




1
MICROBIOLOGIA prof. Antonio Farris Alessandro Bellotto Renato Sedda
CAP. 1 Microrganismi e microbiologia La microbiologia odierna è una scienza dinamica, con ramificazioni di fatto in tutte le scienza della vita. La microbiologia è lo studio dei microrganismi: numeroso gruppo di microscopici organismi tra loro diversi che possono esistere come singola cellula o come raggruppamenti di cellule. Essa studia anche i virus, microscopici ma non cellulari. Le cellule microbiche si distinguono da quelle animali e vegetali, che sono incapaci di vivere da sole in natura e possono esistere solo come parti di un organismo pluricellulare. Al contrario del macrorganismo, i microrganismi sono generalmente in grado di effettuare tutti i processi legati alle funzioni vitali di crescita, di generazione dell’energia e di riproduzione indipendentemente dalle altre cellule. La microbiologia studia i microrganismi ed è considerata una scienza di base e una scienza applicata. Come scienza biologica di base, essa offre alcuni importanti strumenti per capire i meccanismi dei processi vitali; le cellule microbiche condividono molte proprietà biochimiche con quelli degli organismi pluricellulari; questo aspetto, unito al fatto che i microrganismi possono crescere in colture di laboratorio ad alta densità cellulare, per effettuare studi di biochimica e genetica, li rende un modello di studio eccellente per la conoscenza delle funzioni cellulari negli organismi superiori. La cellula del lievito è utilizzata come modello di riferimento nella descrizione della cellula eucariotica. Come scienza applicata, la microbiologia si occupa di problemi pratici di grande importanza in medicina, agricoltura (per esempio la fertilità dei suoli e l’humus) e industria (soprattutto nella trasformazione). (fermentazione alcolica lieviti; fermentazione lattica batteri). Biosfera: spazio dove è possibile la vita, con T e P idonee. Le due condizioni fondamentali perché la vita sia possibile sono le sostanze nutritive e l’energia. La biosfera è un sistema aperto: arriva tutta l’energia necessaria senza che si preoccupi di riutilizzarla, perché la fonte, il sole, continua ad inviarla. L’energia, che viene captata dalle piante e trasformata in energia chimica, attraverso i livelli trofici viene trasformata in energia non più utilizzabile (calore). Per quanto riguarda le sostanze nutritive, la materia è sempre la stessa ed è presente in forme diverse. Le piante (autotrofe) necessitano di sostanze inorganiche, mentre gli eterotrofi di sostanza organica. La degradazione della sostanza organica, nel ciclo della materia, produce H2O, CO2 e sali minerali. I microrganismi fanno parte dell’anello che unisce la sostanza organica a quella inorganica. Altri organismi (mesofauna) fanno parte di tale anello: per esempio il lombrico (anche 14 quintali per ettaro di terreno) fa passare il terreno nel suo corpo degradandone il 25% e sminuzzandone il 75% . Tutti i microrganismi sono ecologicamente utili perché fanno parte del sistema vita; ma per l’interesse dell’uomo possono risultare utili (per es. i batteri acetici nella produzione dell’aceto) o dannosi (per es. i batteri che acidificano il vino e i patogeni). I microrganismi (per lo più eterotrofi) hanno un ruolo centrale nei confronti delle attività umane e di tutti gli aspetti che riguardano la vita sulla terra; in loro assenza nessuna forma vivente avrebbe potuto apparire o potrebbe oggi essere presente; basti pensare che l’ossigeno che respiriamo è il risultato dell’attività microbica (i cianobatteri o alghe azzurre sono i primi organismi aerobi comparsi sulla Terra e sono azoto fissatori che liberano ossigeno).

2
La cellula è l’unità fondamentale della vita. La membrana cellulare è la barriera che separa il compartimento interno dall’esterno e inoltre regola il flusso delle sostanze attraverso essa. Una struttura chiave è il nucleo (presente solo negli eucarioti, con membrana e più cromosomi) oppure il nucleoide (presente nei procarioti, senza membrana e un solo cromosoma circolare), dove sono localizzate le informazioni genetiche (DNA), e il citoplasma, dove sono presenti tutte le strutture deputate alla crescita e al funzionamento cellulare. Tutte le cellule sono costituite da almeno 4 componenti chimici: PROTEINE, ACIDI NUCLEICI, LIPIDI E POLISACCARIDI. La cellula è un sistema aperto che si relaziona con l’ambiente; in condizioni non idonee è in grado di rallentare o di bloccare il metabolismo, inoltre ci sono cellule che in condizioni sfavorevoli creano una struttura di resistenza: la spora. Una volta comparsa la prima cellula (circa 4 miliardi di anni fa) sono seguiti eventi come lo sviluppo e la divisione per formare popolazioni di cellule da cui l’evoluzione ha potuto operare selezioni per migliorare e diversificare, quindi è sorta l’enorme diversità di tipi cellulari tuttora esistenti. Dal momento che le cellule sono costituite dalle 4 classi di macromolecole e condividono aspetti comuni, si può ipotizzare che tutte le cellule discendano da un comune antenato.
I microrganismi ricavano Energia e trasmettono i caratteri alla progenie.

3
Caratteristiche dei sistemi viventi

4
Le cellule vivono in natura in associazione con altre cellule in una sorta di agglomerato chiamato popolazione. Le popolazioni sono quindi composte da gruppi di cellule correlate, generalmente derivate da una singola cellula per successive divisioni cellulari. Il luogo dove la popolazione microbica vive si chiama habitat. In natura raramente le popolazioni vivono isolate, eccetto in alcuni casi particolari, come nella fermentazione alcolica del mosto; esse convivono e interagiscono con altre popolazioni cellulari assemblate nelle comunità microbiche, consistenti di cellule plantoniche in ambienti acquatici, ma molto più spesso adese a formare biofilm su superfici viventi e non viventi. L’efficienza di una comunità microbica in un dato habitat è determinata in gran parte dalle caratteristiche fisico-chimiche dell’ambiente. L’ecosistema è l’insieme degli organismi viventi e delle caratteristiche chimico-fisiche del loro ambiente. Le proprietà di un ecosistema sono spesso sotto il controllo di attività microbiche. Nelle comunità microbiche le diverse specie di microrganismi sono responsabili di molti fenomeni, come per esempio le trasformazioni. Solo alcune trasformazioni sono specifiche, cioè operate da gruppi puri di microrganismi della stessa specie, come la nitrificazione (da ammonio ad N nitrico NO3, utilizzabile dagli organismi autotrofi). (Solo il riso riesce a utilizzare, in piccola parte, anche sali di ammonio NH4
+). Gli ecosistemi naturali sono regolati dall’attività dei microrganismi e per esempio nei pascoli la fertilità è garantita da essi (gli azoto-fissatori possono apportare anche 300 Kg N anno/ha). La mineralizzazione si ottiene anche con gli incendi, con la grossa differenza che si salta l’importante fase intermedia della formazione dell’humus, che è in grado di trattenere l’umidità e le sostanze nutritive, e che determina la struttura e la fertilità del suolo stesso. L’incenerimento, anche dei rifiuti, è il peggiore metodo di degradazione e smaltimento. Le cellule microbiche sono troppo piccole per essere viste a occhio nudo (può distinguere fino ad 1/10 di mm) ed è necessario l’uso del microscopio. In un cm quadrato di pelle ci sono 100mila microrganismi e nella testa sono presenti in maggiore numero. Il numero totale delle cellule microbiche sulla Terra, specificatamente dei procarioti (detti anche batteri), è dell’ordine di 5 x 1030 cellule. Le cellule procariotiche, nella loro infinitesima dimensione, costituiscono la maggior quota di biomassa presente sulla Terra e una fondamentale riserva di nutrienti essenziali per la vita. In un grammo di terra ci possono essere anche 1012 microrganismi. In un ml di mosto in fermentazione, ci sono un miliardo di lieviti.
5
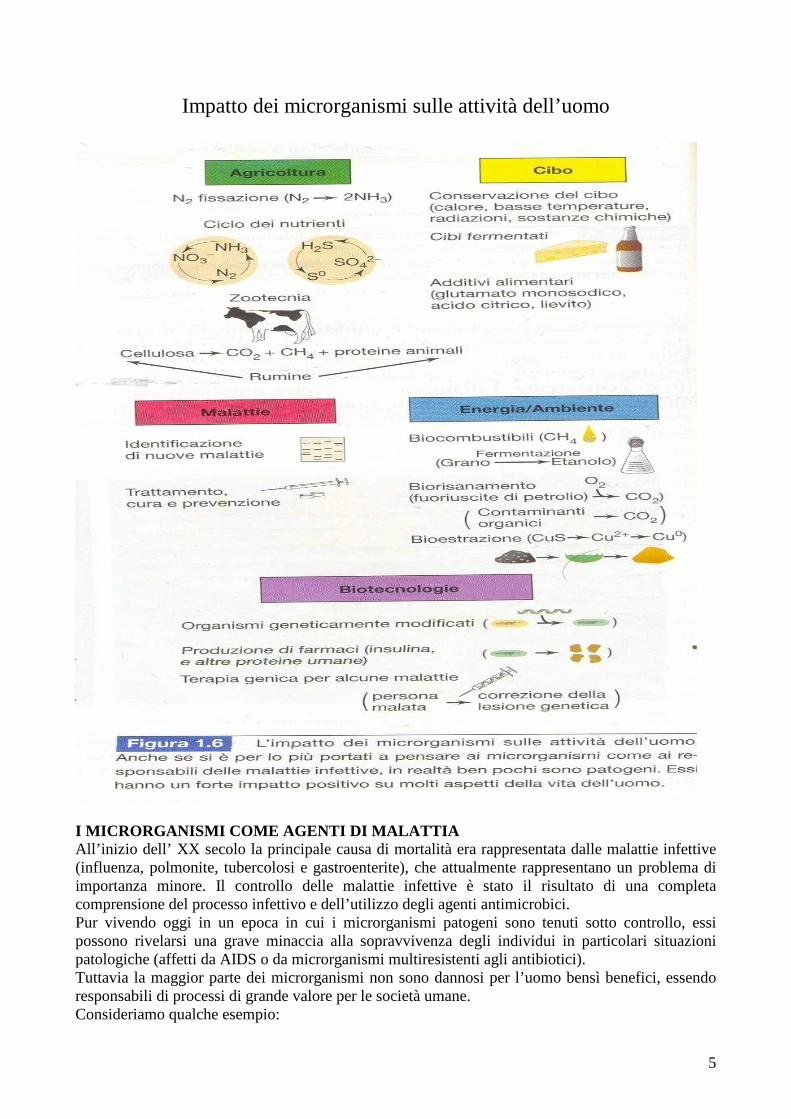
Impatto dei microrganismi sulle attività dell’uomo
I MICRORGANISMI COME AGENTI DI MALATTIA All’inizio dell’ XX secolo la principale causa di mortalità era rappresentata dalle malattie infettive (influenza, polmonite, tubercolosi e gastroenterite), che attualmente rappresentano un problema di importanza minore. Il controllo delle malattie infettive è stato il risultato di una completa comprensione del processo infettivo e dell’utilizzo degli agenti antimicrobici. Pur vivendo oggi in un epoca in cui i microrganismi patogeni sono tenuti sotto controllo, essi possono rivelarsi una grave minaccia alla sopravvivenza degli individui in particolari situazioni patologiche (affetti da AIDS o da microrganismi multiresistenti agli antibiotici). Tuttavia la maggior parte dei microrganismi non sono dannosi per l’uomo bensì benefici, essendo responsabili di processi di grande valore per le società umane. Consideriamo qualche esempio:

6
- I MICRORGANISMI E L’AGRICOLTURA Tutto il nostro sistema dell’agricoltura dipende in gran parte dalle attività microbiche; le leguminose sono piante che vivono in stretta associazione con una specie batterica che forma alle loro radici strutture chiamate noduli; in questi ultimi l’N 2 atmosferico è convertito in un composto azotato fissato che può essere usato dalle piante nel loro sviluppo. Tali piante sono in grado di dare al suolo 200-300 Kg N anno/ha. Altri gruppi microbici di fondamentale importanza in agricoltura sono quelli che intervengono nel processo digestivo dei ruminanti. I microrganismi svolgono inoltre un ruolo chiave nel riciclaggio di importanti nutrienti per la nutrizione delle piante, in particolare C, N, S, rendendoli accessibili alle piante. Tuttavia i microrganismi possono essere anche dannosi per piante e animali provocando infezioni. - I MICRORGANISMI E GLI ALIMENTI I microrganismi ricoprono una funzione fondamentale nell’industria alimentare. Il deterioramento delle derrate alimentari si riflette ogni anno in un danno economico enorme. L’industria alimentare (inscatolamento, congelamento e liofilizzazione) prepara gli alimenti in modo che non vengano danneggiati dai microrganismi. Tuttavia, non tutti i microrganismi sono dannosi per gli alimenti o per coloro che se ne nutrono, per esempio i prodotti lattiero caseari (formaggio, yogurt e burro) vengono lavorati, almeno in parte, dalle attività di alcuni microrganismi. Allo stesso modo crauti, sottaceti, salumi e insaccati devono il loro grado di maturazione all’attività microbica. I prodotti da forno e le bevande alcoliche sono il risultato dell’azione dei lieviti. I microrganismi sono utili nella conservazione della frutta, per esempio sono spruzzati sulla buccia delle arance per evitare lo sviluppo di muffe. Si è sviluppata una branca della microbiologia che studia e seleziona microrganismi detti “starter” per determinate finalità. - MICRORGANISMI, ENERGIA E AMBIENTE Gran parte del gas naturale (metano) è prodotto dall’attività microbica da parte dei batteri metanogeni. I microrganismi fototrofi utilizzano energia luminosa per produrre biomassa (energia conservata in organismi viventi). La biomassa microbica e molti dei prodotti di scarto (domestici, agricoli e di origine animale) possono essere convertiti in “biogas” come metano ed etanolo dall’attività di specifici microrganismi. I microrganismi possono essere utilizzati nel trattamento di composti inquinanti prodotti dalle attività umane (biorisanamento). Ne sono stati isolati molti in grado di degradare petrolio, solventi, pesticidi e altri composti tossici . - MICRORGANISMI E FUTURO Le biotecnologie si riferiscono all’uso di microrganismi (per lo più OGM) in processi industriali di larga scala. Le biotecnologie sono fortemente dipendenti dall’ingegneria genetica, disciplina che si occupa della manipolazione artificiale dei geni e dei loro prodotti. L’insulina umana (un ormone sintetizzato a bassissimo livello nei pazienti diabetici) può essere prodotto microbiologicamente inserendo il gene umano dell’insulina ingegnerizzato in un microrganismo. Quindi è evidente come i microrganismi esercitino una profonda influenza sulle società umane; PASTEUR affermò: “il ruolo dell’infinitamente piccolo, in natura, è infinitamente grande”.

7
LE RADICI STORICHE DELLA MICROBIOLOGIA La scoperta dei microrganismi è legata all’invenzione del microscopio. Hooke descrisse solo i corpi fruttiferi delle muffe nel 1664 (osservate con l’impiego di uno strumento con l’obiettivo collocato alla fine di un soffietto regolabile), ma la prima persona che vide i microrganismi in dettaglio fu l’olandese Anthony Van Leeuwenhoek, estraneo agli ambienti scientifici, che nel 1684 (20 anni dopo) costruì il primo rudimentale microscopio (la lente era montata nella piastrina di ottone adiacente alla punta della vite regolabile del fuoco e permetteva di ingrandire fino a 400 volte); egli riuscì a vedere i batteri e riportò le sue osservazioni in una serie di lettere indirizzate alla Royal Society di Londra; le sue osservazioni furono in seguito confermate da altri scienziati ma la comprensione della natura e dell’importanza dei microrganismi procedevano lentamente. Solo nel secolo XIX entrò in uso e si diffuse rapidamente un microscopio più efficiente. Ciò consentì una più accurata osservazione della cellula batterica e quindi la microbiologia si sviluppò come scienza. Un botanico tedesco Cohen cominciò a interessarsi all’uso del microscopio verso il 1850 per studiare lo sviluppo e la divisione di cellule vegetali e successivamente dei cianobatteri; inoltre si appassionò allo studio delle forme batteriche resistenti al calore, che lo portò alla scoperta del genere Bacillus e al processo di formazione delle spore. Egli introdusse inoltre l’uso del cotone per chiudere tubi e fiasche per evitare la contaminazione dei terreni di coltura sterili. Nel secolo XIX una delle controversie maggiori sorse rispetto alla teoria della generazione spontanea asserita da alcuni scienziati che affermavano che i batteri si generavano spontaneamente da materiale non vivente (es. cibo lasciato all’aria andava in putrefazione). Uno degli oppositori a tale teoria fu il chimico francese Luis Pasteur che dimostrò che i microrganismi trovati nel materiale in putrefazione derivavano dalle cellule microbiche presenti nell’aria e che queste erano presenti su tutti gli oggetti. Pasteur dedusse che trattando l’alimento in modo da distruggere ogni organismo vivente si sarebbe potuto ovviare alla putrefazione. Un semplice esperimento di Pasteur con la fiasca a collo di cigno fu risolutore della controversia intorno alla generazione spontanea. L’eliminazione dei batteri o di altri microrganismi presenti in o su un oggetto è un processo detto sterilizzazione. Koch mise a punto una serie di metodi per ottenere microrganismi in coltura pura in modo da isolare solo un tipo di microrganismi per poterlo studiare. Koch osservò che sulla fetta di patata esposta all’aria e successivamente incubata si sviluppavano colonie batteriche con forme e colori caratteristici; ipotizzò che ogni colonia derivasse da una singola cellula batterica che aveva trovato le condizioni nutrizionali per crescere e moltiplicarsi; ogni colonia rappresentava quindi una colonia pura. Nei substrati liquidi non è possibile evidenziare le singole popolazioni ed è necessario utilizzare un substrato solido; Koch fece crescere i microrganismi su terreni di coltura solidi e impiegò come agente solidificante la gelatina addizionata di nutrienti; la gelatina ha diversi svantaggi primo tra tutti diventa liquida con T maggiore di 22oC. Walter Hesse identificò un’agente solidificante più versatile grazie al suggerimento della moglie che utilizzava l’agar nella preparazione delle confetture di frutta; l’agar è un polisaccaride derivato dalle alghe rosse e rimane solido fino a 48oC.

8
Nel 1887 Petri inventò la piastra doppia; le due parti potevano essere sterilizzate separatamente dal terreno e, in seguito all’aggiunta di terreno fuso in una piastra, quella più grande poteva fungere da coperchio; le colonie che si formavano sulla superficie dell’agar rimanevano esposte all’aria e potevano essere sottoposte a manipolazioni successive. Oggi la piastra di Petri è di vetro sterilizzabile al calore o di plastica monouso. Beijerinch ha formulato il concetto di coltura di arricchimento : propose un metodo selettivo di isolamento, in cui venivano favoriti gruppi microbici specifici rispetto ad altri, solo agendo sulle condizioni colturali e di incubazioni. Egli isolò in coltura pura dei microrganismi del suolo. Sergej Vinogradskij era molto interessato alla microbiologia del suolo e scoprì i microrganismi chemio-autotrofi: isolò i batteri nitrificanti in coltura pura, dimostrando che il processo di nitrificazione (l’ossidazione dell’ NH3 a nitrati) era il risultato del metabolismo batterico e che i microrganismi utilizzavano per vivere l’energia sprigionata dalle reazioni di ossido-riduzione. La coltura di arricchimento in un tubo di vetro, chiuso da un tappo di gomma ad una estremità e all’altra con un tappo di cotone, fa si che nell’ambiente anossico si sviluppano batteri sporigeni come i clostridi, mentre all’altra estremità si sviluppano microrganismi aerobi (azotobacter).
9
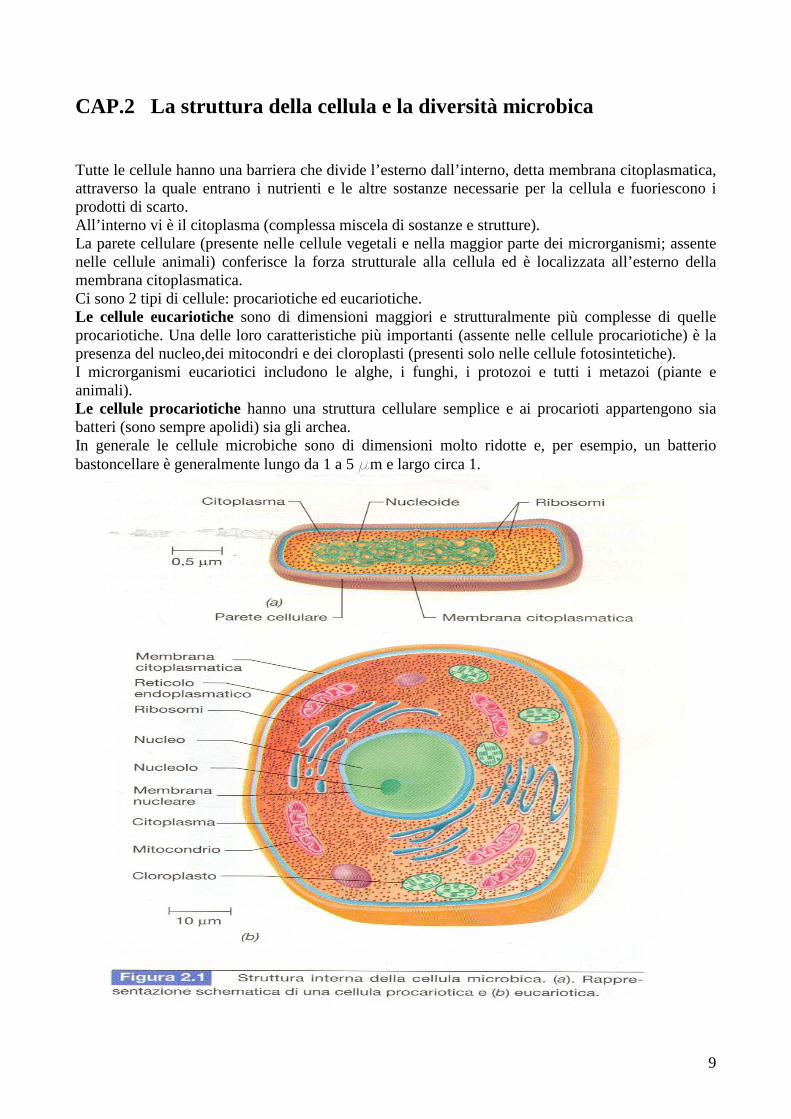
CAP.2 La struttura della cellula e la diversità microbica Tutte le cellule hanno una barriera che divide l’esterno dall’interno, detta membrana citoplasmatica, attraverso la quale entrano i nutrienti e le altre sostanze necessarie per la cellula e fuoriescono i prodotti di scarto. All’interno vi è il citoplasma (complessa miscela di sostanze e strutture). La parete cellulare (presente nelle cellule vegetali e nella maggior parte dei microrganismi; assente nelle cellule animali) conferisce la forza strutturale alla cellula ed è localizzata all’esterno della membrana citoplasmatica. Ci sono 2 tipi di cellule: procariotiche ed eucariotiche. Le cellule eucariotiche sono di dimensioni maggiori e strutturalmente più complesse di quelle procariotiche. Una delle loro caratteristiche più importanti (assente nelle cellule procariotiche) è la presenza del nucleo,dei mitocondri e dei cloroplasti (presenti solo nelle cellule fotosintetiche). I microrganismi eucariotici includono le alghe, i funghi, i protozoi e tutti i metazoi (piante e animali). Le cellule procariotiche hanno una struttura cellulare semplice e ai procarioti appartengono sia batteri (sono sempre apolidi) sia gli archea. In generale le cellule microbiche sono di dimensioni molto ridotte e, per esempio, un batterio bastoncellare è generalmente lungo da 1 a 5 mm e largo circa 1.
10
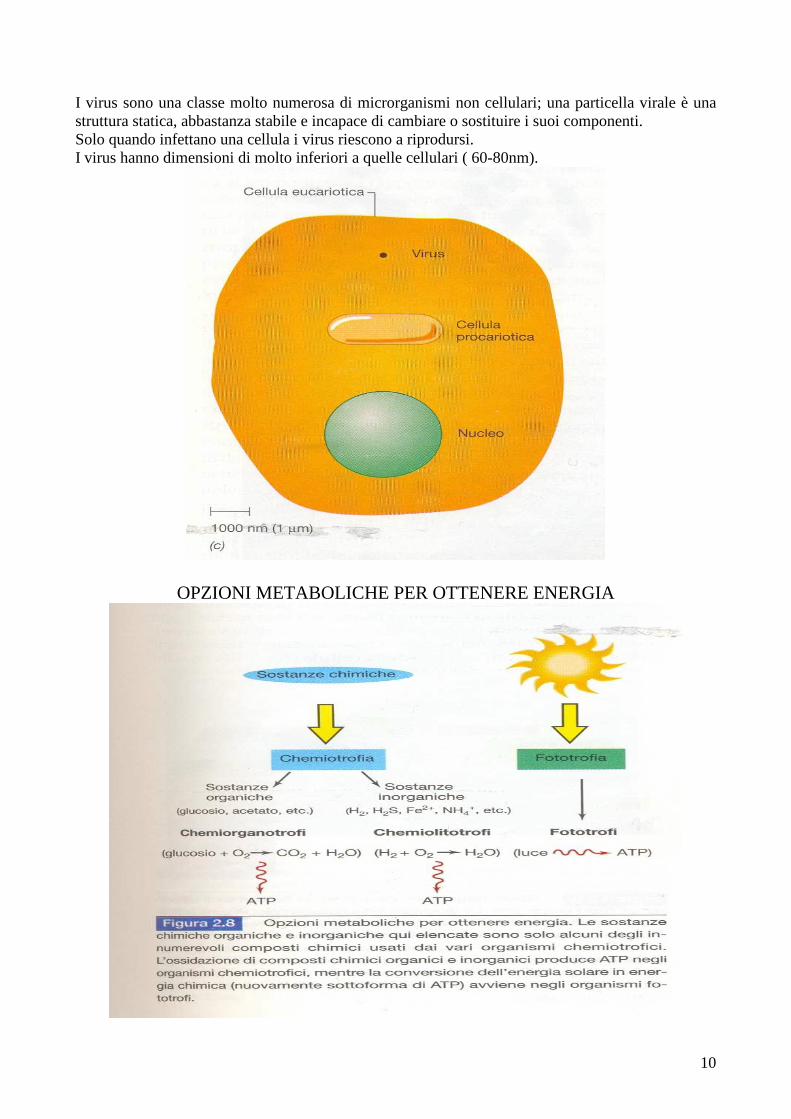
I virus sono una classe molto numerosa di microrganismi non cellulari; una particella virale è una struttura statica, abbastanza stabile e incapace di cambiare o sostituire i suoi componenti. Solo quando infettano una cellula i virus riescono a riprodursi. I virus hanno dimensioni di molto inferiori a quelle cellulari ( 60-80nm).
OPZIONI METABOLICHE PER OTTENERE ENERGIA

