Iguá 4225, CP 11400, Montevideo, Uruguay Pilot Programme for ecosystemic monitoring in Cerro Verde (Uruguay): benthic invertebrates as a tool for conservation. PHASE II Full formal report

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Iguá 4225, CP 11400, Montevideo, Uruguay
Pilot Programme for ecosystemic
monitoring in Cerro Verde (Uruguay):
benthic invertebrates as a tool for
conservation.
PHASE II
Full formal report

Pilot Programme for ecosystemic monitoring in Cerro Verde
(Uruguay): benthic invertebrates as a tool for conservation
PHASE II
MSc. A. Carranza
EXECUTIVE SUMMARY
The aim of this proposal was to complete baseline studies of biodiversity of
benthic invertebrates at Cerro Verde, Uruguay. To this end, biological samples
were taken seasonally, within a stratified sampling design in the soft bottoms,
from the shallow subtidal (i.e. 3 m to 15 m). To date we completed nine extensive
faunistic surveys, collecting more than 70 taxa of benthic invertebrates in the
intertidal and subtidal zones of the study area. We are now working in the
taxonomic determination of the species recorded, with the aid of specialists in the
different taxa and local students. Some previously unreported species of
invertebrates were found, increasing the knowledge about the coastal biodiversity
of the Atlantic coast of Uruguay. Emerging knowledge will be of outmost
importance in the development of managing strategies for this area, which is a
well known feeding zone for several endangered vertebrate species. Products will
allow detection of environmental impacts in a cheap and rapid way. Divulgation of
the preliminary results were driven through presentation of several works in a
local meeting in June 2007. Part of the information gathered during PHASE I was
already published in mainstream scientific journals, and several more papers are
in preparation. In addition, since Cerro Verde is in process of incorporation to the
National System of Protected Areas (SNAP) as a first Pilot Protected Marine Area
of the in Uruguay, the information here collected will be of utmost importance to
strength the scientific basis for appropriate reserve management.

2
BACKGROUND
The increasing need for an adequate environmental management, triggered by the conspicuous degradation of coastal habitats worldwide, has raised awareness of the need for systematic research to asses biodiversity. The need is especially acute for the marine invertebrates, particularly groups that may be sensitive indicators of environmental change. In Uruguay, coastal marine fauna is far from being completely
documented, with little or no systematic effort directed toward taxonomic surveys and inventories of marine invertebrates. This lack of focus is further aggravated by the shortage of experienced field systematists and taxonomists. Worldwide, the importance of detailed taxonomic inventories has been increasingly recognized, since these constitute the basis for the development of ecosystem monitoring programs and the detection of environmental degradation. In this context, fully protected marine reserves
are an emerging tool for marine conservation and management. These areas may provide multiple benefits, including protection of habitat, conservation of biodiversity, insurance about environmental uncertainty and sites for scientific investigation, baseline information, education and recreation. However, data on several key ecological and life history features of marine reserves and communities are needed in order to select the placement and extent of an MPA (marine protected area). In this sense, the aim of this project is to determine benthic invertebrate specific richness in the intertidal and subtidal zone, larval supply from the water column and annual cycles of studied fauna at Cerro Verde, Uruguay, in order to provide a baseline for the evaluation of environmental impacts and to contribute to the design of a MPA.
STUDY AREA
The biodiversity and productivity of the coastal ecoregion "Uruguay-Buenos Aires Shelf" (34°- 41° S), summed to the magnitude of
antrophogenic activities (tourism, urbanisation, industrial and domestic effluents, fisheries), confer to this coastal ecoregion a high priority for conservation. Cerro Verde (33° 57'S, 53°30' W) is a rocky cape on the east coast of Uruguay affected by semidiurnal, low-amplitude tides (range < 0.5 m) that are largely controlled by wind conditions (direction and speed). The rocky platforms have a smooth slope, with a width ranging from 15 to 23 m, and are exposed to different degrees of

3
wave action according to its orientation. These platforms follow a classical zonation scheme, in which three zones can be identified: a high intertidal zone dominated by a cyanobacterial film, a middle intertidal zone dominated by barnacles and a low intertidal and shallow subtidal zone characterized by a dense cover of mussels and/or macroalgae. This site harbours a rich hard-substrata benthic fauna, a yet non-defined number of fish species (e.g. endangered sharks Mustelus schmitti, M. fasciatus, Sphyrna bigelowi) and marine birds, mammals (Otaria bryonia, Arctocephalus australis) and sea turtles (Chelonya mydas). This site has recently been established as the first pilot marine protected area in Uruguay. For this purpose, quantitative data on species abundance, distribution, and temporal variation are of outmost importance in order to asses reserve effectiveness. In this context, our proposal was based on the urgent need for this relevant information. In this context, we developed a two years research programme in the area, focused in benthic invertebrates and its planktonic larval stages. This will allow us to asses benthic organism biodiversity and its dynamic at Cerro Verde, providing a baseline for the evaluation of environmental impacts and to contribute to the management of the MPA.
THE TEAM
Alvar Carranza is Uruguayan, 35 years old, MSc. in Ecology (2006), Facultad de Ciencias, Universidad de la República, is an advanced PhD. student of PEDECIBA (Basic Sciences Development Programme), since 2007. Previous work includes Project PNUD-GEF-RLA 99-G31 (FREPLATA).
Ana Inés Borthagaray is Uruguayan and aged 31 years old. Four-year BSc, honours in Oceanography (2002), Facultad de Ciencias, Universidad de la República, ia also an advanced PhD. student, since 2006. Previous work includes colaboration with PPNUD-GEF-RLA 99-G31 (FREPLATA). Fabrizio Scarabino is specialist in invertebrate taxonomy. He is currently working at the National Museum of Natural history (Montevideo) and National direction
of Aquatic resources (DINARA). Dra. Estela Delgado is helping with crustacean biology. Dr . Gabriel Genzano (Universidad de Mar del Plata, Argentina) helped with identification of Pycnogonids. Michela Borges from Departamento de Zoología, Instituto de Biologia, UNICAMP, Sao Paulo, Brazil, helped with taxonomic determination of Ophiuroideans Angel Segura and Luis Rubio are in charge of data processing and field work. Project “Karumbe-CIID”, an NGO working with sea turtles conservation and ecology in Uruguay provided facilities and contacts with the local community. This NGO is currently developing

4
conservation projects in the Cerro Verde-La Coronilla area. (http://www.karumbe.8k.com/). Facultad de Ciencias, Universidad de la República, in agreement with the organisation I+D (“Investigación y Desarrollo”) is providing facilities for laboratory work. Voucher material is being deposited at the National Museum of Natural History
RESEARCH ACTIVITIES
Field work The Field work was performed with the help of local fishermen and graduate and undergraduate students. Samplings were made onboard the artisanal fishing vessel “Dommy”, during January, May and September 2007. The fishing gears employed consisted in: a) a bottom
trawl net (BTN) with a 9 m horizontal opening, 1.2 m high and a 25 mm stretched mesh in the cod ends and b) a Piccard epibenthic dredge (PEB), with a horizontal aperture of 60cm and 1cm mesh size. In each date some 9 stations were allocated based on a systematic stratified design, between the outer breaker and aproximatelly 10-15 m depth (see map for details). In each station, 10 minutes tows were performed with the BTN, while 5 min tows were performed with the PEB. The exact location of the stations was determined by Garmin-Etrex Global Positioning System (GPS), depth was noticed usign onboard ECOSONDA. Once landed, all benthic animals collected were fixed and taken to laboratory. Fish was also collected, and stomach
contents kept for subsequent dietary analyses. Voucher material is deposited at the National Museum of Natural History, Montevideo
(MNHNM). Laboratory work Organisms fixed were identified and counted in the laboratory. Taxonomic determination is being made for most of the macrofaunal invertebrates collected since no comprehensive inventory of the benthic fauna is available yet. These imply the collaboration with regional researchers’ specialized in particular zoological groups, to avoid misidentifications and/or erroneuous taxonomic determination. It is remarkable that at lest three species were first recorded for the Uruguayan coast by means of this strategy (see attached material).
PRELIMINARY RESULTS
To date, we completed the surveys corresponding to summer, autumn, winter and spring 2007, collecting ca. 50 semi-quantitative samples. During the first survey, (January 2007) we collected 13 macroinvertebrate taxa, being the samples dominated by the crustaceans Loxopagurus loxochelis and Libinia espinosa (45,31 and 26,56% respectively of the collected

5
individuals). Other taxonomic grouops particularly well represented were molluscs (Pachycymbiola brasiliana, Buccinanops cochlidium and B. duartei), cnidarians (Antholoba achates) and Echinodermata (Mellita quinquiesperforata). Two decapod species, Hepatus pudibundus and
Araneus cribarius were also recorded, together with several species of peracariid crustaceans (Isopoda and Amphipoda). Based on prelim nary observations from the samples collected during the remaining surveys, it is estimated that at leas 50 species of benthic macroinvertebrates were recorded. In adittion, individuals of 18 of bony fishes and 5 chondrictians were captured. From these, Mustelus schmitti Sympterygia acuta are currently listed in the IUCN red list as endangered and threatened respectively. From 153 stomach contents analysed to date, representing 17 fish species, 92.6 % presented some content (Urophysis brasiliensis n=23 (100%), Paralonchurus brasiliensis n=18 (94.1%), Micropogonias furnieri n=31 (90.3%), Macrodon ancylodon n=29 (86.2%), Stromateus brasiliensis n=12 (91.7%) y Sympterygia acuta n=18 (94.1%). P. brasiliensis showed a high number of polychatetes, while S. brasiliensis showed
preferences for small peracarid crustaceans (Mysida). The shrimp Artemesia longinaris was the main item in the remaining species, reaching 78.3% occurrence in Urophysis brasiliensis. The presence of amphipods and fish larvae (16% of ocurrente in Macrodon ancylodon) was also registered. The hermit crab Loxopagurus loxochelis was preyed by Mustelus schmitti and Menticirrus americanus. Stellifer rastrifer, a coastal fish species that inhabits the Tropical and Southwestern Atlantic Ocean is reported for the first time from the Uruguayan Coast and its geographic distribution range is extended southward in more than 1200 km. A warm-circulation event in the zone is suggested as a putative factor explaining the occurrence of the fish in the area.
RESULT DISSEMINATION
The lsit of publicatiions to date in peer-reviewed journals and edited books is listed below:
2007: CARRANZA, A, BORGES, M, RODRÍGUEZ, M & BORTHAGARAY, AI. Ophiuroidea (Echinodermata) from La Coronilla-Cerro Verde (Uruguay): a new record for the Uruguayan coast. Biota Neotropica, 7(3): http://www.biotaneotropica.org.br 2007: CARRANZA, A., BORTHAGARAY, A. I. & GENZANO, G. N. Two new records of pycnogonids on the Uruguayan coast. Brazilian Journal of Biology, 67 (2):373-375

6
2007: BORTHAGARAY, A. I. & CARRANZA, A. Mussels as ecosystem engineers: Their contribution to species richness in rocky littoral community. Acta Oecologica 31: 243-250 2007: CARRANZA, A. Pilot programme of ecosystem monitoring in Cerro Verde (Uruguay): two years of malacological assessment. Tentacle.The Newsletter of the IUCN/SSC Mollusc Specialist Group, 15: 22-23 2006: SCARABINO, F., ZAFFARONI, JC., CARRANZA, A., CLAVIJO, C. & NIN, M. Gasterópodos marinos y estuarinos de la costa uruguaya: faunística, distribución, taxonomía y conservación. pp 143-155 En Bases para la conservación y el manejo de la costa uruguaya, Menafra R, Rodríguez-Gallego L, Scarabino F & Conde D (Eds), VIDA SILVESTRE (Sociedad Uruguaya para la Conservación de la Naturaleza), Montevideo 2006: SCARABINO, F., ZAFFARONI, JC., CLAVIJO, C., CARRANZA, A. & NIN, M. Bivalvos marinos y estuarinos de la costa uruguaya: faunística, distribución, taxonomía y conservación. pp 157-169 En Bases para la conservación y el manejo de la costa uruguaya, Menafra R, Rodríguez-Gallego L, Scarabino F & Conde D (Eds), VIDA SILVESTRE (Sociedad Uruguaya para la Conservación de la Naturaleza), Montevideo
In press: CARRANZA, A., SEGURA, A., LÓPEZ, J. & RUBIO, L. Shell use patterns of the hermit crab Loxopagurus loxochelis (Decapoda: Diogenidae) in the Cerro Verde-La Coronilla marine protected area. Comunicaciones de la Sociedad Malacológica del Uruguay In adittion, we presented the following posters and abstracts in local meetings (local schools and university) and presentations in regional or local congresses. 2007: CARRANZA, A. Investigación, monitoreo y conservación: invertebrados bentónicos del area marina protegida de Cerro Verde (Rocha, Uruguay) IV Congreso Nacional de Áreas
Protegidas, Trinidad, Uruguay. 2007: RUBIO, L., SEGURA, A. & CARRANZA, A. La comunidad nectónica en el área marina protegida de Cerro Verde. IV Congreso Nacional de Areas Protegidas ,Trinidad, Uruguay 2005: CARRANZA, A., BORTHAGARAY, A. I. & GENZANO, G. N. Dos nuevos registros de Pycnogonida para aguas Uruguayas. VIII Jornadas de Zoología del Uruguay, Facultad de Ciencias, Montevideo, Uruguay. 2005: CARRANZA, A. & BORTHAGARAY, A. I. Los mejillones como ingenieros ecosistémicos: su contribución a la riqueza especifica en la

7
comunidad del litoral rocoso. II Encuentro de Ecología del Uruguay, Facultad de Ciencias, Montevideo, Uruguay. Final results will also be presented to the Uruguayan committee of the IUCN and all he people implicated, in different ways, in coastal management The field work was supported by The Maurice Laing Foundation Rufford Small Grant for Nature conservation. SCUBA divers Pablo Píriz and Jorge Duran and S. Horta,S. Sauco and Dr. Danilio Calliari are acknowledged for collaboration during field work.
8
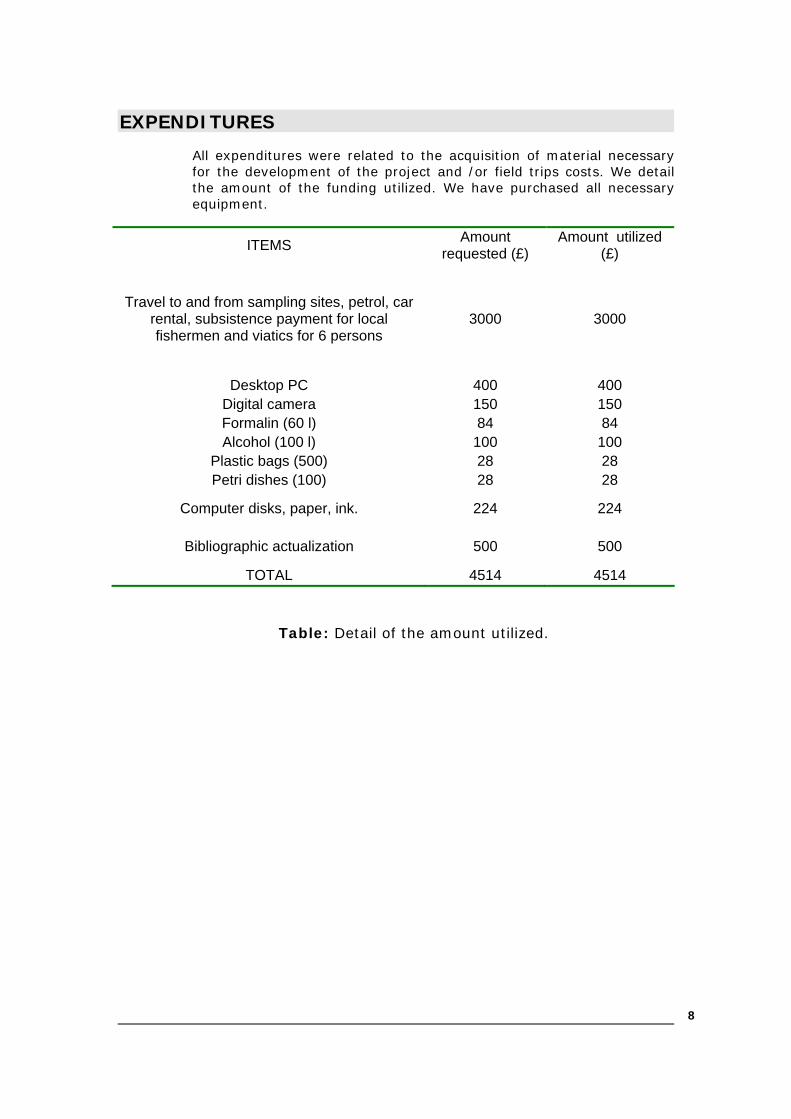
EXPENDITURES
All expenditures were related to the acquisition of material necessary for the development of the project and /or field trips costs. We detail the amount of the funding utilized. We have purchased all necessary equipment.
ITEMS Amount requested (£)
Amount utilized (£)
Travel to and from sampling sites, petrol, car rental, subsistence payment for local fishermen and viatics for 6 persons
3000 3000
Desktop PC 400 400 Digital camera 150 150 Formalin (60 l) 84 84 Alcohol (100 l) 100 100
Plastic bags (500) 28 28 Petri dishes (100) 28 28
Computer disks, paper, ink. 224 224
Bibliographic actualization 500 500
TOTAL 4514 4514
Table: Detail of the amount utilized.

A Biota Neotropica é uma revista eletrônica e está integral e gratuitamente disponível no endereço http://www.biotaneotropica.org.br
Biota Neotropica is an eletronic journal which is available free at the following site http://www.biotaneotropica.org.br
Biota Neotropica é uma revista do Programa BIOTA/FAPESP - O Instituto Virtual da Biodiversidade, que publica resultados de pesquisa original, vinculada ou não ao programa, que abordem a temática
caracterização, conservação e uso sustentável da biodiversidade na região Neotropical.
Biota Neotropica is an electronic, peer-reviewed journal edited by the Program BIOTA/FAPESP: The Virtual Institute of Biodiversity. This journal’s aim is to disseminate the results of original research work,
associated or not to the program, concerned with characterization, conservation and sustainable use of biodiversity within the Neotropical region.
Ophiuroidea (Echinodermata) from La Coronilla-Cerro Verde (Uruguay): a new record for the Uruguayan coast
Carranza, A. et al.
Biota Neotropica, Vol.7 (number 3): 2007; p. 000-000.
A versão on-line completa deste artigo está disponível em:On line version of this paper is available at:http://www.biotaneotropica.org.br/v7n3/pt/abstract?article+bn01607032007
Recebido em/ Data Received 22/01/07 - Versão reformulada recebida em/ Revised 30/05/07 - Publicado em/ Accepted 06/09/07
ISSN 1676-0603 (on-line)

http://www.biotaneotropica.org.br
Ophiuroidea (Echinodermata) from La Coronilla-Cerro Verde (Uruguay): a new record for the Uruguayan coast
Alvar Carranza1, 2, 4, Michela Borges3, Marcel Rodríguez2 & Ana Inés Borthagaray1, 2
Biota Neotropica v7 (n3) – http://www.biotaneotropica.org.br/v7n3/pt/abstract?article+bn01607032007
Data Received 22/01/07 Revised 30/05/07
Accepted 06/09/07
1Investigación & Desarrollo, Iguá 4225, CP 11400, Montevideo, Uruguay, 2Facultad de Ciencias, Iguá 4225, CP 11400, Montevideo, Uruguay
3Departamento de Zoología, Instituto de Biologia, UNICAMP, CP 6109, CEP 13083-970, Campinas, SP, Brasil, http://www.unicamp.br
4Corresponding author: Alvar Carranza e-mail [email protected], http://imasd.fcien.edu.uy/, http://www.fcien.edu.uy/
Abstract
Carranza, A., Borges, M., Rodríguez, M. & Borthagaray, A. I. Ophiuroidea (Echinodermata) from La Coronilla-Cerro Verde (Uruguay): a new record for the Uruguayan coast. Biota Neotrop. Sep/Dez 2007 vol. 7, no. 3 http://www.biotaneotropica.org.br/v7n3/pt/abstract?article+bn01607032007. ISSN 1676-0603.
La Coronilla-Cerro verde has been proposed as the first marine protected area in Uruguay. As part of a detailed benthic biodiversity assessment at the reserve, we analyzed the ophiuroid fauna collected in the intertidal and shallow subtidal during 2005 and 2006. Three species of ophiuroids were identified: Amphioplus lucyae, Amphipholis squamata and Amphiodia sp. Only two species belonging to the genus Amphiodia (A. pulchella and A. planispina) have been previously mentioned for Uruguayan waters, indicating that the unidentified species found in this study constitutes a previously unreported species.
Keywords: Amphiodia, Amphioplus, Amphipholis, Cerro Verde, marine protected area.
Resumen
Carranza, A., Borges, M., Rodríguez, M. & Borthagaray, A. I. Ophiuroidea (Echinodermata) de La Coronilla-Cerro Verde (Uruguay): un nuevo registro para la costa uruguaya. Biota Neotrop. Sep/Dez 2007 vol. 7, no. 3 http://www.biotaneotropica.org.br/v7n3/pt/abstract?article+bn01607032007. ISSN 1676-0603.
La Coronilla-Cerro Verde ha sido propuesta como la primera área marina protegida del Uruguay. Como parte de un detallado análisis de la biodiversidad bentónica dentro de la reserva, se analizó la fauna de ofiuroideos colectada en el intermareal y submareal somero durante 2005 y 2006. Se identificaron tres especies: Amphioplus lucyae, Amphipholis squamata y Amphiodia sp. Solo dos especies del género Amphiodia han sido previamente mencionadas para la costa uruguaya (A. pulchella, y A. planispina) indicando que la especie de este género encontrada en este estudio corresponde a una especie aun no reportada.
Palabras clave: Amphiodia, Amphioplus, Amphipholis, Cerro Verde, área marina protegida.

2 Carranza, A et al. - Biota Neotropica, v7 (n3) - bn01607032007
http://www.biotaneotropica.org.br
Introduction
The increasing need for an adequate environmental management, triggered by the conspicuous degradation of coastal habitats world-wide, has raised awareness of the need for systematic research to asses biodiversity. The need is especially acute for the marine invertebrates, particularly groups that may be sensitive indicators of environmental change (Thomas 1997, Basset et al. 2004). In Uruguay, coastal marine fauna is far from being completely documented, with little or no sys-tematic effort directed toward taxonomic surveys and inventories of marine invertebrates. This lack of focus is further aggravated by the shortage of experienced field systematists and taxonomists (Scarabino 2006). Worldwide, the importance of detailed taxonomic inventories has been increasingly recognized (Gotelli 2004), since these constitute the basis for the development of ecosystem monitoring programs and the detection of environmental degradation.
Among the benthic invertebrates, the Echinodermata comprises nearly 6600 recent species, constituting one of the most important benthic groups worldwide (Hendler et al. 1995). The approximately 2000 species of ophiuroids are among the most widely distributed Echinodermata, occurring from polar regions to the tropics and from the intertidal to the abyssal plain (Hendler et al. 1995). Owing to their ubiquity and abundance, ophiuroids play an important role in energy transfer in benthic ecosystems, linking levels of local trophic webs (Summers & Nybakken 2000).
Previously, the Uruguayan ophiuroidean fauna has been only studied on the continental shelf, from shallow environments (10 m) to 800 m on the slope area (Milstein et al. 1976, Lucchi 1985), with intertidal or shallow subtidal environments receiving little attention. Recently, Scarabino (2006) detailed all invertebrates (except mollusks) reported for Uruguayan shallow waters (i.e. < 50 m), thus providing a basis to distinguish previously unreported species.
In this paper, we analyzed the ophiuroid fauna collected at the in-tertidal and shallow subtidal areas of Cerro Verde. La Coronilla-Cerro verde has been proposed as the first marine protected area in Uruguay, partially due to the existence of several research projects focused in different aspects of its biodiversity. Concerning benthic invertebrates, a detailed faunistic inventory is being performed at the intertidal and subtidal mussel beds and the submarine beach (Carranza & Borthagaray 2005, Carranza et al. 2005, Borthagaray & Carranza in press).
Material and Methods
1. Study area
Cerro Verde (33° 57’S and 53°30’ W) is a rocky cape on the east coast of Uruguay (Figure 1). On each side of the cape there are sandy beach arcs that extend 2-3 km. The coast experiences a semidiurnal tide (range < 0.5 m) with the water level influenced mainly by wind conditions (direction and speed). Winds blow south-west during win-ter and north-east during summer. The rocky platforms have a smooth slope, with a width ranging from 15 to 23 m, and are exposed to dif-ferent degrees of wave action according to their orientation. Three zones, following the classical zonation schemes can be identified [on the platform]: a high intertidal zone dominated by a cyanobacterial film, a middle intertidal zone dominated by barnacles and a low in-tertidal and shallow subtidal zone characterized by a dense cover of mussels and/or macroalgae. Subtidal, deeper (>3-4 m) environments are dominated by sandy sediments (Milstein et al. 1976).
2. Sampling design
Sampling was carried out on intertidal and shallow subtidal (i.e. depth < 1.5 m) rocky platforms of the Cerro Verde area dur-
ing February 2005 and February 2006. Three sampling sites 500 m apart were chosen along the coast: 1) exposed, oriented S-SW, 2) intermediate, and 3) protected, oriented N-NE. Within each site, we randomly selected points at each dominance zone (mid intertidal, low intertidal and shallow subtidal), separated by 2 to 10 m. In each point we placed one quadrat of 20 x 20 cm (0.04 m2) and collected all macrofauna and algae present using a metallic hand scrapper. In the shallow subtidal, three points were sampled by SCUBA divers, all in sandy bottoms: at each point, three samples were taken with a plastic corer of 15 cm diameter, each sample consisting in 5 cores. Organisms were fixed with formalin and, in the laboratory, identified and counted. Voucher material is deposited at Museo Nacional de Historia Natural y Antropología, Montevideo (MNHNM).
Results
A total of 13 specimens of ophiuroids, belonging to 3 species and 1 family (Amphiuridae) were collected. Two specimens were collected in the rocky intertidal and the remaining 11 at the shallow subtidal, in sandy bottoms ranging from 5-7 m depth. A detailed list of the examined material is provided below.
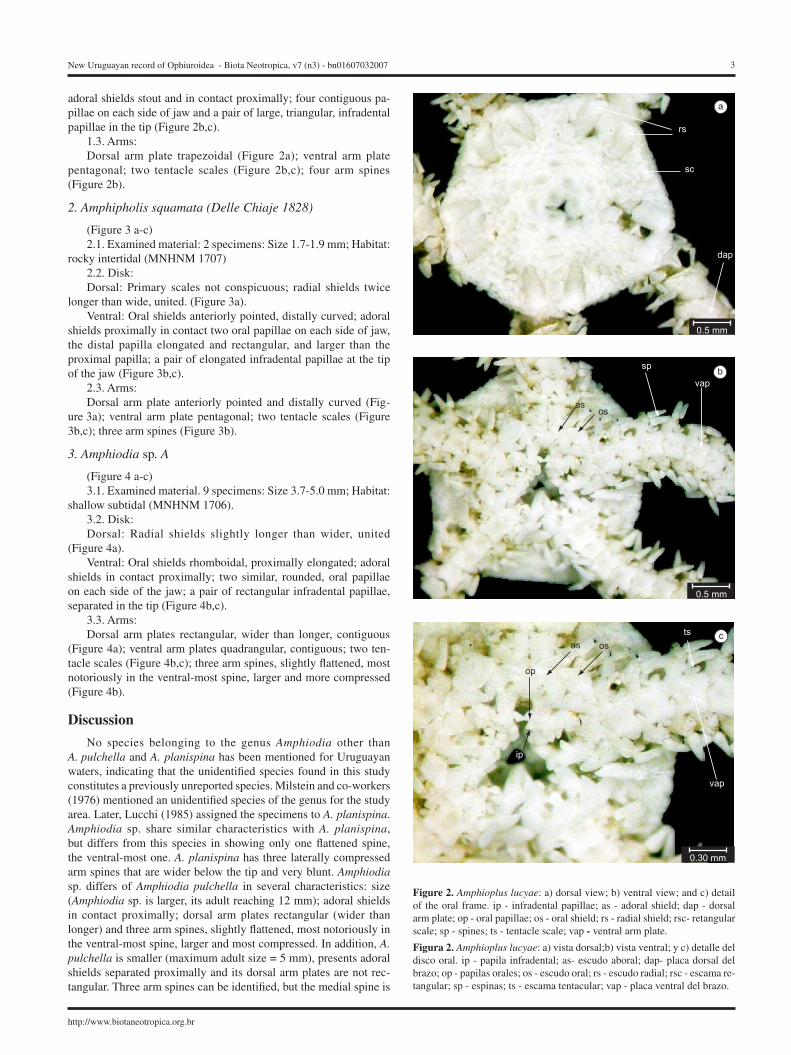
1. Amphioplus lucyae
Tommasi 1971 (Figure. 2 a-c)1.1. Examined material: 2 specimens: Size 2.6 – 3.8 mm; Habitat:
Shallow subtidal. (MNHNM 1705) 1.2. Disk:Dorsal: Marginal interradial region showing a rectangular (= mar-
ginal) scale slightly larger than adjacent ones; radial shield cuneiform, separated and divergent (Figure 2a). Ventral: oral shields elongated;
Argentina
Uruguay
South Atlantic Ocean
Río de la Plata
58°
40°
39°
38°
37°
36°
35°
34°
33°
57° 56° 55° 54° 53° 52°
S1
S2
S3
i3
i2
Cerro Velho
i1
1 Km
Figure 1. Map of South American Atlantic coast, showing the study region in the coast of Uruguay. Sampling sites: i1 - i3: intertidal stations; S1 - S3: subtidal stations.
Figura 1. Mapa de la costa Atlántica de Sudamérica, mostrando el área de es-tudio en la costa Uruguaya. Sitios de muestreo: i1- i3: estaciones intermareales S1- S3: estaciones submareales.
3New Uruguayan record of Ophiuroidea - Biota Neotropica, v7 (n3) - bn01607032007
http://www.biotaneotropica.org.br
adoral shields stout and in contact proximally; four contiguous pa-pillae on each side of jaw and a pair of large, triangular, infradental papillae in the tip (Figure 2b,c).
1.3. Arms:Dorsal arm plate trapezoidal (Figure 2a); ventral arm plate
pentagonal; two tentacle scales (Figure 2b,c); four arm spines (Figure 2b).
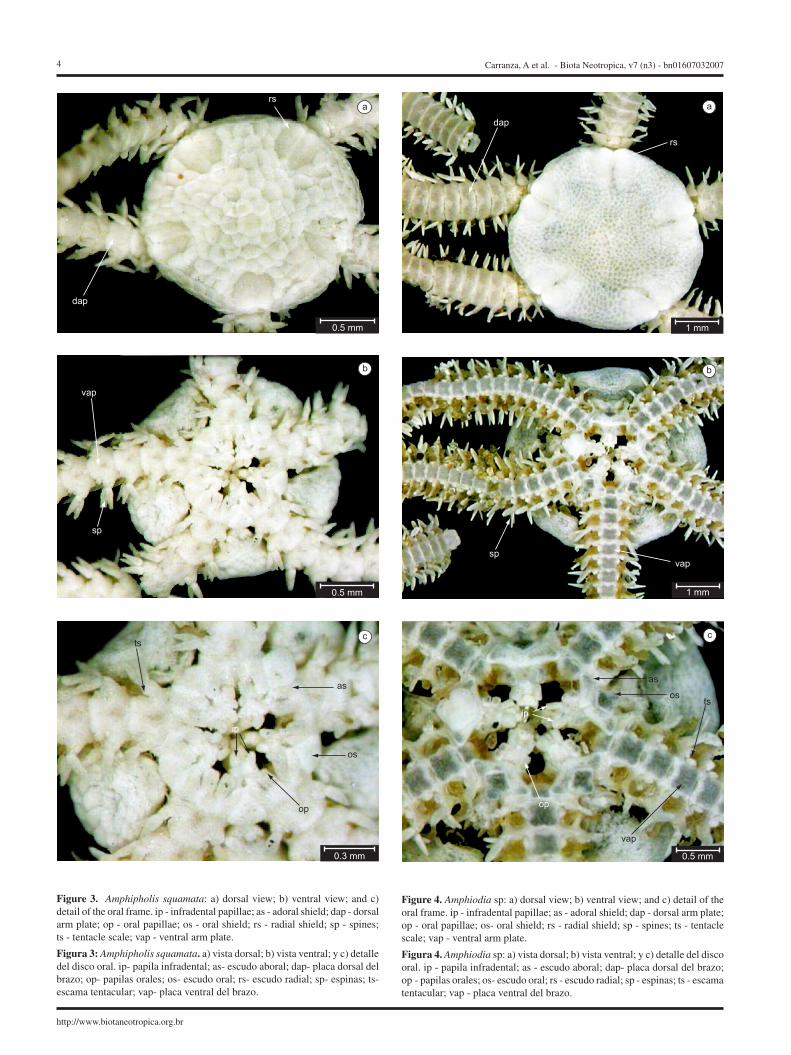
2. Amphipholis squamata (Delle Chiaje 1828)
(Figure 3 a-c)2.1. Examined material: 2 specimens: Size 1.7-1.9 mm; Habitat:
rocky intertidal (MNHNM 1707) 2.2. Disk:Dorsal: Primary scales not conspicuous; radial shields twice
longer than wide, united. (Figure 3a).Ventral: Oral shields anteriorly pointed, distally curved; adoral
shields proximally in contact two oral papillae on each side of jaw, the distal papilla elongated and rectangular, and larger than the proximal papilla; a pair of elongated infradental papillae at the tip of the jaw (Figure 3b,c).
2.3. Arms:Dorsal arm plate anteriorly pointed and distally curved (Fig-
ure 3a); ventral arm plate pentagonal; two tentacle scales (Figure 3b,c); three arm spines (Figure 3b).
3. Amphiodia sp. A
(Figure 4 a-c)3.1. Examined material. 9 specimens: Size 3.7-5.0 mm; Habitat:
shallow subtidal (MNHNM 1706). 3.2. Disk:Dorsal: Radial shields slightly longer than wider, united
(Figure 4a). Ventral: Oral shields rhomboidal, proximally elongated; adoral
shields in contact proximally; two similar, rounded, oral papillae on each side of the jaw; a pair of rectangular infradental papillae, separated in the tip (Figure 4b,c).
3.3. Arms:Dorsal arm plates rectangular, wider than longer, contiguous
(Figure 4a); ventral arm plates quadrangular, contiguous; two ten-tacle scales (Figure 4b,c); three arm spines, slightly flattened, most notoriously in the ventral-most spine, larger and more compressed (Figure 4b).
Discussion
No species belonging to the genus Amphiodia other than A. pulchella and A. planispina has been mentioned for Uruguayan waters, indicating that the unidentified species found in this study constitutes a previously unreported species. Milstein and co-workers (1976) mentioned an unidentified species of the genus for the study area. Later, Lucchi (1985) assigned the specimens to A. planispina. Amphiodia sp. share similar characteristics with A. planispina, but differs from this species in showing only one flattened spine, the ventral-most one. A. planispina has three laterally compressed arm spines that are wider below the tip and very blunt. Amphiodia sp. differs of Amphiodia pulchella in several characteristics: size (Amphiodia sp. is larger, its adult reaching 12 mm); adoral shields in contact proximally; dorsal arm plates rectangular (wider than longer) and three arm spines, slightly flattened, most notoriously in the ventral-most spine, larger and most compressed. In addition, A. pulchella is smaller (maximum adult size = 5 mm), presents adoral shields separated proximally and its dorsal arm plates are not rec-tangular. Three arm spines can be identified, but the medial spine is
Figure 2. Amphioplus lucyae: a) dorsal view; b) ventral view; and c) detail of the oral frame. ip - infradental papillae; as - adoral shield; dap - dorsal arm plate; op - oral papillae; os - oral shield; rs - radial shield; rsc- retangular scale; sp - spines; ts - tentacle scale; vap - ventral arm plate.
Figura 2. Amphioplus lucyae: a) vista dorsal;b) vista ventral; y c) detalle del disco oral. ip - papila infradental; as- escudo aboral; dap- placa dorsal del brazo; op - papilas orales; os - escudo oral; rs - escudo radial; rsc - escama re-tangular; sp - espinas; ts - escama tentacular; vap - placa ventral del brazo.
a
rs
sc
dap
0.5 mm
asos
sp
vapb
0.5 mm
ts
vap
ip
op
as osc
0.30 mm
4 Carranza, A et al. - Biota Neotropica, v7 (n3) - bn01607032007
http://www.biotaneotropica.org.br
Figure 4. Amphiodia sp: a) dorsal view; b) ventral view; and c) detail of the oral frame. ip - infradental papillae; as - adoral shield; dap - dorsal arm plate; op - oral papillae; os- oral shield; rs - radial shield; sp - spines; ts - tentacle scale; vap - ventral arm plate.
Figura 4. Amphiodia sp: a) vista dorsal; b) vista ventral; y c) detalle del disco oral. ip - papila infradental; as - escudo aboral; dap- placa dorsal del brazo; op - papilas orales; os- escudo oral; rs - escudo radial; sp - espinas; ts - escama tentacular; vap - placa ventral del brazo.
rs
dap
a
0.5 mm
vap
sp
b
0.5 mm
as
ts
ip
op
os
c
0.3 mm
Figure 3. Amphipholis squamata: a) dorsal view; b) ventral view; and c) detail of the oral frame. ip - infradental papillae; as - adoral shield; dap - dorsal arm plate; op - oral papillae; os - oral shield; rs - radial shield; sp - spines; ts - tentacle scale; vap - ventral arm plate.
Figura 3: Amphipholis squamata. a) vista dorsal; b) vista ventral; y c) detalle del disco oral. ip- papila infradental; as- escudo aboral; dap- placa dorsal del brazo; op- papilas orales; os- escudo oral; rs- escudo radial; sp- espinas; ts- escama tentacular; vap- placa ventral del brazo.
a
rs
dap
1 mm
b
spvap
1 mm
c
ip
op
as
osts
vap
0.5 mm

