1 הכלורופלסטית הפרוטאזה אפיוןFtsH בצמחיArabidopsis thaliana תואר קבלת לשם חיבור“ לפילוסופיה דוקטור” מאת זלצמן עדי בירושלים העברית האוניברסיטה לסנאט מוגש תשס" ה ספט מבר, 2005

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1
בצמחיFtsHאפיון הפרוטאזה הכלורופלסטית Arabidopsis thaliana
חיבור לשם קבלת תואר
”דוקטור לפילוסופיה“
מאת
עדי זלצמן
מוגש לסנאט האוניברסיטה העברית בירושלים
2005, מבר ספטה"תשס

2
תקציר
FtsHהוא מטלופרוטאזה ממברנלית התלויה ב -ATP ,הוא שייך למשפחת ה-AAA . החלבון פעיל בתור
. נמצא כי הוא מקודד על ידי גן יחיד והוא הפרוטאזה החיונית היחידה בוE. coliבחיידקי . קומפלקס הקסמרי
, אפונה מצמחיFtsH - הצמחי לסט בכלורופ FtsH של קיומו עלהחל מאמצע שנות התשעים הצטבר מידע
תגובה עם נוגדנים כנגדו לפי ו-E. coli מבודד ושובט על בסיס דמיון לחלבון , וארבידופסיס אופיין, תרד
(Lindahl et al., 1996) .FtsH 1 חלבוןהמעורב בפירוק נמצאD בה הפוטוסינתטית ימרכז הראקצימ -PSII
(Lindahl et al., 2000).בצמחים , FtsH מהם התוצרים של תשעה . גנים12 של משפחהודד על ידימק
רק ,יחד עם זאת. (Sakamoto et al., 2003)עשויים להיות מוכוונים לכלורופלסט ושלושה למיטוכונדריה
FtsH1, FtsH2, FtsH 5, FtsH 8 (Sinvany-Villalobo : ארבעה אכן נמצאו באנליזת חלבוני התילקואיד
et al., 2004, Yu et al., 2004) .של כמות החלבון הרבה ביותר היא , בכלורופלסטFtsH2 כשאחריו
בירוק ולבן לכתמים כתוצאה ממוטציות גורם ftsh2אובדן הגן . FtsH5 ,FtsH1 FtsH8,בהתאמה
הירוקים שהאזוריםבעוד (Chen et al., 2000, Takechi et al., 2000). , (variegation) ןלסירוגי
מצביע על זה דבר.כלורופלסטיםכלל הלבנים לא נראו באזורים, םי נורמלישנראיםסטים כלורופלמכילים
להכתמה קלה בלבד גורם FtsH5-חוסר ב, בדומה. בביוגנזה של הכלורופלסטFtsHמעורבות של
(Sakamoto et al., 2002) . רגישות זו מתבטאת . רגישים יותר לאור בעוצמה גבוההבעלי ההכתמההצמחים
פלואורסנציה של כלורופילב הנמדד, PSIIה בפוטנציאל הפעילות של ביריד
: הבאות בשאלותיהתמקדתואת המחקר שלי התחלתי 2000בשנת
?היחסים בין הגנים השונים במשפחההם מה •
?חבצמ FtsH מה תפקידם של חלבוני •
?חופפים או שוניםהשונים האם תפקידי האיזומרים •

3
: וקיבלתי את התוצאות הבאותFtsH -ים בגנים השונים לבמהלך עבודתי חקרתי מוטנט
. FtsH8- ו FtsH2וכך גם , הינם גנים דופליקטים-FtsH5 וFtsH1 כי מראהניתוח פילוגנטי
.במופע החיצוני וברגישות לאור בעוצמה גבוהה WT- מפנוטיפ השונהל גרם לא FtsH8 או FtsH1- בחסר
בתנאי .עלים שוניםבשונה היא אחידה ואינה כי רמת ההכתמה נו מצאftsh2בחינה מעמיקה של המוטנט ב
העלה אך נובט עם פסיגים ירוקים ftsh2כי ונראה ,בשלב ההתפתחותירמת ההכתמה תלויה ,סביבה מבוקרים
רמת ההורקה הולכת ועולה ככל שהעלה נוצר בשלב מאוחר ,בהמשך. האמתי הראשון הינו לבן כמעט לגמרי
במוטנט נמוכה FtsH כמותו של עדייןאך , FtsHה בכמות חלבוני יעלייש , במקביל. יותר בתהליך הגידול
יתלויה בשלב ההתפתחותה FtsHחלבוני יש עלייה בכמות WT- גם בצמחיWT.-מכמותו בבכחמישים אחוז
.כמות החלבון גבוהה יותר, ככל שהעלה עליון יותר :mRNA-ה ברמות היונובעת מהעלי
רמות ,FtsHחסר בו כמעט לגמרי החלבון ש ,בעלה הלבן. התופעה שונהFtsH2 במוטנטים החסרים
הדבר מעיד . ואילו בעלה הירוק הן הנמוכות ביותר, האחרים הן הגבוהות ביותרFtsHחלבוני של -mRNAה
אפשרות של אינטראקציות בין תוצרי הגנים השונים ועל ה, פיצוי בין הגנים השוניםכך שקיים מנגנוןעל
. השונים הגידולישלבב במשפחה
FtsH נמצא מעורב בתיקון חלבוני D1 ממרכז הראקצייה של PSIIבחינת הכמות של חלבוני . בזמן עקת אור
FtsHבהמשך , במשך עקת האור מעלה כי חלה ירידה חדה בכמותם בדקות הראשונות של עקת האור
- ה להעיד על פירוק עקב נזק הנגרם להירידה הזו עשוי. מתרחשת עליה הדרגתית חזרה עד לכמות ההתחלתית
FtsHההתמתנות בקצב צבירת . עצמו והיא עשויה להסביר באופן חלקי את הקינטיקה של צבירת נזקי האור
.דבר המסביר את העלייה בקצב התיקון , המשתתף במערכת התיקון- FtsHנזקי האור מלווה בעליה בכמות
סדרה ויצרתי הכלאתי בין המוטנטים , בקומפלקס הפעיליםמשתתפ FtsHעל מנת לבחון האם שאר חלבוני
היה בעל ,ftsh5- וftsh2 שני הצמחים המוכתמים תוצר הכלאה של, כפולמוטנט , כצפוי. של מוטנטים כפולים
FtsH1מכאן עלתה השאלה האם . משל הוריו יותרבעל הכתמה חמורהוהתקבל צמח חמור יותר פנוטיפ
העדר ברקע של , על הפנוטיפותואם תהינה למוטציית החסר בו השפע, בושתתף קומפלקס הפעיל או מלתורם
FtsH2. התוצאות הראו מופע דומה למופע של מוטנט כפול של ftsh2 ו-ftsh5 . מכאן שגםFtsH1 משתתף

4
כאשר הכלאנו את המוטנט . פקודית לקומפלקסת ותבקומפלקס הפעיל ומהווה גורם התורם מבחינה כמותי
ftsh2המוטנט עם ftsh1או ftsh5 התקבלה העצמה של הפנוטיפ שהיה למוטנטftsh2 .ההחמרה זו היית
האם מה שקובע את עוצמת מנת לבחון על. בקורלציה לכמות החלבון של כל אחד מההומולוגים הללו
הפרטים החסרים אחד מזוג החלבונים הדופליקטיםלאתי בין שני הכ שחסרהFtsH-ההכתמה הוא אכן כמות ה
ושרדו רק הצמחים ההומוזיגוטים נראו קטנים ולבניםftsh8- לftsh2בהכלאה בין . השונים באופן רציפרוקלי
הם הצליחו להתפתח תוך התמינות של אברי הצמח במופע הנראה תקין ואף . על גבי מצע המועשר בסוכרוז
רו חסרות אבקה ועוברים ות נשא הצלק,וללא אבקה נראית לעין, אך האבקנים היו קטנים יותר. לפריחההגיעו
נראו דומים להומוזיגוטיםFtsH8- ובעלי אלל תקין יחיד לFtsH2הצמחים חסרי . לא התפתחו
אך הפסיגים היו ירוקים ובעלים , וגם הם לא התפתחו אלא על מצע הטרוטרופי, ftsh2 ftsh8המוטנטים
בניתוח .ftsh5 עם ftsh1צפתה בהכלאה של תופעה דומה נ .הגבוהים ניתן היה להבחין בכתמים קטנים ירוקים
FtsH2- וב-FtsH8מצאתי כי כתוצאה מהפגיעה ב, בעלים הלבנים באמצעות נוגדניםFtsHשל כמות חלבוני
איננו -FtsH8 וFtsH2 הזוג הנוסף -FtsH5 וFtsH1וגם כאשר חסרים , בכלורופלסטFtsHאין כלל חלבון
. נצבר
- וA "Ftsh1" סוג FtsH קיימים שני סוגים של חלבוני : 'ודל הכפולהמ' את ניתן להציעתוצאות אלו מ
FtsH5 וסוג "B" FtsH2 ו -FtsH8לפחות שני סוגים של דרושים בפלסטידות FtsH , "A"ו-"B" , על מנת
בהעדר אחת מתת היחידות . צמח חיוניוהתפתחותלאפשר ביוגנזה של כלורופלסטים ליצור קומפלקס פעיל ו
. אין ביוגנזה של כלורופלסטים והצמח הופך להיות הטרוטרופי ועקר ,FtsHקומפלקס של הללו לא נצבר ה
. נצברים גם כן חלבוני הסוג השני אינם, כאשר חסר אחד הסוגים
:המסקנות העיקריות העולות מעבודה זו הן
FtsH2 -האחד ענף ה .כלורופלסטליצירת ה כוללת שני ענפים החיוניים FtsHמשפחת הגנים של .1
ענפים אלו נקראים גם סוג .FtsH5 והדופליקט שלו FtsH1 - והענף השני , FtsH8הדופליקט שלו ו
"B" וסוג "A"בהתאמה .

5
2. FtsHחלקו של .לפחות שני מרכיביםהקסמרי בעל בתור הטרוקומפלקס ,ככל הנראה, פועל
. משש תת היחידות2/3- בממוצע כהוא FtsH8 או/ו FtsH2 - רכיבמה
מלווה ברגישות עולה FtsHירידה בכמותו של החלבון . עורב בהגנה מפני נזקי חמצוןקומפלקס זה מ .3
.לאור בעוצמות גבוהות
. ברקמות הצמח משתנה באופן טבעי בהתאם למיקום של העלה בשושנתFtsHכמות החלבון .4
תהיה מתאימה לכמות , ההכתמה–עוצמת הפנוטיפ , נפגע או מושתקFtsHכאשר אחד הגנים של .5
.ובלבד שכמות זו מעל סף מינמלי, ן החסרה החלבו
אשר גורמת לביטוי רב יותר של מספר , מופעלת מערכת משוב, מסויםFtsHכאשר חסר חלבון .6
. ולא רק של החלבון החסר עצמוFtsHחלבוני
כנראה כתוצאה מפגיעה , FtsHבשלבים הראשונים חלה ירידה בכמות החלבון של , בתנאי עקת אור .7
.ול לתרום לצבירת נזקי אור עקב חוסר פירוקם ואי תיקונם של החלבונים שנפגעוהדבר על. בו

6
מבוא
פרוטאוליזה
בעשורים . חיים וגם אותנו בני האדםה-את בעלי, את הצמחים: חלבונים מרכיבים את כל היצורים החיים"
אך מחקרים , ני התא השוניםהאחרונים המחקר הביוכימי התקדם מאוד ונמצא הסבר לאופן הייצור של חלבו
אברהם הרשקו וארווין , חנובר’בתחילת שנות השמונים אהרון צ. מעטים בלבד עסקו בנושא פירוק חלבונים
מערכת (תהליך הפירוק המבוקר : שחו נגד הזרם כאשר חקרו וגילו את תהליך המחזור החשוב ביותר בתאזרו
מתוך נימוקי ועדת ." (ומקבלים פרס נובל לשנה זו בכימיהועל שום כך הם זוכים להערכה רבה , )היוביקוויטין
.)2004פרס נובל כפי שפורסם בשישה באוקטובר
באמצעות בין היתר מבוקרים יםצמחים וחי-י בעלי בתאם והתפתחותיים מורפולוגיי, םפזיולוגייתהליכים
שך חיים הקצר ממשך חיי התא לחלק גדול מחלבוני התא מ. תהליכי סילוק סלקטיביים של חלבונים קצרי חיים
חלבונים המועמדים לפירוק יהיו כאלו העומדים בתנאי אחד או . )Adam, 1996(ומכאן יש לפרקם , עצמו
חלבונים אשר לא הגיעו , חלבונים אשר תנאי סביבה או התפתחות יכתיבו את פירוקם: יותר מהתנאים הבאים
למשל (חלבונים שהמבנה שלהם איננו מושלם , או כאלה שאינם נמצאים בקומפלקס הנורמלי שלהם, ליעדם
אי פירוק וסילוק של החלבונים הללו עשוי להביא . וחלבונים שנפגמו במהלך פעילותם) פאקטור-חוסר של קו
או ,חוסר פעילות של חלבון הפגוע במבנהו או באתרו הפעיל. התא והאורגניזים בכלל, לפגיעה בתפקוד המדור
מצטברים והפרעות במרחב בגלל קיומם של גושי חלבונים ה, פעילות שאינה מתרחשת בזמן או במקום הנכון
בצמחים למעלה מחמישים . יפריעו לתפקוד התקין של התא ולפיכך חייב התא לסלקם, )אגרגטים(בתא
מערכת סילוק החלבונים בתא החי היא חלק . )Vierstra, 1993(אחוזים מכלל החלבונים מוחלפים מדי שבוע
חלק מהפרוטאזות הפועלות בתא הן . אך פרוק איננו הגורם היחיד בבקרה זו, ממנגנוני בקרת האיכות
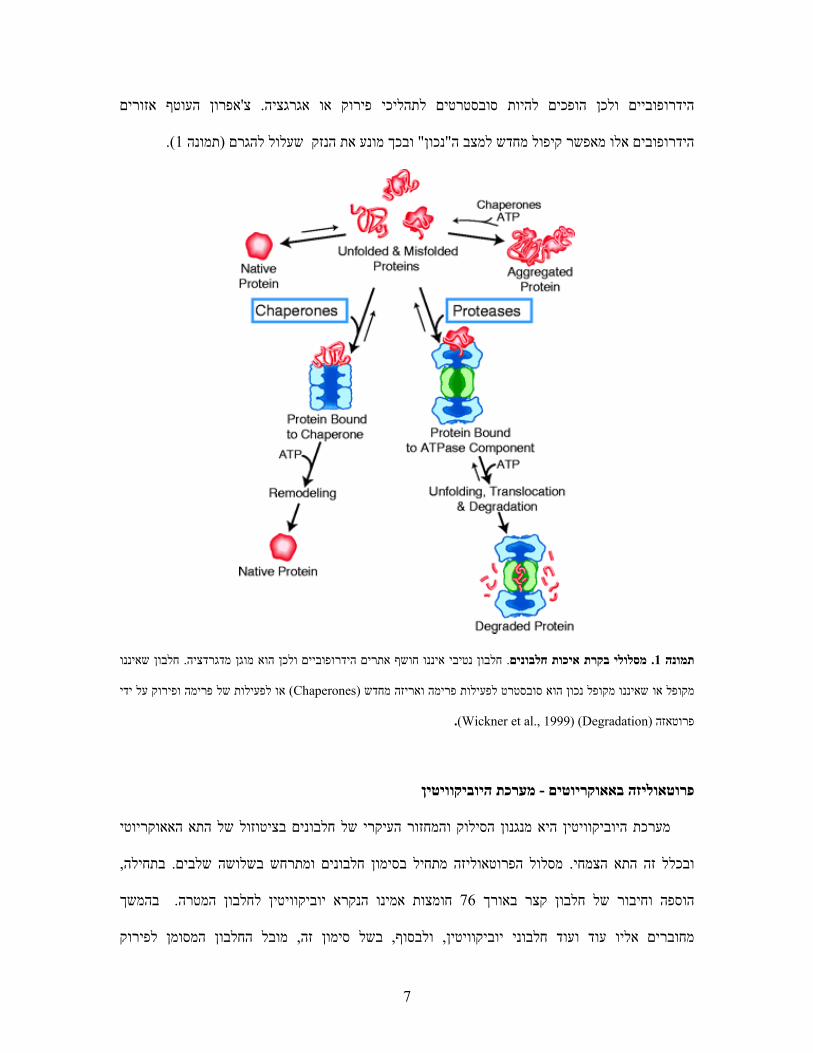
או בתור פרוטאזה כשהן : פרוטאזות כאלה עשויות לפעול באחד משני מסלולים. ATPפרוטאזות תלויות
כשהן פורמות את מבנהו המרחבי של חלבון (chaperone)אפרון'או בתור צ, מפרקות את חלבון המטרה
חושפים אתרים , ים שאינם ארוזים או מקופלים נכוןחלבונ. (folding)המטרה כדי לאפשר לו להתקפל מחדש
7
עוטף אזוריםהאפרון ' צ.ציהסובסטרטים לתהליכי פירוק או אגרגהופכים להיות הידרופוביים ולכן
).1תמונה ( להגרם ובכך מונע את הנזק שעלול" נכון"חדש למצב המאפשר קיפול מ אלוהידרופובים
חלבון שאיננו . חלבון נטיבי איננו חושף אתרים הידרופוביים ולכן הוא מוגן מדגרדציה. מסלולי בקרת איכות חלבונים. 1תמונה
ל ידי לפעילות של פרימה ופירוק ע או(Chaperones)מקופל או שאיננו מקופל נכון הוא סובסטרט לפעילות פרימה ואריזה מחדש
.)Wickner et al., 1999( (Degradation)פרוטאזה
מערכת היוביקוויטין - פרוטאוליזה באאוקריוטים
מנגנון הסילוק והמחזור העיקרי של חלבונים בציטוזול של התא האאוקריוטי מערכת היוביקוויטין היא
, בתחילה. מסלול הפרוטאוליזה מתחיל בסימון חלבונים ומתרחש בשלושה שלבים. ובכלל זה התא הצמחי
בהמשך . לחלבון המטרה חומצות אמינו הנקרא יוביקוויטין76הוספה וחיבור של חלבון קצר באורך
מובל החלבון המסומן לפירוק , בשל סימון זה, ולבסוף, מחוברים אליו עוד ועוד חלבוני יוביקוויטין

8
, הגרעין, הציטופלסמה: תהליך היוביקווטיזציה שתואר עשוי להתרחש במספר מדורים. 26Sבפרוטאוזום
Hershko and) שופות לציטוזול ואף ברשתית האנדופלסמטית שטח הפנים של ממברנות הח
Ciechanover, 1998). בתלויהוא תהליך רב שלבי ה היוביקוויטינזציהתהליך -ATP ומתחיל באנזים E1
לבין AMPפוספואנהידריד בין -על ידי זרוז של הראקציה היוצרת קשר אצטיל, קוויטיןיהמאקטב את היוב
קושר באופן ישיר את היוביקוויטין E1. הקבוצה הקרבוקסילית בגליצין שבקצה הקרבוקסלי של היובקוויטין
אנזים זה מזהה באופן ספציפי את .E1אסטר שנוצר בין הגליצין של היובקוויטין והציסטאין של -בקשר תיאול
אנזים קוניוגציה " נקרא גם - E2.ולרוב ניתן למצוא עותק מקודד אחד או שניים שלו בגנום, היוביקוויטין
אנזימים . רבעים אנזימים כאלהבארבידופסיס למשל ישנם כא. מופיע בתא במספרים גדולים" ליוביקוויטין
או שהם -E3 ומעבירים אותו לE1אלו קושרים באופן ישיר את היוביקוויטין שעבר אקטיבציה על ידי
הקבוצה . כדי לאפשר לראשון להעביר לשני את היוביקוויטין המאוקטבE3- וE1קושרים את האנזימים
טין יקוויאלה מחברים את היוב. וויטין ליגאז הנקראים יוביקE3 אנזימי 700- כוללת כ, האחרונה והגדולה
תהליך . )Smalle and Vierstra, 2004(לחלבון המטרה והם הקובעים את הספציפיות לסובסטרט
ת ארוכה של חלבוני קוויטין אחד למשנהו לשם קבלת שרשריהיוביקוויטנציה עשוי להמשיך תוך חיבור יוב
במקביל עשויה להתרחש גם הוספה של עוד ועוד חלבוני . יוביקוויטיניזציה- נקרא פוליהתהליך ,יובקוויטין
.יובקוויטין לסובסרט שמובל בסופו של התהליך לפירוק בפרוטאוזום
הפרוטאוזום כמערכת מבוקרת
-kDa 2000 זהו קומפלקס ענק של כ.בגלל מקדם השקיעה שלו, פרוטאוזום26S נקרא גם -הפרוטאוזום
יחידות המאורגנות בשני -ומפלקס זה בנוי משלושים ואחת תתק. לפעילותו הפרוטאוליטיתATP-והוא תלוי ב
והשני הוא )CP( roteasePore Cהראשון הוא לב הפרוטאזה ). 2תמונה (S19- והS20-ה, קומפלקסים-תת
בנוי מארבע טבעות זו על זו היוצרות חלל מרכזי CP-ה. )article Pegulatory R )RPהאתר הרגולטורי
שתי הטבעות הקיצוניות , יחידות-כל טבעת בנויה משבע תת. משותף שאליו נחשפים האתרים הפעילים
נוצר האתר הפעיל בקומפלקס. βיחידות- ושתי הטבעות שבמרכז בנויות מתתיαיחידות -ת מתתיומורכב

9
לא מאפשר αהיחידות -הפתח הצר הקיים בתתי. לפי חלל מרכזי אחד הפונות כβ1 ,β2 ,β5היחידות -מתתי
דות אלו מהווה שער לכניסת ייח-גמישות הקצה האמיני של תתי. כניסת חלבונים אלא במצבם הפרום
הוא מסוגל , נמצא באסוציאציה עם קצה אחד או שניים של לב הפרוטאוזום וממוקם עליוRP-ה. החלבונים
על מנת לפרום את ATP-ולהשתמש באנרגיה מ, באתר החיבור עם היוביקוויטיןלהקשר ספציפית לסובסרט
תת יחידות הבונות 17-והוא בנוי מ -AAA הוא משתייך למשפחת ה. להכניסו למרכז הפרוטאוזוםוהסובסרט
.(Base)והבסיס הטבעתי היושב מעל לב הפרוטאוזום , (Lid)המכסה : שני רכיבים
אירגון הקומפלקס הרגולטורי core protease (CP) .(B) אירגון לב הפרוטאוזום 26S .(A) של מבנה ואירגון. 2תמונה
regulatory particle (RP),הבסיס והשער-בצהוב, הזרוע המבקרת את הכניסה– באדום .(C) מציג את המבנה הכולל ואת
.)Vierstra, 2003( פירוק הסובסטרטשרשרת האירועים המביאים ל
למרות שהפרוטאוזום והסובסטרטים המיועדים לפירוק נמצאים לרוב יחד במדור המסיס של התא
המבנה המורכב של הפרוטאוזום והעובדה כי אתריו הפעילים אינם חשופים לסביבה מונעים, )הציטוזול(
מערכת כזו נקראת . פירוק אקראי ומחייבים כניסת הסובסרט אל תוך הקומפלקס
self-compartmentalizing protease )Lupas et al., 1997( . מערכת זו שונה ממערכת אחרת לפירוק
ם הפרוטאזות הן פרוטאזות פעילות במערכת הליזוזו. הליזוזום–חלבונים המתקיימת בתוך גופיף פירוק
(A) 20S CP
(B) 20S RP
(C) 26S proteasom

10
מנוצלת על מנת לפרום את ATP האנרגיה מפירוק ,בפרוטאוזום. והפירוק מבוקר בשלב ההכנסה לאברון
החל , במערכת זו קיימת בקרה במספר שלבים. ולא לשם הפירוק הפרוטאוליטי(unfolding)חלבוני המטרה
- היא גם מהווה את מרכיב המחזור העיקרי לחומצות. משלבי הסימון ביוביקוויטין ועד הפירוק בפרוטאוזום
.האמינו שמשמשות אבני בניין לכלל החלבונים בתא
AAA-משפחת ה
הם משפחה )ctivitiesAious cellular varssociated with ATPase family A )AAA- חלבוני ה
חומצות אמינו 230- מבנה זה כולל רצף של כ.גדולה של חלבונים המוגדרים על פי המבנה הראשוני שלהם
הם משתתפים בפעילויות רבות . ATP-כלומר פעילות החלבונים הללו תלויה ב, ATPaseבעל פעילות של
חלבונים השייכים . (Ogura and Wilkinson, 2001) חילוף חומרים ופרוטאוליזה, בתא כגון חלוקות
פעילה בתאים Clpהפרוטאזה , לדוגמה. למשפחה זו ניתן למצוא הן באאוקריוטים והן בפרוקריוטים
שהן חלבונים ClpX או ClpAיחידות רגולטוריות -מחובר עם תת) הפרוטאזה (ClpPפרוקריוטים כאשר
גם במערכת הפרוטאוזום . )Porankiewicz et al., 1999 ( השייכים למשפחה ATPaseבעלי פעילות של
השייכים למשפחה זו ATPמבוקרת הכניסה או הפתיחה של קומפלקס הפירוק באמצעות חלבונים תלויי
זו משפחה .AAA proteasesחלק נוסף במשפחה הוא קבוצת חלבונים ייחודית הנקראת . Lid-ובונים את ה
. רוטאוליטי על גבי אותו הרצף ואתר פATPase בעלי אתר ATPהכוללת בתוכה קבוצת חלבונים תלויי
הם עשויים . במיטוכונדריה ובכלורופלסט, פעילותם כוללת בקרת פירוקם של חלבונים ממברנלים בבקטריות
אפרון והן כפרוטאזה והם למעשה מערכת בקרת האיכות של החלבונים ובעיקר של חלבונים 'לפעול הן כצ
משפחת . משפחה בפני עצמה- הן משפחה או תתAAA-הפרוטאזות השייכות ל .(Langer, 2000) ממברנלים
. ממברנה- גדולים המעוגנים לממברנות באמצעות רצפים חוצימתארגנת בקומפלקסים AAA proteases-ה
לכוון הקצה הקרבוקסילי והוא אתר ATPase-לחלבונים הללו יש מוטיב אופייני הממוקם אחרי ה, בנוסף
יחידות במבנה דמוי טבעת -המבנה הפעיל שלהם הנו קומפלקס הבנוי משש תת). וטאזהמטלופר(הפרוטאזה
)הטרוהקסמר(יחידות שונות - או בנוי מתת,)הומוהקסמר(יחידה -שעשוי להיות בנוי כולו מאותה תת

11
)Ogura and Wilkinson, 2001( . פרוטאזתLon השייכת למשפחה AAA+ היא פרוטאזה מסיסה
היא חסרה את המוטיב AAAאך שלא בדומה לפרוטאזות . פרוטאז- פרוקריוטית השייכת לקבוצת הסרין
הוא גורם החיוני שATPצעות היא מבוקרת באמ, יחד עם זאת. המעגן לממברנה ולפיכך היא איננה ממברנלית
היא נמצאה מעורבת בפירוק של חלבונים רבים בתאי חיידקים. )Chung et al., 1983(לפעילותה
)Rotanova et al., 2004(.
מערכות פרוטאוליטיות בפרוקריוטים
ATP וחלקן תלויות DegP כגון פרוטאזת ATPחלקן אינן תלויות , בפרוקריוטים נמצאו מספר פרוטאזות
הנה בעובדה שחרף היות תהליך ATP-ת התלויות בחשיבותן הגדולה של הפרוטאזו. FtsH, Lon, Clpכגון
התהליך מבוקר באמצעות קישור או פירוק של , )איננו תלוי אנרגיה(ספונטני , הפרוטאוליזה תהליך אקזוטרמי
ATP ,כלל הפרוטאזות התלויות ב. כלומר הפירוק אינו אקראי אלא מבוקר-ATP ה שייכות למשפחת-
AAA+ .באפופטוזיס וכן בבקרת , בבקרת מחזור בתא, ות לעקות בחיידקיםהן מעורבות בבקרת מגוון תגוב
.)Jenal and Hengge-Aronis, 2003( תגובות של הולכת סיגנל או הפסקתו
פרוטאז-אתר הסרין וATPase-האתר . AAA- ה למשפחת שייכתה היא סרין פרוטאזClpפרוטאזת
מערכת מודל יתהוה, E. coli - לראשונה ב התגלתהClpפרוטאזת . שני פוליפפטידים נפרדיםבנמצאים
רוטאזת מבנה פ .הפרוטאזה אינה חיונית בחיידקים. הקשר בין מבנה לתפקוד במערכות פרוטאוליטיותבחינתל
Clp 26-ה דומה למבנהS יחידה - תת: אוליגומריות- יחידות הומו-הפרוטאזה מורכבת משתי תת .פרוטאוזום
היחידה - תת. ATPaseבעלת פעילות ) ClpA(יחידה רגולטורית - ותת )ClpP(קטליטית מטיפוס סרין
יחידה הרגולטורית ה-תת. ClpPמשבע יחידות , כל אחת בנפרד, המורכבות, הקטליטית מורכבת משתי טבעות
קישור של תת היחידות הרגולטורית והקטליתית האחת .ClpA מורכבת משש יחידות שלבנויה טבעת יחידה ה
שייכת Lon פרוטאזת .)ATP- )Laskowska et al., 1996 בנוכחות סובסטרט ורקאל השניה מתרחש
רו על פעילות שמ Lon- מוטנטים ב חיידקים. נמצאת כפרוטאזה מסיסה בתאי החיידק+AAAלמשפחת
אינה משפיעה על , Lon ו Clpמוטציה כפולה בגנים המקודדים לפרוטאזות פרוטאוליתית כללית ואפילו

12
Hwang et al., 1988; Katayama et al., 1988; Katayama et(ק בתנאים נורמליים של החיידגדילתו
al., 1990(.שתי פרוטאזות אלו מוגדרות כלא חיוניות .
בחיידקיםFtsHפרוטאזת
FtsH ככזאת שלא איפשרה לחיידקים להתקיים מוטציה שזוהתה בחיידקים הקרוי על שםהוא חלבון
ובעל פעילות , הוא קשור לממברנה .Filamentation Temperature-Sensitive H: בטמפרטורות גבוהות
צורתו הפעילה היא קומפלקס הכולל . AAA-proteasesוהוא שייך למשפחת טאז ומטאלופרוATPaseשל
ניתן למצוא אותו בממברנות הפלסמה של חיידקים ובאורגנלות של תאים אאוקריוטים . שש תת יחידות
)Ogura and Wilkinson, 2001( ,בחיידק E. coli הוא הפרוטאזה תלויית הATP-החיונית היחידה .
FtsHזוהה לראשונה בתצפיות בחיידקים שערכו Santos and De Almeida , הם מצאו חיידק . 1975בשנת
: מוטנט זה נשא בפועל שתי מוטציות. הפגוע ביכולת החלוקה שלו בטמפרטורות גבוהות כתוצאה ממוטציה
.)FtsI )Ogura et al., 1991-ב והשניה FtsH-האחת ב
FtsH ב- E. coli בעל משקל מולקולרי של חומצות אמינו היוצרות חלבון647- גן יחיד המקודד להוא
71 kDa , כארבע מאות עותקים של חלבוןFtsH קיימים בתא נורמלי של החיידק)Tomoyasu et al.,
Walker A: ה מוטיבים שמוריםלושב מש וזה מורכATPase-מבנה החלבון כולל את מוטיב ה. )1993
(GPPGTGKT) ו - (IIFIDEID) B ומוטיב נוסף SRH (TN9XRXGR2XR) )Neuwald et al.,
1999( .
FtsH ניתן לזהות ברצף מוטיב בו, עדויות לכך ניתן למצוא ברצף חומצות האמינו. הנו מטלופרוטאזה
חומצת וHEXGHהוא כולל את הרצף . וי האבץ של המטלופרוטאזהייחודי המהווה את האתר הפעיל תל
א ליגאנד ישה, E479, שים חומצות אמינו במורד הרצף לכוון הקצה הקרבוקסלייכשת הנמצא, פת נוסאמינו
סמוך לקצה , יש שני רצפים חוצי ממברנה בתחילת הרצףFtsH-ל. )Saikawa et al., 2002(שלישי לאבץ
- אתר ה. מית של החיידק תוך חציית הממברנה פעמייםרצפים אלו מעגנים אותו לממברנה הפני. האמיני
ATPaseוהוא מעיד על מבנה פעיל בקומפלקס המכיל שש תת יחידות תוך , גובש ומבנהו השלישוני פוענח

13
על , למעשה. )ATP )Kaltwasser et al., 2002- של ה(γ) עם הפוספט השלישי SRH-אינטראקציה של ה
מחקר .)Akiyama and Ito, 2001(חייב להיות לחלבון מבנה של הומואוליגומר , ATP-מנת לפרק את ה
Shotland( מתארגן בתצורה של טבעת FtsH-שנערך באמצעות מיקרוסקופ אלקטרוני הראה כי קומפלקס ה
et al., 1997( . FtsH בדטרגנט ולקבלו כקומפלקס מסיס גדולאך ניתן להמיס אותו , הנו חלבון ממברנלי ,
Akiyama et al., 1995; Akiyama) שש תת יחידותשל הוא בגודל , אשר לפי הנדידה בגרדיאנט סוכרוז
and Ito, 2001) .סטרטיםפעילותו הפרוטאוליטית נלמדה ונמצאו לו מגוון סוב :σ32 , פאקטור השייך
האחראים למעבר CIII- וCII כמו λ' חלבונים הקשורים למסלולי התגובה עם פאז, למערכת התגובה לחום
Ito and( וכן סובסרטים מהונדסים שונים YccAובאופן ייחודי חלבונים ממברנלים כמו , ליזוגניהלשלב
Akiyama, 2005(. ישנן עדיות לאינטקראקציה ביןFtsH לקומפלקס נוסף HflC-HflK , העשוי להקשר
הקשר עצמו חוסם את הגישה אל האתרים הפעילים . ובסטרטים ולשמש מערכת בקרה נוספתבאופן ישיר לס
Kihara(אפשרת דגרדציה מHflC-HflK יפרדותו של תוך הפרעה אלוסטרית לסובסרט ורק הFtsHשל
et al., 1997(. לסקירה נרחבת ראה גם Ito and Akiyama, 2005)(.
FtsHבמיטוכונדריות
. מיוצרים בציטוזול ומועברים במצב פרוס דרך הממברנות אל תוך האורגנלה רוב חלבוני המיטוכונדריה
מחייב , כמו גם קיומם של חלק גדול מהתהליכים המטאבולים על גבי ובתוך הממברנה, המעבר דרך הממברנה
את פרוהיביטין ואת , ABCמערכת זו כוללת בממברנה הפנימית את הנשא . כי בקרת איכות קפדנייםתהלי
.-AAAפרוטאזות
i-AAAאו m-AAA שבמיטוכונדריה נקרא בשמרים FtsH . רוב המידע הקיים נצבר ממחקרים בשמרים
inter membrane) הבין ממברנלי חללעבור ה (i), בהתאם למדור אליו נחשפים האתרים הקטליטים
space) ,ו-)m( עבור חלל המטריקס (matrix) .פרוטאזות-AAAהן קומפלקסים גדולים במיטוכונדריה
Leonhard et al., 1999) Arlt et al., 1996; ( .m-AAA יחידות- בנוי משתי תת, Yta10ו Yta12- ,
. -kDa 2000הבונות הטרואוליגומר בעל משקל מולקולרי של כ, ומטרים זהיםיככנראה ביחסים סטוי

14
Yme1 , לעומת זאת בנוי מתת יחידה אחתi-AAA-ה. קומפלקס זה כולל ככל הנראה גם חלבונים אחרים
(Yta11),1000 במשקל של כ המרכיבה הומואוליגומר kDa- .יחידה זו חוצה את - המונומר של תת
יחד עם זאת כל שאר המוטיבים . m-AAA- או בE. coli-ם כמו ביהממברנה פעם אחת בלבד ולא פעמי
של בעיות אינאקטיבציה של פרוטאזות אלו מביאה לפנוטיפמוטציות הגורמות ל. שהוזכרו שמורים גם כאן
ובין m-AAAישנן עדויות ליצירת קומפלקס משותף בין . )Paul and Tzagoloff, 1995( חמורות בנשימה
i-AAA- וSteglich et al., 1999( .m-AAA( שלהם על מנגנון בקרה משותףדבר המעיד , פרוהיביטין
אלו אינן ארוזות יחידות -כאשר תת, יחידות של קומפלקס הציטוכרומים- תתינמצאו מעורבים בפירוק
. (Weber et al., 1996) בקומפלקס
ורטולוג הומני שלא .)Arlt et al., 1996(אפרונית של החלבונים הללו 'יות לפעילות צוות גם עדקיימ
Yme1) המונומר שלi-AAA ( זוהה בניסוים של קומפלימנטציה)Shah et al., 2000( . של אובדן מוטציה
(paraplegia) בגן זה מלווה במחלות נויורולוגיות סופניות כגון פארפלאגוס (loss of function)פעילות
בכל האורגניזמים שהגנום שלהם מפוענח . )Casari et al., 1998( בפלג גוף תחתוןהמביאה לשיתוקים
.קיימים הומולוגים לגנים אלו
FtsHבציאנובקטריה
ישנה חשיבות רבה בהבנת התהליכים הקשורים ,הנחשבת לאב הקדמון של הכלורופלסט, טריהלציאנובק
בדומה לכלורופלסט יש בה שתי מערכות אור .)Blankenship and Hartman, 1998(לכלורופלסט הצמחי
התהליכים הבסיסיים בשניהם . ATP- וNADPHהאוספות את האור ומתמירות אותו לאנרגיה כימית בצורת
קיימים ביניהם הבדלים מהותיים כמו , למרות זאת. דומים וכך גם הקומפלקסים המעורבים בתהליכים אלה
לעומת קידוד הגנים הכלורופלסטיים ,תא זה- הגנים בחדלמשל הרכב פיגמנטים וקידוד עצמאי של כלל
FtsH- בציאנובקטריה ישנם ארבעה גנים ל. שמתבצע בעיקר בגרעין ביצורים פוטוסינתטיים מתקדמים יותר
Synechocystis sp. PCC 6803 )Nakamuraאלה זוהו לראשונה עם פרסום הגנום של .1-4הממוספרים
et al., 1998(. מוטציות השתקה של שניים מהגנים הללו ,FtsH1ו -FtsH3 , הביאה לפנוטיפ סופני