11
I processi vitali delle cellule sono governati dal loro genoma ; un gene è un segmento di DNA che codifica una proteina . Nelle cellule procariotiche il DNA è presente sottoforma di grande molecola a doppio filamento chiamata cromosoma batterico, aggregato a formare una massa densa definita nucleoide. Il DNA è circolare in molti procarioti e la maggior parte di essi ha un cromosoma singolo (per tale motivo sono geneticamente aploidi). Molti procarioti possiedono inoltre del DNA extracromosomico organizzato in molecole circolari dette plasmidi . Nelle cellule eucariotiche il DNA è presente come molecola lineare all’interno del nucleo, impacchettato a formare i cromosomi. Gli eucarioti hanno 2 copie di ogni gene e quindi sono geneticamente diploidi . Alcune specie di procarioti prosperano in condizioni ambientali estreme: ipertermofili (alta T), psicrofili (bassa T), acidofili (basso pH), alcalofili (alto pH), Barofili (alte pressioni fondali oceanici) e alofili (alte concentrazioni di sale).
CAP.4 Struttura e funzioni cellulari

12
L’esame microscopico dei microrganismi si basa sull’uso del microscopio ottico(nelle applicazioni di routine) e di quello elettronico (per studi più dettagliati). Oltre all’ ingrandimento è importante la risoluzione, cioè la capacità di mostrare due punti adiacenti come distinti. Sebbene l’ingrandimento possa essere aumentato a piacere, la risoluzione è invece limitata e dipende dalle caratteristiche fisiche della luce. La risoluzione del microscopio ottico è circa 0.2mmmmm, invece il microscopio elettronico ha una risoluzione di circa 1000 volte superiore. Si utilizzano vari tipi di microscopi ottici: in campo chiaro, a contrasto di fase, in campo scuro e a fluorescenza. Il microscopio in campo chiaro consiste di 2 serie di lenti (obiettivo e oculare) che lavorano insieme per definire l’immagine; la visualizzazione dei campioni avviene grazie alle differenze di contrasto esistenti tra gli stessi e il mezzo circostante e dovute alla capacità delle cellule di assorbire o disperdere in varia misura la luce. L’ingrandimento di un preparato microscopico è dato dal prodotto degli ingrandimenti dell’obbiettivo e dell’oculare. Il valore massimo di ingrandimento ottenibile con un microscopio ottico è di 1500 volte e mi permette di osservare i mm 1/1000 del mm, mentre con un microscopio elettronico è possibile ottenere ingrandimenti di 1000 volte maggiori (distinguo i nm . un milionesimo del mm). Per aumentare il limite di risoluzione di preparati ad elevato ingrandimento è consigliato l’uso di olio per lenti a immersione da interporre tra l’obiettivo e il preparato, che aumenta la capacità di raccolta della luce da parte di una lente. Uno dei limiti della microscopia in campo chiaro è certamente il contrasto. I coloranti usati per colorare le cellule hanno la funzione di migliorare il contrasto rendendo visibile al meglio il preparato; i coloranti sono composti organici con affinità per specifici componenti cellulari e ne sono un esempio il blu di metilene, il cristal violetto e la safranina. Invece i coloranti differenziali non colorano in maniera uguale tutti i tipi di cellule e tra questi è ampiamente utilizzata la colorazione di Gram. I batteri sono suddivisi in Gram+ (si presentano colorati in viola) e Gram- (colorati in rosso); la diversa reazione alla colorazione di Gram è dovuta a differenze strutturali nella parete cellulare, per cui l’etanolo è in grado di decolorare un batterio Gram- ma non un Gram+. Per l’identificazione di un batterio sconosciuto è indispensabile stabilire per prima cosa se esso sia Gram+ o Gram- (tramite il microscopio a fluorescenza la procedura sarebbe molto più rapida).

13
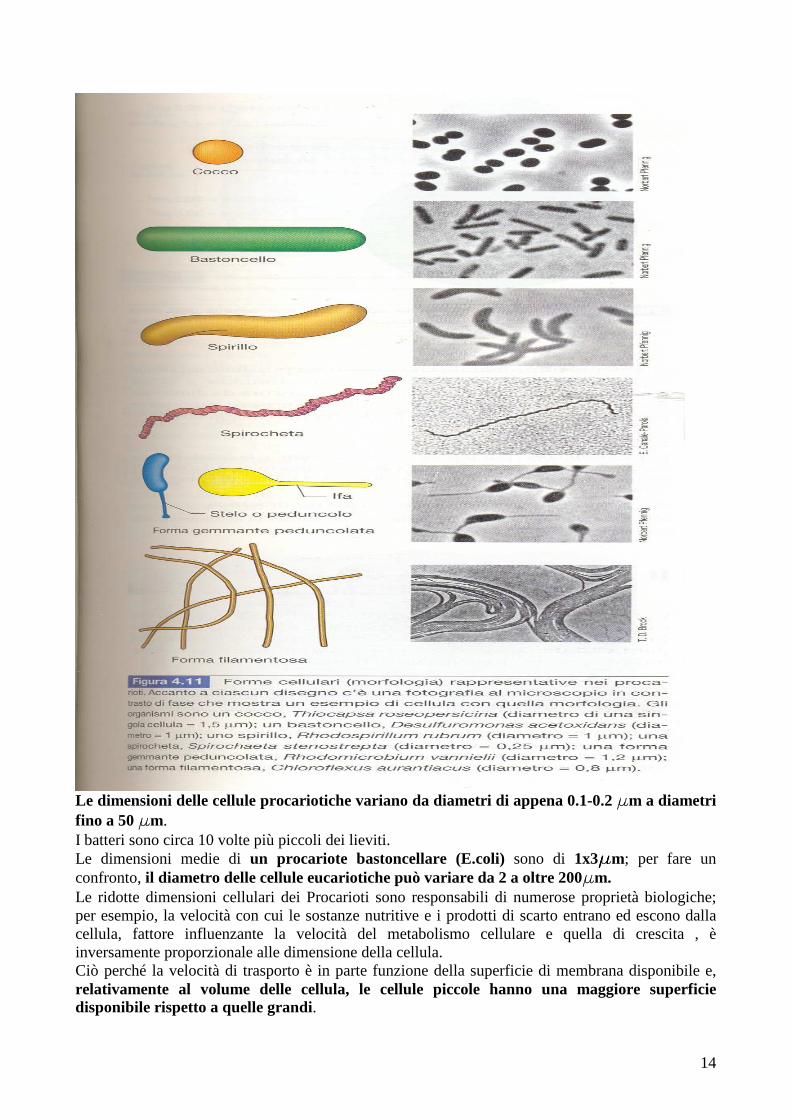
Il microscopio a contrasto di fase è stato sviluppato per migliorare le differenze di contrasto tra le cellule e il mezzo circostante. Il microscopio in campo scuro è un microscopio ottico in cui il sistema di illuminazione è stato modificato in modo che raggiunga il preparato solo lateralmente; l’unica luce che raggiunge la lente è quella dispersa dal campione, quindi esso appare chiaro in campo scuro. Il microscopio a fluorescenza si utilizza nel caso di campioni che emettono fluorescenza, a causa della presenza nella cellula di substrati fluorescenti, come la clorofilla, o in seguito a trattamento con un colorante fluorescente. Il microscopio elettronico a trasmissione è utile per lo studio della struttura interna; in esso un fascio di elettroni sostituisce la radiazione luminosa e le lenti sono costituite da elettromagneti. Il microscopio elettronico a scansione è utile per immagini tridimensionali ma si può visualizzare solo la superficie di un oggetto; il campione, prima di essere attraversato dal fascio di elettroni, viene ricoperto da un sottile strato di un metallo pesante, come per esempio l’oro. MORFOLOGIA E DIMENSIONI DELLE CELLULE BATTERICHE Alcuni gruppi batterici sono facilmente riconoscibili a causa delle loro forme, ne sono esempi i cocchi, i bastoncelli, gli spirilli, le spirochete (batteri spiraliformi), i batteri peduncolati (le cui cellule possiedono estrusioni a forma di lunghi tubi o peduncoli) e i batteri filamentosi (formano lunghe e sottili cellule o catene di cellule).
14
Le dimensioni delle cellule procariotiche variano da diametri di appena 0.1-0.2 mmmmm a diametri fino a 50 mmmmm. I batteri sono circa 10 volte più piccoli dei lieviti. Le dimensioni medie di un procariote bastoncellare (E.coli) sono di 1x3mmmmm; per fare un confronto, il diametro delle cellule eucariotiche può variare da 2 a oltre 200mmmmm. Le ridotte dimensioni cellulari dei Procarioti sono responsabili di numerose proprietà biologiche; per esempio, la velocità con cui le sostanze nutritive e i prodotti di scarto entrano ed escono dalla cellula, fattore influenzante la velocità del metabolismo cellulare e quella di crescita , è inversamente proporzionale alle dimensione della cellula. Ciò perché la velocità di trasporto è in parte funzione della superficie di membrana disponibile e, relativamente al volume delle cellula, le cellule piccole hanno una maggiore superficie disponibile rispetto a quelle grandi.

15
Questo concetto è facilmente esemplificato da una sfera, in cui il volume è funzione del cubo del raggio (V=4/3ppppr3), mentre la superficie è funzione del quadrato del raggio (S=4ppppr2). Il rapporto superficie/volume di una sfera può essere quindi espresso come 3/r. Quindi una cellula con r piccolo avrà un rapporto superficie/volume maggiore di una cellula più grande, e quindi potrà avere scambi più efficienti di sostanze nutritive con l’ ambiente esterno. Questa differenza nel rapporto S/V si manifesta con un tasso di sviluppo più rapido, con la formazione di popolazioni microbiche più ampie in cellule più piccole piuttosto che in cellule più grandi. (il vino invecchia prima nella botte piccola che in quella grande). Tali parametri influenzano a loro volta l’ecologia microbica (un elevato n° di cellule con un metabolismo rapido può portare cambiamenti fisici-chimici fondamentali all’ecosistema anche in un breve periodo di tempo). I nanobatteri sono batteri molto piccoli le cui dimensioni sono dell’ordine di 0.1mmmmm.
LA MEMBRANA CELLULARE La membrana citoplasmatica è una struttura sottile, spessa soltanto 8nm, che circonda completamente la cellula separandone l’interno (il citoplasma) dall’ambiente. Essa è anche una barriera altamente selettiva in quanto permette alla cellula di concentrare al suo interno specifici composti metabolici e di espellere all’esterno sostanze di rifiuto. La struttura generale delle membrane biologiche è un doppio strato lipidico; i fosfolipidi contengono sia una componente idrofobica (acidi grassi) sia una componente idrofilica (glicerolo).
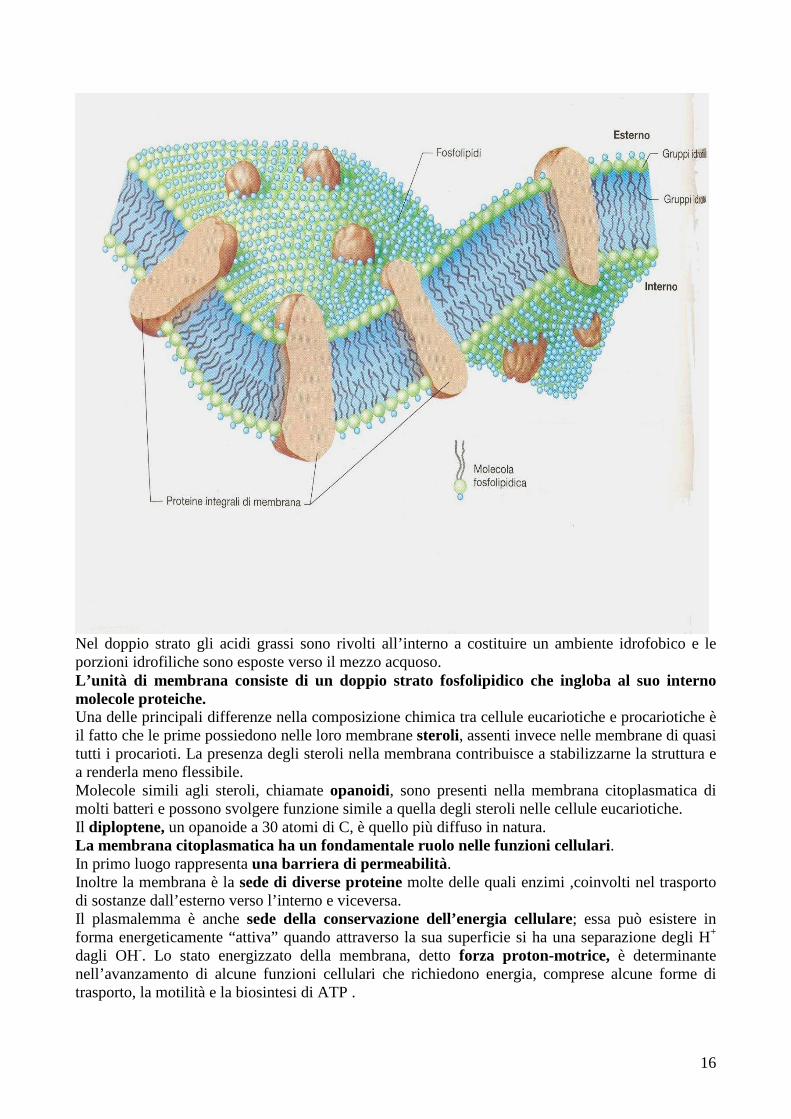
16
Nel doppio strato gli acidi grassi sono rivolti all’interno a costituire un ambiente idrofobico e le porzioni idrofiliche sono esposte verso il mezzo acquoso. L’unità di membrana consiste di un doppio strato fosfolipidico che ingloba al suo interno molecole proteiche. Una delle principali differenze nella composizione chimica tra cellule eucariotiche e procariotiche è il fatto che le prime possiedono nelle loro membrane steroli, assenti invece nelle membrane di quasi tutti i procarioti. La presenza degli steroli nella membrana contribuisce a stabilizzarne la struttura e a renderla meno flessibile. Molecole simili agli steroli, chiamate opanoidi, sono presenti nella membrana citoplasmatica di molti batteri e possono svolgere funzione simile a quella degli steroli nelle cellule eucariotiche. Il diploptene, un opanoide a 30 atomi di C, è quello più diffuso in natura. La membrana citoplasmatica ha un fondamentale ruolo nelle funzioni cellulari. In primo luogo rappresenta una barriera di permeabilità. Inoltre la membrana è la sede di diverse proteine molte delle quali enzimi ,coinvolti nel trasporto di sostanze dall’esterno verso l’interno e viceversa. Il plasmalemma è anche sede della conservazione dell’energia cellulare; essa può esistere in forma energeticamente “attiva” quando attraverso la sua superficie si ha una separazione degli H+ dagli OH-. Lo stato energizzato della membrana, detto forza proton-motrice, è determinante nell’avanzamento di alcune funzioni cellulari che richiedono energia, comprese alcune forme di trasporto, la motilità e la biosintesi di ATP .

17
La natura idrofobica della membrana citoplasmatica fa si che solo alcune piccole molecole idrofobiche possano passare attraverso essa; le molecole d’acque sono sufficientemente piccole per attraversarla ed il trasporto può essere accelerato da trasportatori specifici (acquaporine). Le proteine di trasporto permettono l’accumulo di soluti all’interno della cellula contro un elevato gradiente di concentrazione. Il trasferimento mediato da trasportatori determina una maggiore concentrazione all’interno della cellula e mostra un effetto di saturazione, cioè anche a basse concentrazioni di substrato il tasso di captazione è massimo e la concentrazione del substrato è sufficientemente elevata da saturare il trasportatore. Una peculiarità del meccanismo del trasporto mediato è la sua natura altamente specifica. Inoltre la sintesi delle proteine di trasporto è regolata dalla cellula, cosicché la presenza di specifici trasportatori sulla membrana è funzione sia dei nutrienti presenti sia della loro concentrazione nel mezzo. Nei procarioti ci sono oltre 200 sistemi di trasporto e la traslocazione (trasporto fuori dal plasmalemma) delle proteine avviene attraverso l’attività di proteine chiamate traslocasi. Nei batteri l’esportazione delle proteine è un processo molto importante, in quanto molti enzimi batterici funzionano all’esterno della cellula; per esempio, alcuni enzimi come le amilasi o cellulasi sono escrete direttamente nell’ambiente dove idrolizzano amido e cellulosa trasformandoli in glucosio che viene utilizzato dalla cellula come fonte di carbonio e di energia. Inoltre, molti batteri patogeni secernono nell’ospite, durante il processo infettivo, tossine o altre proteine dannose.

18
LA PARETE CELLULARE DEI PROCARIOTI La concentrazione dei soluti all’interno della cellula sviluppa una notevole pressione che in un batterio come E. coli può raggiungere le 2 atm. I batteri sono provvisti di parete cellulare per sopportare la pressione, che inoltre conferisce alla cellula forma e rigidità. La parete cellulare procariotica può essere messa in evidenza in sezioni sottili in un preparato di microscopia elettronica. I batteri possono essere divisi in 2 gruppi principali (Gram+ e Gram-) utilizzando la colorazione di Gram, che li differenzia in relazione alle diversità strutturali della parete cellulare. Nei Gram- è una struttura complessa e multi-stratificata, mentre nei Gram+ consiste di un singolo tipo di molecola ( il peptidoglicano) ed è molto più spessa. Nella colorazione di Gram un complesso insolubile di cristal-violetto si forma all’interno della cellula: esso viene estratto dall’alcol nei Gram- ma non nei Gram+. Questi ultimi, infatti , hanno una parete cellulare molto spessa che si disidrata in presenza di alcol. Ciò comporta che i pori della membrana si chiudano, impedendo al complesso insolubile cristal-violetto di fuoriuscire.
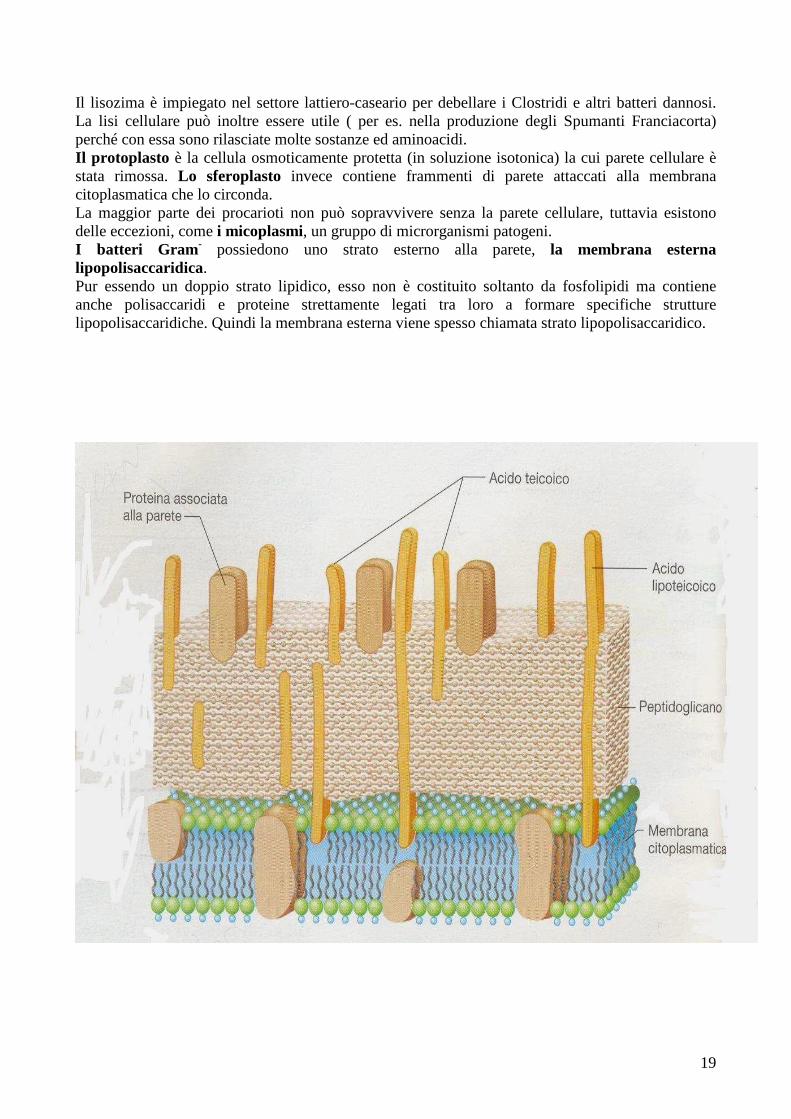
Lo strato di peptidoglicano o mureina è il principale responsabile della rigidità della cellula ed ogni suo strato è una sottile lamina costituita da 2 derivati polisaccaridici, l’ N-acetilglucosamina e l’acido N-acetilmuramico, e da un piccolo gruppo di aminoacidi (L-alanina, D-alanina; acido D-glutamico e Lisina o, in alternativa, acido diaminopimelico). Le catene di glicano (legami bbbb1,4) sono tenute insieme da legami crociati peptidici tra aminoacidi, che conferiscono la robustezza propria della struttura peptidoglicanica. La lunghezza delle catene di peptidoglicano e la quantità di legami crociati sono responsabili della forma caratteristica dei batteri. Nei batteri Gram+ il peptidoglicano rappresenta più del 90% della parete cellulare e molti di essi ne posseggono più di 25 strati. Nei Gram- soltanto il 10% circa della parete è costituito da peptidoglicano (1 o pochi strati), la parte restante è costituita invece dalla membrana esterna. Il peptidoglicano è presente solo nei batteri: sia l’acido N-acetilmuramico sia l’acido diaminopimelico sono del tutto assenti nella parete cellulare degli Archea e degli Eucarioti. Comunque, non tutti i batteri hanno l’acido diaminopimelico nel peptidoglicano; questo aminoacido è presente in tutti i Gram- e in alcune specie di Gram+; in queste ultime troviamo perlopiù Lisina o altri aminoacidi. Il peptidoglicano può essere distrutto da diversi agenti chimici e uno di questi è l’enzima lisozima, una proteina che rompe i legami bbbb1,4 tra l’ N-acetilglucosamina e l’acido N-acetilmuranico del peptidoglicano, indebolendo fortemente la struttura. Quindi l’acqua può entrare nella cellula facendola rigonfiare e infine scoppiare in un processo detto lisi. Il lisozima viene naturalmente isolato dalle secrezioni animali, per esempio le lacrime e la saliva; probabilmente svolge una funzione fondamentale come prima linea difensiva contro le infezioni batteriche.
19
Il lisozima è impiegato nel settore lattiero-caseario per debellare i Clostridi e altri batteri dannosi. La lisi cellulare può inoltre essere utile ( per es. nella produzione degli Spumanti Franciacorta) perché con essa sono rilasciate molte sostanze ed aminoacidi. Il protoplasto è la cellula osmoticamente protetta (in soluzione isotonica) la cui parete cellulare è stata rimossa. Lo sferoplasto invece contiene frammenti di parete attaccati alla membrana citoplasmatica che lo circonda. La maggior parte dei procarioti non può sopravvivere senza la parete cellulare, tuttavia esistono delle eccezioni, come i micoplasmi, un gruppo di microrganismi patogeni. I batteri Gram - possiedono uno strato esterno alla parete, la membrana esterna lipopolisaccaridica. Pur essendo un doppio strato lipidico, esso non è costituito soltanto da fosfolipidi ma contiene anche polisaccaridi e proteine strettamente legati tra loro a formare specifiche strutture lipopolisaccaridiche. Quindi la membrana esterna viene spesso chiamata strato lipopolisaccaridico.

