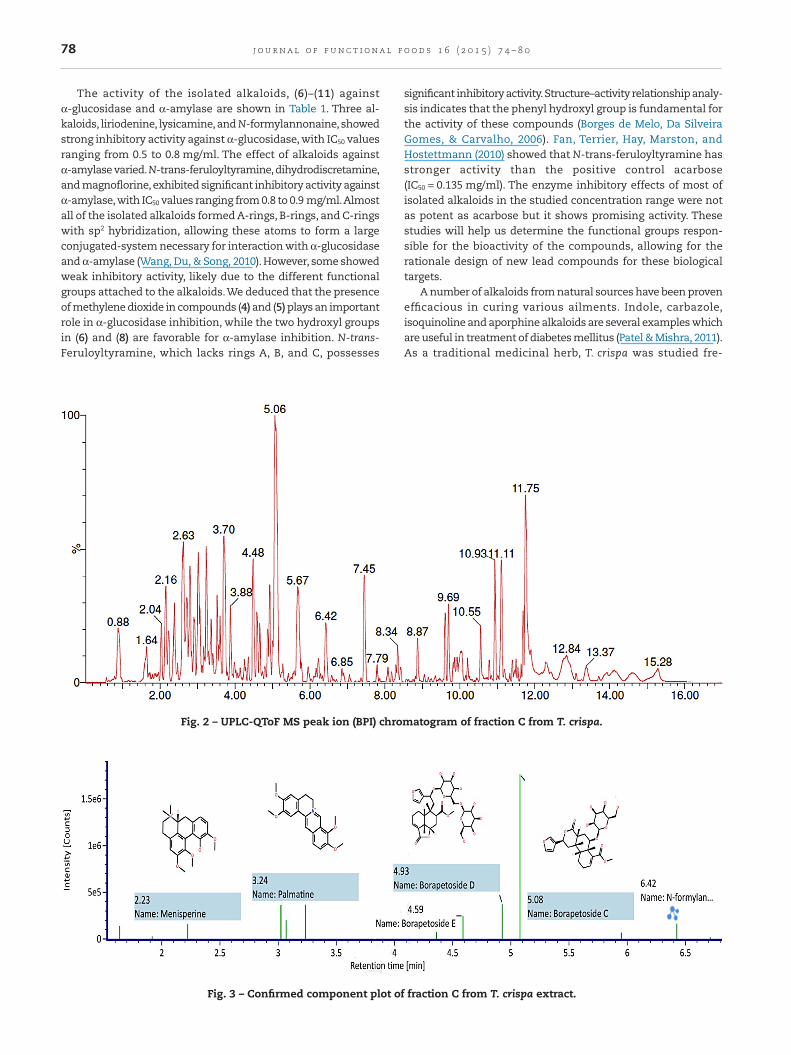
Fig. 2 – UPLC-QToF MS peak ion (BPI) chromatogram of fraction C from T. crispa.
Fig. 3 – Confirmed component plot of fraction C from T. crispa extract.
78 j o u rna l o f f un c t i ona l f o od s 1 6 ( 2 0 1 5 ) 7 4 – 8 0
quently for its beneficial role in diabetes (Noor & Ashcroft, 1989;Ruan et al., 2012, 2013). However mechanisms by which variousoral therapeutic agents achieve antidiabetic action could be linkedto more than one mechanism, such as insulin sensitizing, insulinreleasing, gluconeogenesis inhibiting, and α-glucosidase inhib-iting (Davis & Granner, 2001). As described by Wagner andUlrich-Merzenich (2009), the effect of phytomedicines can be betterevaluated by studying synergistic effects via multi-target effectsor effects on pharmacokinetic or physicochemical properties.Thus, it is worthwhile to evaluate further the effective compo-nents of isolated compounds in vivo rather than make a conclusionbased on enzyme inhibition assay only.