15
)Lethal .(מוטציה ב-FtsH2ה במוטצי הביאה למופע של השתנות הפיגמנטציה ול-FtsH4 לא זוהה כל
העלה כי השתנות זו נגרמת כתוצאה FtsH2-בחקר מעמיק של המוטנט .)Mann et al., 2000(פנוטיפ
אך , שאר רכיבי המערכת הפוטוסינתטית כמו גם פיגמנטים אחרים לא השתנו. 60%- בכPSIמצימצום
עובדה זו מעידה על מעורבות בתיקון .)Mann et al., 2000(התאים היו רגישים לעוצמות אור גבוהות
מחקרים מאוחרים יותר הראו מעורבות בהגנה על המערכת בשלבים . המערכת הפוטוסינתטית ובהגנה עליה
חלק ממערכת התיקון של כוזאת , D1מוקדמים של עקת אור באמצעות דגרדציה של חלבון מרכז הראקציה
PSII יים כמו קלאטורים של מתכות אשר עשויה להיות מעוכבת באמצעים פרמקולוג)Silva et al., 2003(.
מבנה ופעילות - הכלורופלסט הצמחי
לכלורפלסט מבנה אנטומי ייחודי הכולל . תהליך הפוטוסינתזה בו מתרחשנו אברוןיהכלורופלסט הצמחי ה
ממברנת המעטפת הפנימית , ממברנת המעטפת החיצונית: שלושה ממברנלים ושלושה מסיסים, שישה מדורים
החלל הבין ממברנלי התחום על ידי ממברנות .יםליוממברנות התילקואיד הן שלושת המדורים הממברנ
הסטרומה התחומה בממברנת המעטפת הפנימית והתילקואידים וחלל הלומן הפנימי התחום , המעטפת
:זה נהוג לחלק לשתי ראקציותינתאת תהליך הפוטוס. בממברנת התילקואיד הם שלושת המדורים המסיסים
. ריכוז האנרגיה שנקלטה והתמרתה, ור באנטנות פוטוסינתטיות ראקציית האור כוללת קליטת א.האור והחושך
תוך בניית גרדיאנט פרוטונים משני צידי ממברנת , NADPH- ל+NADPהאנרגיה משמשת לחיזור
;ATP )Butler and Kitajima, 1975מניע לייצור -גרדיאנט זה משמש ככוח פרוטון. התילקואיד
Kitajima and Butler, 1975( .ארבעה קומפלקסים . ראקציית האור מתרחשת בממברנות התילקואיד
מערכות ): כולם אוליגומרים המכילים לפחות שבע תת יחידות(ם ממברנליים משתתפים בתהליך אנזימטיי
תהליך הפוטוסינתזה מתחיל בקליטת . -ATPפלקס יייצור הוקומ, קומפלקס הציטוכרומים, תייםהאור אחת וש
-מצוןהאנרגיה הנקלטת משמשת להעברת האלקטרונים מהמים בעלי פוטנציאל ח, האור על ידי הכלורופיל
משמשים במעגל קלווין לקיבוע -ATP וNADPH. בעל הפוטנציאל הגבוה יותר-+NADPחיזור נמוך ל
. המתרחש בסטרומה תהליך ,)ריאקציית החושך(פחמן אטמוספרי וליייצור פחמימות

16
. מסך חלבוני הכלורופלסט-5%פחות מלגנום עצמאי אך הוא מקודד , בדומה למיטוכנדריה, לכלורופלסט
Cline and Henry, 1996; Keegstra(זים בציטוזול תרוב הגדול של החלבונים מקודדים בגרעין ומסונה
and Cline, 1999( .ל מכיוון שעיקר הסינתזה מבוצע בציטוזול מתקיימת בתא מערכת יבוא חלבונים א
שבקצהו האמיני תוספת של (Precursor)תהליך זה מתחיל בסינתוז של חלבון ארוך יותר . הכלורופלסט
חלבון מיועד אם ה.Transit/Signal Peptide (T/SP)פפטיד מוביל בן כמה עשרות חומצות אמינו הנקרא
, )Cline and Henry, 1996( כפול בקצהו האמיניTPהוא מכיל , תילקואידללומן ה, למדור הפנימי ביותר
חלבון זה יקוצץ בשני .)OEE33 )Ko and Cashmore, 1989כמו למשל במקרה של חלבון פירוק המים
והשני ,SPP (Richter and Lamppa, 1999)הראשון עם הכניסה לכלורופלסט על ידי פפטידאזת: שלבים
.TPP (Chaal et al., 1998) על די לומןעם הגעתו ל
פירוק חלבונים בכלורופלסט
פגמו חלבונים אשר נ, פרוטאוליזה בכלורופלסט כוללת פירוק של חלבונים אשר אינם מקופלים כראוי
וחלבונים אשר תהליכי ויסות התפתחותיים ליעדם הגיעו לאחלבונים אשר, עקב נזקי קרינה או נזקים אחרים
מחד , כמות החלבונים הקיימת בנקודת זמן מסויימת מוכתבת משני גורמים .וסביבתיים הכתיבו את פירוקם
חלבוני הכלורופלסט הופכים יציבים ועמידים בפני . )Adam, 1996(ומאידך הפירוק , היייצור והצבירה
רוביסקו בתאי שעורה נשאר יציב למשך שבוע בחושך , לדוגמה. הפירוק מיד עם צבירתם ואריזתם הנכונ
)Huffaker and Peterson, 1974( .במידה שמעוכב ביטויה של אחת מתת היחידות של , יחד עם זאת
. ות מערכת בקרת האיכותהיחידה השניה איננה נצברת ומפורקת ככל הנראה באמצע-תת, רוביסקו
ריכוז האנרגיה והתמרתה לצורך , בכלורופלסט בריאקציית האור מתרחשים תהליכי איסוף אנרגיית אור
ערור הכלורופיל בשלב הראשון של הראקציה עשוי . יצירת פוטנציאל ממברנה באמצעות תהליכי חמצון חיזור
מצב זה איננו ). ם החמצן נמצא באורביטל שאינו סימטריהאלקטרון באטו(להוביל אותו למצב הטריפלטי שלו
חמצן זה הנו רדיקל חופשי ). חסר אלקטרון באורביטל יחיד(סינגלט -יציב ועלול להוביל ליצירה של חמצן

17
או , ) יצירת קשר עם מיםל ידיע(אשר עלול לגרום ליצירה של רדיקלים חופשיים אחרים כמו פרואקסיד
D1 חמצון החלבון . PSII ממרכז הרקציה שלD1 אחרים בסביבתו כגון חלבון לחמצן באופן ישיר גורמים
בשלב זה תחל מערכת התיקון או הפירוק לפעול ותסלק . מוביל לשינוי קונפורמטיבי ולפגיעה בתפקוד החלבון
סביבה שרדיקלים חופשיים נוצרים בה באופן. )Jansen et al., 1999(את החלבונים הפגומים מהממברנה
את החלבונים שבסביבתם ולפגוע מצןדיקלים אינם יציבים ונוטים לחקבוע תחשב כסביבה מסוכנת שכן הר
נמצא כי בשלבים ראשונים של עקת אור חלים בו שינויים . הוא חלבון שנחקר באופן מעמיקD1 . בפעילותם
על מנת לאפשר . )Zer et al., 1994(ידי פוליפפטיד שסונטז מחדש - עלעקב כך הוא מפורק ומוחלף , מבניים
במערכת מתבצעים . 'מערכת התיקון'ת מערכת שלמה הנקראת התקין של תהליך הפוטוסינתזה קיימאת קיומו
סילוק , הוצאה, למלה- למלה אל הסטרומה-הזזה מהגרנה, רמטיביוזיהוי השינוי הקונפ: מספר תהליכים ברצף
בכל אחד מהשלבים מעורבים . למקום הנכון בממברנהזה מחדש והכנסה נתסי, ופירוק של החלבון הפגוע
המערכת . היא מתחילה לפעול תוך דקות ספורות. מערכת זו פועלת במהירות רבה ביותר. מספר חלבונים
מערכת .)Melis, 1999( עשויה לפעול גם בשטפי אור בעוצמה נמוכה ולא רק בשטפים בעוצמות גבוהות
תפקיד חשוב בשמירה על המאזן מבצעות ATPפרוטאזות תלויות כאשר , התיקון מורכבת גם מפרוטאזות
בהעדרו וATP מבוקרים באמצעות פירוק של בפרוטאזות אלהפירוקה תהליכי .מערכת מבוקרתועל התאי
.מזעריתהפעילות הפרוטאוליטית
,Deg, Clp:םיזות אשר לכולן הומולוגים בקטריאליקריות של פרוטאי בכלורופלסט יש שלוש משפחות ע
היא הפרוטאזה הנחקרת ביותר בכלורופלסט Clp . מספר פפטידאזותפועלות בכלורופלסטבנוסף .FtsH-ו
. (Shanklin et al., 1995)וכבר לפני עשור נמצאה כי היא פעילה בקומפלקס הדומה לקומפלקס החיידקי
Clp בחיידקיםהומולוגים יחידות אשר לכולן יש - פרוטאז המורכב מכמה תת-הוא סרין (Shanklin et al.,
1995; Shikanai et al., 2001). Clp תלויה בATP- שלה למבנה צבירת תת היחידותל גםלפעילותה ו
בכלורופלסט איננה חיוניתClpבעבר חשבו כי בדומה לחיידקים . )Halperin et al., 2001(הפעיל
אך עם התקדמות הטכניקות הגנומיות .)Cahoon et al., 2003( ומוטנטים החסרים אותה מצלחים להתקיים
החיונית להתפתחות הנצר בצמחי טבק ClpP1 הן יחידות החיוניות כמו למשל -Clpהתברר כי חלק מה

18
(Kuroda and Maliga, 2003) .Clp חלבונים שונים 20- היא משפחה של פרוטאזות הכוללת למעלה מ
. (Clarke et al., 2005) המשתתפים במגוון תהליכים
גם היא סרין פרוטאזה . )Itzhaki et al., 1998(שובטה וזוהתה בכלורופלסט , נמצאה DegPפרוטאזת
יות על מעורבות אפשרית ו היא משפחה גדולה של פרוטאזות וישנן עדATP . DegP-אך היא איננה תלוייה ב
. )Haussuhl et al., 2001; Nixon et al., 2005( כתוצאה מנזקי קרינהD1בחיתוך הראשוני של
FtsHבצמחים
מצמחי FtsH. בכלורופלסט של הצמחFtsHמאמצע שנות התשעים החל להצטבר מידע לגבי נוכחות של
. ותגובה עם נוגדנים כנגדו-E. coli מושובט על בסיס דמיון לחלבון , בודד, וארבידופסיס אופיין, תרד, אפונה
מחיידקים עם פרקציות שונות מצמחי FtsHבשלב הראשון נערכו ניסויים ביוכימיים בהם הוגבו נוגדנים כנגד
החלבון פרקציונציה נוספת של הכלורופלסט העלתה כי . תרד ואפונה והתקבלה תגובה עם כלורופלסטים
תוך ,חלבון אינטגרלי לממברנת התילקואידוא הטים אךת אינו קשור עם הקומפלקסים הפוטוסינשהגיב
ר מעיד כי הוא עשוי להיות מעורב בפירוק של חלבונים הדב. לסטרומהATPase-אתר הפרוטאזה וה הפניית
עורב עשוי להיות מ FtsH הראו כי in-vitroניסויי . )Lindahl et al., 1996(ממברנלים ומסיסים כאחד
.)Ostersetzer and Adam, 1997( מהמערכת הפוטוסינתטיתRieske FeS (RISP)בפירוק של חלבון
נמצא importבניסויי . ניתן היה לעקוב אחר כניסתו לכלורופלסט, סומן רדיאואקטיביתRISPכאשר החלבון
קציות המסומן עם פרחלבוןההשריית . א חשוף לדגרדציהפלקס והומכי חלק מהחלבון איננו מתארגן בקו
עוכב הזה הפירוק . שונות של הכלורופלסט העלתה כי עיקר הפירוק מתבצע בפרקציית התילקואידים
עדות ראשונה למעורבות היה הדבר .)FtsH )Ostersetzer and Adam, 1997 נוגדנים כנגד הוסיפוכש
-ורב בפירוק חלבון מעFtsH כיהמשך נמצאב. ם בצמח ובכלורופלסטיאפשרית בבקרת תהליכים פיזיולוגי
1D2- של מרכז הראקציה הפוטוסינתטית במערכת אור (PSII) (Lindahl et al., 2000). כאשר נבחנו
מתפרק כאשר הוא מושרה עם , 23kDa, נמצא כי הפרגמנט הגדולD1תוצרי החיתוך הראשוני של חלבון
ניסוי . ת אבץערכיים כדוגמו ומחייב נוכחות של יונים ד ATPפרוק זה נמצא כתלוי, תילקואידים בחושך

19
הפרגמנט במשקל אתק לפר ו בחיידקים ולבחון את פעילות החלבון המנוקה הצביע על יכולתFtsHלבטא את
23kDa .הציעו כי להכל אל FtsH-טיות ת תפקיד חשוב במערכת התיקון של המערכות הפוטוסנ)Lindahl
et al., 2000( .במהלכה נוסף המידע הבא. יה המידע שנצבר עד לתחילת עבודתיהמידע שסקרתי ה :
FtsHחלבוני 12- להמקודדים, היא משפחה של גנים FtsH . עיקר העדיות לכך נצבר עם פענוח הגנום של
התוצרים של תשעה מהם . )Adam et al., 2001(הארבידופסיס והצלבת הנתונים עם מאגרי מידע אחרים
רק ,יחד עם זאת. (Sakamoto et al., 2003) עשויים להיות מוכוונים לכלורופלסט ושלושה למיטוכונדריה
האחת באמצעות , מימדיות שונות- בשתי מערכות הפרדה דוארבעה אכן נמצאו באנליזת חלבוני התילקואיד
.נקודה איזואלקטרית ומשקל מולקולרי והשניה על בסיס משקל מולקולרי של הקומפלקס ושל תת היחידות
,.FtsH1, FtsH2, FtsH5, FtsH8 (Sinvany-Villalobo et al., 2004; Yu et al : הארבעה הם
בצורה שווה FtsHית של החלבונים שהגיבו עם הנוגדן המגיב עם כלל האיזוזימים של בחינה כמות. (2004
FtsH5, כשאחריו בהתאמה FtsH2 ביותר הנו של הגבוההצבירהעור יהראו כי ש) SRH-מזהה את ה(
,FtsH8 FtsH1 )Sinvany-Villalobo et al., 2004(.
FtsH בעלי טבק נמצא כי הוא מעורב בהפעלת מערכת של מוות . מעורב במספר תהליכים פזיולוגים
עדויות לגבי מעורבותו . )Seo et al., 2000( בזמן הפעלת הגנה מפתוגנים (PCD)מתוכנן מראש של תאים
החלו להצטבר כאשר זוהה מוטנט בגן in-vivo הפוטוסינתזה במערכות מערכת שלPSII בתיקון FtsHשל
המוטנט נקרא . (variegation)עלים וגיוונם בלבן ובירוק לסרוגין ה שהראה מופע של הכתמת FtsH2-ל
VARII והמוטציה מופתה לגן FtsH2 .מות החלבון במוטנט זה היתה ירידה בכFtsH. אך בעוד שהאזורים
אלא פלסטידות ,באיזורים הלבנים לא נצפו כלורופלסטים כלל, הירוקים הכילו כלורופלסטים שנראו נורמלים
מוטנט זה רגיש לעוצמות .)Chen et al., 2000; Takechi et al., 2000(פרולמלה -שהכילו מבנים דמויי
של כלורופיל בזמן הארה ירידה בערכי הפלורסנציהבו קיימת ו ,(Photoinhibition)אור גבוהות
(Fv/Fm) . הדבר מעיד על פגיעה בפוטנציאל האלקטרוכימי שלPSII .המוטנט פגוע בכושר , יתרה מכך
מעורב במערכת FtsH2לו למסקנה כי כל אלו הובי. במרכז הראקציה הפוטוסינתטיתD1-הפירוק של חלבון
.)Bailey et al., 2002(אור - בתנאי עקתD1התיקון של חלבון

20
בדומה . )(FtsH5 Sakamoto et al., 2002-זוהה מוטנט בגן ל, במעבדתנונוצעיב במקביל לזיהוי ש
והוא נקרא לעתים נעלמה,אך זו היתה קלה בלבד, גם מוטנט זה הראה ואריגאציה בעליו-FtsH2למוטציה ב
VARI .יו במערכת ובידוד גן זה וביטin-vitroאיששו את הממצאים הקודמים והראו כי יש לו פעילות של
ATPase וגם יכולת פרוטאוליטית שהתבטאה בפירוק של סובסרט כללי - BSA . בחינת המוטנט הראתה כי
גם כאן . VARIIיותר מאשר צמחי הביקורת אך פחות מאשר המוטנט , הוא רגיש לעוצמות אור גבוהות
FtsHייתכן ש כלומר . שבאזורים הירוקיםבמקטעים הלבנים נמצאו פלסטידות במספר דומה לכלורופלסטים
נמצא מעוכב ftsh2 ftsh5מוטנט כפול של . אינו מעורב בחלוקת הפלסטידה אך הוא מעורב בהתמיינותה
הפרדה , ניסוי הפרדה על קולונות .)Sakamoto et al., 2002( בגידולו וחסר יכולת ליצור פרחים פוריים
משקעים האחד את השני ונמצאים -FtsH5ו FtsH2שקעה באמצעות נוגדנים הראו כי בגרדיאנט סוכרוז וה
תוצאות דומות התקבלו מהפרדת הקומפלקסים של . )(Sakamoto et al., 2003יחדיו בהטרוקומפלקס
FtsH באמצעות “green gels” (Yu et al., 2004) .נמצא כי ל, בניסויי קומפלימנטציה שבוצעו השנה-
FtsH1יכולת תיקון של הפנוטיפ הנגרם בהעדר , המבוטא ביתרFtsH5ול FtsH8- יכולת דומה כאשר חסר
FtsH2 אך לא כאשר חסר FtsH5 (Yu et al., 2005) .

21
מטרות המחקר
. וריצוף גנום הארבידופסיס עדיין לא הושלם, בלבד FtsH1בתחילת עבודה זו היה ברשותנו מידע על. 1
, בפירוק חלבונים במערכת הפוטוסינתטית FtsH1 לגבי מעורבותו שלin-vitroמאחר שהיו עדויות מנסויי
.ה של מוטציות השתקה על הצמח השלםמה תהיה ההשפע בחרנו לברר
והחלטנו , עם פירסום הגנום של הארבידופסיס והופעת מאמרים ראשונים על מוטנטים התרחבה השאלה. 2
מי משתתף במבנה הקומפלקס , FtsH-לברר מה תהיה ההשפעה של מוטציות השתקה של הגנים השונים ל
?בכלורופלסט ומה היחסים בין הגנים השונים במשפחה
FtsHתו של ותפתחותיים או הסביבתיים המשפיעים על ביטויו או פעילרצינו למצוא מהם התנאים הה. 3
.בצמח

22
חומרים ושיטות
יחומר צמח
) WT( 'זן הבר' שימשו Wassilewskija -ו Columbia מהזנים Arabidopsis thalianaצמחי
מצע גידול מסוג ב םיחה מבוקריצמחים אלו גודלו בתאי צמ . למוטנטים השונים'רקע'צמחי בתור לניסוים ו
.)יום קצר( 16/8 או )יום ארוך( 8/16חושך של - תחת משטרי אור,קוליטי ורמ50%שהכיל עד " כבול פיני"
CO22+2 טמפרטורה קבועה של בצמחים גודלו ה. מסה וגטטיביתרה פריחה ויצעהתקופות האור הקצרה מנ
. לצרכי הניסוי בהתאם µmol m-2 sec-1 50-150עוצמת האור בתאים הייתה . 70%לחות יחסית של עד ב
CO4 +2 של פרטורההכמנה בטמעברו לפני הנבטה ו, חדרה פרטורתאוחסנו בטמו, מלא יבוש זרעים יובשו
48-72לחות מרבית תוך עטיפת העציצים בניילונית למשך ב בתאי הגידול ונבטוההזרעים .שעות 48למשך
.שעות
טופלו הצמחים בקונפידור וניםכנגד זבוב. בעת הצורךרק בוצעו מחלות טיפולים כנגד מזיקים ו
או , ml/L 30 של בהגמעה"דליה-זהר"מחברת " תוש-ביו"או ב ml m-2 0.1 של בהגמעה "Bayer"מחברת
" אקוגן"מחברת " ספרול"-ו הצמחים בכנגד קמחונות וחלדונות רוסס ."אקוגן" מחברת 2%בריסוס מלתיון
מחברת " 20אופיגל " שעות ב48ד אקרית הקורים או אקרית אדומה אובקו הצמחים למשך כנג. 1%בריכוז
.בהשקיה 0.1% של בריכוז-20:20:20מסוג דישון בוצע אחת לחודש בדשן מורכב מלא ."כימגת"
בתנאים אספטיים וסלקציהגידול ,חיטוי זרעים
בגמר , לצורך חיטויאתנול 70% - בנשטפו ו3פר מסייר ווטמן נ על הונחוצמחים העים אשר נאספו מהזר
צלחות פטרי המכילות מצע בהזרעים נזרעו .ש במשך שעה בתוך מנדף למינריוהשטיפה הם הונחו לייב
0.5XMS, 1% MES-KOH pH 5.7, 3% Sucrose, 1% Agar לסלקציההאנטיביוטיקבמידת הצורך ו ,
)20mg/L Kanamycin( או(BASTA 20μM) . מחדר שהתקבלו מקווי-SALK מצע על גביגודלו

23
במצע המכיל Syngenta או מקווי מחדר של Wisconsinמחדר מאוניברסיטת הקווי . Kanamycinהמכיל
BASTA. סריקות לעמידות ל BASTA- 0.1%של ריסוס ישיר ב בוצעו גם BASTA שגודלו על צמחים
בנפח של סטרילים פלסטיק מיכלית פריחה גודלו ב מוטנטים כפולים שגודלו למטר.החל מגיל שבועיים בעציץ
.ל של מצע גידוml 180על גבי , שני ליטר
FtsH1-בסריקה וזיהוי המחדר
גן המקודד ל ב נסרקה לאיתור קווי מחדר Madison Wisconsinאוניברסיטת לש ספריית המוטנטים
FtsH1-.קווים מוטנטים72960- בספריה זו היו כ (Krysan et al., 1999) .ה-DNA ששימש לבניית
מכיל הכל עציץ , עציצים) קומות( או משורת עציצים או מטור עציצים או ממשטח :תוהספרייה אורגן בקבוצ
בסיום הסריקה מתקבלות שלוש קורדינאטות המובילות לעציץ .יוצג בכל אחת מהקבוצותמכעשרה צמחים
מר שני מקצה הגן יהחוצה ופרי המחדרמר אחד פועל מי כאשר פריPCR הסריקה מבוצעת באמצעות .יחיד
,TAIL-PCR (Thermal Asymmetric Interlaced) )Singer and Burke הטכניקה נקראת .פנימה
ומכל טור ) קומות76כ "בסה( הופק מכל קומה בנפרד DNA :ך אורגנו כ-DNAשלושת מאגרי ה. )2003
7296 צמחים ב 72960סך הכל יוצגו . ) שורות8כ "בסה(בנפרד מכל שורה ו ) טורים12כ "בסה(בנפרד
בכל קבוצה . כל קבוצהב קומות 12-13 המכילות "על"הצמחים חולקו לשש קבוצות .) 8*12*76 (עציצים
הסריקה . 1-12 והטורים במספור A-H השורות סומנו באותיות, מהשורות והטורים בנפרדDNAסודר
בשלב שני בוצעו . ) מבחנות38בסך הכל ( כל שתי קומות יחד נבדקוון בשלב ראש. בשני שלביםהבוצע
PCR התוצאות .)נפרדב הפעם ,י הקומות שאוחדותלש( ראקציות ועוד זוג ראקציות 12+8טורים והשורות ל
הספרייה יוצרה במקור .3 טור A שורה 28 מייצג קומה 28A3 : בשלוש קואורדינטות לדוגמהויוצג
ה עותקים של מכיל ארבע וpSKI015הנקרא ה(Weigel et al., 2000)נארי באמצעות פלסמיד בי
היה לייצר קוים המבטאים ביתר המנחה כאשר הרעיון ,)1תמונה ( בקרבת הגבול הימני 35Sהפרומוטור
T-DNA-ל מהקווים הם קווי השתקה מאחר וה חלק גדו, בפועל.חלבונים שונים בהתאם למיקום המחדר
תוכננו TAIL-PCRעל מנת לבצע סריקה בטכניקת .קודד או באורינטציה לא נכונהתוך אזור מלחדר

24
חומצות 22-29 ובאורך של 65oC לתנאים של FtsH1 לאזור המקודד לגן , גנומיDNA-פריימרים ל
:גרעין
tct cgt ttg tag ctt ctt ctc tct cag aa H1L
H1#2R gtg agt act gaa tcg gtg aca cat atg aa
:ימרים ששימשו לסריקת הספריה ותוכננו לפלסמיד הםיהפר
AD003 : cac gtc ttc aaa gca agt gga ttg atg tg
BAR: agc acg gga act ggc atg acg t
לסריקה וכן לאימותים הייתה PCRתוכנית ה
96oC 5min, (94oC 0.5 min, 65oC 0.5 min, 72oC 4 min) X36, 72oC10 min, 4oC.
EtBr-ונצבעו ב TBE- ב1%ל אגרוז 'גבתוצרי הניסוי הופרדו
pSKI015מפה סכמטית של הפלסמיד . 1 תמונה
ר ששימש לסריקת יימהפר, בארבע עותקים 35Sבקרבת הגבול הימיני מופיע , מפה סכמטית המציגה את מבנה הפלסמיד הבינארי
: תמונה זו נלקחה מתוך. מודגש בקו צהובTDNA- ה. ומסומן בחץ מלאהספריה תוכנן לאזור זה
http://www.weigelworld.org/resources/plasmids/activationtagging/pski015
RB

25
הועברו כלל ,לכן. לא ניתן היה לזהות תוצר יחיד,אלפי קוים בו זמניתל הכל ראקציה בוצעש מאחר
ניילון לממברנת העברה.Southern blots (Southern, 1975)התוצרים לממברנה לביצוע ראקציית
(HybondN+)0.4 באמצעות הוצע בN NaOH הממברנה יובשה וסומנה . בהעברה קפילרית למשך לילה
בתור . (2XSSC, 0.1% SDS, 65oC) בתנאים מחמירים שעות6-16באמצעות גלאי רדיואקטיבי למשך
וסומן H1L-H1#2Rמרים יעות זוג הפרי אשר הוגבר באמצWT מצמחי -PCRגלאי שימש תוצר ה
על חזרתי איתור הקומה הנכונה לאחר .תוצאות הסימון נחשפו למסך זרחן ועברו הדמיה. P 32באמצעות
נזרעו מעציץ זהזרעים . ) גנוטיפים10עציץ בן (שורות והתקבלו שלוש הקורדינאטותל לטורים ופעולות אלה
ריכוז ב"AgrEvo" מחברת BASTA סלקציה בקוטל העשבים וצמחונים בני שבועיים עד שלושה עברו
צמח צורך הגעה לם עברו הכלאה עצמית לי וצמחים חיובי -PCRהצמחים ששרדו נבחנו שוב ב . 0.1%
.הומוזיגוט
יצור מוטנטים כפולים
ל סף פרחים שהיו ע .רציפרוקליות בין זוגות המוטונטיםבוצעו הכלאות , על מנת ליצר מוטנטים כפולים
מהצמח השני ואבקופתיחה מצמחי הורים הומוזיגוטים למוטציות עברו סירוס למניעת האבקה עצמית וה
DNAהזרעים הונבטו ואחרי כשבועיים נלקח עלה בודד ממנו הופק . הפרח סומן ונאספו זרעיו, במקביל
.ר ולגן בהתאמה עם שני זוגות הפריימרים המתאמים למחדPCRששימש לאימות הצלחת ההכלאה באמצעות
T-DNA insertion line לצמחי PCR גנומי לאנליזת DNAהפקת
עד לקבלת רסק מלא כדור זכוכיתבאמצעותנת אפנדורף נכתשה במבח,) mg 100( רקמת עלים צעירים
μl 500 שהכילה DNAפה תמיסת הפקת לרקמה הכתושה הוס. Ultramat (SDI, Australia ( באמצעות
(0.2M Tris HCl pH 9, 0.4M LiCl, 25mM EDTA, 1% SDS) .רובת עורבבה היטב ולה הוסף התע
Phenol: Chloroform: Isoamyl Alcohol התערובת עורבבה היטב . בהתאמה, 25:24:1ביחס של

26
, Isopropanolהפזה העליונה הועברה למבחנה נקיה עם נפח שווה של . 13,000g דקות ב 5וסורכזה
. 13,000g דקות ב 10לאחר מכן סורכזה התערובת במשך . 20ºC- דקות ב 20נחה למשך עורבבה היטב והו
O.D מקדם בליעה אופטית ל פיע נקבע יכוז הדוגמה ר .μl ddH2O 100הנוזל סולק והמשקע הומס ב
260nm.
ל אגרוז' וניקויו בגDNAהפרדת
האגרוז . ל אגרוז'גבי ג לוונטים על הרDNAהופרדו מקטעי ה , DNAלשם תצפית או ניקוי של מקטעי
בופר ההפרדה בוצעה ב) Sambrook et al., 1989ראה (פי הצורך - על) 1.5% - 0.7(ריכוזים שונים היה ב
TBE) 89 mM Tris-borate, 89 mM Boric acid, 20 mM EDTA .( התמיסה הובאה לרתיחה
היא , 60oC - כאשר התקררה התמיסה ל. μg/ml 0.8 לריכוז סופי של EtBrוהוסף לה , באמצעות מיקרוגל
הורחפו בבופר DNAדוגמאות . נמזגה למערכת ההרצה והושארה בטמפרוטורת החדר לצורך התקשותה
ידי הרצתם -והופרדו על) Glycerol, 0.1% Bromo-phenol blue, 0.1% Xylene cyanol 50%(הטענה
DNAמקטעי ה . בי שולחן אור על גUV-ל ל'לאחר ההרצה נחשף הג. במשך שעהV 100במתח של
GFX PCR DNA and gel band(ל אגרוז ' מגDNAהרלוונטיים נוקו באמצעות קיט לניקוי
purification kit ( של"Amersham Pharmacia " .
TAIL-PCR-שיבוט תוצרי ה
לה ישהכל 'חתיכת הג, ל אגרוז ונחתכו ממנו' של צמח יחיד הופרדו בגDNA שבוצע על PCR- תוצרי ה
μl 0.2M Tris HCl pH9, 0.4M LiCl, 25mM EDTA, 1% 500)את המקטע שהוגבר הומסה בבופר
SDS)להוהוסף , התערובת עורבבה היטב Phenol: Chloroform: Isoamyl Alcohol (25:24:1) אחרי
ומצות להשקעת חIsopropanol לה נפח זהה של ףהועברה הפאזה הנוזלית למבחנה חדשה והוס, שסורכזה
בקיט T/A אחרי סירכוז ויבוש הומסו חומצות הגרעין במים ושובטו אל תוך פלסמיד על ידי הקצוות. הגרעין

27
QIAGEN PCR Cloning הפלסמיד שובט לחידקי . על פי הנחיות היצרן DH5α ועבר ריבוי וריצוף
. לקביעת נקודת המחדר
E. coli לחיידקי יצור חיידקים קומפטנטים וטרנספורמציה
(LB זריעת בידוד ומושבה יחידה גודלה בתרבית נוזלית של מצע גידול סטנדרטי נזרעו E. coliחיידקי
(10g Tryptone, 5g Yeast Extract, 10g NaCl, pH 7 250 למשך לילה בטילטול של RPM
-O.D600=0.5ל עד ml 250 וגודלה בנפח של 1:100תמיסת החידקים נמהלה . 37oC של בטמפרטורהו
: בבופר המכילונשטפו הורחפו 5000g, -ב דקות 5קרח והושקעו בסירכוז למשך על ידקים קוררו יהח .0.6
(30mM KOAc, 100mM RbC12, 10mM CaC12, 50mM MnC12, 15% glycerol, pH 5.8) .
של-ml 8החיידקים השטופים הורחפו ב. וסורכזה שוב, דקות נוספות5 קרח למשך להתמיסה הושארה ע
(10mM MOPS KOH pH 6.5, 75mM CaC12, 10MM RbC12, 15% glycerol) . תמיסת החיידקים
ל ע .םנספורמציה באמצעות עקת חור הוקפאה במנות ששימשו לט, בסיומן, דקות15הונחה במי קרח למשך
קוררו ,יל במקב. דקות5הופשרה בקרח למשך 50μlשל מנת חיידקים , מנת לשבט אל החיידקים פלסמידים
תמיסת 20μlעד (החיידקים הוספו לתמיסת הפלסמידים , תנאיםהיה לאותם ליגצהפלסמדים או תוצרי ה
, למשך דקה וחצי42oC -ל העברו ר כךאח, דקות30 קרח למשך לרבבו והונחו לאינקובציה עוע) פלסמיד
והונח להם להתאושש LBבשלב זה הוספה לחיידקים תמיסת גידול . על קרח למשך חמש דקותוררו שובוק
הכילבדרך כללש LBשל מצע סלקטיבילהחיידקים נזרעו ע. דקות40 גידול למשך עד טמפרטורתב
(Ampicilin 100 mg/L) , לצורך סריקת המושבות . המחדר או הפלסמידונבדקה נוכחותוהמושבות נבחנו
אמצעותאו ב, סטריקציה מתאימים חיתוכם באנזימי ר באמצעות נעשההאימותהופקו הפלסמידים מהחיידקים ו
PCR מתאימים לשכפולו של המקטע הרלוונטיה עם פריימרים .

28
Western הפקת חלבונים ואנליזת
,Glycerol 10%( כתישה בבופר ההעמסה ללא צבע באמצעותחלבונים הופקו מצמחי ארבידופסיס
0.5 M Tris HCl pH 6.8, 5M Urea, 8% SDS, 20% β-Mercaptoethanol .( לצורך אנליזת
Western באלקטרופורזה על גבי ,הים זריכוז כלורופיל או משקל דוגמהב ם של חלבונידוגמאות ו הופרד
לאחר מכן .(25mM Tris, 200mM glycine, 0.1% SDS)בבופר הפרדה . 12% בריכוז של AAגל
20%בתוספת וה הפרדפר הזהה לבופר ה בבו60Vהועברו החלבונים לממברנת ניטרוצלולוז במתח קבוע של
Methanol מס בבופרו שה חלב רזה 5%הממברנה טופלה בתמיסה של. דקות90 במשך (TTBS)
(50mM Tris HCl pH7.5, 15mM NaCl, 0.05% Tween-20). והוגבה עם נוגדנים פוליקלונלים
-ל הראשוני עם נוגדן שניוני המחוברבכדי לעקוב אחר הסיגנל האימוני הוגב הנוגדן . )1ראה טבלה מספר (
Alkaline Phosphatase ) BioRad( בסיום פרק הזמן נשטפו הממברנות שלוש .למשך שעתים לפחות
תמיסת פיתוח המכילה ב ,פיתוחלהועברו על מנת לצפות בקומפלקס האמיוני , TTBSפעמים בתמיסת
Nitroblue tetrazolium5 - ו-bromo-4-chloro-3 indolyphosphate בבופר סובסטרטים כ ששימשו
והראקציה הופסקה על ידי .(mM Tris HCl pH9.5, 50mM MgCl, 10mM NaCl 100)ראקציה
.שטיפה במים
קביעת ריכוז כלורופיל
5 μl1-הורחפו ב, ממיצויי כלורופלסטים או תילקואידיםml 80% אצטון )Arnon, 1949 (שימשו ו
ונקבע בהן ריכוז , עמילן ואגרגטים שונים, ת סורכזו להשקעת חלבוניםהמבחנו. קביעת ריכוז כלורופילל
ריכוז הכלורופיל ). 645nm- ו663nm( קביעת בליעת המיצוי האצטוני בשני אורכי גל באמצעותהכלורופיל
: פי הנוסחה-במיצויים השונים חושב על
Chlorophyll concentration (µg /μl) =[20.2 x (A645)+8.02 x (A663)]/5.