20
Un importante proprietà biologica della membrana esterna di molti Gram - è la tossicità per gli animali. Tra i Gram- patogeni per l’uomo vi sono membri dei generi Salmonella, Shigella e Escherichia. Le proprietà tossiche dipendono da un composto tossico (endotossina) presente in una porzione della membrana esterna. A differenza della membrana citoplasmatica ,la membrana esterna dei batteri Gram-, pur essendo essenzialmente un doppio strato lipidico, è parzialmente impermeabile a piccole molecole. Ciò avviene grazie alla presenza di piccole proteine dette porine con funzione di canali attraverso la membrana esterna , permettendo l’entrata e l’uscita di sostanze idrofiliche a basso peso molecolare. Una delle funzioni principali della membrana esterna si ritiene sia quella di impedire che alcuni enzimi, localizzati all’esterno della membrana citoplasmatica in una regione chiamata periplasma, diffondano al di fuori dalla cellula . In E.coli il periplasma ha uno spessore di circa 12-15nm e una consistenza gelatinosa. Il periplasma contiene parecchie proteine, tra cui enzimi idrolitici, e i chemiorecettori, proteine coinvolte nella risposta chemiotattica. LA LOCOMOZIONE MICROBICA Molti procarioti sono dotati di mobilità; questa capacità dipende in genere da una particolare struttura, il flagello (non è visibile al microscopio ottico).

21
Alcune cellule batteriche si muovono sulle superfici solide tramite lo scivolamento e alcuni microrganismi acquatici possono controllare la loro posizione in acqua grazie alla presenza di strutture intracellulari ripiene di gas, dette vescicole gassose. La maggior parte dei procarioti dotati di motilità si muove comunque grazie ai flagelli . Potersi spostare in una nuova posizione può offrire alla cellula nuove opportunità e nuove risorse nutrizionali. I flagelli batterici sono appendici cellulari lunghe e sottili, libere a una estremità e intimamente legate alla cellula dall’altra. Essi sono così sottili (circa 20nm) da essere facilmente visibili solo al microscopio elettronico. I flagelli sono disposti in modo diverso nei diversi batteri. Nella distribuzione polare sono attaccati a uno (batterio monotrico) o entrambi i poli della cellula (batteri anfitrichi ). Talvolta un ciuffo di flagelli può originarsi a una estremità della cellula (batteri loffotrichi ). Nella distribuzione peritrica i flagelli si originano da numerosi punti della superficie della cellula. Il tipo di disposizione dei flagelli, oltre alla funzione nella motilità batterica, trova utilità nella classificazione batterica. I flagelli hanno una struttura elicoidale e presentano, tra due sinuosità adiacenti, una lunghezza d’onda, che è costante per ogni specie. Il filamento dei flagelli batterici è costituito da numerose subunità di una proteina detta flagellina. La regione basale del flagello ha una struttura diversa da quella del filamento. Essa è costituita da una regione più ampia, detta uncino, formata da un singolo tipo di proteina e con la funzione di connettere il filamento alla porzione del motore del flagello. Il motore flagellare, ancorato alla membrana citoplasmatica e alla parete cellulare, è formato da una piccola regione bastoncellare centrale che passa attraverso un sistema di anelli. Nei Gram- la coppia esterna di anelli è ancorata allo strato lipopolisaccaridico e al peptidoglicano della parete cellulare, mentre la coppia più interna è localizzata all’intorno della membrana citoplasmatica. Nei batteri Gram+, privi dello strato lipopolisaccaridico esterno, è presente solo la coppia più interna. Intorno ad essa e ancorate alla membrana citoplasmatica ci sono delle proteine dette Mot , che controllano il motore, che con una torsione fa ruotare il filamento. Un altro gruppo di proteine, dette Fli , funge da invertitore, ribaltando la rotazione del flagello in risposta a segnali intracellulari.

22
Per la sintesi dei flagelli e per il processo di motilità sono necessari numerosi geni: in E.coli ne sono stati identificati oltre 40. Il flagello non cresce dalla base come nel caso dei peli animali, ma dalla punta. Le molecole di flagellina sintetizzate nel citoplasma passano attraverso un canale di 3nm all’interno del filamento e vengono assemblate all’estremità terminale per formare il flagello. Qui si trova una proteina detta Cap che ha il ruolo di organizzare le molecole di flagellina per formare una nuova porzione di filamento.
Ogni singolo flagello è una struttura rigida che non si flette ma si muove per rotazione, come un’ elica. Il corpo basale impartisce il movimento rotatorio del flagello (il principale è antiorario ). L’energia richiesta per la rotazione del flagello proviene dalla forza-motrice. La dissipazione del gradiente protonico attraverso la membrana mediata dal complesso Mot fa ruotare il flagello, ed è stato calcolato che devono essere traslocati circa 1000 H+ affinché possa avvenire un singolo movimento rotatorio. I flagelli non ruotano a velocità costante e la rotazione flagellare può fare avanzare un batterio in un terreno liquido alla velocità di 60 lunghezze cellulari/secondo. Benché ciò significhi un avanzamento pari a 0,00017 Km/h, se compariamo questa velocità con quella di un animale superiore in termini di lunghezze per secondo notiamo che i batteri sono estremamente veloci. L’animale più veloce del mondo, il ghepardo, si muove alla velocità massima di 121 Km/h, equivalente a circa 25lunghezze al secondo. I microrganismi peritrichi in genere si muovono in linea retta, in modo lento e uniforme, mentre i microrganismi polari procedono più rapidamente, muovendosi qua e la a scatti.

23
Alcuni procarioti si muovono attraverso una superficie solida mediante un processo definito scivolamento. Tale tipo di movimento è considerevolmente più lento (circa 10 mm/sec) di quello a propulsione mediato dal flagello. I batteri che si muovono per scivolamento sono filamentosi o bastoncellari. Benché non sia stato ancora ben identificato nessun meccanismo di movimento per scivolamento, vi sono indicazioni che tale fenomeno avvenga secondo modalità diverse. Nei cianobatteri tale movimento è accompagnato dalla secrezione di una sostanza mucosa (slime) di natura polisaccaridica; man mano che viene escreto, lo slime aderisce sulla superficie solida e la cellula viene spinta nel suo avanzamento. In altri batteri il movimento per scivolamento è indotto da un movimento delle proteine della superficie cellulare; in questo modello, le proteine specifiche per la motilità sono ancorate nella membrana citoplasmatica e nella membrana esterna e si ipotizza che sospingano le cellule in avanti e indietro con un meccanismo a ruota dentata. Benché la motilità non sia un carattere presente in tutti i procarioti, essa conferisce un vantaggio selettivo in ambienti particolari. I procarioti spesso incontrano in natura gradienti chimici e fisici, e l’apparato responsabile della motilità nella cellula si è evoluto per rispondere in modo positivo o negativo ad essi inducendo la cellula ad avvicinarsi o ad allontanarsi dalla molecola segnale. Questi movimenti direzionali sono detti tassie e nelle cellule batteriche se ne conoscono diverse; le due più studiate sono la chemiotassi, la risposta ad una sostanza chimica, e la fototassi, la risposta alla luce. I procarioti sono troppo piccoli per percepire un gradiente attraverso la “lunghezza” del loro corpo. Muovendosi, essi devono invece comparare lo stato fisico e chimico dell’ambiente che li circonda con la sensazione registrata pochi secondi prima. In altre parole, i batteri rispondono a un gradiente temporale anziché spaziale delle molecole “segnale” che incontrano nuotando. Le fimbrie e i pili sono simili ai flagelli ma non sono coinvolti nel movimento. Le fimbrie sono notevolmente più corte dei flagelli e molto più numerose, ma come questi sono di natura proteica. Le fimbrie consentono al microrganismo di aderire a superfici come tessuti animali nel caso dei microrganismi patogeni, o di formare pellicole o biofilm sulle superfici. I pili sono strutture simili alle fimbrie, ma generalmente sono più lunghi e presenti sulla cellula in una o più copie. E’ noto che sono coinvolti nel processo di coniugazione nei procarioti; sono inoltre coinvolti nei fenomeni di adesione ai tessuti umani da parte dei microrganismi patogeni.

24
Molti procarioti contengono uno strato superficiale composto da una serie di proteine strutturate bidimensionalmente; questi strati sono chiamati Strato paracristallino o S-layer o strati S. La loro funzione principale è sconosciuta ma è probabile che funzionino almeno come una barriera esterna di permeabilità. Il glicocalice è un materiale ricco in polisaccaridi deposto all’esterno della cellula. Esso varia nei diversi organismi, e a seconda della composizione chimica in uno specifico microrganismo può essere spesso o sottile, rigido o flessibile. Gli strati rigidi sono organizzati come una fitta matrice che non permette il passaggio di particelle e che protegge il microrganismo; questo tipo di struttura è definito capsula. Alcuni batteri capsulati sono responsabili di malattie del vino (“vino che fila come l’olio”) e sono utili nel terreno perché ne migliorano la struttura. Se invece il glicocalice è di tipo flessibile, non è in grado di impedire il passaggio di particelle e questo tipo di struttura è definita strato mucoso (funzione per lo più di adesione). Gli strati polisaccaridici esterni hanno un ruolo importante nell’adesione di alcuni microrganismi patogeni ai loro ospiti. LE ENDOSPORE Alcune specie di batteri producono speciali strutture all’interno delle loro cellule chiamate endospore durante un processo definito sporulazione. Le endospore sono cellule differenziate molto resistenti al calore.(nei Lieviti e quindi negli eucarioti la spora è un gamete prodotto nella meiosi) Batteri che formano le endospore abitano comunemente il suolo ed i generi Bacillus e Clostridium sono tra i batteri sporigeni più studiati. Benché molti altri organismi oltre i batteri formino spore, l’endospora batterica è unica per il suo elevato grado di resistenza al calore. Le endospore sono resistenti anche ad altri agenti dannosi, quali disidratazione, radiazioni, acidi e disinfettanti chimici, e possono restare quiescenti per periodi di tempo estremamente lunghi. Nel 1995 un gruppo di scienziati ha ottenuto la germinazione di spore batteriche che risalivano a circa 25-40milioni di anni fa. Le spore erano conservate nell’intestino di una specie di ape estinta intrappolata in un’ambra risalente a un era geologicamente nota. Ancora più spettacolare fu la scoperta di batteri alofili isolati da cristalli di sale risalenti a oltre 250 milioni di anni fa. Le spore sono impermeabili ai coloranti, proprietà che le evidenzia come regioni incolori in cellule trattate con coloranti basici come il blu di metilene. La struttura di una spora è complessa e costituita da numerosi strati di rivestimento. Lo strato più esterno, detto esosporio, è sottile e delicato. Più internamente si trova la tunica (o parete della spora) che è composta da uno o più strati proteici. Al di sotto di questa si trova la corteccia , uno strato di peptidoglicano lasso, all’interno della quale è situato il core o protoplasto della spora, costituito da strutture convenzionali quali parete cellulare, membrana citoplasmatica, citoplasma, nucleoide etc. Quindi, dal punto di vista strutturale, la spora differisce dalla cellula vegetativa principalmente per gli involucri presenti all’esterno della parete cellulare. Una sostanza chimica caratteristica delle endospore , assente nelle cellule vegetative, è l’acido dipicolinico, che è localizzato a livello del core. Le spore presentano inoltre una elevata concentrazione di ioni calcio, molti dei quali complessati con l’acido dipicolinico.

25
La resistenza della spora è data da tre fattori fondamentali: gli involucri protettivi, la notevole riduzione di acqua (fino al 10%) vivendo in stato di criptobiosi, l’elevata % di calcio complessato con l’acido dipicolinico presente nel core. La germinazione è un processo, che avviene in condizioni idonee, in cui si ha la formazione della nuova cellula vegetativa a partire dall’endospora. Con la formazione dell’endospora, per es. in condizioni di emergenza, una cellula vegetativa viene convertita in una struttura incapace di crescere e molto resistente al calore. Il processo di sporulazione coinvolge una complessa serie di eventi di differenziamento cellulare. La sporulazione batterica non avviene quando le cellule sono in fase esponenziale, ma soltanto quando la crescita cellulare si blocca a causa dell’esaurimento di sostanze nutritive essenziali. Numerosi cambiamenti geneticamente determinanti controllano la transizione da sviluppo vegetativo a sporulazione. Le proteine codificate da questi geni catalizzano tutta quella serie di processi che determinano la trasformazione della cellula vegetativa con attività metabolizzante in un endospore disidratata, metabolicamente inerte ma estremamente resistente. Tramite la sterilizzazione del latte è possibile eliminare le spore. Confrontando la spora e la cellula vegetativa è possibile distinguere vari generi: - genere Clostridium (anaerobio obbligato che in presenza di aria sporifica; nel formaggio causa il “gonfiore tardivo”) ha la spora con diametro maggiore della cellula vegetativa e si trova in posizione centrale. - genere Plectridio ha la spora in posizione polare (deformazione a clava) ed il diametro è maggiore di quello della cellula vegetativa. - genere Bactridio ha la spora con diametro inferiore a quello della cellula vegetativa.
LA SPORULAZIONE

26
CAP.10 Genetica dei microrganismi
MUTAZIONE E RICOMBINAZIONE La mutazione è un cambiamento ereditario della sequenza delle basi dell’acido nucleico del genoma di un organismo. La ricombinazione genetica è il processo mediante il quale segmenti genetici contenuti in due genomi separati vengono messi insieme in un’unica unità. Mentre generalmente la mutazione induce nella cellula cambiamenti genetici di piccola entità, la ricombinazione genetica generalmente comporta cambiamenti più significativi. Interi geni, serie di geni e anche interi cromosomi vengono trasferiti da un organismo all’altro. Anche se i procarioti, diversamente da molti eucarioti, non si riproducono sessualmente, in essi esistono meccanismi di scambio genetico che permettono sia il trasferimento di geni che la ricombinazione. Per riconoscere lo scambio genetico tra due organismi è necessario impiegare marcatori genetici il cui trasferimento possa essere identificato. A questo scopo si usano ceppi mutanti e i cambiamenti sono dovuti a una o più mutazioni del DNA dell’organismo. Queste possono coinvolgere un cambiamento in una o più paia di basi o anche l’inserzione o l’eliminazione di interi geni. Viene detto mutante un organismo il cui genoma è stato oggetto di una mutazione; quindi un mutante differisce dal ceppo parentale nel genotipo, inoltre, le proprietà osservabili del mutante, il suo fenotipo, possono essere diverse rispetto al ceppo parentale. E’ comune riferirsi a un ceppo isolato in natura come a un ceppo di tipo selvatico. A seconda della mutazione, il mutante può avere fenotipo mutante oppure fenotipo uguale a quello dei suoi parentali. Alcune mutazioni sono selezionabili, in quanto conferiscono alcuni vantaggi agli organismi che le possiedono, mentre altre sono non selezionabili, anche se portano a un profondo cambiamento del fenotipo di un organismo. Un esempio di mutazione non selezionabile è quella della perdita di colore in un organismo pigmentato. Tali colonie di solito non hanno ne un vantaggio ne uno svantaggio rispetto alle colonie parentali pigmentate quando crescono in piastre di agar. Si possono identificare queste mutazioni solamente esaminando un gran numero di colonie e cercando quelle differenti attraverso un processo detto screening. Una mutazione selezionabile conferisce al mutante un vantaggio in certe condizioni ambientali, per cui la progenie della cellula mutante è in grado di crescere in modo tale da sostituirsi a quella parentale. Un esempio di mutazione selezionabile è la resistenza ai farmaci: un mutante antibiotico resistente può crescere in presenza di una concentrazione di antibiotico che inibisce o uccide il parentale. E’ relativamente facile riconoscere e isolare i mutanti selezionabili scegliendo le appropriate condizioni ambientali, utilizzando come strumento genetico la selezione. Per certi tipi di mutazioni sono disponibili metodi di screening per grandi numeri di colonie. Per esempio possono essere individuati mutanti nutrizionali con la tecnica della “replica plating” (piastramento per replica).
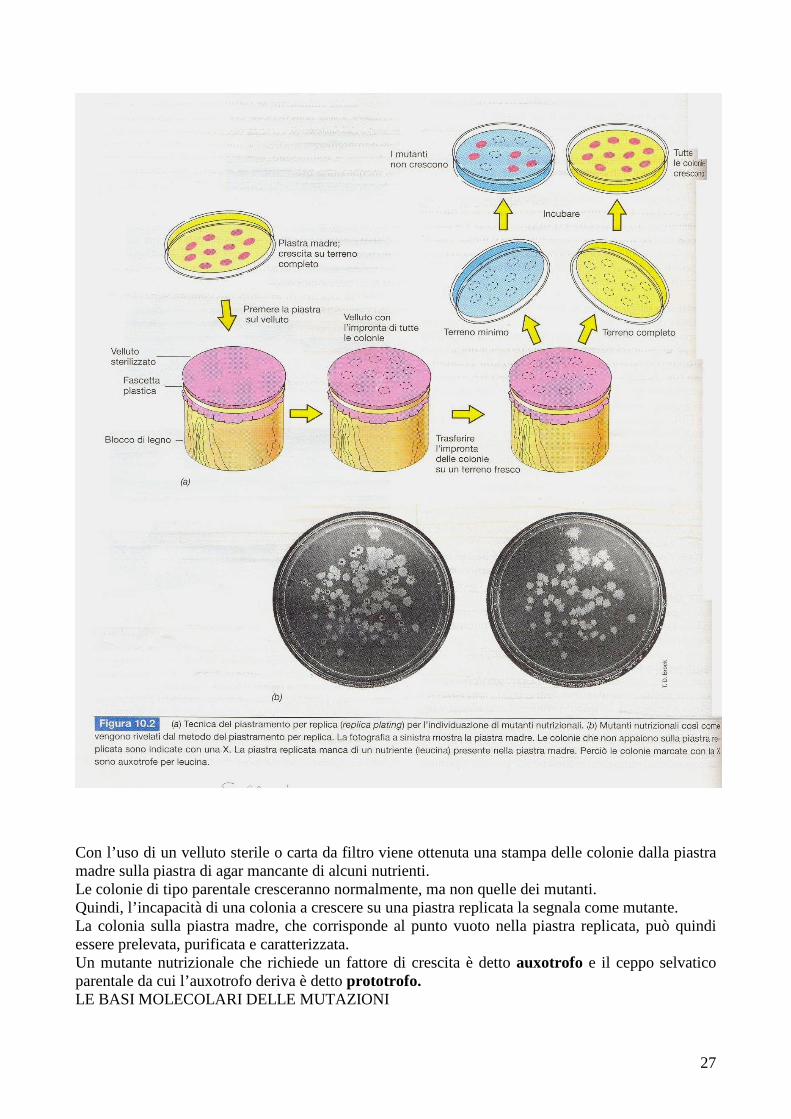
27
Con l’uso di un velluto sterile o carta da filtro viene ottenuta una stampa delle colonie dalla piastra madre sulla piastra di agar mancante di alcuni nutrienti. Le colonie di tipo parentale cresceranno normalmente, ma non quelle dei mutanti. Quindi, l’incapacità di una colonia a crescere su una piastra replicata la segnala come mutante. La colonia sulla piastra madre, che corrisponde al punto vuoto nella piastra replicata, può quindi essere prelevata, purificata e caratterizzata. Un mutante nutrizionale che richiede un fattore di crescita è detto auxotrofo e il ceppo selvatico parentale da cui l’auxotrofo deriva è detto prototrofo. LE BASI MOLECOLARI DELLE MUTAZIONI

28
La mutazione può essere spontanea o indotta; quella spontanea è un evento raro (una mutazione per miliardo di cellule) e può essere il risultato dell’azione di una radiazione naturale (raggi cosmici), che altera la struttura delle basi del DNA. Mutazioni spontanee possono anche avvenire durante la replicazione, come risultato d’errore nell’appaiamento delle basi, fatto che porta a cambiamenti nel DNA replicato. Le mutazioni che coinvolgono una o pochissime paia di basi sono definite mutazioni puntiformi ; queste possono risultare per sostituzione di un paio di basi nel DNA o per un’inserzione o delezione di un paio di basi. Le mutazioni generalizzate riguardano un frammento più o meno grande del genoma.
SOSTITUZIONI DI PAIA DI BASI
Quando in un gene si verifica una mutazione puntiforme nella regione codificante una proteina, è quasi certo che avverrà qualche alterazione nel fenotipo della cellula come conseguenza di una alterazione nella sequenza degli aminoacidi della proteina prodotta. A causa della degenerazione del codice genetico non tutte le mutazione dei geni determinano un cambiamento nella proteina stessa. Per esempio quando il DNA che codifica un singolo codone della tirosina subisce una mutazione: un cambiamento nell’RNA da UAC a UAU non ha alcun effetto, poichè anche UAU è un codone della tirosina. Le mutazioni che inducono tali cambiamenti sono dette mutazioni silenti e tali mutazioni in regioni codificanti cadono sempre nella terza base del codone. I cambiamenti nella prima o seconda base della tripletta possono spesso condurre ad alterazioni nella proteina. Per esempio un cambiamento di base da UAC ad AAC determina un cambiamento nella proteina da tirosina ad asparagina; questa viene definita mutazione missenso, perché la sequenza di aminoacidi nella proteina è cambiata, determinando l’inattivazione o una ridotta attività.

29
Tuttavia, non tutte le mutazioni che determinano una sostituzione di un aminoacido portano a un mancato funzionamento della proteina e ciò dipenderà da dove avviene la sostituzione nella catene polipeptidica e dall’effetto sul ripiegamento e sull’attività catalitica. Un altro possibile risultato della sostituzione di una coppia di basi è la formazione di un codone di stop che può causare una terminazione prematura della traduzione e conseguentemente una proteina incompleta e non funzionale; tali sono dette mutazioni nonsenso . Le mutazioni puntiformi sono reversibili e un revertante è un ceppo in cui il fenotipo selvatico perso nel mutante viene ripristinato. I revertanti possono essere di due tipi; nei revertanti dello stesso sito la mutazione che ripristina l’attività si verifica nel medesimo sito in cui è accaduta la mutazione originale. Se la retromutazione non solo avviene nello stesso sito ma ripristina anche la sequenza di tipo selvatico, viene detta revertante vera. Le delezioni sono mutazioni in cui una regione del DNA viene eliminata. Le inserzioni si verificano per aggiunta di nuove basi al DNA. LA MUTAGENESI (mutazioni indotte dall’uomo) La frequenza con cui avvengono i diversi tipi di mutazione è estremamente variabile; errori nella replicazione del DNA ricorrono con una frequenza di circa 10-7-10-11 per coppia di basi durante un singolo ciclo di replicazione. Mentre la frequenza di mutazioni spontanee è molto bassa, vi sono vari agenti chimici, fisici o biologici che inducono mutazioni e possono aumentarne la frequenza. Tali agenti sono detti mutageni . MUTAGENI CHIMICI Ne esistono varie classi; gli analoghi delle basi, simili nella struttura alle purine e alle pirimidine del DNA, si dimostrano difettosi nell’appaiamento. Quando uno di questi analoghi delle basi viene incorporato nel DNA, il più delle volte la replicazione può avvenire normalmente, ma occasionalmente può verificarsi un errore, con l’incorporazione di una base sbagliata nell’elica copiata. La mutazione si manifesterà durante la successiva segregazione di quest’elica. Altri mutageni chimici causano cambiamenti chimici nelle basi determinando appaiamenti difettosi o altri tipi di modificazioni come ad esempio deaminazione di A e C (acido nitroso), agiscono distruggendo le basi (agenti alchilanti). Tali agenti sono mutageni molto potenti che spesso inducono mutazioni molto più complete. Sia gli analoghi delle basi che gli agenti alchilanti inducono sostituzioni di coppie di basi. Le acridine, un altro gruppo di mutageni chimici che agiscono come agenti intercalanti tendono a inserirsi tra due paia di basi del DNA separandole inducendo mutazioni da scivolamento dello schema di lettura. LE RADIAZIONI Alcuni tipi di radiazioni sono fortemente mutagene. Possiamo dividerle in due categorie: ionizzanti e non ionizzanti ( sono quelle più utilizzate nella genetica microbica ).

30
È stato ormai dimostrato come la radiazione UV a 260 nm è l’agente letale più efficiente, agendo direttamente sul DNA delle cellule colpite. Le radiazioni ionizzanti sono forme di radiazione più potenti che includono lunghezze d’onda corte come raggi X, raggi cosmici e raggi gggg, che causano la ionizzazione dell’acqua e di altre sostanze determinando effetti mutageni indiretti (formazione di radicali liberi tra cui il più importante è l’OH-) che reagiscono e inattivano le macromolecole delle cellule come il DNA, sul quale si possono avere anche effetti permanenti. Le radiazioni ionizzanti vengono usate raramente per i microrganismi a causa della loro pericolosità e della difficile disponibilità.
LA RICOMBINAZIONE GENETICA La ricombinazione genetica è un processo mediante il quale porzioni o intere molecole di DNA diversa origine effettuano degli scambi o si uniscono in una singola molecola. La ricombinazione generale o omologa è quel processo consistente nello scambio genetico tra sequenze omologhe di DNA di due origini differenti. Sequenze omologhe di DNA hanno una sequenza nucleotidica simile ed è per questo che si verifica l’appaiamento di basi in un’ampia regione delle due molecole di DNA; tale tipo di ricombinazione è coinvolta nel processo crossing over. Nella ricombinazione c’è sempre un DNA donatore ed un DNA ricevente; la cellula ricevente in grado di ricevere il DNA è detta competente. Nei batteri la ricombinazione omologa coinvolge la partecipazione di una proteina specifica, la proteina RecA, codificata dal gene recA. Tali proteine sono state identificate in tutti i procarioti, compresi gli archea, così pure nel lievito e negli eucarioti superiori. Il processo della ricombinazione omologa comincia con un taglio (nick), generalmente prodotto da una nucleasi in una delle molecole di DNA. Il filamento spezzato viene divaricato dall’altro filamento da proteine che hanno attività elicasica (srotolamento).