5New Uruguayan record of Ophiuroidea - Biota Neotropica, v7 (n3) - bn01607032007
http://www.biotaneotropica.org.br
dorsoventrally flattened, with a truncate, echinulate tip; the other two spines are bluntly rounded. However, until appropriate comparison with specimens for Amphiodia species that have been reported from Brazil and Argentina we will not be able to suggest that it is an unde-scribed species. According to our samples, this species seems to be the more abundant ophiuroid in the area, with maximum estimated densities of 8.66 ind/m2.
To our knowledge, Tommasi et al. (1988b) constitutes the only previous known record of Amphioplus lucyae for the Uruguayan coast (R/V “Almirante Saldanha”, Station 2865, 34° 33’ S and 53° 12’ W). This species was previously reported for southwestern Brazil (Tommasi 1971, 1999, Monteiro 1987, Borges et al. 2002). Tommasi and co-workers (1988a, 1988b), Monteiro (1987, 1990, 1997) and Borges et al. (2002) reported A. lucyae at depths ranging from 8-600 m. We collected this species only in one subtidal sample (depth 5-7 m), suggesting low population densities at the area (maximum density < 5 ind. /m2).
A. squamata is an euribathic, cosmopolitan species, recorded worldwide with Atlantic records from Florida (USA) to Argentina, occurring from 0 to 1330 m (Hendler et al. 1995), and associated to different biological substrata such as algae, bryozoans, and sponges (Borges & Amaral, 2005). Tommasi and co-workers (1988b) already reported this species for the Uruguayan coast, while Milstein and co-workers (1976) reported specimens from subtidal environments of our study area. Alves & Cerqueira (2000) reported intertidal records for this species, as did Bernasconi (1926) for the Argentinean coast. In this study, this species occurred exclusively in the rocky intertidal, associated with mussel banks dominated by Brachidontes rodriguezii and Perna perna, constituting the first record of this species in this particular habitat for the Uruguayan coast. Most likely, the sediment layer occurring between the mussel layer and the rock surface al-lows the presence of ophiuroids. This species seems to be rare in the mussel banks of the study area, as indicated by the low frequency of occurrence (0.01% of total intertidal samples).
We strongly stressed the need for detailed taxonomic studies on other benthic invertebrates at the study area, aiming to provide a precise taxonomic inventory of the benthic fauna within the reserve area.
Acknowledgments
The field work was supported by The Maurice Laing Foundation Rufford Small Grant for Nature conservation. SCUBA divers Pablo Píriz and Jorge Duran and Dr. Danilio Calliari are acknowledged for collaboration during field work. We also thank F. Scarabino for the valuable comments made on an earlier version of the manu-script. Our thanks to the Departamento de Zoologia, Instituto de Biologia, UNICAMP, for the facilities and assistance with figures. Two anonymous reviewers are acknowledged for their very helpful suggestions.
ReferencesALVES, O.F.S. & CERQUEIRA, W.R.P. 2000. Echinodermata das praias de
Salvador (Bahia, Brasil). Rev. Bras. Zool. 17:543-553.
BASSET, A., SANGIORGIO, F. & PINNA, M. 2004. Monitoring with benthic macroinvertebrates: advantages and disadvantages of body size descriptors. Aquat. Conserv. 14:S43-S58.
BERNASCONI, I. 1926. Una ofiura vivípara de Necochea. An. Mus. Hist. Nat. (Buenos Aires). 34:145-153.
BORGES, M. & AMARAL, A.C.Z. 2005. Classe Ophiuroidea.. In Manual de Identificação dos Invertebrados Marinhos da Região Sudeste-Sul do
Brasil. (A.C.Z. AMARAL, A.E. RIZZO & ARRUDA, E.P. eds). Vol. I. EDUSP Ed. São Paulo. p.238-272
BORGES, M., MONTEIRO, A.M.G. & AMARAL, A.C.Z. 2002. Taxonomy of Ophiuroidea (Echinodermata) from the continental shelf and slope of the Southern and Southeastern brazilian coast. Biota Neotropica, 2:1-69 http://www.biotaneotropica.org.br/v2n2/pt/fullpaper?bn02302022002_1+pt (last accesss in 14/12/2006).
BORTHAGARAY, A.I. & CARRANZA, A. In press. Mussels as ecosystem engineers: Their contribution to species richness in rocky littoral com-munity. Acta Oecol. Int. J. Ecol.
CARRANZA, A. & BORTHAGARAY, A.I. 2005. Los mejillones como ingenieros ecosistémicos: su contribución a la riqueza especifica en la comunidad del litoral rocoso. Publ. Esp. Soc. Zool. Uruguay. Act. VIII. Jorn. Zool. Uruguay. 65 [Abstract]
CARRANZA, A., BORTHAGARAY, A.I. & GENZANO, G.N. 2005. Dos nuevos registros de Pycnogonida para aguas Uruguayas. Publ. Esp. Soc. Zool. Uruguay. Act. VIII. Jorn. Zool. Uruguay. 49 [Abstract].
GOTELLI, N.J. 2004. A taxonomic wish-list for community ecology. Philos. Trans. R. Soc. Lond., B, Biol. Sci. 359:585-597.
HENDLER, G., MILLER, J.E., PAWSON, D.L. & KIER, P.M. 1995. Sea stars, sea urchins and allies: echinoderms of Florida and the Caribbean. Smithsonian Institution Press, Washington.
LUCCHI, C. 1985. Ophiuroidea (Echinodermata) del frente marítimo uru-guayo hasta 800 m de profundidad, con claves para su reconocimiento. Contrib. Depto. Oceanogr. (F.H.C), Montevideo, 2:115-171.
MILSTEIN, A., JUANICÓ, M. & OLAZARRI, J. 1976. Algunas asociaciones bentónicas frente a las costas de Rocha, Uruguay. Resultados de la cam-paña del R/V “Hero”, viaje 72-3a. Com. Soc. Malac. Urug. 4:143-164.
MONTEIRO, A.M.G. 1987. Ophiuroidea (Echinodermata) da região de Ubatuba (SP) - Aspectos morfológicos e ecológicos. Tese de Doutorado, Universidade de São Paulo, São Paulo.
MONTEIRO, A.M.G. 1990. Ophiuroidea (Echinodermata) in the coastal system of the State of São Paulo (Brazil). In Anais II Simpósio de Ecos-sistema da Costa Sul e Sudeste Brasileira: Estrutura, Função e Manejo, v.1. Águas de Lindóia, p.186-201.
MONTEIRO, A.M.G. 1997. Ocorrência de Ophiuroidea (Echinodermata) na plataforma do Estado de São Paulo. In Boletim de Resumos Expandidos VII COLACMAR, Santos, v.2, p.182-183.
SCARABINO, F. 2006. Faunística y taxonomía de invertebrados bentónicos marinos y estuarinos de la costa uruguaya. In Bases para la conservación y el manejo y de la costa uruguaya (R. Menafra, L. Rodríguez-Gallego, F. Scarabino & D. Conde, eds). VIDA SILVESTRE (Sociedad Uruguaya para la Conservación de la Naturaleza), Montevideo, p.113-142.
SUMMERS, A.C. & NYBAKKEN, J. 2000. Brittle star distribution patterns and population densities on the continental slope off Central California (Echinodermata: Ophiuroidea). Deep Sea Res. Part II Top. Stud. Ocea-nogr. 47:1107-1137.
THOMAS, J.D. 1997. Using Marine Invertebrates to Establish Research and Conservation Priorities. In Biodiversity II: Understanding and Protect-ing Our Biological Resources (M. L. Reaka-Kudla, D. E. Wilson, E. O. Wilson, eds.) Joseph Henry Press, London, p.357-370.
TOMMASI, L.R. 1971. Equinodermes do Brasil. I. Sobre algumas novas espécies e outras pouco conhecidas para o Brasil. Boletim do Instituto Oceanográfico, Universidade de São Paulo, 20:1-21;
TOMMASI, L.R., CASTRO, S.M. & SOUSA, E.C. 1988a. Echinodermata coletados durante as campanhas oceanográficas do N/Oc. “Almirante Saldanha” no Atlântico Sul Occidental. Relat. Interno Inst. Oceanogr. Univ. Sao Paulo. 21:1-11
TOMMASI, L.R. 1999. Echinodermata Recentes e Fósseis do Brasil. http://www.bdt.org.br/zoologia/echinodermata/ 12/12/2005
TOMMASI, L.R., CERNEA, M.C.W. & CONDEIXA, M.C.G. 1988b. Equi-nodermes coletados pelo N/Oc. “Almirante Saldanha” entre 26° 59’ S e 38° 39’ S. Relat. Interno Inst. Oceanogr. Univ. Sao Paulo. 22:1-11

Braz. J. Biol., 67(2A): 631-637, 2007 �
ID Artigo: 117-05 envio: 09/03/2007
cubomultimidia publicações e-mail: [email protected]
Two new records of pycnogonids on the uruguayan coast
Carranza, A.a*, Borthagaray, AI.a and Genzano, GN.a,b
aInvestigación & Desarrollo, Facultad de Ciencias Exactas, Universidad de la República, Iguá 4225, CP ��400, Montevideo, Uruguay
bDepartamento de Ciencias Marinas, Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Mar del Plata, Funes 3250 (7600) Mar del Plata, Argentina
*e-mail: [email protected]
Received September 8, 2005 – Accepted May �0, 2005 – Distributed May �, 2007
(With 2 figures)
Abstract
The Pycnogonida from Uruguayan waters are scarcely known, and their reports are markedly discontinuous. In this paper, several individuals of two previously unrecorded pycnogonids at the shallow rocky subtidal and lower intertidal fringes of Cerro Verde (Rocha, Uruguay) are reported. The specimens were assigned to Pycnogonum pamphorum Marcus, �940 and Anoplodactylus petiolatus (Kroyer, �844). This is the first record of these species on the Uruguayan coast and the southernmost record of P. pamphorum, previously recorded only in Santos, Brazil (type locality).
Keywords: Pycnogonida, Pycnogonum, Anoplodactylus, Uruguay.
Dois novos registros de pycnogonida para a costa uruguaia
Resumo
Pouco se conhece da fauna dos Pycnogonida de águas Uruguaias, e seus registros são marcadamente descontínuos. Neste trabalho, são reportados vários indivíduos de dois picnogonídeos não registrados previamente, nas faixas ro-chosas submareal rasa e bordas intermareais baixas de Cerro Verde (Rocha, Uruguai). Os espécimes foram atribuídos às espécies Pycnogonum pamphorum Marcus, �940 e Anoplodactylus petiolatus (Kroyer, �844). Este é o primeiro registro dessas duas espécies para a costa Uruguaia e constitui o registro mais austral para P. Pamphorum, conhecido previamente somente em Santos, Brasil (localidade tipo).
Palavras-chave: Pycnogonida, Pycnogonum, Anoplodactylus, Uruguai.
1. Introduction
Pycnogonids or sea spiders are quite common in many different marine habitats, from the intertidal zone to the abyssal depths. However, they are seldom seen due to their small size and cryptic coloration (Bain, �99�).
On the Uruguayan coast, the pycnogonids are largely unknown, with the available literature referring only to the larger and more conspicuous species, such as the members of the genus Colossendeis. In this vein, C. geofroyi Mañe-Garzón, �944 was originally described from Uruguayan specimens and received further atten-tion by Laramendy (�974), who analyzed material from R/V “Walter Hewig” cruise in the Uruguayan continen-tal shelf. Stock (�966) reported not only this species but also Tanystylum isthmiacum difficile Stock, �955, Nymphon sp. and Pycnogonum elephas (Stock, �966) for the South American Atlantic coast based on material col-lected by the R/V “Calypso”. Additionally, Juanicó and Rodríguez-Moyano (�976) reported the presence of uni-
dentified pycnogonids associated with banks of Mytilus edulis platensis.
However, intertidal pycnogonids have never been re-ported on the Uruguayan coast. In this paper, as part of a project dealing with benthic invertebrates biodiversity, two species found in the rocky intertidal of Cerro Verde (Rocha, Uruguay) are reported.
2. Material and Methods
2.1. Study area
Cerro Verde (33° 57’ S and 53° 30’ W) is a rocky cape on the east coast of Uruguay (Figure �). On each side of the cape there are sandy beach arcs that extend for 2-3 km. The coast experiences a semidiurnal tide (range <0.5 m) being the water level influenced mainly by wind conditions (direction and speed). Winds blow south-west during winter and north-east during summer. The rocky
Carranza, A., Borthagaray, AI. and Genzano, GN.
Braz. J. Biol., 67(2A): 631-637, 20072
platforms have a smooth slope, with a width ranging from �5 to 23 m, and are exposed to different degrees of wave action according to its orientation. In these, three zones, following the classical zonation schemes, can be identified: a high intertidal zone dominated by a cyanobacterial film, a middle intertidal zone dominated by barnacles and a low intertidal and shallow subtidal zone characterized by a dense cover of mussels and/or macroalgae.
2.2. Sampling design
Sampling was carried out on intertidal and shallow subtidal (i.e. depth <�.5 m) rocky platforms of the Cerro Verde area during February 2005. Three sampling sites 500 m apart were chosen along the coast: �) exposed; ori-entated S-SW; 2) intermediate; and 3) protected, orientat-ed N-NE. Within each site, we randomly selected points at each dominance zone (mid intertidal, low intertidal and shallow subtidal), separated by 2 to �0 m. In each point we placed one quadrat of 20 x 20 cm (0.04 m2) and collected all macrofauna and algae present. Organisms were fixed and, in the laboratory, identified and counted. Frequency of occurrence was calculated as number of occurrences/total samples for the dominance zone. Specimens were deposited in the collections of LABIC (Departamento de Ciencias Marinas, FCEyN – UNMdP).
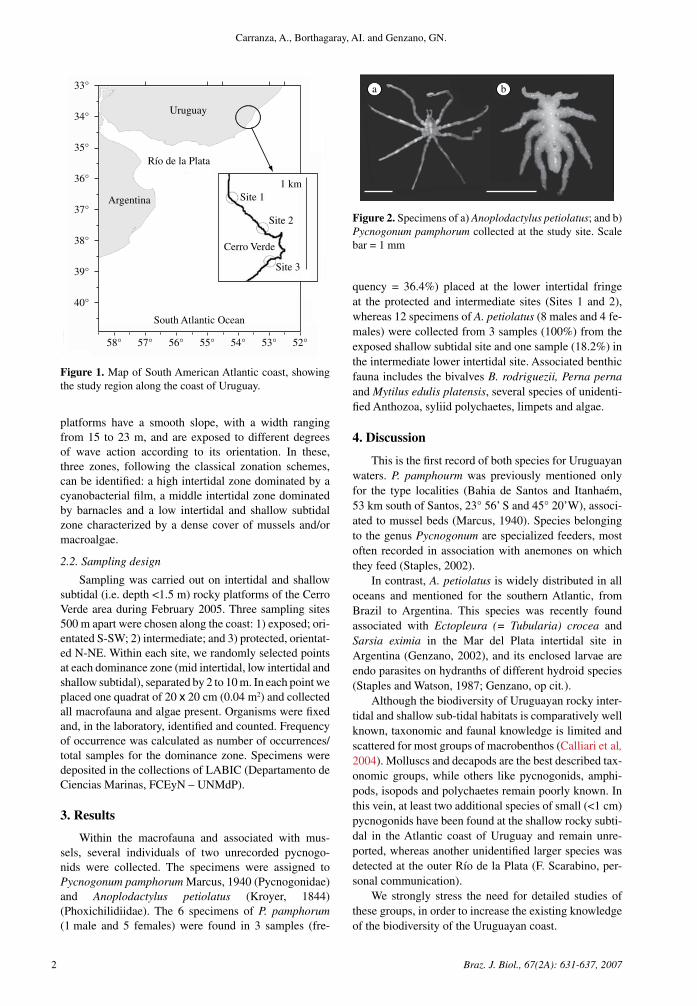
3. Results
Within the macrofauna and associated with mus-sels, several individuals of two unrecorded pycnogo-nids were collected. The specimens were assigned to Pycnogonum pamphorum Marcus, �940 (Pycnogonidae) and Anoplodactylus petiolatus (Kroyer, �844) (Phoxichilidiidae). The 6 specimens of P. pamphorum (� male and 5 females) were found in 3 samples (fre-
quency = 36.4%) placed at the lower intertidal fringe at the protected and intermediate sites (Sites � and 2), whereas �2 specimens of A. petiolatus (8 males and 4 fe-males) were collected from 3 samples (�00%) from the exposed shallow subtidal site and one sample (�8.2%) in the intermediate lower intertidal site. Associated benthic fauna includes the bivalves B. rodriguezii, Perna perna and Mytilus edulis platensis, several species of unidenti-fied Anthozoa, syliid polychaetes, limpets and algae.
4. Discussion
This is the first record of both species for Uruguayan waters. P. pamphourm was previously mentioned only for the type localities (Bahia de Santos and Itanhaém, 53 km south of Santos, 23° 56’ S and 45° 20’W), associ-ated to mussel beds (Marcus, �940). Species belonging to the genus Pycnogonum are specialized feeders, most often recorded in association with anemones on which they feed (Staples, 2002).
In contrast, A. petiolatus is widely distributed in all oceans and mentioned for the southern Atlantic, from Brazil to Argentina. This species was recently found associated with Ectopleura (= Tubularia) crocea and Sarsia eximia in the Mar del Plata intertidal site in Argentina (Genzano, 2002), and its enclosed larvae are endo parasites on hydranths of different hydroid species (Staples and Watson, �987; Genzano, op cit.).
Although the biodiversity of Uruguayan rocky inter-tidal and shallow sub-tidal habitats is comparatively well known, taxonomic and faunal knowledge is limited and scattered for most groups of macrobenthos (Calliari et al, 2004). Molluscs and decapods are the best described tax-onomic groups, while others like pycnogonids, amphi-pods, isopods and polychaetes remain poorly known. In this vein, at least two additional species of small (<� cm) pycnogonids have been found at the shallow rocky subti-dal in the Atlantic coast of Uruguay and remain unre-ported, whereas another unidentified larger species was detected at the outer Río de la Plata (F. Scarabino, per-sonal communication).
We strongly stress the need for detailed studies of these groups, in order to increase the existing knowledge of the biodiversity of the Uruguayan coast.
33°
34°
35°
36°
37°
38°
39°
40°
58° 57° 56° 55° 54° 53° 52°
Uruguay
Río de la Plata
Argentina
1 kmSite 1
Site 2
Site 3
Cerro Verde
South Atlantic Ocean
Figure 1. Map of South American Atlantic coast, showing the study region along the coast of Uruguay.
a b
Figure 2. Specimens of a) Anoplodactylus petiolatus; and b) Pycnogonum pamphorum collected at the study site. Scale bar = � mm

Two new records of pycnogonids for Uruguay
Braz. J. Biol., 67(2A): 631-637, 2007 3
Acknowledgments — The field work was supported by The Maurice Laing Foundation Rufford Small Grant for Nature conservation. A.C. acknowledges financial support from CSIC. A.I.B. thanks the Cleveland Metroparks Zoo for funding received. We also thank F. Scarabino and Marcos Lhano for the valuable comments made on an earlier version of the manuscript.
References
BAIN, BA. Some observations on biology and feeding behavior in two southern California pycnogonids, Bijdr. Dierk., �99�, vol. 6�, no. �, p. 63-64.
CALIARI D., DEFEO O., CERVETO, G., GÓMEZ, M., JIMÉNEZ L., SCARABINO, F., BRAZEIRO, A. and NORBIS, W. Marine life of Uruguay: critical update and priorities for future research. Gayana, 2003, vol. 67, no. 2, p. 34�-370
GENZANO, GN. Associations between pycnogonids and hydroids from the Buenos Aires littoral zone, with observations on the semi-parasitic life cycle of Tanystylum orbiculare (Ammotheidae). Sci. Mar., 2002, vol. 66, no. �, p. 83-92
JUANICÓ, M. and RODRÍGUEZ-MOYANO, M. Composición faunística de la comunidad de Mytilus edulis platensis d´orbigny,
�846 ubicada a unas 55 millas al SE de La Paloma. Com. Soc. Mal. Uruguay, �976, vol. 4, no. 29, p. ��3-��6.
LARAMENDY, ML. Anotaciones sobre Colossendeis geofroyi (Mañe-Garzón, �944). Neotrópica, �974, vol. 20, no. 63, p. �49-�52
MARCUS, E. Os pantopoda brasileiros e os demais sul-americanos. Bol. Fault. Fil., Cien., Letr., �940, vol. �9, no. 4, p. 3-�44.
STAPLES, DA. Pycnogonum (Pycnogonida: Pycnogonidae) from Australia with descriptions of two new species. Mem. Mus. Victoria. 2002, vol. 59, no. 2, p. 54�-553
STAPLES, DA. and WATSON, JE. Associations between pycnogonids and hydroids, In: J. Bouillon, F. Boero, F. Cicogna and P. F. S. Cornelius (eds.), Modern Trends in the Systematics, Ecology, and Evolution of Hydroids and Hydromedusae, Oxford University Press, �987, p. 2�5-226.
STOCK, JH. Résultats scientifiques des campagnes de la “Calypso”. Fascicule VII. Campagne de la Calypso au large des cotes atlantiques de l’Amerique du Sud (�96�-�962). I. 4 Pycnogonida. Ann. Inst. Oceanogr. Monaco, �966, vol. 44, p. 385-406.

ava i lab le at www.sc ienced i rec t . com
journa l homepage : www. e lsev ier . com/ loca te /ac toec
a c t a o e c o l o g i c a 3 1 ( 2 0 0 7 ) 2 4 3 – 2 5 0
Original article
Mussels as ecosystem engineers: Their contribution tospecies richness in a rocky littoral community
Ana Ines Borthagaray*,1, Alvar Carranza1
Investigacion and Desarrollo, Facultad de Ciencias, Igua 4225, CP 11 400, Montevideo, Uruguay
a r t i c l e i n f o
Article history:
Received 23 December 2005
Accepted 17 October 2006
Published online 9 April 2007
Keywords:
Ecosystem engineers
Mussels
Rocky shores
Specific richness
Uruguay
a b s t r a c t
Mussels are important ecosystem engineers in marine benthic systems because they ag-
gregate into beds, thus modifying the nature and complexity of the substrate. In this study,
we evaluated the contribution of mussels (Brachidontes rodriguezii, Mytilus edulis platensis,
and Perna perna) to the benthic species richness of intertidal and shallow subtidal commu-
nities at Cerro Verde (Uruguay). We compared the richness of macro-benthic species be-
tween mussel-engineered patches and patches without mussels but dominated by algae
or barnacles at a landscape scale (all samples), between tidal levels, and between sites dis-
tributed along a wave exposition gradient. Overall, we found a net increase in species rich-
ness in samples with mussels (35 species), in contrast to samples where mussels were
naturally absent or scarce (27 species). The positive trend of the effect did not depend
upon tidal level or wave exposition, but its magnitude varied between sites. Within sites,
a significant positive effect was detected only at the protected site. Within the mussel-
engineered patches, the richness of all macro-faunal groups (total, sessile and mobile)
was positively correlated with mussel abundance. This evidence indicates that the mussel
beds studied here were important in maintaining species richness at the landscape-level,
and highlights that beds of shelled bivalves should not be neglected as conservation targets
in marine benthic environments.
ª 2007 Elsevier Masson SAS. All rights reserved.
1. Introduction
Ecosystem engineering (i.e. the creation, modification and
maintenance of habitats by organisms (Jones et al., 1994) gen-
erates environmental heterogeneity and increases the diver-
sity of habitats at the landscape level (Jones et al., 1997).
Such increases in habitat diversity suggests that ecosystem
engineers can positively affect ecosystem species richness.
However, two conditions must be met to achieve higher
species richness at this spatial scale. First, the engineer spe-
cies must provide conditions not present elsewhere in the
landscape and, second, some species must be able to live
only in the engineered patches (Wright et al., 2002). Only if
the engineer-created patches are sufficiently different from
its surroundings (so that species otherwise excluded from
the landscape can persist) will the addition of an engineer in-
crease species richness via an increase in habitat diversity
(Wright et al., 2002). This newly developed conceptual
* Corresponding author. Tel./fax: þ598 2 5258 61821.E-mail address: [email protected] (A.I. Borthagaray).
1 Both authors contributed equally.1146-609X/$ – see front matter ª 2007 Elsevier Masson SAS. All rights reserved.doi:10.1016/j.actao.2006.10.008

a c t a o e c o l o g i c a 3 1 ( 2 0 0 7 ) 2 4 3 – 2 5 0244
framework is a well-suited tool for management and monitor-
ing issues, since it relates habitat-forming species with pro-
cesses maintaining local and regional biodiversity.
Ecosystem engineers can affect the availability of re-
sources to other organisms either as a direct consequence of
the structure created by them or by the modulation of biotic
or abiotic forces by its structure (Jones et al., 1994, 1997) or
their biological activity (e.g. Commito and Boncavage, 1989).
Shell production and the subsequent creation of habitat by
aquatic molluscs can affect other organisms via three general
mechanisms, namely the provision of substrata for attach-
ment, the provision of refuges to avoid predators or physical
or physiological stress, and the control of the transport of par-
ticles and solutes in the benthic environment (Gutierrez et al.,
2003). Mussels are known to control the above factors and pro-
cesses in marine benthic environments (Frechette et al., 1989;
Crooks and Khim, 1999) suggesting that they can provide other
organisms with unique resources. However, their effects on
the macro-faunal community may depend upon habitat fea-
tures varying along exposure and tidal gradients and with
the spatial scales considered, since a high variability in
the abundance of organisms at spatial scales within and
among shores has been found in several intertidal studies
(Benedetti-Cecchi, 2001a; Benedetti-Cecchi et al., 2001b;
Adami et al., 2004).
Mussel beds are a conspicuous feature of Uruguayan rocky
shores. Brachidontes rodriguezii is the dominant mussel species
in these beds but Mytilus edulis platensis and Perna perna are
also present (Maytia and Scarabino, 1979; Neirotti, 1981).
Although the biodiversity of Uruguayan rocky intertidal
shores is comparatively well known (Caliari et al., 2003), mus-
sel beds have only been studied at the population level, and
specifically in relation to the commercial harvesting of some
species (Riestra et al., 1992). On the other hand, the contribu-
tion of mussels to the structure and species richness of inter-
tidal and subtidal communities has received little attention in
this region. Nevertheless, this is particularly important since
intensive mussel harvesting might result in the loss of other
species relying on critical resources only available at the
mussel-created habitat.
In this paper we assessed the contribution of mussel-
created habitat to the species richness of the benthic intertidal
and shallow subtidal community at a Uruguayan rocky shore.
In particular we quantified differences in macro-benthic spe-
cific richness between mussel-engineered patches (hereafter
namely MEP) and non-mussel-engineered patches (hereafter
namely NMEP) dominated by algae or barnacles and evaluated
the consistency of the engineering effect across environmen-
tal gradients and different spatial scales. Further, we also
focused on how species richness depends upon individual
shell traits and spatial arrangement of shells.
2. Materials and methods
2.1. Study area
Cerro Verde (33�570S, 53�300W) is a rocky cape on the east coast
of Uruguay (Fig. 1) affected by semidiurnal, low-amplitude
tides (range <0.5 m) that are largely controlled by wind
conditions (direction and speed). The rocky platforms have
a smooth slope, with a width ranging from 15 to 23 m, and
are exposed to different degrees of wave action according to
its orientation. These platforms follow a classical zonation
scheme (Stephenson and Stephenson, 1949), in which three
zones can be identified: a high intertidal zone dominated by
a cyanobacterial film; a middle intertidal zone dominated by
barnacles; and a low intertidal and shallow subtidal zone char-
acterized by a dense cover of mussels and/or macro-algae. This
site harbours a rich hard-substrata benthic fauna, a yet unde-
fined number of fish species (e.g. endangered sharks Mustelus
schmitti, M. fasciatus, Sphyrna bigelowi), marine birds, mammals
(Otaria bryonia, Arctocephalus australis) and sea turtles (Chelonya
mydas). It has been proposed as one possible marine protected
area in Uruguay (IUCN Uruguayan Committee, 2002).
2.2. Sampling design
Sampling was carried out on intertidal and shallow subtidal
(i.e. depth <1.5 m) rocky platforms of the Cerro Verde area
during the summer months of 2005 and 2006 to minimize var-
iations due to seasonal changes in climate and sea conditions.
Three sampling sites 500 m apart were chosen along the coast:
(1) wave-exposed; (2) wave-intermediately exposed; and (3)
wave-protected (Fig. 1). Within each site, a variable number
of quadrants of 20 � 20 cm were randomly sampled within
each patch-type (mussel-engineered and non-mussel-
engineered, hereafter MEP and NMEP) and at each one of the
three tidal levels above defined. Not all of the possible combi-
nations of patch type and tidal level were found and the
number of replicates taken within each patch type at a given
Fig. 1 – Map of the South American Atlantic coast, showing
the study region along the coast of Uruguay.

a c t a o e c o l o g i c a 3 1 ( 2 0 0 7 ) 2 4 3 – 2 5 0 245
tidal level varied, but ensured at least a minimum degree of
replication within each condition in order to examine the
main contrasts of interest. Organisms collected were fixed
and identified and counted in the laboratory. In addition, all
the mussels collected were counted, measured (shell length
to the nearest 0.1 mm), oven-dried (40 �C over 48 h) and
weighed to the nearest 0.01 g.
2.3. Data analysis
Each macro-faunal species was assigned to the following cat-
egories according to its occurrence: generalists (present in
both MEP and NMEP); MEP specialists; and NMEP specialists.
Sample-based rarefaction curves were constructed for MEP,
NMEP and total (i.e. landscape) for meaningful standardiza-
tion and comparison of datasets (Gotelli and Entsminger,
2001). Then, we calculated the following parameters in order
to describe and quantify the engineer’s effect: Landscape
Area Engineered (LAE) (calculated as percentage of MEP/total
patches); Relative Habitat Richness (RHR) as engineered
richness/unengineered richness; Landscape Richness En-
hancement (LRE) as [Engineered specialists / (Unengineered
specialists þ Generalists)] � 100]; Landscape Insurance Poten-
tial (LIP) as percentage of generalists; and Habitat Rescue Po-
tential (HRP) as percentage of generalist species whose mean
abundance (density in patch) and incidence (number of
occurrences) was at least two times greater in engineered
patches than in unengineered patches. Statistical significance
of differences in abundance were assessed by means of
Kolmogorov–Smirnov two sample test for independent
samples ( p < 0.05).
The overall effect of patch type on macro-faunal species
richness was assessed by means of a Student’s t-test for inde-
pendent samples ( p < 0.05). The consistency of the engineer
effect along the exposure and tidal gradient was evaluated
by means of the significance of the interaction term in two
separated two-way analysis of variance (ANOVA). Factors for
the first analysis were Site (fixed, three levels) and Patch
type (fixed, two levels), while Tidal Level (fixed, three levels)
and Patch were used for a second analysis. Further, the effects
of Tidal Level, Patch, and Tidal Level � Patch interaction were
evaluated within each Site. Cochran’s C-test was used to
check the assumption of homogeneity of variances and,
when necessary, data were log-transformed to remove het-
erogeneous variances. In cases where homogeneity was not
achieved, we set the critical level to a value equal to the
p-value for variance homogeneity (Underwood, 1997). All anal-
yses were done separately for each one the three faunal
groups (total, sessile and mobile species).
Regression analyses were used to evaluate if species rich-
ness was correlated with mussel density and the mean and
standard deviation of mussel shell length and dry weight at
each sampling quadrant. In addition, we examined the corre-
lations between the abundance of mussels and shell traits in
order to remove density-dependent effects on these variables.
In all cases, possible non-linear responses of the independent
variables were investigated by means of the examination of
the significance of the second-order coefficient of a fitted poly-
nomial function. Otherwise, a linear function was adjusted.
Regression analysis was also done separately for total, sessile
and mobile macro-fauna. Log transformed data (both depen-
dent and independent variables) were used for the regression
analysis due to heterocedasticity.
3. Results
A total of 37 species (or operative taxonomic units) of benthic
invertebrates, distributed in 7 major taxa were found in
the 59 quadrants sampled. These were: 16 crustaceans, 9
molluscs, 3 cnidarians, 4 polychaetes, 2 pycnogoniids, a
nemertean, a ophiuroidean and a platyhelminthe (Table 1).
In addition, three mussel species were present in the assem-
blage: Brachidontes rodriguezii, Perna perna and Mytilus edulis
platensis. Another mytilid, Modiolus carvalhoi, was present as
a single specimen in one quadrant, and considered as a
macro-faunal species. From the analysed samples, 37 were
classified as MEP and 22 as NMEP.
Within the samples more than half of the total sampled
area was engineered (61%), while the maximum Relative Hab-
itat Richness was 1.29. We also found that 10 species were
added to the landscape by the engineer (Landscape Richness
Enhancement, 37%). Generalist species (species present in
the combined engineered and unengineered patches) repre-
sented 67.57% of total species (i.e. Landscape Insurance Poten-
tial). Of these species, 84% showed at least double the
incidence of engineered patches (Habitat Rescue Potential).
Also, 46% of the generalist species showed a mean increase
in abundance from NMEP to MEP, but only 7 species showed
statistically significant differences (Table 1).
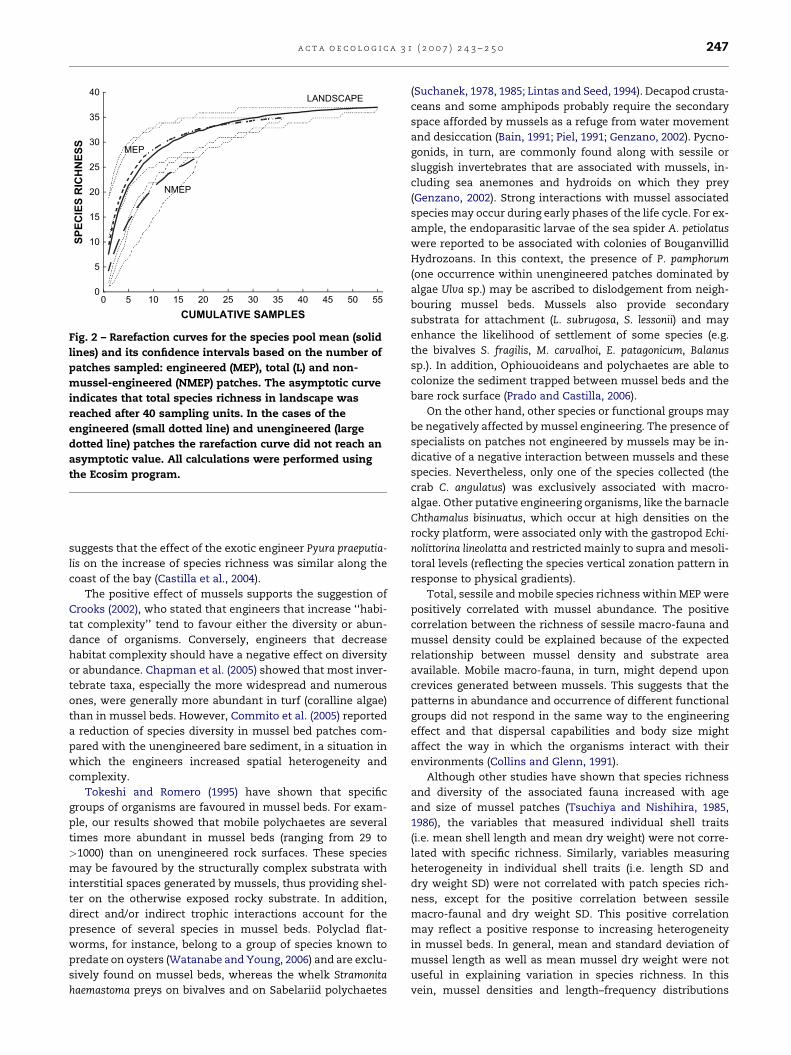
Rarefaction curves showed that total species richness
reached the asymptotic maximum after approximately 40
sampling units (Fig. 2) while species richness at MEP did the
same after 27 samples. However, NMEP did not reach an as-
ymptotic value. The total (landscape) curve lay above MEP
and NMEP curves on all the scales, with the latter displaying
the lowest values across the scales. However, there were no
significant differences among the species richness curves, as
shown by the overlapping of the curves’ 95% confidence inter-
vals (Fig. 2).
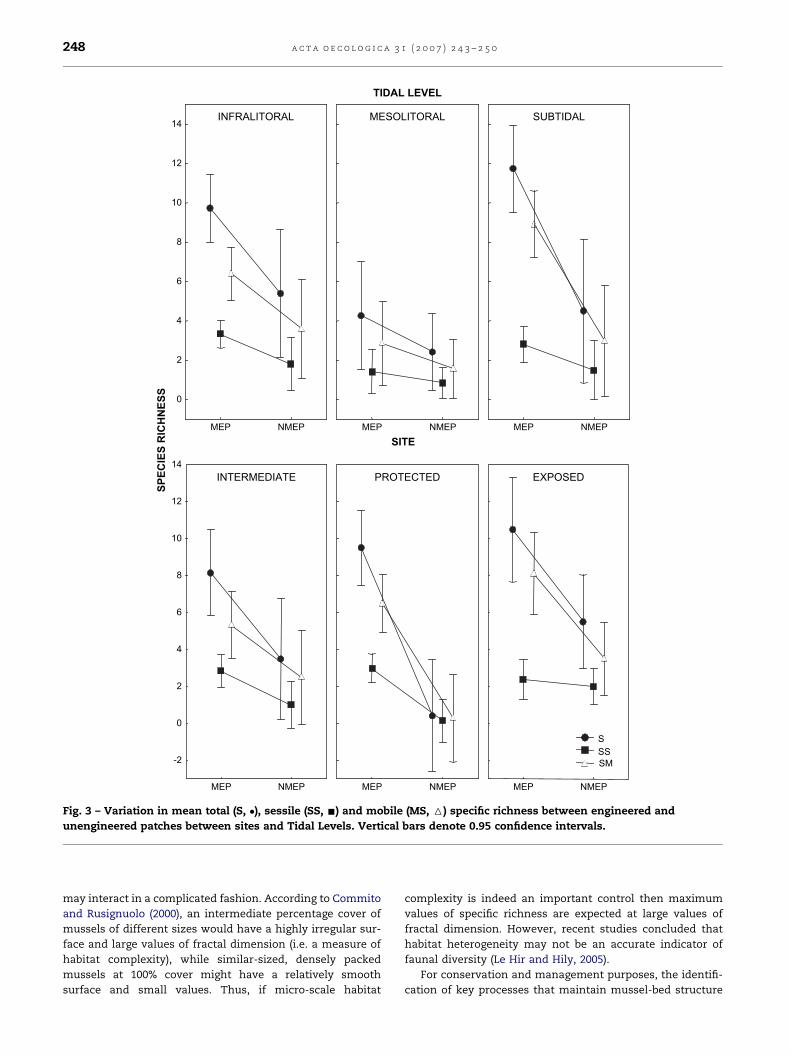
Also, at the landscape scale the total species richness was
significantly higher at MEP compared with NMEP (t(1,57) ¼ 5.25,
p < 0.01). Mussel-engineered patches also showed a signifi-
cantly higher richness of sessile (t(1,57) ¼ 3.88, p < 0.01) and
mobile (t(1,57) ¼ 4.88, p < 0.01) macro-fauna. A significant
Patch � Site interaction was found for total (ANOVA;
F(2,53) ¼ 11.255, p < 0.05), sessile (F(2,53) ¼ 6.20, p < 0.05) and
mobile (F(2,53) ¼ 5.04, p < 0.05) species richness, while Level �Patch interactions were not significant (Fig. 3).
At the site scale in the protected site, a significant patch ef-
fect was detected for all three faunal groups [total (S), sessile
(SS) and mobile (MS) specific richness], but Tidal Level affected
only sessile (F(2,17) ¼ 6.7557, p < 0.05) and total (F(2,17) ¼ 4.6116,
p < 0.05) macro-fauna. At the Exposed site, the effect of tidal
level was significant for Total (F(2,12) ¼ 8.1336, p < 0.05) and
mobile species (F(2,12) ¼ 4.2866, p < 0.05); no patch effects
were detected. Patch effects within the intermediately ex-
posed site could not be estimated due to insufficient samples.
The richness of total (regression analysis, r2 ¼ 0.44,
p < 0.05), mobile (r2 ¼ 0.34, p < 0.05) and sessile macro-faunal