29
2D-PAGE- בהפרדת חלבונים לפי מטען וגודל
,.Kruft et al)כפי שתואר בעבר על יד הפרדת חלבונים לפי נקודה איזואלקטרית של חלבון נעשתה
בבופר וסהומ, כלורופיל μg 100-השקול לכ חלבון μg�700- כ שלדוגמאות .עם התאמות קלות (2001
דקות 30סורכזו למשך הדוגמאות . (8M urea, 1% NP-40, 2% CHAPS, 0.28% DTT) המכיל
אורך בpH 4-7 אשר מורכב מגרדיאנט של AAסטיריפ של על גבי נטו לפי מטען והפרד ואז -13000gב
30- שעות ב6: התוכניתההפרדה בוצעה בשדה חשמלי משתנה על פי . )Amersham-Phrmacia(מ " ס13
V ,660- שעות ב V ,500-שעה ב V ,2,000-שעה ב V8,000- שעות ב-8 ו V 74,500הגעה לל עד Vh .
ף לי על מנת להח-SDSה המועשרת ב דקות כל אחת בתמיס15 למשךבסיום עברו הסטריפים שתי שטיפות
50mM Tris HCl pH 6.8, 6 M urea, 30%) :התמיסה הכילה. לדטרגנט יונישבדוגמהנט את הדטרג
glycerol, 2% SDS). ל 'הונחו הסטריפים על גבי גבסיוםSDS-PAGE .כפי שתואר ההפרדה נעשתה
ל ' ג ,אחרי ההפרדה. שעות6- כmA 35- בכ"משך שעה ואחל mA 25 אך בזרמים של ,בפרק הקודם
.או הועבר לממברנה לבצוע תגובות עם נוגדניםCoomassie Stain - בלילהל החלבונים הושרה
מדידות פלורסנציה בצמחים
יעילות רכיבי אומדן לפוטנציאל ולבתור של כלורופיל הצמחים מן הטיפולים השונים נלקחו למדידת פלורסנצי
צמח הונחו פרוסים ל מספר עלים מכ .PAM (Walz, Germany) באמצעות מכשיר המערכת הפוטוסנתטית
פלורסנציה מינמלית . מים וכוסו בנייר עדשות רטוב המאפשר מעבר של רוב האורמעל גבי נייר ווטמן רטוב
)Fo ( 0.3-תוך הארה ב דקות 10נבדקה לאחר שהעלים עברו הסתגלות לחושך במשך µmol m-2 sec-1 .
ממנורת µmol m-2 sec-1 3000ה של נמדדה אחרי פולס קצר של אור בעוצמ) FM(פלורסנציה מקסימלית
) maximum quantum yield-Fv/Fm(מערכים נמדדים אלה חושב פוטנציאל מקסמלי ). (osramהלוגן
אור בעצמה שלטיפולי עברו העלים ,לחילופין. PSII F0)/ FM- = (FM FV/FMשל מעבר אלקטרונים דרך
) יצור עצמי (mm 18פוליקארבונאט /ילטר מיםת הלוגן לבן דרך פ ממנור µmol m-2 sec-1 1800 עד
30
במקביל . חושךלבכל פרק זמן אחרי הסתגלות חוזרת נמדדה הפלורוסנציה .UVסינון למניעת התחממות ו
.ים אחרים אך לא נמצאו הבדלים מהותיים ולכן הנתונים אינם מוצגיםינתטינבדקו גם מדדים פוטוס
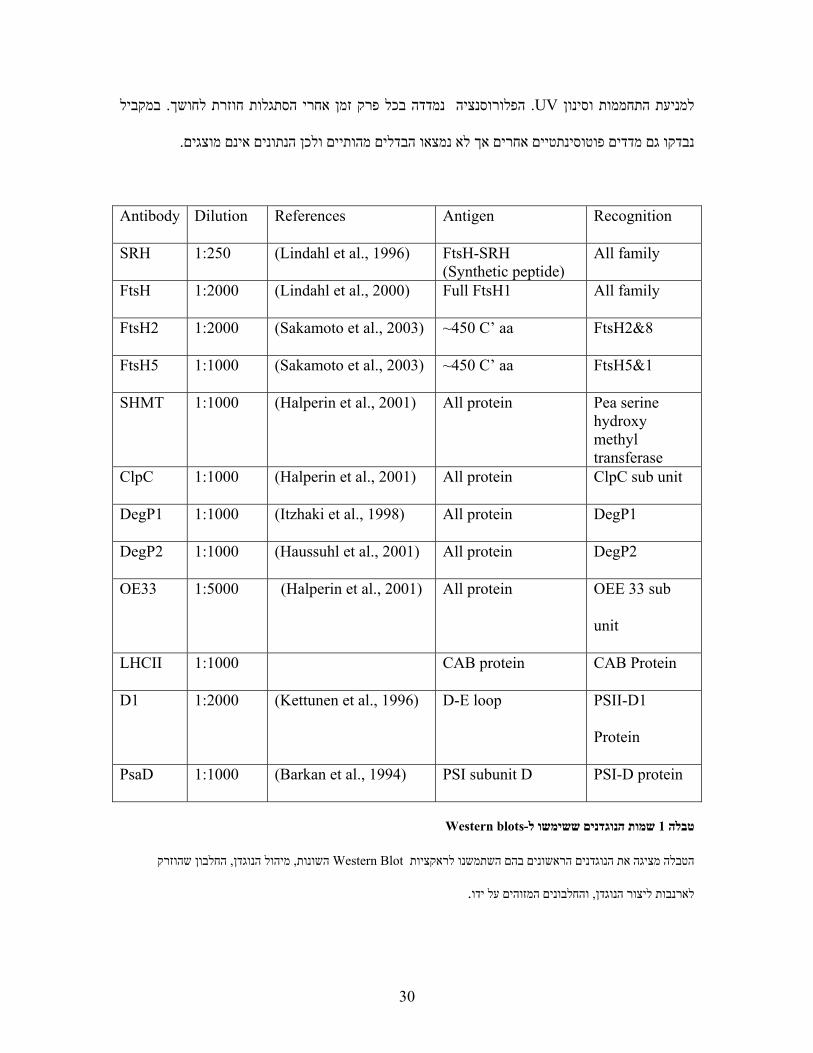
Recognition Antigen References DilutionAntibody
All family
FtsH-SRH(Synthetic peptide)
)Lindahl et al., 1996( 1:250SRH
All family Full FtsH1 )Lindahl et al., 2000( 1:2000FtsH
FtsH2&8 ~450 C’ aa )Sakamoto et al., 2003( 1:2000FtsH2
FtsH5&1 ~450 C’ aa )Sakamoto et al., 2003( 1:1000FtsH5
Pea serine hydroxy methyl transferase
All protein )Halperin et al., 2001( 1:1000SHMT
ClpC sub unit All protein )Halperin et al., 2001( 1:1000ClpC
DegP1 All protein )Itzhaki et al., 1998( 1:1000DegP1
DegP2 All protein )Haussuhl et al., 2001( 1:1000DegP2
OEE 33 sub
unit
All protein )Halperin et al., 2001( 1:5000OE33
CAB Protein CAB protein 1:1000LHCII
PSII-D1
Protein
D-E loop )Kettunen et al., 1996( 1:2000D1
PSI-D protein PSI subunit D )Barkan et al., 1994( 1:1000PsaD
Western blots- שמות הנוגדנים ששימשו ל1טבלה
החלבון שהוזרק , מיהול הנוגדן, השונות Western Blotהטבלה מציגה את הנוגדנים הראשונים בהם השתמשנו לראקציות
.והחלבונים המזוהים על ידו, לארנבות ליצור הנוגדן

31
Reverse Transcription-PCR (RT-PCR) בשיטת mRNAאנליזת
רקמה נטחנו בחנקן נוזלי והועברו Izhaki et al.) 2001 :(300-100 mg לפי הבוצע ם מעליRNAהפקת
Phenol: Chloroform: Isoamyl Alcohol שלml 0.5סטריליות שהכילו ml 1.5למבחנות אפנדורף
mM Tris HCl pH 8.5, 20 mM EDTA, 100 mM 100) בופר מיצויml 0.5 - ו, 25:24:1 ביחס של
NaCl, 1% sarcosine). 13,000 - דקות ב5 במשך ה היטב וסורכזה המבחנות עורבבתכולת g . המקטע
של ביחס ml Phenol: Chloroform: Isoamyl Alcohol 0.5העליון הועבר למבחנות חדשות ועורבב עם
1 הועבר למבחנות שהכילו ן העליומקטעוה g 13,000 - דקות ב 5המבחנות סורכזו שוב במשך . 25:24:1
ml EtOH 96%50 - ו μl 3M Na-Acetate, pH 5.6 .20°- -המבחנות הונחו בC דקות ולאחר 30 למשך
μl 100 - ברלאחר יבוש קצ סולק והמשקע הומס נוזלה. g 13,000 - דקות ב10מכן סורכזו במשך
לאחר כמחצית השעה התקבלה תמיסה . 10U RNAse-free DNAseI (Fermentas)תמיסה המכילה ב
בליעה נקבע לפי -RNAכמות הו, Tri Reagent (Sigma)ה שוב בקיט ת זו נוק, שלם-DNAהנקיה מ
הוספו אליו ng RNA 500בוצעה על Reverse Transcription (RT)-ריאקצית הNanoDrop. -באופטית
1μg oligo-dT primer0.5ו mM dNTP- 60- דקות ב5 התערובת הונחהºCומייד הועברה לקרח .
mM DTT, 2 unit RNaseOUTTM, 200 unit Superscript II reverse 10לראקציה הוספו
transcriptase (Stratagene ) , 42- דקות ב50התערובת הונחהºC 70 - דקות ל15 והועברהºC על מנת
ביןשיות וחמש כמ ומהמיהול נלקחו 5 נמהל התוצר פי RT-בסיום ראקצית ה. סיק את פעילות האנזיםלהפ
μl1-10 לראקציתPCR לגןמרים ספציפייםיעם פרי AtTubulin ) שימשו לקביעת פריימרים אלו ). 2טבלה
למירים שבין הפריי מקטע שהוגברבשמאחר .DNA ווידוא שאין בדוגמאות ולmRNAהכמות היחסית של ה
. על ידי נוכחות של בנד גבוההיואם DNA ניתן היה להבחין בזיהומי , היה אינטרוןטובולין
נערך ניסוי השוואתי לשם קביעת של טובולין בין הדוגמאות השונותmRNA של לאחר קביעת הכמות
הדוגמאות בהן .)2 טבלה( פיים לכל אחד מהגניםיימרים הספצי הפרל ידי עשונים FtsHהכמות היחיסית של
באמצעות והו וז1%תוצרי הראקציה הופרדו בגל אגרוז . טובולין שלmRNAהשתמשנו הכילו כמות זהה של
32
EtBr .תהגל צולם וכומת באמצעות תוכנ ImageJ 1.34 NIH- הכמותי של כל אחד והושווה לעקומי הכיול
.מהגנים
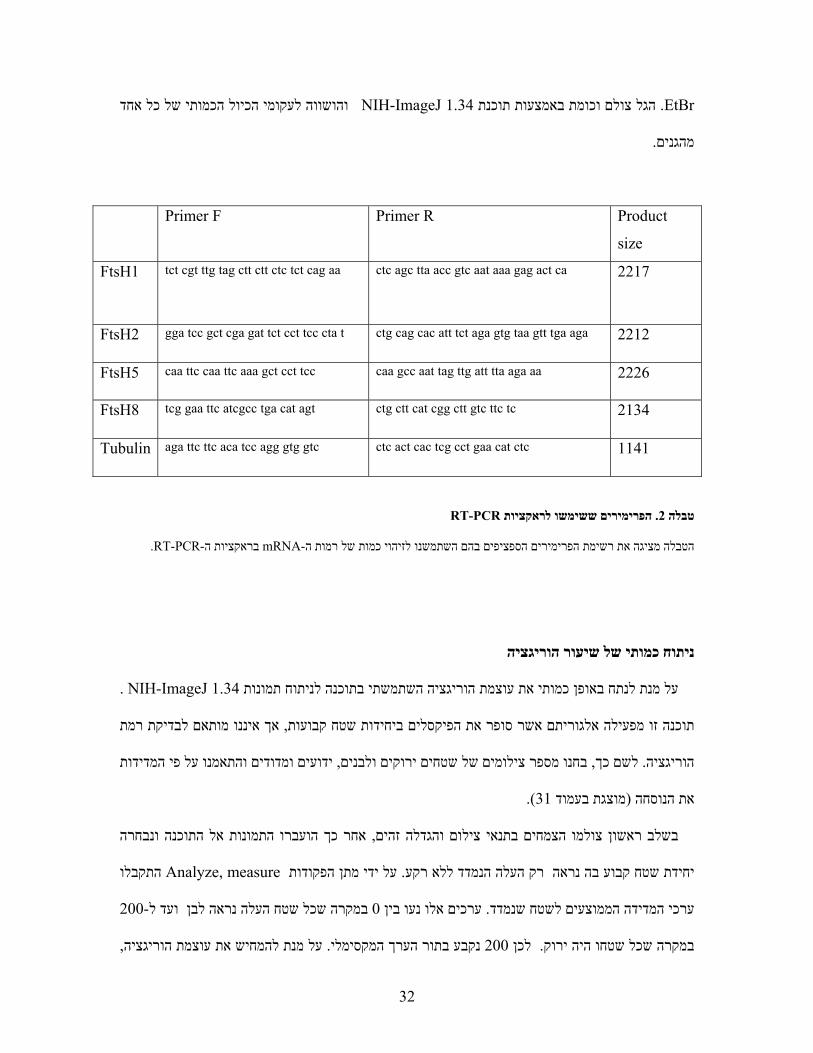
Product
size
Primer R Primer F
2217 ctc agc tta acc gtc aat aaa gag act ca tct cgt ttg tag ctt ctt ctc tct cag aa FtsH1
2212 ctg cag cac att tct aga gtg taa gtt tga aga gga tcc gct cga gat tct cct tcc cta t FtsH2
2226 caa gcc aat tag ttg att tta aga aa caa ttc caa ttc aaa gct cct tcc FtsH5
2134 ctg ctt cat cgg ctt gtc ttc tc tcg gaa ttc atcgcc tga cat agt FtsH8
1141 ctc act cac tcg cct gaa cat ctc aga ttc ttc aca tcc agg gtg gtc Tubulin
RT-PCRהפרימירים ששימשו לראקציות . 2טבלה
.RT-PCR- בראקציות הmRNA- מת הפרימירים הספציפים בהם השתמשנו לזיהוי כמות של רמות השיהטבלה מציגה את ר
עור הוריגציה יניתוח כמותי של ש
.-ImageJ 1.34 NIH באופן כמותי את עוצמת הוריגציה השתמשתי בתוכנה לניתוח תמונות על מנת לנתח
אך איננו מותאם לבדיקת רמת ,ם אשר סופר את הפיקסלים ביחידות שטח קבועותיתתוכנה זו מפעילה אלגור
ידועים ומדודים והתאמנו על פי המדידות ,לבניםו בחנו מספר צילומים של שטחים ירוקים ,לשם כך. הוריגציה
).31ד מוצגת בעמו (את הנוסחה
הועברו התמונות אל התוכנה ונבחרה אחר כך, בשלב ראשון צולמו הצמחים בתנאי צילום והגדלה זהים
התקבלו Analyze, measure מתן הפקודות ל ידיע. יחידת שטח קבוע בה נראה רק העלה הנמדד ללא רקע
200-עד לו לבן העלה נראה שטחכל במקרה ש0 בין נעוערכים אלו . ים לשטח שנמדדעערכי המדידה הממוצ
,יש את עוצמת הוריגציהחעל מנת להמ .ערך המקסימליבתור ה נקבע200 לכן . ירוקשכל שטחו היהבמקרה

33
ים מיצגים ימספרי הבינ, הוא לבן לחלוטין0- וירוק לגמריוא ה10 בה 0-10סקלה בין באופן שרירותי קבענו
.וה את הערך הנמדד על ידי התוכנהו מה X כשהמשתנה,העלבעור השטחים הירוקים יבאופן לינארי את ש
הערך שהתקבל בסקלה שקבענו ו 100 שנמדד היה X ,ירוקו לבן 50%רמת הורגציה היא לדוגמה כאשר
0.98ערך זה מקדם רגרסיה של ל. greening = (1-(200-X)/200)*10: שהתאמנועל פי הנוסחה. 5היה
עשוי את שטח הפנים בלבד הוא מייצגערך זה שמאחר . ריגציהואו המדד אמין לשעור ההורקה הוא ולכן
. רק אם עובי העלה זהה מדד לכמות הכלורופיל ברקמהלהוות
על ידי ריסוק אצטון80% של ml 1 - רקמה טריה ב100mgקה נוספת היתה מיצוי משקל קבוע של ניטכ
,ו הדוגמאות להשקעת עמילן וחלקי רקמה ותאיםבסיום פרק זמן זה סורכז. למשך לילהוהנחה בטילטול בקור
בדומה לבדיקה שערכנו בפרקים הקודמים אך של כמות הכלורופיל ברקמהערכנו קביעה ספקטרומטרית ו
.Chlorophyll concentration (µg /g) =[20.2 x (A645)+8.02 x (A663)]*10הנוסחה הותאמה והיתה
מיקרוסקופיית אור והיסטולוגיה
תפרחות ופרחים : בקצרה. עם התאמות קלות)Li et al., 2003(גמאות למיקרוסקופ הוכנו על פי הדו
60:10:2:40.ביחס FAA (96% EtOH: 40% formaldehyde: acetic acid: dH2O)הושרו בתמיסת
דהידרציה על ידי שטיפות הבהמשך בוצע. סה זו מקבעת את מבנה הרקמהתמי, למשך לילה בחדר קור
הרקמה נחתכה .שאחריהם הרקמה קובע בפארפלסט (EtOH 10-96%) בריכוזים עולים של אלכוהול
החתכים הונחו על גבי זכוכית ונצבעו באמצעות . באמצעות מיקרוטוםμm 7-20לחתכים ברוחב של
Safranin O ובנוסף Fast green,כאשר ה Safranin-צובע את דפנות התאים וה -Fast green צובע את
.פ אורוו דרך מיקרוסקנצפו וצולמ בסיומו (Lin et al., 2004)פי על תהליך הצביעה התבצע.הציטופלסמה

34
תוצאות
זיהוי ואפיון מוטנטים בגנים יחידים.א
T-DNA באמצעות מחדר של ftsh1השתקת הגן
ניסיתי , מהן ההשפעות הפזיולוגיות של העדרו, ואם איננו, חיוני לקיום הצמחFtsH1על מנת לבחון האם
ספריית המוטנטים .אינם מבטאים את הגן ש כך בתוך הגןT-DNAלאתר קווי ארבידופסיס בעלי מחדר של
נסרקה Wisconsin-Madison (Krysan et al., 1999; Sussman et al., 2000)של אוניברסיטת
ותוצרי הניסוי )TAIL-PCR )Singer and Burke, 2003בוצע . 1FtsH-לאיתור קווי מחדר לגן המקודד ל
לא , (DNA-pools)מאחר וכל ראקציה בוצעה עם אלפי קוים בו זמנית ). חומרים ושיטותראה (ל 'הופרדו בג
Southern blot כלל התוצרים לממברנה לביצוע לכן הועברו ). 4Aתמונה (ניתן היה לזהות תוצר יחיד
,לרבות האינטרונים, FtsH1-שימש כל האזור המקודד ל, תור סמןב. )Southern, 1975(בתנאים מחמירים
הממברנה הוגבה עם הסמן ותוצאות הסימון נחשפו . וסומן בזרחן רדיואקטיביPCRאחרי ששוכפל באמצעות
. 4B)תמונה (למסך זרחן ועברו הדמיה
.FtsH1-מוטציית השתקה בגן לסריקת הספריה לאיתור . 4תמונה
התמונה מציגה שלב בזיהוי בו אותרו שתי , טותזיהוי שתי קואורדינ
.(A,3)הקורדינאטות
(A) ריבוי תוצרים ב-TAIL PCR על גבי כפי שמתקבל בסיום ההפרדה
וגודל בבסיסים מצויין בגוף ,"פול"מספר ה. EtBr- הצבוע בל אגרוז'ג
.התמונה
)B( Southern blot : הממברנה הוגבה עם סמן שכולל את האזור
תוצאה . ואקטיביוהתה באמצעות הדמיית זרחן רדיוז -FtsH1המקודד ל
ארבעת הבנדים נוצרו בגלל מבנה (. חיובית התקבלה בשני מקרים
).המחדר
0 1 2 3 A B C D E F DNA pool A
B
bp
1638 1018
1638 1018
500
500

35
ובו נקודה זו מייצגת עציץ יחיד. A,3,28תוצאות הניסוי הובילו לנקודת החיתוך של שלושת הקואורדינטות
בגיל שבין שבועיים לשלושה . שגודלו מעציץ זה נאספו זרעים .)חומרים ושיטותראה ( גנוטיפים שונים 10
-TAILכל הצמחים העמידים נבדקו שוב באמצעות . BASTAקוטל העשבים עברו הצמחים סלקציה ב
PCR .רק מצמחים שהיתה בהם תגובה עם הפריימירים שלFtsH1 ,פרדנאספו זרעים מכל צמח בנ.
זיהוי נקודת המחדר והגנוטיפ
לשם ניקוי מארועי WTביצעתי הכלאה מחזירה עם ,PCR-לצמחים העמידים שהראו תוצאות חיוביות ב
. בסלקציה לעמידות1:3התפצלו וזרעיהם עצמית זו התאפשר לעבור הכלאהתוצרי הכלאהל. חדירה נוספים
מכאן ניתן להסיק כי התרחש ).5תמונה (FtsH1חדר בגן כל הצמחים העמידים הראו תוצאות חיוביות למ
נקודת החדירה נמצאת באקסון בעמדה . שובט אל תוך פלסמיד ורוצף-PCRתוצר ה. רה יחידארוע חדי
לשם אימות חוסר הביטוי RT-PCRביצעתי , לכן. הממוקמת אחרי המוטיבים המוכרים בגן-ATG מ1850
יש צורך FtsHעל מנת ללמוד על תפקידיו של ). 5Bתמונה( למחדר בצמחים שהיו הומוזיגוטיםשל הגן
באות לידי ביטוי רק במצב (Loss of function) מאחר והרבה מוטציות השתקהבצמח הומוזיגוטי למחדר
. BASTA- בוצעו שלושה מחזורי האבקה עצמית לאחר סלקציה בלכן .זה
.ftsh1-זיהוי מוטנט ב. 5תמונה
(A)תוצאות ה PCR- :DNA גנומי מצמחים בודדים
). A-Fאותיות( המתוארים פריימריםהוגב עם זוגות
:התמונה מציגה שלושה גנוטיפים
מגיב עם זוגות -ftshללא מחדר ב WTגנוטיפ . 1
A+B, A+D הפריימרים
למחדר מגיב עם כל זוגות גנוטיפ הטרוזיגוט. 2
. הפריימרים
A+D, C+B עם במגי, גנוטיפ הומוזיגוט למחדר. 3
.A+Bעם אך לא
1 2 3 1 2 3 1 2 3 1 2 3 A+B C+B A+D E+F
A
D
E
F
BC
A
B
3 1 A+B

36
(B) RT-PCR עם פריימרים A+B המתאימים
על צמח הומוזיגוטי למחדר ועל , ftsh1לקצוות של
WT.
,נבדק בתור הומוזיגוט) צאצאים16- לא פחות מ (BASTA- הראה כי כל צאצאיו עמידים לF3צמח שבדור
רק . -PCR ותוצאות חיוביות בBASTA-לאה לעמידות מ: תנאיםחשב הומוזיגוט רק אם ענה על שני צמח נ
והיא 0.99רמת המובהקות היא . זיולוגיות כפי שמתואר בהמשךיצמחים הומוזיגוטים למוטציה עברו בחינות פ
.1:2:1חושבה על פי הסגרגציה שנצפתה לעמידות
הבר איננו שונה באופן מובהק מצמחי ftsh1המוטנט
לעומת צמחי ) קווי המחדר( נבחנו הצמחים המוטנטים , בצמחFtsH1של על מנת לנסות ולהבין מה תפקידו
WT במספר ניסויים :
. שווהאו , קצר, ובמשטר תאורה של יום ארוך, ניסוי לבחינת קצב גידול בתנאי גידול סטנדרטים.1
. וזמן עד לפריחה, גובה,נבחנו משקל טרי
, ימים7-14הצמחים גודלו למשך . פיטוטרוןצוניים ב בתנאי טמפרטורה קיאלהפרמטרים ניסוי לבחינת . 2
. ובמשטרי תאורה זהים,35oC ,30 ,22 ,16: פרטורהבטמ
ויכולת נביטה וגידול , מצע גידול סטנדרטיעל ניסוי לבחינת חיוניות זרעים באמצעות מבחן נביטה .3
נבדקה יכולת הגידול כן כמו . 8 ,7 ,6.5 ,5.8 ,5 ,4.5 של pHבמצעים בעלי חומציות שונה בערכי
. אחרי נביטה בתנאים סטנדרטים10oCבטמפרטורה נמוכה של
.וחושב היחס בין סוגי הכלורופיל השונים, יות והרכב הכלורופילו נמדדו כמ.4
. בכל הפרמטרים שנבדקו לא נמצאו הבדלים מובהקים בין המוטנטים לצמחי הביקורת
FtsH הצביעו על מעורבות אפשרית של )Lindahl et al., 2000( שביצעו -in-vitroמאחר שניסויי ה
נציאל האלקטרוכימי טהחלטתי לבדוק את הפו, PSII-D1במערכת התיקון של מרכז הראקציה הפוטוסנטתית
ערך זה . (Maximum quantum yield - Fv/Fm)פלורסנציה של כלורופיל באמצעות מדידת PSIIשל
לשמששעשויה , מתוך סך האור שנקלטשל אנרגיית האור ייצג את הכמות היחסית המקסימאלית מ
37
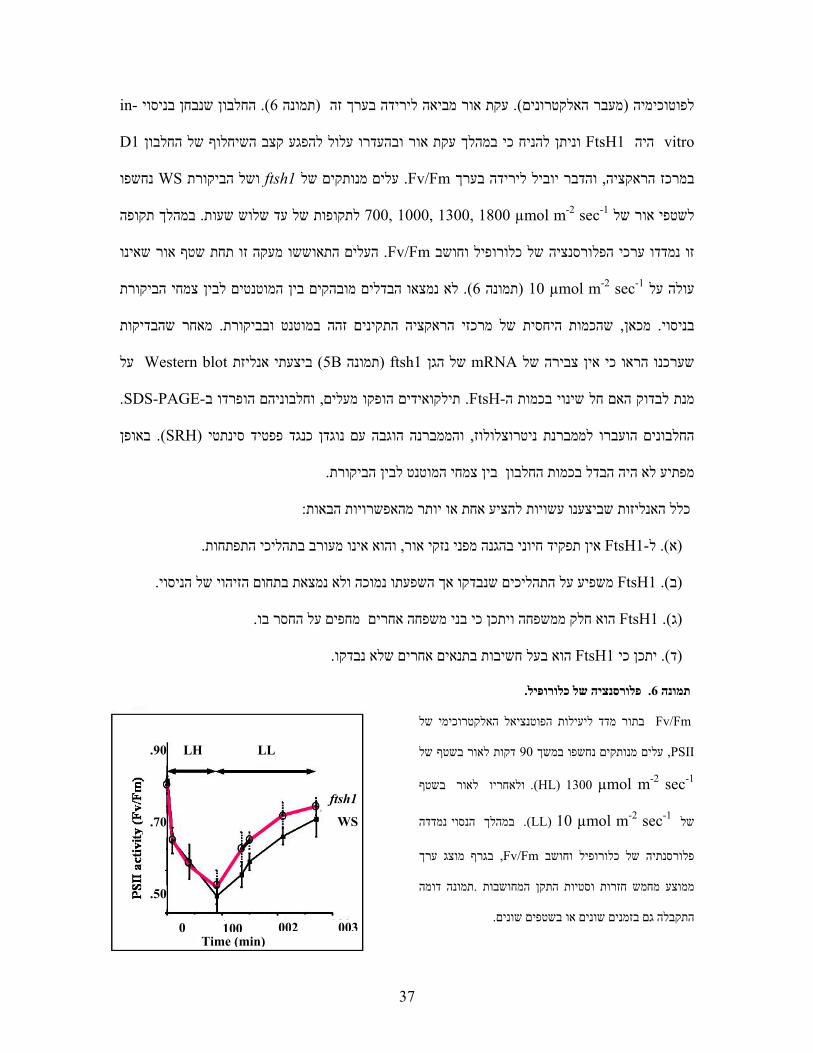
-inהחלבון שנבחן בניסוי ). 6תמונה ( עקת אור מביאה לירידה בערך זה). מעבר האלקטרונים(לפוטוכימיה
vitro היהFtsH1 חלבון עלול להפגע קצב השיחלוף של הואור ובהעדר וניתן להניח כי במהלך עקתD1
נחשפו WS ושל הביקורת ftsh1עלים מנותקים של . Fv/Fmהדבר יוביל לירידה בערך ו, במרכז הראקציה
במהלך תקופה . לתקופות של עד שלוש שעותµmol m-2 sec-1 1800 ,1300 ,1000 ,700לשטפי אור של
תחת שטף אור שאינו העלים התאוששו מעקה זו.Fv/Fmזו נמדדו ערכי הפלורסנציה של כלורופיל וחושב
לא נמצאו הבדלים מובהקים בין המוטנטים לבין צמחי הביקורת ). 6תמונה (µmol m-2 sec-1 10עולה על
מאחר שהבדיקות . הכמות היחסית של מרכזי הראקציה התקינים זהה במוטנט ובביקורתש, מכאן. בניסוי
על Western blotביצעתי אנליזת ) 5Bתמונה (ftsh1 של הגן mRNAשערכנו הראו כי אין צבירה של
. SDS-PAGE-בהופרדו ם וחלבוניה, תילקואידים הופקו מעלים. -FtsHמנת לבדוק האם חל שינוי בכמות ה
באופן . (SRH)והממברנה הוגבה עם נוגדן כנגד פפטיד סינתטי , החלבונים הועברו לממברנת ניטרוצלולוז
.וטנט לבין הביקורתמפתיע לא היה הבדל בכמות החלבון בין צמחי המ
:ו יותר מהאפשרויות הבאות כלל האנליזות שביצענו עשויות להציע אחת א
.הוא אינו מעורב בתהליכי התפתחותו, אין תפקיד חיוני בהגנה מפני נזקי אורFtsH1-ל ).א (
. י משפיע על התהליכים שנבדקו אך השפעתו נמוכה ולא נמצאת בתחום הזיהוי של הניסו FtsH1).ב (
. הוא חלק ממשפחה ויתכן כי בני משפחה אחרים מחפים על החסר בוFtsH1). ג(
. הוא בעל חשיבות בתנאים אחרים שלא נבדקוFtsH1יתכן כי ). ד(
. פלורסנציה של כלורופיל.6תמונה
Fv/Fm בתור מדד ליעילות הפוטנציאל האלקטרוכימי של
PSII , לאור בשטף של דקות90עלים מנותקים נחשפו במשך
(HL) 1300 µmol m-2 sec-1 . ולאחריו לאור בשטף
במהלך הנסוי נמדדה .µmol m-2 sec-1 (LL) 10של
בגרף מוצג ערך , Fv/Fmפלורסנתיה של כלורופיל וחושב
תמונה דומה .ממוצע מחמש חזרות וסטיות התקן המחושבות
. התקבלה גם בזמנים שונים או בשטפים שונים
HL LL
0 100 200 300
ftsh1
WS
0.5
0.7
0.9
Time (min)

38
FtsH איזוזימים מספר הוא חלבון כלורופלסטי בעל
החלבונים . לקואיד הופקו מתילקואידים מבודדים בבופר דנטורטיבי שאינו מכיל דטרגנט יונייחלבוני ת
שהחלבונים מופרדים בה תחילה על פי הנקודה האיזואלקטרית שלהם ואחר , 2D-PAGEהופרדו במערכת
-SRHת הראקציה הראו ארבעה ספוטים שונים שהגיבו עם הנוגדן כנגד תוצאו. כך על פי משקלם המולקולרי
FtsH .ה יותר בולרי גבושניים בעלי משקל מולק-kDa 1.5-2 אפשר להסביר ).7תמונה ( מהשניים האחרים
. מודיפקציות מאוחרות או שמדובר במספר חלבונים המקודדים מגנים שונים: את ריבוי הספוטים בשתי דרכים
,FtsH1, FtsH2:זוהו במעבדתנו כל ארבעת הספוטים והם, פרויקט הגנום של הארבידופסיס השלמת לאחר
FtsH5, FtsH8 ) Sinvany-Villalobo et al., 2004(.
2D-Western blot אנליזת . 7תמונה
ל על פי נקודה איזואלקטרית'הופרדו בג WT חלבוני התילקואיד מצמחי
,SDS-PAGEהופרדו שנית על גבי ,תוצרי הפרדה זו. 4-7של pHבערכי
המזהה את כל האיזוזימיםתוצרי ההפרדה הועברו לממברנה והוגבו עם נוגדן
. התמונה מציגה את רק החלק של הממברנה שהגיב עם הנוגדנים .FtsH2 הספוט החזק ביותר זוהה בהמשך בתור. FtsH של
FtsH עשר גנים-היא משפחה בת שנים
.(Adam et al., 2001) יות לקיומה של משפחת גניםו החלו להצטבר עדהאלהניסוים סמוך לסיום ה
ועוד ארבעה החסרים FtsH גנים המקודדים לחלבוני 12ימים יהאנוטציה הגנומית מציעה כי בארבידופסיס ק
תשעה מתוכם עשויים . )Sokolenko et al., 2002(את מוטיב הקישור לאבץ ולפיכך הם אינם פרוטאזות
ניתוח ). 3טבלה ( )Sakamoto et al., 2003(להיות מוכוונים לכלורופלסט ועוד שלושה למיטוכונדריה
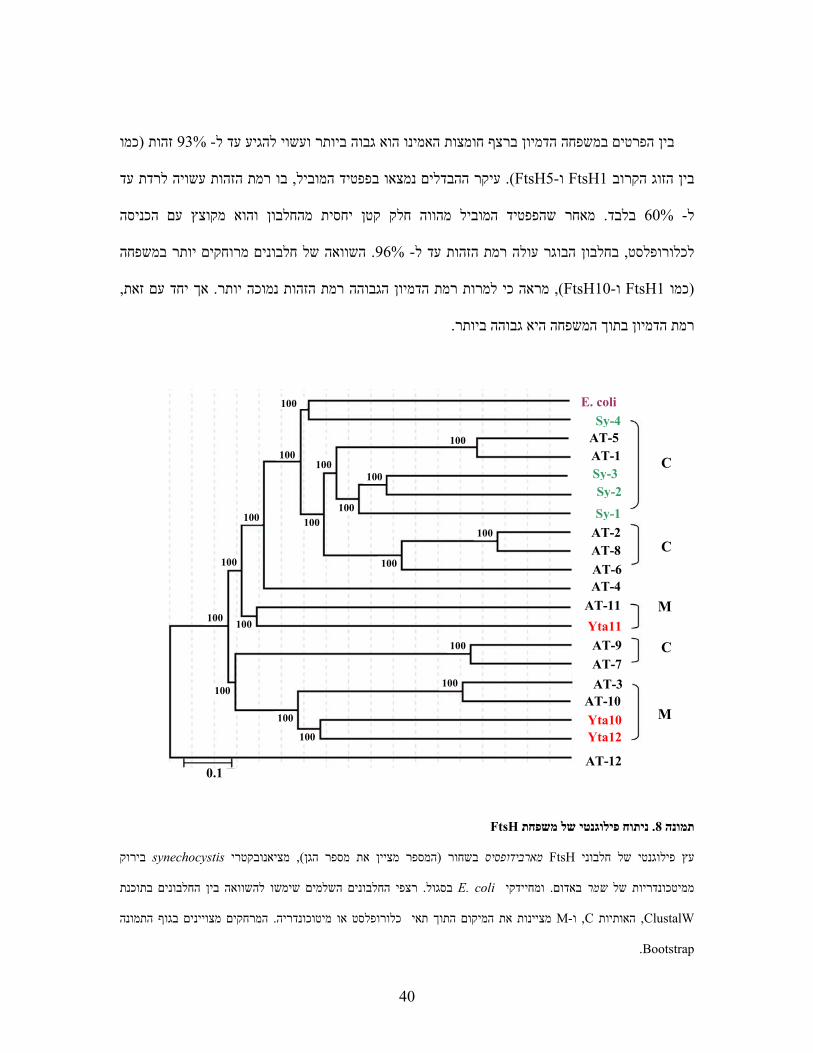
מציע הבחנה בין הענפים ) 8תמונה (פילוגנטי של משפחה זו עם אורטולוגים משמר וציאנובקטריה
: וניתן להבחין במספר פרטים, רצף החלבון השלם שימש לניתוח זה. הכלורופלסטים למיטוכונדריאלים
5 6
86
56
MW kDa
pH
39
גניזימים אחרים מאשר מאור FtsH של ארבידופסיס עשויים להיות דומים יותר לחלבוניFtsHחלבוני
FtsH5 ,FtsH2- וFtsH1 בארבידופסיס מתחלקים לזוגות FtsH’s-רוב ה. אחרים מאותו הצמח-FtsH’sל
ארבעת . הדבר מעיד על דופליקציות מאוחרות באבולוציה, FtsH10- וFtsH9 ,FtsH3- וFtsH8 ,FtsH7-ו
הנראה הדבר מעיד על דופלקציות אינם מתחלקים לזוגות קרובים בתוך המין ככל הנותריםFtsH’s-ה
. מוקדמות יותר באבולוציה
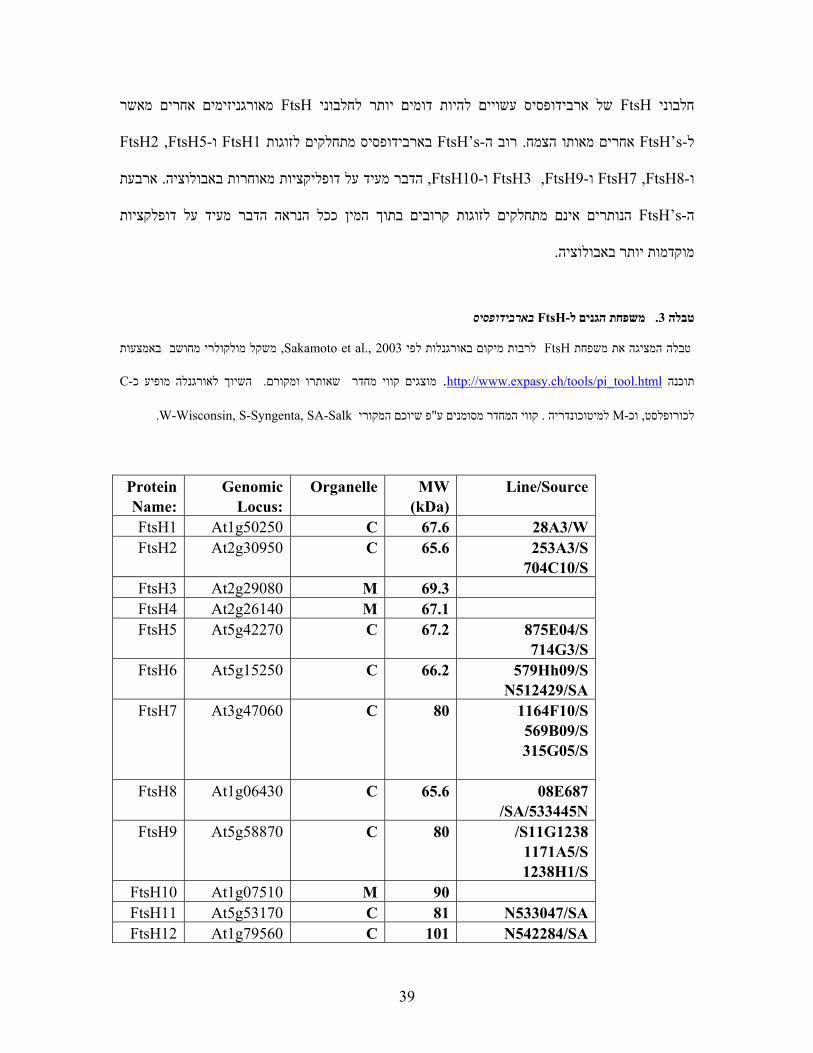
בארבידופסיס FtsH- משפחת הגנים ל.3טבלה
באמצעות משקל מולקולרי מחושב, Sakamoto et al. ,2003 לפי לרבות מיקום באורגנלותFtsHטבלה המציגה את משפחת
C-נלה מופיע כג השיוך לאור.קווי מחדר שאותרו ומקורםמוצגים .html.tool_pi/tools/ch.expasy.www://httpתוכנה
.W-Wisconsin, S-Syngenta, SA-Salk המקורי פ שיוכם"קווי המחדר מסומנים ע. למיטוכונדריה M- וכ,לכורופלסט
Line/Source MW (kDa)
Organelle Genomic Locus:
Protein Name:
28A3/W 67.6 C At1g50250 FtsH1 253A3/S
704C10/S 65.6 C At2g30950 FtsH2
69.3 M At2g29080 FtsH3 67.1 M At2g26140 FtsH4
875E04/S 714G3/S
67.2 C At5g42270 FtsH5
579Hh09/S N512429/SA
66.2 C At5g15250 FtsH6
1164F10/S 569B09/S 315G05/S
80 C At3g47060 FtsH7
687E08 N533445//SA
65.6 C At1g06430 FtsH8
1238G11/S 1171A5/S 1238H1/S
80 C At5g58870 FtsH9
90 M At1g07510 FtsH10 N533047/SA 81 C At5g53170 FtsH11 N542284/SA 101 C At1g79560 FtsH12
40
כמו ( זהות 93% - הדמיון ברצף חומצות האמינו הוא גבוה ביותר ועשוי להגיע עד לבין הפרטים במשפחה
בו רמת הזהות עשויה לרדת עד , עיקר ההבדלים נמצאו בפפטיד המוביל).FtsH5- וFtsH1בין הזוג הקרוב
ם הכניסה מאחר שהפפטיד המוביל מהווה חלק קטן יחסית מהחלבון והוא מקוצץ ע. בלבד60% -ל
השוואה של חלבונים מרוחקים יותר במשפחה . 96% -בחלבון הבוגר עולה רמת הזהות עד ל, לכלורופלסט
, יחד עם זאת אך.נמוכה יותרמראה כי למרות רמת הדמיון הגבוהה רמת הזהות , )FtsH10- וFtsH1כמו (
.רמת הדמיון בתוך המשפחה היא גבוהה ביותר
FtsHנטי של משפחת פילוגניתוח. 8תמונה
בירוקsynechocystis מציאנובקטרי, )המספר מציין את מספר הגן( בשחור מארבידופסיס FtsHעץ פילוגנטי של חלבוני
רצפי החלבונים השלמים שימשו להשוואה בין החלבונים בתוכנת . בסגול E. coliומחיידקי . באדום שמרממיטכונדריות של
ClustalW , האותיותC ,ו-Mהמרחקים מצויינים בגוף התמונה . יינות את המיקום התוך תאי כלורופלסט או מיטוכונדריה מצ
Bootstrap.
0.1 AT-12
Yta12 Yta10
Yta11
AT-10 AT-3 AT-7 AT-9
AT-11 AT-4 AT-6 AT-8 AT-2
AT-1 AT-5
Sy-1
Sy-2 Sy-3
Sy-4 E. coli
C
C
M
C
M
100
100100
100
100
100
100
100
100
100 100
100
100100
100
100
100
100