3.3. Analysis of HPLC data
To investigate the relationship between activities and extractcomposition, the content of the active components in the ex-tracts were determined by UPLC-QToF/MS. QToF/MS providesaccurate MS/MS spectra, due to mass drift compensation andinternal calibration during acquisition.The compounds in frac-tion C and D were identified by their characteristic mass
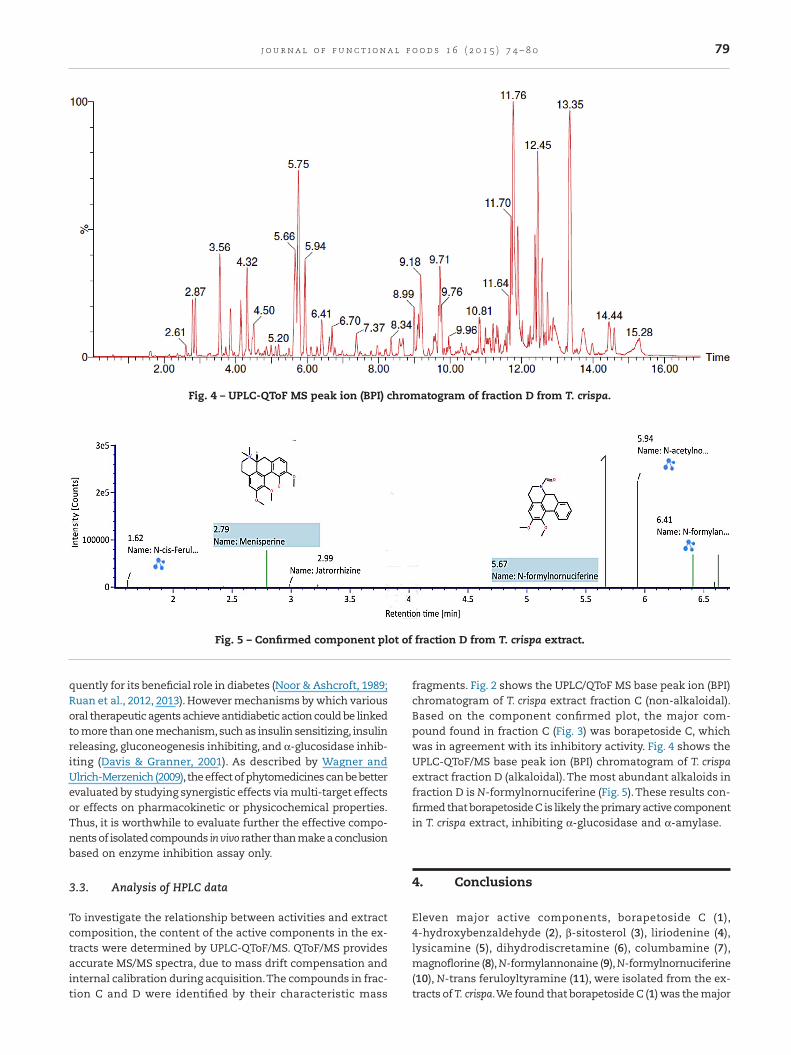
fragments. Fig. 2 shows the UPLC/QToF MS base peak ion (BPI)chromatogram of T. crispa extract fraction C (non-alkaloidal).Based on the component confirmed plot, the major com-pound found in fraction C (Fig. 3) was borapetoside C, whichwas in agreement with its inhibitory activity. Fig. 4 shows theUPLC-QToF/MS base peak ion (BPI) chromatogram of T. crispaextract fraction D (alkaloidal).The most abundant alkaloids infraction D is N-formylnornuciferine (Fig. 5).These results con-firmed that borapetoside C is likely the primary active componentin T. crispa extract, inhibiting α-glucosidase and α-amylase.
4. Conclusions
Eleven major active components, borapetoside C (1),4-hydroxybenzaldehyde (2), β-sitosterol (3), liriodenine (4),lysicamine (5), dihydrodiscretamine (6), columbamine (7),magnoflorine (8), N-formylannonaine (9), N-formylnornuciferine(10), N-trans feruloyltyramine (11), were isolated from the ex-tracts of T. crispa.We found that borapetoside C (1) was the major
Fig. 4 – UPLC-QToF MS peak ion (BPI) chromatogram of fraction D from T. crispa.
Fig. 5 – Confirmed component plot of fraction D from T. crispa extract.