a c t a o e c o l o g i c a 3 1 ( 2 0 0 7 ) 2 4 3 – 2 5 0246
Table 1 – Classification of macro-invertebrate species or operative taxonomic units (OTUs) according to their occurrencesand motility
Occurrence Species OTUs Major taxa
G Cymadusa sp. Amphipoda Motile
G Hyale sp. Amphipoda Motile
G Jassa sp. Amphipoda Motile
G Caprella pennantis Amphipoda Motile
MEP Ampithoe sp. Amphipoda Motile
G Ostreola equestris Bivalvia Sessile
G Modiolus carvalhoi Bivalvia Sessile
MEP Sphenia fragilis Bivalvia Sessile
MEP Entodesma patagonicum Bivalvia Sessile
G Actinia bermudensis Cnidaria Sessile
G Actiniaria spp.* Cnidaria Sessile
MEP Bunodosoma cangicum Cnidaria Sessile
G Balanus improvisus* Cyrripedia Sessile
G Chtamalus bisinuatus Cyrripedia Sessile
G Pachycheles haigae Decapoda Motile
G Cyrtograpsus altimanus Decapoda Motile
G Sesarma ? sp. Decapoda Motile
G Pannopeidae indet. Decapoda Motile
MEP Pilumnus reticulatus Decapoda Motile
NMEP Cyrtograpsus angulatus Decapoda Motile
MEP Amphipholis squamata Echinoidermata Motile
G Siphonaria lesonii Gastropoda Sessile
G Lottia subrugosa* Gastropoda Sessile
MEP Costaoanachis sertulariarum Gastropoda Motile
MEP Stramonita haemastoma Gastropoda Motile
NMEP Echinolittorina lineolata Gastropoda Motile
G Idothea baltica Isopoda Motile
MEP Synidothea marplatensis Isopoda Motile
G Lineus rubens* Nemertea Motile
G Alita succinea Polychaeta Motile
G Halodsynella sp.* Polychaeta Motile
G Syllidae indet.* Polychaeta Motile
G Phragmatopoma sp. Polychaeta Sessile
G Pycnogonum pamphorum* Pycnogonida Motile
MEP Anoplodactylus petiolatus Pycnogonida Motile
G Polycladida indet. Plathelminthes Motile
G Tanaidacea indet. Tanaidacea Motile
Occurrence: (MEP) inhabits only engineered patches; (NMEP) inhabits only rocky substrata not engineered by mussels; (G) inhabits both
(generalist). Motility: (1) mobile able to change spatial location; and (2) sessile are attached to primary or secondary substrata (included vagile
species, i.e. with very low mobility). Species with statistically significant differences in abundance (Kolmogorov–Smirnov two sample test for
independent samples; p < 0.05) between habitat types are denoted with (*).
species (r2 ¼ 0.33, p < 0.05) were positively correlated with
mussel abundance. Sessile macro-faunal specific richness
was positively correlated with the standard deviation of mus-
sel dry weight (r2 ¼ 0.17; p ¼ 0.01). Mean and standard devia-
tion of mussel length and weight were not correlated with
mussel density.
4. Discussion
Our study demonstrated that at a landscape scale, species
richness is increased by the engineering activity of mussel
species, producing shells that introduce complexity into ben-
thic communities. The positive effect of mussel beds in
macro-faunal species richness did not depend upon site and
tidal level. This increase in species richness at the landscape
scale reflects a significant addition of new species to the
intertidal rocky community that otherwise would remain ex-
cluded. However, this effect seems to be scale-dependent,
since differences in mean richness between MEP and NMEP
within sites were not significant in all cases. The same fact
can be observed in the rarefaction curves, where confidence
intervals overlapped at small spatial scales.
We also found a positive effect on the abundances of 60% of
generalist taxa, which showed higher abundances in mussel
beds than in other patches. The same result has been found
in several studies dealing with the positive effect of mussel
mats, but most are restricted to comparisons between bare
soft sediment and mussel beds (Ragnarsson and Raffaelli,
1999; Commito et al., 2005, 2006). Our results are likely to be
extrapolated to other neighbouring rocky shores with similar
characteristics. In this vein, Cerda and Castilla (2001) reported
that macro-invertebrate diversity did not show differences be-
tween sites at a local scale in Antofagasta Bay (Chile), which
a c t a o e c o l o g i c a 3 1 ( 2 0 0 7 ) 2 4 3 – 2 5 0 247
suggests that the effect of the exotic engineer Pyura praeputia-
lis on the increase of species richness was similar along the
coast of the bay (Castilla et al., 2004).
The positive effect of mussels supports the suggestion of
Crooks (2002), who stated that engineers that increase ‘‘habi-
tat complexity’’ tend to favour either the diversity or abun-
dance of organisms. Conversely, engineers that decrease
habitat complexity should have a negative effect on diversity
or abundance. Chapman et al. (2005) showed that most inver-
tebrate taxa, especially the more widespread and numerous
ones, were generally more abundant in turf (coralline algae)
than in mussel beds. However, Commito et al. (2005) reported
a reduction of species diversity in mussel bed patches com-
pared with the unengineered bare sediment, in a situation in
which the engineers increased spatial heterogeneity and
complexity.
Tokeshi and Romero (1995) have shown that specific
groups of organisms are favoured in mussel beds. For exam-
ple, our results showed that mobile polychaetes are several
times more abundant in mussel beds (ranging from 29 to
>1000) than on unengineered rock surfaces. These species
may be favoured by the structurally complex substrata with
interstitial spaces generated by mussels, thus providing shel-
ter on the otherwise exposed rocky substrate. In addition,
direct and/or indirect trophic interactions account for the
presence of several species in mussel beds. Polyclad flat-
worms, for instance, belong to a group of species known to
predate on oysters (Watanabe and Young, 2006) and are exclu-
sively found on mussel beds, whereas the whelk Stramonita
haemastoma preys on bivalves and on Sabelariid polychaetes
NMEP
MEP
LANDSCAPE
0 5 10 15 20 25 30 35 40 45 50 55CUMULATIVE SAMPLES
0
5
10
15
20
25
30
35
40S
PE
CIE
S R
IC
HN
ES
S
Fig. 2 – Rarefaction curves for the species pool mean (solid
lines) and its confidence intervals based on the number of
patches sampled: engineered (MEP), total (L) and non-
mussel-engineered (NMEP) patches. The asymptotic curve
indicates that total species richness in landscape was
reached after 40 sampling units. In the cases of the
engineered (small dotted line) and unengineered (large
dotted line) patches the rarefaction curve did not reach an
asymptotic value. All calculations were performed using
the Ecosim program.
(Suchanek, 1978, 1985; Lintas and Seed, 1994). Decapod crusta-
ceans and some amphipods probably require the secondary
space afforded by mussels as a refuge from water movement
and desiccation (Bain, 1991; Piel, 1991; Genzano, 2002). Pycno-
gonids, in turn, are commonly found along with sessile or
sluggish invertebrates that are associated with mussels, in-
cluding sea anemones and hydroids on which they prey
(Genzano, 2002). Strong interactions with mussel associated
species may occur during early phases of the life cycle. For ex-
ample, the endoparasitic larvae of the sea spider A. petiolatus
were reported to be associated with colonies of Bouganvillid
Hydrozoans. In this context, the presence of P. pamphorum
(one occurrence within unengineered patches dominated by
algae Ulva sp.) may be ascribed to dislodgement from neigh-
bouring mussel beds. Mussels also provide secondary
substrata for attachment (L. subrugosa, S. lessonii) and may
enhance the likelihood of settlement of some species (e.g.
the bivalves S. fragilis, M. carvalhoi, E. patagonicum, Balanus
sp.). In addition, Ophiouoideans and polychaetes are able to
colonize the sediment trapped between mussel beds and the
bare rock surface (Prado and Castilla, 2006).
On the other hand, other species or functional groups may
be negatively affected by mussel engineering. The presence of
specialists on patches not engineered by mussels may be in-
dicative of a negative interaction between mussels and these
species. Nevertheless, only one of the species collected (the
crab C. angulatus) was exclusively associated with macro-
algae. Other putative engineering organisms, like the barnacle
Chthamalus bisinuatus, which occur at high densities on the
rocky platform, were associated only with the gastropod Echi-
nolittorina lineolatta and restricted mainly to supra and mesoli-
toral levels (reflecting the species vertical zonation pattern in
response to physical gradients).
Total, sessile and mobile species richness within MEP were
positively correlated with mussel abundance. The positive
correlation between the richness of sessile macro-fauna and
mussel density could be explained because of the expected
relationship between mussel density and substrate area
available. Mobile macro-fauna, in turn, might depend upon
crevices generated between mussels. This suggests that the
patterns in abundance and occurrence of different functional
groups did not respond in the same way to the engineering
effect and that dispersal capabilities and body size might
affect the way in which the organisms interact with their
environments (Collins and Glenn, 1991).
Although other studies have shown that species richness
and diversity of the associated fauna increased with age
and size of mussel patches (Tsuchiya and Nishihira, 1985,
1986), the variables that measured individual shell traits
(i.e. mean shell length and mean dry weight) were not corre-
lated with specific richness. Similarly, variables measuring
heterogeneity in individual shell traits (i.e. length SD and
dry weight SD) were not correlated with patch species rich-
ness, except for the positive correlation between sessile
macro-faunal and dry weight SD. This positive correlation
may reflect a positive response to increasing heterogeneity
in mussel beds. In general, mean and standard deviation of
mussel length as well as mean mussel dry weight were not
useful in explaining variation in species richness. In this
vein, mussel densities and length–frequency distributions
a c t a o e c o l o g i c a 3 1 ( 2 0 0 7 ) 2 4 3 – 2 5 0248
SP
EC
IE
S R
IC
HN
ES
S
TIDAL LEVEL
MEP NMEP
8
10
12
14INFRALITORAL
MEP
MESOLITORAL
NMEP MEP NMEP
0
2
4
6
SUBTIDAL
SITE
MEP NMEP
-2
0
2
4
6
8
10
12
14INTERMEDIATE
MEP
PROTECTED
NMEP MEP NMEP
EXPOSED
SSSSM
Fig. 3 – Variation in mean total (S, �), sessile (SS, -) and mobile (MS, 6) specific richness between engineered and
unengineered patches between sites and Tidal Levels. Vertical bars denote 0.95 confidence intervals.
may interact in a complicated fashion. According to Commito
and Rusignuolo (2000), an intermediate percentage cover of
mussels of different sizes would have a highly irregular sur-
face and large values of fractal dimension (i.e. a measure of
habitat complexity), while similar-sized, densely packed
mussels at 100% cover might have a relatively smooth
surface and small values. Thus, if micro-scale habitat
complexity is indeed an important control then maximum
values of specific richness are expected at large values of
fractal dimension. However, recent studies concluded that
habitat heterogeneity may not be an accurate indicator of
faunal diversity (Le Hir and Hily, 2005).
For conservation and management purposes, the identifi-
cation of key processes that maintain mussel-bed structure

a c t a o e c o l o g i c a 3 1 ( 2 0 0 7 ) 2 4 3 – 2 5 0 249
are of outmost importance, since these structures control the
local richness of benthic species. The mussel beds studied
here are key structures that add species to the landscape
and it is therefore vital that shelled bivalves are not neglected
as conservation targets. Quantitative research on the relation-
ship between different community traits (species richness
and composition, abundance distribution) and environmental
factors (exposure, habitat complexity, energy input, seascape
configuration, pollution) is needed to understand community
structure and to ensure a proper management of protected
areas.
Acknowledgements
We acknowledge financial support from the Maurice Lang
Foundation through the Rufford Small Grants for Nature Con-
servation. We extend our gratitude to J. Gutierrez for his great
help with an earlier version of the manuscript. A.I.B. thanks
the Cleveland Metroparks Zoo for funding received. A.C. ac-
knowledges CSIC and PEDECIBA for financial support. Marina
and Estela are acknowledged for their support. We also thank
our colleagues D. Caliari, P. Muniz, L. Gimenez, M. Sarassola,
M. Arim, V. Korenko, G. Laufer, C.F. Fagundez, and E. Grosso
from Facultad de Ciencias.
r e f e r e n c e s
Adami, M.L., Tablado, A., Lopez Gappa, J., 2004. Spatial andtemporal variability in intertidal assemblages dominated bythe mussel Brachidontes rodriguezii (d’Orbigny, 1846).Hydrobiologia 520, 49–59.
Bain, B.A., 1991. Some observations on biology and feeding behaviorin two southern California pycnogonids. Bijdr. Dierk. 61, 63–64.
Benedetti-Cecchi, L., 2001a. Variability in abundance of algae andinvertebrates at different spatial scales on rocky sea shores.Mar. Ecol. Prog. Ser. 215, 79.
Benedetti-Cecchi, L., Bulleri, F., Acunto, S., Cinelli, F., 2001b.Scales of variation in the effects of limpets on rocky shores inthe northwest Mediterranean. Mar. Ecol. Prog. Ser. 209, 131.
Caliari, D., Defeo, O., Cerveto, G., Gomez, M., Jimenez, L.,Scarabino, F., Brazeiro, A., Norbis, W., 2003. Marine life ofUruguay: critical update and priorities for future research.Gayana 67, 341–370.
Castilla, J.C., Lagos, N.A., Cerda, M., 2004. Marine ecosystemengineering by the alien ascidian Pyura praeputialis on amid-intertidal rocky shore. Mar. Ecol. Prog. Ser. 268, 119–130.
Cerda, M., Castilla, J.C., 2001. Diversidad y biomasa demacroinvertebrados en matrices intermareales del tunicadoPyura praeputialis (Heller, 1878) en la Bahıa de Antofagasta,Chile. Rev. Chil. Hist. Nat. 74, 841–853.
Collins, S.L., Glenn, S.M., 1991. Importance of spatial andtemporal dynamics in species regional abundance anddistribution. Ecology 72, 654–664.
Commito, J.A., Boncavage, E.M., 1989. Suspension-feeders andcoexisting infauna: an enhancement counterexample. J. Exp.Mar. Biol. Ecol. 125, 33.
Commito, J.A., Celano, E.A., Celico, H.J., Como, S., Johnson, C.P.,2005. Mussels matter: Postlarval dispersal dynamics alteredby a spatially complex ecosystem engineer. J. Exp. Mar. Biol.Ecol. 316, 133–147.
Commito, J.A., Dow, W.E., Grupe, B.M., 2006. Hierarchycal spatialstructure in soft bottom mussel beds. J. Exp. Mar. Biol. Ecol.330, 27–37.
Commito, J.A., Rusignuolo, B.R., 2000. Structural complexity inmussel beds: the fractal geometry of surface topography.J. Exp. Mar. Biol. Ecol. 255, 133–152.
Crooks, J.A., 2002. Characterizing ecosystem-level consequencesof biological invasions: the role of ecosystem engineers.Oikos 97, 153–166.
Crooks, J.A., Khim, H.S., 1999. Architectural vs. biologicaleffects of a habitat altering, exotic mussel, Musculistasenhousia. J. Exp. Mar. Biol. Ecol. 240, 53–75.
Chapman, M.G., People, J., Blokley, D., 2005. Intertidalassemblages associated with natural corallina turf andinvasive mussel beds. Biodivers. Conserv. 14,1761–1776.
Frechette, M., Butman, C.A., Geyer, W.R., 1989. The importanceof boundary layer flows in supplying phytoplankton to thebenthic suspension feeder, Mytilus edulis. Limnol. Oceanogr.34, 19–36.
Genzano, G.B., 2002. Associations between pycnogonids andhydroids from the Buenos Aires littoral zone, withobservations on the semi-parasitic life cycle of Tanystylumorbiculare (Ammotheiidae). Sci. Mar. 66, 83–92.
Gotelli, N.J., Entsminger, G.L., 2001. EcoSim: Null Models Softwarefor Ecology. Acquired Intelligence Inc. & Kesey-Bear,Burlington, VT.
Gutierrez, J.L., Jones, C.G., Strayer, D.L., Iribarne, O., 2003.Molluscs as ecosystems engineers: the role of the shellproduction in aquatic habitats. Oikos 101, 79–90.
IUCN Uruguayan Committee, 2002. Areas Marinas Protegidas:Un Enfoque Bioregional en el Contexto de Manejo CosteroIntegrado. Facultad de Ciencias, Montevideo, Uruguay.
Jones, C.G., Lawton, J.H., Shachak, M., 1994. Organisms asecosystem engineers. Oikos 69, 373–386.
Jones, C.G., Lawton, J.H., Shachak, M., 1997. Positive andnegative effects of organisms as physical ecosystemengineers. Ecology 78, 1946–1957.
Le Hir, M., Hily, C., 2005. Macrofaunal diversity and habitatstructure in intertidal boulder fields. Biodivers. Conserv. 14,233–250.
Lintas, C., Seed, R., 1994. Spatial variation in the fauna associatedwith Mytilus edulis on a wave-exposed rocky shore.J. Molluscan Stud. 60, 165.
Maytia, S., Scarabino, V., 1979. Las comunidades del litoral rocosodel Uruguay: zonacion, distribucion local y consideracionesbiogeograficas. Memorias del Seminario sobre EcologıaBentonica y Sedimentacion de la Plataforma Continental delAtlantico Sur. UNESCO, pp. 149–160.
Neirotti, E., 1981. Estudio comparativo del supralitoral ymesolitoral rocoso en diferentes localidades del estuario delRıo de la Plata. Comun. Soc. Malacol. Urug. 5, 347–370.
Piel, W.H., 1991. Pycnogonid predation on nudibranchs andceratal autotomy. The Veliger 34, 366.
Prado, L., Castilla, J.C., 2006. The bioengineer Perumytiluspurpuratus (Mollusca: Bivalvia) in central Chile: biodiversity,habitat structural complexity and environmentalheterogeneity. J. Mar. Biol. Assoc. U.K. 86, 417–421.
Ragnarsson, S.A., Raffaelli, D., 1999. Effects of the musselMytilus edulis L. on the invertebrate fauna of sediments. J. Exp.Mar. Biol. Ecol. 241, 31–43.
Riestra, G., Gimenez, J.L., Scarabino, V., 1992. Analisis de lacomunidad macrobentonica infralitoral de fondo rocoso enIsla Gorriti e Isla de Lobos (Maldonado, Uruguay). Pub. Com.Tec. Mix. Fren. Mar. 11, 123–127.
Stephenson, T.A., Stephenson, A., 1949. The universal featuresof zonation between tide-marks on rocky coasts. J. Ecol. 37,289–305.

a c t a o e c o l o g i c a 3 1 ( 2 0 0 7 ) 2 4 3 – 2 5 0250
Suchanek, T.H., 1978. The ecology of Mytilus edulis L. in exposedrocky intertidal communities. J. Exp. Mar. Biol. Ecol. 31, 105.
Suchanek, T.H., 1985. Mussels and their role in structuringrocky shore communities. In: Moore, P.G., Seed, R. (Eds.),Ecology of Rocky Coasts. Hodder and Stoughton, pp. 70–96.
Tokeshi, M., Romero, L., 1995. Filling a gap: dynamics of spaceoccupancy on a mussel-dominated subtropical rocky shore.Mar. Ecol. Prog. Ser. 199, 167–176.
Tsuchiya, M., Nishihira, M., 1985. Islands of Mytilus edulis asa habitat for small intertidal animals: effect of island size oncommunity structure. Mar. Ecol. Prog. Ser. 25, 71–81.
Tsuchiya, M., Nishihira, M., 1986. Islands of Mytilus edulis asa habitat for small intertidal animals: effect of Mytilus age
structure on the species composition of the associated faunaand community organization. Mar. Ecol. Prog. Ser. 31, 171–178.
Underwood, A., 1997. Experiments in Ecology: Their LogicalDesign and Interpretation using Analysis of Variances.Cambridge University Press, Cambridge.
Watanabe, J.T., Young, C.M., 2006. Feeding habits andphenotypic changes in proboscis length in the southernoyster drill, Stramonita haemastoma (Gastropoda: Muricidae),on Florida sabellariid worm reefs. Mar. Biol. 148,1021.
Wright, J.P., Jones, C.G., Flecker, A.S., 2002. An ecosystemengineer, the beaver, increases species richness at thelandscape scale. Oecologia 132, 96–101.

BASES para la CONSERVACIÓNy el MANEJO de la
COSTA URUGUAYA
R. MenafraL. Rodríguez-Gallego
F. ScarabinoD. Conde(editores)

La referencia correcta de este libro es:
Menafra R Rodríguez-Gallego L Scarabino F & D Conde(eds) 2006 Bases para la conservación y el manejo de la costauruguaya. VIDA SILVESTRE URUGUAY, Montevideo.i-xiv+668pp
Armado y diagramación: Javier González
Fotografía de portada: Faro de Cabo Polonio (Rocha)Diego Velazco - Aguaclara Fotostock,www.aguaclara.com.uy
Impreso en GRAPHIS Ltda, en el mes de octubre de 2006Nicaragua 2234, Montevideo, UruguayTels.: 409 6821-409 9168. E-mail: [email protected]ósito legal: 339.537/06
ISBN: 9974-7589-2-0
Las opiniones e informaciones contenidas en este libro sonexclusiva responsabilidad de sus autores, y no reflejannecesariamente aquellas de VIDA SILVESTRE URUGUAY,US Fish and Wildlife Service, Facultad de Ciencias, o de lasinstituciones a las cuales los autores están vinculados.

LISTA DE AUTORES PARTICIPANTES
PRÓLOGOOSCAR IRIBARNE
PRÓLOGOULRICH SEELIGER
PREFACIO
AGRADECIMIENTOS
Mapa general de la costa platense-atlántica
Mapa de la costa Oeste del Río de la Plata
Mapa del sector centro-Sur de la costa platense-atlántica
Mapa del sector Este de la costa atlántica
COSTA PLATENSE-ATLÁNTICA
Evolución paleogeográfica y dispersión de los sedimentos del Río de la PlataRICARDO N. AYUP-ZOUAIN
Geología de la costa uruguaya y sus recursos minerales asociadosCÉSAR A. GOSO AGUILAR & ROSSANA MUZIO
Dinámica y fuentes de sedimentos de las playas uruguayasDANIEL PANARIO & OFELIA GUTIÉRREZ
Geomorfología y procesos erosivos en la costa atlántica uruguayaMARÍA ALEJANDRA GÓMEZ PIVEL
Fitoplancton de la zona costera uruguaya: Río de la Plata y Océano AtlánticoGRACIELA FERRARI & LETICIA VIDAL
El impacto de las floraciones algales nocivas: origen, dispersión, monitoreo,control y mitigaciónSILVIA M. MÉNDEZ
Flora y vegetación de la costa platense y atlántica uruguayaEDUARDO ALONSO-PAZ & MARÍA JULIA BASSAGODA
Fauna parasitaria del lobo fino Arctocephalus australis y del león marino Otariaflavescens (Mammalia, Otariidae) en la costa uruguayaDIANA MORGADES, HELENA KATZ, OSCAR CASTRO, DINORA CAPELLINO, LOURDES CASAS,GUSTAVO BENÍTEZ, JOSÉ MANUEL VENZAL & ANTONIO MORAÑA
Zooplancton gelatinoso de la costa uruguayaMARÍA GABRIELA FAILLA SIQUIER
Zooplancton de ambientes costeros de Uruguay: añadiendo piezas alrompecabezasGUILLERMO CERVETTO, DANILO CALLIARI, LAURA RODRÍGUEZ-GRAÑA, GISSELL LACEROT
& RAFAEL CASTIGLIONI
Faunística y taxonomía de invertebrados bentónicos marinos y estuarinos dela costa uruguayaFABRIZIO SCARABINO
Índice
........................................................................................................ i
.............................................................................................................................................................. v
........................................................................................................................................................... vii
............................................................................................................................................................... viii
........................................................................................................................................... x
................................................................................................. xi
................................................................................................... xii
..................................................................... xiii
..................................................................................................... xiv
............................................................................................................................................ 1
............................................................................................................... 9
........................................................................................................................ 21
.................................................................................................................................. 35
............................................................................................................................. 45
....................................................................................................................................................... 57
................................................................................................... 71
............................................................................... 89
............................................................................................................................................... 105
.................................................................................................................................. 97
................................................................................................................................................... 113

Gasterópodos marinos y estuarinos de la costa uruguaya: faunística,distribución, taxonomía y conservaciónFABRIZIO SCARABINO, JUAN CARLOS ZAFFARONI, ALVAR CARRANZA, CRISTHIAN CLAVIJO
& MARIANA NIN
Bivalvos marinos y estuarinos de la costa uruguaya: faunística, distribución,taxonomía y conservaciónFABRIZIO SCARABINO, JUAN CARLOS ZAFFARONI, CRISTHIAN CLAVIJO, ALVAR CARRANZA
& MARIANA NIN
Patrones geográficos de diversidad bentónica en el litoral rocoso de UruguayALEJANDRO BRAZEIRO, ANA INÉS BORTHAGARAY & LUIS GIMÉNEZ
Comunidades bentónicas estuarinas de la costa uruguayaLUÍS GIMÉNEZ
Asociaciones de moluscos bentónicos cuaternarios en la costa uruguaya:implicancias paleoecológicasSERGIO MARTÍNEZ &ALEJANDRA ROJAS
Los recursos pesqueros de la costa de Uruguay: ambiente, biología y gestiónWALTER NORBIS, LAURA PAESCH & OSCAR GALLI
Áreas de cría de peces en la costa uruguayaSUSANA RETTA, GUSTAVO MARTÍNEZ & ADRIANA ERREA
Características biológicas de la corvina (Micropogonias furnieri) en el Río de laPlata y su Frente MarítimoERNESTO CHIESA, OSCAR D. PIN & PABLO PUIG
Abundancia, capturas y medidas de manejo del recurso corvina(Micropogonias furnieri) en el Río de la Plata y Zona Común de PescaArgentino-Uruguaya (1975-2003)OSCAR D. PIN, GUILLERMO ARENA, ERNESTO CHIESA & PABLO PUIG
Herpetofauna de la costa uruguayaRAÚL MANEYRO & SANTIAGO CARREIRA
Biología, ecología y etología de las tortugas marinas en la zona costerauruguayaMILAGROS LÓPEZ-MENDILAHARSU, ANDRÉS ESTRADES, MARÍA NOEL CARACCIO,VICTORIA CALVO, MARTÍN HERNÁNDEZ & VERÓNICA QUIRICI
Conservación y manejo de tortugas marinas en la zona costera uruguayaMARTÍN LAPORTA, PHILIP MILLER, MARIANA RÍOS, CECILIA LEZAMA, ANTONIA BAUZÁ, ANITA
AISENBERG, MARÍA VICTORIA PASTORINO & ALEJANDRO FALLABRINO
Aves de la costa sur y este uruguaya: composición de especies en los distintosambientes y su estado de conservaciónJOAQUÍN ALDABE, SEBASTIÁN JIMÉNEZ & JAVIER LENZI
La franciscana Pontoporia blainvillei (Cetacea, Pontoporiidae) en la costauruguaya: estudios regionales y perspectivas para su conservaciónCAROLINA ABUD, CATERINA DIMITRIADIS, PAULA LAPORTA &MARILA LÁZARO
Revisión preliminar de registros de varamientos de cetáceos en la costauruguaya de 1934 a 2005DANIEL DEL BENE, VIRGINIA LITTLE, RICARDO ROSSI & ALFREDO LE BAS
......................................................................................................................................................... 143
.............................................................................................................................................................. 179
...................................................................................................................... 189
...................................................................................................... 197
.......................................................................................... 211
......................................................................................................................................................... 157
........................................................................... 171
....................................................................................................... 219
...................................................................... 225
................................................................................................................... 233
................................................................................. 247
........................................................................ 259
............................................................................................... 271
.......................................................... 289
.................................................................. 297

Distribución, reproducción y alimentación del lobo fino Arctocephalus australisy del león marino Otaria flavescens en UruguayALBERTO PONCE DE LEÓN & OSCAR D. PIN
Tuberculosis en pinnípedos (Arctocephalus australis y Otaria flavescens) deUruguayMIGUEL CASTRO RAMOS, HELENA KATZ, ANTONIO MORAÑA, MARÍA INÉS TISCORNIA,DIANA MORGADES & OSCAR CASTRO
Interacciones entre lobos marinos y pesca artesanal en la costa de UruguayDIANA SZTEREN & CECILIA LEZAMA
Mamíferos terrestres no voladores de la zona costera uruguayaENRIQUE M. GONZÁLEZ
Vertebrados fósiles de la costa uruguayaANDRÉS RINDERKNECHT
Especies acuáticas exóticas en Uruguay: situación, problemática y manejoERNESTO BRUGNOLI, JUAN CLEMENTE, GUSTAVO RIESTRA, LUCÍA BOCCARDI
& ANA INÉS BORTHAGARAY
Ecología de playas arenosas de la costa uruguaya: una revisión de 25 años deinvestigaciónOMAR DEFEO, DIEGO LERCARI, ANITA DE ÁLAVA, JULIO GÓMEZ, GASTÓN MARTÍNEZ, ELEONORA
CELENTANO, JUAN PABLO LOZOYA, SEBASTIÁN SAUCO, DANIEL CARRIZO & ESTELA DELGADO
Estado actual, propuestas y perspectivas de manejo de las Áreas ProtegidasCosterasJUAN CARLOS GAMBAROTTA
Bases ecológicas y metodológicas para el diseño de un Sistema Nacional deÁreas Marinas Protegidas en UruguayALEJANDRO BRAZEIRO & OMAR DEFEO
COSTA del RÍO de la PLATA
Evolución de la desembocadura del Arroyo Pando (Canelones, Uruguay):¿tendencias naturales o efectos antrópicos?OFELIA GUTIÉRREZ & DANIEL PANARIO
La Bahía de Montevideo: 150 años de modificación de un paisaje costero ysubacuáticoPIERRE GAUTREAU
Monitoreo de cianobacterias en la costa de Montevideo (Uruguay)DANIEL SIENRA & GRACIELA FERRARI
Comunidad componente de tremátodos larvales de Heleobia australis(Mollusca, Cochliopidae) en la costa uruguaya del Río de la PlataOSCAR CASTRO, DANIEL CARNEVIA, ALEJANDRO PERRETTA & JOSÉ MANUEL VENZAL
Composición y ecología de la fauna epígea de Marindia (Canelones, Uruguay)con especial énfasis en las arañas: un estudio de dos años con trampas deintercepciónFERNANDO G. COSTA, MIGUEL SIMÓ & ANITA AISENBERG
Ictioplancton costero de la zona de transición estuarina del Río de la Plata(Uruguay)GABRIELA MANTERO, SUSANA RETTA & MARCELO RODRÍGUEZ
............................................................................................................... 305
......................................................................................................................... 315
........................................................................................................................... 321
............................................................................................................................................. 329
.............................................................................................................................................. 343
......................................................................................................................................... 351
...................................363
........................................................................................................................................ 371
....................................................................................................................... 379
...................................................................................................................... 391
....................................................................................................................................................... 401
......................................................................................................................... 413
.............................................. 421
......................................................................................... 427
.................................................................................. 437

Ecología de un ensamble de anuros en un humedal costero del sudeste deUruguayINÉS DA ROSA, ARLEY CAMARGO, ANDRÉS CANAVERO, DANIEL E. NAYA & RAÚL MANEYRO
Aves de la costa de Montevideo urbano: variación espacial y estacionalMACARENA SARROCA, MATILDE ALFARO, JAVIER LENZI, SEBASTIÁN JIMÉNEZ, CAROLINA ABUD
& DIEGO CABALLERO-SADI
Contaminación de la Bahía de Montevideo y zona costera adyacente y surelación con los organismos bentónicosPABLO MUNIZ, NATALIA VENTURINI & LETICIA BURONE
La pesca artesanal en el Río de la Plata: su presente y una visión de futuroPABLO PUIG
COSTA ATLÁNTICA
Paleolimnología: desarrollo de las lagunas costeras del sudeste de Uruguaydurante el HolocenoFELIPE GARCÍA-RODRÍGUEZ, PETER SPRECHMANN, HUGO INDA, LAURA DEL PUERTO,ROBERTO BRACCO, ADRIANA RODRÍGUEZ, PETER ESTOL & VIRGINIA ACEVEDO
Fisonomía y composición florística de Cabo Polonio (Rocha, Uruguay)SILVANA MASCIADRI, ELOISA FIGUEREDO &LILIANA DELFINO
Estructura y regeneración del Bosque de Ombúes (Phytolacca dioica) de laLaguna de Castillos (Rocha, Uruguay)MARÍA GABRIELA RODRÍGUEZ-GALLEGO
Estructura poblacional y reproducción del tatucito Emerita brasiliensis(Decapoda: Hippidae) en playas de Cabo Polonio (Rocha, Uruguay)ENRIQUE PELUFFO
Invertebrados bentónicos de La Paloma (Rocha, Uruguay)MARIO DEMICHELI & FABRIZIO SCARABINO
Ecología de comunidades de playas de Cabo Polonio (Rocha, Uruguay)LUIS GIMÉNEZ & BEATRIZ YANNICELLI
Transgresiones y regresiones marinas en la costa atlántica y lagunas costerasde Uruguay: efectos sobre los peces continentalesMARCELO LOUREIRO & GRACIELA GARCÍA
Las pesquerías en las lagunas costeras salobres de UruguayGRACIELA FABIANO & ORLANDO SANTANA
La pesca artesanal en la Paloma (Rocha, Uruguay): período 1999-2001ELIZABETH DELFINO, GRACIELA FABIANO & ORLANDO SANTANA
Situación de la administración del recurso lobos y leones marinos en UruguayENRIQUE PÁEZ
Ballena franca (Eubalaeana australis) en la costa atlántica uruguayaMARIANA PIEDRA, PAULA COSTA, PAULA FRANCO FRAGUAS & RAFAEL ÁLVAREZ
Evaluación del turismo de observación de ballenas como una herramientapara la conservación y el manejo de ballena franca austral (Eubalaena australis)RODRIGO GARCÍA & UZI SABAH
................................... 447
......................................................................................................................................... 457
........................................................................................... 467
................................................................................................................................................................. 477
......................................................... 487
.................................................................................... 495
..................................................................................................................... 503
....................................................................................................................................................... 513
................................................................................................................. 523
....................................................................................................................... 535
................................................................................................................. 545
................................................................................................................. 557
............................................................................... 567
............................................................................................................................................................. 577
..................................................... 585
................................................................................................................................. 591

Biodiversidad y calidad de agua de 18 pequeñas lagunas en la costa surestede UruguayCARLA KRUK, LORENA RODRÍGUEZ-GALLEGO, FEDERICO QUINTANS, GISELL LACEROT,FLAVIO SCASSO, NÉSTOR MAZZEO, MARIANA MEERHOFF & JUAN CÉSAR PAGGI
Procesos estructuradores de las comunidades biológicas en lagunas costerasde UruguaySYLVIA BONILLA, DANIEL CONDE, LUIS AUBRIOT, LORENA RODRÍGUEZ-GALLEGO, CLAUDIA PICCINI,ERIKA MEERHOFF, LAURA RODRÍGUEZ-GRAÑA, DANILO CALLIARI, PAOLA GÓMEZ,IRENE MACHADO & ANAMAR BRITOS
Efectos del Canal Andreoni en playas de Rocha: deterioro ambiental y suefecto en la biodiversidadDIEGO LERCARI & OMAR DEFEO
Interfase de conflictos: el sistema costero de Rocha (Uruguay)DANIEL DE ÁLAVA
Importancia de los procesos participativos en la planificación: percepcionesde naturaleza y áreas a proteger en Castillos (Rocha, Uruguay) y su zona deinfluencia costeraDIEGO MARTINO & ANDREA SCHUNK
Aprovechamiento prehistórico de recursos costeros en el litoral atlánticouruguayoHUGO INDA, LAURA DEL PUERTO, CAROLA CASTIÑEIRA, IRINA CAPDEPONT
& FELIPE GARCÍA-RODRÍGUEZ
....................................................... 599
.......................................................................................................................... 611
................................................................................................................................. 631
....................................................................................................................................................... 637
......................................................................................................................... 651
..................................................................................................................................... 661

F SCARABINO, JC ZAFFARONI, C CLAVIJO, A CARRANZA & M NIN
157
Bivalvos marinos y estuarinosde la costa uruguaya:faunística, distribución,taxonomía y conservación
FABRIZIO SCARABINO*, JUAN CARLOS ZAFFARONI, CRISTHIAN
CLAVIJO, ALVAR CARRANZA & MARIANA NIN
RESUMENSe presenta una síntesis y nueva información sobrefaunística, distribución, taxonomía y conservación de lamalacofauna uruguaya de Bivalvia marinos y estuarinoscosteros (0-50 m). La misma está compuesta por más de90 especies, de las cuales solo cuatro son estuarinas (losmitílidos Brachidontes darwinianus, Mytella charruana, elsolecúrtido Tagelus plebeius y el corbuloideo Erodonamactroides) y nueve son marinas eurihalinas. En el Río dela Plata estos mitílidos dominan los fondos consolida-dos mientras que los inconsolidados son dominados porE. mactroides y el máctrido Mactra isabelleana. Elmesodesmátido Mesodesma mactroides y el donácido Donaxhanleyanus son los únicos bivalvos intermareales de lasplayas arenosas atlánticas uruguayas. Aquí los bancosde M. mactroides han colapsado debido a lasobrexplotación y a una mortalidad provocada por unprotozoario. Taxones que contienen especies de talla in-ferior a 15 mm son propuestos como prioridad de inven-tario por su diversidad inexplorada o problemastaxonómicos: Galeommatoidea, Corbulidae, Nuculidae,Lyonsia, Hiatella, Crenella y Abra. La complejidad relativaa las poblaciones de Mytilus requiere asimismo impor-tantes esfuerzos de la caracterización. En el área se ex-plota artesanalmente Mytilus edulis, Perna perna, M.mactroides, D. hanleyanus, T. plebeius y E. mactroides, yPitar rostratus en forma industrial. En todos los casosexisten deficiencias en al menos uno de los siguientesaspectos: conocimiento biológico-pesquero, monitoreo,medidas de manejo y control, indicando la necesidad deenfocar esfuerzos hacia estas actividades. Otras priori-dades de investigación y conservación coinciden con laspropuestas para los gasterópodos de la misma área.
Palabras clave: almejas, ostras, mejillones, Río de la Pla-ta, Uruguay
INTRODUCCIÓNLa fauna uruguaya de Bivalvos marinos y estuarinos
está compuesta por más de 172 especies y subespecies(Scarabino 2003). Este número es inicial dada la canti-dad de material inédito presente en colecciones y las vas-tas zonas y hábitat sin muestrear; varios grupos particu-larmente relevantes o diversos requieren revisionestaxonómicas en forma prioritaria (Scarabino 2003; 2004).
Los bivalvos marinos y estuarinos costeros deUruguay fueron analizados desde un punto de vistamalacológico por una variedad de autores. Sin embargo,
ABSTRACTA synthesis and new data on faunistics, distribution, taxonomyand conservation of the Uruguayan malacofauna of coastalmarine (0-50 m) and estuarine Bivalvia is presented. This iscomposed by more than 90 species, of which only four areestuarine (the mytilids Brachidontes darwinianus, Mytellacharruana, the solecurtid Tagelus plebeius and the corbuloidErodona mactroides) and nine are marine eurihaline. In theRío de la Plata these mytilids dominate the consolidated bot-toms while the unconsolidated ones are dominated by E.mactroides and the mactrid Mactra isabelleana. Muddy At-lantic bottoms are dominated by the nuculid Ennuculauruguayensis, the venerid Pitar rostratus and the corbulidCorbula patagonica. The mesodesmatid Mesodesmamactroides and the donacid Donax hanleyanus are the onlyintertidal bivalves inhabiting the Uruguayan Atlantic sandybeaches. Here the stocks of M. mactroides have colapsed dueto overexploitation and mortality produced by a protozoan.Taxa containing species smaller than 15 mm in size are pro-posed as a priority for the inventory, given their unexploreddiversity or taxonomic problems: Galeommatoidea,Corbulidae, Nuculidae, Lyonsia, Hiatella, Crenella y Abra.The complexity regarding the populations of Mytilus alsorequires strong efforts of research. The following species areexploited in the area (mainly by artisanal gathering): Mytilusedulis, Perna perna, M. mactroides, D. hanleyanus, T. plebeiusy E. mactroides, while Pitar rostratus is exploited by theindustrial fishery. All these resources contain deficiencies inat least one of the following aspects: biological and fisheriesknowledge, monitoring, management and control measures,indicating the need to focus efforts on these activities. Otherresearch priorities and implications for conservation coincidewith those proposed for gastropods in the same area.
Key words: clams, oysters, mussels, Río de la Plata,Uruguay
R MENAFRA, L RODRÍGUEZ-GALLEGO, F SCARABINO & D CONDE (eds) 2006Bases para la conservación y el manejo de la costa uruguaya
VIDA SILVESTREURUGUAY
se destacan por su alcance Barattini & Ureta (1961) yFigueiras & Sicardi (1968; 1969; 1970; 1971; 1980),quienes compilaron la información taxonómica yfaunística incorporando información original. Sinembargo, aunque no fue explicitado por estos autores,muchos de sus registros se basaron en conchillasconjugadas o valvas, por lo que las referencias al hallazgode una especie en determinada playa no implican que lamisma ocupe ese hábitat en particular. Las limitantesque esto plantea al análisis ecológico de la informaciónexistente fueron mencionadas por Sprechmann (1978).