41
עשויה , בארבידופסיס קרובים יותר פילוגנטית לחלבונים ממינים רחוקים FtsHשחלק מחלבוניעובדה ה עצם
וכן לסייע בהבנת ,לסטייםוענפים כלורופ על קיומם של ענפים מיטוכונדרייליםולהעיד על הפונקציה שלהם
הצפויים FtsHחלבוני . בכלורופלסט ובמציאת התהליכים שהם מעורבים בהםFtsHתפקידם של חלבוני
.להגיע לכלורופלסט מתפצלים לשלושה ענפים עקריים כאשר בכל ענף ישנו זוג קרוב
מוטנטים נוספים ואפיון איתור
סרקתי באופן ממחושב את , גנים נוספים במשפחהנה תוצאות ההשתקה שליעל מנת לברר מה תהי
הסריקה בוצעה. SALK ואת ספריית המוטנטים של Syngentaספריית המוטנטים המרוצפים של חברת
בוצע כנגד מאגרי המידע של החברות BLAST-ה. BLASTN בתוכנת DNA-באמצעות השוואת רצפי ה
מאחר , מתוך מאגרי המידע-FtsH1, FtsH2, FtsH5-FtsH9, FtsH11, FtsH12:לש ועם הרצפים
תוצאות סריקה זו מציעות כי ). לא נבדקו המיטוכונדריאלים(לכלורופלסט יים להיות מוכוונים עשושתוצריהם
בסריקה כבר . F2נזרעו ונבחנו בדור , קווים אלו הוזמנו). 3טבלה (קיימים מספר קווי מחדר למשפחה
יש פנוטיפ של כתמים בלבן וירוק לסרוגין ftsh5- וftsh2נטים הראשונית ניתן היה להבחין כי לשני מוט
(Variegation)המוטנט בגן ל. הניתן לזיהוי מיידי-ftsh2חריפה ואילו המוטנט בוריגציה סובל מ -ftsh5
אשר הצמחים בהם נבחנו ftsh8 אוftsh6 או ftsh1מוטציות השתקה אחרות כמו . קלהוריגציהסובל מ
ביקורת אינן מראות פנוטיפ השונה מצמחי ה, למיקום נקודת החדירה בתוך הגןPCRואומתו באמצעות
בנוסף נבדקה רגישותם לשטפי אור ). 34-35ראה עמוד (ftsh1- שתוארו לגבי המוטנט בניסוייםבמגוון
ואופי הגידול לתקופה של מספר שבועות בשטפים של , בזמנים קצרים µmol m-2 sec-1 1800חזקים של עד
20-250 µmol m-2 sec-1 . בכלל הבדיקות שנערכו לא נמצאו הבדלים מובהקים בין המוטנטיםftsh1 או
ftsh6או ftsh8 לבין צמחי ה-WT . נבחנו ולא נמצאו שונים מצמחי 3שאר המוטנטים המופעים בטבלה WT
WT- טיפ השונה ממאחר ולשני מוטנטים נצפה פנו. אך לא בוצע להם ניתוח גנטי מעמיק ולכן אינם מוצגים
. FtsH1, FtsH2, FtsH5, FtsH8ולאור המבנה הפילוגנטי בחרנו להתמקד בארבעת הגנים הללו בלבד

42
mRNA המוטנטים אינם צוברים
בעלי -FtsH’sהם ארבעת ה FtsH1, FtsH2, FtsH5, FtsH8 -הגנים המקודדים להראו כי אנליזות ביטוי
ftsh2-הממצאים הראשוניים הראו של .(Sinvany-Villalobo et al., 2004)ר רמת הביטוי הגבוהה ביות
פנוטיפ , בהתאמה ftsh1, ftsh8 ולשני הקרובים אליהם ,WT-השונה מ יש פנוטיפ נראה לעיןftsh5 -וגם ל
של הגן בו הם פגומים הופק mRNA-לשם אימות כי הצמחים שזוהו אכן אינם מיצרים את ה .WT- הדומה ל
בראקצית cDNA- זה שימש לייצור סך הRNA. חים בני שלושה עד ארבעה שבועות מעלי צמ-RNAכלל ה
RT. לראקצית תוצרי הראקציה נמהלו ושימשוPCR כביקורתטובוליןול עם פריימירים ספציפים לכל גן .
שלו mRNA-אינו צובר את ה ftsh1, ftsh2, ftsh5, ftsh8התוצאות מראות כי כל אחד מהמוטנטים
ניתן להסיק שאינם מכילים את החלבון mRNA-מאחר שהמוטנטים הללו אינם צוברים את ה ).9Aתמונה(
.WT גרם למופע שנראה שונה מצמחי FtsH5- או בFtsH2- רק חסר ב, יחד עם זאת. הספציפי שלהם
וכביקורת לתקינות הפריימרים , Tubulin בדוגמה שימשו הפריימרים של RNAכביקורת לקיומו של
נבדק באמצעות DNA-אי זיהום ב). שתי עמודות ימניות9Aתמונה (WT - מRNA שימש הספציפים
DNA על גבי PCR אשר כללו אינטרון בינהם שיצר בנד גבוהה יותר כאשר בוצע Tubulinהפריימרים של
).9Bתמונה (
השונים של המוטנטים RT-PCR .9תמונה
(A) ראקציית לRT-PCR לפייםפריימרים ספצישימשו- ftsh1,
ftsh2, ftsh5, ftsh8, tubulin . הפריימרים מצויינים לצדי
הפריימרים תואמים לצמחי המוטנט הנמצאים בכל . התמונה
.שורה
(B) שימשו פריימרים לטובולין ו בתור ביקורת- cDNAאו -
DNAמ -WT.
Plant \ Primers
ftsh1
ftsh2
ftsh5
ftsh8
DNA RNA
Tub
Tub
Tub
Tub
Tub
Mutant WT Mutant
B
A

43
FtsH2השפעות התפתחותיות וסביבתיות על ביטוי . ב
ftsh2 - בוריגציהאיפיון ה
הוא . -VARII זה נקרא גם ט מוטנ).10תמונה (וריגציה של פנוטיפ לגורמת FtsH2מוטציית השתקה של
. לסרוגין שנראים נורמליים מקטעים לבנים חסרי כלורופלסטים ומקטעים ירוקים בעלי כלורופלסטיםבעל
בזמן הארה בשטף אור WT- יורדים מהר יותר מאשר בFv/Fmולכן ערכי , ה אור גבושטףהוא רגיש ל
כתופעה מהווה נושא מעניין ביותר אף כי וריגציהה .(Chen et al., 2000; Takechi et al., 2000) גבוהה
מתפתחים חלק מהתאים , העניין הרב הנו בעובדה שאף שהעלה הנו בעל גנוטיפ אחיד. חורג מתחום עבודה זו
מאחר שהמוטציה שגרמה למופע זה היא מוטציה גרעינית ולא . ירוק-וחלק אחר , לבן–בפנוטיפ אחד
הבעיתיות נובעת מן . ההסבר המקובל הנו שונות פנימית, ולרוב, ה זופלסטידית קשה מאוד להסביר תופע
אך הם בעלי שני . דה ששני תאים צמודים באותה הרקמה אינם נבדלים בגנוטיפ הפלסטידי והגרעיניהעוב
.סוגים שונים של פלסטידות
ftsh2וריגציה בצמחי .10תמונה
פגומים , T-DNA ישנו FtsH2-צמחי ארבידופסיס אשר בגן ל
והם מציגים פנוטיפ וריגאנטי , FtsHביכולתם לצבור את חלבוני
ניתן להבחין בכתמים ירוקים או לבנים .VARIIהנקראה גם
.בעלים
העלה , בעוד הפסיג מתפתח בצורה נורמלית ונראה ירוק לחלוטין כי, מראהFtsH2צמחים חסרי בחינת
בו ירוקים יותר ככל שהשלב מתפתחים כשהםשאר העלים ). 11תמונה (תי הראשון כמעט לבן לחלוטין יהאמ
, וריגציה הרמת על מנת לכמת את .כלומר העלים הצעירים יותר יהיו הירוקים יותר. מאוחר יותרהם נוצרים
ראה ( ?צהוריג המידת מה :היאהספציפית ש לשאלהה מתמטית תוך התאמ, לניתוח תמונותבתוכנה שתמשנוה

44
וריגציה בין זוגות העלים משתנה רמת הה כי בתנאי גידול קבועים ומבוקרים מראתוחניה. )שיטות וחומרים
עו העלים שיופי,ט לחלוטין עם כתמים ירוקים קטניםתי הראשון לבן כמעי העלה האמבעוד. השונים בכל פרט
רמת התהליך נראה כמו תהליך הדרגתי בו ככל שהעלה נוצר מאוחר יותר. בהמשך יהיו ירוקים יותר ויותר
.ההורקה שלו גבוהה יותר
FtsH2- הפנוטיפ של עלים שונים בצמח המושתק בגן ל.11תמונה
(A) הפסיג (C) נאספו וצולמו בגודל ) 1-4( וארבעת העלים הראשונים
לבן כמעט ,תי הראשוןיהעלה האמ, בעוד הפסיג ירוק לחלוטין. דומה
.יותר ויותרהעלים הבאים אחריו מתפתחים כשהם ירוקים . לגמרי
(B) ראה חומרים (,וריגציהורקה כמדד לרמת ההכימות של רמת ה
עם סטיות , הערך המוצג מהווה ערך ממוצע של עשר חזרות . )ושיטות
.התקן שלהן
: ם עשויה להיות מוסברת בשני אופני11ההורקה ההדרגתית כפי שהיא מודגמת בתמונה
FtsH’s הביטוי של FtsH2בהעדר ) ב. (ות מאוחר יותר עולה ככל ששלב ההתפתחFtsHהביטוי של ) א(
, שויה תופעה זו להחשב תופעה המוגבלת לעלים הראשוניםיחד עם זאת ע . אחרים מתגבר באופן הדרגתי
ftsh2 - זו גודלו צמחים המוטנטים באפשרותלבדיקת . בעלים העליונים יותר על המתרחש עידהאיננה מו
בתנאים הללו ניתן . טר תאורה של יום קצר ותנאי הזנה מינרלית מיטבייםתחת מש, בתנאים שמעכבים פריחה
ניתוח תופעת ההורקה מלמד כי ככל . לגדל את הצמחים לאורך זמן מבלי שיעברו לשלב הרפרודוקטיבי שלהם
והתופעה ,רמכילים שטחים ירוקים גדולים יות) הצעירים(עליו המאוחרים , שגילו של הצמח מבוגר יותר
.ל תקופת הגידולנמשכת לכ
הנגרמת או מושפעת ממיקום העלה , מתרחש בצורה רציפהאציה בו צמצום הוריג, חותיהאופי ההתפת
ו שעמדתזותפויהה. נו להבין מהו המנגנון המאפשר קיומהי ולפיכך ניס. תכונה אנדוגניתמציע כי זו ,בשושנת

45
- כי יחולו תמורות ברמות הביטוי של הובבסיסם ההנחה, היו שני ההסברים שהוצגו בתחילת הפסקהלמבחן
FtsH’sהשונים .
במהלך ההתפתחותFtsH של mRNA-השינויים ברמות ה
או על ידי הגברת , הם הגורמים להשתנות הוריגציהFtsHכי שינויים בביטוי על מנת לבחון את ההיפותזה
נדוגנית שלהם כמנגנון התפתחותי או על ידי הגברה א, FtsH2- אחרים כפיצוי לחסר בFtsH’sהביטוי של
כולל את אלה שלmRNA- ה כמותניתוח . עלה ראשון ושני, מפסיגיםRNA על RT-PCR בוצע , עצמאי
FtsH1, FtsH2, FtsH5,ו -FtsH8כיוון שאלו היו ה -FtsH’sת התילקואיד נ היחידים שזוהו בממבר
(Sinvany-Villalobo et al., 2004; Yu et al., 2004) .קורת שימש כביTubulin , חלבון הידוע ברמתו
נמוכה בפסיגים מאשר בעלה FtsH’s- של ארבעת הmRNA - הכמות נמצא כי WTבצמחי . הקבועה בתא
של mRNA - הכמותו , FtsH2- וFtsH1- בין עלה ראשון לשני חלה בmRNA-של העלייה . הראשון
FtsH5ו FtsH8- פנל שמאלי12תמונה (יותר בעלה השניולא עולה מגיעה לרמה מירבית בעלה הראשון .(
אך , T-DNA-לבשל היותו קו מחדר , תעתיק שלו עצמו נעדרה. ftshלה בצמחי המוטנט תמונה דומה התקב
ורמות , יותר בעלה הראשון לעומת הפסיגו גבוהות היFtsH8- ו,FtsH1, FtsH5 של mRNA-כמות ה
היו mRNA’s - כל שלשת ה,יתרה מכך). ניי פנל ימ12 תמונה(גבוהות אלו נשארו או ירדו בעלה השני
).12 השוואת פנל ימיני ושמאלי בתמונה (WT- לעומת ה-FtsH2בכמות גבוהה יותר במוטנט חסר
התפתחות הצמח מצטבר עם FtsHשל mRNA -ה .12תמונה
.ftsh2ובמוטנט WT - ב
, (2) ושני, (1) עלה ראשון , (C) בודד מפסיגים RNA-כלל ה
.FtsH2ומצמחי מוטנט חסרי , )פנל שמאלי (WTמצמחי
ותוצרי , RT-PCRראקציית באמצעות רמת התעתיק נבחנה
.ל אגרוז שנצבע באתדיום ברומיד'הראקציה הופרדו על ג
2ftsh WT

46
ה עול FtsH’sשיעתוק שלת הרמ, במערכת הטבעית: יחדימותית מתקיוו האפשר כי שתיסיקן ניתן להאמכ
FtsH’s של mRNA - מפעיל מנגנון פיצוי שמוביל לעליה ברמת ה-FtsH2וחסר ב, עם השלב ההתפתחותי
.ביה אינם ידועיםי ומרכזומשוב מערכת . אחרים
במהלך ההתפתחותFtsHים ברמת החלבון ישינו
. WT (Chen et al., 2000) לעומת צמחיFtsH הראו כי הם צוברים פחות חלבוני -FtsH2צמחי מוטנט ב
FtsHארבעה נוגדנים שונים כנגד ב השתמשנו בה . Western-blotשר תצפית זו בוצעה אנליזת על מנת לא
איזור זה שמור ביותר בתוך .SRH-וכוון אל עבר ה, יטתהראשון הוכן כנגד פפטיד סינ ).13Aתמונה (
י מביטוי הוכנו כנגד חלבון נק-FtsH5 ו,FtsH1, FtsH2הנוגדנים כנגד . ATPase-המשפחה ונמצא באתר ה
מאחר ובמשפחה בכלל ובין הזוגות הקרובים בפרט רמת הזהות ברצף חומצות האמינו . E. coliיתר בחיידקי
מזהה את כלל המשפחה באופן זהה או דומה SRH נוגדן כנגד : ניתן היה לזהות את התגובות הבאות ,גבוהה
באפיניות מזוהים FtsH5- וFtsH1 מזהה את כל המשפחה כאשר FtsH1הנוגדן כנגד החלבון המלא . מאוד
היא הגבוהה FtsH2מאחר והכמות הנצברת של . מזוהים באפיניות נמוכה FtsH8 וגם FtsH2-ו, גבוהה
ה של הסיגנל המתקבל נראת כמעט זהה העוצמ)Sinvany-Villalobo et al., 2004(ביותר לעומת האחרים
על הממברנה יכולים להתקבל . יחדFtsH1- וFtsH5לעומת הסיגנל של , יחדFtsH8 - וFtsH2במקרה של
הנוגדן . FtsH8- ו FtsH2 והתחתון את FtsH1- וFtsH5העליון כולל את , kDa 1.5-2-שני בנדים השונים ב
חומצות האמינו 450-הכולל כהחלק המסיס של החלבון הוכנו כנגד FtsH2 והנוגדן כנגד FtsH5כנגד
כל זוג FtsH8- וFtsH2 או עם FtsH5- וFtsH1הם עשויים להגיב עם . מהקצה הקרבוקסלי של החלבון
). בחומרים ושיטות2ראה טבלה (. )Sakamoto et al., 2003( אך ללא תגובה בין הזוגות ,בהתאמה לנוגדן
כל ארבעת הנוגדנים הראו ירידה משמעותית בכמות החלבון שהם , WT לעומת צמחי ftsh2 בצמחי
יצרנו עקומי כיול לעוצמת , על מנת לקבוע כמותית מהי הירידה בצבירת החלבון. )13Aתמונה (מזהים
התוצאות הראו כי .341ImageJ - NIH. ותתמונ לניתוח תוכנהתוך שימוש ב, הסיגנל המתקבל בראקציה
לכמות החלבון -Western blotניתן היה לזהות תגובה לינארית ב, כלורופיל0.2-6.5μgבתחום שבין
. WTמכמותו בצמחי 50%- היא כ FtsH כמות החלבון של ftsh2מכאן חושב כי בצמחי ). 13Bתמונה (
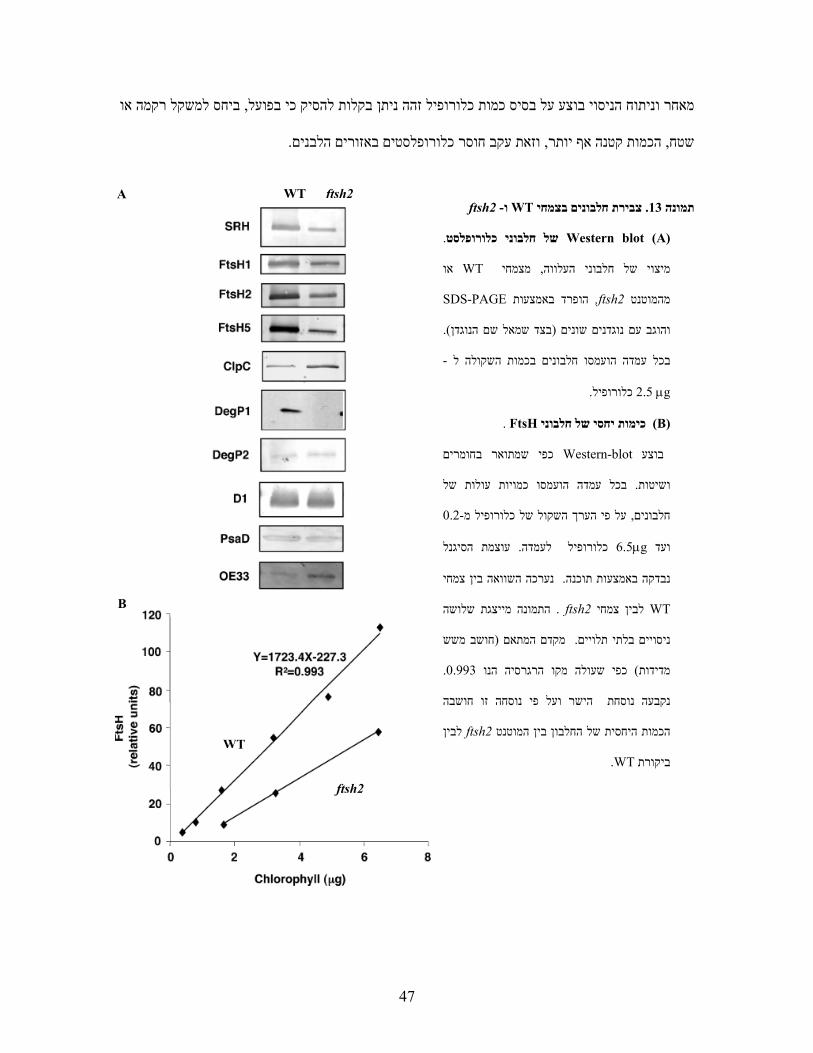
47
ביחס למשקל רקמה או , מאחר וניתוח הניסוי בוצע על בסיס כמות כלורופיל זהה ניתן בקלות להסיק כי בפועל
.וזאת עקב חוסר כלורופלסטים באזורים הלבנים, הכמות קטנה אף יותר, שטח
ftsh2- ו WTצבירת חלבונים בצמחי. 13תמונה
(A) Western blotשל חלבוני כלורופלסט .
או WTמצמחי , מיצוי של חלבוני העלווה
SDS-PAGE הופרד באמצעות ,ftsh2מהמוטנט
. )בצד שמאל שם הנוגדן(והוגב עם נוגדנים שונים
- בכל עמדה הועמסו חלבונים בכמות השקולה ל
2.5 μgכלורופיל .
(B) כימות יחסי של חלבוניFtsH .
חומרים כפי שמתואר בWestern-blot בוצע
יות עולות של ובכל עמדה הועמסו כמ. ושיטות
0.2-על פי הערך השקול של כלורופיל מ, חלבונים
עוצמת הסיגנל . כלורופיל לעמדה6.5μgועד
נערכה השוואה בין צמחי .נבדקה באמצעות תוכנה
WT לבין צמחי ftsh2 . התמונה מייצגת שלושה
חושב משש (דם המתאם מק. ניסויים בלתי תלויים
. 0.993כפי שעולה מקו הרגרסיה הנו ) מדידות
בהנקבעה נוסחת הישר ועל פי נוסחה זו חוש
לבין ftsh2הכמות היחסית של החלבון בין המוטנט
.WTביקורת
B
A WT ftsh2
ftsh2
WT

48
ני מראה כי חלבו, לפי כמות כלורופילאחרי הטענה, ונים כלורופלסטיים אחריםניתוח איכותי של כמות חלב
כמו גם הפרוטאזה המסיסה , ממערכות אור שתיים ואחת בהתאמה-PsaD וD1ית כמו תטהמערכת הפוטוסינ
DegP2 ,חלה עליה בצבירת חלבוני לעומת זאת. אינם משתנים מבחינה כמותית ClpC ובחלבון OE33 ,
בין FtsHני האם משתנה כמות חלבו-השאלה .)13Aתמונה ( DegP1וירידה בכמות הפרוטאזה הלומינלית
.נשארה בעינה, ם לשלב ההתפתחותי שלהםאהעלים השונים בהת
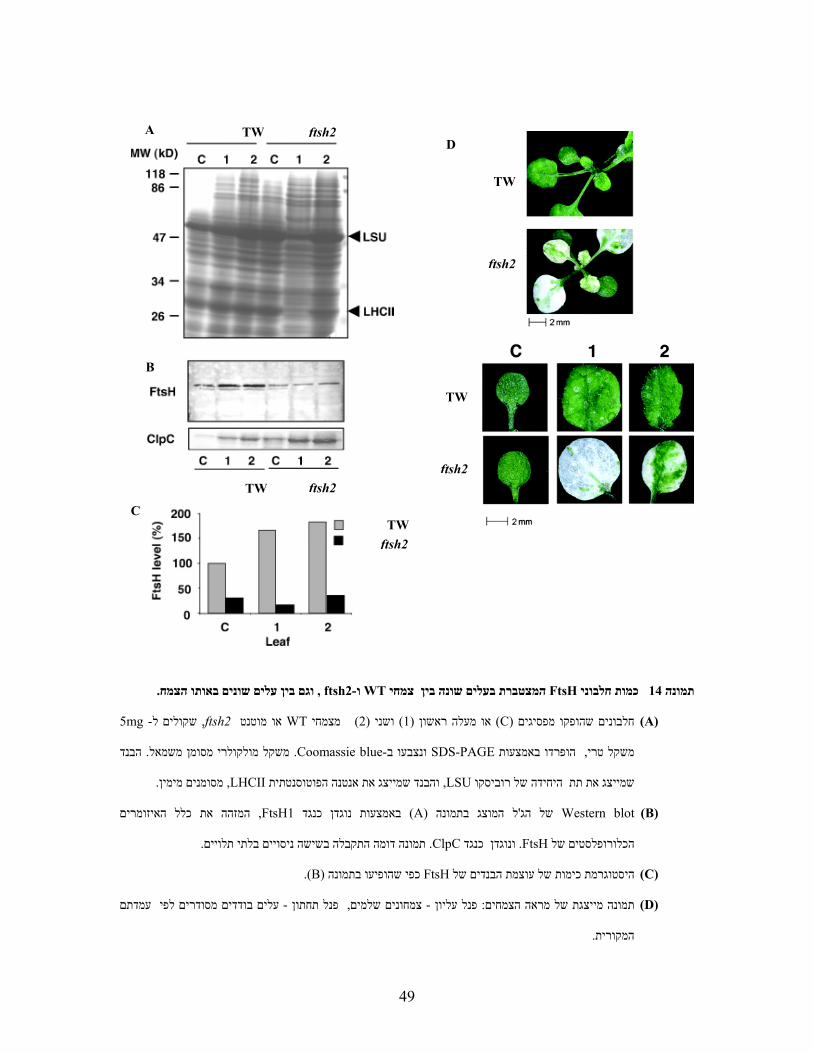
תמונה (ftsh2- וWTמעלה ראשון ושני בשושנת העלים של צמחי , מפסיגיםםהופקו חלבוני, לפיכך
14D .(יוצא . ודפוס החלבונים נראה דומה בין הדוגמאות, מות החלבונים בניסוי זה נורמלה לפי משקל טריכ
כמו תת ,חלבונים כלורופלסטיםבעלה זה . שהיה לבן כמעט לגמרי ftsh2ל הוא העלה הראשון של מן הכל
). 14Aתמונה ( כמעט לגמרי חסרים, LHCIIית טתהאנטנה הפוטוסינו, LSUהגדולה של רוביסקו היחידה
תפתחות נצבר תוך כדי ההWT בצמחי FtsH מציע כי (FtsH) באמצעות הנוגדן FtsHניתוח כמות החלבון
תופעה דומה נצפתה כאשר השתמשתי בנוגדנים כנגד, באופן מעניין. (14B, C)בצורה הדרגתית תמונה
ClpC תת היחידה הרגולטורית של הפרוטאזה הסטרומלית ClpP ,ויתכן כי זו תופעה משותפת לפרוטאזות.
כמות, WT- מאשר בFtsHני ראשית היו פחות חלבו. התופעות שנצפו היו מורכבות יותר ftsh2בצמחי
,יחד עם זאת ). 14Cתמונה (-WT לעומת כמותם בפסיגי ה70%- החלבונים בפסיגים לדוגמה היתה נמוכה בכ
העלה . תייםי אשר עולה בהדרגה בעלים האמFtsHהיתה התאמה בין עוצמת ההורקה לבין כמות חלבוני
זהו אותו עלה שמציג רמות גבוהות ( FtsHתי הראשון הכיל את הכמות הנמוכה ביותר של חלבוני יהאמ
בתנאים . מעלה זה והלאה הכמות עולה באופן הדרגתי). 12בתמונה 2 מלבד FtsH’s-של כל ה mRNAשל
ירוקים לגמרי, לכאורה,הצמח המוטנט יכול אף לפתח עלים הנראים לאורך זמן של כחצי שנה,מעכבי פריחה
. פסיגים צעירים מאשר FtsHיותר 50%-עלים אלו מכילים כ. יםושטחם עשוי לכלול מקטעים לבנים מיזערי
ירידה : ניתן להציע לכך שני הסברים. FtsHעלה הראשון במוטנט מכיל את הכמות הנמוכה ביותר של ה
, היא הגורםסינתזה קשה להניח כי ירידה בmRNA-מאחר וחלה עליה ברמת ה. או עליה בפירוק,סינתזהב
. מבוצעת מחוץ לאורגנלההסינתזמה גם שה
49
. וגם בין עלים שונים באותו הצמח , ftsh2- וWTצמחי המצטברת בעלים שונה בין FtsHכמות חלבוני 14תמונה
)A( חלבונים שהופקו מפסיגים(C) או מעלה ראשון )מצמחי ) 2(ושני ) 1WTאו מוטנט ftsh2 ,5-קולים לשmg
הבנד . מסומן משמאל משקל מולקולרי.Coomassie blue- ונצבעו בSDS-PAGE באמצעות הופרדו ,משקל טרי
. מסומנים מימין,LHCIIוהבנד שמייצג את אנטנה הפוטוסנטתית , LSUהיחידה של רוביסקו שמייצג את תת
)B( Western blotל המוצג בתמונה ' של הג(A)כנגד ן באמצעות נוגד FtsH1, המזהה את כלל האיזומרים
.שישה ניסויים בלתי תלוייםב דומה התקבלהתמונה .ClpCכנגד נוגדן ו. FtsHכלורופלסטים של ה
)C( היסטוגרמת כימות של עוצמת הבנדים שלFtsH כפי שהופיעו בתמונה (B).
)D( עלים בודדים מסודרים לפי עמדתם - פנל תחתון , צמחונים שלמים- פנל עליון : תמונה מייצגת של מראה הצמחים
.המקורית
WT
WT
WT
ftsh2
ftsh2ftsh2
ftsh2
ftsh2
WT
WT
D A
B
C

50
הצטברות של צמצםאשר חוסר בתת יחידה אחת שלו מ, הוא הטרוקומפלקס בכלורופלסט FtsH כמו כן
השוואה בין כמות חלבוני לכן נערכה .(Chen et al., 2000; Yu et al., 2004) היחידות האחרות -תת
FtsH בפסיג ובעלה הראשון של ftsh2 ושלWT . תוצרי פירוק אפשריים במשקלים מולקולרים נמוכים יותר
התהליך שמתרחש מכאן ניתן להניח כי ).15תמונה (נצפו בכמות הרבה ביותר בעלה הראשון של המוטנט
אולי עקב כמויות ( לא צלחו ספקטרומטרית מאסות לאמת תוצאה זאת באמצעות נסיונות.הוא אכן פירוק
).קטנות מידי או מיסוך על ידי חלבונים רבים אחרים
ftsh2 ושל WT בפסיגים ובעלים ראשונים של FtsH תוצרי פירוק של צבירת .15תמונה
גמאות שהוטענו הד .WT או ftsh2 מצמחי (C)סיג או מפ(1) של חלבונים שהופקו מעלה ראשון Western Blotאנליזת
תוצר פירוק , אך הוא מזהה את כלל האיזוזימיםFtsH1הנוגדן ששימש יוצר כנגד החלבון השלם , משקל טרי15mg-שקולות ל
, דומהר על מנת להגיע לפיתוח מהביקורת אך זמן פיתוח התמונה היה ארוך יותFtsHהמונטט מכיל פחות (*-אפשרי מסומן ב
).לפיכך אין להסיק על הבנדים העליונים
FtsH - וכמות הוריגציה האור על רמת ההשפעת
-כל ה. (מספר מחקרים בהוצעה שנפגעה משטף אור גבוה PSIIשל במנגנון תיקון FtsHמעורבותו של
Refהבאים ()(Lindahl et al., 2000; Bailey et al., 2002; Silva et al., 2003; Cheregi et al.,
2005; Nixon et al., 2005)בקצרה מחקרים אלו הראו יכולת פירוק של חלבון PSII-D1 במערכות שונות
אי לכך יתכן כי . לעקות אור מגוונותFtsHורגישויות שונות של מוטנטים הפגומים בחלבוני , FtsHעל ידי
והריסת הכלורופיל רעשויים המקטעים הלבנים במוטנטים נגרמים מהלבנה עקב עקת או
1C ftsh2
1C WT
MW (kDa)
FtsH
FtsH*
86
47
34