79j o u rna l o f f un c t i ona l f o od s 1 6 ( 2 0 1 5 ) 7 4 – 8 0

active compound in these extracts. Furthermore, there was anobvious relationship between the structure and the activity inthe isolated compounds. Additionally, the alkaloidal fractionscould have a synergistic effect via inhibition of α-glucosidaseand α-amylase, as different alkaloid had different activity.Thus,the components of T. crispa are efficient inhibitors of the starch-digesting enzymes α-amylase and α-glucosidase.
Acknowledgements
Support for this work was provided by Universiti MalaysiaPahang via research grant UMP RDU 100322 to MM Yusoffand UMP RDU 120603 to MR Karim in addition to pre-commercialization grant UMP UIC 100303 to MM Yusoff. M Liuwas supported by Universiti Malaysia Pahang’s Doctoral Scheme.The authors are also grateful to IM Choudhary (HEJ Instituteof Chemistry, Karachi, Pakistan) for mass spectra acquisi-tions and to Dr Patel (Waters Pacific Pte Ltd, Singapore) for UPLC-QTof data acquisition.
R E F E R E N C E S
Ali, H., Houghton, P. J., & Soumyanath, A. (2006). α-Amylaseinhibitory activity of some Malaysian plants used to treatdiabetes; with particular reference to Phyllanthus amarus.Journal of Ethnopharmacology, 107, 449–455.
Borges de Melo, E., Da Silveira Gomes, A., & Carvalho, I. (2006).α-Glucosidase and β-glucosidase inhibitors: Chemicalstructure and biological activity. Tetrahedron, 62, 10277–10302.
Braithwaite, M. C., Tyagi, C., Tomar, L. K., Kumar, F., Choonara, Y.E., & Pillay, V. (2014). Nutraceutical-based therapeutics andformulation strategies augmenting their efficiency tocomplement modern medicine. An overview. Journal ofFunctional Foods, 6, 82–99.
Cho, M., Han, J. H., & You, S. (2011). Inhibitory effects of fucansulfates on enzymatic hydrolysis of starch. Food Science andTechnology, 44, 1164–1171.
Davis, S. N., & Granner, D. K. (2001). Oral hypoglycemic agents. InJ. G. Hardman, L. E. Limbird, & A. G. Gilman (Eds.), Goodmanand Gilman’s the Pharmacological Basis of Therapeutics, (10th ed.,pp. 1701–1707). New York, USA: McGraw-Hill International.
Fan, P., Terrier, L., Hay, A. E., Marston, A., & Hostettmann, K.(2010). Antioxidant and enzyme inhibition activities andchemical profiles of Polygonum sachalinensis F.Schmidt ex Maxim(Polygonaceae). Fitoterapia, 81, 124–131.
Fukuda, N., Yonemitsu, M., & Kimura, T. (1993). Isolation andstructure elucidation of the five new furanoid diterpene gly-coside borapetoside C–G. Liebigs Annalen Der Chemie, 491–495.
Husain, K., Jamal, J. A., & Jalil, J. (2012). Phytochemical study ofCananga odorata (Lam) Hook.f. & Thomson & Thoms(Annonaceae). International Journal of Pharmacy andPharmaceutical Sciences, 4, 75–77.
Josse, R. G., Chiasson, J.-L., Ryan, E. D., Lau, D. C. W., Ross, S. A.,Yale, J. F., Leiter, L. A., Maheu, P., Tessier, D., Wolever, T.,Gerstein, H., Rodger, W., Dorman, J. M., Murphy, L. J., Rabasa-Lhoret, R., & Meneilly, G. S. (2003). Acarbose in the treatmentof elderly patients with type 2 diabetes. Diabetes Research andClinical Practice, 59, 37–42.
Khaleque, A., Maith, M. A. W., Huq, M. S., & Basar, K. A. (1970).Tinospora cordifolia. Isolation heptacosanol, β-sitosterol andthree other compounds tinosporine, cordifol and cordifolone.Pakistan Journal of Scientific and Industrial Research, 7, 61.
Kim, J. S., Kwon, C. S., & Son, K. H. (2000). Inhibition ofα-glucosidase and amylase by luteolin, a flavonoid. Bioscience,Biotechnology, and Biochemistry, 64, 2458–2461.