31
Una proteina che lega il DNA a singola elica si associa poi al frammento risultante, seguita dalla proteina RecA, formando un complesso che facilita il riappaiamento con la sequenza complementare nel duplex adiacente, mentre simultaneamente avviene lo scostamento del filamento residente (tale processo è detto invasione del filamento). Dopo l’appaiamento, può avvenire lo scambio di molecole omologhe di DNA, che porta alla formazione di intermedi di ricombinazione. Infine le molecole legate vengono rilasciate dalla nucleasi e dalla DNA-ligasi, per formare due molecole ricombinanti. Tale meccanismo avviene frequentemente in modo spontaneo all’interno della cellula e la probabilità di ricombinazione tra due geni è proporzionale alla loro distanza. Affinché possano emergere nuovi genotipi è essenziale che le due sequenze omologhe siano geneticamente distinte. Le due molecole distinte sono messe insieme come risultato della riproduzione sessuale (processo presente nella maggior parte degli eucarioti). Anche nei procarioti la ricombinazione genetica è molto importante e i frammenti di DNA omologo vengono trasferiti da un cromosoma di una cellula donatrice a quella di una cellula ricevente mediante tre processi:
- trasformazione (DNA “nudo” + cellula) - trasduzione (cellula + cellula e trasferimento tramite fago) - coniugazione (cellula + cellula e trasferimento tramite plasmide)
Dopo il trasferimento avviene la ricombinazione omologa. Poiché nei procarioti viene trasferito solo un frammento di cromosoma, se non avviene ricombinazione, il frammento viene perduto perché non può essere replicato indipendentemente.

32
ORIGINI DELLA GENETICA BATTERICA La scoperta della trasformazione è stata la più significativa e ha portato a mettere in evidenza che il DNA è il materiale genetico. La prima dimostrazione della trasformazione batterica fu ottenuta da Griffith nel 1930. Egli stava lavorando con lo Streptococcus pneumoniae, un batterio che deve la sua abilità di invasione in parte alla presenza di una capsula polisaccaridica. E’ possibile isolare mutanti privi della capsula e quindi incapaci di causare infezioni: i ceppi R, perché le loro colonie appaiono ruvide in agar, in contrasto con l’apparenza liscia dei ceppi capsulati S. Un topo infettato con poche cellule di un ceppo S muore in uno o due giorni di infezione, mentre non si verifica la morte iniettando molti ceppi R. Griffith dimostrò che se erano iniettate cellule S uccise dal calore insieme a cellule R vive, si sviluppava un’infezione letale e i batteri isolati dal topo morto erano di tipo S. Dal momento che si conoscevano diversi tipi di capsule polisaccaridiche isolate da vari ceppi S, fu possibile ripetere l’esperimento con cellule S uccise al calore di un ceppo diverso da quello da cui

33
derivavano i ceppi R. Poiché le cellule S viventi che venivano isolate, avevano sempre la capsula del tipo posseduto dalle cellule S uccise al calore, si concluse che le cellule R erano state trasformate in un nuovo ceppo durante un evento genetico. La spiegazione molecolare della trasformazione dei ceppi di streptococcus è stata trovata da Avery e dai suoi collaboratori all’istituto Rockefeller di New York. Essi dimostrarono che il processo di trasformazione può avvenire in provetta e che l’estratto privo di cellule uccise al calore può indurre trasformazione; la frazione attiva di tale strato fu purificata e si dimostrò che consisteva in DNA. Nel 1953 Watson e Crick presentarono il loro modello della struttura del DNA. La ricerca batteriologica e biochimica di Avery , insieme a quella fisico-chimica di Watson e Crick, consolidarono il concetto che il DNA è il materiale genetico. Mentre la trasformazione batterica è stata il risultato di una scoperta accidentale, la coniugazione batterica è stata dimostrata nel 1946 da Lederberg e Tatum con esperimenti per stabilire se un processo sessuale potesse avvenire nei batteri. Essi svilupparono un metodo che implicava l’ uso di mutanti nutrizionali di E.coli . Fortunatamente questi mutanti erano stati isolati da uno dei pochi ceppi selvatici che contengono naturalmente il plasmide F. L’idea fu quella di mescolare due ceppi con richieste nutrizionali diverse, e piastrare la miscela su un terreno privo dei fattori di crescita a loro necessari. Nessun tipo parentale fu in grado di crescere in questo terreno, mentre era possibile la crescita dei ricombinanti. Si dimostrò che il processo richiedeva un contatto cellula-cellula . La trasduzione batterica venne scoperta da Zinder che lavorava sulla ricombinazione genetica in Salmonella con lo scopo originale del suo lavoro di dimostrare che la coniugazione avveniva in altri organismi oltre che in E.coli. LA TRASFORMAZIONE GENETICA La trasformazione genetica è un processo mediante il quale un DNA libero viene incorporato in una cellula ricevente e determina un cambiamento genetico. Il DNA di un procariote è presente nella cellula come una singola grande molecola, che fuoriesce dalla cellula quando questa è lisata delicatamente. A causa della sua estrema lunghezza (1700mmmmm in Bacillus), la molecola del DNA può rompersi con estrema facilità e ridursi in frammenti di circa 15 kb. Il DNA che corrisponde a un gene medio è di circa 1000 nucleotidi. Quindi ogni frammento di DNA purificato contiene circa 15 geni. Una singola cellula incorpora di solito solo uno o pochi frammenti di DNA, per cui solo una piccola frazione di geni di una cellula può essere trasferita a un’altra durante un singolo evento di trasformazione. Una cellula in grado di assumere una molecola di DNA e di essere trasformata è detta competente. Solamente alcuni ceppi sono competenti e tale capacità sembra essere determinata geneticamente. In una popolazione di batteri solo una cellula su 100 000 è competente e la competenza dipende da diversi fattori:
- tipo del terreno colturale - gli ioni calcio aumentano la competenza - pH - nella fase logaritmica della curva di crescita si ha molta competenza - in bacillus circa il 20% delle cellule divengono competenti - in streptococcus il 100% delle cellule può diventare competente ma solo per pochi minuti
durante il ciclo di crescita.

34
- Il DNA trasformante si lega alla superficie della cellula mediante una proteina (adesina), dopo di che o penetra l’intero frammento a doppio filamento (ciò avviene nei Gram-) o una nucleasi degrada un filamento e l’altro viene acquisito (nei Gram+).
Il DNA penetrato si associa a una specifica proteina di competenza che probabilmente lo protegge dall’attacco delle nucleasi fino a che non raggiunge il cromosoma dove la proteina viene sostituita da RecA. Il DNA viene poi integrato nel genoma del ricevente mediante un processo di ricombinazione.
La competenza può essere indotta artificialmente tramite variazioni nei terreni di coltura, nelle temperature e altri fattori. Tuttavia a tali metodi è preferita la tecnica dell’elettroporazione, che consiste nell’esporre le cellule a campi elettrici pulsanti in modo da aprire piccoli pori nella membrana, facilitando così il passaggio di DNA.

35
Negli eucarioti si usa la pistola a particelle: si spara un proiettile portante tratti di DNA sulla cellula ricevente. LA TRASDUZIONE Nella trasduzione il DNA viene trasferito da una cellula a un’altra attraverso l’intervento di un virus o fago. Tale trasferimento può avvenire in due modi. Nel primo, detto trasduzione generalizzata, qualunque frammento di DNA derivante dal genoma dell’ospite può diventare la componente di DNA di un virus maturo, al posto del genoma del virus. Il secondo, detto trasduzione specializzata, si verifica solo in alcuni virus temperati; il DNA di una specifica regione cromosomica dell’ospite viene integrato direttamente nel genoma del virus, sostituendo, di solito, alcuni geni di questo. La particella virale trasducente risulta difettiva come virus, in entrambe le trasduzioni, poiché i geni batterici hanno sostituito alcuni geni virali indispensabili. La trasduzione generalizzata venne scoperta e studiata nel batterio Salmonella enterica. Quando una popolazione di batteri sensibili viene infettata da un fago, iniziano gli eventi del ciclo litico del fago. Talvolta durante questo processo, gli enzimi responsabili dell’impacchettamento del DNA virale nella testa del fago, impacchettano accidentalmente anche DNA dell’ospite. La particella che ne risulta è detta particella trasducente. Al momento della lisi della cellula tali particelle vengono rilasciate assieme ai virioni normali. Poiché le particelle trasducenti non possono dar luogo a un’infezione virale, (esse non contengono DNA virale) vengono dette difettive. Quando questo lisato viene usato per infettare una popolazione di cellule riceventi, la maggior parte risulta infettata da un virus normale e una piccola aliquota della popolazione riceve particelle trasducenti che iniettano DNA ricevuto dal precedente batterio ospite. Tale DNA non può replicarsi, ma può subire una ricombinazione genetica col DNA del nuovo ospite. I fagi che formano particelle trasducente possono essere virulenti (provocano lisogenia) o temperati (non provocano lisi).

36
La trasduzione specializzata può permettere un trasferimento molto efficiente, a differenza di quella generalizzata (che avviene a bassa frequenza), e può garantire a una piccola regione del cromosoma batterico di venir replicata indipendentemente dal resto. Il primo esempio scoperto di trasduzione specializzata è quella dei geni del galattosio ad opera del fago temperato llll di E.coli. Quando una cellula viene lisogenizzata dal fago l, il genoma fagico si integra nel DNA dell’ospite in un sito specifico in una regione adiacente al gruppo di geni dell’ospite che controllano gli enzimi coinvolti nell’utilizzazione del galattosio (e da quel momento in poi la replicazione del DNA virale risulta totalmente sotto il controllo della cellula ospite). In seguito all’induzione (per esempio con radiazioni U.V.) il DNA virale si separa dal DNA dell’ospite normalmente come una unità. In rare condizioni tuttavia il genoma fagico viene escisso in modo non corretto e alcuni geni del cromosoma batterico adiacenti al fago l integrato (per esempio l’operone del galattosio) sono erroneamente escissi insieme al DNA fagico.

37
Con tecniche di ingegneria genetica è stato possibile costruire un fago lambda trasducente specializzato in grado di contenere geni provenienti da ogni tipo di organismo. Nella trasduzione specializzata solo i geni vicini al punto di integrazione del virus sono trasdotti, ma l’efficienza può essere alta. LA CONIUGAZIONE La coniugazione batterica è un processo di trasferimento di geni che avviene tramite il contatto cellula-cellula e coinvolge un plasmide. Il plasmide è un elemento genetico extracromosomale e l’informazione genetica contenuta in esso non è essenziale alla funzionalità della cellula in tutte le condizioni, ma in alcune può conferire un vantaggio selettivo per la crescita: un esempio è la resistenza agli antibiotici. Il plasmide tipico è una molecola di DNA circolare a doppia elica della dimensione di meno di 1/20 del cromosoma. La maggior parte del DNA plasmidico isolato dalle cellule ha una configurazione superavvolta. I plasmidi che codificano la funzione della coniugazione grazie al contatto cellula-cellula sono detti coniugativi, ma non tutti i plasmidi sono tali. La trasmissibilità mediante coniugazione viene controllata mediante una serie di geni contenuti nei plasmidi detta regione tra. La presenza di questa in un plasmide può avere un’altra importante conseguenza se il plasmide si integra nel cromosoma (tale plasmide è detto episoma), in tal caso il plasmide può mobilizzare il trasferimento del DNA cromosomale da una cellula all’altra. I ceppi batterici che durante la coniugazione trasferiscono una gran quantità di DNA cromosomale sono detti Hfr (alta frequenza di ricombinazione 1000 volte maggiore rispetto ai ceppi F+).
La presenza di plasmidi può avere una profonda influenza nel fenotipo cellulare (produzione di antibiotico, funzioni fisiologiche, resistenza agli antibiotici, virulenza...). In molti casi i plasmidi codificano proprietà fondamentali per il batterio, come la capacità del Rhizobium di interagire con le piante. La costruzione in laboratorio di molti plasmidi artificiali e l’inserimento in essi di geni provenienti da una grande varietà di organismi ha reso possibile il trasferimento di DNA anche attraverso barriere di ogni specie. Un plasmide coniugativo usa la coniugazione per trasferire una copia di se stesso a un nuovo ospite, tuttavia a volte durante la coniugazione, possono essere mobilizzati altri elementi genetici, ovvero altri plasmidi o il cromosoma stesso dell’ospite. La coniugazione venne scoperta proprio grazie al fatto che il plasmide F di E.coli è in grado di mobilizzare il cromosoma dell’ospite. La coniugazione coinvolge un donatore, che contiene un particolare tipo di plasmide coniugativo, e una cellula ricevente, che non lo contiene. I geni che controllano la coniugazione sono contenuti nella regione tra del plasmide e la maggior parte di questi hanno a che fare con la sintesi di una struttura superficiale detta pilo sessuale, che permette l’appaiamento specifico tra la cellula donatrice e quella ricevente. I pili inducono un contatto specifico con un recettore della cellula ricevente e quindi si ritraggono, costringendo così le due cellule a unirsi e, probabilmente in seguito alla fusione delle membrane esterne, il DNA viene trasferito da una cellula all’altra.
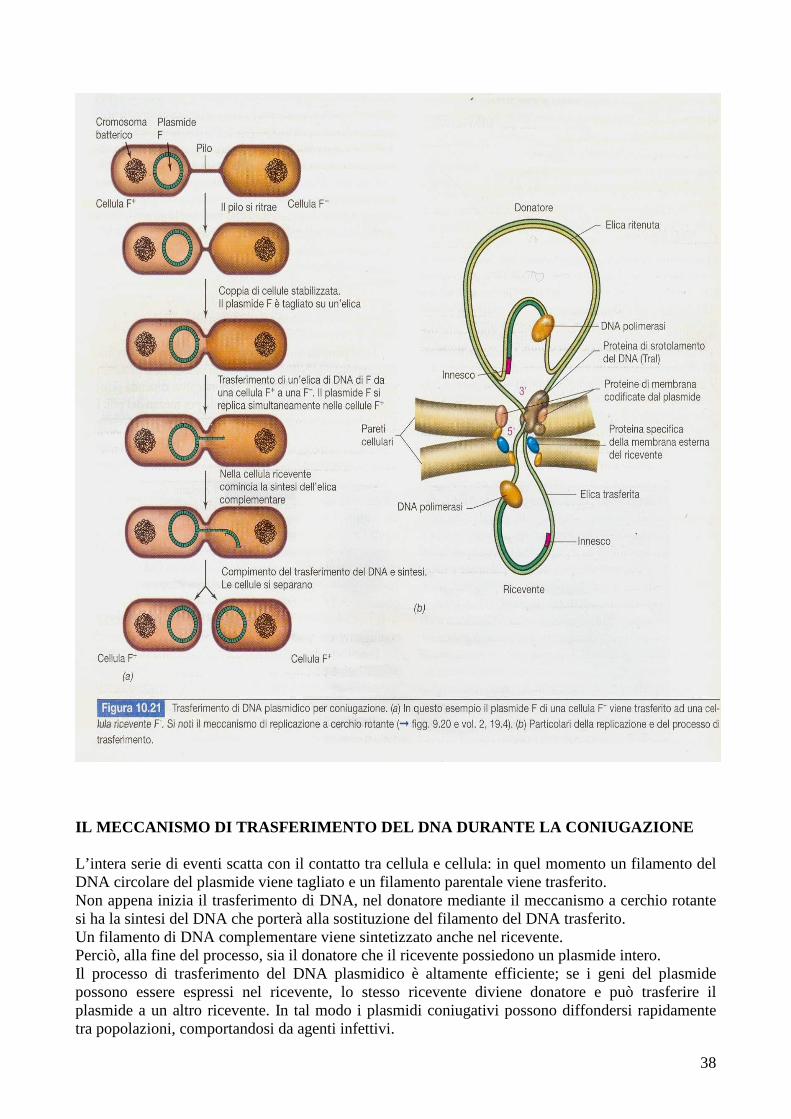
38
IL MECCANISMO DI TRASFERIMENTO DEL DNA DURANTE LA C ONIUGAZIONE L’intera serie di eventi scatta con il contatto tra cellula e cellula: in quel momento un filamento del DNA circolare del plasmide viene tagliato e un filamento parentale viene trasferito. Non appena inizia il trasferimento di DNA, nel donatore mediante il meccanismo a cerchio rotante si ha la sintesi del DNA che porterà alla sostituzione del filamento del DNA trasferito. Un filamento di DNA complementare viene sintetizzato anche nel ricevente. Perciò, alla fine del processo, sia il donatore che il ricevente possiedono un plasmide intero. Il processo di trasferimento del DNA plasmidico è altamente efficiente; se i geni del plasmide possono essere espressi nel ricevente, lo stesso ricevente diviene donatore e può trasferire il plasmide a un altro ricevente. In tal modo i plasmidi coniugativi possono diffondersi rapidamente tra popolazioni, comportandosi da agenti infettivi.

39
Il plasmide F di E.coli può mobilizzare il cromosoma e trasferirlo durante il contatto cellula-cellula. Il plasmide F è un episoma, ovvero un plasmide che può integrarsi nel cromosoma dell’ospite. Se il plasmide F è integrato nel cromosoma , la coniugazione può dar luogo al trasferimento di una larga regione del cromosoma ospite che potrà determinare ricombinazione genetica tra ampie regioni del cromosoma del donatore e del ricevente. Le cellule che possiedono il plasmide F non integrato sono dette F+ e quelle che hanno il plasmide integrato nel cromosoma sono dette Hfr. Le cellule senza plasmide F sono dette F- e agiscono da riceventi. La presenza del plasmide F porta a tre cambiamenti nelle proprietà della cellula:
- capacità di sintetizzare il pilo F - mobilizzazione del DNA per il trasferimento in un’altra cellula - alterazione dei recettori di superficie in modo che la cellula non sia più in grado di
comportarsi come un ricevente nella coniugazione L’integrazione del plasmide F in un cromosoma ospite può avvenire in vari siti specifici, detti IS (sequenza di inserzione), che costituiscono regioni di omologia tra il cromosoma e il plasmide F. Di solito, poiché durante il trasferimento si ha facilmente la rottura del pilo , e di conseguenza del filamento di DNA, solo una parte del cromosoma del donatore viene trasferito, per cui non si può replicare nella cellula ricevente. Pertanto i geni presenti sul cromosoma del donatore sono identificabili solo se avviene ricombinazione tra la porzione del cromosoma del donatore che è entrata e il cromosoma della cellula ricevente. Sebbene i ceppi Hfr trasmettono geni cromosomali ad alta frequenza, essi non convertono le cellule F- in cellule F+ in quanto solo raramente viene trasferito l’intero plasmide F. D’altro lato le cellule F+ convertono in modo efficiente F- in F+ proprio perché viene trasferito l’intero plasmide F. Quindi affinché un ceppo possa diventare F+ o HFR deve penetrare tutto il plasmide. Poiché negli Hfr normalmente l’episoma non entra quasi mai (perché, mentre entra il cromosoma batterico linearizzato, spesso si spezza il pilo lasciando fuori il plasmide), normalmente si lavora con plasmidi non integrati e successivamente si fanno integrare artificialmente. Occasionalmente i plasmidi F integrati possono escindersi dal cromosoma e durante questo processo essere inseriti dei geni cromosomici all’interno del plasmide F libero, questo fenomeno può avvenire in quanto il plasmide integrato e il cromosoma contengono un certo numero di sequenze identiche tra le quali può avvenire la ricombinazione. I plasmidi F contenenti geni cromosomici sono detti plasmidi F’. I plasmidi sono diventati strumenti indispensabili per la biologia molecolare e per lo studio genetico dei microrganismi perché: • Sono molto piccoli ed è conosciuta la loro costituzione genetica. Quindi si sa dove poter inserire
il/i gene/i che vogliamo trasferire. (gli enzimi di restrizione tagliano in determinati siti e le ligasi si occupano dell’inserimento).
• Sono caratterizzati da facilità di moltiplicazione autonoma, quindi si può amplificare molte volte il gene che voglio trasferire.
• Sono facili da trasmettere ad un eventuale ospite, dove può avvenire l’amplificazione. Anche nello studio dei lieviti per avere un gene per un dato carattere, è possibile comprare un plasmide in cui è inserito il gene che interessa. Il plasmide è un vettore di clonaggio. L’E. coli (ospite del plasmide) è importante perché è il batterio più conosciuto (genoma noto) ed è facilmente coltivabile perché si adatta bene in terreni diversi.

40
Tale batterio ha il difetto che alcuni ceppi sono patogeni intestinali, essendo un microrganismo Gram- di origine enterica-intestinale.

41
I LIEVITI
Una cellula eucariotica è strutturalmente più complessa di una procariotica; in questa il DNA non è confinato in un organello specifico, mentre le cellule eucariotiche contengono un nucleo circondato da membrana (in esso il DNA è sotto forma di cromosomi) ed è spesso visibile una struttura detta nucleolo, sede della sintesi dell’ RNA ribosomale. Differenze tra lieviti e batteri :
• I batteri sono procarioti , i lieviti sono eucarioti eterotrofi . • I batteri hanno un cromosoma circolare, i lieviti hanno più cromosomi dentro il nucleo • I batteri hanno una moltiplicazione agamica per scissione binaria (schizomiceti), i lieviti per
gemmazione (blastomiceti), infatti si formano piccole gemme che si ingrandiscono e si separano dalla cellula madre.
• Nella moltiplicazione gamica nei batteri abbiamo i tre sistemi (trasformazione, trasduzione e coniugazione), mentre nei lieviti si ha una riproduzione tramite gameti aploidi (ascospore in grado di coniugare, diverse dalle spore durevoli dei batteri che sono strutture di resistenza diploidi).
I lieviti li dividiamo in tre gruppi: aplonti, diplonti, aplodiplonti.
• Ciclo aplonte La maggior parte del ciclo vitale è nella fase aploide (n). I lieviti, in certe condizioni di crescita, cessano la gemmazione e la cellula vegetativa si trasforma in un asco che contiene quattro spore (con sessualità diversa: + e - oppure a e a). Ogni spora da luogo a n generazioni di cellule vegetative aploidi e con la coniugazione a due a due si formano individui diploidi. Le cellule diploidi (zigoti) sono la generazione breve e vanno subito in meiosi riformando i gameti (spore n ).
• Ciclo diplonte La maggior parte del ciclo vitale è nella fase diploide (2n). Appena si formano le spore non si hanno generazioni ed esse coniugano subito formando zigoti 2n che danno luogo a n generazioni diploidi; successivamente si ha meiosi e si riformano le quattro spore.
• Ciclo aplodiplonte Nel ciclo si ha l’alternarsi di generazioni aploidi e diploidi quindi sia lo zigote 2n che le spore n danno origine alle rispettive generazioni. Tale ciclo è tipico dei lieviti Saccaromyces . I lieviti hanno due diverse forme di cellule aploidi, chiamati tipi sessuali, che possono essere considerate l’analogo dei gameti maschili e femminili. Dall’accoppiamento di due tipi sessuali opposti si forma una cellula diploide da cui si ha la formazione di una struttura contenente quattro gameti, due per tipo sessuale, detta asco. E’ possibile trovare aschi con meno di quattro spore , ciò perché alcune sono morte; per meiosi si formano sempre quattro spore.

42
I lieviti sono funghi unicellulari (la maggior parte di essi è classificata negli Ascomiceti) che si riproducono agamicamente per gemmazione (eccetto il lievito Schizosaccaromyces che usa la scissione come i batteri). Sono ubiquitari e li troviamo nel suolo, nelle acque, negli apparati e tessuti animali, sulla frutta e sulle piante. Hanno diverse funzioni e quella più importante è di fungere da biodegradatori di composti complessi in composti più semplici. In presenza di O2 ossidano e trasformano i composti organici in H2O e CO2; in fase anossica producono prevalentemente alcol e CO2. Quindi ci sono lieviti strettamente aerobi e anaerobi facoltativi (vivono sia in anossia che in presenza di O2). I responsabili delle fermentazioni sono i lieviti. Esistono però anche lieviti patogeni, non quanto i batteri, sia per le piante (marciume acido dell’uva) che per uomini e animali (Candida albicans e Criptococcus neoformans che causa meningite). Sebbene la maggior parte dei lieviti si riproduca e esista come singola cellula, in certe condizioni alcuni possono formare filamenti; in queste specie la forma filamentosa è essenziale per l’espressione della patogenicità (Candida albicans causa infezioni vaginali, orali e polmonari). Tutti i lieviti che producono un velo biancastro sulle superfici liquide sono candide non patogeniche.