51
(Photobleaching).הסבר אלטרנטיבי עשוי להיות כי FtsH מעורב בתהליכי היצירה של התילקואידים ,
על מנת לבחון האם הסקטורים הלבנים . )Chen et al., 2000; Takechi et al., 2000( בעברכפי שהוצע
אם אכן מדובר בתופעה . במשטרי תאורה שוניםוריגציהאפשר לבחון את רמת ה, נגרמים כתוצאה מעקת אור
, וריגציהניתן לצפות כי תהיה התאמה חיובית בין עוצמת האור לבין שיעור ה, הנגרמת מעודף אורשל הלבנה
אם ההלבנה איננה . כך שעור האזורים הלבנים יהיה גדול יותרככל שעוצמת האור תהיה חזקה יותר, כלומר
.וריגציהאזי גדילה בתנאי הארה חזקים יותר לא תשפיע על רמת ה, Photobleachingתוצאה של
בשלושה משטרי תאורה של ftsh2- וWT על מנת לבחון אפשרויות אלו גודלו צמחי ארבידופסיס
, 100 ,50 µmol m-2 sec-1 כי צמחינמצא. כמות הכלורופיל ברקמההכמדד לורגציה שימש. 300- ו WT
מאשר צמחים שגדלו , פחות כלורופיל למשקל רקמה טריה 35%- הגדלים בעוצמת אור גבוהה צברו כ
ftsh2 ציין כי צמחי המוטנטל יש . פחות40%- צמחי המוטנט באותם התנאים צברו כ. בעוצמת אור נמוכה
המגמה הכללית של אובדן הכלורופיל בעוצמות אור גבוהות היתה משותפת . ראשמ מכילים פחות כלורופיל
הצמחים שגדלו בעצמה הגבוהה נראו כהים יותר בגלל צבירת פיגמנטים ). 16Bתמונה (למוטנט ולביקורת
צמחים שגדלו בעוצמת אור בינונית צברו כמות בינונית , בדומה לכך. )16Aתמונה (אחרים כמו אנטוציאנין
.וריגציהם מהותיים בדפוס הילא זוהו שינוי. כלורופילשל
Photobleaching- מ הוא הרחבת השטח הלבן שנגרמת כתוצאהגציהיהגורם לור מאחר שהאפשרות כי
לאורך וריגציה אחרי עלים בודדים בעלי לעקב בחרנו . על ידי צבירה הדרגתית של נזקי קרינה לא נשללה
תלך ותגבר ככל שגילו וריגציהצפוי כי רמת ה, גציהירגתית היא הגורם לוראם צבירה הד. חודש ימים
התמונה נותחה לבדיקת מספר הפיקסלים ליחידת שטח , העלים סומנו וצולמו. הכרונולוגי של העלה יעלה
מתוצאות שני הניסויים הללו ). 16Cתמונה ( לאורך תקופת הניסוי וריגציהקבוע ולא נמצאה עליה ברמת ה
ינות של הכלורופלסט שנצפתה נובעת מבעיה בתהליך היצירה או ההתמוריגציהלהסיק כי תופעת האפשר
כי בתאי , הראו באמצעות מיקרוסקופיה אלקטרונית חודרתSakamoto et al., (2003) . והתילקואידים
ftsh2-הרקמה הלבנה ישנם מספר דומה של פרופלסטידות לעומת הכלורופלסטים של הרקמה הירוקה הן ב
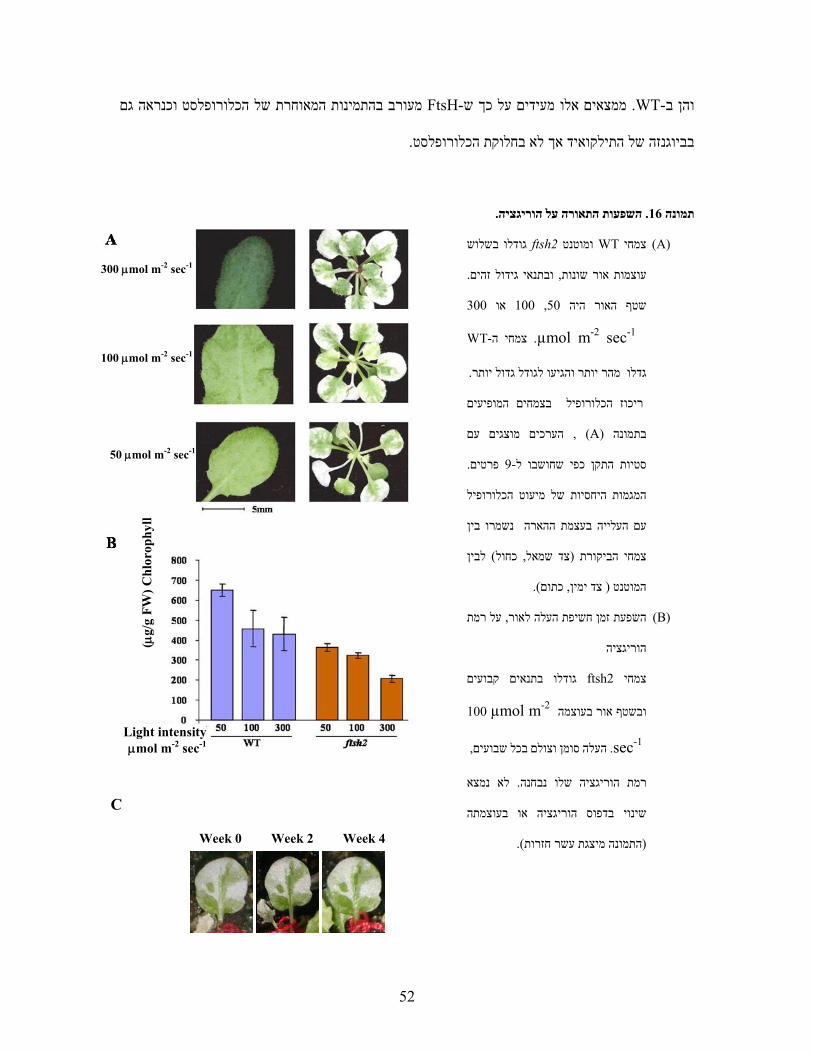
52
וכנראה גם מעורב בהתמינות המאוחרת של הכלורופלסטFtsH-ממצאים אלו מעידים על כך ש. WT- והן ב
.בביוגנזה של התילקואיד אך לא בחלוקת הכלורופלסט
.וריגציה השפעות התאורה על ה.16תמונה
)A( צמחי WT ומוטנטftsh2 גודלו בשלוש
. ובתנאי גידול זהים, עוצמות אור שונות
300 או 100, 50ה יור ה האשטף
µmol m-2 sec-1 .צמחי ה -WT
.גדלו מהר יותר והגיעו לגודל גדול יותר
ריכוז הכלורופיל בצמחים המופיעים
הערכים מוצגים עם , (A)בתמונה
. פרטים9-סטיות התקן כפי שחושבו ל
המגמות היחסיות של מיעוט הכלורופיל
עם העלייה בעצמת ההארה נשמרו בין
לבין ) כחול, צד שמאל(חי הביקורת צמ
).כתום, צד ימין( המוטנט
)B( על רמת , השפעת זמן חשיפת העלה לאור
הוריגציה
גודלו בתנאים קבועים ftsh2צמחי
µmol m-2 100ובשטף אור בעוצמה
sec-1 .יםהעלה סומן וצולם בכל שבוע,
לא נמצא . שלו נבחנהוריגציהרמת ה
יגציה או בעוצמתה ורשינוי בדפוס ה
).תמונה מיצגת עשר חזרותה(
Week 0 Week 2 Week 4
C
50 μmol m-2 sec-1
Light intensityμmol m-2 sec-1
Chl
orop
hyll
(μ
g/g
FW)
100 μmol m-2 sec-1
300 μmol m-2 sec-1

53
FtsHרגיש בעצמו לשטפי קרינה גבוהים
FtsH הראו כי רמות התעתיק של ) (mRNA Sinvany-Villalobo et al., 2004 מבחני ביטוי של
קי זהתאמה של מערכת התיקון מנממצא זה עשוי להעיד על כושר . עולות לאחר חשיפה לשטפי אור גבוהים
mRNA- בעל מנת לבחון האם עליה . הפועלות במערכת זוFtsH של על ידי הגדלת כמות היחידות, קרינה
למשך שלוש µmol m-2 sec-1 1300 לשטף קרינה של WTנחשפו צמחי , באה לידי ביטוי ברמת החלבון
צא הבדל בכמות בתנאים הללו לא נמ. (Western blot) וכמות החלבון נמדדה באמצעות נוגדנים ,שעות
. FtsHכאשר בוצע הניסוי למשך שעה אחת בלבד חלה ירידה בכמות חלבוני , להפתעתנוFtsH.-חלבוני ה
יורדת בשיעור חד במהלך הדקות הראשונות של החשיפה FtsH-בדיקה רציפה למשך כשעה מראה כי כמות ה
בהמשך מתחיל להצטבר .FtsH מחלבוני 60%-כעבור כשלושים דקות נשארו רק כ. לאור בעוצמה גבוהה
מדידות רציפות של הפוטנציאל . 17A)תמונה ( מערכו ההתחלתי 75%החלבון ובתוך כשעה חוזר עד לכדי
, בעלים החשופים לאור בעוצמה גבוהה)Fv/Fm(באמצעות פלורסנציה של כלורופיל , PSIIהמקסימלי של
התגובה , לחצי שעה0בפרק הזמן שבין . אריתלינאיננה מראות כי הירידה בערכי הפוטנציאל איננה קבועה ו
). 17Bתמונה ( בהמשך הירידה מתמתנת .מהיר ביותרהחשת בקצב תרמ
ונראה כי קיימת , אינה מספיקה כדי להסביר תופעה שכזו) כמו אור(גורמית קבועה -עקה פיזיקאלית חד
FtsHחלבוני ת שאין מדד לאיכות למרו. PSII לבין אובדן הפעילות של FtsH התאמה בין אובדן הפרוטאזה
,Adam)הוצע בעבר כפי שה, ניתן להניח בבטחה כי החלבונים שמסולקים מהמערכת אינם תקינים, בניסוי זה
, בביולוגיהות בין פגיעה בחלבון לבין פירוקו אינה תופעה חריגהדד הפרש של דקות בו,יחד עם זאת . (1996
פגיע בחלבון מסויים ועד לשלב שהוא מסולק אחרי שזוהה כחלבון שכן זמן עשוי לחלוף בין הרגע שמתרחשת
. הכולל סינתזה מחד ודגרדציה מאידך,steady stateהקינטיקה המוצגת מייצגת מצב של . פגום שיש לסלקו
אלא , כי עקת האור הנמדדת בצמח נוצרת לא רק מנזק מצטבר למערכות האור מתוצאות אלה אפשר להסיק
.FtsHנגנון התיקון תוך אובדן של גם מפגיעה כלשהי במ
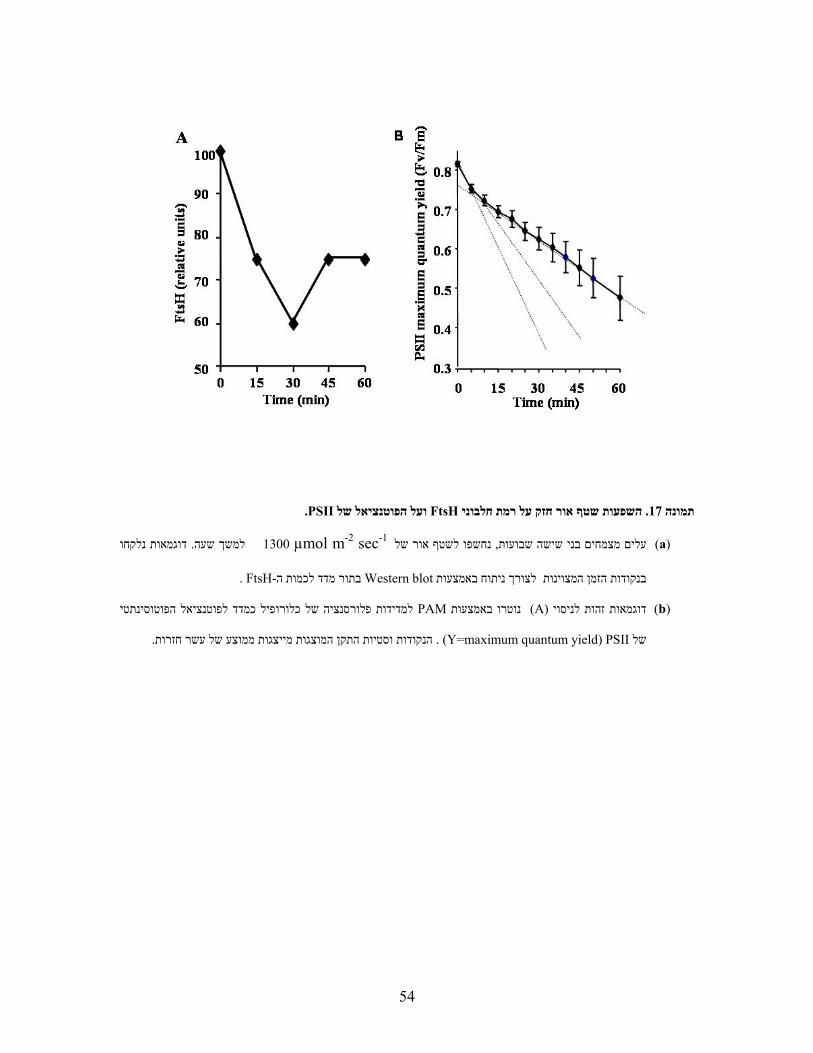
54
. PSII ועל הפוטנציאל שלFtsHהשפעות שטף אור חזק על רמת חלבוני . 17תמונה
)a( 1300 נחשפו לשטף אור של, עלים מצמחים בני שישה שבועות µmol m-2 sec-1 דוגמאות נלקחו . למשך שעה
. -FtsH בתור מדד לכמות הWestern blot לצורך ניתוח באמצעות בנקודות הזמן המצוינות
)b( דוגמאות זהות לניסוי(A)רו באמצעות וט נPAM למדידות פלורסנציה של כלורופיל כמדד לפוטנציאל הפוטוסינתטי
.הנקודות וסטיות התקן המוצגות מייצגות ממוצע של עשר חזרות . PSII (Y=maximum quantum yield)של

55
הכלורופלסטית FtsHהשונות במשפחת יחסים בין תתי היחידות .ג
ftsh2 חמור יותר משל ftsh1 ftsh2 הכפולהמוטנטשל הפנוטיפ
הם בעלי פנוטיפ של -ftsh2 בעוד שקווי מחדר ב,מתפתח ומתפקד בצורה נורמלית ftsh1-קו מחדר ב
, כפי שהראנו. (Chen et al., 2000; Takechi et al., 2000; Sakamoto et al., 2003)וריגציה חמורה
והדבר עשוי לרמז על מנגנון פיצוי WT- גבוהה יותר מאשר בFtsH1 של mRNA- כמות ה ftsh2במוטנט
או שהם בעלי , השונים בכלורופלסטFtsHעל מנת לבחון האם ישנה חפיפה בתפקידיהם של חלבוני . הדדי
F1דור . יו הומוזיגוטים למוטציהדור ההורים ה. בוצעה הכלאה רציפרוקלית בין המוטנטים, תפקידים שונים
נמצאה אוכלוסיה מתפצלת שקרוב לרבע מהצמחים בה הראו מופע של F2אך בדור , -WTלא נראה שונה מה
. ftsh2-וריגציה בדומה ל
.ניתוח הפרופיל הגנטי: ftsh2 ו ftsh1 טים הומוזיגוטים של נהכלאה בין מוט. 18תמונה
)A( התוצאות הפרדה של תוצריPCR- בתמונה הסכמטיות של הגניםימרים המופיעים במפותישבוצעו עם הפר (B) .
)B( 1: בוצעו עם הפריימרים הבאים1-4הראקציות המסומנות . החצים מצביעים על כיוון הפריימר - A+B ,2-B+C ,3-
D+E ,4-D+F .

56
כילו את מחדר כלומר ה( BASTAקוטל העשבים ים ל עמידוורובם הי WT- כמו השאר הצמחים נראו
התפצלות שכזו . ומתוBASTA - מתוכם היו רגישים ל1/16- כרק ). הכולל גן לעמידותT-DNA-ה
הופק מהצמחים העמידים ושימש DNA. הינה התפצלות מנדלית האופיינית לשני גנים בלתי תלויים
) 18Aמונה ת(בניתוח זה נמצאו צמחים הומוזיגוטים לשתי המוטציות . לניתוח הפרופיל הגנטי שלהם
אך הגיבו עם פריימר אחד ftsh2 או ftsh1צמחים אלו לא הגיבו עם פריימרים המתאימים לקצוות הגנים
כל הצמחים העמידים הכילו ). 18A-Bתמונה ( ftsh2 או ftsh1 עם פריימר שני של -T-DNAשל ה
ו ונבחנו מול צמחי הצמחים ההומוזיגוטים בשתי המוטציות גודל. מחדר לפחות באחד משני הגנים הללו
. גדל לאט יותרWT ,ftsh2 נראה וגדל כמו ftsh1בחינת דפוס הגידול מראה כי בעוד . WT -האם וה
ftsh1 ftsh2לכאורה חלה החמרה בפנוטיפ . והוא בעל רמות וריגציה גבוהות יותר, גדל לאט אף יותר
.)19 תמונה( ftsh2שנצפה במוטנט
במוטנט כפול ftsh2 ציה שלוריג החמרת ה.19תמונה
ftsh1ftsh2
, WT ,fsth1 ,ftsh2, : צמחי ארבידופסיס מהקוים
ftsh1ftsh2גודלו בתנאי גידול אופטימלים של יום קצר ,
הצמחים צולמו בגיל חמישה . במשטרי השקיה ודישון מיטביים
, כתמים לבנים וירוקים: מראה וריגציהftsh2המוטנט . שבועות
מוטנט כפול . -WTראה דומה ל נftsh1ואילו המוטנט
.הומוזיגוטי הוא בעל וריגציה חמורה ונראה לבן יותר
,מימין ftsh2 בחלק התחתון של התמונה שני פרטים
ftsh2ftsh1 ההגדלה המצויינת בגוף התמונה מתאימה .משמאל
.לשתי תמונות אלו בלבד
WT
ftsh1
ftsh2
ftsh1 ftsh2
ftsh1 ftsh2 X5
ftsh2 X4
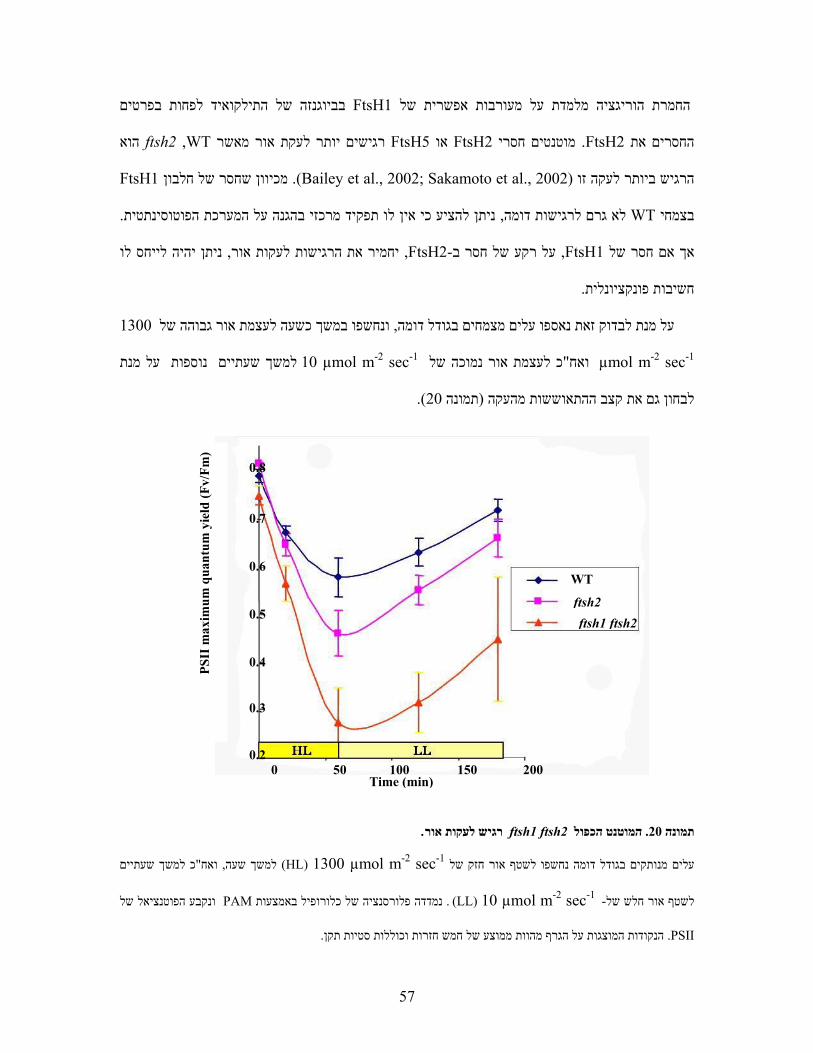
57
ביוגנזה של התילקואיד לפחות בפרטים בFtsH1 החמרת הוריגציה מלמדת על מעורבות אפשרית של
הוא WT ,ftsh2 רגישים יותר לעקת אור מאשר FtsH5 או FtsH2מוטנטים חסרי. FtsH2החסרים את
FtsH1מכיוון שחסר של חלבון . )Bailey et al., 2002; Sakamoto et al., 2002(הרגיש ביותר לעקה זו
. ניתן להציע כי אין לו תפקיד מרכזי בהגנה על המערכת הפוטוסינתטית, לא גרם לרגישות דומהWTבצמחי
ניתן יהיה לייחס לו , ישות לעקות אוריחמיר את הרג, -FtsH2על רקע של חסר ב, FtsH1אך אם חסר של
.פונקציונלית חשיבות
1300ונחשפו במשך כשעה לעצמת אור גבוהה של , על מנת לבדוק זאת נאספו עלים מצמחים בגודל דומה
µmol m-2 sec-1 10כ לעצמת אור נמוכה של "ואח µmol m-2 sec-1 למשך שעתיים נוספות על מנת
).20תמונה (אוששות מהעקה לבחון גם את קצב ההת
. רגיש לעקות אור ftsh2 ftsh1המוטנט הכפול . 20תמונה
כ למשך שעתיים "ואח, למשך שעהµmol m-2 sec-1 (HL) 1300 עלים מנותקים בגודל דומה נחשפו לשטף אור חזק של
ונקבע הפוטנציאל של PAMלורופיל באמצעות נמדדה פלורסנציה של כ. 10 µmol m-2 sec-1 (LL) -לשטף אור חלש של
PSII . של חמש חזרות וכוללות סטיות תקןממוצעהנקודות המוצגות על הגרף מהוות .
0.3
0.2
0.4
0.5
0.6
0.7
0.8
50 150100 2000 Time (min)
WT
ftsh2ftsh2 ftsh1
PSII
max
imum
qua
ntum
yie
ld (F
v/Fm
)

58
של והמוטנט הכפול , כפי שדווח בעבר, לעקות אורWT-מ רגיש יותרftsh2הממצאים מראים כי המוטנט
ftsh1 ftsh2 אליהן ניתן להגיע מבדיקת המוטנטים הבודדיםבניגוד למסקנות). 20תמונה (רגיש אף יותר ,
ככל , יש תפקיד פזיולוגי והוא מעורב הן בהגנה מפני נזקי קרינה-FtsH1בדיקת המוטנט הכפול מעלה כי ל
עולה , לאור הממצאים שהוצגו עד כה. והן בביוגנזה או התמיינות של כלורופלסטים, הנראה במערכת התיקון
על מנת . והאם פעילותם חופפת או משלימה, השונים בכלורופלסט FtsHבוני מהם היחסים שבין חל: השאלה
.לענות על שאלה זו נערכו הכלאות רציפרוקליות בין מספר קווי מחדר
באופן נורמלי מתפתח ומתפקד ftsh1 ftsh8המוטנט
ftsh1 ו -ftsh8 ד מהם בנפרד קווי מחדר בכל אח. 1בארבידופסיס מקודדים שניהם בכרומוזום מספר
מבחינת התפתחות ותפקוד , התפתחו ותפקדו באופן דומה לצמחי הביקורת מבחינת המראה וקצב הגידול
על מנת לבחון האם . )Sakamoto et al., 2003(הכלורופלסטים ומבחינת הרגישות לעצמות אור חזקות
מכיוון . הוכלאו שני קווי המחדר ביניהם, זו כלפי זו(Synergism)לשתי מוטציות אלו יש מימד של אגבור
נמצא כי .ששניהם מקודדים באותו הכרומוזום בוצע ניתוח ממוחשב למבנה הכרומוזום על פי סמנים קרובים
תוצרי ההכלאה הראשונים . ע הכלאה קלאסיתולכן אפשר היה לבצ, שיעור התאחיזה ביניהם נמוך מאוד
בצמחי ).21A-Bתמונה ( לשם זיהוי מוטנט כפול הומוזיגוט למוטציות PCRודורות ההמשך נותחו באמצעות
WTראקציית ה -PCR בניגוד . של כל אחד מהגנים-’3 ו’5 הצליחה רק עם פריימרים שמתאימים לקצוות
פריימר המתאים לגבול . רים לא הצליחו להגביר את המקטעבמוטנט הומוזיגוט כפול אותם הפריימ, לכך
בחינת האוכלוסיה בדור . יחד עם אחד הפריימרים של הגן יצרו הגברה וזוהה בנד-T-DNAהפנימי של ה
F3 ,הן בPCR-אך צמחי המוטנט הכפול לא נמצאו שונים מצמחי . תמכו בזיהוי זה, והן במבחני עמידותWT ,
).21Dתמונה ( והן ברגישות לעקת אור )21Cתמונה ( הן במראה
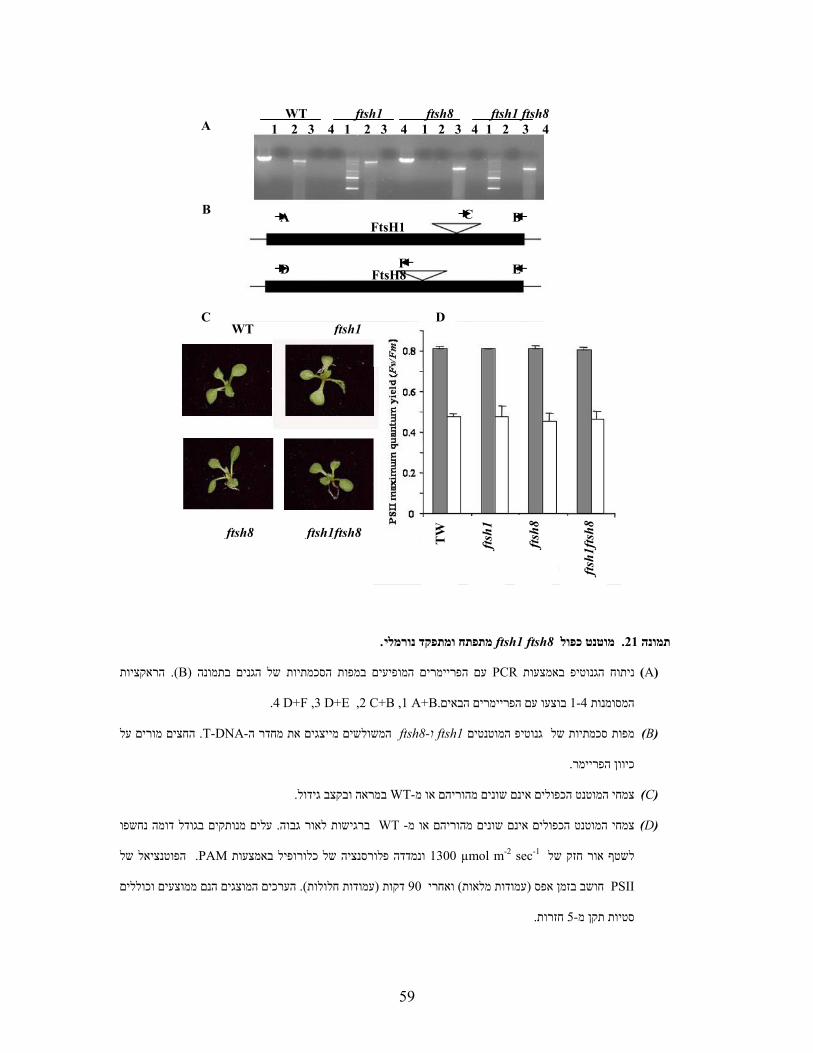
59
.מתפתח ומתפקד נורמלי ftsh1 ftsh8מוטנט כפול . 21 תמונה
)A( ניתוח הגנוטיפ באמצעותPCR בתמונה הסכמתיות של הגניםים המופיעים במפותפריימרעם ה (B) . הראקציות
.A+B ,2 C+B ,3 D+E ,4 D+F 1. בוצעו עם הפריימרים הבאים1-4המסומנות
)B( מפות סכמתיות של גנוטיפ המוטנטיםftsh1ו -ftsh8המשולשים מייצגים את מחדר ה -T-DNA. החצים מורים על
.כיוון הפריימר
)C( צמחי המוטנט הכפולים אינם שונים מהוריהם או מ -WTבמראה ובקצב גידול .
)D( צמחי המוטנט הכפולים אינם שונים מהוריהם או מ-WT מנותקים בגודל דומה נחשפו עלים . ברגישות לאור גבוה
הפוטנציאל של . PAM ונמדדה פלורסנציה של כלורופיל באמצעות µmol m-2 sec-1 1300לשטף אור חזק של
PSII הערכים המוצגים הנם ממוצעים וכוללים ). עמודות חלולות( דקות 90ואחרי ) עמודות מלאות( חושב בזמן אפס
. חזרות5- סטיות תקן מ
WT 1 2 3 4
ftsh1 1 2 3 4
ftsh8 1 2 3 4
ftsh1 ftsh8 1 2 3 4 A
B
C D
CA
D
B
EF
WT ftsh1
ftsh8 ftsh1ftsh8
FtsH1
FtsH8
WT fts
h1 fts
h8
ftsh1
ftsh8

60
סוגל להשלים מחזור חיים מftsh5 ftsh2המוטנט
Chen et al., 2000; Takechi et( מלווה בוריגציה וברגישות לעקות אור ftsh5 או של ftsh2השתקה של
al., 2000; Bailey et al., 2002; Sakamoto et al., 2002( .הכלאה של מוטנט ב-ftsh5 שמקורו בתרבית
בעל וריגציה מוגברת ומעוכב , והוא כצפוי יצר את המוטנט שתוארftsh2 בגן T-DNAעם מוטנט , רקמה
.)Sakamoto et al., 2002(נפגמה יכולתו להשלים את מחזור חייו בגלל הפלת פרחים , בנוסף. בגידולו
ftsh5 נקרא גם VARI בשל היותו בעל מופע וריגנטי חלש )Sakamoto et al., 2002( . ההסבר המקובל
, על פי הסבר זה". חלש" היא נמוכה ולכן חסרונו גורם לפנוטיפ FtsH5לפנוטיפ זה הוא כי כמות החלבון
על מנת לקבוע אם גורם הכמות . WT- למוטנט בו פנוטיפ השונה מ קטנה אף יותר ולכן איןFtsH1כמותו של
ftsh2 ftsh1 הוא הגורם היחיד שמשפיע על הוריגציה רצינו להשוות בין מוטנט כפול של FtsHשל חלבוני
תוצאות ההכלאה נותחו . באופן רציפרוקליftsh5-ל ftsh2לשם כך הכלאנו גם בין . ftsh2 ftsh5 לבין
PCRאשר נבדקו ואומתו באמצעות . נמצאו צמחים הומוזיגוטים לשתי המוטציותF2ובדור , PCRבאמצעות
בעל הוריגציה החריפה מלמד כי רמת הוריגציה הוחמרה ftsh2ניתוח השוואתי שלהם ושל ). 22A-Bתמונה (
של חסר איננה תוצאההאטה בגידולהבכדי לבחון האם . ftsh1 ftsh2רק במעט לעומת זו שבמוטנט הכפול
גם . גודלו הצמחים על גבי מצע המועשר בסוכרוז, במטבוליטים הנגרם כתוצאה מאובדן הכלורופלסטים
בחינת הרגישות ). 22Cתמונה (בתנאים הללו נמצא כי המוטנטים הכפולים מפגרים בגדילתם לעומת הוריהם
אלא רגיש יותר גם ftsh5ו אftsh2-לא רק מ רגיש יותר ftsh2 ftsh5 לעקת אור מעלה כי המוטנט הכפול
.התפתחו ואף העמידו צאצאים, אך יחד עם זאת הצמחים גדלו )22Dתמונה ( ftsh1 ftsh5מהמוטנט הכפול
בהתאם FtsH תורמים שניהם תרומה כמותית קטנה לפעילות FtsH5- וFtsH1מממצאים אלו ניתן לשער כי
מאחר שהמוטנטים . ftsh2 (VARII)וטיפ של חסר שלהם מעצים את הפנ, FtsH2בהעדר . לכמותם הטבעית
מפתחים את כל אברי הצמח ומסוגלים , גדליםftsh1 ftsh8 וגם ftsh1 ftsh2 , ftsh2 ftsh5הכפולים
נראה כי הגורם שיקבע את עצמת הפנוטיפ . כנראה שהחלבונים האלה אינם חיוניים, להשלים מחזור חיים
. ם יהיה כמות החלבון החסרה השוניFtsHבמקרים של חסר של חלבוני
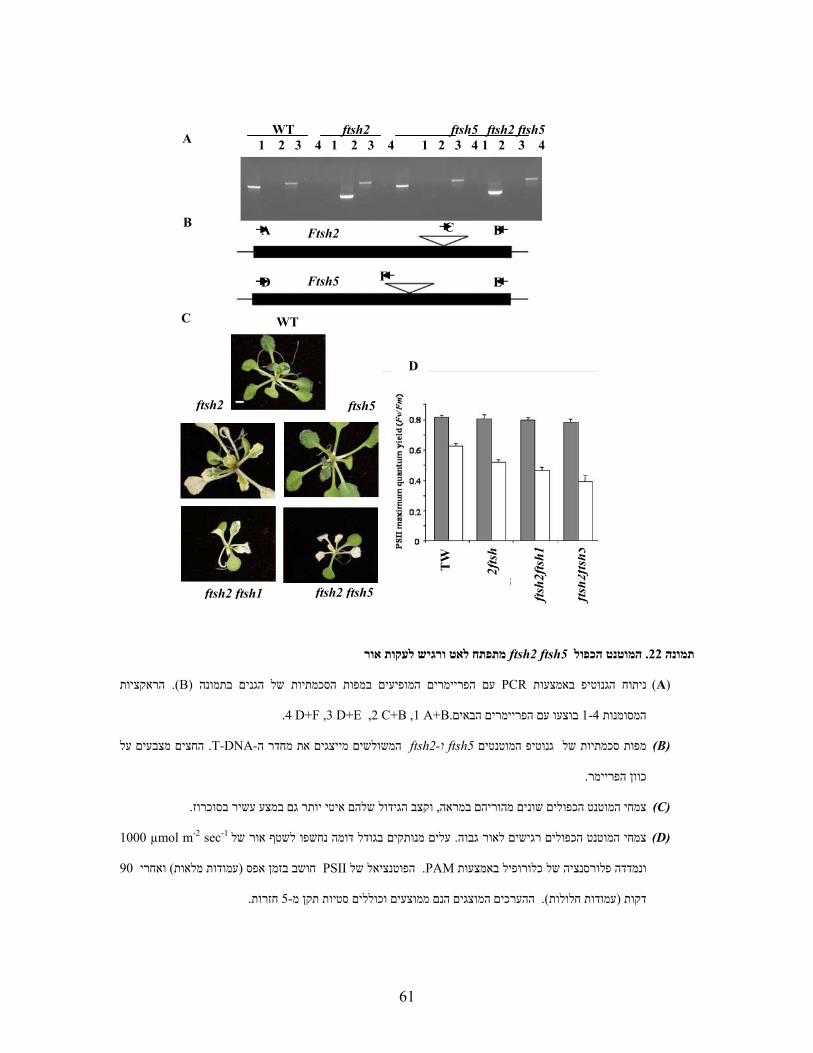
61
מתפתח לאט ורגיש לעקות אור ftsh2 ftsh5המוטנט הכפול . 22תמונה
)A( ניתוח הגנוטיפ באמצעותPCR בתמונה הסכמתיות של הגניםים המופיעים במפותפריימרעם ה (B) . הראקציות
.A+B ,2 C+B ,3 D+E ,4 D+F 1. בוצעו עם הפריימרים הבאים1-4המסומנות
)B( מפות סכמתיות של גנוטיפ המוטנטיםftsh5ו -ftsh2המשולשים מייצגים את מחדר ה -T-DNA. החצים מצבעים על
.כוון הפריימר
)C( וקצב הגידול שלהם איטי יותר גם במצע עשיר בסוכרוז, צמחי המוטנט הכפולים שונים מהוריהם במראה.
)D( 1000שללים מנותקים בגודל דומה נחשפו לשטף אור ע. צמחי המוטנט הכפולים רגישים לאור גבוה µmol m-2 sec-1
90ואחרי ) עמודות מלאות( חושב בזמן אפס PSII הפוטנציאל של .PAMפלורסנציה של כלורופיל באמצעות ונמדדה
. חזרות5- ההערכים המוצגים הנם ממוצעים וכוללים סטיות תקן מ ). עמודות חלולות(דקות
WT 1 2 3 4
ftsh2 1 2 3 4
ftsh5 1 2 3 4
ftsh2 ftsh5 1 2 3 4 A
B
C
D
CA
D
B
EF
WT
ftsh2 ftsh1
ftsh5ftsh2
Ftsh5
Ftsh2
ftsh2 ftsh5
WT
ftsh
2
ftsh2
ftsh1
ftsh2
ftsh5