Kumar, S., Kumar, V., & Prakash, O. (2013). Enzymes inhibitionand antidiabetic effect of isolated constituents from Dilleniaindica. BioMed Research International, 2013, 1–7.
Mitrakou, A., Kelley, D., Mokan, M., Veneman, T., Pangburn, T.,Reilly, J., & Gerich, J. (1992). Role of reduced suppression ofglucose production and diminished early insulin release inimpaired glucose tolerance. New England Journal of Medicine,326, 22–29.
Noor, H., & Ashcroft, S. J. H. (1989). Antidiabetic effects ofTinospora crispa in rats. Journal of Ethnopharmacology, 27, 149–161.
Patel, M. B., & Mishra, S. (2011). Hypoglycemic activity ofalkaloidal fraction of Tinospora cordifolia. Phytomedicine, 18,1045–1052.
Porte, D., Jr., & Kahn, S. E. (2001). Beta-cell dysfunction and failurein type 2 diabetes: Potential mechanisms. Diabetes, 50(Suppl.1), S160–S163.
Ruan, C. T., Lam, S. H., Chi, T. C., Lee, S. S., & Su, M. J. (2012).Borapetoside C from Tinospora crispa improves insulinsensitivity in diabetic mice. Phytomedicine, 19, 719–724.
Ruan, C. T., Lam, S. H., Chi, T. C., Lee, S. S., & Su, M. J. (2013).Hypoglycemic action of borapetoside A from the plantTinospora crispa in mice. Phytomedicine, 20, 667–675.
Sang, S. (2014). Bioavailability and metabolism of bioactivecomponents. Journal of Functional Foods, 7, 1–2.
Shu, X. S., Lv, J. H., Tao, J., Li, G. M., Li, H. D., & Ma, N. (2009).Antihyperglycemic effects of total flavonoids fromPolygonatum odoratum in STZ and alloxan-induced diabeticrats. Journal of Ethnopharmacology, 124, 539–543.
Sogaard-Andersen, L., & Valentin-Hansen, P. (1993). Protein–protein interactions in gene regulation: The cAMP–CRPcomplex sets the specificity of a second DNA-binding protein,the CytR repressor. Cell, 75, 557–566.
Sri Fatmawati, Shimiu, & Kondo, R. (2011). Ganoderol B: A potentα-glucosidase inhibitor isolated from the fruiting body ofGanoderma lucidu. Phytomedicine, 18, 1053–1055.
Subramaniam, R., Asmawi, M. Z., & Sadikun, A. (2008). In vitroα-glucosidase and α-amylase enzyme inhibitory effects ofAndrographis paniculata extract and andrographolide. ActaBiochimica Polonica, 55, 391–398.
Svensson, E. I., Kristoffersen, L., Oskarsson, K., & Bensch, S.(2004). Molecular population divergence and sexual selectionon morphology in the banded demoiselle (Calopteryxsplendens). Heredity, 93, 423–433.
Teeri, T. T. (1991). Engineering of enzymes of carbohydratemetabolism. Current Opinion in Biotechnology, 2, 614–621.
Thamilvaani, M., David, A., Hwee, M. C., & Uma, D. P. (2012).Flavonoids isolated from Syzygium aqueum leaf extract aspotential antihyperglycaemic agents. Food Chemistry, 132,1802–1807.
Wagner, H., & Ulrich-Merzenich, G. (2009). Synergy research:Approaching a new generation of phytopharmaceuticals.Phytomedicine, 16, 97–110.
Wang, H., Du, Y. J., & Song, H. C. (2010). α-Glucosidase andα-amylase inhibitory activities of guava leaves. Food Chemistry,123, 6–13.
Yusoff, M., Hamid, H., & Houghton, P. (2014). Anticholinesteraseinhibitory activity of quaternary alkaloids from Tinosporacrispa. Molecules, 19, 1201–1211.
80 j o u rna l o f f un c t i ona l f o od s 1 6 ( 2 0 1 5 ) 7 4 – 8 0
Related Documents











