ב ביוכימיה: הקורס שם פרנקל שי: המסמך יוצר2021 205-1-9181 : קורס מספר ודבי רן: מרצה תרגול סיכום: החומר סוג

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

שם הקורס: ביוכימיה ב
יוצר המסמך: שי פרנקל
2021
מספר קורס: 205-1-9181מרצה: רן ודבי
סוג החומר: סיכום תרגול
pr_magarac
Text Box
באדיבות מדור אקדמיה, אגודת הסטודנטים, אוניברסיטת בן גוריון.
pr_magarac
Text Box
www.bgu4u.co.il
maagar
Typewritten Text
maagar
New Stamp
1
ביוכימיה ב' מרצים: רז ודבי סיכום: שי פרנקל
למען הסביבה של כולנו , מומלץ שלא להדפיס את הסיכום
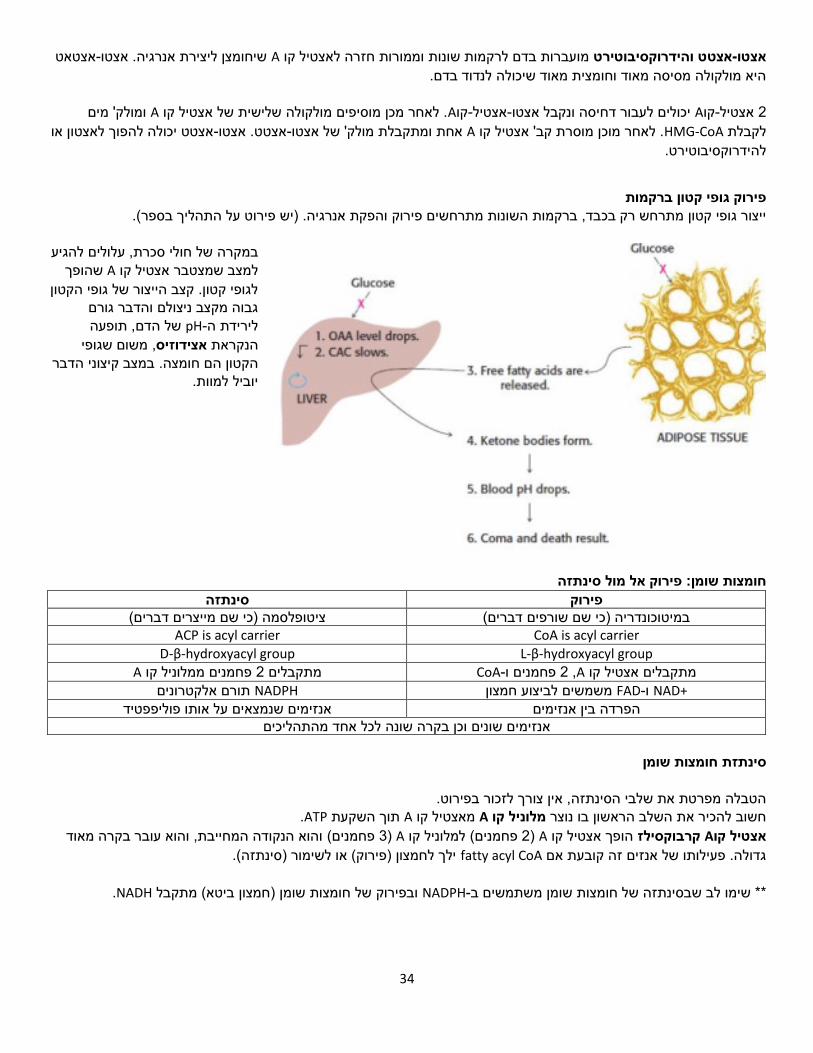
1שיעור( 7/3/18)
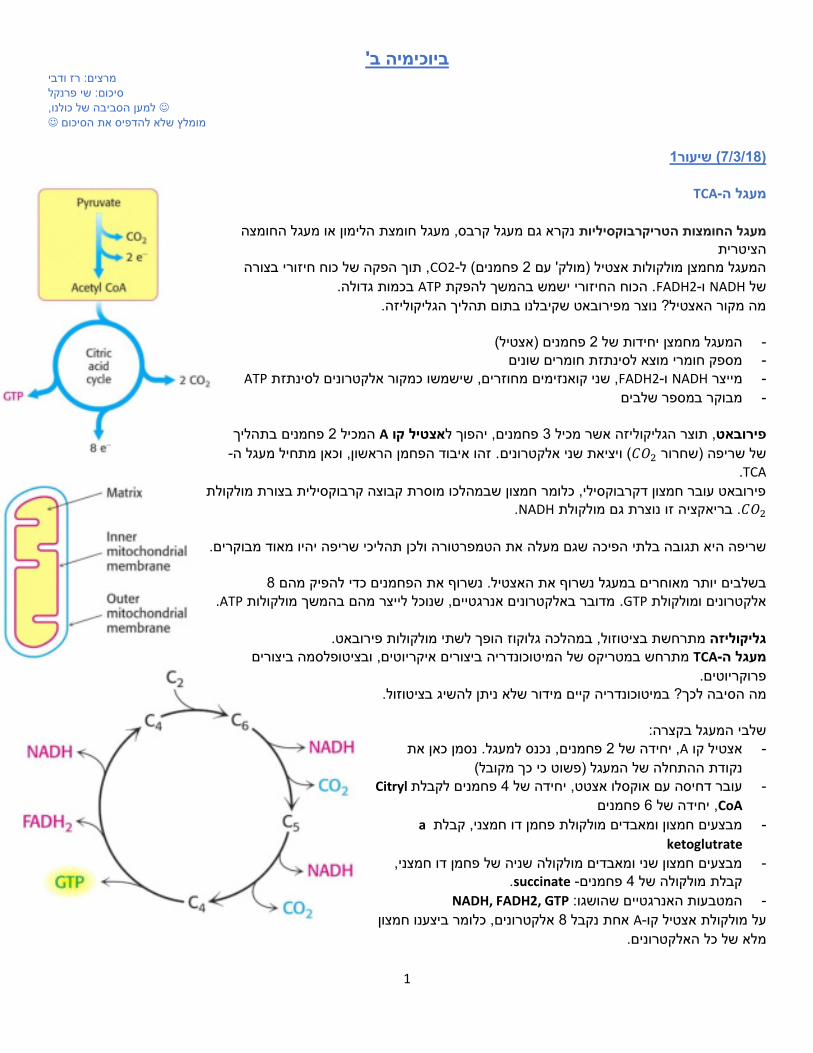
TCA-מעגל ה
מעגל חומצת הלימון או מעגל החומצה נקרא גם מעגל קרבס, מעגל החומצות הטריקרבוקסיליות הציטרית
, תוך הפקה של כוח חיזורי בצורה CO2-פחמנים( ל 2המעגל מחמצן מולקולות אצטיל )מולק' עם בכמות גדולה. ATP. הכוח החיזורי ישמש בהמשך להפקת FADH2-ו NADHשל
מה מקור האצטיל? נוצר מפירובאט שקיבלנו בתום תהליך הגליקוליזה. )אצטיל( פחמנים 2המעגל מחמצן יחידות של -
מספק חומרי מוצא לסינתזת חומרים שונים -
ATPכמקור אלקטרונים לסינתזת ישמשו ש, שני קואנזימים מחוזרים, FADH2-ו NADHמייצר -
מבוקר במספר שלבים -
בתהליך פחמנים 2המכיל Aאצטיל קו פחמנים, יהפוך ל 3מכיל אשר , תוצר הגליקוליזה פירובאט
-מעגל ה. זהו איבוד הפחמן הראשון, וכאן מתחיל יציאת שני אלקטרונים( ו𝐶𝑂2)שחרור של שריפהTCA.
.NADHבריאקציה זו נוצרת גם מולקולת .𝐶𝑂2פירובאט עובר חמצון דקרבוקסילי, כלומר חמצון שבמהלכו מוסרת קבוצה קרבוקסילית בצורת מולקולת
תהליכי שריפה יהיו מאוד מבוקרים.ולכן שגם מעלה את הטמפרטורה שריפה היא תגובה בלתי הפיכה
8נשרוף את האצטיל. נשרוף את הפחמנים כדי להפיק מהם בשלבים יותר מאוחרים במעגל .ATPמדובר באלקטרונים אנרגטיים, שנוכל לייצר מהם בהמשך מולקולות .GTPאלקטרונים ומולקולת
, במהלכה גלוקוז הופך לשתי מולקולות פירובאט.מתרחשת בציטוזולגליקוליזה
ביצורים איקריוטים, ובציטופלסמה ביצורים מתרחש במטריקס של המיטוכונדריה TCA-מעגל ה .פרוקריוטים
מידור שלא ניתן להשיג בציטוזול.קיים במיטוכונדריה מה הסיבה לכך?
:בקצרה שלבי המעגל. נסמן כאן את פחמנים, נכנס למעגל 2, יחידה של Aאצטיל קו -
נקודת ההתחלה של המעגל )פשוט כי כך מקובל(
Citrylלקבלת פחמנים 4יחידה של עובר דחיסה עם אוקסלו אצטט, -
CoA, פחמנים 6יחידה של
a, קבלת מבצעים חמצון ומאבדים מולקולת פחמן דו חמצני -
ketoglutrate
, מבצעים חמצון שני ומאבדים מולקולה שניה של פחמן דו חמצני - .succinate -פחמנים 4קבלת מולקולה של
NADH, FADH2, GTPהמטבעות האנרגטיים שהושגו: -
אלקטרונים, כלומר ביצענו חמצון 8אחת נקבל A-מולקולת אצטיל קו על מלא של כל האלקטרונים.
2
גורלם של תוצרי המעגל: בממברנת המיטוכונדריה. oxidative phosphorylationפקטורים, והם מועברים באמצעות -קו 4-אלקטרונים נמצאים ב 8 -
.ATPלגרדיאנט יש כוח כימי והכוח הכימי הזה מצומד למערכת חלבונים שמייצרת מושג גרדיאנט של פרוטונים. -
Acetyl coenzyme A (Acetyl coA)
מאוד ריאקטיבי, שנוטה לאבד את אטום המימן S-Hשל אצטיל. יש לו קשר יחידה פרוסטטית. מסיסה. משמש כנשא. Aאצטיל קו 'אוספות' NADשישמש בתהליך הנשימה. כך מולקולות NADHליצירת NADאטום המימן שהשתחרר מגיב עם ולהתרכב עם אצטיל.
את הפרוטונים המשתחררים.A: 𝑃𝑦𝑟𝑢𝑣𝑎𝑡𝑒ריאקציה ליצירת אצטיל קו + 𝐶𝑜𝐴 + 𝑁𝐴𝐷+ → 𝐴𝑐𝑒𝑡𝑦𝑙𝐶𝑜𝐴 + 𝐶𝑂2 + 𝑁𝐴𝐷𝐻
(PDC) פירובט דהידרוגנזקומפלקס
סימן שהולך הוצאת מימנים.הידרו = -דה
הליך של חמצון, לקיחה של להתרחש ת מימנים.
הריאקציה המזורזת על ידי אנזים זה היא דהקרבוקסילציה של פירובט.
קומפלקס מאוד דינאמי, יש דומיינים חלבוניים שלמים שנעים בין אזורים בתוך
הקומפלקס.Pyruvate dehydrogenase complex
דות:בנוי ממספר תתי יחיפירובט - E1 - 𝛼3יחידה -
מה שמבצע את - דהידרוגנאז. קרבוקסילציה-הפירובט דה
, TPPפירופוספט )-תיאמין :קופקטור .(B1ויטמין נגזרת של
קומפוננטה שמונעת מהפירובט לברוח, לשוט בתוך התמיסה או לעבור - דיהידרוליפויל טרנסאצטילאז - E2 - 𝛼2𝛽2יחידה -קופקטור: הטרוטטרמר. .Aקואנזים -לקבלת אצטיל A (CoA)העברת קבוצת אצטיל לקופקטור קואנזים מבצעת . דיפוסיה
Lipoamide ,Lioic acid , משמש כזרוע גמישה, קשר שלS-S .קבוצת שיכול לעבור חמצון חיזור, פתיחה וסגירה של הזרוע הזוSH של ליפוליזין היא שמאפשרת את החיבור של האצטיל לקואנזיםA.
.FADקופקטור: מחמצן את הזרוע הליפואמידית. .מזרזת ריאקצית חמצון חיזור – דיהידרוליפויל דהידרוגנאז - E3 - 𝛼𝛽יחידה - קומפלקס גדול, הרבה יותר גדול מריבוזום.
המטרה העיקרית של הקומפלקס היא לייצר את צורון הביניים.
יוטים.התגובה היא לא הפיכה באיקריוטים. כן הפיכה בפרוקר
קרבוקסילציה של פירובאט מתרחשת במיטוכונדריה, אחרי כניסתו של הסובסטרט, פירובאט, מהציטוזול -בתאים אאוקריוטים, הדה Aיל קואנזים טציה בלתי הפכית זו גורמת לכך שאצחמצני. ראק-ופחמן דו Aאצתיל קואנזים -ומתקבלים התוצרים של הריאקציה
חמצני מדפז אל מחוץ למיטוכונדריה ומשם אל מחוץ לתא.-ילכד במיטוכונדריה. הפחמן הדו

3
Aסינתזת אצטיל קו
הוא מולקולה שאינה פוליפפטיד )אינה חומצה אמינית שהיא חלק מהשרשרת החלבונית( ומהווה חלק מהריאקציה פקטור-קו האנזימטית ולפיכך חלק מהאנזים.
?קומפלקס פירובט דהידרוגנזשל ה יהאיך מתרחשת הריאקצ
ליצירת E1פחמנים( עובר חמצון קרבוקסילי ע"י 3) פירובאט -. זהו השלב האיטי ביותר והוא השלב קובע פחמנים( 2אצטיל ) הקצב.
.E1, קופקטור של TPPנקשר אל אצטיל -
עוברת ריאקציה עם E2של קבוצת ליפואמיד על זרוע גמישה -, קשר S-S-פתיחה של קשר המתרחשת , E1שעל האצטיל
עתיר אנרגיה
- S אחד נקשר אל הצטיל ו-S שני נקשר אלH
E2הזרוע הליפואמידית נעה אל -
נקשר אל הצטיל בקשר תיאו אסתרי )קשר עתיר Aקואנזים - Aאנרגיה( והם יוצאים מהקומפלקס כאצטיל קו
- S שני נקשר ל-H נוסף
א נעה אל והי SHקבוצות 2כעת הזרוע הליפואמידית מכילה -E3
- E3 מבצע חיזור של הזרוע הליפואמידית באמצעותFAD ליצירתFADH2
E2נוצר מחדש, הזרוע הליפואמידית חוזרת אל S-Sקשר -
- FADH2 שב-E3 מחזר אתNAD+ ליצירתNADH .FAD .הוא נשא אלקטרונים הפיךNAD לוקח אלקטרון אחד כפרוטון ואלקטרון .+𝐻אחד כמימן שלם )פרוטון+אלקטרון(, מימן נוסף נפלט כיון
הקומפלקס כעת במצבו ההתחלתי וניתן לחזור על הריאקציה -
הרבה משחקים של העברת אלקטרונים.
.LD domain-מתרחשת הכרה של קומפלקס חלבוני שלם, ולא רק של קצה מולקולה. הכרה מאוד ספציפית של הנודד, כל פעם הוא יכול להיכנס רק בכיס אחד. אין אותה הכרה כשהוא מחוזר לעומת כשהוא מחומצן לעומת כשהוא עם LDאותו
אצטיל, וככה נמנעת כניסה רנדומלית שלו.
.substrate channelingהסובסטרט לא עוזב את הקומפלקס בשום שלב, תגובה כזו נקראת
כתוצאה מכך נגרם נזק למוח, שמפיק . PDC-הנמצא ב TPP: גורמת לחוסר בתיאמין, שהוא קריטי ליצירת הקופקטור מחלת בריברי אנרגיה בחמצון אירובי של גלוקוז.
.גם אלכוהוליסטים יכולים לסבול ממצב דומה
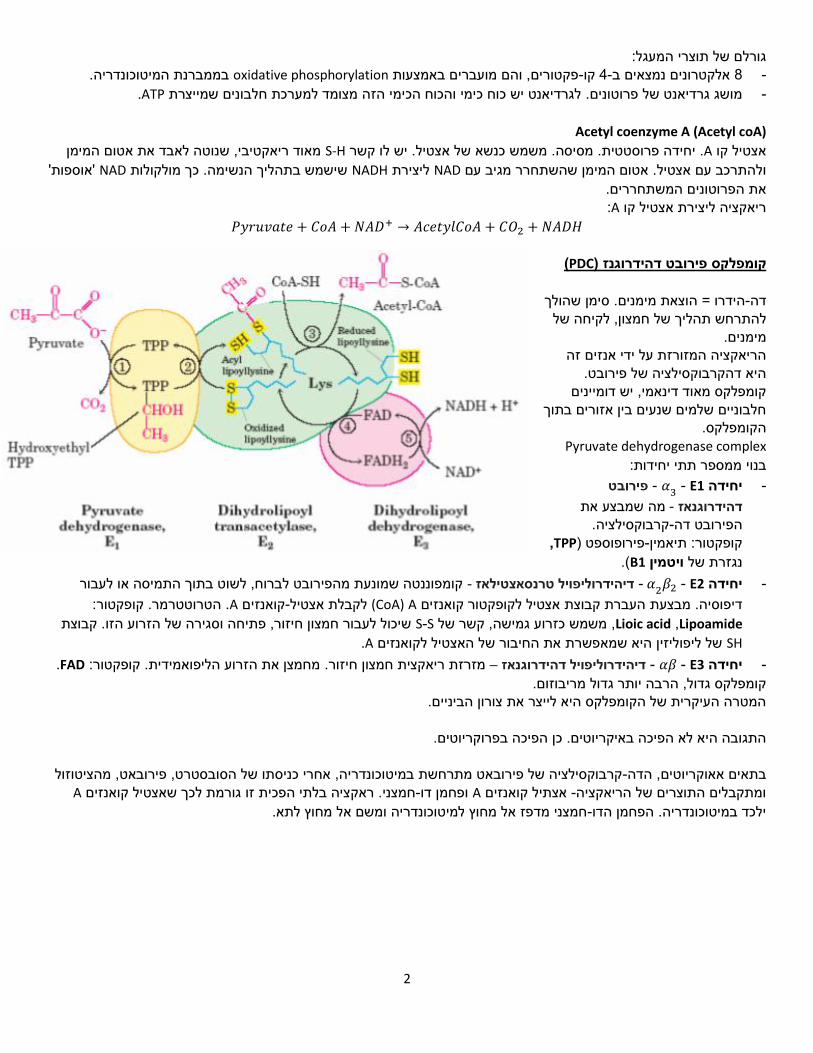
ריאקציות TCA :8-מעגל ה
citrate synthetaseהאנזים: ציטרט, + אוקסלואצטט Aאצטיל קו .1

4
עם המגיב פחמנים, 4מולקולה בעלת ,Oxaloacetateאוקסלואצטט לאוקסלואצטט. Aשלב ראשון ומחייב: דחיסה של אצטיל קו , קבוצת מתיל של האצטיל נקשרת אל קבוצת קרבוניל של בתגובה הראשונה של המעגלפחמנים( 2)בעלת אצטיל קבוצת ה
. האוקסלואצטט בתהליך שמועדף מבחינה אנרגטית , ולכן זהו מעגל.והאחרונה בתגובה השמינית תאוקסלואצטט נוצר' מולק
את כדי לא "לבזבז" סתם את התוצר.תתרחש הידרוליה של הציטרט, וזאוקסלואצטט גורם להתנעת התהליך, בהיעדרו לא Aצטיל קו וגורם לשינוי קונפורמציה שמאפשר את הקשירה של א citrate synthetaseאל האנזים אוקסלואצטט נקשר קודם
(.induced fit -)דוגמה נהדרת ל פחמנים. 6בעלת (citrate)התוצר הוא מולק' ציטרט
Aconitase, האנזים: איזוציטרט ציטרט .2
משום איזומריזציה נדרשת .cis-Aconitate תוצר הביניים דרךהתגובה מתרחשת לפחמן אחר. OH-העברת האיזומריזציה, פחמנים. 6מספר הפחמנים לא משתנה, עדיין שלישוני.הידרוכסיל יושב על פחמן שה
אקוניטט -אקוניטט, ולאחר מכן הידרציה שהופכת את ציס-בשלב הזה מתרחשת דהידרציה שהופכת את ציטרט לציס אקוניטט יכולה להפוך גם חזרה לציטרט.-לאיזוציטרט. ציס
Iso citrate dehydrogenaseהאנזים: ,+CO2 +NADH +𝑯+ גלוטרטאלפא קטו 𝑵𝑨𝑫+ + איזוציטרט .3
זהו (, מולקולה בת חמישה פחמנים. α-Ketoglutarateקטוגלוטרט )-חמצני והופך לאלפא-מאבד פחמן דופחמנים( 6)איזוציטרט -αקרבוקסילציה והעברת אלקטרונים. קצב ריאקציה זו קובע את קצב כל הריאקציה. ניתן להוציא את -שלב ראשון של דה
Ketoglutarate .מהמעגל בשלב זה ליצירת חומצות אמינו
α-Ketoglutarate dehydrogenase, האנזים: A +CO2 +NADHסוקסיניל קו 𝑵𝑨𝑫+ +CoA אלפא קטוגלוטרט + .4
,Aקואנזים ויוצר סוקציניל A חמצני, מתחבר לקואנזים-קטוגלוטרט מאבד פחמן דו-אלפאקרבוקסילציה: -שלב שני של דהSuccinyl CoA, פחמנים. 4בעל
succinyl coA synthase, האנזים: GTP +CoAסוקסינט + A +GDP + Pi יניל קואנזיםססוק .5
. GTP-ל GDP, קשר עתיר אנרגיה מוחלף עם פוספט, האנרגיה מועברת לאנזים שמתעל אותה להפיכת S-CoAשל קשר פירוק .CoAפחמנים )לא היה איבוד פחמנים(, יציאה של 4, מולקולה בעלת (Succinate)ינטססוקיצירת
. ATPוהשני מגיב בריאקציה ליצירת GTP, אחד מגיב בריאקציה ליצירת succinyl coA synthaseיש שני איזואנזימים של Succinate dehydrogenase , האנזים:FADH2+ פומרט FAD + סוקצינט .6
פחמנים. 4בעל Fumarate -פחמנים הופך ל 4סוקסינט בעל
fumarase :האנזים ,(Malateמלט ) פומרט .7
.)כי הוא עושה הידרציה לפומרט( ט הידרטזמרהאנזים נקרא גם פו , הידרציה.השקעת מולקולת מים𝑵𝑨𝑫𝑯+ לואצטטאוקס 𝑵𝑨𝑫+ + מלט .8 + 𝑯+ ,האנזים: malate dehydrogenase
לאוקסלואצטט. L-Malateחמצון של .TCA-להתחלה של מחזור נוסף במעגל ה Aפחמנים(, שניתן לדחוס שוב עם אצטיל קו 4בנקודה זו קיבלנו אוקסלואצטט )
𝐺∆זהו שלב של חמצון עם > . איך התגובה מתרחשת בכל זאת? הריאקציה ממשיכה קדימה בזכות הניצול המהיר של 0 נס מיד למחזור נוסף של המעגל.אוקסלואצטט שנכ -התוצרים
הערות כלליות: נרצה לחמצן כמה שיותר פחמנים עם אלקטרונים חפשיים. -
קשר עתיר אנרגיה משמעו שהוא מכיל הוא קשר עתיר אנרגיה, זהו הקשר של הקואנזים עם האצטיל. Cעם Sהקשר של - אלקטרונים, נוכל להמיר את הקשר ולהשיג אלקטרונים.
ואוקסלואצטט יכולים לשמש פרקורסורים לחומצות האמינו אספרטט וגלוטמט בהתאמה. לכן, אם נרצה לייצר אלפא קטוגלוטרט - של המעגל. 3חומצות אמינו, נוכל להוציא את אלפא קטוגלוטרט בתום שלב
+ 𝐴𝑐𝑒𝑡𝑦 𝐶𝑜𝐴 סטוכיומטריה של המעגל: 3𝑁𝐴𝐷 + 𝐹𝐴𝐷 + 𝐺𝐷𝑃 + 𝑃𝑖 2𝐶𝑂2 + 3 𝑁𝐴𝐷𝐻 + 2𝐻 + + 𝐹𝐴𝐷𝐻2 + 𝐶𝑜𝐴 + 𝐺𝑇𝑃
.Aעל כל אצטיל קו ATPמולקולות 10, כלומר בסה"כ Aעל כל מולקולה של אצטיל קו ATP + 1 GTP 9בסה"כ יש
על כל מולקולת גלוקוז. ATPמולקולות 20ולכן ATPמולק' 10-מולק' פירובאט וכל מולק' פירובאט מובילה ל 2ממולק' גלוקוז נוצרות מולקולות 30-( ולכן בסה"כ על כל מולקולת גלוקוז נקבל כ2ATP+NADH)כלומר ATPמולקולות 4.5הרווחנו גם בגליקוליזה בנוסף,
ATP.
5
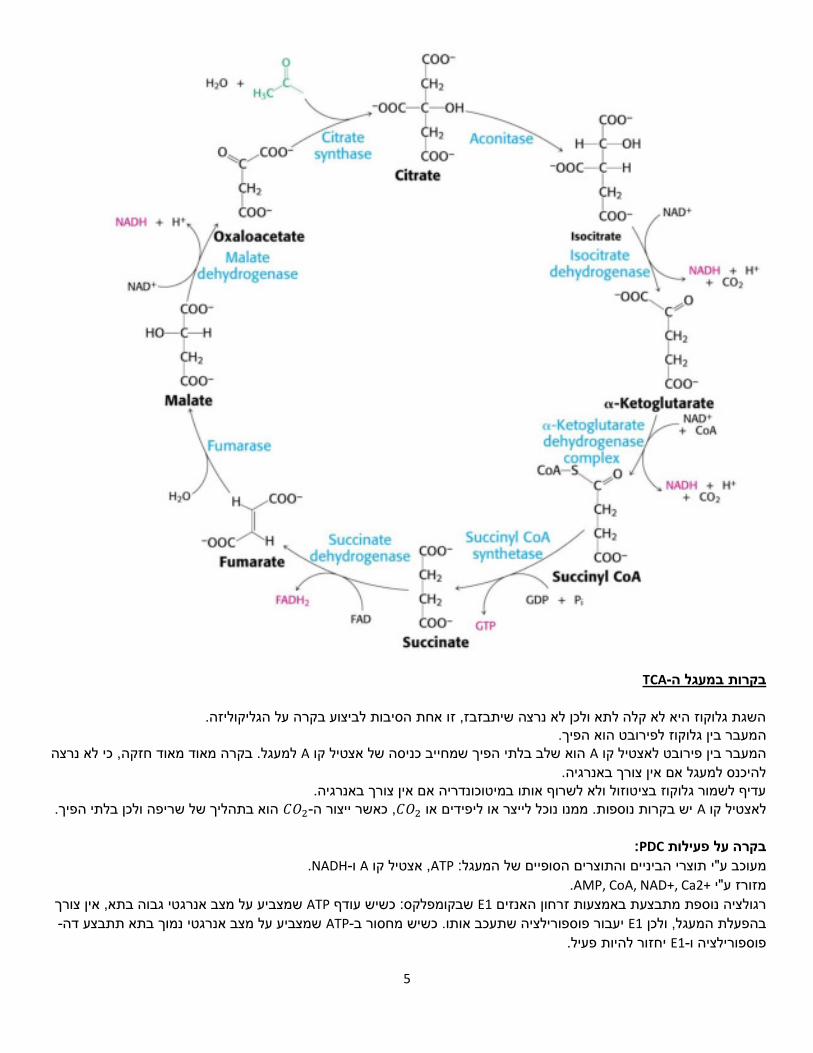
TCA-במעגל הבקרות
השגת גלוקוז היא לא קלה לתא ולכן לא נרצה שיתבזבז, זו אחת הסיבות לביצוע בקרה על הגליקוליזה. מעבר בין גלוקוז לפירובט הוא הפיך.ה
למעגל. בקרה מאוד מאוד חזקה, כי לא נרצה Aהוא שלב בלתי הפיך שמחייב כניסה של אצטיל קו Aהמעבר בין פירובט לאצטיל קו להיכנס למעגל אם אין צורך באנרגיה.
באנרגיה.עדיף לשמור גלוקוז בציטוזול ולא לשרוף אותו במיטוכונדריה אם אין צורך הוא בתהליך של שריפה ולכן בלתי הפיך. 𝐶𝑂2-, כאשר ייצור ה𝐶𝑂2יש בקרות נוספות. ממנו נוכל לייצר או ליפידים או Aלאצטיל קו
:PDCבקרה על פעילות
.NADH-ו A, אצטיל קו ATPתוצרי הביניים והתוצרים הסופיים של המעגל: מעוכב ע"י .+AMP, CoA, NAD+, Ca2מזורז ע"י
שמצביע על מצב אנרגטי גבוה בתא, אין צורך ATP: כשיש עודף שבקומפלקס E1רגולציה נוספת מתבצעת באמצעות זרחון האנזים -שמצביע על מצב אנרגטי נמוך בתא תתבצע דה ATP-יעבור פוספורילציה שתעכב אותו. כשיש מחסור ב E1בהפעלת המעגל, ולכן
יחזור להיות פעיל. E1-פוספורילציה ו
6
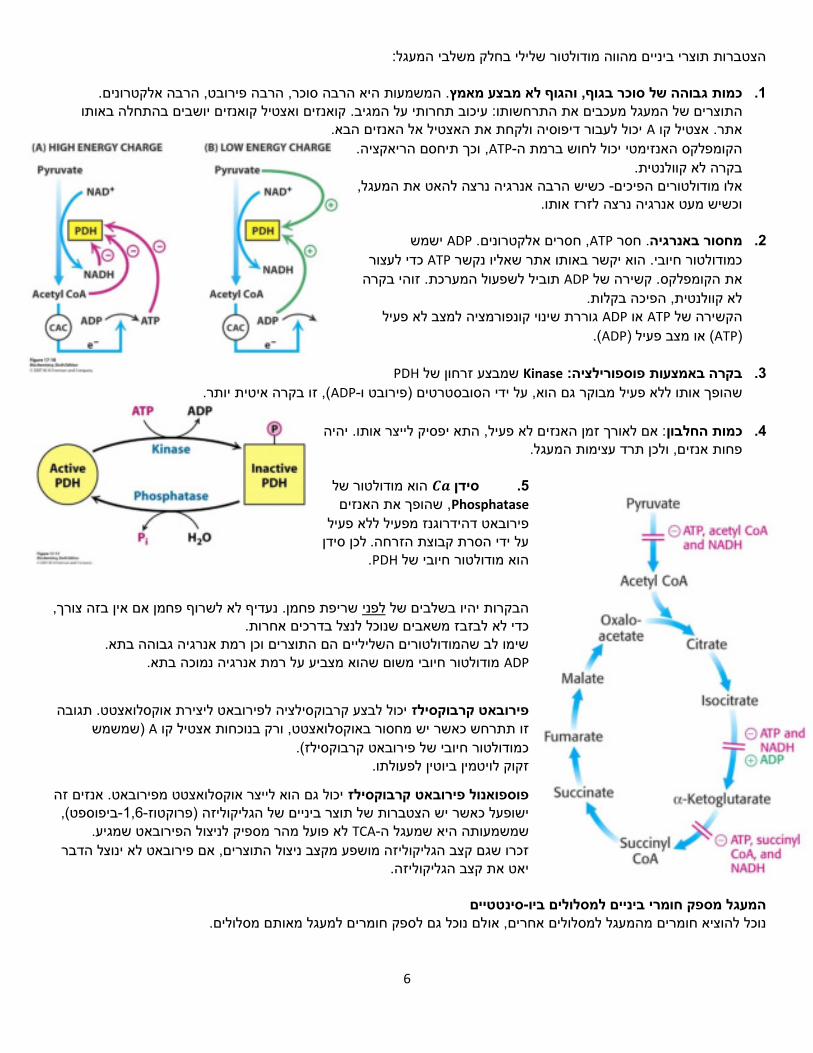
הצטברות תוצרי ביניים מהווה מודולטור שלילי בחלק משלבי המעגל: . המשמעות היא הרבה סוכר, הרבה פירובט, הרבה אלקטרונים. כמות גבוהה של סוכר בגוף, והגוף לא מבצע מאמץ .1
אותו קואנזים ואצטיל קואנזים יושבים בהתחלה בים של המעגל מעכבים את התרחשותו: עיכוב תחרותי על המגיב. התוצר יכול לעבור דיפוסיה ולקחת את האצטיל אל האנזים הבא. Aאתר. אצטיל קו
יחסם הריאקציה. , וכך תATP-הקומפלקס האנזימטי יכול לחוש ברמת ה .בקרה לא קוולנטית
להאט את המעגל, כשיש הרבה אנרגיה נרצה -אלו מודולטורים הפיכים .וכשיש מעט אנרגיה נרצה לזרז אותו
ישמש ADP, חסרים אלקטרונים. ATP. חסר באנרגיהמחסור .2
כדי לעצור ATPכמודולטור חיובי. הוא יקשר באותו אתר שאליו נקשר זוהי בקרה שפעול המערכת.תוביל ל ADPקשירה של את הקומפלקס.
לא קוולנטית, הפיכה בקלות.
גוררת שינוי קונפורמציה למצב לא פעיל ADPאו ATPהקשירה של (ATPאו מצ )( ב פעילADP.)
PDHזרחון של שמבצע Kinaseבקרה באמצעות פוספורילציה: .3
(, זו בקרה איטית יותר.ADP-מבוקר גם הוא, על ידי הסובסטרטים )פירובט ושהופך אותו ללא פעיל
: אם לאורך זמן האנזים לא פעיל, התא יפסיק לייצר אותו. יהיה כמות החלבון .4 המעגל.פחות אנזים, ולכן תרד עצימות
של הוא מודולטור 𝑪𝒂סידן .5Phosphatase שהופך את האנזים ,
מפעיל ללא פעיל פירובאט דהידרוגנז לכן סידן על ידי הסרת קבוצת הזרחה.
.PDHהוא מודולטור חיובי של
שריפת פחמן. נעדיף לא לשרוף פחמן אם אין בזה צורך, לפניהבקרות יהיו בשלבים של .שנוכל לנצל בדרכים אחרות כדי לא לבזבז משאבים
שימו לב שהמודולטורים השליליים הם התוצרים וכן רמת אנרגיה גבוהה בתא.ADP .מודולטור חיובי משום שהוא מצביע על רמת אנרגיה נמוכה בתא
יכול לבצע קרבוקסילציה לפירובאט ליצירת אוקסלואצטט. תגובה פירובאט קרבוקסילז)שמשמש Aלואצטט, ורק בנוכחות אצטיל קו זו תתרחש כאשר יש מחסור באוקס
כמודולטור חיובי של פירובאט קרבוקסילז(. זקוק לויטמין ביוטין לפעולתו.
יכול גם הוא לייצר אוקסלואצטט מפירובאט. אנזים זה פוספואנול פירובאט קרבוקסילז ,ביפוספט(-1,6-)פרוקטוז הגליקוליזהביניים של ישופעל כאשר יש הצטברות של תוצר
הפירובאט שמגיע. ללא פועל מהר מספיק לניצו TCA-שמשמעותה היא שמעגל הזכרו שגם קצב הגליקוליזה מושפע מקצב ניצול התוצרים, אם פירובאט לא ינוצל הדבר
יאט את קצב הגליקוליזה.
סינטטיים-המעגל מספק חומרי ביניים למסלולים ביו לם נוכל גם לספק חומרים למעגל מאותם מסלולים.נוכל להוציא חומרים מהמעגל למסלולים אחרים, או