62
איננו מתקיים בתנאים נורמלייםftsh1 ftsh5המוטנט
בדקנו מה FtsHהאיזוזימים של כמויות שונים בין םעל מנת לבחון האם העצמת הפנוטיפ נגרמת מיחסי
יחד עם ftsh2 ולעומת המוטנטים הכפולים. ftsh5 לעומתftsh5 ftsh1 יהיה הפנוטיפ של המוטנט הכפול
ftsh1 או ftsh5.
:ממצאים שהראו כילאור ה את הבדיקה ערכנו
-WT התפתחה בדומה לftsh1 מוטציה יחידה בגן )א(
(VARI) הראתה פנוטיפ וריגנטי חלשftsh5מוטציה יחידה בגן ) ב(
בהתאמה , ftsh2 היו בעלי פנוטיפ קיצוני יותר משל ftsh5 או ftsh1 עם ftsh2מוטנטים כפולים של ) ג(
.לכמותם באופן טבעי
ניתן היה לצפות כי יתקבל פנוטיפ הדומה FtsH2 קטנה מזו של FtsH5ושל FtsH1 כמותם שלשמאחר
אמתה את PCR באמצעות F1בחינת הגנוטיפ בדור . באופן רציפרוקלי ftsh5- וftsh1הכלאנו בין . -ftsh2ל
על . בתנאי גידול רגילים לא נמצאו צמחים הומוזיגוטים למוטציותF2 אך בסריקה של דור ,הצלחת ההכלאה
-MS שנזרעו על גבי מצע F2נערכו סריקות חדשות לזרעי , לבדוק האם היתה בעיה בהתבססות הצמחיםמנת
agar כמו צמחים שנראו ירוקים: בתנאים הללו זוהו ארבעה פנוטיפים. סוכרוז3% המכילWT) תמונה
23A ( , צמחים שנראו כמו מוטנט החסרFtsH5) 23תמונהB( , צמחים שדמו למוטנטFtsH2וריגציה אך ה
צמחים לבנים לחלוטין שהתקשו להתפתח אפילו , )23Cתמונה (VARIIIנראתה חריפה יותר ונקראו
. למעט הפסיגים בצמחים הלבנים ,הפסיגים של כל הצמחים היו ירוקים). 23Dתמונה (בתנאים תומכים אלו
הורכבה WTראו כמו בדיקת הגנוטיפים באוכלוסיה של הצמחים הללו מעלה כי קבוצת הצמחים הירוקים שנ
):מיצג אלל תקין+ , אות קטנה מיצגת אלל מוטנט(מהגנוטיפים הבאים
WT ,h1+ ,h1h1 ,h1+ h5+ ,h1h1 h5+ ,h5+ . הצמחים שנראו כמוVARI הורכבו מגנוטיפ יחיד h5h5 .
הצמחים הלבנים הכילו את המוטנטים . +h5h5 h1היו בעלי גנוטיפ יחיד " VARIII"הצמחים שנקראו
. h5h5 h1h1 בעלי הגנוטיפ ftsh1 ftsh5ומוזיגוטים הכפוליםהה

63
+h1+ h5 התפצלות הפנוטיפים מהכלאה עצמית של3.2תמונה
)A( אוכלוסיית צמחים שנראו ירוקים כמוWTהיו בעלי הגנוטיפים הבאים :
WT, h1+, h1h1, h1+ h5+, h1h1 h5+, h5+. 2 באורךקו הגודלmm.
)B( ציה קלה היו כולם וריגהצמחים שהראוh5h5.
)C( גציה החריפה היו כולם יהצמחים בעלי הורh5h5 h1+
)D( פהצמחים הלבנים היו בעלי הגנוטי h5h5 h1h1.
)E( תמונת תקריב של עלה ראשון מתמונה A .
)F( תמונת תקריב של עלה ראשון מתמונה B .
)G( תמונת תקריב של עלה ראשון מתמונה C .
)H( נה תמונת תקריב של עלה ראשון מתמו D .
A B
C D
E F G H
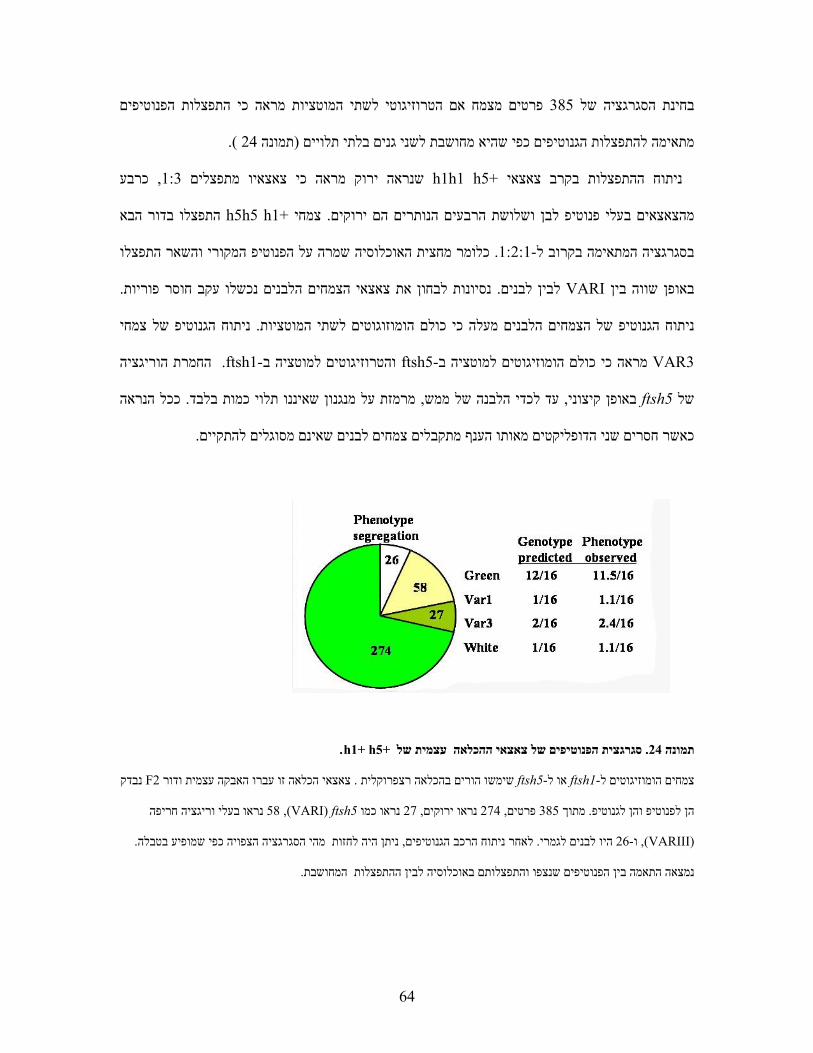
64
פרטים מצמח אם הטרוזיגוטי לשתי המוטציות מראה כי התפצלות הפנוטיפים 385בחינת הסגרגציה של
.) 24 תמונה(מתאימה להתפצלות הגנוטיפים כפי שהיא מחושבת לשני גנים בלתי תלויים
כרבע , 1:3 שנראה ירוק מראה כי צאצאיו מתפצלים +h1h1 h5 ניתוח ההתפצלות בקרב צאצאי
התפצלו בדור הבא +h5h5 h1צמחי . צאצאים בעלי פנוטיפ לבן ושלושת הרבעים הנותרים הם ירוקיםמה
כלומר מחצית האוכלוסיה שמרה על הפנוטיפ המקורי והשאר התפצלו . 1:2:1-בסגרגציה המתאימה בקרוב ל
. חוסר פוריותנסיונות לבחון את צאצאי הצמחים הלבנים נכשלו עקב . לבין לבניםVARIבאופן שווה בין
ניתוח הגנוטיפ של צמחי . ניתוח הגנוטיפ של הצמחים הלבנים מעלה כי כולם הומוזוגוטים לשתי המוטציות
VAR3מראה כי כולם הומוזיגוטים למוטציה ב ftsh5-והטרוזיגוטים למוטציה ב -ftsh1 . החמרת הוריגציה
ככל הנראה . ון שאיננו תלוי כמות בלבדמרמזת על מנגנ, עד לכדי הלבנה של ממש, באופן קיצוניftsh5של
.כאשר חסרים שני הדופליקטים מאותו הענף מתקבלים צמחים לבנים שאינם מסוגלים להתקיים
.+h1+ h5 עצמית של סגרגצית הפנוטיפים של צאצאי ההכלאה . 24תמונה
נבדק F2י הכלאה זו עברו האבקה עצמית ודור צאצא. שימשו הורים בהכלאה רצפרוקלית ftsh5- או לftsh1- הומוזיגוטים לצמחים
נראו בעלי וריגציה חריפה ftsh5 (VARI) ,58 נראו כמו 27, נראו ירוקים274, פרטים385מתוך . הן לפנוטיפ והן לגנוטיפ
(VARIII) ,בטבלהניתן היה לחזות מהי הסגרגציה הצפויה כפי שמופיע, לאחר ניתוח הרכב הגנוטיפים. היו לבנים לגמרי26-ו .
.נמצאה התאמה בין הפנוטיפים שנצפו והתפצלותם באוכלוסיה לבין ההתפצלות המחושבת

65
איננו מפתח כלורופלסטיםftsh2 ftsh8המוטנט הכפול
FtsH8כמות חלבון . )Sakamoto et al., 2003( אינה מלווה בסימנים כלשהם ftsh8 השתקתו של
מהניתוח . )FtsH5 )Sinvany-Villalobo et al., 2004 וגם מכמות FtsH2הנצברת נמוכה מכמות
- A“סוג " ,י ענפיםהפילוגנטי ניתן להסיק כי ארבעת החלבונים הכלורופלסטיים שנצברים מתחלקים לשנ
FtsH1+FtsH5 ,סוג ”ו“B- .FtsH8+FtsH2 סוג " לאור הממצא כי חסר של“Aרצינו לברר , הוא לטאלי
ftsh8 לבין ftsh2בוצעו הכלאות רציפרוקליות בין . B“סוג "האם תופעה דומה תתרחש גם בהשתקה של
ניתן להבחין . F1) של PCR-כלאה בלאחר שאומתה הצלחת הה( נזרע על גבי מצע מועשר בסוכרוז F2ודור
ftsh2 VARIIפנוטיפ הדומה לצמח , )25Aתמונה (-WT דומה ל פנוטיפ ירוק הנראה: בארבעה פנוטיפים
שאחד מהם עשוי לפתח מעין עקבות ירוקים בעלים ) 25C-Dתמונה (שני פנוטיפים לבנים , )25Bתמונה (
. הבוגרים
העלה כי די באלל אחד נורמלי של , ל כל אחד מהפנטופים האלה שPCR)באמצעות (ניתוח הגנוטיפים
FtsH2בכדי לקבל צמח ירוק ללא כל תלות ב -FtsH8 .הפנוטיפ הירוק כלל את הגנוטיפים הבאים, כלומר:
WT ,h8+ ,h8h8, h2+ h8h8, h2+ h8+, . הצמחים בעלי הווריגציה היו כולם זהים לצמח האםh2h2 ,
, השני הומוזיגוט למוטציה-ftsh8האחד הטרוזיגוט למוטציה ב: היו שני גנוטיפיםואילו בצמחים הלבנים
בשלב זה לא מצאנו . הצמחים הלבנים היו בעלי פסיגים לבנים, לרוב. ftsh2-ושניהם הומוזיגוטים למוטציה ב
הגנוטיפים הבדלים בתוך קבוצת הצמחים הלבנים ואילו בשלבי גידול מאוחרים יותר ניתן היה להבחין בין שני
הצליח לפתח מספר תאים ירוקים שהכילו כלורופלסטים בעליו העליונים , +h2h2 h8, הראשון .הלבנים
הצמחים הלבנים , בהעדר סוכר. הצמחים הלבנים הצליחו לגדול רק על מצע מועשר בסוכרוז). 26תמונה (
בסוכרוז היו איטיים במעט קצבי הגידול של צמחים אלו אפילו במצע מועשר . מתו או לא הצליחו להתפתח
.WTמקצבי הגידול של צמחי

66
.+h2+ h8של התפצלות צאצאים של ההכלאה עצמית . 25תמונה
, תוצרי האבקה זו נזרעו על גבי מצע אגר המועשר בסוכרוז, עברו האבקה עצמיתftsh8 לבין ftsh2 תוצרי ההכלאה בין
. שניים שנראו כמו ההורים ושניים לבנים, ניתן היה לזהות ארבעה פנוטיפים שונים. שה שבועותוגודלו למשך כשלו
(A) צמח הנראה כמו ftsh8 .
(B) צמח הנראה כמו ftsH2.
(C-D) צמח לבן .
(E) שלושת העלים הראשונים מצמחים A-C . עלה ראשון של צמח: משמאל לימין ftsh8 , עלה ראשון של צמחftsh2 , עלה
ftsh8-והטרו או הומוזיגוטי ל ftsh2-ראשון של מוטנט כפול הומוזיגוטי ל

67
: כאשר בחנו את התפצלות האוכלוסיה נתגלה כי שיעור הצמחים הלבנים היה קטן בהרבה מהמצפוי
בהתפצלות מנדלית בלתי תלויה של שני גנים או ארבעה אללים השיעור הצפוי של ההומוזיגוטים הכפולים
מכאן שסך , 2/16 והשיעור הצפוי של ההומוזיגוטים למוטציה אחת והטרוזיגוטים לשניה הוא 1/16הוא
).27תמונה (3/16הצמחים הלבנים צפוי היה להיות
. 3/16- ולא ל1/16-מספר הקרוב יותר ל. היו לבנים27 צמחים שגדלו רק 431הממצאים הם כי מתוך
הצמחים הלבנים שהצליחו לגדול על מצע המועשר . 96%ים ועמדו על אחוזי הנביטה בניסוי זה היו נורמלי
אינו מספיק FtsH8קיומו של אלל יחיד נורמלי של . בסוכר הצליחו להתפתח באופן נורמלי אך בקצב איטי
לגדילתו של צמח FtsH2מספיק אלל יחיד של , אך במצב ההפוך, FtsH2בכדי לקיים את הצמח בהעדרו של
.ממצאים אלו עשויים להסביר את היחסים שבין שני דופליקטים אלו. ליבעל מראה נורמ
. מיצרים מעט כלורופלסטיםFtsH8- אך בעלי אלל יחיד לFtsH2 חסרי צמחים. 26תמונה
צמחים אלו היו . שבועות תוך שמירה על טריות המצע12גודלו במשך , צמחים שהונבטו על גבי מצע אגר המועשר בסוכרוז
עם ההתפתחות והופעת העלים העליונים בשושנת ועל עמודי , הצמחים הלבנים בודדו, +h2+ h8 הכלאה עצמית של צאצאי
כמות . +h2h2 h8המכילים כלורופלסטים רק בצמחים בעלי הגנוטיפ , )מסומן בחץ(התפרחת נראו קבצי תאים ירוקים
צמחים שהועברו למצע ללא סוכרים לא . גידול התקיןהכלורופלסטים לא היתה מספקת על מנת לתמוך בצמח ולספק סוכרים ל
.שרדו

68
+h2+ h8 מההכלאה עצמית של אוכלוסיית הצאצאים התפצלות.27תמונה
בשלבי גידול . תוצרי ההפריה זו נזרעו על גבי מצע מועשר בסוכרוז. עברו האבקה עצמיתftsh8 לבין ftsh2תוצרי ההכלאה בין
ניתוח הגנוטיפים של צמחים אלו העלה כי הצמחים הירוקים מכילים כולם . ולבןVARII,ירוק : םצעירים זוהו שלושה פנוטיפי
והכילו שני ftsh2 היו כולם הומוזיגוטים למוטציה VARII- צמחי ה. ftsh8ללא תלות במצבו של , ftsh2לפחות אלל אחד תקין של
. ftsh8 - והכילו אלל או שניים מוטנטים בftsh2למוטציה הצמחים הלבנים היו כולם הומוזיגוטים . ftsh8אללים תקינים של
+h2+h8 של בהכלאה עצמיתשעור הפלת עוברים גבוה
רצינו לברר , חסרו בעיקר המוטנטים הכפולים+h2+ h8 הצאצאים של תהתפצלות אוכלוסיישבמאחר
נראו שלושה סוגי , 28כפי שמציגה תמונה . +h2+ h8נבחנו תרמילים של המוטנט . עד איזה שלב הם שרדו
- והשלישי , זרעים לבנים- השני , מהראשון התפתחו זרעים שנראו נורמלים וצבעם היה ירוק: עוברים
). 28תמונה ( WT- ולא בftsh8 או ftsh2, תמונה דומה לא נצפתה בדור ההורים. עוברים או ביציות מופלות
והעוברים , יתפתחו לצמחים לבנים בוצע חילוץ עוברים בתנאים אספטייםעל מנת לבחון אם הזרעים הלבנים
צמחים אלו היו כולם לבקנים . רק שמונה עוברים הצליחו להתפתח לצמחים. גודלו על גבי מצע המכיל סוכרוז
.ראוי לציין שלאחר הבשלת התרמיל קשה מאוד להבחין בזרעים הלבנים .חסרי כלורופיל

69
+h2+ h8 ושל צמחי WTלים של צמחי תרמ ניתוח.28תמונה
מתפתחים בתרמיל באופן WT- הזרעים מצמחי ה
נורמלי ולא מזוהות בתרמילים הפלות עוברים או
בתרמילים מצמחים בעלי הגנוטיפ . זרעים
h2+ h8+ניתן להבחין באוכלוסיה מתפצלת :
ועוקצי ) בחץ כחול(זרעים לבנים , ירוקים זרעים
כנראה עקב הפלת , שחלה בהם לא מתפתח זרע
).חץ אדום(העובר או הביצית
עקר , ftsh2 ftsh8, המוטנט ההומוזיגוטי הכפול
למרות זאת הם מצליחים לפרוח . מתקיימים בקושי אפילו במצע מועשר בסוכרוזh2h2 h8h8 צמחי
ראים נורמלים ומפתחים את כל האברים הצמחים נ, אם מתעלמים מהעדר הכלורופלסטים). 29A,Bתמונה (
השוואת אברי הרבייה של המוטנט הכפול .עלים ואף עמודי תפרחת ותפרחות, גבעולים, שורשים: שלהם
למעט העובדה כי צבע עלי הגביע לבן ולא ירוק , מעלה כי התפרחות נראות דומותWTלאלה שבצמחי
זיריהם קצרים ולא נראים , המוטנט קטנים יותרבחינת אברי הפרח מעלה כי האבקנים של). 29Cתמונה (
אך על גבי הצלקות של המוטנט , עמודי העלי ובראשם הצלקות נראו דומים). 29Dתמונה (בהם גרגרי אבקה
כנראה שהעדר האבקה מונע הפריה ולכן לא מתפתחים ). 29E - ל29Fהשוואת תמונה (לא זוהו גרגרי אבקה
ניתוח התרמילים הללו מעלה כי ). 29G,Hתמונה (עין תרמיל קטן עמודי העלי מתפתחים למ.תרמילים
.ftsh1 ftsh5תמונה זהה מתקבלת גם במוטנט הכפול . נמצאים בתוכם מבנים דמויי ביציות או עוברים מופלים
WT h2+ h8+ h2+ h8+
h2+ h8+

70
המוטנט הכפול פגום ביכולתו לייצר אברי רבייה תקינים
נבחנו , לנסות לברר מה מתרחש בתוך אברי הרביהעל מנת. עקר זכרית, 29תמונה נראה ב כפי ש, הפרח
האבקנים הצעירים נראו דומים ואף . שגודלו בתנאים זהיםWT ושל ftsh2 ftsh8פרחים של המוטנט הכפול
חתך רוחב בפרח מראה כי העמוד ). 30A,Gתמונה (WT-נראה כי אבקני המוטנט הכילו גרגרי אבקה כמו ב
בעוד שהעמוד המרכזי של המוטנט מכיל ביציות ) 30Bתמונה (ות מופרות מכיל ביציות שנראWT-המרכזי ב
אך קיימת , הביציות מסודרות בשחלה מרכזית על גבי עלי השחלה-WT ב). 30Hתמונה (שאינן מופרות
ונספגים או שאינם מתפתחים כראוי " מתמוססים"ייתכן כי עלי השחלה . בעיה בארגון השחלה במוטנט
המתחילות להתפתח ) 30Dתמונה ( ניתן לזהות ביציות מופרות -WTחינת הביציות בבב). 30C, Iתמונות (
כנראה כי אין -בעוד שבצמחי המוטנט ניתן לזהות ביציות אך הן לא ממשיכות להתפתח לעוברים , לעובר
WT-אך רק בצמחי ה, הן במוטנט והן בביקורת, במאבקים ישנם תאי אבקה שנראים נורמליים. הפריה
במוטנט נוצרת רקמת ניתוק אך ). 30E, Fתמונה (תיחה של המאבק המאפשרת את פיזור האבקה מתרחשת פ
הממצאים האלה מציעים כי מנגנון ). 29K, Lתמונה (היא נשארת מאוחה ולא גורמת לפתיחת האבקן
טים כנראה ללא פגיעה בתאי המין או הספורופי, העקרות הינו מנגנון של עקרות פזיולוגית או מורפולוגית
.בשלבים המוקדמים

71
איננו ftsh2 ftsh8 הכפול המוטנט.29תמונה
פורה
יכול להגיע ftsh2 ftsh8מוטנט הומוזיגוטי ל
לשלב הרפרודוקטיבי שלו ולפרוח אם גדל במצע
עשיר בסוכרוז
אחרי שגודלו ftsh2 ftsh8צמחי ). A-Bתמונה (
. במכלחי פלסטיק עם סוכרוז במשך ארבע חודשים
השוואת ,) משמאלWT, מוטנט מימין, Cנה תמו(
.מראה של תפרחות הצמחים
זיר , השוואת מראה של אבקנים . ) Dתמונה (
.ומאבקים
השוואת עמודי העלי והצלקות ) E, Fתמונה (
סימני האבקה . משמאלWTשל המוטנט מימין
WT-מופעים רק על הצלקת של צמח ה
, )נטלמטה המוט , WTלמעלה , G+Hתמונות (
תרמילים של המוטנט נראים כמבנים קצרים מאוד
.דמויי תרמיל
בתוך ). WT- ה-Iתמונה , המוטנט- Jתמונה (
וביציות או עוברים , WT- התרמילים יש זרעים ב
מופלים
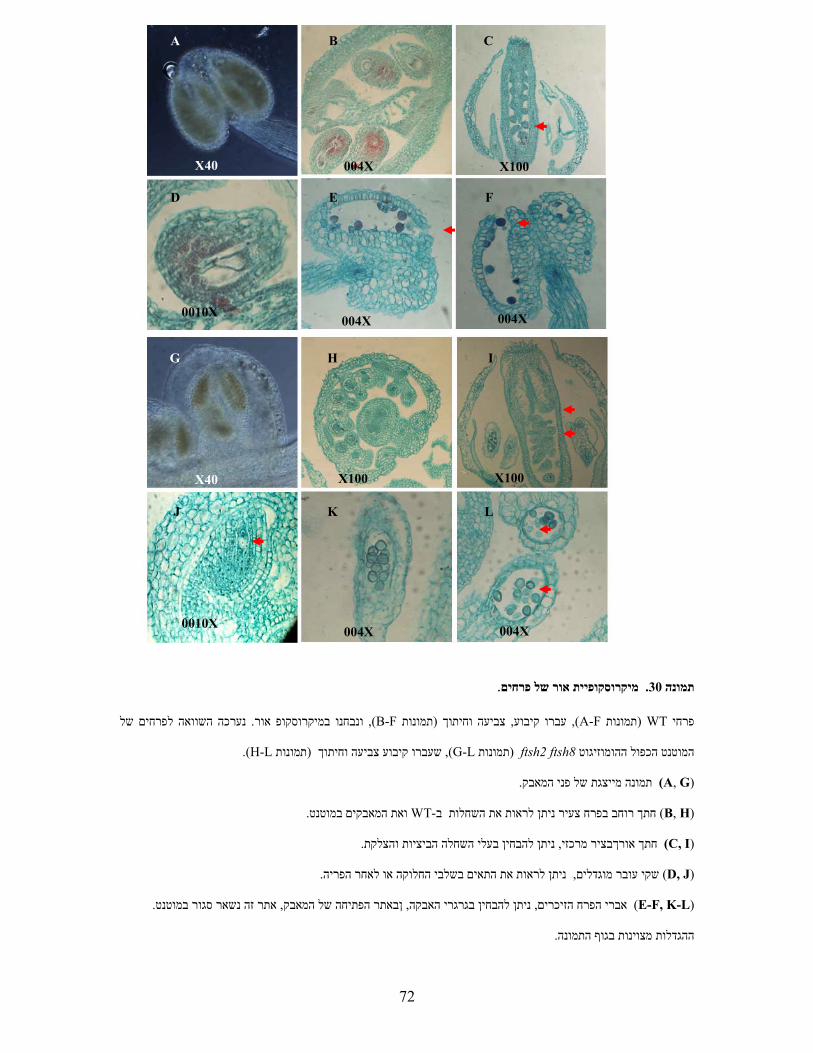
72
. אור של פרחים מיקרוסקופיית.30תמונה
של פרחים ל נערכה השוואה. נבחנו במיקרוסקופ אורו ,)B-Fתמונות (וך וחיתה צביע, עברו קיבוע,)A-Fתמונות ( WTפרחי
).H-Lתמונות ( וחיתוך ה שעברו קיבוע צביע,)G-Lתמונות ( ftsh2 ftsh8ט וגההומוזיכפול המוטנט ה
(A, G)תמונה מייצגת של פני המאבק .
(B, H)חתך רוחב בפרח צעיר ניתן לראות את השחלות ב -WTוטנט ואת המאבקים במ.
(C, I)ניתן להבחין בעלי השחלה הביציות והצלקת, חתך אורךבציר מרכזי.
(D, J)ניתן לראות את התאים בשלבי החלוקה או לאחר הפריה, שקי עובר מוגדלים.
(E-F, K-L)אתר זה נשאר סגור במוטנט, ןבאתר הפתיחה של המאבק, ניתן להבחין בגרגרי האבקה, אברי הפרח הזיכרים.
. לות מצוינות בגוף התמונהההגד
A B C
D E F
G H I
J K L
X40
X40
X100
X100 X100
X400
X400 X400
X400 X400
X1000
X1000

73
בחינה השוואתית של הגנוטיפים השונים
.זרענו את כולם יחד בצלחות פטרי על מצע אגר, על מנת להשוות בין כלל הגנוטיפים שהיו בידנו
). 31A תמונה ( שבועות 3-4יותר הועברו לאחר שבועיים לאדמה והמשיכו לגדול עוד " קלים"הפנוטיפים ה
נערכה בדיקה כמותית ). 31Bתמונה (פנוטיפים החמורים הושארו בצלחות המכילות סוכרוז הצמחים בעלי ה
וגם FtsH8 או של FtsH1אובדן של ). 31Cתמונה (לקביעת כמות הכלורופיל ברקמה בתור מדד לוריגציה
פ של גרם לפנוטיFtsH5 או של FtsH2אובדן של . -WTשל שניהם לא גרם לפנוטיפ מיוחד והם נראו כמו ה
חסר נוסף של , FtsH2-על רקע של חסר ב. בהתאמה, וריגציה ועיכוב בגידול באופן חמור יותר וחמור פחות
FtsH1 או של FtsH5חסר ב. מגביר את מופע הוריגציה בצורה דומה-FtsH1וב -FtsH5או חסר ב - FtsH2
. רק על מצע מועשר בסוכרוזוגדל, מוביל למופע לבן לחלוטין שאיננו מסוגל לבצע פוטוסינתזה FtsH8-וב
מתאפשר תהליך , בהתאמה, FtsH8 או FtsH1 אך קיים אלל אחד של FtsH2 או FtsH5כאשר אין
קיומם של שני אללים מאפשר את הביוגנזה. התפתחות כלורופלסטים בפסיגים וגם בעלים עליונים
.FtsH2 באפיון המוטנט של יההדרגתית של הכלורופלסטים כפי שתיארת
השניזוג המונעת את הצטברות קרובים FtsH's תקה של זוג הש
,.Sakamoto et al) גורם בהתאמה לירידה בכמותו של החלבון השני FtsH5- או בFtsH2-חסר ב
כל הממצאים שלנו מובילים . נצבר בתור הטרוקומפלקס-FtsHתופעה זו מוסברת היטב בכך ש. (2003
FtsH8- וFtsH2-ו, "Aסוג " המהווים את FtsH5- וFtsH .FtsH1 למסקנה כי קימים שני סוגים של
העדר , אם מסקנה זו נכונה. צבירתו ותפקודו, שני הסוגים חיוניים לבניית ההטרוקומפלקס, "Bסוג "המהווים
חיוני לביוגנזה של FtsHהפנוטיפ הלבן מעיד כי . של סוג אחד ימנע את תפקודו או את צבירתו של הסוג השני
? השאלה שעמדה בפנינו הייתה האם נצבר קומפלקס לא פעיל בתא. ופלסטים ולהגנה מפני נזקי קרינהכלור
- על מנת לבדוק זאת הופקו חלבונים מעלי ארבידופסיס שגדלו בתנאים זהים משלושת הגנוטיפים הלבנים ומ
WT .קטיבציה של כל אחד התוצאות המוצגות מראות כי אינא. חלבונים אלו הופרדו וזוהו באמצעות נוגדנים
במקרה שנצבר . הכלורופלסטייםFtsHמהסוגים מונעת לחלוטין צבירה של הזוג השני ושל כלל חלבוני
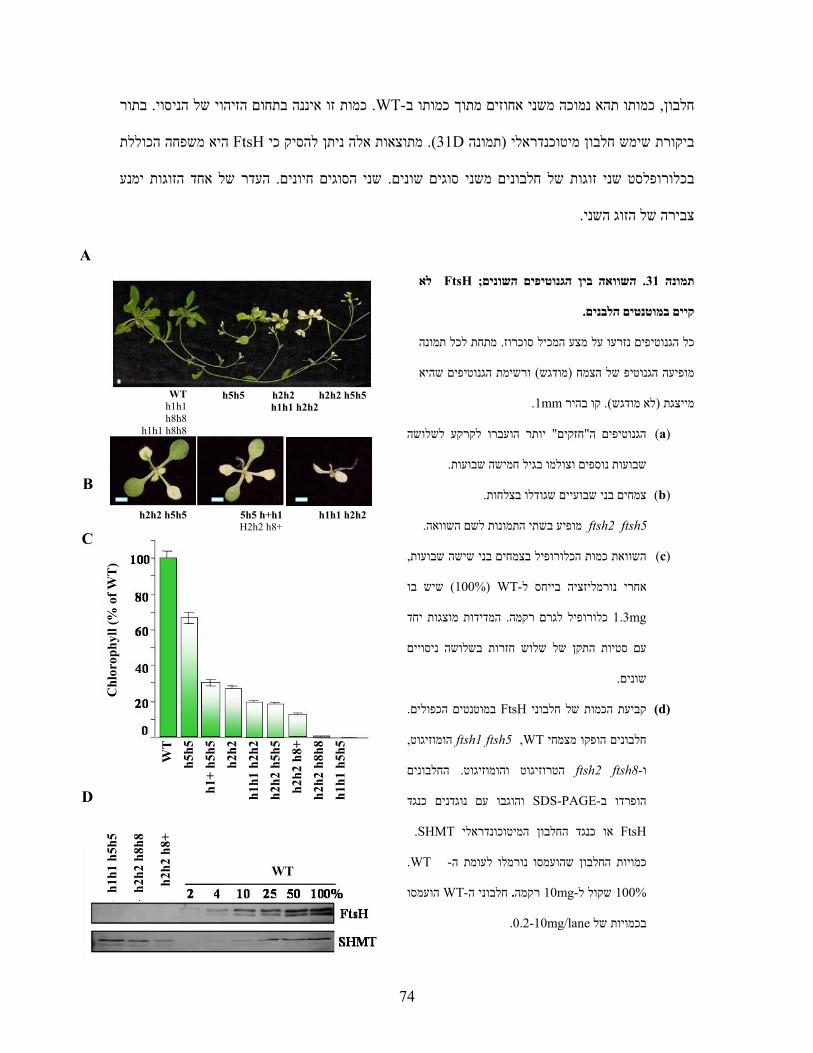
74
בתור . כמות זו איננה בתחום הזיהוי של הניסוי. WT-כמותו תהא נמוכה משני אחוזים מתוך כמותו ב, חלבון
היא משפחה הכוללת FtsH ניתן להסיק כי מתוצאות אלה). 31Dתמונה (ביקורת שימש חלבון מיטוכנדראלי
העדר של אחד הזוגות ימנע .שני הסוגים חיונים. בכלורופלסט שני זוגות של חלבונים משני סוגים שונים
.צבירה של הזוג השני
לא FtsH ; השוואה בין הגנוטיפים השונים .31תמונה
.קיים במוטנטים הלבנים
מתחת לכל תמונה . כרוזכל הגנוטיפים נזרעו על מצע המכיל סו
ורשימת הגנוטיפים שהיא ) מודגש(מופיעה הגנוטיפ של הצמח
.1mmקו בהיר ). לא מודגש(מייצגת
)a( יותר הועברו לקרקע לשלושה " חזקים"הגנוטיפים ה
. שבועות נוספים וצולמו בגיל חמישה שבועות
)b( צמחים בני שבועיים שגודלו בצלחות .
ftsh2 ftsh5 התמונות לשם השוואהמופיע בשתי .
)c( השוואת כמות הכלורופיל בצמחים בני שישה שבועות ,
שיש בו ) WT) 100%- אחרי נורמליזציה בייחס ל
1.3mgהמדידות מוצגות יחד . כלורופיל לגרם רקמה
עם סטיות התקן של שלוש חזרות בשלושה ניסויים
.שונים
)d( קביעת הכמות של חלבוניFtsHכפולים במוטנטים ה .
, הומוזיגוטWT , ftsh1 ftsh5מצמחי הופקו חלבונים
החלבונים .הטרוזיגוט והומוזיגוט ftsh2 ftsh8-ו
והוגבו עם נוגדנים כנגד SDS-PAGE- בהופרדו
FtsH או כנגד החלבון המיטוכונדראלי SHMT.
.WT - נורמלו לעומת היות החלבון שהועמסו וכמ
הועמסו WT- חלבוני ה. רקמה10mg-ל שקול100%
.0.2-10mg/laneיות של בכמו
A
B
C
D
WT h1h1 h8h8
h1h1 h8h8
h5h5
h2h2 h1h1 h2h2
h1h1 h2h2
h2h2 h5h5
h1+ h5h5 H2h2 h8+
h2h2 h5h5
WT
h5h5
h1
+ h5
h5
h2h2
h1
h1 h
2h2
h2h2
h5h
5h2
h2 h
8+ h2
h2 h
8h8
h1h1
h5h
5
Chl
orop
hyll
(% o
f WT
)
h1h1
h5h
5
h2h2
h8h
8
h2h2
h8+
WT

75
דיון
FtsH המודל הכמותי'ו' מודל הסף '– מעורב בתהליכי התפתחות של הכלורופלסט'
ורגישות לשטף אור גבוה , גורמת למופע מוזאיקה של כתמים ירוקים ולבנים לסרוגיןFtsH - במוטציית חסר
)Chen et al., 2000; Takechi et al., 2000( . תופעת הוריגציה היא תופעה מיוחדת של תאים סמוכים
בתור הסבר . או כלורופלסטים או פרופלסטידות: שנצברים בהם שני סוגים של פלסטידותבעלי גנוטיפ אחד
אשר מתחתיו אין יצירת תילקואידים FtsHלתופעה הוצג מודל אשר מסביר כי יש סף מסוים לכמות חלבוני
ההנחה בבסיס המודל הזה היא כי הביטוי , )Chen et al., 2000; Yu et al., 2004(ומתקבלים תאים לבנים
מודל זה אינו מסביר את דפוס הוריגציה שאיננו אקראי . של גן מסויים שונה בין תאים קרובים באופן אקראי
אשר עליו הצעירים הם ftsh5המופע של המוטנט ). 10 תמונה(ת של תאים בקבוצו, אלא מופיע בכתמים
FtsH5בחינת כמות החלבון של . 'מודל הסף'מתאים ל ftsh2בעלי דפוס וריגנטי קל יותר מאשר עלי המוטנט
-FtsH2 )Sinvany אך נמוכה יותר מאשר FtsH8 או FtsH1הראתה כי כמותו רבה יותר מאשר
Villalobo et al., 2004( .סף כמות ה, על פי המודל נראה כי, מכאן-FtsHשניתן להחסיר מ -WT היא
- וגם ב-FtsH1מוטנטים גם ב( וכל כמות שגדולה ממנה תגרום לוריגציה FtsH5כמות דומה לכמות של
FtsH8בבסיס המודל הזה אין הבחנה בין חלבוני ). נטים אינם וריג FtsHתפקידיהם זהים , כלומר, השונים
, A(איננה נכונה שכן ניתן להבחין בשלושה סוגים , כפי שעולה ממצאי עבודה זו, הנחה בסיסית זו. וחופפים
B ,ו-C ( של חלבוניFtsH" : סוגA " - FtsH1ו-FtsH5סוג" ו"B - FtsH2ו -FtsH8 . של בהעדרם
). 29Aתמונה ( מתקבלים צמחים שבפועל אינם אוטוטרופיים אלא אורגניזמים הטרוטרופיים B או Aהסוגים
יתכן כי חלקם פסאודוגנים או שכמותם , שתפקידם ופעילויותיהם אינם ידועים FtsHהכולל חלבוני" -Cסוג "ו
בחינת קווי מחדר . ים אחרים היא מזערית או מתרחשת בתנאים קיצונFtsHותרומתם לפעילות הכוללת של
הוא , גם יחדB- וAהמוכוונים לכלורופלסט מעלה כי כל עוד קיימים בצמח הסוגים FtsHבגנים השונים של
תופעות הוריגאציה והרגישות לאור ייקבעו על פי הכמות הכוללת של יחידות . יהיה מסוגל לתפקד ולחיות
FtsHה מכמות השקולה לכמותו של כאשר הירידה בכמות תהה קטנ. שיתפקדו בתאFtsH5 היא לא תורגש
בחינת כלל המוטנטים במשפחה במצב של מוטציה יחידה מראה -ולראייה . ולא תגרום לפנוטיפ נראה לעין