Bivalvos marinos y estuarinos de la costa uruguaya:faunística, distribución, taxonomía y conservación
158
Por otra parte, Milstein et al. (1976) y Layerle & Scarabino(1984) proporcionaron información fundamental acercade la distribución de los bivalvos del área, principalmen-te en relación al sustrato y la batimetría, y basada enmaterial colectado vivo por rastreo y dragadorespectivamente.
En este trabajo se presenta una puesta a punto delconocimiento sobre faunística, taxonomía y distribuciónde los Bivalvia marinos y estuarinos de la costa urugua-ya (0-50 m), discutiéndose prioridades y perspectivas deinvestigación y conservación. Las limitaciones de estetrabajo son las mismas que las expuestas por Scarabinoet al. (en este volumen).
METODOLOGÍASe consideraron las referencias publicadas (incluyen-
do resúmenes de congresos) desde d´Orbigny (1834-1847), primera obra referida a los bivalvos marinos yestuarinos uruguayos. Tesis inéditas también fueron con-sideradas.
Registros incorrectos o muy dudosos previamentediscutidos no son incluidos aquí, parte de los cuales hansido tratados por Scarabino & Zaffaroni (2004). Se ex-cluyeron Limnoperna fortunei (Mytilidae) y Corbicula spp.(Corbiculidae), especies exóticas dulceacuícolas con ciertatolerancia a la salinidad. La sistemática utilizada es ladetallada por Scarabino (2003).
El material examinado está depositado en las siguien-tes colecciones o consiste en los siguientes materiales (lasabreviaciones colocadas luego de Montevideo correspon-den a las utilizadas en el texto): (1) Museo Nacional deHistoria Natural y Antropología (Montevideo, MNHNM).No ha sido posible analizar detalladamente la totalidaddel material existente ni reanalizar el material estudiadopor Layerle & Scarabino (1984), por la situación actualde la colección; (2) colección LP Barattini, Museo Zooló-gico Municipal “Dámaso A. Larrañaga” (Montevideo,LPB); (3) material obtenido por los primeros autoresdepositado en MNHNM (Montevideo, FS, CC, AC) y (4)colección privada de JC Zaffaroni (Montevideo, JCZ), dela cual material representativo será depositado enMNHNM.
Se consideraron las siguientes divisiones: (1) Río dela Plata (RdlP) y otros ambientes estuariales: costa deMontevideo, Canelones y Maldonado (hasta Punta Ne-gra), así como desembocaduras de arroyos y lagunascosteras, tanto en el RdlP como en la costa atlántica. Seincluyen fondos arenosos, fangosos, rocosos y de fangoconsolidado, 0-10 m. (2) Playas arenosas atlánticas: des-de Punta Negra hasta Barra del Chuy, 0-10 m. (3)Sustratos consolidados atlánticos: Punta Negra a límiteUruguay-Brasil, 0-50 m, incluyendo fondos rocosos, fan-gos consolidados, maderas, bancos someros de mitílidos(BMS) y bancos de Mytilus edulis de profundidad (BMP).(4) Sustratos inconsolidados atlánticos: RdlP exterior alímite Uruguay-Brasil, 10-50 m, existiendo enclaves fan-gosos en zonas más someras que son detalladas.
RESULTADOS Y DISCUSIÓNRdlP y otros ambientes estuariales
Trece especies de bivalvos marinos y estuarinos habi-tan esta zona, de los cuales sólo cuatro son estuarinos yhabitan además las lagunas costeras y desembocadurasde arroyos. Sprechmann (1978) graficó la curva deRemane para la fauna de bivalvos del área, mostrando lacoincidencia con otros estuarios del patrón de distribu-ción y baja diversidad
Sustratos inconsolidadosDos especies son dominantes en estos fondos: Erodona
mactroides (Erodonidae, Fig. 1) y Mactra isabelleana(Mactridae, Fig. 1 y 2) (Carcelles 1941; Figueiras & Sicardi1969; 1970; da Costa 1971; Scarabino et al. 1976;Sprechmann 1978). En el submareal del RdlP, E. mactroidesy M. isabelleana ocupan fondos preferentemente fangosos(FS; Roux & Bremec 1996), coincidiendo ambas especiesen la costa de Montevideo en su amplio rango ecológico ybatimétrico. Hacia el W (desembocadura del Río SantaLucía) E. mactroides resulta dominante, situación inversacon M. isabelleana hacia el E (Costa del Oro) (FS). Lapresencia de esta última en zonas más someras está con-dicionada por la existencia de ambientes más protegidosy con matería orgánica abundante (Scarabino et al. 1976).
La caracterización de la variación observable enMactra de la costa uruguaya no ha sido efectuada y dis-cutida satisfactoriamente. Figueiras & Sicardi (1971)consideran para esta área (incluyendo costa del Río de laPlata) a Mactra petiti, especie descrita en base a materialcolectado en Brasil (d´Orbigny 1846). Aun cuando seconsidera la existencia de ecotipos asociados a distintassalinidades y/o sedimentos (comparar figuras 1 y 2), laconsideración de esta especie nominal para la zona einclusive su sinonimización con M. isabelleana (Aguirre1994) parece prematura.
Erodona mactroides y Tagelus plebeius (Solecurtidae) seencuentran en la desembocadura de Río Santa Lucía y delos arroyos Pando, Solís Chico, Solís Grande, Maldonado,Valizas y Chuy, así como en las lagunas José Ignacio,Garzóny Rocha, donde la primera ocupa una variedadde sustratos (Nión 1979; Bier 1985; Jorcín 1996; 1999;Santana & Fabriano 1999; Muniz & Venturini 2001; Olssonet al. 2003; Passadore & Giménez 2003; Giménez et al.2005; FS; MNHNM). La presencia viviente de T. plebeiusen la costa de Montevideo parecería estar asociada azonas más protegidas (Scarabino et al. 1976).
Erodona mactroides se encuentra también en la Lagu-na del Diario (registrada por d´Orbigny 1846; y presenteaún, P. Etchegaray com. pers.). En esta localidad podríahaber estado presente también T. plebeius cuando la lagu-na se conectaba con la Bahía de Maldonado, de acuerdocon un registro de 1955 de “Barra de la Laguna del Dia-rio” (MNHNM).
Erodona mactroides ha sido explotada comercialmen-te por pescadores artesanales en el Arroyo Solís Grande yen las lagunas de Garzón, Rocha y Castillos (Bier 1985;Santana & Fabiano 1999) mediante arrastre.

F SCARABINO, JC ZAFFARONI, C CLAVIJO, A CARRANZA & M NIN
159
Tagelus plebeius es extraído al menos en las desembo-caduras de los arroyos Pando (Canelones) y Maldonadopara su venta como carnada para pesca (A. Martínezcom. pers.; FS obs. pers.). Se ha registrado su extracciónindividual mediante enterramiento de un gancho fino,
Figura 1. Erodona mactroides, especie dominante en RdlP medio y otrosbiotopos mixohalinos de la costa uruguaya.
Figura 2. Mactra isabelleana; ecotipo de fondos fangosos del RdlP, carac-terizado por su valva alta y de menor tamaño y por su perióstracogrueso cubriendo casi toda la valva.
Figura 3. Mactra isabelleana; ecotipo de fondos arenosos y vinculadosa mayor salinidad, caracterizado por comparativamente más anchay de mayor tamaño y por su perióstraco que cubre solo los márgenesde la valva.
así como la realización de excavaciones importantes dadala profundidad de enterramiento de la especie. Esta últi-ma forma de extracción tendría impacto negativo consi-derable en el hábitat y reclutamiento de la especie.
El concepto de T. plebeius en el área y en toda su áreade distribución debe ser revisado; hay propuestas parala región otras tres especies afines que no han sido ade-cuadamente comparadas desde su descripción: Solecurtusplatensis, Solecurtus mollis y Tagelus poeyi (ver Dall 1901).
Macoma uruguayensis (Tellinidae) y Pitar rostratus(Veneridae) están presentes en la costa de Monevideo yCanelones en forma de ejemplares aislados y poco desa-rrollados (Figueiras & Sicardi 1969; FS; JCZ; F. Ferreyrocom. pers.), siendo especies marinas eurihalinas.
Sustratos consolidadosBrachidontes darwinianus y Mytella charruana (Mytilidae)
ocupan sustratos consolidados en la costa de Montevi-deo, Canelones y Maldonado (hasta Punta Negra)(Martínez y Saez 1872; Klappenbach 1965; Figueiras &Sicardi 1968; Duarte 1971; Scarabino et al. 1976; Maytía& Scarabino 1979; Neirotti 1981; MNHNM), contandocon registros para dentro de la Bahía de Montevideo(Duarte 1971; MNHNM), así como los arroyos Pando,Solís Chico y Solís Grande (MNHMM; Bier 1985;Passadore & Giménez 2003). Mytella charruana posee tam-bién un registro para la Laguna de Rocha (Olsson et al.2003). Ambas especies habitan desde el submareal mássomero hasta una profundidad indeterminada por faltade muestreo.
Mytilus edulis (Mytilidae) es encontrado ocasional-mente en la costa de Montevideo, pero su ocurrencia máspermanente es a partir de la Costa de Oro (Atlántida eIsla de la Tuna) (Barattini & Ureta 1961; Amaro-Padilla1963). En este sentido resulta inusual la cita de Hidalgo(1872) para la costa de Montevideo al mencionarla comoespecie común. Existen registros de ejemplares para Islade Flores e Isla de la Tuna (Canelones) y de bancos paraLas Flores y el área de Piriápolis, incluyendo Punta Colo-rada y Punta Negra (Maldonado) (Amaro 1967).Brachidontes rodriguezii es otro mitílido marino eurihalinopresente en esta zona, con hallazgos hacia el W en LaFloresta, Araminda (Canelones) y Bella Vista(Maldonado) (MNHNM) y prensente el la zona dePiriápolis (Maytía & Scarabino 1979).
Ostrea puelchana (Ostreidae) ha sido detectada alre-dedor de Isla de Flores, inclusive en actividad reproductiva(FS). Información anecótica indica extracción de supues-tos bancos de esta especie en esta localidad.
En San Luis (Canelones), fangos consolidadossubmareales someros se encontraban densamente hora-dados por Petricolaria stellae (Petricolidae) (Pin & Pelufo1983). La reciente observación de este banco durante unabajante (FS 2002) mostró que la mayoría de los ejempla-res se hallaban muertos. Existen también registros paralos balnearios Solís, Bella Vista y Piriápolis (Maldonado)(MNHNM). El desarrollo de esta especie parece estarcondicionada por el avance y retroceso de la arena sobre

Bivalvos marinos y estuarinos de la costa uruguaya:faunística, distribución, taxonomía y conservación
160
el sustrato consolidado que habita. Otros dos bivalvosperforantes marinos son eurihalinos, de acuerdo a regis-tros de Balneario Solís y Bella Vista: Barnea lamellosa yCyrtopleura lanceolata (Pholadidae) (MNHNM).
La ocurrencia de Teredinidae en el Río de la Plata yotros ambientes estuariales ha sido muy escasamenteverificada; Barattini (1951) menciona el hallazgo de Teredonavalis y de Nausitora fusticola (como Bankia brasiliensis) enel RdlP. Calvo (1984) no registró teredínidos en la Bahíade Montevideo. El registro de Teredinidae en la desembo-cadura del Arroyo Maldonado está confirmado por ladestrucción del antiguo puente a causa de estos bivalvos(S. Maytía com. pers.). Calvo (1984) no registróteredínidos en la Bahía de Montevideo.
Playas arenosas atlánticasMesodesma mactroides (Mesodesmatidae) está presen-
te en el intermareal de las playas disipativas (Defeo 1985;González de Baccino 1985) del área de estudio, realizan-do migraciones hacia el submareal somero en los mesesde invierno (Defeo 1985). Los principales bancos de estaespecie (Playa Portezuelo y La Coronilla-Barra del Chuy)han sido sobrexplotados y afectados por una mortan-dad masiva probablemente causada por un protozoarioparásito que impactó a la especie en toda su distribución(Castilla & Defeo 2001; Fiori et al. 2004). Fallas de controlsobre la extracción recreacional y la pesquería artesanal,ambas mínimas por las abudancias actuales, probable-mente afecten la recuperación parcial de estas poblacio-nes. Una síntesis sobre la pesquería de esta especie en lacosta uruguaya fue realizada por Castilla & Defeo (2001).La urbanización de las playas y su consecuente impactoen la dinámica costera (e.g. erosión, anegamiento) proba-blemente estén afectando esta especie (Scarabino 2004).Defeo (1985) mencionó al drenaje de agua dulce prove-niente de la napa freática como uno de los factores deter-minantes de la desaparición del banco de esta especie enPlaya Portezuelo.
Donax hanleyanus (Donacidae) el intermaral de pla-yas de tipo discipativo y reflectivo (Defeo 1985; Gonzálezde Baccino 1985; Defeo et al. 1992; Defeo & de Álava1995). Se ha registrado su reclutamiento puntual en Pla-ya Carrasco (Montevideo) en marzo de 2000 (JCZ). Exis-te extracción recreacional de esta especie en la costa uru-guaya (de Álava & Defeo 1995).
Donax gemmula, habitante del submareal somero(Escofet et al. 1979; FS obs. pers.), fue citada para lacosta uruguaya hasta La Paloma por Figueiras & Sicardi(1980), indicando que es una especie poco frecuente aquí.La búsqueda periódica de micromoluscos en la resaca dela Playa Portezuelo (Maldonado) desde fines de la déca-da de 1970 permitió detectar conchillas (algunas conju-gadas) de esta especie desde 1978 y durante la décadade 1980 (JCZ obs. pers.). Demicheli (1986; 1987a) nodetectó esta especie viviente en esta localidad. En1996-1997 se hallaron algunos ejemplares conjugados con res-tos del animal en Portezuelo y en La Paloma, mientrasque en los años 2000 y 2004 se detectaron numerosos
ejemplares en la resaca de la primera localidad (JCZ obs.pers.). Lo mismo ocurrió durante una mortandad deorganismos bentónicos en la resaca del Puerto de La Pa-loma (2002) (Demicheli & Scarabino en este volumen).Los registros excepcionales de Portezuelo parecen deber-se al desarrollo de esta especie por la escasa descarga delRdlP y la presencia de aguas subtropicales asociadas ala costa (ver Demicheli et al. en prensa). Donax gemmulaha sido hallado en varias oportunidades en el submarealsomero de Barra del Chuy (Defeo com. pers) y su presen-cia aquí parece deberse igualmente a pulsos relaciona-dos con eventos oceanográficos cálidos. No se ha podidoverificar la presencia de Donax sp. registrado por Figueiras& Sicardi (1980) para La Paloma (Rocha).
Solen tehuelchus (Solenidae), Mactra marplatensis(Mactridae), Amiantis purpurata y Tivela zonaria(Veneridae) habitan los fondos submareales de arena finaentre 5 y 10 m, y están presentes también en el enclave dePuerto de La Paloma (Scarabino et al. 1974; Escofet et al.1979; Demicheli & Scarabino en este volumen). Sin em-bargo, S. tehuelchus no es exclusiva de estos fondos(Milstein et al. 1976), mientras que los registros puntua-les de esta especie para Playa Portezuelo y Punta delEste (MNHNM; JCZ) parecen ser excepcionales. Tivelazonaria tiene su límite S de distribución en la mencionadalocalidad de La Paloma. Amiantis purpurata se desarrollóen forma importante dentro de la Bahía de Maldonado almenos durante la década de 1960 (S. Martínez com.pers.), pero actualmente no se encuentra en esta locali-dad. Su presencia al menos ocasional tiene su límite W enPlaya Portezuelo (JCZ; F. Ferreyro com. pers.).
Mactra isabelleana ha sido registrada a partir de 2 m enPlaya Portezuelo (Demicheli 1986; 1987a); habita las pla-yas submarinas en la costa de Maldonado y Rocha (FS).
Sustratos consolidados atlánticosTodas las especies registradas para estos ambientes
han sido verificadas o es muy factible su presencia enmicrosustratos duros sobre fondos de conchilla, areno-sos o fangosos.
Brachidontes rodriguezii (Fig. 4) es el bivalvo dominan-te del intermareal, acompañado por Mytilus edulis y Pernaperna, más típicos del submareal, donde estas especiesforman bancos importantes. La primera especie se ex-tiende hasta por lo menos 15 m, desde Punta Negrahasta Punta de La Coronilla (e. g. Amaro 1967; Milsteinet al. 1976; Maytía & Scarabino 1979; Riestra et al. 1992;FS). Brachidontes darwinianus se restrinje a la zona más Wdel área, i.e. Bahía de Maldonado (Maytía & Scarabino1979; Batallés 1983; Riestra et al. 1992).
Mytilus edulis (Fig. 5) posee bancos muy importantesentre Punta Ballena y José Ignacio (incluyendo Isla Gorritie Isla de Lobos) y es dominante en La Paloma (Amaro1964; 1967; Defeo & Riestra 2000; Demicheli & Scarabinoen este volumen). La distribución, estructura y dinámicapoblacional así como la interacción con otros mitílidos noha sido estudiada en la costa de Rocha, pero planteavarias líneas particularmente relevantes de investigación
F SCARABINO, JC ZAFFARONI, C CLAVIJO, A CARRANZA & M NIN
161
dado la presencia de Perna perna en la costa de Rocha yla diferencias registradas con la costa de Maldonado (verAmaro 1964; 1967; FS). El ecotipo azul se extiende hastapor lo menos 10 m de profundidad (Riestra et al. 1992),desde el submareal más somero. La pesquería de estaespecie fue analizada por Amaro (1967) y Defeo & Riestra(2000). La complejidad evolutiva, taxonómica, ecológicay biogeográfica de esta especie (ver e.g. Seed 1992 yGardner 2004 y referencias allí) no ha sido abordada aúnen la región.
Perna perna (Mytilidae, Fig. 6), que ha tenido unasecuencia de avance y retroceso en la costa uruguaya, seregistra actualmente entre Punta de La Coronilla y Puntadel Este, hasta 8-9 m (ver Klappenbach 1965; Amaro1965; 1967; Maytía & Scarabino 1979; Orensanz et al.2002; Scarabino & Ortega 2004; FS). Orensanz et al. (2002)sugieren la posibilidad de influencia antrópica sumada afatores naturales en la colonización de P. perna en la costauruguaya. Los bancos importantes de esta especie seencuentran actualmente entre Cabo Polonio y Punta deLa Coronilla (FS y AC obs. pers.). La extracción de estaespecie se realizó artesanalmente con fines industrialesentre ca. 15 años a partir de fines de la década de 1950 yluego artesanalmente hasta la desaparición de los ban-cos hacia fines de los ´70. A fines de los ´90 se retomó lapesquería artesanal y la extracción recreacional (Amaro1965; Orensanz et al. 2002; Scarabino & Ortega 2004; A.Pose com. pers; FS obs. pers.)
La presencia escasa de Modiolus carvalhoi (Mytilidae)ha sido verificada puntualmente para La Paloma(Zaffaroni 2000) y durante todo el año en en submarealmás somero de Cerro Verde (AC). Zaffaroni (2000) su-giere una posible influencia antropogénica en la presen-cia de esta especie en la costa uruguaya.
El registro de Mytilaster solisianus (Mytilidae) para lacosta uruguaya (d´Orbigny 1842) podría deberse a unevento excepcional, a un cambio en la distribución histó-rica de esta especie o a un error de procedencia. Su pre-sencia no ha podido ser verificada durante el siglo XX enla costa uruguaya (Klappenbach 1965; FS; AC; JCZ).
Ostreola equestris (Ostreidae) aparece en pozas demarea y bajo piedras en las zonas más someras delsubmareal, siendo registrada hasta 15 m (FS; CC; Milsteinet al. 1976; Batallés et al. 1985; Riestra et al. 1992).
Pododesmus rudis (Anomiidae) y Ostrea puelchanahabitan desde profundidades cercanas a 10 m hasta losBMP (35-50 m) (Ranson 1967; Milstein et al. 1976; Juanicó& Rodríguez-Moyano 1976; Zaffaroni 2000). Se ha men-cionado la presencia del género Crassostrea (Ostreidae) enla costa de Rocha (Figueiras & Sicardi 1968; Milstein et al.1976); su estatus posee elementos que se encuentran bajorevisión (FS en prep.).
Mientras que Plicatula gibbosa (Plicatulidae) ocupamicrosustratos duros de toda el área (Milstein et al. 1976;Juanicó & Rodríguez-Moyano 1976; FS), Pteria hirundo(Pteriidae) ocurre únicamente en los sectores másprofundos, i.e. 30-50 m (Juanicó & Rodríguez-Moyano1976; FS). Ambas especies requieren revisión en toda sudistribución.
Sphenia fragilis (Myidae) y Hiatella sp.? (Hiatellidae),habitantes de la matriz de mitílidos, se hallan desde elsubmareal somero (FS; AC; CC), hasta los BMP (Figueiras& Sicardi 1970; Milstein et al. 1976; Batallés 1983; Layerle& Scarabino 1984; Riestra et al. 1992; Obenat et al. 2001;Scarabino & Zaffaroni 2004; JCZ). La amplia distribuciónde S. fragilis (ver Coan 1999), su desarrollo lecitotrófico ysu presencia en comunidades de fouling sugiere habersido introducida en varias regiones. Sin embargo, suocurrencia en sedimentos cuaternarios del área (Scarabino& Zaffaroni 2004), sugiere un potencial complejo deespecies más que una especie introducida. Lacomposición taxonómica de Hiatella en el AtlánticoSudoccidental requiere revisión (ver Orensanz et al. 2002).Sphenia fragilis, Hiatella sp.? y Phlyctiderma semiaspera
Figura 4. Brachiodontes rodriguezii, mitílido dominante del intermarealrocoso de la costa atlántica.
Figura 5. Mytilus edulis; ecotipo azul.
Figura 6. Perna perna.
Ejemplares pequeños de Aulacomya atra atra(Mytilidae) han sido hallados en la costa de Rocha encoincidencia con períodos fríos en el AtlánticoSudoccidental (ver Scarabino & Ortega 2004).

Bivalvos marinos y estuarinos de la costa uruguaya:faunística, distribución, taxonomía y conservación
162
(Ungulinidae) ocupan también intersticios dentro de rocasperforadas en fondos submareales (FS; JCZ).
El género Lasaea (Galeommatoidea), muy común encomunidades incrustantes en otras zonas templadas delAtlántico, tiene en el área de estudio un solo registropublicado sin precisión de hábitat (Figueiras & Sicardi1980). En la costa uruguaya este género no se haencontrado hasta el momento en los hábitat registradoscomo usuales en otras regiones (e.g. matriz de mítilidos,fouling, FS obs. pers.). La aparente ausencia de este géneroen el registro fósil cuaternario uruguayo y el hallazgo detres valvas en zonas portuarias (Puerto de La Paloma yBahía de Maldonado, JCZ obs. pers.) podría indicar quesu presencia aquí se debe a introducciones puntualesque no prosperan. Esta consideración está claramentelimitada por la falta de relevamientos exhaustivos y seplantea como hipótesis a verificar.
Musculus viator (Mytilidae) vive asociado a inverte-brados coloniales desde el submareal somero (Riestra &Defeo 2000; Obenat et al. 2001; FS).
Lunarca ovalis es el único Arcidae viviente en Uru-guay, pero su presencia en yacimientos cuaternarioscosteros (e.g. Martínez et al. 2001) y la imprecisión delestado de los registros supuestamente actuales (Barattini& Ureta 1961) hacían incierta su presencia en la costauruguaya (Figueiras & Sicardi 1968). Sin embargo, valvascon restos de perióstraco han sido halladas en La Coroni-lla (MNHNM; FS) y algunos ejemplares han sido obteni-dos por pescadores frente a La Paloma (JCZ).
Noetia bisulcata (Noetiidae) y Entodesma patagonicum(Lyonsiidae), típicos de sustratos duros entre 8 y 15 m(Milstein et al. 1976) se hallan en cambio aparentementeausentes de los BMP (Juanicó & Rodríguez-Moyano 1976;Zaffaroni 2000; Demicheli & Scarabino en este volumen).El límite W de las mismas está dado en Punta Negra(Maldonado) (MNHNM; FS; Olazarri & Mones 1967).Obenat et al. (2001) registraron a E. patagonicum en colo-nias del poliqueto Phyllochaetopterus socialis en el RdlPexterior (11-12 m).
Dos géneros conocidos de comunidades incrustantes(Leptopecten y Crenella) son discutidos en el marco de losfondos incosolidados dado el tipo de información dispo-nible actualmente.
Especies perforadorasLos perforadores principalmente mecánicos Petricola
dactylus, Petricolaria stellae (Petricolidae), Cyrtopleuralanceolata, Pholas campechensis, Barnea lamellosa y Netastomadarwinii (Pholadidae) son conocidos para fangos consoli-dados de la costa de Maldonado y Rocha, donde la pri-mera especie ocurre en zonas más profundas y el resto apartir del submareal somero (Turner 1954; 1955; Olazarri1962; Figueiras & Sicardi 1971; Juanicó & Rodríguez-Moyano 1976; Demicheli 1987b; Demicheli & Scarabinoen este volumen; FS; CC).
Los perforantes químicos de rocas son tres: Petricolalapicida (Petricolidae), Lithophaga patagonica (Mytilidae)y Gastrochaena hians (Gastrochaenidae). La primera espe-
Figura 7. Ennucula uruguayensis.
cie es conocida para el rango 1-15 m (Milstein et al. 1976;FS). Mientras que L. patagonica vive entre el submarealsomero y los 35-50 m (BMP), G. hians es conocida de estaúltima localidad y de frente a Punta del Este (20 m)(Milstein et al. 1976; Zaffaroni 1991; JCZ; FS; CC). Obenatet al. (2001) citan a Gastrochaena carcellesi para coloniasdel poliqueto P. socialis en el RdlP exterior (11-12 m). Lasespecies de los tres géneros presentes en la región requie-ren revisión taxonómica.
Se cuenta con registro de cinco especies dePholadoidea perforantes de madera: Martesia fragilis(Pholadidae), Teredo navalis, Teredo bartschi, Lyroduspedicellatus (?), Bankia gouldi y Nausitora fusticola(Teredinidae) (Barattini & Ureta 1961; Klappenbach 1967;Scarabino & Maytía 1968; Calvo 1984). La presencia deM. fragilis en aguas uruguayas podría ser esporádica ono reproductiva por el tipo de registro hasta ahora cono-cido (maderas flotantes).
Sustratos inconsolidados atlánticosUna asociación particularmente recurrente que ocu-
pa fondos de limo y arcilla a partir de 20 m de profundi-dad está constituida por: Ennucula uruguayensis1 (Fig. 7,Nuculidae), Nuculana whitensis (Nuculanidae), Corbulapatagonica (Fig. 8, Corbulidae), Malletia cumingii(Malletiidae), M. uruguayensis, P. rostratus, Periplomacompressum (Periplomatidae), Cardiomya sp.2
(Cuspidariidae) (E. A. Smith 1885; Milstein et al. 1976;Cachés 1980; Layerle & Scarabino 1984; Demicheli &Scarabino en este volumen; FS, JCZ). Todas estas espe-cies tienen sus mayores abundancias en este tipo de fon-do; E. uruguayensis, M. cumingii y P. compressum estánmuy vinculadas a fondos de arcilla y limo mientras queel resto tolera grados variables de arena asociada al fan-go e inclusive C. patagonica ocupa asimismo fondos dearena fina.
1Esta especie ha sido sinonimizada con E. puelcha por diversos autores(e.g. Figueiras 1976 y referencias allí). La colecta y examen preliminarde miles de ejemplares del género de aguas costeras uruguayas eviden-ció diferencias morfológicas constantes y preferencias ecológicas dis-tintas para E. uruguayensis y E. puelcha. El hallazgo en simpatría des-carta inicialmente la posibilidad de que las diferencias halladas co-rrespondan a ecotipos (FS obs. pers.).

F SCARABINO, JC ZAFFARONI, C CLAVIJO, A CARRANZA & M NIN
163
Se vinculan a fondos arenosos o areno-fangosos próxi-mos a la costa pero protegidos (e.g. Bahía de Maldonadoy Puerto de La Paloma): P. semiaspera, Periploma ovatum,Raeta plicatella (Mactridae), Bushia rushii, Thracia similis(Thraciidae) y Venus antiqua (Veneridae) (Pilsbry 1897b;Barattini & Ureta 1961; Figueiras & Sicardi 1969;Demicheli & Scarabino en este volumen; Klappenbach &Ureta 1973; FS; JCZ; CC; MNHNM). Venus antiqua hasido registrada también para la zona de los BMP (Juanicó& Rodríguez-Moyano 1976).
Luego de la revisión de Klappenbach & Ureta (1973),Figueiras & Sicardi (1980) insisten en la existencia deotra especie de Thracia en el área; un análisis detallado deeste aspecto no se ha realizado aún.
Adrana electa (Nuculanidae), Semele proficua, Abrauruguayensis4 (Semelidae), Aequipecten tehuelchus(Pectinidae), Atrina seminuda (Pinnidae) y Mactrajaneiroensis, habitan arenas finas o medias con grado va-riables de fango y conchilla; su presencia en profundida-des menores a 10 m sólo se ha detectado en zona prote-gidas de la costa uruguaya (Pilsbry 1897b; Milstein et al.1976; Juanicó & Rodríguez-Moyano 1976; Layerle &Scarabino 1984; Demicheli & Scarabino en este volumen;FS; JCZ). Trachycardium muricatum (Cardiidae) ocupaestos fondos pero a partir de los 25 m (Juanicó &Rodríguez-Moyano 1976; FS).
Aequipecten tehuelchus posee variaciones en el desa-rrollo y conformación de costillas y resto de la escultura(Figueiras & Sicardi 1980; JCZ; MNHNM). Esto ha pro-movido la identificación errónea de Chlamys noronhensisen el área (Sicardi & Figueiras 1971; Figueiras & Sicardi1980) e inclusive la existencia del concepto Flexopectenfelipponei (Dall 1922; Waller 1991; Waloseek 1984). Esteúltimo ha sido basado en Pectinidae lisos o con pocaescultura; ejemplares de ese tipo han sido detectados enla costa uruguaya exhibiendo a la vez la típica esculturade A. tehuelchus (JCZ; FS), tal como fue detectado porFigueiras & Sicardi (1980) para el caso de la supuestapresencia de C. noronhensis en la costa uruguaya. Auncuando deben analizarse detalladamente las variaciones
Pitar rostratus fue explotada industrialmente a partirde 2002 en forma intermitente; Scarabino et al. (1985)habían destacado la conveniencia de la explotaciónartesanal de esta especie.
Otro grupo se asocia con fondos de arena gruesa ocascajo: Glycymeris longior (Glycymerididae), Semele casali(Semelidae), Eutivela isabelleana3, Transenpitar americana(Veneridae), Diplodonta vilardeboana (Ungulinidae),Macoma brevifrons, Corbula lyoni y Ennucula puelcha (Milsteinet al. 1976; Demicheli & Scarabino en este volumen; FS).Transenpitar americana fue también registrada en fondosde arena fina, entre 20 y 50 m de profundidad (Juanicó &Rodríguez-Moyano 1976; Layerle & Scarabino 1984).
Además de C. patagonica y C. lyoni se han citado parael área otras especies de Corbula, cuyo estatus requiererevisión: Corbula tryoni, Corbula contracta y Corbula cf. nasuta(Figueiras 1973; Figueiras & Sicardi 1980). A estas sesuman Corbula caribaea y su sinónimo propuesto Corbulauruguayensis (Marshall 1928; Carcelles 1944; Figueiras &Sicardi 1970; 1980; Juanicó & Rodríguez-Moyano 1976;Layerle & Scarabino 1984), para las cuales Coan (2002)plantea una estrecha afinidad con C. nasuta.
Mytilus edulis (Fig. 9) forma bancos que se desarro-llan entre 35 y 50 m, sobre arena fangosa (Juanicó &Rodríguez-Moyano 1976; Scarabino & Ortega 2004). Aquídesarrolla características cromáticas y morfométricasdistintas de las poblaciones de sustratos consolidadossomeros (Amaro 1964; FS). La explotación de estos ban-cos se realizó durante los ´70 (Juanicó & Rodríguez-Moyano 1976; Amaro 1979).
Figura 8. Corbula patagonica.
Figura 9. Mytilus edulis; ecotipo marrón asociado a fondos areno-fangosos de la plataforma.
2Layerle & Scarabino (1984) citan a Cardiomya cleryana para el área(40 m); aunque esta especie y su sinónimo propuesto Cardiomya simillimason las especies del género que se han mencionada para la región (e.g.Figueiras & Sicardi 1969; Castellanos 1970; Rios 1994), no se determi-na la especie colectada dado que no se ha efectuado la comparaciónadecuada con estas especies.3El género Eutivela requiere revisión; Fischer-Piette & Fischer II (1942)consideran E. isabelleana distinta de Eutivela dentaria, pero esto no re-sulta evidente de la ilustración del material tipo ni de la comparaciónentre ambas efectuadas por esos autores. Figueiras (1962) y Figueiras& Sicardi (1969; 1980) refieren como E. dentaria para la costa uruguayaa valvas gerónticas y/o desgastadas de E. isabelleana (FS obs. pers.; JCZobs. pers).
4Se ha mencionado para la costa uruguaya a Abra lioica (Barattini &Ureta 1961; Figueiras & Sicardi 1969; Figueiras & Sicardi 1980) perono se ha comparado esta especie con A. uruguayensis; este género re-quiere revisión en el área.