43
I funghi sono caratterizzati dalla presenza di una parete cellulare e dalla produzione di spore. Tre sono i gruppi di maggiore importanza: le muffe , i lieviti e funghi fruttiferi . I funghi mancano di pigmenti fotosintetici e possono essere sia unicellulari (lieviti) sia filamentosi (muffe). Essi sono i maggiori biodegradatori conosciuti in natura e i maggiori riciclatori di sostanza organica nel suolo e in altri ecosistemi. Le muffe producono aflatossine dannose mentre i lieviti ostacolano la formazione di esse. I funghi hanno habitat diversificati e la maggior parte di essi ha habitat terrestre, nel suolo o su materiale vegetale in decomposizione. Molti funghi sono parassiti di piante e sono responsabili della maggior parte delle malattie in esse; alcuni sono anche parasiti di animali sebbene costituiscano agenti patogeni meno importanti dei batteri e dei virus.
MORFOLOGIA DELLE SPORE DEI LIEVITI
Forme :
• A elmetto • Aghiformi (genere Metschnikowia importante nel vino ) • A noce • A saturno • A cappello • Sferoidale Le più conosciute nel Saccaromyces • Ovoidale • Reniforme
MORFOLOGIA DELLE CELLULE DEI LIEVITI
• Cellule sferiche (Saccaromyces cerevisae di cui è conosciuto tutto il genoma) • A forma di limone (dà una gemmazione bipolare tipica di lieviti apiculati, quelli che
iniziano la fermentazione alcolica) • Ellittiche o a pera • Ovali allungate • Rotonde con globuli di grasso (Pulcherrima) • Grandi e allungate, formanti un rudimentale pseudomicelio (Schizosaccaromyces) • Forma allungata stretta in catena ramificata (Candida)
L’ ifa è il filamento di una singola cellula e l’aggregazione di più ife forma il micelio. I lieviti non formano mai vere ife, ma possono formare pseudoife, quando la gemma che si forma non si stacca dalla cellula madre e a sua volta crea un’altra gemma figlia e così via, determinando strutture ramificate (sulla superficie delle salamoie si formano veli biancastri dovuti a lieviti del genere Candida, Pichia e Hansenula che formano pseudoife).

44
LIEVITI DI INTERESSE IN MICROBIOLOGIA ALIMENTARE
Phylum ASCOMYCOTA (la cellula si trasforma in asco che contiene le ascospore) Classe EMYASCOMICETES (aschi liberi) e EUASCOMYCETES (aschi in corpi fruttiferi) Ordine SCHIZOSACCHAROMYCETALES Appartengono famiglie di interesse agro-alimentare (trasformazione prodotti) e alcuni prettamente di interesse enologico
• Famiglia Schizosaccharomicetaceae Unica famiglia di lieviti che si riproduce per scissione, come nei batteri Genere e specie Schizosaccharomyces pombe Schizosaccharomyces octosporus (forma otto spore)
Li troviamo negli alimenti molto zuccherini (miele e succhi di frutta) perché hanno una elevata osmotolleranza. Inoltre nel vino sono presenti nella trasformazione malo-alcolica
Classe EMYASCOMYCETES Ordine SACCHAROMYCETALES (o endomicetales perché le spore sono interne) Tale ordine ha molte famiglie:
• Famiglia Metschnikowiaceae Hanno cellule sferiche e spore aghiformi Genere Metschnikowia Facilmente distinguibile dagli altri lieviti perché all’interno la cellula ha dei granuli di lipidi rifrangenti visibili al microscopio ottico. A livello colturale
produce culchermina (un pigmento carotenoide) presente intorno alla colonia come alone rosso mattone (specie “bella a vedersi”). E’ un lievito negativo nel mosto, dove si trova all’inizio della fermentazione, perché produce poco alcol e molto acido acetico.
• Famiglia Lipomycetaceae Genere Lipomyces
È presente nel suolo ed è uno dei pochi lieviti che forma capsula che aiuta a formare nicchie nel terreno in grado di migliorarne la struttura. La capsula attrae sostanze dannose ed adsorbe fitofarmaci e metalli pesanti, migliorando la qualità del terreno. E’ un lievito usato nella biorimediaton (disinquinamento tramite microrganismi).
• Famiglia Saccharomycetaceae è la più importante nel settore alimentare genere Saccharomyces è uno dei primi lieviti descritti dai ricercatori (1838 da Meyen); il nome significa
“fungo dello zucchero” e partecipa alla sua fermentazione. E’ il lievito per eccellenza coinvolto in tutte le fermentazioni agro-alimentari (vino, birra, pane) dove ci sono zuccheri da fermentare. La specie Saccharomyces cerevisae è di interesse elevatissimo, ha forma ellittica-rotonda, è sporigeno, ha un elevato interesse biotecnologico, si utilizza come starter e conclude la fermentazione nei vini perché ha il più alto potere fermentativo (percentuale di alcol che un lievito riesce a produrre e può arrivare anche al 18%)
genere Debariomyces è simile al Saccaromyces (gemmazione multilaterale, cellule sferiche, sporigeno)
ma si distingue per la superficie delle spore rugosa. Biochimicamente non fermenta

45
come il saccaromyces; lo ritroviamo nei formaggi perché in grado di attaccare i grassi (lievito lipolitico)
specie Debariomyces hansemi è lipolitico ed è importante nella maturazione degli insaccati, infatti insieme ad
alcune muffe si sviluppa all’esterno del budello e partecipa alla maturazione della carne (fermentazione lattica interna)
genere Decchera è dannoso nel settore alimentare perché si può sviluppare nei prodotti fermentati in
bottiglia (momento più delicato); il genere Decchera che è la forma perfetta (produce spore), insieme al genere Bretanomyces che è la forma imperfetta (non produce spore e usa solo via agamica), alterano le bevande alcoliche in bottiglia (come il vino ) producendo un sentore di fenolo
genere Issatchenkia è uno dei pochi lieviti che altera la frutta zuccherina producendo il marciume
acido; le malattie dell’uva sono causate normalmente da muffe, eccetto questo lievito che penetra nell’acino lacerato e produce acido acetico utilizzando lo zucchero.
genere Cluiveromyces è simile al saccharomyces (gemmazione multipolare) ma si distingue perché l’asco
a maturità si rompe facilmente rilasciando delle spore che si agglutinano (avvicinano). E’ un lievito importante per il settore lattiero-caseario (specie Cluiveromyces lactis) nei latti fermentati alcolici (kefir)
generi Pichia e Hansenula (perfetti: producono spore) e Candida (imperfetto; la famiglia delle Candidaceae ha solo lieviti imperfetti )
formano pseudomiceli, hanno metabolismo prevalentemente ossidativo e sono presenti nel settore agro-alimentare quasi sempre sulla superficie dei liquidi (perché hanno metabolismo aerobico); ne sono esempi il velo bianco sulle olive in salamoia, la fioretta del vino che lo porta alla malattia dello spunto.
genere Torulaspora è simile al saccaromyces ma è più piccolo e produce più gemme multilaterali. Si
trova nel vino ed ha un “metabolismo puro” in quanto produce alcol senza produrre sostanze negative (come l’acido acetico, dannoso nelle fermentazioni). L’acido acetico si produce sempre nelle fermentazioni perché è un prodotto secondario ed è importante scegliere quei lieviti che ne producono di meno o per niente (lieviti puri). Tale genere, per il suo peculiare metabolismo, in passato si usava nella spumantizzazione perché ha la capacità di flocculare (riunirsi in fiocchi) creando vini molto più limpidi.
genere Zigosaccharomices le cellule sono ovali e si individuano facilmente perché nella fase coniugativa
formano un ponte (immagine a occhiali; cellule tipo “zigo” unione simmetrica). E’ il lievito più resistente negli ambienti zuccherini (osmofilo) ed è agente dell’unica malattia del miele (la fermentazione); il miele ha una percentuale di zucchero tra il 68-72%, quindi una concentrazione quasi impossibile per la vita dei microrganismi (per conservare le marmellate aumento la concentrazione di zucchero). Le specie Zigosaccharomyces bailii e rauxii vivono anche a concentrazioni di zucchero maggiori del 70% quindi riescono a svilupparsi sulla superficie del miele per poi penetrare all’interno.
Tale genere può alterare anche i succhi di frutta e in enologia formano un velo nei vini Cherry (malvasia di Bosa) dopo la fermentazione, e formano il velo nella vernaccia producendo sostanze che danno un profumo particolare al vino (non è la fioretta che altera negativamente producendo acido acetico).

46
• Famiglia Saccharomycodaceae Sono i lieviti apiculati (forma a limone e gemmazione bipolare) e sono quelli che
danno inizio alla fermentazione alcolica. Genere Hansenihaspora (forma perfetta) e Kloeckera (forma imperfetta) Genere Saccharomycodes Resiste ad elevati stress e ad elevate concentrazioni di anidride solforosa. E’ il
lievito più grosso conosciuto (in rapporto al volume in una cellula più grande entrano meno sostanze). E’ l’unico lievito che altera i mosti muti (mosti di uve fresche mutizzato per aggiunta di alcol di origine vinica oppure reso muto tramite aggiunta di elevate dosi di anidride solforosa).
MICROBIOLOGIA ENOLOGICA LA FERMENTAZIONE SPONTANEA Analizzando microbiologicamente il mosto appena ottenuto osserviamo che in esso è presente la medesima composizione microbiologica presente nell’ecosistema dell’uva (terreno, uva, tralci, attrezzature utilizzate). Quindi troviamo batteri, lieviti e muffe. Le muffe sono i microrganismi maggiormente rappresentati nell’ecosistema “uva” (peronospora, oidio, botritis) e con essi anche i batteri. In un millilitro di mosto appena ottenuto vi è un numero elevato di muffe e batteri (107-108 cellule) mentre è presente un numero limitato di lieviti (104-105 cellule). Tuttavia la fermentazione del mosto esercita un’azione selettiva e alla fine predomineranno i lieviti . I fattori che determinano tale azione selettiva sono:
1. l’assenza di O2 determina l’eliminazione delle muffe (che sono strettamente aerobie) 2. il pH , tendenzialmente acido (circa 3-4), elimina quasi tutti i batteri perché la maggior
parte di essi ha un pH ottimale intorno a 7. Gli unici batteri che rimangono illesi sono i fermenti lattici, che sopravvivono a pH acido
3. la presenza di zuccheri aiuta i lieviti, che li degradano in alcol e CO2. I lieviti principali che si trovano sulla superficie degli acini sono quelli a metabolismo ossidativo o quelli che fermentano con basso potere fermentativo; ci sono pochi saccaromyces (provenienti dall’ambiente “cantina” e con il più alto potere fermentativo) e molti lieviti con basso potere fermentativo:
1. lieviti apiculati: Hanseniaspora e l’omologo imperfetto Kloeckera hanno potere fermentativo 4-6% di alcol e pochi 8% .
2. lieviti del genere Metschinkowia hanno potere fermentativo 2-3% di alcol. In Sardegna sempre presente all’inizio della fermentazione.
3. lieviti con metabolismo ossidativo a carico dell’alcol etilico: Candida, Pichia, Hansenula non producono alcol e se resistono fino alla fine della fermentazione formano la patina bianca e producono acidità rovinando il vino (fioretta in superficie che porta alla malattia dello spunto).
4. Rhodotorula e Criptococcus Muoiono subito

47
DIFFERENZE FRA LIEVITI APICULATI ED ELLITTICI I lieviti ellittici sono quelli di maggiori dimensioni e presentano alcune caratteristiche positive non riscontrabili negli altri lieviti:
1. maggiore resa nella trasformazione dello zucchero in alcol etilico (1.7 g per formare 1 ml di alcol; mentre gli apiculati ne abbisognano 2.1 g ).
2. maggiore resistenza all’alcol etilico (catabolita del lievito e perciò dannoso allo stesso). 3. scarsa produzione di acido acetico che, oltre certi limiti (la legge ne fissa un max),
disturba dal punto di vista organolettico. 4. maggiore resistenza all’anidride solforosa
Il lievito Saccharomyces cerevisiae, di caratteristica forma ellittica , è molto resistente all’alcol etilico ed è il maggiore responsabile della fermentazione della maggior parte dello zucchero; è anche molto resistente all’anidride solforosa. Esiste in un numero elevato di ceppi, cioè di individui derivati (o selezionati) a partire da un’unica cellula. Il lievito Saccharomyces oviformis è in grado di produrre vini con il 18% di alcol; è molto resistente all’anidride carbonica, ragion per cui è utile nella produzione di vini spumanti e per le rifermentazioni in bottiglia . Nella Vernaccia di Oristano forma un velo detto “fiore” (è il micelio del fungo) sulla superficie del vino, e per tale motivo il lievito in questo caso è detto lievito filmogeno (Fleur in Francia e Flor in Spagna). Ha la caratteristica di ossidare l’acido acetico, diminuendo perciò l’acidità volatile; l’acido acetico è in parte ossidato completamente con liberazione di H2O e CO2, mentre in parte è ossidato ad aldeide acetica; in parte è ridotto anche l’alcol ad aldeide acetica: si formano molti acetali che conferiscono al vino un profumo piacevole. È bene che i lieviti ellittici non siano i soli a condurre la fermentazione, in quanto si è constatato che i lieviti apiculati originano prodotti secondari che completano e affinano il profumo del vino. I lieviti apiculati sono poco alcoligeni, producono molto acido acetico e sono scarsamente resistenti all’anidride solforosa. Sono comunque attivi nell’iniziare la fermentazione. L’ambiente dei saccharomyceti è la cantina, ma possono essere presenti anche in vigna; quando inizia la fermentazione del mosto alcuni saccharomyceti penetrano in esso e iniziano a moltiplicarsi (fino ad avere un numero di 100 miliardi di cellule per ml); quasi mai iniziano la fermentazione spontanea perché sono pochi e sono deputati nel portare a termine la fermentazione, utilizzando lo zucchero disponibile, fino a 13o-14o di alcol e alcuni anche fino a 18o. Un lievito produce tanto alcol quanto riesce a resisterne; mediamente un lievito produce 0,6 g di alcol per 1 g di zucchero consumato (rendimento). Ovviamente sono migliori i lieviti che producono più di 0,6 g perché hanno un maggiore rendimento. La quantità di zuccheri presenti nel mosto è quindi molto importante poiché il saccharomyces produce alcol in funzione di essa. Con il 20% di zucchero si produrrà circa il 12% di alcol; con il 30% di zucchero si produrrà circa il 18% di alcol (massimo valore possibile). Per sapere quanto alcol un lievito può produrre bisogna avere un mosto con almeno il 30% di zucchero.
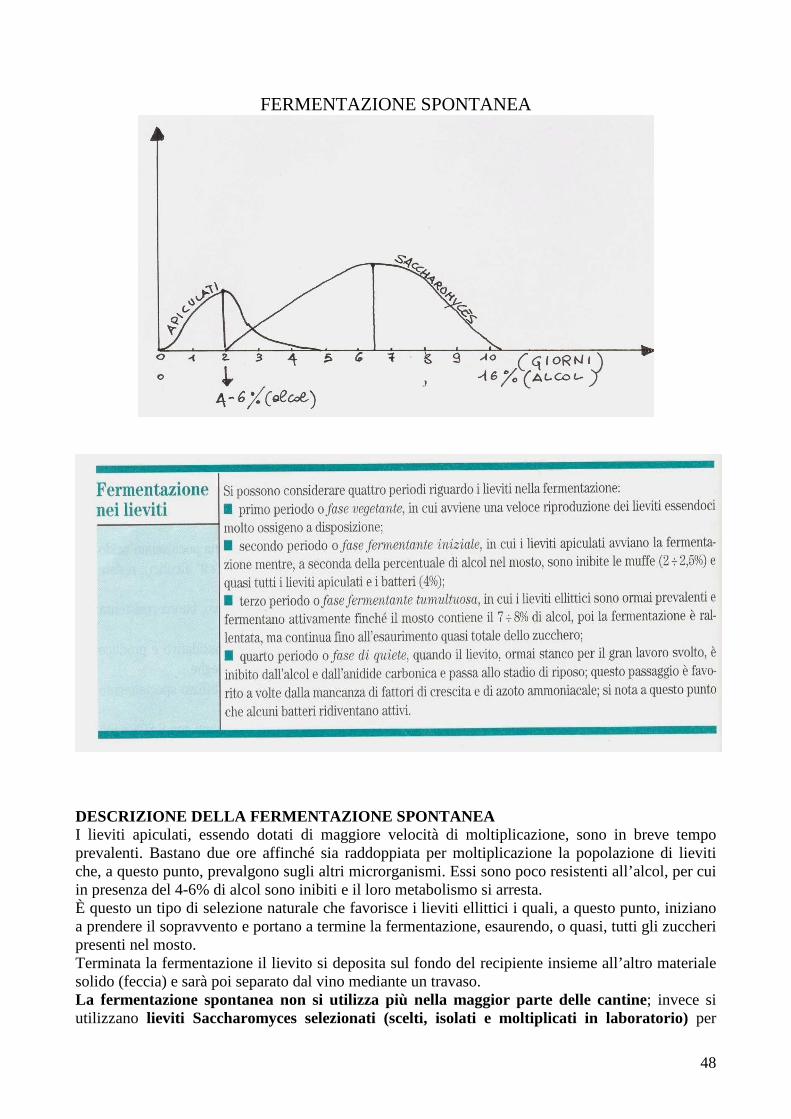
48
FERMENTAZIONE SPONTANEA
DESCRIZIONE DELLA FERMENTAZIONE SPONTANEA I lieviti apiculati, essendo dotati di maggiore velocità di moltiplicazione, sono in breve tempo prevalenti. Bastano due ore affinché sia raddoppiata per moltiplicazione la popolazione di lieviti che, a questo punto, prevalgono sugli altri microrganismi. Essi sono poco resistenti all’alcol, per cui in presenza del 4-6% di alcol sono inibiti e il loro metabolismo si arresta. È questo un tipo di selezione naturale che favorisce i lieviti ellittici i quali, a questo punto, iniziano a prendere il sopravvento e portano a termine la fermentazione, esaurendo, o quasi, tutti gli zuccheri presenti nel mosto. Terminata la fermentazione il lievito si deposita sul fondo del recipiente insieme all’altro materiale solido (feccia) e sarà poi separato dal vino mediante un travaso. La fermentazione spontanea non si utilizza più nella maggior parte delle cantine; invece si utilizzano lieviti Saccharomyces selezionati (scelti, isolati e moltiplicati in laboratorio) per

49
alcuni caratteri, che vengono inoculati fin da subito nel mosto con una concentrazione di 106 cellule per ml (essendo molto numerosi iniziano velocemente la fermentazione e gli apiculati spariscono rapidamente).
STORIA DELLA MICROBIOLOGIA ENOLOGICA
• 1680 Van Leuvenoek è il commerciante di seta che inventò il primo microscopio semplice
per vedere le fibre della seta; osservandole trovò dei piccoli microrganismi, li descrisse in una relazione scritta in olandese e la inviò alla reale società scientifica di Londra; qui la scoperta rimase nell’oblio fino al 1818.
• 1818 Erxleben e Schwan ipotizzarono che i lieviti fossero responsabili della fermentazione dei vini
• 1830 Kutzing scoprì la presenza di lieviti nel mosto • 1838 Meyen descrive i lieviti responsabili nella fermentazione del mosto (Saccharomyces) • 1866 con Pasteur si accetta l’idea che il processo fermentativo è attuato dai lieviti
Saccharomyces; scrisse i primi trattati di microbiologia agraria, di cui ne è fondatore, occupandosi anche del vino, dimostrando che gli agenti di fermentazione si trovano anche sulla superficie dell’acino e che durante la maturazione dell’uva raggiungevano una alta densità di popolazione
• 1870 Rees scopre i saccharomyceti apiculati • 1872 Pasteur dimostra che i lieviti fermentativi del mosto risiedono sulla superficie degli
acini presenti in ambiente naturale (gli acini protetti non fermentano) • 1881 Hansen descrive l’associazione dei lieviti apiculati con la superficie dei diversi frutti • 1890 Hansen isola i lieviti in coltura pura • 1891 Martinand e Rietsch stabiliscono l’associazione lieviti apiculati-uve mature e la
successione apiculati-ellittici • 1896 si osserva che gli apiculati predominano nei terreni dei frutteti • 1908 Hansen utilissa lieviti selezionati nell’industria della birra “sotterrando” la
fermentazione spontanea • 1912 Guillermond scopre che i lieviti in autunno passano dal terreno alla superficie degli
acini • 1935 De Rossi a Perugia è il primo a fare isolamenti sistematici dai mosti (87 mosti umbri),
indicando anche i metodi di isolamento • 1939 con Castelli si dà il via all’isolamento dei mosti delle principali regioni viti-vinicole
d’Italia • 1964 Augusto Capriotti , allievo del prof. Castelli venne in Sardegna creando (attivo da 40
anni) l’istituto di microbiologia di Sassari; isolò dei lieviti dalla Vernaccia (oggi si conoscono i lieviti di tutti i vini della Sardegna)
• 1970 Kunkee e Amerine in California elencarono lieviti trovati sulle uve di varie parti del mondo
Tutti questi studi sono serviti per potere selezionare lieviti da usare nelle diverse produzioni e oggi ogni cantina usa lieviti selezionati per ottenere un prodotto desiderato . Fra i parametri più importanti da considerare nella scelta del lievito vi è l’attività fermentativa (produzione di alcol nell’unità di tempo)

50
Considerando 3 ceppi e la loro produzione di alcol giornaliera osserviamo che il ceppo 1 risulta troppo vigoroso con liberazione di troppa energia (da 1 mole di glucosio si producono 40 Kcal, di cui 14 Kcal sono usate dal lievito e 26 Kcal sono liberate, con conseguente aumento della T che favorisce l’insorgere di batteri termofili e di alcune malattie del vino). I LIEVITI SELEZIONATI
I lieviti selezionati sono colture pure di specie ecologicamente idonee con caratteristiche tali da consentire il conseguimento di risultati programmati.

51
La risposta che può dare un processo in cui si utilizzano microrganismi non è sempre prevedibile; infatti grazie alla variabilità biologica è possibile ottenere risposte diverse. Quindi si è pensato all’utilizzo di colture pure in grado di garantire un determinato risultato finale. Selezionare vuol dire prendere all’interno della popolazione un certo numero di individui e selezionarli per i caratteri che a noi interessano per poi utilizzarli nel modo più opportuno. Ci sono industrie che preparano gli starter sotto forma liquida (la prima che si è sviluppata)
Oggi si utilizzano le colture LSA (lievito secco attivo), perché ,applicato al momento della vendemmia, quello liquido può dare dei problemi. Lo starter sotto forma secca è un lievito in pasta che viene essiccato con aria tiepida in lamiere oscillanti finchè si ha la formazione di palline o filini. Il lievito perde acqua ma mantiene la vitalità e può essere conservato per molti mesi. Il lievito secco al momento dell’utilizzo viene sciolto in acqua tiepida e versato nel recipiente da fermentare (20g/hl). Ci sono così 109 cellule per grammo cioè una quantità superiore a quella che si forma in una fermentazione spontanea.
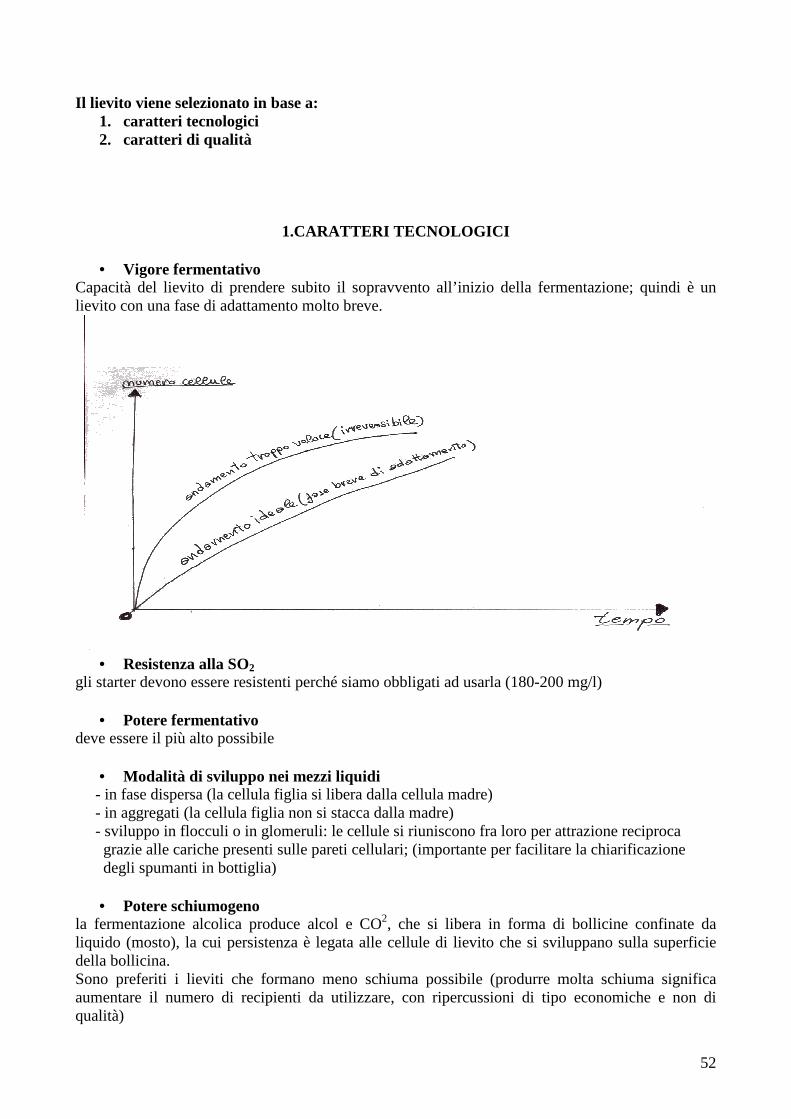
52
Il lievito viene selezionato in base a: 1. caratteri tecnologici 2. caratteri di qualità
1.CARATTERI TECNOLOGICI
• Vigore fermentativo Capacità del lievito di prendere subito il sopravvento all’inizio della fermentazione; quindi è un lievito con una fase di adattamento molto breve.
• Resistenza alla SO2
gli starter devono essere resistenti perché siamo obbligati ad usarla (180-200 mg/l)
• Potere fermentativo deve essere il più alto possibile
• Modalità di sviluppo nei mezzi liquidi - in fase dispersa (la cellula figlia si libera dalla cellula madre) - in aggregati (la cellula figlia non si stacca dalla madre) - sviluppo in flocculi o in glomeruli: le cellule si riuniscono fra loro per attrazione reciproca grazie alle cariche presenti sulle pareti cellulari; (importante per facilitare la chiarificazione degli spumanti in bottiglia)
• Potere schiumogeno la fermentazione alcolica produce alcol e CO2, che si libera in forma di bollicine confinate da liquido (mosto), la cui persistenza è legata alle cellule di lievito che si sviluppano sulla superficie della bollicina. Sono preferiti i lieviti che formano meno schiuma possibile (produrre molta schiuma significa aumentare il numero di recipienti da utilizzare, con ripercussioni di tipo economiche e non di qualità)