76
בהם לא התקבל אף , באמצעות אנטיסנסftsh1תמיכה נוספת מתקבלת מנסיונות שערכנו להשתקת . מצב כזה
FtsH1 שלcDNA-הנסוי נערך באמצעות כל ה). ים אינם מוצגיםנתונ(לא צמח אחד שהכיל את המחדר
ניתן להניח כי חלה השתקה , ובשל העובדה כי הרצף המלא כולל מקטעים ארוכים הזהים בין הגנים השונים
מצב כזה לא מאפשר חיים . הקרוב אליוftsh5 וגם של ftsh1נרחבת של חלבונים במשפחה או השתקה של
.כלל צמחים בניסוי זהלצמח ולכן לא התקבלו
לדוגמה מוטציות . מקנה לצמח סבילות כלפי מוטציות בגן זה או אחר במשפחהFtsH ריבוי הגנים של
ואפילו בשנהם יחד אינן גורמות לירידה מובהקת בכמות חלבוני FtsH8- הן בFtsH1-קיצוניות השתקה הן ב
FtsH ר חלבוני המשפחה מפצים על העדרם ותופסים את יתכן כי שא) הנתונים אינם מוצגים( בכלורופלסט
עשויה להיות , "משפחות גנים"–למנגנון ריבוי עותקים לגן . מקומם הן מבחינה תיפקודית והן צבחינה כמותית
מה שעשוי להפוך ליתרון בתנאים (חשיבות רבה בשמירה על השונות הגנטית לאור אי חשיפת המוטציה
ולא משפחות גנים הופך את האורגניזם לפגיע ביותר מבחינה גנטית וכל קיומם של גנים בודדים). אחרים
במיוחד במינים עם רביה מינית עצמית שעלולים להגיע במהירות יחסית למצב , מוטציה היא משמעותית
.הומוזיגוטי באללים פגועים ולהתקרב להכחדה
ו ופעילותFtsHעים על כמות יהמשפאנדוגניים גורמים
גורם למופע וריגנטי אשר עשוי להיות מושפע מתנאי סביבה כגון טמפרטורה FtsH2-בחסר
(Sakamoto et al., 2002) . אך גם בתנאים קבועים ומבוקרים מופע זה איננו אחיד והוא שונה בזוגות עלים
ירוק ועד עלה או פסיג, עוצמת הפנוטיפ עשויה לנוע מעלה לבן כמעט לחלוטין כמו העלה הראשון. שונים
כל עלה יהיה ירוק יותר מהעלה שמתחתיו ובלבד : בין העלים הוריגציה משתנה באופן הדרגתי. כמעט לחלוטין
ירוק לחלוטין , לעומת זאת, הפסיג). 14 ,11תמונות (שהצמח לא נחשף לתנאי סביבה קיצונים במהלך הגידול
ורוב , כחלק מהעובר, העוברהפסיג מתפתח למעשה בתוך שק. WT וקשה להבחין בינו לבין פסיג של
הוא דומה יותר לעלה האחרון , במידה מסויימת, לפיכך. חלוקות התאים מתרחשות עדיין על גבי צמח האם
. שנוצר מאשר לעלה הראשון

77
התהליך של צמצום הוריגציה הוא תהליך דינמי שחלה בו התרחבות השטחים הירוקים וצמצום השטחים
ביוגנזה של תילקואידים וצבירת , ביוגנזה של כלורופלסטים: תהליכים נוספיםתהליך זה מקביל ל. הלבנים
עשוי להסביר את השינויים באופי הוריגציה בשלבי ' מודל הסף'. הפיגמנטים והקומפלקסים הפוטוסינטתים
FtsHלפי מודל זה ניתן לצפות כי ככל שהעלה ירוק יותר כמות התעתיק והחלבון של . ההתפתחות השונים
מתקבלת התאמה טובה בין רמת ההורקה לבין , כאשר בוחנים את כמות החלבונים, ואכן. ה גדולה יותרתהי
, הנפוציםFtsHכמו כן ניכרת מגמת עליה בכמות התעתיק של כל אחד מארבעת חלבוני . FtsHכמות חלבוני
. בין הפסיג לעלה הראשון ולשני
. כלורופלסטיFtsHש כמות קטנה יותר של תעתיק של מוצאים כי בפסיג יWT- כשבוחנים את צמחי ה
ואכן , בהגברתו בעלים האמתיים' הגיון ביולוגי' מעורב בתיקון המערכת הפוטוסינטתית יש -FtsHמאחר ש
לבין כמות החלבון הנצברת בשלבי , 12מצאתי התאמה חיובית בין רמות התעתיק המוצגות בתמונה
לבין רמת FtsHיש התאמה חיובית בין כמות חלבוני , יתרה מכך. 14ההתפתחות השונים כפי שמוצג בתמונה
כלל . או של המוטנטיםWT-וגם בין עלים שונים של ה) הנתונים אינם מוצגים(ההורקה במוטנטים השונים
אולי , FtsH תפקיד חיוני בעלים וככל שהעלה גבוהה יותר נחוץ יותר -FtsHהממצאים הללו מציעים כי ל
.מסוככים על העלים התחתונים ליוניםמשום שהעלים הע
בעבר הוצע כי תופעת הוריגציה עשויה להיגרם מנזקים פעילים הגורמים להלבנה
(Photobleaching) )Sakamoto et al., 2002( .המבנה המורפולוגי של : קשה לקבל טענה זו מכמה סיבות
הוריגציה איננה , מרכזם מכוסה על ידי העלים שמעליהם- שושנת חושף בעיקר את קצוותיהם לאור עלי ה
ומגמת הגברת הוריגציה בעוצמות אור גבוהות דומה לצימצום , משתנה בעלה במשך חודש בתנאים קבועים
קיים שוניWTממצאים שמראים כי בצמחי , )16תמונה (WT-בכמות הכלורופיל המתרחש במערכת ה
על פי מיקומו של העלה בשושנת העלים FtsHאנדוגני בין העלים המביא לעליה בכמות התעתיק והחלבון של
לפיכך נראה כי תופעת ההשתנות או צימצום הוריגציה לאורך ההתפתחות נגרמת בשל , )14- ו12תמונות (
שאלו עשויים להשפיע למרות. והיא איננה נגרמת מתנאי הסביבהFtsHהעלייה האנדוגנית בכמות חלבוני
.עליה

78
בצמחים . WT- נראה שונה מהמנגנון שבftsh2 במוטנט FtsH המנגנון האנדוגני לשינוי רמת חלבוני
לבין FtsHיש מתאם גבוה בין כמות חלבוני . הפסיג ירוק והעלה הראשון לבן כמעט לחלוטין, FtsH2חסרי
, הכלורופלסטיים התקינים FtsHחד משלושת חלבוני בחינת כמויות התעתיק מעלה כי בכל א. רמת ההורקה
ואילו רמות נמוכות יותר נמצאות , FtsHהרמה הגבוהה ביותר היא דווקא בעלה הלבן שאין בו כמעט חלבון
. לבין כמות החלבוןmRNA-המשמעות היא שקיים מתאם הפוך בין כמות ה. בפסיגים ובעלים שמעליהם
מדכא כנראה FtsHבמודל הזה . 'מודל ההיזון החוזר '-ת מודל שבנינו הדבר עשוי להיות מוסבר באמצעו
הפקטור נצבר FtsH -עקב הפגיעה ב). שאיננו ידוע(פקטור כלורופלסטי , באמצעות פירוק ספציפי
פקטור זה עשוי לצאת מהפלסטידה ולשמש כפקטור שעתוק המפעיל את שעתוקם של חלבוני . בפלסטידה
FtsH ,בפלסטידה ולהפעיל מערך של הובלת סיגנל על להפעלת שעתוקם של חלבוני או לחלופין להשאר
FtsH .עם צבירת חלבוני הFtsH- החדשים מתחיל להתרחש שוב תהליך הפירוק של הפאקטור ונפסק
הופעלה FtsH החסר את החלבון הנפוץ ביותר מבין חלבוני ftsh2במוטנט . FtsHהשעתוק המוגבר של
הביטוי הוא ביטוי מוגבר לעומת מצב שבו מערכת המשוב אינה . וי הופעל כמתוארוהביט, מערכת המשוב
מודל זה הוא מודל תיאורטי ואין כרגע . הוא המכתיב את אופי הביטויFtsHלפיכך חסרונו של . פועלת
ה בחנו למעל, יחד עם זאת. עדויות ביוכימיות המאשרות או שוללות את קיומם של הפקטור הפלסטידי שהוצעו
שונים ומגוונים אשר ברובם הגדול מגמות השינוי ברמות התעתיק של חלבוני Affymetrix ניסויי 250-מ
FtsH עליה ברמת התעתיק של אחד הגנים מלווה בעליה ברמת , לרוב). נתונים אינם מוצגים( הללו הן דומות
.התעתיק של השני ולהפך
" לבנים"בעלים ראשונים , FtsHמצעות נוגדנים נגד כאשר משווים את החלבונים המזוהים בא
. ניתן לזהות במוטנט חלבונים הקטנים משמעותית מהחלבון המקורי, -WTמהמוטנטים לעומת הירוקים מ
. שלא הצליחו להצטבר בקומפלקס FtsHחלבונים אלו עשויים להיות תוצרי פירוק ראשוני של חלבוני
. בעלים השוניםFtsHה קבועים בין תתי היחידות השונות של ממצאים אלו עשויים לרמז על יחסי מבנ

79
רגישים לשטפי אור חזקים FtsHחלבוני
הראתה כי תנאי עקה שונים ובעיקר שטפי אור חזקים גורמים FtsH בחינת רמות התעתיק של חלבוני
בחינת האופי הקינטי של . )FtsH’s )Sinvany-Villalobo et al., 2004 של mRNA-לעליה ברמות ה
מעידה על אפשרות פגיעה לא רק , 17כפי שהוא מוצג בתמונה , (Fv/Fm)הירידה בפוטנציאל הפוטוסינטתי
פגיעה ביכולת התיקון אלא גם על , כתוצאה מעודף קרינהD1במערכת הפוטוסינתטית בחימצונו של חלבון
בשלב הראשון של עקת האור לרוב בשלבים אלו מתחילה גם סינטזה מוגברת של פאקטורים המעורבים
הגברת יכולת התיקון תשתקף בעקום המייצג את . במניעת ניזקי חימצון כמו למשל פיגמנטים קרטנואידים
כיוון , יתמתן עם הזמןFv/Fmי קצב הירידה של ערכ.כך שעם שיפור יכולת התיקון: קצב גרימת הנזק
ניתן להעריך כי קצב התיקון עולה עם הזמן , שעוצמתו מבוקרת והוא קבוע, שהגורם הישיר לנזק הוא האור
מעקב אחר הכמות . ההתמתנות בקצב הפגיעה תחל לאחר דקות ספורות, לרוב. וגורם למיתון קצב גרימת הנזק
כמותם יורדת במהלך מחצית השעה הראשונה של ההארה בכלורופלסט מלמד כיFtsHהכוללת של חלבוני
חלבונים הם סובסטרטים לתהליכי פירוק כאשר הם נפגמים או כאשר הם משנים ). 17תמונה (ועולה בהמשך
מעיד ככל הנראה על כך שעוצמת אור גבוהה מביאה FtsHפרוק חלבוני . )Adam, 1996(את צורתם
זו האחרונה מתחילה מיד . לפגיעה לא רק במערכת הפוטוסינתטית באופן ישיר אלא גם במערכת התיקון שלה
בשלב הראשון קצב הפירוק גבוה . תהליך שאורך דקות, החלפת היחידות הפגומות ושעתוקן מחדש בגרעיןב
בשלבים מאוחרים יותר גובר קצב הייצור וגוברת . FtsHמקצב הייצור וחלה ירידה בכמות החלבון של
ת גם לא מן הנמנע שמערכת המשוב פועל(פעילותה של מערכת התיקון ומתחיל תהליך של התאוששות
ממצאים אלו עשויים לשמש בעתיד כבסיס למחקרים יישומיים בשיפור יעילותה של המערכת ). במקרה זה
בדומה לנסיונות שיפור שנערכים באמצעות . הפוטוסינתטית ובהתאמה לתנאי גידול בעוצמות אור גבוהות
. רכיבי הגנה אחרים של המערכת הפוטוסינתטית
ים והגנה מנזקי קרינה תפקיד בהתפתחות תילקואידFtsH1-ל
FtsH1 הוא החלבון הראשון מבין חלבוני FtsH ארבידופסיס שזוהה ושובט בצמחי)Lindahl et al.,
בשיטה זו "reverse genetics" שלגיה טעל מנת לנסות וללמוד את תפקידיו בצמח בחרנו באסטר. )1996

80
סריקה של , ftsh1אנו חיפשנו קווי מחדר בגן . מחפשים מוטציות בגן ספציפי ובוחנים את הפנוטיפ המתקבל
ממצאים . לא התבטאftsh1בקו זה הגן . ftsh1העלתה רק קו מחדר אחד בגן קווי מחדר 200,000- למעלה מ
באמצעות דגרדציה של PSII הצביעו על מעורבות אפשרית של החלבון במערכת התיקון של in-vitroמנסויי
מוטנטים הומוזיגוטים למוטציית ההשתקה לא נמצאו רגישים , למרות האמור. D1 - חלבון מרכז הראקציה
תפקיד בהגנה מנזקי קרינה FtsH1-ניתן להסיק שאין ל, לכאורה). 6תמונה (צמות אור גבוהות יותר לעו
אחרים ממלאים את מקומו ולכן אין מזהים פנוטיפ FtsHאו לחלופין שחלבוני , ומנזקי עקות אחרות שנבדקו
. למוטנט
ציות השתקה על רקע על פי הפנוטיפ המתקבל כתוצאה ממוטFtsH1 כאשר בוחנים את תפקידיו של
בצמחים FtsH1קיומו של , כלומר. מתגלה תופעה של החמרת הוריגציה כמו גם רגישות לאור. ftsh2המוטנט
תמיכה נוספת לכך ניתן לראות . גורם להגנה מפני נזקי אור ומסייע ביצירת הכלורופלסטיםFtsH2חסרי
תופעת ההגברה של ). 12תמונה (ורת בהשוואה לביק ftsh2 בצמחיFtsH1- הגבוהות שלmRNA-ברמות ה
mRNAי דיכויו של "ע(רפרסיה : של גן מסויים עקב מוטציה בגן אחר עשויה להיות מוסברת בשתי דרכים
אין כרגע ידע ביוכימי משמעותי , יחד עם זאת. או אינדוקציה של גנים במהלך מסלול ביוכימי, )דכאן קיים
שנבחנו בעבודה זו פעילים FtsHכי כלל חלבוני מאחר ונראה . שיכול ללמד על אפשרויות אלו
ניתן להסיק כי הגברת האחד בחסר של השני איננה נגרמת התוצאה ממסלול ביוכימי אלה , בהטרוקומפלקס
כתוצאה ftsh2 כמו גם החמרת הפנוטיפ של FtsH2- וFtsH1הקרבה הפילוגנטית בין . כתוצאה ממשוב
לשניהם יש תפקידים בהגנה מפני נזקי קרינה -ידיהם חופפים מעידה על כך שתפקFtsH1-מפגיעה בגן ל
.לכן נראה כי מערך המשוב והגיבוי ההדדי הוא זה שמתרחש. ומעורבות בביוגנזה של הכלורופלסט
או / ו FtsH2הכולל לפחות את , מתארגנים בכלורופלסט ככל הנראה בהטרוקומפלקסFtsH1י חלבונ
FtsH8 .היא מערכת מאחר ובמערכת הניסוי שin-vivo לא ניתן להבחין בפעילות השונה מפעילות של
. אחריםFtsH’s פעילות ספצפית השונה מפעילותם של FtsH1-קשה ליחס ל, חלבונים אחרים בקומפלקס
יאפשרו למצוא ולזהות , למשל השתקה של אתר פעיל בשאר חלבוני הקומפלקס, יתכן כי ניסוים עתידים
.חד מהאיזוזימיםפעיליות ספצפיות של כל א

81
FtsH יחידות שונות נחוצות לצבירתו- שתי תת - הוא הטרוקומפלקס
12, הכוללת ארבעה בציואנובקטריה, על ידי קבוצות גנים FtsHביצורים פוטוסנתטיים מקודדים חלבוני
בחיידקים יש . )Adam et al., 2005(ולפחות תשעה באורז , )Sokolenko et al., 2002(בארבידופסיס
ריבוי הגנים ביצורים פוטוסינתטיים עשוי להעיד על הצורך הרב בתיקון קומפלקסים . FtsH-לרוב גן אחד ל
ועל , ם בממברנת התילקואיד שנפגעו או שמיועדים לפירוק במהלך ההתמינות לפלסטידות השונותחלבוניי
הגנים עשויים להיות מוכוונים לכלורופלסט 12 מתוך 9, בארבידופסיס. חשיבותו של תהליך הפרוק
)Sakamoto et al., 2003( , וישנן עדויות כי כל התשעה אכן מתבטאים) ספריותEST ונתוני Affymetrix
,FtsH1:ניתן לזהות רק ארבעה חלבונים בממברנות התילקואיד, ד עם זאתיח). הנתונים אינם מוצגים-
FtsH2 ,FtsH5ו - FtsH8 )Sinvany-Villalobo et al., 2004; Yu et al., 2004(.
ואחריו FtsH2 בכלורופלסט מעלה כי הנפוץ ביותר הוא FtsH בחינת הכמות של כל אחד מחלבוני
FtsH8, FtsH1, FtsH5יתכן כי חמשת הנוספים מצויים בכמות נמוכה או שהם נצברים . בהתאמה
ftsh2 יחיד הראתה כי רק לשניים FtsHבחינת מוטנטים אשר אינם מסוגלים לבטא . בפלסטידות אחרות
טים לעומת זאת המוטנ. ftsh2-כאשר הוריגציה חמורה יותר היא ב. יש פנוטיפ ברור של וריגציה, -ftsh5ו
ftsh1 או ftsh8אינם מציגים פנוטיפ השונה מ -WTהניתוח הפילוגנטי מראה . אפילו במוטנט הכפול שלהם
-כאשר מחסירים ממוטנט המושתק ב. FtsH8- ו FtsH2וכן גם , הם גנים דופליקטיםFtsH5- וFtsH1כי
ftsh2 את FtsH1 או FtsH5ההחמרה חריפה מעט . מתקבלים צמחים בהם חלה החמרה של הפנוטיפ המוטנטי
FtsH1 או FtsH5החסרת אלל יחיד של . חסר אך היא משתנה עם תהליך ההתפתחות FtsH5יותר כאשר
תרומה משמעותית לסך FtsH1- וגם לFtsH5- גם ל כי ניתן להסיק מכך .ftsh2איננה משנה את הפנוטיפ של
ftsh1כאשר משתיקים את. לשני והאחדFtsH2אך תפקידיהם חופפים לתפקידיו של , FtsHהפעילות של
אשר אינם שורדים אלא על גבי מצע מועשר , מתקבלים צמחים חסרי יכולת קיום אוטוטרופיתftsh5וגם את
ניתן , FtsH2 נמוכה באופן משמעותי מכמותו של FtsH5 וגם של FtsH1מאחר שכמות החלבון של . בסוכרוז
משעור הערך שהוסף לכל אחד מהם על ידי השתקת היה לצפות כי התרומה לפנוטיפ הנראה תהיה קטנה

82
FtsH2 . ניתן לחזות כי מוטנטים , אם הגורם הכמותי הוא הגורם היחיד המשפיע על עוצמת הפנוטיפ, כלומר
: החסרה ויסתדרו כך FtsHכפולים יהיו בעלי פנוטיפ חמור בהתאמה לכמות
ftsh1 ftsh8< ftsh5 ftsh1 < ftsh2 ftsh8 <ftsh2 ftsh1 < ftsh2 ftsh5 )מהפנוטיפ החמור אל הקל .(
אשר חלבוניהם משתייכים לשני ענפים פילוגנטים ftsh1 ftsh8, ftsh2 ftsh1, ftsh2 ftsh5בעוד הזוגות
שחלבוניהם שייכים לאותו הענף ftsh2 ftsh8=ftsh5 ftsh1שני הזוגות , שונים אכן מתנהגים כצפוי
סוג (גריעת ענף שלם , כלומר. הרבה מעל לצפוי, י פנוטיפ חמור מאודהם בעל, )כל זוג לענף אחר(הפילוגנטי
"A " או סוג"B (" מונעת כנראה את צבירת הקומפלקס של FtsH אפילו אם קיימים עדיין חלבוני FtsH
. תקינים מהענף השני
Sakamoto et al., 2003 הראו כי FtsHם כנראה ביחסי, בצמחי הארבידופסיס נמצא בהטרוקומפלקס
מממצאי עבודה זו ניתן להציע כי . FtsH2או בהומוקומפלקס של , -FtsH2 ל FtsH5סטויכיומטרים שווים בין
FtsHולא יתקיים מצב שבתא יהיו רק . קומפלקסים- הטרו, נכון יותר, אכן מתפקד בתור הטרוקומפלקס
ללא , י במוטנטים הלבנים באמצעות נוגדנים שונים מעלה כ FtsHניתוח חלבוני, יתרה מכך. הומוקומפלקסים
מודל הסף והמודל ) 31Dתמונה (FtsHלא נצברת אף לא אחת מתתי היחידות של , תלות ברקע הגנטי שלהם
שאינם FtsH’sאו של ) יחיד (FtsHהכמותי מסבירים שניהם באופן חלקי את התופעות הנגרמות מחסר של
והצורך הבסיסי בשני הסוגים מהווים את הצלע ניתן לראות כי יחסי המבנה בקומפלקס . שייכים לאותו ענף
).32תמנוה (השלישית החסרה במודל
מודל כמותי- מבנה ופעילות
: בינהםאפיסטטים מעלה כי מתקימים יחסים FtsHבחינת הגנוטיפים של כלל המוטנטים בגנים השונים של
בתוך כל סוג ישנו דופליקט אחד . קסהקומפלעל מנת לצבור את שניהם נחוצים וחיונים " Bסוג "-ו" Aסוג "
כל עוד היחידה . FtsH5 או FtsH2 אלו הם,העדרו מביא לפנוטיפ וריגנטי, נפוץ וכמותו רבה יותרשהוא
גם . לא יהיה בעל משמעות פנוטיפית)באותו הסוג(השני חסר בגן , היא בעלת אלל אחד תקין לפחותהנפוצה
לא גרמה לפנוטיפ השונה ftsh1 ftsh8ני הענפים במוטנט הכפול החסרה של שני החלבונים הפחות נפוצים מש

83
של היחידה השניהמוטצית השתקה מתקבלים פנוטיפים ב, בתוך הסוגנפוצה בהעדר היחידה ה.מצמחי הבר
, וחמור אף מצמחי האם שלהWT-ניתן לזהות פנונטיפ השונה מכבר כאשר בהחסרת האלל הראשון שלה
" gain of function"לכאורה מתרחש תהליך של . האלל השני לפנוטיפ לטאליפנוטיפ זה מחמיר עם החסרת
לפיכך . כאשר החלבון הפחות נפוץ בכל סוג תופס את תפקידו של החלבון הנפוץ בסוג לפחות באופן חלקי
בחסר (והלבנה מלאה , )בחסר אלל יחיד(מוטציות השתקה שלו מראות את הפנוטיפים של החמרת וריגציה
”loss of function”-כתוצאה מ) האלליםשל שני
עשויה ללמד על חשיבותו של כל חלבון מהמשפחה ftsh תצפית כוללת על המוטנטים בגנים השונים של
במקרה .ftsh5<ftsh2<ftsh1 ftsh2<ftsh2 ftsh5 <<ftsh5 ftsh1=ftsh2 ftsh8: על פי הפנוטיפ הנראה
האלל השני וכנראה גם גנים אחרים במשפחה מפצים על , בלבדשל מוטציית השתקה של גן יחיד באלל אחד
מהווים את הגן -ftsh2 וB" ,ftsh5"או " A",במצב ההומוזיגוטי למוטציה בכל אחד מהסוגים. החסר בחלבון
הדופליקטים שלהם שני. והחסרתם תביא לפנוטיפ נראה התלוי בכמותם, העיקרי מבחינה כמותית בתוך הסוג
ftsh1ו -ftsh8מותם פחותה וחסרונם גורם לפנוטים שאינו שונה מ כWT- . בחסרונה של היחידה העיקרית
בהתאמה לכמות , מתקבל פנוטיפ המעצים את הפנוטיפ שהיה קודם, מענף אחד וגם אחת היחידות מהענף השני
את שתי תתי היחידות שלו מתקבל פנוטיפ החמור B אוAאך כאשר מחסירים מסוג אחד . החלבון החסר
- בכלורופלסט הוא הטרו-FtsHמכאן ניתן להציע כי מבנה קומפלקס ה. פנוטיפ לטאלי לבן–הרבה מהצפוי ב
היחסים הסטויכיומטריים כפי שהצוגו , )mAAA )Arlt et al., 1998הקסאמר בדומה להטרוקומפלקס של
עשוים לרמז על הסטיכיומטריה הממוצעת של הקומפלקס (Sinvany-Villalobo et al., 2004) יעל יד
- וFtsH5" ( Aסוג"דות של יחד עם שתי יחיFtsH8)- וB) "FtsH2סוג "ארבע יחידות של : כדלקמן
(FtsH1 .ותמיד , אך הממוצע הכולל יהיה זה שהוצג, לא מן הנמנע שבקומפלקס מסויים היחסים יהיו שונים
כימות עוצמת הפנוטיפ המתבטא בכמות הכלורופיל . ידרשו שני הסוגים על מנת לאפשר את יצירת הקומפלקס
להטרוקומפלקס המשותף כפי FtsH ל אחד מחלבוני מעידה על התרומה הכמותית של כ) 31Cתמונה (בעלה
).32תמונה (שזה מוצג במודל

84
FtsHההטרוקומפלס של מודל מבנה .32 תמונה
מבנהו בדומה ל,הקסמר שמורכב- הטרו. Krzywda et al., (2002(בתור הקסמר על פי נצבר ופעיל רק FtsHכימציע המודל
ןהיבינהסטויכיומטריים היחסים . FtsH8- וB FtsH2- וFtsH5-וA- FtsH1 שתיהן חיוניותש יחידות י משתי תת,מיטוכונדריותב
. להמצא בקומפלקסים היחידות חייבני סוגי תתיעשויים להשתנות אך ש
יש לזכור כי אין זה המצב בכל . לשאלה מדוע יש ריבוי גנים במשפחה אין תשובות מובנות מאליהן
חלה , ועם ההתפתחות לייצורים פוטוסינתטיים רב תאיים, בחיידקים הוא חלבון יחידFtsH: האורגניזמים
לא מן הנמנע כי . סטובעיקר בגנים שתוצריהם עשויים להיות מוכוונים לכלורופל, התרחבות בגודל המשפחה
תנאי הפעילות בכלורופלסט והסביבה הקיצונית בה פעילות זו מתרחשת מחייבים ביטוי רב של חלבונים
עותקים רבים ומניעת הצורך (במקרה כזה צפוי היה כי החלבונים יהיו מקודדים בגנום הפלסטידי . בסביבה זו
הגרעיני היו יתרונות אבולוציוניים לקיומם בצורה משנדדו הגנים הללו מן הגנום הפלסטידי אל הגנום). ביבוא
של ויצירת הטרו קומפלקס המורכב מלפחות שני סוגים-FtsHייתכן כי ההתמינות לשני סוגי ה. של משפחה
עם מגוון תנאי סביבה באמצעות שינוי בהרכב תתי , מאפשרת התמודדות טובה יותר, "”B-ו" A"תתי יחידות
חישוב שערכנו בהתאם למודל . או העשרה בסוג מסויים על חשבון הסוג השניהיחידות בקומפלקס והחלפה
סוגי קומפלקסים שונים 20מעלה כי עשויים להתקיים לפחות ) בלי המשתנה של המיקום בקומפלקס(שהצענו
ומה שמשתנה זה מספר תת היחידות " B"וגם את " A סוג " הם מכילים גם את –העונים על התנאי היסודי
אם מוסיפים גם את משתנה היחסים הסטויכיומטריים בין הקומפלקסים האפשרויות המתקבלות . גמכל סו
FtsH2 or FtsH8
FtsH1 or FtsH5
Structural model for FtsH heterocomplex
B
A
FtsH

85
, במיוחד במינים המתרבים ברביה עצמית, מנגנון ריבוי הגנים עשוי גם לאפשר סבילות למוטציות. רבות מאוד
.גורם המסייע להישרדותו, מאחר שריבוי גנים מאפשר שונות טבעית בתוך הפרט
.)Carol et al., 1999; Moore et al., 2000(בדומה לצמחים לבנים אחרים , בנים עקריםהצמחים הל
טים או מחסר של הגן מנגנון עקרות זה אינו מובן דיו ולא ברור אם העקרות נגרמת מחסר של כלורופלס
בחינת מנגנון העקרות מראה כי יש פגיעה בהתפתחות אברי המין של הפרח וגם במנגנון הפצת . הספציפי
אך לא נבחנה חיוניותן , לא נראה כי הגמטות הצעירות עצמן פגועות, יחד עם זאת). 30 ,29תמונות (הגמטות
מצליחה להתקיים וליצור זרע FtsH -ומפלקס ההעובדה כי זיגוטה החסרה את ק. באמצעות הפרייה חיצונית
מתאימה , )בדומה למצע גידול מועשר בסוכרוז(ובלבד שהיא גדלה על צמח אם המסוגל לתמוך בה ) לבן(
העקרות נגרמת בגלל בעיה , ככל הנראה. לתצפית שמראה כי תאי המין והגמטופיטים נראים תקינים
.ה של הגמטותפיזיולוגית בהפצה ולא בגלל בעיה גנטית ביציר
סיכום
FtsHלא נידונה בעבודה זו והיא כוללת שלושה חלבוני , הראשונה: בצמחים זו משפחה בת שתי קבוצות
FtsHשני סוגים : בה יש שלושה סוגים, של החלבונים כלורופלסטיים, השניה. המוכוונים למיטוכונדריה
מתקיימים יחסי הדדיות בין . ו אינם ידועיםוסוג שלישי שתפקידי) שעבודה זו התמקדה בהם(חיוניים
יביא גם להגברת , גורם שיביא להגברת השיעתוק של אחד הגנים בקבוצה, החלבונים בתוך המשפחה ולרוב
היחסים בין האיזוזימים השונים נובעים ככל הנראה מן העובדה שהם מתפקדים יחד . השיעתוק הגנים האחרים
הם החלבונים היחידים שבפועל FtsH8 אוFtsH2כי , לשם הדיון, גם אם נניח. בהטרוקומפלקס משותף
לפגיעה , בגלל צבירתם המשותפת בקומפלקס, תגרום-FtsH5הרי שפגיעה ב , D1מפרקים את חלבון
inכלומר לא ניתן במערכת . מעורב ישירות בפעילותFtsH5 ולפיכך יראה כאילו גם FtsH2בצבירה של
vivoעילות ולמבנה להפריד בין הפנוטיפ לפ.

86
ככל , הוא מעורבות בפירוק של חלבונים ממברנלים בממברנת התילקואידFtsH תפקידו הברור של
. הוא מעורב בביוגנזה של התילקואיד ונוכחותו חיונית ליצירת הממברנות. הנראה כחלק ממערכת התיקון
.לא יווצרו תילקואידים והצמחים יהיו הטרוטרופים ועקרים, בהעדרו
רגישות זו גורמת להחלפה מהירה . כנראה בגלל חימצון, רגישים לעוצמות אור גבוהותFtsH חלבוני
הקיום של משפחת גנים וריבוי האיזוזימים עשוי להעיד על חשיבותם האיכותית . שלהם ביחידות חדשות
פלסטידות שונות כמו ואולי אף לסוגי, והכמותית וכפי הנראה גם על חיוניות ההתאמה למגוון תנאים
.כרומופלסטידות וכדומה
לברר האם פעילותם , השוניםFtsH מחקרים עתדיים יוכלו לשפוך אור על התפקיד המדוייק של חלבוני
בכך ניתן יהיה . ומיהם הסובסטרטים השונים שהקומפלקס מפרקATPaseפרוטאוליתית או פעילות של
. FtsHמי של פעילות להבין טוב יותר את המנגנון הביוכי

87
רשימת ספרות Adam, Z. (1996). Protein stability and degradation in chloroplasts. Plant Mol Biol 32, 773-783.
Adam, Z., Adamska, I., Nakabayashi, K., Ostersetzer, O., Haussuhl, K., Manuell, A., Zheng, B., Vallon, O., Rodermel, S.R., Shinozaki, K., and Clarke, A.K. (2001). Chloroplast and mitochondrial proteases in Arabidopsis. A proposed nomenclature. Plant Physiol 125, 1912-1918.
Adam, Z., Zaltsman, A., Sinvany-Villalobo, G., and Sakamoto, W. (2005). FtsH proteases in chloroplasts and cyanobacteria. Physiologia Plantarum123, 386-396.
Akiyama, Y., and Ito, K. (2001). Roles of homooligomerization and membrane association in ATPase and proteolytic activities of FtsH in vitro. Biochemistry 40, 7687-7693.
Akiyama, Y., Yoshihisa, T., and Ito, K. (1995). FtsH, a membrane-bound ATPase, forms a complex in the cytoplasmic membrane of Escherichia coli. J Biol Chem 270, 23485-23490.
Alonso, J.M., Stepanova, A.N., Leisse, T.J., Kim, C.J., Chen, H., Shinn, P., Stevenson, D.K., Zimmerman, J., Barajas, P., Cheuk, R., Gadrinab, C., Heller, C., Jeske, A., Koesema, E., Meyers, C.C., Parker, H., Prednis, L., Ansari, Y., Choy, N., Deen, H., Geralt, M., Hazari, N., Hom, E., Karnes, M., Mulholland, C., Ndubaku, R., Schmidt, I., Guzman, P., Aguilar-Henonin, L., Schmid, M., Weigel, D., Carter, D.E., Marchand, T., Risseeuw, E., Brogden, D., Zeko, A., Crosby, W.L., Berry, C.C., and Ecker, J.R. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301, 653-657.
Arlt, H., Tauer, R., Feldmann, H., Neupert, W., and Langer, T. (1996). The YTA10-12 complex, an AAA protease with chaperone-like activity in the inner membrane of mitochondria. Cell 85, 875-885.
Arlt, H., Steglich, G., Perryman, R., Guiard, B., Neupert, W., and Langer, T. (1998). The formation of respiratory chain complexes in mitochondria is under the proteolytic control of the m-AAA protease. EMBO J 17, 4837-4847.
Arnon, D.I., and Whatley, F.R. (1949). Is chloride a coenzyme of photosynthesis? Science 110, 554-556.
Bailey, S., Thompson, E., Nixon, P.J., Horton, P., Mullineaux, C.W., Robinson, C., and Mann, N.H. (2002). A critical role for the Var2 FtsH homologue of

88
Arabidopsis thaliana in the photosystem II repair cycle in vivo. J Biol Chem 277, 2006-2011.
Barkan, A., Walker, M., Nolasco, M., and Johnson, D. (1994). A nuclear mutation in maize blocks the processing and translation of several chloroplast mRNAs and provides evidence for the differential translation of alternative mRNA forms. EMBO J 13, 3170-3
Blankenship, R.E., and Hartman, H. (1998). The origin and evolution of oxygenic photosynthesis. Trends Biochem Sci 23, 94-97.
Butler, W.L., and Kitajima, M. (1975). Fluorescence quenching in photosystem II of chloroplasts. Biochim Biophys Acta 376, 116-125.
Cahoon, A.B., Cunningham, K.A., and Stern, D.B. (2003). The plastid clpP gene may not be essential for plant cell viability. Plant Cell Physiol 44, 93-95.
Carol, P., Stevenson, D., Bisanz, C., Breitenbach, J., Sandmann, G., Mache, R., Coupland, G., and Kuntz, M. (1999). Mutations in the Arabidopsis gene IMMUTANS cause a variegated phenotype by inactivating a chloroplast terminal oxidase associated with phytoene desaturation. Plant Cell 11, 57-68.
Casari, G., De Fusco, M., Ciarmatori, S., Zeviani, M., Mora, M., Fernandez, P., De Michele, G., Filla, A., Cocozza, S., Marconi, R., Durr, A., Fontaine, B., and Ballabio, A. (1998). Spastic paraplegia and OXPHOS impairment caused by mutations in paraplegin, a nuclear-encoded mitochondrial metalloprotease. Cell 93, 973-983.
Chaal, B.K., Mould, R.M., Barbrook, A.C., Gray, J.C., and Howe, C.J. (1998). Characterization of a cDNA encoding the thylakoidal processing peptidase from Arabidopsis thaliana. Implications for the origin and catalytic mechanism of the enzyme. J Biol Chem 273, 689-692.
Chen, M., Choi, Y., Voytas, D.F., and Rodermel, S. (2000). Mutations in the Arabidopsis VAR2 locus cause leaf variegation due to the loss of a chloroplast FtsH protease. Plant J 22, 303-313.
Cheregi, O., Sicora, C., Kos, P.B., Nixon, P.J., and Vass, I. (2005). The FtsH protease is required for the repair of Photosystem II in the cyanbacterium Synechocystis 6803 damaged UV-B radiation. BMC Plant Biology 5, 1-2.
Chung, C.H., Waxman, L., and Goldberg, A.L. (1983). Studies of the protein encoded by the lon mutation, capR9, in Escherichia coli. A labile form of the ATP-dependent protease La that inhibits the wild type protease. J Biol Chem 258, 215-221.