7
2( שיעור14/3/18)
Electron transport and Oxidative Phosphorylation זרחון חמצוני
המשמעותי ביותר בתא. ATP-ייצור ה
הממברנות הבררניות , נחשבת לאחת ממברנות, חיצונית ופנימית. הממברנה הפנימית היא מאוד בררנית 2המיטוכונדריה מכילה
, ולא מאפשרת מעבר אפילו של יונים. הממברנה החיצונית פחות בררנית.ביותר בטבע , קפלים שנכנסים אל תוך הנפח, וכך מגדילים את שטח הפנים של הממברנה.Cristaeבתוך הממברנה הפנימית יש
וכן חמצון של חומצות שומן וחומצות אמינו TCA-הבתוך המטריקס של המיטוכונדריה ישנם על האנזימים הדרושים לביצוע מעגל
)על פירוק חומצות שומן נלמד בהמשך, פירוק חומצות אמינו לא יכלל בקורס השנה(.
:מודל מופשט של מה שנלמדנתבונן ב
והופכים 𝑂2, נשימה. נושמים את החמצן Respirationתהליך שנקרא .אותו למים
, והאלקטרונים TCA-יצרנו במעגל הנשתמש בתורמי אלקטרונים שונים שהאלו יגרמו למעבר פרוטונים אל הצד השני של הממברנה הפנימית תוך כדי פירוק של חמצן למולקולות מים )בהיעדר חמצן המעבר הזה נסתם
ולא ניתן לבצע את התהליך(. .ATPבהמשך נשתמש במולקולות שיצרנו כדי ליצר מהן
, ואלו נשאי האלקטרונים שישמשו FADH2-ו NADHיצרנו TCA-במעגל ה
.כעת ליצירת מפל ריכוזי הפרוטונים משני צידי הממברנה הפנימית של המיטוכונדריהאותנו
The electron transport machinery מולקולות שואפות להתפזר באופן הומוגני, חומר מאזורים מרוכזים מאוד ינוע עם מפל הריכוזים למקום בו ריכוזו נמוך.
ש באלקטרונים כדי לייצר פוטנציאל אלקטרוכימי.נשתמ
Electron donor תורם אלקטרון ל-Elector acceptor למולקולות שונות קל יותר או פחות למסור או לקבל אלקטרון. ריאקציות כאלו . מתרחשות בתגובת שרשרת.
.NAD+, NADP+, FAD, FMNנשאי אלקטרונים אוניברסליים המעורבים בתהליך: NADH שמפיקים אנרגיה( כמו לרוב משמש נשא אלקטרונים בתהליכים קטבולים ,TCA חמצון זרחוני-ו ,)NADPH לרוב משמש נשא
אלקטרונים בתהליכים אנאבולים )דורשים אנרגיה(.
Voltmeter פוטנציאל אלקטרוכימי של מולקולות.מדידת
ון שמעביר את האלקטרונים באופן הדרגתי. אם היו מדלגים על הריאקציות פולטות אנרגיה באופן הדרגתי, ולכן התא פיתח מנגנ
שלבי הביניים, האנרגיה הנפלטת היתה גורמת לפיצוץ התא.
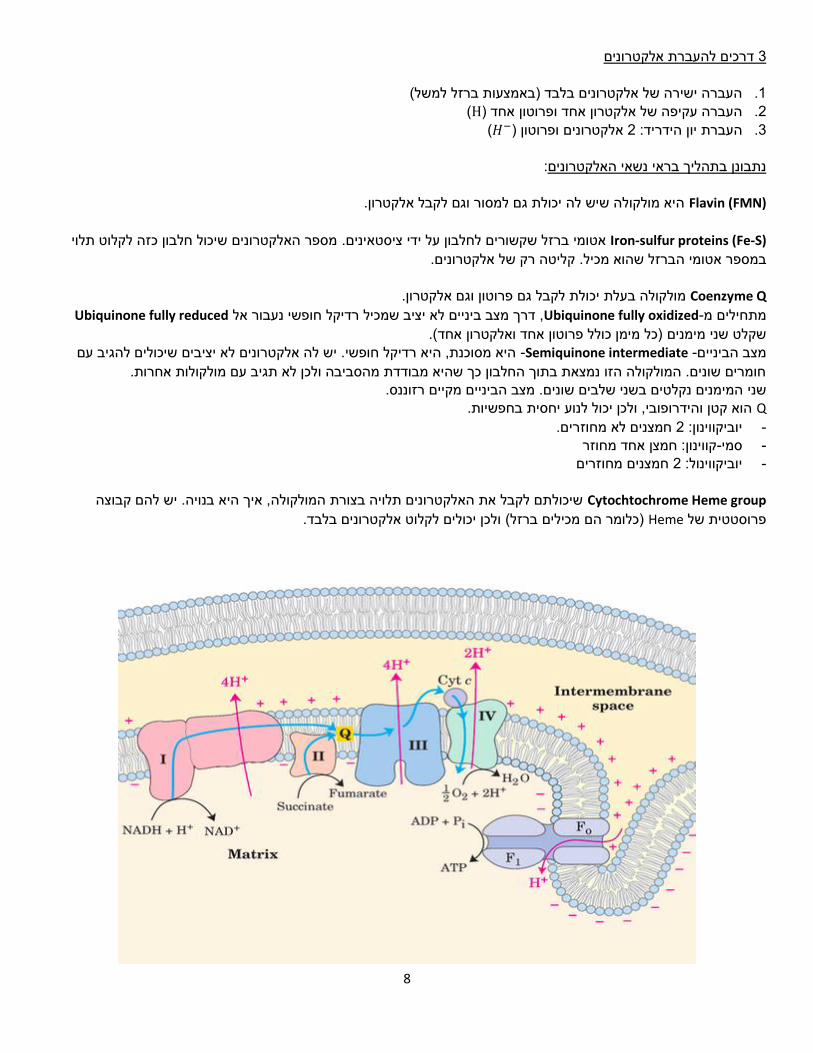
8
דרכים להעברת אלקטרונים 3 העברה ישירה של אלקטרונים בלבד )באמצעות ברזל למשל( .1
(H) העברה עקיפה של אלקטרון אחד ופרוטון אחד .2
(−𝐻ופרוטון ) אלקטרונים 2העברת יון הידריד: .3
:נתבונן בתהליך בראי נשאי האלקטרונים
Flavin (FMN) .היא מולקולה שיש לה יכולת גם למסור וגם לקבל אלקטרון
Iron-sulfur proteins (Fe-S) מספר האלקטרונים שיכול חלבון כזה לקלוט תלוי לחלבון על ידי ציסטאינים. יםברזל שקשוראטומי קליטה רק של אלקטרונים. ברזל שהוא מכיל.במספר אטומי ה
Coenzyme Q ולת לקבל גם פרוטון וגם אלקטרון.מולקולה בעלת יכ
Ubiquinone fully reducedר אל ו, דרך מצב ביניים לא יציב שמכיל רדיקל חופשי נעבUbiquinone fully oxidized-מתחילים מ .)כל מימן כולל פרוטון אחד ואלקטרון אחד( מימניםשקלט שני
היא מסוכנת, היא רדיקל חופשי. יש לה אלקטרונים לא יציבים שיכולים להגיב עם -Semiquinone intermediate -מצב הביניים חומרים שונים. המולקולה הזו נמצאת בתוך החלבון כך שהיא מבודדת מהסביבה ולכן לא תגיב עם מולקולות אחרות.
.המימנים נקלטים בשני שלבים שונים. מצב הביניים מקיים רזוננסשני Q .הוא קטן והידרופובי, ולכן יכול לנוע יחסית בחפשיות חמצנים לא מחוזרים. 2יוביקווינון: - קווינון: חמצן אחד מחוזר-סמי - חמצנים מחוזרים 2יוביקווינול: -
Cytochtochrome Heme group .קבוצה יש להם שיכולתם לקבל את האלקטרונים תלויה בצורת המולקולה, איך היא בנויה ולכן יכולים לקלוט אלקטרונים בלבד. )כלומר הם מכילים ברזל( Hemeפרוסטטית של
9
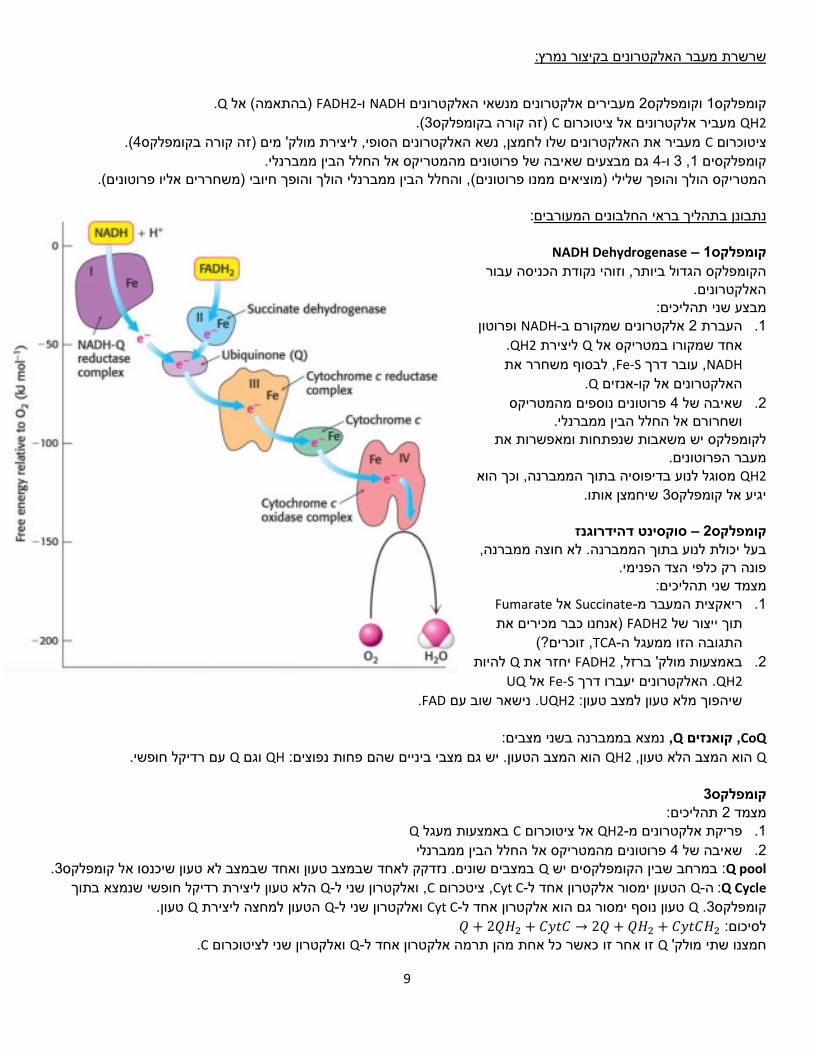
:בקיצור נמרץשרשרת מעבר האלקטרונים
.Q)בהתאמה( אל FADH2-ו NADHמעבירים אלקטרונים מנשאי האלקטרונים 2וקומפלקס 1קומפלקס
QH2 מעביר אלקטרונים אל ציטוכרוםC (.3)זה קורה בקומפלקס (.4מעביר את האלקטרונים שלו לחמצן, נשא האלקטרונים הסופי, ליצירת מולק' מים )זה קורה בקומפלקס Cציטוכרום
גם מבצעים שאיבה של פרוטונים מהמטריקס אל החלל הבין ממברנלי. 4-ו 3, 1קומפלקסים .)משחררים אליו פרוטונים( , והחלל הבין ממברנלי הולך והופך חיובי)מוציאים ממנו פרוטונים( המטריקס הולך והופך שלילי
:תהליך בראי החלבונים המעורביםנתבונן ב
NADH Dehydrogenase – 1קומפלקס
, וזוהי נקודת הכניסה עבור הקומפלקס הגדול ביותר האלקטרונים.
תהליכים:מבצע שני ופרוטון NADH-אלקטרונים שמקורם ב 2העברת .1
.QH2ליצירת Qאחד שמקורו במטריקס אל NADH עובר דרך ,Fe-S לבסוף משחרר את ,
.Qאנזים -האלקטרונים אל קו
פרוטונים נוספים מהמטריקס 4שאיבה של .2 ושחרורם אל החלל הבין ממברנלי.
חות ומאפשרות את לקומפלקס יש משאבות שנפת הפרוטונים. מעברQH2 מסוגל לנוע בדיפוסיה בתוך הממברנה, וכך הוא
שיחמצן אותו. 3יגיע אל קומפלקס
סוקסינט דהידרוגנז – 2קומפלקס. לא חוצה ממברנה, בעל יכולת לנוע בתוך הממברנה
.פונה רק כלפי הצד הפנימי מצמד שני תהליכים:
Fumarateאל Succinate-ריאקצית המעבר מ .1)אנחנו כבר מכירים את FADH2תוך ייצור של
, זוכרים?(TCA-התגובה הזו ממעגל ה
להיות Qיחזר את FADH2באמצעות מולק' ברזל, .2QH2 . דרך יעברו האלקטרוניםFe-S אלUQ
.FADנישאר שוב עם .UQH2שיהפוך מלא טעון למצב טעון:
CoQ קואנזים ,Q, :נמצא בממברנה בשני מצבים Q ,הוא המצב הלא טעוןQH2 :הוא המצב הטעון. יש גם מצבי ביניים שהם פחות נפוציםQH וגםQ .עם רדיקל חופשי
3קומפלקס תהליכים: 2מצמד
Qבאמצעות מעגל Cאל ציטוכרום QH2-פריקת אלקטרונים מ .1
פרוטונים מהמטריקס אל החלל הבין ממברנלי 4שאיבה של .2Q pool הקומפלקסים יש : במרחב שביןQ 3במצבים שונים. נזדקק לאחד שבמצב טעון ואחד שבמצב לא טעון שיכנסו אל קומפלקס .
Q Cycle: ה-Q להטעון ימסור אלקטרון אחד-Cyt C ציטכרום ,C, לואלקטרון שני-Q ליצירת רדיקל חופשי שנמצא בתוך הלא טעון טעון. Qהטעון למחצה ליצירת Q-ואלקטרון שני ל Cyt C-טעון נוסף ימסור גם הוא אלקטרון אחד ל Q. 3קומפלקס𝑄לסיכום: + 2𝑄𝐻2 + 𝐶𝑦𝑡𝐶 → 2𝑄 + 𝑄𝐻2 + 𝐶𝑦𝑡𝐶𝐻2
.Cואלקטרון שני לציטוכרום Q-כאשר כל אחת מהן תרמה אלקטרון אחד לזו אחר זו Qחמצנו שתי מולק'

10
לא נרצה שיגיב עם סביבתו באופן לא מבוקר.משום שמדובר ברדיקל חופשי ו 3הטעון למחצה לצאת מקומפלקס Q-לא ניתן ל
ציטוכרום אוקסידז – 4קומפלקס אלקטרונים. על כל אחד מהאלקטרונים האלו ישאב פרוטון מהמטריקס. 8תורמות Cשל ציטוכרום מולק' 4 (.O2מולק' מים ממולק' אחת של חמצן אטמוספרי ) 2אלקטרונים ישמשו ליצירת 4-פרוטונים ו 4 נוספים יפלטו אל החלל הבין ממברנלי.פרוטונים 4
היא מולקולה מאוד יציבה, נדרשת השקעה משמעותית של אנרגיה כדי לפרק אותה ולייצר מולקולת מים. 𝑂2 מולקולות פירובט. 2כלומר עבור פעמים, 4צריך להתרחש Cyt Cהמעבר של .Cyt C-אלקטרונים נכנסים מליצירת גשר פראוקסידי, ושניים נוספים יגרמו Cu-ו Heme-ומים: שניים ראשונים יעברו למבין האלקטרונים שנתרמו על ידי ציטוכר
פרוטונים שישברו את הגשר שנוצר. 2לשאיבה של
ע"י 2-ו 3ע"י קומפלקס 4, 1ע"י קומפלקס 4פרוטונים: 10לסיכום: על כל מולק' מים אחת שמסונתזת מתרחשת שאיבה של .4קומפלקס
לק' מים היא מאוד מועדפת אנרגטית, ולכן מעברי האלקטרונים האלו ליצירת מו O2אל NADH-אלקטרונים מההעברה של
מתרחשים באופן ספונטני וללא השקעת אנרגיה. מעברי האלקטרונים מצומדים על ידי הקומפלקסים לשאיבה של פרוטונים ימי.מהמטריקס אל החלל הבין ממברנלי וזאת כדי ליצור פוטנציאל אלקטרוכ
.ATPסינתזת ספויילר לשיעור הבא: הפוטנציאל הזה ינוצל ל
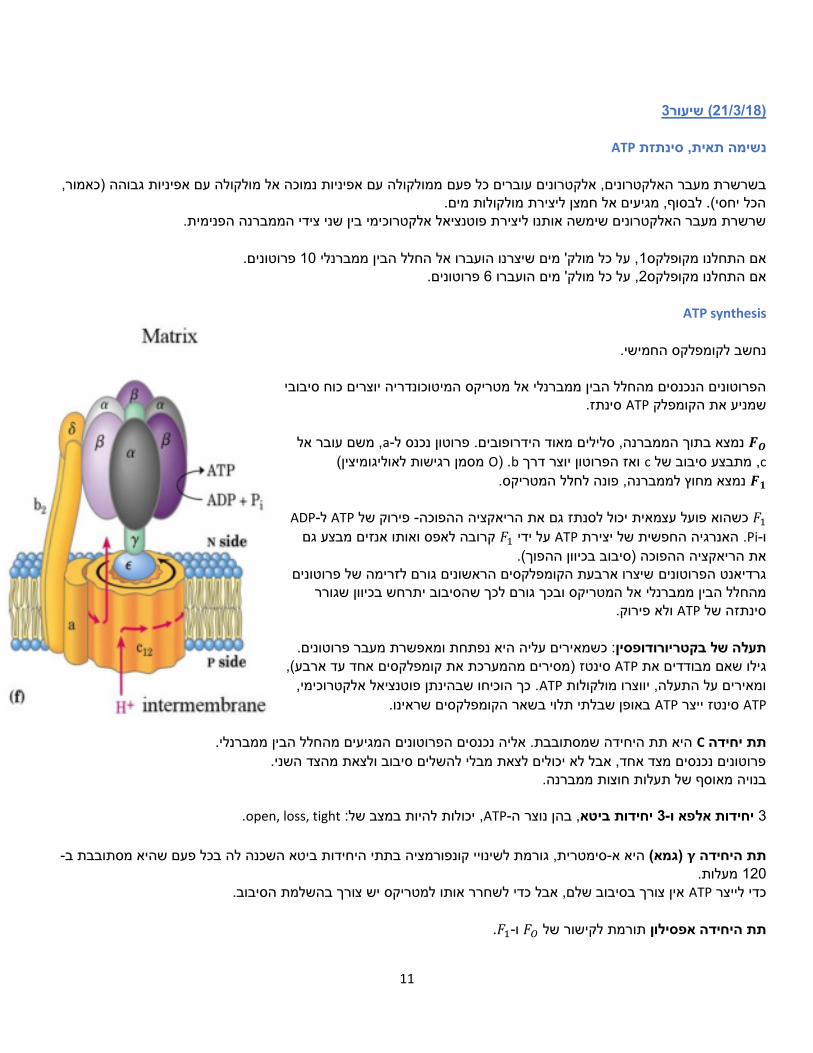
צימוד תהליכים
גורם ATP-, אך לא רק! גם עיכוב של סינתזת הATPלעיכוב של סינתזת נמצא כי עיכוב של שרשרת מעבר האלקטרונים גורם להאטה ואף הפסקה של שרשרת מעבר האלקטרונים.
רעלנים המשפיעים על התהליך:
)הקומפלקס החמישי(. נמצא כי מתן אנטיביוטיקות אלו גורם ATP synthetase-הן אנטיביוטיקות שפוגעות ב אוליגומיצין, נטוריצידין
גם לעיכוב של שרשרת מעבר האלקטרונים )ניתן לזהות זאת על פי הירידה בצריכת החמצן(.
Rotenoneאמיטל , יותר משמעותי משום 1עיכוב של קומפלקס .2המשך פעילותו של קומפלקס, אך מאפשרים את 1חומרים שמעכבים את קומפלקס
את הפקת האנרגיה. ים, רק מאטשממנו מפיקים יותר פרוטונים. החומרים הללו לא הורגים
CO, ציאניד 4ובכך חוסמים את מעבר האלקטרונים אל החמצן, נשא האלקטרונים הסופי. אם קומפלקס 4מעכבים את קומפלקסחומרים ש
מעוכב אחרי זמן קצר גם שלושת הקומפלקסים האחרים יעוכבו משום שתהליך מסירת האלקטרונים לא יושלם. התוצאה היא מוות.
Uncouplersמפרי צימוד , ATP Synthase-אם נעביר את האלקטרונים דרך הנוצר גרדיאנט אלקטרוכימי משני צידי הממברנה הפנימית של המיטוכונדריה.
, מה שבעצם מבטל את כל התהליך.ATP Synthase-מעבר פרוטונים שלא דרך המאפשרים Uncouplers-. הATPנייצר כל מפרי הצימוד הם מולקולות אמפיפטיות שיכולות לחצות את הממברנה.
- DNP ,FCCP יכולים לעבור בדיפוסיה דרך הממברנה ולקחת איתם פרוטונים, כלומר לאפשר העברה של פרוטונים במורד הגרדיאנט
הפרת הצימוד. סינטתז. ATPהאלקטרוכימי שנוצר שלא דרך ולינומיצין -
הפרת הצימוד. מאפשר מעבר יונים חפשי דרך הממברנה. .UCP1, נקרא גם (Thermogeninתרמוגנין ) -
(.ATP, מאפשר מעבר חפשי של פרוטונים חזרה אל המטריקס באופן שגורם ליצירת חום )ולא (brown)נפוץ בשומן חום .(heat)השם תרמוגנין מרמז על כך שיש קשר לייצור של חום
11
3( שיעור18/321/)
ATP, סינתזת נשימה תאית
ממולקולה עם אפיניות נמוכה אל מולקולה עם אפיניות גבוהה )כאמור, בשרשרת מעבר האלקטרונים, אלקטרונים עוברים כל פעם הכל יחסי(. לבסוף, מגיעים אל חמצן ליצירת מולקולות מים.
שרשרת מעבר האלקטרונים שימשה אותנו ליצירת פוטנציאל אלקטרוכימי בין שני צידי הממברנה הפנימית.
פרוטונים. 10, על כל מולק' מים שיצרנו הועברו אל החלל הבין ממברנלי 1מקופלקס התחלנואם פרוטונים. 6, על כל מולק' מים הועברו 2מקופלקסהתחלנו אם
ATP synthesis
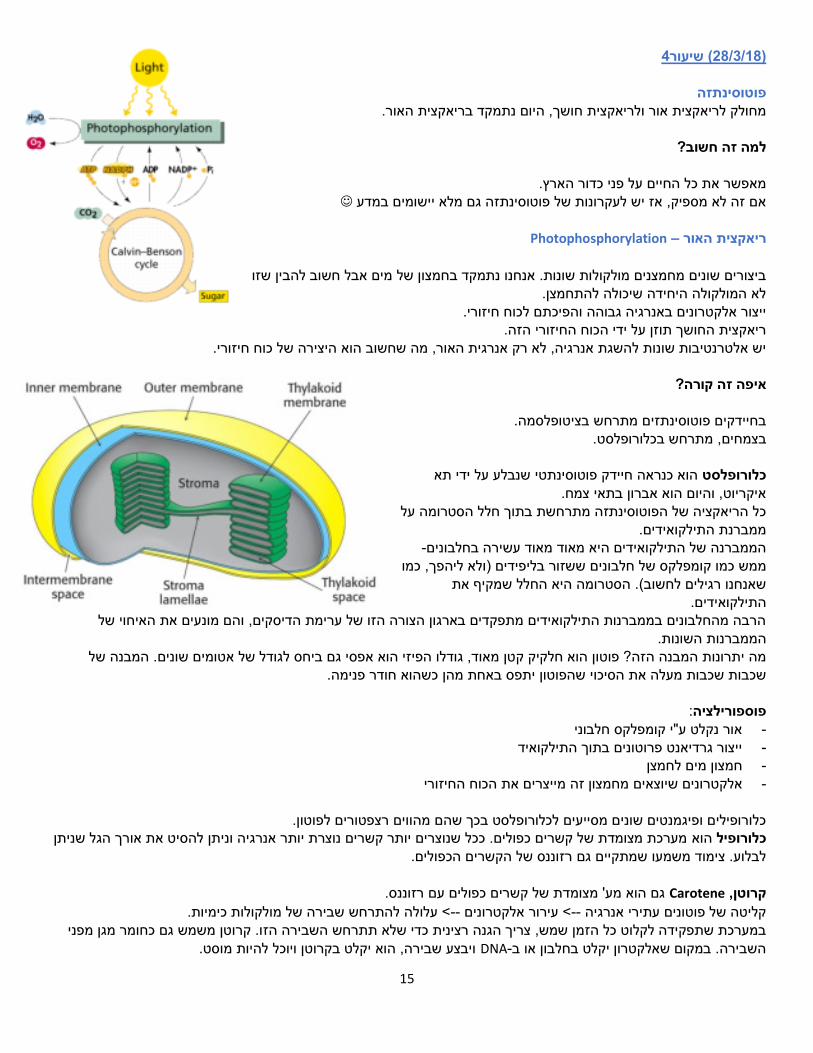
נחשב לקומפלקס החמישי.
הפרוטונים הנכנסים מהחלל הבין ממברנלי אל מטריקס המיטוכונדריה יוצרים כוח סיבובי
סינתז. ATPשמניע את הקומפלק 𝑭𝑶 נמצא בתוך הממברנה, סלילים מאוד הידרופובים. פרוטון נכנס ל-a משם עובר אל ,c מתבצע סיבוב של ,c ואז הפרוטון יוצר דרךb. (O )מסמן רגישות לאוליגומיצין 𝑭𝟏 .נמצא מחוץ לממברנה, פונה לחלל המטריקס 𝐹1 פירוק של -פועל עצמאית יכול לסנתז גם את הריאקציה ההפוכהכשהואATP ל-ADP קרובה לאפס ואותו אנזים מבצע גם 𝐹1על ידי ATPיצירת . האנרגיה החפשית של Pi-ו
.)סיבוב בכיוון ההפוך( את הריאקציה ההפוכהשיצרו ארבעת הקומפלקסים הראשונים גורם לזרימה של פרוטונים גרדיאנט הפרוטונים
גורם לכך שהסיבוב יתרחש בכיוון שגורר מהחלל הבין ממברנלי אל המטריקס ובכך ולא פירוק. ATPסינתזה של
: כשמאירים עליה היא נפתחת ומאפשרת מעבר פרוטונים. תעלה של בקטריורודופסין
מסירים מהמערכת את קומפלקסים אחד עד ארבע(, ז )טסינ ATPגילו שאם מבודדים את . כך הוכיחו שבהינתן פוטנציאל אלקטרוכימי, ATPומאירים על התעלה, יווצרו מולקולות
ATP ז ייצר טסינATP .באופן שבלתי תלוי בשאר הקומפלקסים שראינו
ממברנלי. היא תת היחידה שמסתובבת. אליה נכנסים הפרוטונים המגיעים מהחלל הבין Cתת יחידה פרוטונים נכנסים מצד אחד, אבל לא יכולים לצאת מבלי להשלים סיבוב ולצאת מהצד השני.
בנויה מאוסף של תעלות חוצות ממברנה. .open, loss, tightיכולות להיות במצב של: , ATP-בהן נוצר ה, יחידות ביטא 3-יחידות אלפא ו 3
-בכל פעם שהיא מסתובבת בהשכנה לה בתתי היחידות ביטא פורמציה סימטרית, גורמת לשינויי קונ-היא א (גמא) 𝛄תת היחידה מעלות. 120
אין צורך בסיבוב שלם, אבל כדי לשחרר אותו למטריקס יש צורך בהשלמת הסיבוב. ATPכדי לייצר
.𝐹1-ו 𝐹𝑂תורמת לקישור של תת היחידה אפסילון

12
.b-ו aבנויה משתי תעלות: )דלתא( 𝜹יחידה תת חצאי תעלה שאינם 2מסתיימת באמצע ) aול לצאת אל הצד השני של הממברנה, משום שהתעלה לא יכ aאל פרוטון שנכנס
רציפים(. שנוצר. ATP-וכך מאפשרת את שחרור ה openמעבירה תת יחידה ביטא שבאה איתה במגע לקונפורמציה של
היא יותר c. ברגע שהפרוטון נכנס כל תת היחידה cהפרוטון נמשך אל חומצה אספרטית טעונה שלילית שנמצאת בתת היחידה a-מ
, של החומצה האספרטית הטעונה cיציבה ופחות הידרופובית. הפרוטון מאזן מעט את המטען החשמלי השלילי של תת היחידה יבוב, עד שהסיבוב מושלם ופרוטונים יכולים להתחיל לצאת )המעגל הושלם(. כל פרוטון שנכנס מניע עוד קצת את הס .שלילית
המעגל יקרה שוב ושוב עד שכל הפרוטונים יצאו. לא מסתובבת. cבהיעדר פרוטונים, תת היחידה
.cפרוטונים, כתלות במספר הטבעות ביחידה 14או 10,12דורש כניסה של cסיבוב שלם של יחידה
מסתובבות. השאר סטאטיות.וגמא cרק היחידות
התהליך בנקודות:כל aפרוטון נכנס לחצי תעלה -
cפוגש חומצה אספרטית של תת היחידה -
נוספת מתפנה cנותן כוח סיבובי, יחידת -
טעונה חיובית ש'דוחפת' אותו החוצה ארגיניןכשהסיבוב מושלם והפרוטון מגיע שוב לנקודת ההתחלה שלו במעגל, יש שם -
הבין ממברנלי אל המטריקס הפנימי( פרוטון עבר מהמרווחהתוצאה: ) גורם לסיבוב של גמא cהסיבוב של -
:ATPיצירת
כל סיבוב של גמא גורם לשינויי קונפרומציה ביחידות ביטא:
Open .פתוחות :ADP +Pi .נכנסים אל יחידה ביטא Loos נוצר קירוב בין :ADP ו- Piרם נוצרה מולקולת , כך שאינן יכולות להשתחרר, אולם טATP. Tight נעולות. יצירת :ATP.
.ADP+Piשנוצרה משתחררת, קשירה חוזרת של Open :ATPחזרה אל
יחידות ביטא שהן באותה קונפורמציה. בכל רגע, יש אחת מהן בכל מצב קונפורמציוני. 2בשום שלב אין
קונפורמציה.יחידות אלפא וביטא נשארות במקום, הן לא זזות, רק משנות ות ביטא משינויים מגנה של היחיד -. אלפא משמשת קצת כבופרATPאך הן לא מייצרות ATPגם היחידות אלפא יכולות לקשור
קונפורמציונים, היא 'בולעת' מעט את התזוזות של יחידה ביטא אחת כדי שלא ישפיעו על יחידות הביטא האחרות.
.ATPשל גמא מייצר מולקולת כל סיבוב שלם .ATPמולקולות 3מייצר cלכן, כל סיבוב שלם של יחידה

13
, ADP+Pi-ל ATPמבצעת סיבוב לכיוון הנגדי יתרחש פירוק של cאם יחידה התגובה ההופכית. מתרחש לעיתים במצבים סינטטים.
.ADP-באספקת התלוי ATP-ייצור ה
ATPבקרה על ייצור
, מדד למצב האנרגטי של התא. ATP/ADPיחס
מעידה ATPכמות גבוהה של .ATPבתא ולכן תוביל לסינתזה מוגברת של רמת אנרגיה נמוכה זמין מעידה על ADPכמות גבוהה של .ATPעל רמת אנרגיה גבוהה בתא ותחול האטה בסינתזה
מלווה בהגברה של כל התהליכים שמובילים לנקודה זו: ATPשימו לב שעלייה בסינתזת גליקוליזה בה נוצר פירובאט -
Aחמצון פירובאט לאצטיל קו -
נשרף ליצירת כוח חיזורי Aבו אצטיל קו TCA-מעגל ה -
שרשרת מעבר האלקטרונים במסגרתה הכוח החיזורי משמש ליצירת גרדיאנט אלקטרוכימי -
ניםשרשרת מעבר האלקטרולמגיבים אספקת - ADP טרנסלוקז-נוקלאוטיד-אדניןמוכנס אל תוך המטריקס ע"י האנזים - NADH :NADH נוצר במיטוכונדריה במהלך ה-TCA רוב ה(-NADH ).וגם בציטופלסמה בעת הגליקוליזה
שקיימים בתא. מתרחשת באמצעות מספר 'שאטלים' מהציטופלסמה אל המטריקס של המיטוכונדריה NADHשל ההעברה 1. Malate-aspartate shuttle – 'המערכת הפעילה ביותר. מתחילים ממולקNADH ובסוף מכניסים דרך כמה ריאקציות
.שלם בסוף התהליך, אין אבדן אנרגיה NADHביוכימיות )לא צריך לזכור( מלט אל המיטוכונדריה, ממנו יווצר

14
2. Glycerophosphate shuttle – באבדן אנרגיה ולכן פחות יעיל. קיבלנו וה מלופוספט. -3-שאטל באמצעות גליצרולFADH .בסוף התהליך
קיפולים של ממברנת המיטוכונדריה
סינתז זה לזה, תוך יצירת קפל בממברנה. ATPנגרמים בין היתר משום היצמדות של שני קומפלקסים שונים של
Uncouplersמפרי צימוד:
קולות ש'גונבות' את הפרוטונים שהוצאו אל החלל הבין ממברנלי. כלומר מאפשרים מעבר חפשי של פרוטונים חזרה למטריקס, מול
ז.טסינ ATPשלא דרך ., ומכאן שמםATP-מפרים את הצימוד בין שרשרת מעבר האלקטרונים לתהליך יצירת ה
שני סוגים:
ברנה תוך כדי שהיא קושרת אליה פרוטונים.טיים, מכילים מולקולה שיכולה לעבור את הממתסינ .1
, מולקולה מאוד מסוכנת, נמכרה בעבר כתכשיר להרזיה. עוברת מאזור עשיר בפרוטונים לאזור עני בפרוטונים DNPלמשל, תוך כדי שהיא גוררת איתה פרוטון אל תוך המטריקס. אין קבלת אנרגיה לאחר ביצוע שרשרת מעבר האלקטרונים.
.FCCP דוגמה נוספת היאDNP ו-FCCP נקראותProtonophores.
.(heat) , נחשב לשומן יותר בריא. שחרור הפרוטונים גורם להמרת האנרגיה לחום(brown) בשכבות השומן החוםטבעיים: .2
לתינוקות או חיות קטנות יש הרבה שומן חום, לייצור חום, וככל שמתבגרים אחוז השומן החום יורד. משתמשים בשומן לייצור חום.כשלתינוקות קר, הם
ליצורים בוגרים יש מנגנונים שונים לייצור חום, למשל רעידות של השרירים.
איך זה פועל?UCP1 חומצת שומן ארוכה שנקשרת -הוא חלבון שיכול להעביר פרוטונים מצד אחד לצד שני, אבל זקוק למולקולה נוספתוהופכת אותו לאפיני UCP1-שנקשרת אל ה LCFA-Long Chain Fatty acid ומאפשרת את קליטת הפרוטונים. UCP1-אל ה
יותר לפרוטונים. סינתז. ATPהפרוטונים יעברו כך מצד אחד של הממברנה לצד השני, לא דרך
וחלקן מייצרות חום. ATPזה שומן מאוד עשיר במיטוכונדריות משום שחלקן מייצרות
כתוצאה משרשרת מעבר סיגנלים זו מיוצרים יל להפעלה של אדנילאז ציקלאז.כשקר, ההיפטלמוס במוח שולח סיגנל שמוב .uncouplersחלבוני
נקבל עבור כל נשא אלקטרונים? ATPכמה
.P/Oאו יחס ATP/2eזה נקרא יחס
.ATPמולק' 2.5נקבל NADHעבור .ATPמולק' 1.5)סוקסינט( נקבל FADעבור
נקבל עבור כל אטום חמצן בודד שהופך למולק' מים בשרשרת מעבר האלקטרונים( ATP)החישוב הוא לפי כמה מולק'
בעת מחסור בחמצן
וגורם לירידה במאזן ATPמבצע הידרוליזה של 𝐹1-שרשרת מעבר האלקטרונים יורדת, לא יווצר גרדיאנט פרוטונים, ופעילות האנרגטי של התא.
.𝐼𝐹1מובילה לשפעול של החלבון pH-ליצירת חומצה לקטית. הירידה ב המחסור בחמצן מוביל לנשימה אנאירובית שגורמת ועוצר את פעולתן. סינטז ATP: הוא נקשר אל שתי יחידות של האנזים 𝐹1מעכב את 𝐼𝐹1החלבון
15
4( שיעור18/328/)
פוטוסינתזה חושך, היום נתמקד בריאקצית האור.מחולק לריאקצית אור ולריאקצית
למה זה חשוב?
מאפשר את כל החיים על פני כדור הארץ.
אם זה לא מספיק, אז יש לעקרונות של פוטוסינתזה גם מלא יישומים במדע
Photophosphorylation –ריאקצית האור
מים אבל חשוב להבין שזו חמצון של ביצורים שונים מחמצנים מולקולות שונות. אנחנו נתמקד ב מצן.לא המולקולה היחידה שיכולה להתח
ייצור אלקטרונים באנרגיה גבוהה והפיכתם לכוח חיזורי. .ריאקצית החושך תוזן על ידי הכוח החיזורי הזה
חיזורי.כוח היצירה שליש אלטרנטיבות שונות להשגת אנרגיה, לא רק אנרגית האור, מה שחשוב הוא
איפה זה קורה?
בחיידקים פוטוסינתזים מתרחש בציטופלסמה.
מתרחש בכלורופלסט.בצמחים,
הוא כנראה חיידק פוטוסינתטי שנבלע על ידי תא כלורופלסט איקריוט, והיום הוא אברון בתאי צמח.
כל הריאקציה של הפוטוסינתזה מתרחשת בתוך חלל הסטרומה על ממברנת התילקואידים.
-הממברנה של התילקואידים היא מאוד מאוד עשירה בחלבוניםונים ששזור בליפידים )ולא ליהפך, כמו ממש כמו קומפלקס של חלב
שאנחנו רגילים לחשוב(. הסטרומה היא החלל שמקיף את התילקואידים.
הרבה מהחלבונים בממברנות התילקואידים מתפקדים בארגון הצורה הזו של ערימת הדיסקים, והם מונעים את האיחוי של הממברנות השונות.
לגודל של אטומים שונים. המבנה של סטן מאוד, גודלו הפיזי הוא אפסי גם ביחמה יתרונות המבנה הזה? פוטון הוא חלקיק ק כשהוא חודר פנימה. ןמה תשכבות שכבות מעלה את הסיכוי שהפוטון יתפס באח
:פוספורילציה
אור נקלט ע"י קומפלקס חלבוני - ייצור גרדיאנט פרוטונים בתוך התילקואיד - חמצון מים לחמצן - מחמצון זה מייצרים את הכוח החיזוריאלקטרונים שיוצאים -
כלורופילים ופיגמנטים שונים מסייעים לכלורופלסט בכך שהם מהווים רצפטורים לפוטון.מערכת מצומדת של קשרים כפולים. ככל שנוצרים יותר קשרים נוצרת יותר אנרגיה וניתן להסיט את אורך הגל שניתן הוא כלורופיל
רזוננס של הקשרים הכפולים. לבלוע. צימוד משמעו שמתקיים גם
גם הוא מע' מצומדת של קשרים כפולים עם רזוננס. Caroteneקרוטן, < עלולה להתרחש שבירה של מולקולות כימיות.--< עירור אלקטרונים --קליטה של פוטונים עתירי אנרגיה
ו. קרוטן משמש גם כחומר מגן מפני במערכת שתפקידה לקלוט כל הזמן שמש, צריך הגנה רצינית כדי שלא תתרחש השבירה הז ויבצע שבירה, הוא יקלט בקרוטן ויוכל להיות מוסט. DNA-השבירה. במקום שאלקטרון יקלט בחלבון או ב