53
• Resistenza ai fitofarmaci I fitofarmaci dalla vigna possono arrivare al mosto e i lieviti possono resistere o soccombere, in funzione della quantità residua. E’ bene usare lieviti che resistono bene ai fitofarmaci; in certi casi riescono anche a degradarli (è pericoloso perché non si sa quale potrebbe essere il prodotto di degradazione) o ad assorbirli (il fitofarmaco viene trascinato sul fondo quando il lievito muore)
• Carattere “flor” film superficiale di alcuni lieviti che maturano particolari vini (florizzati) come lo Sherry spagnolo (il più importante nel mondo), la Malvasia e la Vernaccia. Per ottenere tali vini il ceppo starter deve avere il carattere “flor”.
• Fattore “killer” Il lievito può uccidere o inibire altre cellule attraverso la secrezione di una glicoproteina, scoperta nel 1966. Tale fattore è importante e agisce a pH maggiore di 4,8 (anche nel vino, che ha un pH minore, la glicoproteina viene secreta). I lieviti N (neutri) non hanno la tossina killer e non sono inibiti dai lieviti killer. I lieviti K (killer) hanno la proteina killer. I lieviti S (sensibili) non hanno proteine killer e sono inibiti dai lieviti killer. In Sardegna solo lo 0,17% de lieviti ha il fattore killer; tale fattore non è importante in enologia perché non agisce su tutti i lieviti, per esempio sulla Metschinkowia e sugli Apiculati; agisce invece sui Saccharomyces. E’ necessario che il lievito killer sia presente al 70 % rispetto al lievito da inibire per espletare la funzione killer, ma in tali condizioni anche un ceppo normale riesce ad avere il sopravvento.
• Temperatura di fermentazione I lieviti sono mesofili cioè vivono bene con temperature intorno ai 25-35oC; si cercano ceppi starter resistenti alle basse temperature, come quelle delle cantine (16-18 oC) .
2.CARATTERI DI QUALITA’
• azione sull’acido malico: se in eccesso bisogna eliminarlo tramite la fermentazione malo-lattica o quella malo-alcolica.
•••• E’ possibile usare ceppi di Saccharomyces che riescono a degradare l’acido malico durante la fermentazione. Ci sono anche lieviti che producono acido malico a partire dall’acido piruvico e sono utili se voglio mantenere o aumentare la quantità di acido malico nel vino; successivamente bisogna sterilizzare il vino per evitare che i batteri degradino l’acido malico prodotto
•••• produzione di composti solforati: lo zolfo è preso dai solfati derivanti dalla concimazione o dalla SO2. La solforosa prodotta dal lievito è sempre combinata con i composti accettori di solforosa quindi non utile poiché priva di azione antisettica. SO4
- (solfati) HSO3- (ione metasolfito) H2S (idrogeno solforato)
L’idrogeno solforato H2S conferisce al vino odore di uova marce (nei vini freschi posso evitarlo facendo dei travasi). E’ meglio che il lievito non produca composti solforati.
•••• produzione di accettori di solforosa : i lieviti starter ne devono produrre il meno possibile .
•••• purezza fermentativa: capacità del lievito di produrre solo certi composti; il lievito non deve mai produrre acido acetico e deve produrre quantità discrete di glicerolo (12-15 g/l) che nei vini rossi dà morbidezza.

54
IMPIEGO DELL’ANIDRIDE SOLFOROSA
L’anidride solforosa è un gas dal caratteristico odore di fiammifero acceso che si forma dalla combustione dello zolfo. La solforosa non è solo esogena ma viene prodotta nel vino stesso. Il suo uso in enologia è antichissimo ed ancora oggi non si è trovato un valido sostituto (ai tempi di Omero lo zolfo veniva usato per la conservazione degli alimenti nonchè per fini salutistici per esempio le acque sulfuree), alla fine dell’800 fu introdotto nella pratica enologica ed è l’unico antisettico consentito per legge. L’SO2 ha però alcuni effetti collaterali : è irritante , ha sapore sgradevole ,può causare mal di testa e altri disturbi, di conseguenza la legge ne consente un uso limitato. La quantità di solforosa totale non è tutta utile ai fini enologici, quella utile per i nostri scopi è la solforosa libera ( SOLFOROSA LIBERA + SOLFOROSA COMBINATA =SOLFOROSA TOTALE) . Nei mosti e nei vini l’SO2 si ritrova : • In forma libera : in soluzione acquosa o idroalcolica ( come è il vino ) forma vari idrati che
si dissociano liberando ioni H+ .La solforosa libera , cioè non legata ad alcun composto,è un importante antisettico ,questa la possiamo trovare come solforosa molecolare(SO2, che è anche la forma più attiva ), sia come solforosa indissociata (sotto forma di ione bisolfito HSO3
-, ugualmente attiva ma meno rispetto alla molecolare). In genere si tende a far prevalere la forma molecolare agendo sul pH (più è acido e maggiore sarà la componente SO2) e sulla temperatura (più è alta e maggiore sarà la frazione di solforosa molecolare), purtroppo questi due parametri si trovano in contrasto perché la scienza enologica prevede un pH acido (3.2) ma con temperature che non superino certi livelli per i vini bianchi e pH più alti (3.6) e temperature alte (circa 28°C ) per vini rossi ; un altro fattore che determina la quantità finale di solforosa è la quantità di acido piruvico che si trasforma in alcol etilico (con cui l’SO2 non si combina); il passaggio acido piruvico alcol etilico è regolato da enzimi come la piruvato decarbossilasi .Ogni enzima per funzionare ha bisogno di un coenzima( in questo caso la tiamin-pirofosfato detta anche CO-CARBOSSILASI) . Il punto debole della solforosa è la TIAMINA o vitamina B1 che risulta particolarmente sensibile alla solforosa ( distruggendo la tiamina si disattiva il co-enzima con conseguente accumulo di acido piruvico e aumento della quantità di solforosa combinata ) ;la legge consente un aggiunta di tiamina fino ad un massimo di 0,6 mg/l. L’acido solforoso che probabilmente si forma non è però mai stato isolato :
SO2(gas) SO2 acquosa SO2 acquosa + H2O HSO3
- (bisolfito )+ H+
HSO3
- + HSO3- S2O5
2-(metabisolfito o pirosolfito)+ H2O HSO3
- H+ + SO3- (solfito)
Si ottengono quindi bisolfiti , metabisolfiti e , quando il pH è superiore a 5 , anche solfiti .
• In forma legata (combinata ) : l’ SO2 e i suoi derivati si combinano con molecole che presentano funzioni carbonilica, aldeidica e chetonica , come l’aldeide acetica , gli zuccheri, l’acido piruvico ma anche con proteine e polifenoli. La quantità di solforosa combinata è molto inferiore a quella totale .La combinazione dell’anidride solforosa con l’aldeide acetica risulta stabile e definitiva ( irreversibile ) a differenza con gli altri composti formati con le

55
altre sostanze , instabili e reversibili .Secondo alcuni autori la SO2 combinata ( la forma prevalente ) presenta attività antibatterica seppure inferiore a quella libera .
1. acido piruvico , composto secondario della fermentazione , 2. acido aaaa----chetoglutarico, deriva dal metabolismo proteico, 3. aldeide acetica o etanale ,anch’essa prodotto secondario della fermentazione quindi
sempre presente nel vino (se il vino ne contiene più di 10 mg/l saprà di vecchio e verrà detto “vino ossidato”), un vino giovane non deve contenere molta acetaldeide, mentre nei vini invecchiati ne troviamo oltre i 200 mg/l (moscati, cherry). Bisogna anche tenere presente che 70 mg di acetaldeide possono combinare fino a 100 mg di solforosa e che 140 mg/l di acetaldeide combinano la quantità massima di solforosa consentita per legge . Come si è visto la solforosa combinata con CHO è una molecola stabile e irreversibile (costante di dissociazione bassa ) mentre quella combinata all’acido piruvico si può dissociare nuovamente (costante di dissociazione più alta). Proprio da questa propensione a formare composti stabili è importante scegliere il momento adatto di somministrazione per evitare di farla combinare.
La SO2 svolge principalmente azione antisettica selettiva nei confronti dei microrganismi presenti nel mosto ; se usata a basse concentrazioni ( 5-30g/hl) uccide numerosi microrganismi indesiderati senza danneggiare i saccaromiceti . Dosi superiori (130-180g/hl ) rendono invece il mosto infermentescibile (mosto muto). L’efficacia dell’SO2 dipende, oltre che dalla sua concentrazione e dal tipo di microrganismo (i lieviti sono più resistenti dei batteri e tra questi ultimi i bacilli risultano meno sensibili dei cocchi ) , anche dalla loro quantità e dalla fase in cui si trovano ( più i microrganismi sono inerti e più efficace sarà l’azione dell’SO2 ). Anche la temperatura ambiente , l’acidità e la composizione del mosto possono influenzare l’attività dell’antisettico .

56
La SO2 ha numerosi altri effetti:
1. azione solubilizzante nei confronti delle sostanze coloranti e dei polifenoli in genere . L’aumento dell’acidità del mosto e del vino determina una maggiore solubilità dei pigmenti e accresce l’intensità e la vivacità del colore (azione chiarificante con la precipitazione di mucillagine). I vini rossi solfitati hanno quindi un colore più marcato e brillante , dosi eccessive hanno invece azione decolorante .
2. azione acidificante diretta dovuta alla caratteristica acidità delle soluzioni di SO2 e indiretta in quanto , inibendo i batteri lattici , riduce la fermentazione malolattica ( l’acido lattico è meno forte del malico ) .
3. azione defecante l’aumento dell’acidità facilita la flocculazione dei colloidi caricati negativamente .
4. azione antiossidante per le proprietà riducenti della solforosa ( che tende facilmente a ossidarsi a solfato ) , soprattutto nei confronti dei perossidi formatisi dai polifenoli . Inoltre distrugge e inattiva le ossidasi responsabili di un intorbidamento del vino detto “casse ossidasica” (lasciato all’aria il vino inscurisce rapidamente ,con l’aggiunta di SO2 si previene questo difetto. Minore è invece l’effetto nei confronti dei metalli presenti ( Fe,Cu).
5. regola l’andamento della fermentazione può infatti avere un effetto batteriostatico o fungistatico, aumentandone la concentrazione diventa battericida e fungicida , così da inibire lo sviluppo degli apiculati favorendo quello degli ellittici (più è piccolo il microrganismo e maggiore è la superficie di contatto; nel caso dei lieviti ellittici hanno superficie minore quindi tollerano meglio la solforosa ). Ad alte temperature ,inoltre, si fermenta in modo troppo vigoroso e l’aggiunta della solforosa rallenta la fermentazione senza dover raffreddare.
6. fermentazioni più rapide 7. stabilità del colore, poiché la solforosa è riducente quindi capta l’ossigeno prima delle
sostanze fenoliche 8. maggiore stabilità del vino all’aria
Svantaggi della SO2
1. odore e sapore di idrogeno solforato (odore di uova marce) facile da eliminare con un travaso immediato altrimenti si arriva ai mercaptani (sostanza presente nelle fialette puzzolenti), sostanze stabili e difficili da eliminare.
2. difficoltà di invecchiamento 3. azione farmacologica dell’HSO3
- che agisce sui mitocondri.
La SO2 può essere usata in vari modi :
1. allo stato gassoso, 2. allo stato liquido( in bombole; metodo più usato nelle grosse cantine e anche la più
controllabile per la facilità di dosaggio nella somministrazione), 3. allo stato solido ossia come sali ( solfito di sodio, bisolfito i sodio , metabisolfito di sodio e
potassio ) che rappresenta la forma preferita dai piccoli produttori. Sotto questa forma la si usa anche in polvere ( poiché ha una resa del 58% in anidride bisogna raddoppiare circa la quantità da somministrare ).
In enologia la solforosa può essere impiegata dalla conservazione dei vasi vinari , al trattamento delle uve , dalla solfitazione del mosto a quella del vino .Lo stadio miglior rimare comunque l’ammostatura o quello successivo, prima cioè che inizi il processo fermentativo. In linea

57
generale la SO2 viene impiegata a dosi più elevate quando le uve sono alterate , la concentrazione zuccherina alta , il pH basso , la vinificazione in bianco: in quest’ultimo caso infatti la SO2 ostacola anche la fermentazione malo-lattica (la permanenza dell’acido malico mantiene al vino il gusto acidulo) e l’imbrunimento . Per l’Italia i limiti legali di SO2 sono di 160 mg/l per i vini rossi e di 200 mg/l per i bianchi e i rosati. Prima della direttiva europea il limite massimo era fissato a 200 mg/l equivalente a 200 ppm = 20 g/hl per ogni tipo di vino.. L’OMS (organizzazione mondiale della sanità) ha stabilito che la dose massima che un uomo può sopportare è di 0,7 g/kg di peso corporeo .E’ ammesso anche l’acido ascorbico (150 mg/l ) per l’azione antiossidante più immediata a carico dei metalli ( Fe, Cu ) e per creare un ambiente riducente , che favorisce la formazione dell’aroma , meglio se usato prima dell’imbottigliamento e sempre abbinato all’SO2 . Nonostante i numerosi tentativi effettuati sia in campo fisico (radiazioni UV , ultrasuoni, refrigerazione e pastorizzazione) e chimico ( idrogeno solfato , pirocarbonato di etile ecc..) non sono state trovate alternative valide all’SO2 che resta tuttora il miglior additivo in enologia. QUANTITA’ DI SOLFOROSA DA SOMMINISTRARE Uve sane 5-10 g/hl Molto zuccherine 10-15 g/hl Uve alterate 20-30 g/hl Uve troppo calde 15-20 g/hl Uve fredde 3-5 g/hl Uve poco acide 10-15 g/hl Uve acide Uve per vini colorati 20-40 g/hl che favorisce l’estrazione del colore dalle bucce TEMPI DI SOMMINISTRAZIONE
1. direttamente sull’uva 2. subito nel mosto in relazione ai dati precedentemente visti riguardanti la quantità
(allontanando il rischio di legame con l’acetaldeide e l’acido piruvico, più probabile a metà fermentazione), oltre la quale si renderebbe meno efficace l’SO2.
3. aggiunta di SO2 alla fine della fermentazione o prima dell’imbottigliamento, in relazione al destino del vino (solo nei vini da invecchiare, mentre per i novelli non si aggiunge).

58
QUANTITA’ TOTALI DI SOLFOROSA NEI VINI Vini rossi questi vini contengono tannini che son già di per sé antisettici (venivano utilizzati anche nella concia delle pelli ) quindi se ne usano quantità inferiori vini rossi dolci (225 mg/l) Vini bianchi secchi (225 mg/l) Vini rossi secchi hanno pochi zuccheri quindi anche in questo caso se ne usa meno (175 mg/l)
Vini dolci bianchi si usa più solforosa (275 mg/l)
LA VINIFICAZIONE
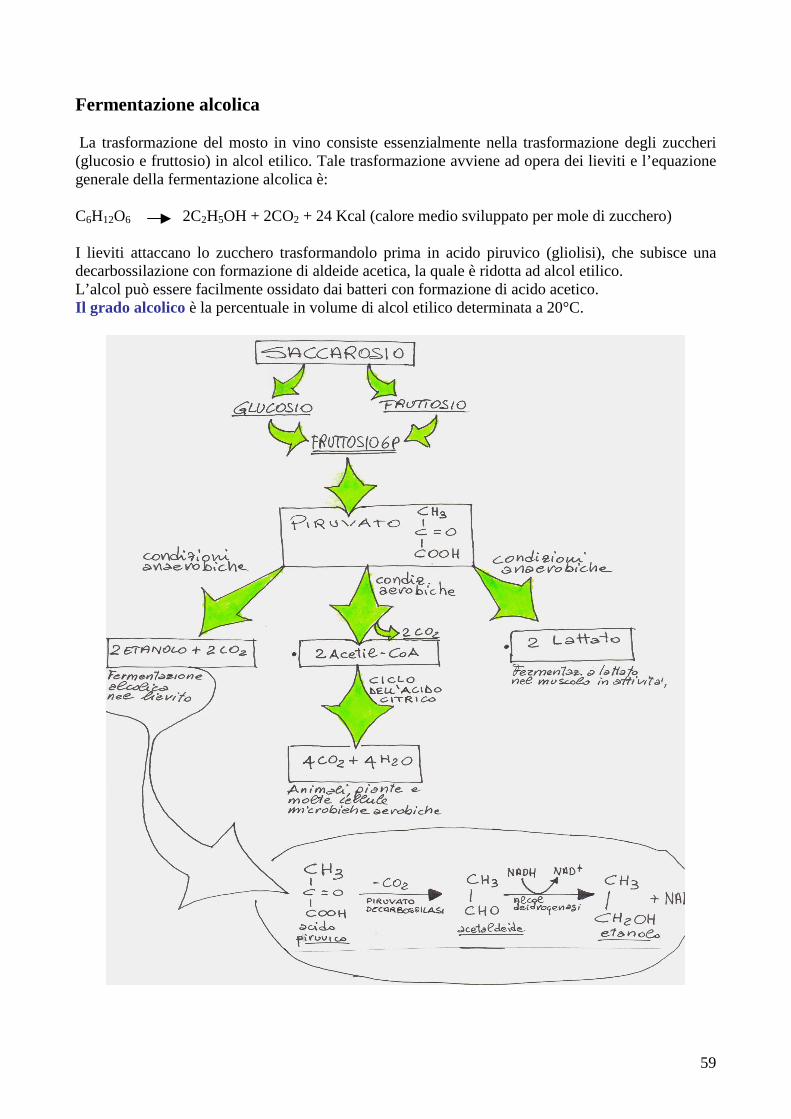
59
Fermentazione alcolica La trasformazione del mosto in vino consiste essenzialmente nella trasformazione degli zuccheri (glucosio e fruttosio) in alcol etilico. Tale trasformazione avviene ad opera dei lieviti e l’equazione generale della fermentazione alcolica è: C6H12O6 2C2H5OH + 2CO2 + 24 Kcal (calore medio sviluppato per mole di zucchero) I lieviti attaccano lo zucchero trasformandolo prima in acido piruvico (gliolisi), che subisce una decarbossilazione con formazione di aldeide acetica, la quale è ridotta ad alcol etilico. L’alcol può essere facilmente ossidato dai batteri con formazione di acido acetico. Il grado alcolico è la percentuale in volume di alcol etilico determinata a 20°C.

60
Fermentazione malolattica ( o fermentazione secondaria) Un batterio lattico è un microrganismo che a partire da qualunque sostanza produce acido lattico. L’acido malico e quello tartarico sono presenti nel vino e contribuiscono all’acidità fissa, mentre l’acidità volatile è data dall’acido acetico. L’acidità fissa non deve superare certi valori: vini rossi 4o/oo ; vini bianchi 3,4 o/oo
In alcuni casi (in climi freddi o se si anticipa la vendemmia) è possibile riscontrare un eccesso di acido malico nel mosto. La diminuzione di acido malico si può ottenere tramite i batteri sferici (cocchi). I fermenti lattici sono bacilli (non ci interessano) o cocchi, e tra quest’ultimi abbiamo gli Streptococchi, con il gen. Streptococcus (sia omofermentanti che eterofermentanti), il gen.Pediococcus (omofermentanti) ed il gen. Leuconostoc (eterofermentanti). La malolattica è una fermentazione batterica, a carico dell’acido malico, i cui agenti sono i batteri lattici che sopportano abbastanza bene bassi valori di pH, contrariamente ai batteri che provocano malattie. Possiamo distinguere i batteri in:
• batteri omofermentanti appartenenti ai generi Lactobacillus e Pediococcus (batteri streptococchi: cocchi in catenelle); producono quasi esclusivamente acido lattico ed operano a pH > 3.4
• batteri eterofermentanti appartenenti al genere Leuconostoc (per la malolattica ci interessa molto la specie Leuconostoc oenos, che si sviluppa nel vino a pH inferiore o uguale a 3.2) che oggi è stato diviso in ulteriori due generi: Leuconostoc (batteri solo del latte) e Oenococcus (batteri del vino). Questi batteri, oltre all’acido lattico, che è il loro prodotto prevalente, producono anche acido acetico e succinico.
Il principale risultato della fermentazione malolattica è la diminuzione dell’acidità fissa di 0,1-0,2 unità di pH; infatti, l’acido lattico è più debole dell’acido malico, sono demoliti anche i malati di Ca e di K, per cui questi metalli si rendono liberi e salificano l’acido tartarico che, in parte, precipiterà; la diminuzione di acidità comporta una maggiore insolubilità dei sali dell’acido tartarico, che precipitano ulteriormente e provocano incrostazioni nelle botti. Si constata inoltre un aumento di acidità volatile dovuto al metabolismo dei batteri eterofermentanti che producono anche acido acetico demolendo acido citrico. Esigenze dei batteri responsabili della fermentazione malolattica : usano solo aminoacidi come fonte di N mentre i lieviti usano anche Sali di ammonio (sostanze inorganiche). Per il C usano anche l’acido malico oltre agli zuccheri. Come fattori di accrescimento usano vitamine (gli omofermentanti richiedono la Tiamina mentre non è richiesta dagli eterofermentanti). Condizioni che favoriscono lo sviluppo della fermentazione malolattica :
• T abbastanza alte 25-45°C (batteri mesofili) • pH circa 3 (tendenzialmente i batteri vivono a pH 5-7) • O2 anaerobi o anaerobi facoltativi • grado alcolico >10° • resistenza alla SO2 sono meno suscettibili alla SO2 quando il pH=3
I batteri lattici sono importanti nella trasformazione della pasta in pane, del latte in formaggio, nelle olive e negli insaccati, ma sono anche agenti di malattie nel vino e sono agenti della fermentazione malolattica nel vino, usando come fonte di energia l’acido malico e utilizzando gli aminoacidi

61
provenienti dalla lisi dei lieviti (perciò i batteri lattici si sviluppano bene alla fine della fermentazione alcolica). Per favorire la fermentazione malolattica occorre ritardare i travasi, in modo che i batteri possano trarre gli alimenti dalla feccia , specialmente gli aminoacidi liberati dai lieviti per autolisi; invece se si vuole prevenire la fermentazione malolattica è utile anticipare il travaso subito dopo la fermentazione. Quando la fermentazione malolattica è completata, conviene effettuare una solfitazione (poiché il vino, ormai meno acido, risulta più soggetto all’azione dei microrganismi) e un travaso (per separare il nuovo deposito). Questa fermentazione è desiderata oltre che per i grandi vini rossi ,che si vogliano poco acidi e morbidi, anche nei climi freddi, dove si ha abbondanza di acido malico. Meccanismo della fermentazione malolattica 1g di acido malico 0,77g di acido lattico e 0,33g di CO2
L’enzima più importante è l’enzima malico che è un complesso di enzimi NADP+-dipendenti necessita di cationi bivalenti come Mg2+ , pH=7, temperature intorno a 35°C e tollera condizioni alcoliche elevate. L’enzima non è costitutivo ma adattativo (si forma in presenza di acido malico). Prodotti secondari liberati dai batteri: - acido acetico (aumenta dopo la fermentazione) - diacetile e acetoino (composti aromatici) - glicerina (è dolce)