Bivalvos marinos y estuarinos de la costa uruguaya:faunística, distribución, taxonomía y conservación
164
y adjuntar elementos genéticos para comprender estasvariaciones, se rechaza aquí el concepto de F. felipponeicomo especie distinta de A. tehuelchus, considerándolocomo basado en mutaciones cuyo estudio es de graninterés evolutivo. J.M. Orensanz & V. Scarabino (com.pers.), habiendo examinado miles de ejemplares dePectinidae de los golfos patagónicos, coinciden con esteconcepto.
La presencia de Mactra en esta zona y profundidadesta dada fundamentalmente por Mactra marplatensis,registrada a lo largo de todo el rango batimétrico y aso-ciada a arena y arena fangosa (Juanicó & Rodríguez-Moyano 1976; Layerle & Scarabino 1984; FS). Sin embar-go, Mactra isabelleana también esta presente en algunaslocalidades (FS). Por otra parte, Mactra sp. ha sido consi-derada históricamente como Mactra patagonica (e.g.Figueiras & Sicardi 1969; Castellanos 1970; Rios 1994),pero el material tipo de esta especie difiere totalmentedel concepto atribuido normalmente (Aguirre 1994; FSobs. pers.). Mactra sp. es una nueva especie en proceso dedescripción cuya ocurrencia viviente es muy puntual yvinculada a arenas fangosas (Juanicó & Rodríguez-Moyano 1976; Layerle & Scarabino 1984; MNHNM). Lasconchillas de esta especie son extraordinariamente abun-dantes en fondos y playas de la zona costera uruguaya;el contraste con su presencia viviente podría deberse a unparticular potencial de conservación de sus conchillas o auna restricción progresiva de la distribución de esta es-pecie en la costa uruguaya.
Semele modesta podría estar representada por unapseudopoblación dada su distribución anfiatlántica pro-ducto probablemente de planctotrofía larval y su esca-sez en la costa uruguaya (Boss 1969; FS). Se cuenta condos registros vivos de esta especie para la zona (JCZ obs.pers.), aunque no existen datos ni evidencia indirecta deltipo de fondo que ocupa. Valvas sueltas son frecuentesen el sector E de la Bahía de Maldonado (Klappenbach1968; JCZ; FS) aunque han sido halladas también en JoséIgnacio y La Paloma.
Panopea abbreviata (Hiatellidae) fue hallada vivienteen la zona de BMP (Juanicó & Rodríguez-Moyano 1976;MNHNM), aunque valvas conjugadas juveniles han sidodetectadas en la Bahía de Maldonado (JCZ), indicandouna población en el lugar. La profundidad de enterra-miento de esta especie probablemente contribuya a laescasez de registros.
Dos de las especies de Crassatellidae mencionadaspara el área (Pilsbry 1897a; 1897b; Figueiras & Sicardi1969; Ituarte 1998), Crassinella lunulata y Crassinellamaldonadoensis, han sido asociadas respectivamente aarenas medias y arenas finas bien clasificadas (Layerle &Scarabino 1984), pero su presencia viviente en otros tiposde fondo ha sido verificada (FS). La primera ha sidoregistrada hasta ca. 40 m y la segunda hasta ca. 30 m(Layerle & Scarabino 1984). Crassinella marplatensis fueregistrada sin precisión para La Paloma (Figueiras &Sicardi 1980; Demicheli & Scarabino en este volumen),
aunque también existe una cita de Obenat et al. (2001)para agregados del poliqueto P. socialis en el RdlP exte-rior (11-12 m).
Cinco especies de Tellininae fueron halladas en el áreade estudio (Barattini 1951; Barattini & Ureta 1961; Boss1966; 1968; Figueiras & Sicardi 1969; Milstein et al. 1976;Figueiras & Sicardi 1980; JCZ; FS); Tellina sp. (Figueiras& Sicardi 1980) corresponde a Tellina iheringi (JCZ obs.pers.). Tellina petitiana, Tellina gibber y T. iheringi ocupanarenas finas o medias con grado variable de fango y enun amplio rango de profundidades (Layerle & Scarabino1984; Demicheli & Scarabino en este volumen; FS). Lapresencia de Tellina alerta (Figueiras & Sicardi 1980) pa-recería más restricta dentro de la zona costera, al igualque Tellina sandix, conocida únicamente para el Puertode La Paloma (pero con ocurrencia es incostante)(Figueiras & Sicardi 1969; JCZ) y para frente a Punta deLa Coronilla (Milstein et al. 1976).
Para Strigilla carnaria (Tellinidae), cuyo concepto a lolargo de su distribución debe ser revisado mediante mor-fología fina y genética molecular, existe poca informa-ción sobre su hábitat en la costa uruguaya, habiéndoseladetectado en la resaca del Puerto de La Paloma, JoséIgnacio y Playa Portezuelo (Layerle & Scarabino 1984;Demicheli & Scarabino en este volumen; JCZ; MNHN).
Ctena pectinella (Lucinidae) es frecuente en forma devalvas sueltas en fondos de 20-50 m (FS) y en la resacade Bahía de Maldonado (JCZ; FS) pero su presencia vi-viente fue registrada únicamente en tres oportunidadesen profundidades cercanas a 30 m (Layerle & Scarabino1984; JCZ). Esta ocurrencia y el hecho de que la costauruguaya es el límite S de distribución de esta especiepodría indicar que su presencia aquí no ha sido perma-nente en los últimos miles de años.
Warrana besnardi (Condylocardiidae) ha sido ubicadaen la resaca de La Paloma y Cabo Polonio (conchillas)(Klappenbach 1963; Figueiras & Sicardi 1980; JCZ) yviviente en dos puntos del área (15-18 m) (Layerle &Scarabino 1984).
Para el Carditidae Pleuromeris sanmartini no existenregistros de material coletado vivo en el área de estudiopero sus conchillas conjugadas son usuales en La Palo-ma, Bahía de Maldonado y Playa Portezuelo(Klappenbach 1970b; Figueiras & Sicardi 1980; JCZ). Estoúltimo también ocurre con Carditamera plata, para la cualsin embargo existen dos registros vivientes (JCZ; FS) parael área, probablemente por falta de muestreos.
Se han mencionado dos especies de Nucula(Lamellinucula) (Nuculidae) para el área: Nucula semiornatay Nucula marshalli (Marshall 1928; Figueiras 1976), cuyoestatus requiere revisión. Sin embargo, sólo se han detec-tado conchillas, lo que sugeriría una distribución restrin-gida en el área.
Lyonsia (Lyonsiidae) cuenta con el registro de Lyonsiaalvarezii (Olazarri & Mones 1967; Figueiras & Sicardi1968; Layerle & Scarabino 1984; Demicheli & Scarabinoen este volumen), pero se ha detectado heterogeneidad

F SCARABINO, JC ZAFFARONI, C CLAVIJO, A CARRANZA & M NIN
165
nicas especialmente dirigidas y efectivas (i.e. malla 0.5mm, rastra Warén, enmalladores bentónicos, buceo, as-piradora, lavado de sustratos). Tanto esos grupos comootros que incluyen especies de mayor talla han sufridohistóricamente de falta de atención de los investigadoreso del uso de metodologías inadecuadas (Mytilidae,Nuculanidae, Mactridae). Parte de estos grupos se en-cuentran bajo estudio. El uso de técnicas de biologíamolecular sobre grupos ya “estabilizados” en términosde taxonomía alfa (e.g. Veneridae y Pholadidae) resultaasimismo fundamental para la detección de potencialesespecies crípticas. El estatus de las poblaciones de Mytilusdebe recibir particulares esfuerzos de investigación dadala complejidad involucrada y la relevancia económica yecológica del género.
Otras prioridades y perspectivas de investigacióncoinciden con las propuestas por Scarabino et al. (en estevolumen) para los gasterópodos bentónicos de la mismaárea, así como para el resto de los invertebradosbentónicos (ver Scarabino en este volumen).
Implicancias para la conservación y el manejoEn el área se explota M. edulis, P. perna, M. mactroides,
D. hanleyanus, T. plebeius y E. mactroides, principalmenteen forma artesanal, aunque también existe explotaciónrecreacional en las primeras cuatro especies. Pitar rostratusse ha explotado industrialmente, pero el desarrollo deuna pesquería artesanal es recomendable. En este senti-do, en todos los casos existen fallas en al menos uno delos siguientes aspectos: conocimiento biológico-pesquero(estructura y dinámica), monitoreo, medidas de manejoy control, indicando la necesidad de enfocar esfuerzoshacia estas actividades.
El impacto de del gasterópodo invasor Rapana venosasobre poblaciones de bivalvos del área (Scarabino et al.1999) no ha sido evaluado pero podría ser importantedado el gran desarrollo de esta especie, requiriendo porlo tanto especial consideración.
Otras implicancias o actividades de conservación ymanejo coinciden con las propuestas para losgasterópodos de la misma área (ver Scarabino et al. eneste volumen).
AGRADECIMIENTOSLas siguientes personas e instituciones contribuye-
ron a este trabajo brindando: acceso a colecciones y usode equipamiento (A. Toscano, E. Cohen, MUNHINA,
Tabla 1. Número de especies de Bivalvia marinos y estuarinos presen-tes en el área según subclase y unidades ambientales consideradas.RdlP y ae=Río de la Plata y otros ambientes estuariales; Paa=Playasarenosas atlánticas; Fia=Sustratos inconsolidades atlánticos;SCA=Sustratos consolidados atlánticos.
dentro de este concepto en el área (Pilsbry 1897a; FS;JCZ), apuntando a la necesidad de una revisión en todoel Atlántico Sudoccidental. El género ocupa en el áreafondos preferentemente de arena media-gruesa y conchillay a partir de 15 m (Layerle & Scarabino 1984; FS).
Limatula sp. (Limidae) ha sido hallada en el área enbase a valvas sueltas en la Bahía de Maldonado y LaPaloma (JCZ; FS). Para esta última localidad fue regis-trada como Limatula pygmaea (Figueiras & Sicardi 1968),especie de Antarctolima que ocupa la plataforma externaen Uruguay (MNHNM; FS; Carcelles 1947).
Mysella spp. (Galeommatoidea) es común en el áreay en diversos fondos (Figueiras & Sicardi 1980; Layerle &Scarabino 1984 como Erycina sp.; FS; JCZ). Se han halla-do valvas sueltas de otras dos especies indeterminadasde Galeommatoidea en la Bahía de Maldonado (JCZ).Kellia suborbicularis ha sido mencionada para dos locali-dades del área por Figueiras & Sicardi (1980) y Layerle &Scarabino (1984); el concepto de esta especie debe revisarseen toda su supuesta área distribución.
Leptopecten bavayi (Pectinidae) es escasamente cono-cida en el área; su distribución parece situarse en profun-didades intermedias de la misma (15-40 m) (Klappenbach1970a; Milstein et al. 1976; JCZ; M. Aguilera com. pers.).
Crenella divaricata (Mytilidae) ha sido mencionada parael área a partir de ca. 20 m (Figueiras & Sicardi (1970b;Layerle & Scarabino 1984). Orensanz et al. (2002) desta-caron la necesidad de revisión de esta especie a lo largode su distribución, donde la falta de estudios morfológicosy taxonómicos así como la asociación de este género acomunidades de fouling implica la posibilidad de espe-cies introducidas no reconocidas aún.
CONCLUSIONESHasta el momento se han registrado más de 90 espe-
cies en el área, ninguna de ellas endémica (pero verScarabino et al. en este volumen). La composición deespecies es similar a la regiones inmediatas (litorales dela Provincia de Buenos Aires, Argentina y del Estado deRio Grande do Sul, Brasil) (ver Castellanos 1970; Rios1994). Como ha sido planteado por varios autores (e.g.Maytía & Scarabino 1979; Layerle & Scarabino 1984), setrata de una fauna compuesta por elementossubtropicales y templados. Tellinidae (8 spp.) y Veneridae(7) son las familias con más especies en el área.
El RdlP y otros ambientes estuariales son habitadospor 13 especies (Tabla 1) de las cuales cuatro son propia-mente estuarinas; Erodona mactoides y Mactra isabelleanadominan estos ambientes, estando la primera general-mente asociada a fondos más someros.
Prioridades y perspectivas de investigaciónEl mayor desafío taxonómico se encuentra en los gru-
pos cuyas dimensiones son generalmente inferiores a 15mm (Corbulidae, Nuculidae, Crenella, Hiatella, Lyonsia yAbra), así como taxa con diverso grado de asociación ysimbiosis con poliquetos y crustáceos (Galeommatoidea).Esto implica la necesidad de búsquedas utilizando téc-
Subclase RdlP y oae Paa Sia Sca
Protobranchia - - 7 -
Pteriomorphia 5 - 6 31
Heterodonta 8 8 42 4
Total 13 8 55 35

Bivalvos marinos y estuarinos de la costa uruguaya:faunística, distribución, taxonomía y conservación
166
Centro Interdisciplinario para el Desarrollo -CID-,Karumbé y DINARA), amplia colaboración durantemuestreos (E. Chiesa, Y. Marín, A. Masello, P. Puig, R.Bird, A. Casella, L. Delfino, S. Maytía, M. Laporta, A.Estrades, A. Fallabrino, Karumbé) y observaciones reali-zadas (M. Aguilera, O. Defeo, A. Martínez, S. Martínez,V. Scarabino, Lobo Orensanz, F. Ferreyro). A todos ellosnuestro sincero agradecimiento. J. González asistió con elmanejo de las fotografías. Una parte importante de losmuestros se realizaron en el marco del ProyectoECOPLATA III. Agradecemos a la Fundación WhitleyLaing (Rufford Small Grants for Nature Conservation)por la financiación otorgada a AC para la realización demuestreos. Este trabajo se pudo realizar gracias al apo-yo del CID y Karumbé, y especialmente a M. Amato,quien proporcionó facilidades indispensables.
REFERENCIASAguirre ML 1994 Revisión de las especies de Mactra Linné (Bivalvia)
del Cuaternario bonaerense (Argentina). Aspectos ecológicosy de distribución. Revista Española de Paleontología 9(2):131-144
Amaro-Padilla J 1964 La familia Mytilidae en el Uruguay. Revistadel Instituto de Investigaciones Pesqueras 1(3):227-241. Mon-tevideo
Amaro-Padilla J 1965 La familia Mytilidae en el Uruguay (Segundaparte). Revista del Instituto de Investigaciones Pesqueras1(4):323-339. Montevideo
Amaro-Padilla J 1967 El mejillón de la Bahía de Maldonado. Revis-ta del Instituto de Investigaciones Pesqueras 2(1):81-93. Mon-tevideo
Amaro J 1979 Mytilus edulis platensis-Pinnotheres maculatus, uncaso de comensalismo con incidencia en la tecnología de lasconservas de mariscos. Pp 273-278 In: Memorias del Semina-rio sobre Ecología Bentónica y Sedimentación de la Platafor-ma Continental del Atlántico Sur, UNESCO, ORCYT
Barattini LP 1951 Malacología uruguaya. Enumeración sistemáti-ca y sinonímica de los moluscos del Uruguay. PublicacionesCientíficas del SOYP (Servicio Oceanográfico y de Pesca)(6):179-293. Montevideo
Barattini LP & EH Ureta 1961 (“1960”) La fauna de las costas deleste (invertebrados). Publicaciones de Divulgación Científica,Museo “Dámaso Antonio Larrañaga”, 195 pp. Montevideo
Batallés LM 1983 La comunidad de mejillón Mytilus edulis platensis(d´Orbigny, 1846) de Punta del Chileno (Maldonado Uruguay):distribución, composición y estructura de la población. Tesisde Licenciatura en Oceanografía Biológica, Facultad de Huma-nidades y Ciencias (Universidad de la República), Montevi-deo. 73 pp (Inédita)
Batallés LM García V & A Malek 1985 Observaciones sobre lazonación en el litoral rocoso de la costa uruguaya. I. Recono-cimiento de los niveles superiores del sistema litoral: CaboPolonio (Depto. de Rocha, Uruguay). Contribuciones del Depto.de Oceanografía de la Facultad de Humanidades y Ciencias2(2):42-50. Montevideo
Bier R 1985 Estudio de la macrofauna bentónica del curso inferiordel Arroyo Solís Grande (Canelones-Maldonado, Uruguay).Tesis de Licenciatura en Oceanografía Biológica, Facultad deHumanidades y Ciencias (Universidad de la República), Mon-tevideo. i-v+140 pp (Inédita)
Boss KJ 1966 The subfamily Tellinidae in the Western Atlantic. Thegenus Tellina (Part I). Johnsonia 4(45):217-272
Boss KJ 1968 The subfamily Tellinidae in the Western Atlantic. Thegenera Tellina (Part II) and Tellidora. Johnsonia 4(46):273-344
Boss KJ 1969 The genus Strigilla in the Western Atlantic. Johnsonia4(47):345-368
Cachés MA 1980 Nota sobre la biología de los depósitos fangososcircalitorales frente a Punta del Este, Uruguay. Boletim doInstituto Oceanográfico 29(2):73-74. San Pablo
Calvo G 1984 Ataques de organismos perforantes a 6 especies demaderas expuestas al medio marino. Contribuciones (Departa-mento de Oceanografía, Facultad de Humanidades y Cien-cias) 1(3):1-7, 8 tablas, 4 fig. Montevideo
Carcelles A 1941 “Erodona mactroides” en el río de la Plata. Physis19:11-21, 1 lám. Buenos Aires
Carcelles A1944 Catálogo de los moluscos marinos de PuertoQuequén (República Argentina). Revista del Museo de La Plata(NS) Zoología 3:233-309, 15 lám
Carcelles A 1947 Notas sobre algunos bivalvos argentinos.Comunicaciones Zoológicas del Museo de Historia Natural deMontevideo 2(41):1-10, 1 lám
Castellanos ZA de 1970 (“1967”) Catálogo de los moluscos marinosbonaerenses. Anales de la Comisión de Investigación Científicade la Provincia de Buenos Aires 8:1-365
Castilla J & O Defeo 2001 Latin American benthic shellfisheries:emphasis on comanagement and experimental practices. Re-views in Fish Biology and Fisheries 11:1-30
Coan EV 1999 The eastern Pacific species of Sphenia (Bivalvia,Myidae). The Nautilus 113 (4):103-120
Coan EV 2002 The eastern Pacific Recent species of the Corbulidae(Bivalvia). Malacologia 44(1):47-105
da Costa MB 1971 Importância paleoecológica e estratigráfica deErodona mactroides Daudin (Mollusca, Bivalvia). Iheringia (SerieGeologia) (4):3-18, 1 lám. Porto Alegre
Dall WH 1901 Synopsis of the Solenidae of North America andAntilles. Proceedings of the United States National Museum22(1185):107-112
Dall WH 1922 Two new bivalves from Argentina. The Nautilus36(2):58-59
Defeo O 1985 Aspectos biocenológicos y de dinámica de poblaciónde “almeja amarilla”, Mesodesma mactroides (Deshayes, 1854)en la zona de la Barra del Chuy, depto. de Rocha, Uruguay. I.Biocenología. Contribuciones del Depto. de Oceanografía dela Facultad de Humanidades y Ciencias 2(4):76-98. Montevi-deo
Defeo O & A de Álava 1995 Effects of human activities on long-term trends in sandy beach populations: the wedge clam Donaxhanleyanus in Uruguay. Marine Ecology Progress Series 123:73-82
Defeo O G Riestra 2000 El mejillón Mytilus edulis platensis encostas del departamento de Maldonado: propuesta para laordenación de la pesquería. Pp 58-72 In: Rey (ed) Recursospesqueros no tradicionales: moluscos bentónicos marinos.Proyecto URU/92/003. INAPE-PNUD, Montevideo
Defeo O Jaramillo E & A Lyonnet 1992 Community structure andintertidal zonation of the macroinfauna in the Atlantic coast ofUruguay. Journal of Coastal Research 8:830-839
Demicheli MA 1986 (“1984”) Estudios exploratorios del infralitoralde las playas arenosas uruguayas. I. Playa Portezuelo.Comunicaciones de la Sociedad Malacológica del Uruguay 6(47):235-241, 4 mapas
Demicheli MA 1987a (“1985”) Estudios exploratorios del infralitoralde las playas arenosas uruguayas: II. Datos complementariossobre Playa Portezuelo. Comunicaciones de la SociedadMalacológica del Uruguay 6(48):287-290, 1 mapa

F SCARABINO, JC ZAFFARONI, C CLAVIJO, A CARRANZA & M NIN
167
Demicheli MA 1987b (“1985”) Estudios exploratorios del Infralitoralde las playas arenosas uruguayas: III, Playa Anaconda.Comunicaciones de la Sociedad Malacológica del Uruguay6(49):301-309, 3 mapas
d’Orbigny A 1834-1847 Voyage dans l’Amérique méridionale (leBrésil, la République orientale de l’Uruguay, la RépubliqueArgentine, la Patagonie, la République du Chili, la Républiquede Bolivia, la République du Pérou), exécuté pendant les années1826, 1827, 1828, 1829, 1830, 1831, 1832 et 1833, 5(3):Mollusques: i-xliii, 1-758, lám 1-85. París/Estrasburgo
Duarte E 1971 Lo nuestro en “Los moluscos del Viaje al Pacífico”de Hidalgo y Martínez (Partes I-II y III). Comunicaciones de laSociedad Malacológica del Uruguay 3(20):83-89
Escofet A Gianuca N Maytía S & V Scarabino 1979 Playas areno-sas del Atlántico Sudoccidental entre los 29º y 43º S: conside-raciones generales y esquema biocenológico. Pp 245-258 In:Memorias del Seminario sobre Ecología Bentónicas y Sedi-mentación de la Plataforma Continental del Atlántico Sur (Mon-tevideo, 9-12 de mayo de 1978). UNESCO ORCYT
Figueiras A 1962 Sobre nuevos hallazgos de moluscos subfósilesde la Transgresión Querandina. Comunicaciones de la Socie-dad Malacológica del Uruguay 1(3):53-68
Figueiras A 1973 Confirmación de la validez y existencia de Corbulatryoni E. A. Smith, 1880 viviente y fósil en la FormaciónQuerandina de Uruguay y nota sobre Corbula (Caryocorbula)nasuta Sowerby, 1833. Comunicaciones de la SociedadMalacológica del Uruguay 3(25):299-305
Figueiras A 1976 (“1975”) Revisión de las especies uruguayas dela Subclase Paleotaxodonta Korobkov, 1954 (Mollusca -Bivalvia). Comunicaciones de la Sociedad Malacológica delUruguay 4(28):59-92
Figueiras A & OE Sicardi 1968 Catálogo de los moluscos marinosdel Uruguay (Parte II). Comunicaciones de la SociedadMalacológica del Uruguay 2(15):255-273, 2 lám
Figueiras A & OE Sicardi 1969 Catálogo de los moluscos marinosdel Uruguay (Parte III). Comunicaciones de la SociedadMalacológica del Uruguay, 2(16/17):355-376 lám 3 y 4
Figueiras A & OE Sicardi 1970 Catálogo de los moluscos marinosdel Uruguay. Parte IV. Comunicaciones de la SociedadMalacológica del Uruguay 2(18):407-421 láms. 5 y 6
Figueiras A & OE Sicardi 1971 (“1970”) Catálogo de los moluscosmarinos del Uruguay. Adiciones y correcciones a la ClasePELECYPODA. Comunicaciones de la Sociedad Malacológicadel Uruguay 3(19):15-22 lám 7
Figueiras A & OE Sicardi 1980 (“1979”) Catálogo de los moluscosmarinos del Uruguay. Parte X. Revisión actualizada de losmoluscos marinos del Uruguay con descripción de las espe-cies agregadas. Sección I - Polyplacophora - Scaphopoda -Bivalvia. Comunicaciones de la Sociedad Malacológica delUruguay 5(37):107-157, 2 lám
Fiori S Vidal-Martínez V Simá-Alvarez R Rodríguez-Canul RAguirre-Macedo ML & O Defeo 2004 Field and laboratoryobservations of the mass mortality of the yellow clamMesodesma mactroides in South America: the case of Isla delJabalí, Argentina. Journal of Shellfish Research 23(2):451-455
Fischer-Piette E & PH Fischer II1942 Révision des espècesvivantes de Tivela et Eutivela du Muséum National d´HistoireNaturelle. Journal de Conchyliologie 4série 39 85(1):5-49
Gardner JA 2004 A historical perspective of the genus Mytilus(Bivalvia: Mollusca) in New Zealand: multivariate morphometricanalyses of fossil, midden and contemporary blue mussels.Biological Journal of the Linnean Society 82(3):329-344
González de Baccino R 1985 (“1984”) Estudio de una comunidadde almeja amarilla (Mesodesma mactroides Deshayes, 1854)
en la Playa de Portezuelo, Depto. de Maldonado, Uruguay.Comunicaciones de la Sociedad Malacológica del Uruguay6(46):193-206
Ituarte CF 1998 Argentine species of Crassinella Guppy, 1874(Bivalvia: Crassatellidae), and comments on other Southwest-ern Atlantic species. The Veliger 41(2):186-194
Jorcin A 1996 Distribución, abundancia y biomasa de Erodonamactroides (Mollusca: Bivalvia, Daudin 1801) en la Laguna deRocha (Rocha, Uruguay). Revista Brasileira de Biologia56(1):155-162
Jorcin A 1999 Temporal and spatial variability in the macro-zoobenthic community along a salinity gradient in the CastillosLagoon (Uruguay). Archiv für Hydrobiologie 146(3):369-384
Juanicó M & M Rodríguez-Moyano 1976 (“1975”) Composiciónfaunística de la comunidad de Mytilus edulis platensis d’Orbigny,1846, ubicada a unas 55 millas al SE de La Paloma.Comunicaciones de la Sociedad Malacológica del Uruguay4(29):113-116
Klappenbach MA 1963 Sobre Cuna (Moll. Pelecypoda) y génerosrelacionados en el Atlántico y costa de Africa del Sur. Boletimdo Instituto Oceanográfico de São Paulo 12(3):11-19, 1 lám
Klappenbach MA 1965 Lista preliminar de los Mytilidae brasileñoscon claves para su determinación y notas sobre su distribu-ción. Anais da Academia Brasileira de Ciencias 37:327-352.Rio de Janeiro
Klappenbach MA 1967 La familia Pholadidae en aguas uruguayas,con la mención de Martesia (Martesia) fragilis Verrill & Bush(1890) para la malacofauna de nuestro país. Revista del Insti-tuto de Investigaciones Pesqueras 2(1):163-168. Montevideo
Klappenbach MA 1968 Semele martinii (Reeve, 1853) of SouthernBrazil and Uruguay. The Veliger 10(3):273
Klappenbach MA 1970a Notas malacológicas, II. ComunicacionesZoológicas del Museo de Historia Natural de Montevideo10(132):1-7
Klappenbach MA 1970b Nueva especie de Pleuromeris (Pelecypoda,Carditidae) obtenida en la costa atlántica de Uruguay. Journalde Conchyliologie 108(2):35-38
Klappenbach MA & EH Ureta 1973 La familia Thraciidae (Moll.Pelecypoda) en el Atlántico sudamericano. Trabajos del V Con-greso Latinoamericano de Zoología 1:123-128. Montevideo
Layerle C & V Scarabino 1984 Moluscos del frente marítimouruguayo entre los 9 y 78 m de profundidad: análisisbiocenológico. Contribuciones (Departamento de Oceanogra-fía, Facultad de Humanidades y Ciencias) 1(9):1-17. Montevi-deo
Marshall WB 1928 New fresh-water and marine bivalve shellsfrom Brazil and Uruguay Proceedings of the United StatesNational Museum 74(2762):1-7, 4 láms
Martínez S Ubilla M Verde M Perea D Rojas R Guérèquiz R & GPiñeiro 2001 Paleoecology and geochronology of Uruguayancoastal marine Pleistocene deposits. Quaternary Research55:246-254
Martínez y Saez F 1872 (“1869”) Moluscos del Viaje al Pacíficoverificado de 1862 a 1865 por una comisión de nauralisasenviada por el Gobierno Español. Parte segunda, Bivalvosmarinos. Imprenta de Miguel Ginesta, Madrid. 78 pp, 8 lám
Maytía S & V Scarabino 1979 Las comunidades del litoral rocosodel Uruguay: zonación, distribución local y consideracionesbiogeográficas. Pp 149-160 In: Memorias del Seminario sobreEcología Bentónicas y Sedimentación de la Plataforma Conti-nental del Atlántico Sur (Montevideo, 9-12 de mayo de 1978).UNESCO, ORCYT
Milstein A Juanicó M & J Olazarri 1976 Algunas asociacionesbentónicas frente a las costas de Rocha, Uruguay. Resulta-

Bivalvos marinos y estuarinos de la costa uruguaya:faunística, distribución, taxonomía y conservación
168
dos de la campaña del R/V “Hero”, viaje 72-3A. Comunicacio-nes de la Sociedad Malacológica del Uruguay 4(50):143-164
Milstein A Juanicó M & J Olazarri 1976 Algunas asociacionesbentónicas frente a las costas de Rocha, Uruguay. Resulta-dos de la campaña del R/V “Hero”, viaje 72-3A. Comunicacio-nes de la Sociedad Malacológica del Uruguay 4(50):143-164
Neirotti E 1981 Estudio comparativo de supralitoral y mesolitoralrocoso en diferentes localidades del estuario del Río de laPlata. Comunicaciones de la Sociedad Malacológica del Uru-guay 5(40):347-370
Nion H 1979 Zonación del macrobentos en un sistema lagunarlitoral oceánico. Pp 225-235 In: Memorias del Seminario sobreEcología Bentónicas y Sedimentación de la Plataforma Conti-nental del Atlántico Sur (Montevideo, 9-12 de mayo de 1978).UNESCO, ORCYT
Obenat S Ferrero L & E Spivak 2001 Macrofauna associated withPhyllochaetopterus socialis aggregations in the southwesternAtlantic. Vie et Milieu 51(3):131-139
Olazarri J 1962 Ampliación de la distribución de Pholascampechiensis Gmelin. Comunicaciones de la SociedadMalacológica del Uruguay 1(3):51-52
Olazarri J & A Mones 1967 Las especies uruguayas de LyonsiaTurton, 1822 (Pelecypoda, Pandoracea). Comunicaciones dela Sociedad Malacológica del Uruguay 2(13):113-116
Olsson D Pesamosca P Saona G Forni F Vizziano D Verocai J& W Norbis 2003 Análisis del contenido estomacal de la corvina(Micropogonias furnieri) (Desmarest, 1823) en la Laguna deRocha-Uruguay. Publicación Especial de la Sociedad Zoológi-ca del Uruguay (Actas de las VII Jornadas de Zoología delUruguay, I Encuentro de Ecología del Uruguay):125
Orensanz JM Schwindt E Pastorino G Bortolus A Casas GDarrigrán G Elías R López-Gappa JJ Obenat S Pascual MPenchaszadeh P Piriz ML Scarabino F Spivak ED & EAVillarino 2002 No longer the pristine confines of the worldocean: a survey of exotic marine species in the southwesternAtlantic. Biological Invasions 4:115-143
Passadore C & L Giménez 2003 Composición y variación intra-anual de la macroinfauna de la desembocadura del arroyoPando. Publicación Especial de la Sociedad Zoológica del Uru-guay (Actas de las VII Jornadas de Zoología del Uruguay, IEncuentro de Ecología del Uruguay):125
Pilsbry HA 1897a List of mollusks collected in Maldonado Bay,Uruguay, by Dr. Wm. H. Rush, U. S. N. The Nautilus 11(1):6-9
Pilsbry HA 1897b New species of mollusks from Uruguay. Pro-ceedings of the Academy of Natural Sciences of Philadelphia49:290-298, lám 6 y 7
Pin O & FG Peluffo 1983 Hallazgo y descripción de una poblaciónde Petricola pholadiformis (Lamarck, 1818) (Heterodonta,Petricolidae) [en] Balneario San Luis (Canelones, Uruguay).VIII Simposio Lationamericano sobre Oceanografía Biológica(Montevideo, 28de noviembre-2 de diciembre de 1983),Resúmenes:124
Ranson G 1967 Mollusques lamellibranches: Ostreidae. RésultatsScientifiques des Campagnes de la «Calypso» (8). Annales del’Institute Océanographique 45(2):189-191. París
Riestra G & O Defeo 2000 La comunidad macrobentónica asociadaal mejillón Mytilus edulis platensis en costas del Departamentode Maldonado: variación espacio-temporal e incidencia del im-pacto pesquero. Pp 17-57 In: Rey (ed) Recursos pesqueros notradicionales: moluscos bentónicos marinos. Proyecto URU/92/003. INAPE-PNUD, Montevideo
Riestra G Giménez JL & V Scarabino 1992 Análisis de la comu-nidad macrobentónica infralitoral de fondo rocoso en Isla Gorriti
e Isla de Lobos (Maldonado, Uruguay). Frente Marítimo 11:123-127. Montevideo
Rios EC 1994 Seashells of Brazil. Museu Oceanográfico da FundaçaoUniversidade do Rio Grande, 329 pp, 102 lám
Roux A & C Bremec 1996 Comunidades bentónicas relevadas enlas transecciones realizadas frente al Río de la Plata (35º15´S),Mar del Plata (38º10´S) y Península Valdés (42º35´S), Argen-tina. INIDEP Informe Técnico (11):13 pp. Mar del Plata
Santana O & G Fabiano 1999 Medidas y mecanismos de adminis-tración de los recursos de las lagunas costeras del litoral atlánticodel Uruguay (Lagunas José Ignacio, Garzón, de Rocha y deCastillos). Plan de Investigación Pesquera URU/92/003. INAPE-PNUD, Montevideo. 165 pp+apéndice
Scarabino F 2003 Lista sistemática de los Bivalvia marinos yestuarinos vivientes de Uruguay. Comunicaciones de la So-ciedad Malacológica del Uruguay 8(80-81):229-259
Scarabino F 2004 Conservación de la malacofauna uruguaya.Comunicaciones de la Sociedad Malacológica del Uruguay 8(82/83):267-273
Scarabino F & L Ortega 2004 Registros uruguayos de Aulacomyaatra atra (Bivalvia: Mytilidae): rol de condiciones oceanográficasanómalas y de dispersión por feofitas flotantes. Comunicacio-nes de la Sociedad Malacológica del Uruguay 8(84/85-86/87):299-304
Scarabino F & JC Zaffaroni 2004 Estatus faunístico de veinteespecies de moluscos citadas para aguas uruguayas. Comu-nicaciones Zoológicas, Museo Nacional de Historia Natural yAntropología 13(202):1-15. Montevideo
Scarabino F Menafra R & P Etchegaray 1999 Presencia de Rapanavenosa (Valenciennes, 1846) (Gastropoda: Muricidae) en el Ríode la Plata. Boletín de la Sociedad Zoológica del Uruguay(Actas de las V Jornadas de Zoología del Uruguay) 11 (Segun-da Epoca):40. Montevideo
Scarabino V & S Maytía 1968 Teredinidae (Moll. Pelecypoda) delUruguay (I). Comunicaciones de la Sociedad Malacológica delUruguay 2(15):321-325
Scarabino V Defeo O & L Barea 1985 Invertebrados bentónicosaccesibles a la actividad pesquera nacional. Actas de las Jor-nadas de Zoología del Uruguay:54-56
Scarabino V Maytía S & M Cachés 1976 (“1975”) Carta bionómicalitoral del departamento de Montevideo I. Niveles superioresdel sistema litoral. Comunicaciones de la Sociedad Malacológicadel Uruguay 4(29):117-126, 3 lám
Scarabino V Maytía S & JC Faedo 1974 Zonación biocenológicade las playas arenosas del Depto. de Rocha (Uruguay), conespecial referencia a la presencia de Ocypode quadrata(Fabricius, 1787) (Decapoda, Brachyura). Boletín de la Comi-sión Nacional de Oceanografía 1(1):42-52, 2 lám 1 mapa.Montevideo
Seed R 1992 Systematics evolution and distribution of musselsbelonging to the genus Mytilus: an overview. AmericanMalacological Bulletin 9(2):123-137
Sicardi OE & A Figueiras 1971 Presencia de Chlamys noronhensis(E. A. Smith, 1885) en aguas uruguayas. Comunicaciones dela Sociedad Malacológica del Uruguay 3(20):97-98
Smith EA 1885 Report on the Lamellibranchiata collected by theH.M.S Challenger during the years 1873-76. Report of theScientific Results of the Voyage H.M.S. Challenger during theyears 1873-76...Zoology 13:i-viii+1-341, 25 lám
Sprechmann P 1978 The paleoecology and paleogeography of theUruguayan coastal area durng the Neogene and Quaternary.Zitteliana 4:3-72 lám 1-6
Turner RD 1954 The family Pholadidae in the Western Atlantic andthe Eastern Pacific. Part I - Pholadinae. Johnsonia 3(33):1-63

F SCARABINO, JC ZAFFARONI, C CLAVIJO, A CARRANZA & M NIN
169
Turner RD 1955 The family Pholadidae in the Western Atlantic andthe Eastern Pacific. Part II - Martesiinae, Jouannetiinae andXylophaginae. Johnsonia 3(34):65-160
Waller TR 1991 Evolutionary relationships among commercial scal-lops (Mollusca: Bivalvia: Pectinidae). Pp 1-73 In: Shumway(ed) Scallops: biology, ecology an aquaculture. Developmentsin Aquaculture and Fisheries Science 21
Walossek D 1984 Taxonomie und Verbreitung von Chlamyspatagonica (King & Broderip 1832) und Anmekungen zuweiteren Chlamys-Arten von der Südspitze Süd-Amerikas (Mol-lusca, Bivalvia, Pectinidae). Verhandlungen desnaturwissenschaftlichen Vereins in Hamburg (NF) 27:207-276
Zaffaroni JC 1991 (“1989”) Eulimella bermudensis (Dall & Bartsch,1911) y Gastrochaena hians (Gmelin, 1791), primera cita paraaguas uruguayas. Comunicaciones de la Sociedad Malacológicadel Uruguay 7(56/57):123-128
Zaffaroni JC 2000 (“1998”) Presencia de Modiolus carvalhoi(Mollusca, Pelecypoda) en aguas uruguayas. Comunicacionesde la Sociedad Malacológica del Uruguay 8(74/75):101-103

Bivalvos marinos y estuarinos de la costa uruguaya:faunística, distribución, taxonomía y conservación
170

BASES para la CONSERVACIÓNy el MANEJO de la
COSTA URUGUAYA
R. MenafraL. Rodríguez-Gallego
F. ScarabinoD. Conde(editores)

La referencia correcta de este libro es:
Menafra R Rodríguez-Gallego L Scarabino F & D Conde(eds) 2006 Bases para la conservación y el manejo de la costauruguaya. VIDA SILVESTRE URUGUAY, Montevideo.i-xiv+668pp
Armado y diagramación: Javier González
Fotografía de portada: Faro de Cabo Polonio (Rocha)Diego Velazco - Aguaclara Fotostock,www.aguaclara.com.uy
Impreso en GRAPHIS Ltda, en el mes de octubre de 2006Nicaragua 2234, Montevideo, UruguayTels.: 409 6821-409 9168. E-mail: [email protected]ósito legal: 339.537/06
ISBN: 9974-7589-2-0
Las opiniones e informaciones contenidas en este libro sonexclusiva responsabilidad de sus autores, y no reflejannecesariamente aquellas de VIDA SILVESTRE URUGUAY,US Fish and Wildlife Service, Facultad de Ciencias, o de lasinstituciones a las cuales los autores están vinculados.