16
יעילות פוטוסינתזה כתלות באורך גל
תחומים ור בנתבונן בפרופילי הבליעה של הכלורופילים השונים: מאפשרים לבלוע א. 400-500או 600-700באזור של הרוב כתום(.-)צבע אדום 650-700 וכן 450-500ספציפיים,
השמש נותנת אור לבן שמכיל את כל אורכי הגל, אולם בגלל האטמוספירה היא לא בעוצמה שווה על כל הספקטרום.
איך מתרחשת בליעה? איך תופסים פוטון כך שיגיע אל מרכז הריאקציה?
האלקטרון.: המקום בו מקפיצים את מרכז הריאקציה כלורופילים. 2הסיכוי לתפוס את הפוטון במרכז הריאקציה הוא מאוד נמוך.
הוא יקפוץ ממקום אחד -יש מערכת שלמה של הכלורופלסטים והכלורופילים והפיגמנטים, כך שלא משנה איפה נקלוט את הפוטון לשני עד שיגיע אל מרכז הריאקציה.
פתרון מבני של הקומפלקס הזה. נים המשתתפים בפוטוסינתזה.גילו את המבנה השלם של הקולט 2016-רק ב
כל הפיגמנטים, כל הכלורופילים, הם לא מעוגנים באופן רנדומלי אלא בצורה מאוד ספציפית שתאפשר את הקפצת האנרגיה
מדת, עד שתגיע לבור האנרגטי שהוא מרכז הריאקציה.ממערכת מצומדת אל מערכת מצו
לתפוס עד ארבעה פוטונים.ון ניתן בכל רגע נתככל שיש יותר אנרגיה האור מקפיץ את האלקטרון למצב מעורר במערכת המצומדת.
תתרחש קפיצה יותר גבוהה.
: פוטון יקלט רק אם הוא יכול לתת קפיצה שלמה של רמת אנרגיה לא קליטה ראשוניתנרגיה באמצעות יותר ולא פחות, צריך לקפוץ ביחידה שלמה. לכן, נפתח מגוון של רמות א
קולטנים שונים.
אם כשהאלקטרון מעורר, הוא יכול לחזור למצבו ההתחלתי תוך שחרור של חום או אור. תהיה ליד מולקולה שתשמש כבור אנרגטי קטן, האלקטרון המעורר יעבור אליה.
Quantum tunneling
הודאות.-אישרדינגר: רק כאשר דוגמים את המערכת מגלים את מצבה. עקרון
ברגע שהפוטון נכנס למערכת יש אנרגיה בתוך הכלורופיל.
אנחנו רוצים להשתמש באנרגיה הזו, ולא נרצה שתשתחרר בצורת חום )היא גם עלולה לשרוף את הצמח(.
את ענן הסתברותי מסויים. ברגע שעברנו מולקולה איפסנו גל הוא הסתברות מסויימת של מציאת האנרגיה במקום מסויים. ההסתברות שהגל יהיה במולקולה הקודמת, וכעת שוב נוצר ענן הסתברותי לאן תעבור האנרגיה. כל פעם יש קפיצה עם הענן
ההיסתברותי ממולקולה ממולקולה. כך עד שהפוטון יקלט במרכז הריאקציה, שמה שמיוחד בו הוא שמרכז הריאקציה משמש כבור ית היא לא תעבור למקום אחר.אנרגטי. האנרגיה יותר נמוכה ולכן הסתברות
, הוא עובר מנשא בעל אנרגיה Quantum tunneling-אם הגענו למרכז הריאקציה והקפצנו אלקטרון, גם האלקטרון הזה עובר ב
. לאלקטרון יש מסה, ולכן בוודאות יתרחש איבוד של אנרגיה.גבוהה לנשא בעל אנרגיה נמוכה יותר
שימו לב: פעם בכל אנרגיה שיאבד אלקטרון כמו)אלקטרון( מסה בעל לחלקיק בניגוד QT בביצוע אנרגיה יאבד לא)פוטון( מסה חסר חלקיק .ויורד הולך פוטנציאל בעלי לנשאים מועברים שהם ככל שלהם מהאנרגיה מאבדים אלקטרונים, שלמדנו כפי. QT שיבצע
העברה בין נשאים.של אנרגיה משום שאין בעת הקפצת האלקטרון במרכז הריאקציה אין איבוד
הפרש ואז העזר, בפיגמנטי א קיים רצף בין תחומי קליטת הפוטוןרק אם ל QT-יכול להתרחש איבוד אנרגיה בעת מעבר של פוטון ב .האנרגיה ישתחרר כחום

17
הוא לא אקראי. סידור החלבונים בתוך התילקואידים
בדיפוסיה?נשאלת השאלה, אם המבנה כל כך צפוף, איך נספק מספיק מים
במערכת של מים, פרוטונים לא חייבים לעבור דיפוסיה מצד לצד. מתקבל רזוננס דרך קשרי המימן במים, מה שלוקח פחות זמן. רזוננס של קשרים בין מולקולות מים. לא מבצעים דיפוסיה של יון אלא הקפצה של קשרים שהיא הרבה יותר מהירה.
כנראה שלא. נס הוא לאו דווקא הפרוטון שיוצא,הפרוטון שנכלסט. זה מתרחש כך גם בציטוכרום וגם בכלורופ
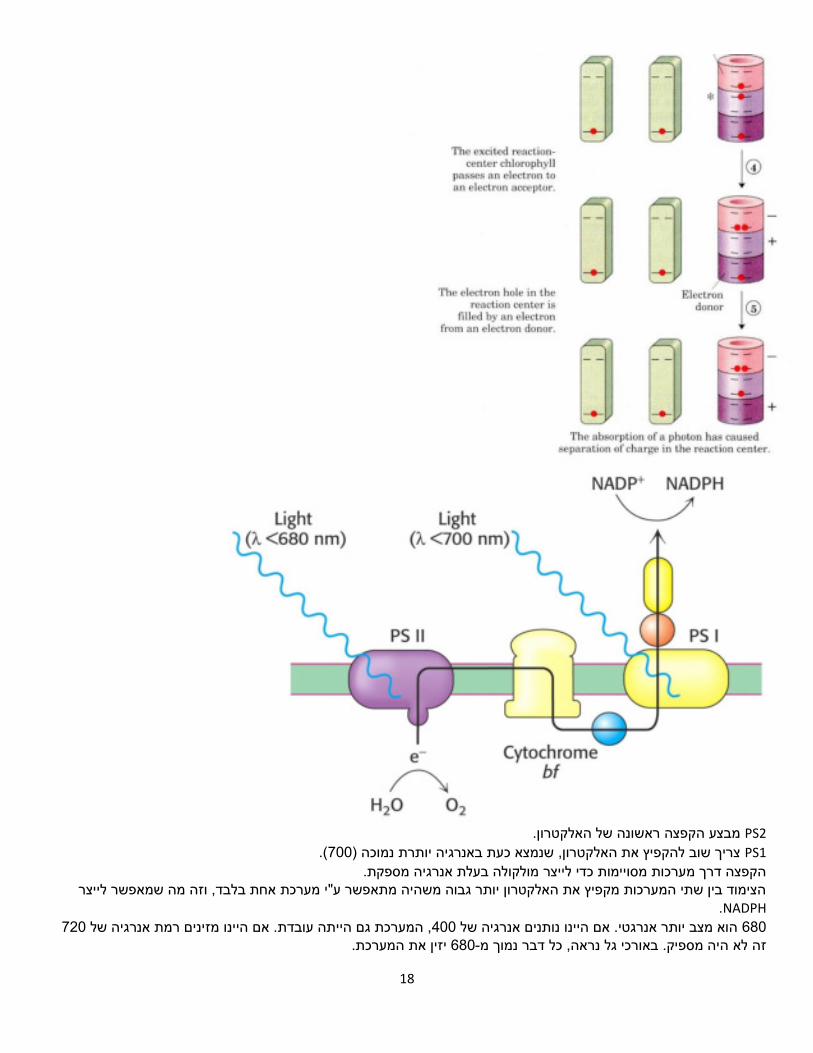
18
PS2 .מבצע הקפצה ראשונה של האלקטרון PS1 (.700נמוכה ) תצריך שוב להקפיץ את האלקטרון, שנמצא כעת באנרגיה יותר
מולקולה בעלת אנרגיה מספקת.הקפצה דרך מערכות מסויימות כדי לייצר הצימוד בין שתי המערכות מקפיץ את האלקטרון יותר גבוה משהיה מתאפשר ע"י מערכת אחת בלבד, וזה מה שמאפשר לייצר
NADPH. 720, המערכת גם הייתה עובדת. אם היינו מזינים רמת אנרגיה של 400הוא מצב יותר אנרגטי. אם היינו נותנים אנרגיה של 680
יזין את המערכת. 680-היה מספיק. באורכי גל נראה, כל דבר נמוך מ זה לא
19
. PS1תהיה הזנה רק של 700-ל 680בטווח שבין
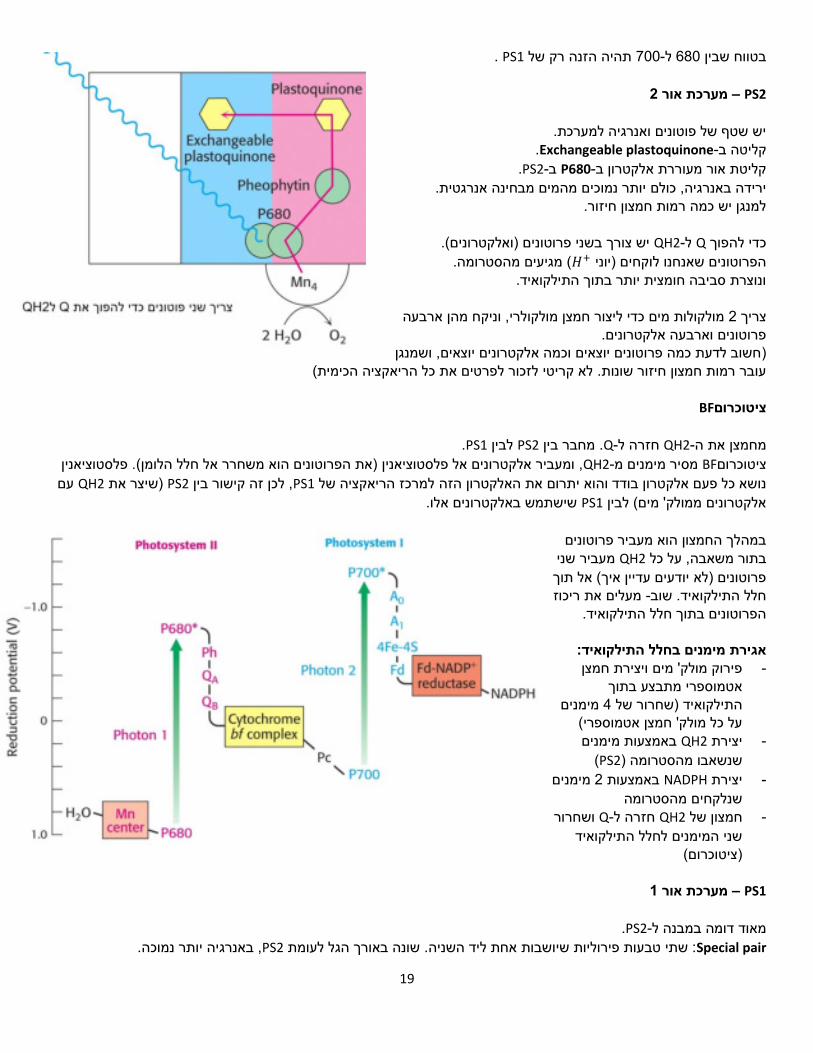
PS2 – 2מערכת אור
יש שטף של פוטונים ואנרגיה למערכת. .Exchangeable plastoquinone-קליטה ב
.PS2-ב P680-קליטת אור מעוררת אלקטרון ב .יותר נמוכים מהמים מבחינה אנרגטיתירידה באנרגיה, כולם
למנגן יש כמה רמות חמצון חיזור.
.)ואלקטרונים( וטוניםריש צורך בשני פ QH2-ל Qכדי להפוך הסטרומה.( מגיעים מ+𝐻הפרוטונים שאנחנו לוקחים )יוני
ונוצרת סביבה חומצית יותר בתוך התילקואיד.
ארבעה ןמולקולרי, וניקח מהמולקולות מים כדי ליצור חמצן 2צריך פרוטונים וארבעה אלקטרונים.
חשוב לדעת כמה פרוטונים יוצאים וכמה אלקטרונים יוצאים, ושמנגן ) (לא קריטי לזכור לפרטים את כל הריאקציה הכימית עובר רמות חמצון חיזור שונות.
BFציטוכרום
.PS1לבין PS2מחבר בין .Q-חזרה ל QH2-מחמצן את ה
. פלסטוציאנין)את הפרוטונים הוא משחרר אל חלל הלומן( , ומעביר אלקטרונים אל פלסטוציאניןQH2-מסיר מימנים מ BFציטוכרוםעם QH2)שיצר את PS2, לכן זה קישור בין PS1למרכז הריאקציה של זהה ןיתרום את האלקטרו נושא כל פעם אלקטרון בודד והוא
שישתמש באלקטרונים אלו. PS1אלקטרונים ממולק' מים( לבין
במהלך החמצון הוא מעביר פרוטונים מעביר שני QH2 לבתור משאבה, על כ
פרוטונים )לא יודעים עדיין איך( אל תוך מעלים את ריכוז -חלל התילקואיד. שוב
הפרוטונים בתוך חלל התילקואיד.
אגירת מימנים בחלל התילקואיד:פירוק מולק' מים ויצירת חמצן -
ספרי מתבצע בתוך אטמומימנים 4התילקואיד )שחרור של
על כל מולק' חמצן אטמוספרי(
באמצעות מימנים QH2יצירת - (PS2שנשאבו מהסטרומה )
מימנים 2באמצעות NADPHיצירת - שנלקחים מהסטרומה
ושחרור Q-חזרה ל QH2חמצון של -שני המימנים לחלל התילקואיד
)ציטוכרום(
PS1 – 1מערכת אור
.PS2-דומה במבנה ל מאודSpecial pair.שונה באורך הגל לעומת : שתי טבעות פירוליות שיושבות אחת ליד השניהPS2 .באנרגיה יותר נמוכה ,
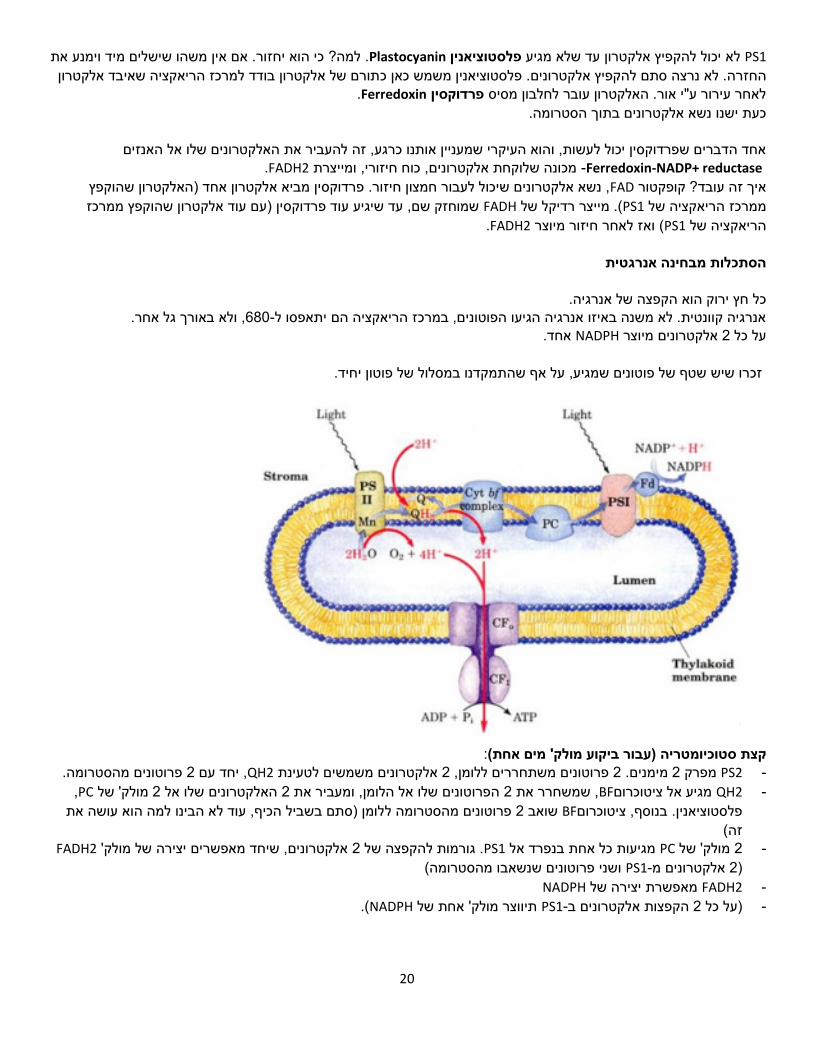
20
PS1 פלסטוציאניןלא יכול להקפיץ אלקטרון עד שלא מגיע Plastocyanin ?כי הוא יחזור. אם אין משהו שישלים מיד וימנע את . למהתורם של אלקטרון בודד למרכז הריאקציה שאיבד אלקטרון פלסטוציאנין משמש כאן כ להקפיץ אלקטרונים. החזרה. לא נרצה סתם
.Ferredoxin פרדוקסיןלאחר עירור ע"י אור. האלקטרון עובר לחלבון מסיס כעת ישנו נשא אלקטרונים בתוך הסטרומה.
רגע, זה להעביר את האלקטרונים שלו אל האנזיםאחד הדברים שפרדוקסין יכול לעשות, והוא העיקרי שמעניין אותנו כ
Ferredoxin-NADP+ reductase- מכונה שלוקחת אלקטרונים, כוח חיזורי, ומייצרתFADH2. האלקטרון שהוקפץ , נשא אלקטרונים שיכול לעבור חמצון חיזור. פרדוקסין מביא אלקטרון אחד )FADאיך זה עובד? קופקטור
)עם עוד אלקטרון שהוקפץ ממרכז שמוחזק שם, עד שיגיע עוד פרדוקסין FADH. מייצר רדיקל של (PS1ממרכז הריאקציה של .FADH2ואז לאחר חיזור מיוצר ( PS1הריאקציה של
הסתכלות מבחינה אנרגטית
כל חץ ירוק הוא הקפצה של אנרגיה.
, ולא באורך גל אחר.680-יתאפסו ל אנרגיה קוונטית. לא משנה באיזו אנרגיה הגיעו הפוטונים, במרכז הריאקציה הם אחד. NADPHאלקטרונים מיוצר 2על כל
יחיד.על אף שהתמקדנו במסלול של פוטון זכרו שיש שטף של פוטונים שמגיע,
:(אחת )עבור ביקוע מולק' מים קצת סטוכיומטריה
- PS2 אלקטרונים משמשים לטעינת 2פרוטונים משתחררים ללומן, 2מימנים. 2מפרקQH2 פרוטונים מהסטרומה. 2, יחד עם
- QH2 מגיע אל ציטוכרוםBF מולק' של 2האלקטרונים שלו אל 2הפרוטונים שלו אל הלומן, ומעביר את 2, שמשחרר אתPC ,לומן )סתם בשביל הכיף, עוד לא הבינו למה הוא עושה את פרוטונים מהסטרומה ל 2שואב BF. בנוסף, ציטוכרוםפלסטוציאנין
זה(
FADH2אלקטרונים, שיחד מאפשרים יצירה של מולק' 2. גורמות להקפצה של PS1מגיעות כל אחת בנפרד אל PCמולק' של 2 - ושני פרוטונים שנשאבו מהסטרומה( PS1-אלקטרונים מ 2)
- FADH2 מאפשרת יצירה שלNADPH
(.NADPHתיווצר מולק' אחת של PS1-אלקטרונים בהקפצות 2)על כל -

21
מעבר אלקטרונים מעגלי
.NADPHיתרחש כאשר יש עודף של , NADPH. אין סינתזה של PS1-ו PS2מתרחש עדיין במעורבות של
מוגברת מעט. ATPעדיין נוצר גרדיאנט פרוטונים ויצירת
:השוואה בין פוטוסינתזה לבין פוספורלציה חמצונית האנזימים שונים, אבל בשניהם יש ציטוכרומים שיודעים ליצור -
גרדיאנט
האלקטרונים מתקבלים משמש לעומת משריפה של מזון -
מתאפשר גם ללא אור ATPייצור
מפרי צימוד
. הפוטוסינתזה לא +K+/H. משחלף ATPמערכב סינתזת ניגריסין
במשך היום. ATP, אך הצמח ימות משום שלא יצליח לייצר עתיפג במשך הלילה, צמח נושם בדיוק כמו בע"ח או אדם.
לא משנה את הגרדיאנט. ולינומיצין
מולקולה ניטרלית שעוברת דרך הממברנה ויכולה לתפוס אמוניה
פרוטון וכך להפר את הצימוד.
מעכבי מעבר אלקטרונים
DCMU : לא מונע את הקפצת האלקטרונים, אבל מונע ייצור שלNADPH לו להגיע הלאה ןאת האלקטרון בדרך ולא נות תופסמשום ש
.BFמציטוכרום
)תמיד במבחן יש שאלה על מעכב. הכוונה היא תמיד לשאלה מה קורה לצמח אחרי זמן מה. כלומר אם בשניות הראשונות לא פן מיידי.(מעניין אותנו מה שקורה אחרי כמה דקות ולא באו -מתרחש עיכוב אבל אחרי כמה דקות הצמח ימות
לא לא יווצר גרדיאנט פרוטונים , לאחר זמן מה: יפסק מעבר האלקטרונים הלינארים והמעגלי PS2אם מעכבים את פעילות
.ATP-ו NADPHיווצרו עדיין יקפיץ אלקטרונים, אולם לא יווצר PS2מרכז הריאקציה של : BFלבין ציטוכרום PS2אם מעכבים את מעבר האלקטרונים בין
.ATP-ו NADPHלא יווצרו גרדיאנט פרוטונים PS1-תוצרי הביניים ילכו ויצטברו עד שכל החלבונים שקודמים ללאחר זמן מה כל המערכת תיתקע. :PS1אם מעכבים את פעילות
בשרשרת יעוכבו. זה יקרה אחרי זמן ארוך.
במהלך שינוי אור מהיר
הצמח מגן על עצמו?איך צמח חי בסביבה משתנה, רגע יש הרבה שמש ואז עובר ענן ויש פחות שמש.
.פיקוביליזוםשכל הזמן מנסות לקלוט פוטונים. למשל, כשיש ענן. PSיש אנטנות יותר גדולות משל ה
עלולה לעלות מהר מדי והצמח ואיך נוכל לעצור את הפוטונים במקרה שנקלטים יותר מדי? אם נתפסים יותר מדי פוטונים הטמפ' ישרף.
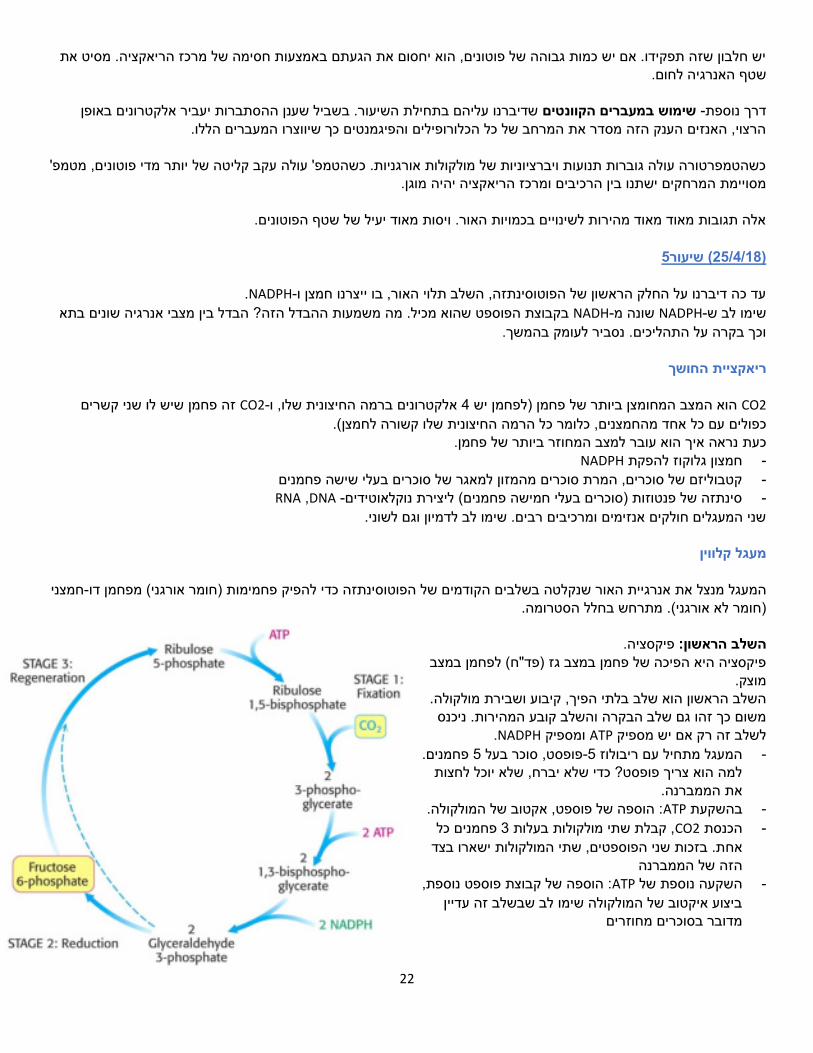
22
יש חלבון שזה תפקידו. אם יש כמות גבוהה של פוטונים, הוא יחסום את הגעתם באמצעות חסימה של מרכז הריאקציה. מסיט את שטף האנרגיה לחום.
נים באופן שדיברנו עליהם בתחילת השיעור. בשביל שענן ההסתברות יעביר אלקטרו שימוש במעברים הקוונטים -דרך נוספת
הרצוי, האנזים הענק הזה מסדר את המרחב של כל הכלורופילים והפיגמנטים כך שיווצרו המעברים הללו.
כשהטמפרטורה עולה גוברות תנועות ויברציוניות של מולקולות אורגניות. כשהטמפ' עולה עקב קליטה של יותר מדי פוטונים, מטמפ' ומרכז הריאקציה יהיה מוגן.מסויימת המרחקים ישתנו בין הרכיבים
אלה תגובות מאוד מאוד מהירות לשינויים בכמויות האור. ויסות מאוד יעיל של שטף הפוטונים.
5( שיעור184/25/)
.NADPH-וחמצן עד כה דיברנו על החלק הראשון של הפוטוסינתזה, השלב תלוי האור, בו ייצרנו הבדל בין מצבי אנרגיה שונים בתא בקבוצת הפוספט שהוא מכיל. מה משמעות ההבדל הזה? NADH-שונה מ NADPH-שימו לב ש
וכך בקרה על התהליכים. נסביר לעומק בהמשך.
החושךריאקציית
CO2 אלקטרונים ברמה החיצונית שלו, ו 4פחמן )לפחמן יש הוא המצב המחומצן ביותר של-CO2 זה פחמן שיש לו שני קשרים אחד מהחמצנים, כלומר כל הרמה החיצונית שלו קשורה לחמצן(.כפולים עם כל
נראה איך הוא עובר למצב המחוזר ביותר של פחמן. כעת NADPHחמצון גלוקוז להפקת - קטבוליזם של סוכרים, המרת סוכרים מהמזון למאגר של סוכרים בעלי שישה פחמנים -
DNA ,RNA -סינתזה של פנטוזות )סוכרים בעלי חמישה פחמנים( ליצירת נוקלאוטידים -
שני המעגלים חולקים אנזימים ומרכיבים רבים. שימו לב לדמיון וגם לשוני.
מעגל קלווין
חמצני-דו ןמפחמ ()חומר אורגני פחמימות להפיק כדי הפוטוסינתזה של הקודמים בשלבים שנקלטה האור אנרגיית את מנצל המעגל מתרחש בחלל הסטרומה. .)חומר לא אורגני(
פיקסציה. שלב הראשון:ה
פיקסציה היא הפיכה של פחמן במצב גז )פד"ח( לפחמן במצב מוצק.
בלתי הפיך, קיבוע ושבירת מולקולה. שלב השלב הראשון הוא ניכנס משום כך זהו גם שלב הבקרה והשלב קובע המהירות.
.NADPHומספיק ATPלשלב זה רק אם יש מספיק פחמנים. 5פופסט, סוכר בעל -5המעגל מתחיל עם ריבולוז -
למה הוא צריך פופסט? כדי שלא יברח, שלא יוכל לחצות את הממברנה.
: הוספה של פוספט, אקטוב של המולקולה. ATPבהשקעת -פחמנים כל 3, קבלת שתי מולקולות בעלות CO2הכנסת -
ת. בזכות שני הפוספטים, שתי המולקולות ישארו בצד אח הזה של הממברנה
: הוספה של קבוצת פוספט נוספת, ATPהשקעה נוספת של -ביצוע איקטוב של המולקולה שימו לב שבשלב זה עדיין
מדובר בסוכרים מחוזרים
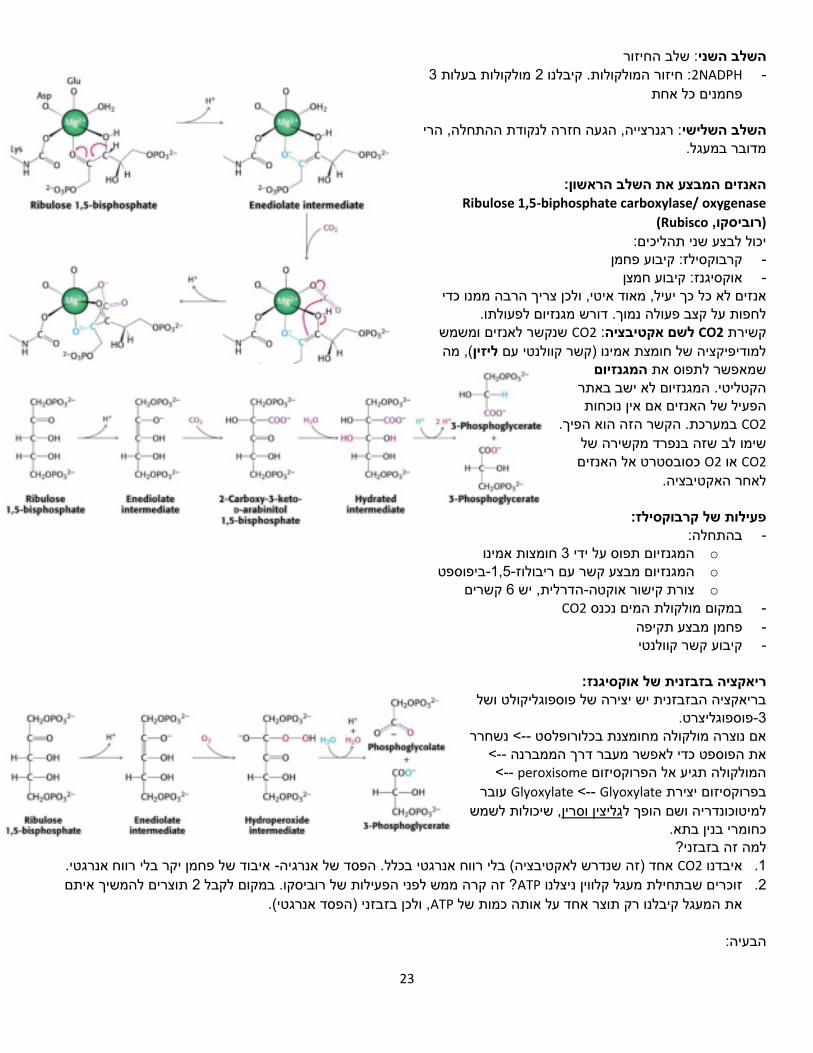
23
: שלב החיזורהשלב השני- 2NADPH 3מולקולות בעלות 2: חיזור המולקולות. קיבלנו
פחמנים כל אחת
: רגנרצייה, הגעה חזרה לנקודת ההתחלה, הרי השלב השלישי מדובר במעגל.
:הראשוןהמבצע את השלב האנזים
Ribulose 1,5-biphosphate carboxylase/ oxygenase
(Rubisco ,רוביסקו)
יכול לבצע שני תהליכים: פחמן קרבוקסילז: קיבוע -
חמצןקיבוע אוקסיגנז: -
כך יעיל, מאוד איטי, ולכן צריך הרבה ממנו כדי אנזים לא כל לחפות על קצב פעולה נמוך. דורש מגנזיום לפעולתו.
משמש ושנקשר לאנזים CO2: לשם אקטיבציה CO2קשירת (, מה ליזיןלמודיפיקציה של חומצת אמינו )קשר קוולנטי עם
המגנזיוםשמאפשר לתפוס את הקטליטי. המגנזיום לא ישב באתר
האנזים אם אין נוכחות הפעיל שלCO2 .במערכת. הקשר הזה הוא הפיך
שימו לב שזה בנפרד מקשירה של CO2 אוO2 כסובסטרט אל האנזים
לאחר האקטיבציה.
:קרבוקסילזפעילות של בהתחלה: -
o חומצות אמינו 3המגנזיום תפוס על ידי
o ביפוספט-1,5-המגנזיום מבצע קשר עם ריבולוז
o קשרים 6הדרלית, יש -אוקטהצורת קישור
CO2במקום מולקולת המים נכנס -
פחמן מבצע תקיפה -
קיבוע קשר קוולנטי -
ריאקציה בזבזנית של אוקסיגנז:
בריאקציה הבזבזנית יש יצירה של פוספוגליקולט ושל פוספוגליצרט.-3
נשחרר <--אם נוצרה מולקולה מחומצנת בכלורופלסט < --את הפוספט כדי לאפשר מעבר דרך הממברנה
< -- peroxisomeהמולקולה תגיע אל הפרוקסיזום עובר Glyoxylate -- >Glyoxylateבפרוקסיזום יצירת
, שיכולות לשמש גליצין וסריןלמיטוכונדריה ושם הופך ל כחומרי בנין בתא. למה זה בזבזני?
בלי רווח אנרגטי. יקר ןאיבוד של פחמ -אנרגיההפסד של בלי רווח אנרגטי בכלל. ( אחד )זה שנדרש לאקטיבציה CO2איבדנו .1
תוצרים להמשיך איתם 2? זה קרה ממש לפני הפעילות של רוביסקו. במקום לקבל ATPזוכרים שבתחילת מעגל קלווין ניצלנו .2 .)הפסד אנרגטי( , ולכן בזבזניATPאת המעגל קיבלנו רק תוצר אחד על אותה כמות של
הבעיה:

24
לקבע בעייתי כי נרצה זה . )כתוצאה ממבנה האנזים( ככל שהטמפרטורה עולה, פעילותו כאוקסיגנז תגבר על פעילותו כקרבוקסילז .פד"ח ולא חמצן, המטרה היא להוסיף פחמן
הפתרון: מקצב הריאקציה של האוקסיגנז. 4קצב הריאקציה של הקרבוקסילז גבוה פי .1 שגורם להפסקת ריאקציית האוקסיגנז.המגנזיום, מה מהאנזים, וכך לשחרור CO2-של הגורמת לשחרור CO2ירידה ברמת .2
ואם כבר נוצר חמצון, מה נעשה עם המולקולה?
וכעת נחזור לריאקציה הכן יעילה, :ריאקציה של קרבוקסילז
משמאל, תרשים מפורט יותר של השלב הראשון, שלב הפיקסציה. באופן מלאכותי )כשאין אור בסביבה(, יתרחש קיבוע. NADPH-ו ATPאם נספק לתא שימו לב:
ברגע שיש זמינות של אנרגיה יתרחש קיבוע, כי במעגל קלווין אין שום צורך באור, רק בזמינות המרכיבים.
.transketolase and aldolase, מבוצע על ידי שני אנזימים: השלב השלישי, שלב הרגנרציה
פחמנים, ולכן צריך את שני 5פחמנים למולק' של 6-ו 3אנחנו צריכים להפוך מולק' של .והאל האנזימים
Transketolase )נקודה חשובה ודע להעביר שני פחמנים בין קטוז לבין אלדוז. י )טרנסרטולז (.E1, ביחידה בפירובט קרבוקסילז TPP)ראינו TPP-היא שהאנזים משתמש ב
Aldolase )ביפוספט לשתי -1,6-שובר פרוקטוזלמדנו על פעילותו בגליקוליזה: )אלדולז. GAP-ו DHAPמולקולות:
כיוונית, -מדובר בתגובה דוולכן יתקיים שיווי משקל
בהתאם לכמות המגיבים מה והתוצרים שיש בתא.
בהקשר של שמעניין אותנו יכול Aldolaseמעגל קלווין:
nלהפוך מולקולה בעלת ., וליהפךפחמנים n+3למולקולה בעלת פחמנים
5ויוצרים מהם שתי מולק' של GAPפוספט ו-6-פרוקטוזמולק' 2בתהליך רב שלבי לוקחים )פירוט על פוספט שיכול להיכנס שוב למעגל קלווין 5פחמנים כל אחת, אחת מהן היא ריבוז
.כך במעגל הפנטוז פוספט(
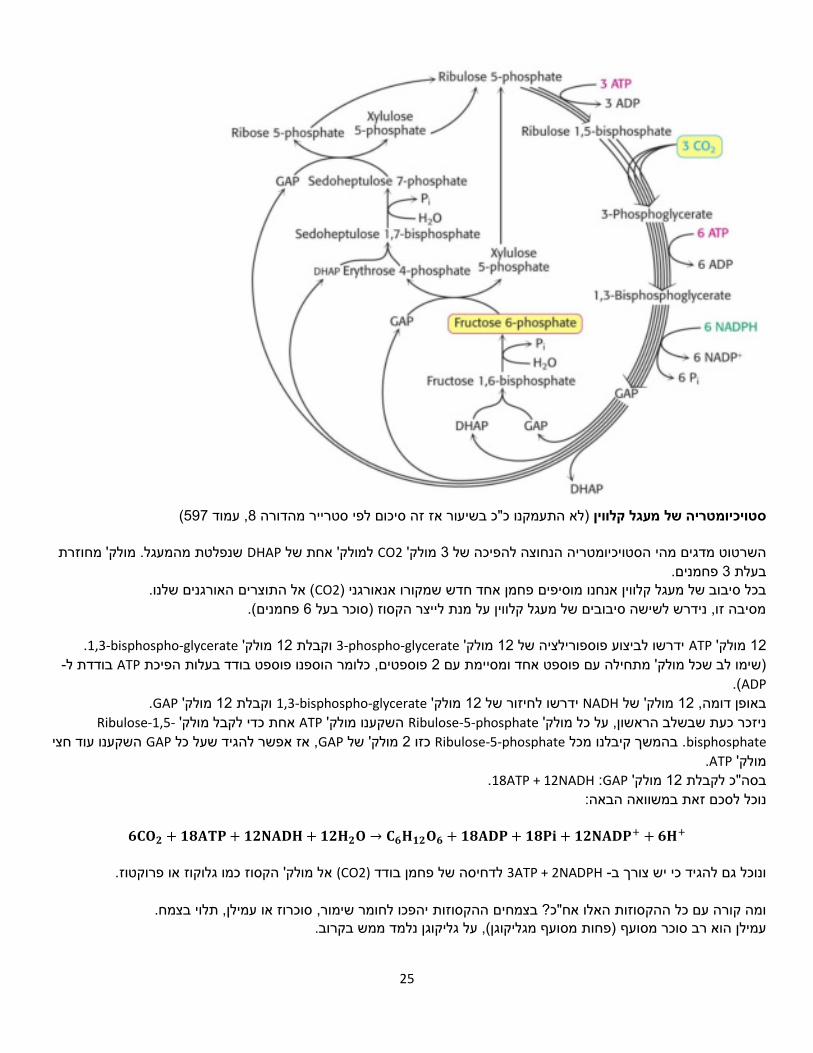
25
(597, עמוד 8)לא התעמקנו כ"כ בשיעור אז זה סיכום לפי סטרייר מהדורה סטויכיומטריה של מעגל קלווין
שנפלטת מהמעגל. מולק' מחוזרת DHAPלמולק' אחת של CO2מולק' 3השרטוט מדגים מהי הסטויכיומטריה הנחוצה להפיכה של פחמנים. 3בעלת
( אל התוצרים האורגנים שלנו.CO2פחמן אחד חדש שמקורו אנאורגני )בכל סיבוב של מעגל קלווין אנחנו מוסיפים .פחמנים( 6מסיבה זו, נידרש לשישה סיבובים של מעגל קלווין על מנת לייצר הקסוז )סוכר בעל
.bisphospho-glycerate-1,3מולק' 12 וקבלת phospho-glycerate-3 מולק' 12ידרשו לביצוע פוספורילציה של ATPמולק' 12
-בודדת ל ATPהפיכת פוספטים, כלומר הוספנו פוספט בודד בעלות 2)שימו לב שכל מולק' מתחילה עם פוספט אחד ומסיימת עם ADP).
.GAPמולק' 12וקבלת bisphospho-glycerate-1,3מולק' 12 לחיזור שלידרשו NADHמולק' של 12באופן דומה, -Ribulose-1,5אחת כדי לקבל מולק' ATPהשקענו מולק' Ribulose-5-phosphateניזכר כעת שבשלב הראשון, על כל מולק'
bisphosphate בהמשך קיבלנו מכל .Ribulose-5-phosphate מולק' של 2כזוGAP אז אפשר להגיד שעל כל ,GAP השקענו עוד חצי .ATPמולק'
.GAP :18ATP + 12NADHמולק' 12בסה"כ לקבלת הבאה: נוכל לסכם זאת במשוואה
𝟔𝐂𝐎𝟐 + 𝟏𝟖𝐀𝐓𝐏 + 𝟏𝟐𝐍𝐀𝐃𝐇 + 𝟏𝟐𝐇𝟐𝐎 → 𝐂𝟔𝐇𝟏𝟐𝐎𝟔 + 𝟏𝟖𝐀𝐃𝐏 + 𝟏𝟖𝐏𝐢 + 𝟏𝟐𝐍𝐀𝐃𝐏+ + 𝟔𝐇+
אל מולק' הקסוז כמו גלוקוז או פרוקטוז. (CO2)לדחיסה של פחמן בודד 3ATP + 2NADPH -ונוכל גם להגיד כי יש צורך ב
.סוכרוז או עמילן, תלוי בצמח, ההקסוזות יהפכו לחומר שימורם בצמחי ומה קורה עם כל ההקסוזות האלו אח"כ? )פחות מסועף מגליקוגן(, על גליקוגן נלמד ממש בקרוב. עמילן הוא רב סוכר מסועף

26
C-3 plants פחמנים. 3רוב הצמחים, מתקיימים ממולקולות של
.מזופיל, תאי מהאוויר CO2מעגל קלווין מתרחש באותם תאים בהם נקלט C3בצמחי בתוך התא. CO2היתרון: לא צריך לשנע
ואז יורדת היעילות של האנזים רוביסקו, מפני ששתי המולקולות יכולות לשמש כסובסטרט ויש O2נקלט גם CO2החסרון: יחד עם ביניהן תחרות.
כדי לייצר הקסוז אחד. ATPמולקולות 18נדרשות צמחים אלו ב
C-4 plants מהצמחים. 4%
.ולחיםצמחים שחיים קרוב יותר לאזור קו המשווה, אזורים חמים הם צמחים טרופיים, C4צמחי באזורים חמים יש בעיה עם רוביסקו: הוא יבצע יותר אוקסיגנציה, וזו התגובה הפחות רצויה.
CO2-ה( ואז מעביר את Mesophyllבתאי מזופיל ) CO2-הצמח קולט את ה :CO2תרחשת אגירה של כדי להתמודד עם החום ( שם יתרחש מעגל קלווין. Bandle sheathשנקלט אל תאי נדן הצרור )
נדחסות CO2מולק' 4פחמנים, 4. שאטל של Pyruvate-הופכת ל Malateכנשאית של פחמן דו חמצני. Malateשימוש במולקולה טט היא זו שמועברת ע"י מלט אל תאי , ומולק' אוקסלואצ(C4פחמנים, לכן השם הוא צמחי 4)מולק' של יחד ליצירת אוקסלואצטט
נדן הצרור.
כמעט ובכלל. O2היתרון: היעילות של רוביסקו עולה מאוד, משום שאין תחרות של מתא אחד אל תא אחר עולה באנרגיה. CO2-החסרון: השאטל של ה כדי ליצור הקסוז אחד. ATPמולקולות 30בצמחים אלו נדרשות
בעיה של איבוד מים משום שהם באזור טרופי, ולכן אין בעיה של פתיחת פיוניות במהלך היום.שימו לב שלצמחים אלו אין
CAM plants
מהצמחים. 7% O2-ו CO2כדי למנוע איבוד של מים בעת פתיחת הפיוניות של הצמח )דרכן נקלטים , ויבשים צמחי מדבר ומותאמים לאזורים חמים
.CO2יות סגורות במהלך היום ובלילה אוגרים אך גם מתאדים מים( הם משאירים את הפיונשנקלט אל מולק' אורגניות, ובלילה CO2-מחברים במהלך היום את ה CAM( צמחי C4במקום לבצע שאטל אל תא אחר )כמו
משחררים אותו כדי שיוכל להיכנס אל מעגל קלווין.
בז הרבה יותר אנרגיה כדי לייצר את אותה כמות סוכר.הוא פחות מותאם לתנאי קור משום שהוא יבז CAM-ו C4 יאבולוציונית, צמח (. )עוד השלכה מסוכנת של ההתחממות הגלובלית , ואלו הם מרבית מיני הצמחים הקיימיםלא מותאם לאזורים חמים C3צמח
שזה טיפה מוזר כי ארכיאות לא -נבחין כי הוא קיים בבקטריות, אקריוטים וגם בארכיאות אם נתבונן בעץ האבולוציה של רוביסקו
מבצעות פוטוסינתזה. מחזק את ההשערה שהוא התפתח אבולוציונית לתפקיד אחר.
יש סט שלם ושונה של חלבונים לקיבוע פחמן שלא קשור למעגל קלווין ולאנזים רוביסקו.

27
נתזה?מה קורה בתא שלא מבצע פוטוסי
או, בתא צמח בשעות הלילהבאותם בדיוק התהליך ההפוך לרגנרציה של מעגל קלווין, וגם משתמש
כדי לייצר פנטוזים, נוכל לפרק מאגרים של הקסוזים או לייצר האנזימים.כל התהליכים האלו קורים פנטוזים באמצעות אנרגיה וכוח חיזורי.
בציטוזול.
כונדריה הם פשוט ישרפו לייצור אנרגיה.אם פנטוזות והקסוזות יהיו במיטו בציטוזול אנחנו בונים דברים ובמיטוכונדריה אנחנו שורפים דברים. -ככלל
מסלול פנטוז פוספט )בתרשים שמשמאל(
בהתאם לצרכי התא. NADPH-תפקידו העיקרי לספק פנטוזות ו Oxidative step - דהידרוגנציה .1 פוספט-6-חמצון של גלוקוז -
הטבעתפתיחה של -
חמצון נוסף -
הידרוגנזות, שני תהליכי חמצון, למה? כי אנחנו במצב -יש צורך בשתי דה .COHשל
CO2 .הוא חמצון מלא של פחמן, ולא נגיע אליו בצעד אחד בלבד
דהידרוגנז הוא נקודת הבקרה של התהליך, -פוספט-6-גלוקוזהאנזים מבצע את החמצון הראשון.
.NADPHוכן יצירת שתי מולקולות CO2תוך פליטה של פוספט -5-פוספט הופך לריבולוז-6-בשלב החמצון גלוקוז
Phosphopentose) פוספט יכול לעבור אח"כ איזומריזציה ע"י אותו אנזים שמשתתף במעגל קלווין-5-ריבולוזאיזומריזציה:
isomeraseספט.פו-5-ולהפוך לריבוז איזומראז(-פנטוז-, פוספו 2. Non-Oxidative step
, מולקולות אלו יכולות להיכנס לגליקוליזה או לגלוקונאוגנזה בהתאם לצרכי התא.GAP-פוספט ו-6-הפקת גלוקוז .Transealdolase-ו transketolaseשלב זה מזורז ע"י
פקטור.-כקו TPPשימו לב! טרנסקטולזות דורשים בשלב הלא מחמצן.פוספט, ולכן יש צורך גם -5-יותר משהם זקוקים לריבוז NADPH-רוב התאים זקוקים רוב הזמן ל
.(גליקוליזה וגלוקונאוגנזה) מטבוליים אחרים /מסלוליםמשום שניתן לקחת את תוצריו למעגליםהשלב הזה הוא גם צומת מטבולית,
סוכרים שמגיעים מהמזון .6וגם סוכרי 5גם סוכרי
לסוכרי 5או 4רגנרציה של סוכרי שמקורם במזון, יותר משאנחנו צריכים, לכן חשובה היכולת לבצע 5לעיתים יהיו לנו הרבה סוכרי .(6או להשתמש בהם )למשל גליקוליזה מתחילה גם היא מסוכר שנוכל לאגור 6
קטולז. -המעברים הללו מבוצעים על ידי טרנס פחמנים מהתזונה הולכים לרוב לגליקוליזה. 5סוכרי
פוספט?-6המטבוליזם בתא קובע מה יעשה בגלוקוזאיך
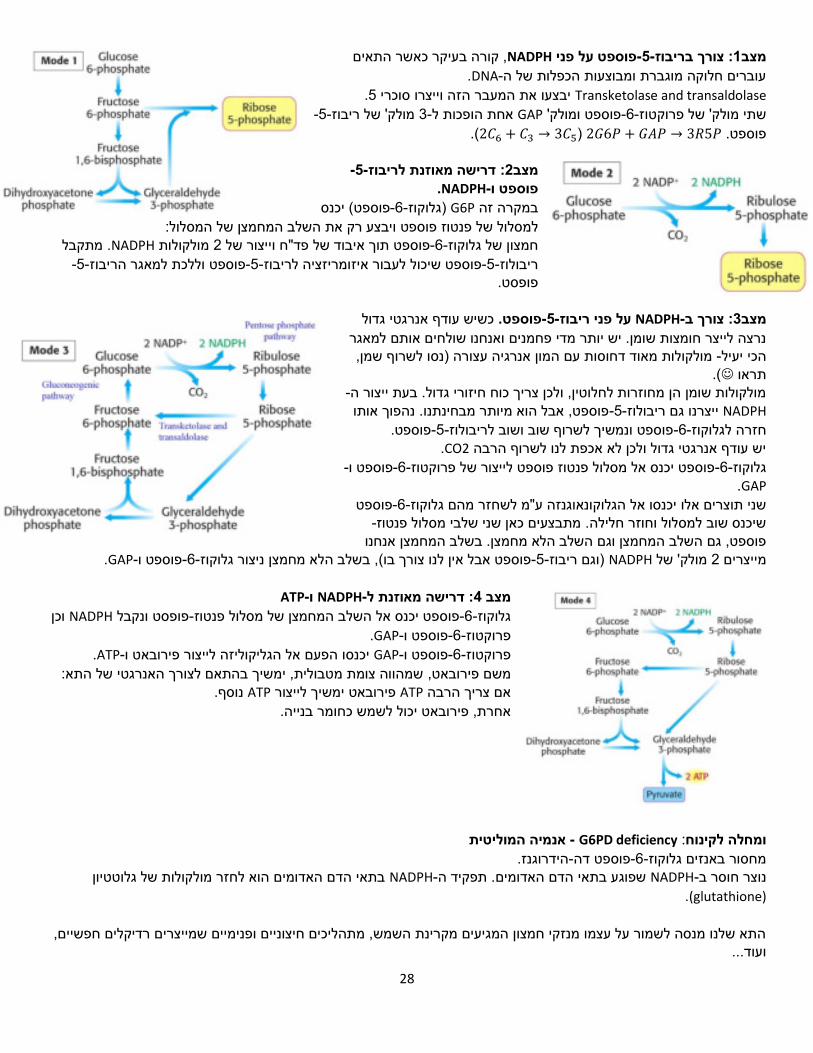
28
, קורה בעיקר כאשר התאים NADPHעל פני פוספט -5-צורך בריבוז: 1מצב .DNA-עוברים חלוקה מוגברת ומבוצעות הכפלות של ה
Transketolase and transaldolase 5יבצעו את המעבר הזה וייצרו סוכרי. -5-מולק' של ריבוז 3-אחת הופכות ל GAPפוספט ומולק' -6-שתי מולק' של פרוקטוז
2𝐺6𝑃 פוספט. + 𝐺𝐴𝑃 → 3𝑅5𝑃 (2𝐶6 + 𝐶3 → 3𝐶5).
-5-דרישה מאוזנת לריבוז: 2מצב . NADPH-פוספט ו
פוספט( יכנס -6-)גלוקוז G6Pבמקרה זה :ויבצע רק את השלב המחמצן של המסלול למסלול של פנטוז פוספט
מתקבל . NADPHמולקולות 2תוך איבוד של פד"ח וייצור של פוספט-6-חמצון של גלוקוז-5-ללכת למאגר הריבוזפוספט ו-5-לעבור איזומריזציה לריבוזפוספט שיכול -5-ריבולוז .פופסט
כשיש עודף אנרגטי גדול .פוספט-5-ריבוזעל פני NADPH-צורך ב: 3מצב
נרצה לייצר חומצות שומן. יש יותר מדי פחמנים ואנחנו שולחים אותם למאגר ף שמן, ומולקולות מאוד דחוסות עם המון אנרגיה עצורה )נסו לשר -הכי יעיל
.(תראו
-מולקולות שומן הן מחוזרות לחלוטין, ולכן צריך כוח חיזורי גדול. בעת ייצור הNADPH פוספט, אבל הוא מיותר מבחינתנו. נהפוך אותו -5-ריבולוז ייצרנו גם פוספט.-5-פוספט ונמשיך לשרוף שוב ושוב לריבולוז-6-לוקוזגחזרה ל
.CO2יש עודף אנרגטי גדול ולכן לא אכפת לנו לשרוף הרבה -פוספט ו-6-פוספט יכנס אל מסלול פנטוז פוספט לייצור של פרוקטוז-6-גלוקוז
GAP. פוספט -6-הגלוקונאוגנזה ע"מ לשחזר מהם גלוקוזשני תוצרים אלו יכנסו אל
-מתבצעים כאן שני שלבי מסלול פנטוז שיכנס שוב למסלול וחוזר חלילה.פוספט, גם השלב המחמצן וגם השלב הלא מחמצן. בשלב המחמצן אנחנו
.GAP-פוספט ו-6-בשלב הלא מחמצן ניצור גלוקוז, פוספט אבל אין לנו צורך בו(-5-)וגם ריבוז NADPHמולק' של 2מייצרים
ATP-ו NADPH-דרישה מאוזנת ל: 4מצב וכן NADPHפופסט ונקבל -פוספט יכנס אל השלב המחמצן של מסלול פנטוז-6-גלוקוז
.GAP-פוספט ו-6-פרוקטוז . ATP-אל הגליקוליזה לייצור פירובאט ויכנסו הפעם GAP-פוספט ו-6-פרוקטוז
שמהווה צומת מטבולית, ימשיך בהתאם לצורך האנרגטי של התא: משם פירובאט, נוסף. ATPפירובאט ימשיך לייצור ATPאם צריך הרבה
אחרת, פירובאט יכול לשמש כחומר בנייה.
אנמיה המוליטית - G6PD deficiency :ומחלה לקינוח
הידרוגנז.-פוספט דה-6-מחסור באנזים גלוקוזבתאי הדם האדומים הוא לחזר מולקולות של גלוטטיון NADPH-שפוגע בתאי הדם האדומים. תפקיד ה NADPH-נוצר חוסר ב
(glutathione) .
התא שלנו מנסה לשמור על עצמו מנזקי חמצון המגיעים מקרינת השמש, מתהליכים חיצוניים ופנימיים שמייצרים רדיקלים חפשיים, ועוד...

29
גליצין. הן בעלות קשר שיכול לעבור גם חמצון וגם חיזור. מולקולות אלו הן -ציסטאין-שמכילות גלוטמטמולק' גלוטטיון הן מולקולות חלק ממנגנון ההגנה של התא כנגד רדיקלים חפשיים. אם הקשר עובר חמצון האלקטרונים יעברו אל הרדיקל.
עלול להיות )קורה לאנשים, פגם גנטי די נפוץ( וגנז דהידר-פוספט-6-בהיעדר האנזים גלוקוזלכן בתאי דם אדומים אין מיטוכונדריה,
עודף של רדיקלים.
זה יכול .לא טוב – רדיקלים מפרקים קשרים ויכולים לגרום להרס של חלבונים, אזורים הידרופוביים חשופים, אגרגציותנזקי חמצון: להרוס ולפוצץ את תא הדם האדום.
.)מסיבה זו אנמיה המוליטית יחסית נפוצה באפריקה( ד מלריהפינת האופטימיות: המחלה הזו מקנה יתרון כנג
רגישות לפול.המחלה הזו מקנה פינת הלא קשור:
6( שיעור185/2/)
מטבוליזם של חומצות שומן
ליפידים או שומנים הם חומרים אורגניים שאינם מסיסים במים.: הגדרה פיסיקלית
:תפקידים מרכזיים של חומצות שומן
אבני בניין .1
קישור קוולנטי לחלבונים .2
הורמונים ומולקולות שמאפשרות קשר בין תאי .3
דלק, ייצור אנרגיה .4
𝑝𝐾𝑎הידרופובית עם קצה קרבוקסילי. (=CH)קרבונית -שרשרת הידרו: חומצות שומן מבנה = 4.8.
שהוא הידרופילי, וכך נוצר מצב אמפיפטי )גם חלק הידרופובי וגם חלק הידרופילי( -COOלעיתים קרובות קצה
היא חומצת שומן לא רוויההיא חומצת שומן שאין בה אף קשר כפול בין פחמנים )רוויה במימנים(, ובהתאם לכך חומצת שומן רוויה כזו שמכילה קשר כפול אחד לפחות בין פחמנים.
דה את טמפ' הרתיחה ומעלה את המסיסות, והוספת קשרים כפולים עושה את ההפך.הוספת קשרים כפולים מורי פחמנים מהקשר הקודם בשרשרת )אם ישנו(. 3קשרים כפולים בחומצת שומן לרוב נמצאים במצב של ציס, ויהיו במרחק של
למה חומצות שומן משמשות כחומר לשימור אנרגיה בגוף?
גם חיידקים מייצרים חומצות שומן.
הפקת אנרגיה מתרחשת בתהליך של . מאודשמדובר במולקולה מחוזרת הוא חומר שימור יתרון אחד לשימוש בחומצות שומן כ חמצון )שריפה(, ככל שחומר יותר מחוזר ניתן להעביר אותו יותר חמצונים ולהפיק ממנו יותר אלקטרונים.
לגרם, הבדל משמעותי.ק"ל ק 4ים וחלבונים יש אנרגיה של לסוכרקילו קלוריות לגרם בעוד ש 9ש אנרגיה של לחומצות שומן י
לשמר אותן.בכדי ולכן צריך פחות מים חומצות שומן הן מאוד הידרופוביותיתרון נוסף הוא שמים הם נפח אותה אנרגיה בפחות נפח ופחות מסה. גרם קליקוגן. 1פחות מים מאחסון של גרם חומצות שומן דורש 1אחסון של
ה מעדיפה אנשים עם פחות נפח ומסה, יותר קל להם לצוד ולברוח וכו'.האבולוצים שלא ניתן להפיק מהם אנרגיה. מבוזבז משו (.צידמעודדת ולא כותבת הסיכום אוהבת אנשים בכל הנפחים והמסותכך לפי המרצה, )
איך חומצות שומן נארזות?
מאפשר לארוז את חומצות השומן משום שהוא מנטרל את המטען שלהן. חומר מאוד חשוב. גליצרול ניתן לחמצן אותו, ואם נחמצן לקחנו ממנו אנרגיה וקיבלנו תוצרים של פירוק גליקוליזה מהם ניתן ליצור פנטוזות, הקסוזות ועוד.

30
שלוש חומצות שומן.ל המחוברת בקשר אסתרי ליצרולהשומן הפשוט ביותר, והוא מורכב ממולקולה אחת של גהוא טריגליצרול נקרא גם טריאציל גליצרול או טריגליצריד.
לגליצרול ושלוש חומצות שומן. טריגליצרידיםיודע לפרק ולהרכיב את המבנים הללו, את ה אנזים ליפאזה
מאיפה מגיע השומן?
מגיע מהמזוןרובו .1
בגוף: חומצה לינולאית וחומצה לינולנית.שתי חומצות שומן חיוניות שלא ניתן לסנתז
זמינות של אנרגיה בצמוד לאיברים שצורכים אנרגיה –מאגרי שומן בגוף, בתוך תאי שומן, בעיקר ליד השרירים .2
ייצור של שומן .3
עיכול שומן מהמזון
שומן לפיסות קטנות יותר. מיסים את השומנים הנצרכים במזון, מפרידים גושים של מ, מכילים נגזרות של כולסטרול מיצי מרה .1
)קצת כמו פעולה של סבון(. לגליצרול וחומצות שומןטריגליצרידים את ה יםמפרקליפזות יעודיים לכך .2
הדק, שם הם יארזו מחדש לטריגליצרידים נספגים במעיוגליצרול חומצות שומן .3 .Chylomicronsליצירת ApoC2-ו כולסטרול ואריזה נוספת מבוצעת על ידי אריזה חוזרת בטריגליצרידים,בתוך המעי: .4
5. Chylomicrons ומשם אל הרקמות השונותאל זרם הדם דרך נוזל הלימפה יוצאים
6. Lipoprotein lipase מאוקטב על ידיApoC2 ומשחרר שוב את המבנה של הטריגליצרידים לגליצרול וחומצות שומן
חומצות השומן נכנסות אל התאים .7
לבניין התא או מחומצנות להפקת דלקחומצות שומן נאגרות, משמשות .8
שימו לב שהתהליך של אריזה ופירוק חוזר על עצמו שוב ושוב במהלך העברת חומצות השומן בתוך הגוף.
מתי הכבד יפריש חומצות שומן?
(ן/אדרנלין בשריר ובכבד, גלוקגון בכבד בלבדסיגנל הורמונלי נקלט ומשפעל את אדנילאט ציקלאז )אפינפרי .1
cAMPיצור .2
PKAאקטיבציה של .3
4. PKA ליפז -גליצרול-מזרחן את האנזים צריאציל(triacylglycerol lipase) וגורם לו לפרק טריגליצרידים לגליצרול וחומצות שומן
הגליצרול נשאר בכבד והולך לגלוקונאוגנזה .5
חומצות השומן יופרשו אל זרם הדם )עם חלבון נשא שנקרא סרום אלבומין( .6
רת של כולסטרול, הידרופובי. נכנס בתוך גושי השומן ויוצר פלואידיות, מפר מעט את הסדר כדי שאנזימים יוכלו הוא נגז גליקולט להיכנס ולעבוד.
ברוב העיקרי הליפידי המרכיב הם פוספוגליצרידים. גליצרול עשוי שלד בעל פוספוליפידטריגליצריד, הוא גליצרופוספוליפיד
.הביולוגיות הממברנות
Adipose cell –תאי שומן תפקידם העיקרי הוא לאגור שומן. השומן מגיע כחומצות שומן מזרם הדם.תאים אדיפוציטים.
, וכן (גליצרול-מפרקים טריאצילוגם גליצרול-יוצרים טריאציל)גם ליפזות שמזרזים הידרוליזה של טריגליצרידיםהם מכילים ומתי לקלוט שומן.רצפטורים שונים שאומרים לתא מתי לשחרר
בתוך תאי השומן, השומנים הטבעיים מאוחסנים כטיפות של טריגליצרידים מוקפות פוספוליפידים, על שטח הפנים של הטיפות יש חלבון המגביל גישה אל חומצות השומן. -פריליפין
מה קורה כשחומצות שומן יוצאות מתא שומן?
המתאים המאותת לתא השומן כי יש צורך בשחרור שומנים. הטריגליצרידים יובלו רמונים שיעבירו את הסיגנל ראשית, יש צורך בהו אז אל הרקמות השונות.
אדנילאטציקלאזמשפעלים את ואפינפרין גלוקוגןההורמונים .7

31
cAMPייצור .8
PKAאקטוב .9
Aפריליפין פוספורילציה ל .10
חומצות שומן וגליצרולים ללגשת לפני השטח של הטיפות ולבצע הידרוליזה של טריגליצריד ליפזותגורם ל .11
חומצות שומן משתחררות אל זרם הדם .12
שיעשה שאטל לחומצות השומן אל תאי השריר. Serum Albuminסרום אלבומין נקשרות אל חומצות השומן .13
חיוביים אליהם הן נקשרות כך שכל שרשרת -חומצות שומן, יש בו אזורים אלקטרו 10כל מונומר של החלבון יכול לקשור במקום אחר כך שבהמשך הן יהיו עדיין קלות מאוד להמסה ולשימוש מיידי בתאים. מעוגנת
אסור שהוא יהיה במבנה שעובר אגרגציה במגע עם חומצות שומן )אם זה קורה בדם אנחנו עלולים למות מזה די מהר.. לא ן אזור הידרופובי חשוף וגדול שעלול (. אז מה מונע את האגרגציה? רוב החלבון הוא אלקטרו שלילי, מה שיוצר דחייה. אימומלץ
ליצור אגרגציה. מהאנרגיה שמופקת מהטריגליצריד( 5%)רק פוספט וילך לגליקוליזה, כך תופק אנרגיה גם ממנו-3-יהפוך לגליצרול גליצרול .14
כעת יש חומצות שומן בתוך השריר. מה עושים איתן?
של החומצות ההידרופוביות. י ממברנות, כלומר שוב יש מעברחומצת השומן תיכנס אל המיטוכונדריהפחמנים שורפים 2פחמנים להפקת אנרגיה. אילו 2פחמנים, וכל פעם אנחנו שורפים 2חומצות שומן נשברות לחלקים של
.CO2ונשרף והופך להיות TCA-שנכנס אל מעגל ה Aבמיטוכונדריה? אצטיל קו אילו חומצות שומן הולכות להפקת אנרגיה ואילו מהן הולכות לבניית שיהיה ברור -חומצות השומן צריכות להיות מאוקטבות
ממברנות למשל.
פירוק חומצות שומן
של חומצת השומן אקטיבציה -שלב ראשון סינטתז.-Aקו-קורה בציטופלסמה על ידי האנזים אציל
, דרך אציל, זה קשר עתיר אנרגיה. Oמייצרים קשר דרך .PPiמשתחרר
HS-CoA תוקף ומחליף את הקשר עתיר האנרגיה בקשר עתיר אנרגיה אחר.
נעה קדימה מכוח האנטרופיה, מעלים את מספר הריאקציה שלילי. 𝐺∆המולקולות בתא,
PPi מתפרק אח"כ לשני פוספטים ומצטרף אל מאגר הפוספטים בתא.
שנמצאות בציטופלסמה.התוצאה של השלב הראשון היא חומצות שומן מאוקטבות )מוכנות לשריפה(
הכנסה אל המיטוכונדריה -שלב שני
מחליף את האצטיל אציל קרניטין הבעיה היא שלא ניתן להעביר קואנזים את הממברנה. פעיל בתגובה זו. 1טרנספראז-אציל-האנזים קרניטין .A-קו
שלו הוא , בעל צד אחד שלילי וצד אחד חיובי, כלומר המטען הכללי zwitterionקרניטין הוא אפס והוא יכול לעבור דרך הממברנה מבלי להשפיע על פוטנציאל הממברנה.
פחמנים יכולות לעבור את הממברנה עצמאית, וחומצות שומן 12חומצות באורך של עד
, אלו הן רוב חומצות (Carnitine Shuttle) מעבורת קרניטיןפחמנים ויותר יעזרו ב 14בנות השומן.
Fatty acyl-CoA יכולה להיכנס לתוך המיטוכונדריה או לשמש לסינתזת ממברנות. אם
חומצת השומן מיועדת לעבור חמצון במיטוכונדריה יתרחש שלב נוסף של מעבורת קרניטין: .הפנימית של המיטוכונדריהריאקציה זו מתרחשת על גבי הממברנה
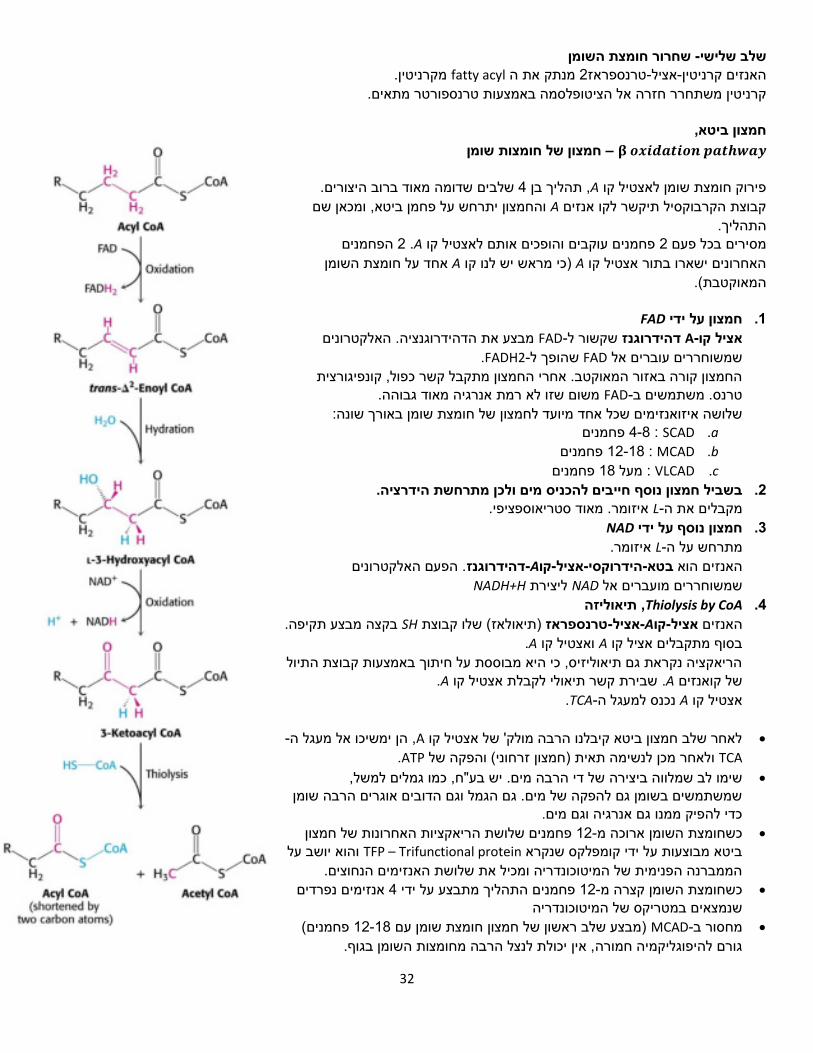
32
שחרור חומצת השומן -שלב שלישי מקרניטין. fatty acylמנתק את ה 2טרנספראז-אציל-קרניטיןהאנזים
אל הציטופלסמה באמצעות טרנספורטר מתאים. חזרהקרניטין משתחרר
חמצון של חומצות שומן – 𝛃 𝒐𝒙𝒊𝒅𝒂𝒕𝒊𝒐𝒏 𝒑𝒂𝒕𝒉𝒘𝒂𝒚 ,חמצון ביטא
שלבים שדומה מאוד ברוב היצורים. 4, תהליך בן Aפירוק חומצת שומן לאצטיל קו והחמצון יתרחש על פחמן ביטא, ומכאן שם Aקבוצת הקרבוקסיל תיקשר לקו אנזים
התהליך.הפחמנים A. 2פחמנים עוקבים והופכים אותם לאצטיל קו 2 בכל פעם מסירים
אחד על חומצת השומן A)כי מראש יש לנו קו Aהאחרונים ישארו בתור אצטיל קו .המאוקטבת(
FADעל ידי חמצון .1
מבצע את הדהידרוגנציה. האלקטרונים FAD-שקשור ל דהידרוגנז A-אציל קו .FADH2-שהופך ל FADשמשוחררים עוברים אל
החמצון קורה באזור המאוקטב. אחרי החמצון מתקבל קשר כפול, קונפיגורצית משום שזו לא רמת אנרגיה מאוד גבוהה. FAD-טרנס. משתמשים ב
של חומצת שומן באורך שונה:שלושה איזואנזימים שכל אחד מיועד לחמצון a. SCAD :4-8 פחמנים
b. MCAD :12-18 פחמנים
c. VLCAD פחמנים 18: מעל
בשביל חמצון נוסף חייבים להכניס מים ולכן מתרחשת הידרציה. .2
איזומר. מאוד סטריאוספציפי. L-מקבלים את ה
NADעל ידי חמצון נוסף .3
איזומר. L-מתרחש על ה. הפעם האלקטרונים דהידרוגנז-Aקו-אציל-הידרוקסי-בטאהאנזים הוא
NADH+Hליצירת NADשמשוחררים מועברים אל 4. Thiolysis by CoAתיאוליזה ,
בקצה מבצע תקיפה. SHשלו קבוצת )תיאולאז( טרנספראז-אציל-Aקו-אצילאנזים ה
.Aואצטיל קו Aים אציל קו בסוף מתקבלהריאקציה נקראת גם תיאוליזיס, כי היא מבוססת על חיתוך באמצעות קבוצת התיול
.Aשבירת קשר תיאולי לקבלת אצטיל קו .Aשל קואנזים .TCA-נכנס למעגל ה Aאצטיל קו
לאחר שלב חמצון ביטא קיבלנו הרבה מולק' של אצטיל קוAהן ימשיכו אל מעגל ה ,-
TCA ולאחר מכן לנשימה תאית )חמצון זרחוני( והפקה שלATP. של די הרבה מים. יש בע"ח, כמו גמלים למשל, ביצירהשימו לב שמלווה
שמשתמשים בשומן גם להפקה של מים. גם הגמל וגם הדובים אוגרים הרבה שומן כדי להפיק ממנו גם אנרגיה וגם מים.
ריאקציות האחרונות של חמצון פחמנים שלושת ה 12-כשחומצת השומן ארוכה מוהוא יושב על TFP – Trifunctional proteinביטא מבוצעות על ידי קומפלקס שנקרא
הממברנה הפנימית של המיטוכונדריה ומכיל את שלושת האנזימים הנחוצים.
אנזימים נפרדים 4פחמנים התהליך מתבצע על ידי 12-כשחומצת השומן קצרה מ יטוכונדריהשנמצאים במטריקס של המ
מחסור ב-MCAD פחמנים( 12-18)מבצע שלב ראשון של חמצון חומצת שומן עם , אין יכולת לנצל הרבה מחומצות השומן בגוף.גורם להיפוגליקמיה חמורה