62
Effetti della fermentazione malolattica:
• diminuzione dell’acidità totale • aumento del pH • produzione di CO2 • torbidità • il gusto diventa più gentile (diminuisce l’asprezza) • liberazione di prodotti secondari (acido acetico, di acetile e acetoino) • miglioramento o peggioramento della qualità
Un vantaggio della fermentazione malolattica è che quando si ha un eccesso di acido malico posso eliminarlo. Posso favorire tale fermentazione posticipando il travaso (i batteri lattici hanno così sostanze nutritive a disposizione), innalzando la T e dando poca SO2; una presenza eccessiva di tannini ostacola la fermentazione malolattica. La fermentazione malolattica è utile nei vini rossi (soprattutto nelle zone fredde) perché rende il vino più maturo e armonico (l’acido malico ha un gusto aspro), mentre va inibita nei vini bianchi, dove bisogna sempre cercare di mantenere una certa acidità per assicurare la stabilità delle caratteristiche organolettiche relative al colore e alla fragranza di sapore. Quindi nei vini bianchi si opta per dei travasi veloci, basse T, maggiore dose di SO2, presenza di lieviti e filtrazione con pori minori di 0,2 mm. Se si vuole ottenere la fermentazione malolattica si può inoculare il vino con del vino preso in fase di fermentazione malolattica, oppure oggi si può indurre anche con batteri lattici selezionati che vivono bene a pH 3.2 e con T basse, resistenti all’alcol, alla SO2 e all’antagonismo con alcuni lieviti. Il momento migliore per fare avvenire la fermentazione malolattica è in funzione del vino che si vuole ottenere; alcuni vini non sono commerciabili se non avviene tale fermentazione. I vini novelli non subiscono la malolattica perché devono esprimere il profumo dell’uva MALATTIE DEI VINI I difetti dei vini sono alterazioni di carattere chimico-fisico che interessano le caratteristiche organolettiche, in particolare l’odore e il sapore. Le malattie dei vini sono alterazioni di natura microbiologica; per evitare le malattie è bene effettuare una attenta pulizia dei locali e degli attrezzi, inoltre bisogna tenere il vino in un ambiente fresco, non al contatto con l’aria. Le malattie possono essere causate da lieviti (fioretta spunto) oppure da batteri (tutte le altre malattie).I vini più soggetti a malattie sono quelli poveri in alcol, in acidità fissa e in tannino e ricchi in N e zucchero. Le malattie possono essere distinte in funzione dell’O2 in:
- aerobiche: dovute a microrganismi di superficie a contatto con l’aria (fioretta, spunto-acescenza)
- anaerobiche: dovute a microrganismi presenti nella massa interna del vino (filante, agrodolce, girato e amaro)
Le malattie possono essere distinte in funzione del composto attaccato: zucchero agrodolce e filante acido tartarico girato glicerina amaro alcol etilico fioretta e spunto-acescenza

63
•••• Fioretta Causata da lieviti aerobi o in fase aerobica, del genere Candida (non sporigeno), Pichia (sporigeno) e Hansemula (sporigeno). Il fattore predisponente la malattia è l’O2.
Se lascio la botte scolma, i lieviti agenti della malattia attaccano l’alcol etilico (producendo H2O e CO2) e formano in superficie una massa bianca a forma di fiorellini. Il colore del vino assume riflessi giallognoli. Per prevenire la fioretta, che rende il vino non commerciabile, bisogna usare il tappo colmatore ed evitare il contatto con l’aria. Dopo la fioretta, di solito, avviene l’acescenza tramite l’Acetobacter e il Gluconobacter che utilizzano l’alcol etilico e producono acido acetico.
•••• Spunto-acescenza Causata da batteri aerobi (Acetobacter e Gluconobacter) che attaccano l’alcol etilico producendo acido acetico; nel vino si osserva un velo superficiale iridescente e viscido, inoltre si ha uno sgradevole odore di aceto. Per prevenire tale malattia bisogna evitare il contatto con l’aria e utilizzare il tappo colmatore.
•••• Agrodolce o fermentazione lattico-mannitica Non colpisce i vini già ottenuti ma si può verificare durante la fermentazione o in un vino rimasto dolce. Tale malattia è causata da batteri lattici (anaerobi) ed è causata da un’alta temperatura di fermentazione (che causa una limitata attività dei lieviti, favorendo i batteri lattici termofili) o dalla rifermentazione con temperature maggiori di 37oC ; quindi la temperatura è il fattore predisponente ed è bene mantenerla bassa per evitare tale malattia. I composti attaccati dalla malattia sono gli zuccheri e si formano mannite ( è un polialcol dolce che deriva dal fruttosio), acido acetico, acido lattico (dal glucosio) e CO2 . Agro(acido acetico)-dolce(mannite).
•••• Girato o sobbollimento o cerchione Colpisce i vini rossi, viene attaccato dai batteri anaerobi l’acido tartarico e si forma acido acetico, acido propionico, acido lattico e CO2 . Nel vino rosso si nota uno scolorimento mentre nel vino bianco un incupimento; sulla superficie si forma un velo iridescente che forma un cerchione e si formano all’interno del vino nubecole sericee che precipitano sul fondo trasportando le sostanze coloranti, quindi il liquido diventa opalescente. I vini molto colpiti da tale malattia sono quelli poco alcolici e poco acidi, quindi l’alcol è il fattore predisponente. Per prevenire effettuare travasi tempestivi.
•••• Amaro I batteri anaerobi attaccano la glicerina trasformandola in aldeide acrilica (responsabile del sapore amaro della frittura) che si condensa con le sostanze coloranti del vino e precipita. Quindi si ha uno scolorimento, gusto amaro, odore di burro e putrido. Il fattore predisponente è la glicerina e i vini deboli (poco alcolici 10-11% e pH alto > 3,6). Per prevenire bisogna tenere sotto controllo il vino con la solforosa ed effettuare travasi tempestivi.
•••• Filante Tale malattia colpisce per lo più i vini bianchi e si manifesta con una viscosità accentuata e il vino quando viene versato “fila” come l’olio. Inoltre si può avere un retrogusto acre e rancido e un intorbidamento. Gli agenti sono i batteri lattici in catenelle che hanno la capsula (catenelle + mucosa = viscosità). Tale malattia si verifica in vini in cui si ha presenza di zuccheri residui e molto N disponibile (i batteri lo utilizzano per formare la capsula); il filante colpisce i vini della pianura (ricchi di sostanza organica e quindi di N), mentre i vini ottenuti da terreni in collina ( più poveri di

64
sostanza organica) sono migliori, perché la produzione di sostanze aromatiche è una risposta allo stress. Il filante si cura rompendo le catenelle mediante sbattimento e filtrazione.
MALATTIE DEI VINI (Schema generale)
DIFETTI MALATTIE ( di carattere chimico-fisico come la rottura di colore ) (alterazioni microbiologiche) DEI PRODOTTI ATTACCATI DA LIEVITI DA BATTERI da (fioretta ,spunto) (tutte le altre ) microrganismi aerobi e anaerobi zucchero alcol ac. tartarico glicerina fioretta acescenza tutte le altre agrodolce filante fioretta acescenza girato amaro

65
MICROBIOLOGIA DEL TERRENO
Ciclo dell’azoto L’azoto entra a far parte degli aminoacidi e degli acidi nucleici. Generalmente le piante assumono l’N dal suolo o dall’acqua sotto forma ionica, mentre quello atmosferico è sotto forma biatomica (N2) in cui i due atomi di azoto sono tenuti insieme da un triplo legame che richiede molta energia per essere scisso (molto stabile), per cui le piante non sono in grado di assorbire l’azoto direttamente dall’atmosfera ma serve un processo che trasformi l’azoto biatomico in azoto ionico * detto biofissazione (il ciclo dell’azoto è una continua serie di processi naturali con i quali l’azoto passa dall’aria al suolo, ai vegetali e in ultimo agli animali e quindi torna all’aria o al suolo attraverso i processi di demolizione della sostanza organica e/o della denitrificazione).
• Nitrificazione : i batteri nitrificanti possono essere compresi in due gruppi:
1. quelli appartenenti al genere Nitrosomonas che ossidano l’ammoniaca a nitrito (NO2
-) secondo la reazione NH4++1/2 O2 H2O+NO2
- + 2H3O+ con
liberazione di 273,9 kj mol-1 2. quelli appartenenti al genere Nitrobacter che ossidano nitrito a nitrato secondo la
reazione : NO2-+1/2 O2 NO3
- con liberazione di 76,7 kj mol-1 • Denitrificazione : esistono ben determinate specie batteriche che possono operare un
processo di respirazione anaerobica nel quale invece dell’ossigeno utilizzano il nitrato come accettare finale di elettroni. Tale processo viene definito come riduzione disassimilativa del nitrato durante la quale i batteri riducono il nitrato a nitrito ma non oltre. Altri batteri invece possono ridurre il nitrato a protossidido d’azoto (ed infine ad azoto molecolare che infine si libera nell’atmosfera). La denitrificazione mette in grado i batteri denitrificanti di ottenere una completa ossidazione del substrato organico e quindi un rendimento di crescita ottimale anche in assenza di ossigeno. La denitrificazione, infatti, è un processo respiratorio a tutti gli effetti; tuttavia essendo qui il nitrato, e non l’ossigeno, l’accettore finale di elettroni, è un processo di respirazione anaerobica.

66
• Azoto fissazione L’aria contiene l’80% di azoto ed è considerata la grande “riserva” oltre che valvola di sicurezza del sistema. L’azoto dell’aria, per effetto delle scariche elettriche, può essere scisso per combinarsi immediatamente con l’ossigeno a formare ossidi di azoto che, sciogliendosi nell’acqua durante le piogge, arrivano nel suolo sotto forma di nitrati. Direttamente dall’aria l’azoto può essere fissato, con un determinato dispendio energetico, da determinati batteri e alghe azzurre; i primi lo possono fare da soli ( fissazione libera operata da batteri quali quelli appartenenti al genere Azotobacter, che vivono in suoli neutri; il genere Beijerinckia, che si adatta bene anche a suoli acidi, tant’è che si è isolato in foreste equatoriali insieme al genere Derxia; il genere Pseudomonas, tutti aerobici, e il genere Clostridium , anaerobici),vivendo nel suolo da soli o in simbiosi come nel caso del genere Rhizobium che vive nelle radici delle leguminose formando dei noduli radicali. Relativamente di recente si è scoperto che anche dei funghi Attinomiceti sono in grado di formare noduli radicali negli ontani (genere Alnus) e di fissare azoto atmosferico ( finora sono state individuate circa 160 specie di dicotiledoni appartenenti a vari generi con noduli di Attinomiceti e la maggior parte di esse vivono in terreni poveri dove l’azoto inorganico disponibile è molto scarso. Queste alghe dispongono di cellule particolari dove avviene il processo di fissazione chiamate eterocisti . Dette cellule hanno bassi contenuti di ossigeno per favorire l’azione di un enzima particolare, la nitrogenasi, che catalizza la reazione.
Questo enzima per funzionare correttamente necessita che sussista:
1. la presenza di tracce di Fe e Mo per la sintesi dell’enzima 2. presenza di processi metabolici che liberano elettroni per la riduzione dell’azoto e di
ATP per il fabbisogno energetico 3. presenza di ioni magnesio 4. temperature compatibili con la crescita di batteri mesofili 5. assenza di ossigeno,che inattiva irreversibilmente l’enzima
La fissazione biologica è alquanto costosa in termini energetici; bisogna infatti rompere il triplo legame della molecola biatomica dell’azoto per formare, con l’aggiunta dell’idrogeno dell’acqua, due molecole di ammoniaca (NH3) . Le leguminose nei noduli radicali devono fornire un minimo di 10g di glucosio (pari a circa 40 Kcal) perché i batteri o gli Attinomiceti fissino 1g di azoto con una resa del 10% . A questo proposito gli organismi liberi hanno rese inferiori necessitando talvolta di 100g di glucosio per 1g di azoto con una efficienza dell’1%. Piante e Cianobatteri fissano il 60% dell’azoto totale fissato per anno sulla terra ; il 15% deriva dai lampi e dalle radiazioni UV ed il rimanente 25% deriva dai processi industriali. In sintesi il diverso stato di ossidazione che l’azoto può assumere, e quindi il suo diverso ruolo per i microrganismi, fa si che continuamente esso possa essere trasformato da una forma ad un’altra mediante reazioni catalizzate che, nel loro insieme, formano un processo ciclico :
• la riduzione assimilativa del nitrato operata dagli organismi azoto autotrofi trasforma (riduce) nitrato(+5) in ammoniaca (+3); così come la fissazione biologica dell’azoto operata dai procarioti trasforma (riduce) l’azoto molecolare in ammoniaca;
• l’ammoniaca così prodotta , o assorbita dall’esterno come tale, viene incorporata senza cambiamento dello stato di ossidazione , nei composti organici per la sintesi di materiale cellulare azotato;
• la degradazione della materia organica azotata ad opera dei decompositori trasforma l’azoto organico in ammoniaca
• la nitrificazione operata dai batteri nitrificanti trasforma ( ossida ) l’ammoniaca (-3) in nitrito (+3) , ed il nitrito in nitrato (+5);
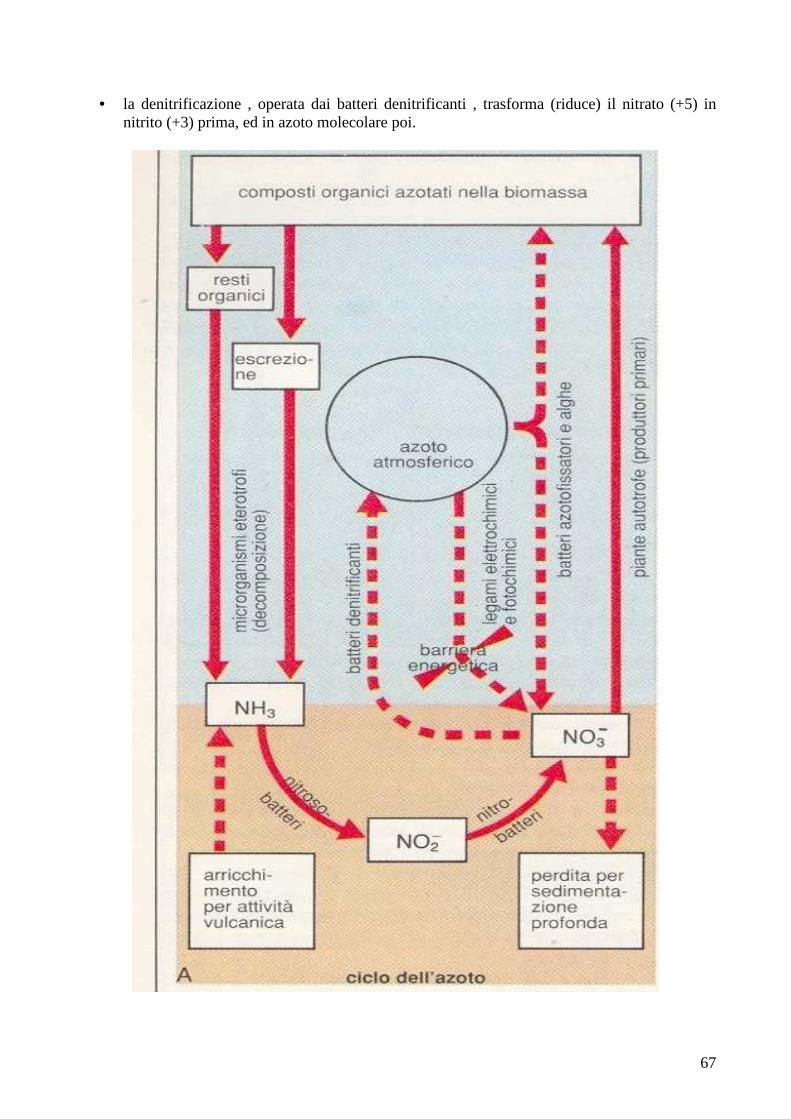
67
• la denitrificazione , operata dai batteri denitrificanti , trasforma (riduce) il nitrato (+5) in nitrito (+3) prima, ed in azoto molecolare poi.

68
Ciclo del carbonio
Più semplice, rispetto a quello dell’azoto, perché alla fine del ciclo tutto si trasforma in CO2 e H2O. La cellulosa e l’amido sono le sostanze prodotte in maggiore quantità durante l’anno. La prima è un polimero del glucosio formato da molecole legate da ponti b: i cellulosolitici degradano le lunghe molecole di cellulosa interessando dapprima i legami idrogeno che legano le catene laterali per poi spezzare le catene di glucosio fino all’idrolisi completa (glucosio singolo). Tra questi microrganismi ne possiamo elencare di obbligati aerobi: Ordine MYXOBATTERIALES genere Citophaga genere Sporocitofaga genere Solangium e microrganismi non obbligati: Ordine PSEUDOMONALES genere Pseudomonas Ordine genere Celvibio genere Celfacicula Tra gli anaerobi troviamo anche microrganismi del genere Clostridium . La cellulosa ha un ruolo importante anche nell’azoto fissazione (è infatti un consiglio quello di sotterrare sempre le stoppie piuttosto che bruciarle); la cellulosa,infatti ,va a costituire l’HUMUS prima di essere degradata, questo ha grosse capacità di trattenere acqua e sostanze nutritive, tutte prerogative di un buon terreno e che è impossibile avere in mancanza di sostanza organica.
La quantità di carbonio fissata sulla terra attraverso la fotosintesi è impressionante: le stime variano da circa 70 a 120 miliardi di tonnellate l’anno; attualmente l’atmosfera ne contiene soltanto circa un

69
millesimo dell’1% del totale; ogni anno più del 13% del carbonio contenuto nell’atmosfera è utilizzato nella fotosintesi e circa la stessa quantità è scambiata con la CO2 disciolta negli oceani. La quantità di CO2 presente nell’aria è rimasta abbastanza stabile a circa 280 ppm per la maggior parte degli ultimi mille anni, a partire dal 1850 circa la quantità di CO2 nell’atmosfera è aumentata esponenzialmente fino a circa 352 ppm nel 1990. La ragione principale di questo aumento è attribuito alla combustione dei prodotti fossili (si consideri che quando si brucia 1 litro di petrolio è come se si bruciassero 23,5 tonnellate di sostanza organica; all’aumento avrebbe però anche contribuito il disboscamento ed in particolare gli incendi delle foreste tropicali. Gli ecosistemi stabili, come le foreste pluviali tropicali, introducono nell’atmosfera, attraverso la respirazione e la decomposizione, una quantità di CO2 quasi uguale a quella che sottraggono quando sono disboscati ed incendiati trasferendo dalla biosfera all’atmosfera il carbonio accumulato nella loro biomassa. In questo discorso è da considerare una grossa aggravante l’ effetto serra che ha effetto a livello globale sulla biosfera; viene infatti postulato che, per effetto dell’emissione della CO2 nell’aria, si verifichi una diminuzione dell’entità dell’emissione calorica della terra verso lo spazio e di conseguenza la temperatura terrestre aumenterebbe.

70
Ciclo dello zolfo
Lo zolfo è un costituente dei due aminoacidi solforati cisteina e metionina e di numerosi altri composti cellulari quali coenzimi e prodotti secondari delle piante. La maggior parte dello zolfo funzionale è presente nelle cellule in forma ridotta come gruppo solfuro (S2-) e disolfuro (-S-S-). Lo zolfo è coinvolto poi in numerosi enzimi e metallo proteine (come le ferro-solfo proteine e le rame-proteine). La carenza di zolfo è un evento raro in natura, dato che la maggior parte dei suoli sono ricchi di solfato (SO4) che rappresenta la forma chimica sotto cui lo zolfo viene assorbito dalle piante. Comunque la mancanza di zolfo impedisce la formazione degli aminoacidi solforati provocando un arresto della sintesi proteica. Anche per lo zolfo esiste un ciclo in natura per descrivere il quale si possono fare le stesse premesse fatte per il ciclo dell’azoto: lo zolfo può esistere in numerosi stati di ossidazione che vanno da -2, come nel solfuro S2-, a +6 come nel solfato SO4
-, ognuna delle quali può essere utilizzata per scopi fisiologici a volte diversi dagli organismi viventi; infatti:
• il solfato rappresenta la principale fonte di zolfo per le piante, batteri e funghi che lo riducono a solfuro incamerandolo nella materia organica come gruppo sulfidrilico;
• mediante un processo di degradazione anaerobica e di mineralizzazione della materia organica, definita come desulfuricazione, i gruppi mercapto dei composti organici solforati, tipo metionina e cisteina, vengono spezzati dalle sulfurasi con formazione di idrogeno solforato;
• il solfato può essere inoltre utilizzato dai batteri solfato riduttori quali Desulfovibrio desulfuricans che lo utilizzano come accettare ultimo di idrogeno in un processo di respirazione anaerobica riducendolo a idrogeno solforato;
• i batteri fototropici anaerobici, quali Chromatium e Chlorobium , ossidano l’idrogeno solforato prodotto nei sedimenti anossici a zolfo elementare e a solfato. Se però l’idrogeno solforato si forma in condizioni di aerobiosi, esso viene ossidato abioticamente a zolfo mediante reazione con l’ossigeno presente;
• lo zolfo verrà ossidato dai solfobatteri chemiosintetici aerobi del genere Thiobacillus a solfato.

71
MICROBIOLOGIA DEL LATTE
Il latte costituisce un ottimo terreno di crescita per i microrganismi poiché racchiude in se sostanze sia semplici che complesse e proprio in base a questa capacità di dare “vitto e alloggio” persino ad agenti patogeni sono state varate leggi atte a tutelare i consumatori di latte prevedendo tecniche di pastorizzazione in grado al contempo di eliminare gran parte di questi microrganismi e di lasciare pressoché inalterate le qualità organolettiche dell’alimento. Le sostanze che si sono trovate e studiate nel latte sono:
• Vitamine del gruppo B
• Acido lattico
• Acido citrico
• Sostanze azotate solubili quali urea e aminoacidi liberi
• Sostanze in sospensione colloidale: 1. caseina b 2. caseina g 3. caseina a 4. enzimi: o fosfatasi alcalina usata come test per l’avvenuta caseificazione del latte o proteasi o lipasi o perossidasi: utile per verificare il maltrattamento del latte; si possono infatti
trovare valori alti di perossidasi in latti contenuti nelle cisterne per il trasporto se riempite a metà, quando cioè queste tendono a scuotere il latte favorendone l’urto con le pareti del recipiente.
• Immunoglobuline
• Componenti in emulsione: come grassi a catena lunga o corta
• Leucociti o cellule somatiche; presenti in quantità cospicue quando l’animale è malato o ha turbe
• Grassi; 3% per latte di vacca, 2.8-3.5% per latte di pezzata nera
• Lattosio; quantità simili nelle tre specie
• Proteine; 3-5% per latte bovino 5-6% per latte ovino 4% per latte caprino

72
Fattori che influenzano la produzione e la composizione del latte
Fattori fisiologici o periodo di lattazione o ciclo di lattazione o produzione di colostro o latti anomali (lenti) o periodo secco
Fattori alimentari
o livello alimentare o composizione della razione: alimentazione con fieno o pascolo asciutto fa produrre latti
ricchi in proteine ma con grassi a catena corta; alimentazione con fieno di alta qualità fa produrre latti con gusto più persistente nel tempo,
o azioni specifiche
Fattori climatici o stagione o temperatura o illuminazione o condizioni metereologiche: con vento e grandine si è notato un aumento della
concentrazione lipidica nel latte Fattori genetici
o razza o soggetto o selezione o ereditarietà
Fattori zootecnici diversi
o animale o mungitura (manuale o meccanica) o stabulazione o condizioni di raccolta del latte
Fattori sanitari
o stato di salute dell’animale o stato di salute della mammella: bisogna garantire la salubrità del latte e di conseguenza
dell’animale che lo produce; lo stesso latte è definito come il prodotto di una mungitura completa di un animale in buone salute e con un buon stato di alimentazione.