LISTA DE AUTORES PARTICIPANTES
PRÓLOGOOSCAR IRIBARNE
PRÓLOGOULRICH SEELIGER
PREFACIO
AGRADECIMIENTOS
Mapa general de la costa platense-atlántica
Mapa de la costa Oeste del Río de la Plata
Mapa del sector centro-Sur de la costa platense-atlántica
Mapa del sector Este de la costa atlántica
COSTA PLATENSE-ATLÁNTICA
Evolución paleogeográfica y dispersión de los sedimentos del Río de la PlataRICARDO N. AYUP-ZOUAIN
Geología de la costa uruguaya y sus recursos minerales asociadosCÉSAR A. GOSO AGUILAR & ROSSANA MUZIO
Dinámica y fuentes de sedimentos de las playas uruguayasDANIEL PANARIO & OFELIA GUTIÉRREZ
Geomorfología y procesos erosivos en la costa atlántica uruguayaMARÍA ALEJANDRA GÓMEZ PIVEL
Fitoplancton de la zona costera uruguaya: Río de la Plata y Océano AtlánticoGRACIELA FERRARI & LETICIA VIDAL
El impacto de las floraciones algales nocivas: origen, dispersión, monitoreo,control y mitigaciónSILVIA M. MÉNDEZ
Flora y vegetación de la costa platense y atlántica uruguayaEDUARDO ALONSO-PAZ & MARÍA JULIA BASSAGODA
Fauna parasitaria del lobo fino Arctocephalus australis y del león marino Otariaflavescens (Mammalia, Otariidae) en la costa uruguayaDIANA MORGADES, HELENA KATZ, OSCAR CASTRO, DINORA CAPELLINO, LOURDES CASAS,GUSTAVO BENÍTEZ, JOSÉ MANUEL VENZAL & ANTONIO MORAÑA
Zooplancton gelatinoso de la costa uruguayaMARÍA GABRIELA FAILLA SIQUIER
Zooplancton de ambientes costeros de Uruguay: añadiendo piezas alrompecabezasGUILLERMO CERVETTO, DANILO CALLIARI, LAURA RODRÍGUEZ-GRAÑA, GISSELL LACEROT
& RAFAEL CASTIGLIONI
Faunística y taxonomía de invertebrados bentónicos marinos y estuarinos dela costa uruguayaFABRIZIO SCARABINO
Índice
........................................................................................................ i
.............................................................................................................................................................. v
........................................................................................................................................................... vii
............................................................................................................................................................... viii
........................................................................................................................................... x
................................................................................................. xi
................................................................................................... xii
..................................................................... xiii
..................................................................................................... xiv
............................................................................................................................................ 1
............................................................................................................... 9
........................................................................................................................ 21
.................................................................................................................................. 35
............................................................................................................................. 45
....................................................................................................................................................... 57
................................................................................................... 71
............................................................................... 89
............................................................................................................................................... 105
.................................................................................................................................. 97
................................................................................................................................................... 113

Gasterópodos marinos y estuarinos de la costa uruguaya: faunística,distribución, taxonomía y conservaciónFABRIZIO SCARABINO, JUAN CARLOS ZAFFARONI, ALVAR CARRANZA, CRISTHIAN CLAVIJO
& MARIANA NIN
Bivalvos marinos y estuarinos de la costa uruguaya: faunística, distribución,taxonomía y conservaciónFABRIZIO SCARABINO, JUAN CARLOS ZAFFARONI, CRISTHIAN CLAVIJO, ALVAR CARRANZA
& MARIANA NIN
Patrones geográficos de diversidad bentónica en el litoral rocoso de UruguayALEJANDRO BRAZEIRO, ANA INÉS BORTHAGARAY & LUIS GIMÉNEZ
Comunidades bentónicas estuarinas de la costa uruguayaLUÍS GIMÉNEZ
Asociaciones de moluscos bentónicos cuaternarios en la costa uruguaya:implicancias paleoecológicasSERGIO MARTÍNEZ &ALEJANDRA ROJAS
Los recursos pesqueros de la costa de Uruguay: ambiente, biología y gestiónWALTER NORBIS, LAURA PAESCH & OSCAR GALLI
Áreas de cría de peces en la costa uruguayaSUSANA RETTA, GUSTAVO MARTÍNEZ & ADRIANA ERREA
Características biológicas de la corvina (Micropogonias furnieri) en el Río de laPlata y su Frente MarítimoERNESTO CHIESA, OSCAR D. PIN & PABLO PUIG
Abundancia, capturas y medidas de manejo del recurso corvina(Micropogonias furnieri) en el Río de la Plata y Zona Común de PescaArgentino-Uruguaya (1975-2003)OSCAR D. PIN, GUILLERMO ARENA, ERNESTO CHIESA & PABLO PUIG
Herpetofauna de la costa uruguayaRAÚL MANEYRO & SANTIAGO CARREIRA
Biología, ecología y etología de las tortugas marinas en la zona costerauruguayaMILAGROS LÓPEZ-MENDILAHARSU, ANDRÉS ESTRADES, MARÍA NOEL CARACCIO,VICTORIA CALVO, MARTÍN HERNÁNDEZ & VERÓNICA QUIRICI
Conservación y manejo de tortugas marinas en la zona costera uruguayaMARTÍN LAPORTA, PHILIP MILLER, MARIANA RÍOS, CECILIA LEZAMA, ANTONIA BAUZÁ, ANITA
AISENBERG, MARÍA VICTORIA PASTORINO & ALEJANDRO FALLABRINO
Aves de la costa sur y este uruguaya: composición de especies en los distintosambientes y su estado de conservaciónJOAQUÍN ALDABE, SEBASTIÁN JIMÉNEZ & JAVIER LENZI
La franciscana Pontoporia blainvillei (Cetacea, Pontoporiidae) en la costauruguaya: estudios regionales y perspectivas para su conservaciónCAROLINA ABUD, CATERINA DIMITRIADIS, PAULA LAPORTA &MARILA LÁZARO
Revisión preliminar de registros de varamientos de cetáceos en la costauruguaya de 1934 a 2005DANIEL DEL BENE, VIRGINIA LITTLE, RICARDO ROSSI & ALFREDO LE BAS
......................................................................................................................................................... 143
.............................................................................................................................................................. 179
...................................................................................................................... 189
...................................................................................................... 197
.......................................................................................... 211
......................................................................................................................................................... 157
........................................................................... 171
....................................................................................................... 219
...................................................................... 225
................................................................................................................... 233
................................................................................. 247
........................................................................ 259
............................................................................................... 271
.......................................................... 289
.................................................................. 297

Distribución, reproducción y alimentación del lobo fino Arctocephalus australisy del león marino Otaria flavescens en UruguayALBERTO PONCE DE LEÓN & OSCAR D. PIN
Tuberculosis en pinnípedos (Arctocephalus australis y Otaria flavescens) deUruguayMIGUEL CASTRO RAMOS, HELENA KATZ, ANTONIO MORAÑA, MARÍA INÉS TISCORNIA,DIANA MORGADES & OSCAR CASTRO
Interacciones entre lobos marinos y pesca artesanal en la costa de UruguayDIANA SZTEREN & CECILIA LEZAMA
Mamíferos terrestres no voladores de la zona costera uruguayaENRIQUE M. GONZÁLEZ
Vertebrados fósiles de la costa uruguayaANDRÉS RINDERKNECHT
Especies acuáticas exóticas en Uruguay: situación, problemática y manejoERNESTO BRUGNOLI, JUAN CLEMENTE, GUSTAVO RIESTRA, LUCÍA BOCCARDI
& ANA INÉS BORTHAGARAY
Ecología de playas arenosas de la costa uruguaya: una revisión de 25 años deinvestigaciónOMAR DEFEO, DIEGO LERCARI, ANITA DE ÁLAVA, JULIO GÓMEZ, GASTÓN MARTÍNEZ, ELEONORA
CELENTANO, JUAN PABLO LOZOYA, SEBASTIÁN SAUCO, DANIEL CARRIZO & ESTELA DELGADO
Estado actual, propuestas y perspectivas de manejo de las Áreas ProtegidasCosterasJUAN CARLOS GAMBAROTTA
Bases ecológicas y metodológicas para el diseño de un Sistema Nacional deÁreas Marinas Protegidas en UruguayALEJANDRO BRAZEIRO & OMAR DEFEO
COSTA del RÍO de la PLATA
Evolución de la desembocadura del Arroyo Pando (Canelones, Uruguay):¿tendencias naturales o efectos antrópicos?OFELIA GUTIÉRREZ & DANIEL PANARIO
La Bahía de Montevideo: 150 años de modificación de un paisaje costero ysubacuáticoPIERRE GAUTREAU
Monitoreo de cianobacterias en la costa de Montevideo (Uruguay)DANIEL SIENRA & GRACIELA FERRARI
Comunidad componente de tremátodos larvales de Heleobia australis(Mollusca, Cochliopidae) en la costa uruguaya del Río de la PlataOSCAR CASTRO, DANIEL CARNEVIA, ALEJANDRO PERRETTA & JOSÉ MANUEL VENZAL
Composición y ecología de la fauna epígea de Marindia (Canelones, Uruguay)con especial énfasis en las arañas: un estudio de dos años con trampas deintercepciónFERNANDO G. COSTA, MIGUEL SIMÓ & ANITA AISENBERG
Ictioplancton costero de la zona de transición estuarina del Río de la Plata(Uruguay)GABRIELA MANTERO, SUSANA RETTA & MARCELO RODRÍGUEZ
............................................................................................................... 305
......................................................................................................................... 315
........................................................................................................................... 321
............................................................................................................................................. 329
.............................................................................................................................................. 343
......................................................................................................................................... 351
...................................363
........................................................................................................................................ 371
....................................................................................................................... 379
...................................................................................................................... 391
....................................................................................................................................................... 401
......................................................................................................................... 413
.............................................. 421
......................................................................................... 427
.................................................................................. 437

Ecología de un ensamble de anuros en un humedal costero del sudeste deUruguayINÉS DA ROSA, ARLEY CAMARGO, ANDRÉS CANAVERO, DANIEL E. NAYA & RAÚL MANEYRO
Aves de la costa de Montevideo urbano: variación espacial y estacionalMACARENA SARROCA, MATILDE ALFARO, JAVIER LENZI, SEBASTIÁN JIMÉNEZ, CAROLINA ABUD
& DIEGO CABALLERO-SADI
Contaminación de la Bahía de Montevideo y zona costera adyacente y surelación con los organismos bentónicosPABLO MUNIZ, NATALIA VENTURINI & LETICIA BURONE
La pesca artesanal en el Río de la Plata: su presente y una visión de futuroPABLO PUIG
COSTA ATLÁNTICA
Paleolimnología: desarrollo de las lagunas costeras del sudeste de Uruguaydurante el HolocenoFELIPE GARCÍA-RODRÍGUEZ, PETER SPRECHMANN, HUGO INDA, LAURA DEL PUERTO,ROBERTO BRACCO, ADRIANA RODRÍGUEZ, PETER ESTOL & VIRGINIA ACEVEDO
Fisonomía y composición florística de Cabo Polonio (Rocha, Uruguay)SILVANA MASCIADRI, ELOISA FIGUEREDO &LILIANA DELFINO
Estructura y regeneración del Bosque de Ombúes (Phytolacca dioica) de laLaguna de Castillos (Rocha, Uruguay)MARÍA GABRIELA RODRÍGUEZ-GALLEGO
Estructura poblacional y reproducción del tatucito Emerita brasiliensis(Decapoda: Hippidae) en playas de Cabo Polonio (Rocha, Uruguay)ENRIQUE PELUFFO
Invertebrados bentónicos de La Paloma (Rocha, Uruguay)MARIO DEMICHELI & FABRIZIO SCARABINO
Ecología de comunidades de playas de Cabo Polonio (Rocha, Uruguay)LUIS GIMÉNEZ & BEATRIZ YANNICELLI
Transgresiones y regresiones marinas en la costa atlántica y lagunas costerasde Uruguay: efectos sobre los peces continentalesMARCELO LOUREIRO & GRACIELA GARCÍA
Las pesquerías en las lagunas costeras salobres de UruguayGRACIELA FABIANO & ORLANDO SANTANA
La pesca artesanal en la Paloma (Rocha, Uruguay): período 1999-2001ELIZABETH DELFINO, GRACIELA FABIANO & ORLANDO SANTANA
Situación de la administración del recurso lobos y leones marinos en UruguayENRIQUE PÁEZ
Ballena franca (Eubalaeana australis) en la costa atlántica uruguayaMARIANA PIEDRA, PAULA COSTA, PAULA FRANCO FRAGUAS & RAFAEL ÁLVAREZ
Evaluación del turismo de observación de ballenas como una herramientapara la conservación y el manejo de ballena franca austral (Eubalaena australis)RODRIGO GARCÍA & UZI SABAH
................................... 447
......................................................................................................................................... 457
........................................................................................... 467
................................................................................................................................................................. 477
......................................................... 487
.................................................................................... 495
..................................................................................................................... 503
....................................................................................................................................................... 513
................................................................................................................. 523
....................................................................................................................... 535
................................................................................................................. 545
................................................................................................................. 557
............................................................................... 567
............................................................................................................................................................. 577
..................................................... 585
................................................................................................................................. 591

Biodiversidad y calidad de agua de 18 pequeñas lagunas en la costa surestede UruguayCARLA KRUK, LORENA RODRÍGUEZ-GALLEGO, FEDERICO QUINTANS, GISELL LACEROT,FLAVIO SCASSO, NÉSTOR MAZZEO, MARIANA MEERHOFF & JUAN CÉSAR PAGGI
Procesos estructuradores de las comunidades biológicas en lagunas costerasde UruguaySYLVIA BONILLA, DANIEL CONDE, LUIS AUBRIOT, LORENA RODRÍGUEZ-GALLEGO, CLAUDIA PICCINI,ERIKA MEERHOFF, LAURA RODRÍGUEZ-GRAÑA, DANILO CALLIARI, PAOLA GÓMEZ,IRENE MACHADO & ANAMAR BRITOS
Efectos del Canal Andreoni en playas de Rocha: deterioro ambiental y suefecto en la biodiversidadDIEGO LERCARI & OMAR DEFEO
Interfase de conflictos: el sistema costero de Rocha (Uruguay)DANIEL DE ÁLAVA
Importancia de los procesos participativos en la planificación: percepcionesde naturaleza y áreas a proteger en Castillos (Rocha, Uruguay) y su zona deinfluencia costeraDIEGO MARTINO & ANDREA SCHUNK
Aprovechamiento prehistórico de recursos costeros en el litoral atlánticouruguayoHUGO INDA, LAURA DEL PUERTO, CAROLA CASTIÑEIRA, IRINA CAPDEPONT
& FELIPE GARCÍA-RODRÍGUEZ
....................................................... 599
.......................................................................................................................... 611
................................................................................................................................. 631
....................................................................................................................................................... 637
......................................................................................................................... 651
..................................................................................................................................... 661

F SCARABINO, JC ZAFFARONI, A CARRANZA, C CLAVIJO & M NIN
143
Gasterópodos marinos yestuarinos de la costauruguaya: faunística,distribución, taxonomía yconservación
FABRIZIO SCARABINO*, JUAN CARLOS ZAFFARONI, ALVAR
CARRANZA, CRISTHIAN CLAVIJO & MARIANA NIN
RESUMENSe presenta una síntesis y nueva información sobrefaunística, distribución, taxonomía y conservación de lamalacofauna uruguaya de Gastropoda marinos yestuarinos costeros bentónicos (0-50 m). La misma estácompuesta por al menos 140 especies, de las cuales solosiete son estuarinas (incluyendo las exóticas Rapana venosay Myosotella myosotis). Los géneros Olivancillaria (Olividae)y Buccinanops (Nassariidae) son particularmente diver-sos en especies y están distribuidos en una variedad defondos inconsolidados. En el Río de la Plata y y otrosambientes estuariales el género Heleobia (Cochliopidae)está ampliamente distribuido y es considerado aquí comomáxima prioridad de investigación. Otros grupos priori-tarios son aquellos generalmente compuestos pormicrogasterópodos que poseen diversos grados de aso-ciación y simbiosis con poríferos, antozoos, poliquetos,crustáceos, equinodermos y briozoos (Epitoniidae,Eulimidae, Triphoridae, Cerithiopsidae, Pyramidellidaey Nudibranchia), así como Trochoidea y Vitrinellidae. Seinsiste en la necesidad de nuevas técnicas de muestreo yenfoques de investigación, formación de colecciones yrecursos humanos como elementos particularmente crí-ticos para llenar vacíos de información. El manejo ade-cuado del agua de lastre y del químico tributiltina (TBT),la reglamentación y control de la extracción de algunasespecies, la implementación de áreas protegidas y el en-foque ecosistémico en pesquerías son considerados departicular relevancia para la conservación de esa fauna.
Palabras clave: Mollusca, caracoles, Heleobia, Uruguay,Río de la Plata.
INTRODUCCIÓNAún cuando los moluscos son mejor conocidos que
cualquier otro phylum de invertebrados presente en Uru-guay, y a pesar de su enorme relevancia, el conocimientode los mismos es producto de esfuerzos aislados y dis-pares, lo que determina grandes vacíos de conocimientotaxonómico y faunístico (Scarabino 1999; 2004a; 2004b).El estudio básico de los gasterópodos costeros de Uru-guay tiene una historia importante (ver Scarabino 2004b),pero no queda excluido de ese contexto.
Barattini & Ureta (1961) y Figueiras & Sicardi (1971;1972; 1973a; 1973b; 1974; 1980) compilaron y ampliaronla información básica existente sobre los gasterópodosmarinos y estuarinos de Uruguay, sirviendo de base y
ABSTRACTA synthesis and new data on faunistics, distribution, taxonomyand conservation of the Uruguayan malacofauna of coastalbenthic marine (0-50 m) and estuarine Gastropoda is pre-sented. It is composed by at least 140 species, of which onlyseven are estuarine (including the exotics Rapana venosa andMyosotella myosotis). The genera Olivancillaria (Olividae)and Buccinanops (Nassariidae) are particularly well repre-sented and distributed in varied unconsolidated bottoms. Inthe the Río de la Plata and other estuarine environments thegenus Heleobia (Cochliopidae) is widely distributed and isconsidered here as a top research priority. Other prioritygroups are those generally composed by microgastropodsthat have varied degrees of association and symbiosis withporiferans, anthozoans, polychaetes, crustaceans, echinodermsand bryozoans (Epitoniidae, Eulimidae, Triphoridae,Cerithiopsidae, Pyramidellidae and Nudibranchia), as wellon Trochoidea and Vitrinellidae. The need for new samplingtechniques and research approaches is stressed, as well as thegeneration of collections and human capacity building asparticularly critical elements for filling gaps of information.The adequate management of ballast water and tributyltin(TBT), the regulation and control of extraction of some spe-cies, the implementation of protected areas and ecosystemapproach of bottom fisheries are considered particularly rel-evant for the conservation of this fauna.
Key words: Mollusca, snails, Heleobia, Uruguay, Ríode la Plata
R MENAFRA, L RODRÍGUEZ-GALLEGO, F SCARABINO & D CONDE (eds) 2006Bases para la conservación y el manejo de la costa uruguaya
VIDA SILVESTREURUGUAY
estímulo para el desarrollo de nuevos conocimientos yrecursos humanos. Algunas limitaciones de la informacióncontenida en estos trabajos y de aportes posteriores sonmencionadas por Sprechmann (1978) y Scarabino et al.(en este volumen) en relación a bivalvos e igualmenteaplicable a gasterópodos.
En este trabajo se presenta una puesta a punto delconocimiento faunístico, de distribución y taxonómicode los Gastropoda marinos y estuarinos bentónicos de lacosta uruguaya, discutiéndose prioridades y perspecti-vas de investigación, así como implicancias y otras con-sideraciones sobre conservación y manejo.
La falta de bases de datos actualizadas y electróni-cas del material presente en colecciones, la vasta canti-

Gasterópodos marinos y estuarinos de la costa uruguaya:faunística, distribución, taxonomía y conservación
144
dad de material recientemente obtenido y la dispersiónde algunas muestras limitan el alcance de este trabajo.
METODOLOGÍASe consideraron las referencias publicadas en los últi-
mos 172 años (incluyendo resúmenes de congresos), yaque King & Broderip (1832) marcan el punto de partidadel conocimiento de los gasterópodos uruguayos. Tesisinéditas también fueron consideradas. Registros incorrec-tos o muy dudosos ya discutidos no se mencionan eneste trabajo y una parte de los mismos figura en Scarabino& Zaffaroni (2004) y Scarabino (2004b). La sistemáticaconsiderada es la usada por Scarabino (2004b), a excep-ción de Littorinidae, Naticidae y Cylichnidae, que sigue aWilliams et al. (2003), Pastorino (2005b) y Valdés &Camacho-García (2004), respectivamente.
El material examinado está depositado en las siguien-tes colecciones o consiste en los siguientes materiales (lasabreviaciones colocadas luego de Montevideo correspon-den a las utilizadas en el texto): 1) Museo Nacional deHistoria Natural y Antropología (Montevideo, MNHNM).No ha sido posible reanalizar el material estudiado porLayerle & Scarabino (1984), depositado en esa colección,por la situación actual de la misma; 2) colección LPBarattini, Museo Zoológico Municipal “Dámaso A.Larrañaga” (Montevideo, LPB); 3) material obtenido porF. Scarabino (FS), A. Carranza (AC) y C. Clavijo (CC)depositado en MNHNM y 4) colección privada de J.C.Zaffaroni (JCZ, Montevideo), de la cual material repre-sentativo será depositado en MNHNM.
Se consideraron las siguientes divisiones: 1) Río de laPlata (RdlP) y otros ambientes estuariales: costa de Mon-tevideo, Canelones y Maldonado (hasta Punta Negra),así como desembocaduras de arroyos y lagunas costeras,tanto en el RdlP como en la costa atlántica. Se incluyenfondos arenosos, fangosos, rocosos y de fango consoli-dado, 0-10 m; 2) Playas arenosas atlánticas: desde Pun-ta Negra hasta Barra del Chuy, 0-10 m; (3 Sustratosconsolidados atlánticos: Punta Negra a límite Uruguay-Brasil, 0-50 m, incluyendo fondos rocosos, fangos conso-lidados, bancos someros de mitílidos (BMS) y bancos deMytilus edulis de profundidad (BMP); y 4) Sustratosinconsolidados atlánticos: RdlP exterior a límite Uru-guay-Brasil, 11-50 m, existiendo enclaves fangosos enzonas más someras que son detalladas. Los grupos deespecies mayormente formados por microgasterópodosque poseen vínculo partícular con otros invertebradosbentónicos son discutidos separadamente.
Géneros o especies mencionadas por primera vez parala costa uruguaya son indicadas con el símbolo “¤”. Elregistro detallado y obligatoriamente complementario deestas citas será objeto futuro de publicación.
RESULTADOS Y DISCUSIÓNRdlP y otros ambientes estuariales
Cuatro especies de Heleobia (Cochliopidae) han sidodescritas para la costa de Montevideo: Heleobia australis,Heleobia charruana, Heleobia conexa y Heleobia isabelleana
(d´Orbigny 1840; Gaillard 1974). Salvo esta última, lasotras especies nominales se encontrarían desde elintermareal (todos los sustratos) hasta los fangossubmareales (Figueiras 1964; Gaillard 1974; Scarabino etal. 1976; Venturini et al. 2003; FS). Heleobia spp. es recu-rrente en ambientes con baja salinidad de toda la costauruguaya (todos los sustratos), con registros paraPiriápolis, Punta del Este y Puerto de La Paloma, losarroyos Pando, Solís Grande, Maldonado y Valizas y laslagunas Garzón, José Ignacio y Rocha (Gaillard 1974;Nion 1979; Bier 1985; Pintos et al. 1991; Jorcin 1999;Muniz & Venturini 2001; Passadore & Giménez 2003;Giménez et al. 2005; Demicheli & Scarabino en este volu-men; FS). Heleobia robusta, descrita de las lagunas costerasde Rio Grande do Sul (Brasil) (da Silva & Veitenheimer-Mendes 2004a), probablemente habite también las lagu-nas costeras uruguayas. La variabilidad de las especiespropuestas, distribución e identidad de los registros parala costa uruguaya (generalmente “Heleobia australis”) nohan sido adecuadamente caracterizados o verificados(ver también Castro et al. en este volumen).
Parodizia uruguayensis, recientemente atribuida aPyramidellidae (da Silva & Veitenheimer-Mendes 2004b;Scarabino 2004b), se halla viviente en la costa de Monte-video sobre sustrato fangoso (Scarabino & Masello 1996).La descripción original de esta especie (Pereira de Medina1959) y conchillas halladas en la resaca de la PlayaCarrasco (JCZ) indican que otro Pyramidellidae, hasta elmomento confundido con P. uruguayensis, se encuentraviviente en la zona.
La exótica Myosotella myosotis (Ellobiidae) habitaríala desembocadura del Arroyo del Potrero (Maldonado)y/o hábitat similares de la Ensenada del Potrero(Orensanz et al. 2002). Sin embargo no ha sido halladaviva in situ aún y su presencia es conocida por conchillasdetectadas en la Playa Portezuelo (Zaffaroni 1994;MNHNM).
Cylichnella bidentata (Cylichnellidae) había sido men-cionado para la costa de Montevideo (Playa Carrasco,Figueiras & Sicardi 1974), pero no había sido confirmadaviviente en esta zona, existiendo la posibilidad de que setratara de material subfósil (Sprechmann 1978). Sin em-bargo, ha sido registrada viviente en fondos fangosospróximos a la Isla de Flores (FS).
Los Volutidae Pachycymbiola brasiliana y Zidonadufresnei ocurren marginalmente en el RdlP. Mientras quela primera muestra cierta tolerancia a las bajas salinidadesdel estuario (salinidad a partir de 16), el RdlP parecerepresentar un ambiente subóptimo para la segunda(Carranza et al. en prensa).
El Muricidae invasor Rapana venosa fue detectado enesta zona en 1999 ocupando fondos submareales fango-sos y microsustratos duros que colonizó rápidamente(Scarabino et al. 1999; Carranza et al. en prensa; FS); apartir de 2004 invadió zonas más someras de la costa Ede Canelones y costa de Maldonado ocupando una granvariedad de sustratos (Karumbé com. pers; J. Choccacom. pers.; A. Reboledo com. pers.).

F SCARABINO, JC ZAFFARONI, A CARRANZA, C CLAVIJO & M NIN
145
Buccinanops deformis1 (Nassariidae) es frecuente enfondos fangosos y arenosos frente a Montevideo y Cane-lones (Barattini 1951; Figueiras & Sicardi 1968; FS), mien-tras que la presencia de Buccinanops cochlidium2 es muypuntual (MNHNM; FS).
Siphonaria lessoni (Siphonariidae) esta presente en laspuntas rocosas del área de Piriápolis (Maytía & Scarabino1979) aunque su mayor incursión hacia el W es probable.
El registro de Anachis moleculina (i. e. Costoanachissertulariarum) (Columbellidae) para la Laguna de Castillos(Jorcin 1999) debe ser verificada.
Los registros de Burch & Burch (1964) paraOlivancillaria contortuplicata (Olividae) y Beu (1978) paraFusitriton magellanicus magellanicus (Ranellidae) conlocalidad “Montevideo” son claramente incorrectos,basadas en la referencia del envío del material. Ambasson especies estrictamente marinas conocidas paraambientes muy distantes en la costa y plataformauruguayas respectivamente.
Playas arenosas atlánticasOlivancillaria vesica (Olividae) y Buccinanops duartei
(Nassariidae) viven en la zona de rompiente y se acercanhacia la zona de barrido en los meses de verano, en pla-yas disipativas (Scarabino et al. 1974; Escofet et al. 1979;Demicheli 1987; Riestra et al. 1997; Defeo et al. 1992;1997; FS; AC). Olivancillaria vesica es más tolerante que B.duartei a intermedias, sin llegar a presentarse en playastotalmente reflectivas; el límite de la distribuciónsubmareal de O. vesica es desconocido, mientras se po-seen registros de B. duartei para 5 m de profundidad(AC). La primera tiene su límite de distribución SW enPlaya Brava (Punta del Este), mientras que sólo existenregistros de conchillas al S de Playa Anaconda para lasegunda (FS; JCZ; Scarabino 1984).
A diferencia de las dos especies anteriores,Olivancillaria contortuplicata, Olivancillaria teaguei y Olivellaformicacorsii (Olividae) (Fig. 1) son muy escasas actual-mente y tienen su distribución restringida a la franja Ba-rra del Chuy-La Coronilla, habitando asimismo la zonade barrido y zona de rompiente, aunque se carece deinformación detallada sobre su presencia en la playasubmarina (Klappenbach 1962; 1964a; Figueiras &Sicardi 1973; Riestra et al. 1997; Defeo et al. 1997; Scarabino2004a).
1 B. globulosus probablemente es un ecotipo marino y/o de fondosarenosos de B. deformis, habiédose detectado ejemplares intermedios; elmaterial tipo de esta última esta compuesto por una conchilla de cadamorfotipo (FS obs. pers.). La cita de B. duartei para el Río de la Plata(Giberto et al. 2004) esta basada en este especie.
2 Buccinanops gradatus fue erróneamente listada como especie dis-tinta por Scarabino (2004b); Allmon (1990) estableció su sinonimiacon B cochlidium.
3 Esta especie ha sido referida para Uruguay y la Provincia deBuenos Aires (Argentina) como Parvanachis paessleri (e. g. Castellanos& Fernández 1967; Figueiras & Sicardi 1973a). Costa (2005) ilustra elmaterial tipo de esta especie (sinonimizándola con Parvanachisisabellei); no parece existir un nombre disponible para Parvanachis sp.,que a pesar de la variabilidad considerada para P. isabellei (ver Costa2005), se separa netamente de esta última siendo inclusive simpátricas.
Tienen presencia ocasional en la zona de barrido yrompiente de playas disipativas los gasterópodosOlivancillaria urceus, Olivancillaria uretai y el ConoideaTerebra gemmulata (Escofet et al. 1979; Scarabino 1984;FS), que poseen poblaciones en el submareal, donde sesuman Olivancillaria deshayesiana, Olivella puelcha, Olivellaplata, Buccinanops uruguayensis, Buccinanops monilifer yBuccinanops cochlidium (Scarabino et al. 1974; Escofet et al.1979; Demicheli 1987; Demicheli & Scarabino en estevolumen). Estas especies ocurren en el banco arenoso delPuerto de La Paloma en el submareal más somero, pro-ducto de condiciones hidrodinámicas particulares dellugar (Demicheli & Scarabino en este volumen).Pachycymbiola brasiliana también ocupa el submareal delas playas arenosas (Escofet et al. 1979), con registros apartir de 5 m (A. Segura com. pers.; Riestra et al. 2000).
Buccinanops globulosus (ver B. deformis) vive en la Pla-ya Portezuelo a partir de 4 m (Demicheli 1986).
Parvanachis sp.3 (Columbellidae) ha sido registradoviviente en Playa Anaconda y playas de Cabo Polonio(Figueiras & Sicardi 1973a), probablemente producto desu arrastre desde el submareal somero, donde esta espe-cie habita en el litoral de Rio Grande do Sul (Brasil) (FSobs. pers.).
Phalium granulatum (Tonnoidea), tiene su límite surde distribución en Uruguay con pseudopoblaciones quepueden no ser permanentes en la costa de Rocha (FS;Demicheli com. pers.).
La presencia permanente de R. venosa en el submarealsomero de Playa Mansa (Bahía de Maldonado) (M.Zarucki com. pers.) es incierta, pudiéndose tratar de ejem-plares arrastrados.
Sustratos inconsolidados atlánticosLos fondos de arena mayormente fina y fangosa al-
bergan más especies de gasterópodos que los fondosfangosos costeros.
Buccinanops uruguayensis, B. monilifer y Olivancillariacarcellesi habitan el intervalo 10-25 m (Klappenbach 1965;Milstein et al. 1976; FS), aunque esta última ha sido de-tectada también en fondos protegidos más someros(ver Demicheli & Scarabino en este volumen), así comoen profundidades de ca. 30 m (JCZ). Estas especies estanasociadas con arenas y arenas fangosas (FS).
Figura 1. Olivacillaria contortuplicata, Olivancillaria teaguei y Olivellaformicacorsii (de izquierda a derecha), olívidos de distribución restrin-gida en la costa uruguaya con aparentes problemas de conservación.Tomados de Scarabino (1984).