33
חומצות שומן ארוכות במיוחד עוברות חמצון בפרוקסיזום
מצב ציסשנמצא ב אם יש קשר כפולא טרנס, ולכן אם חומצת השומן בלתי רוויה ומכילה קשר כפול כלשהו סביר שהיא תכיל רוב הקשרים הכפולים הם במצב ציס ול
אז כשנגיע אל הקשר הבלתי רוויכי הוא פועל רק על קשרי טרנס, האנזים שמבצע את ההידרציה לא יוכל לפעול קשר מסוג ציס.. זה קורה קשרים כפולים עוקבים או יותר( 2 ( ורדוקטאז )אם ישמעביר קשר ממצב ציס למצב טרנס)נצטרך להשתמש באיזומראז
לפני ההידרציה.
מה קורה עם חומצות שומן לא זוגיות?. לא (פחמנים 3) Aניל קו אופרופיפחמנים( ו 2אחד ) Aפחמנים. במקרה כזה נקבל אצטיל קו 5הכל כרגיל עד שמגיעים למצב של
זה שנוכל לחתוך לשני זוגות. Aפחמנים ואחרי כמה שלבים, מקבלים סוקסיניל קו 4מוסיפים פחמן למצב של נעמיק בנושא זה. מהפירוק כך שזה שווה את זה. ATPדורש השקעת אנרגיה, אבל נרוויח מספיק
קצת סטויכיומטריה
βפעמים 7פחמנים. נבצע 16מוצג חישוב עבור חומצה בעלת oxidation פעמים 7ולכן נקבלFADH2 פעמים 7-וNADH + H+.
פחמנים(. ועכשיו נספור 16)כי Aפעמים אצטיל קו 8נקבל בסה"כ ATP לכל :FAD לכל 1.5נקבל ,NADH לכל אצטיל קו 2.5נקבל ,A
במעגל קרבס. 10נקבל
שניצלנו ממש בהתחלה. ATP-ה 2בסוף, אלו 2-חשוב להוריד את ה פחמנים. 16עבור חומצת שומן אחת בעלת ATPמולק' 106בסה"כ:
גופי קטון
, אלו חומרים מאוד גופי קטוןהמופק בכבד יכול לעבור המרה ל Aאצטיל קו
מסיסים בדם ובשתן. הכבד ממיר חומצות שומן לאנרגיה מסיסה, בניגוד לחומצת שומן שהיא אנרגיה לא מסיסה.
, תהליך לא הפיך. אצטוןכשהגוף שורף חומצות שומן מתקבלת נשימה של אצטון הוא היחיד מבין גופי הקטון שננשף החוצה.
34
אצטאט -שיחומצן ליצירת אנרגיה. אצטו Aמועברות בדם לרקמות שונות וממורות חזרה לאצטיל קו ט והידרוקסיבוטירטאצט-אצטו היא מולקולה מסיסה מאוד וחומצית מאוד שיכולה לנדוד בדם.
ומולק' מים A. לאחר מכן מוסיפים מולקולה שלישית של אצטיל קו Aקו-אצטיל-יכולים לעבור דחיסה ונקבל אצטו Aקו-אצטיל 2
אצטט יכולה להפוך לאצטון או -אצטו אצטט.-אצטות מולק' של אחת ומתקבל A. לאחר מוכן מוסרת קב' אצטיל קו HMG-CoAלקבלת להידרוקסיבוטירט.
פירוק גופי קטון ברקמות )יש פירוט על התהליך בספר(. והפקת אנרגיה. פירוק ים, ברקמות השונות מתרחשייצור גופי קטון מתרחש רק בכבד
של חולי סכרת, עלולים להגיע במקרה
שהופך Aלמצב שמצטבר אצטיל קו לגופי קטון. קצב הייצור של גופי הקטון
גבוה מקצב ניצולם והדבר גורם , תופעה של הדם pH-לירידת המשום שגופי ,אצידוזיסהנקראת
במצב קיצוני הדבר הקטון הם חומצה. יוביל למוות.
פירוק אל מול סינתזה: חומצות שומן
סינתזה פירוק ציטופלסמה )כי שם מייצרים דברים( דברים( במיטוכונדריה )כי שם שורפים
CoA is acyl carrier ACP is acyl carrier
L-β-hydroxyacyl group D-β-hydroxyacyl group Aפחמנים ממלוניל קו 2מתקבלים CoA-פחמנים ו A ,2אצטיל קו מתקבלים
NAD+ ו-FAD משמשים לביצוע חמצון NADPH תורם אלקטרונים אנזימים שנמצאים על אותו פוליפפטיד אנזימיםהפרדה בין
אנזימים שונים וכן בקרה שונה לכל אחד מהתהליכים
סינתזת חומצות שומן
הטבלה מפרטת את שלבי הסינתזה, אין צורך לזכור בפירוט. .ATPתוך השקעת Aמאצטיל קו Aמלוניל קו חשוב להכיר את השלב הראשון בו נוצר
והוא הנקודה המחייבת, והוא עובר בקרה מאוד פחמניםA (3 )למלוניל קו פחמניםA (2 )הופך אצטיל קו קרבוקסילז Aאצטיל קו ילך לחמצון )פירוק( או לשימור )סינתזה(. fatty acyl CoAגדולה. פעילותו של אנזים זה קובעת אם
.NADH מתקבל( ביטא חמצון) שומן חומצות של ובפירוק NADPH-ב משתמשים שומן חומצות של שבסינתזה לב שימו**

35
:הוא מעגלי השלב השני .ACPאל הנשא Aמועבר מהנשא קו Aמלוניל קו .ACPאל הנשא Aמועבר מהנשא קו Aאצטיל קו
פחמנים. 2קרבוקסילציה כדי להפוך למולק' של -עובר דה ACPמלוניל פחמנים(. 4)מולק' בעלת ACPאצטואצטיל נדחסים יחד ליצירת ACPאצטיל )אחרי קרבוקסילציה( + ACPמלוניל : דחיסה .1
.fatty acid synthaseזה קורה ע"י האנזים ACP-הידרוקסיבוטיריל-D-3לקבלת NADPH ע"י חיזור .2
: דחיסה החוצה של מולק' מיםדהידרציה .3
NADPHנסוף ע"י חיזור .4
ACPבוטיריל קיבלנו .5
נוספת. ACPהמעגל חוזר חלילה עם דחיסה של מולק' מלוניל לבסוף קבלת חומצת שומן.
בין המיטוכונדריה והציטוזול Aהעברת אצטיל קו
אל הציטופלסמה כדי לייצר שם חומצות שומן. Aצריך להביא אצטיל קו ואוקסלואצטט נדחסים יחד לציטרט )בשלב Aבמיטוכונדריה אצטיל קו
(. ציטרט יכול לחצות את הממברנה.TCA-הראשון של מעגל ה ואוקסלואצטט. Aבציטוזול, ציטרט יכול להתפרק לאצטיל קו )בדיוק ההפך ממה NADPHאוקסלואצטט יכול להפוך למלט תוך ניצול
.(TCA-שקורה בפירובאט יכול גם הוא , וNADPHמלט להפוך לפירובאט תוך פליטה של
לחצות את הממברנה.
בקרה על סינתזת חומצות שומן ריכוז גבוה של אוקסלואצטט במיטוכונדריה יכול לגרום לכך שנעבור מפירוק )דגרדציה( לסינתזה של חומצות שומן.
יה.מספר רב של פעמים, כלומר יש הרבה אנרג TCA-ריכוז גבוה של אוקסלואצטט משמעו שהפעלנו את מעגל ה
קרבוקסילז Aאצטיל קו , מולקולה בעלת שלושה פחמנים.Aמלוניל קווקרבונט ותוך השקעת אנרגיה יוצר Aלוקח אצטיל קו
)קבוצה שנמצאת בקצה של ליזין(. ביוטיןקרבונט עובר אקטיבציה על ידי יון
36
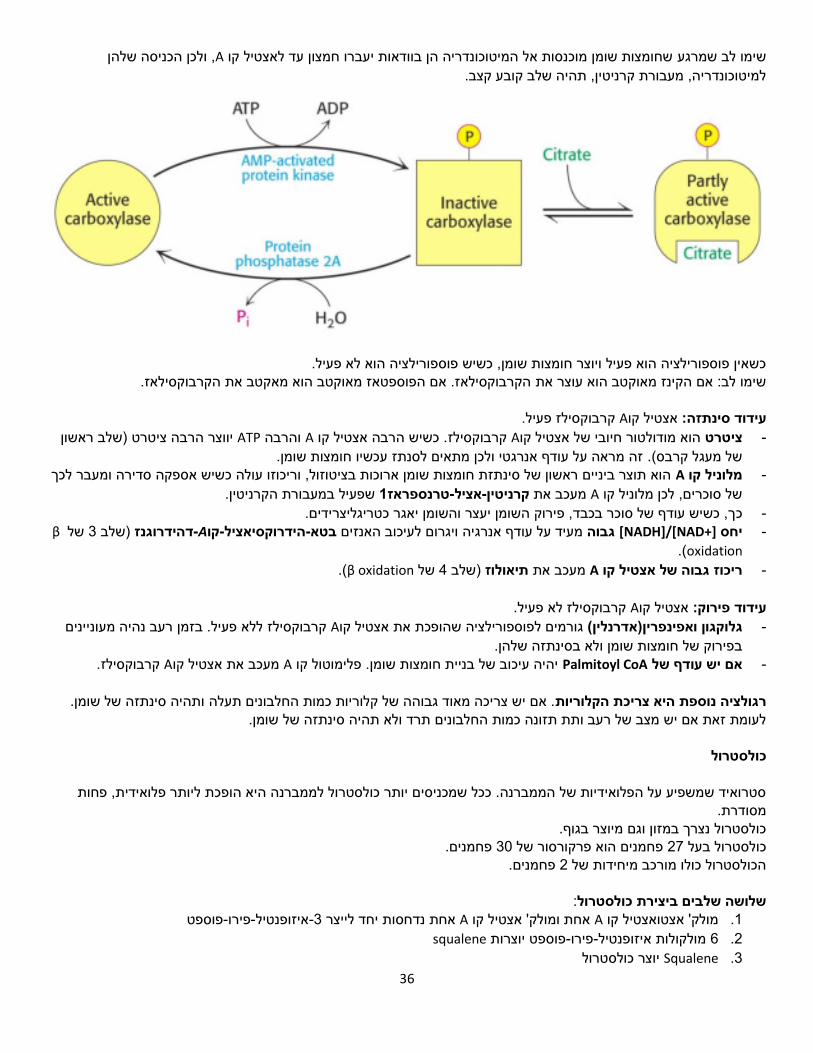
, ולכן הכניסה שלהן Aשחומצות שומן מוכנסות אל המיטוכונדריה הן בוודאות יעברו חמצון עד לאצטיל קו שימו לב שמרגע למיטוכונדריה, מעבורת קרניטין, תהיה שלב קובע קצב.
כשאין פוספורילציה הוא פעיל ויוצר חומצות שומן, כשיש פוספורילציה הוא לא פעיל.
הקרבוקסילאז. אם הפוספטאז מאוקטב הוא מאקטב את הקרבוקסילאז.שימו לב: אם הקינז מאוקטב הוא עוצר את
פעיל. קרבוקסילז Aאצטיל קועידוד סינתזה: )שלב ראשון ציטרטיווצר הרבה ATPוהרבה A. כשיש הרבה אצטיל קו קרבוקסילז Aאצטיל קוהוא מודולטור חיובי של ציטרט -
זה מראה על עודף אנרגטי ולכן מתאים לסנתז עכשיו חומצות שומן. .של מעגל קרבס(הוא תוצר ביניים ראשון של סינתזת חומצות שומן ארוכות בציטוזול, וריכוזו עולה כשיש אספקה סדירה ומעבר לכך Aמלוניל קו -
שפעיל במעבורת הקרניטין. 1טרנספראז-אציל-קרניטיןמעכב את Aשל סוכרים, לכן מלוניל קו ך, כשיש עודף של סוכר בכבד, פירוק השומן יעצר והשומן יאגר כטריגליצרידים.כ - βשל 3)שלב דהידרוגנז-Aקו-הידרוקסיאציל-בטאמעיד על עודף אנרגיה ויגרום לעיכוב האנזים גבוה [+NAD]/[NADH]יחס -
oxidation.)
(.β oxidationשל 4)שלב תיאולוזמעכב את Aריכוז גבוה של אצטיל קו -
לא פעיל. קרבוקסילז Aאצטיל קועידוד פירוק:
ללא פעיל. בזמן רעב נהיה מעוניינים קרבוקסילז Aאצטיל קוגורמים לפוספורילציה שהופכת את )אדרנלין(ן ואפינפריןגלוקגו - בפירוק של חומצות שומן ולא בסינתזה שלהן.
. קרבוקסילז Aאצטיל קומעכב את Aיהיה עיכוב של בניית חומצות שומן. פלימוטול קו Palmitoyl CoAאם יש עודף של -
. ותהיה סינתזה של שומן. אם יש צריכה מאוד גבוהה של קלוריות כמות החלבונים תעלה צריכת הקלוריותרגולציה נוספת היא .סינתזה של שומןולא תהיה לעומת זאת אם יש מצב של רעב ותת תזונה כמות החלבונים תרד
כולסטרול
כת ליותר פלואידית, פחות סטרואיד שמשפיע על הפלואידיות של הממברנה. ככל שמכניסים יותר כולסטרול לממברנה היא הופ
מסודרת. כולסטרול נצרך במזון וגם מיוצר בגוף.
פחמנים. 30פחמנים הוא פרקורסור של 27כולסטרול בעל פחמנים. 2הכולסטרול כולו מורכב מיחידות של
:שלושה שלבים ביצירת כולסטרול
פוספט-פירו-איזופנטיל-3לייצר ות יחדנדחסאחת Aאחת ומולק' אצטיל קו Aמולק' אצטואצטיל קו .1
squaleneפוספט יוצרות -פירו-מולקולות איזופנטיל 6 .2
3. Squalene יוצר כולסטרול
37
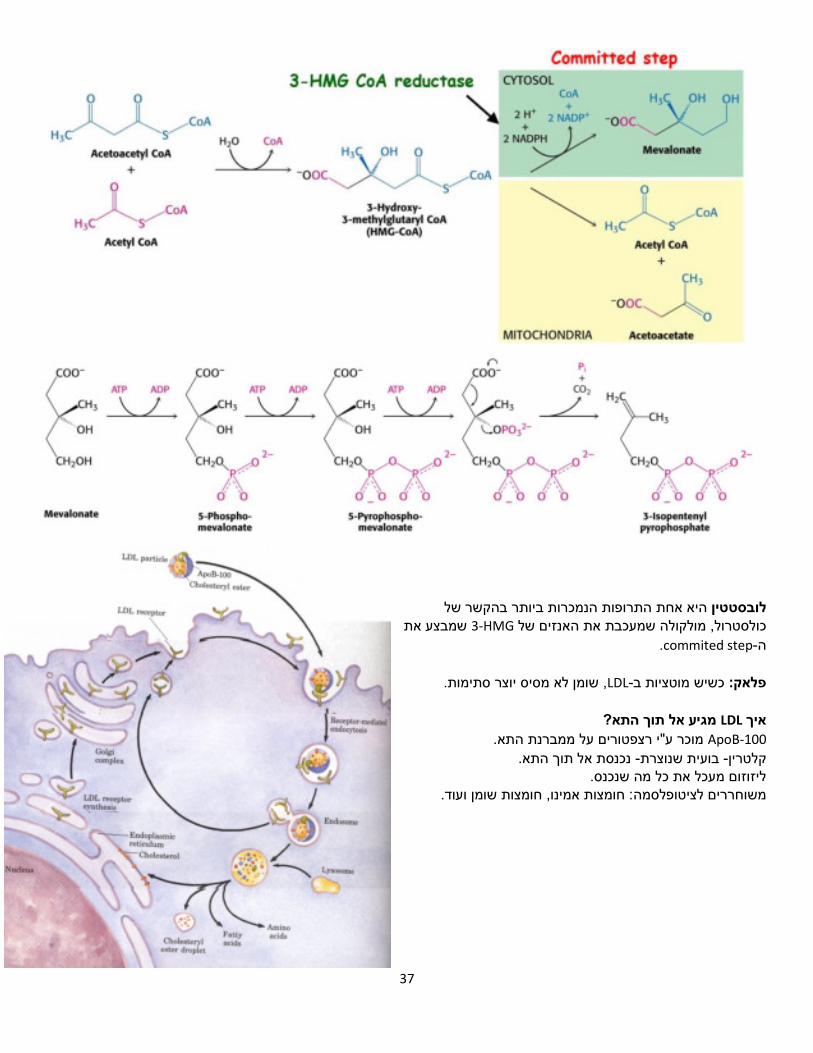
התרופות הנמכרות ביותר בהקשר של היא אחת טיןלובסטשמבצע את HMG-3את האנזים של מולקולה שמעכבתכולסטרול,
.commited step-ה
, שומן לא מסיס יוצר סתימות.LDL-ב כשיש מוטציות פלאק:
מגיע אל תוך התא? LDLאיך ApoB-100 .מוכר ע"י רצפטורים על ממברנת התא
ך התא.נכנסת אל תו -בועית שנוצרת -קלטרין ליזוזום מעכל את כל מה שנכנס.
: חומצות אמינו, חומצות שומן ועוד.משוחררים לציטופלסמה

38
7( שיעור185/9/)
מטבוליזם של גליקוגן
גליקוגן הוא פולימר מסועף של גלוקוז. חומר תשמורת בבע"ח.
הגליקוגן מאוחסן בתא בגרנולות )העיגולים השחורים האלה שרואים בצילומים(.
סינתזה של גליקוגן
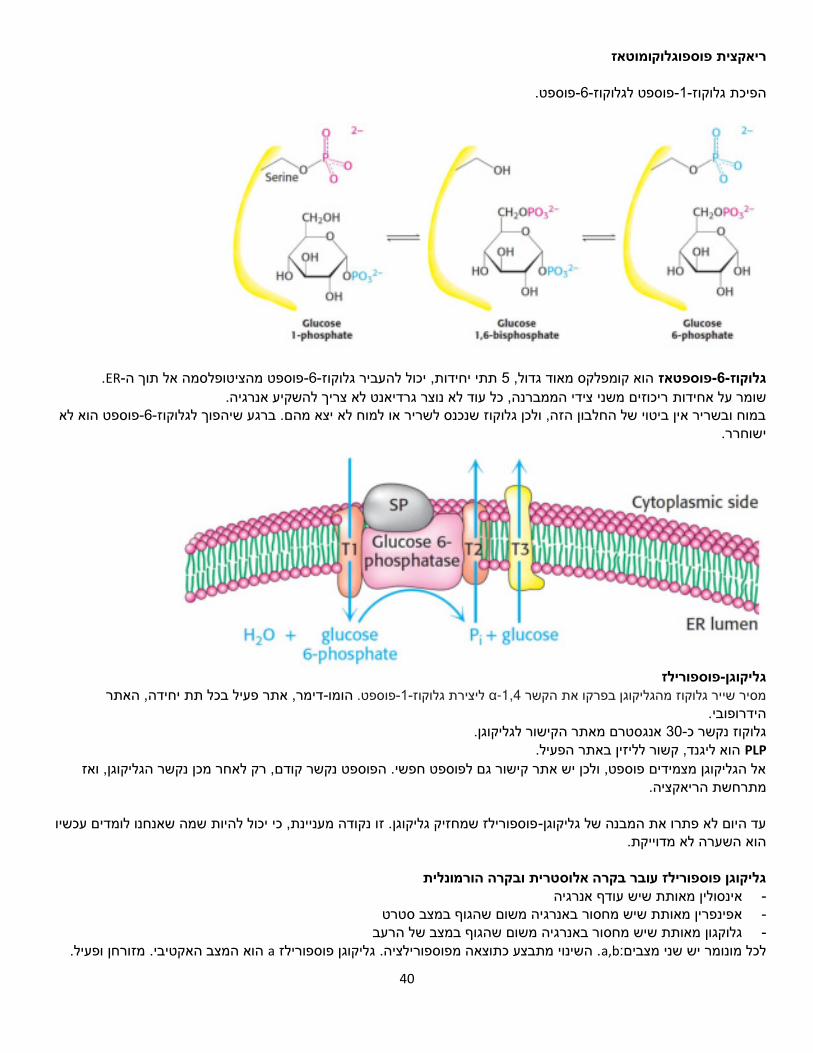
הפוספט מטרתו כרגיל להשאיר את הגלוקוז בתא, וזהו קשר עתיר , זו נקודת המוצא.פוספט-1-גלוקוזממאגר של מתחילים אנרגיה.
UTP היא מקבילה שלATP פוספט עם -6-מבחינה אנרגטית. מאקטבים את גלוקוזUTP .וכך המולק' הזו מיועדת לסינתזה .(PPi) ומקבלים פירופוספט גלוקוז-UDPמקבלים
UDP-מונומרים. 4, שיכול להוסיף אותו רק לשרשרת שכבר יש בה סינטז-גליקוגן נקשר אל גלוקוז
היחידות הללו קשורות 8יחידות גלוקוז ליצירת פריימר. 8סוכר שאליו אנחנו מוסיפים -חלבון המכיל דיאנזים די גדול, הוא גליקוגניןנוקלאציה: נקודת גירעון, יצירת התחלה קטנה בבת אחת יתרחשו שתי נוקלאציות. -. החלבון עובד כדימרטירוזיןבקשר קוולנטי ל
שממנה תצמח שאר המולקולה. יצירת גרעין ראשוני.רכנו גליקוגנין בונה על עצמו את הפריימר ונשאר קשור אליו, כלומר בסוף כל הגליקוגן עדיין קשור לגליקוגנין )ללא קשר לכמה הא
כך קיבלנו מולק' גליקוגן מסועפת כאשר במרכזה מולק' של גליקוגנין. אותו(.
39
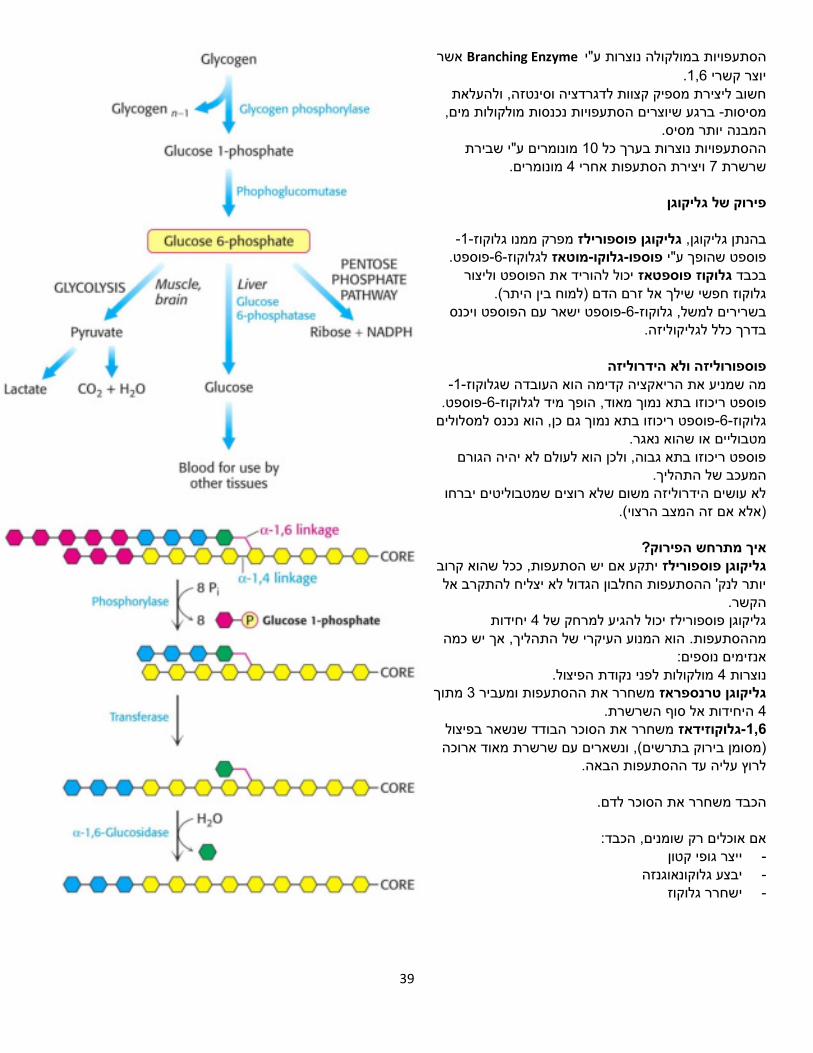
אשר Branching Enzymeע"יבמולקולה נוצרות הסתעפויות .1,6יוצר קשרי
חשוב ליצירת מספיק קצוות לדגרדציה וסינטזה, ולהעלאת ברגע שיוצרים הסתעפויות נכנסות מולקולות מים, -מסיסות
מסיס. המבנה יותרמונומרים ע"י שבירת 10בערך כל ההסתעפויות נוצרות
מונומרים. 4ויצירת הסתעפות אחרי 7שרשרת
פירוק של גליקוגן
-1-מפרק ממנו גלוקוז גליקוגן פוספורילזבהנתן גליקוגן, פוספט.-6-לגלוקוזמוטאז -גלוקו-פוספופוספט שהופך ע"י
יכול להוריד את הפוספט וליצור גלוקוז פוספטאזבכבד (.בין היתר אל זרם הדם )למוחגלוקוז חפשי שילך
פוספט ישאר עם הפוספט ויכנס -6-בשרירים למשל, גלוקוז בדרך כלל לגליקוליזה.
פוספורוליזה ולא הידרוליזה
-1-מה שמניע את הריאקציה קדימה הוא העובדה שגלוקוז פוספט.-6-ד לגלוקוז, הופך מינמוך מאודפוספט ריכוזו בתא
פוספט ריכוזו בתא נמוך גם כן, הוא נכנס למסלולים -6-גלוקוז מטבוליים או שהוא נאגר.
פוספט ריכוזו בתא גבוה, ולכן הוא לעולם לא יהיה הגורם המעכב של התהליך.
לא עושים הידרוליזה משום שלא רוצים שמטבוליטים יברחו )אלא אם זה המצב הרצוי(.
הפירוק?איך מתרחש
יתקע אם יש הסתעפות, ככל שהוא קרוב גליקוגן פוספורילזיותר לנק' ההסתעפות החלבון הגדול לא יצליח להתקרב אל
הקשר.יחידות 4גליקוגן פוספורילז יכול להגיע למרחק של
הוא המנוע העיקרי של התהליך, אך יש כמה מההסתעפות. אנזימים נוספים:
ת הפיצול.מולקולות לפני נקוד 4נוצרות מתוך 3משחרר את ההסתעפות ומעביר גליקוגן טרנספראז
היחידות אל סוף השרשרת. 4 הבודד שנשאר בפיצולמשחרר את הסוכר גלוקוזידאז-1,6
, ונשארים עם שרשרת מאוד ארוכה )מסומן בירוק בתרשים( לרוץ עליה עד ההסתעפות הבאה.
הכבד משחרר את הסוכר לדם.
, הכבד: שומניםאם אוכלים רק
ייצר גופי קטון -
יבצע גלוקונאוגנזה -
ישחרר גלוקוז -
40
ריאקצית פוספוגלוקומוטאז
פוספט. -6-פוספט לגלוקוז-1-הפיכת גלוקוז
.ER-פוספט מהציטופלסמה אל תוך ה-6-תתי יחידות, יכול להעביר גלוקוז 5הוא קומפלקס מאוד גדול, פוספטאז-6-גלוקוז ריכוזים משני צידי הממברנה, כל עוד לא נוצר גרדיאנט לא צריך להשקיע אנרגיה.שומר על אחידות
פוספט הוא לא -6-במוח ובשריר אין ביטוי של החלבון הזה, ולכן גלוקוז שנכנס לשריר או למוח לא יצא מהם. ברגע שיהפוך לגלוקוז ישוחרר.
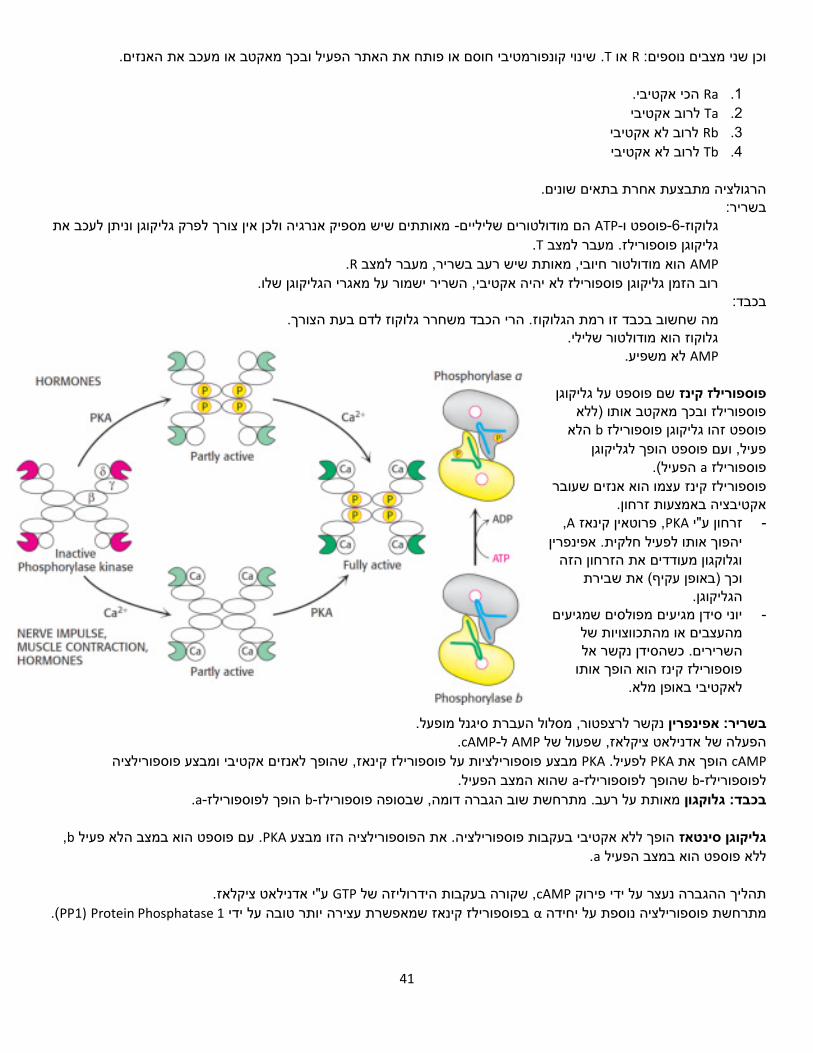
פוספורילז-גליקוגן
דימר, אתר פעיל בכל תת יחידה, האתר -הומו פוספט.-1-ליצירת גלוקוז α- 1,4הקשרמסיר שייר גלוקוז מהגליקוגן בפרקו את הידרופובי.
אנגסטרם מאתר הקישור לגליקוגן. 30-גלוקוז נקשר כPLP .הוא ליגנד, קשור לליזין באתר הפעיל
ר מכן נקשר הגליקוגן, ואז אל הגליקוגן מצמידים פוספט, ולכן יש אתר קישור גם לפוספט חפשי. הפוספט נקשר קודם, רק לאח מתרחשת הריאקציה.
פוספורילז שמחזיק גליקוגן. זו נקודה מעניינת, כי יכול להיות שמה שאנחנו לומדים עכשיו -עד היום לא פתרו את המבנה של גליקוגן
הוא השערה לא מדוייקת.
גליקוגן פוספורילז עובר בקרה אלוסטרית ובקרה הורמונלית עודף אנרגיהאינסולין מאותת שיש -
אפינפרין מאותת שיש מחסור באנרגיה משום שהגוף במצב סטרט -
גלוקגון מאותת שיש מחסור באנרגיה משום שהגוף במצב של הרעב -
מזורחן ופעיל. הוא המצב האקטיבי. aגליקוגן פוספורילז . השינוי מתבצע כתוצאה מפוספורילציה.a,bלכל מונומר יש שני מצבים:
41
שינוי קונפורמטיבי חוסם או פותח את האתר הפעיל ובכך מאקטב או מעכב את האנזים.. Tאו Rוכן שני מצבים נוספים:
1. Ra .הכי אקטיבי 2. Ta לרוב אקטיבי
3. Rb לרוב לא אקטיבי 4. Tb לא אקטיבילרוב
הרגולציה מתבצעת אחרת בתאים שונים.
בשריר:מאותתים שיש מספיק אנרגיה ולכן אין צורך לפרק גליקוגן וניתן לעכב את -הם מודולטורים שליליים ATP-פוספט ו-6-גלוקוז
.Tמעבר למצב גליקוגן פוספורילז.AMP הוא מודולטור חיובי, מאותת שיש רעב בשריר, מעבר למצבR.
, השריר ישמור על מאגרי הגליקוגן שלו.רוב הזמן גליקוגן פוספורילז לא יהיה אקטיבי :בכבד
זו רמת הגלוקוז. הרי הכבד משחרר גלוקוז לדם בעת הצורך. מה שחשוב בכבד גלוקוז הוא מודולטור שלילי.
AMP .לא משפיע
שם פוספט על גליקוגן פוספורילז קינזפוספורילז ובכך מאקטב אותו )ללא
הלא bפוספט זהו גליקוגן פוספורילז פעיל, ועם פוספט הופך לגליקוגן
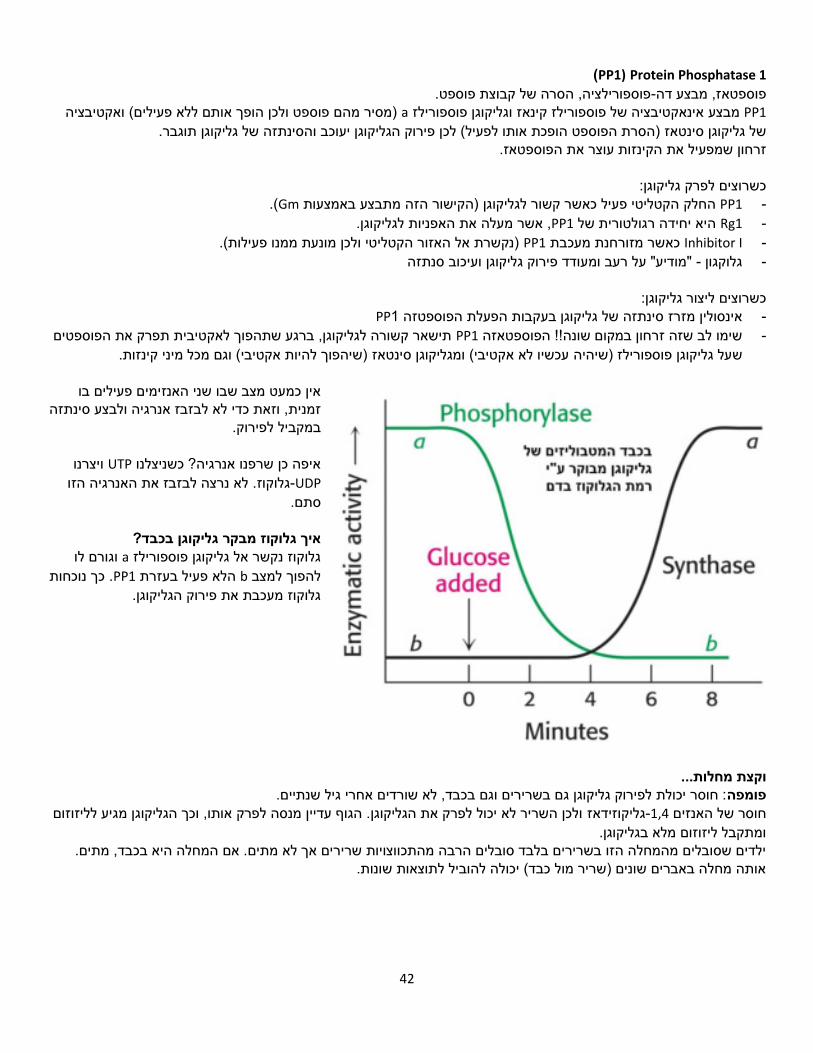
הפעיל(. aפוספורילז קינז עצמו הוא אנזים שעובר פוספורילז
אקטיבציה באמצעות זרחון. ,A, פרוטאין קינאז PKAזרחון ע"י -
אפינפרין יהפוך אותו לפעיל חלקית.וגלוקגון מעודדים את הזרחון הזה
וכך )באופן עקיף( את שבירת הגליקוגן.
מפולסים שמגיעים יוני סידן מגיעים -מהעצבים או מהתכווצויות של
כשהסידן נקשר אל השרירים. פוספורילז קינז הוא הופך אותו
לאקטיבי באופן מלא.
נקשר לרצפטור, מסלול העברת סיגנל מופעל. בשריר: אפינפרין
.cAMP-ל AMPהפעלה של אדנילאט ציקלאז, שפעול של cAMP הופך אתPKA .לפעיל PKA פוספורילציה מבצע לאנזים אקטיבי ושהופך פוספורילציות על פוספורילז קינאז, מבצע
שהוא המצב הפעיל. a-לפוספורילזשהופך b-לפוספורילז .a-הופך לפוספורילז b-מתרחשת שוב הגברה דומה, שבסופה פוספורילזמאותת על רעב. בכבד: גלוקגון
, bעם פוספט הוא במצב הלא פעיל .PKAהופך ללא אקטיבי בעקבות פוספורילציה. את הפוספורילציה הזו מבצע גליקוגן סינטאז
.aללא פוספט הוא במצב הפעיל
ע"י אדנילאט ציקלאז. GTP, שקורה בעקבות הידרוליזה של cAMPתהליך ההגברה נעצר על ידי פירוק (.(Protein Phosphatase 1 PP1בפוספורילז קינאז שמאפשרת עצירה יותר טובה על ידי α הפוספורילציה נוספת על יחידמתרחשת
42
Protein Phosphatase 1 PP1))
פוספורילציה, הסרה של קבוצת פוספט.-מבצע דה ,פוספטאזPP1 גליקוגן פוספורילז ואינאקטיבציה של פוספורילז קינאז מבצעa )ואקטיבציה )מסיר מהם פוספט ולכן הופך אותם ללא פעילים
.תוגבר והסינתזה של גליקוגןלכן פירוק הגליקוגן יעוכב )הסרת הפוספט הופכת אותו לפעיל( ז טאשל גליקוגן סינ זרחון שמפעיל את הקינזות עוצר את הפוספטאז.
כשרוצים לפרק גליקוגן:
- PP1 הקישור הזה מתבצע באמצעות לגליקוגן קשור כאשר פעיל הקטליטי החלק(Gm.) - Rg1 היא יחידה רגולטורית שלPP1 לגליקוגן האפניות את מעלה, אשר. - Inhibitor I מעכבת מזורחנת כאשר PP1 )נקשרת אל האזור הקטליטי ולכן מונעת ממנו פעילות(.
עודד פירוק גליקוגן ועיכוב סנתזה"מודיע" על רעב ומ -גלוקגון -
כשרוצים ליצור גליקוגן:
1PP הפוספטזה הפעלת בעקבות גליקוגן של סינתזה מזרז אינסולין -
תישאר קשורה לגליקוגן, ברגע שתהפוך לאקטיבית תפרק את הפוספטים PP1שימו לב שזה זרחון במקום שונה!! הפוספטאזה - )שיהפוך להיות אקטיבי( וגם מכל מיני קינזות. עכשיו לא אקטיבי( ומגליקוגן סינטאזעל גליקוגן פוספורילז )שיהיה ש
אין כמעט מצב שבו שני האנזימים פעילים בו
זמנית, וזאת כדי לא לבזבז אנרגיה ולבצע סינתזה במקביל לפירוק.
ויצרנו UTPאיפה כן שרפנו אנרגיה? כשניצלנו
UDP-.לא נרצה לבזבז את האנרגיה הזו גלוקוז סתם.
איך גלוקוז מבקר גליקוגן בכבד?וגורם לו aגלוקוז נקשר אל גליקוגן פוספורילז
כך נוכחות .PP1הלא פעיל בעזרת bלהפוך למצב גלוקוז מעכבת את פירוק הגליקוגן.
וקצת מחלות... : חוסר יכולת לפירוק גליקוגן גם בשרירים וגם בכבד, לא שורדים אחרי גיל שנתיים.פומפה
גיע לליזוזום ולכן השריר לא יכול לפרק את הגליקוגן. הגוף עדיין מנסה לפרק אותו, וכך הגליקוגן מגליקוזידאז -1,4חוסר של האנזים ומתקבל ליזוזום מלא בגליקוגן.
המחלה היא בכבד, מתים.ילדים שסובלים מהמחלה הזו בשרירים בלבד סובלים הרבה מהתכווצויות שרירים אך לא מתים. אם אותה מחלה באברים שונים )שריר מול כבד( יכולה להוביל לתוצאות שונות.