73
Microrganismi del latte
Studiando il latte dal punto di vista microbiologico si è notata l’assenza di archeabatteri mentre è molto facile la proliferazione di certi generi di lieviti e muffe (eubatteri). Tra questi quelli molto studiati e conosciuti appartengono ai generi Lactobacillus, Streptococcus, Staphilococchi , Enterobatteri ecc. Le principali vie di contatto dei microrganismi con il latte è
1. la contaminazione mammaria: endogena: direttamente tramite il sangue, l’animale malato trasmette così il patogeno direttamente nel latte che sta producendo. Tra questo tipo di microrganismi troviamo l’agente della difterite, di cui l’animale è portatore sano;l’agente della brucellosi che per via ematica si trasmette al latte (genere Brucella abartus nei bovini, Brucella helitensis negli ovini, Brucella suis nei caprini);l’agente della tubercolosi (Microbacterium tubercolosis bovis);l’agente della gastroenterite (enterobatteri quali Salmonella, prolens, E.coli ); certi stafilococchi (come Staphilococcus aurens che convive col mungitore che fa da vettore di trasmissione attraverso la mungitura manuale trasferendo il patogeno all’animale); l’agente del carbonchio (Bacillus anthracis); l’agente del vaiolo e del tifo; le mastiti (in genere più latte un animale produce e più è suscettibile all’insorgenza di mastiti; inoltre anche errori nella mungitura meccanica ,come quello di adoperare mungitrici per bovini su ovini, possono creare disfunzioni nel tessuto mammario dovute ad esempio da Staphilococcus aurens che si manifesta con l’insorgenza di pustole localizzate anche esteramente); esogena: Staphilococchi e Enterobatteri penetrano nella mammella in modo ascendente partendo dalla cute e risalendo attraverso l’orifizio del capezzolo proliferano come colonie di batteri saprofiti patogeni.
2. contaminazione esterna alla mammella : Clostridi butirrici (conferiscono il tipico gonfiore ai formaggi) trasmessi attraverso gli insilati, i
pellettati ne contengono un quantitativo notevole, per questo è necessario migliorare la qualità sanitaria degli insilati in genere; si pensi che per la produzione del Parmigiano reggiano le vacche non vengono alimentate con insilati per evitare in alcun modo il gonfiore tardivo.
Qualità batteriologica del latte
1. allevamento: attenzioni particolari direttamente in sala di mungitura; 2. l’animale: il suo stato di salute, di nutrizione e d’igiene; 3. mungitura: lavaggio della mammella, eliminazione dei primi getti di latte, cura dell’igiene
del mungitore, pulizia dei materiali impiegati nella mungitura; secondo la legge n°46 del 1992 si obbliga la presenza della sala di mungitura per chi vuole commercializzare latte (naturalmente se lo produce anche), inoltre obbliga la presenza in essa di un lavandino che eroghi acqua potabile;
4. raccolta e del latte: filtrazione, pulizia dei contenitori e raffreddamento; 5. trasporto del latte: raccolta, raffreddamento e durata del viaggio;

74
Igiene dell’animale
I fattori che influenzano la contaminazione microbica del latte legati alla mungitura sono:
1. condizioni igieniche della sala di mungitura: la mungitura meccanica migliora la qualità del latte solo se gestita bene altrimenti si rischia un netto peggioramento qualitativo (come nel caso in cui si usino acque non potabili per il lavaggio della macchina di mungitura o una non perfetta pulizia della stessa con presenza di residui e incrostazioni );
2. condizioni igieniche dell’animale:pulito se latte con carica batterica = 8 000 germi/ml, non pulito se latte con carica batterica = 17 000 germi/ml
3. condizioni igieniche della mammella: parametro molto importante; lavata se latte con carica batterica = 40 000 germi/ml; non lavata se latte con carica batterica = 1 900 000 germi/ml (lavando la mammella prima dell’operazione di mungitura si può ridurre la carica batterica nel latte fino al 97,9%);
4. igiene mani del mungitore: in una mano non perfettamente pulita si possono trovare in media 45 000 000 di germi;
5. influenza tecnica di mungitura: nei primi getti si possono trovare fino a 2 000 000 germi/ml di latte; tolti i primi getti si riduce a 100 000 germi/ml, con una riduzione del 62% della carica microbica; questo se accompagnato alla pulizia della mammella si può tradurre in un latte la cui carica non supera i 10 000 germi/ml; oltretutto con l’eliminazione dei primi getti è anche possibile individuare la presenza di mastiti (grumi di caseina se mastiti subcriniche, oppure grumi di sangue se mastiti croniche);
6. recipienti sporchi: si rischia un incremento da 200 000 a 1 000 000 germi/ml;
Classificazione dei microrganismi del latte
Lactobacillus Streptococcus Leuconostoc Pediococcus Propioniobacterium Micrococus Brevibacterium Staphylococcus Bacillus (anthracis)-----------------------viene inattivato a 55°C, a 110°C si inattivano le sue spore Clostridium Enterobacteriaceae Pseudomonas Corynebacterium Bifidobacterium Lieviti Muffe e batteriofagi

75
Caratteristiche del latte al momento della raccolta o dell’ammissione allo stabilimento Valori da attribuire al latte vaccino:
• punto crioscopico: -0.520°C • peso specifico a 20°C > 1.028g/l • contenuto proteico > 28g/l • estratto secco magro > 8.50%
produzione latte alimentare:
• carica batterica totale a 30°C < 100 000 germi per ml • contenuto in cellule somatiche sempre per ml < 400 000
produzione di prodotti a base di latte:
• carica batterica totale a 30°C < 400 000 germi per ml • contenuto di cellule somatiche per ml < 500 000 • formaggi da latte crudo CBT < 400 000
I microrganismi presenti nel latte sono divisibili in due gruppi: microrganismi saprofiti non patogeni (Lattobacilli, streptococchi, Escherichia coli, Pseudomonas, Clostridi) e microrganismi patogeni (agenti del tifo, della difterite, del colera, della brucellosi). Il latte contiene persino sostanze ad attività antibatterica, quali lattenine, ma il loro potere battericida si esaurisce nel giro di poche ore lasciando così spazio alla proliferazione batterica. La contaminazione del latte può avvenire attraverso due vie: una contaminazione endogena che si ha quando un animale malato trasmette i propri germi patogeni al latte che produce ; quando l’affezione è semplicemente locale (a livello della mammella) i microrganismi possono contaminare il latte durante il suo passaggio nel canale del capezzolo . persino l’animale sano può possedere una microflora di tipo saprofitico a livello della mammella (Micrococcus e Corynebacterium); una contaminazione esogena provocata dal contatto del latte con la pelle dell’animale, i recipienti e gli strumenti di raccolta del latte (mungitura), l’ambiente, il foraggio, il personale addetto e gli strumenti usati per la refrigerazione e il trasporto. La refrigerazione provoca una selezione della popolazione microbica: a temperatura ambiente si moltiplicano più rapidamente i batteri acidificanti e i lieviti (nel giro di poche ore). A 4°C, temperatura di refrigerazione, si sviluppano microrganismi psicrofili ad attività lipolitico e proteolitica ma il loro sviluppo è evidente solo dopo 24 ore dalla mungitura. BATTERI DEL LATTE I batteri che interessano le trasformazioni lattiero-casearie sono i batteri lattici

76
I batteri possono essere Gram+ , in grado di fermentare i carboidrati ad acido lattico, non possiedono citocromi, sono anaerobi ma tollerano la presenza di ossigeno, sono immobili e asporigeni. Streptococchi e Leuconostoc hanno forma sferica (tipica dei cocchi) mentre i lattobacilli hanno forma bastoncellare. Gli Streptococchi sono omofermentanti perché la loro fermentazione è di tipo omolattica (trasformazione del lattosio in acido lattico al 90%). Tra gli Streptococchi quelli lattici sono i più comuni sono:
• Streptococcus lactis e Streptococcus cremonis: entrambi responsabili dell’acidificazione spontanea del latte
• Streptococcus diacetylactis : consente anche la fermentazione dell’acido citrico a diacetile, caratteristica sostanza aromatica del burro
• Streptococcus thermophilus : importante per la maturazione dei formaggi a pasta cotta I Lattobacilli si dividono in omofermentanti (termofili e mesofili e producono maggiori quantità di acido lattico rispetto agli Streptococchi ed hanno una lieve ma più spiccata attività proteolitica) ed eterofermentanti (responsabili della fermentazione eterolattica producendo acido lattico solo per il 50% e per il restante 50% etanolo, CO2 ed altre sostanze aromatizzanti , come l’acido acetico). L’ attività proteolitica è sfruttata nella maturazione del formaggi :
• Lactobacillus helveticus e Lactobacillus lactis : importanti per la maturazione dei formaggi a pasta cotta, sono termofili (temperature massime di sviluppo rispettivamente a 48°C e 52°C)
• Lactobacillus casei e Lactobacillus plantarum : importanti nella maturazione dei formaggi a pasta dura non cotta
• Lactobacillus bulgaricus: termofilo, la sua attività fortemente acidificante viene sfruttata nella produzione dello yogurt
• Leuconostoc citrovorum: eterofermentante responsabile della fermentazione della crema con produzione di diacetile.
Oltre a quelli lattici troviamo batteri che favoriscono la maturazione dei formaggi e, al contrario, batteri che ne possono essere causa di alterazioni:
• Escherichia coli e Coli aerogenes: batteri enterici coliformi di origine fecale che fermentano lattosio ad acido lattico, acido acetico e gas quali CO2 e idrogeno (questi ultimi sono conosciuti come la causa di una caratteristica alterazione dei formaggi molli detta “gonfiore precoce”dovuta ad una fermentazione acido mista);
• Batteri propionici : di origine fecale, fermentano latte ad acido lattico tramite fermentazione propionica con produzione di acido propionico, acido acetico, CO2; questa fermentazione gasogena è causa del “gonfiore tardivo” nei formaggi a pasta cotta , alterazione che si manifesta durante la salatura e la maturazione che provoca la formazione di un’occhiatura anomala o di grosse cavità saltuarie (nell’Emmental questa fermentazione è invece molto ricercata).
• Clostridium : batteri anaerobi presenti nei foraggi e insilati, responsabili infatti della contaminazione del latte prodotto in inverno, cioè quando il bestiame viene alimentato solo con foraggi; sono responsabili delle fermentazioni di lattosio e lattati ad acido butirrico,CO2 e H2 causando l’alterazione tipica di formaggi a pasta cotta dette “gonfiori tardivi”(fermentazione butirrica).
• Micrococcus luteus: esempio di batterio proteolitico, favorisce la maturazione dei formaggi perché attaccano la caseina, idrolizzandola, e gli zuccheri acidificando il latte con formazione di acido lattico

77
• Clostridium lentoputrescens: batterio proteolitico responsabile della produzione di odore e aroma sgradevoli
• Bacillus subtilis: batterio proteolitico aerobico, sporigeno e molto resistente alle temperature (difficile sterilizzazione del latte che li contiene).
• Pseudomonas aeruginosa e Escherichia coli: non sporigeni, si sviluppano bene alle temperature di refrigerazione, hanno la capacità di prendere il sopravvento sui batteri lattici.
• Pseudomonas fluorescens: il più comune batterio lipolitico (idrolizza trigliceridi). Lieviti e muffe appartengono al regno dei protisti superiori quindi sono eucarioti; la loro contaminazione nel latte non è quasi mai rilevante dal punto di vista quantitativo (nonostante il latte rappresenti per loro un ottimo terreno di coltura).
• Saccharomyces fragilis , Saccharomyces lactis, Morula kephyr , Candida pseudotropicalis (varietà lactosa): sono tutti lieviti capaci di dare origine a fermentazione alcolica utilizzando come substrato il lattosio, largamente utilizzati per i latti fermentati
• Lieviti a fermentazione gasogena: che inquinano con facilità formaggi molli e yogurt (gonfiore precoce, causato questo anche da coliformi).
• Oospora lactis, Cladosporium butyri, Scopulariopsis brevicaule: muffe responsabili dell’irrancidimento del burro (provengono da soffitti e pareti degli ambienti in cui si lavora il burro);
• Penicillium: si evidenzia con la formazione di tipiche chiazze verdi su burro conservato per tempi lunghi o impastato in maniera non razionale;
• Alternaria e Cladosporium: formano chiazze scure nel burro; • Oospora: forma chiazze arancioni sempre nel burro; • Penicillium roqueforti : in formaggi a muffa verde intera (come Gorgonzola e
Roquefort), muffa proteolitica che causa un rapido innalzamento del coefficiente di maturazione(CM);
• Penicilliun camemberti: formaggi a muffa bianca esterna. Processi di risanamento e conservazione del latte Il Latte per essere messo in commercio subisce dopo la raccolta un insieme di processi :
• Filtrazione: per eliminare le particelle più grossolane o centrifugazione che permette una migliore separazione dello sporco in sospensione;
• Standardizzazione del contenuto in grasso: in Italia il latte intero deve avere un contenuto minimo in grasso pari al 3.2% , quello parzialmente scremato l’1.8%, quello scremato non più dello 0.5%
• Omogeneizzazione e deodorizzazione: trattamento meccanico effettuato anche dopo il risanamento termico atta a ridurre le dimensioni dei globuli di grasso diminuendo di conseguenza la velocità con cui questi affiorano. Consiste nello spingere il latte a forti pressioni (150-200atm) contro una valvola munita di orifizio molto piccolo che conduce il latte in una camera a pressione atmosferica (1 atm ) ;

78
• Risanamento: distruggendo i microrganismi patogeni eventualmente presenti .Oggi per questa operazione si utilizza il calore capace di inattivare enzimi (come le lipasi) e di avere azione battericida;Tra queste tecniche è diffusa la pastorizzazione e la sterilizzazione:
o Pastorizzazione : condotta a temperatura inferiore a quella di ebollizione, in grado ugualmente di
distruggere tutti i microrganismi patogeni come Mycobacterium tubercolosis, Brucella abortus e le loro tossine (sopravvivono alla pastorizzazione spore e microrganismi termoresistenti). Naturalmente l’efficacia della pastorizzazione dipende anche dalla carica batterica iniziale e dalla durata del trattamento termico o meglio dal rapporto tempo/temperatura ( 25’ a 62°C, 15’’ a 72°C, 6’’ a 74°C )
o Pastorizzazione bassa o lenta: in vasche a doppia parete con agitatori, per evitare la salita della schiuma, il latte raggiunge la temperatura di 65°C per 30’.
o Pastorizzazione rapida: si porta il latte a temperature comprese tra i 72-85°C per tempi inferiori al minuto grazie ad apparecchi in cui il latte scorre per strati sottilissimi dell’ordine del millimetro (l’ideatore di questo metodo oggi molto in uso nelle centrale è stato l’italiano Stassano negli anni ’20 da cui il nome del processo di stassanizzazione). In genere un buon latte pastorizzato ha una carica di saprofiti mesofili non superiore alle 30 000 colonie/ml. Per essere poi sicuri che il latte non sia stato sottoposto a maltrattamenti termici si ricercano tracce di perossidasi che in un latte ben pastorizzato dovrebbe essere attiva.
o Sterilizzazione: oggi consiste nel metodo U.H.T. (Ultra-High Temperature), consta in trattamenti continui con capacità lavorativa di 20 000 litri/ora, prevedendo temperature di 140-150°C per pochi secondi ;
Trattamenti alternativi al riscaldamento
• Battofugazione: sottoponendo il latte a centrifugazione (8000-10 000 giri/minuto) consentendo l’eliminazione fisica del 99.9% dei microrganismi presenti;
• Attinizzazione: utilizza radiazioni U.V. per la loro azione battericida ; offre anche il vantaggio della trasformazione di composti stereoidei in tiamina (vitamina D), però porta al rancidimento della frazione lipidica ed è infine un metodo troppo costoso;
• Antisettici: la legge italiana vieta l’uso di qualunque tipo di antisettico (l’acqua ossigenata è usata in modo indiretto per sterilizzare le confezioni, nel latte però è presente una perossidasi attiva che la degrada);
• Freddo: tramite refrigerazione, mantenendo il latte a temperatura di 4°C,e congelazione utilizzando catene del freddo molto ben organizzate;
• Ultrafiltrazione: utilizza membrane filtranti con pori del diametro di qualche Angstrom (oggi si usano solo per la produzione di formaggi freschi e a pasta molle);
• Osmosi inversa: con la fuoriuscita dell’acqua dalla soluzione più concentrata (dall’alimento) Preparazione di latti fermentati Sono prodotti dal latte a cui viene aggiunte colture specifiche, ovviamente prive di germi patogeni, mantenute vive fino alla vendita del prodotto: Yogurt Latte fermentato più diffuso in Italia, si produce a partire da latte fresco dopo una drastica pastorizzazione (90°C) che, oltre ad abbassare la carica microbica, stabilizza il futuro coagulo che viene omogeneizzato. Per elevare il residuo secco all’estero si aggiunge quasi sempre latte in polvere (in Italia questa procedura è vietata); si procede poi all’insemenzamento con Lactobacillus bulgaricus e Streptococcus thermophilus e si pone nel maturatore a 45°C per 6-8 ore; dopo l’incubazione subisce una blanda omogeneizzazione (laminazione) che evita la separazione del

79
siero e migliora la sua consistenza. Il prodotto così ottenuto viene conservato a 4°C , avrà un grado di acidità pari a 40 SH, pH=4.6 , contenuto in acido lattico pari all’1%, lattosio al 3.5% , contenuto di batteri lattici intorno a 106-109/grammo. Kefir Bevanda moderatamente alcolica , alcol < 1%, e gassosa (CO2) ottenuta fermentando a 20°C latte di cammella, di pecora o di capra. Kumys Prodotto con latte di giumenta o di asina, ma anche con latte vaccino, che subiscono la fermentazione acido-alcolica. Gioddu o mizzuraddu o miciuratu Prodotto in Sardegna a partire dal latte di pecora . Oggi viene insemenzato con la stessa microflora dello Yogurt e subisce quindi una fermentazione acida.
CREMA E BURRO
CREMA Derivato del latte ottenuto tramite centrifugazione o per affioramento in modo da ottenere un prodotto arricchito in materia grassa pur contenendo gli stessi costituenti del latte (deve contenere almeno il 10% di grasso). Affioramento: sfrutta la legge di Stokes:
Il latte viene lasciato a riposare in vasche di acciaio inossidabile con il fondo concavo , alte almeno 15cm e con capacità di almeno 70 litri mantenendo una temperatura costante di 12-15°C per un tempo di 12-15 ore (tempo di affioramento). L’affioramento è un processo spontaneo dovuto alla differenza di densità tra fase grassa e fase acquosa ; la sua velocità è, secondo la legge di Stokes, direttamente proporzionale alle dimensioni del globulo di grasso ed è facilitato dalla presenza di agglutinine sulla membrana che possono agglutinare anche i globuli tra loro.

80
Centrifugazione Si ottiene una crema con titolo maggiore di grasso, è più dolce, ha lo stesso pH del latte. Risanamento Tramite pastorizzazione e uperizzazione con temperature più alte rispetto a quelle utilizzate per il latte a causa dell’effetto protettivo che il grasso ha nei confronti dei microrganismi. Il trattamento consente inoltre l’inattivazione delle lipasi . BURRO Prodotto che si ottiene esclusivamente dalla crema di latte dopo zangolatura e sufficiente separazione del latticello fino all’ottenimento di un contenuto minimo di grasso pari all’82% . Il termine “burro” da solo spetta al prodotto che si ottiene da crema di latte vaccino . Tecniche di lavorazione La crema utilizzata per la produzione di burro si può ottenere per affioramento o per scrematura, per affioramento la panna è acida e ricca di microrganismi che fermentando producono aromi diversificati conferendo caratteri organolettici apprezzabili e ricercati; panna ottenuta per centrifugazione verrà pastorizzata e insemenzata con batteri con funzioni aromatizzanti quali: Streptococcus diacetilactis, Leuconostoc citrovorum e paracitrovorum, Betacoccus citrovorum e paracitrovorum . L’acidificazione è consentita dalla fermentazione del lattosio ad acido lattico, fino ad un pH=4.7-5.0 . L’abbassamento del pH facilita l’azione dei Betacocchi .I batteri aromatizzanti fermentano il lattosio ad acido citrico consentendo la produzione di sostanze aromatiche come diacetile e aldeide acetica. La fase di maturazione interessa la formazione delle sostanze aromatiche ed avviene ad una temperatura di circa 15°C per alcune ore, consentendo anche il raffreddamento del grasso e la sua parziale cristallizzazione., ma soprattutto questa temperatura consente l’ attività fermentativa dei microrganismi, limitandone invece quella riproduttiva.

81
Difetti del burro Difetti di carattere organolettico riguardano principalmente fattori di ordine microbiologico o chimico; come l’irrancidimeto idrolitico dovuto a muffe e batteri , oppure l’irrancidimento ossidativo dovuto invece alla presenza di ossigeno catalizzato da metalli e favorito dalla luce. Come conseguenza delle ossidazioni si ha il particolare gusto metallico, di pesce o di cotto. Sapori sgradevoli potrebbero anche provenire direttamente dal latte (sapore amaro e sapore di cavolo). Sapore di formaggio può essere causato dalla degradazione della caseina della crema per la presenza di Pseudomonas putrefaciens oppure dall’azione di batteri lattici sulla caseina . Sapore di muffa potrebbe essere indice di una eccessiva proliferazione di muffe che si sviluppano anche a basse temperature. Conservazione del burro Si conserva in involucri di carta vegetale , plastificata o alluminata per proteggerlo dalla contaminazione esterna microbiologica, dalla luce e dall’ossigeno. In genere la contaminazione microbiologica trova nel burro un terreno di coltura poco favorevole. Purtroppo anche a basse temperature si possono avere proliferazioni batteriche psicrofile, solamente al di sotto dei -6°C l’attività microbica sul burro viene ostacolata mentre le reazioni chimiche sono solo rallentate
IL FORMAGGIO
Nella scelta dei latti da destinare alla produzione di formaggio devono essere tenuti in considerazione alcuni fattori. Ai batteri lattici compete la funzione acidificante indispensabile per la formazione della cagliata e per lo spurgo del siero , di conseguenza si deve permettere loro uno sviluppo ottimale (i batteri lattici sono molto esigenti dal punto di vista nutrizionale come frazioni azotate, vitamine ed alcuni oligoelementi) . Anche la presenza di eccessiva quantità di ossigeno non facilita i batteri lattici ed inoltre l’aumento dell’acidità dovuto alla produzione di acido lattico seleziona solo i batteri meno sensibili ( a pH<5 lo Streptococcus lactis può subire dei danni e una perdita della sua attività). Inoltre con la refrigerazione prolungata del latte a 48 ore si può avere lipolisi e liberazione degli acidi grassi dai trigliceridi fattore inibente l’attività acidificante e proteolitica dei batteri lattici. Produzione del formaggio Il fine primo è la produzione della cagliata e il controllo della sua disidratazione . La cagliata non è altro che il coagulo proteico , essenzialmente a base di caseina, in grado di determinare o meno una buona caseificazione. Per definizione il formaggio o cacio è il prodotto che si ricava dal latte intero o parzialmente o totalmente scremato , oppure dalla crema in seguito a coagulazione acida o presamica anche facendo uso di fermenti e sale da cucina.

82
Durante la fase di insemenzamento vengono aggiunte al latte colture microbiche specifiche in grado di acidificare la cagliata attraverso la fermentazione omolattica ed eterolattica e di fornire enzimi che caratterizzeranno la maturazione.
• Cocchi: hanno azione rapida ma comportano bassi livelli di acidificazione (Streptococcus thermophilus nella crescenza)
• Bastoncini lattici: Lactobacillus bulgaricus con acidificazione più lenta ma più spinta. INNESTI SPONTANEI Sviluppo spontaneo della microflora tipica del latte prodotto in un dato luogo e in una tipica stagione o dal siero delle precedenti caseificazioni. È quello preferito perché permette di acquisire uno specifico aroma evitando la standardizzazione del prodotto e perché è resistente all’attacco di determinati virus capaci altresì di distruggere colture microbiche selezionate. LATTOINNESCO Impiegato per la produzione di formaggi a pasta molle nei quali si deve avere una rapida acidificazione: se viene richiesto molto spurgo, come per la Crescenza o il Bel paese, l’innesco è costituito prevalentemente da Streptococcus thermophilus. Se si richiede invece una azione spinta e una certa demineralizzazione , come per il Gorgonzola, si utilizza anche il Lactobacillus bulgaricus nonché una certa microflora mesofila. SIEROINNESCO Impiegato per la produzione di formaggi a pasta cotta come Parmigiano Reggiano e Grana Padano in cui si richiede una acidificazione lenta e spinta; si usano solitamente colture naturali costituite da lactobacilli termofili quali Lactobacillus bulgaricus, L.helveticus e L.lactis (importante anche la loro azione proteolitica). Il siero ha una azione privilegiante i cocchi piuttosto che i bastoncellari. INNESTI FUNGINI Muffe del tipo Penicillium roqueforti e camemberti, Aspergillus che vengono aggiunti al momento di formazione della cagliata come spore che germinano in seguito (durante la fase di maturazione ). Gli enzimi che si sviluppano sono in grado di svolgere spiccata azione proteolitica e lipolitico contribuendo alla formazione dell’odore e del sapore caratteristico del Gorgonzola.

83
ALTERAZIONI DEI FORMAGGI
• GONFIORE PRECOCE: causato da Coli-aerogenes come Escherichia coli e Aerobacter aerogenes oppure da lieviti. Presenti in latti molto inquinati non pastorizzati oppure in latti contenenti antibiotici . Essi fermentano il lattosio producendo acido lattico,CO2 e idrogeno; questa alterazione in genere compare entro le prime 12 ore dalla formazione della cagliata, durante la stufatura dei formaggi molli e la salatura dei formaggi duri.
• GONFIORE TARDIVO: fermentazione a carico dell’acido lattico prodotto durante la
maturazione del formaggio (tipico infatti di formaggi a media o lunga stagionatura). I microrganismi responsabili sono:
o Batteri butirrici fermentano acido lattico in butirrico , acido acetico, anidride carbonica e idrogeno;
o Batteri propionici producono acido propionico, acido acetico e anidride carbonica, questi sono batteri responsabili della tipica occhiatura dell’Emmental e del Gruviera; nel Grana viene aggiunta formaldeide e lisozima per frenarne l’azione cosa che non è invece consentita nella produzione del Parmigiano Reggiano;
• COLATURA O RAMMOLLIMENTO: causa di spurgo, acidificazione e salatura
insufficienti che consentono quindi lo sviluppo di Oidium lactis con proprietà proteolitiche con fuoriuscita di pasta alterata dalla crosta;
• AMARO: dovuto ad alcuni streptococchi a rapida azione proteolitica che causano
l’accumulo di peptidi amari in seguito alla demolizione della b-caseina .Come rimedi si usano pastorizzazioni basse e utilizzo di fermenti selezionati.
• MARCIUME: alterazioni putrefattive proteolitiche operate da Clostridium sporogenes ,
con pH elevati, eccessiva umidità e lavorazioni difettose,

84

85

86
BIBLIOGRAFIA Sicheri, G., Tecnologie agrarie volume 2, Hoepli, Milano, 2000 Bastasin, P., Ceresa, L., Industrie agroalimentari, Lucidano, Milano, 1999 Cappelli, P., Vannucci, V., Chimica degli alimenti Conservazione e trasformazioni, Zanichelli, Bologna, 1998 Madigan, Michael T., Martinko, J., Parker, J., Brock, Biologia dei microrganismi volume1 e 2, Ambrosiana, Milano, 2003 Sechi Nicola, dispense corso di ecologia, 2005-04-08 Farris,A., appunti lezioni a.a. 2004/2005

87





