Gasterópodos marinos y estuarinos de la costa uruguaya:faunística, distribución, taxonomía y conservación
146
Tabla 1. Cefalaspídeos registrados en el área de estudio.
Klappenbach (1965) cita a O. uretai para 10-15 m peroeste dato parecería incorrecto de acuerdo a los hallazgosposteriores.
Tanto próximos a la costa (10-30 m) como hacia 35-50 m fueron halladas O. urceus, T. gemmulata, Conus clenchi(Conidae) y Notocochlis isabelleana (Naticidae), mientrasque Solariella patriae (Trochidae), Amalda josecarlosi(Olividae), Nassarius tango, Nassarius scissuratus(Nassariidae), “Pleurotomella” aguayoi, Carinodrilliabraziliensis, “Drillia” rioensis y Terebra doellojuradoi(Conoidea) ocuparían únicamente los sectores más pro-fundos de estos fondos (Carcelles 1953; Mol et al. 1967;Figueiras & Sicardi 1973a; 1980; Amaro 1974; Juanicó &Rodríguez-Moyano 1976; Rodríguez-Moyano 1976;Milstein et al. 1976; Layerle & Scarabino 1984; verScarabino 2004b; JCZ; MNHNM). La distribución de es-tas especies en la plataforma interna uruguaya es pococonocida y claramente heterogénea, siendo necesario de-terminar las variables que determinan la misma.
La asociación descrita por Cachés (1980) para fon-dos fangosos incluye a los siguientes gasterópodos: B.cochlidium, Parvanachis sp., Polystira formosissima(Conoidea), Prunum martini (Marginellidae), Z. dufresnei,así como Tectonatica pusilla (Naticidae) (Watson 1886;ver Klappenbach 1967; FS; Zaffaroni en prensa) comovarios cefalaspídeos (FS, Tabla 1). Esta asociación exis-tía como un enclave dentro del Puerto de La Paloma(Demicheli & Scarabino en este volumen) aunque sólo sedetectó a B. cochlidium y Parvanachis sp. Ninguna de es-tas especies es exclusiva de dicha asociación; todas tie-nen amplia distribución en el área a partir de 20 m yocupan también fondos areno-fangosos (FS).
Tonna galea y Cymatium parthenopeum, dos Tonnoideaque tienen su límite S de distribución atlántica en Uru-guay, habitan a partir de ca. 20 m, sobre fondosarenofangosos o fangosos (Juanicó & Rodríguez-Moyano1976; Carranza et al. en prensa).
Pachycymbiola brasiliana, Z. dufresnei y B. cochlidiumhabitan fondos arenosos o fangosos y han sido registra-das para todo el rango de profundidades, inclusive haciael submareal de zonas protegidas (Watson 1886; Carcelles& Parodiz 1939; Juanicó & Rodríguez-Moyano 1976;Milstein et al. 1976; Riestra et al. 2000; Nion 1979; Fabianoet al. 2000; Carranza et al. en prensa). Pachycymbiolabrasiliana fue explotada artesanalmente en la costa deRocha (ca. 11-12 m) en forma muy intensa en 1991-1993,al punto de su sobrexplotación (Fabiano et al. 2000; Riestraet al. 2000; Masello 2000). Zidona dufresnei fue explotadaindustrialmente desde 1994 en la zona más profunda delárea (Fabiano et al. 2000). Las profundidades a la cual sedesarrolló la pesca en el área coinciden con la zonas demayor ocurrencia de cada especie (P. brasiliana: 10-20 myZ. dufresnei: 30-40 m) (AC y FS obs. pers.).
Adelomelon beckii (Volutidae) es una especieestenoterma y estenohalina que usualmente habita pro-fundidades mínimas de 30 m, aunque puntualmente sela ha registrado en zonas má someras (Juanicó &Rodríguez-Moyano 1976; Scarabino 2004; Carranza et al.en prensa; FS).
La presencia de R. venosa en estos fondos está por elmomento vinculada al RdlP exterior (FS), y a la zona dePiriápolis y Punta Ballena a partir de 2005 (J. Chocca,com. pers.; A. Reboledo com. pers.); su avance haciazonas más oceánicas es incierta y los hallazgos de pues-
Familia Especie Hallazgo/referencia Observaciones
Acteonidae Acteon sp.Hallazgo viviente en 35º30´S-55º00´W (M.Aguilera, JCZ; MNHNM)
A comparar con Acteon pelecais
Retusidae Pyrunculus caelatus
Conchillas en resaca de playas de Maldonado yRocha (Barattini 1951; Figueiras & Sicardi 1974);registro viviente en 19 m (Layerle & Scarabino1984)
Retusidae Volvulella persimilis
Conchillas en resaca de playas de Maldonado yRocha (Cachés 1973; Figueiras & Sicardi 1974;JCZ); amplia distribución en el área (11-41 m,fondos arenosos, areno-fangosos y fangosos;Cachés 1973; Layerle & Scarabino 1984; FS)
Referencia de Layerle & Scarabino(1984) puede incluir tambiénVolvulella sp.
Retusidae Volvulella sp.Conchillas en resaca de playas de Maldonado yRocha (Figueiras & Sicardi 1974; 1980; JCZ)
CylichnidaeCylichnella bidentata
Conchillas en resaca de playas de Montevideo,Maldonado y Rocha (Figueiras & Sicardi 1974);distribución relativamente amplía en el área (11-40 m, Layerle & Scarabino 1984); distribuciónviviente en estuario confirmada y en fondosfangosos y areno-fangosos (FS)
Citada por Barattini & Ureta (1961)como Cylichna crispula (LPB)
Cylichnidae Acteocina candei
Conchillas en resaca de playas de Maldonado yRocha (Barattini & Ureta (1961; Zaffaroni 1991);amplia distribución viviente en el área (18-44 m,Layerle & Scarabino 1984). Fondos fangosos yareno-fangosos (FS)
Referencia de Layerle & Scarabino(1984) podría incluir a Acteocinarecta
Cylichnidae Acteocina rectaConchillas en resaca de Playa Portezuelo yBahía de Maldonado (Zaffaroni 1991)
Concepto en necesidad de revisión
Philinidae Philine spp.?25-40 m, fango (Layerle & Scarabino 1984; FS);pocos registros puntuales

F SCARABINO, JC ZAFFARONI, A CARRANZA, C CLAVIJO & M NIN
147
tas completas realizados antes de 2004 en Bahía deMaldonado, así como de puestas vacías en 1999 y 2006para la Barra de Maldonado y La Paloma (Scarabino etal. 1999; FS) parecerían corresponder a materiales arras-trados.
Los registros de Phalium labiatum iheringi dentro delárea son escasos y generalmente basados en conchillasvacías y rodadas (Barattini & Ureta 1961, como Semicassisachatina; Figueiras & Sicardi 1972; Juanicó & Rodríguez-Moyano 1976; Demicheli & Scarabino en este volumen);su presencia en el área y el concepto de esta subespecie(Abbott 1968) requieren revisión.
El único registro del nudibranquio Marionia cucullata(Tritoniidae) para el área corresponde a Berg (1884; verOdhner 1926); muestreos realizados en el mismo lugarde ese hallazgo (estación 321 del “Challenger”, fondofangoso) y en el resto del área no permitieron colectarnuevamente esta especie, que en cambio ha sido captu-rada a mayor profundidad y en fondos areno-fangosos,fuera del área de estudio (FS).
Natica sp. ha sido mencionada por Figueiras & Sicardi(1980). Se trata de una especie claramente distinta de N.isabelleana; se han obtenido pocos ejemplares frente a lacosta de Rocha (JCZ). La correspondencia de esta especiecon el registro de Layerle & Scarabino (1984) es incierta.
El estatus de Halistylus circumstriatus con respecto aHalistylus columna (Trochidae) (Pilsbry 1897a), no ha sidorevisado. Se trata de tróquidos asociados a granulometríagruesa y probablemente también fondos rocosos, al igualque “Photinula” blakei (Trochoidea) y el columbélidoAesopus sp. (ver Costa 2005), con registros vivientes has-ta los 25-30 m (Layerle & Scarabino 1984; FS).
Varias especies con larvas teleplánicas reclutan apa-rentemente en forma ocasional en fondos de al menos 20m de la costa uruguaya. En este sentido, se ha detectadofrente a La Paloma el Ovulidae Pseudocyphoma intermedium(Figueiras & Sicardi 1972) y Tonna berthae (¤, JCZ).
Al menos cinco especies de Olivella habitan estos fon-dos: Olivella riosi, Olivella orejasmirandai, Olivella plata,Olivella puelcha y Olivella tehuelcha (Castellanos &Fernández 1965; Figueiras & Sicardi 1973b; Klappenbach1991a; Demicheli & Scarabino en este volumen; FS; JCZ).Klappenbach (1991b) aclaró el estatus de las dos últi-mas, que generalmente ocurren en sedimentos distintos.Mientras O. puelcha junto a O. plata poseen preferenciapor arena fina y eventualmente fangosa, O. tehuelcha ocu-pa fondos de arena gruesa con distintos niveles deconchillas (FS; Milstein et al. 1976). Existe poca informa-ción sobre O. riosi y O. orejasmirandai, aunque se trata deespecies habitantes de fondos más profundos que el res-to de la especies y con grados importantes de fango(Klappenbach 1991a; FS; JCZ). Figueiras & Sicardi (1980)y Layerle & Scarabino (1984) mencionan especies inde-terminadas de Olivella, parte de las cuales podrían co-rresponderse con O. riosi y O. orejasmirandai.
Se han mencionado dos especies de Caecum (Caecidae)desprovistas de escultura anillada para la costa urugua-ya (Klappenbach 1964b; Figueiras & Sicardi 1980; Layerle
& Scarabino 1984; Scarabino & Zaffaroni 2004); mien-tras Figueiras & Sicardi (1980) las refieren como Caecumcf. achironum y Caecum aff. antillarum, Layerle & Scarabino(1984) las refieren definitivamente a estas especies. Laidentidad de la primera coincide con Caecum someri (¤), yla de la segunda con Caecum striatum (¤), de acuerdo conlas ilustraciones y comparaciones de Gomes & Absalão(1996) y Absalão & Gomes (2001). Ambas especies habi-tan fondos generalmente arenosos (FS; Obenat et al. 2001),existiendo para C. striatum registros para hasta profun-didades de ca. 30 m.
Barleeia rubrooperculata y Rissoa cruzi (Rissoidea) hansido referidas para los sectores más profundos del área(Figueiras & Sicardi 1980; Layerle & Scarabino 1984) ypara alguna localidad puntual más somera que debeconfirmarse. Layerle & Scarabino (1984) refieren para 27m frente al RdlP exterior a Alvania sp. Figueiras & Sicardi(1980) refieren como Cingula (Microdochus) sp. a conchillashalladas en la resaca de la costa de Maldonado y Rocha;no se ha podido corresponder esta referencia con hallaz-gos posteriores de microgasterópodos en el área.
La presencia de conchillas únicamente desgastadasde Finella dubia (Scaliolidae) en la resaca de Portezuelo yBahía de Maldonado (JCZ) hacen incierta su presenciaviviente en el área, la cual fue reportada por Figueiras &Sicardi (1980), registrando también su hallazgo en lacosta de Rocha. Sin embargo, Layerle & Scarabino (1984)mencionan un único registro viviente para ca. 40 m quedebe ser confirmado.
Spirotropis patagonica (Conoidea), ampliamente dis-tribuida en el área, ha sido referida viviente a partir de 15m; se asocia con fondos arenosos y areno-fangosos(Layerle & Scarabino 1984; FS).
La presencia viviente y el concepto del Columbellidaereferido como Mitrella rubra para la costa uruguaya enbase a conchillas halladas en la resaca de Maldonado yRocha (Figueiras & Sicardi 1980; JCZ; MNHNM) soninciertas y claramente requieren revisión.
Un skeneiforme indeterminado (ver Scarabino &Zaffaroni 2004) ha sido detectado en la resaca de LaPaloma, Bahía de Maldonado y Portezuelo (Figueiras &Sicardi 1980; JCZ).
Los registros de Aclis sp. para tres localidades delárea (Figueiras & Sicardi 1980; Layerle & Scarabino 1984)deben ser revisados y pueden no estar basados en ejem-plares de Aclididae dado el avance de los conceptos rela-cionados a este grupo (ver e.g. Bouchet & Warén 1986;Warén 1993; 1994; Van der Linden & Moolenbeek 2004).
Se han detectado cuatro especies de Vitrinellidae enel área (ver Figueiras & Sicardi 1980; Layerle & Scarabi1980; JCZ; FS), aunque es muy factible la existencia deotras dada su pequeña talla y hábitos crípticos (verBieler & Mikkelsen 1988 y referencias allí). Tres de estasespecies han sido halladas vivientes a partir de ca. 20 men el área, requiriendo esfuerzos particulares de caracte-rización morfológica y taxonómica.
Macromphalina argentina (Vanikoridae) ha sido repor-tada viviente en el área de una única localidad a 36 m de

Gasterópodos marinos y estuarinos de la costa uruguaya:faunística, distribución, taxonomía y conservación
148
profundidad (Layerle & Scarabino 1984); conchillas hansido registradas en la resaca de La Paloma y Bahía deMaldonado (Figueiras & Sicardi 1980).
Sustratos consolidados atlánticosTodas las especies registradas que habitan a partir
del submareal han sido verificadas o es muy factible supresencia en microsustratos duros sobre fondos deconchilla, arenosos o fangosos.
Las poblaciones de Littorinidae que se hallan presen-tes en el intermareal alto y hasta supralitoral entre laPunta de La Coronilla (Rocha) y Punta del Este (Playa delos Ingleses) han sido atribuidas a Echinolittorina ziczac(Bequaert 1943; Barattini & Ureta 1961; Figueiras &Sicardi 1972; Maytía & Scarabino 1979; Scarabino 1984;Batallés et al. 1985). Bandel & Kadolsky (1982) conside-ran para Uruguay (Cabo Polonio) a Echinolittorinalineolata. La existencia del “complejo ziczac”, donde tresespecies se distinguen en base a diferencias sutiles(Absalão & Roberg 1999; Absalão & Louro 2002, perover Andrade et al. 2003), hacen necesario la revisión delos Littorinidae de Uruguay.
Lottia subrugosa (Lottiidae) y S. lessoni son las lapasintermareales que han sido identificadas en sustratosduros de la costa atlántica uruguaya, entre Punta Balle-na y Punta de La Coronilla, incluyendo Isla Gorriti e Islade Lobos (d´Orbigny 1840; Métivier 1967; Amaro-Padilla1967; Maytía & Scarabino 1979; Neirotti 1981; Batallés etal. 1985). Sin embargo, no se ha efectuado hasta el mo-mento una revisión de la primera especie, que puedecomprender dos (Figueiras & Sicardi 1980; Scarabino2004b).
La lapa submareal Diodora patagonica (Fissurellidae)habita en oquedades y en la cara inferior de piedras des-de el submareal somero hasta un límite de profundidadprobablemente inferior a 30 m (Scarabino 1984; Demicheli& Scarabino en este volumen; FS).
El Muricidae Stramonita haemastoma está presente conpoblaciones reproductivas desde el submareal somerohasta los BMP. Su presencia y reproducción ha sido veri-ficada para Punta de La Coronilla, Cerro Verde (PuntaLoberos), Santa Teresa, Punta del Diablo, Cabo Castillo,Cabo Polonio y La Paloma, Barra de Maldonado y LaVirgen (Punta del Este) ( Juanicó & Rodríguez-Moyano1976; Milstein et al. 1976; Batallés et al. 1985; Demicheli& Scarabino en este volumen; FS; CC; AC; P. Miller, com.pers; P. Etchegaray, com. pers.). La presencia deStramonita floridana es ocasional (Figueiras & Sicardi 1973;1980; FS; JCZ) y probablemente ocurre en la costa uru-guaya en forma de pseudopoblaciones.
La situación de R. venosa en fondos inconsolidadosatlánticos se aplica asimismo para fondos consolidados;la detección de ejemplares de esta especie dentro de laBahía de Maldonado se efectuó en 2004 (R. Fontainacom. pers; JCZ; FS).
Hanetia haneti (Muricidae) ocurre con ejemplares ais-lados en el submareal más somero (BMS) y probable-mente no se distribuye más allá de 20 m (Pilsbry 1897b;
Batallés 1983; Riestra et al. 1992; Demicheli & Scarabinoen este volumen; FS; AC). Sus puestas son halladas en laresaca de zonas rocosas entre Punta de La Coronilla yPlaya de los Ingleses (FS; CC).
Bostrycapulus odites (Calyptraeidae) vive desde elsubmareal somero hasta 35-50 m (BMP) (Parodiz 1939;Milstein et al. 1976; Juanicó & Rodríguez-Moyano 1976;Layerle & Scarabino 1984; FS). Todas las referencias pre-vias han sido referidas como Crepidula aculeata; se sigueaquí a Collin (2005), quien restringe Bostrycapulus aculeatuspara el Atlántico Noroccidental. Bostrycapulus odites hasido registrado en Brasil, Argentina y Sudáfrica. Su pre-sencia en Uruguay requiere ser confirmada medianteanálisis moleculares y de morfología fina (protoconcha);no se descarta asimismo la existencia de otras especiesdel género en el área dado su potencial de dispersióncomo fouling.
Costoanachis sertulariarum (Columbellidae) es comúnen fondos submareales someros, principalmente en lacosta de Rocha, donde en verano es frecuente hallar suspuestas sobre una variedad de microsustratos, en parti-cular hidrozoarios y algas (Zaffaroni 2000; Demicheli &Scarabino en este volumen; FS; CC; AC). Su presencia deextendería hasta al menos ca. 25 m (Layerle & Scarabino1984; FS). Parvanachis isabellei ha sido hallado en elsubmareal somero de Cerro Verde (FS); su talla reduciday su asociación han aparentemente limitado el númerode registros vivientes en el área.
Tegula patagonica (Trochidae) se extiende desde elsubmareal (por los menos 1 m) hasta los BMP (Milsteinet al. 1976; Juanicó & Rodríguez-Moyano 1976; Demicheli& Scarabino en este volumen).
La liebre de mar Aplysia brasiliana (Anaspidea) ocu-rre en forma abundante en años con influencia inusual deaguas cálidas (JCZ obs. pers.; M. Demicheli com. pers.).Información proporcionada por pescadores indica quepor lo menos algunos individuos sobreviven durante elinvierno en la costa de Rocha, habiéndose encontradoejemplares en Valizas y Punta de La Coronilla (P. Miller,com. pers.; C. Romero, com. pers.).
La única especie de nudibranquio citada para el áreay hábitat es Polycera marplatensis (Riestra et al. 1992). Sinembargo, al menos otras tres especies de nudibranquioshan sido halladas y se encuentran en estudio (FS).
Fissurellidea megatrema (Fissurellidae), Calliostomajucundum, Calliostoma carcellesi, Calliostoma coppingeri(Calliostomatidae), Cabestana felipponei (Ranellidae) yFusinus frenguellii (Fasciolariidae) han sido ubicados enBMP o zonas cercanas (Juanicó & Rodríguez-Moyano1976; Figueiras & Sicardi 1980). De estas especies, C.coppingeri, C. jucundum y C. felipponei así como Muricopsisnecocheana (Muricidae), ocuparían preferentementesustratos consolidados a partir de ca. 20 m de profundi-dad (M. Aguilera com. pers.; FS). Urosalpinx cala(Muricidae) posee una distribución similar pero a partirde zonas más someras (Zaffaroni 2000; M. Aguilera com.pers.). Lucapinella henseli (Fissurellidae) es mayormenteconocida de conchillas halladas en la resaca (Figueiras &

F SCARABINO, JC ZAFFARONI, A CARRANZA, C CLAVIJO & M NIN
149
Sicardi 1971; JCZ; FS; MNHNM) y se posee un únicoregistro viviente para el área, aunque sin detalle de pro-cedencia (JCZ). Todas estas especies conforman un gru-po cuya distribución en el área es muy poco conocidadebido a la falta de muestreo en fondos consolidados.Calliostoma militare sería también una de estas especies,aunque particularmente enigmática en la zona: se cono-cen unas pocas conchillas de Bahía de Maldonado y LaPaloma (MNHNM; FS; JCZ).
Trophon pelseneeri y Trophon patagonicus (Muricidae)habitan los BMP, aunque su presencia en estos no esexclusiva (Juanicó & Rodríguez-Moyano 1976; Figueiras& Sicardi 1980; Pastorino 2005a; FS; JCZ).
Las lapas del género Crepidula (Calyptraeidae), queposeen particular variabilidad fenotípica y diversidadcríptica (ver Collin 2000; Simone et al. 2000; Simone 2006y referencias allí), no han sido estudiadas en detalle enaguas uruguayas. Crepidula protea está presente desde elsubmareal somero hasta al menos 22 m (Milstein et al.1976; Hoagland 1983; Layerle & Scarabino 1984; FS).Ejemplares de morfología similar a Crepidula argentina yCrepidula intratesta se hallan en los BMP y en conchillaspagurizadas por Dardanus insignis (Decapoda: Anomura)a partir de 20 m (Juanicó & Rodríguez-Moyano 1976;Hoagland 1983, como Crepidula plana; FS).
Calyptraea centralis (Calyptraeidae), citada sin detallepor Figueiras & Sicardi (1980) para Uruguay, ha sidodetectada viviente en fondos de conchilla en Isla de Lo-bos (10 m) y frente a José Ignacio (25 m) (FS) y susconchillas son frecuentes en la resaca de La Paloma yPunta del Este (JCZ). Esta especie no ha sido revisadataxonómicamente y podría formar un complejo de espe-cies no reconocidas aún. Los registros de Trochita pileoluspara el área (Juanicó & Rodríguez-Moyano 1976; Layerle& Scarabino 1984) podrían estar basados en C. centralis,tal como lo plantean para al menos parte de su conceptoprevio Figueiras & Sicardi (1980).
Microgasterópodos asociados a invertebradosbentónicos (fondos consolidados e inconsolidados)
De acuerdo con lo conocido para esta familia (verKosuge 1966), la presencia de Triphoridae en la zona(Figueiras & Sicardi 1972) se vincularía a poríferos, loscuales se encuentran mayormente asociados a fondosconsolidados. Aun cuando no se ha muestreadoespecíficamente, es incierta su presencia en el área ya quesólo se han detectado conchillas vacías siempre con cier-to grado de desgaste (JCZ; FS), pudiéndose tratar dematerial subfósil. Bouchet (1984) discute las limitacio-nes para la determinación de Triphoridae únicamentesobre la base de conchillas. Triphora medinae y “Triphorapulchella” han sido mencionadas para el Puerto de LaPaloma, así coma para Playa Portezuelo y Bahía deMaldonado en el segundo caso (Figueiras & Sicardi 1972;JCZ). “Triphora pulchella“ (non Cerithium pulchellum C. B.Adams, 1850, un Cerithiopsidae) es un nombre aplicadoincorrectamente a Triphoridae del Atlántico Occidental.Rojas & Martínez (1999) asignan materiales propios pro-
venientes de Punta Rasa, coincidentes en morfología ge-neral con los registrados por Piñeiro et al. (1992), aMarshallora nigrocinta.
Los Cerithiopsinae ocuparían el mismo hábitat quelos Triphoridae (ver Marshall 1978), pero en cambio sehan detectado en buen estado conchillas de dos especiesen Maldonado y Rocha (JCZ; MNHNM). Una fue men-cionada para el Cuaternario por Piñeiro et al. (1992) comoCerithiopsis aff. emersoni. La otra especie corresponde a lamencionada sin detalle para la costa uruguaya porFigueiras & Sicardi (1972) como Cerithiopsis greeni. Estaidentidad debe confirmarse en el área, al igual que suestatus en todo el Atlántico Sudoccidental. El materialregistrado por Layerle & Scarabino (1984) comoAtaxocerithium pullum para 22 y 43 m podría correspon-der a alguna de estas dos especies por razonesbiogeográficas. Seila adamsii (Cerithiopsidae) ha sido men-cionada sin detalle para la costa uruguaya por Figueiras& Sicardi (1980). El material colectado de este géneroconsiste en conchillas halladas en la resaca de PlayaPortezuelo, Bahía de Maldonado y La Paloma (JCZ;MNHNM) y es asignado preliminarmente a esta especie,cuyo concepto debe revisarse dada la probable existenciade un complejo de especies (ver Rolan & Fernandes 1990).
Los Epitoniidae son taxonómica y morfológicamentediversos en el área y consecuentemente han sido pocoestudiados o merecen revisiones detalladas. Su caracte-rística asociación con cnidarios, especialmente Actiniaria(ver e.g. Robertson 1981; 1983 y referencias allí), implicasu presencia en diversos fondos, aunque tienden a serinusuales en aquellos estrictamente fangosos (limo-arci-lla) (FS). Epitonium georgettinum y Epitonium striatellumson las especies de mayor talla y propias del AtlánticoSudoccidental, y al igual que Epitonium denticulatum,Epitonium arnaldoi, Epitonium candeanum, Epitonium aff.unifasciatum y Epitonium albidum han sido mencionadospara la costa uruguaya (Clench & Turner 1951; 1952;Figueiras & Sicardi 1972; Layerle & Scarabino 1984; verScarabino 2004b). Salvo E. arnaldoi (para la cual sin em-bargo también se ha propuesto su sinonimización conEpitonium angulatum, especie del Caribe), estas últimasespecies han sido descritas para el Caribe y costa deUSA. Es necesario revisar el concepto actual de las mis-mas a lo largo de su distribución propuesta en el Atlán-tico Occidental. Gyroscala xenicima (¤, Epitoniidae) pare-ce tener reclutamientos ocasionales en el área segun unospocos registros de conchillas para La Paloma y Portezuelo(JCZ; MNHNM). La distribución de esta especie es lacosta uruguaya es coincidente con lo conocido para otrasáreas donde ha sido registrada (ver García 2003).
Figueiras & Sicardi (1972; 1980) registraronsomeramente varias especies de Eulimidae pertenecien-tes a Eulima y Melanella, en general basados en conchillasen mal estado (JCZ). Unos pocos registros adicionales deestos géneros provienen del área a partir de 20 m (Layerle& Scarabino 1984; JCZ; FS). Figueiras & Sicardi (1980)agregan el hallazgo de conchillas de una especie que atri-buyen a Sabinella en la resaca de Punta Ballena. Se ha

Gasterópodos marinos y estuarinos de la costa uruguaya:faunística, distribución, taxonomía y conservación
150
registrado Microeulima sp. (¤) en muestras de fondoextraidas en Isla de Lobos y en la resaca de la Bahía deMaldonado (FS; JCZ). Cachés (1973) citó Eulima bifasciataen el contenido estomacal de asteroideos capturados enel Banco del Plata (30 millas al S de Punta del Este),sobre fondo arenoso. Los Eulimidae, típicos parásitos deequinodermos, están pobremente conocidos a nivel mun-dial en relación a su abundancia; los registros uruguayosrequieren revisión basados en material colectado vivoque permita un estudio más adecuado (Warén 1992;Warén com. pers.).
Los Pyramidellidae son particularmente diversos ypoco estudiados en el área (ver Figueiras & Sicardi 1974;1980). Robertson & Mau-Lastovicka (1979) y referenciasallí, discuten a asociación de las especies de esta familiacon otros macroinvertebrados bentónicos. El géneroTurbonilla (s. l.) está representado en la costa uruguayapor las siguientes especies previamente citadas: Turbonillaaracruzencis, Turbonilla atypha, Turbonilla dispar, Turbonillamacaensis, Turbonilla rushii, Turbonilla sanmatiensis yTurbonilla uruguayensis, así como por numerosas otrasbajo estudio (Pilsbry 1897b; Bush 1900; Figueiras &Sicardi 1974; 1980; Castellanos 1982; Layerle & Scarabino1984; Pimenta & Absalão 2001; 2004; JCZ). Turbonillafarinatiae (¤) posee una amplia distribución en el área,mientras que Turbonilla midasi (¤) ha sido registrada pun-tualmente frente a Rocha y Turbonilla paralaminata (¤)para Bahía de Maldonado (conchillas) (JCZ). Otras citaspara el área (T. interrupta, T. americana y T. fasciata) sonerróneas y/o sus conceptos se encuentran en revisión(Pimenta & Absalão 2001a; JCZ). Boonea jadisi y Booneaseminuda han sido registradas en la costa de Maldonadoy Rocha (Figueiras & Sicardi 1974; 1980), incluyendoejemplares vivientes (JCZ). Se han hallado tambiénconchillas coincidentes con Chrysallida multituberculata (¤)(Bahía de Maldonado y Portezuelo) y Chrysallida dux (¤)(La Paloma y Cabo Polonio). Eulimastoma canacaliculatumhabita una variedad de fondos y sus conchillas son usualesen la resaca de Maldonado y Rocha (Figueiras & Sicardi1980; Layerle & Scarabino 1984; JCZ; FS). Scarabino et al.(2003) registraron Pyramidellidae para el submareal so-mero de Cerro Verde (Rocha), que corresponden aEulimastoma. Careliopsis styliformis es únicamente conoci-da en el área a través de conchillas ubicadas en la Bahíade Maldonado (Zaffaroni 1991; FS; ver Pimenta &Absalão 2001b); Eulimella argentina fue registrada en lamisma condición para La Paloma (Figueiras & Sicardi1974). Iselica anomala (Amathinidae) es únicamente co-nocida de conchillas vacías halladas en la resaca de losdepartamentos de Maldonado y Rocha (JCZ; FS).
CONCLUSIONESA nivel nacional son pocas y generalmente prelimina-
res las revisiones taxonómicas y los relevamientosmalacofaunísticos (ver Scarabino et al. 2003).
El rango de profundidad aquí considerado está ocu-pado mayormente por especies de origen subtropical ytemplado, tal como lo establecieron Escofet et al. (1979),
Maytía & Scarabino (1979) y Layerle & Scarabino (1984).Hasta el momento no han sido detectadas en el áreaespecies propias, estando la mayoría de la malacofaunacompartida con regiones inmediatas (litorales de la Pro-vincia de Buenos Aires, Argentina y del Estado de RioGrande do Sul, Brasil) (ver Castellanos 1970; Rios 1994).El pobre conocimiento de numerosos grupos, en especialaquellos de hábitos o distribución más restringida, per-mite plantear como hipótesis de trabajo el hallazgo futu-ro de especies endémicas.
Hasta el momento se han registro al menos 140 espe-cies en el área. Los Olividae (14 spp.) y Nassariidae (7)estan bien representados debido a la diversidad deOlivancillaria (7) y Buccinanops (5), géneros representadosen una variedad de fondos inconsolidados del área yendémicos del Atlántico Sudoccidental.
El RdlP y otros ambientes estuariales son habitadospor 14 especies (Tabla 2) de las cuales seis son propia-mente estuarinas; el género Heleobia domina estos am-bientes.
Tabla 2. Número de especies de Gastropoda presentes en el área segúnalgunos clados y unidades ambientales consideradas. Rdlp y oae=Ríode la Plata y otros ambientes estuariales; Paa=Playas arenosas atlán-ticas; Sia=Sustratos inconsolidades atlánticos; Fac=Sustratos atlánti-cos consolidados; Maib=Microgasterópodos asociados a invertebra-dos bentónicos.
Prioridades y perspectivas de investigaciónDesde un punto de vista taxonómico y faunístico, la
mayor prioridad de investigación de los Gastropoda ma-rinos y estuarinos de la costa uruguaya se encuentra enlos grupos cuyas dimensiones son generalmente inferio-res a 10 mm.
El género Heleobia posee una gran relevancia ecológicaen el área (ver e.g. Castro et al. en este volumen; Muniz etal. en este volumen; Giménez en este volumen; Bonilla etal. en este volumen), lo que sumado a las consideracio-nes efectuadas aquí, lo indican como prioridad máxima.
También son prioridad taxa con diverso grado deasociación y simbiosis con poríferos, antozoos, poliquetos,crustáceos, equinodermos y briozoos (Epitoniidae,Eulimidae, Triphoridae, Cerithiopsidae y Pyramidellidaey Nudibranchia), así como los Trochoidea y Vitrinellidae.Esto implica la necesidad de búsquedas específicas enlos mencionados microhábitat, que se vinculan en mayormedida a sustratos duros, utilizando técnicas especial-mente dirigidas y efectivas (i.e. malla 0.5 mm, rastraWarén, enmalladores bentónicos, buceo, aspiradora, la-vado de sustratos).
Taxón RdlP y oae Paa Sai Sac Maib
Patellogastropoda - - - 1 -
Vetigastropoda - - 5? 8 -
Cerithoidea - - 1 - -
Littorinimorpha 4 - 15? 6? 5
Ptenoglossa - - 2? - 18
Neogastropoda 5 18 27 10 -
Heterobranchia 5 - 9 6 17
Total 14 18 55 31 40

F SCARABINO, JC ZAFFARONI, A CARRANZA, C CLAVIJO & M NIN
151
El uso de técnicas de biología molecular sobre gru-pos ya “estabilizados” en términos de taxonomía alfa(e.g. Fissurellidae, Olividae y Volutidae) es asimismofundamental para detectar posibles especies crípticas.
La necesidad de establecer el estado vivo o muertodel material reportado en cualquier estudio es básico,dado el gran potencial de preservación de las conchillas yla posibilidad consecuente de establecer rangos ecológicoso inclusive faunísticos falsos. Este último se refiere a laposibilidad de considerar especies como vivientes enaguas uruguayas, basados en conchillas subfósiles otransportadas por grampones de la feofita Macrocystispyrifera (ver Scarabino & Zaffaroni 2004; Scarabino &Ortega 2004; Scarabino 2004b).
Los fondos entre 30 y 50 m representan zonas demayor interés para el hallazgo de nuevas especies, mien-tras que el área comprendida entre Montevideo y PuntaNegra, así como el Río de la Plata exterior, lo es paracaracterizar la eurihalinidad de todas las especies. Elrelevamiento del sector José Ignacio-La Paloma permiti-ría inventariar una zona particularmente poco conociday de interés biogeográfico.
Otras prioridades y perspectivas de investigaciónpara el grupo en cuestión han sido discutidas para elresto de los invertebrados bentónicos del área (verScarabino en este volumen).
Implicancias para la conservación y el manejoScarabino (2004a) ha planteado la dificultad de eva-
luación del estado de conservación de la malacofaunamarina y estuarina uruguaya dada la escasez de estu-dios faunísticos, taxonómicos, biológicos y ecológicos,así como la falta de estudios nacionales sobre el impactoantrópico en zonas inter-submareales someras estuarinasy rocosas. Destaca asimismo la necesidad de laimplementación de áreas marinas protegidas y de enfo-ques multiespecíficos y ecosistémicos para las pesque-rías de arrastre, aun cuando se carece de informaciónsobre el impacto de estas últimas en la malacofauna.
Son también de relevancia mayor el manejo adecua-do de aguas de lastre y del químico tributiltina (TBT),por sus implicancias respectivas para la contaminaciónbiológica y de masculinización de las poblaciones degasterópodos (Carlton 1999; Penchaszadeh et al. 2001).
Defeo et al. (1997) analizaron el impacto del CanalAndreoni sobre los gasterópodos de la playa arenosa LaCoronilla-Barra del Chuy, el cual afecta negativamente atoda la comunidad de invertebrados de esta última (verLercari & Defeo en este volumen y Defeo et al. en estevolumen). Para esta zona Scarabino (2004a) reportó unadeclinación poblacional para O. contortuplicata, y la nece-sidad de evaluar el estatus de conservación de O. teagueiy O. formicacorsii. Pese al mencionado impacto, el o losfactores involucrados en la declinación de O. contortuplicatason inciertos, pudiendo estar involucrados también unamortandad masiva y la intensa explotación por parte decoleccionistas (Scarabino 2004a).
Adelomelon beckii es intensamente explotado con finesornamentales aunque sin ninguna regulación. Esta situa-ción, junto a los particulares aspectos biológicos de estaespecie (i.e. crecimiento lento, baja densidad, desarrollodirecto), hacen muy proclives al colapso a sus poblacio-nes. Resulta entonces urgente implementar medidas demanejo para este volútido (Scarabino 2004a).
La sobrexplotación de P. brasiliana en la costa de Ro-cha es producto de varias carencias vinculadas a la ad-ministración de las pesquerías de Volutidae en Uruguay(ver Fabiano et al. 2000; Riestra et al. 2000; Masello 2000),la mayoría de las cuales aún subsisten.
Otras implicancias o actividades de conservación ymanejo necesarias para este grupo coinciden con las dis-cutidas por Scarabino (en este volumen) para el resto delos invertebrados bentónicos de la costa uruguaya.
AGRADECIMIENTOSLas siguientes personas e instituciones contribuye-
ron a este trabajo brindando: acceso a colecciones y usode equipamiento (A. Toscano, E. Cohen, MUNHINA,Centro Interdisciplinario para el Desarrollo -CID-,Karumbé y DINARA), oportunidades y amplia colabo-ración durante muestreos (P. Puig, Y. Marín, E. Chiesa,A. Masello, O. Defeo, L. Delfino, S. Maytía, M. Laporta,A. Estrades, A. Fallabrino) y observaciones realizadas(M. Aguilera, J. Chocca, M. Demicheli, P. Etchegaray, R.Fontaina, P. Miller, A. Reboledo, C. Romero, A. Segura,M. Zarucki, Karumbé). A todos ellos nuestro sincero agra-decimiento. FS agradece a V. Scarabino, P. Bouchet y A.Warén, quienes ayudaron a través de su experiencia a lapercepción de varios aspectos aquí planteados. A. Waréntambién confirmó la identificación de Microeulima sp. Partede los muestreos considerados fueron efectuados en elmarco del Proyecto ECOPLATA III. A. C. agradece a laCSIC (Comisión Sectorial de Investigación Científica), aPEDECIBA y a la Fundación Whitley Laing (RuffordSmall Grants for Nature Conservation) por las financia-ciones que permitieron la recopilación y análisis de infor-mación existente de relevancia para este artículo, así comola realización de muestreos. Este trabajo se pudo realizargracias al apoyo del CID y Karumbé, y especialmente M.Amato, quien proporcionó facilidades indispensablespara su concreción.
REFERENCIASAbbott RT 1968 The helmet shells of the World (Cassidae). Part I.
Indo-Pacific Mollusca 2(9):7-202Absalão RS & RF Louro 2002 Variação morfológica da genitália
masculina nas espécies do complexo Littorina ziczac (Mollusca,Gastropoda, Littorinidae) procedentes da Ilha Grande, Rio deJaneiro, Brasil. Biociências 10(1):115-125. Porto Alegre
Absalão RS & RAP Roberg 1999 Complexo Littorina ziczac (Gmelin)(Mollusca, Gastropoda, Caenogastropoda) no litoral fluminense:análise morfométrica, distribuição vertical e bioquímica. Re-vista Brasileira de Zoologia 16(2):381-395
Absalão RS & RS Gomes 2001 The species usually reported in thesubgenus Brochina (Caecum, Caecidae, Caenogastropoda) from

Gasterópodos marinos y estuarinos de la costa uruguaya:faunística, distribución, taxonomía y conservación
152
Brazil and some relevant type specimens from western Atlantic.Bolletino Malacologico 37(1-4):9-22
Allmon WD 1990 Review of the Bullia Group (Gastropoda:Nassariidae) with comments on its evolution, biogeography,and phylogeny. Bulletins of American Paleontology 99(335):179 pp
Amaro J 1974 Noticia sobre una campaña exploratoria a bordo de“Striker” durante el verano de 1971-1972. Boletín de la Comi-sión Nacional de Oceanografía 1(1):15-17. Montevideo
Amaro-Padilla J 1967 El mejillón de la Bahía de Maldonado. Revis-ta del Instituto de Investigaciones Pesqueras 2(1):81-93. Mon-tevideo
Andrade SCS Magalhães CA & VN Solferini 2003 Patterns ofgenetic variability in Brazilian littorinids (Mollusca): amacrogeographic approach. Journal of Zoological Systematics& Evolutionary Research 41(4):249-255
Bandel K & D Kadolsky 1982 Western Atlantic species ofNodilittorina (Gastropoda: Prosobranchia): comparativemorphology and its functional, ecological, phylogenetic andtaxonomic implications. The Veliger 25:1-42
Barattini LP 1951 Malacología uruguaya. Enumeración sistemáti-ca y sinonímica de los moluscos del Uruguay. PublicacionesCientíficas del SOYP (Servicio Oceanográfico y de Pesca)(6):179-293. Montevideo
Barattini LP & EH Ureta 1961 (“1960”) La fauna de las costas deleste (invertebrados). Publicaciones de Divulgación Científica,Museo “Dámaso Antonio Larrañaga”, 195 pp. Montevideo
Batallés LM 1983 La comunidad de mejillón Mytilus edulis platensis(d´Orbigny, 1846) de Punta del Chileno (Maldonado Uruguay):distribución, composición y estructura de la población. Tesisde Licenciatura en Oceanografía Biológica, Facultad de Huma-nidades y Ciencias (Universidad de la República), Montevi-deo. 73 pp (Inédita)
Batallés LM García V & A Malek 1985 Observaciones sobre lazonación en el litoral rocoso de la costa uruguaya. I. Recono-cimiento de los niveles superiores del sistema litoral: CaboPolonio (Depto. de Rocha, Uruguay). Contribuciones del Depto.de Oceanografía de la Facultad de Humanidades y Ciencias2(2):42-50. Montevideo
Bequaert JC 1943 The genus Littorina in the Western Atlantic.Johnsonia 1(7):1-27
Bergh R 1884 Report of the Nudibranchia dredged by H.M.S.Challenger during the years 1873-1876. Report of the ScientificResults of the Voyage H.M.S. Challenger during the years1873-76 under the command of Captain George S. Nares, R.N., F. R. S. and the late Captain Frank Tourle Thomson, Zoology,10:1-154, 14 láms.
Beu AG 1978 The marine fauna of New Zealand: the molluscangenera Cymatona and Fusitriton (Gastropoda, FamilyCymatiidae). New Zealand Oceanographic Institute Memoir(65):43 pp.
Bieler R & PM Mikkelsen 1988 Anatomy and reproductive biologyof two Western Atlantic species of Vitrinellidae, with a case ofprotandrous hermaphroditism in the Rissoacea. The Nautilus102(1):1-29
Bier R 1985 Estudio de la macrofauna bentónica del curso inferiordel Arroyo Solís Grande (Canelones-Maldonado, Uruguay).Tesis de Licenciatura en Oceanografía Biológica, Facultad deHumanidades y Ciencias (Universidad de la República),Montevideo. i-v+140 pp (Inédita)
Bouchet P 1984 Les Triphoridae de Mediterranee et de la procheatlantique (Mollusca, Gastropoda). Lavori S.I.M (Atti dei Simposio“Sistematica dei Prosobranchi del Mediterraneo”) (Bologna, 24-26 de setiembre de 1982) 21:5-8. Milano
Bouchet P & A Warén 1986 Revision of the northeastern Atlanticbathyal and abyssal Aclididae, Eulimidae, Epitoniidae (Mollusca,Gastropoda). Bolletino Malacologico, Supplemento 2:299-576
Burch JQ & RL Burch 1964 The genus Agaronia J. E. Gray, 1839.The Nautilus 77(4):110-114 lám 6 y 7
Bush KJ 1900 Description of new species of Turbonilla of theWestern Atlantic fauna, with notes on those previously known.Proceedings of the Academy of Natural Sciences ofPhiladelphia:145-177 lám 8
Cachés MA 1973 Volvulella persimilis (Morch, 1875), Eulima bifasciatad’Orbigny, 1842 y Stylocheilus citrinus (Rang, 1828), tres nue-vas especies para aguas uruguayas. Comunicaciones de laSociedad Malacológica del Uruguay 3(25):295-298, 1 lám
Cachés MA 1980 Nota sobre la biología de los depósitos fangososcircalitorales frente a Punta del Este, Uruguay. Boletim doInstituto Oceanográfico 29(2):73-74. San Pablo
Carcelles AR 1953 Nuevas especies de gastrópodos marinos delas Repúblicas Oriental del Uruguay y Argentina. Comunica-ciones Zoológicas del Museo de Historia Natural de Montevi-deo 70(4):1-16, 5 lám
Carcelles AR & JJ Parodiz 1939 Dorsaninae argentinas y urugua-yas. Physis 17(49):745-769. Buenos Aires
Carranza A Scarabino F & L Ortega En prensa. Distribution oflarge benthic gastropods in the Uruguayan continental shelfand the Río de la Plata estuary. Journal of Coastal Research
Castellanos ZA de 1970 (“1967”) Catálogo de los moluscos mari-nos bonaerenses. Anales de la Comisión de InvestigaciónCientífica de la Provincia de Buenos Aires 8:1-365
Castellanos ZA de 1982 Los Pyramidellidae de la República Argen-tina (Moll. Entomotaeniata). Comunicaciones del Museo Argen-tino de Ciencias Naturales “Bernardino Rivadavia”, Hidrobiología2(7):61-85. Buenos Aires
Castellanos ZA de & D Fernández 1965 Consideraciones sobrelas especies del género Olivella y una nueva especie para laArgentina (Mollusca, Gastropoda). Neotrópica 11 (36):101-104.La Plata
Castellanos ZA de & D Fernández 1967 Nuevos aportes a losPyrenidae de Argentina (Moll. Gastropoda). Neotrópica 13(41):49-52. La Plata
Clench WJ & RD Turner 1951 The genus Epitonium in the WesternAtlantic. Part I. Johnsonia 2(30):249-287
Clench WJ & RD Turner 1952 The genera Epitonium (Part II),Depressiscala, Cylindriscala, Nystiella and Solutiscala in theWestern Atlantic. Johnsonia 2(31):289-356
Collin R 2000 Phylogeny of the Crepidula plana (Gastropoda:Calyptraeidae) cryptic species complex in North America.Canadian Journal of Zoology 78:1500-1514
Collin R 2005 Development, phylogeny, and taxonomy ofBostrycapulus (Caenogastropoda; Calyptraeidae), an ancientcryptic radiation. Zoological Journal of the Linnean Society144:75-101
Costa PMS 2005 Estudo taxonômico dos representantes da FamíliaColumbellidae Swainson, 1840 (Mollusca, Caenogastropoda)da costa brasileira. Tesis de Doctorado, Universidade Federaldo Rio de Janeiro-Museu Nacional, Rio de Janeiro. I-xx+291 pp(Inédita)
da Silva MCP & IL Veitenheimer-Mendes 2004a Nova espécie deHeleobia (Rissooidea, Hydrobiidae) da planicie costeira do suldo Brasil. Iheringia (Série Zoología) 94(1):89-94. Porto Alegre
da Silva MCP & IL Veitenheimer-Mendes 2004b Transferência dogênero monotípico Parodizia Medina de Bithyniidae (Gastropoda,Prosobranchia) para Pyramidellidae (Gastropoda,Heterobranchia). Revista Brasileira de Zoologia 21(2):277-280