43
8( שיעור185/16/)
Integration of metabolism
התבוננות ברמת האורגניזם. מה משותף לתהליכים שלמדנו עד כה? , יצירת אנרגיהוכו'( UTP ,GTP)או ATPמטבוליזם קטבולי תכליתו בעיקר יצירת -
ניצול או יצירה של כוח מחזר, כוח מחמצן - יצירת אבני בניין -
:עקרונות משותפים למסלולים מטבוליים 1. ATP = energy
להרבה מהתהליכים שראינו בקורס.אנרגיה
2. ATP מגיע מחמצון של דלק
למדנו טרםשומן, סוכר, חומצות אמינו, חלבונים )את שני האחרונים -דלק יכול להגיע ממגוון מקורות ובמגוון צורות - בקורס זה(
וכך לקבל את הכמות TCA-הוא צומת מטבולית. נוכל לחמצן אותו במעגל ה A, אצטיל קו Aתמיד נחמצן לאצטיל קו - האפשרית פר פחמן ATPהגדולה ביותר של
. הם ברמות אנרגיה שונות משום NADH, FADH2 -נשאי אלקטרוניםאלא מייצרים קודם ATPלא מייצרים ישירות - שהאלקטרונים בקליפה החיצונית שלהם ברמות אנרגיה שונות
, המעבר שלהם מרמה אחת לאחרת נעשית quantum tunnelingהאלקטרונים עוברים בין קבוצות שונות ועושים - בקפיצה, כל מולק' שקולטת אלקטרון היא ברמת אנרגיה נמוכה מהמולק' שמסרה את האלקטרון
. הכוונה היא שבחלק ו לרוב הולכים לחמצן )אבל לא תמיד, לא כולם ולא תמיד נושמים חמצן(האלקטרונים האל - נו, כמובן שזה תלוי בנוכחות של חמצן.מהמקרים חמצן יהיה נשא האלקטרונים הסופי של
שרוצה למסור אלקטרונים לתהליכי חיזור. כתורמת אלקטרונים, חומר חיזורי תמשמש NADPHהמולק' .3
הפרדה בין חומר שמחזר לחומר שמחמצן. .ברוב תהליכי הסינתזה, תהליכים של יצירת מולקולות, נעשה שימוש בחומר חיזורי
ממולקולות קטנותמולקולות גדולות מסונתזות .4
סוכרים, חלבונים.חומצות שומן, מולק' קטנות מגיעות ממולקולות גדולות שפירקנו, למשל מולק' גדולות שמגיעות מהמזון מפורקות לתתי יחידות בסיסיות, ומהן
הגוף בונה את הפולימרים הנחוצים לו.5. Biosynthesis != Degradation
תמיד קיימת הפרדה בין תהליך של יצירה לתהליך של פירוק כדי שלעולם לא מבצעים או פירוק או יצירה, לא ביחד. - יתרחשו באותו הזמן, ממניעים של בזבוז אנרגיה.
כדי להבטיח שהתגובה הרצויה תתרחש בזמן הרצוי, שימוש בצימוד תגובות. -
בקרה על תהליכים מטבוליים
לרוב נבקר את פעולת האנזימים. קשה לבקר ריכוזי חומרים.
בקרה אלוסטרית .1
באמצעות בקרה , שינוי קונפורמטיבי של חלבון מתרחש במהירות.נעדיף בקרה אלוסטרית משום שהיא מאוד מהירה - אלוסטרית ניתן להשרות תגובות מהירות ומשמעותיות של אנזימים למטבוליטים שונים
, כדי לא להתחיל בכלל את בתהליך שמעורבים בו מספר אנזימים, נרצה שעיקר הבקרה תהיה על האנזים הראשון -שרשרת התהליכים. לפעמים תהיה בקרה גם על אנזים באמצע המסלול, המשמעות היא לרוב שאנזים זה הוא צומת
בין תהליכים מטבוליים שונים.
לרוב נראה בקרה אלוסטרית מאוד חזקה על אנזימים שמבצעים את השלב הבלתי הפיך הראשון במסלול מטבולי - כלשהו.
קוולנטיתבקרה .2
.קשר קוולנטי קשה יותר ליצור וקשה יותר לשחרר לעומת אינטראקציה אלוסטרית -
טריגר לטווח ארוך יותר. בקרה קוולנטית היא ארוכה יותר. -

44
בכל הגוף. OFFאו ONסיגנל יחיד יכול ליצור תגובה של -
רמות אנזימים .3
אם יש צריכה מוגברת של אנזים מסוים, הגוף ייצר ממנו יותר -
4. Compartmentalization
תהליכים שונים מתרחשים באברונים שונים, ויש גם תהליכים חוצי אברונים -
ייצור אנרגיה: מעגל החומצה הציטרית, ייצור גופי קטון )תהליך ו שריפת חומרים יש הרבה תהליכים של במיטוכונדריה - של העברת אנרגיה(
מעבר הלוך ושוב בין האברונים: ייצור אוריאה -
5. Organ specialization
הכבד פועל אחרת מהמוח או מהשרירים -
לכל איבר יש תפקיד ומתפקיד זה נגזרת הפעילות המטבולית של אותו איבר -
איברים שונים יבדלו בסוגי אנזימים, רמות אנזימים, סוגי בקרות -
בין תהליכים שונים יםצמתים מרכזי
, TCAגנזה, חיבור בין גליקוליזה, גלוקונאו יםצמתים שמהוו השלוש
פוספורלציה חמצונית, סינתזה של חומצות שומן, פירוק של חומצות שומן ועוד...
פוספט-6-גלוקוז
רוב התאים לא משחררים גלוקוז.
אחד האיברים היחידים שמשחררים גלוקוז הוא הכליה.. חמצון מבוצע על הוא חד כיווני, ביצענו חמצון פוספט-5-ריבוזהמעבר ל
-)זכרו: חמצון מבוצע ע"י דה הידרוגנז.-דה-פוספט-6-גלוקוזידי .הידרוגנז(
, פוספט-6-פרוקטוזאם יש צורך באנרגיה, ניתן ע"י איזומריזציה להפוך ל .כיווני-. עד כאן התהליך הוא דופירובאטומשם ניתן להגיע ל
)עברנו פוספט-1-גלוקוזכשיש הרבה אנרגיה, ניתן ע"י מוטאז להפוך ל .גליקוגןביפוספט( שיאגר כ-1,6-של גלוקוז intermidiateדרך
פירובאט
בשרירים, זו דרך להעביר עומס מהשרירים אל לקטאטנייצר
. הייצור של לקטאט +NADומייצר ATPהמעבר הזה מייצר הכבד. בשריר הוא בעייתי משום שזו חומצה ולכן מתרחש הפינוי החוצה
מהתאים. נוצר גם הוא מפירובאט ויכול לסייע בהפקת אנרגיה. אלנין
.Aאצטיל קו להפוך באופן בלתי הפיך לגם יכול פירובאט
Aאצטיל קו
.)חץ דו כיווני( חומצות שומןלהופך משתחרר CO2-, תהליך בלתי הפיך משום שTCA-ב CO2-שריפה ל
החוצה מהתא )והגוף(.
הפרופיל המטבולי של איברים
הכבד

45
מכיל מאגר של גליקוגן, משחרר גלוקוז לדם. יש מאגר של טריאציל גליצרול, וחלבונים שניתן לפרק בתור אנרגיה )מאגר ביניים, לחלוקה לתאים(
, מייצר דלק למוח ולשרירים. מהווה בקרה חשובה על ניתוב החומרים הנספגים במעי. , לא נח לרגעמאוד אקטיבי מבחינה מטבולית עים מהמעי עוברים קודם כל בכבד. הכבד קובע את ריכוז כל החומרים בדם. כל הורידים שמגי
שני שליש מהגלוקוז שנכנס לגוף וכל המונוסכרידים האחרים שאינם גלוקוז נספגים בכבד. השאר עובר לשאר האיברים.מעותי יותר של גלוקוז בכבד בשונה משאר התאים, הכבד לא מנצל הרבה גלוקוז. עודף של גלוקוז בכבד הופך לגליקוגן ועודף מש
שחרור גלוקוז מהכבד יבוצע ע"י פירוק הגליקוגן שבכבד או ע"י גלוקונאוגנזה. יהפוך לחומצות שומן. . הכבד יכול לצרוך חומצות אמינו להפקת אנרגיה.הכבד הופך חומצות שומן לגופי קטון וגליצרול לסוכר בצום
שניתן לייצר ממנה גלוקוז. גליצרולפירוק טריאציל גליצרול מניב מולק' הכבד לא מכיל אנזימים שצורכים גופי קטון, כך שהוא יכול רק לייצר אותם אך לא לנצל אותם.
המוח
מכיל קצת גלוקוז, לצריכה. אין אגירה של שום דבר, צריך לספק למוח הכל משאר האיברים. מסיבה זו כל עצירה של אספקה למוח
היא קטלנית. 60%לא במצב רעב, גלוקוז הוא המזון העיקרי למוח. המוח יצרוך אותו בלי הפסקה דרך הדם. במצב מנוחה : כשהגוף גלוקוז
מהאנרגיה מיוצרת בגליקוליזה. 60-70% מהגלוקוז בדם הולך למוח.לתא תחרות בין תא כבד לתא שרירב אין אנזים שמשחרר גלוקוז החוצה מהתא, גלוקוז שנכנס לשריר לא יצא ממנו.בתאי שריר
מוח, המוח יהיה זה ש'יזכה' בגלוקוז.מחומצות שומן, גופי הקטון גופי קטון. במצב של רעב נשרף שומן והכבד מייצר חומצות שומן לא יכולות לשמש כמקור מזון למוח
. והוא יכול להפיק מהם אנרגיה יגיעו אל המוח
השריר
פוספט.-6-מעבר מאוד מהיר לגלוקוז זמין ביותר. גלוקוזגדולים, כדי שיהיה גליקוגן מכיל מאגרי .שומןהשריר מכיל גם מאגר של
.פירוק שרירים יכול להציל )לזמן קצר...(חמור מאוד . במצב של רעב חלבוניםהמאגר הגדול ביותר של השריר הוא של מהדם. גלוקוזהשריר קולט בדומה למוח, אין אנזים שמשחרר גלוקוז החוצה מהתא, גלוקוז שנכנס לשריר לא יצא ממנו.
.פרצי אנרגיה קצרים גלוקוז יהיה מקור האנרגיה העיקרי, בפרצי אנרגיה ארוכים יפורק גם שומןב: צריך הפקת אנרגיה מאוד מעגל קורי
מהירה דרך גליקוליזה. אין זמן ולכן פירובאט הופך TCA-להיכנס ל
ללקטאט )חומצה ששורפת בשריר( הדם יוצאים אל לקטאט ואלנין ואלנין.
ויכולים לעבור אל הכבד, דרך -6-גלוקונאוגנזה להפוך חזרה לגלוקוזפוספט שישוחרר לזרם הדם ויוכל
להגיע חזרה אל השריר.
הלב
מסתמך שריר שמתכווץ כל הזמן, כי צריך כמויות גדולות TCAמאוד על
. המשמעות היא שהלב צורך ATPשל ה , וכן כמות גדולכל הזמן Aאצטיל קו
.חמצןשל .אין בלב מאגרים כללבגלל הפעולה השוטפת,
.Aיעיל להפיק מהן אצטיל קו ופחות פירוק של חומצות שומן הוא מאוד איטי לכן לא מומלץ להגיע למצב של היצרות עורקים. אם חמצן לא מגיע ללב, זה קטלני תוך זמן קצר מאוד.

46
תאי שומן
של חומצות שומן. מכילים את המאגר הכי גדול בגוףיאגר בגוף )עודף של גלוקוז שמגיע מהכבד( הגוף נועד לאגור שומן, וכל עודף אנרגטי
פירוק שומן יכול לשמש ליצירת גלוקוז. כשומן, טריאציל גליצרול.תא שומן גם בתור תא מספק אנרגיה, אם הוא רוקן מכל השומן שנאגר בו, התא עצמו
יפורק וישמש כתאי בנין.יגרמו לפירוק ושחרור )גלוקגון למשל( הורמונים במצב רעב ליפזות שמופעלות ע"י
.של חומצות שומן לדם בקרות מאוד עדינות על שחרור וקליטה של חומצות שומן.
, מה קורה מבחינה מטבולית?רעבאכילה ואז במצב של
(Fed-starv-fed) .גליקוגןיצירה של מופרש אינסולין רמת הגלוקוז בדם עולה אכילה
G3P (פוספט-3-גליצרול )Fatty acyl CoA ויאגרו בתאי השומן. טריגליצרידיםליהפכו , ירידה בגלוקונאוגנזה.גליקוליזהבעלייה
.ריכוז הגלוקוז בדם יורדלאחר הארוחה, ככל שעובר זמן
מתחילה. גלוקגוןהפרשת אינסולין נפסקת והפרשת המאזן המטבולי משתנה:
סינתזה של גליקוגן וחומצות שומן יורדת. מתחילה לעלות. גלוקונאוגנזהגליקוליזה יורדת,
בארוחה הבאה, בשלב הראשון הכבד לא יקח שום סוכר מהדם, כדי שכל התאים יוכלו למלא את מאגרי הגליקוגן הריקים שלהם.
וכך ממלא את מאגר הגליקוגן שלו. הכבד נשאר במצב של גלוקונאוגנזה
צום של כמה ימים, צום מטבולי. ואם הצום ארוך יותר?
כתלות ברמת הפעילות. יש לגוף מאגר של בין קלוריות 1600-6000בגוף יש הרבה קילוקלוריות עודפות. במהלך יום משתמשים חודש לשלושה חודשים )לא כשאין מים, ללא מים ניתן לשרוד בין יומיים לשלושה(.
מאגר הסוכר נגמר תוך יום בערך. מאגר הדלק ברובו איננו של סוכר. ך )הכבד ממשיך ליצר(.רימים ריכוז הסוכר בדם ירד ויגיע למצב קבוע בע 6תוך
תוך יומיים ריכוז גופי הקטון וחומצות השומן בדם עולה מאוד, משום שהכבד מתחיל לייצר גופי קטון.הפוך לגלוקוז, מתאי השומן רק הגליצרול יכול להפוך לגלוקוז. גם חומצות אמינו יכולות להפוך שימו לב שחומצות שומן לא יכולות ל
לגלוקוז. מייצר גלוקוז וגופי קטוןהכבד -
ליפזות משחררות טריגליצרידים מתאי השומן -
עוצרת את Aו רמה גבוהה של אצטיל ק בדם עולה. Aבשריר: מעבר מצריכת גלוקוז לצריכת חומצות שומן. ריכוז אצטיל קו - לא צורך פירובאט TCA-הידרוגנז, וה-הקומפלקס פירובאט דה
ד כדי להפוך לגלוקוזים שיכולים להפוך לגלוקוז עוברים לכברכל התוצ -
יותר גופי קטון ופחות ריכוזם בדם עולה, והמוח צורך גלוקונאוגנזה שנמשכת לאורך זמן מביאה למצב של הרבה גופי קטון, - . עדיין יש תהליכים שתלויים בלעדית בגלוקוז.ם של צום האנרגיה של המוח מיוצרת בעיקר מגופי קטוןימי 3. לאחר גלוקוז
Aיכול להפוך בקלות לאצטיל קו )אחד מגופי הקטון( אצטואצטט -
מה קובע כמה זמן אדם יכול לשרוד בצום?
כמות השומן שיש לאותו אדם.
גנטי שבנוי לאגור. פעם אנשים לא היו מחזיקים את מאגרי השומן האלה זמן רב, לא מסיבה זו לגוף יש נטייה לצבור שומן, מנגנון היה שפע כמו היום.

47
במצב של פעילות גופנית
.ATPגם כאן יש צריכה של אנרגיה, אבל זה מצב שונה של צריכת אנרגיה ונתייחס בעיקר לצריכת הצריכה במצב אנאירובי שונה מהצריכה במצב אירובי.
ראשונות:בשניות ה- ATP )שכבר נמצא בשריר )אין הרבה
ATPקריאטין פוספט שיוצר מיידית -
שניות ראשונות: 10עד - הפיכה ללקטט ואז הלקטט עובר למעגל קורי. אלא לא חמצון מלא בנשימה אנאירובית: פירוק גליקוגן בשריר -
בשתי הדקות הראשונות: - CO2ומייצר TCAפירוק גליקוגן בשריר בנשימה אירובית: השריר מבצע -
לאחר מכן: -
פירוק גליקוגן בכבד -
פעילות גופנית ברמה של שעות: -
CO2וביצוע נשימה אירובית לקבלת Aפירוק חומצות שומן לאצטיל קו -
ככל שהריצה ארוכה יותר, הקצב יהיה איטי יותר משום שהפקת האנרגיה לא מתרחשת בקצב של צריכת האנרגיה.
במצב של צריכת אלכוהול
.NADHיפרק וייצר הידרוגנז-דה-אלכוהולחייבים לפרק את הרעל הזה, ולכן תחילה עוצר תהליכים רבים: בעיקר פועל כסמן שיש בגוף הרבה אנרגיה. זה בעייתי כי אין בגוף באמת הרבה NADHריכוז גבוה של -
אנרגיה. עצרת ההפיכה של לקטאט לפירובאט. אם מצטבר גלוקונאוגנזה עוצרת )משום שיש הרבה אנרגיה( ולכן נ גבוה NADHריכוז -
הרבה לקטאט זה הופך את הדם לחומצי ופוגע בתאים.
)כי מסמן עודף אנרגיה( מעודד סינתזה של חומצות שומן גבוה NADHריכוז -
הידרוגנזות במעגל זה, גם איזו ציטרט דה הידרוגנז וגם -הוא מודולטור שלילי של שני דה NADH-מאט משום ש TCA-מעגל ה - אלפא קטוגלוטרט דה הידרוגנז.
. )המולק' שמגנות על תאי דם אדומים מרדיקלים חפשיים( מחזור של גלוטטיוןמנגנון נוסף שמנוטרל ע"י אתנול הוא -

48
נפגעת ההגנה של הגוף כנגד רדיקלים חפשיים של גלוטטיון. מים לחיזורדם אדו , שמשמש בתאיNADPHפירוק האלכוהול צורך - (.oxidative stressונזקי חמצון )
.Hypoglycemia and lactic acidosis. הרס של הכבדלאט לאט מתרחש -
9( שיעור185/23/)
, תעלות, נשאים ומשאבותממברנות
מליפידים, הידרופוביים.עשויות על תכונותיה, עד כמה היא פלואידית, עד כמה היא מימית או קשיחה.הרכב הממברנה ישפיע
טמפרטורה
לפני שמקפיאים תא צריך להחדיר חומרים שלא יאפשרו טמפ' נמוכה תגרום לממברנה להיות מאוד קשיחה, ועלולה להישבר. לממברנה להישבר בעליית נפח המים.
תפקד כראוי.טמפ' גבוהה תגרום לממברנה להיות מאוד מימית, ולא ל
שומנים שומנים רוויים לא מאפשרים יצירת הסתעפויות, הממברנה מאוד קשיחה.
שומנים בלתי רוויים יאפשרו ממברנה מרווחת יותר.כולסטרול שולט גם הוא בפלואידיות של הממברנה, מולק' שומנית שנכנסת בין הליפידים. בטמפ' גבוהות הליפידים נמשכים אל
יותר צמודים. בטמפ' נמוכות הליפידים יתרחקו מהכולסטרול. הכולסטרול ואז יהיו
מעבר דרך הממברנה חמצן, פד"ח ועוד יכולות לחצות את הממברנה באופן ספונטני. -
מולק' קטנות ולא טעונות כמו מים וגליצרול יכולות לעבור את הממברנה גם הן -
מולק' גדולות וטעונות לא יוכלו לחצות את הממברנה בדיפוסיה -
אם חומרים אלה חוצים את הממברנה ספונטנית או לא. לשערשונים מחוץ ובתוך התא וכך חומריםניתן להתבונן בריכוזי
נעות בהתאם למפל הריכוזים, ממקום בו ריכוזן גבוה למקום בו ריכוזן נמוך באופן טבעי.מולקולות .פאסיבית מעבר בדיפוסיהאם נראה שמחוץ לתא הריכוז מאוד גבוה או מאוד נמוך, נדע כי אין
דיפוסיה פאסיבית לוקחת הרבה יותר זמן מהעברה אקטיבית.
כמות האנרגיה שתידרש תלויה במספר גורמים: הריכוז ההתחלתי של המולקולות -
מטען חשמלי -
דחייה חשמלית חזקה יותר תדרוש השקעה של יותר אנרגיהבו. לממברנה יש פוטנציאל חשמלי בצידה הפנימי ובצידה החיצוני, ויש לממברנה יש פוטנציאל חשמלי, וניתן למדוד את השינויים
לתכונה זו חשיבות בתהליכים רבים.
49
בליעה. :פאגוציטוזה
: שתייה, בחינת פינוצטוזההחומרים שיש מחוץ לתא
באופן לא מאוד מבוקר.אנדוציטוזה שמתווכת ע"י
: מתרחשת בעקבות רצפטור קליטה של סיגנל ברצפטור.
סיכום דרכי העברה דרך הממברנה:
העברה פאסיבית מתרחשת ללא השקעת אנרגיה.
: מולק' שפשוט חוצות את הממברנה. דיפוסיה פשוטה
: תעלות, חורים בממברנה שמאפשרים דיפוסיה פאסיבית מעבר של מולק' בהתאם למפל הריכוזים ללא השקעת אנרגיה.
העברה באמצעות : (Facilitated diffusion)דיפוסיה מזורזת נשא מסוג יוניפורטר, ללא השקעת אנרגיה, בהתאם למפל
הריכוזים.
:העברה אקטיבית מתרחשת תוך השקעת אנרגיה.
בתא משמש להעברות דרך הממברנה. ATP-מה 25%בערך זה המון!
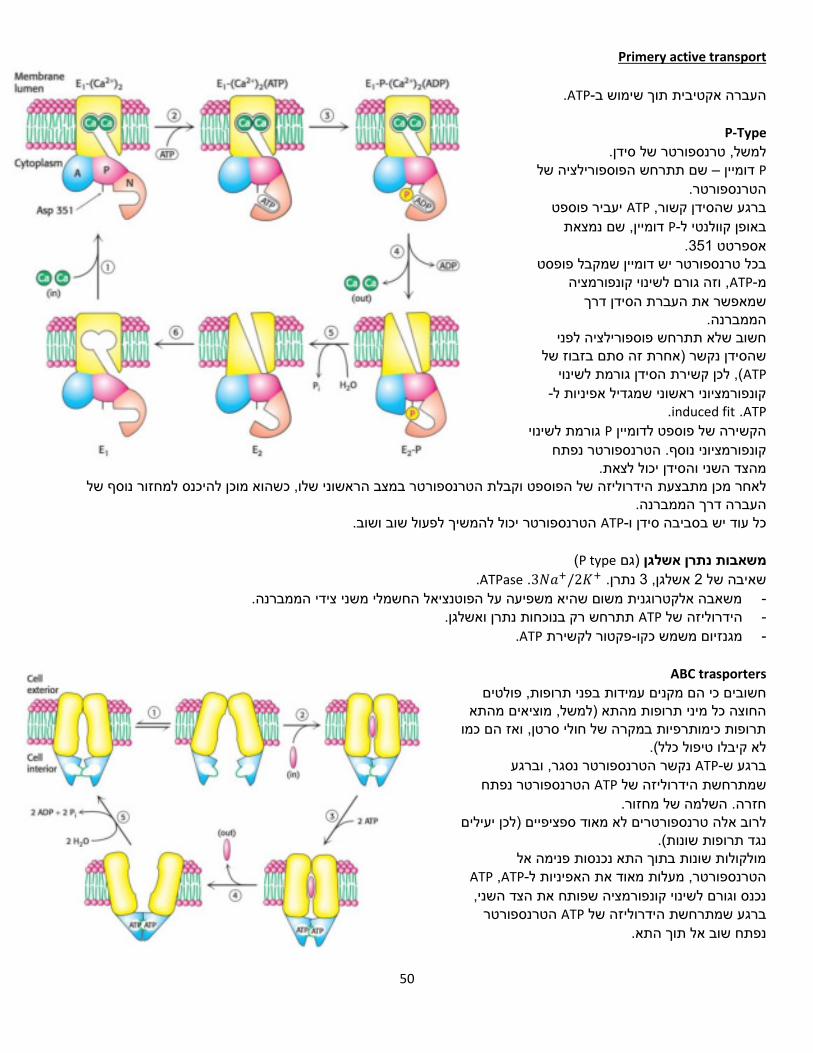
50
Primery active transport
.ATP-העברה אקטיבית תוך שימוש ב
P-Type טרנספורטר של סידן. למשל,
P שם תתרחש הפוספורילציה של –דומיין הטרנספורטר.
יעביר פוספט ATPברגע שהסידן קשור, , שם נמצאת דומיין P-באופן קוולנטי ל
. 351אספרטט בכל טרנספורטר יש דומיין שמקבל פופסט
, וזה גורם לשינוי קונפורמציה ATP-משמאפשר את העברת הסידן דרך
הממברנה.חש פוספורילציה לפני חשוב שלא תתר
שהסידן נקשר )אחרת זה סתם בזבוז של ATP לכן קשירת הסידן גורמת לשינוי ,)
-קונפורמציוני ראשוני שמגדיל אפיניות לATP. induced fit.
לשינוי מתגור Pהקשירה של פוספט לדומיין הטרנספורטר נפתח קונפורמציוני נוסף.
מהצד השני והסידן יכול לצאת.הידרוליזה של הפוספט וקבלת הטרנספורטר במצב הראשוני שלו, כשהוא מוכן להיכנס למחזור נוסף של לאחר מכן מתבצעת
העברה דרך הממברנה. הטרנספורטר יכול להמשיך לפעול שוב ושוב. ATP-סידן ובסביבה כל עוד יש
(P typeגם ) משאבות נתרן אשלגן
.3𝑁𝑎+/2𝐾+. ATPaseנתרן. 3אשלגן, 2של שאיבה משאבה אלקטרוגנית משום שהיא משפיעה על הפוטנציאל החשמלי משני צידי הממברנה. -
תתרחש רק בנוכחות נתרן ואשלגן. ATPהידרוליזה של -
.ATPפקטור לקשירת -מגנזיום משמש כקו -
ABC trasporters
בפני תרופות, פולטים חשובים כי הם מקנים עמידותהחוצה כל מיני תרופות מהתא )למשל, מוציאים מהתא
תרופות כימותרפיות במקרה של חולי סרטן, ואז הם כמו .לא קיבלו טיפול כלל(
ברגע נקשר הטרנספורטר נסגר, ו ATP-ברגע שהטרנספורטר נפתח ATPשמתרחשת הידרוליזה של חזרה. השלמה של מחזור.
טרנספורטרים לא מאוד ספציפיים )לכן יעילים לרוב אלה נגד תרופות שונות(.
מולקולות שונות בתוך התא נכנסות פנימה אל ATP ,ATP-הטרנספורטר, מעלות מאוד את האפיניות ל
נכנס וגורם לשינוי קונפורמציה שפותח את הצד השני, הטרנספורטר ATPברגע שמתרחשת הידרוליזה של
נפתח שוב אל תוך התא.
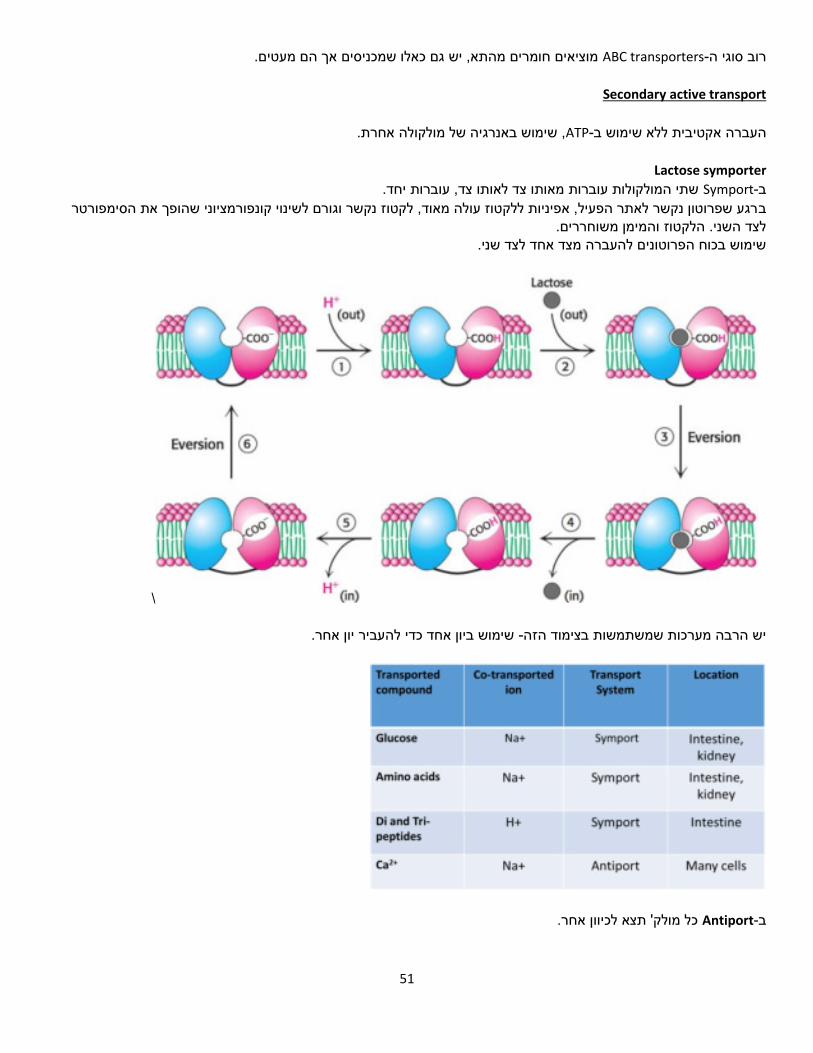
51
מוציאים חומרים מהתא, יש גם כאלו שמכניסים אך הם מעטים. ABC transporters-רוב סוגי ה
Secondary active transport
, שימוש באנרגיה של מולקולה אחרת.ATP-אקטיבית ללא שימוש ב העברה
Lactose symporter
שתי המולקולות עוברות מאותו צד לאותו צד, עוברות יחד. Symport-ברגע שפרוטון נקשר לאתר הפעיל, אפיניות ללקטוז עולה מאוד, לקטוז נקשר וגורם לשינוי קונפורמציוני שהופך את הסימפורטר ב
לצד השני. הלקטוז והמימן משוחררים. שימוש בכוח הפרוטונים להעברה מצד אחד לצד שני.
\
להעביר יון אחר.שימוש ביון אחד כדי -יש הרבה מערכות שמשתמשות בצימוד הזה
כל מולק' תצא לכיוון אחר. Antiport-ב