89
Clarke, A.K., MacDonald, T.M., and Sjogren, L.L.E. (2005). The ATP-dependent Clp protease in chloroplasts of higher plants. Physiol Plant 123, 406-412.
Cline, K., and Henry, R. (1996). Import and routing of nucleus-encoded chloroplast proteins. Annu Rev Cell Dev Biol 12, 1-26.
Halperin, T., Ostersetzer, O., and Adam, Z. (2001). ATP-dependent association between subunits of Clp protease in pea chloroplasts. Planta 213, 614-619.
Haussuhl, K., Andersson, B., and Adamska, I. (2001). A chloroplast DegP2 protease performs the primary cleavage of the photodamaged D1 protein in plant photosystem II. EMBO J 20, 713-722.
Hershko, A., and Ciechanover, A. (1998). The ubiquitin system. Annu Rev Biochem 67, 425-479.
Huffaker, R.C., and Peterson, L.W. (1974). Protein turnover in plants and possible means of Its Regulation. Annu Rev of Plant Physiol 25, 363-392.
Hwang, B.J., Woo, K.M., Goldberg, A.L., and Chung, C.H. (1988). Protease Ti, a new ATP-dependent protease in Escherichia coli, contains protein-activated ATPase and proteolytic functions in distinct subunits. J Biol Chem 263, 8727-8734.
Ito, K., and Akiyama, Y. (2005). Cellular functions, mechanism of action, and regulation of FtsH Protease. Annua Rev of Microbiol 253, 212-224.
Izhaki, A., Swain, S.M., Tseng, T.S., Borochov, A., Olszewski, N.E., and Weiss, D. (2001). The role of SPY and its TPR domain in the regulation of gibberellin action throughout the life cycle of Petunia hybrida plants. Plant J 28, 181-190.
Itzhaki, H., Naveh, L., Lindahl, M., Cook, M., and Adam, Z. (1998). Identification and characterization of DegP, a serine protease associated with the luminal side of the thylakoid membrane. J Biol Chem 273, 7094-7098.
Jansen, M.A., Mattoo, A.K., and Edelman, M. (1999). D1-D2 protein degradation in the chloroplast. Complex light saturation kinetics. Eur J Biochem 260, 527-532.
Jenal, U., and Hengge-Aronis, R. (2003). Regulation by proteolysis in bacterial cells. Curr Opin Microbiol 6, 163-172.
.

90
Kaltwasser, M., Wiegert, T., and Schumann, W. (2002). Construction and application of epitope- and green fluorescent protein-tagging integration vectors for Bacillus subtilis. Appl Environ Microbiol 68, 2624-2628.
Katayama, Y., Kasahara, A., Kuraishi, H., and Amano, F. (1990). Regulation of activity of an ATP-dependent protease, Clp, by the amount of a subunit, ClpA, in the growth of Escherichia coli cells. J Biochem 108, 37-41.
Katayama, Y., Gottesman, S., Pumphrey, J., Rudikoff, S., Clark, W.P., and Maurizi, M.R. (1988). The two-component, ATP-dependent Clp protease of Escherichia coli. Purification, cloning, and mutational analysis of the ATP-binding component. J Biol Chem 263, 15226-15236.
Keegstra, K., and Cline, K. (1999). Protein import and routing systems of chloroplasts. Plant Cell 11, 557-570.
Kettunen, R., Tyystjarvi, E., and Aro, E.M. (1996). Degradation pattern of photosystem II reaction center protein D1 in intact leaves. The major photoinhibition-induced cleavage site in D1 polypeptide is located amino terminally of the DE loop. Plant Physiol 111, 1183-1190.
Kihara, A., Akiyama, Y., and Ito, K. (1997). Host regulation of lysogenic decision in bacteriophage lambda: transmembrane modulation of FtsH (HflB), the cII degrading protease, by HflKC (HflA). Proc Natl Acad Sci U S A 94, 5544-5549.
Kitajima, M., and Butler, W.L. (1975). Quenching of chlorophyll fluorescence and primary photochemistry in chloroplasts by dibromothymoquinone. Biochim Biophys Acta 376, 105-115.
Ko, K., and Cashmore, A.R. (1989). Targeting of proteins to the thylakoid lumen by the bipartite transit peptide of the 33 kd oxygen-evolving protein. EMBO J 8, 3187-3194.
Kruft, V., Eubel, H., Jansch, L., Werhahn, W., and Braun, H.P. (2001). Proteomic approach to identify novel mitochondrial proteins in Arabidopsis. Plant Physiol 127, 1694-1710.
Krysan, P.J., Young, J.C., and Sussman, M.R. (1999). T-DNA as an insertional mutagen in Arabidopsis. Plant Cell 11, 2283-2290.
Krzywda, S., Brzozowski, A.M., Verma, C., Karata, K. ,Ogura, T., and Wilkinson,
A.J. (2002). The crystal structure of the AAA domain of the ATP-dependent protease FtsH of Escherichia coli at 1.5 A resolution. Structure 10, 1073-1083.

91
Kuroda, H., and Maliga, P. (2003). The plastid clpP1 protease gene is essential for plant development. Nature 425, 86-89.
Langer, T. (2000). AAA proteases: cellular machines for degrading membrane proteins. Trends in Biochemical Sciences 25, 247-251.
Laskowska, E., Kuczynska-Wisnik, D., Skorko-Glonek, J., and Taylor, A. (1996). Degradation by proteases Lon, Clp and HtrA, of Escherichia coli proteins aggregated in vivo by heat shock; HtrA protease action in vivo and in vitro. Mol Microbiol 22, 555-571.
Leonhard, K., Stiegler, a., Neupert, W., and Langer, T. (1999). Chaperone-like activity of the AAA domain of the yeast Yme1 AAA protease. Nature 398, 348-351.
Li, X., Qian, Q., Fu, Z., Wang, Y., Xiong, G., Zeng, D., Wang, X., Liu, X., Teng, S., Hiroshi, F., Yuan, M., Luo, D., Han, B., and Li, J. (2003). Control of tillering in rice. Nature 422, 618-621.
Lin, C.-S., Lin, C.-C., and Chang, W.-C. (2004). Effect of thidiazuron on vegetative tissue-derived somatic embryogenesis and flowering of bamboo Bambusa edulis. Plant Cell, Tissue and Organ Culture 76, 75-82.
Lindahl, M., Tabak, S., Cseke, L., Pichersky, E., Andersson, B., and Adam, Z. (1996). Identification, characterization, and molecular cloning of a homologue of the bacterial FtsH protease in chloroplasts of higher plants. J Biol Chem 271, 29329-29334.
Lindahl, M., Spetea, C., Hundal, T., Oppenheim, A.B., Adam, Z., and Andersson, B. (2000). The thylakoid FtsH protease plays a role in the light-induced turnover of the photosystem II D1 protein. Plant Cell 12, 419-431.
Lupas, A., Flanagan, J.M., Tamura, T., and Baumeister, W. (1997). Self-compartmentalizing proteases. Trends Biochem Sci 22, 399-404.
Mann, N.H., Novac, N., Mullineaux, C.W., Newman, J., Bailey, S., and Robinson, C. (2000). Involvement of an FtsH homologue in the assembly of functional photosystem I in the cyanobacterium Synechocystis sp. PCC 6803. FEBS Lett 479, 72-77.
Melis, A. (1999). Photosystem-II damage and repair cycle in chloroplasts: what modulates the rate of photodamage in vivo? Trends Plant Sci 4, 130-135.

92
Nakamura, Y., Kaneko, T., Hirosawa, M., Miyajima, N., and Tabata, S. (1998). CyanoBase, a www database containing the complete nucleotide sequence of the genome of Synechocystis sp. strain PCC6803. Nucleic Acids Res 26, 63-67.
Moore, M., Harrison, M.S., Peterson, E.C., and Henry, R. (2000). Chloroplast Oxa1p homolog albino3 is required for post-translational integration of the light harvesting chlorophyll-binding protein into thylakoid membranes. J Biol Chem 275, 1529-1532.
Neuwald, A.F., Aravind, L., Spouge, J.L., and Koonin, E.V. (1999). AAA+: A class of chaperone-like ATPases associated with the assembly, operation, and disassembly of protein complexes. Genome Res 9, 27-43.
Nixon, P.J., Barker, M., Boehm, M., de Vries, R., and Komenda, J. (2005). FtsH-mediated repair of the photosystem II complex in response to light stress. J Exp Bot 56, 357-363.
Ogura, T., and Wilkinson, A.J. (2001). AAA+ superfamily ATPases: common structure--diverse function. Genes Cells 6, 575-597.
Ogura, T., Tomoyasu, T., Yuki, T., Morimura, S., Begg, K.J., Donachie, W.D., Mori, H., Niki, H., and Hiraga, S. (1991). Structure and function of the ftsH gene in Escherichia coli. Res Microbiol 142, 279-282.
Ostersetzer, O., and Adam, Z. (1997). Light-stimulated degradation of an unassembled Rieske FeS protein by a thylakoid-bound protease: the possible role of the FtsH protease. Plant Cell 9, 957-965.
Ostersetzer, O., Tabak, S., Yarden, O., Shapira, R., and Adam, Z. (1996). Immunological detection of proteins similar to bacterial proteases in higher plant chloroplasts. Eur J Biochem 236, 932-936.
Paul, M.F., and Tzagoloff, A. (1995). Mutations in RCA1 and AFG3 inhibit F1-ATPase assembly in Saccharomyces cerevisiae. FEBS Lett 373, 66-70.
Porankiewicz, J., Wang., J., and Clarke, A.K. (1999). New insights into the ATP-dependent Clp protease: Escherichia coli and beyond. Mol Microbiol 32, 449-458
Richter, S., and Lamppa, G.K. (1999). Stromal processing peptidase binds transit peptides and initiates their ATP-dependent turnover in chloroplasts. J Cell Biol 147, 33-44.

93
Rotanova, T.V., Melnikov, E.E., Khalatova, A.G., Makhovskaya, O.V., Botos, I., Wlodawer, A., and Gustchina, A. (2004). Classification of ATP-dependent proteases Lon and comparison of the active sites of their proteolytic domains. Eur J Biochem 271, 4865-4871.
Saikawa, N., Ito, K., and Akiyama, Y. (2002). Identification of glutamic acid 479 as the gluzincin coordinator of zinc in FtsH (HflB). Biochemistry 41, 1861-1868.
Sakamoto, W., Tamura, T., Hanba-Tomita, Y., and Murata, M. (2002). The VAR1 locus of Arabidopsis encodes a chloroplastic FtsH and is responsible for leaf variegation in the mutant alleles. Genes Cells 7, 769-780.
Sakamoto, W., Zaltsman, A., Adam, Z., and Takahashi, Y. (2003). Coordinated regulation and complex formation of yellow variegated1 and yellow variegated2, chloroplastic FtsH metalloproteases involved in the repair cycle of photosystem II in Arabidopsis thylakoid membranes. Plant Cell 15, 2843-2855.
Sambrook, P.N., Cohen, M.L., Eisman, J.A., Pocock, N.A., Champion, G.D., and Yeates, M.G. (1989). Effects of low dose corticosteroids on bone mass in rheumatoid arthritis: a longitudinal study. Ann Rheum Dis 48, 535-538.
Santos, D., and De Almeida, D.F. (1975). Isolation and characterization of a new temperature-sensitive cell division mutant of Escherichia coli K-12. J Bacteriol 124, 1502-1507.
Seo, S., Okamoto, M., Iwai, T., Iwano, M., Fukui, K., Isogai, A., Nakajima, N., and Ohashi, Y. (2000). Reduced levels of chloroplast FtsH protein in tobacco mosaic virus-infected tobacco leaves accelerate the hypersensitive reaction. Plant Cell 12, 917-932.
Shah, Z.H., Hakkaart, G.A., Arku, B., de Jong, L., van der Spek, H., Grivell, L.A., and Jacobs, H.T. (2000). The human homologue of the yeast mitochondrial AAA metalloprotease Yme1p complements a yeast yme1 disruptant. FEBS Lett 478, 267-270.
Shanklin, J., DeWitt, N.D., and Flanagan, J.M. (1995). The stroma of higher plant plastids contain ClpP and ClpC, functional homologs of Escherichia coli ClpP and ClpA: an archetypal two-component ATP-dependent protease. Plant Cell 7, 1713-1722.
Shikanai, T., Shimizu, K., Ueda, K., Nishimura, Y., Kuroiwa, T., and Hashimoto, T. (2001). The chloroplast clpP gene, encoding a proteolytic subunit of ATP-dependent protease, is indispensable for chloroplast development in tobacco. Plant Cell Physiol 42, 264-273.

94
Shotland, Y., Koby, S., Teff, D., Mansur, N., Oren, D.A., Tatematsu, K., Tomoyasu, T., Kessel, M., Bukau, B., Ogura, T., and Oppenheim, A.B. (1997). Proteolysis of the phage lambda CII regulatory protein by FtsH (HflB) of Escherichia coli. Mol Microbiol 24, 1303-1310.
Silva, P., Thompson, E., Bailey, S., Kruse, O., Mullineaux, C.W., Robinson, C., Mann, N.H., and Nixon, P.J. (2003). FtsH is involved in the early stages of repair of photosystem II in Synechocystis sp PCC 6803. Plant Cell 15, 2152-2164.
Singer, T., and Burke, E. (2003). High-throughput TAIL-PCR as a tool to identify DNA flanking insertions. Methods Mol Biol 236, 241-272.
Sinvany-Villalobo, G., Davydov, O., Ben-Ari, G., Zaltsman, A., Raskind, A., and Adam, Z. (2004). Expression in multigene families. Analysis of chloroplast and mitochondrial proteases. Plant Physiol 135, 1336-1345.
Smalle, J., and Vierstra, R.D. (2004). The ubiquitin 26S proteasome proteolytic pathway. Annu Rev Plant Biol 55, 555-590.
Sokolenko, A., Pojidaeva, E., Zinchenko, V., Panichkin, V., Glaser, V.M., Herrmann, R.G., and Shestakov, S.V. (2002). The gene complement for proteolysis in the cyanobacterium Synechocystis sp. PCC 6803 and Arabidopsis thaliana chloroplasts. Curr Genet 41, 291-310.
Southern, E.M. (1975). Detection of specific sequences among DNA fragments separated by gel electrophoresis. J Mol Biol 98, 503-517.
Steglich, G., Neupert, W., and Langer, T. (1999). Prohibitins regulate membrane protein degradation by the m-AAA protease in mitochondria. Mol Cell Biol 19, 3435-3442.
Sussman, M.R., Amasino, R.M., Young, J.C., Krysan, P.J., and Austin-Phillips, S. (2000). The Arabidopsis knockout facility at the University of Wisconsin-Madison. Plant Physiol 124, 1465-1467. Takechi, K., Sodmergen, Murata, M., Motoyoshi, F., and Sakamoto, W. (2000). The YELLOW VARIEGATED (VAR2) locus encodes a homologue of FtsH, an ATP-dependent protease in Arabidopsis. Plant Cell Physiol 41, 1334-1346.
Tomoyasu, T., Yamanaka, K., Murata, K., Suzaki, T., Bouloc, P., Kato, A., Niki, H., Hiraga, S., and Ogura, T. (1993). Topology and subcellular localization of FtsH protein in Escherichia coli. J Bacteriol 175, 1352-1357.
Vierstra, R.D. (1993). Illuminating Phytochrome Functions (There Is Light at the End of the Tunnel). Plant Physiol 103, 679-684.

95
Vierstra, R.D. (2003). The ubiquitin/26S proteasome pathway, the complex last chapter in the life of many plant proteins. Trends Plant Sci 8, 135-142.
Weber, E.R., Hanekamp, T., and Thorsness, P.E. (1996). Biochemical and functional analysis of the YME1 gene product, an ATP and zinc-dependent mitochondrial protease from S. cerevisiae. Mol Biol Cell 7, 307-317.
Wickner, S., Maurizi, M.R., and Gottesman, S. (1999). Posttranslational quality control: folding, refolding, and degrading proteins. Science 286, 1888-1893.
Yu, F., Park, S., and Rodermel, S.R. (2004). The Arabidopsis FtsH metalloprotease gene family: interchangeability of subunits in chloroplast oligomeric complexes. Plant J 37, 864-876.
Yu, F., Park, S., and Rodermel, S.R. (2005). Functional Redundancy of AtFtsH Metalloproteases in Thylakoid Membrane Complexes. Plant Physiol. (in press)
Zer, H., Prasil, O., and Ohad, I. (1994). Role of plastoquinol oxidoreduction in regulation of photochemical reaction center II D1 protein turnover in vivo. J Biol Chem 269, 17670-17676.
.

96
This work was carried out under the supervision of Prof. Zach Adam
The Robert H. Smith Institute of Plant Sciences and Genetics in
Agriculture
Faculty of Agricultural, Food and Environmental Quality Sciences
The Hebrew University of Jerusalem

97
Characterization of the chloroplast FtsH protease in Arabidopsis
thaliana plants
FtsH is an ATP-dependent membrane metalloprotease belonging to the big AAA
(ATPase associated with various cellular activities) family. It is the only essential ATP-
dependent protease in Escherichia coli. FtsH forms hexamer complexes in E. coli and in
other plastids from eukaryotic cells. Chloroplastic FtsH was first identified
immunologically in the late 1990s from spinach leaves, and characterized as an integral
thylakoid membrane protein whose expression is light-dependent (Lindahl et al., 1996).
The FtsH protein is bound to the stroma-exposed lamellae, with its ATP- and Zn2+-
binding domains exposed to the soluble stroma. The first indication of chloroplastic
FtsH's physiological function came from an in-vitro study. After import into isolated
chloroplasts, unassembled Rieske Fe-S protein was rapidly degraded by a protease which
appeared to be FtsH (Ostersetzer and Adam 1997). Another in-vitro study suggested that
FtsH is involved in the degradation of the D1 protein from the PSII reaction center. This
degradation could be inhibited by removing FtsH from the membrane, and restored by
adding recombinant FtsH (Lindahl et al. 2000). Genes that encode FtsH proteins
constitute a small gene family in higher plants. The Arabidopsis nuclear genome contains
12 such genes (Adam et al., 2001; Sokolenko et al. 2002). Of these, the products of three
(FtsH3, 4 and 10) are targeted to the mitochondria, whereas the other nine (FtsH1, 2, 5-9,
11 and 12) are capable of entering the chloroplast, as revealed by transient expression
assay of GFP-fusions (Sakamoto et al. 2003). However, only four of the isozymic

98
proteins have been found in the native chloroplast under two different 2-D experimental
systems (Sinvany-Villalobo et al. 2004; Yu et al. 2004).
I began my research in 2000, with the main question: What are the relationships
between the different isomers? What is the function of FtsH proteins in chloroplasts?
Does the function of one isomer correspond to those of others?
As a research strategy, we used reverse genetics to try and establish connections
between genomic null mutations at specific loci and phenotype. We used T-DNA
insertion lines to cause gene disruptions: this is a powerful tool for obtaining knock-out
mutants that help ascertain the biological function of the numerous uncharacterized open
reading frames (ORFs). At the time, we only had FtsH1, and I used its primers to screen
the UW-Madison Arabidopsis knock-out facility. I found one insertion line in the ftsh1
locus. Seeds for this line were organized in pools of 10; I found a single plant to serve as
the donor for back-crosses with the WT, and after a few selfing generations I obtained a
T-DNA homozygous plant. By sequences of TAIL-PCR product, the point of insertion
was revealed, but when FtsH protein level was examined, no difference was observed
with the WT, even if the mRNA level was undetectable by RT-PCR. I attempted to
determine this null mutant's effect on photosynthesis. But in all of the experiments,
results were similar to the WT. I concluded that FtsH1 has no vital function under these
experimental conditions. I searched for other FtsH insertion lines by BLAST against the
Syngenta database, and found insertions to all chloroplast FtsHs. Mutations in two of
these, FtsH2 and 5, demonstrated a visible phenotype of variegated leaves, with the
phenotype of the FtsH2 mutant being more pronounced. Moreover, the degree of
variegation appeared to be dependent upon developmental stage and environmental

99
factors, suggesting an intricate relationship between the different gene products. To
explore this, developmental and light effects on the accumulation of FtsH protease were
studied in WT and FtsH2-mutant plants. Whereas cotyledons of the mutant were
indistinguishable from those of the WT, its first true leaves were almost completely
white. Subsequent leaves contained increasing proportions of green sectors. Analysis of
the mRNA of the four FtsH genes, in the cotyledons and first and second leaves of WT
and mutant plants, revealed that: (i) transcript level increases during development, and
(ii) transcript level in the mutant is higher than in the WT. FtsH protein level in the
mutant was ca. 50% of that found in the WT, whereas the levels of other thylakoid
proteins were the same. In individual leaves, the level of FtsH protein increased during
development as well. Exposure of seedlings to different light intensities did not affect the
degree of variegation, suggesting that it is due to a defect in chloroplast development
rather than photobleaching. Examination of FtsH protein during exposure to high light
revealed a decrease in its level, concomitant with a decrease in PSII potential, suggesting
that the kinetics of photoinhibition reflects not only photodamage to PSII and induction
of protective mechanisms, but also a decrease in repair capacity due to a reduction in the
level of FtsH protease.
The chloroplasts contain four FtsH proteins. Mutants in two of these, FtsH2 and 5,
demonstrated a visible phenotype of variegated leaves; mutants in FtsH 1 and 8 had no
visible phenotype. The FtsH2 and 5 mutants also demonstrated higher sensitivity to
photoinhibition than WT plants. To further explore the function of FtsH protease, we
generated double mutants of the four aforementioned genes. Null mutants lacking both
FtsH1 and FtsH8 developed and functioned normally, whereas the phenotype of the ftsh2

100
mutant was aggravated in the absence of either FtsH5 or FtsH1: double mutants were
more variegated and had a lower proportion of normal chloroplasts. Homozygous double
mutants of FtsH2 and FtsH8 were unable to develop normal chloroplasts and could be
maintained only if supplemented with sucrose. A similar effect on viability and
chloroplast development was observed in FtsH1 and FtsH5 double mutants. Furthermore,
H2+ H8+ sets of double mutants had a higher proportion of aborted seeds, and seeds
that were unable to germinate properly, suggesting that FtsH might be involved in early
stages of plant development, thylakoid formation and chloroplast development. The
combinations of genes leading to the most severe phenotypes suggest that FtsH
complexes are composed of two types of subunits, each encoded by two duplicated
genes. The duplicated genes are interchangeable, but both types of subunit must be
present to obtain a functional complex. A model accounting for the differential effects of
the loss of the different genes is presented.
Major conclusions of this work:
1. Functional complexes of the FtsH gene family are composed of 'Type A'---FtsH1
and FtsH5, and 'Type B'---FtsH2 and FtsH8 subunits. Both types are essential.
2. FtsH functions as a six-unit complex, an average four units of which are FtsH2
and/or FtsH8.
3. FtsH take part in the repair of PSII following photoinhibition. During the initial
period of photoinhibition, the amount of FtsH itself decreases.

101
4. The accumulation of FtsH protease during development is a dynamic process, and
the presence of a number of FtsH genes allows them to effectively compensate for
one another.
5. If one subunit of FtsH is missing, feedback leads to upregulation of the other
FtsHs.

102
List of contents
Abstract 2-5
Introduction 6-20
Proteolysis 6
Proteolysis in eukaryote – the Ubiquitin system 7
Degradation by the Proteasome as controlled system
The AAA Family 10
Proteolysis in prokaryote 11
FtsH proteases in bacteria cells 12
Mitochondrial FtsH 13
Cyanobacteria FtsH 15
Plant chloroplast-structure and function 15
Proteolysis in chloroplast 16
Plant FtsH 18
Research goals 21
Method and materials 22-33
Plant material 22
Seeds sterilization, aseptic conditions and selection 22 growth
Screening and identification of FtsH1 null mutant 23
Crosses for double mutant 25
Isolation of DNA from T-DNA insertion line and PCR analysis 25
Gel Analysis of DNA 26
Cloning of TAIL-PCR product 26

103
Prepare competent E. coli cells and transformation 27
Protein prep and immunoblot analysis 28
Estimating Chlorophyll Concentration 28
2D-PAGE 29
Chlorophyll-fluorescence measurement 29
Semiquantitative RT-PCR 31
Quantification of variegation level 32
Histology and light microscopy 33
Results 34-74
Part A. Characterization of singel FtsH mutant 34-42
Null mutant of ftsh1 by T-DNA insertion line 34
Genotype analyses of FtsH1 null mutant 35
The null mutant ftsh1 is similar to WT 36
The chloroplast FtsH is few isomers 38
FtsH is family with 12 genes member 38
Characterization of other FtsH mutant 41
The mutant fail to accumulate mRNA 42
Part B. Developmental and environmental effects on the accumulation of FtsH 43-54
The ftsh2 variegation 43
Changes in FtsH mRNA during development 45
Changes in FtsH protein during development 46

104
Effect of light intensity on the degree of variegation and FtsH level 50
FtsH protein is sensitive to high light intensity 53
Part C. The chloroplast FtsH subunit relation 54-74
ftsh1 ftsh2 double mutant demonstrate severe variegation phenotype 55
ftsh 1 ftsh8 double mutant is similar to WT 58
ftsh1 ftsh2 double mutant is able to complete is life cycle 60
ftsh1 ftsh5 double mutant cannot survive under normal conditions 62
ftsh2 ftsh8 double mutant has no chloroplast 65
h2+ h8+ has high rate of embryo abortion 68
ftsh2 ftsh8 double mutant is sterile 69
The double mutant has abnormal flower organs 70
Comparison between different genotypes 73
Null mutant of FtsH gene duplication prevent the accumulation of other FtsH’s 73
DISCUSSION 75-86
FtsH involve in chloroplast biogenesis – the threshold and quantiting models 75
The influences of endogenic factors on the FtsH quantity and activity 76
FtsH is sensitive to high light intensity 78
FtsH1 involve in thylakoid formation and protection from high light 79
FtsH is hetrocomplex required tow sub unit for it accumulation and function 80
Structure and function the model 81
Summation 86

105
Characterization of the chloroplast FtsH protease in
Arabidopsis thaliana plants
Thesis submitted for the degree of “Doctor of Philosophy”
By
Adi Zaltsman
Submitted to the Senate of the Hebrew University of Jerusalem
September, 2005

106
עבודה זו נעשתה בהדרכתו של פרופסור צח אדם
,סמית ' רוברט הל שםמכון למדעי הצמח וגנטיקה בחקלאות עב
ה של המזון ואיכות הסביב,פקולטה למדעי החקלאותה
.ליםירוש, האוניברסיטה העברית

107
תודות
שעות רבות של , עבודה זו מסכמת מספר שנים של מחקר אינטנסיבי. פרק חשוב בחיי הגיע אל סיומו
לאורך המחקר ידענו . דיונים ארוכים וניסויים מורכבים, עבודה על שולחן המעבדה או מול צג המחשב
.מורדות ועליות וענינו על שאלות מדעיות שונות
: התאפשר הודות לאנשים רבים שליווי אותי לאורך הדרך כל זאת
על. עידו ורונית, יפעת, יוני, גלית, אורית, מזל, מרי, ארי, עינת: תודה לחברי וחברותי למעבדה
ולזו שניצחה ; אורן ותמי, מגי: לאלו שליוו אותי בתחילת דרכי וכבר עזבו ; עשהשכלתם לסייע ולהסתיי
. לאה-על כולם בעזרה ובידע
, לנעמי, לדודו, לאלון: תודה לחוקרי המכון שדלתות משרדם ומעבדתם היו תמיד פתוחות בפני
. וכן לצוות המנהלי של המכון והפקולטה. לחיים לאמנון ולאלי,לנאווה
Richardולפרופסור . תודה לפרופסור יצחק אוהד על הסיוע בלימוד המערכת הפוטוסינתטית
Vierstraתו למשך חודש ואפשר לי לחפש ולמצוא את המוטנט ב על שקיבל אותי למעבד-FtsH1. וכן
. יעל-לעורכת העבודה
, שלי שותפתי לדרך: למי ששילמו ועדיין משלמים את המחיר של דרך העבודה והלמידה שלי, וכמובן
ולאלה שאפשרו את הקיום הכלכלי שלנו בבחירתם הנדיבה להעניק לי . ובן וגיל שנולדו במהלך הדרך
".קרן הורוביץ "- מלגה את ה
ביכולת , בפרשנות, בסבלנות, בידע- לא יכולתי לבחור מנחה טוב ממנו -תודה מיוחדת שמורה לצח
תמיד הדרך הנכונה הגם אם זו לא היית, לנהל דיונים מדעיים ובנכונות לאפשר לי לבחור את הדרך
.בעיניו
.בלעדיכם כל זה לא היה מתרחש

108
יניםיתוכן ענ
2-5 תקציר
6-20מבוא
6 פרוטאוליזה
7 מערכת היוביקוויטין –פרוטאוליזה באאוקריוטים
8 מערכת מבוקרת -הפרוטאוזום
AAA 10-משפחת ה
11מערכות פרוטאוליטיות בפרוקריוטים
12 בחיידקים FtsHפרוטאזת
FtsH 13 במיטוכונדריות
FtsH15 בציאנובקטריה
15 מבנה ופעילות –ופלסט הצמחי הכלור
16פירוק חלבונים בכלורופלסט
FtsH 18 בצמחים
21מטרות המחקר
22-33 חומרים ושיטות
22חומר צמחי
22גידול בתנאים אספטיים וסלקציה , חיטוי זרעים
FtsH1 23-סריקה וזיהוי המחדר ב
25ייצור מוטנטים כפולים
T-DNA insertion line 25 מצמחי PCR גנומי לאנליזת DNAהפקת
26 ל אגרוז ' וניקויו בגDNAהפרדת
TAIL-PCR 26-שיבוט תוצרי ה

109
E. coli 27ייצור חיידקים קומפוטנטים וטרנספורמציה לחיידקי
Western blot 28הפקת חלבונים ואנליזת
28קביעת ריכוז כלורופיל
2D-PAGE 29-הפרדת חלבונים לפי מטען וגודל ב
30 מדידות פלורסנציה בצמחים
RT-PCR 31 בשיטת mRNAאנליזת
32ניתוח כמותי של שיעור הווריגציה
33 מיקרוסקופית אור והיסטולוגיה
34-74 תוצאות
34-42 זיהוי ואפיון מוטנטים בגנים יחידים . א
T-DNA 34 באמצעות מחדר של ftsh1השתקת הגן
35זיהוי נקודת המחדר והגנוטיפ
36 איננו שונה באופן מובהק מצמחי הבר ftsh1המוטנט
FtsH 38 הוא חלבון כלורופלסטי בעל מספר איזוזימים
FtsH38עשר גנים - היא משפחה בת שנים
41איתור ואפיון מוטנטים נוספים
mRNA 42המוטנטים אינם צוברים
FtsH2 43-55השפעות התפתחותיות וסביבתיות על ביוטוי . ב
ftsh2 43-אפיון הווריגציה ב
45 במהלך ההתפתחות FtsH של mRNA-השינוים ברמות ה
46שינויים ברמות החלבון במהלך ההתפתחות
FtsH 50-השפעות האור על רמת הווריגציה וכמות ה
FtsH 53רגיש לשטפי קרינה גבוהים

110
55-75 הכלורופלסטית FtsHיחסים בין תתי היחידות השונות במשפחת . ג
ftsh2 55 חמור יותר משל ftsh2 ftsh1הפנוטיפ של המוטנט הכפול
58 באופן נורמלימתפתח ומתפקד ftsh1 ftsh8המוטנט
60 מסוגל להשלים מחזור חיים ftsh5 ftsh2המוטנט
62 נאים נורמליים איננו מתקיים בת ftsh1 ftsh5המוטנט
65 איננו מפתח כלורופלסטים ftsh2 ftsh8המוטנט
h2+ h8+ 68שיעור הפלת עוברים גבוה בהכלאה עצמית של
69 עקר , ftsh2 ftsh8, המוטנט ההומוזיגוטי הכפול
70המוטנט הכפול פגום ביכולתו לייצר אברי רבייה תקינים
73בחינה השוואתית של הגנוטיפים השונים
73 קרובים מונעת את הצטברות הזוג השני FtsHלבוני השתקה של זוג ח
75-86דיון
FtsH75' המודל הכמותי'ו' סףמודל ה': מעורב בתהליכי התפתחות של הכלורופלסט
76ופעילותו FtsH גורמים אנדוגנים המשפיעים על כמות
79 רגישים לשטפי אור חזקים FtsHחלבוני
79מנזקי קרינה תפקיד בהתפתחות תילקואידים והגנה FtsH1-ל
FtsH81 שתי תת יחידות שונות נחוצות לצבירתו : הוא הטרוקומפלקס
82 מודל כמותי –מבנה ופעילות
85 סיכום
87-94רשימת ספרות
97-101 תקציר אנגלית
107תודות

111
108-110 תוכן עניינים
112 רשימת פירסומים
113 קיצורים
114טרמינולוגיה
115 רשימת תמונות

112
:רסומיםפ
:ממצאי עבודה זו פורסמו במאמרים הבאים
Sakamoto, W., Zaltsman, A., Adam, Z., and Takahashi, Y. (2003). Coordinated
regulation and complex formation of yellow variegated1 and yellow variegated2, chloroplastic FtsH metalloproteases involved in the repair cycle of photosystem II
in Arabidopsis thylakoid membranes. Plant Cell 15, 2843-2855.
Zaltsman, A., Feder, A., and Adam, Z. (2005). Developmental and light effects on the accumulation of FtsH protease in Arabidopsis chloroplasts - implications for
thylakoid formation and photosystem II maintenance. Plant J 42, 609-617.
Zaltsman A., Ori N. and Adam Z. (2005). Two Types of FtsH Protease Subunits Are Required For Chloroplast Biogenesis and Photosystem II Repair in Arabidopsis.
Plant Cell (in press)
:ובמאמר הסקירה
Adam, Z., Zaltsman, a., Sinvany-Villalobo, G., and Sakamoto, W. (2005). FtsH proteases in chloroplasts and cyanobacteria. Physiol Plant 123, 386-391.

113
Shortening קיצורים
2D - PAGE - two dimensional polyacrylamide gel electrophoresis
35S - Cauliflower mosaic virus promoter
AA - Acryl Amid
ctivitiesAssociated with various cellular ATPases A -AAA
AP - Alkaline phosphatase
ATP - Adenosine-5'-triphosphate
bp - Base pairs
cDNA - Complimentary DNA
DNA - Dioxy-ribonucleic acid
F0 - non-variable chlorophyll fluorescence
Fm - maximal chlorophyll fluorescence
FtsH- Filamentation temperature-sensitive H
Fv - variable chlorophyll fluorescence
Fv/Fm - Maximum quantum efficiency of PSII primary photochemistry
HY - Hypersensitive
kDa- Kilo Dalton
mRNA - messenger RNA.
MS- Murashige and Skoog.
MW - Molecular-Weight

114
odulatedMmplitude Aulse P -PAM
PCD - Programmed cell death
PCR - Polymerase chain reaction
PCI – Phenol Chloroform Isoamyl Alcohol
PSI -Photosystem
RNA - Ribo nucleoic acid
RPM – Run per minute
RT - Reverse transcriptase
SDSPAGE - sodium dodecyl sulfate polyacrylamide gel electrophoresis
SP- Signal peptide
SRH - Second region of homology
TP - Transit peptide
רמינולוגיה ט
FtsH- FtsH protein
ftsh - FtsH gene
ftsh - FtsH null mutant
ftsh1xftsh2 - cross of ftsh1 null with ftsh2 null
ftsh1 ftsh2 – double mutant of ftsh1 and ftsh2 homozygous to both
h2h2 h8+ - double mutant that has 2 null ftsh2 alleles and 1 null ftsh8 allele

115
h2h2 h1h1- allele arrangement of the mutant ftsh2 ftsh1
רשימת תמונות
עמודמספר התמונה ושמה
7מסלולי בקרת איכות של חלבונים . 1
26S 9מבנה ואירגון של . 2
pSKI015 24מפה סכמטית של הפלסמיד . 3
FtsH1 34-סריקת ספריה לאיתור מוטציית השתקה בגן ל. 4
ftsh1 35-זיהוי מוטנט ב. 5
37 פלורסנציה של כלורופיל . 6
2D-Western blot 38אנליזת . 7
FtsH 40ניתוח פילוגנטי של משפחת . 8
9 .Rt-PCR 42 למוטנטים השונים
ftsh2 43וריגציה בצמחי . 10
FtsH2 44-הפנוטיפ של העלים השונים בצמח המושתק בגן ל. 11
ftsh2 45 ובמוטנט WT- מצטבר עם התפתחות הצמח ב FtsH שלmRNA-ה. 12
ftsh2 47- וWT צבירת חלבונים בצמחי. 13
49 וגם בין עלים שונים באותו צמח, ftsh2- וWT המצטברת בעלים שונה בין צמחי FtsHכמות חלבוני . 14
ftsh2 50 ושל WTשל בפסיגים ובעלים ראשונים FtsHצבירת תוצרי פירוק אפשריים של . 15
52השפעות התאורה על הווריגציה . 16
PSII 54 ועל הפונציאל של FtsHהשפעת שטף אור חזק על רמת חלבוני . 17
55ניתוח הפרופיל הגנטי : ftsh2- וftsh1הכלאה בין מוטנטים הומוזיגוטים של . 18

116
ftsh2 ftsh1 56 במוטנט הכפול ftsh2החמרת הווריגציה של . 19
57רגיש לעקות אור ftsh2ftsh1המוטנט הכפול . 20
59 מתפתח ומתפקד באופן נורמלי ftsh1 ftsh8המוטנט הכפול . 21
61 לעקות אור מתפתח לאט ורגיש ftsh2 ftsh5המוטנט הכפול .22
h1+ h5+ 63התפצלות הפנוטיפים בהכלאה עצמית של . 23
h1+ h5+ 64סגרגצית הפנוטיפים של צאצאי ההכלאה העצמית של . 24
h2+ h8+ 66התפצלות הצאצאים של הכלאה עצמית של . 25
67 מייצרים מעט כלורופלסטים FtsH8- אך בעלי אלל יחיד לFtsH2צמחים חסרי . 26
h2+ h8+ 68אוכלוסית הצאצאים מהכלאה עצמית של התפצלות .27
h2+ h8+ 69ושל צמחי WT ניתוח תרמילים של צמחי . 28
71 איננו פורה ftsh2 ftsh8המוטנט הכפול . 29
72מיקרוסקופית אור של פרחים . 30
74ם הלבנים לא קיים במוטנטיFtsH: השוואה בין הגנוטיפים השונים. 31
FtsH 84מודל מבנה ההטרוקומפלקס של . 32

117
Related Documents