F SCARABINO, JC ZAFFARONI, A CARRANZA, C CLAVIJO & M NIN
153
d’Orbigny A 1834-1847 Voyage dans l’Amérique méridionale (leBrésil, la République orientale de l’Uruguay, la RépubliqueArgentine, la Patagonie, la République du Chili, la Républiquede Bolivia, la République du Pérou), exécuté pendant les années1826, 1827, 1828, 1829, 1830, 1831, 1832 et 1833, 5(3):Mollusques:i-xlii 1-758 lám 1-85. París/Estrasburgo
Defeo O Brazeiro A & G Riestra 1997 (“1996”) Impacto de ladescarga de un canal artificial en la biodiversidad degasterópodos en una playa de arena de la costa atlánticauruguaya. Comunicaciones de la Sociedad Malacológica delUruguay 8(70/71):13-18
Defeo O Jaramillo E & A Lyonnet 1992 Community structure andintertidal zonation of the macroinfauna in the Atlantic coast ofUruguay. Journal of Coastal Research 8:830-839
Demicheli MA 1986 (“1984”) Estudios exploratorios del infralitoralde las playas arenosas uruguayas. I. Playa Portezuelo. Comu-nicaciones de la Sociedad Malacológica del Uruguay 6 (47):235-241, 4 mapas
Demicheli MA 1987b (“1985”) Estudios exploratorios del Infralitoralde las playas arenosas uruguayas: III, Playa Anaconda. Co-municaciones de la Sociedad Malacológica del Uruguay6(49):301-309, 3 mapas
Escofet A Gianuca N Maytía S & V Scarabino 1979 Playas areno-sas del Atlántico Sudoccidental entre los 29º y 43º S: conside-raciones generales y esquema biocenológico. Pp 245-258 In:Memorias del Seminario sobre Ecología Bentónicas y Sedi-mentación de la Plataforma Continental del Atlántico Sur (Mon-tevideo, 9-12 de mayo de 1978). UNESCO ROSTLAC
Fabiano G Riestra G Santana O Delfino E & R Foti 2000 Consi-deraciones sobre la pesquería del caracol fino Zidona dufresnei(Mollusca, Gastropoda) en el Uruguay: Período 1996-1998. Pp114-142 In: Rey (ed) Recursos pesqueros no tradicionales:moluscos bentónicos marinos. Proyecto URU/92/003. INAPE-PNUD, Montevideo
Figueiras A 1964 La malacofauna dulceacuícola del Uruguay.Ensayo de catálogo sistemático y sinonímico. Comunicacio-nes de la Sociedad Malacológica del Uruguay 1(7):161-202
Figueiras A & OE Sicardi 1968 Catálogo de los moluscos marinosdel Uruguay (Parte II). Comunicaciones de la SociedadMalacológica del Uruguay 2(15):255-273, 2 lám
Figueiras A & OE Sicardi 1971 (“1970”) Catálogo de los moluscosmarinos del Uruguay. Parte V. Comunicaciones de la SociedadMalacológica del Uruguay 3(19):25-347 lám 8
Figueiras A & OE Sicardi 1972 (“1971”) Catálogo de los moluscosmarinos del Uruguay. Parte VI. Comunicaciones de la Socie-dad Malacológica del Uruguay 3(21):101-127 lám 9-11
Figueiras A & OE Sicardi 1973a (“1972”) Catálogo de los moluscosmarinos del Uruguay. Parte VII. Comunicaciones de la Socie-dad Malacológica del Uruguay 3(22):169-186 lám 12-13
Figueiras A & OE Sicardi 1973b Catálogo de los moluscos mari-nos del Uruguay. Parte VIII. Comunicaciones de la SociedadMalacológica del Uruguay 3(25):259-286 lám 14-17
Figueiras A & OE Sicardi 1974 Catálogo de los moluscos marinosdel Uruguay. Parte IX. Comunicaciones de la SociedadMalacológica del Uruguay 3(26):323-352 lám 18-21
Figueiras A & OE Sicardi 1980 Catálogo de los moluscos marinosdel Uruguay. Parte X. Revisión actualizada de los moluscosmarinos del Uruguay con descripción de las especies agrega-das. Sección II - Gastropoda y Cephalopoda. Comunicacionesde la Sociedad Malacológica del Uruguay 5(38):179-272 lám 3-6
Gaillard MC 1974 Una nueva Littoridina de albufera. Neotrópica20(62):104-106. La Plata
García EF 2003 Gyroscala turnerae (Gastropoda: Epitoniidae), awestern Atlantic junior synonym of the Indo-Pacific Gyroscalaxenicima (Melvill & Standen, 1903). American Conchologist31(2):21-25
Giberto DA Bremec CS Acha EM & H Mianzan 2004 Large-scalespatial patterns of benthic assemblages in the SW Atlantic: theRío de la Plata estuary and adjacent shelf waters. Estuarine,Coastal and Shelf Science (61):1-13
Giménez L Borthagaray A Rodríguez M Brazeiro A & CDimitriadis 2005 Scale-dependent patterns of macrofaunaldistribution in soft-bottom sediment intertidal habitats along alarge-scale estuarine gradient. Helgoland Marine Research59:224-236
Gomes RS & RS Absalão 1996 Lista comentada e ilustrada dosCeacidae (Mollusca, Prosobranchia, Mesogastropoda) daOperação Oceanográfica GEOMAR XII. Revista Brasileira deZoologia 13(2):513-531
Hoagland RE 1983 Ecology and larval development of Crepidulaprotea (Prosobranchia: Crepidulidae) from southern Brazil: anew type of egg capsule for the genus. The Nautilus 97(3):105-109
Jorcin A 1999 Temporal and spatial variability in the macro-zoobenthic community along a salinity gradient in the CastillosLagoon (Uruguay). Archiv für Hydrobiologie 146(3):369-384
Juanicó M & M Rodríguez-Moyano 1976 (“1975”) Composiciónfaunística de la comunidad de Mytilus edulis platensis d’Orbigny,1846, ubicada a unas 55 millas al SE de La Paloma.Comunicaciones de la Sociedad Malacológica del Uruguay4(29):113-116
King PP & WJ Broderip 1832 Description of the Cirrhipeda,Conchifera and Mollusca, in a collection formed by the officersof H.M.S Adventure and Beagle employed between the years1826 and 1830 in surveying the Southern Coasts of SouthAmerica, including the Straits of Magalhaens and the Coast ofTierra del Fuego. Zoological Journal 5:332-349
Klappenbach MA 1961 Una nueva especie de Buccinanops de lacosta atlántica uruguaya y sudbrasileña (Moll. Gast.). Neotrópica7(24):87-91. La Plata
Klappenbach MA 1962 Nuevo subgénero y especie de Olivella dela costa atlántica del Uruguay. Archiv für Molluskenkunde 91(1/3):95-98
Klappenbach MA 1964a A new species of Olivancillaria fromUruguay and Brazil. The Nautilus 77(4):132-134
Klappenbach MA 1964b La familia Caecidae (Moll. Gastr.) enaguas uruguayas. I. Dos especies descritas por el Marqués DeFolin. Comunicaciones de la Sociedad Malacológica del Uru-guay 1(6):145-149, 1 fig.
Klappenbach MA 1965 Consideraciones sobre el género Olivancillariad’Orbigny, 1840 (Moll. Gastr.) y descripción de dos nuevasespecies de aguas argentinas y uruguayas. ComunicacionesZoológicas del Museo de Historia Natural de Montevideo8(104):1-10, 2 lám
Klappenbach MA 1967 Nuevos hallazgos de Polystira formosissima(E. A. Smith, 1915) (Moll. Gastropoda) en la costa atlánticasudamericana. Neotrópica 13(41):73-76. La Plata
Klappenbach MA 1991a Olivella riosi (Mollusca, Gastropoda,Olividae) nueva especies obtenida en aguas de la plataformacontinental uruguaya y del extremo sur del Brasil.Comunicaciones Zoológicas del Museo de Historia Natural deMontevideo 12(175):1-9
Klappenbach MA 1991b (“1989”) Notas sobre Olivella Swainson,1831. I. Comentarios sobre nomenclatura en dos especies deOlivella (Moll. Gastr.) del Atlántico Sur. Comunicaciones de laSociedad Malacológica del Uruguay 7(56/57):117-122

Gasterópodos marinos y estuarinos de la costa uruguaya:faunística, distribución, taxonomía y conservación
154
Kosuge S 1966 The family Triphoridae and its systematic position.Malacologia 4(2):297-324
Layerle C & Scarabino 1984 Moluscos del frente marítimo uruguayoentre los 9 y 78 m de profundidad: análisis biocenológico.Contribuciones (Departamento de Oceanografía, Facultad deHumanidades y Ciencias) 1(9):1-17. Montevideo
Marshall BA 1978 Cerithiopsidae (Mollusca: Gastropoda) of NewZealand, and a provisional classification of the family. NewZealand Journal of Zoology 5:47-120
Masello A 2000 Análisis histórico de la pesquería de caracol en elUruguay. Período 1991-1997. Pp 82-113 In: Rey (ed) Recursospesqueros no tradicionales: moluscos bentónicos marinos.Proyecto URU/92/003. INAPE-PNUD, Montevideo
Maytía S & V Scarabino 1979 Las comunidades del litoral rocosodel Uruguay: zonación, distribución local y consideracionesbiogeográficas. Pp 149-160 In: Memorias del Seminario sobreEcología Bentónicas y Sedimentación de la PlataformaContinental del Atlántico Sur (Montevideo, 9-12 de mayo de1978). UNESCO, ORCYT
Métivier B 1967 Mollusques Prosobranches: Fissurellidae, Acmaeidaeet Patellidae. Campagne de la Calypso au large des côtesatlantiques de l´Amérique du Sud (1961-1962). I. Annales del’Institute Océanographique 47:115-125, 1 lám
Milstein A Juanicó M & J Olazarri 1976 Algunas asociacionesbentónicas frente a las costas de Rocha, Uruguay. Resulta-dos de la campaña del R/V “Hero”, viaje 72-3A. Comunicacio-nes de la Sociedad Malacológica del Uruguay 4(50):143-164
Mol JJ van Tursch B & M Kempf 1967 Les Conidae du Bresil.Etude basée en partie sur les spécimens recueillis par la Calypso.Annales de l’Institut Océanographique 45(2):233-255, lám 5-10. Paris
Muniz P & N Venturini 2001 Spatial distribution of themacrozoobenthos in the Solís Grande stream estuary (Canelo-nes-Maldonado, Uruguay). Brazilian Journal of Biology 61(3):409-420
Neirotti E 1981 Estudio comparativo de supralitoral y mesolitoralrocoso en diferentes localidades del estuario del Río de laPlata. Comunicaciones de la Sociedad Malacológica del Uruguay5(40):347-370
Nion H 1979 Zonación del macrobentos en un sistema lagunarlitoral oceánico. Pp 225-235 In: Memorias del Seminario sobreEcología Bentónicas y Sedimentación de la PlataformaContinental del Atlántico Sur (Montevideo, 9-12 de mayo de1978). UNESCO, ORCYT
Obenat S Ferrero L & E Spivak 2001 Macrofauna associated withPhyllochaetopterus socialis aggregations in the southwesternAtlantic. Vie et Milieu 51(3):131-139
Odhner NH 1926 Die Opisthobranchien. Further Zoological Resultsof the Swedish Antarctic Expedition, 1901-1903, under thedirection of Dr. Otto Nordeskjold 2:1-100, 3 lám
Orensanz JM Schwindt E Pastorino G Bortolus A Casas GDarrigrán G Elías R López-Gappa JJ Obenat S Pascual MPenchaszadeh P Piriz ML Scarabino F Spivak ED & EAVillarino 2002 No longer the pristine confines of the worldocean: a survey of exotic marine species in the southwesternAtlantic. Biological Invasions 4:115-143
Parodiz JJ 1939 Las especies de Crepidula de las costas argenti-nas. Physis 17(49):685-709, 1 lám. Buenos Aires
Passadore C & L Giménez 2003 Composición y variación intra-anual de la macroinfauna de la desembocadura del arroyoPando. Publicación Especial de la Sociedad Zoológica del Uru-guay (Actas de las VII Jornadas de Zoología del Uruguay, IEncuentro de Ecología del Uruguay):125
Pastorino G 2005a A revision of the genus Trophon Montfort, 1810(Gastropoda: Muricidae) from southern South America. TheNautilus 119(2):55-82
Pastorino 2005b The Recent Naticidae (Mollusca: Gastropoda)from the Patagonian coast. The Veliger 47(4):225-258
Penchaszadeh PE Averbuj A & M Cledón 2001 Imposex in gastro-pods from Argentina (south-western Atlantic). Marine PollutionBulletin 42(9):790-791
Pereira de Medina N 1959 “Syrnolopsinae” en el Río de la Plata,República Oriental del Uruguay. Neotrópica 5(17):51-55. LaPlata
Pilsbry HA 1897a List of mollusks collected in Maldonado Bay,Uruguay, by Dr. Wm. H. Rush, U. S. N. The Nautilus 11(1):6-9
Pilsbry HA 1897b New species of mollusks from Uruguay.Proceedings of the Academy of Natural Sciences ofPhiladelphia 49:290-298 lám 6 y 7
Pimenta AD & RS Absalão 2001a The genera Bacteridium Thiele,1929 and Careliopsis Mörch, 1875 (Gastropoda: Pyramidellidae)from the east coast of South America. Bolletino Malacologico37(1-4):41-48
Pimenta AD & RS Absalão 2001b Taxonomic revision of the speciesof Turbonilla Risso, 1826 (Gastropoda, Heterobranchia,Pyramidellidae) with type localities in Brazil, and description ofa new species. Basteria 65(1/3):69-88
Pimenta AD & RS Absalão 2004 Fifteen new species and ten newrecords of Turbonilla Risso, 1826 (Gastropoda, Heterobranchia,Pyramidellidae) from Brazil. Bolletino Malacologico 39(5-8):113-140
Piñeiro G Scarabino F & M Verde 1992 Una nueva localidadfosilífera del Holoceno marino del Uruguay (Punta Rasa, De-partamento de Maldonado). Boletín de la Sociedad Zoológicadel Uruguay (Actas de las III Jornadas de Zoología del Uru-guay) 7(2ª serie):61-62
Riestra G Giménez JL & V Scarabino 1992 Análisis de la comu-nidad macrobentónica infralitoral de fondo rocoso en Isla Gorritie Isla de Lobos (Maldonado, Uruguay). Frente Marítimo 11:123-127. Montevideo
Riestra G Defeo O & A Brazeiro 1997 (“1996”) Dinámica de lazonación de gasterópodos en una playa disipativa micromareal.Comunicaciones de la Sociedad Malacológica del Uruguay 8(70-71):1-6
Riestra G Fabiano G & O Santana 2000 El caracol negro Adelomelonbrasiliana como recurso no tradicional de importancia para elpaís: análisis socioeconómico de la pesquería y medidasprecautorias de manejo. Pp 82-92 In: Rey (ed) Recursospesqueros no tradicionales: moluscos bentónicos marinos.Proyecto URU/92/003. INAPE-PNUD, Montevideo
Rios EC 1994 Seashells of Brazil. Museu Oceanográfico da FundaçãoUniversidade do Rio Grande, 329 pp, 102 lám
Roberston R 1981 (“1980”) Epitonium millectostatum and Coralliophilaclathrata: two prosobranch gastropods symbiotic with Indo-Pacific Palythoa (Coelenterata: Zoanthidae). Pacific Science34(1):1-17
Robertson R 1983 Observations on the life history of the wentletrapEpitonium albidum in the West Indies. American MalacologicalUnion 1:1-12
Robertson R & T Mau-Lastovicka 1979 The ectoparasitism ofBoonea and Fargoa (Gastropoda: Pyramidellidae). BiologicalBulletin 157:320-333
Rodríguez-Moyano M 1976 (“1975”) Drillia braziliensis (E. A. Smith,1915), nueva especie del género Drillia Gray, 1838 para aguasuruguayas. Comunicaciones de la Sociedad Malacológica delUruguay 4(28):93-94

F SCARABINO, JC ZAFFARONI, A CARRANZA, C CLAVIJO & M NIN
155
Rojas A & S Martínez 1999 Restricción actual en la distribucióngeográfica de algunos gastrópodos y bivalvos del Cuaternariodel Uruguay. Publicación Especial de la Sociedad Zoológica delUruguay (Actas de las V Jornadas de Zoología del Uruguay):38
Rolán E & F Fernandes 1990 The genus Seila A. Adams, 1861(Mollusca, Gastropoda, Cerithiopsidae) in the Atlantic Ocean.Apex 5(3-4):17-30. Bruxelles
Scarabino F 1999 Inventario de los moluscos bentónicos del Ríode la Plata y su Frente Marítimo: estado actual del conocimien-to, necesidades y perspectivas de estudio. Boletín de la So-ciedad Zoológica del Uruguay (Actas de las V Jornadas deZoología del Uruguay) 11 (Segunda Epoca):39
Scarabino F 2004a Conservación de la malacofauna uruguaya.Comunicaciones de la Sociedad Malacológica del Uruguay 8(82/83):267-273
Scarabino F 2004b Lista sistemática de los Gastropoda marinos yestuarinos vivientes de Uruguay. Comunicaciones de la So-ciedad Malacológica del Uruguay 8(84/85-86/87):305-346
Scarabino F & A Masello 1996 Dos moluscos interesantes encon-trados en un sector de la costa oeste de Montevideo: Parodiziauruguayensis (Gastropoda) y Limnoperna fortunei (Bivalvia).Conferencia Internacional ECOPLATA´96: Hacia el desarrollosostenible de la zona costera del Río de la Plata (Montevideo,25-27 de noviembre de 1996). Resúmenes de trabajos cientí-ficos presentados por el Proyecto ECOPLATA: 27
Scarabino F & L Ortega 2004 Registros uruguayos de Aulacomyaatra atra (Bivalvia: Mytilidae): rol de condiciones oceanográficasanómalas y de dispersión por feofitas flotantes. Comunicacio-nes de la Sociedad Malacológica del Uruguay 8(84/85-86/87):299-304
Scarabino F & JC Zaffaroni 2004 Estatus faunístico de veinteespecies de moluscos citadas para aguas uruguayas. Comu-nicaciones Zoológicas (Museo Nacional de Historia Natural yAntropología) 13(202):1-15. Montevideo
Scarabino F Clavijo M & M Laporta 2003 Moluscos del intermareal-submareal somero rocoso de Cerro Verde, Departamento deRocha. 2as Jornadas de Conservación y Uso Sustentable dela Fauna Marina (Montevideo, 1-3 de octubre de 2003), Libro deResúmenes:26
Scarabino F Menafra R & P Etchegaray 1999 Presencia de Rapanavenosa (Valenciennes, 1846) (Gastropoda: Muricidae) en el Ríode la Plata. Boletín de la Sociedad Zoológica del Uruguay(Actas de las V Jornadas de Zoología del Uruguay) 11 (Segun-da Epoca):40
Scarabino V 1984 Clave para el reconocimiento de moluscoslitorales del Uruguay, I. Gastropoda. Contribuciones del Depar-tamento de Oceanografía (Facultad de Humanidades y Cien-cias) 1(2):12-22. Montevideo
Scarabino V Maytía S & JC Faedo 1974 Zonación biocenológicade las playas arenosas del Depto. de Rocha (Uruguay), conespecial referencia a la presencia de Ocypode quadrata(Fabricius, 1787) (Decapoda, Brachyura). Boletín de la Comi-sión Nacional de Oceanología 1(1):42-52, 2 lám, 1 mapa. Mon-tevideo
Scarabino V Maytía S & M Cachés 1976 (“1975”) Carta bionómicalitoral del departamento de Montevideo I. Niveles superioresdel sistema litoral. Comunicaciones de la Sociedad Malacológicadel Uruguay 4(29):117-126, 3 lám
Simone LRL 2006 Morphological and phylogenetic study of theWestern Atlantic Crepidula plana complex (Caenogastropoda,Calyptraeidae), with the description of three new species fromBrazil. Zootaxa (1112):1-64
Simone LRL Pastorino G & PE Penchaszadeh 2000 Crepidulaargentina (Gastropoda: Calyptraeidae), a new species fromthe littoral of Argentina. The Nautilus 114:127-141
Sprechmann P 1978 The paleoecology and paleogeography of theUruguayan coastal area during the Neogene and Quaternary.Zitteliana 4:3-72 lám 1-6
Van der Linden J & RG Moolenbeek 2004 A survey of Graphisspecies from the West Indies with the description of two newspecies. Gloria Maris 43(2-3):1-13. Antwerpen
Valdés A & YE Camacho-García 2004 “Cephalaspidean”heterobranchs (Gastropoda) from the Pacific coast of CostaRica. Proceedings of the California Academy of Sciences55(26):459-497
Venturini N Muniz P & M Rodríguez 2003 Macrobenthic subtidalcommunities in relation to sediment pollution: the phylum-levelmeta-analysis approach in a south-eastern costal region ofSouth America. Marine Biology 144:119-126
Warén A 1992 Comments on and descriptions of eulimid gastro-pods from Tropical West America. The Veliger 35(3):177-194
Warén A 1993 New and little known Mollusca from Iceland andScandinavia. Part 2. Sarsia 78:159-20
Warén A 1994 Systematic position and validity of Ebala Gray,1847 (Ebalidae fam. n., Pyramidelloidea, Heterobranchia).Bolletino Malacologico 30(5-9):203-210
Watson RB 1886 Report on the Scaphopoda and Gasteropoda (sic)collected by the H.M.S. Challenger during the years 1873-76.Report of the Scientific Results of the Voyage H.M.S. Chal-lenger during the years 1873-76 under the command of Cap-tain George S. Nares, R. N., F. R. S. and the late CaptainFrank Tourle Thomson, Zoology, 15:i-v+1-756, 50 lám
Williams ST Reid DG & DT Littlewood 2003 A molecular phylogenyof the Littorininae (Gastropoda: Littorinidae): unequal evolu-tionary rates, morphological paralellism, and biogeography ofthe Southern Ocean. Molecular Phylogeny and Systematics28(1):60-86
Zaffaroni JC 1990 (“1988”) Presencia de Acteocina recta (d’Orbigny,1841) (Gastropoda: Cephalaspidea) en aguas uruguayas. Co-municaciones de la Sociedad Malacológica del Uruguay 7(54/55):75-77
Zaffaroni JC 1991 (“1989”) Eulimella bermudensis (Dall & Bartsch,1911) y Gastrochaena hians (Gmelin, 1791), primera cita paraaguas uruguayas. Comunicaciones de la Sociedad Malacológicadel Uruguay 7(56/57):123-128
Zaffaroni JC 1994 (“1989”) Ovatella myosotis (Draparnaud, 1801)en aguas uruguayas. Comunicaciones de la SociedadMalacológica del Uruguay 7(62/63):271-272
Zaffaroni JC 2000 (“1998”) Presencia de Modiolus carvalhoi(Mollusca, Pelecypoda) en aguas uruguayas. Comunicacionesde la Sociedad Malacológica del Uruguay 8(74/75):101-103
Zaffaroni JC En prensa. Tectonatica pusilla (Say, 1822) (Mollusca,Gastropoda): primera cita para aguas y depósitos holocenosuruguayos. Comunicaciones de la Sociedad Malacológica delUruguay

Gasterópodos marinos y estuarinos de la costa uruguaya:faunística, distribución, taxonomía y conservación
156




RESULTADOS
En el muestreo piloto realizado en Enero de 2007 se colectaron 13 taxa de macroinvertebradosbentónicos, siendo dominantes los crustáceos Loxopagurusloxochelis y Libinia spinosa(45,31 y 26,56% de los individuos colectados). Otros grupos bien representados fueron los moluscos (Pachycymbiola brasiliana, Buccinanops cochlidium y B. duartei), los cnidarios (Antholoba achates) y los equinodermos (Mellita quinquiesperforata). Además, se colectaron dos especies de decápodos, Hepatuspudibundus y Araneus cribariusy varias especies de peracáridos aun no identificados.
INVESTIGACIÓN, MONITOREO Y CONSERVACIÓN: INVERTEBRADOS BENTÓNICOS DEL AREA MARINA PROTEGIDA DE CERRO VERDE (ROCHA, URUGUAY)
Carranza, A. Investigación & Desarrollo - Facultad de Ciencias, Iguá 4225, C.P. 11400, Montevideo, URUGUAY
OBJETIVOS
Realizar un detallado inventario faunistico de la comunidad de invertebradosbentonicos en el area marina protegida de Cerro Verde
IMPLICACIONES PARA LA CONSERVACIÓN Y EL
MANEJO DEL AMP
La información obtenida durante este proyecto permitirá conocer en profundidad la diversidad de invertebrados bentónicos en el área y su variabilidad espacial y temporal, proveyendo una herramienta útil para el monitoreo y manejo del AMP.
MÉTODOS
Muestreos sistemáticos a bordo de embarcaciones artesanales, empleando los artes de pesca utilizados comúnmente por los pescadores locales (i.e. red camaronera) así como mediante el empleo de rastras epibentónicas.
Agradecimientos:
Tripulación del B/P “Domy”; Sub-Prefectura del Chuy
Actiniaria bermudensis Bunodosoma cangicum
Hepatus pudibundus Picnogonum pamphorum
Libinia spinosa
Antholoba achactes sobre Pachycymbiola brasiliana yBuccinanops cochlidium

METODOLOGMETODOLOGÍÍAA
En el marco del proyecto En el marco del proyecto ““PilotPilot programmeprogramme forforecosystemicecosystemic monitoringmonitoring in in Cerro Verde (Uruguay): Cerro Verde (Uruguay): benthicbenthic invertebratesinvertebrates as a as a tooltool forfor conservationconservation”” se se estestáán realizando muestreos n realizando muestreos estacionalesestacionales de la de la comunidad comunidad íícticactica. El dise. El diseñño o de muestreo se basa en de muestreo se basa en transectastransectas equidistantes equidistantes con lances de pesca tras la con lances de pesca tras la rompiente, a 5 y a 10 rompiente, a 5 y a 10 metros de profundidad. metros de profundidad. Los mismos se realizan a Los mismos se realizan a bordo de las chalanas de bordo de las chalanas de pesca artesanal de Punta pesca artesanal de Punta del Diablo utilizando una del Diablo utilizando una red tipo camaronera (t = red tipo camaronera (t = 1010´). El primer muestreo se ). El primer muestreo se realizrealizóó en enero de 2007, en enero de 2007, los peces fueron los peces fueron separados a nivel separados a nivel especifico y los individuos especifico y los individuos fueron medidos y pesados. fueron medidos y pesados. AdemAdemáás se extrajeron los s se extrajeron los estestóómagos para realizar magos para realizar estudios trestudios tróóficos ficos posterioresposteriores
La comunidad nectónica en el Área Marina Protegida de Cerro Verde
Luis Rubio1, Angel Segura1,2 & Alvar Carranza1.Facultad de Ciencias. Universidad de la República.
Instituto de Investigaciones Pesqueras. Facultad de Veterinaria. Universidad de la República.
INTRODUCCIÓN
Recientemente la zona de Cerro Verde se incorporó al Sistema Nacional de Áreas Protegidas (SNAP) como la primer Área Marina Protegida (AMP). Los pescadores de Punta del Diablo explotan recursos dentro del AMP. Por otro lado, las áreas marino-costeras son conocidas como zona de cría de peces. Los hábitos tróficos de los peces y la composición de la comunidad bentónica permite relacionar ambas comunidades a nivel ecosistémico. Sin embargo, no existen para ésta zona trabajos que describan la comunidad íctica.
RESULTADOSEn el muestreo de enero de 2007 se registraron un total de 23 especies (Tabla 1), en su mayoría con talla inferior a la de primera madurez. La comunidad estuvo dominada por algunas especies muy representadas y muchas especies poco abundantes (Figura 2).
OBJETIVOSOBJETIVOS
Describir la comunidad Describir la comunidad íícticactica de la zona costera de la zona costera en el AMP de Cerro en el AMP de Cerro Verde, y su variaciVerde, y su variacióón n espacioespacio--temporal.temporal.
Describir los hDescribir los háábitos bitos trtróóficos de las principales ficos de las principales especies de peces especies de peces presentes en la zona.presentes en la zona.
Agradecimientos:
Tripulación del B/P “Domy” ; Sub-prefectura del Chuy
IMPLICACIONES PARA LA IMPLICACIONES PARA LA CONSERVACICONSERVACIÓÓN Y EL N Y EL
MANEJO DEL AMPMANEJO DEL AMP
ÉÉste es el primer proyecto ste es el primer proyecto de investigacide investigacióón enfocado n enfocado en la comunidad en la comunidad íícticactica de de la zona y su variacila zona y su variacióón n espacioespacio--temporal. temporal.
En el primer muestreo se En el primer muestreo se registrregistróó un alto nun alto núúmero de mero de especies. La gran especies. La gran proporciproporcióón de juveniles n de juveniles aporta evidencia a favor de aporta evidencia a favor de la hipla hipóótesis de la franja tesis de la franja costera como costera como áárea de crrea de críía a de peces, aumentando el de peces, aumentando el interinteréés en proteger su s en proteger su biodiversidad. biodiversidad.
El proyecto se estEl proyecto se estáállevando a cabo con la llevando a cabo con la cooperacicooperacióón de los n de los pescadores artesanales de pescadores artesanales de Punta de Diablo. Es crucial Punta de Diablo. Es crucial la participacila participacióón de la n de la comunidad local en las comunidad local en las investigaciones para que investigaciones para que las las AMPsAMPs tengan una tengan una viabilidad a largo plazo.viabilidad a largo plazo.
Es fundamental continuar Es fundamental continuar con estos monitoreos y con estos monitoreos y comprender los cambios comprender los cambios estacionalesestacionales de las de las comunidades para poder comunidades para poder sugerir posibles medidas sugerir posibles medidas de manejo basadas en el de manejo basadas en el ecosistema. ecosistema.
Tabla 1. Nº de individuos y biomasa de las especies capturadas en Cerro Verde en enero de 2007
Código Especie Nº individuos Biomasa (g)1 Macrodon ancylodon 143 114642 Paralonchurus brasiliensis 40 1330,33 Micropogonias furnieri 36 769,14 Urophysis brasiliensis 26 908,15 Sympterygia acuta 12 57656 Stromateus Brasiliensis 12 103,27 Menticirrhus americanus 7 323,48 Sympterygia Bonaparti 6 2016,49 Peprilus paru 5 35
10 Mustelus schmitti 3 273,211 Porichthys porossisimus 3 222,412 Lagocephalus laevigatus 3 9213 Oncopterus Darwinni 3 18,214 Symphurus spp 2 2215 Prionotus punctatus 2 11,116 Anchoa marinii 2 5,617 Paralichtys orbignianus 1 80018 Myliobatis godei 1 22519 Conger Orbignianus 1 22520 Squatina Gugenheim 1 17521 Psamobatis 1 10,822 Monacanthus ciliatus 1 1,823 Trichurus Lepturus 1 0,3
0
20
40
60
80
100
120
140
160
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23
Código (especie)
Nºi
ndiv
iduo
s
Figura 2. Composición de la comunidad íctica de Cerro Verde (enero 2007).

Este es el primer registro de ambas especies para la costa uruguaya y el registro mas austral de P. pamphorum, solo conocido de su localidad tipo (Bahía de Santos, Brasil). A. petiolatus, en cambio, ha sido reportado para la zona de Santos (Brasil), Mar del Plata, Tierra del Fuego y Canal de Beagle (Argentina) y para la costa sur chilena. Se insiste en el estudio adecuado de la biodiversidad y taxonomía de los grupos menos conocidos de invertebrados de la costa uruguaya, los cuales permanecen pobremente estudiados.
827826825
824823821
Dos nuevos registros de Pycnogonida para aguas uruguayas
Carranza, A.1, Borthagaray1, A. I. & Genzano, G. N. 2
1Investigación & Desarrollo - Facultad de Ciencias, Iguá 4225, C.P. 11400, Montevideo, URUGUAY; 2 Departamento de Ciencias Marinas, Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Mar del Plata, Funes 3250 (7600)
Mar del Plata, ARGENTINA
Chelicerata y Pycnogonida
Los quelicerados constituyen probablemente el grupo mas homogéneo de artrópodos, aunque hay problemas de definición respecto a si los picnogonidos constituyen un grupo de quelicerados, o bien un grupo de artrópodos independiente. Generalmente, los picnogonidos se clasifican como el grupo hermano o como un grupo interno de los quelicerados. Sin embargo, aunque en algunos casos se ha dudado de su condición artropodiana, varios análisis moleculares que han incluido datos de picnogonidos indican que constituyen claramente un grupo de artrópodos, probablemente grupo hermano de los quelicerados, o bien grupo hermano del resto de artropodos, dependiendo de los parámetros utilizados en los analisis basados unicamente en datos moleculares. La posicion de los picnogonidos parece ser inestable en dichos analisis moleculares basados en genes ribosomicos.
DISCUSIÓN
INTRODUCCION
Los Pycnogonida de aguas uruguayas han recibido hasta el presente escasa atención. En particular, los antecedentes existentes se refieren a material colectado por los cruceros de los B.I “Calypso” y “Walter Hewig” en la plataforma continental y a la descripción de Colosendeis geofroyi (Mañé-Garzón, 1944). Sin embargo, la fauna de pycnogonidos del litoral rocoso permanece mayormente inexplorada.
Temperatura
Salinidad
Algunas hipótesis de las relaciones entre Pycnogonida y el resto de los artrópodos
Agradecimientos:
Este trabajo fue financiado por
Rufford Small Grants for Nature
Conservation
RESULTADOS
En tal sentido, en este trabajo se reporta el hallazgo de varios ejemplares de Pycnogonida en el litoral rocoso de Cerro Verde (Rocha, Uruguay). Los especimenes fueron asignados a Pycnogonum pamphorum Marcus, 1940 (Pycnogonidae) y Anoplodactylus petiolatus(Kroyer, 1844) (Phoxichilidiidae). Los 6 especimenes de P. pamphorum (1 macho y 5 hembras) se encontraron en 3 muestras (frecuencia =36.4%) en el intermareal inferior en los sitios 1 y 2 mientras que 12 especimenes de A. petiolatus (8 machos y 4 hembras) se colectaron en tres muestras (100%) del submareal somero expuesto y una muestra (18.2 %) del intermarealinferior del sitiointermedio. La macrofauna bentónicaasociada incluyó a losbivalvos B. rodriguezii, Perna perna y Mytilus edulis platensis, variasespecies de Anthozoa, poliquetos y lapas
Pycnogonum pamphorumMarcus, 1940
Anoplodactyluspetiolatus (Kroyer, 1844)
Expuesto
Protegido
Intermedio
Puntos de Muestreo
Related Documents