52
transportPassive
K+ Channel ספציפיות.מאוד
ראשית, מתאימה רק ליונים טעונים חיובית, יונים טעונים שלילית לא יכנסו לתעלה זו.מטען: .1 כל היונים הגדולים מרדיוס מסוים לא יוכלו לעבור בתעלה זו. גודל: .2מולק' מים פולארית יכולה לעטוף יון כך שיהיה לו 'נוח' איך ניצור ספציפיות גם עבור יונים שקטנים מרדיוס זה?כוח דהידרציה: .3
.salvation cageבסביבה מימית, זו תופעה שנקראת המים. הסרת הקשרים של -כדי להכניס את היון עלינו לייצר סביבה שמים לא יכולים לעבור בה, כוח של דהידרציה
ואלין יוצר בדיוק את המצב הזה. לאשלגן יותר נוח עם ח' אמינו אלו מאשר עם המים.-גליצין-טירוזין-רצף של גליצין אין כאן באמת השקעת אנרגיה משום שהקשר של האשלגן עם התעלה מחזיר את האנרגיה שהשקענו בביצוע הדהידרציה.
, לאחר דהידרציה המרחק שנוצר בין ח' האמינו בתוך התעלה salvation cageנתרן למשל היא מולק' קטנה יותר מיון אשלגן עם הוא כבר גדול מדי עבור יון הנתרן והנתרן לא יעבור.
של חומצות אמינו שותף בביצוע הדהידרציה בתעלות אשלגן. TVGYGהרצף
)אם צריך להשקיע יותר מדי אנרגיה, דהידרציהולפי כוח )רק קטן מרדיוס מסוים( לפי גודל)רק מטען חיובי(, לפי מטען ספציפיות למשל עבור מולק' קטנה מדי, זה פשוט לא יקרה(.
איך נוציא את האשלגן מהתעלה אם כל כך נוח לו שם? מנגנון שדוחף את האשלגן החוצה. בתעלה זו יש מקום לארבע מולק' אשלגן
חשמלית.שטעונות חיובית ולכן הן דוחפות אחת את השניה, משום הדחייה המולק' ראשונה שנכנסת תתמקם בנוחות, בחלק פנימי יחסית. מולק' שניה של אשלגן תשלח את הראשונה עוד קצת פנימה, וכך
הלאה עד שהראשונה שנכנסה לתעלה תידחף החוצה ממנה.
Membrane depolarization
Voltage dependent channels
תעלות שתלויות בפוטנציאל של הממברנה.למשל, תעלה סגורה כך שיונים לא יכולים לעבור מצד אחד לצד השני. כשיש עודף של יונים חיוביים, אחת הזרועות של התעלה
תנוע פנימה לתוך הממברנה כתוצאה מהדחייה שלה מהיונים החיוביים, ובכך תגרום לפתיחה של התעלה. מצד אחד של הממברנה גורם לפתיחת התעלה.מטען חשמלי כלומר
שיטה לאינאקטיבציה מהירה מאוד של תעלה. :Ball and chainשיטת
כשהממברנה נפתחת, הכדור יכול לסגור את התעלה. שלושה מצבים:
- Closed
- Open
- Inactivated
תעלה מאוד מבוקרת.
תעלות נתרן אשלגןכשפותחים ראשית את תעלת הנתרן, נכנס הרבה מאוד נתרן בבת אחת,
של תעלת הנתרן. אקטיבציה-ונגרמת דה אחרי פתיחת תעלת הנתרן, תעלת אשלגן נפתחת לאט יותר וגם נסגרת לאט יותר.
אשלגן , קודם כל ריכוז נתרן מתחיל לעלות ועולה במהירות גבוהה. לאחר מכן ריכוז B)אפשר לראות את זה בתרשים בתמונה סגורה, פתוחה ואח"כ לא -מונים לאורך זמן של מצב התעלהאפשר לראות סי C. בתמונה עולה בצורה מתונה יותר ולאורך יותר זמן
(.ball and chainפעילה משום השימוש במנגנון

53
: תיאור של העברת סיגנל באמצעות מתח חשמלי, שמימיןבשרטוט פולאריזציה.-פולאריזציה ומיד לאחר מכן רה-דה
יש היום שיטות כמותיות למדידת פוטנציאל הממברנה.
, משום דחייה ממברנה שלא יאפשר מעבר של יוניםיש פוטנציאל משלב מסוים לא יתאפשר יותר מעבר של יונים מצד לצד חשמלית.
וישמר ריכוז לא שווה משני צידי הממברנה.
Nerst equation פוטנציאל מהו למצוא כדי, הריכוזים בין היחסים של ln-ב כופלים. היוני המשקל שיווי פוטנציאל
.חשמלי למתח מתייחס Ex שהערך להבין חשוב. מ"בשו יהיו היונים שבו הממברנה מחוץ היונים יחס יהיה מה לנבא נרסט משוואת ובאמצעות, ממברנה על מתח להפעיל גם אפשר .הממברנה ובתוך
Z 2,+1,+1-, היון של ולנטיות מייצג. ומחוץ לממברנה, כל בהצבה בנוסחה, מה שמעניין אותנו בעיקר זה ריכוז היון בתוך הממברנה
השאר קבועים.
Goldman equation בתא יש יותר מיון אחד, ולפעמים נצטרך להתחשב בכמה יונים שונים בעת ההצבה בנוסחה.
נוסחה זו מתחשבת בריכוזים של שני יונים שונים, ומתקבל פוטנציאל קרוב יותר לפוטנציאל שקיים בתאים. כאשר עוסקים במספר יונים כשלכל יון יש מקדם חדירות.נרצה לדעת מהו מתח הממברנה
בכלור שהוא יון שלילי: נרשום במונה את הבפנים ובמכנה את הבחוץ. ביונים חיוביים כמו אשלגן נכתוב ההפך )ריכוז בחוץ חלקי ריכוז בפנים(.
ההבדלים הם משום שיונים חיוביים ושליליים משפיעים אחרת.
0.058 [ ]log
[ ]
I
II
XEx
z X
[ ] [ ] [ ]0.058log
[ ] [ ] [ ]
K outside Na outside Cl inside
K inside Na inside Cl outside
P K P Na P ClVm
P K P Na P Cl

54
Ligand activated channels במקרה של נוירונים למשל, כשרוצים להעביר סיגנל מתא אחד אל תא שני.
סיגנלים )ליגנד( נקשרים אל רצפטורים בתא, לפעמים הרצפטורים הללו הם תעלות שבמגע עם הליגנד משנות קונפורמציה ונפתחות.
הליגנד )רצפטור(.מכילות אתר תוך ממברנלי שהוא המעבר עצמו, ואתר חוץ ממברנלי אליו נקשר
Acetylcholine Receptor (AchR) Ligand
תעלה לא מאוד ספציפית ליונים, יכולה להכניס גם נתרן וגם אשלגן. שתי יחידות אלפא שיכולות לקשור אצטילכולין.
ת לאצטילכולין וכך התעלה סגורה, קישור אצטילכולין גורמת לשינוי קונפורמציה. קשירה של אצטילכולין ראשון מגבירה את האפיניו במהירות יקשר גם אצטילכולין שני. ברגע ששניים קשורים התעלה תבצע סיבוב מלא ותיפתח.
אצטילכולין משמש בעיקר בהעברת סיגנל עצבי: אצטיל כולין הוא נוירוטרנסמיטר שמשתחרר אל הסינפסה )המרווח בין קצה אקסון
כולין נקשר אל רצפטור של תעלת נתרן תלוית ליגנד וגורם לפתיחת התעלה של תא עצב לבין תא עצב אחר או תא שריר(. אצטיל פולריזציה של -פולריזציה )עלייה בפוטנציאל הממברנה, הפיכה ליותר חיובי(, דה-וכניסת נתרן לתא. כניסת הנתרן גורמת לדה
לאפס )אחרי שירד מתחת לאפס(.אשלגן יחזירו את פוטנציאל הממברנה חזרה -לשחרור של סידן. משאבות נתרן מתהממברנה גור
של תעלות אצטילכולין, ואז.. מתים. חשוב לסגור את התעלות האלו. הנשק כימי כמו גז חרדל וכדומה, מעכב את הסגיררוב הרעלים שמפרישים עקרבים, נחשים, עכבישים וכו' פוגעים בתעלות. פוגעים במערכת העצבית של בע"ח שהורעל וכך אין
ולא מתאפשרת הזזה של השרירים.הפעלת סיגנלים שילוב בין סיגנל חשמלי וסיגנל כימי.פועלים עפ"י י עצב את
Optogenetics Light activated channels שאורך גל מסוים גורם להן לאקטיבציה.תעלות אופטוגנטיקה.
גורמים לבע"ח לבטא את התעלות האלה, ואז באמצעות אור גורמים להעברת סיגנל. אפשר לתת סיגנל ספציפי. ספציפיות מאוד גבוהה, ניתן לבטא את התעלות הללו רק בתאים מסוימים.נותן
אופטוגנטיקה עושה שימוש בשתי תעלות:
.פולריזציה-דה מאפשרת מעבר של קטיונים )יוני נתרן( , Channelrhodopsinתעלת רודופסין, -
אקטיבציה באמצעות אור כחול.
.)חזרה למצב התקין( פולריזציה-היפר מאפשרת מעבר של יוני כלור , Halorhodopsinתעלת כלור, -
אקטיבציה באמצעות אור צהוב.
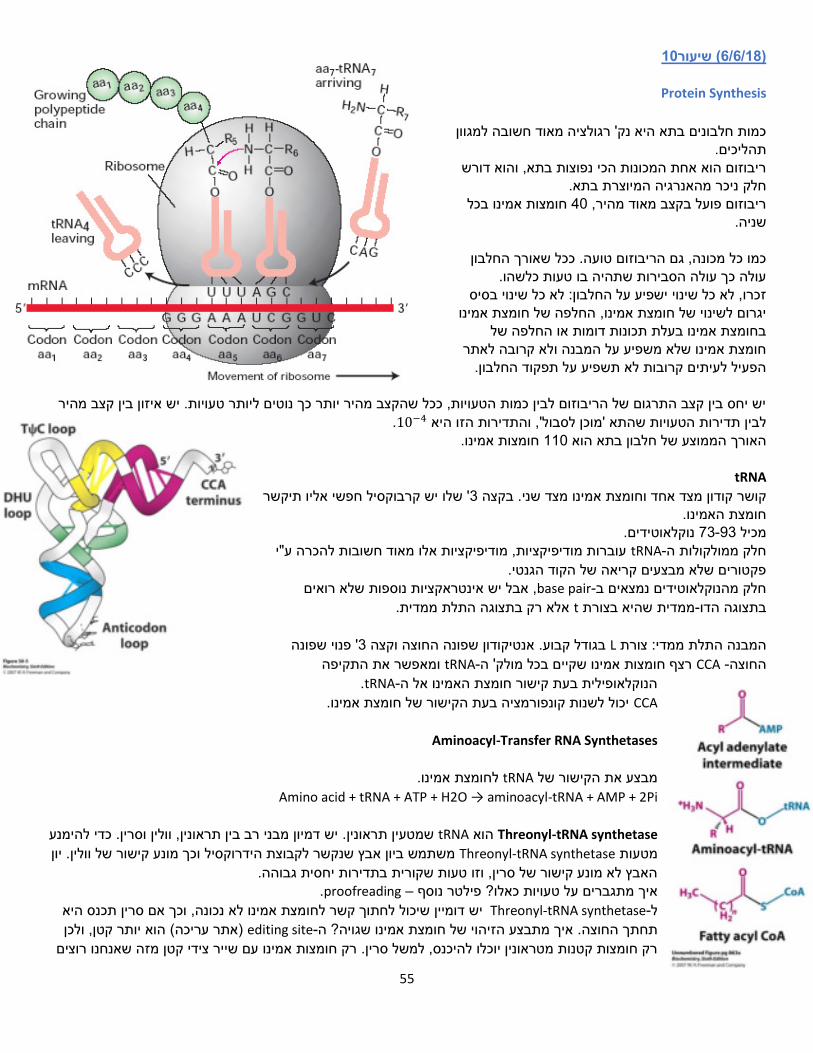
55
10( שיעור186/6/)
Protein Synthesis
כמות חלבונים בתא היא נק' רגולציה מאוד חשובה למגוון תהליכים.
ריבוזום הוא אחת המכונות הכי נפוצות בתא, והוא דורש ניכר מהאנרגיה המיוצרת בתא. חלק
חומצות אמינו בכל 40ריבוזום פועל בקצב מאוד מהיר, שניה.
כמו כל מכונה, גם הריבוזום טועה. ככל שאורך החלבון
עולה כך עולה הסבירות שתהיה בו טעות כלשהו.זכרו, לא כל שינוי ישפיע על החלבון: לא כל שינוי בסיס
החלפה של חומצת אמינו יגרום לשינוי של חומצת אמינו, בחומצת אמינו בעלת תכונות דומות או החלפה של
חומצת אמינו שלא משפיע על המבנה ולא קרובה לאתר הפעיל לעיתים קרובות לא תשפיע על תפקוד החלבון.
איזון בין קצב מהיר יש יחס בין קצב התרגום של הריבוזום לבין כמות הטעויות, ככל שהקצב מהיר יותר כך נוטים ליותר טעויות. יש
.10−4ירות הטעויות שהתא 'מוכן לסבול', והתדירות הזו היא לבין תד חומצות אמינו. 110הממוצע של חלבון בתא הוא האורך
tRNA ' שלו יש קרבוקסיל חפשי אליו תיקשר 3קודון מצד אחד וחומצת אמינו מצד שני. בקצה קושר
חומצת האמינו. נוקלאוטידים. 73-93מכיל
עוברות מודיפיקציות, מודיפיקציות אלו מאוד חשובות להכרה ע"י tRNA-חלק ממולקולות ה פקטורים שלא מבצעים קריאה של הקוד הגנטי.
, אבל יש אינטראקציות נוספות שלא רואים base pair-חלק מהנוקלאוטידים נמצאים ב אלא רק בתצוגה התלת ממדית. tשהיא בצורת ממדית -בתצוגה הדו
' פנוי שפונה 3אנטיקודון שפונה החוצה וקצה בגודל קבוע. Lהמבנה התלת ממדי: צורת
ומאפשר את התקיפה tRNA-רצף חומצות אמינו שקיים בכל מולק' ה CCA -החוצה .tRNA-הנוקלאופילית בעת קישור חומצת האמינו אל ה
CCA .יכול לשנות קונפורמציה בעת הקישור של חומצת אמינו
Aminoacyl-Transfer RNA Synthetases
לחומצת אמינו. tRNAמבצע את הקישור של Amino acid + tRNA + ATP + H2O → aminoacyl-tRNA + AMP + 2Pi
Threonyl-tRNA synthetase הואtRNA יש דמיון מבני רב בין תראונין, וולין וסרין. כדי להימנע ראונין. תשמטעין
וכך מונע קישור של וולין. יון משתמש ביון אבץ שנקשר לקבוצת הידרוקסיל Threonyl-tRNA synthetaseמטעות האבץ לא מונע קישור של סרין, וזו טעות שקורית בתדירות יחסית גבוהה.
.proofreading –פילטר נוסף על טעויות כאלו?מתגברים איך מצת אמינו לא נכונה, וכך אם סרין תכנס היא חוקשר לדומיין שיכול לחתוך יש Threonyl-tRNA synthetase -ל
הוא יותר קטן, ולכן )אתר עריכה( editing site-האיך מתבצע הזיהוי של חומצת אמינו שגויה? תחתך החוצה.רק חומצות אמינו עם שייר צידי קטן מזה שאנחנו רוצים רק חומצות קטנות מטראונין יוכלו להיכנס, למשל סרין.
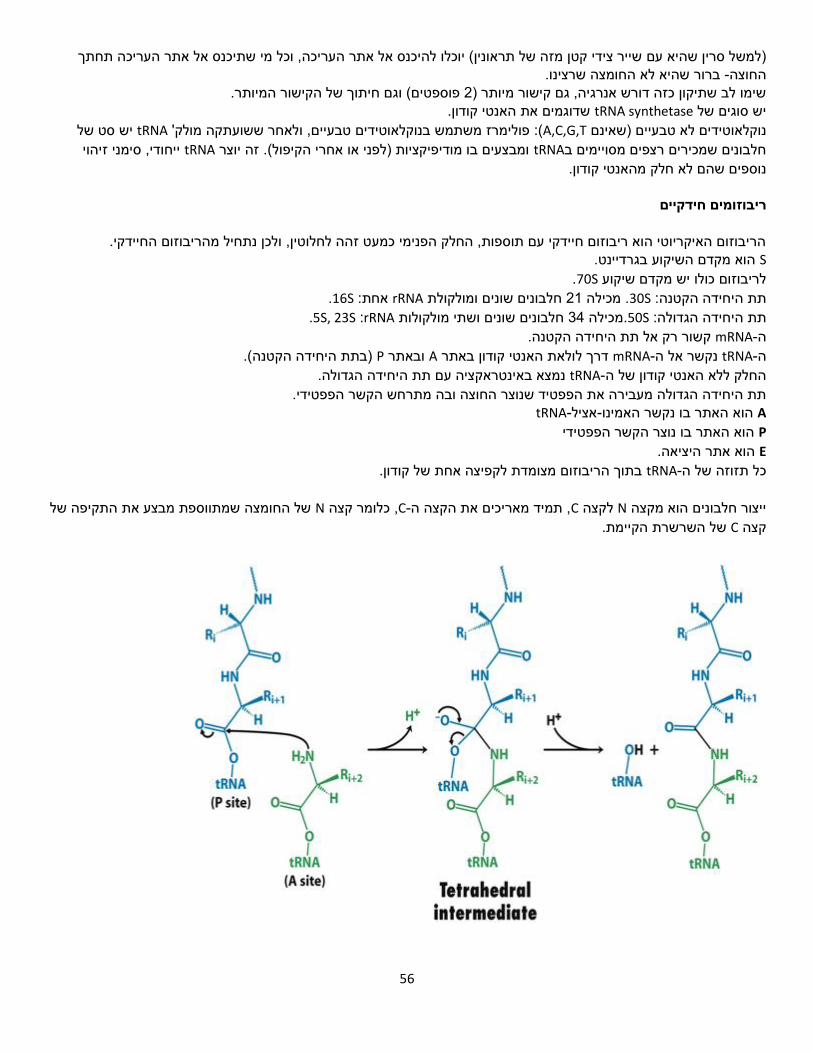
56
תיכנס אל אתר העריכה תחתך שי כל משייר צידי קטן מזה של תראונין( יוכלו להיכנס אל אתר העריכה, ו )למשל סרין שהיא עם ברור שהיא לא החומצה שרצינו. -החוצה
.פוספטים( וגם חיתוך של הקישור המיותר 2שימו לב שתיקון כזה דורש אנרגיה, גם קישור מיותר ) האנטי קודון. שדוגמים את tRNA synthetaseיש סוגים של
יש סט של tRNAלאחר ששועתקה מולק' ו, טבעייםפולימרז משתמש בנוקלאוטידים : (A,C,G,T)שאינם נוקלאוטידים לא טבעייםייחודי, סימני זיהוי tRNAזה יוצר ומבצעים בו מודיפיקציות )לפני או אחרי הקיפול(. tRNAחלבונים שמכירים רצפים מסויימים ב
מהאנטי קודון.נוספים שהם לא חלק
ריבוזומים חידקיים
הריבוזום האיקריוטי הוא ריבוזום חיידקי עם תוספות, החלק הפנימי כמעט זהה לחלוטין, ולכן נתחיל מהריבוזום החיידקי.S .הוא מקדם השיקוע בגרדיינט
. 70Sלריבוזום כולו יש מקדם שיקוע .16Sאחת: rRNAלת חלבונים שונים ומולקו 21. מכילה 30Sתת היחידה הקטנה: .rRNA :5S, 23Sחלבונים שונים ושתי מולקולות 34מכילה .50Sתת היחידה הגדולה:
קשור רק אל תת היחידה הקטנה. mRNA-ה )בתת היחידה הקטנה(. Pובאתר Aדרך לולאת האנטי קודון באתר mRNA-נקשר אל ה tRNA-ה
נמצא באינטראקציה עם תת היחידה הגדולה. tRNA-החלק ללא האנטי קודון של ה תת היחידה הגדולה מעבירה את הפפטיד שנוצר החוצה ובה מתרחש הקשר הפפטידי.
A אציל-הוא האתר בו נקשר האמינו-tRNA P הוא האתר בו נוצר הקשר הפפטידי E .הוא אתר היציאה
בתוך הריבוזום מצומדת לקפיצה אחת של קודון. tRNA-כל תזוזה של ה
של החומצה שמתווספת מבצע את התקיפה של N, כלומר קצה C-, תמיד מאריכים את הקצה הCלקצה Nייצור חלבונים הוא מקצה של השרשרת הקיימת. Cקצה

57
החדש. tRNA-תוקף, והשרשרת עוברת אל ה Aהחדש שנכנס אל אתר tRNA-אמינואצילאמין חפשי של ה אסתרי.קשר אמידי הוא יותר יציב מקשר
במיקומים ובזוויות מתאימות ולות המעורבותהקשרים, התפקיד שלו הוא לייצב את כל המולק תבתקיפה וביציר מעורבהריבוזום לא כך שתתרחש התקיפה.
, אם הקישור של הקודון והאנטי קודון לא תקין הוא יוצא החוצה, יודעים אם הקישור תקין לפי הגובה של Aנכנס אל אתר tRNA-כש צור גובה שונה(.בתוך הריבוזום )קישור לא מתאים י tRNA-ה
שצריכים להיות מוכרים ע"י קודונים שונים 61יש שונים יש בתא? tRNAכמה שונות. tRNAמולק' 61, אבל אין בתא tRNAאנטיקודונים של
לא חייב להיות בהתאמה מלאה.איך זה אפשרי? הנוקלאוטיד השלישי בקודון שימו לב שלרוב כשחומצת אמינו אחת מקודדת על ידי יותר מקודון אחד הקודונים
.wobblingהאחרון, זה נקרא השלישי והשונים נבדלים זה מזה בזהות הנוקלאוטיד קודון שמאפשרת להכיר כל -היא מודיפיקציה בעמדה האחרונה של האנטי אינוזין
לעיתים קרובות העמדה האחרונה קודון.-מיני קודונים שונים באמצעות אותו אנטי של האנטי קודון תהיה אינוזין אך לא תמיד.
הפרוקריוטי פעולת הריבוזום
:אתחול
.’3אל קצה ’5ומתחילה לנוע על גביה במהירות די גבוהה מקצה mRNAתת יחידה קטנה של הריבוזום נקשרת אל מולק' הראשון, זה רצף שמוכר ע"י תת היחידה הקטנה של AUG-לפני ה רצף שיין דלגרנובחיידקים קיים
הריבוזום ובעקבות הכרה זו נגרמת האטה של הסריקה. (IF1, IF3) עם פקטורים שונים mRNA-אל הנקשרת קודם כל 30Sהקטנה תת היחידה בחיידקים - ביצוע סריקה עד להגעה לרצף שיין דלגרנו -
:הקיימים בפרוקריוטיםפקטורים -
- IF1 : ,30נקשר אל תת היחידה הקטנהSומונע מאמינואציל , של הריבוזום-tRNA להיקשר, להיקשר גם היא 50S. בנוסף, מונע מתת היחידה הגדולה, IF2-מגדיל את האפיניות לאליה.
וליצור את הקומפלקס השלם
- IF2 :IF2 קושרGTP .לאחר מכן ונקשר גם הוא אל תת היחידה הקטנה של הריבוזוםfMET-
tRNA נקשר אלIF2ו ,-IF2 מעביר אותו אל האתרP .בעת הקשירה של תת של הריבוזוםישנה קונפורמציה וישתחרר, וכך IF2בעקבותיו GTPהיחידה הגדולה יתבצע פירוק של
.יאפשר את יצירת הקומפלקס
- IF3 :נחוץ להיקשרות של תת היחידה הקטנה אל אזור תחילת התרגום ב-mRNA
פקטורים נוספיםבאיקריוטים ישנם -
מתיונין )ח' האמינו הראשונה בעת תרגום חלבון -, מכיל פורמילPאתר בתוך ראשון tRNAכעת ישנו - בפרוקריוטים(.
מתווספת 50Sתת היחידה הגדולה -
נעילה של הקומפלקס -
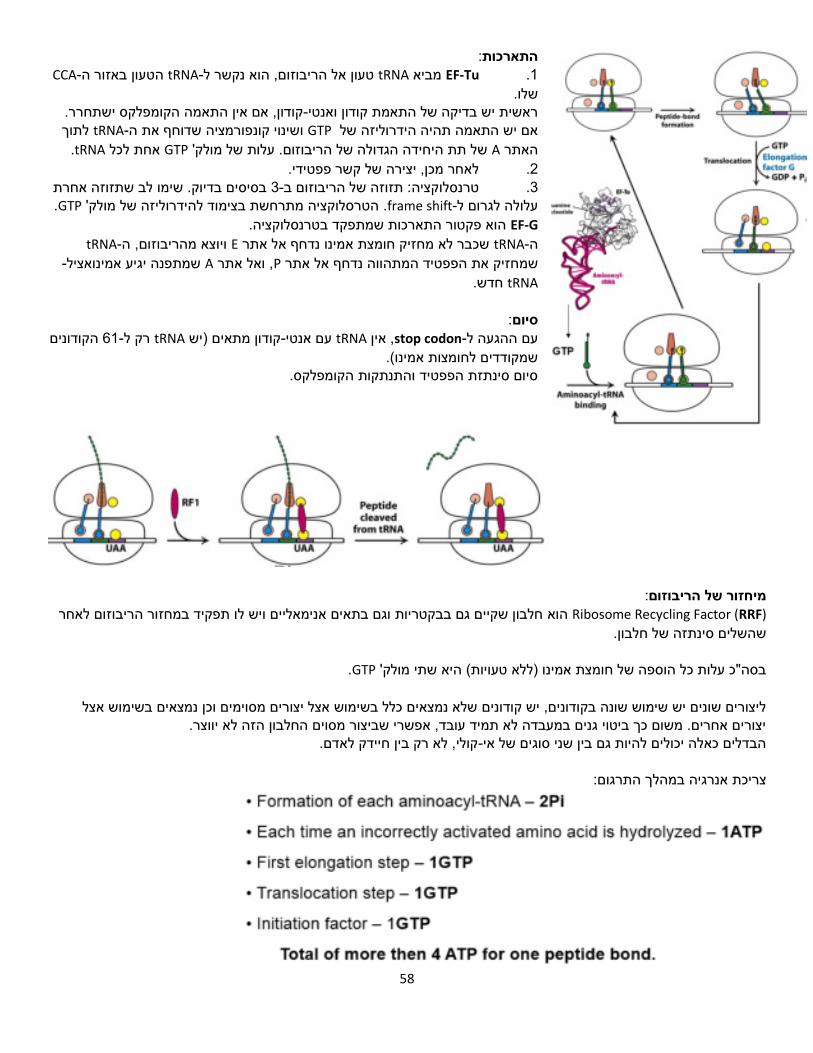
58
:התארכות1. EF-Tu מביאtRNA אל הריבוזום, הוא נקשר לטעון-tRNA באזור ה הטעון-CCA
.שלוקודון, אם אין התאמה הקומפלקס ישתחרר. -ראשית יש בדיקה של התאמת קודון ואנטי
לתוך tRNA-ושינוי קונפורמציה שדוחף את ה GTPאם יש התאמה תהיה הידרוליזה של .tRNAאחת לכל GTPשל תת היחידה הגדולה של הריבוזום. עלות של מולק' Aהאתר
לאחר מכן, יצירה של קשר פפטידי. .2בסיסים בדיוק. שימו לב שתזוזה אחרת 3-קציה: תזוזה של הריבוזום בטרנסלו .3
.GTPהטרסלוקציה מתרחשת בצימוד להידרוליזה של מולק' .frame shift-עלולה לגרום לEF-G .הוא פקטור התארכות שמתפקד בטרנסלוקציה
tRNA -ויוצא מהריבוזום, ה Eשכבר לא מחזיק חומצת אמינו נדחף אל אתר tRNA-ה -שמתפנה יגיע אמינואציל A, ואל אתר Pחזיק את הפפטיד המתהווה נדחף אל אתר שמ
tRNA .חדש
:סיום
הקודונים 61-רק ל tRNAקודון מתאים )יש -עם אנטי tRNA, אין stop codon-עם ההגעה ל שמקודדים לחומצות אמינו(.
סיום סינתזת הפפטיד והתנתקות הקומפלקס.
:חזור של הריבוזוםימ
Ribosome Recycling Factor (RRF) חלבון שקיים גם בבקטריות וגם בתאים אנימאליים ויש לו תפקיד במחזור הריבוזום לאחר הוא שהשלים סינתזה של חלבון.
.GTPשתי מולק' היאבסה"כ עלות כל הוספה של חומצת אמינו )ללא טעויות(
שלא נמצאים כלל בשימוש אצל יצורים מסוימים וכן נמצאים בשימוש אצל ליצורים שונים יש שימוש שונה בקודונים, יש קודונים
יצורים אחרים. משום כך ביטוי גנים במעבדה לא תמיד עובד, אפשרי שביצור מסוים החלבון הזה לא יווצר. קולי, לא רק בין חיידק לאדם.-הבדלים כאלה יכולים להיות גם בין שני סוגים של אי
התרגום:צריכת אנרגיה במהלך
59
ריבוזומים אנושיים
תהליך התרגום בבני אדם )ובעלי חיים באופן כללי( הוא מורכב יותר. - mRNA באיקריוטים שונה מ-mRNA בפרוקריוטים, הוא עובר עריכה ויש לוpolyA ו-cap 5’. טרם תחילת התרגום. mRNA-באיקריוטים יש קשירה מעגלית של ה - ם של תחילת התרגום, בפרוקריוטים יש סט שלם של פקטורים לתחילת התרגום.פקטורי 3אתחול: בבקטריות יש רק - .tRNA-מתיונין-ולא פורמיל tRNA-מתיוניןבאיקריוטים התרגום מתחיל ב -
. יש רצף מיוחד ששונה בין פרוקריוטים ואיקריוטים והוא מאותת להשהיית ER-הבדל נוסף הוא בהכנסת חלבונים לממברנה, ל - ימשך תהליך התרגום. ER-. עם הגעת הריבוזום לER-הריבוזום אל ה התרגום עד להגעה של
.טרנסלוקון: יכול להכניס קיפול של חלבון אל תוך הממברנה או אל פנים האברון )דרך הממברנה(
אנטיביוטיקה
ואחרות. אנטיביוטיקה חומר שמפריע למטבוליזם של החיידק, כשהריבוזום הוא אחת המטרות המרכזיות של אנטיביוטיקות כאלו יכולה להיות מיוצרת באופן טבעי או מלאכותי.
ציקלין-טטרה
קולי ומשתמשים בה במעבדות שונות.-אנטיביוטיקה שהורגת אייעילה נגד מגוון רחב של מיקרואורגניזמים.
Chloramphnicolכלורמפניקול .מיוצרת באופן סינטטי. יעילה נגד מגוון רחב של מיקרואורגניזמים
Chloramphenicol resistance is achieved by production of chloramphenicol acetyltransferase which modifies the hydroxyl
positions 1 and 3.
ח' האמינו )החדשה, 2ומעכבת את האנזים שמזרז את יצירת הקשר הפפטידי בין 50Sכלורמפניקול נקשרת לתת היחידה הגדולה האחרונה בפפטיד המתהווה(. והחומצה
Macrolides
נפוץ בהרבה תרופות.מקרולידים, מודיפיקציות שונות על טבעת הלקטון.

60
של הריבוזום וחוסם באופן כלשהו את מעבר בריבוזום יש מנהרה דרכה עובר הפפטיד. המקרוליד נקשר במרחק קצר מאתר _ הפפטיד.
:עמידות וסלקטיביות
מקרולידים.אדם לא קושרים בני מספיק אטום אחד שיתנגש במקרוליד והמקרוליד לא יקשר, והחיידק יהיה עמיד לכך.
למיטוכונדריה יש ריבוזום מיטוכונדריאלי, הוא דומה יותר לריבוזום החיידקי ברצף )זה המקור ממנו הוא בא(. בריכוזים גבוהים של
חולשה ועייפות.אנטיביוטיקה זה עלול להשפיע על המיטוכונדריה, הרגשה של 11( שיעור13/06/18)
The proteasome
למה צריך להיפטר מחלבונים?
בקרת איכות, חלבונים פגומים יכולים להביא למצב של טוקסיות בתא. .1
מיחזור: אם אין צורך בחלבונים מסויימים, עדיף לפרק אותם לחומצות אמינו ולהשתמש בהן לבניין חלבונים נחוצים. .2
מזהה חלבונים זרים ומפרקת אותם.המע החיסונית: .3
שמירה על יחס סטוכיומטרי .4
בקרת איכות
לתא חלבונים שלא מקופלים נכון יכולים להיות רעילים -
חלבונים שאין בהם צורך כרגע, אם הם נשארים הם ימשיכו לפעול -, כדי cell cycle-באופן שלא נחוץ לנו. דוגמה קלאסית לכך היא ה
לשלב צריך חלבונים שונים וצריך לאפשר לתא להתקדם משלב לנטרל את פעולת החלבונים שכבר לא נחוצים לנו
מקופלים נכון, השאר זקוקים ER-מהחלבונים שנכנסים אל ה 20%רק
לעזרת שפרונים כדי להתקפל.
Cell cycle ם צורך.חלבונים שונים מופיעים בשלבים שונים של חיי התא, לא נרצה להשאיר בתא חלבונים שכבר אין בה
פקטורים של בהשפעתלמשל קינאז שפועל גם יש פקטורים שגורמים למיטוזה, בסוף המיטוזה נרצה להפטר מפקטורים אלו. , ולפי פקטורים של השלבים השונים הוא יזרחן G1פקטורים של שלב בהשפעת מיטוזה וגם
חלבונים אחרים.יהיו פעילים בו זמנית, אחרת יהיה הדגש כאן הוא שלא נרצה שחלבונים של שלבי תא שונים
מצב קצת כאוטי בתא. במעבר משלב לשלב על התא לפרק באופן פעיל את החלבונים שרלוונטים לשלב הקודם.
רוב החלבונים שמקדמים את מחזור התא משלב לשלב הם קינזות או אנזימים שעושים
יוביקינציה )נלמד בהמשך השיעור(.
.proteinB-ו proteinA: למשל חלבונים שפועלים בקומפלקסחלבונים מסוג זה שלא נקשרו בקומפלקס )לא מצאו בן זוג( משאירים את אתר הדגרדציה
שלהם חשוף וכך אפשר לפרק אותם לחומצות אמינו.
לחץ למולקולרית , ואין חלבונים בודדים ששטים בתא בחוסר מעש )הגרסה ה1:1בדרך זו נוודא שחלבונים אלו מיוצרים ביחד של (.חתונה

61
: במצב של הרעבה הגוף צריך להתמודד כך שלא נתמוטט, ומתחיל לפרק מקורות אנרגיה זמינים. בהתחלה אלו חזור של אנרגיהימליפידים ובהמשך אלו יהיו גם חומצות אמינו. פירוק חומצות אמינו במצב של הרעבה מאפשר גם ייצור של חלבונים נחוצים וגם
ז יפורקו החלבונים שהכי פחות חשובים לתא באותו רגע(.לצורך הפקת אנרגיה )א
Antigen presenting system .כל תא שמייצר חלבונים צריך להציג מדגם : המערכת החיסונית צריכה להבחין בין עצמי לבין זר, MHC class2-ו MHC class1משתמשים בחלבונים שנקראים לשם הצגה זו לרוב מהחלבונים שהוא מייצר מחוץ לממברנת התא.
מציגים מחוץ לממברנת התא חלקי חלבונים שמיוצרים בתוך התא. כך המע' החיסונית יכולה יושבים על הממברנה וחלבונים ששני לזהות תאים שמציגים חלבונים של חיידק או וירוס ולתקוף את התא.
האם מדובר בעצמי או בזר.שמציגים פפטידים אל מחוץ לתא וקובעים MHC-תאי המע' החיסונית ניגשים אל ה וירוס שהדביק את התא גורם לתא לייצר את החלבונים של אותו הוירוס, וחלק מהם יעברו פירוק ויוצגו אל מחוץ לתא.
אורך חיי חלבון
- Genetic causes :הקוד הגנטי מתורגם לרצף של חלבון שמכיל בתוכו את המידע לגבי אורך חיי התא
- Quality :באיכות גבוהה לא חייבים לפרק אותו, אם הוא פגום אז הוא יפורק איכות: אם החלבון
- Functional causes :צורך: אם יש צורך בחלבון הוא לא יפורק, ואם הוא סיים את תפקידו ואין בו צורך אז הוא יפורק
1. PEST Domain חייבות להיות בסדר זה, זה פשוט אזור בחלבון שמכיל . לא Pro(P), Glu(E), Ser(S), Thr(T)דומיין שמכיל את חומצות האמינו הבאות:
את חומצות האמינו הללו בריכוז מאוד גבוה. חלבונים בעלי אתר זה עוברים דגרדציה הרבה יותר מהר.
מצאו שעם הסרת רצף זה מהחלבון החלבון נשאר בתא לאורך זמן משמעותית רב יותר.2. N-end rule
חצית חיים של החלבון. של החלבון קיים סימון לזמן מ Nבקצה 20זמן מחצית החיים הוא שייר צד יציבים Met(M), Ser(S), Ala(A), Thr(T), Val(V), Gly(G) אם מופיעות ח' האמינו -
VG MAST שעות.
או דקות 3זמן מחצית החיים הוא שיירי צד לא יציבים Phe(F), Leu(L), Asp(D), Lys(K), Arg(R) האמינו אם מופיעות ח' - DR KLF פחות.
CHX ,חומר שמעכב סינתזת חלבונים.ציקלוהקסמיד :
נקשר בתוך הריבוזום ותוקע אותו כך שלא מתאפשר המשך התרגום. אנטיביוטיקה כנגד ריבוזום איקריוטי. נוצר ריבוזום עם שרשרת חלקית של חומצות אמינו.
ולא יתקבלו חלבונים חדשים. mRNA-הריבוזום עדיין יהיה בתחילת הנרצה לעצור את הסינתזה החדשה של חלבונים. נדגום כל זמן מה את כמות החלבון וכך נדע מהו Ribosome profilingאם נבצע
רצף הדגרדציה )משום שאין ייצור, רק פירוק של חלבונים(. החלבון מתרחשת בשלב מאוחר יותר. שהירידה בכמות Ribosome profiling-רואים ב PEST Domain-כשמסירים את ה
איך מפרקים חלבונים?
לחתוך חלבונים.פרוטאזות. חלבונים שיודעים סוגי פרוטאזות:
שימוש בהתקפה נוקלאופילית כדי לחתוך את הסובסטרט. : האתר הפעיל שלהם מכיל סרין.סרין פרוטאזות -
בהידרוליזה לפירוק הקומפלקס, וחומצות אמינו ישתחררו. מצב ביניים בו האנזים קשור לשרשרת של החלבון, ואז שימוש כתלות ברצף )צריך לזהות רצף ספציפי( יתבצע חיתוך.
שני סוגים של סרין פרוטאזות: בסרין פרוטאזות סרין היא חומצת האמינו הנוקלאופילית.

62
: אתר חיתוך עם שייר צד ארומטיכימוטריפסין -
חיובית: אתר חיתוך עם שייר צד טעון טריפסין -
אספרטאט. אחת תורמת ואחת מקבלת 2: האתר הפעיל מכיל אספרטאט פרוטאזות -
פרוטון.
פרוטון יעבור מהאספרטאט הראשונה אל בהנתן פפטיד או חלבון שנרצה לחתוך, הפפטיד, זה יערער את הקשר הפפטידי. לאחר מכן הפרוטון יעבור מהפפטיד אל
לפירוק של הקשר הפפטידי.האספרטאט השניה, וההידרוליזה הזו תגרום הקשר הפפטידי שיפורק יהיה ממוקם בין שתי האספרטאט.
הפרוטון הראשוני מגיע ממים. - Zinc פרוטאזות :
מאפשר תקיפה נוקלאופילית. (++Zn) אבץיון -
מבצע את התקיפהשנמצאת באתר הפעיל ממולקולת מים חמצן -
, וכך מסייע בתגובה זושביצעה את התקיפה ממולקולת המים פרוטוניםקושר באתר הפעיל גלוטמט -
מעורבות בדגרדציה של מטריקס חוץ תאי ברקמה.
:ציסטאין פרוטאזות -
לרוב מחוץ לתא.
S, ונותר באתר הפעיל ההיסטידיןשל ציסטאין מאבד פרוטון לטובת שייר (SH) הסולפט -
תוקף נוקלאופילית את הקשר הפפטידי( S)יון סולפט -
חשוב לדעת:
סוגי פרוטאזות בעלי ספציפיות שונה ואתרים פעילים שונים, ולכן נבדלים זה מזה בחלבונים אותם הם יכולים לחתוך.הרבה לפרוטאזות יש הרבה תפקידים חשובים, בין היתר גם במצב של הרג תא.
אנזים אחר חותך את הרצף , כשיש להן רצף שמעכב אותן. כדי להפעיל את הפרוטאזה pre-proteinפרוטאזות לרוב יגיעו במצב של
המעכב. המטרה היא שלא לסכן את התא, הרי פרוטאזות יכולות לפרק את כל החלבונים בתא ולכן עליהן להיות מאוד מבוקרות.
?איך בוחרים מי יעבור דגרדציה
:ציה של חלבוןדסמנים שמובילים לדגר
פוספורילציה-דה -
נזק לחלבון -
לדגרדציה חיתוך החלבון כך שיחשף אזור שמסמן -
אוקסידציה שמסמנת דגרדציה -
קיפול שגוי של החלבון כך שיחשף אזור שמסמן לדגרדציה -
המולקולה יוביקיטין
, זהו חלבון שמסמן חלבונים לדגרדציה.ע"י שני חוקרים בטכניון שזכו על זה בפרס נובל י מספר שניםהתגלתה לפנמולקולה ש
חומצות אמינו. מה שמיוחד בה הוא שהיא מתחברת גם לחלבון המטרה )קישור קוולנטי( וגם לעצמה וכך 76 -מולקולה מאוד קטנה יוצרת שרשראות של יוביקיטין.
יחידות יכולה לסמן 4באורך לפחות רק שרשרת של יוביקיטיןמשמשת רק לסיגנל. יוביקיטין יחידה היא לא סמן לדגרדציה, מולק' יוביקיטין יחידות, ועדיין הוא לא יעבור דגרדציה משום שהן לא 100)למשל, יכול להיות חלבון שמחוברות אליו .לדגרדציה
בשרשרת(. לרוב היוביקיטין נקשרת אל מולק' ליזין חשופות בחלבון.

63
כמה אתרים שונים שיכולים לשמש לחיבור של יוביקיטין אחת -ביקיטיןסוגים שונים של חיבורים בין מולקולות יויש כמה לרוב שולחת 48למשל שרשרת של ליזיןיש משמעות שונה לשימוש בחיבורים שונים, מדובר בקוד שטרם פוענח. ליוביקיטין אחרת.
שימו לב שיש גם .זה של התאלשיפור תרגום/מערכת חיסונית/אנדוציטו היא לרוב סיגנל כלשהו 63לדגרדציה ושרשרת של ליזין שמסמנות דגרדציה. 63שרשראות של ליזין
אבל זה יותר מורכב מזה... משערים שיש קידוד שמסמן גם את קצב הדגרגציה ועוד. יש שרשראות שבנויות מרצף של חיבורים (, ואנחנו לא יודעים לפענח אותן.63ואז ליזין 11ואז ליזין 48באזורים שונים )ליזין
מאוד ארוכה של יוביקיטין גורמת לחלבון לנוע לאט יותר בהרצה בג'ל.שרשרת
: יוביקיטין בודדת מחוברת לחלבון.מונויוביקיטינציה : הרבה יוביקיטינים בודדות מחוברות לחלבון.יוביקיטינציה-מולטי
: שרשרת של יוביקיטין מחוברת לחלבון.יוביקיטינציה-פולי
עמים שרשרת היוביקיטין תהיה במקום פנימי בחלבון, ואז אמנם הוא מכיל את הסמן לפ: מיקום השרשרת במבנה החלבוןלדגרגדציה אך הסמן לא יזוהה והחלבון לא יפורק. אם החלבון יעבור דנטורציה ויהיה בקיפול שאיננו הקיפול הטבעי שלו שרשרת
יפורק.יזוהה על ידי מערכת היוביקיטין והיוביקיטין תהיה חשופה והחלבון עמים שינוי הקונפורמציה שיגרום לשרשרת היוביקיטין להיות חשופה יתרחש בעקבות זרחון.לפ
תהליך הקישור של יוביקיטין לחלבון:, שאר ATPליוביקיטין. זה השלב היחיד שדורש AMP, קשירת קוולנטית של ATPמתרחשת תוך השקעה של אדנילציה ליוביקיטין
ת היוביקיטין הפעיל.אהריאקציות יהיו תלויות באנזימים שיתפסו ציה אחרת.ד( אלא במערכת דגרATPאם אין מספיק אנרגיה והתא במצוקה, כנראה שלא יהיה שימוש במערכת היוביקיטין )שצורכת
E1 הוא האנזים שמבצע את האקטיבציה של יוביקיטין. יש לו ציסטאין באתר הפעיל ואתר שקושרATP לא באופן קוולנטי(. ה(-ATP .ATP-. שחרור הE1תעבור הידרוליזה ותתרחש קשירה של היוביקיטין לציסטאין של
זהו תהליך רב שלבי ומאוד מבוקר. פי.למה כל כך הרבה? כדי שזה יהיה מאוד ספצי .E1, E2, E3המערכת משתמשת באנזימים .E2אל E1-מ Ubiמעבירים את
E2 נקשר אלE3. E3 חלבון המטרה וגם את קושר כסובסטרט אתE2 כשהוא קשור אלUbi .מאוקטבת
:E3שני סוגים של רטוט(ש)זה מה שרואים ב ואז מעביר אותו לסובסטרט Ubiאחד תופס את -
אל הסובסטרט מבלי לתפוס אותו Ubiהשני מעביר את -
המאוקטב הועבר אל הסובסטרט. Ubi-כך או כך, ה מאבד את האפיניות שלו לחלבון, והחלבון משוחרר. E3כשמגיעים לשרשרת ארוכה מספיק
(.Ubiשלב הראשוני של האקטיבציה של מזרז את E1-)שימו לב של

64
:E1, E2, E3בקרה כתוצאה משימוש בשלושת האנזימים מידה של סלקטיביות שנובעת מהקומבינטוריה של שלושת האנזימים.
E1 מזהה כמה סוגים שונים שלE2 ,E2 מזהה כמה סוגים שונים שלE3. השילוב בין סוגי האנזימים יקבע מה יהיה הטיפול בחלבון.
E3 מאוד ספציפי בנוגע ל-E2 והוא לא יקשור סוגים אחרים של שהוא יכול לקשור ,E2. E3 ים את עצמו למגוון של סובסטרטים.מתא
פוספורילציה כדי לחקור את נושא הדגרדציה של חלבונים. ניתן לבצע עיכוב של הפרוטאוזום
ראו כי חלבון שעשו לו פוספורילציה נשאר בכמות גבוהה בתא, הכמות שלו יציבה. הפוספט מגן על החלבון מפני פירוק.
גליקוזילציה להוסיפו לחלבונים זה סימן שיש מספיק אנרגיה ואין צורך במיחזור של חומרים בתא.כשיש מספיק סוכר כדי
כך הוספת סוכר יש לסוכרים גם תפקיד, כמובן, אך הם מהווים סמן מטבולי בתא שאין צורך בפירוק חלבונים לשם הפקת אנרגיה. מגנה על החלבון מפני פירוק.
פרוטאוזום
רוב החלבונים המסיסים ורוב החלבונים במסלול פירוק חלבונים זה יפורקו התאיים.
ת קטליטית של פרוטאז, פירוק חלבונים.פעילובצורה של גליל, מבנה יחסית סימטרי, החלבון יכול להיכנס או לצאת בנוי
משני הצדדים, אך ברגע שנכנס מצד אחד הוא חייב לצאת מהצד השני.
יוביקיטין החלבון יקשר כשחלבון מסומן בשרשרת של לפחות ארבע מולקולות אל הפרוטאוזום ויעבור פירוק.
במבט מעל התעלה סגורה ולא יכולים להיכנס אליה פפטידים.
20S ה-core. עושה את הפירוק עצמו ושומרת על תתי יחידות ביטא בחלק הפנימי של המבנה.
מסוים של חומצות אמינו וגם במידה מה לפי תתי יחידות ביטא הן מעין מספריים שנמצאות בתוך הפרוטאוזום וחותכות לפי אורך ספציפיות )חלקן דומות קצת לטריפסין פרוטאזות(.
הפרוטאוזום מיוצר כך שהאתרים הקטליטיים פונים כלפי פנים ולא יפעלו על אף חלבון שלא יעבור את האתרים הרגולטוריים. מהצד השני של הפרוטאוזום יצאו פפטידים קצרים.
פרוטאזות מזהים רצפים שונים.סוגים שונים של יש שני מעגלים של תתי יחידות ביטא, כל מעגל בנוי משבעה סוגים שונים של תתי יחידות ביטא.
אין אף יחידה ביטא פעילה לפני שנוצר המבנה של החבית, לפני שהמבנה הזו נוצר תתי היחידות ביטא מכילות שרשרת שמונעת את פעילותן.
19S cap :היא זו שקושרת את החלבון ומאפשרת את כניסתו לפרוטאוזום.מקנה ספציפיות לפרוטאוזום יחידה רגולטורית .
יש צורך לפתוח את החלבון מהקיפול שלו טרם כניסתו אל הפרוטאוזום. -להיכנס לתוך יחידה שגורמת לפתיחה של החלבון חזרה למבנה ראשוני, רצף של חומצות אמינו שיכולות, cap-שפרונים בתוך ה
לפעילותם וחלק לא. לא כל השפרונים יכולים ATPחלק מהשפרונים דורשים הפרוטאוזום. מרגע זה יתחיל הפירוק של החלבון. לפעול על כל החלבונים.
שרשרת של יוביקיטין.זיהוי - ישנם אנזימים שמשחררים את היוביקיטין חזרה לתא, לשימוש חוזר. -
ה של הפרוטאוזום ומאפשר כניסה של חלבון פנימההוא זה שמבצע את הפתיח cap-ה -
הסוגים של תתי יחידות ביטא, אפשר שיעוכבו רק חלק מהן ואז בשלב מסוים חלבון 7מעכבי פרוטאוזום: לא חייבים לעכב את כל כלשהו יתקע בתוך הפרוטאוזום וימנע ממנו להמשיך לפעול.
20 S Proteasome
(yeast) closed state
two views PDB 1JD2

65
, עוד לא ברור מה תפקידם, רק יודעים שהם לא מבצעים דגרדציה לחלבונים עם שרשרת 19S-capמצאו פרוטאוזומים שהם חסרי .11S-capאחר, capיוביקיטין. יש להם
:ליזוזום
במסלול פירוק זה יפורקו רוב החלבונים הממברנליים. רוב החלבונים מגיעים מאנדוציטוזה.
חלק מהחלבונים הם מהציטוזול.
חלבונים ולפרק אותם בתוכן בהפעלת חומציות ופרוטאזות.: יכולות לבלוע וסיקולות
12( שיעור20/06/18)
אפופטוזיס ומוות תאי
למה תאים צריכים למות?
כימי: חשיפה לכימיכלים רעילים שגורמים נזק לתא נזק -
נזק פיזי: תאים מתים בעקבות מכה חזקה -
יוצא החוצה ולא ניתן להשתמש בו. כל ATP: מוות לא מסודר, מוות בתאונה. למשל פיצוץ של התא: ממברנות שבורות, נקרוזיס הרכיבים בתא נשברים באופן אקראי.
תאים שכנים יכולים להיפגע )למשל, מפרוטאזות שיוצאות מהתא המפורק ומתפזרות באזור(.
התא, אין שבירה של הממברנות התאיות. : הריגה מכוונת של תא. מכווצים אתאפופטוזיס .ATPאפופטוזיס דורש אנרגיה, דורש
המת כדי שחלקיו לא יסתובבו בגוף: התא הופך לבועיות קטנות שמסומנות סימון למע' החיסונית לאסוף את התא )פאגוציטוזה( בדגלים שהמע' החיסונית יכולה לזהות.
.DNA-פירוק של ה וון סיטואציות.ניתן להשתמש באפופטוזיס במג
מוות אלטרואיסטי: מוות מסודר שלא פוגע בסביבה.
אפופטוזיס בנקודות: , כל האברונים מתכווצים.כיווץ של התא .1
מטרים לכל תא בגוף(. 9. זו מולקולה מאוד גדולה )בערך והכרומטין DNA-ה חיתוךוכיווץ .2
ורדיקלים חפשיים יגרמו לתא להגיע לנק' האל חזור ATPשינויים במיטוכונדריה: המיטוכונדריה עוזרת למות התא. שחרור .3 של המוות התאי.
יצירת בועיות: הממברנה נחתכת כך שכל חתיכה עוטפת חלק ממה שהיה התא השלם. חתיכות קטנות שיוכלו להיבלע .4 .בהמשך
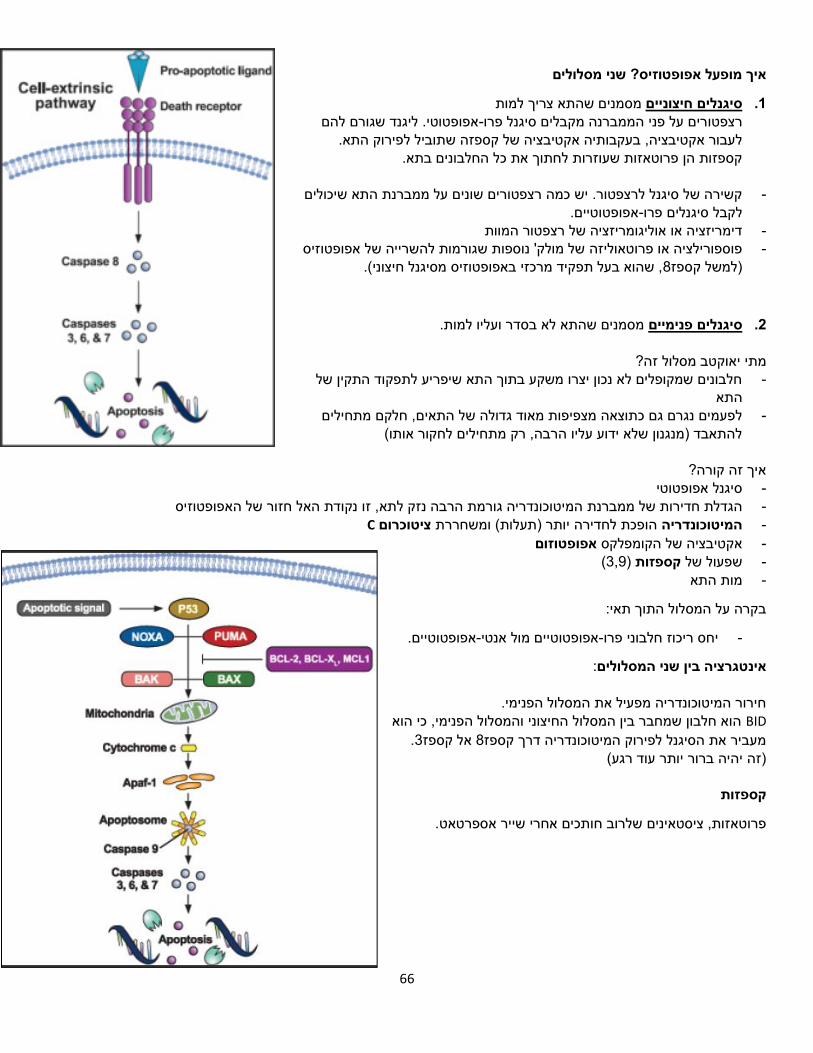
66
שני מסלולים איך מופעל אפופטוזיס?
מסמנים שהתא צריך למות סיגנלים חיצוניים .1
אפופטוטי. ליגנד שגורם להם -רים על פני הממברנה מקבלים סיגנל פרורצפטו שתוביל לפירוק התא.קספזה לעבור אקטיבציה, בעקבותיה אקטיבציה של
קספזות הן פרוטאזות שעוזרות לחתוך את כל החלבונים בתא.
קשירה של סיגנל לרצפטור. יש כמה רצפטורים שונים על ממברנת התא שיכולים - אפופטוטיים.-ים פרולקבל סיגנל
דימריזציה או אוליגומריזציה של רצפטור המוות -
של מולק' נוספות שגורמות להשרייה של אפופטוזיספוספורילציה או פרוטאוליזה - .(, שהוא בעל תפקיד מרכזי באפופטוזיס מסיגנל חיצוני8)למשל קספז
.מסמנים שהתא לא בסדר ועליו למות סיגנלים פנימיים .2
מסלול זה?יאוקטב מתי
חלבונים שמקופלים לא נכון יצרו משקע בתוך התא שיפריע לתפקוד התקין של - התא
לפעמים נגרם גם כתוצאה מצפיפות מאוד גדולה של התאים, חלקם מתחילים - להתאבד )מנגנון שלא ידוע עליו הרבה, רק מתחילים לחקור אותו(
איך זה קורה?
סיגנל אפופטוטי -
המיטוכונדריה גורמת הרבה נזק לתא, זו נקודת האל חזור של האפופטוזיסהגדלת חדירות של ממברנת - Cציטוכרום משחררת הופכת לחדירה יותר )תעלות( ו המיטוכונדריה -
אפופטוזוםאקטיבציה של הקומפלקס - (3,9) קספזותשפעול של - מות התא -
בקרה על המסלול התוך תאי:
וטיים.אפופט-אפופטוטיים מול אנטי-יחס ריכוז חלבוני פרו -
:אינטגרציה בין שני המסלולים
חירור המיטוכונדריה מפעיל את המסלול הפנימי.BID הוא חלבון שמחבר בין המסלול החיצוני והמסלול הפנימי, כי הוא
. 3זפאל קס 8זפמעביר את הסיגנל לפירוק המיטוכונדריה דרך קס )זה יהיה ברור יותר עוד רגע(
קספזות
שלרוב חותכים אחרי שייר אספרטאט.פרוטאזות, ציסטאינים

67
אנזימים, כמו כל שאר הפרוטאזות.-מגיעים כפרו
לאחר שעוברים אקטיבציה הם יוצרים קומפלקס והופכים לפעילים.
הן הרבה יותר ספציפיות, לא (Initiator) קספזות מתחילות
(.9-, ו8, 2יכולות לפרק את כל מרכיבי התא )אפופטוזיס. ניתן לחזור אחורה השפעול שלהן עדיין לא מחייב
ולעצור את התהליך.
מפרקות חלבונים רבים בתא (Effector) קספזות אפקטוריות (.7-, ו6, 3בצורה הרבה פחות ספציפית )
ברגע שהפעלנו את האפקטוריות, אין דרך חזרה, מתחיל והתא בוודאות ימות. DNA-פירוק של ה
(Inflammatory)
T (1,4,5.)מטורציה של תאי פעילות במיוחד במע' החיסונית,
מנגנונים. 3איך מפעילים את הקספזות? )בדוגמה שבמצגת, חיבור של דומיין כדי שאפשר יהיה לייצר קומפלקס-אנזים שחותך את הפרו: חיתוך פרוטאוליטי .1
טוטי כלשהו.אפופ-החיתוך מתבצע ע"י קספז או ע"י חלבון פרו (.P20-ו P10הדומיינים
2. Induced proximity : .אותו סוג )לדוגמה, קספזות מ-רצפטור שיכול לגייס כמה פרוגורם גם הוא בסופו של דבר לחיתוךם עוד לא נחתך מהם איקשר ויתנו חלבון פעיל, אפילו . כשהם קרובים זה לזה האתר הפעיל(8במצגת מדובר על קספז
הקספזות עושות אקטיבציה לעצמן. באופן סינטטי. P20-ו P10יצרנו חיבור של הדומיינים דומיין.-הפרו
ביצירה של 9קספז: ייצור קומפלקס, למשל עם אוליגומריזציה .3 .ATPהוא חלבון שדורש Apaf1. האפופטוזום
הוא זה שאליו יקשרו שאר חלבוני הקומפלקס. Apaf1 החלבוןדרוש ATP-יתחבר לקומפלקס הזה הוא יהיה פעיל. ה 9כשקספז
. האפופטוזום הוא הקומפלקס הכי Cעם ציטוכרום Apaf1לקישור של ידוע, אבל כבר התגלו קומפלקסים נוספים.
מה עושות קספזות פעילות?
יכול 9חיתוכים שגורמים לאקטיבציה: למשל אקטיבציה של קספזות. קספז - .3לחתוך את קספז
חיתוך חלבונים והפיכתם ללא פעילים -
הרס פונקציונליות: הרס של מבנים שלמים )כמו חיתוכים שגורמים ל - המיקרוטובולים בתא(
מסונתז, CAD-, נוקלאז. תוך כדי שCADהוא מעכב של DNA :ICADהרס -ICAD נקשר אליו, עוזר לו לעבור קיפול נכון ומעכב אותו. ברגע
CADשמשחררים את
(ICAD נוצרת דימריזציה של )עובר חיתוךCAD חיתוך של , שיבצע בגרעין .DNA-ה
CAD חותך את ה-DNA לכן הבין הנוקלאוזומים ,-DNA נחתך ב-pattern מסוים.

68
Bcl-2משפחת חלבוני
-אפופטוטים )תופסים חלבונים פרו-כוללת גם חלבונים שהם אנטי Bcl-2 משפחתאפופטוטיים -אפופטוטיים ולא מאפשרים להם לבצע את תפקידם( וגם חלבונים שהם פרו
)הפעלתם תגרום למוות תאי(, יש להם דומיינים דומים.הוא החלבון הראשון שהתגלה bcl2 החלבוןשימו לב לקטע המאוד מאוד מבלבל כאן:
)חשוב להבין אם שואלים אתכם על החלבון או על אפופטוטי-והוא אנטי חה זובמשפ המשפחה(.
למשל, כדי לפרק את המיטוכונדריה יש לחורר את הממברנה המיטוכונדריאלית.
שמשחררות מהממברנה המיטוכונדריאלית חומרים מפעילים תעלות אפופטוטיים: -פרו BCL2)שחותך את BCL-Xsוכן BAXלמשל .ורדיקלים חפשיים Cשונים דוגמת ציטוכרום
(3ע"י קספז .BCL-2למשל אפופטוטיים: יקשרו אל המשאבות וינטרלו את פעולתם.-אנטי
אפופטוטיים.-אפופטוטיים לבין החלבונים האנטי-השאלה האם התא ימות או לא תלויה באיזון שבין החלבונים הפרו
:אפופטוזיסחקר מתודולוגיות שונות ב
, ליפידים שנמצאים לרוב בממברנה ישתנה הרכב הממברנהכשתא עומד לעבור אפופטוזיס הפנימית יצאו אל הממברנה החיצונית. זהו סיגנל למע' החיסון לבלוע את התא הזה.
כלומר, בתאים אפופטוטיים תשתנה האוריינטציה של הליפידים בממברנה.
Tunel assayה חיתוךאחר במעבדה : מעקב-DNA מבוצע בשיטה של סימון קצוות פתוחים ב-DNA נוכל לראות . בחלבון פלורסנטי אף פעם לא אמור להיות עם קצוות פתוחים. DNA-עובר חיתוך. ה DNA-המון נקודות פלורסנטיות כשה
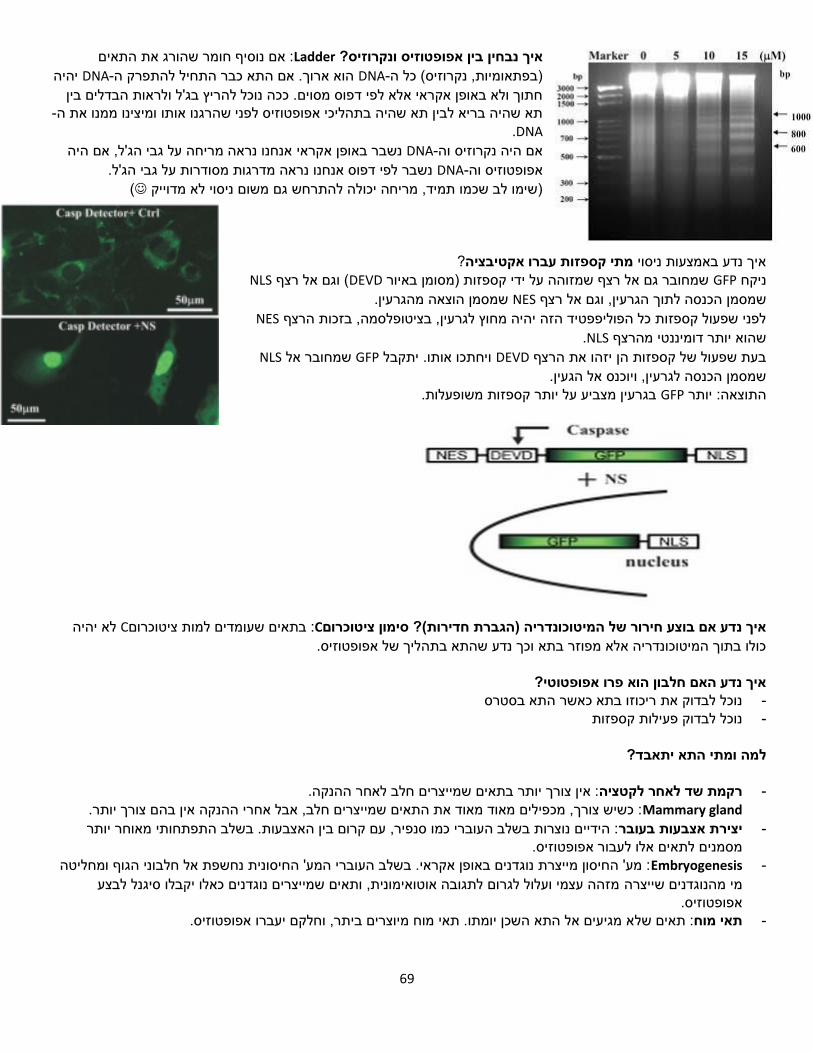
69
: אם נוסיף חומר שהורג את התאים Ladder איך נבחין בין אפופטוזיס ונקרוזיס?יהיה DNA-הוא ארוך. אם התא כבר התחיל להתפרק ה DNA-כל ה)בפתאומיות, נקרוזיס(
חתוך ולא באופן אקראי אלא לפי דפוס מסוים. ככה נוכל להריץ בג'ל ולראות הבדלים בין -תא שהיה בריא לבין תא שהיה בתהליכי אפופטוזיס לפני שהרגנו אותו ומיצינו ממנו את ה
DNA. היה אנחנו נראה מריחה על גבי הג'ל, אםנשבר באופן אקראי DNA-אם היה נקרוזיס וה
נשבר לפי דפוס אנחנו נראה מדרגות מסודרות על גבי הג'ל. DNA-הו אפופטוזיס ()שימו לב שכמו תמיד, מריחה יכולה להתרחש גם משום ניסוי לא מדוייק
?מתי קספזות עברו אקטיבציהאיך נדע באמצעות ניסוי NLS( וגם אל רצף DEVDשמחובר גם אל רצף שמזוהה על ידי קספזות )מסומן באיור GFPניקח
שמסמן הוצאה מהגרעין. NESשמסמן הכנסה לתוך הגרעין, וגם אל רצף NESלפני שפעול קספזות כל הפוליפפטיד הזה יהיה מחוץ לגרעין, בציטופלסמה, בזכות הרצף
.NLSשהוא יותר דומיננטי מהרצף NLSשמחובר אל GFPויחתכו אותו. יתקבל DEVDבעת שפעול של קספזות הן יזהו את הרצף
שמסמן הכנסה לגרעין, ויוכנס אל הגעין. בגרעין מצביע על יותר קספזות משופעלות. GFPהתוצאה: יותר
לא יהיה Cם למות ציטוכרוםבתאים שעומדי :Cסימון ציטוכרוםאיך נדע אם בוצע חירור של המיטוכונדריה )הגברת חדירות(?
כולו בתוך המיטוכונדריה אלא מפוזר בתא וכך נדע שהתא בתהליך של אפופטוזיס.
איך נדע האם חלבון הוא פרו אפופטוטי? נוכל לבדוק את ריכוזו בתא כאשר התא בסטרס -
נוכל לבדוק פעילות קספזות -
מתי התא יתאבד?למה ו בתאים שמייצרים חלב לאחר ההנקה.: אין צורך יותר רקמת שד לאחר לקטציה -
Mammary gland.כשיש צורך, מכפילים מאוד מאוד את התאים שמייצרים חלב, אבל אחרי ההנקה אין בהם צורך יותר : : הידיים נוצרות בשלב העוברי כמו סנפיר, עם קרום בין האצבעות. בשלב התפתחותי מאוחר יותר יצירת אצבעות בעובר -
ר אפופטוזיס.מסמנים לתאים אלו לעבו
- Embryogenesis מע' החיסון מייצרת נוגדנים באופן אקראי. בשלב העוברי המע' החיסונית נחשפת אל חלבוני הגוף ומחליטה :מזהה עצמי ועלול לגרום לתגובה אוטואימונית, ותאים שמייצרים נוגדנים כאלו יקבלו סיגנל לבצע הנוגדנים שייצרה מי מ
אפופטוזיס. א מגיעים אל התא השכן יומתו. תאי מוח מיוצרים ביתר, וחלקם יעברו אפופטוזיס.: תאים שלתאי מוח -

70
סוגים נוספים של מוות תאי מתוכנן: הכרומטין והתא כולו הופך לוסיקולות.בעיה בדחיסה של , Autophagyמוות -- Comnificationדי לקבל את שכבת העור שמגנה , תהליך במסגרתו יש צורך שתאים ימותו לשם פונקציונליות. למשל תאי עור. כ
הם מוותרים על הגרעין ועל אברונים נוספים ויוצרים שכבת הגנה של תאים מתים בחלק -עלינו יש צורך שתאי העור ימותוהחיצוני של העור. שימו לב שבמוות זה התא הופך לשטוח ועם הרבה קרטין כדי שיהיה חזק, אבל התא לא מתפרק. בעצם
התא הזה הוא מצב של מוות )לא יכול להתחלק ולבצע שלל תהליכים(. המצב הפונקציונלי של
אפופטוזיס מאפשר לנו להיפטר מתאים לא רצויים: תא שהודבק בוירוס -
תגובה אוטואימונית - DNAתא עם נזקי -
תא סרטני -
איך?
- Granzymesפרוטאזות שמשוחררים על ידי גרנולות של תאי -סריןות תאים. : חירור ממברנT ציטוטוקסים ותאיNK. זו דרך של מע' החיסון לחסל תאים סרטניים או תאים שנדבקו בוירוס )וזוהו ככאלה על ידי מע' החיסון(.
: התא יכול לנסות לתקן, ואם התיקון לא יהיה מספק או שהנזק מאוד רחב יתרחש אפופטוזיס. אם יש יותר DNA-עודף נזק ל - יס, ויתרחש נקרוזיס.מדי נזק התא לא יוכל להפעיל אפופטוז
כדי לא לאפשר לוירוס יומת : וירוס הוא לא עצמאי, הוא משתלט על מערכות התא כדי לשרוד. תא שהודבק בוירוס אבולוציה-קו -מה אפופטוטיים בניסיון לגרום לתא להמשיך לשרוד וכך להתרבות.-יש וירוסים שפיתחו מנגנונים אנטי להדביק תאים נוספים.
אפופטוטיים.-אפופטוטיים לבין החלבונים האנטי-יזון בין החלבונים הפרויקרה? זה תלוי בא
שימו לב שאם הנזק עדיין הפיך התא יכול לחזור בו ולא למות, זה תלוי בכמה נזק נגרם וכמה זמן נמשך הסיגנל לאפופטוזיס.
אפופטוזיס כמנגנון הגנה מפני מחלות:
שמתו. : עודף תאים Tissue atrophyתגובה מוגזמת
רקמה לא מתפקדת -
עור דק מאוד -
ועוד.. -
: תאים שהיו צריכים למות ולא הומתו Hyperlasiaתגובה מועטת
מצב סרטני -
מצב אוטואימוני -
ועוד.. -
ף !-ו-ס-ה בהצלחה במבחן
Related Documents











